
Various Natural and Anthropogenic Factors Responsible for ...
Upload
khangminh22Category
view
3download
0
ANTHROPOGENIC DEBRIS AND PATHOLOGY IN AQUATIC BIRDS
By
Erika R. Holland
B.Sc. (Hons.), Acadia University, 2015
Thesis submitted in partial fulfillment of the requirements for
the Degree of Master of Science (Biology)
Acadia University
October 2017
© by Erika Reigh Holland, 2017



ii
This thesis is accepted in its present form by the Division of Research and Graduate
Studies as satisfying the thesis requirements for the degree Master of Science (Biology).
.................................................
I, Erika R. Holland, grant permission to the University Librarian at Acadia University to
reproduce, loan or distribute copies of my thesis in microform, paper or electronic
formats on a non-profit basis. I, however, retain the copyright in my thesis.
Erika Holland
_ _
Dr. Mark Mallory
_
Dr. Dave Shutler
_ _________________
Date

iii
TABLE OF CONTENTS
TABLE OF CONTENTS ...................................................................................................................... iii LIST OF TABLES .............................................................................................................................................. iv LIST OF FIGURES .............................................................................................................................................. v ABSTRACT .......................................................................................................................................................... vi ACKNOWLEDGMENTS ............................................................................................................................... vii PREFACE .............................................................................................................................................................. ix
CHAPTER ONE: GENERAL INTRODUCTION TO ENVIRONMENTAL ANTHROPOGENIC DEBRIS ............................................................................................................. 1
Introduction ............................................................................................................................................................ 1 Plastics in Marine Environments ..................................................................................................................... 4 Plastics in Freshwater Environments ............................................................................................................. 7
Microbeads: A growing concern ................................................................................................................ 9 My Research ........................................................................................................................................................ 12
Structure of this thesis ................................................................................................................................. 13 CHAPTER TWO: PLASTICS AND OTHER ANTHROPOGENIC DEBRIS IN FRESHWATER BIRDS FROM CANADA ................................................................................. 15
Introduction .......................................................................................................................................................... 15 Material and Methods ....................................................................................................................................... 19
Sampling .......................................................................................................................................................... 19 Processing, separation, sorting and identifying .................................................................................. 19 Statistical analyses........................................................................................................................................ 21
Results .................................................................................................................................................................... 22 Discussion ............................................................................................................................................................. 24 Conclusions .......................................................................................................................................................... 27
CHAPTER THREE: ANTHROPOGENIC DEBRIS AND PATHOLOGY OF FULMARS AND SHEARWATERS BEACHED ON SABLE ISLAND, NOVA SCOTIA, CANADA ............................................................................................................................. 33
Introduction .......................................................................................................................................................... 33 Materials and Methods ..................................................................................................................................... 37
Sample collection and processing ............................................................................................................ 37 Statistical analyses........................................................................................................................................ 38
Results .................................................................................................................................................................... 39 Sooty Shearwater .......................................................................................................................................... 40 Great Shearwater .......................................................................................................................................... 40 Northern Fulmar ........................................................................................................................................... 41
Discussion ............................................................................................................................................................. 42 CHAPTER FOUR: GENERAL DISCUSSION AND FUTURE DIRECTIONS .............. 52
Future Actions ..................................................................................................................................................... 52 Appendices ........................................................................................................................................................... 86

iv
LIST OF TABLES
Table 1. Sample sizes, body mass, the frequency, and mean number of pieces for plastic,
metal, and any debris…………………………………………...………………………..30
Table 2. Amount and types of debris recovered from the nine species that ingested
debris.…………………………………………………………………………………….32
Table 3. Overview of the number and corresponding percentages of age, sex, and season
collected, for four species of Procellariiformes collected beached on Sable Island, Nova
Scotia, between 2000 and 2012.……………………………………...………………….47
Table 4. Pathology, ranked from most to least common, in 318 procellariids collected
beached on Sable Island, Nova Scotia, Canada, between 2000 and 2012……………….48
Table 5. Average body mass, and average mass of anthropogenic debris particles found
in the gizzards and proventriculi of four species of Procellariiformes collected beached on
Sable Island, Nova Scotia, between 2000 and 2012..………………………………........49

v
LIST OF FIGURES
Figure 1. Omnivore and piscivore digestive tracts showing physiological differences in
structure.…………...………………………………………………..……………………14
Figure 2. Sample sites used for this study...…………………………………..…………29
Figure 3. Monthly collection breakdown of procellariid carcasses found beached on
Sable Island, Nova Scotia, Canada, between 2000-2012...………………………………50
Figure 4. Plumage morphs of beached northern fulmars (Fulmarus glacialis) collected
from Sable Island, Nova Scotia, Canada, between 2000-2012 and associated ingested
masses of anthropogenic debris (g) removed from the gizzard and proventriculus of
birds………………………………………………………………………………………51

vi
ABSTRACT
Plastics in marine and freshwater environments are a global environmental issue.
Plastic ingestion is associated with a variety of deleterious health effects for wildlife, and
is a focus of much international research and monitoring. However, little research has
focused on the ramifications of plastic debris for freshwater organisms. In chapter 2 I
quantified plastic and other anthropogenic debris in 350 individuals of 17 freshwater and
one marine bird species collected across Canada. I determined prevalence of
anthropogenic debris in freshwater birds’ to be 11.4% across all species studied.
In chapter 3 I determined the prevalence of anthropogenic debris and their
consequences in seabirds, using four species of procellariid seabirds found beached on
Sable Island, Nova Scotia, Canada between 2000 and 2012. Pooling the four species, no
differences in prevalence of different pathologies was found relative to age or sex, with
birds having (in descending order of occurrence) emaciation, autolysis, parasite infection,
inflammation, trauma, bacterial infection, drowned, tumors, tissue necrosis, impaction,
myopathy, or pneumonia. Northern fulmars washed up dead of emaciation less frequently
(69/115; 60%) than sooty (36/48; 75%) or great (67/86; 78%) shearwaters. Pathology in
northern fulmars (Fulmarus glacialis), sooty (Ardenna grisea) and great shearwaters
(Ardenna gravis) appeared unrelated to mass of debris ingested or body mass, and
similarly body mass was not significantly related to mass of debris ingested.
This work established that anthropogenic debris is a genuine concern for
management of the health of freshwater and marine ecosystems, provided a baseline for
the prevalence of anthropogenic debris ingestion in freshwater birds in Canada, and
presented the first in-depth look at pathology of birds beached on Sable Island.

vii
ACKNOWLEDGMENTS
First and foremost, thank you to my advisors and co-authors Drs. Mark Mallory
and Dave Shutler, without whom this thesis would not have been possible. Thanks also to
Dr. Pierre-Yves Daoust and Zoe Lucas for providing data for chapter two. Thanks are
also due to my advisory committee, composed of Drs. Mark Mallory, Dave Shutler, and
Steve Mockford. A million thanks to Danielle Quinn, Trevor Avery, and the entire R-Bar
crew for your essential help with stats in R. I also thank my family and friends for their
continuous and unending support, patience, and humour.
A huge thank you to all the individuals who contributed digestive tracts to my
research: Leo and Maria Brink, Peggy Crawford, Matthew English, Mark Gloutney, John
Goas, Randy Goreham, Matt Gregoire, Jeff Krete, Julie Mallett, Regan Maloney, Nic
McLellan, Michael O'Brien, Kirsten Pearson, Eric Reed, Greg Robertson, Ken Tucker,
Becky Whittam, Paul Woodard, and most of all, Laurie Wilson. Thanks also to Brook
Beauliua, Ashton Baich, Robin Dornan, Laura McGinnis, Sarah Mitchell, Jackie Morris,
Alexandra Nesnidalova, Dione Rousseau, and Lindsay Trainor for their help in lab with
freshwater bird dissections. Special thanks to Matthew English for his assistance with
dissection methods, and to Stephanie White for her assistance finding local hunters and
lab assistants through the Annapolis Valley Ducks Unlimited Canada Chapter.
Financial support for this project was provided by a Natural Sciences and
Engineering Research Council of Canada Alexander Graham Bell Canada Graduate
Scholarship - Master’s Program (CGS M), The Nova Scotia Innovation & Research
Graduate Scholarship (Master's) in Ocean Science & Tech/Life Sciences, an Alden B.

viii
Dawson Scholarship, an Acadia Graduate Scholarship, a Ducks Unlimited
Canada/Acadia Research Partnership Grant, and the Natural Sciences and Engineering
Research Council of Canada (NSERC; Grant numbers 418551-2012 and 2015-05617).
Collection and possession of waterfowl were made under appropriate permits from the
Canadian Wildlife Service (Service Permit no. SS2802).

ix
PREFACE
Chapters 2 and 3 of this thesis were written with the intent of being published in
peer-reviewed journals along with co-authors. Thus, I have chosen to retain the more
inclusive pronouns “we” and “our” for those chapters.
Chapter 2 has previously been published in a peer-reviewed journal, although it
has been updated since this publication. The original publication may be referenced in
other works as:
Holland, E., Mallory, M.L., Shutler, D., 2016. Plastics and other anthropogenic debris in
freshwater birds from Canada. Science of the Total Environment 571, 251–258.
doi:http://dx.doi.org/10.1016/j.scitotenv.2016.07.158


1
CHAPTER ONE: GENERAL INTRODUCTION TO ENVIRONMENTAL
ANTHROPOGENIC DEBRIS
Introduction
Extending over three quarters of the Earth’s surface, aquatic ecosystems play a
critical role in planetary life support (Secretariat of the Convention on Biological
Diversity, 2012). Humans rely on aquatic systems for food, water, industry and
agriculture, and millions of jobs (Food and Agriculture Organization of the United
Nations, 2016; Millennium Ecosystem Assessment, 2005; Secretariat of the Convention
on Biological Diversity, 2012). However, human actions, specifically pollution by
anthropogenic debris such as plastics, constitute a major threat to aquatic environments,
the species therein, and the resources they provide (Derraik, 2002).
Humans have been releasing plastics into their environment since the early 1900s
(Bijker et al., 1987; Klar et al., 2014), and plastics in marine environments have been
identified as an emerging environmental issue at a global level, with concentrations
increasing rapidly (United Nations Environment Programme, 2014). Since commercial
production began in the 1950s, the plastics industry has grown by an average of 8.7% per
year, and in 2013, global production rose to 299 million tonnes (Plastics Europe, 2013,
2015).
Of all anthropogenic debris released into our waterways, plastic debris may
overall be the most problematic due to its negative effects on wildlife (Ashton et al.,
2010; Rios et al., 2010; Wright et al., 2013) and slow breakdown time (Derraik, 2002).

2
Plastic entanglement of marine organisms can lead to general debilitation, entanglement-
related injuries, impaired diving for feeding or surfacing for breath, starvation, and
eventual death (Mascarenhas et al., 2004; Gregory, 2009; Votier et al., 2011). Marine
plastic waste can mimic natural food, leading to debris ingestion by aquatic organisms
being more than proportional to their availability. However, plastics fail to provide
nutrition and once ingested can lead to weakness, false feelings of satiation, irritation of
the stomach lining, digestive tract blockage, internal bleeding, abrasion, ulcers, failure to
put on fat stores necessary for migration and reproduction, and death through starvation
(Moore, 2008; Wright et al., 2013). Plastic debris also vector heavy metal contaminants
and high concentrations of organochlorines such as polychlorinated biphenyls (PCBs),
dichlorodiphenyl trichloroethane (DDT) and polycyclic aromatic hydrocarbons (PAHs)
(Teuten et al., 2007; Ashton et al., 2010; Rios et al., 2010). Once plastics are discharged
into an aquatic environment they can persist for anywhere from three to 50 years prior to
complete disintegration, and complete mineralization may take hundreds or thousands of
years (Gregory, 1978; Derraik, 2002; Driedger et al., 2015).
Although plastic debris is the most prevalent and most researched form of aquatic
anthropogenic debris it is not the only material with reported deleterious effects (van
Franeker and Meijboom, 2002). Fragments of metals, rubber, cloth, glass, Styrofoam, and
other anthropogenic products often find their way into aquatic ecosystems, resulting in
seabird ingestion (Auman et al., 1997; Moser and Lee, 1992; Tourinho et al., 2009; van
Franeker and Meijboom, 2002). However, ingestion rates among a variety of species are
higher for plastic debris than other anthropogenic materials (van Franeker and Meijboom,
2002). This may be due to higher exposure, as plastics often make up the majority (80-

3
92%) of total anthropogenic material present in aquatic ecosystems and along shorelines
(Lucas, 1992; Driedger et al., 2015).
To date little research has focused on the ramifications of plastic and other
anthropogenic debris for freshwater organisms. Research on freshwater ecosystems is
needed to put into effect crucial laws and regulations governing debris release into
waterways. These regulations are needed to stop potentially harmful biomagnification up
the food chain, reducing future plastic and debris contaminant contact by humans through
their freshwater food sources.
This introduction reviews the current state of knowledge for marine
anthropogenic debris and research, including species harmed by plastic exposure, and
introduces the growing field of freshwater debris research, with a specific focus on the
rising issue of microbeads. I suggest future analyses on freshwater organisms are
important to maintaining healthy populations, and decreasing anthropogenic debris
contact. Research into freshwater plastic and other anthropogenic debris is significant
because little is currently known about anthropogenic debris exposure and ingestion in
freshwater organisms. More data are required to determine potentially negative effects on
humans through biomagnification from wild-caught avian foods. Research has shown
that for North Sea cod (Gadus morhua), plastic ingestion appears to be a pathway for
intestinal tract leaching, and subsequent cod exposure to nonylphenol (NP) and bisphenol
A (BPA), although concentrations acquired are negligible (Koelmans et al., 2014).
Research on short-tailed shearwaters (Puffinus tenuirostris) and great shearwaters
(Puffinus gravis) has shown that avian tissues can acquire polybrominated diphenyl ether
(PBDE) flame retardants and polychlorinated biphenyls (PCBs) from ingested plastic

4
debris (Ryan et al., 1988; Tanaka et al., 2013, 2015), which could put humans consuming
contaminated tissues at risk of bioaccumulation. Likewise, northern fulmar (Fulmarus
glacialis) eggs of the Faroe Islands contain high levels of brominated flame retardants
(BFRs) and PBDEs, and may be a source of lipid-soluble persistent pollutants to the
Faroese peoples (Karlsson et al., 2006). PBDEs ingested by humans can have adverse
health effects, including cancer, thyroid hormone disruption, anti-androgen action,
neurodevelopmental deficits, and developmental neurotoxicity (McDonald, 2002; Costa
et al., 2008; Talsness et al., 2009). Humans ingesting PCBs through animal fats may find
that the accumulation of these fat-soluble substances can initiate suppression of the
immune system, alter thyroid and reproductive function, increase risk of developing
cardiovascular and liver disease and diabetes, while pregnant women exposed to PCBs
are at risk of giving birth to infants of low birth weight (Cheek et al., 1999; Holladay and
Smialowicz, 2000; Carpenter, 2006). PCB exposure in infants may also reduce IQ and
alter behaviour (Carpenter, 2006).
Plastics in Marine Environments
Marine environments from the equator to the poles contain plastic debris from
their surface waters to their deep-sea sediments (Kanehiro et al., 1995; Derraik, 2002;
Van Cauwenberghe et al., 2013; Obbard et al., 2014; Woodall et al., 2014; Taylor et al.,
2016). Due to plastic’s initial buoyancy and eventual settlement, surface-feeding birds
and dabbling ducks may be particularly susceptible to plastic debris ingestion (Ng and
Obbard, 2006; Eriksen et al., 2014; Ivar do Sul and Costa, 2014). Research on plastics in
marine environments has identified plastic’s initial buoyancy (specific gravity of 0.9;

5
Snyder and Vakos, 1966) as detrimental, because this buoyancy often leads to dispersal
over long distances (Derraik, 2002; Eriksen et al., 2014) and increased availability to
surface-feeding species (Day et al., 1985; Moser and Lee, 1992). However, over time
plastic debris in aquatic environments undergoes fouling due to biofilms, hitchhiking
organisms, and absorption of pollutants, causing plastics to become negatively buoyant
and sink into sediment (Ye and Andrady, 1991; Barnes et al., 2009; Frias et al., 2010;
Driedger et al., 2015).
Entanglement and ingestion of marine anthropogenic debris negatively affects all
seven known species of sea turtle (100%), 45% of all species of marine mammals, and
56% of seabird species (Secretariat of the Convention on Biological Diversity, 2012; Gall
and Thompson, 2015). Studies on seabird plastic ingestion have been conducted since the
1960s, when Kenyon and Kridler (1969) found that Laysan albatross (Phoebastria
immutabilis) fledglings were consuming plastic. Estimates from the 1990s were that 35%
of the world's seabird species were negatively affected by marine plastics (Laist, 1997).
Recent studies predict that, unless effective waste management practices are put into
place, 99% of all seabird species will be ingesting plastics by 2050 (Wilcox et al., 2015).
Once consumed it can take a bird one month to a year to pass ingested plastics (Day,
1980; Ryan and Jackson, 1987; van Franeker and Law, 2015). Despite estimates of slow
plastic expulsion times, the majority of studies fail to find plastics in seabird digestive
tracts or feces (studies on albatross, fulmars, shearwaters, storm-petrels, gulls, auklets,
and puffins; Day, 1980; Pettit et al., 1981; Rothstein, 1973). This could in part be due to
adults of certain species feeding their chicks through regurgitation, and thus passing their

6
plastic load on to their offspring (Pettit et al., 1981; Laist, 1987). Due to this unloading,
young birds often suffer mortality due to plastic (Derraik, 2002).
Some seabirds consume specific plastic shapes and colours more frequently,
perhaps mistaking them for prey items (Day et al., 1985; Moser and Lee, 1992; Shaw and
Day, 1994). Pale particles are often overrepresented in fulmars, shearwaters, petrels,
phalaropes, gulls, kittiwakes, and terns (Rothstein, 1973; Day, 1980; Moser and Lee,
1992; Lavers and Bond, 2016), although this could be due to the higher availability of
light-colored plastic particles in the environment (Moser and Lee, 1992). Selective
ingestion also ties into foraging style, and availability of certain plastics in certain niches
(Ryan, 1987). For example, Moser and Lee (1992) reported aerial plunging birds had the
lowest percentage of plastic in their gizzard (2%), pursuit diving species were mid-range
(8%), and surface seizing (12%) and pattering seabirds (14%) had the highest. Similarly,
Day et al., (1985) found that aerial plunging birds contained no plastic, pattering (9%)
and surface seizing (16%) seabirds were midrange, and pursuit diving species were
highest (26%), indicating that surface-feeding species are among the most likely to have
plastics in their diets.
In the Maritimes, research into plastic pollution begin in 1983, when Gregory
(1983) reported small virgin plastic pellets and granules in litter on the shores of Nova
Scotia and Sable Island. In 1989, it was determined that 62% of persistent marine litter in
Halifax Harbor originated from recreation and land-based sources (Ross et al., 1991).
Most notable research done on microplastic pollution centers around the Gully Trough, a
marine protected area located 200 kilometers off Nova Scotia, on the edge of the Scotian
Shelf (GTA Consultants Inc., 1999; Government of Canada, 2010), and Sable Island, a

