
ANTI-TUMOR ACTIVITY OF FRUIT EXTRACTS OF MOMORDICA DIOICA ROXB
Upload
independentCategory
view
2download
0
192 MARGARETA WELANDER PhysioL Plant. 30: 192-199. 1974
Enzyme Activities in Urtica dioica: Effects of Daylength and Leaf Ageon Glutamate Dehydrogenase, Aspartate Aminotransferase
and Alanine Aminotransferase
By
MARGARETA WELANDER
Department of Plant Physiology, Solvegatan 35, Fack, S-220 07 Lund 7, Sweden
(Received June 16, 1973)
Abstract
Enzymes, important to protein synthesis, were investigated inyoung and old leaves of Urtica dioiea. The plants, divided intotwo groups, were exposed to either 18-hour or 12-hour photo-periods. One group of plants from each pholoperiodic regime wassubjected to an irradiance of 28 W x m" •', and the other group ofplants to 42 W X m ^. The enzymes investigated were glutamatedehydrogenase (GDH), aspartate aminotransferase (glutamate-oxaloacetate transaminase, GOT), and alanine aminotransferase(glutamate-pyruvate transaminase, GPT). GDH and GOT weredetermined by means of electrophoretic separation on poly-acrylamide and spectrophotometric measurements. GPT wasdetermined only by the latter method.
Plants exposed to 18-hour photoperiods showed much higherGDH activity than did those exposed to 12-hour photoperiods.The activity of GDH also increased with leaf age. Besides oneuniform NAD*-dependent GDH, two other NAD^-independentenzymes, showing GDH activity, were identified on polyacryl-amide gel electrophoresis. The distribution of NADH andNAD^-dependent GDH activity between young and old leaveswas similar under different growth conditions.
The activity of GOT was insensitive to environmental changes.The results regarding GPT indicate that this enzyme respondedto different photoperiods in the same way as GDH. A correlationcoefficient of 0.928 was obtained for the relationship betweenGDH and GPT activity.
Introduction
The aim of the present investigation was to study the effectof daylength and leaf age on enzymes important to proteinsynthesis and on protein content. The enzymes studied areglutamate dehydrogenase (E.C. 1.4.1.2; GDH), aspartateaminotransferase (E.G. 2.6.1.1) also named glutamate-oxaloacetate transaminase (GOT) and alanine aminotrans-
ferase (E.C. 2.6.1.2) also named glutamate-pyruvatetransaminase (GPT). GDH is of importance for the incor-poration of inorganic nitrogen into amino acids. GOT andGPT are important in transaminating processes to convertand produce amino acids.
The effects on the protein metabolism due to environ-mental change and stress have been reported in severalstudies. In experiments on Mentha piperita. Steward eT al.(1959) demonstrated that storage of soluble nitrogen inleaves was higher under short-day conditions than underlong-day conditions. This was accompanied by relativesuppression of protein synthesis. The main question iswhether the decrease in the protein content is due to proteo-lytic activity or to a lower rate of synthesis. Some evidenceshowing that a lower rate of synthesis is of importance, wasgiven for leaves from Perilla friitescens (Kannangara andWoolhouse 1968).
Changes in the transaminase activity in leaves of Loliumtemiilentum, exposed to either short or long days, wereestablished by Hedley and Stoddart (1971, 1972).
Isoenzymes of GDH were investigated by Yue (1969) inseveral different plants. The number of isoenzymes and theirelectrophoretic mobility varied between the different plantsstudied. Pahlich and Joy (1971) stated that, in pea roots,the enzyme showed both NADH- and NAD'^-dependentGDH activity, although the ratio of these activities was notconstant. However, the fundamental information aboutGDH effects on the regulatory mechanism in higher plants,is not sufficient.
Abbreviations. GDH: glutamic acid dehydrogenase; GOT:glutamate-oxaloacetate transaminase; GPT: glutamate-pyruvatetransaminase; NAD^, NADH: nicotinamide adenine dinucleo-tide oxidized and reduced; PVP: polyvinylpyrrolidone; TCA:trichloroacetic acid.

