
Wild Edible Plants of the Sikkim Himalaya: Marketing, Value Addition and Implications for Management
Upload
khangminh22Category
view
0download
0
Isolation and Characterization of Extracts from Wild Edible and Non-edible
Mushrooms in Zimbabwe
By
Tsungai Reid
Thesis submitted in fulfilment of the requirements for the degree of Doctor of
Philosophy in Biochemistry
Department of Biochemistry
Faculty of Science
University of Zimbabwe
July 2019
Supervisor: Professor Takafira Mduluza
Co-supervisor: Doctor Chenjerayi Kashangura

ii
DECLARATION
I hereby declare that the material contained in this thesis is my own original work and
has not been submitted for a degree in any other university.
Tsungai Reid

iii
ABSTRACT
Poor nutrition and an increasing emergence of infectious diseases, particularly in developing
countries, represent major threats to human health. Mushrooms are known to possess
numerous bioactivities such as antioxidant and antimicrobial activities. However, the role of
local Zimbabwean mushrooms in human health remains largely unexplored. In this study, ten
local mushrooms, namely; Amanita zambiana, Boletus edulis, Cantharellus heinemannianus,
C. miomboensis, C. symoensii, Lactarius kabansus, Amanita species, Coprinus species,
Ganoderma lucidum and Trametes strumosa, collected from selected areas of Zimbabwe,
were characterised for nutritional, secondary metabolites and biological activity. The main
research hypothesis was that the extracts of different wild edible and non-edible mushrooms
of Zimbabwe contained nutritional, antibacterial and anti-HIV components. Determination of
protein content of mushroom powder was carried out using the Kjeldhal method while the
qualitative analysis of carbohydrates was done using Molisch’s and Benedict’s tests. The
lectin content was detected by the haemagglutination activity of mushrooms against sheep and
goat erythrocytes. The total protein content ranged from 9.3% ± 0.99 to 30.8% ± 1.27.
Amanita sp., B. edulis and L. kabansus were able to agglutinate both sheep and goat
erythrocytes. Amanita zambiana showed high levels of both carbohydrates and reducing
sugars. Crude extracts from 50 mushrooms,were obtained using hot water, cold water,
methanol, ethanol or acetone were used to determine the protein and total phenol contents
using the Folin Ciocalteu (FC) and Lowry C methods, respectively. Boletus edulis had high
protein and total phenolic content (2.02 mg ± 0.1 and 503.70 mg ± 20.7, respectively). The
antibacterial effect of the crude extracts against Escherichia coli, Salmonella typhi,
Staphylococcus aureus and Streptococcus pneumoniae was determined using the agar disc
diffusion method. The extracts exhibited antibacterial properties against the four bacteria
tested. Sixteen of the extracts that showed high levels of bacterial growth inhibition were
selected for further characterization. A total of 131 compounds (CP1 – CP131) were isolated
using Preparative Thin Layer Chromatography from the 16 extracts. Thirteen of the isolated
compounds exhibited high inhibitory activity against the growth of S. typhi (82 to 99.8%).
One of the compounds (CP50) inhibited S. aureus growth (87.5%). Identification of
compounds responsible for the high antibacterial activity was carried out using LC-MS. The
tepernoids (boviquinone 4, cavipetin D, goshonoside, lucidenic acid M, 26-methyl nigranoate
and notoginsenoside); phospholipid (C16 sphinganine) and fatty acid derivatives (11-amino-
undecanoic acid, z-13-oxo-9-octadecanoic acid, palmitic amide, sorbitan oleate and
stearamide) were identified as compounds partly responsible for the antibacterial activity
observed. The effect of the crude extracts against HIV replication was determined using the
anti-HIV-1c reverse transcriptase (RT) and HIV-1c p24 ELISA assays. The cold water extract
of L. kabansus demonstrated the highest level of HIV-1 RT inhibitory activity (92.6%), whilst
the hot water extracts from Coprinus species and C. heinemannianus exhibited high potent
levels of HIV-1c p24 inhibitory activity, with IC50 values of 24.3 µg/ml and 33.8 µg/ml,
respectively. This study revealed for the first time the presence of bioactive compounds in the
local Zimbabwean mushrooms studied. The hypothesis that the extracts of different wild
edible and non-edible mushrooms of Zimbabwe contained nutritional, antibacterial and anti-
HIV components was proven to be true. The information obtained will potentially enable
development of more efficient foods as medicine in the country, based on the wealth of
information generated on the health-promoting properties of the ten mushrooms studied. From
this study, the development of anti-bacterial and anti-HIV therapeutic agents from the local
mushrooms is recommended, due to the presence of antimicrobial compounds identified in
the mushroom extracts. The study also recommends the development of edible local
mushrooms such as A. zambiana, B. edulis and L. kabansus into functional food products due
to their high nutritive value.

iv
Dedicated with love to
my dear husband, Andrew John Cheyne Reid and
our lovely children, Simbarashe Michael, Tinashe Anesu, Rumbidzwaiishe Esther,
Joshua Munashe and Nyashadzaishe Mercy

v
ACKNOWLEDGEMENTS
I am deeply grateful to my Heavenly Father, the Almighty God, for the gift of life, His
unfailing love to me and in whose strength l could do all things (Phillipians 4:13). I love you
LORD and without You l am nothing.
Many people have helped me along the journey, for this work to be a reality and although l
may not name everyone here, l am so thankful. My sincere appreciation goes to my mentor
and supervisor, Professor Takafira Mduluza. Thank you Prof. for your guidance, expert
advice and support throughout this project and for constantly pushing me to achieve my
career goals. Thank you to my co-supervisor, Dr Chenjerayi Kashangura, for your advice,
support and expertise in the mushroom field that l could tap into anytime. To my project
mentors, Professor Mudadi Albert Benhura and Dr Catherine Chidewe, l am most grateful for
your valuable input, especially during the early stages of my research.
To Dr Sikhulile Moyo, Dr Simani Gaseitsiwe and Mr Terrence Mohammed at the Botswana
Harvard Partnership HIV Reference Laboratory (BHP), thank you so much for all your
support, for hosting me and generously affording me the opportunity to do part of my research
work at your institution. I also extend my gratitude to Dr Lucy Mupfumira and Mr Wonderful
Choga and many hospitable staff members at BHP for your great help and support. I am
thankful to Dr Melvin Leteane and Boingotlo Raphane from the University of Botswana for
all your assistance and laboratory skills on HIV research that you taught me while at BHP.
I acknowledge the support of Professor James Hakim, Dr Tariro Makadzange, Professor Val
Robertson and Professor Dexter Tagwireyi, through your wisdom and mentorship. To the
Chairman of the Biochemistry Department, Professor Stanely Mukanganyama, the Chief
Technician, Mrs Elizabeth Chinyanga, my work colleagues, Dr Farisai Chidzwondo, Dr Fiona
Robertson, and the entire staff in the Department, l am very thankful to you all for your input
and support. To Professor Christopher Chetsanga and Professor Idah Sithole-Niang, thank
you for the molecular techniques and skills you taught me during my earlier academic studies
and research. Thank you again Professor Chetsanga for your unwavering support, wisdom and
mentorship.

vi
I acknowledge and greatly appreciate the financial support l received from the Letten
Foundation and University of Zimbabwe Research Grant to enable this study to become a
reality. Thank you so much Professor Babill Stray-Pederson for believing in me. To Mrs
Auxillia Mazhambe, thank you for your support and facilitation. Thank you to the Letten
Foundation family for all your input.
I want to acknowledge the impact that my spiritual parents, Pastors Tom (my dear dad) and
Bonnie Deuschle (my dear mum), have had on my life. Your teachings, your message of
reformation over the years and how you and your family model every aspect of what you
teach us, keeps inspiring me to strive to achieve more and never give up. Pastor Bonnie, your
prophetic voice and actions have been a source of encouragement to me all the way. I love
you and thank God for you and family. To my dear mum Refiloe, siblings, family (Reid
family) in New Zealand and grandmums Esther and Eva, thank you so much for all your
support and encouragement.
Last but certainly not least I would like to express my heartfelt gratitude to my dear husband,
Andrew John Cheyne for your kindness, patience, love, encouragement and unwavering
support. Thank you for being my number one cheer leader. Thank you for releasing me to do
my research work even at odd hours and in distant lands, while you minded our family and
the hectic schedules, with your own busy schedule. You are amazing! To my lovely sons and
daughters: Simbarashe, Tinashe, Rumbidzwaiishe, Joshua and Nyashadzaishe; thank you so
much for your kindness, love and support. Your keen interest in what l was doing and
cheering me on meant a lot and l truly appreciate. You and your dad are truly a great blessing
to me. I treasure and love you and thank God for the precious gift of having all of you in my
life.

vii
TABLE OF CONTENTS
Declaration……………………………………………………………………………... (ii)
Abstract………………………………………………………………………………… (iii)
Dedication……………………………………………………………………………… (iv)
Acknowledgements…………………………………………………………………….. (v)
Table of Contents………………………………………………………………………. (vii)
List of Figures………………………………………………………………………….. (xii)
List of Tables…………………………………………………………………………… (xiv)
List of Abbreviations………………….……………………………………………….. (xvi)
List of Appendices...………………………………………………………...………...... (xix)
CHAPTER ONE: INTRODUCTION
1.1 BACKGROUND INFORMATION……………………………………………. 2
1.2 PROBLEM STATEMENT AND JUSTIFICATION…………………………… 3
1.3 OBJECTIVES OF THE STUDY……………………………………………….. 5
1.3.1 Main Objective of the Study…………………………………………….. 5
1.3.2 Specific Objectives of the Study………………………………………… 5
1.4 RESEARCH QUESTIONS……………………………………………………… 6
1.4.1 Other Research Questions……………………………………………….. 6
1.5 RESEARCH HYPOTHESES …………………………………………………… 6
1.5.1 Other Research Hypotheses……………………………………………… 6
CHAPTER TWO: LITERATURE REVIEW
2.1 MUSHROOM BIOLOGY AND CLASSIFICATION………………………….. 8
2.1.1 Classification of Mushrooms……………………………………………. 9
2.1.2 Types of Mushrooms ……………………………………………………. 12
2.1.2.1 Edible Mushrooms……………………………………………….. 12
2.1.2.2 Non-edible Mushrooms………………………………………….. 13
2.2 HUMAN HEALTH BENEFITS OF MUSHROOMS AND THEIR EXTRACTS 14
2.2.1 Mushrooms as Food and Medicine………………………………………. 14

viii
2.2.1.1 Mushrooms as Source of Proteins………………………………… 15
2.2.1.2 Mushrooms as Source of Carbohydrates…………………………. 16
2.2.1.3 Phenolic Compounds and Antioxidant Properties of Mushrooms… 17
2.2.2 Mushroom Extracts as Medicine……………………………………........ 18
2.2.2.1 Antibacterial Properties of Mushrooms………………………….. 19
2.2.2.2 Antiviral Properties of Mushrooms………………………………. 21
2.2.2.3 Mushroom Lectins and their Role in Human Health.……………. 22
2.3 CHARACTERIZATION OF MUSHROOM EXTRACTS……………………… 23
2.3.1 Methods of Extraction……………………………………………………. 23
2.3.1.1 Choice and Effect of the Extracting Solvent……………………… 24
2.3.2 Methods of Seperation of Constituents from Mushroom Extracts……….. 26
2.3.2.1 Thin Layer Chromatography (TLC)……………………………... 26
2.3.2.2 High Performance Liquid Chromatography (HPLC)……………… 27
2.3.3 Methods of Identification of Compounds………………………………... 27
2.3.3.1 Ultra Violet (UV)-Visible Spectroscopy………………………….. 28
2.3.3.2 Liquid Chromatography- Mass Spectroscopy (LC-MS)…………... 28
2.3.4 Classes of Secondary Metabolites in Mushrooms……………………….. 29
2.3.4.1 Alkaloids…………………………………………………………. 29
2.3.4.2 Glycosides………………………………………………………… 30
2.3.4.3 Flavonoids………………………………………………………… 30
2.3.4.4 Phenolics………………………………………………………….. 31
2.3.4.5 Terpenoids………………………………………………………… 32
2.3.4.6 Steroids and Sterols……………………………………………….. 32
2.3.5 Methods of Detecting Antibacterial Activity…………………………….. 32
2.3.5.1 Agar Disc and Well Diffusion Methods…………………………. 33
2.3.6 Methods of Detecting Anti-HIV Activity…………………………..…….. 34
2.3.6.1 MTT or XTT Assay………………………………….……………. 35
2.3.6.2 HIV-1 p24 Expression Assay..………………………..……............ 36
2.3.6.3 HIV-1 Reverse Transcriptase (RT) Inhibition Assay…………….. 36
2.4 MUSHROOMS FOUND IN ZIMBABWE…………………………………………... 37

ix
CHAPTER THREE: MATERIALS AND METHODS
3.1 COLLECTION, IDENTIFICATION AND PREPARATION OF
MUSHROOMS…………………………………………………………………... 42
3.2 CHARACTERIZATION OF MUSHROOMS…………………………………… 42
3.2.1 Quantitative Determination of the Total Protein Content………………… 42
3.2.2 Determination of Carbohydrates………………………………………….. 43
3.2.3 Determination of Lectins…………………………………………………. 44
3.2.3.1 Extraction of Mushroom Crude Protein…………………………… 44
3.2.3.2 Preparation of the Red Blood Cells……………………………… 44
3.2.3.3 Haemaggutination Assay………………………………………….. 44
3.3 CHARACTERIZATION OF MUSHROOM CRUDE EXTRACTS…………….. 45
3.3.1 Preparation of Crude Mushrooms Extracts……………………………….. 45
3.3.2 Determination of Protein Content………………………………………… 46
3.3.3 Determination of Total Phenolic Content……………………………........ 46
3.3.4 Determination of Antibacterial Activity of Crude Extracts….…………… 46
3.3.4.1 Analysis of Mushroom Crude Extracts Showing High
Antibacterial Activity by Spectrophotometry……………………….......... 47
3.3.4.2 Analysis of Crude Extracts by TLC……………………………….. 47
3.3.4.3 Selection of Mobile Phase for Preparative Thin Layer
Chromatography (PTLC) ………………………………….………….…... 48
3.3.4.4 Isolation of Compounds from Mushroom extracts by PTLC……… 48
3.3.4.5 Screening of Isolated Fractions for Antibacterial Activity………. 49
3.3.4.6 Identification of Antibacterial Compounds of Mushroom
Extracts by Non-targeted LC - MS………………………………………… 50
3.3.5 Determination of Anti-HIV Activity……………………………………… 51
3.3.5.1 Determination of Anti-HIV-1 Reverse Transcriptase Activity…..... 51
3.3.5.2 Cytotoxicity Assay of the Mushroom Extracts…………………….. 52
3.3.5.3 Determination of the in vitro Anti-HIV-1 Activity Using
HIV p24 Expression Assay………………………………………… 53
3.3.5.4 Assay for HIV-1c Induced Cytopathic Effect…………………. ….. 54
3.3.5.5 Analysis of the Anti-HIV Mushroom Crude Extracts by
Non-targeted LC – MS…………………………………………….. 55

x
3.4 STATISTICAL ANAYLSIS……………………………………………………… 55
CHAPTER FOUR: RESULTS
4.1 COLLECTION, IDENTIFICATION AND PREPARATION OF
MUSHROOMS…………………………………………………………………… 57
4.2 CHARACTERIZATION OF MUSHROOMS…………..……………………….. 58
4.2.1 Quantitative Determination of the Total Protein Content…………………... 58
4.2.2 Qualitative Determination of Carbohydrates……………..………………… 59
4.2.3 Determination of Lectins…………………………………………………… 61
4.3 CHARACTERIZATION OF MUSHROOM EXTRACTS…………………….. 63
4.3.1 Determination of Protein Content of the Crude Extracts…………………. 63
4.3.2 Determination of Total Phenolic Content………………………………… 64
4.3.3 Determination of the Antibacterial Activity of Mushroom
Crude Extracts………….............................................................................. 66
4.4 CHARACTERIZATION OF MUSHROOM EXTRACTS SHOWING HIGH
ANTIBACTERIAL ACTIVITY….......................................................................... 74
4.4.1 Analysis of the Extracts using Absorption Spectroscopy and TLC………. 74
4.4.2 Separation of Mushroom Crude Extracts by TLC………………………… 78
4.4.3 Isolation of Components of Mushroom Crude Extracts by PTLC……….. 84
4.4.4 Determination of the Antibacterial Activity of the Isolated Fractions……. 84
4.4.5 Identification of the Most Potent Antibacterial Compounds of Mushroom
Extracts by LC – MS…………………………………………………....... 86
4.5 DETERMINATION OF THE ANTI-HIV ACTIVITY OF MUSHROOM
CRUDE EXTRACTS…….………………………………………………………. 88
4.5.1 Determination of Anti-HIV-1 Reverse Transcriptase Activity…………… 88
4.5.2 Cytotoxicity Assay of Mushroom Extracts …….………………………… 90
4.5.3 Determination of the In Vitro Anti-HIV-1 Activity using HIV p24
Antigen Expression Assay………………………………………………... 92
4.5.4 Analysis of the Anti-HIV Mushroom Crude Extracts by Non-Targeted
LC – MS………………………………………………………………….. 94

xi
CHAPTER FIVE: DISCUSSION
5.1 CHARACTERIZATION OF MUSHROOM FRUITING BODIES……………... 97
5.1.1 Quantitative Determination of the Total Protein Content………………… 97
5.1.2 Qualitative Determination of Carbohydrates……………………………... 98
5.1.3 Determination of Lectins…………………………………………………. 99
5.2 CHARACTERIZATION OF MUSHROOM CRUDE EXTRACTS…………….. 99
5.2.1 Determination of Protein Content of Crude Extracts……………………... 99
5.2.2 Determination of Total Phenolic Content of Crude Extracts …………...... 100
5.2.3 Determination of Antibacterial Activity of Crude Extracts ……………… 101
5.2.4 Characterization of Mushroom Extracts Showing High Antibacterial
Activity……………………………………………………………………. 105
5.2.4.1 Analysis of the Extracts Using Absorption Spectroscopy and
TLC……………………………………………………………….. 105
5.2.4.2 Separation of Mushroom Compounds by TLC and Isolation
of Components of Mushroom Extracts by PTLC……………….. 106
5.2.4.3 Determination of Antibacterial Activity of the
Isolated Compounds………………………………………..……… 107
5.2.4.4 Analysis of Antibacterial Components of Mushroom Extracts by
LC - MS ………………………………………………………...... 108
5.2.5 Determination of the Anti-HIV Activity of Mushroom Crude Extracts…. 112
5.2.5.1 Determination of Anti-HIV-1 Reverse Transcriptase Activity……. 112
5.2.5.2 Cytotoxicity Assay of Mushroom Extracts………………………… 113
5.2.5.3 Determination of the in vitro Anti-HIV-1 Activity Using p24
Antigen Expression Assay……………………………………….. 113
5.2.5.4 Analysis of the Anti-HIV Mushroom Crude Extracts by
LC – MS…………………………………………………………... 115
6.0 CONCLUSION ……………………………………………………………….. 117
6.1 RECOMMENDATIONS…………………………………………………………. 119
7.0 REFERENCES………………………………………………………………….. 121
8.0 APPENDICES…………………………………………………………………… 137

xii
LIST OF FIGURES
Figure No. Title Page No.
Figure 2.1: Agaricus bisporus 8
Figure 2.2: Amanita fulvoalba 9
Figure 2.3: Overview of the six main classes of secondary metabolites with
representative examples 30
Figure 2.4: Agar disc diffusion assay showing zones of inhibitions of bacterial
growth 34
Figure 2.5: Antiretroviral targets in HIV Life Cycle 35
Figure 2.6: HIV-1 ELISA p24 antigen assay 36
Figure 2.7: Different types of mushrooms commonly found in Zimbabwe and
used in this study 39
Figure 4.1: Sliced pieces of Cantharellus miomboensis during the drying process 58
Figure 4.2: Percentage total protein content of nine different mushrooms 59
Figure 4.3: Analysis of carbohydrates showing positive tests for (A) Benedict’s
test for reducing sugars and (B) Molisch’s test for carbohydrates 60
Figure 4.4: Haemagglutination assay of three of the ten mushroom species
with sheep erythrocytes 62
Figure 4.5: Protein content of mushrooms extracted by methanol, ethanol,
acetone, cold water and hot water 64
Figure 4.6: Total phenolic content of mushrooms extracted by methanol,
ethanol, acetone, cold water and hot water 66
Figure 4.7: Representative UV spectra obtained from acetone (Ac) and water (H2O)
extracts of mushrooms 75
Figure 4.8: Representative UV spectra obtained from ethanol (Eth) extracts of
mushrooms 75
Figure 4.9: Representative UV spectra obtained from methanol (Meth) extracts of
mushrooms 76
Figure 4.10: Representative UV spectra obtained from different solvent extracts of
the same mushroom 77
Figure 4.11: Representative chromatograms of mushrooms extracts developed in
TEM solvent and sprayed with vanillin - sulphuric acid 78

xiii
Figure 4.12: Representative assay for anti-HIV RT activity of mushroom crude
extracts 89
Figure 4.13: HIV-1 Reverse transcriptase inhibitory activity of crude extracts from
edible and non-edible mushrooms 90
Figure 4.14: Representative curve used to derive the CC50 values of the different
mushroom extracts 91
Figure 4.15: Anti-HIV activity of the hot water extracts of Coprinus sp. and C.
heinemannianus, showing a dose dependent inhibitory activity 94
Figure 5.1: Structure of lucidenic acid M 109
Figure 5.2: Structure of cavipetin D 110
Figure 5.3: Structure of palmitic amide 110
Figure 5.4: Structure of phytosphingosine 111

xiv
LIST OF TABLES
Table No. Title Page No.
Table 2.1: Simplified classification of the major groups of fungi according to
Ainsworth, 1971 11
Table 2.2: Effect of solvents in extracting different components from
biological sources 25
Table 4.1: Different types of mushrooms collected from the woodlands of
Zimbabwe 57
Table 4.2: Qualitative analysis of carbohydrate content in mushrooms using
Molisch’s test and Benedict’s tests 61
Table 4.3: Haemagglutination assay of the ten mushroom species with sheep
and goat erythrocytes 62
Table 4.4: Specificity activity of crude extracts of mushrooms showing
haemagglutination activity 63
Table 4.5: Antibacterial activities of methanol extracts of mushrooms on
test organisms 67
Table 4.6: Antibacterial activities of ethanol extracts of mushrooms on test
organisms 68
Table 4.7: Antibacterial activities of acetone extracts of mushrooms on test
organisms 69
Table 4.8: Antibacterial activities of cold water extracts of mushrooms on test
organisms 70
Table 4.9: Antibacterial activities of hot water extracts of mushrooms on test
organisms 71
Table 4.10: Extracts exhibiting high antibacterial activity (9 – 14 mm zones
of inhibition) which were selected for further study 73
Table 4.11: Absorption spectrum peaks obtained from the selected crude extracts
of mushrooms that showed high antibacterial activity 74
Table 4.12: The Rf values of fractions separated from selected mushroom extracts
using different TLC mobile phases 80
Table 4.13: Number of fractions obtained after scrapping bands from each of the
separated crude extract on the PTLC plate 84

xv
Table 4.14: Percentage inhibition of growth of S. typhi and S. aureus by the
fourteen potent compounds isolated from different mushroom crude
extracts using PTLC 86
Table 4.15: Compounds identified from seven of the most potent components
isolated from the antibacterial crude extracts of mushrooms 87
Table 4.16: The CC50 values of mushroom extracts showing varying levels of
inhibitory effects to the growth of PBMCs 92
Table 4.17: Inhibitory effects of different mushroom extracts on HIV-1 replication 93
Table 4.18: Cytotoxicity and anti-HIV-1 activity of the hot water extracts of
Coprinus species and C. heinemannianus using HIV-1c (MJ4) in
PBMC cells 94

xvi
LIST OF ABBREVIATIONS
A. bisporus Agaricus bisporus
ANOVA Analysis of variance
AZT Azidothymidine
A. zambiana Amanita zambiana
ABTS 2,2'-Azino-bis (3-ethylbenzothiazoline-6-sulfonic acid)
Amanita sp. Amanita species
Anti-DIG-POD Anti-digoxigenin peroxidase
B. edulis Boletus edulis
BSA Bovine serum albumin
CC50 50% cytotoxic concentration
CD4 Cluster of differentiation 4
CFU Colony forming units
CM Chloroform: methanol
CO2 Carbon dioxide
C. heinemannianus Cantharellus heinemannianus
C. miomboensis Cantharellus miomboensis
C. symoensii Cantharellus symoensii
DNA Deoxyribonucleic acid
DMSO Dimethyl sulphoxide
DDDP DNA-dependant-DNA polymerase
DW Dry weight
EEW Ethyl acetate:ethanol:water
EMW Ethyl acetate:methanol:water
E. coli Escherichia coli
EC50 Half maximal effective concentration
ELISA Enzyme linked immunosorbent assay
EDTA Ethylenediaminetetraacetic acid
FBS Fetal bovine serum
FC Folin – Ciocalteu
G. lucidum Ganoderma lucidum
GAE Gallic acid equivalent
GC – MS Gas chromatography – mass spectroscopy

xvii
HAU Hemagglutination unit
HIV-1c Human immunodeficiency virus type 1 subtype C
HIV-1 gp120 HIV-1 surface glycoprotein 120
HPLC High performance liquid chromatography
HRP Horse radish peroxidase
IL-1 Interleukin 1
IL-2 Interleukin 2
IR Infra-red
LC–DAD Liquid chromatography – diode array detection
LC – MS Liquid chromatography – mass spectroscopy
LC – NMR Liquid chromatography – nuclear magnetic resonance spectroscopy
L. edodes Lentinus edodes
L. kabansus Lactarius kabansus
MAE Microwave assisted extraction
Mg Milligram
m/z Mass-to-charge ratio
MTT 3-(4,5-dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide
Oligo-dT Oligonucleotide-deoxythymine
PBMCs Peripheral blood mononuclear cells
PBS Phosphate buffered saline
PHA Phytohaemagglutinin
Poly A Poly adenine
Psig Per square inch gauge
PTLC Preparative thin layer chromatography
Q-TOF Quadrupole-time of flight mass spectrometer
RPMI-1640 Roswell Park Memorial Institute-1640 medium
RNA Ribonucleic acid
RNase H Ribonleclease H
Rf Retention factor
S. aureus Staphylococcus aureus
S. pneumoniae Streptococcus pneumoniae
SD Standard deviation
SI Selectivity index

xviii
TEM Toluene:ethylacetate
TLC Thin layer chromatography
TLC-DB Thin layer chromatography – direct bioautography
TMB 3,3',5,5'-Tetramethylbenzidine
T. strumosa Trametes strumosa
UAE Ultrasound assisted extraction
UV Ultra violet
V Voltage
v/v Volume per volume
w/v Weight per volume
XTT Sodium 3-[1-(phenylamino)-carbonyl]- 3,4-tetrazoliumbis (4-methoxy-
6-nitro) benzene-sulfonic acid hydrate

xix
LIST OF APPENDICES
Appendix No. Title Page No.
Appendix 8.1: Publications arising from the work in this thesis 137
Appendix 8.2: Preparation of reagents 138
Appendix 8.3: Percentage inhibition of bacterial growth by components
isolated from different mushroom extracts using PTLC 139
Appendix 8.4: LC-MS Chromatograms of isolated components of
mushroom samples that exhibited high anti-bacterial
activity analyzed in positive mode with a column 142
Appendix 8.5: LC-MS profiles of some of the mushroom crude extracts
that exhibited high anti-HIV activity analyzed in positive
mode with a column 143

1
CHAPTER ONE:
INTRODUCTION

2
1.0 INTRODUCTION
1.1 BACKGROUND INFORMATION
Mushrooms are a group of fungi that belong to the higher phyla Ascomycota and
Basidiomycota, with distinctive fruiting bodies and reproductive structures. The fruiting
bodies especially basidiomes are usually umbrella shaped structures that produce spores in
large numbers (Camassola, 2013; Florence and Balasundaran, 2000). Mushrooms lack
chlorophyll, unlike green plants and obtain nutrition from non-living organinc matter or living
plants in their surroundings. Some mushrooms are edible, such as Cantharellus miomboensis
while other mushrooms are extremely poisonous, such as Amanita phalloides. A number of
mushroom varieties have global economic importance, through being all year round cultivated
delicacies considered as food and medicine. These include button (Agaricus bisporus) and
shiitake (Lentinus edodes) mushrooms which are considered as the two most cultivated
mushrooms worldwide (Reagile, 2011, Stamets, 2000).
Since ancient times, mushrooms have been recognized as functional foods (foods containing
health-giving additives) and as a source for the development of medicines and nutraceuticals
throughout the world, including being prescribed for the treatment of various human
microbial diseases (Alves et al., 2012; Gbolagade and Fasidi, 2005; Halpern, 2007; Prasad et
al., 2015; Ziarati and Ghasemynezhad – Shanderman, 2015). Medicinal mushroom
application can be through concentrates or powdered forms on hot water extracts and
essences, which are applied as alternative medicine regularly in Korea, China, Japan and
eastern Russia (Prasad et al., 2015). Species like Inonotus obliquus, Coprinus comatus,
Ganoderma lucidum, Fomitopsis officinalis, Piptoporus betulinus and Fomes fomentarius
have been applied in the treatment of gastrointestinal disorders, diabetes, haemorroids,
bronchial asthma and different types of cancers (Ardigo, 2016; Nowacka et al., 2015).

3
Furthermore, many varieties of mushrooms such as Agaricus bisporus, A. brasiliensis, A.
subrufescens, Lentinus edodes, Pleurotus florida and Tricholoma giganteum, contain
biologically active compounds that have been reported to exert immunomodulating,
hepatoprotective (Moukha et al., 2011), antifibrotic, anti-inflammatory, antidiabetic (Moon
and Lo, 2013), antioxidant (Chowdhury et al., 2015; Gan et al., 2013), antiviral (Wang and
Ng, 2004), antimicrobial (Padmavathy et al., 2014; Tehrani et al., 2012) and anticancer
properties (Durgo et al., 2013; Geethangili et al., 2013; Moukha et al., 2011). These
compounds include phenolic compounds, terpenes, flavonoids (Ramesh and Pattar, 2010),
polysaccharides, triterpenoids (Chang and Wasser, 2012; Reis et al., 2012), glycopeptides,
ribonucleases and lectins (Moukha et al., 2011).
The number of mushroom species identified all over the world is estimated at about 140 000
and of these only 22 000 have been investigated (Faridur et al., 2010; Hawksworth, 2001;
Nowacka et al., 2015). Despite their potential and enormous diversity in tropical ecosystems,
many species of mushrooms have not been tapped, particularly in the field of medicine
(Prasad et al., 2015). Thus, considering that many varieties of mushrooms are already a
valued source of active ingredients, this study embarked on assessing the nutritional and
chemical composition, as well as the biological potential, of ten mushroom species growing in
the wild in Zimbabwe. The mushrooms used in this study included both edible and non-edible
species.
1.2 PROBLEM STATEMENT AND JUSTIFICATION
In recent years, there has been an upsurge of interest in mushrooms in various countries not
only as a health food but also due to the presence of biologically active compounds with
potential therapeutic properties (Prasad et al., 2015). Poor nutrition and an increasing

4
emergence of infectious diseases, caused by pathogenic bacteria, fungi and viruses, represent
major threats to human health (Lindahl and Grace, 2015; Shridhar et al., 2015). There is a
general agreement that many chronic health problems worldwide, relate mainly to unhealthy
food choices. Obesity and dehydration and ailments like cardiac problems, diabetes mellitus
and arthritis are on the increase, especially in developing countries (Shridhar et al., 2015).
Drug resistance continues to present a large and growing problem in treatment of infections
that account for most of Africa’s disease burden, including Human Immunodefficiency Virus
(HIV) infections, respiratory and diarrheal diseases (Avert Newsletter, 2018; Kharsany and
Karim, 2016; Okon et al., 2013; Padmavathy et al., 2014; Sangeeth et al., 2014). Sub-Saharan
Africa continues to have the highest burden of HIV/Acquired Immune Deficiency Syndrome
(AIDS) worldwide and although the clinical management of HIV infection has greatly
improved, resistance to antiretroviral drugs has emerged (Hamers et al., 2013; Wainberg et
al., 2011; WHO Report, 2012). Hence, there is a need for continuous search and development
of novel antimicrobial/antiviral substances from different biological sources to minimize the
threat of further antimicrobial/antiretroviral resistance (Padmavathy et al., 2014; Shah et al.,
2014). Due to the production of a large variety of secondary metabolites with interesting
biological actions, mushrooms are reservoirs of valuable chemical resources and can be used
as a source for nutritional supplements and biotherapeutics (Prasad et al., 2015). However,
despite the enormous therapeutic potential of mushrooms, there is very little research and
awareness on local mushrooms as a healthy food and as an important source of biologically
active substances with medicinal value in most of the African countries. Although Zimbabwe
is rich in mushroom diversity, very little work has been carried out on profiling the
nutritional, secondary metabolite composition and therapeutic value of mushrooms in
Zimbabwe. In addition, the potential of mushrooms as a source of new drugs is still largely
unexplored. Nowadays, it is highly desirable to characterize and search for natural

5
constituents with health benefits due to the burden of civilization diseases (including HIV)
affecting humans. Thus, the main goal of this study was to determine the nutritional and
chemical composition of mushrooms, as well as the antibacterial and anti-HIV potential of the
aqueous and organic extracts from selected wild mushrooms found in Zimbabwe. The
chemical composition and intensity of the therapeutic effect of the extracts has been reported
to be dependent upon mushroom species, extracting solvent, concentration of mushroom
extract and the organism being tested (Alves et al., 2012; Pushpa and Purushothama, 2010).
In this study, different extracting media were also employed. The information obtained in this
study will inform further studies needed for a better understanding of the health-promoting
properties of mushroom constituents. These properties will enhance the use mushrooms in
preventing and treating human diseases.
1.3 OBJECTIVES OF THE STUDY
1.3.1 Main Objective of the Study
The main objective of the study was to characterize mushrooms and their extracts through
analysis of the nutritional and secondary metabolite content, as well as the antibacterial and
anti-HIV properties of selected wild edible and non-edible mushrooms found in Zimbabwe.
1.3.2 Specific Objectives of the Study
The specific objectives of the study were to:
1.3.2.1 determine the nutritional and secondary metabolite composition of wild edible and
non-edible mushrooms found in Zimbabwe.
1.3.2.2 determine the antibacterial and anti-HIV effects of extracts of wild edible and non-
edible mushrooms.

6
1.3.2.3 isolate, characterize and identify signature compounds from mushroom extracts
exhibiting high antimicrobial activity.
1.3.2.4 investigate the effect of various solvents in extracting bioactive compounds from
mushrooms.
1.4 MAIN RESEARCH QUESTION
Do extracts of different wild edible and non-edible mushrooms of Zimbabwe possess
nutritional, antibacterial and anti-HIV properties?
1.4.1 OTHER RESEARCH QUESTIONS
1.4.1.1 Do different solvents affect the composition and bioactivity of crude extracts and
compounds obtained from different mushrooms?
1.4.1.2 Do wild edible and non-edible mushrooms found in Zimbabwe contain considerable
levels of chemicals and secondary metabolites?
1.5 MAIN RESEARCH HYPOTHESIS
Extracts of different wild edible and non-edible mushrooms of Zimbabwe contain nutritional,
antibacterial and anti-HIV properties.
1.5.1 OTHER RESEARCH HYPOTHESES
1.5.1.1 Different solvents affect the composition and bioactivity of crude extracts and
compounds obtained from different mushrooms.
1.5.1.2 Wild edible and non-edible mushrooms found in Zimbabwe contain considerable
levels of chemical and secondary metabolites.

7
CHAPTER TWO:
LITERATURE REVIEW

8
2.0 LITERATURE REVIEW
2.1 MUSHROOM BIOLOGY AND CLASSIFICATION
Mushrooms are broadly defined as macrofungi with distinctive spore-bearing fruiting bodies
that can be either epigeous (grow above ground) or hypogeous (below ground) and are large
enough to be seen with the naked eye and to be picked by hand (Chang and Miles, 1992;
Stamets, 2000). Most types of mushrooms are commonly found in the form of umbrella-
shaped fruiting body with pileus (cap) under which spores are produced and have a stipe
(stem), for example Lentinus edodes. In addition, some species possess an annulus (ring), for
example Agaricus bisporus (Figure 2.1), or a volva (cup), for example Amanita fulvoalba
(Figure 2.2), or have both the annulus and the volva. The annulus may disappear with age
while the volva is usually buried in the ground (Chang and Miles, 1992). However,
mushrooms can also be in a wide variety of forms where some of them look like pliable cups,
golf balls, or small clubs, with some resembling corals while others are yellow or orange
jellylike globes (Chang, 2008). Usually, the forms that deviate from the usual umbrella-shape
have more specific common names, such as puffballs, stinkhorn and morels.
Figure 2.1: Agaricus bisporus (Source: Gry and Andresson, 2014)
gills
annulus
stipe
pileus

9
Figure 2.2: Amanita fulvoalba (Source: Mighell et al., 2019)
Mushrooms have two phases of growth: the reproductive phase (fruit bodies) and the
vegetative phase (mycelia). Only the fruiting body of the mushroom can be seen whereas the
rest of the mushroom remains in the substrate as mycelium (Wani et al., 2010). The mycelium
comprises a system of branching threads and cordlike strands that branch out through the soil,
compost, wood log or other lignocellulosic material on which the fungus is growing. After a
period of growth, and when the conditions such as moisture and temperature are right, the
established mycelium produces the fruiting structure, which is the mushroom (Chang and
Miles, 1992; Sanchez, 2010; Stamets, 2005).
2.1.1 Classification of Mushrooms
Historically, mushrooms were classified among the so-called lower plants in the division
Thallophyta by Linnaeus. This was largely due to the relatively simple, anatomically
uncomplicated structural attributes (lack of true roots, stems, leaves, flowers and seeds) of the
mushrooms. The presence of a cell wall related them to plants rather than to animals. Modern
studies have established that mushrooms, together with other fungi, have features of their
own, which are sufficiently and significantly distinct to place them in a separate fungal
kingdom. Unlike green plants, mushrooms do not contain chlorophyll and so they cannot
volva

10
manufacture their own nutrients from simple inorganic materials like water and carbon
dioxide. They exploit foods from complex organic materials stored in dead or living tissues of
plants and animals (Ren, 2014; Sharp, 2011).
Mushrooms belong to two subdivisions in the fungal kingdom (Mycota) which belongs to the
Domain Eucarya, namely, Basidiomycetes and Ascomycetes (Table 2.1). The spores for these
two groups are located in a special structure or cell. In Ascomycetes, the sexual spores
(ascospores) are produced inside a club-shaped or cylindrical cell known as the ascus, while
in Basidiomycetes, the sexual spores (basidiospores) are borne externally on a club-shaped
cell known as a basidium (Dickinson, 1979). Basidiomycetes include gilled fungi (cultivated
button mushroom, Agaricus), bracket fungi, boletes and the brightly coloured Cantharellus
mushroom species (Sharp, 2011; 2014).
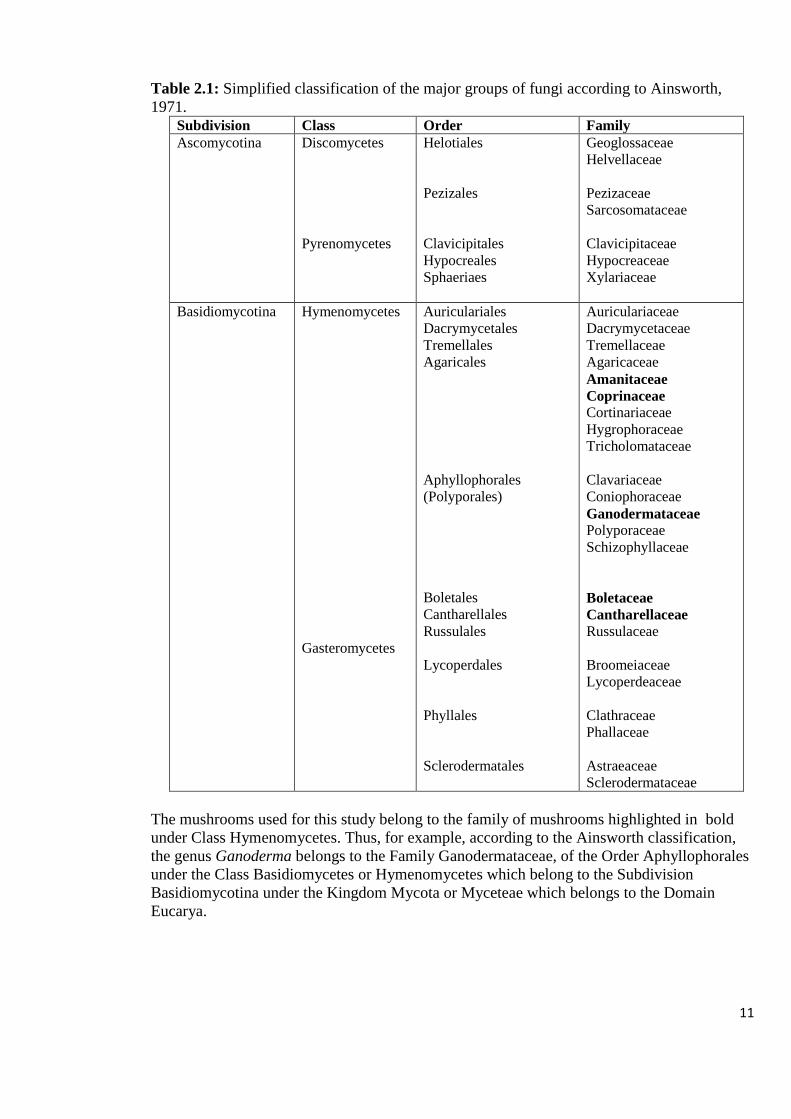
11
Table 2.1: Simplified classification of the major groups of fungi according to Ainsworth,
1971.
Subdivision Class Order Family
Ascomycotina Discomycetes
Pyrenomycetes
Helotiales
Pezizales
Clavicipitales
Hypocreales
Sphaeriaes
Geoglossaceae
Helvellaceae
Pezizaceae
Sarcosomataceae
Clavicipitaceae
Hypocreaceae
Xylariaceae
Basidiomycotina Hymenomycetes
Gasteromycetes
Auriculariales
Dacrymycetales
Tremellales
Agaricales
Aphyllophorales
(Polyporales)
Boletales
Cantharellales
Russulales
Lycoperdales
Phyllales
Sclerodermatales
Auriculariaceae
Dacrymycetaceae
Tremellaceae
Agaricaceae
Amanitaceae
Coprinaceae
Cortinariaceae
Hygrophoraceae
Tricholomataceae
Clavariaceae
Coniophoraceae
Ganodermataceae
Polyporaceae
Schizophyllaceae
Boletaceae
Cantharellaceae
Russulaceae
Broomeiaceae
Lycoperdeaceae
Clathraceae
Phallaceae
Astraeaceae
Sclerodermataceae
The mushrooms used for this study belong to the family of mushrooms highlighted in bold
under Class Hymenomycetes. Thus, for example, according to the Ainsworth classification,
the genus Ganoderma belongs to the Family Ganodermataceae, of the Order Aphyllophorales
under the Class Basidiomycetes or Hymenomycetes which belong to the Subdivision
Basidiomycotina under the Kingdom Mycota or Myceteae which belongs to the Domain
Eucarya.