7
long, crescent-shaped, sandy island located 160 km off the coast of mainland Nova Scotia
and 53 km west of the Gully Trough (Lucas et al., 2012). Studies done on Sable Island
have assessed both coastal debris and seabird plastic ingestion.
Studies on seabird plastic ingestion often rely on culling live birds, or collecting
samples opportunistically from birds found dead on beaches. These beached birds offer
an opportunistic window into natural seabird mortality (although these individuals are
often emaciated; Barrett et al., 2007), and rates of anthropogenic debris ingestion (Pierce
et al., 2004).
Plastics in Freshwater Environments
Although most historic research focuses on plastics in marine environments,
increasing work is focusing on freshwater environmental plastic contamination.
Freshwater bodies may have comparable plastic concentrations to marine waters. The
most commonly detected plastics are < 5 mm fragments of polyethylene and
polypropylene polymers (Zbyszewski and Corcoran, 2011; Eriksen et al., 2013;
Zbyszewski et al., 2014; Corcoran et al., 2015). Corcoran et al. (2015) found up to nine
microplastic particles (0.5-3 mm in size) per 2 cm, to depths of 8 cm, in Lake Ontario
sediments, indicating deposition and accumulation over approximately the past 38 years.
Over 80% of anthropogenic litter found along the Great Lake shorelines is composed of
plastics (Driedger et al., 2015). Anthropogenic debris entering freshwater and marine
ecosystems can come from a variety of sources, the majority of which originate from
land-based activities and waterway use (Ocean Conservancy, 2010). Freshwater plastic
waste commonly includes microplastic beads (polyethylene and polypropylene

8
microspheres widely used in cosmetics as exfoliating agents; Eriksen et al., 2013) from
consumer products, pellets from the plastic manufacturing industry, and waste from
beach-goers, shipping activities, and fishing (Driedger et al., 2015). The distribution of
plastics in bottom sediments of the Great Lakes is essentially unknown, because the
majority of what is known about microplastic pollution in freshwater bodies focuses
primarily on surface waters (Castañeda et al., 2014; Driedger et al., 2015). However,
deep-sea sediments at 1100 to 5000 m contain microplastic pollution of up to an average
of one particle per 25 cm2 (Van Cauwenberghe et al., 2013) and in Tokyo Bay, Japan,
plastics made up 80-85% of seabed debris (Kanehiro et al., 1995). Sediments of the St.
Lawrence River have microbead pollution comparable in density to marine microplastic
concentrations (Castañeda et al., 2014).
Aside from environmental concentrations, data from freshwater organisms
suggest that plastic ingestion levels are also comparable to those in marine ecosystems
(Denuncio et al., 2011; Castañeda et al., 2014). Franciscana dolphins (Pontoporia
blainvillei) in estuarine habitats had higher levels of plastic ingestion (34.6%) than their
marine counterparts (19.2%; Denuncio et al., 2011), suggesting that plastic ingestion by
freshwater organisms could be a larger issue than previously thought. Likewise, up to
33% of estuarine catfish (Cathorops spixii, Cathorops agassizii and Sciades herzbergii)
from the southwestern Atlantic and 7.9% of estuarine drums (Stellifer brasiliensis and
Stellifer stellifer) of the Goiana Estuary in Brazil consumed plastics (Possatto et al., 2011;
Dantas et al., 2012). Studies focusing on freshwater ecosystems have found plastic debris
affected some green algae (Scenedesmus obliquus) and zooplankton (Daphnia magna)
(Besseling et al., 2014), as well as gudgeons (Gobio gobio; Sanchez et al., 2014), yellow

9
perch (Perca flavescens; Moseman, 2015), and could potentially also affect benthivorous
fishes [preliminary results on benthic round gobys’ (Neogobius melanostomus) digestive
tracts suggested the presence of microbeads] and macroinvertebrates (Castañeda et al.,
2014).
Red phalaropes (Phalaropus fulicarius) and red-necked phalaropes (P. lobatus),
which spend a portion of their life consuming freshwater zooplankton, also consume
plastic debris (Day et al., 1985; Moser and Lee, 1992; Bayly, 1993). English et al. (2015)
examined three waterfowl species wintering in Atlantic Canada: mallards (Anas
platyrhynchos), American black ducks (A. rubripes) and common eiders (Somateria
mollissima). They found a surprisingly high prevalence of plastics in ducks’ stomachs
[46.1% (6/13) of mallards, 6.9% (6/87) of black ducks, and 2.1% (1/48) of eiders
contained plastic]. However, it is unknown whether the birds acquired plastic debris in
freshwater or marine locations. Likewise Gil-Delgado et al. (2017) reported plastic in the
faeces of three species of waterfowl: shelduck (Tadorna tadorna), European coot (Fulica
atra), and mallard. Shelduck faeces had a 43.8% prevalence of plastic remnants,
European coots 60%, and mallards 45% (Gil-Delgado et al., 2017).
Microbeads: A growing concern
Studies on freshwater species potential acquisition of plastics are important,
especially in light of research on freshwater microbead concentrations. Microbeads have
gained popularity in recent years and their increased use has led to an unforeseen increase
of these plastics entering aquatic ecosystems (Castañeda et al., 2014; Doughty and
Eriksen, 2014). Their relatively small diameter (< 1 mm) means many wastewater

10
treatment plants cannot remove them, unless they employ fine- or micro-screens,
microfiltration, sand filtration, or mixed media filtration (Castañeda et al., 2014; Doughty
and Eriksen, 2014; Driedger et al., 2015). In New York State, USA, sampling showed
that not employing these filtration methods led to six out of seven wastewater treatment
plants discharging microbeads into rivers, lakes, and oceans (Schneiderman, 2014). An
example of the effectiveness of these methods can be seen when comparing Lake Huron,
USA’s 2,779 plastic particles km2 due to high plastic recycling rates, and high
wastewater treatment plant retention efficiencies, to Lake Hovsgol, Mongolia, which
lacking a modern waste management system has 20,264 plastic particles/km2 (Free et al.,
2014).
To date, relatively little research has been conducted on microbeads. However,
public outcry has meant that some major companies have banned, or are planning to ban,
microbeads, but lax regulations mean that loopholes still exist (Newman et al., 2013;
International Joint Commission Canada and United States, 2016). To combat this, the
Great Lakes and St. Lawrence Cities Initiative (GLSLCI), a binational coalition of
mayors and municipal officials, has adopted a resolution calling for industries to phase
out microbeads (Great Lakes and St. Lawrence Cities Initiative, 2014). GLSLCI further
calls for bans on developing new products containing microbeads, for companies to phase
out the use of all existing products containing microbeads, and for provincial, state, and
federal bans on microbead usage in consumer products (Great Lakes and St. Lawrence
Cities Initiative, 2014). In response, the United States recently passed the Microbead
Free Waters Act of 2015 (House Report No. 114-371, 2015) and the Canadian
government added microbeads to the Schedule 1 list of toxic substances under the

11
Canadian Environmental Protection Act (Environment and Climate Change Canada,
2010; Government of Canada, 2015). The proposed Microbeads in Toiletries Regulations
would prohibit the sale of products that contain microbeads by 2019 (Government of
Canada, 2016). The Belgian, Netherlands, Austrian and Swedish delegations to the
European Union, supported by the Luxembourg delegation, are pushing for a European
ban on microbeads (Council of the European Union, 2014) and the UK government is
preparing to ban the sale and manufacture of cosmetics and personal care products
containing microbeads (UK Government, 2016). In Australia, New South Wales and
South Australia are leading a phase-out of microbeads in their jurisdictions, to be
completed by 2018 (Australian Government, 2016). Legislation banning the use of
microbeads in consumer goods may be a huge and necessary step, because per capita
consumption of polyethylene microplastic beads in personal care products for the U.S.
population is approximately 2.4 mg per person each day, leading to the U.S. alone
discharging 263 tonnes per year of polyethylene microplastic (Gouin et al., 2011). Large
concentrations of these plastics are problematic because microbeads are expected to
persist for centuries before re-entering normal biogeochemical cycles (Leslie, 2014).
Although efforts are being made to clean ocean waters of plastics and help
mitigate increasing concentrations (Kershaw et al., 2011), lack of data, primarily through
the absence of work documenting the scale and scope of this problem, has meant that no
similar initiatives exist for freshwater bodies (Ryan et al., 2009). Inadequate initiatives
and legislation governing freshwater plastic debris could be due to the inadequacy of
standardized measures for quantifying plastic debris (Driedger et al., 2015).
Complications derived from different methods of reporting plastic debris concentrations

12
in freshwater and marine environments could also affect initiatives, because there is no
internationally agreed upon classification system for plastic debris (Driedger et al., 2015).
However, the following size classes, defined by mesh filter sizes, are often used:
microscopic plastic debris (0.45 μm to 333 μm), microplastics (> 333 μm to 5 mm), and
macroplastic (> 5 mm) (Arthur et al., 2008; Andrady, 2011; Driedger et al., 2015).
My Research
I researched anthropogenic debris ingestion in freshwater and marine birds, with a
specific focus on microplastics and other micro-debris. Although numerous similar
studies have been conducted on marine birds, little research has focused on the
ramifications of increasing debris concentrations to freshwater species. I also conducted
research into beached bird pathology, and the relationship of pathological findings to
ingested debris, among fulmars and shearwaters from Sable Island, Nova Scotia, Canada.
These studies were primarily dissection-based, with the gastrointestinal tract of
individuals sampled being dissected in a lab. The digestive tracts of birds are relatively
simple, consisting of an esophagus, an acid-proteolytic stomach (consisting of the
proventriculus and gizzard), a tubular small intestine, and a very short colon (Hilton et
al., 2000). For these studies only the proventriculus and gizzards were dissected.
Piscivorous birds, such as seabirds and loons, have a larger proventriculus than gizzard,
while omnivorous waterfowl have larger gizzards (Fig. 1).

13
Structure of this thesis
Chapter 2 presents data on anthropogenic debris in the diets of freshwater birds
collected from across Canada. Digestive tracts of freshwater ducks, geese, and loons were
acquired from hunters and government employees for subsequent in-lab dissections and
dietary analyses. All methods were consistent with van Franeker and Meijboom (2002)
and sampling was carried out in accordance with Environment Canada’s Canadian
Wildlife Service Migratory Birds Regulations. We quantified debris in 350 individuals of
17 freshwater and one marine bird species collected across Canada. Collaboration with
other studies and organizations (e.g. Ducks Unlimited Canada and Nova Scotia’s
Department of Natural Resources) ensured that sampled birds served many purposes.
Chapter 3 presents data on the pathology of four species of seabirds (Cory’s
shearwater, Calonectris borealis, sooty shearwater, Puffinus griseus, great shearwater,
Puffinus gravis, and northern fulmar, Fulmarus glacialis) collected beached on Sable
Island, Nova Scotia, Canada. Beached birds offer an opportunistic window into seabird
mortality, pathology, and anthropogenic interactions (Camphuysen and Heubeck, 2001;
Roletto et al., 2003; Balseiro et al., 2005; van Franeker et al., 2011), although individuals
collected in this manor are often emaciated (Barrett et al., 2007). Beached birds often
contain ingested anthropogenic debris, yet to date little is known of the interaction among
debris, pathology, body mass, age and sex. Chapter 3 evaluated these factors, and
analyzed pathological findings among the four species.
Chapter 4 concludes the thesis by summarizing the research and results, making
further recommendations for future research, and discussing possible management
implications for this work, with a specific focus on plastic debris.

14
Figure 1. Overhead image of omnivore (duck; left) and piscivore (loon; right) digestive
tracts showing anatomical differences in structure. Images taken by author.

15
CHAPTER TWO: PLASTICS AND OTHER ANTHROPOGENIC DEBRIS IN
FRESHWATER BIRDS FROM CANADA
Holland, E., Mallory, M.L., Shutler, D., 2016. Plastics and other anthropogenic debris in freshwater birds
from Canada. Science of the Total Environment 571, 251–258. doi:http://dx.doi.org/10.1016/j.scitotenv.2016.07.158
Introduction
Humans have been releasing plastic debris into the environment since the early
1900s (Bijker et al., 1987). Originally thought to be little more than an eyesore, we now
know that the very properties that make plastics ideal for human use (i.e., being
lightweight and strong, and having a durable physical configuration) also make plastics
serious environmental hazards (Laist, 1987; Derraik, 2002). The ubiquity of
anthropogenic debris in the environment, such as plastic and waste metal, raises concerns
regarding its ingestion by animals, and so has been particularly well studied for animals
living in aquatic habitats (Rochman et al., 2014). Anthropogenic debris is problematic
due to its negative effects on wildlife, including entanglement and ingestion (Derraik,
2002; Wright et al., 2013; Provencher et al., 2014). Plastic debris also has an affinity for
certain non-essential trace elements and persistent organic pollutants (POPs), such as
polychlorinated biphenyls (PCBs) and dichlorodiphenyltrichloroethane (DDT; Ashton et
al., 2010; Bakir et al., 2014). Once plastics are discharged into aquatic environments,
they can persist for up to 50 years, and their complete mineralization may take hundreds
or thousands of years (Gregory, 1978; Derraik, 2002; Driedger et al., 2015).
Entanglement and ingestion of marine anthropogenic debris negatively affects all seven
known species of sea turtle (100%), about half of all species of marine mammals (45%),

16
and one-fifth of all species of seabirds (21%); these numbers represent a 40% increase
(from 247 to 663 affected species) from 1997 (Secretariat of the Convention on
Biological Diversity, 2012). As of 2015, 56% of seabird species were affected by marine
anthropogenic debris (Gall and Thompson, 2015), with predictions that by 2050, 99% of
seabird species will be affected (Wilcox et al., 2015) and the mass of plastics in the
oceans will outweigh fish (Neufeld et al., 2016). Whereas much is known about effects of
plastic debris on marine birds, virtually no comparable data are available for freshwater
species.
Freshwater bodies can have comparable anthropogenic debris concentrations to
marine waters (Castañeda et al., 2014; Lechner et al., 2014; Driedger et al., 2015). In the
Great Lakes of North America, over 80% of anthropogenic shoreline debris is composed
of plastics (Castañeda et al., 2014; Driedger et al., 2015) and sediments of the St.
Lawrence River have microbead (polyethylene and polypropylene microspheres widely
used in cosmetics as exfoliating agents; Eriksen et al., 2013) pollution comparable in
magnitude to marine microplastic concentrations [Castañeda et al., 2014; microplastics
defined by Moore (2008) and Arthur et al. (2008) as plastic fragments < 5 mm].
Likewise, a multiyear study on the Danube River in Austria quantified discharges of
1,533 tonnes of plastics per year into the Black Sea (Lechner et al., 2014), although the
majority turned out to be industrial microplastics from a plastic-producing company
(Lechner and Ramler, 2015). A similar study in Mongolia found that Lake Hovsgol has
plastic particle concentrations reaching 20,264 particles/km2 (Free et al., 2014), and a
recent study on two lakes in central Italy (Lake Bolsena and Lake Chiusi) found 2.68 to
3.36 particles/m3 and 0.82 to 4.42 particles/m3, respectively, in surface waters (Fischer et

17
al., 2016). These studies suggest that not only are plastics a major problem in marine
settings, they are also an issue in freshwater ecosystems.
Studies focusing on organisms in freshwater ecosystems have found dietary
plastic debris in green algae (Scenedesmus obliquus) and zooplankton (Daphnia magna)
(Besseling et al., 2014), as well as planktivorous fish (Sanchez et al., 2014; Moseman,
2015). Plastics could also potentially affect benthivorous fishes and macroinvertebrates
[preliminary results on benthic round goby’s (Neogobius melanostomus) digestive tracts
suggest the presence of microbeads (Castañeda et al., 2014) while reports on benthic
gudgeon (Gobio gobio) found ingested polymer fibers and pellets (Sanchez et al., 2014)].
Migratory birds, such as red phalaropes (Phalaropus fulicarius) and red-necked
phalaropes (P. lobatus), which eat freshwater zooplankton, also consume plastic debris
(Day et al., 1985; Moser and Lee, 1992). English et al. (2015) examined mallard (Anas
platyrhynchos), American black duck (A. rubripes), and common eider (Somateria
mollissima) wintering in Atlantic Canada, and found an 11.5% prevalence of
anthropogenic debris in 140 birds. However, it was not known whether those birds
acquired debris in freshwater or marine locations due to the long residency time of
dietary plastics (from two months to a year; Connors and Smith, 1982; Ryan and Jackson,
1987) and known movement patterns of these ducks between marine and freshwater
ecosystems in this area (English, 2016).
Encounters between organisms and marine debris have been reported since the
1960s, with the first study on seabird plastic ingestion on Laysan albatross (Phoebastria
immutabilis) conducted in 1966 (Kenyon and Kridler, 1969; Gall and Thompson, 2015).
Between 1969-1977 and 1988-1990, a significant increase (up to 26.3%) was recorded in

18
the frequency of seabird plastic ingestion (Robards et al., 1995). If trends in freshwater
waterfowl ingestion of debris mirror seabird historical trends, we may see a similar
increase in waterfowl debris consumption over time. This is problematic due to the
negative consequences of consuming debris. Debris fails to provide nutrition proportional
to its mass or volume, and can lead to weakness, false feelings of satiation, irritation of
the stomach lining, digestive tract blockage, internal bleeding, abrasion, ulcers, failure to
put on fat stores necessary for migration and reproduction, absorption of toxins, and
potential death through starvation (Moore, 2008; Wright et al., 2013; Zhao et al., 2016).
Surface-feeding birds and dabbling ducks may be particularly susceptible to
plastic ingestion due to the initial buoyancy of plastic. Plastics eventually sink over time
from biofilm fouling and hitchhiking organisms (Barnes et al., 2009; Frias et al., 2010;
Driedger et al., 2015). However, after settling, they remain available to benthic
organisms, and those that feed on benthos, and thus can return to food webs (Wright et
al., 2013). Due to biomagnification through debris consumed by fish, piscivorous birds
may also be at risk (Day et al., 1985; Castañeda et al., 2014; Sanchez et al., 2014;
Moseman, 2015). Additionally, urban birds are at an increased risk of ingesting debris
because of a greater density of plastic near industrial centers (Zbyszewski et al., 2014).
We undertook this study to bridge a knowledge gap on anthropogenic debris
ingestion by freshwater birds. We asked the following questions: 1) What is the
prevalence of anthropogenic debris in freshwater birds? 2) Is there geographic variation
in prevalence? 3) Are there differences among species in prevalence and does this relate
to their foraging niches? 4) Is prevalence related to body mass? 5) What are the
characteristics of ingested particles (i.e., type, color and size)?