Physiol. Plant. 30. 1974 ENZYME ACTIVITIES IN URTICA DIOICA 193
Materials and Methods
Plant material
Urtica dioica L. (wild, south of Sweden) was used in allthe experiments. The seeds were sown in soil in boxes.Seedlings, 2-3 cm high, were planted in pots, 10 plants ineach. Two weeks later, 10 g of Weibull's Enpeka Specialfertilizer (12% N, 12% P2O5, 19% K2O and micro-nutrients) was applied to each pot. Cultivation of the plantswas performed in climate chambers under controlled condi-tions. The plants, divided into two groups, were exposed toeither 18-hour (long day) or 12-hour (short day) photo-periods. One group of plants from each photoperiodicregime was subjected to the irradiance of 28 W x m"^ (10,000lux), and the other group of plants to 42 Wx m"^ (15,000 lux).The light energy was supplied from cool-white fluorescentlamps (F 48 PG17/CWC, General Electric). The daytemperature was 22°C, and the night temperature ]2°C.Daylengths and irradiances were chosen so that the sameamount of light energy was received at 28 W x m"^ during18 hours, as at 42W x m"^ during 12 hours. The plants]werewatered daily with distilled water. Plants exposed to longdays were harvested 30 days after planting, and plants ex-posed to short days 10 days later. The cultivations wereperformed twice.
Sampling and extraction
The youngest (not fully expanded) leaves and the oldestones (without deficiency symptoms) of each plant wereused. The leaves were sampled simultaneously in order toavoid diurnal fluctuations in the enzyme level. All subse-quent operations were carried out at 0-5°C.
Four grams fresh weight of leaf material was frozen inliquid nitrogen and pulverized with a mortar and pestle.The pulverized material was mixed with 3 g of Polyclar AT(polyvinylpyrrolidone PVP) and 20 ml bufl"er consisting of0.1 M Tris-HCl, and 0.005 M mercaptoethanol at pH 7.5.The sample was ground with an Ultra-Turrax apparatus,which was rinsed with 10 ml bufler. The slurry was squeezedthrough double layers of cheese cloth. The filtrate wascentrifuged at 1000^ for 10 minutes, at 0°C. The precipitatewas discarded, and the resulting supernatant was centri-fuged at 48,000^ for 30 minutes. The proteins of the super-natant were then purifled from low-molecular weight com-ponents by passing through a Sephadex G25 column. Avolume of 20 ml of the purified sample was concentratedagainst aquacide I for 90 minutes. To this protein extractwas added dimethylsulfoxide to 25 % and mercaptoethanolto a final concentration of 0.005 M. By the addition of di-methylsulfoxide and mercaptoethanol it was then possibleto store the sample at -20°C without any loss of enzymeactivity.
Enzyme determinations
Qualitative and quantitative enzyme assays were carriedout by means of polyacrylamide gel electrophoresis andspectrophotometric measurements. All enzyme determina-tions were repeated. Good correlation was found betweenelectrophoretic and spectrophotometric measurements.
Polyacrylamide gel electrophoresis. The proteins in theextract were separated on 7 % polyacrylamide gels at pH 8.3,according to the method used by Davis (1964). A volume of100 n\, corresponding to 200-400 ng protein, was appliedto the top of the running gel.
The electrode buffer was mixed with bromphenol bluefor detection of the front after staining. The electrophoresiswas conducted at 2 mA per tube for 90 minutes. Theenzymes were detected by incubation in solutions containinga specific substrate and stain for each enzyme. For all thereactions, test solutions without substrate or cofactor servedas controls. The GDH activity was determined in the direc-tion of a-ketoglutarate production using NAD""" as a co-enzyme (Fine and Costello 1963). GOT was visualized byusing aspartate and a-ketoglutarate as substrates and fastviolet B as dye (Brewbaker et al. 1968). No method on gelswas found for GPT. After being stained, the gels werephotographed. Densitometer tracings of the gels wereobtained by use of a lin/log densitometer (Kipp & ZonenDD691-E). A red filter with maximum transmittance at580-650 nm was used for scanning the enzyme patterns ofGDH. A blue filter with maximum transmittance at 375-425 nm was used for GOT. Quantitative calculations fromdensitometer curves were carried out by measuring the curvearea. The densitometer curve area was directly proportionalto the amount of sample applied to the gel within the rangebetween 50 and 150 fi\ protein extract corresponding to50-400 fig protein (Figure 1).