12
2.1.2 Types of Mushrooms
Mushrooms can be roughly divided into four categories: (i) those which are fleshy and edible,
for example Agaricus bisporus; (ii) mushrooms which are considered to have medicinal
applications, for example, Ganoderma lucidum; (iii) those which are proven to be, or
suspected to be poisonous, for example Amanita phalloides and (iv) those in a miscellaneous
category which includes a large number of mushrooms whose properties remain less well-
defined. However, many types of mushrooms are not only edible, but also possess tonic and
medicinal properties such as Lentinus edodes and Agaricus species (Miles and Chang, 2004).
2.1.2.1 Edible Mushrooms
Edible mushrooms are the fleshy and edible fruit bodies of several species of macrofungi. The
fruiting bodies of edible mushrooms are mainly consumed in their fresh or dried form
(Cheung, 2013). The edibility of mushrooms may be defined by criteria that includes absence
of poisonous effects on humans and palatability. Edible mushrooms include many fungal
species that are either harvested wild or cultivated (Cheung, 2013; Ching et al., 2011). Some
mushrooms that are toxic when raw are said to be edible when cooked. For example, Amanita
muscaria is edible if parboiled to leach out toxins. However, because there is no known test
by which to tell if a mushroom is edible or not, a mushroom should never be eaten unless it
has been accurately identified and the edibility of the species is known (Moukha et al., 2011).
In Southern Africa, roadside sellers only offer "safe species" and most market places are a
reliable means of obtaining known, edible wild mushrooms (Boa, 2004).
Mushrooms are not easy to separate into different categories of beng medicinal and even
being edible because many of the common edible species are also beneficial in the prevention
and treatment of various human diseases. The medicinal properties of these mushrooms is

13
linked to their richness in bioactive compounds, such as phenolic compounds, polyketides,
terpenes, steroids, beta-carotenes, and vitamins A and C (Buruleanu et al., 2018; Rai et al.,
2005; Zhang et al., 2016). The edible class of mushrooms that show potential medicinal and
functional properties includes Cantharellus, Lentinus, Auricularia, Hericium, Grifola and
Pleurotus species (Prasad et al., 2015).
2.1.2.2 Non-edible Mushrooms
Non-edible mushrooms include species that are not palatable while some from this category
are extremely poisonous. The Amanita phalloides group is an example of mushrooms that
cause the most dangerous type of mushroom poisoning. The toxins involved belong to the
phallotoxin and amatoxin complexes (Duffy, 2008). Several Coprinus species, such as C.
micaceus and C. atramentarius, when consumed with an alcoholic drink, produce unpleasant
symptoms which include reddening of the face, increased rate of heartbeat and, in some cases,
vomiting and diarrhoea (Chang and Miles, 1992). Several non-edible mushrooms such as
Ganoderma and Trametes, have been studied and used for their medicinal properties (Cheung,
2013; Ching et al., 2011; Prasad et al., 2015). Ganoderma mushroom lacks gills on its
underside and releases its spores through pores, leading to its morphological classification as
a Polypore. Ganoderma is documented to show strong potency, not only as an immune
response booster but also as an anti-bacterial, anti-parasitic, anti-tumour and anti-
inflammation mushroom (Haoses-Gorases and Goraseb, 2013; Kamra and Bhatt, 2012; Prasad
and Wesely, 2008). Bioactive molecules have also been isolated from non-edible species such
as the ones belonging to the Polyporaceae, Xylariaceae, Thelephoraceae and Paxillaceae
families (Reis et al., 2011).

14
2.2 HUMAN HEALTH BENEFITS OF MUSHROOMS AND THEIR EXTRACTS
2.2.1 Mushrooms as Food and Medicine
Wild edible mushrooms have been collected and consumed by humans worldwide for
thousands of years, with the Chinese growing Auricularia auricular, the wood ear mushroom,
around AD 600 (Boa, 2004; Kues and Liu, 2000; Moon and Lo, 2013; Wang and Xu, 2014).
During the early days of civilization, mushrooms were consumed mainly for their palatability
and unique flavors. However, the current use of mushrooms has now changed because of a lot
of research that has been done on the chemical composition of mushrooms, which revealed
that mushrooms can be used as a diet to combat diseases (Wani et al., 2010). Thus, interest in
consumption of edible mushrooms, both wild and cultivated, has increased remarkably over
the past few decades in many countries, due to the increasing awareness of their nutritional
value (Wani et al., 2010). Mushrooms are high in protein, low in fat and provide low energy
content, factors that make them an excellent food for low-caloric diets. Some mushrooms are
even consumed for medicinal purposes as they contain valuable bioactive components, for
example, Ganoderma lucidum (Buruleanu et al., 2018; Ching et al., 2011; Moon and Lo,
2013; Wang and Xu, 2014; Ziarati and Rabizadeh, 2013).
When in season, the mushrooms provide a notable contribution to diets in Central and
Southern Africa (Cheung, 2008). They are a significant source of nutrition for rural people in
particular, as well as a delicacy for some people (Boa, 2004). Mushrooms as functional foods
are used as nutrient supplements to enhance immunity in the form of tablets. Due to low
starch content and low cholesterol, they suit diabetic and heart patients. The food value of
mushrooms has been reported to compare favourably with meat, egg and milk food sources
(Wani et al., 2010). Recently, various studies have been conducted to increase the application

15
of mushrooms in processed foods. For example, mushrooms can be added to the products
directly as functional ingredients in various baked products (Moon and Lo, 2013).
2.2.1.1 Mushrooms as Source of Proteins
Proteins are the most critical component contributing to the nutritional value of food and is an
important constituent of dry matter of mushrooms. Mushrooms have good nutritional value,
particularly as a source of protein that can enrich human diets, especially in some developing
countries where animal protein may not be available or is expensive (Boda et al., 2012; Wani
et al., 2010). The protein content of fresh mushrooms is 3.7% while the crude protein content
in percent dry weight (% DW) of edible mushrooms range from 15 to 35 % as compared to
7.3 % in rice, 12.7 % in wheat, 38.1 % in soybean, 1.4 % in cabbage, 9.4 % in corn, 12 - 14%
in poultry meat (Soriano, 2010) and 20 – 25% in red meat (Williams, 2007). Thus, in terms of
the amount of crude protein, mushrooms rank well above most other foods, including milk
(2.9 – 3.3 %), which is an animal product. In addition, mushroom protein contains all the
essential amino acids required by humans. The crude protein content varies greatly among the
mushroom species, depending on the size of the pileus and their stage of development (Boa,
2004; Boda et al., 2012; Chang, 2008; Cheung, 2008; Mattila, 2000; Wani et al., 2010).
Mushrooms produce many kinds of proteins with biological activities, including lectins,
antifungal, antiviral and antibacterial proteins, polyphenol oxidase and ribonucleases (Tehrani
et al., 2012; Xu et al., 2014).
Due to their high amount of proteins, mushrooms can be used to bridge the protein
malnutrition gap (Boda et al., 2012). In underdeveloped countries where protein malnutrition
has taken epidemic proportions, Food and Agricultural Organization (FAO) has recommended
the intake of mushroom foods to address the condition (Boda et al., 2012; Wani et al., 2010).

16
The digestibility of mushroom protein has been reported to be as high as 72 to 83 %.
Mushrooms are regarded as an ideal protein source for vegetarian diets, since they contain
some essential amino acids which are found in animal proteins, as well as for old age people
who are unable to chew meat (Wani et al., 2010).
2.2.1.2 Mushrooms as Source of Carbohydrates
Mushrooms contain different amounts of carbohydrates ranging from 51 – 88 % on dry
weight basis (Cheung, 2008). Free sugars amount to about 11 % while mannitol, also referred
to as mushroom sugar, constitutes about 80 % of the total free sugars. Fresh mushrooms are
reported to contain 0.9 % mannitol, 0.28 % reducing sugar, 0.59 % glycogen and 0.91 %
hemicellulose (Waktola and Temesgen, 2018; Wani et al., 2010). Raffinose, sucrose, glucose,
fructose and xylose are reportedly dominant in some mushrooms (Wani et al., 2010).
Several polysaccharides and protein-bound polysaccharides with immunomodulatory and
antitumor activities have been isolated from a variety of mushrooms (Linderquist et al.,
2005). The β-glucans, pleuron from Pleurotus ostreatus and lentinan from Lentinus edodes,
have been reported to increase intestinal mucosal resistance to inflammation and decrease the
occurrence of ulcers in the intestine (Linderquist et al., 2005). Lentinan and another
polysaccharide, schizophyllan from mushroom Schizophyllum, may activate lymphocytes (T
and B cells), macrophages and natural killer cells (Wiater et al., 2011). Polysaccharide-K
(PSK) and polysaccharopeptide (PSP) are polysaccharide-proteins extracted from the
mushroom Corilus versicolor. KrestinTM, which is the trade name for PSK, displays
biological activities that include stimulation of functional maturation of macrophages,
inhibition of the cytophatic effect of HIV infection and an ability to scavenge reactive oxygen
species (Chang and Buswell, 1996).

17
2.2.1.3 Phenolic Compounds and Antioxidant Properties of Mushrooms
Mushrooms are considered to be a natural and good source of antioxidants, chemical
compounds that protect cells from the damage caused by unstable molecules known as
reactive oxygen species. Reactive oxygen species generated during oxidative phosphorylation
by NADPH oxidase, are normal components of healthy cells and also mediators of the first
defensive actions of cells (Kozarski, 2015). However, overproduction of reactive oxygen
species and oxygen-derived free radicals creates oxidative stress, which may induce many
diseases, such as rheumatoid arthritis, atherosclerosis, diabetes, cancer and aging. The
antioxidants are an important defense of the body against free radicals. Fruit bodies and
mycelia of several mushrooms have been reported to show high levels of antioxidant activity
(Buruleanu et al., 2018; Wani et al., 2010).
Different studies have revealed a positive correlation between the total phenolic content in the
mushroom extracts and their antioxidative properties, such as, inhibition of lipid peroxidation
by L. edodes and the radical scavenging and chelating effect on ferrous ions by methanolic
extracts of mushrooms Dictyophora indusiata and Grifola frondosa (Cheung, 2008; Gan et
al., 2013; Ramesh and Pattar, 2010). Phenolic compounds represent a large group of
secondary plant metabolites which now attract great interest due to their benefits for human
health. There are studies that have shown that the antioxidant activity exhibited by phenolic
compounds offers protection against chronic degenerative diseases, cardiovascular diseases,
diabetes mellitus, and neurodegenerative diseases (Buruleanu et al., 2018). Phenolic acids,
such as trans-cinnamic acid, hydroxbenzoic acid, protocatechuic acid and caffeic acid have
been reported in A. bisporus and L. edodes (Wani et al., 2010).

18
The alcoholic extracts of Coprinus comatus were shown to be more effective in scavenging
activity on hydroxyl radicals than hot water extracts (Li et al., 2010). The naturally occurring
antioxidant components included total phenols, tocopherols, flavonoids and polysaccharides.
Tyrosinase from A. bisporus has reportedly shown antioxidant activity. The high potential of
mushrooms as a dietary source of phenolic antioxidants can be used to enhance the low
antioxidant status in the human body (Cheung, 2008), a condition prevailing during certain
infections like HIV. Furthermore, the presence of antioxidant and anti-inflammatory
compounds in mushrooms might be clinically relevant in the management of heart and
circulation health complications (Moon and Lo, 2013).
2.2.2 Mushroom Extracts as Medicine
Scientific studies confirmed recently that bioactive compounds from many edible mushrooms
are involved in lowering the cholesterol levels and protecting against various disorders
including tumors (Ruiz-Rodriguez, 2009; Valverde, 2015). Some medicinal mushrooms
exhibit cardiovascular, anticancer, antiviral, antibacterial, antiparasitic, anti-inflammatory and
antidiabetic properties (Buruleanu et al., 2018; Feng et al., 2016). The potential of wild
mushrooms as sources of antibiotics was reported in 1941 (Sudirman, 2010; Moukha et al.,
2011). In the reported study, extracts of fruiting bodies and mycelia culture from over 200
species were tested. Several compounds that inhibit the growth of a large spectrum of
saprophytic and phytopathogenic fungi and bacteria were isolated from Basidiomycetes. The
study on polypores, such as several species of Ganoderma, Trametes versicolor, T. marianna,
T. cingulata, and Laetiporus sulphureus and gilled mushrooms, such as P. ostreatus, Lentinus
connatus, and Lentinus edodes showed either the antibacterial, anti-Candida, antiviral or
cytotoxic activities (Sudirman, 2010; Moukha et al., 2011).

19
Several mushroom species belonging to the Polyporaceae family are now being regarded as
potential sources of valuable medicines (Prasad and Wesely, 2008). Nortriterpenoids isolated
from Ganoderma showed a wide range of biological activities such as, anti-tumor, anti-
inflammatory, neurotrophic, hepatoprotective and anti-HIV-1 protease activities (Chen et al.,
2017). Aqueous extracts from Pleurotus pulmonarius var sajor caju proved effective in renal
failure while the hot water extracts from several mushrooms exhibited antitumor effects. An
antitumor polysaccharide, named lentinan, was isolated from the shiitake fruiting bodies. It
was reported that mushrooms cure epilepsy, wounds, skin diseases, heart ailments,
rheumatoid arthritis, diarrhea, dysentery, cold, anesthesia, liver and gall bladder diseases
(Afiukwa et al., 2013; Pala and Wani, 2011; Valverde et al., 2015). Most of the mushroom
extracts are now available as therapeutic drugs in China (Ganeshpurkar et al., 2010; Wani et
al., 2010). Puffballs have been used in urinary infections while Pleurotus tuberregium
mushroom has been used for curing headache, high blood pressure, smallpox, asthma, colds
and stomach ailments. Mushroom health supplements can be marketed in the form of
powders, capsules or tablets made of dried fruiting bodies, extracts of mycelium with
substrate, or extracts from liquid fermentation (Wani et al., 2010).
2.2.2.1 Antibacterial Properties of Mushrooms
Both the edible and non-edible wild mushrooms have antibacterial properties. Inhibition of
microbial growth by mushroom extracts is due to the presence of bioactive components in the
mushrooms (Ramesh and Pattar, 2010). Crude organic and aqueous extracts from Ganoderma
have been reported to inhibit in vitro growth of Escherichia coli, Staphylococcus aureus,
Bacillus cereus, Neisseria meningitides, Alcaligenes faecalis and Proteus vulgaris, bacteria
known to cause wound infections, intestinal and urinary-genital tract infections and skin
infections (Kamra and Bhatt, 2012; Prasad and Wesely, 2008; Shikongo et al., 2013).

20
Aqueous, ethanol, methanol and xylene extracts of A. bisporus and Pleurotus pulmonarius var
sajor caju have been reported to exhibit antibacterial activity against E. coli, Enterobacter
aerogenes, Pseudomonas aeruginosa and Klebsiella pneumoniae. Most of the species of
Agrocybe perfecta, Hexagonia hydnoides, Irpex lacteus and Tyromyces duracinus showed
antimicrobial activity against bacteria and yeasts (Chaudhary and Tripathy, 2015). Pleurotus
ostreatus, commonly known as white oyster mushroom, has been reported to possess
compounds that inhibit the growth of E. coli, Bacillus megaterium, S. aureus, K. pneumoniae
isolates and species of Streptococcus and Enterococcus (Sala Uddin et al., 2015).
Some selected mushroom metabolites were reported to have high inhibitory activity against
Gram-positive organisms, including acid fast bacterium Mycobacterium smegnatis
(Gbolagade and Fasidi, 2005). Crude extracts that were obtained from Portuguese wild
mushrooms including Cantharellus cibarius, Hypholoma fasciculare and Ramaria botrytis
showed antibacterial activity against Gram-positive bacteria (Barros et al., 2008). Two
triterpenes, trichomycins A and B, separated from Tricholoma species exhibited antibacterial
activity against S. aureus and S. pneumoniae. A glucosylceramide isolated from Pleurotus
citrinopileatus was found to be active against E. coli and S. aureus (Chomcheon et al., 2013).
Several species in the genera Cantharellus, Lentinus, Russula, Agaricus and Pleurotus have
shown antimicrobial properties against Bacillus, Enterococcus, Streptococcus,
Staphylococcus and Micrococcus species (Alves et al., 2012; Khan and Tania, 2012; Pushpa
and Purushothama, 2010). The European Ganoderma has been reported to inhibit growth of
most bacteria especially methillicin-resistant S. aureas (Linderquist et al., 2005). Different
mushroom species vary in their antimicrobial activity. The intensity of the antimicrobial effect
is dependent upon mushroom species, concentration of the mushroom extract and the
organism being tested against (Ramesh and Pattar, 2010).

21
2.2.2.2 Antiviral Properties of Mushrooms
The HIV type 1 pandemic afflicts approximately 34 million people worldwide (Jadaun et al.,
2016; Tran et al., 2011; WHO, 2018). Side effects such as hypersensitivity, lactic acidosis,
bleeding and anaemia, and uneven access to anti-retroviral drugs remain considerable
therapeutic challenges (Leteane et al., 2012; Mataftsi et al., 2010; Tran et al., 2011;
Yunihastuti et al., 2014). A variety of mushrooms have been reported to possess strong anti-
HIV properties. Proteins, peptides and polysaccharopeptides from mushrooms have been
reported to be capable of inhibiting human immunodeficiency virus type 1 (HIV-1) reverse
transcriptase and protease, the two key enzymes in the life cycle of the HIV (Roupae et al.,
2012). The laccase enzyme, produced by fungi of the genera Ganoderma and Lentinus, was
reported to inhibit the reverse transcriptase (RT) of HIV-1 in in vitro cell-free models (Orozco
et al., 2016). The melanin-glucan complex obtained from Fomes fomentarius mushroom
showed higher anti-HIV activity in comparison with the drug zidovudine in vitro (Friedman,
2016).
Medicinal mushrooms such as Tricholoma giganteum, Hericium erinaceum, Russula
paludosa, Pleurotus eryngii, G. lucidum and L. edodes have shown to contain in their
extracts, ribosome inactivating proteins, lectins, ubiquitin-like proteins and laccases with
strong antiviral effects (Orozco et al., 2016). Lectins from A. bisporus have shown inhibitory
activity against HIV-1 reverse transcriptase. Some triterpenes from G. lucidum are effective
as antiviral agents against HIV-1 (Linderquist et al., 2005). Flammulina velutipes contains a
ribosome inactivating protein that inhibits HIV-1 reverse transcriptase. Some types of
mushrooms such as maitake mushrooms increase CD4+ cell counts, enhancing the activity of
T-helper cells and reducing symptoms and secondary illnesses caused by HIV (Linderquist et
al., 2005; Wani et al., 2010). A methyl gallate compound with anti-HIV activities has been

22
isolated from the mushroom Pholiota adiposa (Wang et al., 2014). In studies conducted in
Zambia, Tanzania and Namibia, the effectiveness of Ganoderma in the therapy of HIV/AIDS
patients and opportunistic infections was reported. The overall health of the patients was
significantly improved compared to the control groups, while significant increases were noted
for body weight, appetite, as well as the CD4+ cell count (Haoses-Gorases and Goraseb,
2013). Lentinan sulphate obtained from Lentinus species has also reportedly inhibited HIV
(Wani et al., 2010).
2.2.2.3 Mushroom Lectins and their Role in Human Health
Lectins are proteins or glycoproteins of non-immune origin which have the ability to bind
specifically and reversibly to complex carbohydrates that are abundant on cell surfaces,
resulting in agglutination of cells or precipitation of glycoconjugates. The detection of lectins
relies on the ability of lectins to agglutinate red blood cells and lectins inhibition by a specific
sugar, which is a major attribute of these proteins (Dhamodharan and Mirunalini, 2011; Sun et
al., 2014; Zhang et al., 2014). Accumulation of lectins in crude extracts of mushrooms can be
detected by hemagglutination assay using human (A, B and O blood groups) and animal
(goose, rabbit, rat and sheep) red blood cells. At least 60 mushroom lectins have been
identified (Santhiya and Jansi, 2013). The high content of lectins in mushrooms has been
detected in diverse species of genera Lactarius, Russula, Boletus, Phallus, Amanita and
Hygrophorus. Mushroom lectins are highly affected by the environment, such as time of
harvest, geographic location and part of mushroom where the lectin was isolated from. The
same mushroom species can have different types of lectins depending on the environment
where the mushrooms were collected (Dhamodharan and Mirunalini, 2011; Zhang et al.,
2014).

23
In recent years, mushroom lectins have drawn the attention of many researchers, mainly due
to the discovery of some of these lectins displaying an array of functions such as
antimicrobial, antitumor, immune-enhancing, anti-insect, antiviral, mitogenic and anti-HIV-1
reverse transcriptase activities (Dhamodharan and Mirunalini, 2011; Eghianruwa et al., 2011;
Koyama et al., 2002; Santhiya and Jansi, 2013; Zhang et al., 2014;). Agaricus bisporus lectin
has exhibited antiproliferative action against human colon cancer and breast cell lines (Patel
and Goyal, 2012). Mushroom lectins that are specific to mannose have antiviral activity. The
Pholiota adiposa lectin exhibited HIV inhibitory activity by targeting the reverse transcriptase
and also antiproliferative activity towards hepatoma Hep G2 cells (Zhang et al., 2009).
Volvariella volvacea lectin possesses antitumor activity to sarcoma S-180 cells while Boletus
lectins have been found to have mitogenic activity towards tumour cells (Bovi et al., 2011;
Sun et al., 2014; Zheng et al., 2007), antimicrobial activity as well as inducing IL-1 and IL-2
(Licastro et al., 1993). Mushroom lectins specific to mucin have been found to have
antimicrobial activity and antiproliferative activity (Lutsik-Kordovsky et al., 2001). These
findings clearly indicate that mushrooms are a valuable source of lectins for drug discovery.
2.3 CHARACTERIZATION OF MUSHROOM EXTRACTS
2.3.1 Methods of Extraction
Characterization of mushroom extracts begins with the pre-extraction and extraction
procedures. The basic pre-extraction steps include washing and drying of mushrooms or
freeze drying and grinding to powder to obtain a homogenous product which increases the
contact of sample surface with the solvent system. Proper action must be taken to ensure that
potential active constituents are not lost, distorted or destroyed during the preparation of the
extracts (Altemimi et al., 2017; Sasidharan et al., 2011). Extraction is a crucial step in the
analysis of mushrooms, because it is necessary to extract the desired chemical constituents

24
from the mushroom samples for further separation and characterization. Solvent extraction
has been the most widely used method for the recovery of active compounds from natural
sources, particularly phytochemical constituents, although other technologies such as
microwave-assisted extraction (MAE), ultrasound-assisted extraction (UAE) and supercritical
fluid extraction are gaining recognition (Zhang et al., 2018). The methods are aimed at
increasing the extract yields at lower cost. In addition, modifications on the methods are
continuously being developed (Altemimi et al., 2017; K l a u s et al., 2009; Shen et al., 2017).
However, the yields and bioactive efficacy of the extracts obtained is strongly affected by the
polarity of the solvent as well as the chemical nature of the isolated compounds (Anwar and
Przybylski, 2012).
2.3.1.1 Choice and Effect of the Extracting Solvent
One of the most important factors affecting the efficient extraction of bioactive compounds
from natural sources is the extraction solvent, since the compounds can range from very polar
to non-polar. The nature of the extraction solvent and varying chemical characteristics and
polarities of chemical compounds can result in different extraction yields and biological
activities of the mushroom (Azwanida, 2015; Doughari, 2012; Gan et al., 2013; Ngo et
al.,2017). Polar compounds are easily extracted using polar solvents while non-polar
compounds will be easily extracted by the non-polar solvents. Thus, extraction of the
mushroom samples can be carried out using different solvents because of the diversity of the
chemical nature of their components and the different solubilities in different solvents
(Altemimi et al., 2017; Anwar and Przybylski, 2012; Nur Syukriah et al., 2014).
Water and other organic solvents such as methanol, ethanol, acetonitrile, acetone, hexane and
diethyl ether have usually been applied in the extraction of bioactive compounds from plants

25
and mushrooms (Table 2.2). Polar solvents such as water, methanol, ethanol, acetone and
their aqueous mixtures, are mostly recommended for the extraction of polyphenols (Anwar
and Przybylski, 2012; Dailey and Vuong, 2015; Tatiya et al., 2011). Water is a common
medium for biochemical reactions and has been shown to be capable of extracting different
classes of active compounds depending on the temperature used (Askin et al., 2007; Shen et
al., 2017). However, although a higher temperature may increase extraction efficiency, it may
also result in degradation of temperature-sensitive antimicrobials. Thus, the chemical profile
of the extracts obtained with different solvents can vary and result in variations of their
antimicrobial properties (Shen et al., 2017).
Table 2.2: Effect of solvents in extracting different components from biological sources
Water Ethanol Methanol Chloroform Ether Acetone
Anthocyanins
Starches
Tannins
Saponins
Terpenoids
Polypeptides
Lectins
Tannins
Polyphenols
Polyacetylenes
Flavonol
Terpenoids
Sterols
Alkaloids
Anthocyanins
Terpenoids
Saponins
Tannins
Xanthoxyllines
Totarol
Quassinoids
Lactones
Flavones
Phenones
Polyphenols
Terpenoids
Flavonoids
Alkaloids
Terpenoids
Coumarins
Fatty acids
Phenols
Flavonols
Although ethanol and methanol have similar polarities, methanol extracted more secondary
metabolites than ethanol. Chloroform and acetone extracted the least compounds (Source:
Tiwari et al., 2011)

26
2.3.2 Methods of Separation of Constituents from Mushroom Extracts
Once extracted from the source, the bioactive components have to be separated (Doughari,
2012). Separation of biologically active constituents from crude extracts for the process of
identification and characterization still remains a big challenge due to the fact that mushroom
and plant extracts usually occur as a combination of various types of bioactive compounds or
phytochemicals with different polarities. Common techniques used in the isolation of these
bioactive compounds include Thin Layer Chromatography (TLC), Paper Chromatography,
Column Chromatography, Flash Chromatography and High Performance Liquid
Chromatography (HPLC). The pure compounds are then used for their identification and
biological activity (Altemimi et al., 2017; Ingle et al., 2017).
2.3.2.1 Thin Layer Chromatography (TLC)
Although there is a wide range of chromatographic methods for isolation of crude extracts,
thin layer chromatography remains a valid and simple analytical procedure for qualitative
detection and quantitative determination of components of mushroom extracts (Masoko,
2007; Ingle et al., 2017). The TLC method is an adsorption chromatography where samples
are separated based on the interaction between a thin layer of adsorbent attached on the plate.
The technique is mostly employed for the separation of low molecular weight compounds
(Ingle et al., 2017). The method does not require expensive instrumentation, is easy to run,
quick, reproducible and samples do not require extensive purification prior to analysis. The
adsorbent layer of silica gel-G has been used with a variety of mobile phase solvent systems
(Pyka, 2014). Preparative or semi-preparative TLC techniques can be used to obtain larger
amounts of the fraction or compound of interest. Preparative TLC plates with a thickness of 1
mm can be prepared using the same stationary and mobile phases, to isolate the bioactive
components (Altemimi et al., 2017). The sample is applied as a wide band and developed

27
under selected chromatographic conditions. After solvent evaporation, bands of desired
bioactive compounds are scraped off together with silica gel, and then they are eluted with
appropriate solvent (Choma and Jesionek, 2015).
2.3.2.2 High Performance Liquid Chromatography (HPLC)
High performance liquid chromatography (HPLC) is a versatile, robust, and widely used
technique for the isolation of natural products. The technique is gaining popularity among
various analytical techniques as the main choice for fingerprinting study for the quality
control of biological sources (Sasidharan et al., 2011). The resolving power of HPLC is
ideally suited to the rapid processing of such multicomponent samples on both an analytical
and preparative scale. Chemical separations can be accomplished using HPLC by utilizing the
fact that certain compounds have different migration rates given a particular column and
mobile phase. The extent or degree of separation is mostly determined by the choice of
stationary phase and mobile phase. Generally the identification and separation of
phytochemicals can be accomplished using isocratic system (using single unchanging mobile
phase system) (Sasidharan et al., 2011). However, gradient elution, in which the proportion of
organic solvent to water is altered with time, is used when more than one sample component
is being studied (Altemimi et al., 2017; Sasidharan et al., 2011)
2.3.3 Methods of Identification of Compounds
The identification of the bioactive compounds isolated from crude extracts can be obtained
using chromatographic techniques such as HPLC and TLC as well as different varieties of
spectroscopic techniques such as UV-visible, Infrared (IR), Nuclear Magnetic Resonance
(NMR), and mass spectroscopy (Altemimi et al., 2017; Buruleanu et al., 2018). Spectroscopy
is based on passing electromagnetic radiation through an organic molecule that absorbs some

28
of the radiation. By measuring the amount of absorption of electromagnetic radiation, a
spectrum, specific to certain bonds in a molecule, can be produced. Depending on these
spectra, the structure of the organic molecule can be identified (Altemimi et al., 2017).
2.3.3.1 Ultra Violet (UV) - Visible Spectroscopy
The isolated compounds can be visualized on a TLC plate by physical (colour or fluorescence
of a compound in Ultra Violet [UV] light) and chemical (coloured reactions of separated
compounds with visualizing reagents) methods (Pyka, 2014). After spraying the TLC plates
with reagents such as vanillin-sulphuric acid, many different compounds could be observed.
Analysis of the compounds is done by comparing the distance traveled relative to the solvent
front called retention factor (Rf value) on the TLC against a reference value of a standard
(Altemimi et al., 2017; Pyka, 2014; Sasidharan et al., 2011). In TLC fingerprinting, the data
that can be recorded using a high performance TLC (HPTLC) scanner includes the recording
of the chromatogram, retention time of individual peaks, the colour of the separated bands and
their absorption spectra. The information generated can be used in the identification of a
compound. The UV-visible spectroscopy can be performed for qualitative analysis and for
identification of certain classes of compounds in both pure and biological mixtures, due to
aromatic molecules that are strong chromophores in the UV range. The technique was
reportedly used to determine the total phenolic extract (280 nm), flavones (320 nm), phenolic
acids (360 nm) and total anthocyanins (520 nm) from a plant (Altemimi et al., 2017).
2.3.3.2 Liquid Chromatography - Mass Spectroscopy (LC-MS)
Recent approaches of applying a combination of chromatography and spectrometry
techniques such as Liquid Chromatography with Photo Diode Array Detection (LC–DAD),
Gas Chromatography - Mass Spectrometry (GC-MS), LC-MS and LC- NMR are increasingly

29
providing the additional spectral information, which is very helpful for the qualitative analysis
and structure determination of novel compounds. In mass spectrometry, organic molecules are
bombarded with either electrons or lasers and thereby converted to charged ions, which are
highly energetic (Altemimi et al., 2017). A mass spectrum gives a plot of the relative
abundance of a fragmented ion against the ratio of mass/charge of these ions. Using mass
spectrometry, the relative molecular mass (molecular weight) of a compound can be
determined with high accuracy and an exact molecular formula can be determined (Altemimi
et al., 2017). Liquid chromatography coupled with mass spectrometry (LC-MS) facilitates
rapid and accurate identification of chemical compounds in extracts, especially when a pure
standard is not available. Recently, LC-MS has been extensively used for the analysis of
phenolic compounds (Altemimi et al., 2017; Sasidharan et al., 2011).
2.3.4 Classes of Secondary Metabolites Found in Mushrooms
2.3.4.1 Alkaloids
Alkaloids are a group of naturally occurring chemical compounds that mostly contain basic
nitrogen atoms. The compounds have basic properties and are alkaline in reaction. Most
alkaloids are readily soluble in alcohol and though they are sparingly soluble in water, their
salts are usually soluble (Kumar, 2014). These nitrogenous compounds function in the
defence of plants against herbivores and pathogens, and are widely exploited as
pharmaceuticals, stimulants, narcotics, and poisons due to their potent biological activities.
Alkaloids are found in certain types of fungi, such as psilocybin which occurs in the fungus of
the genus Psilocybe (De Geyter, 2012; Doughari, 2012).

30
2.3.4.2 Glycosides
A glycoside (Figure 2.3) is a molecule in which a sugar (including polysaccharides) is bound
to a non-carbohydrate part, usually a small organic molecule. The sugar group is referred to as
the glycone and the non-sugar group as the aglycone (De Geyter, 2012). Alcohol, glycerol or
phenol represents aglycones. Glycosides are classified on the basis of the type of sugar
component, chemical nature of the aglycone or pharmacological action. Chemically, the bitter
principles contain the lactone group that may be diterpene lactones or triterpenoids. Extracts
of biological sources that contain cyanogenic glycosides are reportedly used as flavouring
agents in many pharmaceutical preparations (Doughari, 2012).
Alkaloids Fatty acids Glycosides
Example: nicotine Example: butyric acid Example: salicin
Peptides Phenols Terpenes
Example: glycylglycine Example: phenol Example: isoprene
Figure 2.3: Overview of the six main classes of secondary metabolites with representative
examples (De Geyter, 2012)
2.3.4.3 Flavonoids
Flavonoids constitute the largest group of polyphenolic substances, exhibiting extraordinary
diversity and variation. Their chemical structure is built upon a C6-C3-C6 skeleton and the

31
three-carbon bridge is usually cyclised with oxygen. Flavonoids are known to be synthesized
in response to microbial infection. Naturally occurring flavonoids and their glycosides have
been reported to possess various biological properties including anti-inflammatory,
antimicrobial, antiallergic, antioxidant, cytotoxic, antitumour, antidiabetic antiviral and
anticancer activity (Abugria and McElhenney, 2013; Bylka et al., 2004; Cushnie and Lamb,
2005; Djouossi et al., 2015; Kumar and Pandey, 2013). Studies on flavonoids by spectroscopy
have revealed that most flavones and flavonols exhibit two major absorption bands: band I
(320–385 nm) representing the B ring absorption, while band II (250–285 nm) corresponds to
the A ring absorption (Kumar and Pandey, 2013). The antimicrobial activity of total flavonoid
compounds and the antibacterial activities of 2,4-dihydroxybenzoic, protocatechuic, vanillic,
and p-coumaric acids from different wild mushrooms have been reported (Alves et al., 2012;
Chowdhury et al., 2015). Flavonoids occur as aglycones, glycosides, and methylated
derivatives (Kumar and Pandey, 2013). Some flavonoides exhibit anti-HIV-1 effect in vitro
and can interact at different steps in the life cycle of HIV-1, including viral entry, reverse
transcriptase, integrase, and viral protease (Sudsai et al., 2017).
2.3.4.4 Phenolics
Phenolics, phenols or polyphenolics are characterized by at least one aromatic ring (C6) and
one or more hydroxyl groups. Phenols range from simple tannins to more complex flavonoids
that give plants much of their red, blue, yellow, and white pigments (De Geyter, 2012).
Different studies have reported the bioactive potential of phenolic acids, including their
application in the control of human pathogenic infections. Phenolics are classified into (i)
phenolic acids, (ii) flavonoid polyphenolics (flavonones, flavones, xanthones and catechins)
and (iii) non-flavonoid polyphenolics (Doughari, 2012; De Geyter, 2012; Taofiq et al., 2015;
2016).

32
2.3.4.5 Terpenoids
Terpenoids are highly flammable unsaturated hydrocarbons, found in plants, animals and
macrofungi (Duru and Cayan, 2015). They are named based on the number of isoprene units
(from which they are built), for example, monoterpenes (10 carbons), sesquiterpenes (15
carbons), diterpenes (20 carbons), sesterterpenes (25 carbons), triterpenes (30 carbons), and
tetraterpenes (40 carbons). Terpenoids isolated from mushrooms have been associated with
various pharmacological activities like anticancer, antimalarial, antiviral, antibacterial and
anti-inflammatory activities (De Geyter, 2012; Duru and Cayan, 2015; Taofiq et al., 2016).
2.3.4.6 Steroids and Sterols
Steroids and sterols are produced from terpenoid precursors and include cholesterol, vitamin
D and (steroidal) saponins. A steroid is a type of terpenoid compound that contains a specific
arrangement of four rings that are joined to each other. Sterols are special forms of steroids,
with a hydroxyl group at position-3 and a skeleton derived from cholestane (De Geyter,
2012). Steroids are derived either from the sterol lanosterol (animals and fungi) or from
cycloartenol (plants). Steroids have been reported to play several biological functions such as
anti-tumor, antioxidant, immune function as well as prevention of certain diseases (De Geyter,
2012; Taofiq et al., 2016).
2.3.5 Methods of Detecting Antibacterial Activity
A number of antimicrobial test methods are available to determine the susceptibility of
bacteria to antimicrobial agents, but they vary in their sensitivity and efficacy. The method
selected should ideally be simple, rapid, reproducible, flexible, cost-effective and maximize
high sample throughput in order to cope with a varied number of extracts and fractions
(Bailey, 2013; OIE Terrestrial, 2012). The currently available screening methods for the

33
detection of antimicrobial activity of natural products include bioautographic, diffusion and
dilution methods. The bioautographic and diffusion methods are referred to as qualitative
techniques since these methods will only give an idea of the presence or absence of
compounds with antimicrobial activity. On the other hand, dilution methods are regarded as
quantitative assays when they are used to determine the minimal inhibitory concentration
(Valgas et al., 2007).
2.3.5.1 Agar Disc and Well Diffusion Methods
The disk diffusion method is regarded as the gold standard for confirming the susceptibility of
bacteria (Khan et al., 2019). In this method, the isolated colony of the target organism is
suspended into growth media, and standardized through a turbidity test. The standardized
suspension is then inoculated onto the solidified Muller Hinton agar plate. Paper discs
impregnated with specific antibiotics or the test substances are placed on the surface of the
inoculated agar plate (Hudzicki, 2009; Khan et al., 2019). The plates are incubated overnight
at 37 °C as the disc containing the antibiotic or test substance is allowed to diffuse through the
solidified agar, resulting in the formation of an inhibition zone. Thereafter, the zones of
inhibition around each paper disc are measured (Hudzicki, 2009; Khan et al., 2019). In agar
well diffusion method, the antimicrobials present in the test extract are allowed to diffuse out
into the medium and interact in a plate, freshly seeded with the test organisms. The resulting
zones of inhibition will be uniformly circular as there will be a confluent lawn of growth. The
diameter of zone of inhibition can be measured in millimeters (Valgas et al., 2007).

34
Figure 2.4: Agar disc diffusion assay showing zones of inhibition of bacterial growth (Photo
taken by T. Reid)
2.3.6 Methods of Detecting Anti-HIV Activity
In the life cycle of HIV, there are various steps that can be targeted for therapeutic
intervention (Figure 2.5). Preliminary screening of crude extracts and/or isolated compounds
for anti-HIV potential can be carried out with simple cell-based assays such as MTT (3-(4,5-
dimethylthiazol-2-yl)-2,5- diphenyltetrazolium bromide) or XTT (Sodium 3-[1-
(phenylamino)-carbonyl]- 3,4-tetrazoliumbis (4-methoxy-6-nitro) benzene-sulfonic acid
hydrate) assay and HIV p24 expression using human T-cell lines. Target specific studies can
be achieved through evaluating the effect of the test samples on HIV-1 gp120/CD4
interactions and enzymes involved in the replication of HIV-1 using techniques such as
ELISA, flourometric and spectrophotometric methods (Narayan and Rai, 2016; Rege et al.,
2015).
zone of inhibition

35
Figure 2.5: Antiretroviral targets in HIV life cycle (Adapted from Rege et al., 2015). Steps in
the life cycle of HIV, that can be targeted by antiretroviral drugs, include the entry of the HIV
virus into the host genome cell (entry inhibitors), reverse transcription of viral RNA (reverse
transcriptase inhibitors), integration of the double stranded DNA into the host cellular DNA
(integrase inhibitors), protein synthesis and viral assembly (protease inhibitors).
2.3.6.1 MTT or XTT assays
Anti-HIV and cytotoxic effects of natural products or isolated compounds can be evaluated
simultaneously with Human T-cell lines such as Jurkat, CEM-SS, MT4, H9 and PBMCs using
MTT or XTT assays. The results can be expressed as 50 % cytotoxic concentration (CC50), 50
% effective concentration (EC50) and the selectivity index (SI) can be calculated as CC50/EC50
ratio (Chen et al., 2013). Thus, SI reflects both antiviral activity and eventual toxicity of the
test material. A high SI value indicates low toxicity of the test compound and high activity
against the virus. The MTT or XTT assays do not provide detailed information on the
mechanism of action of anti-HIV compounds but allows an estimation of the in vitro
therapeutic index for compounds being considered for further development studies.

36
2.3.6.2 HIV-1 p24 Expression Assay
The effect of crude extracts or isolated compounds on HIV-1 replication can be tested by viral
core protein p24 expression in cell-free supernatants using commercial ELISA kit, according
to the manufacturers’ instructions. The culture supernatant is incubated in microtitre wells
pre-coated with anti-p24 antibody and assayed for p24 antigen by biotin-labelled anti-p24
antibody followed by streptavidin-peroxidase conjugate (Figure 2.6). Horse Radish
Peroxidase (HRP) in the presence of peroxide converts tetramethylbenzidine (TMB) substrate
to a bluish reaction product. Upon acidification by HCl or H2SO4 a bright yellowish color is
generated enabling detection and quantification of p24 present in the supernatant by
absorbance at 450 nm.
Figure 2.6: HIV-1 ELISA p24 antigen assay (Adapted from http://i-
base.info/guides/testing/appendix-3-how-hiv-tests-work). The p24 viral antigen in the test
sample is captured onto the immobilized antibody specific for the p24 gene product of HIV-1,
during incubation. The captured antigen is then reacted with human anti-HIV-1 antibody
conjugated with biotin and then streptavidin-peroxidase is added. When TMB is added, colour
develops as the bound enzyme reacts with the substrate.
2.3.6.3 HIV-1 reverse transcriptase (RT) inhibition assay
The HIV-1 reverse transcriptase (RT) is of tremendous medical interest as it is the target
enzyme for one of the best known anti-HIV drug, AZT, which acts by causing chain
termination of the polymerase reaction (University of Pretoria, https://repository.up.ac.za/). It
is a multifunctional enzyme with 3 enzymatic activities. Firstly, the polymerase domain
p24
Ab
p24 antigen
HRP
conjugate-biotin
TMB

37
transcribes viral RNA to viral DNA. Secondly, in the course of reverse transcription, an
intermediary RNA/ DNA hybrid is formed and RT through its ribonuclease H (RNase H)
domain degrades the RNA component of the hybrid. Thirdly, RT carries out DNA-dependent-
DNA-polymerase (DDDP) activity, producing complementary DNA strands. The completion
of each of these processes is required for the formation of competent viral DNA capable of
integrating into the genome of the infected cell. Hence, RT enzyme is considered as one of the
most important targets for antiretroviral substances (Bessong et al., 2005, Li et al., 2008). The
effect of natural products on RT enzyme can be evaluated by using non-radioactive ELISA
and/or by ribonuclease H activity (Rege et al., 2015).
2.4 Mushrooms Found in Zimbabwe
In Zimbabwe, a wide range of mushroom species have colonized almost every available niche
in the environment, from the hot, dry Lowveld and Zambezi Valley to the cool, wet Eastern
parts of the country. The country’s Miombo woodlands of Msasa (Brachystegia spiciformis)
and Mnondo (Julbernardia globiflora) are particularly rich in mushrooms (Sharp, 2011).
During the rainy season, which is generally between November and March, spores that have
been released by mushrooms from the previous season begin to germinate and produce
hyphae, which develops into a mycelium mat and eventually fruiting bodies. Over 120 wild
edible and non-edible mushroom species have been described in Zimbabwe out of the
estimated several hundreds of mushroom species (Sharp, 2011; 2014). The most common
wild mushrooms include: Termite fungi (Termitomyces), Chanterelles (Cantharellus),
Lactarius, Russula, Boletes (Boletus) or ‘sponge fungi’and Amanita species (Sharp, 2011).
The largest mushroom found in Zimbabwe so far is a bracket fungi (Lenzites elegans)
measuring almost 1.60 m in diameter. Termitomyces titanicus, Zimbabwe’s largest gilled

38
fungus, is according to the Guinness Book of Records, the largest mushroom in the world.
The cap of the mushroom has a diameter of up to one metre. The largest tubed fungus (bolete)
in Zimbabwe is Phlebopus colossus, known as “Dindindi” in Shona and can weigh up to 3.5
kg and measure more than 35 cm in diamenter (Sharp, 2011). Non-edible mushrooms such as
Ganoderma are also found in Zimbabwe, even after the rainy season (Mabveni, 2004;
Ryvarden et al., 1994). Other mushroom species common in Zimbabwe and generally in
Southern Africa include Coprinus micaceus, Podaxis pistillaris, Coriolopsis polyzona,
Leucoagaricus bisporus, Phlebopus sudanicus, Coprinus micaceus, Coprinus plicatilis,
Pycnoporus sanguineus, Laccaria placate, Lactarius deliciosus, Lactarius hepaticus,
Panaeolus papilionaceus, Scleroderma citrinum as well as Laccaria amethystine
(Branch, 2001; Gryzenhout, 2010; Reagile, 2011).
Many wild mushrooms are used as a seasonal domestic food source contributing significantly
to Zimbabwean household food security, with sustainance of livelihoods achieved through
roadside selling of edible mushrooms (Ryvarden et al., 1994). Amanita zambiana (Figure
2.7), commonly known as the “Zambian Slender Caesar”, is an edible basidiomycete fungus
in the genus Amanita which is commonly sold in markets and plays a major role in the
achievement of healthy diets during the rainy season. Amanita zambiana was first described
scientifically by British mycologists Pegler and Piearce, 1980 from Zambia. Piearce had
published an illustration of the species three years earlier, but without a description (Branch,
2001; Ryvarden et al., 1994). The Chanterelles (Figure 2.7) are fairly common mushrooms
and also one of the most highly prized edible ones. The mushrooms are a good source of
potassium and vitamins C and D, so adding these to one’s diet is very useful (Sharp, 2011).
The Boletes, Chanterelles and edible Amanita species, have a ready market for export
(Chiroro, 2004).