19
Material and Methods
Sampling
Ducks, geese, and loons occupying freshwater habitats were collected from across
Canada (Fig. 2); 40 common eiders (a marine sea duck) were also acquired as a
comparison group. All birds were from hunter kills, airport culls or collisions, and
predation, and were shipped frozen to Acadia University where dissections were
performed. We recorded species, date, location, and if available, sex, age, and body mass
(g). Birds were kept frozen at -22°C until dissection and analysis, and allowed to thaw for
one to two days prior to dissection.
Processing, separation, sorting and identifying
Methods followed the recommendations of van Franeker and Meijboom (2002)
and van Franeker (2004) for quantifying anthropogenic debris ingestion by seabirds. To
avoid contamination, work surfaces were thoroughly cleaned with a 1/3 to 2/3 bleach and
water mixture and all tools were cleaned under running tap water between each specimen.
Gloves, lab coats, and facemasks were worn throughout the study. Sample analysis was
standardized, with the primary author inspecting all samples. Thawed digestive tracts
were opened over their full length, and contents carefully flushed with cold tap water
above a 0.5-mm mesh sieve to ensure that no small particles were left behind on organ

20
walls (particles smaller than 0.5 mm were detected due to debris’ ability to adhere to
larger dietary particles). All material was rinsed under running tap water (van Franeker
and Meijboom, 2002). Proventriculus and gizzard tissues were examined for
inflammation, abrasion, or swelling from exposure to debris. Care was taken to note any
indications of damage from birdshot. Proventriculus and gizzard contents were
transferred to a Petri dish, and inspected under a dissecting microscope (AmScope SM-
2BZ) as follows:
a) Anthropogenic debris was identified (per Desforges et al., 2015 and Zhao et al.,
2016) if: i) no cellular or organic structures were visible; ii) fibers were
uniformly thick over their length and not tapered at the end, bendable, or soft; iii)
colored items were homogeneously colored and of hues not usually occurring in
food; iv) debris had unnatural edges of obvious anthropogenic origin. All
potential microscopic anthropogenic debris was re-examined with extra care and
under higher magnification (4.5× zoom objective). Once classified as artificial
material, particles were transferred into Petri dishes and ready for step b). If there
was an indication of birdshot damage to digestive organs, shot was assumed to
have been hunting-related, and not consumed, and was therefore not included in
analysis as contaminant debris.
b) Anthropogenic debris was classified following van Franeker and Meijboom
(2002): plastic (fragments and other), non-plastic rubbish (thread-like, foil, paint
chips, glass, and rubber), and metal (birdshot and metal fragments).

21
Anthropogenic debris was classified as either light (clear-white, yellow, light
green-blue, pink-tan) or dark (red-orange, dark green, dark blue-purple, brown-
black; Day et al., 1985; Moser and Lee, 1992). After sorting contents under a
dissection microscope, items were counted for each bird. Each particle was
measured (in mm) for length, width, and height or diameter (only for round
debris, and measured where widest). Debris was photographed for subsequent
analyses. Once confirmed, anthropogenic debris items were removed from a
sample, and remaining items were categorized as natural.
Statistical analyses
Previous studies using power analyses have generally found that a minimum of 18
to 40 or more individuals per species are required for reliable estimates of intraspecific
prevalence (van Franeker and Meijboom, 2002; Provencher et al., 2015), so we used a
minimum sample size of 18 individuals to include a species in interspecific comparisons.
We used Fisher exact tests for most comparisons because >20% of the cells had expected
counts of less than five (Cochran, 1954). We used logistic regression to test if debris
presence was related to interspecific and intraspecific variation in body mass. Statistical
analyses were performed in program R (version 3.2.2; R Core Team, 2015) using the
“dplyr” package (Wickham and Francois, 2015). Values are reported as mean ± SD
unless otherwise stated.

22
Results
We obtained data from 350 birds of 18 species, including three herbivorous geese,
eight omnivorous dabbling ducks, three omnivorous diving ducks, one carnivorous sea
duck, and three piscivorous loons (Table 1; all diving duck and loon species sampled are
partially marine species that breed on inland freshwater bodies. All individuals of these
species in this study were culled inland). Because we recovered no debris from the
proventriculus (nor have others, e.g., Day et al., 1985; Moser and Lee, 1992; Robards et
al., 1995), hereafter we focus only on the gizzard. We found 110 items of anthropogenic
debris, ranging in size from 50 μm to 5 mm (hence all plastic debris would be classified
as microplastics; Arthur et al., 2008; Moore, 2008; Ivar do Sul and Costa, 2014), in 10 of
18 (55%) species, and 40 of 350 (11.4%) birds, with an average of 0.31 (± 3.1) items per
bird. Sample sizes for the eight species that did not have ingested anthropogenic debris
were all < 15, so their anthropogenic debris prevalence of zero should be interpreted
cautiously. There was no difference in debris ingestion among foraging niches (Table 1;
Fisher exact test, p > 0.99).
Of the 10 species with ingested debris, five had the minimum recommended
samples of 18 (Table 1). These species had similar prevalences of anthropogenic debris
(Fisher exact test, p = 0.50). For species with the recommended minimum sample sizes
[we had mass data for < 12 Canada geese (Branta canadensis) and American wigeon
(Anas americana)], interspecific variation in body mass was not associated with
anthropogenic debris ingestion (logistic regression, t1, 99 = 0.5, p = 0.64) nor was
intraspecific variation in body mass associated with debris ingestion (three species, all N

23
for individuals with body mass > 24, all t < 0.34, all p > 0.60]. A more detailed
breakdown is presented in Table 2 (also see Appendix A), but is not amenable to
statistical analysis.
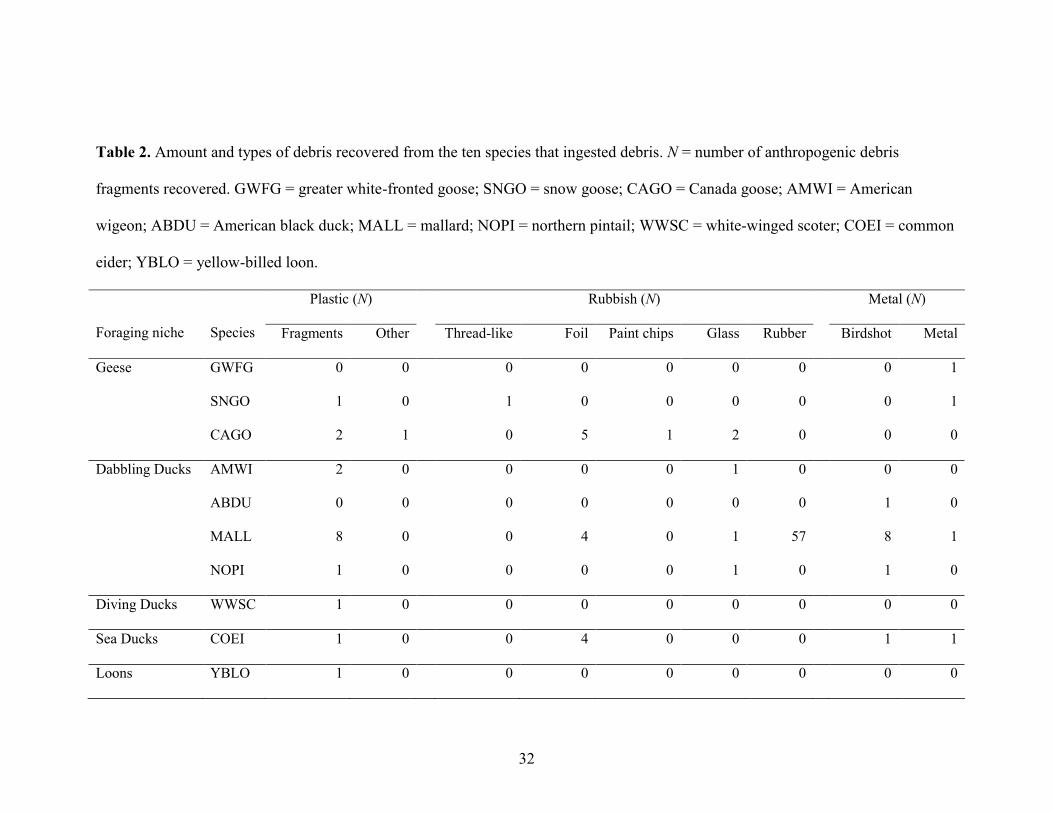
Plastic debris was present in eight species and 15 of 350 (4.3%) birds; prevalence
did not differ among species with sufficient samples (Fisher exact test, p = 0.85). Non-
plastic rubbish was present in six species, and ingested by 13 of 350 (3.7%) birds. Non-
plastic rubbish prevalence did differ among species with sufficient samples [Fisher exact
test, p = 0.01; Canada goose (7/43, 16.3%), American wigeon (1/32, 3.1%), mallard
(3/120, 2.5%), common eider (1/40, 2.5%), snow goose (Chen caerulescens; 1/47,
2.1%)]. Metal debris was present in six species, and ingested by 12 of 350 (3.4%) birds,
and prevalence did not differ among species with sufficient samples (Fisher exact test, p
= 0.48). All ingested birdshot was a non-toxic alternative to lead shot, such as steel, based
on uncrushed, rounded shapes of recovered pellets. One northern pintail (Anas acuta) had
17 lead birdshot pellets in its gizzard, but was presumably hunted with this shot. Pellet
composition was determined visually and with a simple crush test. Lead pellets tend to be
deformed and fragmented upon impact with soft tissues and bone, whereas steel shot
usually remains round (Wilson, 1999; Peitzman et al., 2012). The remaining metal
fragments were metalworking waste (swarf).
Prevalences of anthropogenic debris in British Columbia (23/145, 15.9%), Nova
Scotia (6/74, 8.1%), Northwest Territories (5/66, 7.6%), Newfoundland (3/29, 10.3%),
Ontario (1/19, 5.3%) and New Brunswick (0/13) (Fig. 2) were not statistically different
(Fisher exact test, p = 0.30). Of four common eiders without accompanying information
on origin, one had ingested debris.

24
Ingested particle coloration was classified for all debris except birdshot (7/39,
17.9%), because we assume birds encounter this by accident and it becomes retained as if
it were grit (a mixture of mineral, rock, and hardened food fragments retained to aid in
fibrous food digestion; Thomas et al., 1977). Light-colored anthropogenic debris was
more commonly ingested (27/32, 84.2%) than dark colored (5/32, 15.6%). Clear and
white debris were the most common colors ingested by all species (11/32, 34.4%). Gold
was the second most commonly ingested color (4/32, 12.5%), and black was the least
common (1/32, 3.1%). All other colors (light yellow, light green-blue, dark green, dark
blue-purple) were ingested with the same frequency (2/32, 6.3%).
Discussion
Prevalence of anthropogenic debris in freshwater birds and the marine common
eider comparison group presented in this study (11.4%) provides compelling evidence
that freshwater and marine organisms currently face similar threats from anthropogenic
debris ingestion. Although our sample was mostly limited to commonly hunted or culled
species, our results suggest that anthropogenic debris ingestion by freshwater birds is
likely to apply to a wider range of species, because anthropogenic debris ingestion was
found in some species for which we had small samples. We expect that sampling of other
waterfowl and freshwater bird species (such as herons and kingfishers) will likely reveal
ingestion of anthropogenic debris.
Seabird plastic ingestion is assumed to occur because plastics mimic natural food
items (Day et al., 1985; Moser and Lee, 1992). Although freshwater birds may mistake
plastic debris as food, the high prevalence of birdshot (which they encounter

25
opportunistically after it sinks into sediment, and does not appear to mimic any known
food items) suggests freshwater birds may be retaining birdshot as grit (Thomas et al.,
1977; Moore et al., 1998). They could also acquire small, broken pieces of plastic debris
in this manner. Some seabirds consume specific plastic shapes and colors more
frequently, and debris ingestion ties into foraging niche and availability of certain plastics
in particular habitats (Day, 1980; Day et al., 1985; Ryan, 1987; Moser and Lee, 1992).
Surface-feeding species are most likely to have plastics in their diets, perhaps due to
polyethylene’s specific gravity of 0.9, enabling plastics to float at the water’s surface
(Snyder and Vakos, 1966; Day et al., 1985; Moser and Lee, 1992). Although our study
supports previous evidence that birds preferentially ingest lighter colored debris
(Rothstein, 1973; Day, 1980; Moser and Lee, 1992; Lavers and Bond, 2016), we cannot
evaluate whether this reflects selective uptake without knowing availability in the
environment. However, we did not find evidence that debris ingestion was related to
foraging niche.
Anthropogenic debris ingestion by freshwater birds should also be an important
issue to waterfowl hunters. In 2013 alone, approximately 189,844 individuals across
Canada hunted approximately 2,286,951 waterfowl (Environment and Climate Change
Canada, 2016; Gendron and Smith, 2016). Given that debris can vector various
contaminants (Teuten et al., 2007; Ashton et al., 2010; Rios et al., 2010) this may put
hunters such as Aboriginal peoples, who rely most heavily on wild foods (Van Oostdam
et al., 1999; Johansen et al., 2001; El-Hayek, 2007), at risk of consuming contaminated
tissues. Quantifying plastic-associated toxins is crucial to understanding potentially

26
hidden effects of anthropogenic debris ingestion on Aboriginal peoples and other
vulnerable groups, and to developing future avian conservation plans.
In contrast to marine anthropogenic debris, microscopic anthropogenic debris in
freshwater ecosystems may be perceived as an environmental issue that is closer to home,
hopefully resulting in more scientific and public attention. One example of this is the
recent passing of the Microbead Free Waters Act of 2015 in the United States (House
Report No. 114-371, 2015). Microbead use has increased in recent years, and their
relatively small diameter (< 1 mm) means many wastewater treatment plants cannot
remove them, leading to an increase of these plastics entering aquatic ecosystems
(Castañeda et al., 2014; Doughty and Eriksen, 2014; Driedger et al., 2015). The resultant
increase in microbead concentrations in waterways led to public outcry, and shortly
thereafter many major companies banned microbeads (Newman et al., 2013). Although
care was taken in our study to find microbeads, none were recorded. This could be due to
the sieves used. Our finest was a 0.5 mm mesh, the same size used by (Castañeda et al.,
2014) to sieve microbeads from river sediment. However, despite our similar sieve size,
we found that running a microbead-containing product (Clean & Clear® morning burst®
facial scrub) through our sieve permitted passage of smaller beads, retaining only less
frequent larger beads. Similarly, larger beads could have been ground down over time in
gizzards, permitting their passage through our sieve. Therefore our microbead findings
should be interpreted cautiously, because waterfowl are likely ingesting them.
Our results on the prevalence of anthropogenic debris ingestion in waterfowl
indicate that it is occurring at similar rates to historic trends in in some marine birds
(Laist 1997) and current rates in some freshwater birds (English et al., 2015). Future

27
studies examining gut contents of freshwater birds should adopt a screening method for
anthropogenic debris similar to that of van Franeker and Meijboom (2002) and van
Franeker (2004), and collection points for examination of harvested bird digestive tracts
should be established. This could be essential in monitoring anthropogenic debris
ingestion over a number of years to reliably assess trends.
Conclusions
Our study adds to the limited but mounting evidence (Moser and Lee, 1992;
Denuncio et al., 2011; Besseling et al., 2014; Sanchez et al., 2014; English et al., 2015;
Moseman, 2015; Fischer et al., 2016) that anthropogenic debris may be a threat to aquatic
biota in freshwater environments. We found debris in 55% of species collected from
freshwater habitats in Canada, including from remote sites as far as 63ºN. However, there
was no suggestion of patterns in anthropogenic debris ingestion relative to body mass,
geographic location of capture, or foraging niche. This was surprising, because we
expected that birds collected near urban or industrial centers (where debris may occur at
higher densities; Zbyszewski et al., 2014) or those foraging as carnivores or omnivores
might be more likely to ingest anthropogenic debris. We did not acquire many samples
from the Great Lakes region, where research has shown significant pollution by plastic
(Eriksen et al., 2013; Castañeda et al., 2014; Driedger et al., 2015), and thus we expect
that greater sampling effort of birds wintering there will reveal higher prevalence of
ingested plastic and other debris. Consequently, we suggest that our data represent a
conservative baseline of anthropogenic debris ingestion in waterbirds in Canada and we

28
expect that additional studies will confirm debris ingestion in other species, as has been
shown in marine birds (Provencher et al., 2015).
Although there is evidence that anthropogenic debris is a threat to aquatic biota,
there is still a need for long term monitoring to provide input for conservation
management, to strengthen the basis for educational campaigns, and to provide scientists
with better evidence that could be used to increase efforts to mitigate the problem
(Derraik, 2002). Our baseline data provide insights suggesting that this may have to occur
sooner than expected to prevent waterfowl debris ingestion levels from reaching the
levels currently observed in seabirds.

29
Figure 2. Sample sites used for this study. Circles indicate collection locations. BC:
British Colombia, NT: Northwest Territories, ON: Ontario, NB: New Brunswick, NS:
Nova Scotia, NL: Newfoundland.

30
Table 1. Sample sizes, body mass ± SE (g), the frequency, and mean number of pieces ± SE for plastic, metal, and any debris. N =
number of specimens dissected. Means include birds without ingested debris. GWFG = greater white-fronted goose; SNGO = snow
goose; CAGO = Canada goose; GADW = gadwall; AMWI = American wigeon; ABDU = American black duck; MALL = mallard;
BWTE = blue-winged teal; NOSH = northern shoveler; NOPI = northern pintail; GWTE = green-winged teal; LESC = lesser scaup;
WWSC = white-winged scoter; LTDU = long-tailed duck; COEI = common eider; RTLO= red-throated loon; COLO = common loon;
YBLO = yellow-billed loon.
Mean body
mass ± SE (g)
Ingestion frequency (%) Mean pieces of debris/bird ± SE
Foraging niche Species N Plastic Rubbish Metal Plastic Rubbish Metal
Geese GWFG 2 1616 ± 203 0.0 0.0 50.0 0.00 ± 0.00 0.00 ± 0.00 0.50 ± 0.71
SNGO 47 2119 ± 423 2.1 2.0 2.1 0.02 ± 0.15 0.02 ± 0.15 0.02 ± 0.15
CAGO 43 3584 ± 241 4.7 14.0 0.0 0.05 ± 0.30 0.21 ± 0.51 0.00 ± 0.00
Dabbling Ducks GADW 2 836 ± N/A 0.0 0.0 0.0 0.00 ± 0.00 0.00 ± 0.00 0.00 ± 0.00
AMWI 32 738 ± 102 6.3 3.1 0.0 0.06 ± 0.25 0.03 ± 0.18 0.00 ± 0.00
ABDU 5 1242 ± N/A 0.0 0.0 20.0 0.00 ± 0.00 0.00 ± 0.00 0.25 ± 0.50
MALL 120 1185 ± 178 5.0 2.5 5.0 0.07 ± 0.34 0.52 ± 5.21 0.08 ± 0.37
BWTE 1 N/A 0.0 0.0 0.0 0.00 ± N/A 0.00 ± N/A 0.00 ± N/A

31
Mean body
mass ± SE (g)
Ingestion frequency (%) Mean pieces of debris/bird ± SE
Foraging niche Species N Plastic Rubbish Metal Plastic Rubbish Metal
NOSH 1 N/A 0.0 0.0 0.0 0.00 ± N/A 0.00 ± N/A 0.00 ± N/A
NOPI 10 762 ± 50 10.0 10.0 10.0 0.10 ± 0.32 0.00 ± 0.00 0.20 ± 0.63
GWTE 15 325 ± N/A 0.0 0.0 0.0 0.00 ± 0.00 0.00 ± 0.00 0.00 ± 0.00
Diving Ducks LESC 1 N/A 0.0 0.0 0.0 0.00 ± N/A 0.00 ± N/A 0.00 ± N/A
WWSC 16 N/A 6.3 0.0 0.0 0.06 ± 0.25 0.00 ± 0.00 0.00 ± 0.00
LTDU 4 N/A 0.0 0.0 0.0 0.00 ± 0.00 0.00 ± 0.00 0.00 ± 0.00
Sea Ducks COEI 40 1825 ± 126 2.5 2.5 5.0 0.02 ± 0.16 0.10 ± 0.63 0.05 ± 0.22
Loons RTLO 7 N/A 0.0 0.0 0.0 0.00 ± 0.00 0.00 ± 0.00 0.00 ± 0.00
COLO 1 N/A 0.0 0.0 0.0 0.00 ± N/A 0.00 ± N/A 0.00 ± N/A
YBLO 3 N/A 33.3 0.0 0.0 0.33 ± 0.58 0.00 ± 0.00 0.00 ± 0.00
32
Table 2. Amount and types of debris recovered from the ten species that ingested debris. N = number of anthropogenic debris
fragments recovered. GWFG = greater white-fronted goose; SNGO = snow goose; CAGO = Canada goose; AMWI = American
wigeon; ABDU = American black duck; MALL = mallard; NOPI = northern pintail; WWSC = white-winged scoter; COEI = common
eider; YBLO = yellow-billed loon.
Foraging niche
Species
Plastic (N) Rubbish (N) Metal (N)
Fragments Other Thread-like Foil Paint chips Glass Rubber Birdshot Metal
Geese GWFG 0 0 0 0 0 0 0 0 1
SNGO 1 0 1 0 0 0 0 0 1
CAGO 2 1 0 5 1 2 0 0 0
Dabbling Ducks AMWI 2 0 0 0 0 1 0 0 0
ABDU 0 0 0 0 0 0 0 1 0
MALL 8 0 0 4 0 1 57 8 1
NOPI 1 0 0 0 0 1 0 1 0
Diving Ducks WWSC 1 0 0 0 0 0 0 0 0
Sea Ducks COEI 1 0 0 4 0 0 0 1 1
Loons YBLO 1 0 0 0 0 0 0 0 0