3.0 -
25 150
protein extract
Figure 1. The densitometer curve area (relative units) as a functionof the amount of protein applied to the gel. (•) Aspartate amino-transferase, (•) glutamate dehydrogenase (NAD"^-dependent).

194 MARGARETA WELANDER Physiol. Plant. 30. 1974
Spectrophotometric measurements. A Beckman DK2spectrophotometer was used for all reactions describedbelow. The activity of GDH was determined in the directionof glutamate formation usingNADHasacoenzyme(Schmidt1963). Instead of triethanolamine buffer (with EDTA),Tris-HCl buffer was used, and EDTA was excluded, sincethis substance causes a decrease in the enzymatic activity(Lefranc, Welander and Vieira da Silva 1972). Calculationsof the GDH activity were made according to Biicher(Schmidt 1963).
The GOT activity was measured with aspartate and a-ketogiutarate as substrates. Keto acid production wasdefined by formation of dinitrophenylhydrazones andmeasurement of absorbance at 546 nm (Bergmeyer andBernt 1962). GTP was determined in the same way as GOT,but alanine was used as substrate instead of aspartate.
For .soluble protein determination, the method employedby Potty (1969) was used after TCA-precipitation of theproteins. Kjeldahl analysis of the total nitrogen could notbe used because PVP (included in the grinding buffer) isnitrogenous. Phenols give a positive reaction with theLowry method (1951) so that the method was not applicable,since most plant material contains large amounts of phenols.
The non-protein a-amino nitrogen was determined colori-metrically according to Moore and Stein, as described byLexander et al. (1970). The total protein nitrogen of the drymatter was determined according to Kjeldahl after TCAprecipitation.
ResultsFigure 2 indicates that the amount of protein per gram
dry weight was larger in young leaves than in old ones,whereas the amount of non-protein a-amino nitrogen in-creased with leaf age. The amount of protein per gram dryweight and non-protein a-amino nitrogen seemed to berather insensitive to both daylength and incident lightenergy. Small changes, however, could be detected withincreasing total light energy.
GDH separated on polyacrylamide electrophoresis andvisualized as described in Materials and Methods showedseveral bands on the gels (Figure 3). ln the absence of sub-strate, sodium glutamate, no bands were visible. When thecofactor NAD+, necessary for GDH activity, was omitted,two bands were still visible. These two bands representother enzymes able to oxidize glutamic acid. Such NAD"*"-
r.
300
c 200
336- 1 I lA 1
50A 756 336 50Ato ta l l i gh t e n e r g y W h - m - d a y " '
15
10
oc
756
Hgiire 2. The effect of daylength and total light energy on (A) theamount of protein per gram dry weight, (B) the content of non-protein ot-aniino nitrogen expressed as percentage of total nitrogen.Each point is a mean of three replicates. The values correspond tocultivation II. ( ) short clay, young leaves; (1 ) long day, youngleaves; (•) short day, old leaves; (•) long day, old leaves.
front
1 2 3Figure 3. Glutamate dehydrogenase (N AD"'^-dependent) separatedby polyacrylamide gel electrophoresis. (1) Total reaction medium,(2) without NAD+, (3) without glutamic acid.
Figure 4 (next page). Photographs and densitometric traces of glutamate dehydrogenase (NAD^-dependent) after polyaerylamide gelelectrophoresis. The gels received 100 /i\ protein extract each. The values correspond to cultivation 1.
