
Pharmacological evaluation of extracts from Buxus macowanii ...

ANTIOXIDANT PROPERTIES OF OREGANO (OHGANUM VULGARE) LEAF EXTRACTS
GIOVANNA CERVATO', MARTA CARABELLI, SILVIA GERVASIO, ANDREA CITTERA, ROBERTA CAZZOLA and BENVENUTO CESTARO
Dipartimento di Scienze Precliniche L. I. T.A. Vialba Universita degli Studi di Milano
Italia
Received for Publication March 1,2000 Accepted for Publication June 22,2000
ABSTRACT
We tested the antioxidant properties of both aqueous and methanolic extracts of oregano (origanum vulgare). They proved to be eflective in the inhibition of all phases of the peroxidative process: first neutralizing free radicals (superoxide anion, hydroxyl radical and I , I-diphenyl-2-picrylhydraql radical), then blocking peroxidation catalysis by iron (through iron-chelating and iron-oxidizing properties), and finally through interruption of lipid-radical chain reactions (chain-breaking activity). Their anti-glycosylation activity was also effective. The glycosylation of lipoproteins is directly related to their peroxidation. The amount of extract used in our experiments was obtained from 0.1-1 mg of dried leaves, amounts far less than those normally used in the Mediterranean diet.
INTRODUCTION
Lipid peroxidation is a multistep reaction that can result in destruction of cellular membranes. The reaction starts when a free radical (which can be an oxygen radical) reacts with the double allylic bond of a polyunsaturated fatty acid to form a conjugate diene. This reacts with oxygen and forms a lipid hydroperoxide. Iron can promote the breaking of this compound and the formation of lipid peroxyl and alkoxyl radicals. These radicals can cause hrther reactions with other polyunsaturated fatty acids and thus propagate the chain reactions as long as substrates are available.
An antioxidant is any substance that delays or inhibits peroxidative reactions.
'Correspondence to: Giovanna Cervato, Dipartimento di Scienze Precliniche L.I.T.A. Vialba, V. G.B. Grassi, 74, 20157 Milano, Italia, TEL: +39-2-38210442; FAX: +39-2-38210440; E-mail: [email protected]
Journal of Food Biochemistry 24 (2000) 453-465. All Righis Reserved. "Copyright 2000 by Food & Nutrition Press, Inc., Trumbull, Connecticui 453

454 G. CERVATO ETAL.
Antioxidants can affect different stages of an oxidative sequence. An antioxidant can be specific or have multiple sites of action. A number of specific reactions can be used to monitor the different molecular mechanisms of an antioxidant.
As peroxidative processes are involved in many degenerative physiopathologic events (aging, cancer, diabetes, atherosclerosis, etc.), a good pro-/antioxidant balance is very important for health (Gutteridge 1995). Oxidative status can be influenced by dietary compounds, both positively by antioxidant components of foods (Kubow 1992) and negatively by food peroxidized during the cooking processes (Aruoma et al. 1996). Therefore, there is increasing interest for the natural dietary antioxidants, not only vitamins, but also nonessential dietary antioxidants (Decker 1995). Among the latter, polyphenols have assumed great importance, and in a recent review Aruoma (Aruoma 1997) proposed the use of antioxidant vegetable extracts both as food preservatives and as prophylactic agents for some human diseases.
Spices have a high content of polyphenols, so they presumably have antioxidant properties. The Mediterranean diet is particularly rich in spices, one of which is oregano (Origanum vulgare). Lagouri and Boskou (1996) reported the presence of five major phenolic compounds with antioxidant properties in hexane and methanol extracts of oregano (Origanum vulgare). The compounds identified were protocatechinic acid and his phenyl glucoside, caffeic acid, rosmarinic acid and a phenolic derived of rosmarinic acid. Kikuzaki and Nakatani (1989) identified tocopherol homologues, in the dichloroethane extract of oregano.
Here we investigate the different antioxidant mechanisms, of aqueous and methanol extracts of oregano.
To test these antioxidant properties we used diverse in vitro models which served to elucidate the different molecular action mechanisms. In particular, we tested radical-scavenging, chain-breaking, iron-chelating and iron-oxidizing activities. Moreover, we tested for possible inhibitory effects of oregano extracts in ex-in vivo models of both plasma lipoprotein peroxidation and plasma albumin glycos ylation.
MATERIALS AND METHODS
Materials
Analytical grade chemicals, distilled solvents and double-distilled water were used. Dipalmitoylphosphatidylcholine (DPPC), arachidonic acid (AA), deoxyribose (DR), nitrobluetetrazolium chloride (NBT), xanthine (X) and xanthine oxidase (XO), superoxide dismutase (SOD), quercetin, mannitol, trolox, a- tocopherol, 1,l ,diphenyl-2-picryl-hydrazyl radical (DPPH), were purchased from Sigma Chemicals Co. (St. Louis, MO). The dried oregano (Origanum vulgare)

ANTIOXIDANT ACTIVITY OF OREGANO EXTRACTS 455
leaves used were a commercial herbal product.