33
CHAPTER THREE: ANTHROPOGENIC DEBRIS AND PATHOLOGY OF
FULMARS AND SHEARWATERS BEACHED ON SABLE ISLAND, NOVA
SCOTIA, CANADA
Introduction
Throughout the world seabirds are frequently found dead on beaches, with
recorded reports from Europe as early as the late 19th century (Gray, 1871; Stephen and
Burger, 1994; Furness and Camphuysen, 1997). These “beached bird” carcasses can
provide an opportunistic window into seabird diets and causes of death (Barrett et al.,
2007). However they may be biased, often revealing that beached birds are primarily
emaciated individuals which have likely starved to death (Barrett et al., 2007). Autopsies
of beached birds can provide information on causes of death, diet prior to death, and
other deleterious health conditions (Camphuysen and Heubeck, 2001; Roletto et al.,
2003; van Franeker et al., 2011). Likewise, beached bird surveys can record any large
species die offs or sudden influxes of dead oiled birds (Camphuysen and Heubeck, 2001;
Roletto et al., 2003; Balseiro et al., 2005). This information is important for monitoring
disease outbreaks and quantifying impacts of human activities (Camphuysen and
Heubeck, 2001; Balseiro et al., 2005).
Although the majority of seabird mortality is likely natural, and related to
biotoxins, infectious diseases, emaciation, trauma, and predation, anthropogenic causes of
mortality may also be responsible (Newman et al., 2007). One commonly recorded
anthropogenic cause of seabird death is entanglement in and ingestion of marine

34
anthropogenic debris (Moser and Lee, 1992; Newman et al., 2007). With respect to the
latter, debris, most commonly plastic, is problematic due to concerns about its toxicity,
digestive tract blockage, internal abrasion, and attendant starvation (Moore, 2008; Wright
et al., 2013). Written records of humans releasing anthropogenic debris into the
environment exist since the early 1900s (Klar et al., 2014), and studies on seabird debris
ingestion have been conducted since the 1960s (Kenyon and Kridler, 1969). Surface-
feeding seabirds, such as fulmars and shearwaters, may be particularly susceptible to
plastic ingestion because plastics can float at the water surface and appear to be food
(e.g., specific gravity of polyethylene is 0.9; Day et al., 1985; Snyder and Vakos, 1966).
Fulmar and shearwater species are long-lived (> 40 years; Michel et al., 2003),
philopatric (86% per annum rate of nest-site return; Macdonald, 1977), largely pelagic
seabirds, which only come to land only to breed (Powers, 1984). However, many pass by
islands on the Atlantic Coast during migration, and birds that die at sea occasionally wash
onto shores of islands (Bond et al., 2014). Sable Island is situated 160 km east of Halifax,
Nova Scotia (Canada; 43.9337° N, 59.9149° W). Sable Island is 45 km long, crescent-
shaped, and sandy with over 64 species of beached seabirds collected since the 1970s
(Lucas et al., 2012). Sable Island is a potentially highly valuable monitoring site because
large numbers of Procellariiformes are encountered dead regularly on the beaches (Lucas
et al. 2012). To date, no information exists on pathologies of birds beached on Sable
Island, because most published studies deal with anthropogenic causes of death, such as
oiling and anthropogenic debris ingestion (Bond et al., 2014; Lucas and MacGregor,
2006). Although beached bird surveys have been conducted on Sable Island since the
1970s, surveys to determine persistent coastal litter only started in the early 1980s

35
(Gregory, 1983; Lucas, 1992). These surveys found that the majority of total
anthropogenic material present on Sable Island’s beaches was plastic (Gregory, 1983;
Lucas, 1992). In fact, up to 93% of northern fulmars (Fulmarus glacialis) beached on
Sable Island have ingested plastic (Bond et al., 2014).
Across their broad range, high rates of plastic ingestion by northern fulmar are
well documented, so much so that these birds are used as an indicator species on the state
of the North Sea ecosystem (van Franeker, 2004). Current Ecological Quality Objective
targets have been established in the North Sea of no more than 10% of fulmars having >
0.1 g of ingested plastic (OSPAR Commission, 2010). However, relatively recent surveys
showed that 48–78% of fulmars exceed this goal (van Franeker et al., 2011).
Northern fulmars have variability in plumage coloration, and this coloration may
be related to their breeding success and life history (Hatch, 1991). A four color phase
system is used to classify fulmar plumage morphs: single light, double light, single dark,
and double dark (O’Reilly, 1818; Fisher, 1952; van Franeker and Wattel, 1982).
However, in practice the main distinction is made between the double light color phase
and the darker color phases (van Franeker, 2004; van Franeker and Luttik, 2008). Double
light individuals make up nearly 100% of southern subspecies in the temperate breeding
range, and a variable proportion (0-10%) of Arctic populations (van Franeker, 2004; van
Franeker and Luttik, 2008). Previous studies have found no statistical differences in
stomach plastic content between the different color morphs (Trevail et al., 2015).
However, these studies examined only individuals from the single light, single dark, and
double dark color phases (Trevail et al., 2015). The lightest fulmar morphs (double light)
have a more southerly distribution (Hatch, 1991); thus they are likely exposed to more

36
debris, and therefore may consume debris at a greater frequency (van Franeker and Law,
2015).
Although not as well studied as northern fulmars, plastic ingestion among
shearwater species has also been documented, with 83% of Cory’s shearwater
(Calonectris borealis), 72% of sooty shearwater (Ardenna grisea), and 88% of great
shearwater (Ardenna gravis) having ingested plastic (Rodríguez et al., 2012; Bond et al.,
2014). Comparisons to northern fulmar show that great shearwater did not differ in the
proportion of ingested debris, whereas sooty shearwaters had a lower frequency of debris
ingestion (Bond et al., 2014).
Beached birds of all four of the preceding seabird species were collected on Sable
Island, Nova Scotia, Canada. Sooty shearwaters, great shearwaters, and northern fulmars
are found beached regularly on Sable Island, whereas Cory’s shearwaters rarely range
into Canadian waters (Lucas et al., 2012). Between 1993 and 2009, only 14 of the latter
species were recorded beached on Sable Island (Lucas et al., 2012), and during our
collection period only three birds were collected. Due to this small sample size reliable
analyses could not be run on this species independently of the others.
The purpose of this study was to quantify pathological findings of beached birds
of these species, as well as to asses the time of year, gender, and relative age of birds
beaching on Sable Island, determine if pathology was associated with these variables, and
compare nutritional condition of these birds in relation to mass of anthropogenic debris
ingested.

37
Materials and Methods
Sample collection and processing
Intact procellariid carcasses were collected from the entire Sable Island shoreline
between July 2000 and January 2012. The majority of birds were collected during
beached bird surveys conducted every 25-45 days (weather and beach conditions
permitting), with the longest interval between surveys being 96-days (additional details in
Lucas et al., 2012). The remainder of birds were gathered opportunistically when
encountered on the beach (Lucas et al., 2012). Carcasses were frozen and stored on the
island prior to shipment to the Department of Pathology and Microbiology, Atlantic
Veterinary College, University of Prince Edward Island in Charlottetown, Prince Edward
Island, Canada. Carcasses were then dissected at the Atlantic Veterinary College, where
the upper gastrointestinal tract was removed, ingested anthropogenic material was
quantified, and pathology was determined (see Bond et al., 2014 for further detail).
Gender and age (immature or adult) were determined based on the criteria
described by van Franeker (2004). Carcasses were examined for external lesions, debris
entanglement, or other signs of trauma. Emaciation was characterized by severe atrophy
of the pectoral muscles, complete absence of subcutaneous and/or abdominal fat deposits,
and serious atrophy of the pericardial fat. Samples for histopathology were collected from
the brain, heart, lung, gizzard, liver, kidney, bursa, adrenal, ovary or testis, and skeletal
system. Liver, kidneys, lungs, intestine, and duodenum swabs were submitted for

38
bacteriological investigations to the Department of Pathology and Microbiology at the
Atlantic Veterinary College.
Statistical analyses
Previous studies using power analyses have generally found that a minimum of 18
to 40 or more individuals per species are required for reliable estimates of intraspecific
prevalence of debris ingestion (van Franeker and Meijboom, 2002; Provencher et al.,
2015). Chi-squared and Fisher exact tests (the latter when expected values were less than
5 for any cell) were used to test for relationships between the explanatory variables age
and gender and the response variable pathology, categorized as emaciated or ‘other’
(representing all pathology other than emaciation and pooled because of small samples
sizes) (Cochran, 1954; Larntz, 1978). Generalised Linear Models (GLM; Venables and
Ripley, 2002) were used to test for relationships between pathology (emaciated or
‘other’), individual mass (heavily scavenged or soiled carcasses were not included), and
mass of ingested debris, for all species and within species if sample sizes were sufficient.
We evaluated whether the mass of debris consumed was negatively associated
with body condition, through a principle components analysis (PCA). To do this we first
calculated a correlation matrix of structural measurements which was used to generate PC
scores of “size” (Wold et al., 1987) for northern fulmars on five structural variables:
culmen, bill depth, total head, tarsus, and wing (Mallory and Forbes, 2005). On the
remaining three species, metrics of structural measurements were not recorded. We used
PC1 scores for each fulmar as a measure of body size and regressed mass on PC1 to

39
determine body condition (Kirk and Gosler, 1994). This metric of condition was then
regressed against mass of debris ingested (Blanco et al., 1997).
Means are reported ± standard deviation unless noted otherwise. Statistical
analyses were performed in R (version 3.2.2; R Core Team, 2015) using the “dplyr” and
“broom” packages (Wickham and Francois, 2015; Robinson, 2016).
Results
We gathered data from 318 seabird carcasses (Table 3). The majority of birds
recovered were northern fulmars (55%) and great shearwaters (28%), and were primarily
immature birds of all species (222/318; 70%). Most birds were collected between March
and July when regular surveying due to stable beach conditions was possible (Fig. 3).
Winter conditions on Sable Island often result in narrow beach profiles, eroded peat,
irregular beach topography, decreased visibility, post-storm flooding, and an increased
prevalence of beach debris, all of which affect detectability of beached birds and
increased the intervals between surveys (Lucas et al., 2012)
Pathological symptoms (defined in Table 4) were found for 251 (79%) of the 318
collected individuals, with most birds displaying symptoms of emaciation (54%),
followed by autolysis (8%), parasite infection (3%), inflammation (3%), trauma (3%),
bacterial infection (2%), drowning (2%), tumors (1%), tissue necrosis (1%), egg and
rectal impaction (1%), myopathy (1%), or pneumonia (1%) (some additional pathological
and histological details are given in Appendix B). Northern fulmars washed up dead of

40
emaciation less frequently (69/115; 60%) than sooty (36/48; 75%) or great (67/86; 78%)
shearwaters (χ21 = 8.36, p = 0.02; Cory’s shearwater not tested due to small sample size).
Sooty Shearwater
Of 50 sooty shearwaters, pathological symptoms were found in for 48 (96%;
Table 4). Pathology was not related to sex (Fisher's exact test; p = 0.44) or age (p > 0.99).
Likewise, pathology was not related to the mass of debris (0.07 ± 0.17 g; Bond et al.,
2014) an individual had ingested (R2 = 0.01, F1, 45 = 0.50, p = 0.48), or body mass (R2 =
0.07, F1, 37 = 2.80, p = 0.10). Mass of debris ingested was also not related to body mass
(R2 < 0.01, F1, 38 = 0.1, p = 0.78; Table 5).
Great Shearwater
Pathology was found in 86 of 89 (97%) of great shearwaters (Table 4). Pathology
was not related to sex (χ21 < 0.1, p = 0.91) or age (p = 0.78). Likewise, pathology was not
related to the mass of debris (0.17 ± 0.33 g; Bond et al., 2014) an individual had ingested
(R2 < 0.01, F1, 78 = 0.37, p = 0.54), or individual mass (R2 = 0.02, F1, 75 = 1.19, p = 0.28).
Mass of debris ingested was also not related to mass of the individual (R2 < 0.01, F1, 71 =
0.3, p = 0.58; Table 5).

41
Northern Fulmar
Of 176 carcasses of northern fulmars, pathology was found in 115 (65%; Table
4). Pathology was not related to sex (χ21 = 0.2, p = 0.68) or age (χ2
1 < 0.1, p = 0.91).
Likewise, pathology was not related to the mass of debris (1.09 ± 1.93 g; Bond et al.,
2014) an individual had ingested (R2 < 0.01, F1, 113 = 0.02, p = 0.88), or individual mass
(R2 = 0.04, F1, 73 = 3.32, p = 0.07), and mass of debris ingested was not related to mass of
the individual (R2 < 0.01, F1, 113 = 0.1, p = 0.74; Table 5).
The first principal component (PC1) of the five morphometric variables had
loadings ranging from -0.51 to -0.35 (eigenvalue 2.83) and accounted for 57% of the total
original variance, more than expected by chance (Jackson, 1993). Residuals saved from
regressions on PC1 against body mass showed that body condition was not related to
mass of debris ingested (F1, 91 = 0.1, p = 0.80).
Fulmar plumage was grouped into two categories, double light and single dark,
because no single light birds were collected, and only one double dark individual was
collected. Double light birds made up the majority of beached birds (169/176; 96%).
There was no statistically significant difference in mass of debris ingested by fulmars of
these two morphs (Fig. 4; ANOVA F1, 174 = 1.5, p = 0.23).

42
Discussion
This is the first systematic pathological investigation of beached seabirds from
Sable Island, and builds on the detailed surveys on age and type of beached birds that
have previously been conducted (Lucas et al., 2012). The birds we analyzed had limited
physiological or tissue trauma, perhaps less than one might associate with high levels of
anthropogenic debris ingestion (Pierce et al., 2004). This suggests that post-mortem
dietary analyses of beached birds are a useful approach to monitoring ingestion amounts
and rates, but that ascertaining causes of mortality, or attributing them to debris, is more
difficult. This is especially true because most beached birds collected have likely died of
starvation.
The northern fulmar is an indicator species of the state of the North Sea
ecosystem within the current Ecological Quality Objectives (van Franeker, 2004; OSPAR
Commission, 2010), and thus it would be beneficial to know common causes of
mortality. Although three of the species studied (northern fulmar, Cory’s shearwater, and
great shearwater) are listed as Least Concern by the IUCN (BirdLife International, 2016a,
2016b, 2016c), the sooty shearwater is listed as Near Threatened, with a population trend
designated as decreasing (BirdLife International, 2016d). This designation should prompt
further studies into common causes of death in this species, both natural and
anthropogenic.
Northern fulmars were emaciated less often than sooty and great shearwaters. It’s
unlikely this is due to age, because 84% of sooty shearwaters were juvenile, a larger
proportion than northern fulmars (70%) and great shearwater (69%). A contributor to this

43
disparity may be species movements and home ranges, because northern fulmars are
transatlantic migrants, breeding in the Canadian Arctic and wintering in the northwest
Atlantic Ocean (Mallory et al., 2008), whereas both shearwater species are
transequatorial migrants that breed in the southern hemisphere (Huettmann and Diamond,
2000). As a group, the shearwater’s longer migratory pathways may be associated with
greater energetic stresses and more frequent emaciation. Fulmars could also encounter
conditions that lead them to die suddenly and not become emaciated. This may include
high organochloride tissue contamination. Fulmars have higher rates of plastic ingestion
than shearwaters, and fulmar muscle, fat, and eggs contain PCBs, DDTs,
hexachlorobenzene (HCB), and polybrominated diphenyl ethers (PBDEs) (Fängström et
al., 2005). Further research into the differences in rates of emaciation between these three
species is warranted.
For Sable Island, mortality rates and pathology of marine birds were not available
for comparison to our results. However, the main pathological findings, emaciation and
autolysis, were not unexpected in a sample composed of primarily immature beached
birds (Barrett et al., 2007). These findings attest to the challenges birds face surviving at
sea, especially weak, young, and inexperienced animals. Most parasitic and bacterial
infections in these birds had been recorded previously in these four species, as had many
cases of inflammation, trauma, and tumors (Appendix C). Avian influenza, a disease of
recent concern (Lang et al., 2016), was not found in any of the specimens that we
examined. Similarly, we did not detect avian cholera, despite recent outbreaks of this
disease in larids of this region (Wille et al., 2016).

44
Pathological symptoms we reported have been recorded in other seabird species.
Autolysis is often reported in beached birds, although in many cases it has likely occurred
post-mortem (Stephen and Burger, 1994; Harris et al., 2006). Capture myopathy has been
reported in many seabird species (Newman et al., 1997) but has not previously been
noted in beached birds. Multi-organ, including hepatic, necrosis has been recorded in
wedge-tailed shearwater (Puffinus pacificus), although we found no previous reports of
tissue necrosis for sooty shearwater, great shearwater, or northern fulmar (Work and
Rameyer, 1999). Likewise, there are no mentions of sarcocystosis in great shearwater and
northern fulmar in the literature; however, the protozoan Sarcocystis has been reported in
common murre (Uria aalge) (Muzaffar and Jones, 2004). Trematodes and pododermatitis
in northern fulmar have not previously been reported; however other seabird species have
had these symptoms (Haman et al., 2013; Bogstad et al., 2014). Septicemia has been
recorded in many seabird species previously, but always in relation to rehabilitation
(Lieske et al., 2002; Steele et al., 2005). Mycobacteriosis has not previously been
recorded in seabirds, but has been in dunlin (Calidris alpina) and oystercatchers
(Haematopus ostralegus) (Pennycott, 2016).
Some novel pathological findings we report may have been observed before (e.g.,
unpublished findings, or those outside of indexed peer-reviewed journals and
publications in languages other than English). Some of these pathologies may have been
difficult to detect because beached bird surveys often yield decaying and/or heavily
scavenged carcasses, making necropsies difficult (Stephen and Burger, 1994). Carcass
mass can also be difficult to accurately ascertain in cases where birds are wet, covered in
sand, or scavenged.