Part-figures Daylength and irradiance Leafage protein/100 /d extract
4A4B4C4D4E4F4G4H
short day 28 W x m""̂long day 28 W x m"^short day 42 W x m~̂long day 42 W x m'^short day 28 W x m~̂long day 28 W x m"-̂short day 42 W x m~̂long day 42 W x rn"-̂
youngyoungyoungyoungoldoldoldold
28512311115517690
198115

Physiol. Plant. 30. 1974 ENZYME ACTIVITIES IN URTICA DIOICA 195
08
0.8
0.6
OD
0.4
02
0
1A
H 1
/
I I I
196 MARGARETA WELANDER Physiol. Plant. 30. 1974
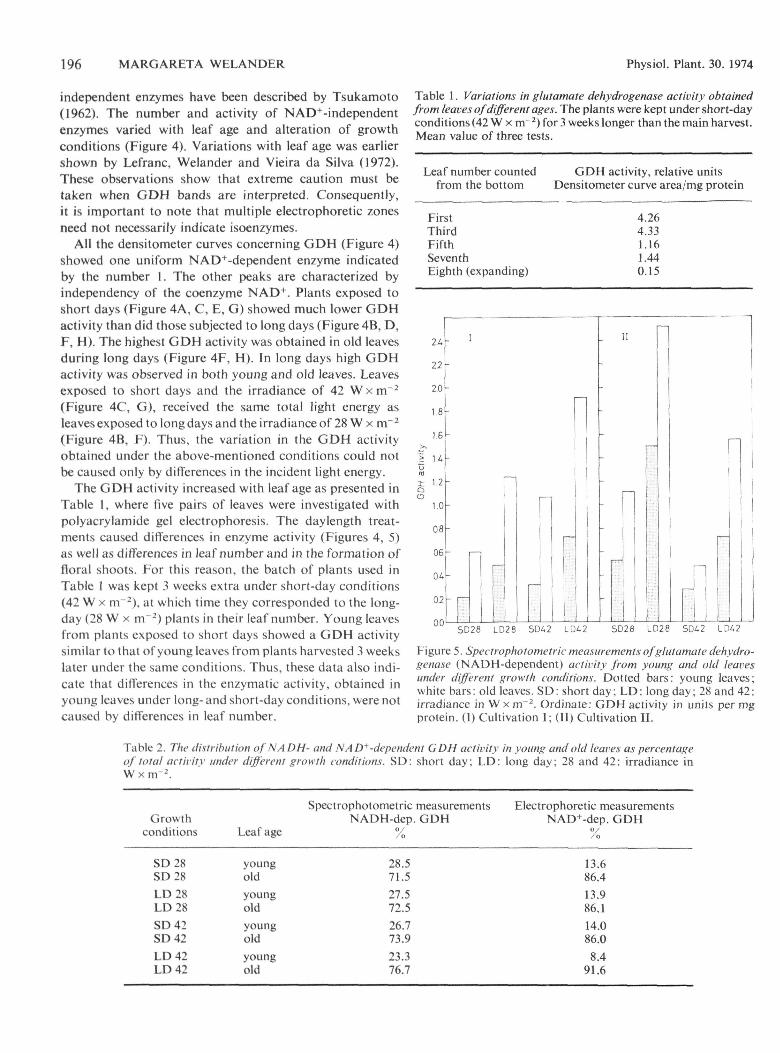
independent enzymes have been described by Tsukamoto(1962). The number and activity of NAD+-independentenzymes varied with leaf age and alteration of growthconditions (Figure 4). Variations with leaf age was earliershown by Lefranc, Welander and Vieira da Silva (1972).These observations show that extreme caution must betaken when GDH bands are interpreted. Consequently,it is important to note that multiple electropboretic zonesneed not necessarily indicate isoenzymes.
All the densltometer curves concerning GDH (Figure 4)showed one uniform NAD'^-dependent enzyme indicatedby the number 1. The other peaks are characterized byindependency of the coenzyme NAD" .̂ Plants exposed toshort days (Figure 4A, C, E, G) showed much lower GDHactivity than did those subjected to long days (Figure 4B, D,F, H). The highest GDH activity was obtained in old leavesduring long days (Figure 4F, H). In long days high GDHactivity was observed in both young and old leaves. Leavesexposed to short days and the irradiance of 42 W x m~̂(Figure 4C, G), received the same total light energy asleaves exposed to long days and the i rradiance of 28 W x m~̂(Figure 4B, F). Thus, the variation in the GDH activityobtained under the above-mentioned conditions could notbe caused only by differences in the incident light energy.