Methods
Oregano Extracts. Dry oregano leaves were extracted with either water or methanol.
Water Extraction. One gram dry oregano leaves was minced in a small water volume, diluted to 200 mL and boiled until 100 mL of volume, then stirred for 24 h.
Methanol Extract. One gram dry oregano leaves were stirred in 100 mL. methanol for 24 h at room temperature (Gugliucci and Stahl 1995).
Reactivity with l,l-diphenyl-2-picrylhydrazyl (DPPH) Radical. DPPH is a stable free radical that, dissolved in ethanol, has a blue-violet color. In the presence of a radical scavenger, DPPH is converted to 1,l -diphenyl-2-picrylhydrazine with yellow color. The decrease of color monitors the radical scavenger activity. To 3 mL. of 60 pM DPPH in ethanol were added different amounts of oregano (water or methanol) extracts, and absorbancy was monitored at 517 nm. The decrease of AS,,,,,,, is an index of scavenger activity of oregano extracts (Constantin et al. 1990). Oregano extract scavenger activity was compared with the activity of the same amounts of quercetin, a strong antioxidant and the major dietary flavonoid (Hollman et al. 1997).
Hydroxyl Radical Formation Inhibition. Hydroxyl radical formation was measured according to Halliwell et al. (1987). Deoxyribose (DR) (2.8 mM) in 20 mM phosphate buffer pH 7.4 was oxidized by hydroxyl radical formed by the reaction of 100 pM iron (111) 100 pM ascorbate and 1 mM hydrogen peroxide, in presence or absence of oregano extracts at 37C for 1 h. The oxidation products were made to react with thiobarbituric acid (TBA) 0.25 mM in 7% percloric acid at lOOC for 10 min, and the pink colored products were read at 532 nm after the addition of ethano1:ether 3: 1 by vol. Hydroxyl radical scavenger inhibits the oxidation of DR and, thus, thiobarbituric acid reactive substance (TBARS) formation. The scavenger activity of oregano extracts was compared with that of mannitol (commonly used as hydroxyl radical scavenger) in comparable amounts.
Superoxide Radical Formation Inhibition. Superoxide radicals formed by the xanthine/xanthine oxidase (X/XO) system were measured by their ability to oxidize nitrobluetetrazolium (NBT, yellow) to formazane (blue). The reaction occurs in 3 mL of 60 mM phosphate buffer, pH 7.4 in which there is 30 mM xanthine, 30 mM EDTA, 3 mM NBT. Addition of 5 U of xanthine oxidase starts the reaction leading

456 G. CERVATO ETAL.
to superoxide anion and, thus, formazane formation. The scavenging property of oregano extracts was measured by their ability to inhibit this reaction (Aruoma ef al. 1989) and was monitored as decrease of a 560 nm in the presence of extracts with respect to the control without any antioxidant. Superoxide dismutase (SOD) was used as a reference.
Iron-binding, Iron-oxidizing and Chain-breaking Properties in Aqueous Phase Assay. Modulating reciprocal concentrations of iron and ascorbate used for DR oxidation, it is possible to distinguish different antioxidant activities against oxygen radical formation (Gutteridge and Quinlan 1992). DR (1 mM) was oxidized (in presence or absence of oregano extract or comparative antioxidant) by 1 mM hydrogen peroxide, and (1) for iron-binding determination: 0.75 mh4 ascorbate; (2) iron-oxidizing: 0.1 mM ferric chloride and 0.0125 mM ascorbate; (3) chain- breaking: 0.1 mM ferric chloride and 0.75 mM ascorbate. Reaction was made at 37C for 1 h and then the formation of TBARS was monitored as reported above. The activity of oregano extract was compared against the activity of transferrin as Fe chelator, quercetin as Fe oxidant, Trolox as hydrosoluble analogue of tocopherol and, thus, hydrosoluble chain breaking.
Inhibition of the Peroxidation of Arachidonic Acid (AA) Inserted Into Multilayered Vesicles (MLV) of Dipalmitoylphosphatidylcholine (DPPC). The peroxidation of AA inserted in multilamellar liposome of DPPC (AA 20% mol/mol) was induced by adding a mixture of ferrous sulfate and ascorbic acid (50 pM and 500 pM final concentration, respectively) to the liposomal suspension in presence or absence of oregano extract or vitamin E. Peroxidation was performed at room temperature and was stopped by adding hydroquinone (25 mM final concentration). TBARS were monitored as an index of AA peroxidation (Cervato et al. 1988). Vitamin E, a very strong hydrophobic chain-breaking antioxidant, was used as a reference. Vitamin E acts inside the vesicle membrane and inhibits propagation of peroxidative reactions.