45
Data were clearly biased towards immature and emaciated individuals as well as
double light birds. The majority of birds were collected in spring and summer due to
decreased winter sampling, so reliable tests could not be run on these data. Due to our
sample sizes, ages, seasons and year were pooled, which could have obscured patterns.
For all three species with sufficient data, pathology was neither related to the
mass of debris ingested nor body mass, and mass of debris ingested was unrelated to
body mass. This was surprising because previous studies have found that heavier birds
were more likely to contain ingested plastics (Spear et al., 1995; Verlis et al., 2013).
Body condition could only be calculated for fulmars, and despite some prior studies
finding that birds with higher amounts of ingested plastic had reduced body condition
(e.g., Ryan, 1987), we found no support for this prediction. However, this is consistent
with Avery-Gomm et al. (2016) who found no connection between dovekie (Alle alle)
body condition and mass of plastic ingested. Our findings could be due to debris amounts
being below thresholds known to affect a bird, although data for these thresholds are hard
to determine. Current thresholds are only available in the literature for northern fulmars
and the Ecological Quality Objectives (OSPAR Commission, 2010; Bond and Lavers,
2013). These objectives are arbitrary, as no biologically meaningful level has yet been
established (OSPAR Commission, 2010).
Because anthropogenic debris is increasing in the world’s oceans, even species
with northern ranges or in seemingly remote locations are being exposed (Provencher et
al., 2015; Trevail et al., 2015; Amélineau et al., 2016). Recent reports of microplastics
have been confirmed in Arctic waters in concentrations comparable to those of other
oceans, and recent studies have documented plastic ingestion by Arctic birds (Provencher

46
et al., 2015; Trevail et al., 2015; Amélineau et al., 2016). Studies of beached bird
mortalities, pathology, and debris ingestion are important for tracking trends among
endangered and non-threatened species. Without these baseline studies, it is hard to
determine if threats facing wild populations are sustainable long term, or emerging and
deadly to long-term species survival. Because Sable Island is the only site in Atlantic
Canada where large numbers of Procellariiformes are encountered regularly (Lucas et al.,
2012), it could be an important monitoring site for marine anthropogenic debris, seabird
mortality, and pathology.

47
Table 3. Overview of the number and corresponding percentages of age, sex, and season collected, for four species of
Procellariiformes collected beached on Sable Island, Nova Scotia, between 2000 and 2012. Seasons delimited by seasonal equinoxes.
COSH = Cory’s shearwater; GRSH = great shearwater; N = Number collected; NOFU = northern fulmar; SOSH = sooty shearwater;
Unk. = Unknown.
N (%)
Immature Adult Age unknown Season
Species Male (%)
Female (%)
Unk. (%)
Male (%)
Female (%)
Male (%)
Female (%)
Unk. (%)
Spring (%)
Summer (%)
Fall (%)
Winter (%)
Unk. (%)
COSH 3 (1) - - - 1 (33) 2 (67) - - - 2 (67) - 1 (33) - - SOSH 50 (16) 24 (48) 16 (32) 2 (4) 3 (6) 2 (4) - - 3 (6) 32 (64) 18 (36) - - - GRSH 89 (28) 35 (39) 26 (29) 1 (1) 7 (8) 6 (7) 9 (10) - 5 (6) 12 (13) 75 (84) 1 (1) - 1 (1) NOFU 176 (55) 52 (30) 55 (31) 10 (6) 19 (11) 10 (6) 11 (6) 8 (5) 11 (6) 107 (61) 13 (7) 19 (11) 34 (19) 3 (2) Total 318 (100) 111 (35) 97 (31) 13 (4) 30 (9) 20 (6) 20 (6) 8 (3) 19 (6) 152 (48) 106 (33) 22 (7) 34 (11) 4 (1)

48
Table 4. Pathology, ranked from most to least common, in 318 procellariids collected
beached on Sable Island, Nova Scotia, Canada, between 2000 and 2012. Percentage of
total by species displayed in brackets. “-” indicates no symptoms were found.
PATHOLOGY
Cory’s Shearwater
(%)
Sooty Shearwater
(%)
Great Shearwater
(%)
Northern Fulmar
(%)
Total
(%) Starvation 1 (33) 36 (72) 67 (75) 69 (39) 173 (54) Unknown - 2 (4) 3 (3) 61 (35) 66 (21) Autolysis 2 (67) 7 (14) 5 (6) 12 (7) 26 (8) Parasite Infection - - 4 (4) 6 (3) 10 (3) Sarcocystosis - - 2 2 4 Intestinal cestodiasis - - - 2 2 Parasitic proventriculitis - - 2 - 2 Pulmonary nematodiasis - - - 1 1 Renal parasitism (trematodes) - - - 1 1 Inflammation - - 1 (1) 7 (4) 8 (3) Air sacculitis - - - 2 2 Myocarditis - - - 1 1 Oophoritis and peritonitis - - - 1 1 Periarterial abscesses - - - 1 1 Pericarditis and air sacculitis - - - 1 1 Podpdermatitis - - - 1 1 Proventriculitis and serositis - - 1 - 1 Trauma - - 3 (3) 5 (3) 8 (3) Bacterial Infection - 1 (2) 2 (2) 4 (2) 7 (2) Cellulitis - - 2 1 3 Chronic ventricular ulcer - 1 - - 1 Mycobacteriosis - - - 1 1 Orchitis and funiculitis - - - 1 1 Septicemia - - - 1 1 Drowned - 3 (6) 2 (2) - 5 (2) Tumour - - - 4 (2) 4 (1) Fibromas - - - 2 2 Fibrochodroma - - - 1 1 Neoplasia - - - 1 1 Tissue Necrosis - 1 (2) 1 (1) 1 (1) 3 (1) Fat necrosis - 1 - 1 2 Hepatic necrosis - - 1 - 1 Impaction - - - 3 (2) 3 (1) Egg impaction - - - 2 2 Rectal Impaction - - - 1 1 Myopathy - - 1 (1) 2 (1) 3 (1) Pneumonia - - - 2 (1) 2 (1)

49
Table 5. Average body mass, and average mass of anthropogenic debris particles found
in the gizzards and proventriculi of four species of Procellariiformes collected beached on
Sable Island, Nova Scotia, between 2000 and 2012.
Body mass (g) Mass of debris (g) Species x̅ ± SD x̅ ± SD
Cory’s shearwater Calonectris borealis
686.83 ± 257.03 0.00 ± 0.00
Sooty shearwater Ardenna grisea
513.23 ± 69.05 0.07 ± 0.17
Great shearwater Ardenna gravis
528.70 ± 92.64 0.17 ± 0.33
Northern fulmar Fulmarus glacialis
665.24 ± 133.41 1.09 ± 1.93

50
Figure 3. Monthly collection breakdown of procellariid carcasses found beached on
Sable Island, Nova Scotia, Canada, between 2000-2012. Years combined. GRSH = great
shearwater; NOFU = northern fulmar; SOSH = sooty shearwater. Made in R using the
“ggplot2” package (Wickham, 2009).

51
Figure 4. Plumage morphs of beached northern fulmars (Fulmarus glacialis) collected
from Sable Island, Nova Scotia, Canada, between 2000-2012 and associated ingested
masses of anthropogenic debris (g) removed from the gizzard and proventriculus of birds.
Masses on y-axis log (x + 1)-transformed.

52
CHAPTER FOUR: GENERAL DISCUSSION AND FUTURE DIRECTIONS
Future Actions
A better understanding of the prevalence and effect of anthropogenic debris on
freshwater organisms, such as waterfowl, may help justify stronger regulations
controlling the manufacture and use of microbeads and microplastics, and improve
legislation to prevent their release into our waterways. Although there is evidence that
anthropogenic debris is a threat to aquatic biota, there is still a need for long term
monitoring to provide input for conservation management, strengthen the basis for
educational campaigns, and provide scientists with better evidence that could be used to
increase efforts to mitigate the problem (Derraik, 2002).
Although much is known about marine plastic debris, future research needs to
focus on the effects of freshwater debris. Freshwater environments are as polluted as their
marine counterparts (Eerkes-Medrano et al., 2015), and human plastic contaminant
exposure from either habitat could lead to potential long-term negative human health
effects. Future analyses on freshwater organisms are crucial to limiting exposure, and
decreasing human contaminant contact, as little is currently known about plastic exposure
and ingestion in freshwater organisms, and potential subsequent biomagnification to
humans. A specific focus on freshwater birds would be timely due to recent studies
finding plastics in commonly consumed bird species (English et al., 2015; Holland et al.,
2016).

53
Due to the ability of aquatic microplastics to vector heavy metal contaminants and
pollutants, it is likely that through biomagnification and subsequent aquatic animal
consumption, human health will be affected (Teuten et al., 2007; Ashton et al., 2010;
Rios et al., 2010). In laboratory settings zooplankton consume microplastics (Cole et al.,
2013; Wright et al., 2013; Cole et al., 2014) and recently, microplastics have been found
in digestive tracts of mesopelagic and epipelagic planktivorous fishes and in demersal
estuarine fish species (Boerger et al., 2010; Possatto et al., 2011; Dantas et al., 2012). Of
fish sold in markets for human consumption in California, USA, and Makassar,
Indonesia, 25-28% had anthropogenic debris present in their digestive tracts (Rochman et
al., 2015). Microplastics have also been found in the soft tissues of marine bivalves
(Mytilus edulis and Crassostrea gigas) commercially cultured for human consumption,
and resultant human dietary exposure from these two species could amount to 11,000
microplastic pieces consumed per year (Van Cauwenberghe and Janssen, 2014).
In regards to determining secondary exposure to plastics through contaminated
food species tissues, necropsies and stable isotope analyses of waterfowl would be
beneficial. Waterfowl are among one of the most culturally and economically significant
groups of hunted wildlife in Canada with most being taken from freshwater sites. In 2015
alone, an estimated 186,201 individuals across Canada hunted waterfowl (numbers from
Migratory Game Bird Hunting Permit stubs returned to Environment and Climate Change
Canada, 2016) and collected 2,167,973 birds from across Canada (Gendron and Smith,
2016).
Due to the large number of waterfowl sustenance hunters and their heavy focus on
hunting in freshwater environments, mitigation efforts to control plastic and other

54
potentially harmfully anthropogenic materials into the environment should be enacted.
We now know plastics are consumed by freshwater birds (English et al., 2015; Holland et
al., 2016; Gil-Delgado et al., 2017). Although efforts are being made to clean ocean
waters of plastics and help mitigate increasing concentrations (Kershaw et al., 2011), lack
of standardized research has meant that no similar initiatives exist for freshwater bodies
(Ryan et al., 2009). Problematically, legislation regarding marine dumping is often
ignored or not enforced, leading to little decrease in plastic debris release in marine
waters (Derraik, 2002), and thus seabirds continue to be exposed to debris.
Although more is known about seabird plastic ingestion, research needs to be
done to determine potential connections between seabird mortality, pathology, and plastic
ingestion. To determine which mortalities are natural, thorough investigations into
common causes of seabird mortality need to be conducted. Beached bird surveys are a
potential opportunistic sampling method, although these surveys often yield a
disproportionate number of immature and emaciated individuals. Our research on Sable
Island provides a much-needed baseline on seabird pathology in Nova Scotian waters.
This research also shows that seabird plastic ingestion continues to be a very real threat,
and illustrates that one population’s relationship with plastic may not apply to another.
To decrease aquatic anthropogenic debris concentrations in general, and plastic in
particular, stricter enforcement of existing laws and increased efforts at education are
required. Previous research has shown that education starting at a young age is effective
in bringing about positive change (Orr, 2004; Upham et al., 2009). Awareness of issues
and participation in their mitigation at a young age can not only help change
environmentally destructive habits, it can also lead to shared awareness with families and

55
communities, working as a catalyst for change (Derraik, 2002; Checkoway and Gutierrez,
2006). However, the most effective method will likely be a combination of legislative
and education-based ecological and environmental consciousness (Derraik, 2002).
Although past initiatives have been lacking, stronger regulations, enforcement, and public
education could make a positive difference.
Given that there is evidence that plastic pollution is a threat to aquatic
biodiversity, there is still a need for long term monitoring to provide input for
conservation management, strengthen the basis for educational campaigns, and provide
scientists with better evidence that could be used to increase efforts to mitigate the
problem (Derraik, 2002). It is imperative that research commence immediately, because
due to the long life of plastics, even if production and disposal of plastics was stopped
today, existing debris would continue to cause harm for many decades (Derraik, 2002).
Complications derived from different methods of reporting plastic debris concentrations
in freshwater and marine environments could hamper initiatives, because there is no
internationally agreed upon classification system for plastic debris (Driedger et al., 2015).
A better understanding of the prevalence and effect of microplastics on freshwater
organisms, such as waterfowl, may help justify stronger regulations against the
manufacture and use of microbeads and microplastics, and improve legislation to prevent
their release into our waterways. Our baseline data provide insights that this may have to
occur sooner than expected to prevent freshwater waterfowl debris ingestion levels from
reaching the levels currently observed in seabirds.
Steps to control plastic in the environment have been made, with plastic product
bans (e.g. polystyrene, bottled water and plastic bags) sweeping the world, while public

56
awareness and involvement increases (Wabnitz and Nichols, 2010). Likewise, increases
in knowledge of the harm debris can cause has led to subsequent increases in research
and legislation. As research into the newly arising field of freshwater anthropogenic
debris continues, we hopefully will continue to see public outcry over plastic waste, and
see renewed interest in renewable and ecologically friendly options.
References
Amélineau, F., Bonnet, D., Heitz, O., Mortreux, V., Harding, A.M.A., Karnovsky, N.,
Walkusz, W., Fort, J., Grémillet, D., 2016. Microplastic pollution in the
Greenland Sea: Background levels and selective contamination of planktivorous
diving seabirds. Environmental Pollution 219, 1131–1139.
doi:10.1016/j.envpol.2016.09.017
Andrady, A.L., 2011. Microplastics in the marine environment. Marine Pollution Bulletin
62, 1596–1605. doi:10.1016/j.marpolbul.2011.05.030
Arthur, C., Baker, J., Bamford, H., 2008. Proceedings of the international research
workshop on the occurrence, effects, and fate of mircroplastic marine debris.
National Oceanic and Atmospheric Administration Technical Memorandum
NOS-OR&R-30, 1–49.
Ashton, K., Holmes, L., Turner, A., 2010. Association of metals with plastic production
pellets in the marine environment. Marine Pollution Bulletin 60, 2050–2055.
doi:10.1016/j.marpolbul.2010.07.014

57
Auman, H.J., Ludwig, J.P., Giesy, J.P., Colborn, T., 1997. Plastic ingestion by Laysan
albatross chicks on Sand Island, Midway Atoll, in 1994 and 1995. Albatross
Biology and Conservation, 239-244.
Australian Government, 2016. Federal Government strengthens efforts to tackle plastic
waste [Media Release] [WWW Document]. The Hon. Greg Hunt MP Minister for
the Environment. URL
https://www.environment.gov.au/minister/hunt/2016/pubs/mr20160229a.pdf
(accessed 3.17.17).
Avery-Gomm, S., Valliant, M., Schacter, C.R., Robbins, K.F., Liboiron, M., Daoust, P.-
Y., Rios, L.M., Jones, I.L., 2016. A study of wrecked dovekies (Alle alle) in the
western North Atlantic highlights the importance of using standardized methods
to quantify plastic ingestion. Marine Pollution Bulletin 113, 75–80.
doi:10.1016/j.marpolbul.2016.08.062
Bakir, A., Rowland, S.J., Thompson, R.C., 2014. Enhanced desorption of persistent
organic pollutants from microplastics under simulated physiological conditions.
Environmental Pollution 185, 16–23. doi:10.1016/j.envpol.2013.10.007
Balseiro, A., Espí, A., Márquez, I., Pérez, V., Ferreras, M.C., Marín, J.F.G., Prieto, J.M.,
2005. Pathological features in marine birds affected by the Prestige’s oil spill in
the north of Spain. Journal of Wildlife Diseases 41, 371–378. doi:10.7589/0090-
3558-41.2.371
Barnes, D.K.A., Galgani, F., Thompson, R.C., Barlaz, M., 2009. Accumulation and
fragmentation of plastic debris in global environments. Philosophical

58
Transactions of the Royal Society B: Biological Sciences 364, 1985–1998.
doi:10.1098/rstb.2008.0205
Barrett, R.T., Camphuysen, K.C.J., Anker-Nilssen, T., Chardine, J.W., Furness, R.W.,
Garthe, S., Hüppop, O., Leopold, M.F., Montevecchi, W.A., Veit, R.R., 2007.
Diet studies of seabirds: A review and recommendations. International Council
for the Exploration of the Sea Journal of Marine Science 64, 1675–1691.
doi:10.1093/icesjms/fsm152
Bayly, I.A.E., 1993. The fauna of athalassic saline waters in Australia and the Altiplano
of South America: Comparisons and historical perspectives, in: Hurlbert, S.H.
(Ed.), Saline Lakes V, Developments in Hydrobiology. Springer Netherlands,
Bohn Stafleu van Loghum, Houten, Netherlands, pp. 225–231. doi:10.1007/978-
94-011-2076-0_18
Bellizzi, M., Duerr, R.S., 2013. Northern fulmar (Fulmarus glacialis) rehabilitation
during “wrecks” in California, in: The 44th Annual International Association for
Aquatic Animal Medicine Conference. The Marine Mammal Center, Sausalito,
California.
Besseling, E., Wang, B., Lürling, M., Koelmans, A.A., 2014. Nanoplastic affects growth
of S. obliquus and reproduction of D. magna. Environmental Science &
Technology 48, 12336–12343. doi:10.1021/es503001d
Bijker, W.E., Hughes, T.P., Pinch, T. (Eds.), 1987. The social construction of Bakelite:
toward a theory of invention, in: The Social Construction of Technological
Systems: New Directions in the Sociology and History of Technology. MIT Press,
Cambridge, Massachusetts, USA, pp. 159–187.