39
A B
C D
E F
Figure 2.7: Different types of mushrooms commonly found in Zimbabwe and used in this
study. The mushrooms were collected from Harare under the Miombo woodlands. A: Amanita
zambiana, B: Boletus edulis, C: Cantharellus heinemannianus, D: Cantharellus miomboensis,
E: Cantharellus symoensii, F: Ganoderma lucidum (Photos taken by T. Reid)
In this study different mushroom genera from Zimbabwe were investigated to determine their
bioactive capabilities against bacteria and HIV infection. The results obtained will to aid in
the development of an integrated balanced diet that includes mushroom food with medicinal

40
role. Information for further development of effective anti-infective therapies based on
mushooms will be obtained as well.

41
CHAPTER THREE:
MATERIALS AND
METHODS

42
3.0 MATERIALS AND METHODS
3.1 COLLECTION, IDENTIFICATION AND PREPARATION OF MUSHROOMS
A total of ten different mushrooms, namely; Amanita zambiana, Boletus edulis, Cantharellus
heinemannianus, Cantharellus miomboensis, Cantharellus symoensii, Lactarius kabansus
(edible mushrooms), Amanita species, Coprinus species, Ganoderma lucidum and Trametes
strumosa (non-edible mushrooms), were collected from the woodlands of Zimbabwe. The
mushrooms, with the exception of G. lucidum, were collected during the rainy season,
between January and March. The edible mushrooms selected for this study, are the most
common mushrooms consumed in Zimbabwe, when they are available during the rainy
season. Although Ganoderma lucidum and Trametes strumosa have been widely studied
elsewhere, not much research has been done on the local mushroom species. Identification of
the mushrooms was done on the basis of morphological characteristics, including colour of
the mushroom cap and spore print. Final identification was done by comparing the visual
appearance and the recorded characters of mushroom species with standard mushroom
collection guides by Sharp (2011; 2014) and Ryvarden et al., (1994). The fresh mushrooms
were sliced into thin strips and sun dried for seven days. The dried mushrooms were then
ground to powder using an electrical grinder (Siebtechnik Steel Pulverizer 2, GmbH,
Germany).
3.2 CHARACTERIZATION OF MUSHROOMS
3.2.1 Quantitative Determination of the Total Protein Content
Total protein content of the mushrooms was determined using Kjeldhal method (Rana, 2016).
One and half grams each of the dried mushroom samples were digested in a Kjeldahl
digestion flask by boiling with 25 ml of concentrated sulfuric acid and 10 g of Kjeldahl
catalyst tablet until the mixture was clear. After cooling the flasks to ambient temperature,

43
400 ml of cold water and 100 ml of 40 % sodium hydroxide were added. The contents were
distilled until 200 ml of solution was collected, and mixed with 50 ml of boric acid with
indicator. The solution was titrated with 0.1 mol/L HCl, which had been standardized with
sodium carbonate. The protein content was calculated using Formulae 1 and 2:
% Nitrogen = (A × [HCl] × 0.014× 100) ∕ weight of sample in grams,
Formula 1
where: A is the titration volume of 0.1 mol/L hydrochloric acid minus volume of blank.
% Crude Protein (CP) = % Nitrogen × Protein Factor (6.25)
Formula 2
The experiment was carried out once, in duplicate.
3.2.2 Determination of Carbohydrates
Analysis of carbohydrates in the mushrooms was done using Benedict’s test for reducing
sugars and Molisch’s test for carbohydrates (Carbohydrate Lab Report, 2016). For Benedict’s
test, 2 ml of Benedict’s solution was mixed with 0.5 ml of an aqueous mushroom extract in a
boiling tube. The tube was placed in boiling water bath for 5 minutes and then set aside to
cool to ambient temperature. A red or yellow precipitate indicated a positive reaction. For
Molisch’s test, 2 drops of ethanolic α-naphthol solution were added to 1 ml of an aqueous
mushroom extract. After mixing, 3 ml of concentrated sulphuric acid was carefully added, a
purple colour at the interface indicated a positive result for carbohydrates. Glucose was used
as a positive test for both the tests while sucrose and starch were used as positive tests for the
Molisch’s test. Distilled water was used in place of the mushroom extract for the negative
control.

44
3.2.3 Determination of Lectins
3.2.3.1 Extraction of mushroom crude protein
Half a gram of the powdered form of the mushrooms under study were each mixed with
freshly prepared 0.15 M NaCl in the ratio 1:10 w/v. The mixture was incubated at 4 ℃ for 48
hours in the cold room. After incubation, the mixture was centrifuged at 5 200 rpm using a
Japson centrifuge (Centrifuge Machine Digital, Jambu Pershad & Sons, Ambala, India) for 35
minutes. The supernatant was collected and used in haemagglutination assay.
3.2.3.2 Preparation of the red blood cells
Preparation of red blood cells was carried out according to the methods by Albores et al.,
2014 and Singh et al., 2013, with modifications. A sterile syringe was used to draw two sets
of blood from the sheep and goat (5 ml each per set). One set of blood was mixed with 0.7 ml
of Alsever solution in a 10 ml falcon tube and the other set was poured into falcon tubes
containing EDTA. A volume of 2 ml of blood was drawn out from each tube and poured into
a 50 ml falcon tube. The volume of each tube was made up to 30 ml by adding 0.9 % saline
azide. The blood was centrifuged at 3 500 rpm for 15 minutes using a Japson centrifuge
(Centrifuge Machine Digital, Jambu Pershad & Sons, Ambala, India) kept at 4 ℃ in the cold
room. The supernatant was discarded and 0.9 % saline azide was added up to the 30 ml mark.
The washing process was repeated twice and the resultant pellet was resuspended in 20 ml of
0.9 % saline azide to give a 5 % red blood cell suspension.
3.2.3.3 Haemaggutination assay
Lectins in the crude mushroom extracts were detected by their haemagglutination activity
against sheep and goat erythrocytes (Albores et al., 2012; Singh et al., 2013). The
haemagglutination assay was carried out in 96-well, U-bottom Sarstedt microtiter plates. A

45
serial two-fold dilution of the mushroom extract in microtiter (25 µl) was mixed with 50 µl of
a 5 % suspension of sheep and goat red blood cells in 0.9 % saline azide solution at room
temperature. The first two rows containing 0.9 % saline azide and erythrocytes in a plate were
the negative control. The results were recorded after one hour (60 minutes) when the
erythrocytes in the negative control had fully sedimented. The hemagglutination titer, defined
as the reciprocal of the highest dilution exhibiting agglutination, represented one
hemagglutination unit. Specific activity was determined as the number of hemagglutination
units/mg mushroom. The assays were carried out in duplicate.
3.3 CHARACTERIZATION OF MUSHROOM EXTRACTS
3.3.1 Preparation of Crude Mushroom Extracts
The ten different mushrooms were sliced into thin strips prior to sun drying for seven days
and ground to powder using an electrical grinder (Siebtechnik Steel Pulverizer 2, GmbH,
Germany). Dried mushroom powder (ranging from a tenth of a gram to a gram, depending on
the available quantity of each mushroom type) was extracted with 15 ml of sterile cold water,
absolute methanol, ethanol or acetone at 25 ℃ and 150 rpm for 24 hours using an Orbital floor
incubator shaker (Gallenkamp, UK). Hot water extracts were obtained by boiling the same
amounts of mushrooms in 15 ml of distilled water for 10 minutes and then allowing the
suspension to cool to room temperature. All the suspensions were then filtered using
Whatman no. 1 filter paper, dried under a stream of non-sterile air and reconstituted to 10
mg/ml in sterile water for water extracts or 100 % dimethyl sulfoxide for the rest of the
extracts. The reconstituted crude extracts were filter sterilized using sterile syringe filters
(Filter-Bio, 0.22 µm, Filter Bioscience Membrane Technology Co., Ltd, China). A total of
fifty different extracts were obtained and coded CME 1-50 which were then stored at 4 ℃ for
further use.

46
3.3.2 Determination of Protein Content
Protein content in each mushroom extract was determined using the Folin Ciocalteu (FC) and
Lowry C reagents method with bovine serum albumin as the standard (Owusu-Apenten,
2002). Briefly, 50 µl of each sample was diluted to 250 µl using sterile water and mixed with
250 µl of 0.5 M NaOH. Two and a half millilitres of freshly prepared Lowry C reagent was
added, after which 250 µl of the FC reagent was added. After 30 minutes incubation,
absorbance was measured at 720 nm using a spectrophotometer (SpectronicR 20 GenesysTM,
Spectronic Instruments, USA).
3.3.3 Determination of Total Phenolic Content
Total phenolic content in each mushroom extract was determined using the Folin and
Ciocalteu (FC) reagent method with gallic acid as the standard (Gan et al., 2013). Briefly,
40 µl of each sample was diluted to 200 µl using sterile water or 100 % dimethyl sulfoxide
and mixed with 200 µl of Folin and Ciocalteu’s phenol reagent, diluted 1 : 9 ml in distilled
water. After 6 minutes incubation, 200 µl of 7.5 % sodium carbonate was added to the
mixture and adjusted to 2 ml with distilled water. The reaction was kept in the dark for 60
minutes, after which the absorbance was measured at 725 nm using a spectrophotometer
(SpectronicR 20 GenesysTM, Spectronic Instruments).
3.3.4 Determination of Antibacterial Activity of Crude Extracts
Antibacterial effect of the mushroom extracts against E. coli, S. typhi, S. aureus and S.
pneumoniae was determined using the agar disc diffusion method. E. coli and S. aureus were
obtained from Cimas Medical Aid Society Laboratory, S. pneumoniae from Lancet
Laboratory and S. typhi from the University of Zimbabwe. The bacterial strains were sub-
cultured on Muller Hinton agar to obtain pure cultures. Briefly a suspension containing

47
1 x 106 cfu/ml of bacteria, adjusted using McFarland turbidity standard assay (Hudzicki,
2009), was inoculated into Mueller Hinton Agar (Mast Group Ltd., Merseyside, U.K.). The
discs (6 mm) were dipped in 200 µg/ml of mushroom extract for one hour, dried and placed
on the inoculated agar. Negative controls were prepared with the same solvents used to
dissolve the sample extracts. Kanamycin (50 μg/ml) and vancomycin (30 μg/ml) were used as
positive controls for the tested bacteria. After 2 hours incubation at 4 ℃, inoculated plates
were incubated at 37 ℃ for 18 hours in an incubator (Gallenkamp, UK). At the end of the
incubation period, the zones of inhibition around the discs were measured.
3.3.4.1 Further analysis of mushroom crude extracts showing high antibacterial activity
by spectrophotometry
Based on the results obtained from the antibacterial assays (Section 3.3.4), sixteen extracts,
that showed high potency against the tested bacteria were selected for further characterization.
The absorption spectra, employing wavelengths from 200 nm to 800 nm for the mushroom
extracts, were obtained using the UV-Visible spectrophotometer (UV-1601, Shimadzu,
Japan).
3.3.4.2 Analysis of crude extracts by TLC
The sixteen extracts selected for further study were analysed by TLC, using toluene : ethyl
acetate: methanol (40 : 0.5 : 0.5 v/v) as the mobile phase. Five microliters of the mushroom
extract was spotted onto a TLC plate (Silica Gel 60 F254 plate, Analtech, USA) and left to dry
for five minutes. The TLC plates were dipped in 100 ml of the mobile phase in a developing
tank and left to run for I hour. Visualization of the spots was done by viewing the plates under
UV lamp (P.W. Allen and Co., United Kingdom). Staining with vanillin sulphuric acid

48
solution, a universal staining reagent, and heating the plates at 110 ºC for five minutes was
carried out to determine the compound profile of the selected extracts.
3.3.4.3 Selection of mobile phase for preparative thin layer chromatography (PTLC)
Six different types of mobile phases were used to separate the components of the selected
sixteen mushroom extracts and to analyse the components using TLC. The data obtained was
used to determine the best solvent to use for preparative TLC for each extract. The mobile
phases used were: (i) chloroform : methanol (CM1) 9.5 : 0.5 v/v, (ii) chloroform : methanol
(CM2) 9 : 1 v/v, (iii) ethyl acetate : ethanol : water (EEW) 8 : 2 : 0.2 v/v, (iv) toluene : ethyl
acetate (TE) 9.5 : 0.5 v/v, (v) toluene : ethyl acetate : methanol (TEM) 4 : 0.5 : 0.5 v/v and
(vi) ethyl acetate : methanol : water (EMW) 100 : 13.5 : 10 v/v. Seven microliters of
mushroom extract was spotted onto a TLC plate (Silica Gel 60 F254 plate, Analtech, USA) and
left to dry for five minutes. The TLC plates were dipped in 100 ml of the mobile phase in a
developing tank and left to run for 30 minutes. Visualization of the spots was done by
viewing the plates under UV lamp (P.W. Allen and Co., United Kingdom) and staining with
vapour from iodine crystals.
3.3.4.4 Isolation of compounds from mushroom extracts by PTLC
Two mobile phases, namely, TEM and CM1, were selected for separation of components of
the sixteen mushroom extracts by PTLC. The selection was based on the number of bands and
good resolution from TLC plates. Two hundred and fifty microliters of each mushroom
extract was carefully spread onto a preparative TLC plate (Silica Gel GF F254, 1 000 micron,
Analtech, Inc., USA) and left to dry. The plates were dipped in 100 ml of the mobile phase in
a developing tank and left to run for 50 minutes. Visualization of the spots was done by
viewing the plates under UV lamp (P.W. Allen and Co., United Kingdom) and staining with

49
iodine crystals vapour. The separated bands were each scrapped off the silica plates and
extracted with 15 ml of 100 % ethyl acetate, filtered through Whatman no. 1 filter paper and
dried under a stream of sterile air. The dried extracts were re-suspended in methanol or
dimethyl sulphoxide at a concentration of 10 mg/ml for fraction yields that were 5 mg and
above, and 1 - 8 mg/ml of the same solvents for yields that were below 5 mg. A total of one
hundred and thirty one samples were obtained and coded CP1 to CP131.
3.3.4.5 Screening of isolated fractions for antibacterial activity
The isolated samples were screened for their antimicrobial activity against the same bacteria
previously inhibited by the crude exracts, namely, S. typhi, S. aureus and S. pneumoniae,
using the MTT cell viability assay. Briefly, a suspension containing 1 x 106 cfu/ml of bacteria,
adjusted using McFarland turbidity standard assay (Hudzicki, 2009), was mixed with four
dilutions of each mushroom fraction in wells of a 96-well microtitre plate and made up to 250
µl using Muller Hinton nutrient broth (Sigma-Aldrich, Germany). The final concentrations of
the diluted mushroom fractions were 200, 400, 600 and 800 µg/ml. Negative controls were
prepared with the same solvents used to dissolve the fractions as well as the media alone with
the cells. Ampicillin (50 µg/ml) was used as a positive control for the tested bacteria. The
plates were then incubated at 37 ℃ for 18 hours. Twenty-five microlitres of 5 mg/ml MTT
was then added into each well and the plates were incubated at 37 ℃ for one hour, after which
absorbance was read at 560 nm using an ELISA plate analyser (Micro Plate Read, Global
Diagnostics and Medical Solutions, Belgium). Percentage inhibition of each fraction was
calculated using formula 3:
% inhibition = [(A560 control – A560 sample)/A560 control] x 100
Formula 3

50
3.3.4.6 Identification of antibacterial compounds of mushroom extracts by non-targeted
LC – MS
Seven of the isolated fractions that exhibited high percentage inhibition against the growth of
S. typhi and S. aureus were used for further analysis using an Agilent HPLC 1260 System
(Agilent, USA) coupled to an Agilent 6530 Accurate-Mass Quadrupole Time-of-Flight (Q-
TOF) mass spectrometer (Agilent, USA). The HPLC 1260 System was equipped with a
binary pump, autosampler and thermostated column compartment. The separation of samples
was performed on an Agilent Poroshell 120 C18 column (100 x 2.1 mm, 2.7 µm, Agilent
Technologies) at a column temperature of 40 ℃. The flow rate was 0.25 ml/min and the
mobile phase consisted of 0.1 % (v/v) formic acid in sterile water (A) and 0.1 % (v/v) formic
acid in acetonitrile (B). The following gradient program was used: 0 – 3 min, 80 – 20 % B; 3
– 7 min, 70 – 30 % B; 7 –10 min, 65 – 35 % B; 10 – 13 min, 60 – 40 % B; 13 – 17 min, 40 –
60 % B; 17 – 20 min, 100 % B; 20 – 25 min, 80 –20 % B. The sample injection volume was 5
µl and the mass detection was operated in positive and negative ion modes with parameters
set as follows: drying gas flow rate, 8 L/min; gas temperature, 350 °C; pressure of nebulizer,
35 psig; nozzle voltage, 1 000 V; fragmentor voltage, 175 V; skimmer voltage, 65 V and a
scan range of m/z 100 – 1 000. The mushroom fractions were also analysed in the positive
mode and negative mode without a column.
Mass spectrometry analysis was performed on an Agilent 6530 Q-TOF spectrometer (Agilent,
USA) fitted with a jet stream electrospray ionisation source. Data was analysed using the
Mass Hunter Qualitative Analysis software package (Agilent Technologies). Blanks using
each of the solvent extraction systems were analysed using the Find by Molecular Feature
algorithm in the software package to generate a compound list of molecules with abundance
greater than 500 counts. This was used as an exclusion list to eliminate background

51
contaminant compounds from the analysis of the fractions. Each fraction was then analysed
using the Find the Molecular Feature function to generate a putative list of compounds in the
extracts. Compound lists were then screened against three accurate mass databases; a database
of known plant compounds of therapeutic importance generated specifically for this study
(800 compounds); the Metlin metabolomics database (24,768 compounds); and the Forensic
Toxicology Database by Agilent Technologies (7,509 compounds). Empirical formula for
unidentified compounds was determined using the Find Formula function in the software
package.
3.3.5 Determination of Anti-HIV Activity
3.3.5.1 Determination of anti-HIV-1 reverse transcriptase activity
The dried powder from ten different wild mushrooms (Section 3.1) were extracted with either
sterile cold water, distilled boiling water, methanol, ethanol or acetone as in Section 3.3.1.
The suspensions were filtered, dried under a stream of non-sterile air and reconstituted to 10
mg/ml in sterile distilled water. The reconstituted crude extracts were filter sterilized using
sterile syringe filters (Filter-Bio, 0.22 µm, Filter Bioscience Membrane Technology Co., Ltd).
The effect of the mushroom extracts against HIV-1 reverse transcriptase (RT) activity was
carried out as instructed in the protocol supplied with the assay kit (HIV-1 RT assay kit,
Roche, Germany). Briefly, a 60 µl reaction mixture, consisting of RT solution (20 µl or 4 ng)
diluted in lysis buffer, mushroom test sample (20 µl of 300 µg/ml) and the template/primer
hybrid poly (A) x oligo (dT)15, was prepared in a micro-centrifuge tube. After one hour
incubation at 37 ℃, the reaction mixture was transferred into each well of a 96-well plate of
the assay kit. The plate was incubated at 37 ℃ for one hour. Washing buffer (250 µl) was
added to each well and the wells were washed five times. Antibody (anti-DIG-POD)(200 µl)
was added to each well and the plate was incubated at 37 ℃ for one hour, after which the

52
wells were washed. In the final step, 200 µl of peroxidase substrate, ABTS substrate solution,
was added and the plate was left at room temperature for 20 minutes. The absorbance of the
sample was measured at 405 nm using an ELISA plate analyser (Micro Plate Read, Global
Diagnostics and Medical Solutions, Belgium). Two control samples, one with no RT solution
and the other with sterile distilled water in place of mushroom sample, were included. The
resulting signal intensity was directly proportional to the actual RT activity. The inhibitory
activity of the mushroom extracts was calculated as percent inhibition compared to a control
sample without the mushroom extracts. The assays were carried out in duplicate.
3.3.5.2 Cytotoxicity assay of the mushroom extracts
The general cytotoxicity (CC50) of the fifty different mushroom extracts was determined by a
tetrazolium (MTT) viability assay. The human peripheral blood mononuclear cells (PBMCs)
were adjusted to 1 x 105 cells/ ml in RPMI-1640 medium supplemented with 10 % FBS. A
volume of 100 µl of PBMC suspension was transferred into each well of a 96-well plate and
left to adhere for 24 hours at 37 ℃/5 % CO2. Cells were then exposed for 48 hours to serial
dilutions (0.001 µg/ml to 1 000 µg/ml) of mushroom extracts in RPMI-1640 at 37 ℃/5 %
CO2. After incubation, 25 µl of MTT was added into each well and the plates were left at 37
℃/5 % CO2 for 4 hours. The medium was then carefully removed and the MTT crystals were
dissolved in 100 µl of 100 % dimethyl sulphoxide. Two independent experiments were
carried out in triplicate. The plates were read at 450 nm in an ELISA plate reader (Labsystems
Multiskan, USA). The CC50 values were calculated with Graph pad Prism (Version 5.03,
GraphPad Software Inc.).

53
3.3.5.3 Determination of the in vitro anti-HIV-1 activity using HIV p24 expression assay
Twenty extracts, exhibiting low or no cytotoxic effects to the PBMCs, were selected for the
anti-HIV assay. The PBMCs were obtained from HIV-1 seronegative donors of blood donated
to the Botswana/Harvard Partnership Laboratory, Gaborone. The HIV-1c (MJ4) molecular
clone was used to infect the PBMCs. The MJ4 is an infectious HIV-1 subtype C molecular
clone from a Southern African (Botswana) isolate (Ndung’u et al., 2001). Five millilitre vials
containing PBMCs were removed from the liquid nitrogen tank and immediately placed on
dry ice. Ten millilitres of RPMI growth media, warmed at 37 ℃ was mixed with 10 % fetal
bovine serum (FBS), 2 µl of benzoate and streptomycin. One millilitre of the media was
pipetted out and added dropwise into the thawed PBMC vial. All the contents in the vial were
then drawn out and poured into the tube containing 10 ml of growth media. The tube was
spun at 400 rpm for 10 minutes and the supernatant was carefully decanted. Another 10 of
growth media, without benzoate, was added and the pocess was repeated. The pellet was
resuspended in 5 ml growth media and incubated at 37 ℃ and 5 % CO2 overnight.
The determination of the in vitro anti-HIV activity of mushroom extracts was carried out
according to the method by Leteane et al., 2012, with slight modifications. PBMCs cultured
overnight, at 37 ℃ and 5 % CO2, were stimulated with 1 µg/ml of phytohemagglutinin (PHA)
and 100 U/ml of interleukin 2. The PHA stimulated cells were infected for 3 hours with HIV-
1c (MJ4) molecular clone, at a multiplicity of infection (M.O.I.) of 0.1, in the presence or
absence of the selected mushroom extracts at concentrations of 50, 100 and 150 µg/ml. The
cells were then washed 3 times with PBS and cultured for seven days in 1x AIM-V (Gibco,
USA) growth medium containing PHA. After 7 days of incubation, the HIV-1c (MJ4) p24
antigen from PBMC culture supernatant was quantified by enzyme-linked immunosorbent
assay (ExpressBio Life Sciences, USA) according to the manufacturer’s instructions.

54
Briefly, 100 µl of the supernatant was transferred into the wells of an HIV p24 antigen plate
containing 100 µl of 1x AIM-V (Gibco, USA) growth medium and 20 µl of lysis buffer and
incubated at 37 ℃/5 % CO2 for an hour. The supernatant was removed and the plates were
washed five times with wash buffer. One hundred microlitres of detector antibody was added
into each well and incubated at 37 ℃ for one hour. After aspiration of the detector antibody,
the plate was rinsed again five times. Streptavidin HRP (horse radish peroxidase)(100 µl)
conjugate was added into each well and the plate was incubated at room temperature for 30
minutes. The conjugate was aspirated from the wells and the plates washed five times.
Substrate solution (100 µl) was dispensed into each well and the plate was left at room
temperature protected from direct sunlight for 30 minutes. The reaction was stopped by
adding stop solution and absorbance was read at 450 nm. Azidothymidine (AZT) (20 µg/ml)
and dimethyl sulphoxide were used as the positive and negative controls, respectively. The
percentage inhibition of each extract was calculated using formula 4:
% inhibition = [(A450 control – A450 sample) /A450 control] x 100
Formula 4
3.3.5.4 Assay for HIV-1c induced cytopathic effect
The protective effect of the selected mushroom extracts against HIV-1c induced
cytopathogenicity was measured using the MTT viability assay described in Section 3.3.5.2.
3.3.5.5 Analysis of the anti-HIV mushroom crude extracts by non-targeted LC – MS
Three of the mushroom crude extracts that had the highest percentage inhibition against HIV-
1 RT (89.9 - 92.6 %) and two extracts that had the highest percentage inhibiton against HIV-1
replication from the HIV-1 p24 ELISA assay, were further analysed using an Agilent HPLC
1260 System coupled to an Agilent 6530 Accurate-Mass Quadrupole Time-of-Flight (Q-TOF)

55
mass spectrometer. The same procedure described in section 3.3.4.6 was carried out.
3.4 STATISTICAL ANALYSIS
Experimental values are given as means ± standard deviation (SD). Graph-Pad Prism was
used to analyse the data. Statistical significance was determined by one way variance analysis
(ANOVA) (Version 5.03, GraphPad Software Inc.). Differences at P < 0.05 were considered
to be significant.

56
CHAPTER FOUR:
RESULTS

57
4.0 RESULTS
4.1 COLLECTION, IDENTIFICATION AND PREPARATION OF MUSHROOMS
A total of ten different mushrooms, both edible and non-edible, were collected from the
woodlands of Zimbabwe (Table 4.1). After drying for seven days (Figure 4.1), the
mushrooms were ground to powder using an electrical grinder (Siebtechnik Steel Pulverizer 2,
GmbH, Germany).
Table 4.1: Different types of mushrooms collected from the woodlands of Zimbabwe
Latin name Local Shona Name Edibility
Amanita zambiana Nhedzi Edible
Amanita sp. - Non-edible
Boletus edulis Dindindi Edible
Cantharellus miombooensis Chihombiro Edible
Cantharellus symoensii Jongwe/Firifiti Edible
Cantharellus heinemannianus Tsvuketsvuke Edible
Coprinus sp. - Non-edible
Ganoderma lucidum Howa danda Non-edible
Lactarius kabansus Nzeveyambuya Edible
Trametes strumosa Howa danda Non-edible

58
Figure 4.1: Sliced pieces of Cantharellus miomboensis during the drying process.
4.2 CHARACTERIZATION OF MUSHROOMS
4.2.1 Quantitative Determination of the Total Protein Content
The percentage total protein content of the different mushrooms are shown in Figure 4.2 . The
percentage values ranged from 9.3 % ± 1.0 to 30.8 % ± 1.3. Boletus edulis, Coprinus sp.,
Lactarius kabansus and Amanita zambiana had very high protein content (30.8 % ± 1.3, 30.7
% ± 4.4, 30.1 % ± 0.3 and 28.6 ± 2.4, respectively), followed by Cantharellus symoensii,
Cantharellus heinemannianus and Cantharellus miomboensis with percentage values of 22.9
% ± 0.6, 21.1 % ± 2.4 and 16.8 ± 0.3, respectively. Ganoderma lucidum and T. strumosa had
the lowest protein content values (9.3 % ± 1.0 and 10.0 % ± 1.1, respectively).
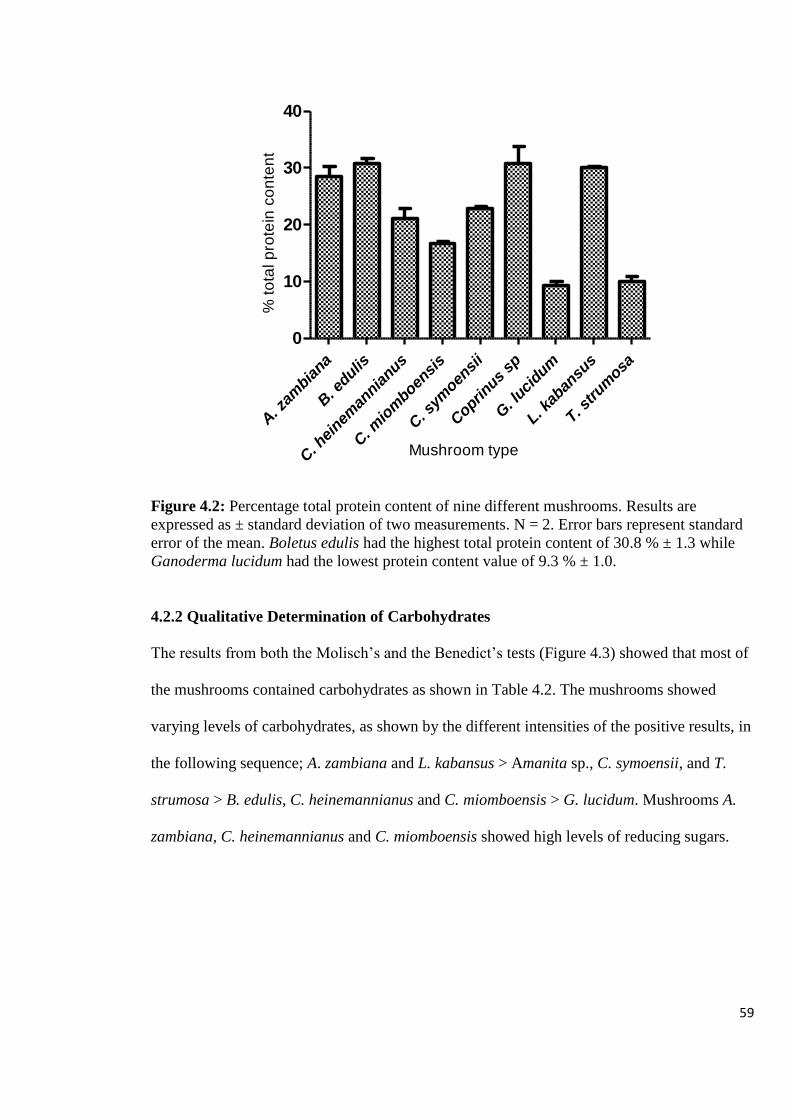
59
A. z
ambia
na
B. e
dulis
C. h
einem
annia
nus
C. m
iom
boensi
s
C. s
ymoen
sii
Copri
nus sp
G. l
ucidum
L. kab
ansu
s
T. strum
osa
0
10
20
30
40
% t
ota
l p
rote
in c
on
ten
t
Mushroom type
Figure 4.2: Percentage total protein content of nine different mushrooms. Results are
expressed as ± standard deviation of two measurements. N = 2. Error bars represent standard
error of the mean. Boletus edulis had the highest total protein content of 30.8 % ± 1.3 while
Ganoderma lucidum had the lowest protein content value of 9.3 % ± 1.0.
4.2.2 Qualitative Determination of Carbohydrates
The results from both the Molisch’s and the Benedict’s tests (Figure 4.3) showed that most of
the mushrooms contained carbohydrates as shown in Table 4.2. The mushrooms showed
varying levels of carbohydrates, as shown by the different intensities of the positive results, in
the following sequence; A. zambiana and L. kabansus > Amanita sp., C. symoensii, and T.
strumosa > B. edulis, C. heinemannianus and C. miomboensis > G. lucidum. Mushrooms A.
zambiana, C. heinemannianus and C. miomboensis showed high levels of reducing sugars.

60
1 2 3 4 1 2 3 4 5 6 7
Figure 4.3: Analysis of carbohydrates showing positive tests for (A) Benedict’s test for
reducing sugars and (B) Molisch’s test for carbohydrates. A1: C. miomboensis, A2: T.
strumosa, A3: A. zambiana, A4: Glucose (positive control), B1: Sucrose (positive control),
B2: C. symoensii, B3: C. heinemannianus, B4: B. edulis, B5: G. lucidum, B6: Coprinus sp.,
B7: Starch (5%) (positive control)
Red
precipitate
for positive
results
Purple
colour
for
positive
results
B A

61
Table 4.2: Qualitative analysis of carbohydrate content in mushrooms using Molisch’s and
Benedict’s tests
Mushroom Molisch’s test
(carbohydrates)
Benedict’s test (reducing
sugars)
A. zambiana ++++ +++
Amanita sp. +++ +
B. edulis ++ ++
C. miomboensis ++ +++
C. heinemannianus ++ +++
C. symoensii +++ ++
Coprinus sp. ++ ++
G. lucidum + ++
L. kabansus ++++ +
T. strumosa +++ ++
Glucose (positive control) +++
Starch (positive control) ++++
(+) Extracts showed positive tests. All the mushrooms tested positive for both the
carbohydrates and reducing sugars. The increasing number of positive signs corresponds to
the increasing intensity of the colour or precipitate.
4.2.3 Determination of Lectins
The results of the haemagglutination assays of the ten different mushrooms in this study using
two types of red blood cells, namely, sheep and goat, are shown in Figure 4.4 and Table 4.3.
The results showed that three of the ten mushrooms, namely Amanita sp., B. edulis and L.
kabansus were able to agglutinate both the sheep and goat red blood cells.

62
Figure 4.4: Haemagglutination assay of three of the ten mushroom species with sheep
erythrocytes showing representative results. The negative control in the first two rows (A-B)
had 0.9% saline azide. Rows C-D, E-F and G-H contain Amanita species, T. strumosa and A.
zambiana, respectively. Amanita species showed haemagglutination activity.
Table 4.3: Haemagglutination assay of the ten mushroom species with sheep and goat
erythrocytes
Mushroom Sheep RBCs Goat RBCs
A. zambiana - -
Amanita sp. + +
B. edulis + +
C. miomboensis - -
C. heinemannianus - -
C. symoensii - -
Coprinus sp. - -
G. lucidum - -
L. kabansus + +
T. strumosa - -
(+) denotes haemagglutination activity, (-) no haemagglutination activity observed. Three
mushrooms, namely, Amanita sp., Boletus edulis and Lactarius kabansus, showed
haemagglutination activity.
Negative control
Amanita sp. showing
haemagglutination
activity
T. strumosa
A. zambiana
D
G
F
H
C
E
A
B

63
The specific haemagglutination activity of the three mushrooms varied, with B. edulis having
the highest specific activity of 617 HAU/ mg mushroom, and L. kabansus having the lowest
activity of 5 HAU/ mg mushroom as shown in Table 4.4. However, although the three
mushroom lectins could agglutinate both sheep and goat erythrocytes, Amanita sp. and L.
kabansus showed a little higher activity with sheep erythrocytes, while B. edulis had much
higher activity with goat erythrocytes.
Table 4.4: Specificity activity of crude extracts of mushrooms showing haemaggutination
activity
Mushroom Specific activity (HAU/ mg mushroom)
Sheep RBCs Goat RBCs
Amanita sp. 39 10
B. edulis 154 617
L. kabansus 77 5
The hemagglutination titer, defined as the reciprocal of the highest dilution exhibiting
agglutination, represented one hemagglutination unit (HAU). Specific activity was determined
as the number of hemagglutination units/mg mushroom. Boletus edulis had the highest
specific activity, showing high lectin content.
4.3 CHARACTERIZATION OF MUSHROOM EXTRACTS
4.3.1 Determination of Protein Content of the Crude Extracts
Results from the protein analysis in Figure 4.5 showed significantly high yields from the cold
water extracts. However, a few boiled extracts gave higher yields, while some of the protein
yields, although lower than the cold water extracts, were not significantly different. Protein
yields from methanol extracts followed the water extracts while ethanol and acetone gave
similar low yields. A zambiana, C symoensii, C. heinemannianus, G. lucidum, L. kabansus
had similar trends: cold water > hot water > methanol > ethanol > acetone, whilst Amanita sp,
B. edulis, Coprinus sp., and T. strumosa had a similar trend, apart from hot water having
greater absolute values than cold water. C. miomboensis had a high acetone protein content
value.
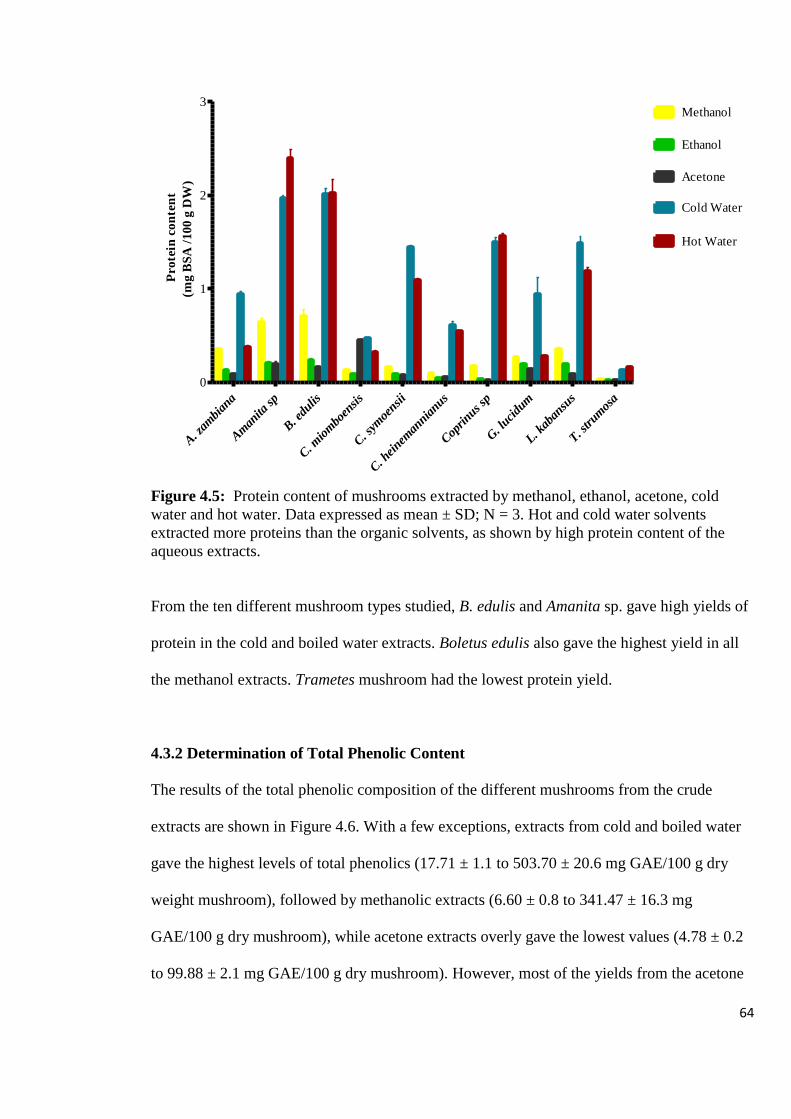
64
A. z
ambi
ana
Am
anita
sp
B. e
dulis
C. m
iom
boen
sis
C. s
ymoe
nsii
C. h
einem
annia
nus
Cop
rinus
sp
G. l
ucidu
m
L. kab
ansu
s
T. strum
osa
0
1
2
3Methanol
Ethanol
Acetone
Cold Water
Hot Water
Pro
tein
co
nte
nt
(mg B
SA
/100 g
DW
)
Figure 4.5: Protein content of mushrooms extracted by methanol, ethanol, acetone, cold
water and hot water. Data expressed as mean ± SD; N = 3. Hot and cold water solvents
extracted more proteins than the organic solvents, as shown by high protein content of the
aqueous extracts.
From the ten different mushroom types studied, B. edulis and Amanita sp. gave high yields of
protein in the cold and boiled water extracts. Boletus edulis also gave the highest yield in all
the methanol extracts. Trametes mushroom had the lowest protein yield.
4.3.2 Determination of Total Phenolic Content
The results of the total phenolic composition of the different mushrooms from the crude
extracts are shown in Figure 4.6. With a few exceptions, extracts from cold and boiled water
gave the highest levels of total phenolics (17.71 ± 1.1 to 503.70 ± 20.6 mg GAE/100 g dry
weight mushroom), followed by methanolic extracts (6.60 ± 0.8 to 341.47 ± 16.3 mg
GAE/100 g dry mushroom), while acetone extracts overly gave the lowest values (4.78 ± 0.2
to 99.88 ± 2.1 mg GAE/100 g dry mushroom). However, most of the yields from the acetone