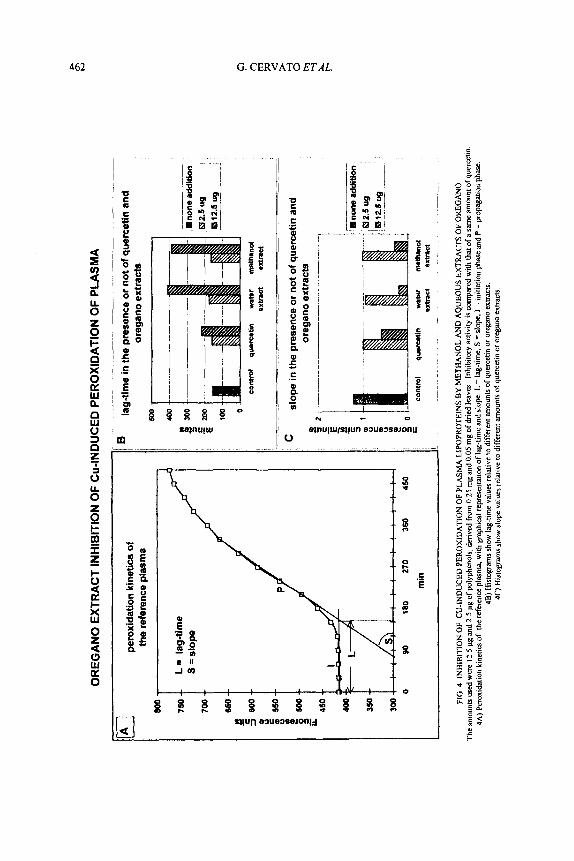
Plasmatic Peroxidation Inhibition. Inhibition of plasmatic peroxidation was measured as increment of lag-time with fluorescence kinetic method (Cervato et af. 1999). To 0.6 mL of plasma from volunteers was added 0.6 mL of 1 mM copper sulfate. Kinetics of peroxidation was monitored as increment of fluorescence (A,, 360 nm and A,, 430 nm) due to Schiff base formation through the carbonyl groups of aldehydes (end products of lipid peroxidation) and plasmatic protein amino groups. Lag-time (L in the graph A of Fig. 4) is the latency time of plasma peroxidation, and is expressed in min. It was calculated as the intercept of tangents of the initiation phase (I in the graph A of Fig. 4) and propagation phase curves (P in the graph A of Fig. 4). Slope (S in the graph A of Fig. 4) is the velocity of reaction in the propagation phase and is expressed in fluorescence units (FU) for min. Quercetin was used as reference antioxidant.

ANTIOXIDANT ACTIVITY OF OREGANO EXTRACTS 457
Protein Glycosylation Inhibition. Glycation of bovine serum albumin in the presence of glucose was determined according to Morimitsu et al. (1995) in the presence or absence of oregano extracts or aminoguanidine. A mixture of glucose (500 mg) and albumin (50 mg) was dissolved in 5 mL of phosphate saline buffer (0.15 M, pH 7.2) and incubated at 60C. Sample of reaction solution were taken every 24 h for measurement of amino-carbonyl reaction. Measurement of the formation of advanced glycosylation end products (AGES) was assessed by monitoring their characteristic fluorescence (Aex 370 nm and Am 440 nm. Aminoguanidine in the same amounts was used for comparison.
Total Phenol Determination. Total phenols were measured by reaction with phosphomolybdic-phosphotungstic acid (Folin-Ciocalteau reagent of Merck) (Singleton and Draper 1964), using quercetin as reference standard. Reaction occurs in the presence of NaCO, (2% in 0.1N NaOH), CuSO, (0.5%) and Na-K tartrate (1%). Absorbance was read at 750 nm.
Statistical Analysis. Experiments were performed in triplicate and repeated five times. The variance of results was evenly distributed within a 5% range. Scavenger activity or inhibition activity was expressed as percent of the reaction without antioxidant.
RESULTS AND DISCUSSION
Extracts of dried oregano leaves were obtained with water or methanol. The extraction conditions were vigorous. Both extractions were performed for 24 h with stirring. Water extraction was performed at lOOC until water volume was reduced by half, and then at room temperature. Methanol extraction was done at room temperature. Figure 1 shows that final dry weight and polyphenol content of extracts were quite similar for both methods. Extracted material was about 1/4 of initial weight; 115 of dry weight showed polyphenol reactivity.