The GDH activity increased with leafage as presented inTable 1, where five pairs of leaves were investigated withpolyacrylamide gel electrophoresis. The daylength treat-ments caused differences in enzyme activity (Figures 4, 5)as well as difTerences in leaf number and in the formation offloral shoots. For this reason, the batch of plants used inTable 1 was kept 3 weeks extra under short-day conditions(42 W X m̂ •̂ ), at which time they corresponded to the long-day (28 W X m"'̂ ) plants in their leaf number. Young leavesfrom plants exposed to short days showed a GDH activitysimilar to that ofyoung leaves from plants harvested 3 weekslater under the same conditions. Thus, these data also indi-cate that differences in the enzymatic activity, obtained inyoung leaves under long- and short-day conditions, were notcaused by differences in leaf number.
Table 1. Variations In glutamate dehydrogenase activity obtainedfrom leaves of different ages. The plants were kept under short-dayconditions (42 W x m~ )̂ for 3 weeks longer than the main harvest.Mean value of three tests.
Leaf number countedfrom the bottom
GDH activity, relative unitsDensitometer curve area/mg protein
FirstThirdFifthSeventhEighth (expanding)
4.264.331.161.440.15
l.u-
2.2 -
2.0-
1 8 -
16
12
1.0
08
06
Oi
02
00SD28 LD28 SD42 LDi2 SD28 LD28 SD62
Figure 5. Spectrophotometric measurements of glutamate dehydro-genase (NADH-dependent) activity from young and old leavesunder different growth conditions. Dotted bars: young leaves;white bars: old leaves. SD: short day; LD: long day; 28 and 42:irradiance in W x m -̂̂ . Ordinate: GDH activity in units per mgprotein. (I) Cultivation 1; (II) Cultivation II.
Table 2. The distribution of NA DH- and N.4 D*-dependent GDH activity in young and old leaves as percentageof total actiiitv under different growth conditions. SD: short day; LD: long day; 28 and 42: irradiance inW X m-^
Growthconditions
SD28SD28LD28LD28SD42SD42LD42LD42
Leaf age
youngoldyoungoldyoungoldyoungold
Spectrophotometric measurementsNADH-dep. GDH
O /.' O
28.571.527.572.526.773.923.376.7
Electrophoretic measurementsNAD+-dep. GDH
o//o
13.686.413.986.114.086.08.4
91.6
Physiol. Plant. 30. 1974 ENZYME ACTIVITIES IN URTICA DIOICA 197
0,6.
0.4.
02
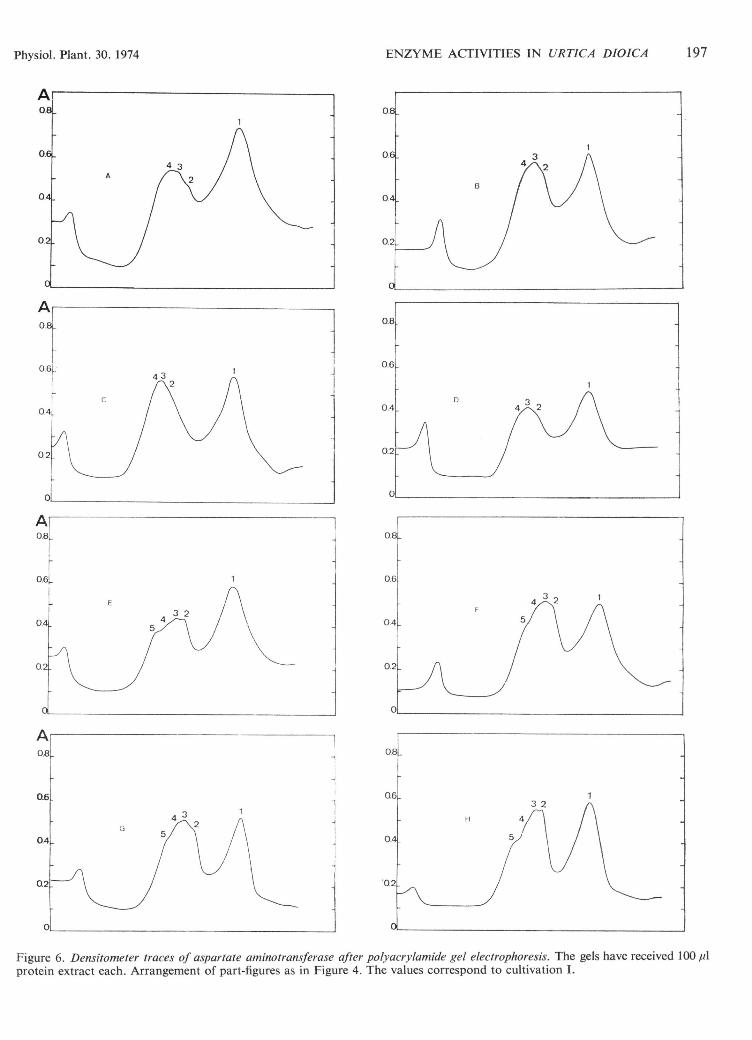
Figure 6. Densitometer traces of aspartate aminotransferase after polyacrylamide gel electrophoresis. The gels have received 100 /Aprotein extract each. Arrangement of part-figures as in Figure 4. The values correspond to cultivation I.