65
and ethanol extracts were not significantly different. Cantharellus symoensii, C.
heinemannianus, Coprinus sp., and L. kabansus had similar trends: cold water > hot water >
methanol > ethanol > acetone, whilst Amanita sp. had a similar trend, apart from hot water
having a greater absolute value than cold water. Amanita zambiana and G. lucidum had
similar above trends as well, with the exception of methanol having a greater absolute value
than hot water. Boletus edulis had high hot water and methanol phenol content whilst C.
miomboensis had a high acetone phenol content value. Statistical analysis by two way
ANOVA showed that there was significant difference in the effect of solvents in extracting
total phenols (4 df, F = 7.815, P-value = 0.000122) and that the total phenolic composition
was also dependant on the mushroom type (9 df, F = 4.984, P value = 0.000224). From the ten
different mushroom types studied, Boletus was observed to have the highest total phenolic
compounds followed by Amanita sp., while T. strumosa had the lowest phenolic content.
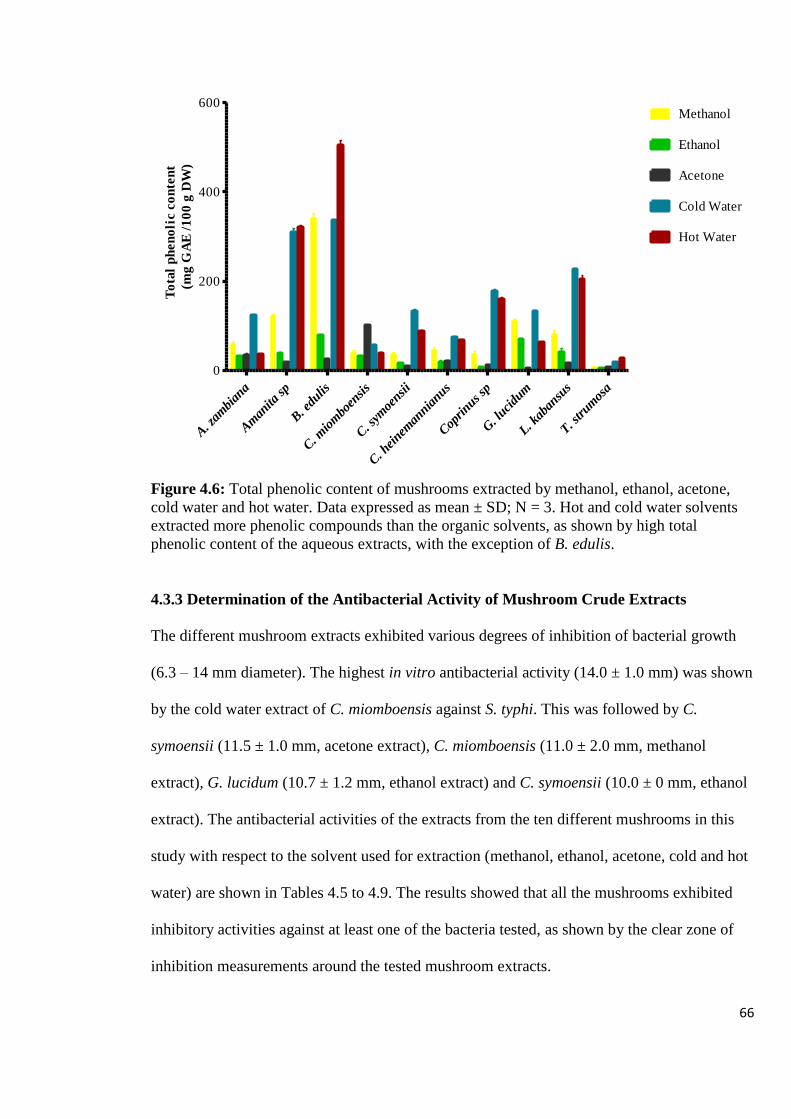
66
A. z
ambi
ana
Am
anita
sp
B. e
dulis
C. m
iom
boen
sis
C. s
ymoe
nsii
C. h
einem
annia
nus
Cop
rinus
sp
G. l
ucidu
m
L. kab
ansu
s
T. strum
osa
0
200
400
600Methanol
Ethanol
Acetone
Cold Water
Hot Water
To
tal
ph
en
oli
c c
on
ten
t
(
mg
GA
E /
10
0 g
DW
)
Figure 4.6: Total phenolic content of mushrooms extracted by methanol, ethanol, acetone,
cold water and hot water. Data expressed as mean ± SD; N = 3. Hot and cold water solvents
extracted more phenolic compounds than the organic solvents, as shown by high total
phenolic content of the aqueous extracts, with the exception of B. edulis.
4.3.3 Determination of the Antibacterial Activity of Mushroom Crude Extracts
The different mushroom extracts exhibited various degrees of inhibition of bacterial growth
(6.3 – 14 mm diameter). The highest in vitro antibacterial activity (14.0 ± 1.0 mm) was shown
by the cold water extract of C. miomboensis against S. typhi. This was followed by C.
symoensii (11.5 ± 1.0 mm, acetone extract), C. miomboensis (11.0 ± 2.0 mm, methanol
extract), G. lucidum (10.7 ± 1.2 mm, ethanol extract) and C. symoensii (10.0 ± 0 mm, ethanol
extract). The antibacterial activities of the extracts from the ten different mushrooms in this
study with respect to the solvent used for extraction (methanol, ethanol, acetone, cold and hot
water) are shown in Tables 4.5 to 4.9. The results showed that all the mushrooms exhibited
inhibitory activities against at least one of the bacteria tested, as shown by the clear zone of
inhibition measurements around the tested mushroom extracts.
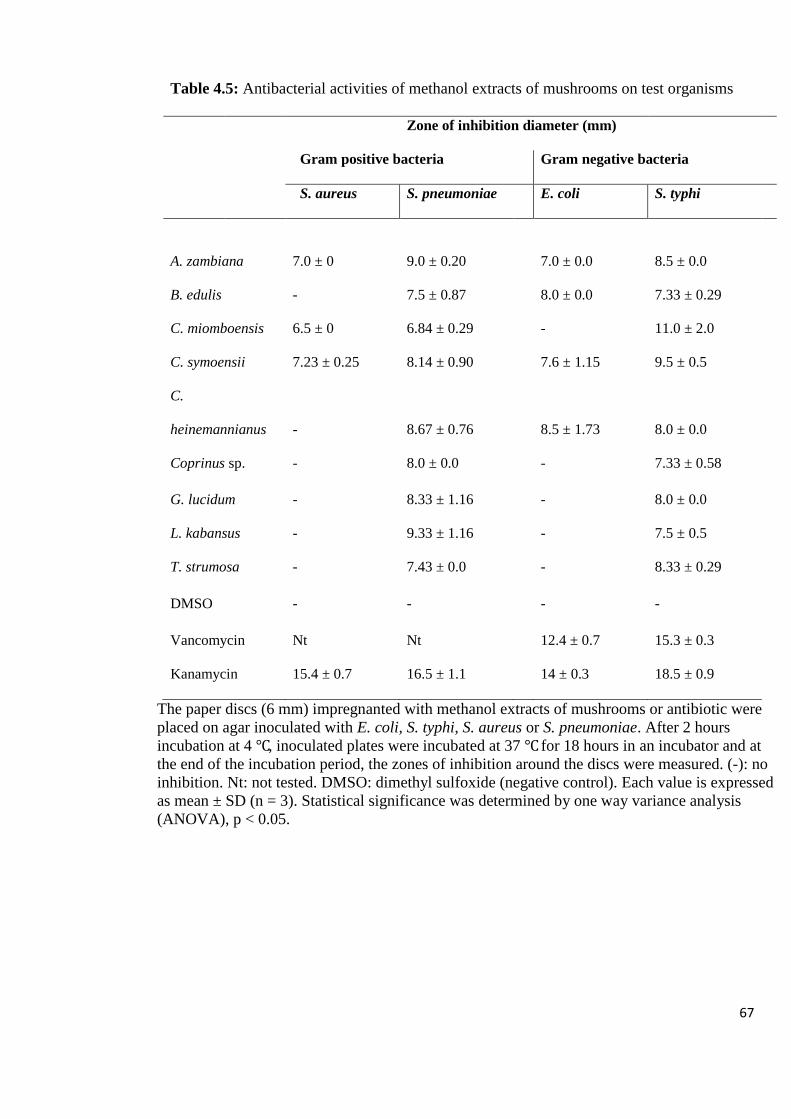
67
Table 4.5: Antibacterial activities of methanol extracts of mushrooms on test organisms
Zone of inhibition diameter (mm)
Gram positive bacteria Gram negative bacteria
S. aureus S. pneumoniae E. coli S. typhi
A. zambiana 7.0 ± 0 9.0 ± 0.20
7.0 ± 0.0 8.5 ± 0.0
B. edulis - 7.5 ± 0.87
8.0 ± 0.0 7.33 ± 0.29
C. miomboensis 6.5 ± 0 6.84 ± 0.29 - 11.0 ± 2.0
C. symoensii 7.23 ± 0.25 8.14 ± 0.90
7.6 ± 1.15 9.5 ± 0.5
C.
heinemannianus - 8.67 ± 0.76
8.5 ± 1.73 8.0 ± 0.0
Coprinus sp. - 8.0 ± 0.0
- 7.33 ± 0.58
G. lucidum - 8.33 ± 1.16
- 8.0 ± 0.0
L. kabansus - 9.33 ± 1.16
- 7.5 ± 0.5
T. strumosa - 7.43 ± 0.0
- 8.33 ± 0.29
DMSO - - - -
Vancomycin Nt Nt 12.4 ± 0.7 15.3 ± 0.3
Kanamycin 15.4 ± 0.7 16.5 ± 1.1 14 ± 0.3 18.5 ± 0.9
The paper discs (6 mm) impregnanted with methanol extracts of mushrooms or antibiotic were
placed on agar inoculated with E. coli, S. typhi, S. aureus or S. pneumoniae. After 2 hours
incubation at 4 ℃, inoculated plates were incubated at 37 ℃ for 18 hours in an incubator and at
the end of the incubation period, the zones of inhibition around the discs were measured. (-): no
inhibition. Nt: not tested. DMSO: dimethyl sulfoxide (negative control). Each value is expressed
as mean ± SD (n = 3). Statistical significance was determined by one way variance analysis
(ANOVA), p < 0.05.
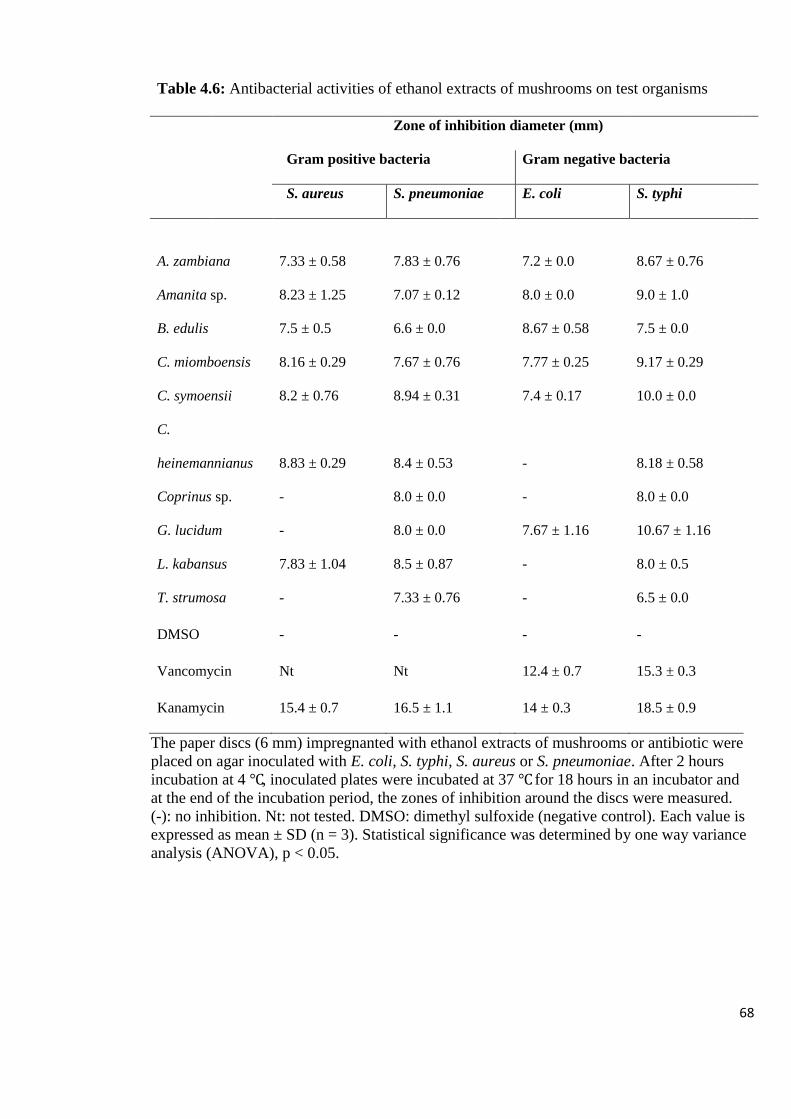
68
Table 4.6: Antibacterial activities of ethanol extracts of mushrooms on test organisms
Zone of inhibition diameter (mm)
Gram positive bacteria Gram negative bacteria
S. aureus S. pneumoniae E. coli S. typhi
A. zambiana 7.33 ± 0.58 7.83 ± 0.76
7.2 ± 0.0 8.67 ± 0.76
Amanita sp. 8.23 ± 1.25 7.07 ± 0.12
8.0 ± 0.0 9.0 ± 1.0
B. edulis 7.5 ± 0.5 6.6 ± 0.0
8.67 ± 0.58 7.5 ± 0.0
C. miomboensis 8.16 ± 0.29 7.67 ± 0.76
7.77 ± 0.25 9.17 ± 0.29
C. symoensii 8.2 ± 0.76 8.94 ± 0.31
7.4 ± 0.17 10.0 ± 0.0
C.
heinemannianus 8.83 ± 0.29 8.4 ± 0.53
- 8.18 ± 0.58
Coprinus sp. - 8.0 ± 0.0
- 8.0 ± 0.0
G. lucidum - 8.0 ± 0.0
7.67 ± 1.16 10.67 ± 1.16
L. kabansus 7.83 ± 1.04 8.5 ± 0.87
- 8.0 ± 0.5
T. strumosa - 7.33 ± 0.76
- 6.5 ± 0.0
DMSO - - - -
Vancomycin Nt Nt 12.4 ± 0.7 15.3 ± 0.3
Kanamycin 15.4 ± 0.7 16.5 ± 1.1 14 ± 0.3 18.5 ± 0.9
The paper discs (6 mm) impregnanted with ethanol extracts of mushrooms or antibiotic were
placed on agar inoculated with E. coli, S. typhi, S. aureus or S. pneumoniae. After 2 hours
incubation at 4 ℃, inoculated plates were incubated at 37 ℃ for 18 hours in an incubator and
at the end of the incubation period, the zones of inhibition around the discs were measured.
(-): no inhibition. Nt: not tested. DMSO: dimethyl sulfoxide (negative control). Each value is
expressed as mean ± SD (n = 3). Statistical significance was determined by one way variance
analysis (ANOVA), p < 0.05.
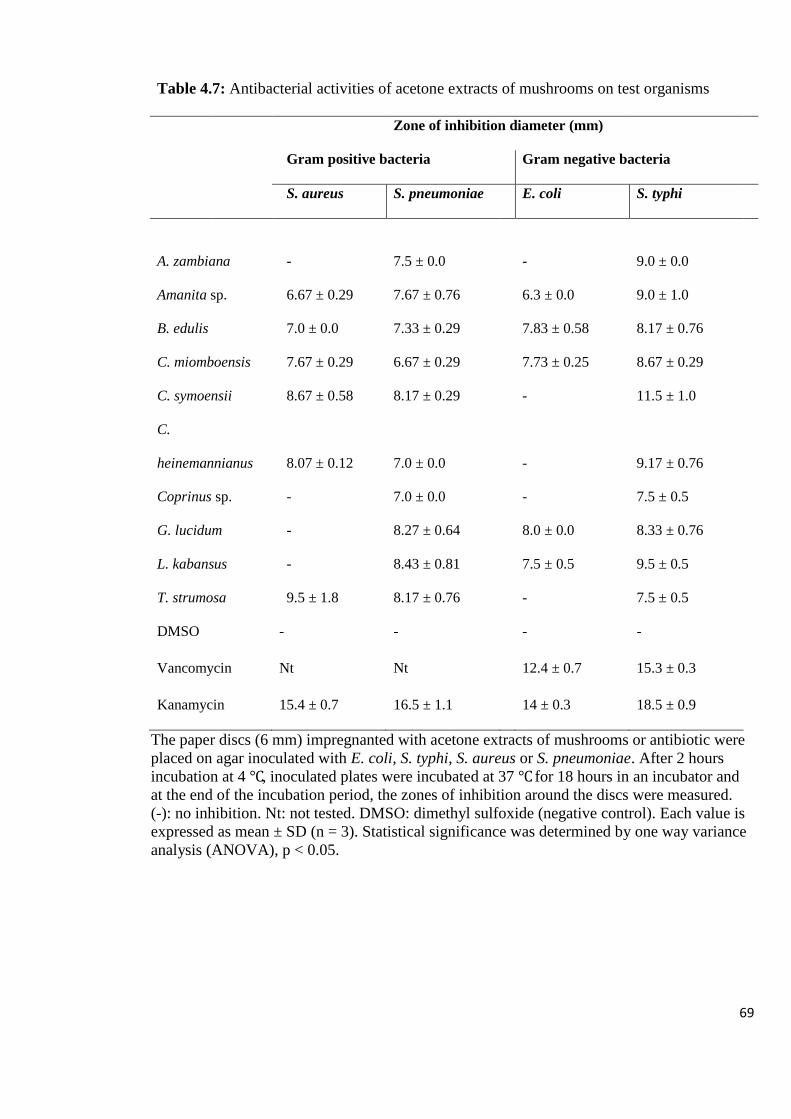
69
Table 4.7: Antibacterial activities of acetone extracts of mushrooms on test organisms
Zone of inhibition diameter (mm)
Gram positive bacteria Gram negative bacteria
S. aureus S. pneumoniae E. coli S. typhi
A. zambiana - 7.5 ± 0.0
- 9.0 ± 0.0
Amanita sp. 6.67 ± 0.29 7.67 ± 0.76
6.3 ± 0.0 9.0 ± 1.0
B. edulis 7.0 ± 0.0 7.33 ± 0.29
7.83 ± 0.58 8.17 ± 0.76
C. miomboensis 7.67 ± 0.29 6.67 ± 0.29
7.73 ± 0.25 8.67 ± 0.29
C. symoensii 8.67 ± 0.58 8.17 ± 0.29
- 11.5 ± 1.0
C.
heinemannianus 8.07 ± 0.12 7.0 ± 0.0
- 9.17 ± 0.76
Coprinus sp. - 7.0 ± 0.0
- 7.5 ± 0.5
G. lucidum - 8.27 ± 0.64
8.0 ± 0.0 8.33 ± 0.76
L. kabansus - 8.43 ± 0.81
7.5 ± 0.5 9.5 ± 0.5
T. strumosa 9.5 ± 1.8 8.17 ± 0.76
- 7.5 ± 0.5
DMSO - - - -
Vancomycin Nt Nt 12.4 ± 0.7 15.3 ± 0.3
Kanamycin 15.4 ± 0.7 16.5 ± 1.1 14 ± 0.3 18.5 ± 0.9
The paper discs (6 mm) impregnanted with acetone extracts of mushrooms or antibiotic were
placed on agar inoculated with E. coli, S. typhi, S. aureus or S. pneumoniae. After 2 hours
incubation at 4 ℃, inoculated plates were incubated at 37 ℃ for 18 hours in an incubator and
at the end of the incubation period, the zones of inhibition around the discs were measured.
(-): no inhibition. Nt: not tested. DMSO: dimethyl sulfoxide (negative control). Each value is
expressed as mean ± SD (n = 3). Statistical significance was determined by one way variance
analysis (ANOVA), p < 0.05.
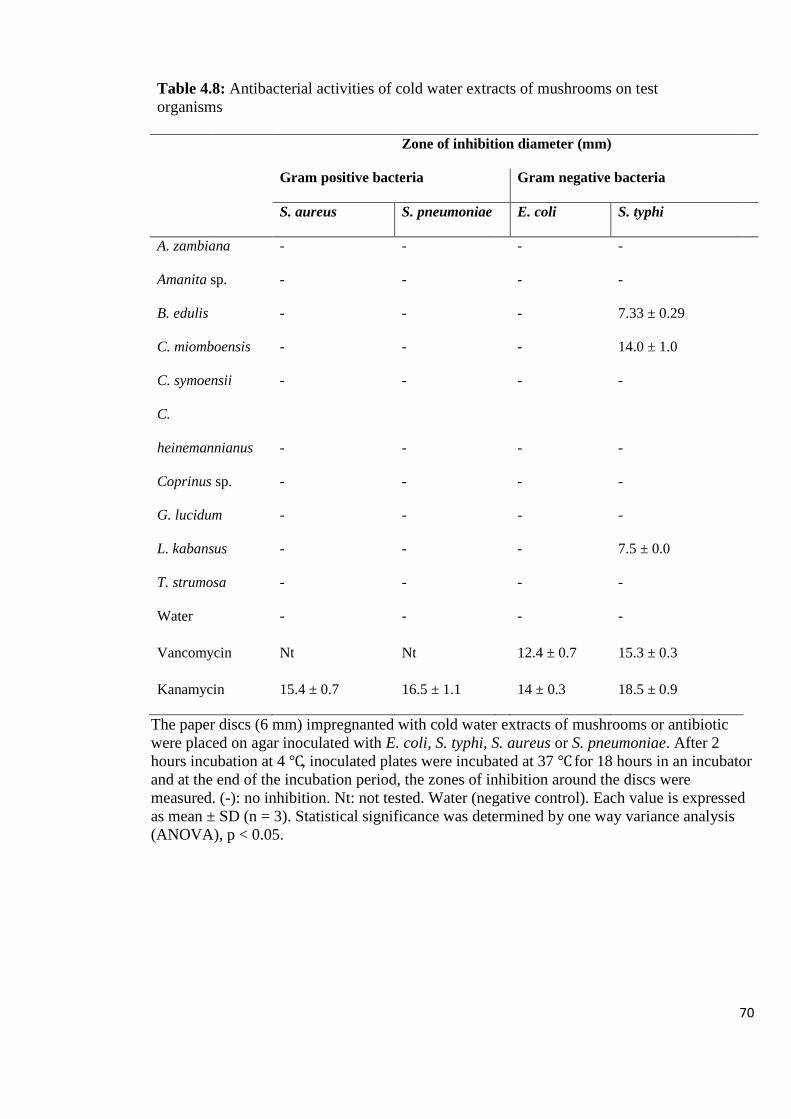
70
Table 4.8: Antibacterial activities of cold water extracts of mushrooms on test
organisms
Zone of inhibition diameter (mm)
Gram positive bacteria Gram negative bacteria
S. aureus S. pneumoniae E. coli S. typhi
A. zambiana - -
- -
Amanita sp. - -
- -
B. edulis - -
- 7.33 ± 0.29
C. miomboensis - -
- 14.0 ± 1.0
C. symoensii - -
- -
C.
heinemannianus - -
- -
Coprinus sp. - -
- -
G. lucidum - -
- -
L. kabansus - -
- 7.5 ± 0.0
T. strumosa - -
- -
Water - - - -
Vancomycin Nt Nt 12.4 ± 0.7 15.3 ± 0.3
Kanamycin 15.4 ± 0.7 16.5 ± 1.1 14 ± 0.3 18.5 ± 0.9
The paper discs (6 mm) impregnanted with cold water extracts of mushrooms or antibiotic
were placed on agar inoculated with E. coli, S. typhi, S. aureus or S. pneumoniae. After 2
hours incubation at 4 ℃, inoculated plates were incubated at 37 ℃ for 18 hours in an incubator
and at the end of the incubation period, the zones of inhibition around the discs were
measured. (-): no inhibition. Nt: not tested. Water (negative control). Each value is expressed
as mean ± SD (n = 3). Statistical significance was determined by one way variance analysis
(ANOVA), p < 0.05.
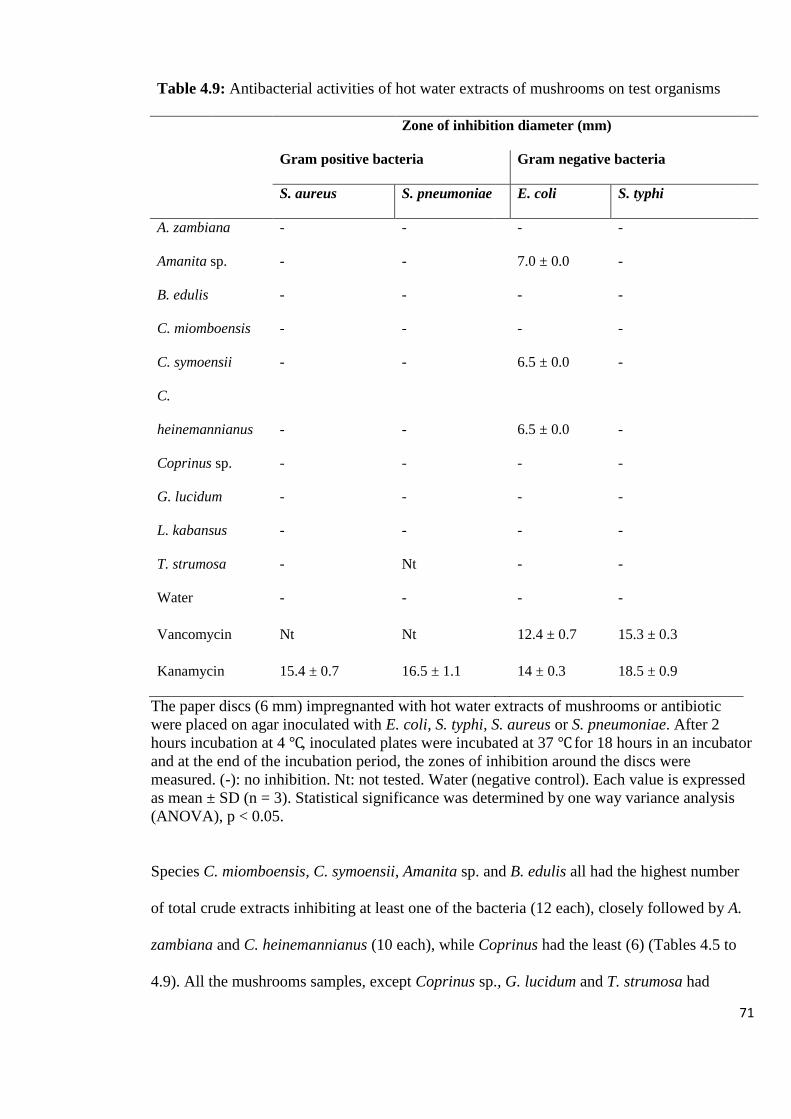
71
Table 4.9: Antibacterial activities of hot water extracts of mushrooms on test organisms
Zone of inhibition diameter (mm)
Gram positive bacteria Gram negative bacteria
S. aureus S. pneumoniae E. coli S. typhi
A. zambiana - -
- -
Amanita sp. - -
7.0 ± 0.0 -
B. edulis - -
- -
C. miomboensis - -
- -
C. symoensii - -
6.5 ± 0.0 -
C.
heinemannianus - -
6.5 ± 0.0 -
Coprinus sp. - -
- -
G. lucidum - -
- -
L. kabansus - -
- -
T. strumosa - Nt
- -
Water - - - -
Vancomycin Nt Nt 12.4 ± 0.7 15.3 ± 0.3
Kanamycin 15.4 ± 0.7 16.5 ± 1.1 14 ± 0.3 18.5 ± 0.9
The paper discs (6 mm) impregnanted with hot water extracts of mushrooms or antibiotic
were placed on agar inoculated with E. coli, S. typhi, S. aureus or S. pneumoniae. After 2
hours incubation at 4 ℃, inoculated plates were incubated at 37 ℃ for 18 hours in an incubator
and at the end of the incubation period, the zones of inhibition around the discs were
measured. (-): no inhibition. Nt: not tested. Water (negative control). Each value is expressed
as mean ± SD (n = 3). Statistical significance was determined by one way variance analysis
(ANOVA), p < 0.05.
Species C. miomboensis, C. symoensii, Amanita sp. and B. edulis all had the highest number
of total crude extracts inhibiting at least one of the bacteria (12 each), closely followed by A.
zambiana and C. heinemannianus (10 each), while Coprinus had the least (6) (Tables 4.5 to
4.9). All the mushrooms samples, except Coprinus sp., G. lucidum and T. strumosa had

72
inhibitory effect on all the four bacteria tested. Species C. heinemannianus and C. symoensii
had the highest effect on inhibition of bacteria as indicated by having the most extracts which
had high inhibitory properties ranging from 8-14 mm [15.4 % (8) each] followed by G.
lucidum [13.5 % (7)], while B. edulis, Coprinus species and T. strumosa had the least [5.8 %
(3 each)].
The organic solvent, ethanol, was the most effective in extracting antibacterial compounds as
shown by its highest number of bacterial growth inhibiting crude extracts (33), followed by
acetone (31) and methanol (28). In addition, ethanol extracts showed the strongest
antibacterial activity (8-14 mm) among the five solvents used, followed by methanol and
acetone. Water extracts exhibited the lowest number of antibacterial activity, despite having
the extract with the highest inhibitory effect. Ethanol, acetone and methanol extracts were all
effective against all the four bacteria, indicating the broad spectrum of antibacterial activity of
the extracts. Gram negative bacteria were more susceptible to the extracts than gram positive
bacteria (52 and 46 extracts, respectively). Among the four bacteria tested, S. typhi was the
most susceptible as indicated by its highest number of inhibitions as well as the highest
number of the most potent extracts in the 8-14 mm diameter range.
Based on the results obtained from the antibacterial assays (Section 4.3.3), the sixteen extracts
that showed high inhibitory levels against the tested bacteria were selected for further
characterization (Table 4.10).

73
Table 4.10: Extracts selected for further study, exhibiting high antibacterial activity (9 – 14
mm zones of inhibition) at a concentration of 200 µg/ml
Mushroom type Solvent used for
extraction
Zone of inhibition
(mm)
Bacteria inhibited
C. miomboensis Cold water 14.0 ± 1.0 S. typhi
C. symoensii Acetone 11.5 ± 1.0 S. typhi
C. miomboensis Methanol 11.0 ± 2.0 S. typhi
G. lucidum Ethanol 10.7 ± 1.2 S. typhi
C. symoensii Ethanol 10.0 ± 0 S. typhi
C. symoensii Methanol 9.5 ± 0.5 S. typhi
L. kabansus Acetone 9.5 ± 0.5 S. typhi
T. strumosa Acetone 9.5 ± 1.8 S. aureus
Amanita sp. Methanol 9.3 ± 1.2 S. pneumoniae
L. kabansus Methanol 9.3 ± 1.2 S. pneumoniae
C. heinemannianus Acetone 9.2 ± 0.8 S. typhi
C. miomboensis Ethanol 9.2 ± 0.3 S. typhi
A. zambiana Methanol 9.0 ± 0.2 S. pneumoniae
A. zambiana Acetone 9.0 ± 0 S. typhi
Amanita sp. Ethanol 9.0 ± 1.0 S. typhi
Amanita sp. Acetone 9.0 ± 1.0 S. typhi
Total number of extracts 16
Acetone had the highest number of extracts that showed high antibacterial activity while hot
water had none. S. typhi was the most susceptible bacteria as it was inhibited by most of the
extracts selected (12 out of 16 extracts).

74
4.4 CHARACTERIZATION OF MUSHROOM EXTRACTS SHOWING
ANTIBACTERIAL ACTIVITY
4.4.1 Analysis of the Extracts using Absorption Spectroscopy and TLC
The UV spectra of mushroom extracts that exhibited antibacterial activity showed absorbance
peaks ranging from 227 – 308 nm; 734 – 745 nm and 586 nm as shown in Table 4.11.
Table 4.11: Absorption spectrum peaks obtained from the selected crude extracts of
mushrooms that showed high antibacterial activity
Mushroom sample Absorbance Peak (nm)
Methanol Ethanol Acetone Cold water
Amanita sp. 266, 638 236, 259, 586 227, 259
G. lucidum
234, 308
T. strumosa
228, 740
C. heinemmanianus
272
C. symoensii 308 279 235, 745
C. miomboensis 276 295
228, 734
L. kabansus 256
231, 738
A. zambiana 257, 735
264
The absorbance peak values obtained varied from one extract to another showing that the
secondary metabolite profile of each mushroom extract was different.

75
The absorbance profile obtained from the different mushrooms using the same extraction
solvent type varied with each mushroom species (Figures 4.7 – 4.9).
Figure 4.7: Representative UV spectra obtained from acetone (Ac) and water (H2O) extracts
of mushrooms. Am: Amanita sp., Tr: T. strumosa, Az: A. zambiana, Lk: L. kabansus, Cs: C.
symoensii, Ch: C. heinemannianus and Cm: C. miomboensis
Figure 4.8: Representative UV spectra obtained from ethanol (Eth) extracts of mushrooms.
Am: Amanita sp., Cs: C. symoensii, Cm: C. miomboensis and G: G. lucidum
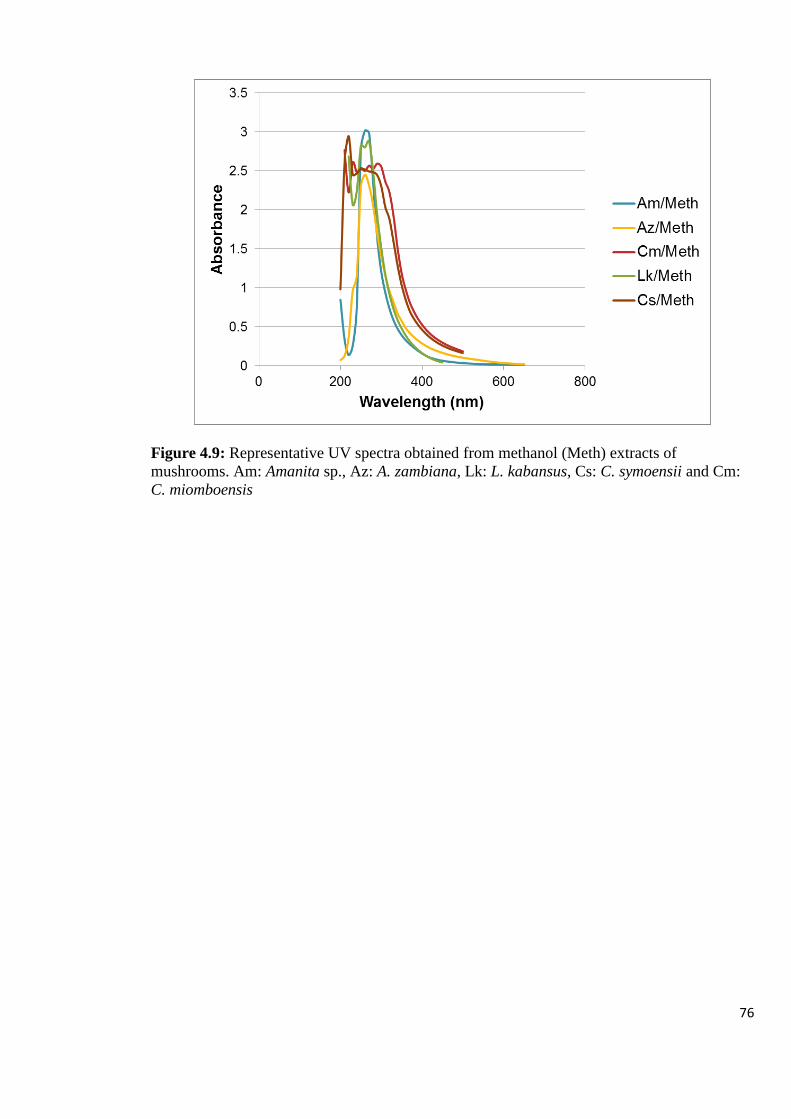
76
Figure 4.9: Representative UV spectra obtained from methanol (Meth) extracts of
mushrooms. Am: Amanita sp., Az: A. zambiana, Lk: L. kabansus, Cs: C. symoensii and Cm:
C. miomboensis

77
Each mushroom type also exhibited a unique absorption profile from the different solvents as
shown in Figure 4.10.
A B
C D
E
Figure 4.10: Representative UV spectra obtained from different solvent extracts of the same
mushroom. (A): methanol (Meth) and acetone (Ac) extracts of L. kabansus (Lk), (B):
methanol, acetone and ethanol (Eth) extracts of Amanita sp. (Am), (C) ethanol, acetone and
methanol extracts of C. symoensii (Cs), (D): ethanol, cold water (Cw) and methanol extracts
of C. miomboensis (Cm), (E): methanol and acetone extracts of A. zambiana (Az)

78
Analysis of the mushroom crude extracts by TLC and staining with vanillin - sulphuric acid
revealed different profiles as shown in Figure 4:11 .
1 2 3 4 5 6 7 8
Figure 4.11: Representative chromatograms of mushrooms extracts developed in Toluene
Ethyl acetate Methanol (TEM) solvent and sprayed with vanillin-sulphuric acid. (1): acetone
extract of L. kabansus, (2): methanol extract of A. zambiana, (3): ethanol extract of G.
lucidum, (4): methanol extract of C. symoensii, (5): methanol extract of C. miomboensis, (6):
acetone extract of T. strumosa, (7): acetone extract of L. kabansus and (8): cold water extract
of C. miomboensis
4.4.2 Separation of Mushroom Crude Extracts by TLC
The retention factor (Rf) values that were obtained from the mushroom extracts using the six
different mobile systems are shown in Table 4.12. The TEM mobile phase gave the highest
total number of separated bands, followed by CM1 with a total of 93 and 92 bands,
respectively. The EEW mobile phase gave the least number of bands (42). The TEM mobile
phase was selected for the PTLC separation of methanol extracts of Amanita species, A.
zambiana, L. kabansus, C. miomboensis and C. symoensii; acetone extracts of A. zambiana,
C. heinemannianus, L. kabansus and T. strumosa and the ethanol extracts of C. symoensii and

79
G. lucidum. The CM1 mobile phase was selected for the PTLC separation of the remaining
extracts, namely, the water extract of C. miomboensis, the acetone extracts of Amanita species
and C. symoensii and the ethanol extracts of Amanita species and C. miomboensis. These two
mobile phases were selected for separation of components of the 16 mushroom extracts
(Table 4.10) by PTLC based on the high number of bands obtained and the good resolution of
bands.
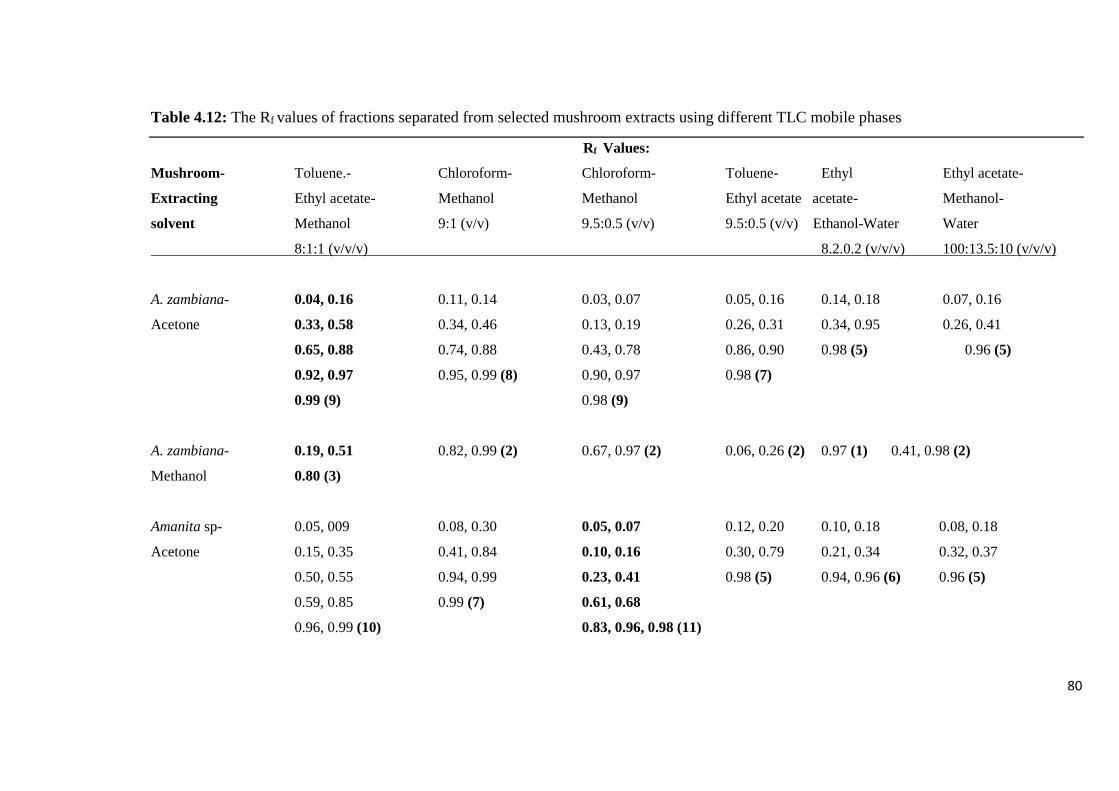
80
Table 4.12: The Rf values of fractions separated from selected mushroom extracts using different TLC mobile phases
Rf Values:
Mushroom- Toluene.- Chloroform- Chloroform- Toluene- Ethyl Ethyl acetate-
Extracting Ethyl acetate- Methanol Methanol Ethyl acetate acetate- Methanol-
solvent Methanol 9:1 (v/v) 9.5:0.5 (v/v) 9.5:0.5 (v/v) Ethanol-Water Water
8:1:1 (v/v/v) 8.2.0.2 (v/v/v) 100:13.5:10 (v/v/v)
A. zambiana- 0.04, 0.16 0.11, 0.14 0.03, 0.07 0.05, 0.16 0.14, 0.18 0.07, 0.16
Acetone 0.33, 0.58 0.34, 0.46 0.13, 0.19 0.26, 0.31 0.34, 0.95 0.26, 0.41
0.65, 0.88 0.74, 0.88 0.43, 0.78 0.86, 0.90 0.98 (5) 0.96 (5)
0.92, 0.97 0.95, 0.99 (8) 0.90, 0.97 0.98 (7)
0.99 (9) 0.98 (9)
A. zambiana- 0.19, 0.51 0.82, 0.99 (2) 0.67, 0.97 (2) 0.06, 0.26 (2) 0.97 (1) 0.41, 0.98 (2)
Methanol 0.80 (3)
Amanita sp- 0.05, 009 0.08, 0.30 0.05, 0.07 0.12, 0.20 0.10, 0.18 0.08, 0.18
Acetone 0.15, 0.35 0.41, 0.84 0.10, 0.16 0.30, 0.79 0.21, 0.34 0.32, 0.37
0.50, 0.55 0.94, 0.99 0.23, 0.41 0.98 (5) 0.94, 0.96 (6) 0.96 (5)
0.59, 0.85 0.99 (7) 0.61, 0.68
0.96, 0.99 (10) 0.83, 0.96, 0.98 (11)
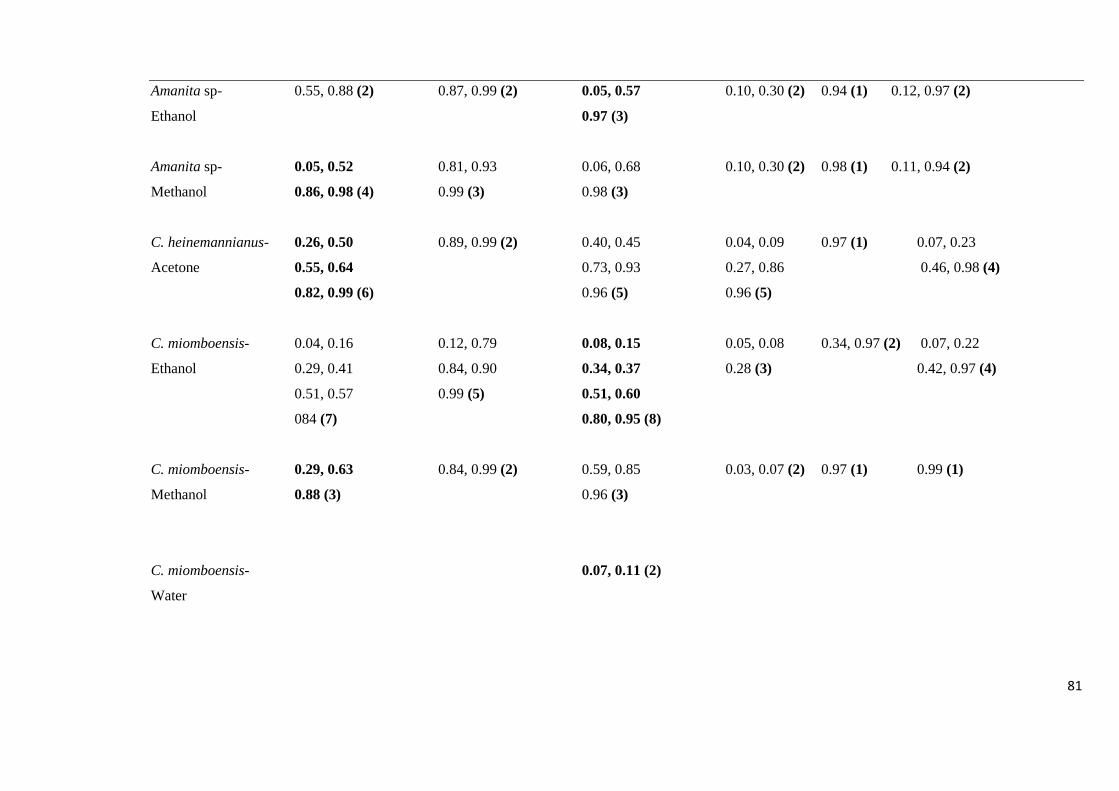
81
Amanita sp- 0.55, 0.88 (2) 0.87, 0.99 (2) 0.05, 0.57 0.10, 0.30 (2) 0.94 (1) 0.12, 0.97 (2)
Ethanol 0.97 (3)
Amanita sp- 0.05, 0.52 0.81, 0.93 0.06, 0.68 0.10, 0.30 (2) 0.98 (1) 0.11, 0.94 (2)
Methanol 0.86, 0.98 (4) 0.99 (3) 0.98 (3)
C. heinemannianus- 0.26, 0.50 0.89, 0.99 (2) 0.40, 0.45 0.04, 0.09 0.97 (1) 0.07, 0.23
Acetone 0.55, 0.64 0.73, 0.93 0.27, 0.86 0.46, 0.98 (4)
0.82, 0.99 (6) 0.96 (5) 0.96 (5)
C. miomboensis- 0.04, 0.16 0.12, 0.79 0.08, 0.15 0.05, 0.08 0.34, 0.97 (2) 0.07, 0.22
Ethanol 0.29, 0.41 0.84, 0.90 0.34, 0.37 0.28 (3) 0.42, 0.97 (4)
0.51, 0.57 0.99 (5) 0.51, 0.60
084 (7) 0.80, 0.95 (8)
C. miomboensis- 0.29, 0.63 0.84, 0.99 (2) 0.59, 0.85 0.03, 0.07 (2) 0.97 (1) 0.99 (1)
Methanol 0.88 (3) 0.96 (3)
C. miomboensis- 0.07, 0.11 (2)
Water

82
C. symoensii- 0.04, 0.20 0.13, 0.81 0.05, 0.07 0.05, 0.10 0.08, 0.15 0.09, 0.23
Acetone 0.30, 0.40 0.81, 0.88 0.11, 0.13 0.27, 0.59 0.32, 0.97 (4) 0.44, 0.99 (4)
0.55, 0.63 0.93, 0.99 (6) 0.16, 0.18 0.77, 0.99 (6)
0.70, 0.87 0.28, 0.33
0.96 (9) 0.46, 0.57
0.61, 0.66
0.79, 0.96, 0.98 (15)
C. symoensii- 0.18, 0.27 0.11, 0.82 0.18, 0.61 0.08, 0.27 (2) 0.32, 0.97 (2) 0.07, 0.21
Ethanol 0.38, 0.49 0.87, 0.94 0.71, 0.91 0.43, 0.99 (4)
0.55, 0.63 0.99 (5) 0.96 (5)
0.80, 0.99 (8)
C. symoensii- 0.19, 0.28 0.10, 0.80 0.51, 0.62 0.06 0.97 (1) 0.08, 0.20
Methanol 0.49, 0.55, 0.83 (5) 0.98 (3) 0.82, 0.96 (4) 0.26 (2) 0.41, 0.99 (4)
G. lucidum- 0.24, 0,46 0.08, 0.21 0.13, 0.62 0.10, 0.21 0.42, 0.94 0.40, 0.84
Ethanol 0.55, 0.60 0.27, 0.78 0.74, 0.84 0.30, 0.98 (4) 0.98 (3) 0.98 (3)
0.66, 0.85 0.91, 0.99 (6) 0.90, 0.97
0.92, 0.99 (8) 0.99 (7)
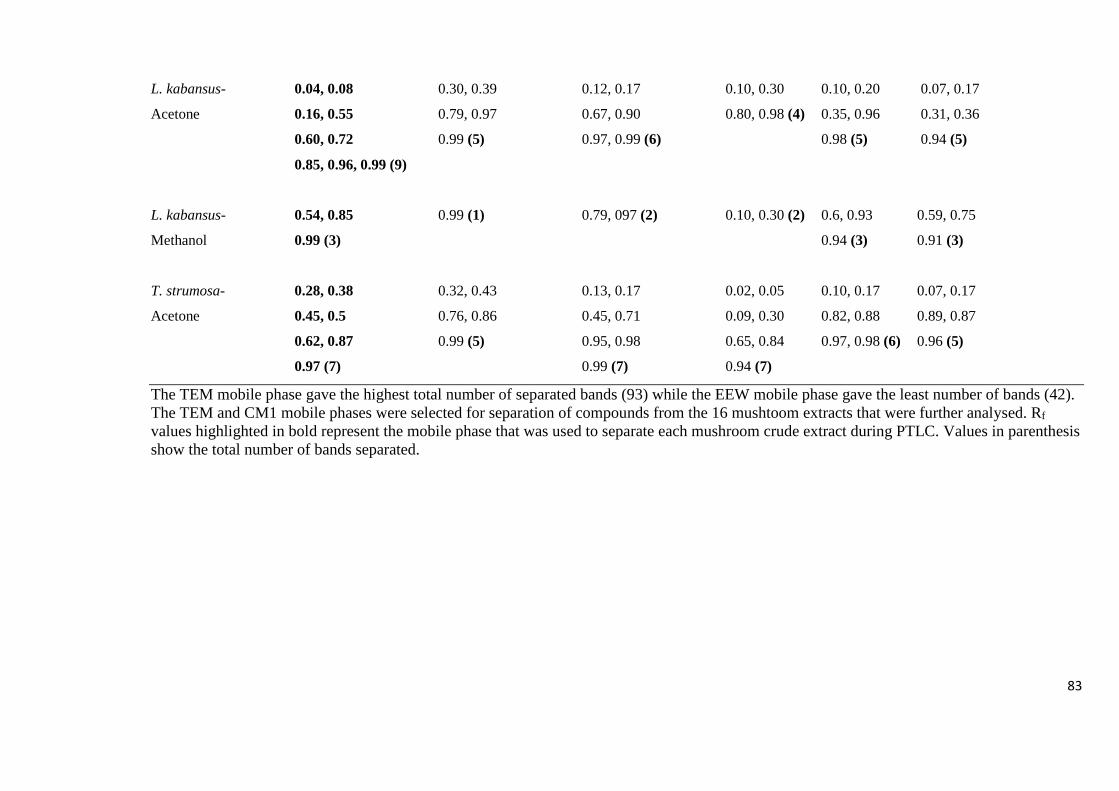
83
L. kabansus- 0.04, 0.08 0.30, 0.39 0.12, 0.17 0.10, 0.30 0.10, 0.20 0.07, 0.17
Acetone 0.16, 0.55 0.79, 0.97 0.67, 0.90 0.80, 0.98 (4) 0.35, 0.96 0.31, 0.36
0.60, 0.72 0.99 (5) 0.97, 0.99 (6) 0.98 (5) 0.94 (5)
0.85, 0.96, 0.99 (9)
L. kabansus- 0.54, 0.85 0.99 (1) 0.79, 097 (2) 0.10, 0.30 (2) 0.6, 0.93 0.59, 0.75
Methanol 0.99 (3) 0.94 (3) 0.91 (3)
T. strumosa- 0.28, 0.38 0.32, 0.43 0.13, 0.17 0.02, 0.05 0.10, 0.17 0.07, 0.17
Acetone 0.45, 0.5 0.76, 0.86 0.45, 0.71 0.09, 0.30 0.82, 0.88 0.89, 0.87
0.62, 0.87 0.99 (5) 0.95, 0.98 0.65, 0.84 0.97, 0.98 (6) 0.96 (5)
0.97 (7) 0.99 (7) 0.94 (7)
The TEM mobile phase gave the highest total number of separated bands (93) while the EEW mobile phase gave the least number of bands (42).
The TEM and CM1 mobile phases were selected for separation of compounds from the 16 mushtoom extracts that were further analysed. Rf
values highlighted in bold represent the mobile phase that was used to separate each mushroom crude extract during PTLC. Values in parenthesis
show the total number of bands separated.