59
BirdLife International, 2016a. Fulmarus glacialis: BirdLife International: The IUCN Red
List of Threatened Species 2016: e.T22697866A89663126 [WWW Document].
Fulmarus glacialis. doi:10.2305/IUCN.UK.2016-
3.RLTS.T22697866A89663126.en
BirdLife International, 2016b. Calonectris borealis. The IUCN Red List of Threatened
Species 2016: e.T22732244A86237651 [WWW Document]. Calonectris borealis.
doi:10.2305/IUCN.UK.2016-3.RLTS.T22732244A86237651.en
BirdLife International, 2016c. Ardenna gravis. The IUCN Red List of Threatened Species
2016: e.T22698201A93669258 [WWW Document]. Ardenna gravis.
doi:10.2305/IUCN.UK.2016-3.RLTS.T22698201A93669258.en
BirdLife International, 2016d. Ardenna grisea: BirdLife International: The IUCN Red
List of Threatened Species 2016: e.T22698209A95225016 [WWW Document].
Ardenna grisea. doi:10.2305/IUCN.UK.2016-3.RLTS.T22698209A95225016.en
Blanco, G., Tella, J.L., Potti, J., 1997. Feather mites on group-living red-billed choughs:
A non-parasitic interaction? Journal of Avian Biology 28, 197–206.
doi:10.2307/3676970
Boerger, C.M., Lattin, G.L., Moore, S.L., Moore, C.J., 2010. Plastic ingestion by
planktivorous fishes in the North Pacific Central Gyre. Marine Pollution Bulletin
60, 2275–2278. doi:10.1016/j.marpolbul.2010.08.007
Bogstad, B., Drevetnyak, K.V., Byrkjedal, I., Dolgov, A.V., Gjøsæter, H., Johannesen,
E., Mehl, S., Høines, Å., Shevelev, M.S., Smirnov, O.V., 2014. Infectious
organisms [WWW Document]. BarentsPortal: Barents Sea Environmental Status
(A Norwegian-Russian Collaboration). URL

60
http://barentsportal.com/barentsportal/index.php/en/more/general-description-of-
the-ecosystem/89-biotic-components/155- (accessed 3.22.17).
Bond, A.L., Lavers, J.L., 2013. Effectiveness of emetics to study plastic ingestion by
Leach’s storm-petrels (Oceanodroma leucorhoa). Marine Pollution Bulletin 70,
171–175. doi:10.1016/j.marpolbul.2013.02.030
Bond, A.L., Provencher, J.F., Daoust, P.Y., Lucas, Z.N., 2014. Plastic ingestion by
fulmars and shearwaters at Sable Island, Nova Scotia, Canada. Marine Pollution
Bulletin 87, 68–75. doi:10.1016/j.marpolbul.2014.08.010
Camphuysen, C.J., Heubeck, M., 2001. Marine oil pollution and beached bird surveys:
The development of a sensitive monitoring instrument. Environmental Pollution
112, 443–461.
Carpenter, D.O., 2006. Polychlorinated biphenyls (PCBs): Routes of exposure and effects
on human health. Reviews on Environmental Health 21, 1–24.
doi:10.1515/REVEH.2006.21.1.1
Castañeda, R.A., Avlijas, S., Simard, M.A., Ricciardi, A., Smith, R., 2014. Microplastic
pollution in St. Lawrence River sediments. Canadian Journal of Fisheries and
Aquatic Sciences 71, 1767–1771. doi:10.1139/cjfas-2014-0281
Checkoway, B.N., Gutierrez, L.M., 2006. Youth participation and community change:
An introduction. Journal of Community Practice 14, 1–9.
doi:10.1300/J125v14n01_01
Cheek, A.O., Kow, K., Chen, J., McLachlan, J.A., 1999. Potential mechanisms of thyroid
disruption in humans: Interaction of organochlorine compounds with thyroid

61
receptor, transthyretin, and thyroid-binding globulin. Environmental Health
Perspectives 107, 273–278.
Cochran, W.G., 1954. Some methods for strengthening the common χ2 tests. Biometrics
10, 417–451. doi:10.2307/3001616
Cole, M., Lindeque, P., Fileman, E., Halsband, C., Goodhead, R., Moger, J., Galloway,
T.S., 2013. Microplastic ingestion by zooplankton. Environmental Science &
Technology 47, 6646–6655. doi:10.1021/es400663f
Cole, M., Webb, H., Lindeque, P.K., Fileman, E.S., Halsband, C., Galloway, T.S., 2014.
Isolation of microplastics in biota-rich seawater samples and marine organisms.
Scientific Reports 4, 4528. doi:10.1038/srep04528
Connors, P.G., Smith, K.G., 1982. Oceanic plastic particle pollution: Suspected effect on
fat deposition in red phalaropes. Marine Pollution Bulletin 13, 18–20.
doi:10.1016/0025-326X(82)90490-8
Corcoran, P.L., Norris, T., Ceccanese, T., Walzak, M.J., Helm, P.A., Marvin, C.H., 2015.
Hidden plastics of Lake Ontario, Canada and their potential preservation in the
sediment record. Environmental Pollution 204, 17–25.
doi:10.1016/j.envpol.2015.04.009
Costa, L.G., Giordano, G., Tagliaferri, S., Caglieri, A., Mutti, A., 2008. Polybrominated
diphenyl ether (PBDE) flame retardants: Environmental contamination, human
body burden and potential adverse health effects. Acta Biomed 79, 172–183.
Council of the European Union, 2014. 3363rd Council meeting [Press Release] [WWW
Document]. Environment. URL

62
http://www.consilium.europa.eu/uedocs/cms_data/docs/pressdata/en/envir/146374
.pdf (accessed 3.29.17).
Dantas, D.V., Barletta, M., Costa, M.F. da, 2012. The seasonal and spatial patterns of
ingestion of polyfilament nylon fragments by estuarine drums (Sciaenidae).
Environmental Science and Pollution Research 2, 600–606. doi:10.1007/s11356-
011-0579-0
Day, R.H., 1980. The occurrence and characteristics of plastic pollution in Alaska’s
marine birds (MSc Thesis). University of Alaska, Fairbanks, Alaska, USA.
Day, R.H., Wehle, D.H.S., Coleman, F.C., 1985. Ingestion of plastic pollutants by marine
birds, in: Proceedings of the Workshop on the Fate and Impact of Marine Debris.
National Oceanic and Atmospheric Administration, Honolulu, Hawaii, USA, pp.
344–386.
Denuncio, P., Bastida, R., Dassis, M., Giardino, G., Gerpe, M., Rodríguez, D., 2011.
Plastic ingestion in Franciscana dolphins, Pontoporia blainvillei (Gervais and
d’Orbigny, 1844), from Argentina. Marine Pollution Bulletin 62, 1836–1841.
doi:10.1016/j.marpolbul.2011.05.003
Derraik, J.G., 2002. The pollution of the marine environment by plastic debris: A review.
Marine Pollution Bulletin 44, 842–852.
Desforges, J.P.W., Galbraith, M., Ross, P.S., 2015. Ingestion of microplastics by
zooplankton in the Northeast Pacific Ocean. Archives of Environmental
Contamination and Toxicology 69, 320–330. doi:10.1007/s00244-015-0172-5
Doughty, R., Eriksen, M., 2014. The case for a ban on microplastics in personal care
products. Tulane Environmental Law Journal 27, 277.

63
Driedger, A.G.J., Dürr, H.H., Mitchell, K., Van Cappellen, P., 2015. Plastic debris in the
Laurentian Great Lakes: A review. Journal of Great Lakes Research 41, 9–19.
doi:10.1016/j.jglr.2014.12.020
Eerkes-Medrano, D., Thompson, R.C., Aldridge, D.C., 2015. Microplastics in freshwater
systems: A review of the emerging threats, identification of knowledge gaps and
prioritisation of research needs. Water Research 75, 63–82.
doi:10.1016/j.watres.2015.02.012
El-Hayek, Y.H., 2007. Mercury contamination in Arctic Canada: Possible implications
for Aboriginal health. Journal on Developmental Disabilities 13, 67–89.
English, M.D., 2016. Diet and body condition of American black ducks (Anas rubripes)
overwintering in Atlantic Canada (MSc Thesis). Acadia University, Wolfville,
Nova Scotia.
English, M.D., Robertson, G.J., Avery-Gomm, S., Pirie-Hay, D., Roul, S., Ryan, P.C.,
Wilhelm, S.I., Mallory, M.L., 2015. Plastic and metal ingestion in three species of
coastal waterfowl wintering in Atlantic Canada. Marine Pollution Bulletin 98,
349–353. doi:10.1016/j.marpolbul.2015.05.063
Environment and Climate Change Canada, 2016. Permit Sales [WWW Document]. URL
https://www.ec.gc.ca/reom-mbs/default.asp?lang=en&n=C9046964 (accessed
3.19.17).
Environment and Climate Change Canada, 2010. Environment and Climate Change
Canada - Acts & Regulations - Toxic Substances List - Schedule 1 [WWW
Document]. URL http://www.ec.gc.ca/lcpe-

64
cepa/default.asp?lang=En&n=0DA2924D-1&wsdoc=4ABEFFC8-5BEC-B57A-
F4BF-11069545E434 (accessed 3.17.17).
Eriksen, M., Lebreton, L.C.M., Carson, H.S., Thiel, M., Moore, C.J., Borerro, J.C.,
Galgani, F., Ryan, P.G., Reisser, J., 2014. Plastic pollution in the world’s oceans:
More than 5 trillion plastic pieces weighing over 250,000 tons afloat at sea. PLoS
ONE 9, e111913. doi:10.1371/journal.pone.0111913
Eriksen, M., Mason, S., Wilson, S., Box, C., Zellers, A., Edwards, W., Farley, H., Amato,
S., 2013. Microplastic pollution in the surface waters of the Laurentian Great
Lakes. Marine Pollution Bulletin 77, 177–182.
doi:10.1016/j.marpolbul.2013.10.007
Fängström, B., Athanasiadou, M., Athanassiadis, I., Bignert, A., Grandjean, P., Weihe,
P., Bergman, Å., 2005. Polybrominated diphenyl ethers and traditional
organochlorine pollutants in fulmars (Fulmarus glacialis) from the Faroe Islands.
Chemosphere 60, 836–843. doi:10.1016/j.chemosphere.2005.01.065
Fischer, E.K., Paglialonga, L., Czech, E., Tamminga, M., 2016. Microplastic pollution in
lakes and lake shoreline sediments – A case study on Lake Bolsena and Lake
Chiusi (central Italy). Environmental Pollution 213, 648–657.
doi:10.1016/j.envpol.2016.03.012
Fisher, J., 1952. The colour-phases of the fulmar, in: The Fulmar, Collins New Naturalist.
Collins, London, England, pp. 266–288.
Food and Agriculture Organization of the United Nations, 2016. The state of world
fisheries and aquaculture: Contributing to food security and nutrition for all. Food
and Agriculture Organization of the United Nations, Rome, Italy.

65
Free, C.M., Jensen, O.P., Mason, S.A., Eriksen, M., Williamson, N.J., Boldgiv, B., 2014.
High-levels of microplastic pollution in a large, remote, mountain lake. Marine
Pollution Bulletin 85, 156–163. doi:10.1016/j.marpolbul.2014.06.001
Frias, J.P.G.L., Sobral, P., Ferreira, A.M., 2010. Organic pollutants in microplastics from
two beaches of the Portuguese coast. Marine Pollution Bulletin 60, 1988–1992.
doi:10.1016/j.marpolbul.2010.07.030
Furness, R.W., Camphuysen, K.C., 1997. Seabirds as monitors of the marine
environment. ICES Journal of Marine Science 54, 726–737.
Gall, S.C., Thompson, R.C., 2015. The impact of debris on marine life. Marine Pollution
Bulletin 92, 170–179. doi:10.1016/j.marpolbul.2014.12.041
Gaynor, A.M., Fish, S., Duerr, R.S., Cruz, F.N.D., Pesavento, P.A., 2015. Identification
of a novel papillomavirus in a northern fulmar (Fulmarus glacialis) with viral
production in cartilage. Veterinary Pathology 52, 553–561.
doi:10.1177/0300985814542812
Gendron, M.H., Smith, A.C., 2013. National Harvest Survey web site. Bird Populations
Monitoring, National Wildlife Research Centre, Canadian Wildlife Service,
Ottawa, Ontario [WWW Document]. National Harvest Survey. URL
http://www.ec.gc.ca/reom-mbs/enp-nhs/index.cfm?do=a&lang=e (accessed
3.19.17).
Gendron, M.H., Smith, A.C., 2016. National Harvest Survey web site. Bird Populations
Monitoring, National Wildlife Research Centre, Canadian Wildlife Service,
Ottawa, Ontario. [WWW Document]. National Harvest Survey. URL

66
http://www.ec.gc.ca/reom-mbs/enp-nhs/index.cfm?do=def&lang=e (accessed
3.19.17).
Gil-Delgado, J.A., Guijarro, D., Gosálvez, R.U., López-Iborra, G.M., Ponz, A., Velasco,
A., 2017. Presence of plastic particles in waterbirds faeces collected in Spanish
lakes. Environmental Pollution 220, 732–736. doi:10.1016/j.envpol.2016.09.054
Gouin, T., Roche, N., Lohmann, R., Hodges, G., 2011. A thermodynamic approach for
assessing the environmental exposure of chemicals absorbed to microplastic.
Environmental Science & Technology 45, 1466–1472. doi:10.1021/es1032025
Government of Canada, 2016. Canada Gazette – Microbeads in Toiletries Regulations
[WWW Document]. Microbeads in Toiletries Regulations. URL
http://www.gazette.gc.ca/rp-pr/p1/2016/2016-11-05/html/reg2-eng.php (accessed
3.17.17).
Government of Canada, 2015. Microbeads [WWW Document]. URL
http://www.chemicalsubstanceschimiques.gc.ca/plan/approach-approche/microb-
eng.php (accessed 3.17.17).
Government of Canada, F. and O.C., 2010. The Gully [WWW Document]. URL
http://www.inter.dfo-mpo.gc.ca/Maritimes/Oceans/OCMD/Gully/Gully-MPA
(accessed 3.17.17).
Grant, G.S., Trail, P.W., Clapp, R.B., 1994. First specimens of sooty shearwater,
Newell’s shearwater, and white-faced storm-petrel from American Samoa.
Notornis 41, 215–217.
Gray, R., 1871. The birds of the west of Scotland, including the Outer Hebrides. T.
Murray, Glasgow, Scotland.

67
Great Lakes and St. Lawrence Cities Initiative, 2014. Great Lakes and St. Lawrence
Cities Initiative Resolution 03-2014M [WWW Document]. Microbeads. URL
https://glslcities.org/wp-content/uploads/2015/05/2014_3_microbeads.pdf
(accessed 3.29.17).
Gregory, M.R., 1978. Accumulation and distribution of virgin plastic granules on New
Zealand beaches. New Zealand Journal of Marine and Freshwater Research 12,
399–414. doi:10.1080/00288330.1978.9515768
Gregory, M.R., 1983. Virgin plastic granules on some beaches of Eastern Canada and
Bermuda. Marine Environmental Research 10, 73–92. doi:10.1016/0141-
1136(83)90011-9
Gregory, M.R., 2009. Environmental implications of plastic debris in marine settings –
entanglement, ingestion, smothering, hangers-on, hitch-hiking and alien
invasions. Philosophical Transactions of the Royal Society B: Biological Sciences
364, 2013–2025. doi:10.1098/rstb.2008.0265
GTA Consultants Inc., 1999. Socio-economic profile of the Sable Gully. GTA
Consultants Inc., Halifax, Nova Scotia, Canada.
Haman, K.H., Norton, T.M., Ronconi, R.A., Nemeth, N.M., Thomas, A.C., Courchesne,
S.J., Segars, A., Keel, M.K., 2013. Great shearwater (Puffinus gravis) mortality
events along the eastern coast of the United States. Journal of Wildlife Diseases
49, 235–245. doi:10.7589/2012-04-119
Harris, R.J., Tseng, F.S., Pokras, M.A., Suedmeyer, B.A., Bogart, J.S.H., Prescott, R.L.,
Newman, S.H., 2006. Beached bird surveys in Massachusetts: The seabird
ecological assessment network (SEANET). Marine Ornithology 34, 115–122.

68
Harvey, J., Nevins, H.M., Hatch, S., Adams, J., Hill, J., Ames, J., Parkin, J., Newton, K.,
Roletto, J., Mortenson, J., Hall, J., Hass, T., 2004. An unusual mortality of
northern fulmar (Fulmarus glacialis), in: Sanctuary Currents Symposium.
Monterey Bay National Marine Sanctuary, Seaside, California, USA.
Hatch, S.A., 1991. Evidence for color phase effects on the breeding and life history of
northern fulmars. The Condor 93, 409–417. doi:10.2307/1368957
Hilton, G.M., Furness, R.W., Houston, D.C., 2000. A comparative study of digestion in
North Atlantic seabirds. Journal of Avian Biology 31, 36–46. doi:10.1034/j.1600-
048X.2000.310106.x
Holladay, S.D., Smialowicz, R.J., 2000. Development of the murine and human immune
system: Differential effects of immunotoxicants depend on time of exposure.
Environmental Health Perspectives 108, 463–473.
Holland, E., Mallory, M.L., Shutler, D., 2016. Plastics and other anthropogenic debris in
freshwater birds from Canada. Science of the Total Environment 571, 251–258.
doi:http://dx.doi.org/10.1016/j.scitotenv.2016.07.158
House Report No. 114-371, 2015. H. Rept. 114-371 - Microbead-free waters act of 2015
[WWW Document]. URL https://www.congress.gov/congressional-report/114th-
congress/house-report/371/1 (accessed 3.19.17).
Huettmann, F., Diamond, A.W., 2000. Seabird migration in the Canadian northwest
Atlantic Ocean: Moulting locations and movement patterns of immature birds.
Canadian Journal of Zoology 78, 624–647. doi:10.1139/cjz-78-4-624

69
Humphries, G.R.W., 2014. Using long term harvest records of sooty shearwaters (Tītī;
Puffinus griseus) to predict shifts in the Southern Oscillation (PhD Thesis).
University of Otago, University of Otago, Dunedin, New Zealand.
International Joint Commission Canada and United States, 2016. Microplastics in the
Great Lakes workshop report. International Joint Commission Canada and United
States Final Report, 1–31.
Ivar do Sul, J.A., Costa, M.F., 2014. The present and future of microplastic pollution in
the marine environment. Environmental Pollution 185, 352–364.
doi:10.1016/j.envpol.2013.10.036
Jackson, D.A., 1993. Stopping rules in principal components analysis: A comparison of
heuristical and statistical approaches. Ecology 74, 2204–2214.
doi:10.2307/1939574
Jessup, D.A., Miller, M.A., Ryan, J.P., Nevins, H.M., Kerkering, H.A., Mekebri, A.,
Crane, D.B., Johnson, T.A., Kudela, R.M., 2009. Mass stranding of marine birds
caused by a surfactant-producing red tide. PLoS ONE 4, e4550.
doi:10.1371/journal.pone.0004550
Johansen, P., Asmund, G., Riget, F., 2001. Lead contamination of seabirds harvested with
lead shot – implications to human diet in Greenland. Environmental Pollution
112, 501–504.
Kalmar, I.D., Dicxk, V., Dossche, L., Vanrompay, D., 2014. Zoonotic infection with
Chlamydia psittaci at an avian refuge centre. The Veterinary Journal 199, 300–
302. doi:10.1016/j.tvjl.2013.10.034

70
Kanehiro, H., Tokai, T., Matuda, K., 1995. Marine litter composition and distribution on
the sea-bed of Tokyo Bay [Japan]. Fisheries Engineering (Japan) 31, 195–199.
Karlsson, M., Ericson, I., van Bavel, B., Jensen, J.K., Dam, M., 2006. Levels of
brominated flame retardants in Northern Fulmar (Fulmarus glacialis) eggs from
the Faroe Islands. Science of The Total Environment 367, 840–846.
doi:10.1016/j.scitotenv.2006.02.050
Kenyon, K.W., Kridler, E., 1969. Laysan albatrosses swallow indigestible matter. The
Auk 86, 339–343. doi:10.2307/4083505
Kershaw, P., Katsuhiko, S., Lee, S., Samseth, J., Woodring, D., Smith, J., 2011. Plastic
debris in the ocean, UNEP Year Book. United Nations Environment Programme,
Nairobi, Kenya, Africa.
Kirk, D.A., Gosler, A.G., 1994. Body condition varies with migration and competition in
migrant and resident South American vultures. The Auk 111, 933–944.
doi:10.2307/4088825
Klar, M., Gunnarsson, D., Prevodnik, A., Hedfors, C., Dahl, U., 2014. Everything you
(don’t) want to know about plastics. Swedish Society for Nature Conservation 1–
197.
Koelmans, A.A., Besseling, E., Foekema, E.M., 2014. Leaching of plastic additives to
marine organisms. Environmental Pollution 187, 49–54.
doi:10.1016/j.envpol.2013.12.013
Laist, D.W., 1997. Impacts of marine debris: Entanglement of marine life in marine
debris including a comprehensive list of species with entanglement and ingestion