198 MARGARETA WELANDER Physiol. Plant. 30. 1974
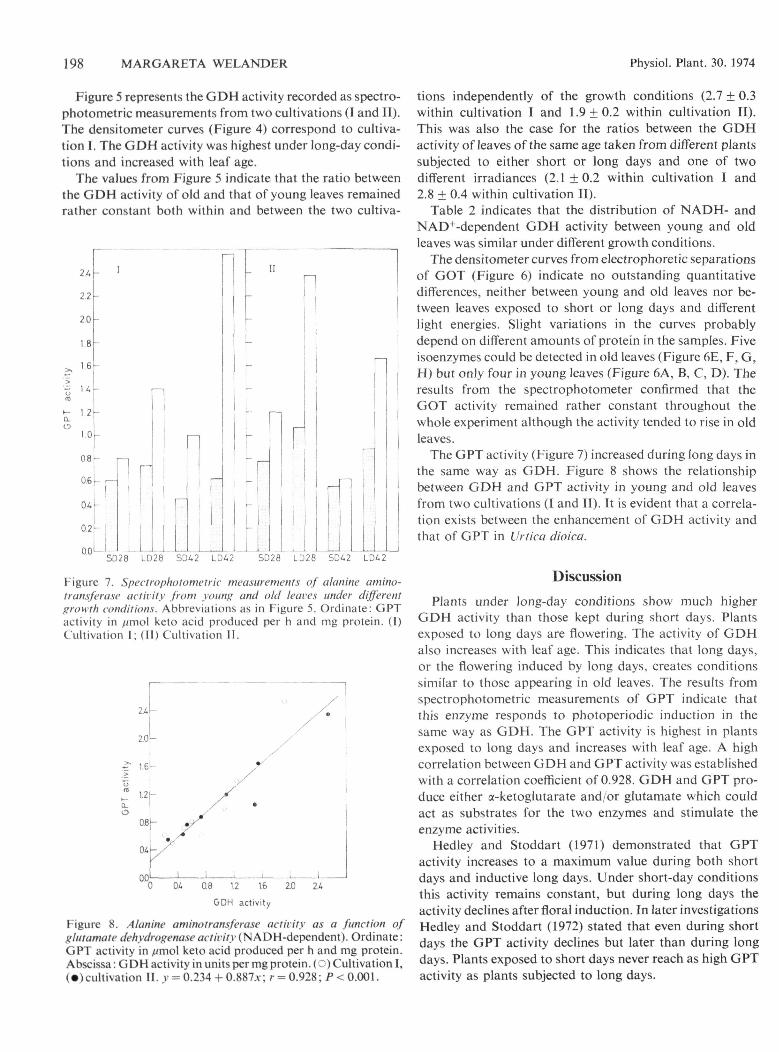
Figure 5 represents the GDH activity recorded as spectro-photometric measurements from two cultivations (I and II).The densitometer curves (Figure 4) correspond to cultiva-tion I. The GDH activity was highest under long-day condi-tions and increased with leaf age.
The values from Figure 5 indicate that the ratio betweenthe GDH activity of old and that ofyoung leaves remainedrather constant both within and between the two cultiva-
O
2A
2 2 -
2,0-
1.8-
16-
1 2
1.0
08
06
02
0.0
1
[\1—
i—
1-r
r
I
—
SD28 LD28 SDi2 LDi2 SD28 LD28 SDi2 LDi2
Figure 7. Spectrophotometric measurements of alanine amino-transferase activity from young and old leaves under differentgrowth conditions. Abbreviations as in Figure 5. Ordina te : G P Tactivity in //mol keto acid produced per h and mg protein. (I)Cultivation I; (II) Cultivation II.