Antioxidant activity tests were performed using the extracts derived from 1 mg and 0.1 mg of dried oregano leaves, corresponding to 50 pg and 5 pg of polyphenols. Different experimental models were used, as it is known that an antioxidant can act both in the initiation phase and during the propagation phase of the peroxidative process (Lewin and Popov 1994). In the initiation phase it could neutralize or inhibit the formation both of oxygen free radicals (superoxide anion or hydroxyl radical) and of other radicals. The antioxidant could also act on reactions catalyzed by transition metals, either by their chelation or by altering their oxidative status. Lastly, it could stop chain reaction propagation promoted by lipidic radicals (chain-breaking). The activity of oregano extracts was compared with the activity of standard antioxidants commonly used by other investigators,

458 G. CERVATO ETAL.
OREGANO (ORIGANUM VULGARE) POLYPHENOLS EXTRACTION
1- dried oregano leaves -1
water (200 mL at IOOC. I evaporation to a
niethanol (I00 inl. at 22C. no
signiftcanl evaporation)
65.35 rng
FIG. 1. WATER AND METHANOL EXTRACTION OF DRIED OREGANO LEAVES
at concentrations quite similar to those of maximal activity and comparable amounts of polyphenol as our samples.
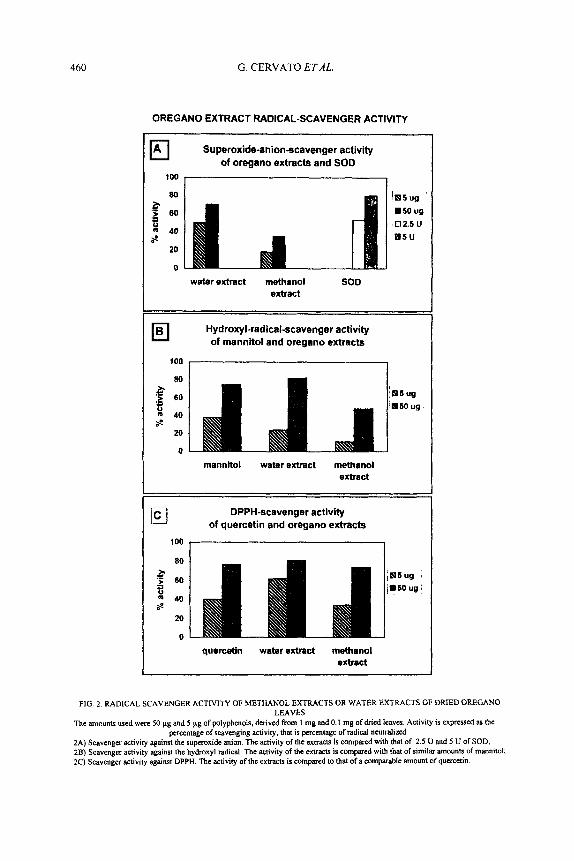
Figure 2 shows the activity of dned oregano leaf methanol and aqueous extracts as scavengers of different radicals.
Figure 2A shows superoxide radical scavenger activity of the extracts as compared to superoxide dismutase activity. It is evident that the aqueous extract is far more active than the methanol extract and comparable to that of SOD. Fifty pg of polyphenols in the aqueous extract derived from dried oregano leaves scavenged 70% of the superoxide anion, as compared to only 35% scavenged by an equal quantity of the methanolic oregano extract. Thus the 50 pg of the aqueous extract polyphenols were almost as effective as 5 U of SOD, which scavenged 80%.
As reported in Fig. 2B, aqueous extract activity is superior to that of methanolic extract also as a hydroxyl radical scavenger. A 50 pg aliquot of aqueous extract polyphenols scavenged 78% of hydroxyl radical, which was the same activity demonstrated for an equal amount of mannitol.

ANTIOXIDANT ACTIVITY OF OREGANO EXTRACTS 459
To test oregano scavenger activity on a generic radical, we used the DPPH radical. The result is reported in Fig. 2C. As a reference standard we used quercetin. At the higher concentration (50 pg of polyphenols) the activity of both of the two extracts was comparable to quercetin (about 80%). On the contrary, it is of interest to note that at the small concentration (5 pg of polyphenols) the aqueous extract was much more active (62%) than either quercetin (40%) or the methanol extract (37%). This would seem to indicate a greater reactivity of the polyphenols of the aqueous oregano extract for DPPH than of methanol extract, polyphenols or quercetin.