84
4.4.3 Isolation of Components of Mushroom Crude Extracts by PTLC
After separation of the sixteen crude extracts by PTLC, each of the bands was scrapped off
together with silica gel and eluted with ethyl acetate. A total of one hundred and thirty one
fractions (CP1 to CP131) were obtained from the 16 extracts as shown in Table 4.13.
Table 4.13: Number of fractions obtained after scrapping bands from each of the separated
crude extract on the PTLC plate
Mushroom crude extract Number of fractions obtained
C. miomboensis – methanol 7 (CP1 to CP7)
C. symoensii – methanol 8 (CP8 to CP15)
A. zambiana – methanol 7 (CP16 to CP22)
C. heinemannianus – acetone 8 (CP23 to CP30)
A. zambiana – acetone 9 (CP31 to CP38)
C. symoensii – ethanol 9 (CP39 to CP47)
T. strumosa – acetone 8 CP48 to CP55)
Amanita sp. – methanol 7 (CP56 to CP62)
L. kabansus – acetone 9 (CP63 to CP71)
G. lucidum – ethanol 8 (CP72 to CP80)
L. kabansus – methanol 10 (CP81 to CP90)
C. symoensii – acetone 7 (CP91 to CP97)
C. miomboensis – water 6 (CP98 to CP103)
C. miomboensis – ethanol 11 (CP104 to CP114)
Amanita sp. – acetone 8 (CP115 to CP122)
Amanita sp. – ethanol 9 (CP123 to CP131)
Total number of fractions 131
The ethanol extract of C. miomboensis had the highest number of compounds separated (11),
closely followed by the methanol extract of L. kabansus (10).
4.4.4 Determination of the Antibacterial Activity of the Isolated Fractions
The compounds (CP1 – CP131) isolated from the mushroom crude extracts exhibited varying
degrees of inhibition of bacterial growth ranging from 0 to 99.8 % (Appendix 8.3). Thirteen
of the isolated compounds exhibited very high inhibitory activity against growth of S. typhi

85
ranging from 86.6 to 99.8 %, while one of the components (CP50) exhibited high inhibitory
activity against growth of S. aureus (87.5 %), as shown in Table 4.14. The highest bacterial
inhibitory activities were observed from the compounds isolated from the ethanol extract of
G. lucidum (CP73) and the acetone extract of C. symoensii (CP94), each with 99.8%
inhibitory activity against S. typhi, closely followed by the compound from acetone extract of
L. kabansus (CP70) (99.7 %) against S. typhi. However, CP70 was the most potent isolate as
it exhibited the lowest IC50 value of 206 µg/ml, followed by CP94 and the compound isolated
from the ethanol extract of C. symoensii (CP44) with IC50 values of 223 µg/ml and 245 µg/ml,
respectively.

86
Table 4.14: Percentage inhibition of growth of S. typhi and S. aureus by the fourteen potent
compounds isolated from different mushroom crude extracts using PTLC
Isolate
no.
Rf
value
% inhibition IC50
(µg/ml)
Source of
Isolate
Bacteria
tested 200 µg/ml 400 µg/ml 600 g/ml 800 µg/ml
CP25 0.26 15.4 ± 5.7 41.3 ± 9.9 99.0 ± 0 98.7 ± 0 413 Acetone extract
of C.
heinemmanianus
S. typhi CP26 0.55 7.1 ± 10.0 11.3 ± 16.0 82.1 ± 0 97.4 ± 0 505
CP27 0.63 10.8 ± 2.0 62.2 ± 0 88.4 ± 0 95.7 ± 1.8 350
CP39 0.09 6.2 ± 5.0 38.6 ± 9.1 69.7 ± 3.2 86.6 ± 1.3 462 Ethanolic extract
of C. symoensii
S. typhi
CP44 0.50 33.0 ± 0 86.5 ± 5.2 92.7 ± 0 92.9 ± 0.1 245
CP50 0.20 11.4 ± 0.0 13.5 ± 0.1 57.6 ± 0.1 87.5 ± 0.3 564 Acetone extract
of T. strumosa
S. aureus
CP70 0.76 29.3 ± 29.6 98.7 ± 0.3 99.7 ± 0 99.7 ± 0 206 Acetone extract
of L. kabansus
S. typhi
CP73 0.20 0.0 ± 0 2.3 ± 3.0 78.3 ± 29 99.8 ± 0.2 573 Ethanolic extract
of G. lucidum
S. typhi
CP76 0.38 13.9 ± 18.5 10.0 ± 14.1 95.5 ± 5.3 98.3 ± 1.4 467
CP77 0.43 2.4 ± 2.1 41.1 ± 4.4 97.4 ± 1.4 94.9 ± 3.7 415
CP78 0.56 0.0 ± 0 0.6 ± 0.9 97.5 ± 2.4 97.7 ± 0 506
CP92 0.07 0.0 ± 0 0.0 ± 0 38.0 ± 16 98.4 ± 0.2 619 Acetone extract
of C. symoensii
S. typhi
CP94 0.55 16.1 ± 22.7 98.2 ± 0 99.7 ± 0 99.8 ± 0 223
CP98 0.03 4.3 ± 6.0 93.3 ± 5.6 95.7 ± 2.2 96.9 ± 0 291 Cold water
extract of C.
miomboensis
S. typhi
Ampicillin
(50 µg/ml)
96.9 ± 0.4 S. typhi
91.3 ± 0.5 S. aureus
The negative control was used to calculate the percentage inhibition of bacterial growth by
each extract. The highest bacterial inhibitory activities were observed from the compounds
isolated from the ethanol extract of G. lucidum (CP73) and the acetone extract of C. symoensii
(CP94), each with 99.8% inhibitory activity against S. typhi, closely followed by the
compound from acetone extract of L. kabansus (CP70) (99.7 %) against S. typhi. However,
CP70 was the most potent isolate as it exhibited the lowest IC50 value of 206 µg/ml.
4.4.5 Identification of the Most Potent Antibacterial Compounds of Mushroom Extracts
by LC-MS
Analysis of seven of the most potent isolated mushroom components (Section 4.4.4) using an
Agilent HPLC 1260 System coupled to an Agilent 6530 Accurate-Mass Quadrupole Time-of-
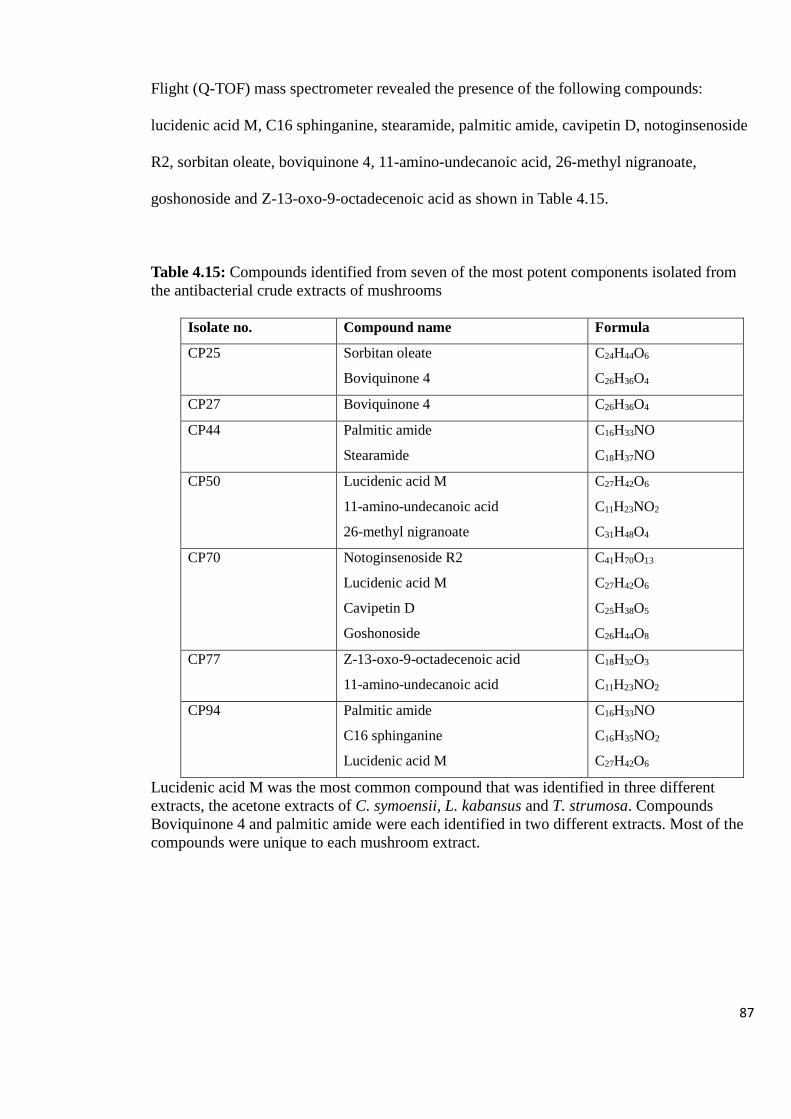
87
Flight (Q-TOF) mass spectrometer revealed the presence of the following compounds:
lucidenic acid M, C16 sphinganine, stearamide, palmitic amide, cavipetin D, notoginsenoside
R2, sorbitan oleate, boviquinone 4, 11-amino-undecanoic acid, 26-methyl nigranoate,
goshonoside and Z-13-oxo-9-octadecenoic acid as shown in Table 4.15.
Table 4.15: Compounds identified from seven of the most potent components isolated from
the antibacterial crude extracts of mushrooms
Isolate no. Compound name Formula
CP25 Sorbitan oleate
Boviquinone 4
C24H44O6
C26H36O4
CP27 Boviquinone 4 C26H36O4
CP44 Palmitic amide
Stearamide
C16H33NO
C18H37NO
CP50 Lucidenic acid M
11-amino-undecanoic acid
26-methyl nigranoate
C27H42O6
C11H23NO2
C31H48O4
CP70 Notoginsenoside R2
Lucidenic acid M
Cavipetin D
Goshonoside
C41H70O13
C27H42O6
C25H38O5
C26H44O8
CP77 Z-13-oxo-9-octadecenoic acid
11-amino-undecanoic acid
C18H32O3
C11H23NO2
CP94 Palmitic amide
C16 sphinganine
Lucidenic acid M
C16H33NO
C16H35NO2
C27H42O6
Lucidenic acid M was the most common compound that was identified in three different
extracts, the acetone extracts of C. symoensii, L. kabansus and T. strumosa. Compounds
Boviquinone 4 and palmitic amide were each identified in two different extracts. Most of the
compounds were unique to each mushroom extract.

88
4.5 DETERMINATION OF THE ANTI-HIV ACTIVITY OF MUSHROOM CRUDE
EXTRACTS
4.5.1 Determination of Anti-HIV-1 Reverse Transcriptase Activity
The HIV-1 RT inhibition ratio of mushroom extracts from the ten wild edible and non-edible
mushrooms used in this study ranged from 0.7 % to 92.6 % as shown in Figures 4.12 to 4.13 .
The cold water extracts from all the mushrooms gave the highest levels of RT inhibitory
activity, which was all above 50%, followed by the boiled water extracts, while acetone
extracts gave the lowest values. The cold water extract of the mushroom L. kabansus
demonstrated the highest inhibition ratio of 92.6 %, closely followed by the cold water
extracts of Amanita sp., B. edulis, T. strumosa, A. zambiana, C. heinemannianus and C.
miomboensis with inhibition ratios of 91.3 %, 89.9 %, 89.6 %, 88 %, 86.2 % and 81.2 %,
respectively. From the ten different mushroom types studied, Amanita sp. and C. miomboensis
had the most number of extracts exhibiting high anti-RT activity.

89
1 2 3 4 5 6 7 8
Figure 4.12: Representative assay for anti-HIV RT activity of mushroom crude extracts. A1-
B1: negative control, C1-D1: positive control (0.2 ng/µl), columns 2-8: different mushroom
crude extracts (100 µg/ml) in duplicate wells for each extract. The resulting signal intensity,
which was measured spectrophotometrically, was directly proportional to the actual RT
activity. The more intense green colour represented extracts that were less effective in
inhibiting HIV RT than the lighter coloured wells.
A
B
C
D
E
G
F
H
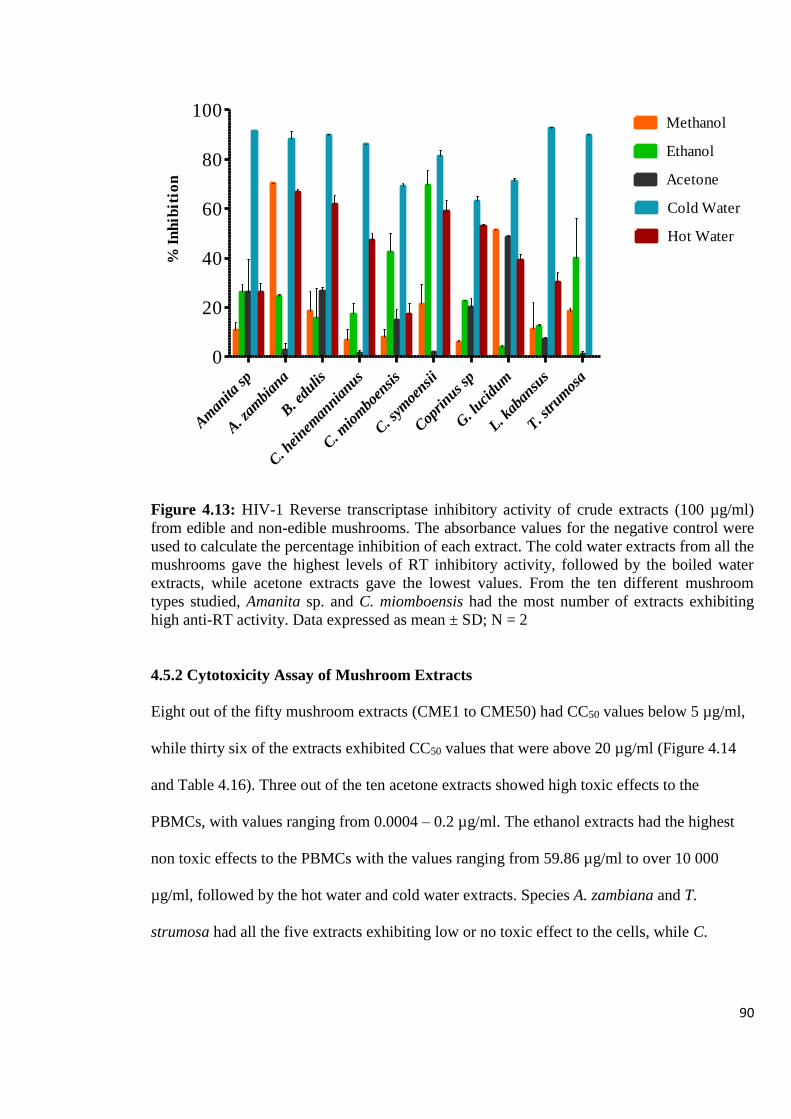
90
Am
anita
sp
A. z
ambi
ana
B. e
dulis
C. h
einem
annia
nus
C. m
iom
boen
sis
C. s
ymoe
nsii
Cop
rinus
sp
G. l
ucidu
m
L. kab
ansu
s
T. strum
osa
0
20
40
60
80
100Methanol
Ethanol
Acetone
Cold Water
Hot Water
% I
nh
ibit
ion
Figure 4.13: HIV-1 Reverse transcriptase inhibitory activity of crude extracts (100 µg/ml)
from edible and non-edible mushrooms. The absorbance values for the negative control were
used to calculate the percentage inhibition of each extract. The cold water extracts from all the
mushrooms gave the highest levels of RT inhibitory activity, followed by the boiled water
extracts, while acetone extracts gave the lowest values. From the ten different mushroom
types studied, Amanita sp. and C. miomboensis had the most number of extracts exhibiting
high anti-RT activity. Data expressed as mean ± SD; N = 2
4.5.2 Cytotoxicity Assay of Mushroom Extracts
Eight out of the fifty mushroom extracts (CME1 to CME50) had CC50 values below 5 µg/ml,
while thirty six of the extracts exhibited CC50 values that were above 20 µg/ml (Figure 4.14
and Table 4.16). Three out of the ten acetone extracts showed high toxic effects to the
PBMCs, with values ranging from 0.0004 – 0.2 µg/ml. The ethanol extracts had the highest
non toxic effects to the PBMCs with the values ranging from 59.86 µg/ml to over 10 000
µg/ml, followed by the hot water and cold water extracts. Species A. zambiana and T.
strumosa had all the five extracts exhibiting low or no toxic effect to the cells, while C.
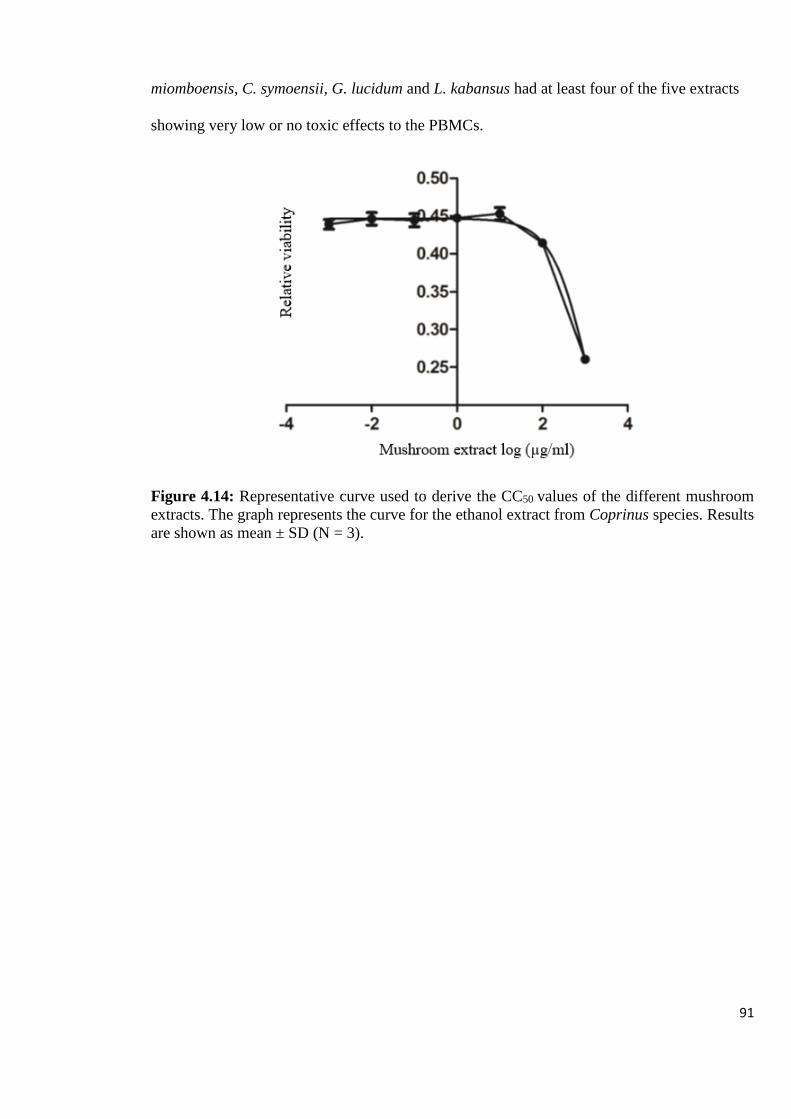
91
miomboensis, C. symoensii, G. lucidum and L. kabansus had at least four of the five extracts
showing very low or no toxic effects to the PBMCs.
Figure 4.14: Representative curve used to derive the CC50 values of the different mushroom
extracts. The graph represents the curve for the ethanol extract from Coprinus species. Results
are shown as mean ± SD (N = 3).

92
Table 4.16: The CC50 values of mushroom extracts showing varying levels of inhibitory
effects to the growth of PBMCs
CC50 (µg/ml) of mushroom extracts
Mushroom Acetone Ethanol Methanol Cold water Hot water
Amanita sp. 0.0004 123.4 0.03 19.7 80.82
A. zambiana 111.6 >10 000 >1 000 890.4 784.7
B. edulis 6.48 718.5 >10 000 >10 000 16.8
C. heinemannianus 0.006 NC 2 591 126.1 >10 000
C. miomboensis >1 000 149.8 46.84 4.07 208.6
C. symoensii 142.8 59.86 5.81 547.1 494.7
Coprinus sp. 7.1 1 184 0.11 0.27 >10 000
G. lucidum 0.2 >10 000 138.5 >10 000 1 740
L. kabansus 331.2 224.2 2 612 167.3 0.3
T. strumosa 153.1 3 444 1 009 57.14 2 425
The ethanol extracts had the highest non toxic effects to the PBMCs with the values ranging
from 59.86 µg/ml to over 10 000 µg/ml, followed by the hot water and cold water extracts.
Species A. zambiana and T. strumosa had all the five extracts exhibiting low or no toxic effect
to the cells. NC – data did not converge.
4.5.3 Determination of the in vitro Anti-HIV-1 Activity using HIV-1 p24 Antigen
Expression Assay
Twenty extracts of the CME1 to CME50 set, with CC50 values ranging from 80.82 to over 10
000 µg/ml were selected for the anti-HIV-1 activity on PBMCs. The percentage inhibition of
HIV-1c infection by the selected mushroom crude extracts ranged from 16.8 % to 97.8 % as
shown in Table 4.17. Two of the twenty extracts, namely, the boiled water extracts from
Coprinus species and C. heinemannianus, exhibited high inhibition of HIV-1c infection (92.8
± 1.7% and 97.8 % ± 0, respectively), with IC50 values of 24.3 µg/ml and 33.8 µg/ml,
respectively and no cellular cytotoxicity to the cells (Table 4.18).

93
Table 4.17: Inhibitory effects of different mushroom extracts on HIV-1 replication
Mushroom Extracting solvent HIV-1 p24 inhibition (%)
50 µg/ml 100 µg/ml 150 µg/ml
C. miomboensis Ethanol 42.8 ± 2.4 41.1 ± 0.0 52.3 ± 0.9
B. edulis Ethanol 38.5 ± 3.7 38.9 ± 1.5 32.3 ± 10.9
T. strumosa Ethanol 24.7 ± 2.4 45.8 ± 8.3 46.8 ± 1.7
C. heinemannianus Methanol 16.8 ± 6.4 29.2 ± 5.2 32.5 ± 6.1
L. kabansus Methanol 49.3 ± 5.2 54.5 ± 22.0 40.3 ± 9.2
T. strumosa Methanol 17.1 ± 0.5 33.9 ± 2.8 40.4 ± 10.7
G. lucidum Methanol 71.7 ± 0 67.0 ± 0 73.3 ± 0
B. edulis Methanol 52.4 ± 6.3 48.0 ± 3.4 59.0 ± 1.0
L. kabansus Cold water 39.5 ± 20.9 59.8 ± 16.8 59.1 ± 6.9
C. symoensii Cold water 45.6 ± 0 65.5 ± 0 59.0 ± 2.9
C. heinemannianus Cold water 44.0 ± 5.6 48.8 ± 7.7 39.6 ± 5.3
B. edulis Cold water 48.7 ± 6.0 51.8 ± 7.2 34.8 ± 1.4
G. lucidum Cold water 68.5 ± 0 60.1 ± 18.8 54.9 ± 2.8
Amanita species Hot water 43.4 ± 3.2 - 39.5 ± 12.0
T. strumosa Hot water 49.6 ± 22.4 49.9 ± 19.9 38.7 ± 9.7
A. zambiana Hot water 67.9 ± 7.7 37.6 ± 14.6 68.7 ± 24.1
C. symoensii Hot water 52.2 ± 12.6 50.5 ± 5.3 59.5 ± 19.6
C. heinemannianus Hot water* 65.6 ± 0.2 54.3 ± 1.9 97.8 ± 0
G. lucidum Hot water 54.2 ± 0 60.1 ± 28.4 73.3 ± 0
Coprinus species Hot water* 75.8 ± 16.7 92.6 ± 1.4 92.8 ± 1.7
Azidothymidine (20 µg/ml) 85.4 ± 2.6
(-) results not converging. Results are shown as mean ± SD (n = 2). The absorbance values
obtained from the negative control were used to calculated the percentage inhibition for each
extract. Rows highlighted in bold indicated extracts that exhibited high inhibitory effects of
above 90 %. Samples marked with asterisk* were selected for analysis of compound profile by
LC – MS.
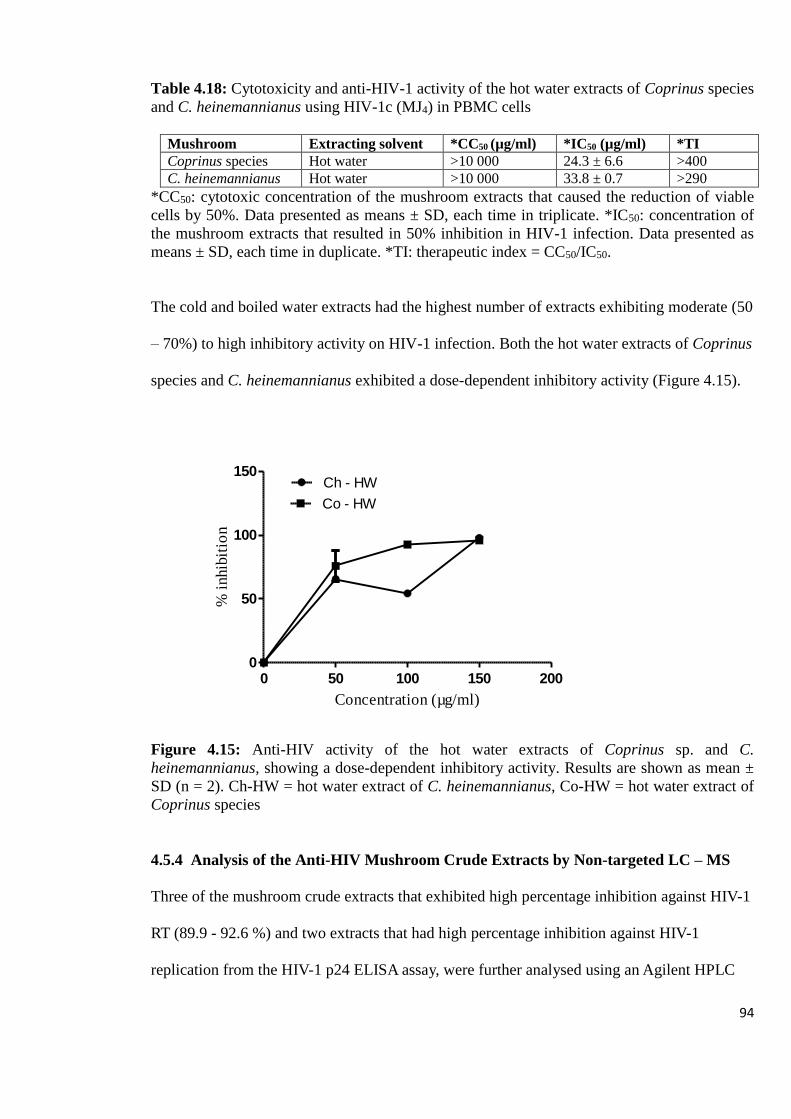
94
Table 4.18: Cytotoxicity and anti-HIV-1 activity of the hot water extracts of Coprinus species
and C. heinemannianus using HIV-1c (MJ4) in PBMC cells
Mushroom Extracting solvent *CC50 (µg/ml) *IC50 (µg/ml) *TI
Coprinus species Hot water >10 000 24.3 ± 6.6 >400
C. heinemannianus Hot water >10 000 33.8 ± 0.7 >290
*CC50: cytotoxic concentration of the mushroom extracts that caused the reduction of viable
cells by 50%. Data presented as means ± SD, each time in triplicate. *IC50: concentration of
the mushroom extracts that resulted in 50% inhibition in HIV-1 infection. Data presented as
means ± SD, each time in duplicate. *TI: therapeutic index = CC50/IC50.
The cold and boiled water extracts had the highest number of extracts exhibiting moderate (50
– 70%) to high inhibitory activity on HIV-1 infection. Both the hot water extracts of Coprinus
species and C. heinemannianus exhibited a dose-dependent inhibitory activity (Figure 4.15).
0 50 100 150 2000
50
100
150Ch - HW
Co - HW
Concentration (µg/ml)
% i
nhib
itio
n
Figure 4.15: Anti-HIV activity of the hot water extracts of Coprinus sp. and C.
heinemannianus, showing a dose-dependent inhibitory activity. Results are shown as mean ±
SD (n = 2). Ch-HW = hot water extract of C. heinemannianus, Co-HW = hot water extract of
Coprinus species
4.5.4 Analysis of the Anti-HIV Mushroom Crude Extracts by Non-targeted LC – MS
Three of the mushroom crude extracts that exhibited high percentage inhibition against HIV-1
RT (89.9 - 92.6 %) and two extracts that had high percentage inhibition against HIV-1
replication from the HIV-1 p24 ELISA assay, were further analysed using an Agilent HPLC

95
1260 System coupled to an Agilent 6530 Accurate-Mass Quadrupole Time-of-Flight (Q-TOF)
Mass Spectrometer. Analysis of the crude extracts revealed the presence of a variety of
secondary metabolites which included alkaloids, penicillins, carboxylic acid esters, fatty acids
and fatty acid derivatives, diterpenes, diterpenoid alkaloids, amino acid derivatives,
sesqueterpenoids, terpene derivatives, and phospholidids.

96
CHAPTER FIVE:
DISCUSSION

97
5.1 CHARACTERIZATION OF MUSHROOM FRUITING BODIES
5.1.1 Quantitative Determination of the Total Protein Content
Nutritionally, edible mushrooms are highly valued due to high protein content and other
nutrients than most of the plants. Some investigators have equated the amino acid
composition of mushrooms to that of animal proteins (Boda et al., 2012; Reis et al., 2011).
The results obtained confirm that the edible mushrooms used in this study are good sources of
protein. Overall, the edible mushrooms such as B. edulis, L. kabansus and A. zambiana had
high levels of protein content reaching 30.8 % ± 1.3 when compared to non-edible
mushrooms such as G. lucidum (9.3 % ± 1.0) and T. strumosa (10.0 % ± 1.1) as shown in
Figure 4.2. Proteins are the workhorses of the cell that carry out all the key functions such as
digesting food, building tissue, transporting oxygen in the blood to the body tissues, dividing
cells and powering muscles. A good protein source can prevent the occurrence of protein
deficiency diseases such as kwashiorkor and marasmus. Malnutrition, with its constituents of
protein and micronutrient deficiencies, is a major health burden in developing countries. It is
the most important risk factor for illness and death globally, affecting hundreds of millions of
pregnant women and young children (Henchion et al., 2017; Luchuo et al., 2013; Schonfeldt
et al., 2013). In Zimbabwe, around 650,000 children under 5 years (27 %) suffer from chronic
malnutrition (stunting) (USAID, 2018). Thus, local edible mushrooms such as A. zambiana,
B. edulis, Cantharellus species and L. kabansus can be used in the development of mushroom
based protein products that are able to minimize protein-energy malnutitrion, particularly
stunting in children. These mushrooms can be used to enrich human diets, especially in
developing countries where animal protein may not be readily available and out of the reach
of most household budgets (Teklit, 2015).

98
Mushrooms have been reported to contain protein levels ranging from 15 % to 35 % dry
weight (Masamba and Kazombo-Mwale, 2010). The values obtained from this study ranged
from 9.3 % ± 1.0 to 30.8 % ± 1.3 dry weight, which compare favourably well with the general
protein content of mushroom species reported in previous studies (Masamba and Kazombo-
Mwale, 2010). In addition, the three Cantharellus species (C. heinemannianus, C.
miomboensis and C. symoensii), had different levels of protein content (21.1 % ± 2.4, 16.8 %
± 0.3 and 30.7% ± 4.8, respectively). Similar studies where mushrooms of the same genus
gave different protein content values on a dry weight basis have been reported (Masamba and
Kazombo-Mwale, 2010).
5.1.2 Qualitative Determination of Carbohydrates
Carbohydrates are a major constituent of mushroom dry matter (usually about 50 - 60 %)
comprising of various compounds; monosaccharides and their derivatives, oligosaccharides
(commonly called sugars) and polysaccharides (Kalac, 2012). The results from both the
Molisch’s and the Benedict’s tests showed that most of the mushrooms contained varying
levels of carbohydrates, based on the different intensities of the positive reactions (Table 4.2).
Species Amanita, C. symoensii, L. kabansus and T. strumosa showed high levels of
carbohydrates while, C. heinemannianus and C. miomboensis showed high levels of reducing
sugars. High levels of both carbohydrates and reducing sugars were exhibited by A. zambiana
when compared to the other mushrooms studied. To a certain extent, the aforementioned
mushroom species can in addition to providing protein nutrition (section 5.1.1), augment the
energy providing portions of household diets to result in a greater chance of balanced diets in
Zimbabwean communities and lower tier developing countries. Several reports have also
highlighted the variation in the carbohydrate content in different species of mushrooms (Boda
et al., 2012; Cheung, 2008).

99
5.1.3 Determination of Lectins
Mushrooms produce many kinds of proteins with important biological activities, including
lectins and ribonucleases. In this study, the lectin activity of the ten different wild
mushrooms, using two types of red blood cells, namely sheep and goat, were studied. The
results showed that three of the ten mushrooms, namely Amanita sp., B. edulis and L.
kabansus contained varying levels of lectin content with B. edulis having the highest specific
activity of 617 HAU/ mg mushroom, and L. kabansus having the lowest activity of 5 HAU/
mg mushroom (Table 4.4). In other related studies, high content of lectins in mushrooms has
also been detected in B. edulis, Lactarius and Amanita sp. (Dhamodharan and Mirunalini,
2011; Singh and Bhari, 2014). Lectins obtained from mushrooms are reportedly able to bind
to abnormal and cancer cells and label these cells for destruction by the human body’s
immune system (Hassan et al., 2015). Thus, the mushrooms under study, particularly B.
edulis and L. kabansus, could be an excellent source of cancer-fighting macromolecules and
can be used in the human diet to promote health. Extracts from the mushrooms can be
obtained and commercialized as dietary supplements for their ability to enhance immune
function and antitumor activity.
5.2 CHARACTERIZATION OF MUSHROOM CRUDE EXTRACTS
5.2.1 Determination of Protein Content of Crude Extracts
Results from the protein analysis in Figure 4.5 showed high yields of protein from the cold
and boiled water extracts. The high protein yields observed in the boiled water extracts could
be due to increase in protein availability during boiling as a result of hydrolysis of insoluble
protein to soluble protein. Protein yields from methanol extracts followed the aqueous
extracts while ethanol and acetone gave similar low yields. The mushroom species Amanita
gave significantly high yields of protein in the cold and boiled water extracts, while B. edulis

100
had high yields in the cold and boiled water extracts, as well as in all the methanol extracts.
Thus, both cold and boiled water would be the solvents of choice in the development of
mushroom protein extracts that could be used as healthy dietary supplements. The results also
indicate that boiling mushrooms is the method of choice for cooking the edible mushrooms
under study, due to the enhanced availability of the protein content. The yields of protein
obtained correlated with the results of the total protein content by the Kjeldhal method
(section 4.2.1), particluary for the B. edulis mushroom which gave the highest yield and T.
strumosa which gave the lowest yield in both the assays. It has been reported that the amount
of protein varies from species to species in the same genus as observed in the two different
species of the genus Cantharellus (C. cibarius, 1.057 mg/ml of protein and C. subcibarius,
1.567 mg/ml of protein) (Boda et al., 2012). Similarly, the three different species of the genus
Cantharellus gave different protein yields (Figure 4.5).
5.2.2 Determination of Total Phenolic Content of Crude Extracts
Phenolic compounds are crucial due to their free radical scavenging activity, hence, they
protect the human body from cell damage by reactive oxygen species (Gan et al., 2013). Total
phenolic compounds have been reported as the major naturally occurring antioxidant
compounds in the wild edible mushrooms (Wang and Xu, 2014). In this study, crude
mushroom extracts were prepared from the mushrooms using cold water, boiling water,
methanol, ethanol and acetone, and total phenolic compounds were determined. High levels of
total phenolics were observed in both the cold and boiled water extracts (Figure 4.6), when
compred to the organic extracts. Similar trends, where cold water extracts had high total
phenolic yields followed by hot water extracts, while acetone extracts gave the least yields,
were observed by Wang and Xu (2014). The high values in water extracts could be explained
by the high polarity of water compared to the other organic solvents, hence, more polar

101
compounds dissolving in water. From the ten different mushroom types studied, B. edulis had
the highest total phenolic compounds followed by Amanita species. A diet rich in antioxidants
protects cells from free radicals, helping the body cope with the normal oxidative stress that
damages healthy cells. The ability of phenolic compounds to protect against several
degenerative disorders, including brain dysfunction, cancer and cardiovascular diseases, is
related to their capacity to act as antioxidants (Zhang et al., 2015). Thus, local edible
mushrooms such as B. edulis can be used as healthy foods for the prevention of a range of
illnesses including cancer, diabetes and arteriosclerosis. Powder formulations of these
mushrooms could be prepared and used in diets as antioxidants.
5.2.3 Determination of Antibacterial Activity of Crude Extracts
The results of the antibacterial activities of methanol, ethanol, acetone, cold and hot water
extracts from ten different mushrooms used in this study highlighted that all the mushrooms
possess inhibitory activities against at least one of the bacteria tested. This was shown on
clear zones of inhibition around the tested mushroom extracts. It has been reported that
mushroom species possess different constituents and in different concentrations which
account for their differential antimicrobial activity (Akyuz et al., 2010; Padmavathy et al.,
2014). In this study, the highest in vitro antibacterial activity was shown by the cold water
extract of C. miomboensis against S. typhi (14 mm zone of inhibition). This was followed by
the acetone extract of C. symoensii, the methanol extract from C. miomboensis and the ethanol
extracts of G. lucidum and C. symoensii (section 4.3.3). Extracts of these species, using the
identified optimal solvents for extraction, can potentially be used for antibacterial medicinal
purposes similar to antibiotics. In a separate study, the edible mushroom species C. cibarius
showed antimicrobial activity against some Gram-positive and Gram-negative bacteria (Rahi
and Malik, 2016).

102
Species C. miomboensis, Amanita sp. and B. edulis all had the highest number of total extracts
inhibiting at least one of the bacteria tested (section 4.3.3). Species C. heinemannianus and C.
symoensii had the highest effect on inhibition of bacteria as indicated by having the most
extracts which had high inhibitory properties ranging from 8-14 mm [15.4 % (8) each],
followed by G. lucidum [13.5% (7)]. This shows that C. heinemannianus, C. symoensii and G.
lucidum extracts contain compounds that have more inhibitory potential against the bacteria
studied than the rest of the mushroom extracts used in this study. In similar studies carried out
by Quereshi et al. (2010), methanol, ethanol, acetone and cold water extracts of G. lucidum
from India showed antimicrobial activity against the S. aureus, S. typhi and E. coli bacterial
culture collections. From this study, the methanol extracts showed no inhibition to S. aureus
and E. coli, while the ethanol and acetone extracts inhibited growth of both E. coli and S.
typhi but did not inhibit growth of S. aureus. Ethanol extracts of G. lucidum from Turkey
inhibited growth of E. coli while the methanol extract showed no inhibition (Celik et al.,
2014). In another study, acetone and ethanol extracts of C. cibarius collected in Turkey,
exhibited antibacterial activity against E. coli and S. aureus but showed no inhibition against
S. typhi (Dulger et al., 2004). Results of another study in Nigeria showed that methanol and
ethanol extracts of C. cibarius inhibited E. coli and S. typhi growth but showed no inhibition
against S. aureus and S. pneumoniae (Aina et al., 2012). Similarly, the research findings in
this study showed that methanol, ethanol and acetone extracts of the three Cantharellus
species studied exhibited various degrees of inhibition against the four bacteria tested. This
shows that different species of mushrooms exhibit different antimicrobial activity due to a
number of factors such as the presence of different antimicrobial components, type of the
extracting medium, geographical location of the mushroom and the type of organism being
tested.