0.008 12 1.6
GDH activity
2.0 2J.
Figure 8. Alanine aminotransferase activity as a function ofglutamate dehydrogenase activity (NADH-dependent). Ordinate;GPT activity in //mol keto acid produced per h and mg protein.Abscissa: GDH activity in units per mg protein. (O) Cultivation I,(•)cultivation II. >• = 0.234 -h 0.887A-; r = 0.928; P < 0.001.
tions independently of the growth conditions (2.7 ± 0.3within cultivation I and 1.9 ±0.2 within cultivation II).This was also the case for the ratios between the GDHactivity of leaves of the same age taken from different plantssubjected to either short or long days and one of twodifferent irradiances (2.1 ±0.2 within cultivation I and2.8 ± 0.4 within cultivation II).
Table 2 indicates that the distribution of NADH- andNAD'^-dependent GDH activity between young and oldleaves was similar under different growth conditions.
The densitometer curves from electrophoretic separationsof GOT (Figure 6) indicate no outstanding quantitativedifferences, neither between young and old leaves nor be-tween leaves exposed to short or long days and differentlight energies. Slight variations in the curves probablydepend on different amounts of protein in the samples. Fiveisoenzymes could be detected in old leaves (Figure 6E, F, G,H) but only four in young leaves (Figure 6A, B, C, D). Theresults from the spectrophotometer confirmed that theGOT activity remained rather constant throughout thewhole experiment although the activity tended to rise in oldleaves.
The GPT activity (Figure 7) increased during long days inthe same way as GDH. Figure 8 shows the relationshipbetween GDH and GPT activity in young and old leavesfrom two cultivations (I and II). It is evident that a correla-tion exists between the enhancement of GDH activity andthat of GPT in UrTica dioica.
Discussion
Plants under long-day conditions show much higherGDH activity than those kept during short days. Plantsexposed to long days are flowering. The activity of GDHalso increases with leaf age. This indicates that long days,or the flowering induced by long days, creates conditionssimilar to those appearing in old leaves. The results fromspectrophotometric measurements of GPT indicate thatthis enzyme responds to photoperiodic induction in thesame way as GDH. The GPT activity is highest in plantsexposed to long days and increases with leaf age. A highcorrelation between GDH and GPT activity was establishedwith a correlation coefficient of 0.928. GDH and GPT pro-duce either a-ketoglutarate and/or glutamate which couldact as substrates for the two enzymes and stimulate theenzyme activities.
Hedley and Stoddart (1971) demonstrated that GPTactivity increases to a maximum value during both shortdays and inductive long days. Under short-day conditionsthis activity remains constant, but during long days theactivity declines after floral induction. In later investigationsHedley and Stoddart (1972) stated that even during shortdays the GPT activity declines but later than during longdays. Plants exposed to short days never reach as high GPTactivity as plants subjected to long days.

Physiol. Plant. 30. 1974 ENZYME ACTIVITIES IN URTICA DIOICA 199
There is a continuous "turnover" of proteins in leaves(Steward and Durzan 1965). During the ageing of the leavesthere is a sequence in the build-up of the individual amidesand amino acids in the soluble phase of the leaf (Pate 1966).Well-defined rhythms in the enzyme activity and in theamounts of specific metabolites create the impression thatageing may involve progressive alteration in the balance ofanabolic and catabolic compounds. Figure 2 indicates thataccumulation of free amino acids increases with leafage. Inexperiments with Xanthium pennsylvanium, Krizek,Mcllrath and Vergara (1966) demonstrated that the kind ofleaf senescence that follows a photoperiodically inducedtransition from vegetative to reproductive conditionsoccurs whether or not the plants had a capacity to produceflowers or fruits. This type of leaf senescence seems to resultfrom photoperiodic efi"ects on the leaves themselves.