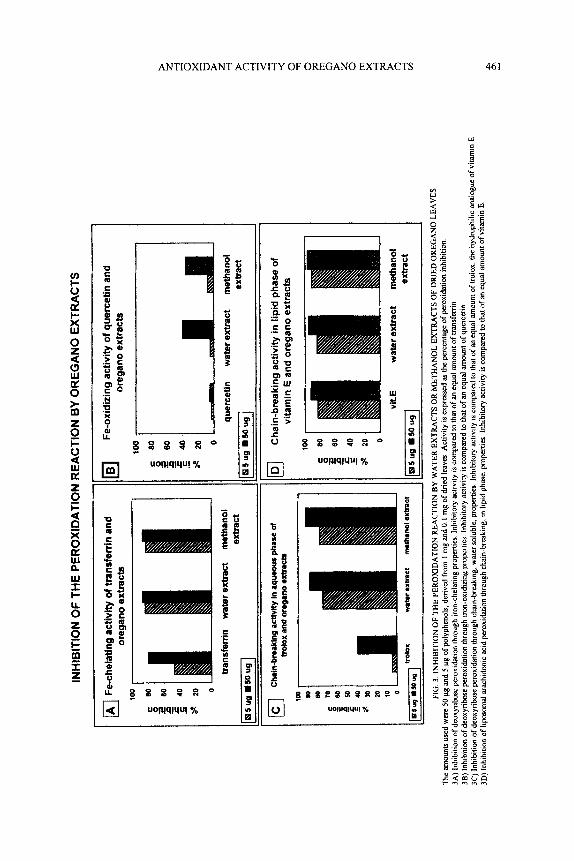
Figure 3 shows the results of experiments monitoring the antioxidant activity of oregano extracts in the peroxidation propagation phase. Here, antioxidant activity is expressed as % of peroxidation reaction inhibition.
In Fig. 3A the iron chelating activity of oregano extracts is shown, using as standard a comparable amount of transferrin. About 80% of iron was chelated in the presence of 5 and 50 pg of polyphenols, both aqueous and methanolic. This activity was similar to that of 50 pg of transferrin. In small quantities (5 pg), transferrin showed far less activity. Thus, oregano polyphenols seem to have a greater affinity for iron than transferrin. Iron oxidizing activity is reported in Fig. 3B. The standard used was quercetin, which in our experimental condition, however, showed very little activity (less than 4-5%). The oregano extracts (both aqueous and methanol) had a far greater effect, especially when we used 50 pg of the polyphenols (40% mhibition of peroxidation). Chain-breaking activity (Fig. 3C and 3D) was monitored both as deoxyribose peroxidation activity (hydrophilic molecule peroxidation), and as peroxidation of arachidonic acid inserted into multilamellar dipalmitoylphosphatidylcholine liposomes (hydrophobic substrate peroxidation). From the data shown in Fig. 3C, it is clear that the efficacy of the oregano extracts in the inhibition of deoxyribose peroxidation was greater than that of an equal amount of trolox (hydrophilic vitamin E analog). The extremely high mhibitory capacity of the two extracts was comparable. Inhibition of arachidonic acid peroxidation (Fig. 3D) was very good, and for both of the extracts was comparable to that of vitamin E.
We also tested the extracts’ antioxidant activity in an ex-in vivo model, Cu- induced peroxidation of plasma lipoproteins. In Fig. 4A the peroxidation kinetics, monitored by fluorescence, are represented by a sigmoidal curve from which it is possible to derive two parameters related to the peroxidative process: the lag-time (180 min for the reference plasma used), which is indicative of plasma susceptibility to peroxidation; and the slope (1.2 FU/min for the reference plasma), which represents the velocity of the propagation reaction.
The addition of oregano extracts to the plasma remarkably increased lag-time values (Fig. 4B), similarly for both aqueous and methanolic extracts and about twice that of comparable amounts of quercetin (used as reference standard). The addition of the oregano extracts affected the slope values in a similar manner (Fig.
460 G. CERVATO E T A L .
OREGANO EXTRACT RADICAL-SCAVENGER ACTIVITY
1w
80 ' 60
5 40 s
20
0
Superoxide-anion-scavenger activity of oregano extracts and SOD
weter extract methanol SOD extract
185 ug m5Oug
, 0 2 . 5 U
115 u
Hydroxyl-radical-scavenger activity of mannitol and oregano extracts
100 1 80
@ 60 8 5 ug m 50 ug
40
20
0
s
mannitol water extract methanol extract
100
80
f 60
40 s 20
0
.;
DPPH-scavenger activity of quercetin and oregano extracts
I
querceth water extract methanol extract
FIG 2. RADICAL SCAVENGER ACTIVITY OF METHANOL EXTRACTS OR WATER EXTRACTS OF DRIED OREGANO LEAVES
The amounts used were SO pg and 5 pg of polyphenols, derived from 1 mg and 0.1 mg of dried leaves. Activity is expressed as the percentage of scavenging activity, that is percentage of radical neutralized
ZA) Scavenger activity against the superoxide anion. The activity of the extracts is compared with that of 2 5 U and 5 U of SOD, 2B) Scavenger activity against the hydroxyl radical. The activity of the exvacts is compared with that of similar amounts of mannitol, 2C) Scavenger activity against DPPH The activity of the extracts is compared to that of a comparable amount of quercetin.