71
records, in: Coe, J.M., Rogers, D.B. (Eds.), Marine Debris. Springer New York,
New York, NY, pp. 99–139. doi:10.1007/978-1-4613-8486-1_10
Laist, D.W., 1987. Overview of the biological effects of lost and discarded plastic debris
in the marine environment. Marine Pollution Bulletin 18, 319–326.
doi:10.1016/S0025-326X(87)80019-X
Lang, A.S., Lebarbenchon, C., Ramey, A.M., Robertson, G.J., Waldenström, J., Wille,
M., 2016. Assessing the role of seabirds in the ecology of influenza A viruses.
Avian Diseases 60, 378–386. doi:10.1637/11135-050815-RegR
Larntz, K., 1978. Small-sample comparisons of exact levels for chi-squared goodness-of-
fit statistics. Journal of the American Statistical Association 73, 253–263.
doi:10.2307/2286650
Lavers, J.L., Bond, A.L., 2016. Selectivity of flesh-footed shearwaters for plastic colour:
Evidence for differential provisioning in adults and fledglings. Marine
Environmental Research 113, 1–6. doi:10.1016/j.marenvres.2015.10.011
Lechner, A., Keckeis, H., Lumesberger-Loisl, F., Zens, B., Krusch, R., Tritthart, M.,
Glas, M., Schludermann, E., 2014. The Danube so colourful: A potpourri of
plastic litter outnumbers fish larvae in Europe’s second largest river.
Environmental Pollution 188, 177–181. doi:10.1016/j.envpol.2014.02.006
Lechner, A., Ramler, D., 2015. The discharge of certain amounts of industrial
microplastic from a production plant into the River Danube is permitted by the
Austrian legislation. Environmental Pollution 200, 159–160.
doi:10.1016/j.envpol.2015.02.019

72
Leslie, H.A., 2014. Review of microplastics in cosmetics (No. Report R14/29). Institute
for Environmental Studies [IVM], Amsterdam, Netherlands.
Li, L., Pesavento, P.A., Gaynor, A.M., Duerr, R.S., Phan, T.G., Zhang, W., Deng, X.,
Delwart, E., 2015. A gyrovirus infecting a sea bird. Archives of Virology 160,
2105–2109. doi:10.1007/s00705-015-2468-1
Lieske, C.L., Ziccardi, M.H., Mazet, J.A., Newman, S.H., Gardner, I.A., 2002.
Evaluation of 4 handheld blood glucose monitors for use in seabird rehabilitation.
Journal of Avian Medicine and Surgery 16, 277–285.
Lucas, Z., 1992. Monitoring persistent litter in the marine environment on Sable Island,
Nova Scotia. Marine Pollution Bulletin 24, 192–199. doi:10.1016/0025-
326X(92)90529-F
Lucas, Z., Horn, A., Freedman, B., 2012. Beached bird surveys on Sable Island, Nova
Scotia, 1993–2009, show a decline in the incidence of oiling. Proceedings of the
Nova Scotian Institute of Science 47, 91–129.
Lucas, Z., MacGregor, C., 2006. Characterization and source of oil contamination on the
beaches and seabird corpses, Sable Island, Nova Scotia, 1996–2005. Marine
Pollution Bulletin 52, 778–789. doi:10.1016/j.marpolbul.2005.11.023
Macdonald, M.A., 1977. Adult mortality and fidelity to mate and nest-site in a group of
marked fulmars. Bird Study 24, 165-168.
doi:http://dx.doi.org/10.1080/00063657709476552
Mallory, M.L., Akearok, J.A., Edwards, D.B., O’Donovan, K., Gilbert, C.D., 2008.
Autumn migration and wintering of northern fulmars (Fulmarus glacialis) from

73
the Canadian high Arctic. Polar Biology 31, 745–750. doi:10.1007/s00300-008-
0417-0
Mallory, M.L., Forbes, M.R., 2005. Sex discrimination and measurement bias in northern
fulmars Fulmarus glacialis from the Canadian Arctic. Ardea 93, 25–36.
Mallory, M.L., Mclaughlin, J.D., Forbes, M.R., 2007. Breeding status, contaminant
burden and helminth parasites of northern fulmars Fulmarus glacialis from the
Canadian high Arctic. Ibis 149, 338–344. doi:10.1111/j.1474-919X.2006.00636.x
Mascarenhas, R., Santos, R., Zeppelini, D., 2004. Plastic debris ingestion by sea turtle in
Paraı́ba, Brazil. Marine Pollution Bulletin 49, 354–355.
doi:10.1016/j.marpolbul.2004.05.006
McDonald, T.A., 2002. A perspective on the potential health risks of PBDEs.
Chemosphere 46, 745–755.
Michel, P., Ollason, J.C., Grosbois, V., Thompson, P.M., 2003. The influence of body
size, breeding experience and environmental variability on egg size in the
northern fulmar (Fulmarus glacialis). Journal of Zoology 261, 427-432.
doi:10.1017/S0952836903004291
Millennium Ecosystem Assessment, 2005. Ecosystems and human well-being.
Moore, C.J., 2008. Synthetic polymers in the marine environment: A rapidly increasing,
long-term threat. Environmental Research 108, 131–139.
doi:10.1016/j.envres.2008.07.025
Moore, J.L., Hohman, W.L., Stark, T.M., Weisbrich, G.A., 1998. Shot prevalences and
diets of diving ducks five years after ban on use of lead shotshells at Catahoula

74
lake, Louisiana. The Journal of Wildlife Management 62, 564.
doi:10.2307/3802330
Moseman, E., 2015. Micro-plastic bioaccumulation in yellow perch (Perca flavescens) of
Lake Champlain, in: Center for Earth and Environmental Science Student Posters.
Center for Earth and Environmental Science, Plattsburgh, New York, USA.
Moser, M.L., Lee, D.S., 1992. A fourteen-year survey of plastic ingestion by western
north Atlantic seabirds. Colonial Waterbirds 15, 83. doi:10.2307/1521357
Mougin, J.L., Jouanin, C., Roux, F., Zino, F., 2000. Fledging weight and juvenile
survival of Cory’s shearwaters Calonectris diomedea on Selvagem Grande.
Ringing & Migration 20, 107–110. doi:10.1080/03078698.2000.9674231
Muzaffar, S.B., Jones, I.L., 2004. Parasites and diseases of the auks (Alcidae) of the
world and their ecology – a review. Marine Ornithology 32, 121–146.
Nemeth, N., Keel, K., Yabsley, M., 2011. Case W10-367, in: The 39th Annual South
Eastern Veterinary Pathology Conference (SEVPAC). The University of Georgia
Veterinary Diagnostic & Investigational Laboratory, Athens, Georgia.
Nemeth, N.M., Yabsley, M., Keel, M.K., 2012. Anisakiasis with proventricular
perforation in a greater shearwater (Puffinus gravis) off the coast of Georgia,
United States. Journal of Zoo and Wildlife Medicine 43, 412–415.
doi:10.1638/2011-0193.1
Neufeld, L., Stassen, F., Sheppard, R., Gilman, 2016. The new plastics economy:
Rethinking the future of plastics. World Economic Forum 1–36.
Newman, S., Chmura, A., Converse, K., Kilpatrick, A., Patel, N., Lammers, E., Daszak,
P., 2007. Aquatic bird disease and mortality as an indicator of changing

75
ecosystem health. Marine Ecology Progress Series 352, 299–309.
doi:10.3354/meps07076
Newman, S., Watkins, E., Farmer, A., 2013. How to improve EU legislation to tackle
marine litter. Institute for European Environmental Policy, London. 1–65.
Newman, S.H., Piatt, J.F., White, J., 1997. Hematological and plasma biochemical
reference ranges of Alaskan seabirds: their ecological significance and clinical
importance. Colonial Waterbirds 20, 492–504. doi:10.2307/1521600
Ng, K.L., Obbard, J.P., 2006. Prevalence of microplastics in Singapore’s coastal marine
environment. Marine Pollution Bulletin 52, 761–767.
doi:10.1016/j.marpolbul.2005.11.017
Obbard, R.W., Sadri, S., Wong, Y.Q., Khitun, A.A., Baker, I., Thompson, R.C., 2014.
Global warming releases microplastic legacy frozen in Arctic Sea ice. Earth’s
Future 2, 2014EF000240. doi:10.1002/2014EF000240
Ocean Conservancy, 2010. Trash travels: From our hands to the sea, around the glove,
and through time. [WWW Document]. International Coastal Cleanup. URL
http://act.oceanconservancy.org/site/DocServer/icc2010report_regional_Mex.pdf?
docID=6004 (accessed 3.29.17).
O’Reilly, B., 1818. Greenland, the adjacent seas, and the North-west Passage to the
Pacific Ocean, illustrated in a voyage to Davis’s strait, during the summer of
1817. London, Baldwin, Craddock and Joy, London, England.
Orr, D.W., 2004. Earth in Mind: On Education, Environment, and the Human Prospect.
Island Press, Washington, DC, USA.

76
OSPAR Commission, 2010. The OSPAR system of ecological quality objectives for the
North Sea, a contribution to OSPAR’s quality status report 2010. OSPAR
Commission, London, London, England.
Peitzman, A.B., Fabian, T.C., Rhodes, M., Yealy, D.M., Schwab, C.W., 2012. Chapter 3:
Patterns of penetrating injury, in: The Trauma Manual: Trauma and Acute Care
Surgery. Lippincott Williams & Wilkins, pp. 16–22.
Pennycott, T., 2016. Seabirds. Images of pathology (miscellaneous diseases 2) 1994-
2013. University of Edinburgh, Royal (Dick) School of Veterinary Studies,
Midlothian, Scotland.
Pettit, T.N., Grant, G.S., Whittow, G.C., 1981. Ingestion of plastics by Laysan albatross.
The Auk 98, 839–841.
Pierce, K.E., Harris, R.J., Larned, L.S., Pokras, M.A., 2004. Obstruction and starvation
associated with plastic ingestion in a northern gannet Morus bassanus and a
greater shearwater Puffinus gravis. Marine Ornithology 32, 187–189.
Plastics Europe, 2015. Plastics – the facts 2014/2015. An analysis of European plastics
production, demand and waste data. Plastics Europe, Brussels, Belgium.
Plastics Europe, 2013. Plastics – the facts 2013: An analysis of European latest plastics
production, demand, and waste data. Plastics Europe, Brussels, Belgium.
Pokras, M.A., 1988. Captive management of aquatic birds. Association of Avian
Veterinarians Today 2, 24. doi:10.2307/30134115
Possatto, F.E., Barletta, M., Costa, M.F., Ivar do Sul, J.A., Dantas, D.V., 2011. Plastic
debris ingestion by marine catfish: An unexpected fisheries impact. Marine
Pollution Bulletin 62, 1098–1102. doi:10.1016/j.marpolbul.2011.01.036

77
Powers, K.D., 1984. Pelagic distributions of marine birds off the northeastern United
States. US Department of Commerce, National Oceanic and Atmospheric
Administration, National Marine Fisheries Service, Northeast Fisheries Center,
Manomet, Massachusetts, USA.
Provencher, J.F., Bond, A.L., Hedd, A., Montevecchi, W.A., Muzaffar, S.B., Courchesne,
S.J., Gilchrist, H.G., Jamieson, S.E., Merkel, F.R., Falk, K., Durinck, J., Mallory,
M.L., 2014. Prevalence of marine debris in marine birds from the North Atlantic.
Marine Pollution Bulletin 84, 411–417. doi:10.1016/j.marpolbul.2014.04.044
Provencher, J.F., Bond, A.L., Mallory, M.L., 2015. Marine birds and plastic debris in
Canada: A national synthesis and a way forward. Environmental Reviews 23, 1–
13. doi:10.1139/er-2014-0039
R Core Team, 2015. R: A language and environment for statistical computing [WWW
Document]. The R Project for Statistical Computing. URL https://www.r-
project.org/ (accessed 3.19.17).
Reyes-Arriagada, R., Campos-Ellwanger, P., Schlatter, R.P., Baduini, C., 2007. Sooty
shearwater (Puffinus griseus) on Guafo Island: The largest seabird colony in the
world? Biodiversity and Conservation 16, 913–930. doi:10.1007/s10531-006-
9087-9
Rios, L.M., Jones, P.R., Moore, C., Narayan, U.V., 2010. Quantitation of persistent
organic pollutants adsorbed on plastic debris from the Northern Pacific Gyre’s
“eastern garbage patch.” Journal of Environmental Monitoring 12, 2226.
doi:10.1039/c0em00239a

78
Robards, M.D., Piatt, J.F., Wohl, K.D., 1995. Increasing frequency of plastic particles
ingested by seabirds in the subarctic North Pacific. Marine Pollution Bulletin 30,
151–157.
Robinson, D., 2016. broom: Convert statistical analysis objects into tidy data frames. R
package version 0.4.1.9000 [WWW Document]. GitHub. URL
https://github.com/tidyverse/broom (accessed 3.21.17).
Rochman, C.M., Hentschel, B.T., Teh, S.J., 2014. Long-term sorption of metals is similar
among plastic types: Implications for plastic debris in aquatic environments.
PLoS ONE 9, e85433. doi:10.1371/journal.pone.0085433
Rochman, C.M., Tahir, A., Williams, S.L., Baxa, D.V., Lam, R., Miller, J.T., Teh, F.-C.,
Werorilangi, S., Teh, S.J., 2015. Anthropogenic debris in seafood: Plastic debris
and fibers from textiles in fish and bivalves sold for human consumption.
Scientific Reports 5, 1–10. doi:10.1038/srep14340
Rodríguez, A., Rodríguez, B., Nazaret Carrasco, M., 2012. High prevalence of parental
delivery of plastic debris in Cory’s shearwaters (Calonectris diomedea). Marine
Pollution Bulletin 64, 2219–2223. doi:10.1016/j.marpolbul.2012.06.011
Roletto, J., Mortenson, J., Harrald, I., Hall, J., Grella, L., 2003. Beached bird surveys and
chronic oil pollution in central California. Marine Ornithology 31, 21–28.
Ross, J.B., Parker, R., Strickland, M., 1991. A survey of shoreline litter in Halifax
Harbour 1989. Marine Pollution Bulletin 22, 245–248. doi:10.1016/0025-
326X(91)90919-J
Rothstein, S.I., 1973. Plastic particle pollution of the surface of the Atlantic Ocean:
evidence from a seabird. Condor 75, 5.

79
Ryan, P.G., 1987. The effects of ingested plastic on seabirds: Correlations between
plastic load and body condition. Environmental Pollution 46, 119–125.
doi:10.1016/0269-7491(87)90197-7
Ryan, P.G., Connell, A.D., Gardner, B.D., 1988. Plastic ingestion and PCBs in seabirds:
Is there a relationship? Marine Pollution Bulletin 19, 174–176. doi:10.1016/0025-
326X(88)90674-1
Ryan, P.G., Jackson, S., 1987. The lifespan of ingested plastic particles in seabirds and
their effect on digestive efficiency. Marine Pollution Bulletin 18, 217–219.
doi:10.1016/0025-326X(87)90461-9
Ryan, P.G., Moore, C.J., van Franeker, J.A., Moloney, C.L., 2009. Monitoring the
abundance of plastic debris in the marine environment. Philosophical
Transactions of the Royal Society B: Biological Sciences 364, 1999–2012.
doi:10.1098/rstb.2008.0207
Sanchez, W., Bender, C., Porcher, J.M., 2014. Wild gudgeons (Gobio gobio) from French
rivers are contaminated by microplastics: Preliminary study and first evidence.
Environmental Research 128, 98–100. doi:10.1016/j.envres.2013.11.004
Schneiderman, E.T., 2014. Unseen threat: How microbeads harm New York waters,
wildlife, health and environment. New York State Attorney General, New York,
New York, USA.
Secretariat of the Convention on Biological Diversity, 2012. Impacts of marine debris on
biodiversity: Current status and potential solutions, CBD technical series.
Secretariat of the Convention on Biological Diversity, Montreal, Quebec, Canada.

80
Shaw, D.G., Day, R.H., 1994. Colour- and form-dependent loss of plastic micro-debris
from the North Pacific Ocean. Marine Pollution Bulletin 28, 39–43.
doi:10.1016/0025-326X(94)90184-8
Snyder, J.A., Vakos, H.C., 1966. High shear strength blends of high and low density
polyethylene. US3231636 A.
Spear, L.B., Ainley, D.G., Ribic, C.A., 1995. Incidence of plastic in seabirds from the
tropical pacific, 1984–1991: Relation with distribution of species, sex, age,
season, year and body weight. Marine Environmental Research 40, 123–146.
Steele, C.M., Brown, R.N., Botzler, R.G., 2005. Prevalences of zoonotic bacteria among
seabirds in rehabilitation centers along the Pacific Coast of California and
Washington, USA. Journal of Wildlife Diseases 41, 735–744.
Stephen, C., Burger, A.E., 1994. A comparison of two methods for surveying mortality of
beached birds in British Columbia. The Canadian Veterinary Journal 35, 631–
635.
Talsness, C.E., Andrade, A.J.M., Kuriyama, S.N., Taylor, J.A., vom Saal, F.S., 2009.
Components of plastic: Experimental studies in animals and relevance for human
health. Philosophical Transactions of the Royal Society B: Biological Sciences
364, 2079–2096. doi:10.1098/rstb.2008.0281
Tanaka, K., Takada, H., Yamashita, R., Mizukawa, K., Fukuwaka, M., Watanuki, Y.,
2015. Facilitated leaching of additive-derived PBDES from plastic by seabirds’
stomach oil and accumulation in tissues. Environmental Science & Technology
49, 11799–11807. doi:10.1021/acs.est.5b01376

81
Tanaka, K., Takada, H., Yamashita, R., Mizukawa, K., Fukuwaka, M., Watanuki, Y.,
2013. Accumulation of plastic-derived chemicals in tissues of seabirds ingesting
marine plastics. Marine Pollution Bulletin 69, 219–222.
doi:10.1016/j.marpolbul.2012.12.010
Taylor, M.L., Gwinnett, C., Robinson, L.F., Woodall, L.C., 2016. Plastic microfibre
ingestion by deep-sea organisms. Scientific Reports 6. doi:10.1038/srep33997
Teuten, E.L., Rowland, S.J., Galloway, T.S., Thompson, R.C., 2007. Potential for plastics
to transport hydrophobic contaminants. Environmental Science & Technology 41,
7759–7764. doi:10.1021/es071737s
Thomas, G.J., Owen, M., Richards, P., 1977. Grit in waterfowl at the Ouse Washes,
England. Wildfowl 28, 136–138.
Tourinho, P.S., Ivar do Sul, J.A., Fillmann, G., 2010. Is marine debris ingestion still a
problem for the coastal marine biota of southern Brazil? Marine Pollution Bulletin
60, 396-401. doi:https://doi.org/10.1016/j.marpolbul.2009.10.013
Trevail, A.M., Gabrielsen, G.W., Kühn, S., van Franeker, J.A., 2015. Elevated levels of
ingested plastic in a high Arctic seabird, the northern fulmar (Fulmarus glacialis).
Polar Biology 38, 975–981. doi:10.1007/s00300-015-1657-4
UK Government, 2016. Microbead ban announced to protect sealife [WWW Document].
URL https://www.gov.uk/government/news/microbead-ban-announced-to-
protect-sealife (accessed 3.17.17).
United Nations Environment Programme, 2014. UNEP Yearbook: Emerging Issues in
our global environment, UNEP Year Book. United Nations Environment
Programme, Nairobi, Kenya, Africa.