103
Extracts obtained from ethanol gave the highest number of bacterial growth inhibition (33),
followed by acetone (31) and methanol (28). In addition, ethanolic extracts showed the
strongest antibacterial activity (8 - 14 mm) among the five extracts against the bacterial
strains, followed by methanol and acetone. Water extracts exhibited the lowest number of
antibacterial activity, despite having the extract with the highest inhibitory effect. This
indicates that the active compounds from the mushrooms studied, which inhibit the growth of
susceptible bacteria, may dissolve better in the organic solvents than in aqueous solvents.
These results are consistent with already reported literature that extracts from organic solvents
give more consistent antimicrobial activity than water extracts (Kamra and Bhatt, 2012;
Tiwari et al., 2011).
It is interesting to note that, although cold and hot water extractions gave highest values of
total phenolic compounds (Figure 4.6), these had the least effect on most bacteria. This shows
that the antibacterial activity in the mushroom extracts depends not only on the presence of
phenolic compounds, but also on the presence of various other secondary metabolites.
Ethanol, acetone and methanol extracts were all effective against all the four bacteria tested,
indicating the broad spectrum of antibacterial activity of the crude extracts. However, Gram-
negative bacteria were slightly more susceptible to the extracts than Gram-positive bacteria
(52 and 46 extracts, respectively). Several different classes of antibiotics block steps in the
synthesis of peptidoglycan, resulting in cells being more susceptible to osmotic lysis.
Although all bacteria have an inner cell wall, Gram-negative bacteria have a unique outer
membrane which prevents certain drugs and antibiotics from penetrating the cell. Thus,
antibiotics that affect the cell wall will impair Gram-positive bacteria and not Gram-negative
bacteria. The results obtained in this study suggest that the antibacterial extracts obtained may

104
act by affecting not just the cell wall, but other cell growth mechanisms like protein synthesis,
bacterial DNA replication and transcription.
Among the four bacteria tested, S. typhi was the most susceptible bacteria as shown by its
highest number of inhibitions as well as the highest number of most potent extracts in the 8 -
14 mm diameter range. A decline in the number of multidrug resistant clinical isolates (S.
typhi) has been reported (Madhulika et al., 2004). Thus, this study shows that the S. typhi
isolate studied, may be a phage type that is susceptible to most antibiotics. The antibacterial
activity of the ethanolic, methanolic and acetone extracts against E. coli, S. typhi, S. aureus
and S. pneumoniae is of great importance in the human healthcare system. Streptococcus
pneumoniae is the most common cause of community acquired pneumoniae (CAP) in
children while E. coli accounts for more than 70 % of the infections of the urinary tract
worldwide (Blossom et al., 2006; Sangeeth et al., 2014). Species S. typhi is the cause of
typhoid fever, which has been an epidemic in Zimbabwe. The S. aureus is the most common
cause of bacterial infections and abscesses of skin, joints and bones (Stanely et al., 2013).
Resistance to antibiotics has been reported in S. aureus, S. pneumoniae, S. typhi and E. coli
(Blossom et al., 2006; Okonko et al., 2009; Rowe et al., 1997; Sangeeth et al., 2014; Stanely
et al., 2013). The bacterial strains used were clinical isolates. Species E. coli and S. aureus are
mostly encountered in urinary tract infections while isolated cases of S. typhi are common.
Thus, the antibacterial activity found in the mushroom extracts can be further investigated for
future use in the development of therapeutic agents to treat infections caused by these
bacteria. The species with the highest antibacterial activity have potential application in
clinical situations where patients are not responding to orthodox antibiotic regimes.
Application of a cocktail of the identified antibacterial extracts may potentially succeed in
minimising occurrence of multi-drug resistant strains of bacterial pathogens [such as the

105
extended-spectrum β-lactamase-producing and gentamicin resistant Enterobacteriaceae
(Magwenzi et al., 2017) and the methicillin-resistant Staphylococcus aureus (Nikaido, 2009)]
that arise through selective pressures of the continual antibiotic environment. Thus, bacteria
that would normally develop antibiotic resistance mutations in the single antibiotic
environment, would face lethality from another mushroom extract component in the
mushroom extract cocktail.
5.2.4 Characterization of Mushroom Extracts Showing High Antibacterial Activity
During mushroom growth, a variety of primary and secondary metabolites can be
accumulated as intracellular and extracellular products, including phenolics, polyketides,
terpenoids, steroids, antibacterial or antifungal proteins, and fatty acids (Shen et al., 2017).
Different mushroom species usually have characteristic metabolite profiles, although they
may show similar antimicrobial activities. Many phenolic compounds, especially the low-
molecular weight phenolic compounds, have been identified in various mushrooms, and their
antimicrobial properties have been demonstrated (Shen et al., 2017). Analysis of the
mushroom crude extracts using absorption spectroscopy confirmed that each extract had a
unique secondary metabolite profile.
5.2.4.1 Analysis of the extracts using absorption spectroscopy and TLC
The UV spectra of mushroom extracts showed absorbance peaks ranging from 227 – 308 nm;
734 – 745 nm and 586 nm (Table 4.11). The absorbance peaks from the same extraction
solvent type varied with each mushroom type. Although the spectrum of the mushrooms
extracted with the same solvent showed some similarities, changes could be seen in the peak
heights at similar wavelengths for the different extracts. This may be a result of different
compounds being present in each of the extracts. Each mushroom type also exhibited different

106
absorption peaks from the different solvents. Some chemical components appeared to be very
similar among the extracts. For example, the ethanol and acetone extracts of Amanita species
were characterised by strong absorption at the wavelength of 259, while the acetone extract of
T. strumosa and the cold water extract of C. miomboensis had a strong absorption at a
wavelength of 228 nm. However, each crude extract had unique characteristic absorption
peaks to reveal its fingerprint feature. The results indicate that the crude extracts obtained
contain a variety of compounds depending on the type of extracting solvent and the type of
mushroom. Absorption spectrum results also indicate that there may be high levels of
phenolic compounds in most of the extracts as these compounds can exhibit maximum
absorption at wavelengths of 220 - 310 nm (Carvalho et al., 2015). Analysis of the extracts
by TLC and staining with vanillin sulphuric acid further confirmed the unique profile of each
extract. These generated profiles can also aid in the identification of unknown mushroom
tissues in conjunction with other morphological and genetic techniques to assist in forensic
cases. This can further be used in distinguishing between almost identical poisonous and non-
poisonous mushroom species resulting from environmental phenotypic plasticity and
determining the effect of the environment on the genome.
5.2.4.2 Separation of mushroom compounds by TLC and Isolation of components of
mushroom extracts by PTLC
The different retention factors (Rf) indicated the presence of a variety of compounds in each
extract ranging from 6 – 11 compounds that could be visualized, depending on the type of
extracting medium and the mushroom (section 4.4.2). The acetone extracts had the highest
number of compounds separated, with an average of 9 bands per sample for the TEM mobile
phase, indicating that acetone extracted more compounds when compared to the other
solvents. It was noted that PTLC yielded more bands than the ones obtained during analytical

107
TLC using the same mobile phases. This difference may be due to the quantity of the samples
spotted on the plate, being too low to be detected during the analytical TLC while these
became more visible when larger amounts of samples were applied.
5.2.4.3 Determination of Antibacterial Activity of the Isolated Compounds
The isolated components (CP1 – CP131) of the mushroom crude extracts exhibited varying
degrees of inhibition of bacterial growth ranging from 0 to 99.8 % (section 4.4.4). This
suggests that the bioactive products which are contained in mushrooms are in different
concentrations which exhibit varying degrees of antimicrobial activity. This result is in
agreement with the findings of Chu (2013) where fractions obtained from different
mushrooms were reported to show varying inhibitory effects on growth of the microorganism.
Thirteen of the isolated components exhibited very high percentage inhibition activity against
growth of S. typhi ranging from 86.6 to 99.8 %, while one of the components (CP50)
exhibited high inhibitory activity against growth of S. aureus (87.5 %), as shown in Table
4.14. The compound CP70 was the most potent isolate as it exhibited the lowest IC50 value of
206 µg/ml, followed by the compounds CP94 and CP44, with IC50 values of 223 µg/ml and
245 µg/ml, respectively.
Some of the mushroom crude extracts that had shown high potency against the bacteria under
study did not yield any components that had high antibacterial activity when tested against the
same bacteria. A number of reasons for this lack of potency may be possible, one of which
may be that the compounds responsible for the actibacterial activity may have been acting in
synergy, hence separating the compounds reduced their potency. Karmegam et al. (2012)
reported that medicinal plant extracts used in combination exhibited higher antibacterial
activity against E. coli and Bacillus cereus than the individual extracts. The other reason may

108
be that the concentrations of the compounds of interest were too low to exert antibacterial
activity.
5.2.4.4 Analysis of Antibacterial Components of Mushroom Extracts by LC-MS
Analysis of seven of the most potent isolated mushroom components using LC-MS revealed
the presence of a number of terpenoid derivatives namely; lucidenic acid M, cavipetin D,
notoginsenoside R2, boviquinone 4, 26-methyl nigranoate and goshonoside (Table 4.15). The
compound lucidenic acid M (Figure 5.1) was identified in 3 of the isolates obtained from the
acetone extracts of C. symoensii, L. kabansus and T. strumosa. The results indicate that the
acetone organic solvent extracted the same compound in the three different mushrooms,
which was not present in the ethanol extract of the same mushroom, C. symoensii (CP44).
Thus, the results are in agreement with the reported findings that the bioactive compounds
isolated vary depending on the type of solvent used for extraction and the mushroom type
(Shen et al., 2017). Lucidenic acid M, which is also known as lucidenate M, is a member of a
class of organic compounds known as triterpenoids (Chen et al., 2017). Mushroom terpenoids
are a large group of secondary metabolites belonging to terpenes with different functional
groups, which have been identified in various mushroom genera/species, including
sesquiterpenoids from Lentinus conatus, Lactarius sp. and Flammulina velutipes, and
triterpenoids from Ganoderma sinense and Ganoderma pfeifferi (Shen et al., 2017). The

109
Figure 5.1: Structure of Lucidenic acid M
diverse biological activity of lucidenic acid, including its antibacterial activity has been
reported (Basnet et al., 2017; Chen et al., 2017; Hsu and Yen, 2014).
Notoginsenoside R2 and 26-methyl nigranoate are also both members of the triterpenoid class
of compounds (Lee et al., 2010; Sun et al., 1996), which were found in the isolates from the
acetone extracts of L. kabansus and T. strumosa, respectively. Cavipetin D (C25H38O5) and
boviquinone 4 are both members of compounds known as diterpenoids. Cavipetin D (Figure
5.2) is a constituent of edible mushrooms and has been reported to exhibit antibacterial
activity (Shen et al., 2009). In this study, cavipetin D was identified in the component isolated
from the acetone crude extract of L. kabansus, an edible mushroom, confirming reports that
the compound is found in edible mushrooms. Boviquinone 4 has also been reported in
mushrooms (Velisek and Cejpek, 2011). Goshonoside, identified in the acetone extract of L.
kabansus, is a diterpenoid glycoside.

110
Figure 5.2: Structure of Cavipetin D
Another class of compounds that were identified in the isolates were the fatty acid derivatives
namely, palmitic amide, stearamide, sorbitan oleate, 11-amino-undecanoic acid (a fatty acid
amine) and Z-13-oxo-9-octadecenoic acid. Palmitic amide was common in two of the isolates:
CP44 and CP94, while stearamide was found present in CP44. Palmitic amide (Figure 5.3)
and stearamide are both primary fatty acid amides coming from palmitic acid and stearic acid,
respectively. Primary fatty acid amides (R-CO-NH2) are a class of compounds that have only
recently been isolated and characterized from biological sources. Stearamide is often used in
the synthesis of organic chemicals and surfactants and is reported to have antibacterial activity
(Ahmed et al., 2017). Sorbitan oleate belongs to the class of organic compounds known as
fatty acid esters and was isolated from the acetone extract of C. heinemannianus. The
compound is an emulsifier and clarification agent in food preparations (Grant et al., 2006).
The compound Z-13-oxo-9-octadecenoic acid was isolated from the ethanol extract of G.
lucidum and its antimicrobial activity has been reported (Idan et al., 2015; Prost et al., 2005).
Figure 5.3: Structure of Palmitic amide

111
The compound C16 sphinganine, which was identified in the acetone extract of C. symoensii,
is a sphingolipid metabolite derived from the same group of compounds as phytosphingosine.
Phytosphingosine (Figure 5.4) is a phospholipid and sphingolipid metabolites such as
sphingosine and ceramide are highly bioactive compounds that are involved in diverse cell
processes, such as cell-cell interaction, cell proliferation, differentiation, and apoptosis. The
compound is one of the most widely distributed natural sphingoid bases, which is abundant in
fungi and plants and is known to inhibit the growth of both Gram-positive and Gram-negative
bacterial strains (Fischer et al., 2012).
Figure 5.4: Structure of Phytosphingosine
Analysis of the isolates further indicated that the majority of the components isolated from the
mushroom crude extracts by preparative TLC contained more than one compound (Table
4.15). This may mean that the compounds present in the fractions have similar polarities,
hence the mobile phase used and the time for the PTLC run may not have been enough to
separate the different compounds. However, each mushroom component exhibited a unique
profile of compounds, although some similar compounds were found in more than one
fraction. The results thus show that a variety of secondary metabolites, that is, tepernoids,
phospholipids and fatty acids isolated from the selected mushrooms are part of the key
compounds responsible for the antibacterial activity observed. These compounds may be
acting in synergy or as individual compounds.

112
5.2.5 Determination of the Anti-HIV Activity of Mushroom Crude Extracts
5.2.5.1 Determination of anti-HIV-1 Reverse Transcriptase Activity
Although anti-HIV-1 treatment has achieved a lot of success by suppressing viral replication
to undetectable levels and has improved the quality of life of HIV-1 infected individuals,
complete, long-term suppression of HIV-1 replication is still a major challenge due to the
rapid emergence of drug resistance. Hence, continuous search for new anti-HIV-1 agents and
novel targets is still an urgent priority as part of a global strategy to combat the spread of
HIV-1 infection. The HIV-1 encodes three enzymatic proteins, reverse transcriptase (RT),
integrase, and protease, which are critical for its replication. The RT is critical during the
early steps of the viral replication cycle since the enzyme is necessary for reverse
transcription of the viral genome (Dahake et al., 2013; Jadaun et al., 2016; Leteane et al.,
2012; Orozco et al., 2016; Tietjen et al., 2016; Zhiming et al., 2016;). In this study, fifty
aqueous and organic crude extracts from the ten mushrooms selected were screened for
inhibitory properties against HIV-1 reverse transcriptase (RT). The cold water extracts of the
mushrooms L. kabansus, Amanita species, B. edulis, T. strumosa, A. zambiana and C.
heinemannianus, demonstrated high inhibition ratios of 92.6%, 91.3 %, 89,9 %, 89.6 %, 88 %
and 86.2, respectively (Figure 4.13). The high levels of RT inhibition by the cold water
extracts, indicate that the active compounds from the mushrooms studied, that inhibit the
HIV-1 RT activity, dissolve better in the aqueous solvents than in organic solvents. From the
ten different mushroom types studied, Amanita species and C. miomboensis had the most
number of extracts exhibiting high anti-RT activity.
Of note is the observation that cold water extracts gave the highest values of total phenolic
compounds in the total phenol assay (section 4.3.2). Thus, the high levels of RT inhibitory
activity observed in this study may indicate a correlation between total phenolic compounds

113
and anti-HIV-1 RT activity. Hot water extracts of A. zambiana, B. edulis, C. miomboensis and
Coprinus sp. had inhibitory values that were above 50 %. Similar trends were reported
elsewhere, where hot water extracts of different edible and medicinal mushroom species
screened for HIV-1 RT inhibitory activity exhibited over 50 % inhibition (Wang et al., 2007).
The results obtained from this assay showed that most of the mushrooms selected for this
study are potential candidates for development of anti-HIV therapeutic agents. The
advantages of developing HIV-1 RT inhibitors based on mushrooms are the availablility in
large quantities of the basidiomes, and the amenability of the majority of the species to large-
scale fermentation to yield mycelia. Additionally, natural products extracted from
mushrooms, especially edible medicinal fungi exhibit lower toxicity and fewer side effects
than chemical drugs (Orozco et al., 2016).
5.2.5.2 Cytotoxicity Assay of Mushroom Extracts
The results obtained from the general cytotoxicity assay of the fifty mushroom extracts
showed that the mushrooms under study generally had low toxic effects or were not toxic to
the cells. However, the type of solvent may have an effect on the level of toxicity of some of
the extracts. In this study, the ethanol extracts had the highest non toxic effects to the PBMCs
with CC50 values ranging from 59.86 µg/ml to over 10 000 µg/ml, followed by the hot and
cold water extracts.
5.2.5.3 Determination of the in vitro Anti-HIV-1 Activity using p24 Antigen Expression
Assay
Twenty mushroom crude extracts with CC50 values ranging from 80.82 µg/ml to over 10 000
µg/ml were evaluated for their effect on HIV-1 replication by measuring the levels of p24
antigen. The results obtained from the assay showed that some of the mushrooms under study

114
contain bioactive compounds that have the potential to prevent entry of HIV-1 viral particles
into the human cells. Four of the twenty extracts exhibited percentage inhibition of HIV-1c
infection ranging from above 70%, while two of the extracts, namely, the boiled water
extracts from Coprinus species and C. heinemannianus, exhibited percentage inhibition of
HIV-1c infection of over 90 % (Table 4.17) and section 4.5.3. Therapeutic indices (TI) of the
hot water extracts of Coprinus species and C. heinemannianus were calculated as a ratio of
CC50 to IC50. A TI of >200 by both extracts strongly suggest that these mushroom types and
their extracts are good candidates for further anti-HIV studies. The cold and boiled water
extracts had the highest number of extracts exhibiting moderate to high inhibitory activity on
HIV-1 infection. The results tally with the observation made from the anti-HIV-1 RT assay
(section 4.5.1), that the active compounds from the mushrooms studied, which inhibit the
HIV-1 infection, dissolve better in the aqueous solvents than in organic solvents.
The extracts of Ganoderma mushroom expressed moderate levels of inhibition of HIV-1
infection (up to 73.3 %). From the studies conducted by Haoses-Gorases and Goraseb (2013)
in Zambia, Tanzania and Namibia, results showed that the use of Ganoderma mushroom
significantly improved the health status of HIV/AIDS patients. The findings show that the
mushrooms studied, in particular, C. heinemannianus, Coprinus species and G. lucidum,
contain compounds that may inhibit the early stages of HIV infection. These identified
mushroom extracts may assist as alternative therapeutic interventions, as part of a holistic
integrated therapeutic management of HIV, which includes other therapeutic interventions
currently being employed.

115
5.2.5.4 Analysis of the Anti-HIV Mushroom Crude Extracts by LC-MS
Three of the mushroom crude extracts that exhibited high percentage inhibiton against HIV-1
RT (section 4.5.1) and two extracts that had high percentage against HIV-1 replication from
the HIV-1 p24 ELISA assay (Table 4.17), were further analysed using an Agilent HPLC 1260
System coupled to an Agilent 6530 Accurate-Mass Quadrupole Time-of-Flight (Q-TOF) mass
spectrometer. Analysis of the crude extracts revealed the presence of a variety of secondary
metabolites which included alkaloids, penicillins, carboxylic acid esters, fatty acids and fatty
acid derivatives, amino acid derivatives, diterpenes, diterpenoid alkaloids, sesqueterpenoids
and other terpene derivatives, and phospholidids. The results obtained are in agreement with
several reports of the anti-HIV activities of a variety of these secondary metabolites obtained
from mushrooms (Adotey et al., 2011; De Silva et al., 2013; El Dine et al., 2008; Finimundy
et al., 2014; Narayan and Rai, 2016; Rahi and Malik, 2016; Wang et al., 2007). For example,
phytosphingosine and C16 sphinganine were both identified in the cold water extracts of B.
edulis and the hot water extract of Coprinus species. Phytosphingosine was further identified
in the cold water extract of L. kabansus. Both compounds whose anti-HIV activities have
been reported, belong to the class of compounds known as phospholipids. The sphinganine-
and phytosphingosine-based compounds have been reported to possess anti-HIV-1 activity by
disrupting the early and late HIV-1 membrane fusion mechanisms (Ashkenazi et al., 2012;
Klug et al., 2014). Sesquiterpenoids and triterpenes, which were common in most of the crude
extracts analysed, have reportedly exhibited anti-HIV-1 activity (El Dine et al., 2008; Wang et
al., 2014).

116
CHAPTER SIX:
CONCLUSION

117
6.0 CONCLUSION
In this study, the nutritional and phytochemical composition of ten wild edible and non-edible
mushroom species found in Zimbabwe, were analysed. The antibacterial and anti-HIV
potential of the bioactive constituents of the selected mushrooms were evaluated. Although
there are many studies on edible and wild mushroom species globally, the reports are based
on species that are country - specific or are popular for consumption in certain geographical
locations. Very little or no work, to the author’s best knowledge, has been carried out in
Zimbabwe to characterize and compare the nutritive value and bioactive composition among
local wild edible and non-edible mushrooms. Only few disparate information related to the
ten mushroom species used in this study can be found. In addition, the role of a large variety
of mushrooms in human health is still a largely unexplored area of research. Characterization
of mushrooms provides information on the biological properties of mushrooms occurring in
Zimbabwe and potentially provides a stepping stone towards the development of new natural
infection - fighting strategies to control viral and microbial infections.
The mushrooms used in this study, especially edible ones, contained considerable amounts of
proteins and carbohydrates which are vital in supplementing nutrition to humans. The
nutritive contents with respect to protein and carbohydrates varied amomg the ten mushroom
species. The mushrooms studied exhibited high levels of total phenolic compounds, which
play a key role in the antioxidant activity of the mushrooms, potentially contributing to cancer
prevention strategies. Some of the mushrooms studied also contained lectins with varying
levels of specific activity. Thus, local wild mushrooms are a valuable source of lectins for
further research into drug development. Furthermore, the mushrooms can be developed into
functional foods or medicines for prevention and treatment of chronic diseases. From this
study, it can be concluded that B. edulis is the mushroom of choice for augumenting a healthy

118
and balanced diet due to the high levels of proteins, lectins and total phenolic content as well
as moderate levels of carbohydrates exhibited by the mushroom.
Extracts of natural materials are a good source for the characterization and development of
compounds with various biological activities. By screening extracts of natural sources such as
mushrooms, it is possible to discover a multitude of structurally diverse drugs and enzyme
inhibitors. Based on the results obtained from the present study, it can be concluded that wild
edible and non-edible mushrooms found in Zimbabwe are rich in resources that possess a
multitude of biological activities. The mushrooms possess compounds than can be used as
antibacterial agents against some common bacterial infections. The tepernoids, phospholipids
and fatty acids were identified as some of the compounds responsible for the antibacterial
activity of mushrooms. From the study, C. symoensii, C. miomboensis and L. kabansus are
recommended as the mushrooms of choice for the development of antibacterial agents against
S. typhi due to the high number and effectiveness of the most potent crude extracts and
isolated compounds that the mushrooms exhibited. Similarly, L kabansus and Amanita
species are the mushrooms of choice for screening for antibacterial agents against
Streptococcus pneumoniae while T. strumosa can be further investigated for the antibacterial
activity against S. aureus.
The replicative cycle of HIV comprises a number of steps that could be considered as targets
for therapeutic intervention, hence any effective treatment of HIV- 1 infection should target as
many aspects of viral life cycle as possible. From this study, it can be concluded that the local
mushrooms have compounds that are able to inhibit the entry as well as the reverse
transcription stages of the HIV replication cycle. Thus, these mushroom extracts may have
potential for either prophylactic or therapeutic intervention in HIV infection. The anti-HIV

119
activity of the mushroom extracts may be attributed to the secondary metabolites present in
the mushrooms. From the anti-HIV-1 assays, it can be concluded that mushroom species L.
kabansus, Amanita species and B. edulis are the mushrooms of choice for the development of
anti-HIV agents that inhibit reverse transcriptase activity due to their high percentage
inhibition. Similarly, C. heinemannianus and Coprinus species as well as G. lucidum, are the
best candidates for development of anti-HIV agents that can block the entry stage of HIV.
This study confirmed the reports that the chemical profile of the extracts as well as the
antibacterial and anti-HIV effects of mushrooms vary depending on the type of mushroom
and the extracting solvent medium used. Of the five solvents tested, water was determined to
be the solvent of choice for isolation of anti-HIV-1 compounds from the mushrooms studied,
while ethanol and acetone were determined to be the solvents of choice for isolation of
antibacterial compounds.
6.1 RECOMMENDATIONS
This study represents a novel starting point for future studies in which extracts of the selected
mushrooms and more can be used in different fields, such as medicine and pharmaceuticals. It
is, therefore, recommended that further purification, identification and determination of the
mechanism of action of the antibacterial and anti-HIV compounds be carried out to assist in
the optimization of the compounds activity. Isolation and identification of more bioactive
compounds from the local mushrooms is necessary as this will contribute towards the
development of new therapeutic agents against bacterial and HIV infections. It is also
recommended that the compounds identified in this study namely, lucidenic acid M, cavipetin
D, notoginsenoside R2, boviquinone 4, 26-methyl nigranoate, goshonoside, C16 sphinganine,
palmitic amide, stearamide, sorbitan oleate, 11-amino-undecanoic acid and Z-13-oxo-9-

120
octadecenoic acid, be further purified and each compound’s antibacterial effect be tested,
either individually or in combination with other known therapeutic agents to evaluate their
potency as well as assess whether there is enhanced treatment and prevention of drug
resistance. In addition the possibility of compounds acting in synergy needs to be explored.

121
7.0 REFERENCES
Abugria DA, McElhenney WH. (2013). Extraction of total phenolic and flavonoids from
edible wild and cultivated medicinal mushrooms as affected by different solvents. Journal of
Natural Product and Plant Resources, 3: 37 - 42.
Adotey G, Quarcoo A, Holliday JC, Fofie S, Saaka B. (2011). Effect of immunomodulating
and antiviral agent of medicinal mushrooms (Immune Assist 24/7TM) on CD4+ T-lymphocyte
counts of HIV-infected patients. International Journal of Medicinal Mushrooms, 13: 109 -
113.
Afiukwa CA, Oko AO, Afiukwa JN, Ugwu OPC, Ali FU, Ossai EC. (2013). Proximate and
mineral element compositions of five edible wild grown mushroom species in Abakaliki,
southeast Nigeria. Research Journal of Pharmaceutical, Biological and Chemical Sciences,
4(2): 1056-1064.
Ahmed S, Liu H, Ahmad A, Akraw W, Abdelrahman EKN, Ran F, Ou W, Dong S, Cai
Q, Zhang Q, Li X, Hu S, Hu X. (2017). Characterization of anti-bacterial compounds from
the seed coat of Chinese windmill palm tree (Trachycarpus fortunei). Frontiers in
Microbiology, 8: 1894.
Aina DA, Jonathan SG, Olawuyi OJ, Ojelabi DO, Durowoju BM. (2012). Antioxidant,
antimicrobial and phytochemical properties of alcoholic extracts of Cantharellus cibarius – a
Nigerian musroom. NewYork Science Journal, 5: 114 - 120.
Ainsworth GC. (1971). Ainsworth and Bisby’s dictionary of the fungi. 6th ed.
Commonwealth Mycology Institute. Kew, Surrey, England.
Akyuz M, Onganer AN, Erecevit P, Kirbag, S. (2010). Antimicrobial activity of some
edible mushrooms in the Eastern and Southern Anatolia region of Turkey. Gazi University
Journal of Science, 23: 125 - 130.
Albores S1, Mora P, Cerdeiras MP, Fraguas LF. (2014). Screening for lectins from
basidiomycetes and isolation of Punctularia atropurpurascens lectin. Journal of Basic
Microbiology, 54: 89 – 96.
Altemimi A, Lakhssassi N, Baharlouei A, Watson DG, Lightfoot DA. (2017). Review:
Phytochemicals: extraction, isolation and identification of bioactive compounds from plant
extracts. Plants, 6: 1 - 23.
Alves MJ, Ferreira ICFR, Dias J, Teixeira V, Martins A, Pintado M. (2012). A review on
antimicrobial activity of mushroom (Basidiomycetes) extracts and isolated compounds.
Planta Medica, 78: 1707 - 1718.
Anwar F, Przybylski R. (2012). Effect of solvent extraction on total phenolics and
antioxidant activity of extracts from flaxseed (Linum usitatissimum L.). Acta scientiarum
polonorum. Technologia Alimentaria, 11: 293 - 302.
Ardigo W. (2016). Healing with medicinal mushrooms: A practical handbook. Youcanprint
Self-Publishing, Tricase, Italy, 1 - 388.

122
Ashkenazi A, Viard M, Unger L, Blumenthal R, Shai Y. (2012). Sphingopeptides:
dihydrosphingosine-based fusion inhibitors against wild-type and enfuvirtide-resistant HIV-1.
The FASEB Journal, 26: 4628 - 4636.
Askin R, Sasaki M, Goto, M. (2007). Sub- and supercritical fluid extraction of bioactive
compounds from Ganoderma lucidum. Proceedings of International Symposium on EcoTopia
Science (ISETS07), 574 - 578.
Avert Newsletter. (2018). HIV and AIDS in East and Southern Africa regional overview.
Global Information and Education on HIV and AIDS.
Azwanida NN. (2015). A review on the extraction methods used in medicinal plants,
principle, strength and limitation. Medicinal and Aromatic Plants, 4: 196.
Bailey TW. (2013). Antimicrobial assays: Comparison of conventional and fluorescence-
based methods. Journal of Purdue Undergraduate Research, 3: 81.
Barros L, Venturini BA, Baptista P, Estevinho LM, Ferreira ICFR. (2008). Chemical
composition and biological properties of Portuguese wild mushrooms: a comprehensive
study, Journal of Agricicultural and Food Chemistry, 56: 3856 - 3862.
Basnet BB, Liu L, Bao L, Liu H. (2017). Current and future perspective on antimicrobial
and anti-parasitic activities of Ganoderma sp.: An update. Mycology: An International
Journal on Fungal Biology, 8: 111 - 124.
Bessong PO, Obi CL, Andr´eola M-L, Rojas LB, Pouys´egu L, Igumbor E, Meyer JJM,
Quideau S, Litvak S. (2005). Evaluation of selected South African medicinal plants for
inhibitory properties against human immunodeficiency virus type 1 reverse transcriptase and
integrase. Journal of Ethnopharmacology, 99: 83 - 91.
Blossom DB, Namayanja-Kaye G, Nankya-Mutyoba J, Mukasa JB, Bakka H,
Rwambuya S, Windau A, Bajaksouzian S, Walker CJ, Joloba ML, Kityo C,
Mugyenyi P, Whalen CC, Jacobs MR, Salata RA. (2006). Orophryngeal
colonization by Streptococcus pneumoniae among HIV-infected adults in Uganda:
assessing prevalence and antimicrobial susceptibility. International Journal of Infectious
Diseases, 10: 458 - 464.
Boa E. (2004). Wild edible fungi: a global overview of their use and importance to people.
Non-Wood Forest Products, No. 17, FAO, Forestry Department, Rome, Italy.
Boda RH, Wani AH, Zargar MA, Ganie BA, Wani BA, Ganie SA. (2012). Nutritional
values and antioxidant potential of some edible mushrooms of Kashmir valley. Pakistan
Journal of Pharmaceutical Sciences, 25: 623 - 627.
Bovi M, Carrizo ME, Capaldi S, Perduca M, Chiarelli LR, Galliano M, Monaco HL. (2011). Structure of a lectin with antitumoral properties in king bolete (Boletus edulis)
mushrooms. Glycobiology, 21: 1000 - 1009.

123
Branch M. (2001). First field guide to mushrooms of South Africa. Struik Nature Publishers,
Century City, South Africa, 1 - 57.
Buruleanu LC, Radulescu C, Georgescu AA, Danet FA, Olteany RL, Nicolescu CM,
Dulama ID. (2018). Spectroscopy: Statistical characterization of the phytochemical
characteristics of edible mushroom extract. Analytical Letters, 51: 1039 - 1059.
Bylka W, Matlawska J, Pilewski NA. (2004). Natural flavonoids as antimicrobial agents.
Journal of American Nutraceutical Association, 7: 24 - 31.
Camassola M. (2013). Mushrooms – The incredible factory for enzymes and metabolites
productions. Fermentation Technology, 2: 1.
Carbohydrate Lab Report. (2016). Science, chemistry. Topics, Sample Papers and Articles
Online for Free, https://studymoose.com/carbohydrates-lab-report-essay
Carvalho DO, Curto AF, Guido LF. (2015). Determination of phenolic content in different
barley varieties and corresponding malts by liquid chromatography-diode array detection-
electrospray ionization tandem mass spectrometry. Antioxidants, 4: 563 - 576.
Celik GY, Onbasli D, Altinsoy B, Alli H. (2014). In vitro antimicrobial and antioxidant
properties of Ganoderma lucidum extracts grown in Turkey. European Journal of Medicinal
Plants, 4: 709 - 722.
Chang ST. (2008). Overview of mushroom cultivation and utilization as functional foods. In:
P.C.K. Cheung (ed), Mushrooms as Functional Foods, Wiley, Hoboken, New Jersey, USA,
1 - 33.
Chang ST, Buswell JA. (1996). Mushroom nutriceuticals, World Journal of
Microbiology and Biotechnology, 12: 473 - 476.
Chang ST, Miles PG. (1992). Mushroom biology – a new discipline. Mycologist, 6: 64 - 65.
Chang ST, Wasser SP. (2012). The role of culinary-medicinal mushrooms on human welfare
with a pyramid model for human health. International Journal of Medicinal Mushrooms, 14:
95 - 134.
Chaudhary R, Tripathy A. (2015). Isolation and identification of bioactive compounds
from Irpex lacteus wild fleshy fungi. Journal of Pharmaceutical Sciences and Research, 7:
424 - 434.
Chen L, Zhujun AO, Jayappa KD, Kobinger G, Liu SP, Wu G, Wainberg MA, Yao X.
(2013). Characterization of antiviral activity of benzamide derivative AH0109 against HIV-1
infection. Antimicrobial agents and chemotherapy, 57: 3547 - 3554.
Chen XQ, Chen LX, Zhao J, Tang YP, Li SP. (2017). Nortriterpenoids from the fruiting
bodies of the mushroom Ganoderma resinaceum. Molecules, 22: 1073.
Cheung PCK. (2008). Nutritional value and health benefits of mushrooms. In: P.C.K.
Cheung (ed), Mushrooms as Functional Foods, Wiley, Hoboken, New Jersey, USA, 71 - 110.

124
Cheung PCK, (2013). Mini-review on edible mushrooms as source of dietary fiber:
Preparation and health benefits. Food Science and Human Wellness, 2: 162 – 166
Ching CL, Abdullah N, Shuib AS. (2011). Characterization of antihypertensive peptides
from Pleurotus cystidiosus O.K. MILLER (Abalone Mushroom), Proceedings of the 7th
International Conference on Mushroom Biology and Mushroom Products (ICMBMP7), 314 -
323.
Chiroro CK. (2004). Mushroom industry in Zimbabwe. Mushroom Growers’ Handbook
1: Oyster mushroom cultivation, 204 - 211.
Choma IM, Jesionek W. (2015). TLC-Direct bioautography as a high throughput method for
detection of antimicrobials in plants. Chromatography, 2: 225 - 238.
Chomcheon P, Kheawkum B, Sriwiset P, Dulsamphan S, Dulsamphan, C. (2013).
Antibacterial activity of crude extracts from edible mushrooms Pleurotus citrinopileatus and
Tricholoma crassum Berk. Thailand Journal of Pharmaceutical Sciences, 37: 107 - 111
Chowdhury MMH, Kubra K, Ahmed SR. (2015). Screening of antimicrobial, antioxidant
properties and bioactive compounds of some edible mushrooms cultivated in Bangladesh.
Annals of Clinical Microbiology and Antimicrobials, 14: 8.
Chu HS. (2013). Identification of inhibitory compounds in medicinal mushrooms against
pathogenic bacteria and spoilage yeast. Thesis submitted to the Faculty of the Virginia
Polytechnic Institute and State University in partial fulfillment of the requirements for the
degree of MSc in Life Sciences in Food Science and Technology. Blacksburg, VA.
Chu W, Liu D, Guo XH, Yu B, Wu H, Zhang HH, Wu JX, Jiang CL, Kong W, Yu XH. (2015). Antiviral activity of a zymolytic grain based extract on Human Immunodeficiency
Virus Type 1 in vitro. Evidence-Based Complementary and Alternative Medicine, 2015: 9.
Cushnie T, Lamb AJ. (2005). Antimicrobial activity of flavonoids. International Journal of
Antimicrobial Agents, 26: 343 - 356.
Dahake R, Roy S, Patil D, Rajopadhye S, Chowdhary A, Deshmukh RA. (2013). Potential
anti-HIV activity of Jatropha curcas Linn. leaf extracts. Journal of Antivirals and
Antiretrovirals, 5: 160 - 165.
Dailey A, Vuong QV. (2015). Effect of extraction solvents on recovery of bioactive
compounds and antioxidant properties from macadamia (Macadamia tetraphylla) skin waste.
Cogent Food & Agriculture, 1: 1.
De Geyter E. (2012). Toxicity and mode of action of steroid and terpenoid secondary plant
metabolites against economically important pests insects in agriculture. PhD dessertation,
Faculty of Bioscience Engineering, Ghent University, Ghent, Belgium.
De Silva DD, Rapior S, Sudarman E, Stadler M, Xu J, Alias SA, Hyde KD. (2013).
Bioactive metabolites from macrofungi: ethnopharmacology, biological activities and
chemistry. Fungal Diversity, 62:1 – 40.

125
Dhamodharan G, Mirunalini S. (2011). Identification of mushroom lectin and its medicinal
properties. Pharmacologyonline, 1: 170 - 184.
Dickinson C, Lucas J. (1979). The encyclopedia of mushrooms. Orbis Publishing Limited,
London.
Djouossi MG, Tamokou JD, Ngnokam D, Kuiate JR, Tapondjou LA, Harakat D,
Voutquenne-Nazabadioko L. (2015). Antimicrobial and antioxidant flavonoids from the
leaves of Oncoba spinosa Forssk. (Salicaceae). BMC Complementary and Alternative
Medicine, 15: 134.
Doughari JH. (2012). Phytochemicals: extraction methods, basic structures and mode of
action as potential chemotherapeutic agents. In: Rao V (ed), Phytochemical - A global
perspective of their role in nutrition and health, ISBN:978-953-51-02960.
Duffy TJ. Toxic fungi of Western North America, March 2008, http:// www.mykoweb.com.
Dulger B, Gonuz A, Gucin F. (2004). Antimicrobial activity of the macrofungus
Cantharellus cibarius. Pakistan Journal of Biological Sciences, 7: 1535 - 1539.
Durgo K, Koncar M, Drazenka K, Belscak-Cvitanovic A, Franekic J, Jakopovich I,
Jakopovich N, Jakopovich B. (2013). Cytotoxicity of blended versus single medicinal
mushroom extracts on human cancer cell lines: Contribution of polyphenol and
polysaccharide content. International Journal of Medicinal Mushrooms, 15: 435 – 448.
Duru ME, Çayan GT. (2015). Biologically active terpenoids from mushroom origin: A
Review. Records of Natural Products, 9: 456 - 483.
Eghianruwa Q, Odekanyin O, Kuku A. (2011). Physicochemical properties and
acute toxicity studies of a lectin from the saline extract of the fruiting bodies of the shiitake
mushroom, Lentinula edodes (Berk). International Journal of Biochemical and Molecular
Biology, 2: 309 - 317.
El Dine RS, El Halawany AM, Ma CM, Hattori M. (2008). Anti-HIV-1 protease activity of
lanostane triterpenes from the Vietnamese mushroom Ganoderma colossum. Journal of
Natural Products, 71: 1022 - 1026.
Faridur RM, Rezaul KM, Farhadul IM, Rowshanul HM, Tofazzal HM. (2010).
Phytochemical and cytotoxic investigation on oyster mushroom (Pleurotus ostreatus).
International Research Journal of Pharmacy, 1: 342 - 345.
Feng T, Cai JL, Li XM, Zhou ZY, Li ZH, Liu JK. (2016). Chemical constituents and their
bioactivities of mushroom Phellinus rhabarbarinus. Journal of Agricultural and Food
Chemistry, 64: 1945 - 1949.
Finimundy TC, Dillon AJP, Henriques JAP, Ely MR. (2014). A review on general
nutritional compounds and pharmacological properties of the Lentinula edodes mushroom.
Food and Nutrition Sciences, 5: 1095 - 1105.

126
Fischer CL, Drake DR, Dawson DV, Blanchette DR, Brogden KA, Wertza PW. (2012).
Antibacterial activity of sphingoid bases and fatty acids against Gram-positive and Gram-
negative bacteria. Antimicrobial Agents and Chemotherapy, 56: 1157 - 1161.
Florence EJM, Balasundaran M. (2000). Mushroom cultivation using forest litter
and waste wood. Kerala Forest Research Institute Report 195.
Friedman, M. (2016). Mushroom polysaccharides: Chemistry and antiobesity, antidiabetes,
anticancer, and antibiotic properties in cells, rodents, and humans. Foods, 5: 80.
Gan CH, Narul AB, Asmah R. (2013). Antioxidant analysis of different types of
edible mushrooms (Agaricus bisporus and Agaricus brasiliensis). International Food
Research Journal, 20: 1095 - 1102.
Ganeshpurkar A, Rai G, Jain AP. (2010). Review Article: Medicinal mushrooms: Towards
a new horizon. Pharmacognosy Reviews, 4(8): 127 – 135.
Gbolagade JS, Fasidi IO. (2005). Antimicrobial activities of some selected Nigerian
mushrooms. African Journal of Biomedical Research, 8: 83 - 87.
Geethangili M, Rao YK, Tzeng YN. (2013). Development and validation of HPLC-DAD
separation method for determination of bioactive anthrocon medicinal mushroom Antrodia
camphorate. International Journal of Applied Science and Engineering, 11: 195 - 201.
Grant J, Cho J, Allen C. (2006). Self-assembly and physiochemical and rheological
properties of a polysaccharide-surfactant system formed from the cationic biopolymer
chitosan and nonionic sorbitan esters. Langmuir, 22: 4327 - 4335.
Gry J, Andersson C. (2014) Mushrooms traded as food. Vol. 2. Nordic Council of
Ministries.
Gryzenhout M. (2010). Mushrooms of South Africa. Struik Nature Publishers, Cape Town,
South Africa, 1- 144.
Halpern G.M. (2007). Healing mushrooms: Mushrooms - Therapeutic use. Aloha Medicinals
Inc., Square One publishers, New York, USA.
Hamers RL, Sigaloff KCE, Kityo C, Mugyenyi P, Rinke de WTF. (2013). Emerging HIV-
1 drug resistance after roll-out of antiretroviral therapy in Sub - Saharan Africa. Current
Opinions in HIV AIDS, 8: 19 - 26.
Haoses-Gorases L, Goraseb M. (2013) The health ameliorating benefits of
Ganoderma mushroom nutriceuticals of people living with HIV/Aids in Namibia.
Journal of Medicine and Medical Science Research, 2: 63 - 71.
Hassan MAA, Rouf R, Tiralongo E, May TW, Tiralongo J. (2015). Mushroom Lectins:
Specificity, Structure and Bioactivity Relevant to Human Disease. International Journal of
Molecular Sciences, 16(4): 7802–7838.