INH
IBIT
ION
OF T
HE
PE
RO
XID
AT
ION
RE
AC
TIO
N B
Y O
RE
GA
NO
EX
TR
AC
TS
Fee
hel
ath
g a
ctiv
ky o
f tra
nsfe
rrin
and
or
egan
o ex
trac
ts
Fe-o
xidi
zing
act
ivity
of
quer
cetin
and
10
0,
1 II
100
oreg
ano
extr
acts
I
auer
cetin
w
ater
ext
ract
m
etha
nol
tran
sfen
in
wat
erex
trac
t m
etha
nol
II
Cha
in-b
reak
ing
activ
ity in
lipi
d ph
ase
of
vita
min
E a
nd o
rega
no e
xtra
cts
Ch8
in-b
reak
lng
ac
tmy
In aqueous *re
of
tm)o
x an
d 0ra
g.m
ext
ract
s
mh
r *
me
t m
ttun
ol e
xhac
t ex
trac
t
FIG
3 I
NH
IBIT
ION
OF
THE
PERO
XID
ATI
ON
REA
CTIO
N BY
WA
TER
EXTR
AC
TS O
R M
ETH
AN
OL
EXTR
AC
TS O
F D
RIE
D O
REG
AN
O L
EAV
ES
The
amou
nIs
used
wer
e SO
pg
and
5 pg
of p
olyp
heno
ls, d
eriv
ed fr
om I
mg
and
0 I
mg
of d
ried
leav
es A
ctiv
ity is
exp
ress
ed as
the
perc
enta
ge o
f per
oxid
atio
n in
hibi
tion
3A) I
nhib
ition
of d
eoxy
ribos
e pe
roxi
datio
n th
roug
h iro
n-ch
elat
ing
prop
ertic
s In
hibi
tory
act
ivity
is c
ompa
red
to th
at o
f an
equa
l am
ount
of t
rans
feni
n 30
) In
hibi
tion
of d
eoxy
ribos
e pe
roxi
datio
n th
roug
h iro
n-ox
idiz
ing
prop
ertie
s In
hibi
tory
act
ivity
is c
ompa
red
to th
at o
f an
cqua
l am
ount
of q
uerc
etin
3C) I
nhib
ition
of d
eoxy
ribos
e pe
roxi
data
on th
roug
h ch
ain-
brea
king
, wat
er so
lubl
e, p
rope
rties
In
hibi
tory
act
ivity
IS co
mpa
red
to th
at o
f an
equa
l am
ount
of t
rolo
x. th
e hy
drop
hilic
anal
ogue
of v
itam
in E
3D
) Inh
ibiti
on o
f lip
osom
al a
rach
idon
ic a
cid
pero
xida
tion
thro
ugh
chai
n-br
eaki
ng.
in li
pid
phas
e, p
rope
rties
In
hibi
tory
act
ivity
is c
ompa
red
to th
at o
f an c
qual
am
ount
of
vita
min
E
P
N
m
OR
EG
AN
O E
XTR
AC
T IN
HIB
ITIO
N O
F C
U-IN
DU
CE
D P
ER
OX
IDA
TIO
N O
F P
LAS
MA
pero
xida
tion
kine
tics o
f th
e re
fere
nce
plas
ma
1
0 Bo
480
270
360
450
min
I
~ -
-__
---
lag-
time
in th
e pr
esen
ce o
r n
ot o
f que
rcet
in a
nd
or
egan
o ex
trac
ts
__
__
__
W
non
e ad
dltlo
n 8
2.5
ug
Q 1
2.5
up
_-
_~
cont
col
quer
cehn
w
ak
r m
etha
nol
extr
act
extr
act
- - __
____
_ -
__
_-_
__
_
__
_
__
. __
__
__
__
__
__
-__
slop
e in
the
pres
ence
or
not o
f que
rcet
in a
nd
or
egan
o ex
trac
ts
-.