82
Upham, P., Whitmarsh, L., Poortinga, W., Purdam, K., Darnton, A., McLachlan, C.,
Devine-Wright, P., 2009. Public attitudes to environmental change: A selective
review of theory and practice (A research synthesis for the living with
environmental change research programme). Living With Environmental Change,
Cardiff, Manchester, UK.
Van Cauwenberghe, L., Janssen, C.R., 2014. Microplastics in bivalves cultured for
human consumption. Environmental Pollution 193, 65–70.
doi:10.1016/j.envpol.2014.06.010
Van Cauwenberghe, L., Vanreusel, A., Mees, J., Janssen, C.R., 2013. Microplastic
pollution in deep-sea sediments. Environmental Pollution 182, 495–499.
doi:10.1016/j.envpol.2013.08.013
van Franeker, J.A., 2004. Save the North Sea fulmar-Litter-EcoQO manual part 1:
Collection and dissection procedures. Alterra, Wageningen Alterra-rapport 672,
38.
van Franeker, J.A., Blaize, C., Danielsen, J., Fairclough, K., Gollan, J., Guse, N., Hansen,
P.-L., Heubeck, M., Jensen, J.K., Olsen, B., Olsen, K.-O., Pedersen, J., Stienen,
E.W.M., Turner, D.M., 2011. Monitoring plastic ingestion by the northern fulmar
Fulmarus glacialis in the North Sea. Environmental Pollution 159, 2609–2615.
doi:10.1016/j.envpol.2011.06.008
van Franeker, J.A., Law, K.L., 2015. Seabirds, gyres and global trends in plastic
pollution. Environmental Pollution 203, 89–96. doi:10.1016/j.envpol.2015.02.034
van Franeker, J.A., Luttik, R., 2008. Colour and size variation in the northern fulmar
Fulmarus glacialis on Bear Island, Svalbard. Circumpolar Studies 4, 39–58.

83
van Franeker, J.A., Meijboom, A., 2002. LITTER NSV, Marine litter monitoring by
northern fulmars (a pilot study). Wageningen, Alterra, Green World Research,
Alterra-rapport Alterra-rapport 401, 72.
van Franeker, J.A., Wattel, J., 1982. Geographical variation of the Fulmar Fulmarus
glacialis in the North Atlantic. Ardea 70, 31–44.
Van Oostdam, J., Gilman, A., Dewailly, E., Usher, P., Wheatley, B., Kuhnlein, H., Neve,
S., Walker, J., Tracy, B., Feeley, M., Jerome, V., Kwavnick, B., 1999. Human
health implications of environmental contaminants in Arctic Canada: A review.
Science of the Total Environment 230, 1–82.
Venables, W.N., Ripley, B.D., 2002. Random and mixed effects, in: Modern Applied
Statistics with S. Springer, pp. 271–300.
Verlis, K.M., Campbell, M.L., Wilson, S.P., 2013. Ingestion of marine debris plastic by
the wedge-tailed shearwater Ardenna pacifica in the Great Barrier Reef, Australia.
Marine Pollution Bulletin 72, 244–249. doi:10.1016/j.marpolbul.2013.03.017
Votier, S.C., Archibald, K., Morgan, G., Morgan, L., 2011. The use of plastic debris as
nesting material by a colonial seabird and associated entanglement mortality.
Marine Pollution Bulletin 62, 168–172. doi:10.1016/j.marpolbul.2010.11.009
Wabnitz, C., Nichols, W.J., 2010. Plastic pollution: An ocean emergency. Marine Turtle
Newsletter 129, 1–4.
Walther, I., Daoust, P.Y., Lucas, Z., 2008. Plastic ingestion in northern fulmars
(Fulmarus glacialis) in the western North Atlantic. Wildlife Health Centre
Newsletter 13, 5–6.

84
Wickham, H., 2009. ggplot2: Elegant graphics for data analysis, Use R! Springer
International Publishing, New York, New York, USA.
Wickham, H., Francois, R., 2015. dplyr: A grammar of data manipulation. R package
version 0.5.0 [WWW Document]. URL https://cran.r-
project.org/web/packages/dplyr/index.html (accessed 3.19.17).
Wilcox, C., Van Sebille, E., Hardesty, B.D., 2015. Threat of plastic pollution to seabirds
is global, pervasive, and increasing. Proceedings of the National Academy of
Sciences 112, 11899–11904. doi:10.1073/pnas.1502108112
Wille, M., McBurney, S., Robertson, G.J., Wilhelm, S.I., Blehert, D.S., Soos, C.,
Dunphy, R., Whitney, H., 2016. A pelagic outbreak of Avian Cholera in North
American gulls: Scavenging as a primary mechanism for transmission? Journal of
Wildlife Diseases 52, 793–802. doi:10.7589/2015-12-342
Wilson, A.J., 1999. Gunshot injuries: What does a radiologist need to know?
RadioGraphics 19, 1358–1368. doi:10.1148/radiographics.19.5.g99se171358
Wold, S., Esbensen, K., Geladi, P., 1987. Principal component analysis. Chemometrics
and Intelligent Laboratory Systems 2, 37–52. doi:10.1016/0169-7439(87)80084-9
Woodall, L.C., Sanchez-Vidal, A., Canals, M., Paterson, G.L.J., Coppock, R., Sleight, V.,
Calafat, A., Rogers, A.D., Narayanaswamy, B.E., Thompson, R.C., 2014. The
deep sea is a major sink for microplastic debris. Royal Society Open Science 1,
140317–140317. doi:10.1098/rsos.140317
Work, T.M., Rameyer, R.A., 1999. Mass stranding of wedge-tailed shearwater chicks in
Hawaii. Journal of Wildlife Diseases 35, 487–495. doi:10.7589/0090-3558-
35.3.487

85
Wright, S.L., Thompson, R.C., Galloway, T.S., 2013. The physical impacts of
microplastics on marine organisms: A review. Environmental Pollution 178, 483–
492. doi:10.1016/j.envpol.2013.02.031
Ye, S., Andrady, A.L., 1991. Fouling of floating plastic debris under Biscayne Bay
exposure conditions. Marine Pollution Bulletin 22, 608–613. doi:10.1016/0025-
326X(91)90249-R
Yorio, P., Caille, G., 1999. Seabird interactions with coastal fisheries in Northern
Patagonia: Use of discards and incidental captures in nets. Waterbirds 22, 207.
doi:10.2307/1522209
Zbyszewski, M., Corcoran, P.L., 2011. Distribution and degradation of fresh water plastic
particles along the beaches of Lake Huron, Canada. Water, Air & Soil Pollution
220, 365–372. doi:10.1007/s11270-011-0760-6
Zbyszewski, M., Corcoran, P.L., Hockin, A., 2014. Comparison of the distribution and
degradation of plastic debris along shorelines of the Great Lakes, North America.
Journal of Great Lakes Research 40, 288–299. doi:10.1016/j.jglr.2014.02.012
Zhao, S., Zhu, L., Li, D., 2016. Microscopic anthropogenic litter in terrestrial birds from
Shanghai, China: Not only plastics but also natural fibers. Science of The Total
Environment 550, 1110–1115. doi:10.1016/j.scitotenv.2016.01.112

86
Appendices
Appendix A. Details on each bird that ingested anthropogenic debris in this study. N = number of anthropogenic debris fragments
recovered per-bird. GWFG = greater white-fronted goose; SNGO = snow goose; CAGO = Canada goose; AMWI = American wigeon;
ABDU = American black duck; MALL = mallard; NOPI = northern pintail; WWSC = white-winged scoter; COEI = common eider;
YBLO = yellow-billed loon (data on all birds, including those that did not ingest plastic, are available from the authors).
Plastic (N) Rubbish (N) Metal (N) Foraging niche
Species Location Sex Age Fragments Other Thread-like
Foil Paint chips
Glass Rubber Birdshot Metal
Geese GWFG 49°11'41 N 123°10'48 W
M Juvenile 0 0 0 0 0 0 0 0 1
SNGO 49°11'41 N 123°11'02 W
M Juvenile 0 0 1 0 0 0 0 0 0
SNGO 49°11'41 N 123°10'48 W
N/A Juvenile 1 0 0 0 0 0 0 0 0
SNGO 49°04'34 N 123°09'37 W
F Juvenile 0 0 0 0 0 0 0 0 1
CAGO 44°50'29 N 65°17'22 W
N/A Unknown 0 0 0 0 1 0 0 0 0
CAGO 49°11'41 N 123°11'02 W
M Adult 0 0 0 2 0 0 0 0 0
CAGO 49°10'27 N 121°54'38 W
M Adult 0 0 0 2 0 0 0 0 0
CAGO 49°10'27 N 121°54'38 W
F Adult 0 0 0 1 0 0 0 0 0
CAGO 49°10'12 N 121°54'36 W
F Adult 0 0 0 0 0 1 0 0 0
CAGO 49°10'12 N 121°54'36 W
F Adult 0 0 0 0 0 1 0 0 0

87
Plastic (N) Rubbish (N) Metal (N) Foraging niche
Species Location Sex Age Fragments Other Thread-like
Foil Paint chips
Glass Rubber Birdshot Metal
CAGO 44°50'29 N 65°17'22 W
N/A Unknown 2 0 0 0 0 0 0 0 0
CAGO 44°50'29 N 65°17'22 W
N/A Unknown 0 1 0 0 0 0 0 0 0
Dabbling Ducks
AMWI 49°06'22 N 122°05'31 W
F Adult 1 0 0 0 0 0 0 0 0
AMWI 48°59'34 N 122°13'30 W
M Adult 0 0 0 0 0 1 0 0 0
AMWI 49°04'35 N 123°09'39 W
M Adult 1 0 0 0 0 0 0 0 0
ABDU 44°48'06 N 65°23'57 W
N/A Unknown 0 0 0 0 0 0 0 1 0
MALL 44°48'05 N 65°23'57 W
N/A Unknown 0 0 0 0 0 0 0 3 0
MALL 49°12'50 N 121°46'57 W
M Adult 3 0 0 0 0 0 0 0 0
MALL 49°12'50 N 121°46'57 W
M Adult 0 0 0 0 0 0 0 0 1
MALL 49°09'27 N 122°34'48 W
F Adult 0 0 0 4 0 0 0 0 0
MALL 49°12'50 N 121°46'58 W
M Adult 0 0 0 0 0 0 0 1 0
MALL 49°12'50 N 121°46'57 W
M Adult 0 0 0 0 0 0 0 1 0
MALL 49°06'41 N 123°04'53 W
F Adult 1 0 0 0 0 0 0 0 0
MALL 49°12'50 N 121°46'58 W
F Adult 0 0 0 0 0 0 57 0 0
MALL 49°12'50 N 121°46'58 W
M Adult 0 0 0 0 0 1 0 0 0
MALL 49°16'00 N 121°43'12 W
F Adult 1 0 0 0 0 0 0 0 0
MALL 49°12'50 N 121°46'58 W
N/A Unknown 0 0 0 0 0 0 0 2 0
88
Plastic (N) Rubbish (N) Metal (N) Foraging niche
Species Location Sex Age Fragments Other Thread-like
Foil Paint chips
Glass Rubber Birdshot Metal
MALL 62°44'06 N 115°42'35 W
N/A Unknown 1 0 0 0 0 0 0 0 0
MALL 62°44'06 N 115°42'35 W
N/A Unknown 0 0 0 0 0 0 0 1 0
MALL 62°44'06 N 115°42'35 W
N/A Unknown 1 0 0 0 0 0 0 0 0
MALL 45°06'26 N 64°39'19 W
F Adult 1 0 0 0 0 0 0 0 0
NOPI 49°04'44 N 123°02'43 W
M Adult 0 0 0 0 0 1 0 1 0
NOPI 49°06'41 N 123°04'48 W
F Adult 1 0 0 0 0 0 0 0 0
Diving Ducks
WWSC 43°19'31 N 79°47'56 W
N/A Unknown 1 0 0 0 0 0 0 0 0
Sea Ducks COEI 49°48'44 N 54°07'07 W
M Adult 1 0 0 0 0 0 0 0 0
COEI 49°48'44 N 54°07'07 W
F Adult 0 0 0 0 0 0 0 0 1
COEI 49°48'44 N 54°07'07 W
M Juvenile 0 0 0 0 0 0 0 1 0
COEI Unknown
F Adult 0 0 0 1 0 0 0 0 0
Loons YBLO 63°26'04 N 109°11'10 W
N/A Juvenile 1 0 0 0 0 0 0 0 0

89
Appendix B. Summary of pathology and microbiology findings of note from 39 procellariids collected beached on Sable Island, Nova
Scotia, Canada, between 2000 and 2012. GRSH = great shearwater; Imm. = Immature; NOFU = northern fulmar; NSF = No
significant findings; Sp. = Species; SOSH = sooty shearwater.
Sp. Date (D/M/Y) Age Sex Weight
(g) Gross lesions Histological lesions Debris (g)
SOSH 03/06/2008 Imm Female 524.2 Emaciation Chronic multifocal coelomic fat necrosis 0.0087
SOSH 04/07/2010 Imm. Male 441.5 Chronic ventricular ulcer; emaciation Intralesional nematodes 0.0000
GRSH 17/06/2005 Adult Female NA Emaciation Moderate muscular sarcocystosis 0.0000
GRSH 20/06/2006 Imm. Male 764.5 Chronic locally extensive cellulitis, esophageal perforation (fish bone); emaciation
NSF
0.0798
GRSH 23/06/2006 NA NA 492.1 Emaciation Mild muscular sarcocystosis 0.0781 GRSH 01/07/2006 Adult Female 506.4 Emaciation Focally extensive subacute myopathy 0.1327 GRSH 23/08/2006 NA Male 568.3 Emaciation Acute multifocal hepatic necrosis 0.0957
GRSH 29/06/2007 Imm. Female 736.8 Coelomic cavity infection Proventricular parasitic infection, transmural proventriculitis, fibrinous perihepatitis, fibrinous air sacculitis
0.1346
GRSH 01/09/2010 Imm Male NA Trauma, multifocal renal hemorrhages; emaciation NSF 0.1663
GRSH 07/07/2011 Imm. Male 528.4 Emaciation Focal chronic parasitic proventriculitis 0.0395
GRSH 07/07/2011 Imm. Males 471.9 Proventricula perforation; emaciation
Focal chronic transmural proventriculitis, chronic fibrinous serositis
0.0182
GRSH 14/09/2011 Imm Male 717.9 Trauma fibrinopurulent cellulitis 0.0000 GRSH NA Adult Male 511.7 Chronic focal cellulitis; emaciation NSF 0.9740 NOFU 01/05/2004 Adult Male 751.2 Emaciation Moderate muscular sarcocystosis 0.1780 NOFU 01/05/2004 Imm. Male 847.6 Emaciation Hepatic neoplasia 0.3370 NOFU 04/05/2004 Adult Female NA Egg impaction, autolysis; emaciation NSF 4.2050 NOFU 22/10/2004 NA Male 402.7 Emaciation Chronic pericarditis, air sacculitis 0.0590 NOFU 04/04/2005 Imm. Male NA NSF Chronic focal myocarditis 4.4720
NOFU 16/04/2005 Adult Male NA Multifocal subcutaneous fibromas; emaciation NSF 1.3760
NOFU 01/06/2005 NA Male 650.0 Focal subcutaneous fibroma Cartilaginous metaplasia 0.7040

90
Sp. Date (D/M/Y) Age Sex Weight
(g) Gross lesions Histological lesions Debris (g)
NOFU 21/12/2005 Adult Female 677.2 NSF Acute multifocal pectoral myopathy, mild (multi-) focal granulomatous hepatitis
0.0773
NOFU 22/12/2005 Imm. Male 659.6 Emaciation Moderate acute pectoral myopathy with mineralization.
0.0000
NOFU 13/04/2006 Imm Male NA NSF Chronic intracoelomic fat necrosis 3.6307
NOFU 04/05/2006 Imm. Female 604.7 NSF Mild renal parasitism (trematodes), mild muscular sarcocystosis, and mild subacute multifocal (pectoral) myositis
0.4839
NOFU 26/02/2007 Imm. Female 630.7 NSF acute septicemia 0.0096 NOFU 01/03/2007 Imm. NA 869.0 NSF Sarcosystis 0.0000 NOFU 13/06/2007 Imm. Male 567.3 Emaciation Severe intestinal cestodiasis 0.1583 NOFU 28/02/2007 NA Male 881.4 Subacute periarterial abscesses NSF 0.3080 NOFU 29/04/2007 Imm. Female NA Emaciation Focal mycotic air sacculitis. 0.0871
NOFU 04/01/2008 Imm Female NA Chronic focal traumatic proventriculitis (fish hook) NSF 0.9322
NOFU 30/05/2008 Imm. Female 604.6 Emaciation Chronic focal pyogranulomatous air sacculitis. 0.0000 NOFU 06/06/2008 Imm. Male 540.9 Intestinal cestodiasis NSF 0.0600 NOFU 07/04/2009 Imm. Male NA Emaciation Mild pulmonary nematodiasis 0.5993 NOFU 09/04/2009 Imm. NA NA Podpdermatitis, emaciation NSF 0.7583 NOFU 27/09/2010 Imm. Male NA Fibrochodroma; emaciation NSF 0.1042 NOFU 15/05/2011 Imm. NA 603.5 NSF Hepatic and splenic mycobacteriosis 2.5595
NOFU 05/06/2011 Adult Male 745.3 Severe unilateral chronic necrotizing orchitis and funiculitis; emaciation NSF 0.1817
NOFU 05/06/2011 Adult Female NA Chronic fibrinous oophoritis and peritonitis; emaciation NSF 0.3575
NOFU 20/01/2012 Imm. Male 557.2 Emaciation Chronic focal cellulitis 0.0222

91
Appendix C. Pathology of four procellariid species collected beached on Sable Island, Nova Scotia, Canada, between 2000-2012, and
comparisons to previous pathological descriptions in the literature. “This study” indicates pathological reports new to these species,
determined by a literature review searching species name and the pathological symptom. “-” indicates no symptoms were found.
Pathology Cory’s Shearwater
Sooty Shearwater
Great Shearwater
Northern Fulmar
Starvation Mougin et al., 2000
Grant et al., 1994 Pierce et al., 2004 Harvey et al., 2004
Autolysis This study Humphries, 2014 This study Harris et al., 2006 Parasite Infection Sarcocystosis - - This study This study Intestinal cestodiasis - - - Mallory et al., 2007 Parasitic proventriculitis - - Nemeth et al., 2011 - Pulmonary nematodiasis - - - Mallory et al. ,2007 Renal parasitism
(trematodes) - - - This study
Inflammation Air sacculitis - - - Jessup et al., 2009 Myocarditis - - - Kalmar et al., 2014 Oophoritis and peritonitis - - - This study Periarterial abscesses - - - This study Pericarditis and air
sacculitis - - - Jessup et al., 2009
Pododermatitis (bumblefoot)
- - - This study
Proventriculitis and serositis
- - Nemeth et al., 2012 -
Trauma - - Haman et al., 2013 van Franeker and Meijboom, 2002

92
Pathology Cory’s Shearwater
Sooty Shearwater
Great Shearwater
Northern Fulmar
Bacterial Infection Cellulitis - - This study Li et al., 2015 Chronic ventricular ulcer - This study - - Mycobacteriosis - - - This study Orchitis and funiculitis - - - This study Septicemia - - - This study Drowned - Reyes-Arriagada et al., 2007 Yorio and Caille, 1999 - Tumour Fibromas - - - Pokras, 1988 Fibrochodroma - - - Pokras, 1988 Neoplasia - - - Gaynor et al., 2015 Tissue Necrosis Fat necrosis - This study - This study Hepatic necrosis - - This study - Impaction Egg impaction - - - Walther et al., 2008 Rectal Impaction - - - van Franeker and
Meijboom, 2002 Myopathy - - This study This study Pneumonia - - - Bellizzi and Duerr, 2013
Copyright © 2022 FDOKUMEN




