127
Hawksworth DL. (2001). Mushrooms: The extent of unexplored potential. International
Journal of Medical Mushrooms, 3: 333-337.
Henchion M, Hayes M, Mullen AN, Fenelon M, Tiwari B. (2017). Future protein supply
and demand: Strategies and factors influencing a sustainable equilibrium. Foods, 6(7): 53.
Hsu CL, Yen GC. (2014). Ganoderic acid and lucidenic acid (triterpenoid). Enzymes, 36: 33
- 56.
http://i-base.info/guides/testing/appendix-3-how-hiv-test-work
Hudzicki J. (2009). Kirby-Bauer Disk Diffusion Susceptibility Test Protocol. American
Society for Microbiology.
Idan SA, Al-Marzoqi AH, Hameed IH. (2015). Spectral analysis and anti-bacterial activity
of methanolic fruit extract of Citrullus colocynthis using gas chromatography-mass
spectrometry. African Journal of Biotechnology, 14: 3131 - 3158.
Ingle KP, Deshmukh AG, Padole DA, Dudhare MS, Moharil MP, Khelurkar VC. (2017).
Phytochemicals: Extraction methods, identification and detection of bioactive compounds
from plant extracts. Journal of Pharmacognosy and Phytochemistry, 6: 32 - 36.
Jadaun P, Khopkar P, Kulkarni S. (2016). Repurposing phytochemicals as anti-HIV
agents. Journal of Antivirals and Antiretrovirals, 8: 4.
Kalac P. (2012). Chemical composition and nutritional value of European species of wild
growing mushrooms. In: Andres S, Baumann N. (eds), Mushrooms: Types, properties and
nutrition, Nova Science Publishers, Inc., 129 - 152.
Karmegam N, Jayakumar M, Karuppusamy S. (2012). Synergistic antibacterial activity of
four medicinal plants collected from Dharapuram Taluk of Tiruppur district, South India.
Journal of Plant Sciences, 7: 32 – 38.
Kamra A, Bhatt AB. (2012). Evaluation of antimicrobial and antioxidant activity of
Ganoderma lucidum extracts against human pathogenic bacteria. International Journal of
Pharmacy and Pharmaceutical Sciences, 4(2): 359 - 362.
Khan A, Tania M. (2012). Nutritional and medicinal importance of Pleurotus
mushrooms: An overview. Food Reviews International, 28: 313 - 329.
Khan ZA, Siddiqui MF, Seungkyung P. (2019). Review: Current and emerging methods of
antibiotic susceptibility testing. Diagnostics, 9(2): 49;
https://doi.org/10.3390/diagnostics9020049
Kharsany ABM, Karim QA. (2016). HIV infection and AIDS in Sub-Saharan Africa:
Current status, challenges and opportunities. Open AIDS Journal, 10: 34 – 48.
Klaus AS, Kozarski MS, Niksic MP. (2009). Influence of bioactive compounds extracted
from mushroom Ganoderma lucidum on B and T cells. Proceedings for Natural Sciences,
116: 217 - 223.

128
Klug YA, Ashkenazi A, Viard M, Porat Z, Blumenthal R, Shai Y. (2014). Early and late
HIV-1 membrane fusion events are impaired by sphinganine lapidated peptides that target the
fusion site. Biochemical Journal, 461: 213 - 222.
Koyama Y, Katsuno Y, Miyoshi N, Hayakawa S, Mita T, Muto H, Isemura S,
Aoyagi Y, Isemura M. (2002). Apoptosis induction by lectin isolated from the
mushroom Boletopsis leucomelas in U937 cells. Bioscience, Biotechnology and Biochemistry,
66: 784 - 789.
Kozarski M, Klaus A, Jakovljevic D, Todorovic N, Vunduk J, Petrovic P, Niksic M,
Vrvic MM, van Griensven L. (2015). Antioxidants of edible mushrooms. Molecules, 20(10):
19489 – 19525.
Kues U, Liu Y. (2000). Fruiting body production in basidiomycetes. Applied Microbiology
and Biotechnology, 54: 141 - 152.
Kumar S. (2014). A Review: Alkaloidal drugs. Asian Journal of Pharmaceutical Science and
Technology, 4(3): 107 – 119.
Kumar S, Pandey AK. (2013). Chemistry and biological activities of flavonoids: An
overview. The Scientific World Journal, 6: 176.
Lee SA, Hong SK, Suh CI, Oh MH, Park JH, Choi BW, Park SW, Paik SY. (2010). Anti-
HIV-1 efficacy of extracts from medicinal plants. The Journal of Microbiology, 48 (2): 249 -
252.
Leteane MM, Ngwenya BN, Muzila M, Namushe A, Mwinga J, Musonda R,
Moyo S, Mengestu YB, Abegaz BM, Andrae-Marobela K. (2012). Old plants
newly discovered: Cassia sieberiana D.C. and Cassia abbreviate Oliv. root extracts
inhibit in vitro HIV-1c replication in peripheral blood mononuclear cells (PBMCs) by
different modes of action. Journal of Ethnopharmacology, 141: 48 - 56.
Li B, Lu F, Suo X, Nan H, Li B. (2010). Antioxidant properties of cap and stipe from
Coprinus comatus. Molecules, 15(3): 1473 – 1486.
Li YR, Liu QH, Wang HX, Ng TB. (2008). A novel lectin with potent antitumor, mitogenic
and HIV-1 reverse transcriptase inhibitory activities from the edible mushroom Pleurotus
citrinopileatus. Biochimica et Biopyhsica Acta, 1780: 51 – 57.
Licastro F, Morini MC, Kretz O, Dirheimer G, Creppy EE, Stirpe F. (1993). Mitogenic
activity and immunological properties of bolesatine, a lectin isolated from the mushroom
Boletus satanus Lenz. International Journal of Biochemistry, 25: 789 - 792.
Lindahl JF, Grace F. (2015). The consequences of human actions on risks for infectious
diseases. Infections Ecology and Epidemiology, 5: 10.3402.
Linderquist U, Neidermeyer THJ, Julich W. (2005). Review: The pharmacological
potential of mushrooms. Evidence-Based Complementary and Alternative Medicine, 2: 285 -
299.

129
Luchuo EB, Paschal KA, Ngia G, Njem PK, Yelena S, Nsah B, Ajime TT. (2013).
Review: Malnutrition in sub – Saharan Africa: burden, causes and prospects. Pan African
Medical Journal. 15:120.
Lutsik-Kordovsky MD, Stasyk TV, Stoika RS. (2001). Analysis of cytotoxicity of lectin
and non-lectin proteins from Amanita mushrooms. Experimental Oncology, 23: 43 - 45.
Mabveni ARS. (2004). Mushroom cultivation in Zimbabwe. Mushroom Growers’
Handbook 1: Oyster mushroom cultivation, 212 - 219.
Madhulika U, Harish BN, Parija SC. (2004). Current pattern in antimicrobial susceptibility
of Salmonella typhi isolate in Pondicherry. Indian Journal of Medical Research, 120: 111 -
114.
Magwenzi MT, Gudza-Mugabe M, Mujuru HA, Dangarembizi-Bwakura M, Robertson
V, Aiken AM. (2017). Carriage of antibiotic-resistant Enterobacteriaceae in hospitalised
children in tertiary hospitals in Harare, Zimbabwe. Antimicrobial Resistance & Infection
Control, 6:10.
Masamba KG, Kazombo-Mwale R. (2010). Determination and comparison of nutrient and
mineral contents between cultivated and indigenous edible mushrooms in Central Malawi.
African Journal of Food Science, 4: 176 - 179.
Masoko P. (2007). Characterization of antifungal compounds isolated from Combretum and
Terminalia species (Combretaceae). DPhil Thesis, University of Pretoria, South Africa.
Mataftsi M, Skoura L, Sakellari D. (2010). Review Article: HIV infection and
periodontal diseases: an overview of the post-HAART era. Oral Diseases, 17: 13 - 25.
Mattila P. (2000). Functional properties of edible mushrooms. Nutrition, 16: 694 - 696.
Mighell KS, Henkel TW, Koch RA, Goss A, Aime MC. (2019). New species of Amanita
subgen. Lepidella from Guyana. Fungal Systematics and Evolution, 3: 1 – 12.
Miles PG, Chang ST. (2004). Mushroom: Cultivation, nutritional value, medicinal effect,
and environmental impact. 2nd ed., CRC Press: Boca Raton, FL, USA, 1 - 26.
Moon B, Lo YM. (2013). Conventional and novel applications of edible mushrooms in
today’s food industry. Journal of Food Processing and Preservation, 38: 2146 - 2153.
Moukha S, Ferandon C, Mobio T, Creppy E. (2011). Safety evaluation of Agaricus
subrufescens varieties and their products of therapeutic interest or for disease prevention.
Proceedings of the 7th International Conference on Mushroom Biology and Mushroom
Products (ICMBMP7), 285 - 296.
Narayan CL, Rai RV. (2016). Anti-HIV-1 Activity of Ellagic acid Isolated from Terminalia
paniculata. Free Radicals and Antioxidants, 6: 1.

130
Ndung’u T, Renjifo B, Essex M. (2001). Construction and analysis of an infectious human
immunodeficiency virus type 1 subtype C molecular clone. Journal of Virology, 75: 4964 -
4972.
Ngo TV, Scarlett CJ, Bowyer MC, Ngo PD and Vuong QV. (2017). Impact of different
extraction solvents on bioactive compounds and antioxidant capacity from the root of Salacia
chinensis L. Journal of Food Quality, 2007: 1 - 8.
Nikaido H. (2009). Multidrug Resistance in Bacteria. Annual Review of Biochemistry, 78:
119 – 146.
Nowacka N, Nowak R, Drozd M, Olech M, Los R, Malm A. (2015). Antibacterial,
antiradical potential and phenolic compounds of thirty-one polish mushrooms. PLoS ONE 10:
e0140355.
Nur Syukriah, A. R.,Liza, M. S., Harisun, Y. & Fadzillah, A. A. M., (2014). Effect of
Solvent Extraction on Antioxidant and Antibacterial Activities from Quercusin fectoria
(Manjakani), International Food Research Journal, 21(3): 1031-1037.
OIE Terrestrial Manual. (2012). Guidelines 2 .1. Laboratory methodogies for bacterial
antimicrobial susceptibility testing. 1 - 11.
Okon KO, Shittu AO, Usman H, Adamu N, Balogun ST, Adesina OO. (2013).
Epidermiology and antibiotic susceptibility pattern of methicillin-resistant Staphylococcus
aureus recovered from tertiary hospitals in Northeastern, Nigeria. Journal of Medicine and
Medical Sciences, 4: 214 - 220.
Okonko IO, Donbraye-Emmanuel OB, Ijandipe LA, Ogun AA, Adedeji AO. Udeze AO. (2009). Antibiotics sensitivity and resistance patterns of uropathogens to nitrofurantoin and
nalidixic acid in pregnant women with urinary tract infections in Ibadan, Nigeria. Middle-East
Journal of Scientific Research, 4: 105 - 109.
Orozco LP, Florez-Sampedro L, Zapata W, Mejia AI, Arboleda C, Rugeles MT. (2016).
In vitro anti-HIV activity of the enzymatic extract enriched with laccase produced by the
fungi Ganoderma sp. and Lentinus sp. Vitae, 23(2): 109 - 118.
Owusu-Apenten RK. (2002). Protein analysis: Quantitative eff ects on processing. pp97–122.
New York: Marcel Dekker, Inc.
Padmavathy M, Sumathy R, Manikandan N, Kumuthakalavalli R. (2014). Antimicrobial
activity of mushrooms against skin infection causing pathogens. Research in Biotechnology,
5: 22 - 26.
Pala SA, Wani AH. (2011). Mushrooms: The entities with multifarious medicinal properties.
Journal of Pharmacy Research, 4(12): 4721-4726.
Patel S, Goyal A. (2012) Recent developments in mushrooms as anti-cancer therapeutics: a
review. Biotechnology, 2:1 – 15.

131
Pegler DN, Piearce GD. (1980). The edible mushrooms of Zambia. Kew Bulletin, 35: 475–
491.
Prasad S, Rathore H, Sharma S, Yadav AS. (2015). Medicinal mushrooms as a source of
novel functional food. International Journal of Food Science, Nutrition and Diet, 4: 221 –
225.
Prasad Y, Wesely WEG. (2008). Research article: Antibacterial activity of the bio-
multidrug (Ganoderma lucidum) on multidrug resistant Staphylococcus aureus (MRSA).
Advanced Biotechnology, 6: 9 - 16.
Prost I, Dhondt S, Rothe G, Vicente J, Rodriguez MJ, Kift N, Carbonne F, Griffiths G,
Esquerre-Tugaye MT, Rosahi S, Castresana C, Hamberg M, Fournier J. (2005).
Evaluation of the antimicrobial activities of plant oxylipins supports their involvement in
defense against pathogens. Plant Physiology, 139(4): 1902 - 1913.
Pushpa H, Purushothama KB. (2010). Antimicrobial activity of Lyophyllum decastes
an edible wild mushroom. World Journal of Agricultural Sciences, 6: 506 - 509.
Pyka, A (2014). Review article: Detection progress of selected drugs in TLC. BioMed
Research International, doi: 10.1155/2014/732078.
Quereshi S, Pandey AK, Sandhu SS. (2010). Evaluation of antibacterial activity of different
Ganoderma lucidum extracts. People’s Journal of Scientific Research,. 3: 9 - 13.
Rahi K, Malik D. (2016). Diversity of mushrooms and their metabolites of nutraceutical and
therapeutic significance. Journal of Mycology, doi: 10.1155/2016/7654123.
Rai M, Tidke G, Wasser SP. (2005). Review article: Therapeutic potential of mushrooms.
Natural Product Radiance, 4(4): 246 – 257.
Ramesh C, Pattar MG. (2010). Antimicrobial properties, antioxidant activity and
bioactive compounds from six wild edible mushrooms of western ghats of Karnataka,
India. Pharmacognosy Research, 2: 107 - 112.
Rana R. (2016). Nutritive analysis of wild edible mushroom Cantharellus lutescens Fries,
colleted from North West Himalayan region. International Journal of Innovative Research in
Science, Engineering and Technology, 5(2): 2624 – 2630.
Reagile M. (2011). Screening, isolation and characterization of lectins extracted from
mushrooms indigenous to Southern Africa. MSc Research Dessertation, University of
Limpopo, South Africa.
Rege A, Dahake R, Roy S, Chowdhary A. (2015). Screening of natural products for anti-
HIV potential: An in vitro approach. Juniper Online Journal of Immuno Virology, 1: 1-7.
Reis FS, Barros L, Martins A, Ferreira ICFR. (2012). Chemical composition and
nutritional value of the most widely appreciated cultivated mushrooms: an inter-species
comparative study. Food Chemical Toxicology, 50: 191 - 197.

132
Reis FS, Pereira E, Barros L, Sousa MJ, Martins A, Ferreira ICFR. (2011). Biomolecule
profiles in inedible wild mushrooms with antioxidant value. Molecules, 16: 4328 - 4338.
Ren L. (2014). Anticancer ability of mushroom polysaccharides. A thesis submitted in
fulfilment of the requirements for the degree of Doctor of Philosophy in Food Science,
School of Chemical Sciences, The University of Auckland, New Zealand.
Roupae P, Margetta C, Taylor P, Kause D, Noakee M. (2012). Anti-viral
properties: Mushrooms and health. CSIRO Food and Nutritional Sciences, Australia.
Rowe B, Ward LR, Threlfall EJ. (1997). Multidrug-resistant Salmonella typhi: A
worldwide epidermic. Clinical Infectious Diseases, 24: 106 - 109.
Ruiz-Rodriguez A, Santoyo S, Soler-Rivas C. (2009). Antioxidant propertiesof edible
mushrooms. Functional Plant Science and Biotechnology, Global Science Books
Ryvarden L, Piearce GD, Masuka AJ. (1994). An introduction to the larger fungi
of South Central Africa. Baobab Books. ISBN 0-908311-52-4.
Sala Uddin GM, Sarwar HM, Moniru IM, Asaduzzaman M, Jahan BI, Ruhul AM. (2015) Evaluation of Antimicrobial, Antioxidant and Cytotoxic Property of Pleurotus
ostreatus Mushroom, International Research Journal of Biological Sciences, 4(1): 29-33
Sanchez C. (2010). Mini-Review: Cultivation of Pleurotus ostreatus and other edible
mushrooms. Applied Microbiology and Biotechnology, 85: 1321.
Sangeeth K, Rajesh KR, Indrapriyadharsini R. (2014). Antibiotic resistance pattern of
Escherichia coli causing urinary tract infection with an emphasis on fluoroquinolone
resistance. Global Journal of Medicine and Public Health, 3: 2277 - 2284.
Santhiya M, Jansi, M. (2013). Screening of wild mushroom Amanita species for
occurrence of lectins and their partial purification by RP-HPLC. Middle-East Journal of
Scientific Research, 14: 456 - 460.
Sasidharan S, Chen Y, Saravanan D, Sundram KM, Latha LY. (2011). Extraction,
isolation and characterization of bioactive compounds from plant extracts. African Journal of
Traditional, Complementary and Alternative Medicines, 8: 1 - 10.
Schönfeldt HC, Pretorius B, Hall N. (2013). The impact of animal source food products on
human nutrition and health. South African Journal of Animal Science, 41(3): 394 – 412.
Shah P, Modi HA, Shukla MD, Lahiri SK. (2014). Preliminary phytochemical analysis and
antibacterial activity of Ganoderma lucidum collected from Dang District of Gujarat, India.
International Journal of Current Microbiology and Applied Sciences, 3(3): 246 - 255.
Sharp C. (2011). A pocket guide to mushrooms in Zimbabwe: Some common species.
Directory Publishers, Bulawayo, 1: 1 - 111.

133
Sharp C. (2014). A pocket guide to mushrooms in Zimbabwe: Other common species.
Directory Publishers, Bulawayo, 2: 1 - 111.
Shen HS, Shao S, Chen JC, Zhou T. (2017). Antimicrobials from mushrooms for assuring
food safety. Comprehensive Reviews in Food Science and Food Safety, 16: 316 - 329.
Shen JW, Ruan Y, Ma BJ. (2009). Doterpenoids of macromycetes. Journal of Basic
Microbiology, 49: 242 - 255.
Shikongo LT, Chimwamurombe PM, Lotfy HR, Kandawa-Schulz M. (2013).
Antimicrobial screening of crude extracts from the indigenous Ganoderma lucidum
mushrooms in Namibia. African Journal of Microbiological Research, 7: 4812 - 4816.
Shridhar G, Rajendra N, Murigendra H, Shridevi P, Prasad M, Mujeeb MA, Arun S,
Neeraj D, Vikas S, Suneel D, Vijay K. (2015). Modern diet and its impact on human health.
Journal of Nutrition and Food Sciences, 5: 6.
Singh R, Bhari R. (2014). Current status of microbial lectins in biomedical research. In:
Singh RS, Pandey A, Larroche C. (eds), Advances in biotechnology, IK International
Publishing House Pvt. Ltd., India, 315 - 362.
Singh RS, Jain P, Kaur HP. (2013). Characterization and antimicrobial activity of lectins
from Penicillium sp. Indian Journal of Experimental Biology, 51: 984 – 991.
Soriano J. (2010). Chemical composition and nutritional content of raw poultry meat.
10.1002/9780470504451. In book: Handbook of Poultry Science and Technology, 1: 467 –
489.
Stamets P. (2000). Growing gourmet and medicinal mushrooms. 3rd Ed, Ten Speed Press
Publishers, Berkeley, USA, 1 - 574.
Stamets P. (2005). Mycelium running. 1st Ed, Ten Speed Press Publishers, Berkeley, USA, 1
- 343.
Stanely CN, Ugboma HAA, Ibezim EC, Attama AA. (2013). Prevalence and
antibiotic susceptibility of Staphylococcus aureus and other Staphylococcal infections in
pregnant women attending antenatal clinic in a tertiary hospital in Port Harcourt, Nigeria.
Journal of Infectious Diseases and Therapy, 1: 125.
Sudirman LI. (2010). Partial purification of antimicrobial compounds isolated from mycelia
oftTropical Lentinus cladopus LC4. HAYATI Journal of Biosciences, 17: 63 - 67.
Sudsai T, Leajae S, Dangmanee N, Chatgat W, Chaniad P, Tewtrakul S. (2017).
Antibacterial and anti-HIV-1 integrase properties of isolated compounds from Boesenbergia
kingie. Songklanakarin Journal of Science and Technology, 39: 131 - 135.

134
Sun HD, Qiu SX, Lin LZ, Wang ZY, Lin ZW, Pengsuparp T, Pezzuto JM, Fong HHS,
Cordell GA, Farnsworth NR. (1996). Nigranoic acid, a triterpenoid from Schisandra
sphaerandra that inhibits HIV-1 reverse transcriptase. Journal of Natural Products, 59: 525
- 527.
Sun J, Ng T-B, Wang H and Zhang G. (2014) A novel hemagglutinin with
antiproliferative activity against tumor cells from the hallucinogenic mushroom Boletus
speciosus. BioMed Research International, 2014: 340467.
Taofiq O, Calhelha RC, Heleno SA, Barros L, Martins A, Santos-Buelga C, Queiroz
MJRP, Ferreira ICFR. (2015). The contribution of phenolic acids to the anti-inflammatory
activity of mushrooms: Screening in phenolic extracts, individual parent molecules and
synthesized glucuronated and methylated derivatives. Food Research Interntional, 76: 821 -
827.
Taofiq O, Heleno SA, Calhelha RC, Alves MJ, Barros L, Barreiro MF, Gonzalez-
Paramas AM, Ferreira ICFR. (2016). Development of mushroom-based cosmeceutical
formulations with anti-inflammatory, ant-tyrosinase, antioxidant, and antibacterial
properties. Molecules, 21: 1372.
Tatiya AU, Tapadiya GG, Kotecha S, Surana SJ. (2011). Effect of solvents on total
phenolics, antioxidant and antimicrobial properties of Bridelia retusa Spreng. stem bark.
Indian Journal of Natural Products and Resources, 2: 442 - 447.
Tehrani MHH, Fakhrehoseini E, Nejad MK, Mehregan H, Hakemi-Vala M. (2012).
Search for proteins in the liquid extract of edible mushroom, Agaricus bisporus,
and studying their antibacterial effects. Iranian Journal of Pharmaceutical Research, 11:
145 - 150.
Teklit GA. (2015). Chemical composition and nutritional value of the most widely used
mushrooms cultivated in Mekelle Tigray Ethiopia. Journal of Nutrition and Food Science,
5: 408.
Tietjen I, Gatonye T, Ngwenya BN, Namushe A, Simonambanga S, Muzila M,
Mwimanzi P, Xiao J, Fedida D, Brumme ZL, Brockman MA, Andrae-Marobela K. (2016). Croton megalobotrys Müll Arg. and Vitex doniana (Sweet): Traditional medicinal
plants in a three-step treatment regimen that inhibit in vitro replication of HIV-1. Journal of
Ethnopharmacology, 191: 331 - 340.
Tiwari P, Kumar BKM, Kaur G, Kaur H. (2011). Phytochemical screening and
extraction: A Review. Internationale Pharmaceutica Scientica, 1: 98 - 106.
Tran TH, Baz RE, Cuconati A, Arthos J, Jain P, Khan ZK. (2011). A novel high-
throughput screening assay to identify inhibitors of HIV-1 gp120 protein interaction with
DC-SIGN. Journal of Antivirals and Antiretrovirals, 3: 49 - 54.
University of Pretoria. Anti-HIV activity of the isolated compounds.
https://repository.up.ac.za

135
USAID. (2018). Zimbabwe: Nutrition Profile.
Valgas C, de Souza SM, Smânia EFA, Smânia Jr. A. (2007). Screening methods
todetermine antibacterial activity of natural products. Brazilian Journal of Microbiology, 38:
369 - 380.
Valverde ME, Hernandez-Perez T, Paredes-Lopez O. (2015) Edible mushrooms:
Improving human health and promoting quality life. International Journal of Microbiology,
2015: doi.org/10.1155/2015/376387.
Velisek J, Cejpek K. (2011). Pigments of higher fungi: A review. Czech Journal of Food
Sciences, 29: 87 - 102.
Wainberg MA, Zaharatos J, Brenner BG. (2011). Review article: Development of
antiretroviral drug resistance. The New England Journal of Medicine, 365: 637 - 646.
Waktola G, Temesgen T. (2018). Application of mushroom as food and medicine.
Advances in Biotechnology & Microbiology. 11(4): doi: 10.19080/AIBM.2018.11.555817.
Wang H, Ng TB. (2004). Purification of a novel low-molecular mass laccase with HIV-1
reverse transcriptase inhibitory activity from the mushroom Tricholoma giganteum.
Biochemical and Biophysical Research Communications, 315: 450 - 454.
Wang J, Wang HX, Ng TB. (2007). A peptide with HIV-1 reverse transcriptase inhibitory
activity from the medicinal mushroom Russula paludosa. Peptides, 28: 560 - 565.
Wang Y, Xu B. (2014). Distribution of antioxidant activities and total phenolic contents in
acetone, ethanol, water and hot water extracts from 20 edible mushrooms via sequential
extraction. Austin Journal of Nutrition and Food Sciences, 2: 1009 - 1013.
Wang CR, Zhou R, Ng TB, Wong JH, Qiao WT, Liu F. (2014). First report on isolation
of methyl gallate with antioxidant, anti-HIV-1 and HIV-1 enzyme inhibitory activities from
a mushroom (Pholiota adiposa). Environmental Toxicology and Pharmacology, 37: 626 -
637.
Wani BA, Bodha RH, Wani AH. (2010). Review: Nutritional and medicinal importance of
mushrooms. Journal of Medicinal Plants Research, 4: 2598 - 2604.
WHO Drug Resistance Report. (2012). World Health Organisation, ISBN 978 92 4
1503938.
WHO Fact Sheet. (2018). HIV/AIDS: Key facts.
Wiater A, Paduch R, Pleszczynska M, Prochniak K, Choma A, Kandefer-Szerszen M,
Szczodrak J. (2011). -(13)-D-Glucans from fruiting bodies of selected macromycetes
fungi and the biological activity of their carboxymethylated products. Biotechnology Letters,
33: 787 - 795.

136
Williams PG. (2007). Nutritional composition of red meat. Nutrition & Dietetics. 64(4):
S113–S119.
Xu CJ, Wang YX, Niu BN, Li YB, Wang XM, Lu SL. (2014). Isolation and
characterization of a novel lectin with mitogenic activity from Pleurotus ferulae. Pakistan
Journal of Pharmaceutical Science, 27: 983 - 989.
Yunihastuti E, Widhani A, Karjadi TH. (2014). Review article: Drug hypersensitivity in
human immunodeficiency virus-infected ptient: challenging diagnosis and management. Asia
Pac Allergy, 4: 54 - 67.
Zhang GQ, Sun J, Wang HX, Ng TB. (2009). A novel lectin with antiproliferative activity
from the medicinal mushroom Pholiota adiposa. Acta Biochimica Polonica, 56: 415 - 429.
Zhang JJ, Li Y, Zhou T, Xu DP, Zhang P, Li S, Li HB. (2016). Review: Bioactivities and
health benefits of mushrooms mainly from China. Molecules, 21: 938;
doi:10.3390/molecules21070938
Zhang QW, Lin LG, Ye WC. (2018). Techniques for extraction and isolation of natural
products: a comprehensive review. Chinese Medicine, 13: 20.doi:10.1186/s13020-018-0177-x
Zhang W, Tian G, Geng X, Zhao Y, Ng TB, Zhao L, Wang H. (2014). Isolation and
characterization of a novel lectin from the edible mushroom Stropharia rugosoannulata.
Molecules, 19: 19880 - 19891.
Zhang YJ, Gan RY, Li S, Zhou Y, Li AN, Xu DP, Li HB. (2015). Review: Antioxidant
phytochemicals for the prevention and treatment of chronic diseases. Molecules, 20: 21138 –
21156.
Zheng S, Li C, Ng TB, Wang HX. (2007). A lectin with mitogenic activity from the edible
wild mushroom Boletus edulis, Process Biochem, 42: 1620-1624
Zhiming F, Liu YI, Zhang Q. (2016). A potent pharmacological mushroom: Pleurotus
eryngii. Fungal Genomics and Biology, 6: 1.
Ziarati P, Ghasemynezhad – Shanderman SS. (2015). Mineral contents in Pleurotus
(Oyster mushroom): Association of cooking method. International Journal of Plant, Animal
and Environmental Sciences, 4: 496 – 501.
Ziarati P, Rabizadeh H. (2013). Safety and nutritional comparison of fresh, cooked and
frozen mushroom (Agaricus bisporus). International Journal of Farming and Allied Sciences,
2: 1141 - 1147.

137
8.0 APPENDICES
8.1 PUBLICATONS ARISING FROM THE WORK PRESENTED IN THIS THESIS
8.1.1 Reid T, Kashangura C, Chidewe C, Benhura MA, Mduluza T. (2016). Antibacterial
properties of wild edible and non-edible mushrooms found in Zimbabwe. African
Journal of Microbiology Research, 10(26): 977-984.
8.1.2 Reid T, Munyanyi M, Mduluza T. (2017). Effect of cooking and preservation on
nutritional and phytochemical composition of the mushroom Amanita zambiana.
Food Science and Nutrition, 5(3): 538–544.
8.1.3 Reid T, Kashangura C, Chidewe C, Benhura MA, Stray-Pedersen B, Mduluza T.
Characterization of anti-Salmonella typhi compounds from mushroom extracts in
Zimbabwe. International Journal of Medicinal Mushrooms, Manuscript accepted for
publication.

138
8.2 REAGENTS
0.9% Saline azide
8.766 g NaCl,
2 g NaN3,
0.9895 g MnCl2 and
0.735 CaCl2
Fill to 1 L with distilled water.
Alsever solution
0.05 g glucose,
0.80 g Sodium citrate and
0.42 g NaCl
Fill to 100 ml with distilled water.
Vanillin-sulphuric reagent
1 g vanillin
100 ml ethanol
1.5 ml concentrated sulphuric acid .
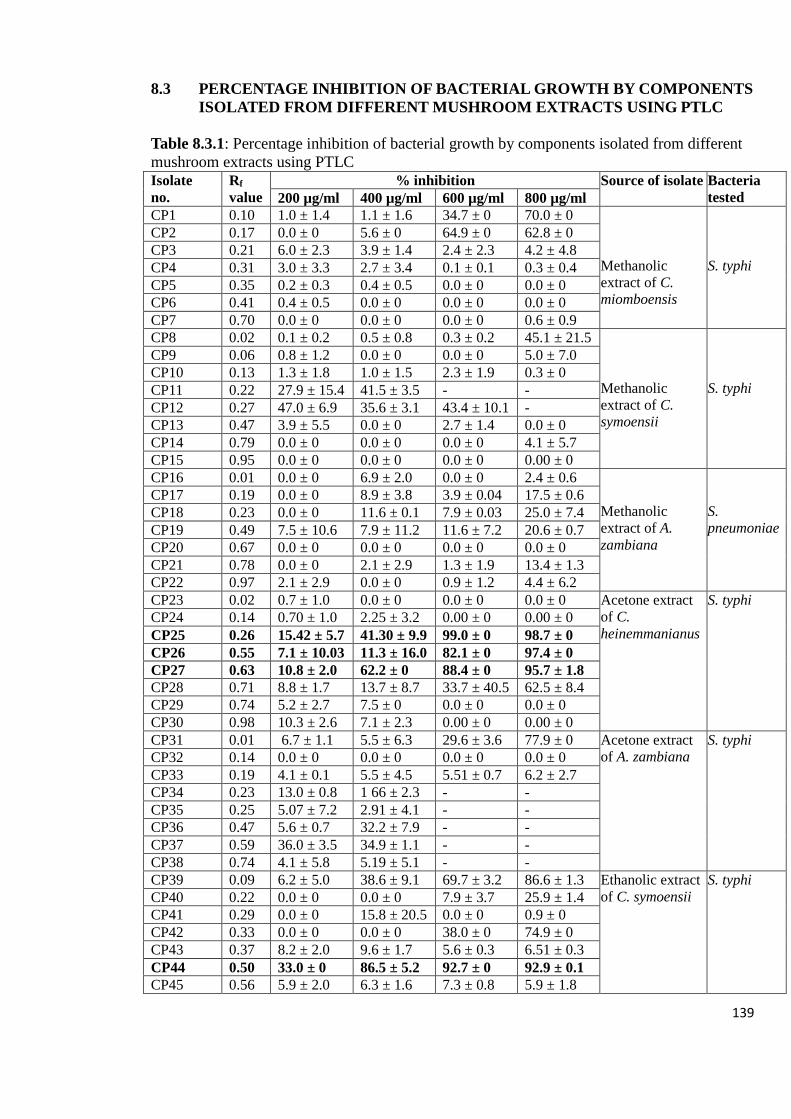
139
8.3 PERCENTAGE INHIBITION OF BACTERIAL GROWTH BY COMPONENTS
ISOLATED FROM DIFFERENT MUSHROOM EXTRACTS USING PTLC
Table 8.3.1: Percentage inhibition of bacterial growth by components isolated from different
mushroom extracts using PTLC
Isolate
no.
Rf
value
% inhibition Source of isolate Bacteria
tested 200 µg/ml 400 µg/ml 600 µg/ml 800 µg/ml
CP1 0.10 1.0 ± 1.4 1.1 ± 1.6 34.7 ± 0 70.0 ± 0
Methanolic
extract of C.
miomboensis
S. typhi
CP2 0.17 0.0 ± 0 5.6 ± 0 64.9 ± 0 62.8 ± 0
CP3 0.21 6.0 ± 2.3 3.9 ± 1.4 2.4 ± 2.3 4.2 ± 4.8
CP4 0.31 3.0 ± 3.3 2.7 ± 3.4 0.1 ± 0.1 0.3 ± 0.4
CP5 0.35 0.2 ± 0.3 0.4 ± 0.5 0.0 ± 0 0.0 ± 0
CP6 0.41 0.4 ± 0.5 0.0 ± 0 0.0 ± 0 0.0 ± 0
CP7 0.70 0.0 ± 0 0.0 ± 0 0.0 ± 0 0.6 ± 0.9
CP8 0.02 0.1 ± 0.2 0.5 ± 0.8 0.3 ± 0.2 45.1 ± 21.5
Methanolic
extract of C.
symoensii
S. typhi
CP9 0.06 0.8 ± 1.2 0.0 ± 0 0.0 ± 0 5.0 ± 7.0
CP10 0.13 1.3 ± 1.8 1.0 ± 1.5 2.3 ± 1.9 0.3 ± 0
CP11 0.22 27.9 ± 15.4 41.5 ± 3.5 - -
CP12 0.27 47.0 ± 6.9 35.6 ± 3.1 43.4 ± 10.1 -
CP13 0.47 3.9 ± 5.5 0.0 ± 0 2.7 ± 1.4 0.0 ± 0
CP14 0.79 0.0 ± 0 0.0 ± 0 0.0 ± 0 4.1 ± 5.7
CP15 0.95 0.0 ± 0 0.0 ± 0 0.0 ± 0 0.00 ± 0
CP16 0.01 0.0 ± 0 6.9 ± 2.0 0.0 ± 0 2.4 ± 0.6
Methanolic
extract of A.
zambiana
S.
pneumoniae
CP17 0.19 0.0 ± 0 8.9 ± 3.8 3.9 ± 0.04 17.5 ± 0.6
CP18 0.23 0.0 ± 0 11.6 ± 0.1 7.9 ± 0.03 25.0 ± 7.4
CP19 0.49 7.5 ± 10.6 7.9 ± 11.2 11.6 ± 7.2 20.6 ± 0.7
CP20 0.67 0.0 ± 0 0.0 ± 0 0.0 ± 0 0.0 ± 0
CP21 0.78 0.0 ± 0 2.1 ± 2.9 1.3 ± 1.9 13.4 ± 1.3
CP22 0.97 2.1 ± 2.9 0.0 ± 0 0.9 ± 1.2 4.4 ± 6.2
CP23 0.02 0.7 ± 1.0 0.0 ± 0 0.0 ± 0 0.0 ± 0 Acetone extract
of C.
heinemmanianus
S. typhi
CP24 0.14 0.70 ± 1.0 2.25 ± 3.2 0.00 ± 0 0.00 ± 0
CP25 0.26 15.42 ± 5.7 41.30 ± 9.9 99.0 ± 0 98.7 ± 0
CP26 0.55 7.1 ± 10.03 11.3 ± 16.0 82.1 ± 0 97.4 ± 0
CP27 0.63 10.8 ± 2.0 62.2 ± 0 88.4 ± 0 95.7 ± 1.8
CP28 0.71 8.8 ± 1.7 13.7 ± 8.7 33.7 ± 40.5 62.5 ± 8.4
CP29 0.74 5.2 ± 2.7 7.5 ± 0 0.0 ± 0 0.0 ± 0
CP30 0.98 10.3 ± 2.6 7.1 ± 2.3 0.00 ± 0 0.00 ± 0
CP31 0.01 6.7 ± 1.1 5.5 ± 6.3 29.6 ± 3.6 77.9 ± 0 Acetone extract
of A. zambiana
S. typhi
CP32 0.14 0.0 ± 0 0.0 ± 0 0.0 ± 0 0.0 ± 0
CP33 0.19 4.1 ± 0.1 5.5 ± 4.5 5.51 ± 0.7 6.2 ± 2.7
CP34 0.23 13.0 ± 0.8 1 66 ± 2.3 - -
CP35 0.25 5.07 ± 7.2 2.91 ± 4.1 - -
CP36 0.47 5.6 ± 0.7 32.2 ± 7.9 - -
CP37 0.59 36.0 ± 3.5 34.9 ± 1.1 - -
CP38 0.74 4.1 ± 5.8 5.19 ± 5.1 - -
CP39 0.09 6.2 ± 5.0 38.6 ± 9.1 69.7 ± 3.2 86.6 ± 1.3 Ethanolic extract
of C. symoensii
S. typhi
CP40 0.22 0.0 ± 0 0.0 ± 0 7.9 ± 3.7 25.9 ± 1.4
CP41 0.29 0.0 ± 0 15.8 ± 20.5 0.0 ± 0 0.9 ± 0
CP42 0.33 0.0 ± 0 0.0 ± 0 38.0 ± 0 74.9 ± 0
CP43 0.37 8.2 ± 2.0 9.6 ± 1.7 5.6 ± 0.3 6.51 ± 0.3
CP44 0.50 33.0 ± 0 86.5 ± 5.2 92.7 ± 0 92.9 ± 0.1
CP45 0.56 5.9 ± 2.0 6.3 ± 1.6 7.3 ± 0.8 5.9 ± 1.8
140
CP46 0.78 17.8 ± 0.9 2.1 ± 0.7 0.00 ± 0 0.0 ± 0
CP47 0.97 14.6 ± 9.1 6.3 ± 8.9 63.9 ± 47.7 67.7 ± 38.0
CP48 0.11 21.2 ± 1.4 21.7 ± 0.8 22.9 ± 0.3 24.3 ± 0.7 Acetone extract
of T. strumosa
S. aureus
CP49 0.14 10.9 ± 0.1 17.3 ± 0.4 22.4 ± 0.3 24.8 ± 0.01
CP50 0.20 11.4 ± 0.0 13.5 ± 0.1 57.6 ± 0.1 87.5 ± 0.3
CP51 0.24 15.7 ± 0.2 22.4 ± 0.5 22.6 ± 0.3 24.2 ± 0.1
CP52 0.27 0.0 ± 0 1.8 ± 0.1 7.5 ± 0.2 6.2 ± 6.9
CP53 0.33 1.4 ± 0.01 9.1 ± 0.4 10.3 ± 0.1 10.6 ± 0.3
CP54 0.46 0.0 ± 0 0.0 ± 0 3.6 ± 0.02 6.8 ± 0.2
CP55 0.73 0.0 ± 0 4.6 ± 0.1 5.6 ± 0.2 17.7 ± 0.3
CP56 0.01 0.00 ± 0 0.00 ± 0 0.00 ± 0 0.00 ± 0 Methanolic
extract of
Amanita sp
S.
pneumoniae CP57 0.09 0.0 ± 0 4.1 ± 0.01 3.0 ± 0.04 0.00 ± 0
CP58 0.22 0.0 ± 0 4.0 ± 0.04 4.6 ± 0.1 4.0 ± 0
CP59 0.25 0.0 ± 0 0.0 ± 0 0.00 ± 0 0.00 ± 0
CP60 0.44 0.0 ± 0 0.0 ± 0 0.00 ± 0 6.8 ± 0.01
CP61 0.73 0.0 ± 0 0.0 ± 0 10.7 ± 0.4 7.6 ± 0
CP62 0.98 0.0 ± 0 0.0 ± 0 16.9 ± 0.04 36.4 ± 0.2
CP63 0.01 9.7 ± 0.8 1.6 ± 2.2 16.5 ± 6.0 6.4 ± 4.6 Acetone extract
of L. kabansus
S. typhi
CP64 0.10 27.0 ± 3.7 24.1 ± 3.4 23.3 ± 14.6 25.2 ± 16.2
CP65 0.24 39.4 ± 4.1 52.5 ± 3.3 - 82.0 ± 2.5
CP66 0.28 25.1 ± 7.1 27.8 ± 4.6 14.8 ± 2.6 15.7 ± 2.7
CP67 0.31 0.0 ± 0 39.1 ± 10.2 - -
CP68 0.49 0.0 ± 0 2.0 ± 2.9 - -
CP69 0.56 21.2 ± 1.2 26.1 ± 0.2 29.2 ± 7.7 28.3 ± 8.4
CP70 0.76 29.3 ± 29.6 98.7 ± 0.3 99.7 ± 0.01 99.7 ± 0
CP71 0.98 2.4 ± 3.3 18.0 ± 25.4 50.1 ± 24.8 79.7 ± 1.6
CP72 0.03 1.1 ± 0.3 10.1 ± 14.3 1.2 ± 1.7 5.0 ± 7.1 Ethanolic extract
of G. lucidum
S. typhi
CP73 0.20 0.0 ± 0 2.3 ± 3.0 78.3 ± 28.7 99.8 ± 0.2
CP74 0.26 0.0 ± 0 0.0 ± 0 0.0 ± 0 68.4 ± 15.3
CP75 0.29 57.1 ± 2.8 46.9 ± 4.5 - -
CP76 0.38 13.9 ± 18.5 10.0 ± 14.1 95.5 ± 5.3 98.3 ± 1.4
CP77 0.43 2.4 ± 2.1 41.1 ± 4.4 97.4 ± 1.4 94.9 ± 3.7
CP78 0.56 0.0 ± 0 0.6 ± 0.9 97.5 ± 2.4 97.7 ± 0
CP79 0.79 4.3 ± 1.2 1.51 ± 2.1 0.00 ± 0 9.3 ± 13.2
CP80 0.99 38.1 ± 6.6 54.7 ± 14.5 75.7 ± 1.3 -
CP81 0.01 0.0 ± 0 0.0 ± 0 0.0 ± 0 0.0 ± 0 Methanolic
extract of L.
kabansus
S.
pneumoniae CP82 0.15 0.0 ± 0 0.0 ± 0 29.6 ± 0.1 45.0 ± 0.1
CP83 0.25 0.0 ± 0 0.0 ± 0 0.00 ± 0 0.0 ± 0
CP84 0.29 0.0 ± 0 37.7 ± 0.03 18.1 ± 0.1 27.8 ± 0.1
CP85 0.33 0.0 ± 0 39.9 ± 1.1 35.6 ± 0.02 39.6 ± 0.7
CP86 0.46 0.0 ± 0 0.0 ± 0 0.00 ± 0 0.0 ± 0
CP87 0.53 0.0 ± 0 0.0 ± 0 0.00 ± 0 17.5 ± 0.03
CP88 0.73 0.0 ± 0 0.0 ± 0 11.6 ± 0.03 28.3 ± 0.4
CP89 0.90 0.0 ± 0 0.0 ± 0 0.00 ± 0 0.00 ± 0
CP90 0.99 0.0 ± 0 0.0 ± 0 1.3 ± 1.8 19.5 ± 1.0
CP91 0.04 0.0 ± 0 0.0 ± 0 0.00 ± 0 9.3 ± 13.2 Acetone extract
of C. symoensii
S. typhi
CP92 0.07 0.0 ± 0 0.0 ± 0 38.0 ± 15.9 98.4 ± 0.2
CP93 0.28 0.0 ± 0 4.9 ± 6.9 7.2 ± 10.2 8.3 ± 11.7
CP94 0.55 16.1 ± 22.7 98.2 ± 0 99.7 ± 0 99.8 ± 0.01
CP95 0.59 0.8 ± 1.1 0.0 ± 0 0.0 ± 0 0.0 ± 0
CP96 0.88 0.6 ± 0.9 7.4 ± 10.5 10.5 ± 14.8 16.5 ± 19.7

141
CP97 0.96 7.2 ± 10.2 6.7 ± 9.4 4.1 ± 5.8 5.0 ± 4.0
CP98 0.03 4.3 ± 6.0 93.3 ± 5.6 95.7 ± 2.2 96.9 ± 0 Cold water
extract of C.
miomboensis
S. typhi
CP99 0.08 0.0 ± 0 2.2 ± 3.2 6.6 ± 9.3 7.7 ± 11.0
CP100 0.13 7.8 ± 0.6 0.0 ± 0 0.0 ± 0 0.0 ± 0
CP101 0.20 0.0 ± 0 15.5 ± 4.5 64.9 ± 0.4 -
CP102 0.31 4.4 ± 4.5 0.0 ± 0 0.0 ± 0 0.6 ± 0.9
CP103 0.98 15.9 ± 7.4 35.7 ± 6.4 48.2 ± 13.9 79.0 ± 0
CP104 0.02 0.0 ± 0 0.0 ± 0 0.0 ± 0 44.6 ± 63.1 Ethanolic extract
of C.
miomboensis
S. typhi
CP105 0.15 8.7 ± 4.2 7.2 ± 1.2 0.0 ± 0 0.0 ± 0
CP106 0.20 0.0 ± 0 0.0 ± 0 0.0 ± 0 0.0 ± 0
CP107 0.26 0.0 ± 0 0.0 ± 0 0.0 ± 0 0.0 ± 0
CP108 0.47 0.0 ± 0 0.0 ± 0 0.0 ± 0 8.3 ± 11.7
CP109 0.54 0.0 ± 0 0.0 ± 0 0.0 ± 0 1.4 ± 2.0
CP110 0.60 0.0 ± 0 0.0 ± 0 0.0 ± 0 0.8 ± 1.1
CP111 0.67 53.2 ± 4.1 76.7 ± 1.0 76.8 ± 2.2 -
CP112 0.75 0.0 ± 0 0.0 ± 0 3.5 ± 3.4 11.9 ± 13.6
CP113 0.86 0.0 ± 0 1.2 ± 1.7 18.9 ± 2.5 15.0 ± 6.8
CP114 0.96 0.0 ± 0 9.2 ± 10.8 11.0 ± 0.5 8.4 ± 11.9
CP115 0.02 0.0 ± 0 0.0 ± 0 0.0 ± 0 0.0 ± 0 Acetone extract
of Amanita sp
S. typhi CP116 0.09 0.0 ± 0 0.0 ± 0 0.0 ± 0 0.0 ± 0
CP117 0.15 0.0 ± 0 0.0 ± 0 26.3 ± 9.0 26.7 ± 13.4
CP118 0.35 1.1 ± 1.6 0.0 ± 0 17.7 ± 3.7 15.0 ± 6.6
CP119 0.49 0.0 ± 0 0.0 ± 0 0.0 ± 0 0.0 ± 0
CP120 0.62 0.0 ± 0 2.9 ± 4.03 7.5 ± 5.06 12.9 ± 2.2
CP121 0.82 23.3 ± 32.9 48.5 ± 68.6 48.8 ± 69.0 49.0 ± 69.3
CP122 0.96 0.0 ± 0 0.0 ± 0 0.00 ± 0 1.3 ± 1.9
CP123 0.02 0.0 ± 0 0.0 ± 0 1.64 ± 2.3 5.4 ± 7.6 Ethanolic extract
of Amanita sp
S. typhi
CP124 0.08 1.7 ± 2.4 12.2 ± 0.9 - -
CP125 0.17 0.0 ± 0 0.0 ± 0 8.9 ± 12.5 8.3 ± 11.7
CP126 0.36 1.4 ± 2.0 0.0 ± 0 0.0 ± 0 0.0 ± 0
CP127 0.55 0.0 ± 0 0.7 ± 1.1 2.6 ± 3.6 10.3 ± 14.6
CP128 0.60 0.0 ± 0 5.2 ± 7.3 3.0 ± 4.3 0.0 ± 0
CP129 0.72 0.0 ± 0 8.0 ± 5.1 33.8 ± 11.2 61.2 ± 3.5
CP130 0.86 0.0 ± 0 4.2 ± 3.6 0.5 ± 0.7 12.7 ± 0.5
CP131 0.98 0.0 ± 0 3.6 ± 5.1 8.1 ± 11.5 29.0 ± 12.7
Ampicillin
(50 µg/ml)
96.9 ± 0.4 S. typhi
91.3 ± 0.5 S. aureus
92.4 ± 0.7 S. pneumoniae
The negative control values were used to calculate the percentage inhibition of bacterial
growth by each extract.