I - __
_
F
h
Y k
t-
FIG
4 IN
HIB
ITIO
N O
F C
U-I
ND
UC
ED P
ERO
XID
ATI
ON
OF
PLA
SMA
LIP
OPR
OTE
INS
BY M
ETH
AN
OL
AN
D A
QU
EOU
S EX
TRA
CTS
OF
OR
EGA
NO
4A)
Pero
xida
tion
kine
tics o
f th
e re
fere
nce
plm
a. w
ith g
raph
ical
repr
esen
tatio
n of
lag-
time and
slopc
L =
lag-
time,
S =
slop
e, I
= in
itiat
ion
phas
e an
d P
=pro
paga
tion
phas
e 4B) H
isto
gram
s sho
w la
gtim
e va
lues
rela
tive
to d
iffer
ent a
mou
nts o
f que
rcet
in o
r ore
gano
exba
cts
4C)
His
togr
ams s
how
slo
p va
lues
rela
tive
to d
iffer
ent a
mou
nts o
f que
rcet
in o
r ore
gano
ext
ract
s
The
amou
nts u
sed
wer
e 12
5 p
g an
d 2
5 pg
of p
olyp
heno
ls, d
enve
d from 0
25 mg a
nd 0
05
mg
of dricd
leav
es
Inhi
bito
ry a
ctiv
ity is
com
pare
d w
ith th
at o
f a s
ame
amou
nt o
f que
rcet
in
ANTIOXIDANT ACTIVITY OF OREGANO EXTRACTS 463
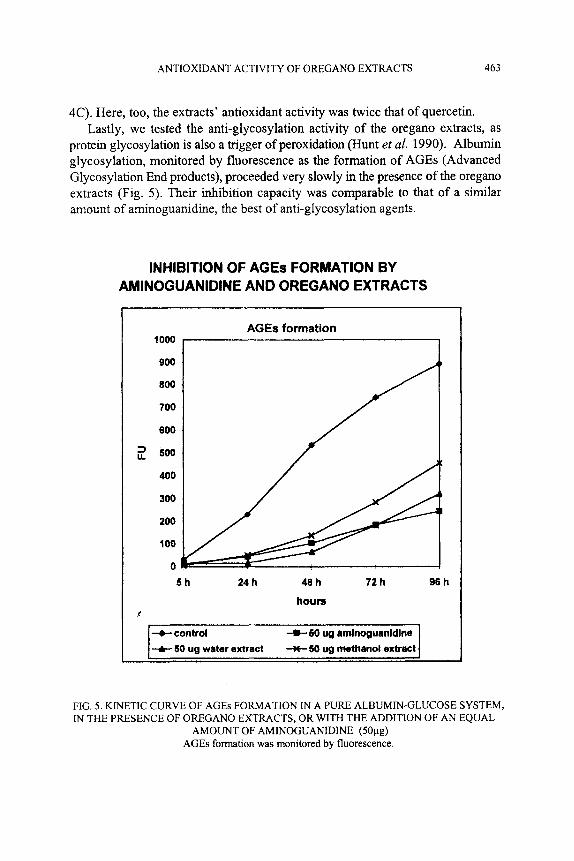
4C). Here, too, the extracts’ antioxidant activity was twice that of quercetin. Lastly, we tested the anti-glycosylation activity of the oregano extracts, as
protein glycosylation is also a trigger of peroxidation (Hunt et al. 1990). Albumin glycosylation, monitored by fluorescence as the formation of AGEs (Advanced Glycosylation End products), proceeded very slowly in the presence of the oregano extracts (Fig. 5). Their inhibition capacity was comparable to that of a similar amount of aminoguanidine, the best of anti-glycosylation agents.
INHIBITION OF AGEs FORMATION BY AMINOGUANIDINE AND OREGANO EXTRACTS
1000
900
800
700
600
500
400
300
200
100
0
AGEs formation
Sh 24 h 48 h 72 h 96 h
hours i
-4- control +SO UQ aminoguanldine -A- 50 ug water extract -X- 50 ug methanol extract
FIG. 5. KINETIC CURVE OF AGES FORMATION IN A PURE ALBUMIN-GLUCOSE SYSTEM, IN THE PRESENCE OF OREGANO EXTRACTS, OR WITH THE ADDITION OF AN EQUAL
AMOUNT OF AMINOGUANIDINE (50pg) AGES formation was monitored by fluorescence.

464 G. CERVATO ETAL.
CONCLUSIONS
This data demonstrates that the oregano extracts (both the aqueous and methanol), have a very high polyphenol content and strong but differing antioxidant properties. They proved effective in the prevention of all phases of the peroxidative process: first through the neutralization of free radicals, then through blockage of peroxidation catalysis by iron (and presumably by other transition metals), and finally through interruption of lipid radical chain reactions (chain- breaking activity). Of equal importance was their anti-glycosylation effect, essential for the preservation of the plasma proteins and their efficacy.
The fact that several antioxidant mechanisms exist makes oregano a particularly efficient blocker of an array of pro-oxidant events. Their radical-scavenger activity is optimal for blocking alimentary free radicals, their chain- breaking activity protects against peroxidized foods, and their iron-chelating activity defends the body from the deleterious action of free iron.
It is also very important to note that the water extraction conditions used in our experiments are comparable to cooking conditions, and that the amounts of the extracts used in our experiments were derived from 0.1 - 1 mg of dried leaves, quantities far less than those normally used in the diet.