The GOT activity does not respond to environmentalchanges. This seems to reflect a rather general state of theGOT activity. Hedley and Stoddart (1971) demonstratedthat Lolium temulentum, exposed to inductive photo-periods, maintains a constant GOT activity.
The investigation was supported by a grant from the Bank ofSweden Tercentenary Fund.
References
Bergmeyer, H. U. & Bernt, E. 1962. Glutamat-oxalacetat Trans-aminase, Giutamat-pyruvat Transaminase. — In Methodender enzymatischen Analyse (H. U. Bergmeyer, ed.), pp. 837-853. Verlag Chemie GmbH Weinheim/Bergstr.
Brewbaker, I. L., Upadhya, M. D., Makinen, Y. & MacDonald,T. 1968. Isoenzympolymorphism in flowering plants. III. Gelelectrophoretic methods and applications. — Physiol. Plant.21:930-940.
Davis, B. J. 1964. Disc electrophoresis. II. Method and applica-tion to human serum proteins. — Ann. N.Y. Acad. Sci. 121:404-427.
Fine, I. H. & Costello, L. A. 1963. The use of starch electro-phoresis in dehydrogenase studies. — In Methods in Enzym-ology (S. P. Colowick & N. O. Kaplan, eds.) 6: 968.
Hedley, C. L. & Stoddart, J. L. 1971. Factors influencing alanineaminotransferase activity in leaves of Lolium temulentum (L).— J. Exp. Bot. 22:239-248.
1972. Patterns of protein synthesis in Lolium temulentum(L).—/6(V/. 23:490-501.
Kannangara, C. G. & Woolhouse, H. W. 1968. Changes in theenzyme activity of soluble protein fractions in the course offoliar senescence in Perilla frutescens (L.) Britt. — NewPhytol. 67: 533-542.
Krizek, D. J., Mcllrath, W. J. & Vergara, B. S. 1966. Photo-periodic induction of senescence in Xanthium plants. —Science 151: 95-96.
Lefranc, M., Welander, M. & Vieira da Silva, J. B. 1972. L'activiteglutamate deshydrogenase dans les tissus foliaires du Cheno-podium quinoa Wild. — C.R. Acad. Sci., Paris 274: 689-692.
Lexander, K., Carlsson, R., Schalen, V., Simonsson, A. &Lundborg, T. 1970. Quantities and qualities of leaf proteinconcentrates from wild species and crop species grownunder controlled conditions. — Ann. Appl. Biol. 66: 193-216.
Lowry, O. H., Rosebrough, N. J., Farr, A. L. & Randall, R. J.1951. Protein measurement with the folin phenol reagent. —J. Biol. Chem. 193: 265-275.
Pahlich, E. & Joy, K. W. 1971. Glutamate dehydrogenase frompea roots; Purification and properties of the enzyme. — Can.J. Biochem. 49: 127-138.
Pate, J. S. 1966. Photosynthesizing leaves and nodulated roots asdonors of carbon to protein of the shoot of the field pea(Pisum arvense L.). — Ann. Bot. 30: 93-109.
Potty, W. H. 1969. Determination of proteins in the presence ofphenols and pectins. — Anal. Biochem. 29: 535-539.
Schmidt, E. 1963. Glutamic dehydrogenase. — In Methods ofEnzymatic Analysis (H. U. Bergmeyer, ed.), pp. 752-756.New York.
Steward, F. C. & Durzan, D. J. 1965. Metabolism of nitrogenouscompounds. — In Plant Physiology (F. C. Steward, ed.) 4a,Ch. 4. Academic Press, New York.
— Crane, F., Millar, K., Zachrius, R. M., Rabson, R. &Margolis, D. 1959. Nutritional and environmental effects onthe nitrogen metabolism of plants. — Symp. Soc. Exp. Biol.Number XIII. Utilization of nitrogen and its compounds byplants, pp. 148-176.
Tsukamoto, A. 1962. Elavoproteins in the leaves of higher plantscatalyzing the oxidation of L-glutamate. — Plant Cell Physiol.3: 293-307.
Yue, S. B. 1969. Isoenzymes of glutamate dehydrogenase inplants. — Plant Physiol. 44: 453-457.

Copyright © 2022 FDOKUMEN




