142
8.4 LC-MS CHROMATOGRAMS OUTPUT OF ISOLATED COMPONENTS OF
MUSHROOM SAMPLES
Figure 8.4.1: LC-MS output of chromatograms of isolated components of mushroom
samples that exhibited high anti-bacterial activity analyzed in positive mode with a column.
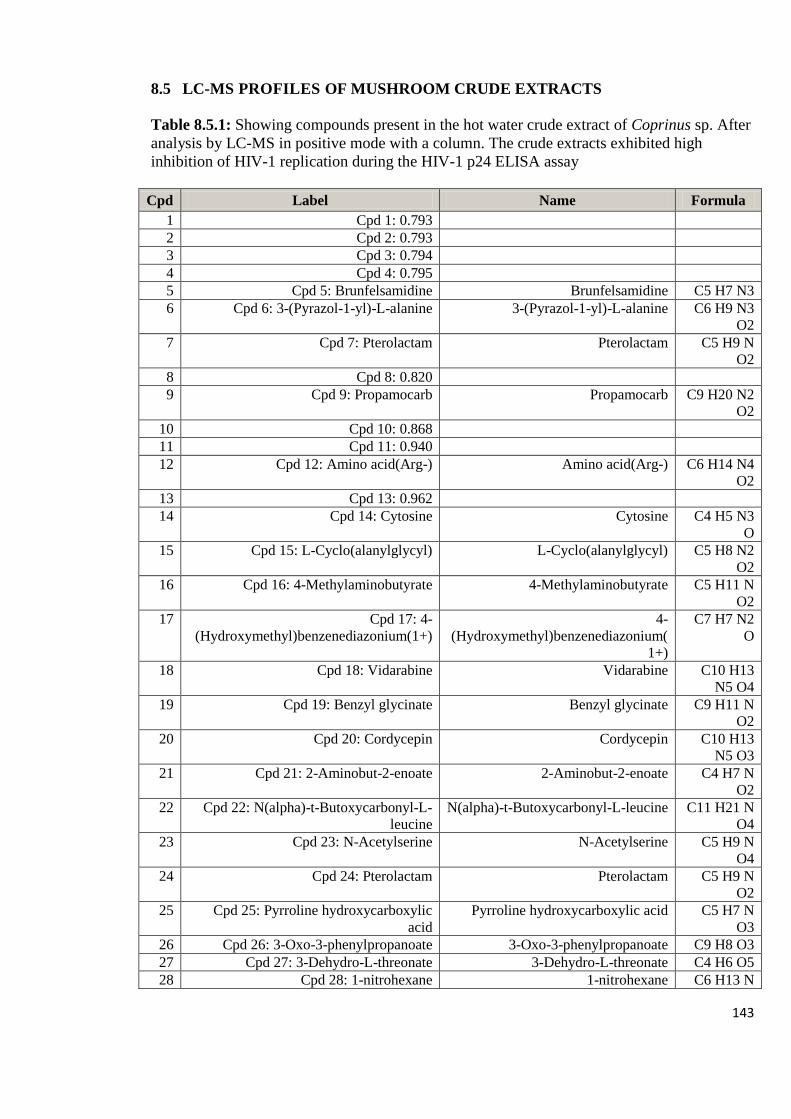
143
8.5 LC-MS PROFILES OF MUSHROOM CRUDE EXTRACTS
Table 8.5.1: Showing compounds present in the hot water crude extract of Coprinus sp. After
analysis by LC-MS in positive mode with a column. The crude extracts exhibited high
inhibition of HIV-1 replication during the HIV-1 p24 ELISA assay
Cpd Label Name Formula
1 Cpd 1: 0.793
2 Cpd 2: 0.793
3 Cpd 3: 0.794
4 Cpd 4: 0.795
5 Cpd 5: Brunfelsamidine Brunfelsamidine C5 H7 N3
6 Cpd 6: 3-(Pyrazol-1-yl)-L-alanine 3-(Pyrazol-1-yl)-L-alanine C6 H9 N3
O2
7 Cpd 7: Pterolactam Pterolactam C5 H9 N
O2
8 Cpd 8: 0.820
9 Cpd 9: Propamocarb Propamocarb C9 H20 N2
O2
10 Cpd 10: 0.868
11 Cpd 11: 0.940
12 Cpd 12: Amino acid(Arg-) Amino acid(Arg-) C6 H14 N4
O2
13 Cpd 13: 0.962
14 Cpd 14: Cytosine Cytosine C4 H5 N3
O
15 Cpd 15: L-Cyclo(alanylglycyl) L-Cyclo(alanylglycyl) C5 H8 N2
O2
16 Cpd 16: 4-Methylaminobutyrate 4-Methylaminobutyrate C5 H11 N
O2
17 Cpd 17: 4-
(Hydroxymethyl)benzenediazonium(1+)
4-
(Hydroxymethyl)benzenediazonium(
1+)
C7 H7 N2
O
18 Cpd 18: Vidarabine Vidarabine C10 H13
N5 O4
19 Cpd 19: Benzyl glycinate Benzyl glycinate C9 H11 N
O2
20 Cpd 20: Cordycepin Cordycepin C10 H13
N5 O3
21 Cpd 21: 2-Aminobut-2-enoate 2-Aminobut-2-enoate C4 H7 N
O2
22 Cpd 22: N(alpha)-t-Butoxycarbonyl-L-
leucine
N(alpha)-t-Butoxycarbonyl-L-leucine C11 H21 N
O4
23 Cpd 23: N-Acetylserine N-Acetylserine C5 H9 N
O4
24 Cpd 24: Pterolactam Pterolactam C5 H9 N
O2
25 Cpd 25: Pyrroline hydroxycarboxylic
acid
Pyrroline hydroxycarboxylic acid C5 H7 N
O3
26 Cpd 26: 3-Oxo-3-phenylpropanoate 3-Oxo-3-phenylpropanoate C9 H8 O3
27 Cpd 27: 3-Dehydro-L-threonate 3-Dehydro-L-threonate C4 H6 O5
28 Cpd 28: 1-nitrohexane 1-nitrohexane C6 H13 N

144
O2
29 Cpd 29: Isonicotinic acid Isonicotinic acid C6 H5 N
O2
30 Cpd 30: Pirbuterol Pirbuterol C12 H20
N2 O3
31 Cpd 31: 1,8-Diazacyclotetradecane-2,9-
dione
1,8-Diazacyclotetradecane-2,9-dione C12 H22
N2 O2
32 Cpd 32: 3'-O-Methyladenosine 3'-O-Methyladenosine C11 H15
N5 O4
33 Cpd 33: 1.031
34 Cpd 34: 1.043
35 Cpd 35: Lysyl-Tyrosine Lysyl-Tyrosine C15 H23
N3 O4
36 Cpd 36: Arginyl-Phenylalanine Arginyl-Phenylalanine C15 H23
N5 O3
37 Cpd 37: Pyruvophenone Pyruvophenone C9 H8 O2
38 Cpd 38: 1.115
39 Cpd 39: Alanyl-Isoleucine Alanyl-Isoleucine C9 H18 N2
O3
40 Cpd 40: Benzaldehyde Benzaldehyde C7 H6 O
41 Cpd 41: 1.117
42 Cpd 42: Pantothenic Acid Pantothenic Acid C9 H17 N
O5
43 Cpd 43: Quinacetol Quinacetol C11 H9 N
O2
44 Cpd 44: 5-Phenyl-1,3-oxazinane-2,4-
dione
5-Phenyl-1,3-oxazinane-2,4-dione C10 H9 N
O3
45 Cpd 45: N(alpha)-t-Butoxycarbonyl-L-
leucine
N(alpha)-t-Butoxycarbonyl-L-leucine C11 H21 N
O4
46 Cpd 46: 1.118
47 Cpd 47: 1.118
48 Cpd 48: Bethanidine Bethanidine C10 H15
N3
49 Cpd 49: L-isoleucyl-L-proline L-isoleucyl-L-proline C11 H20
N2 O3
50 Cpd 50: Pterolactam Pterolactam C5 H9 N
O2
51 Cpd 51: 2'-Aminoacetophenone 2'-Aminoacetophenone C8 H9 N O
52 Cpd 52: Benzenepropanenitrile Benzenepropanenitrile C9 H9 N
53 Cpd 53: 2(1H)-Quinolinone 2(1H)-Quinolinone C9 H7 N O
54 Cpd 54: quinaldine quinaldine C10 H9 N
55 Cpd 55: 1.161
56 Cpd 56: 1.166
57 Cpd 57: 1.172
58 Cpd 58: Pinacidil Pinacidil C13 H19
N5
59 Cpd 59: L-1,2,3,4-Tetrahydro-beta-
carboline-3-carboxylic acid
L-1,2,3,4-Tetrahydro-beta-carboline-
3-carboxylic acid
C12 H12
N2 O2
60 Cpd 60: Anisomycin Anisomycin C14 H19 N
O4
61 Cpd 61: 1.182
62 Cpd 62: 1.182
63 Cpd 63: Marimastat Marimastat C15 H29

145
N3 O5
64 Cpd 64: 1.211
65 Cpd 65: 1.219
66 Cpd 66: D-Lombricine D-Lombricine C6 H15 N4
O6 P
67 Cpd 67: 1.225
68 Cpd 68: 1.225
69 Cpd 69: 1.226
70 Cpd 70: 1.239
71 Cpd 71: 1,3,8-Trihydroxy-4-methyl-2,7-
diprenylxanthone
1,3,8-Trihydroxy-4-methyl-2,7-
diprenylxanthone
C24 H26
O5
72 Cpd 72: 1.267
73 Cpd 73: 1.286
74 Cpd 74: 1.289
75 Cpd 75: 1.313
76 Cpd 76: 1.319
77 Cpd 77: 1.319
78 Cpd 78: 1.330
79 Cpd 79: Coutaric acid Coutaric acid C18 H27
N3 O4
80 Cpd 80: Ximelagatran Ximelagatran C24 H35
N5 O5
81 Cpd 81: 1.433
82 Cpd 82: 1.511
83 Cpd 83: Leucyl-leucyl-norleucine Leucyl-leucyl-norleucine C18 H35
N3 O4
84 Cpd 84: 1.654
85 Cpd 85: 1.655
86 Cpd 86: 1.791
87 Cpd 87: Tebuconazole Tebuconazole C16 H22 Cl
N3 O
88 Cpd 88: 2.064
89 Cpd 89: 2.088
90 Cpd 90: 2.104
91 Cpd 91: 2.108
92 Cpd 92: Kanzonol V Kanzonol V C24 H24
O4
93 Cpd 93: Phenylacetonitrile Phenylacetonitrile C8 H7 N
94 Cpd 94: 2(1H)-Quinolinone 2(1H)-Quinolinone C9 H7 N O
95 Cpd 95: Lupinate Lupinate C13 H18
N6 O3
96 Cpd 96: 5(S),14(R)-Lipoxin B4 5(S),14(R)-Lipoxin B4 C20 H32
O5
97 Cpd 97: Isocolumbin Isocolumbin C20 H22
O6
98 Cpd 98: 5(S),14(R)-Lipoxin B4 5(S),14(R)-Lipoxin B4 C20 H32
O5
99 Cpd 99: 5(S),14(R)-Lipoxin B4 5(S),14(R)-Lipoxin B4 C20 H32
O5
100 Cpd 100: 8.445
101 Cpd 101: 5(S),14(R)-Lipoxin B4 5(S),14(R)-Lipoxin B4 C20 H32
O5
102 Cpd 102: 8.747
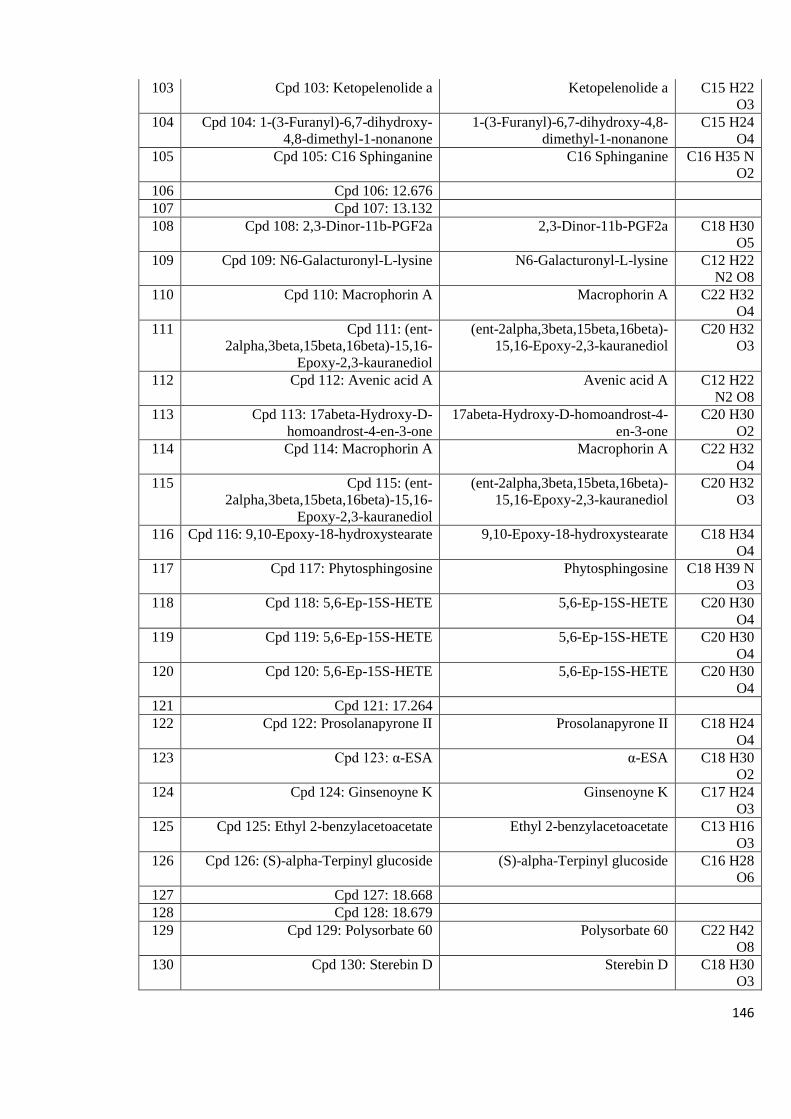
146
103 Cpd 103: Ketopelenolide a Ketopelenolide a C15 H22
O3
104 Cpd 104: 1-(3-Furanyl)-6,7-dihydroxy-
4,8-dimethyl-1-nonanone
1-(3-Furanyl)-6,7-dihydroxy-4,8-
dimethyl-1-nonanone
C15 H24
O4
105 Cpd 105: C16 Sphinganine C16 Sphinganine C16 H35 N
O2
106 Cpd 106: 12.676
107 Cpd 107: 13.132
108 Cpd 108: 2,3-Dinor-11b-PGF2a 2,3-Dinor-11b-PGF2a C18 H30
O5
109 Cpd 109: N6-Galacturonyl-L-lysine N6-Galacturonyl-L-lysine C12 H22
N2 O8
110 Cpd 110: Macrophorin A Macrophorin A C22 H32
O4
111 Cpd 111: (ent-
2alpha,3beta,15beta,16beta)-15,16-
Epoxy-2,3-kauranediol
(ent-2alpha,3beta,15beta,16beta)-
15,16-Epoxy-2,3-kauranediol
C20 H32
O3
112 Cpd 112: Avenic acid A Avenic acid A C12 H22
N2 O8
113 Cpd 113: 17abeta-Hydroxy-D-
homoandrost-4-en-3-one
17abeta-Hydroxy-D-homoandrost-4-
en-3-one
C20 H30
O2
114 Cpd 114: Macrophorin A Macrophorin A C22 H32
O4
115 Cpd 115: (ent-
2alpha,3beta,15beta,16beta)-15,16-
Epoxy-2,3-kauranediol
(ent-2alpha,3beta,15beta,16beta)-
15,16-Epoxy-2,3-kauranediol
C20 H32
O3
116 Cpd 116: 9,10-Epoxy-18-hydroxystearate 9,10-Epoxy-18-hydroxystearate C18 H34
O4
117 Cpd 117: Phytosphingosine Phytosphingosine C18 H39 N
O3
118 Cpd 118: 5,6-Ep-15S-HETE 5,6-Ep-15S-HETE C20 H30
O4
119 Cpd 119: 5,6-Ep-15S-HETE 5,6-Ep-15S-HETE C20 H30
O4
120 Cpd 120: 5,6-Ep-15S-HETE 5,6-Ep-15S-HETE C20 H30
O4
121 Cpd 121: 17.264
122 Cpd 122: Prosolanapyrone II Prosolanapyrone II C18 H24
O4
123 Cpd 123: α-ESA α-ESA C18 H30
O2
124 Cpd 124: Ginsenoyne K Ginsenoyne K C17 H24
O3
125 Cpd 125: Ethyl 2-benzylacetoacetate Ethyl 2-benzylacetoacetate C13 H16
O3
126 Cpd 126: (S)-alpha-Terpinyl glucoside (S)-alpha-Terpinyl glucoside C16 H28
O6
127 Cpd 127: 18.668
128 Cpd 128: 18.679
129 Cpd 129: Polysorbate 60 Polysorbate 60 C22 H42
O8
130 Cpd 130: Sterebin D Sterebin D C18 H30
O3
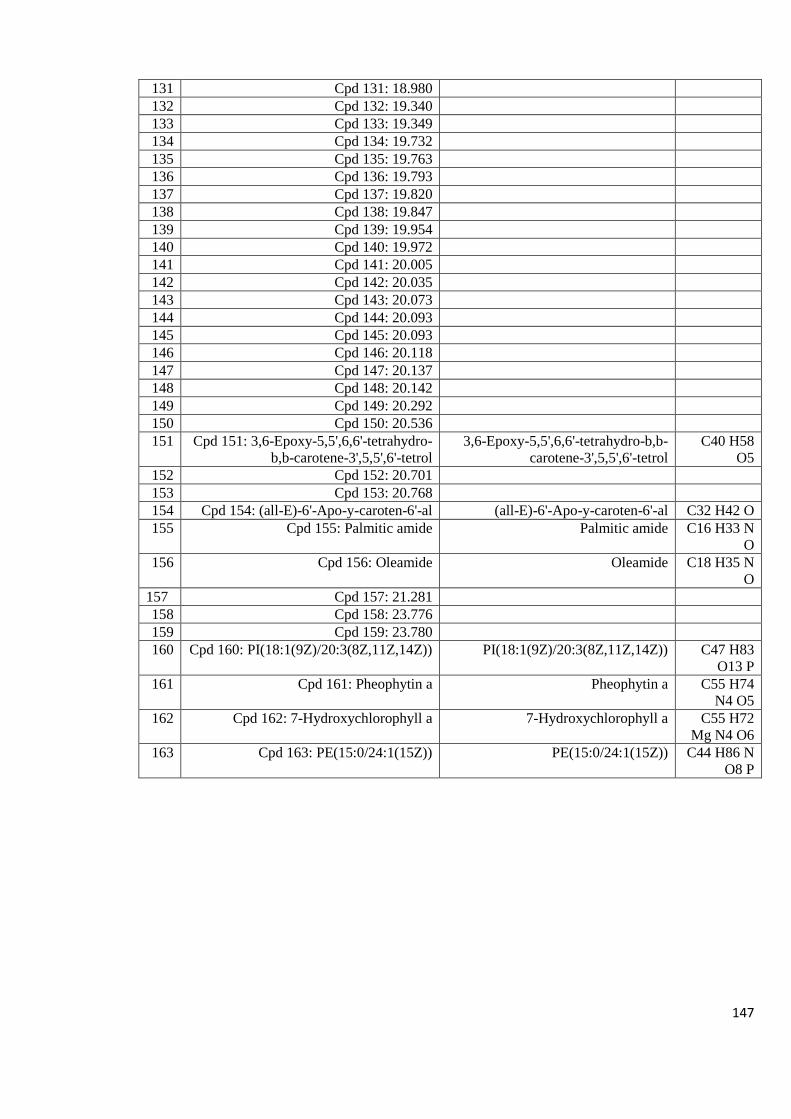
147
131 Cpd 131: 18.980
132 Cpd 132: 19.340
133 Cpd 133: 19.349
134 Cpd 134: 19.732
135 Cpd 135: 19.763
136 Cpd 136: 19.793
137 Cpd 137: 19.820
138 Cpd 138: 19.847
139 Cpd 139: 19.954
140 Cpd 140: 19.972
141 Cpd 141: 20.005
142 Cpd 142: 20.035
143 Cpd 143: 20.073
144 Cpd 144: 20.093
145 Cpd 145: 20.093
146 Cpd 146: 20.118
147 Cpd 147: 20.137
148 Cpd 148: 20.142
149 Cpd 149: 20.292
150 Cpd 150: 20.536
151 Cpd 151: 3,6-Epoxy-5,5',6,6'-tetrahydro-
b,b-carotene-3',5,5',6'-tetrol
3,6-Epoxy-5,5',6,6'-tetrahydro-b,b-
carotene-3',5,5',6'-tetrol
C40 H58
O5
152 Cpd 152: 20.701
153 Cpd 153: 20.768
154 Cpd 154: (all-E)-6'-Apo-y-caroten-6'-al (all-E)-6'-Apo-y-caroten-6'-al C32 H42 O
155 Cpd 155: Palmitic amide Palmitic amide C16 H33 N
O
156 Cpd 156: Oleamide Oleamide C18 H35 N
O
157 Cpd 157: 21.281
158 Cpd 158: 23.776
159 Cpd 159: 23.780
160 Cpd 160: PI(18:1(9Z)/20:3(8Z,11Z,14Z)) PI(18:1(9Z)/20:3(8Z,11Z,14Z)) C47 H83
O13 P
161 Cpd 161: Pheophytin a Pheophytin a C55 H74
N4 O5
162 Cpd 162: 7-Hydroxychlorophyll a 7-Hydroxychlorophyll a C55 H72
Mg N4 O6
163 Cpd 163: PE(15:0/24:1(15Z)) PE(15:0/24:1(15Z)) C44 H86 N
O8 P
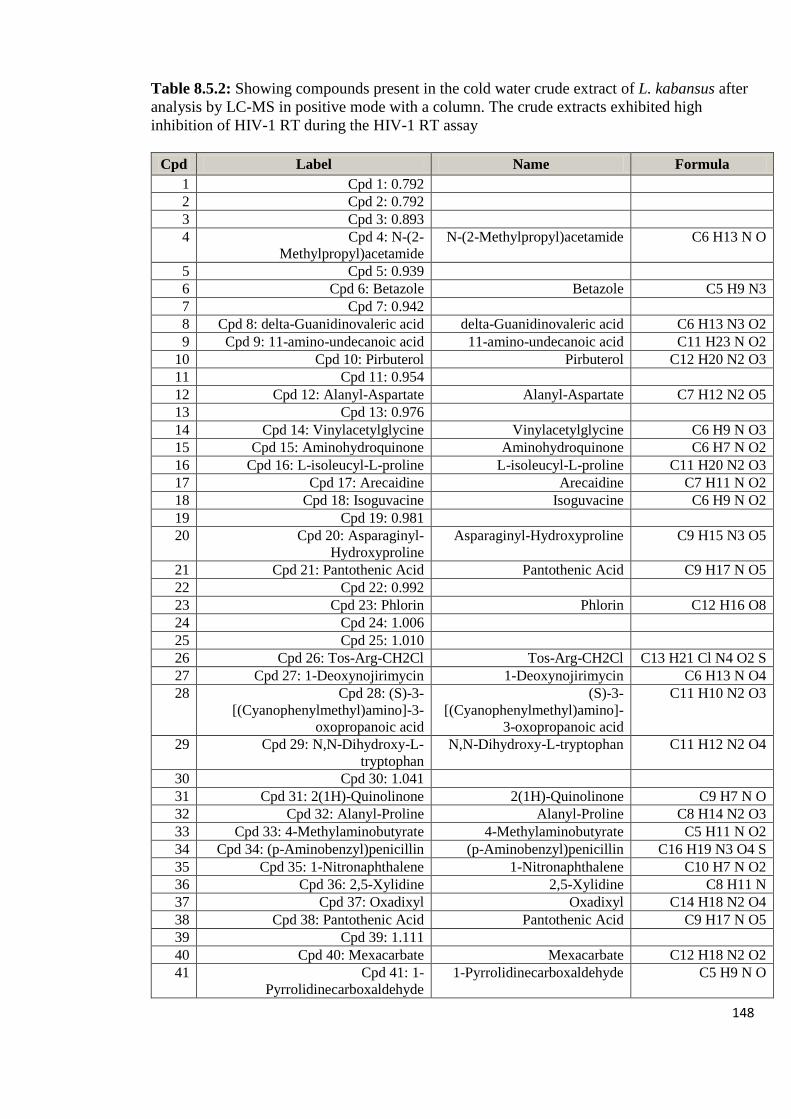
148
Table 8.5.2: Showing compounds present in the cold water crude extract of L. kabansus after
analysis by LC-MS in positive mode with a column. The crude extracts exhibited high
inhibition of HIV-1 RT during the HIV-1 RT assay
Cpd Label Name Formula
1 Cpd 1: 0.792
2 Cpd 2: 0.792
3 Cpd 3: 0.893
4 Cpd 4: N-(2-
Methylpropyl)acetamide
N-(2-Methylpropyl)acetamide C6 H13 N O
5 Cpd 5: 0.939
6 Cpd 6: Betazole Betazole C5 H9 N3
7 Cpd 7: 0.942
8 Cpd 8: delta-Guanidinovaleric acid delta-Guanidinovaleric acid C6 H13 N3 O2
9 Cpd 9: 11-amino-undecanoic acid 11-amino-undecanoic acid C11 H23 N O2
10 Cpd 10: Pirbuterol Pirbuterol C12 H20 N2 O3
11 Cpd 11: 0.954
12 Cpd 12: Alanyl-Aspartate Alanyl-Aspartate C7 H12 N2 O5
13 Cpd 13: 0.976
14 Cpd 14: Vinylacetylglycine Vinylacetylglycine C6 H9 N O3
15 Cpd 15: Aminohydroquinone Aminohydroquinone C6 H7 N O2
16 Cpd 16: L-isoleucyl-L-proline L-isoleucyl-L-proline C11 H20 N2 O3
17 Cpd 17: Arecaidine Arecaidine C7 H11 N O2
18 Cpd 18: Isoguvacine Isoguvacine C6 H9 N O2
19 Cpd 19: 0.981
20 Cpd 20: Asparaginyl-
Hydroxyproline
Asparaginyl-Hydroxyproline C9 H15 N3 O5
21 Cpd 21: Pantothenic Acid Pantothenic Acid C9 H17 N O5
22 Cpd 22: 0.992
23 Cpd 23: Phlorin Phlorin C12 H16 O8
24 Cpd 24: 1.006
25 Cpd 25: 1.010
26 Cpd 26: Tos-Arg-CH2Cl Tos-Arg-CH2Cl C13 H21 Cl N4 O2 S
27 Cpd 27: 1-Deoxynojirimycin 1-Deoxynojirimycin C6 H13 N O4
28 Cpd 28: (S)-3-
[(Cyanophenylmethyl)amino]-3-
oxopropanoic acid
(S)-3-
[(Cyanophenylmethyl)amino]-
3-oxopropanoic acid
C11 H10 N2 O3
29 Cpd 29: N,N-Dihydroxy-L-
tryptophan
N,N-Dihydroxy-L-tryptophan C11 H12 N2 O4
30 Cpd 30: 1.041
31 Cpd 31: 2(1H)-Quinolinone 2(1H)-Quinolinone C9 H7 N O
32 Cpd 32: Alanyl-Proline Alanyl-Proline C8 H14 N2 O3
33 Cpd 33: 4-Methylaminobutyrate 4-Methylaminobutyrate C5 H11 N O2
34 Cpd 34: (p-Aminobenzyl)penicillin (p-Aminobenzyl)penicillin C16 H19 N3 O4 S
35 Cpd 35: 1-Nitronaphthalene 1-Nitronaphthalene C10 H7 N O2
36 Cpd 36: 2,5-Xylidine 2,5-Xylidine C8 H11 N
37 Cpd 37: Oxadixyl Oxadixyl C14 H18 N2 O4
38 Cpd 38: Pantothenic Acid Pantothenic Acid C9 H17 N O5
39 Cpd 39: 1.111
40 Cpd 40: Mexacarbate Mexacarbate C12 H18 N2 O2
41 Cpd 41: 1-
Pyrrolidinecarboxaldehyde
1-Pyrrolidinecarboxaldehyde C5 H9 N O
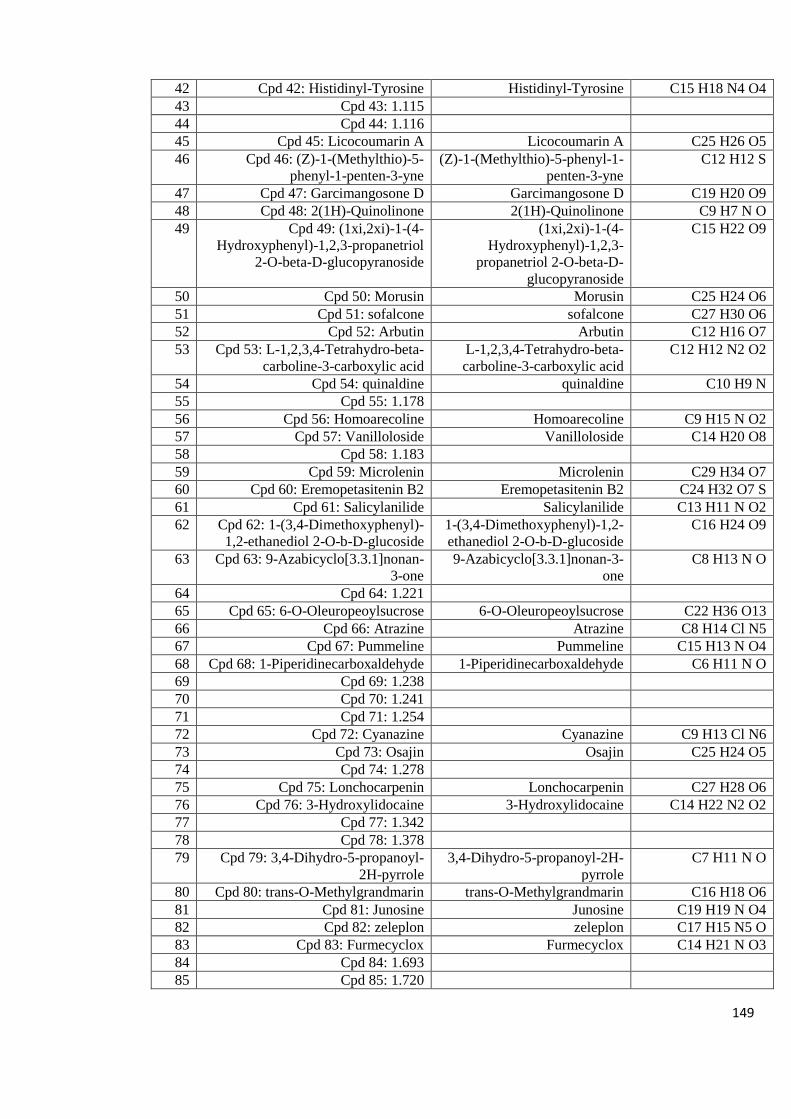
149
42 Cpd 42: Histidinyl-Tyrosine Histidinyl-Tyrosine C15 H18 N4 O4
43 Cpd 43: 1.115
44 Cpd 44: 1.116
45 Cpd 45: Licocoumarin A Licocoumarin A C25 H26 O5
46 Cpd 46: (Z)-1-(Methylthio)-5-
phenyl-1-penten-3-yne
(Z)-1-(Methylthio)-5-phenyl-1-
penten-3-yne
C12 H12 S
47 Cpd 47: Garcimangosone D Garcimangosone D C19 H20 O9
48 Cpd 48: 2(1H)-Quinolinone 2(1H)-Quinolinone C9 H7 N O
49 Cpd 49: (1xi,2xi)-1-(4-
Hydroxyphenyl)-1,2,3-propanetriol
2-O-beta-D-glucopyranoside
(1xi,2xi)-1-(4-
Hydroxyphenyl)-1,2,3-
propanetriol 2-O-beta-D-
glucopyranoside
C15 H22 O9
50 Cpd 50: Morusin Morusin C25 H24 O6
51 Cpd 51: sofalcone sofalcone C27 H30 O6
52 Cpd 52: Arbutin Arbutin C12 H16 O7
53 Cpd 53: L-1,2,3,4-Tetrahydro-beta-
carboline-3-carboxylic acid
L-1,2,3,4-Tetrahydro-beta-
carboline-3-carboxylic acid
C12 H12 N2 O2
54 Cpd 54: quinaldine quinaldine C10 H9 N
55 Cpd 55: 1.178
56 Cpd 56: Homoarecoline Homoarecoline C9 H15 N O2
57 Cpd 57: Vanilloloside Vanilloloside C14 H20 O8
58 Cpd 58: 1.183
59 Cpd 59: Microlenin Microlenin C29 H34 O7
60 Cpd 60: Eremopetasitenin B2 Eremopetasitenin B2 C24 H32 O7 S
61 Cpd 61: Salicylanilide Salicylanilide C13 H11 N O2
62 Cpd 62: 1-(3,4-Dimethoxyphenyl)-
1,2-ethanediol 2-O-b-D-glucoside
1-(3,4-Dimethoxyphenyl)-1,2-
ethanediol 2-O-b-D-glucoside
C16 H24 O9
63 Cpd 63: 9-Azabicyclo[3.3.1]nonan-
3-one
9-Azabicyclo[3.3.1]nonan-3-
one
C8 H13 N O
64 Cpd 64: 1.221
65 Cpd 65: 6-O-Oleuropeoylsucrose 6-O-Oleuropeoylsucrose C22 H36 O13
66 Cpd 66: Atrazine Atrazine C8 H14 Cl N5
67 Cpd 67: Pummeline Pummeline C15 H13 N O4
68 Cpd 68: 1-Piperidinecarboxaldehyde 1-Piperidinecarboxaldehyde C6 H11 N O
69 Cpd 69: 1.238
70 Cpd 70: 1.241
71 Cpd 71: 1.254
72 Cpd 72: Cyanazine Cyanazine C9 H13 Cl N6
73 Cpd 73: Osajin Osajin C25 H24 O5
74 Cpd 74: 1.278
75 Cpd 75: Lonchocarpenin Lonchocarpenin C27 H28 O6
76 Cpd 76: 3-Hydroxylidocaine 3-Hydroxylidocaine C14 H22 N2 O2
77 Cpd 77: 1.342
78 Cpd 78: 1.378
79 Cpd 79: 3,4-Dihydro-5-propanoyl-
2H-pyrrole
3,4-Dihydro-5-propanoyl-2H-
pyrrole
C7 H11 N O
80 Cpd 80: trans-O-Methylgrandmarin trans-O-Methylgrandmarin C16 H18 O6
81 Cpd 81: Junosine Junosine C19 H19 N O4
82 Cpd 82: zeleplon zeleplon C17 H15 N5 O
83 Cpd 83: Furmecyclox Furmecyclox C14 H21 N O3
84 Cpd 84: 1.693
85 Cpd 85: 1.720
150
86 Cpd 86: 1.911
87 Cpd 87: 1.952
88 Cpd 88: 1.980
89 Cpd 89: 2.010
90 Cpd 90: 2.060
91 Cpd 91: Lumichrome Lumichrome C12 H10 N4 O2
92 Cpd 92: N4-Phosphoagmatine N4-Phosphoagmatine C5 H15 N4 O3 P
93 Cpd 93: Ptelatoside A Ptelatoside A C19 H26 O10
94 Cpd 94: Lymecycline Lymecycline C29 H38 N4 O10
95 Cpd 95: Ononin Ononin C22 H22 O9
96 Cpd 96: 3-Methylene-indolenine 3-Methylene-indolenine C9 H7 N
97 Cpd 97: 3.523
98 Cpd 98: 3.523
99 Cpd 99: Hexyl glucoside Hexyl glucoside C12 H24 O6
100 Cpd 100: 3.812
101 Cpd 101: 4.459
102 Cpd 102: hexamethylene
bisacetamide
hexamethylene bisacetamide C10 H20 N2 O2
103 Cpd 103: 5.514
104 Cpd 104: Heptopargil Heptopargil C13 H19 N O
105 Cpd 105: 8.079
106 Cpd 106: Isocolumbin Isocolumbin C20 H22 O6
107 Cpd 107: 8.742
108 Cpd 108: Ferimzone Ferimzone C15 H18 N4
109 Cpd 109: Isogingerenone B Isogingerenone B C22 H26 O6
110 Cpd 110: 2-Methylbenzaldehyde 2-Methylbenzaldehyde C8 H8 O
111 Cpd 111: Styrene Styrene C8 H8
112 Cpd 112: Isogingerenone B Isogingerenone B C22 H26 O6
113 Cpd 113: Sapidolide A Sapidolide A C14 H18 O5
114 Cpd 114: 13.016
115 Cpd 115: Ginsenoyne K Ginsenoyne K C17 H24 O3
116 Cpd 116: N6-Galacturonyl-L-lysine N6-Galacturonyl-L-lysine C12 H22 N2 O8
117 Cpd 117: 17.266
118 Cpd 118: 17.897
119 Cpd 119: Tributyl phosphate Tributyl phosphate C12 H27 O4 P
120 Cpd 120: Prosolanapyrone II Prosolanapyrone II C18 H24 O4
121 Cpd 121: (S)-alpha-Terpinyl
glucoside
(S)-alpha-Terpinyl glucoside C16 H28 O6
122 Cpd 122: 18.672
123 Cpd 123: Polysorbate 60 Polysorbate 60 C22 H42 O8
124 Cpd 124: 18.985
125 Cpd 125: 19.032
126 Cpd 126: 19.046
127 Cpd 127: 19.047
128 Cpd 128: 19.058
129 Cpd 129: Goshonoside F2 Goshonoside F2 C26 H44 O8
130 Cpd 130: 19.344
131 Cpd 131: 19.352
132 Cpd 132: 19.707
133 Cpd 133: 13-Demethylspirolide C 13-Demethylspirolide C C42 H61 N O7
134 Cpd 134: 19.738
135 Cpd 135: 19.751

151
136 Cpd 136: 19.768
137 Cpd 137: 19.772
138 Cpd 138: 19.791
139 Cpd 139: 19.799
140 Cpd 140: 19.828
141 Cpd 141: Iriomoteolide 1a Iriomoteolide 1a C29 H46 O7
142 Cpd 142: 19.852
143 Cpd 143: 19.959
144 Cpd 144: 19.975
145 Cpd 145: 20.009
146 Cpd 146: 20.039
147 Cpd 147: 20.071
148 Cpd 148: 20.096
149 Cpd 149: 20.098
150 Cpd 150: Methyl (7Z,9Z,9'Z)-6'-apo-
y-caroten-6'-oate
Methyl (7Z,9Z,9'Z)-6'-apo-y-
caroten-6'-oate
C33 H44 O2
151 Cpd 151: 20.119
152 Cpd 152: 20.134
153 Cpd 153: 20.142
154 Cpd 154: 20.253
155 Cpd 155: Pipericine Pipericine C22 H41 N O
156 Cpd 156: Docosatetraenoyl
Ethanolamide
Docosatetraenoyl
Ethanolamide
C24 H41 N O2
157 Cpd 157: 20.537
158 Cpd 158: 20.539
159 Cpd 159: Docosatrienoic Acid Docosatrienoic Acid C22 H38 O2
160 Cpd 160: 20.542
161 Cpd 161: PEP-16:0/18:1(11Z)) PEP-16:0/18:1(11Z)) C39 H76 N O7 P
162 Cpd 162: 20.646
163 Cpd 163: 20.666
164 Cpd 164: 20.722
165 Cpd 165: 20.729
166 Cpd 166: 20.751
167 Cpd 167: 20.770
168 Cpd 168: 20.774
169 Cpd 169: 20.783
170 Cpd 170: 20.798
171 Cpd 171: Palmitic amide Palmitic amide C16 H33 N O
172 Cpd 172: Oleamide Oleamide C18 H35 N O
173 Cpd 173: Caffeoylcycloartenol Caffeoylcycloartenol C39 H56 O4
174 Cpd 174: 21.285
175 Cpd 175: 21.298
176 Cpd 176: 23.764
177 Cpd 177: 23.776
178 Cpd 178:
PI(18:1(9Z)/20:3(8Z,11Z,14Z))
PI(18:1(9Z)/20:3(8Z,11Z,14Z)) C47 H83 O13 P
179 Cpd 179: 7-Hydroxychlorophyll a 7-Hydroxychlorophyll a C55 H72 Mg N4 O6
180 Cpd 180: Pheophytin a Pheophytin a C55 H74 N4 O5
181 Cpd 181: PE(15:0/24:1(15Z)) PE(15:0/24:1(15Z)) C44 H86 N O8 P
Copyright © 2022 FDOKUMEN




