As the oregano extracts proved effective in such minimal quantities, the amount of this herb necessary for good protection against peroxidation is easy to reach in a normal Mediterranean diet, notwithstanding a low bioavailability of flavonoids and polyphenols (Hollman et al. 1997).
As a nutritional ingredient which aids in preserving foods both during cooking and during digestion, oregano is highly recommended as an additive for use in the food industry.
REFERENCES
ARUOMA, 0.1. 1997. Extracts as antioxidant prophylactic agents. INFORM 8,
ARUOMA, O.I., LAUGHTON, M.J. and HALLIWELL, B. 1989. Carnosine, homocarnosine and anserine: could they act as antioxidants in vivo? Biochem.
ARUOMA, O.I., SPENCER, J.P.E., ROSSI, R., AESCHBACH, R., KHAN, A., MAHMOOD, N., MIJNOZ, A., MURCIA, A., BUTLER, J. and HALLIWELL, B. 1996. An evaluation of the antioxidant and antiviral action of extracts of rosemary and Provencal herbs. Food Chem. Toxicol. 34,449-456.
CERVATO, G., VIANI, P., CAZZOLA, R. and CESTARO, B. 1999. A fluorescence method for determination of plasma susceptibility to lipid peroxidation. Clin. Biochem. 32, 171 -1 77.
1236-1242.
J. 264, 863-869.

ANTIOXIDANT ACTIVITY OF OREGANO EXTRACTS 465
CERVATO, G., VIANI, P., MASSERINI, M., DI IORIO, C. and CESTARO, B. 1988. Studies on peroxidation of arachidonic acid in different liposomes below and above phase transition temperature. Chem. Phys. Lipids 49, 135-139.
CONSTANTIN, M., BROMONT, C., FICKAT, R. and MASSINGHAM, R. 1990. Studies on the activity of bepridil as a scavenger of free radicals. Biochem. Pharmacol. 40, 1615-1622.
DECKER, E.A. 1995. The role of phenolics, conjugated linoleic acid, camosine, and pyrroquinoline quinone as nonessential dietary antioxidants. Nutr. Rev. 53,
GUGLIUCCI, A. and STAHL A.J.C. 1995. Low density lipoprotein oxidation is mhibited by extracts of ilex paraguariensis. Biochem. Mol. Biol. Int. 35,47-56.
GUTTERIDGE, J.M.C. 1995. Lipid peroxidation and antioxidants as biomarkers of tissue damage. Clin.Chem. 41, 1819-1828.
GUTTERIDGE, J.M.C. and QUINLAN, G.J. 1992. Antioxidant protection against organic and inorganic oxygen radicals by normal human plasma: The important primary role for iron-binding and iron-oxidising proteins. Biochim. Biophys. Acta 1159, 248-254.
HALLIWELL, B., GUTTERIDGE, J.M.C. and ARUOMA, 0.1. 1987. The deoxyribose method: a simple “test-tube” assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem. 165, 2 15-2 19.
HOLLMAN, P.C.H., VAN TRIJP, J.M.P., BWSMAN, M.N.C.P., VAN DER GAAG, M.S., MENGELERS, M.J.B., DE VRIES, J.H.M and KATAN, M.B. 1997. Relative bioavailability of the antioxidant flavonoid quercetin from various foods in man. FEBS Lett. 418, 152-156.
HUNT, J.V., SMITH, C.C.T. and WOLFF, S.P. 1990. Autoxidative glycosylation and possible involvement of peroxides and free radicals in LDL modification by glucose. Diabetes 39, 1420-1424.
KIKUZAKI, H. and NAKATANI, N. 1989. Structure of a new antioxidative phenolic acid from origano (Origanum vulgare L.). Agric. Biol. Chem. 53, 519- 524.
KUBOW, S. 1992. Routes of formation and toxic consequences of lipid oxidation products in foods. Free Rad. Biol. Med. 12, 63-8 1.
LAGOURI, V. and BOSKOU, D. 1996. Nutrient antioxidants in origano. Int. J. Food Sci. Nutr. 47,493-497.
LEWIN, G. and POPOV, I. 1994. The antioxidant system of the organism. Theoretical basis and practical consequences. Med. Hypothesis 42,269-275.
MORIMITSU, Y., YOSHIDA, K., ESAKI, S and HIROTA, A. 1995. Protein glycation mhibitors from Thyme (Thymus vulgaris). Biosci. Biotech. Biochem.
SINGLETON, V.L. and DRAPER, D.E. 1964. The transfer of polyphenolic compounds from grape seeds into wines. Am. J. Enol. Viticult. 15, 34-40.
49-58.
59, 20 18-202 1.

Copyright © 2022 FDOKUMEN




















