
Analysis of genetic similarities among species of Fragaria , Potentilla , and Duchesnea found in...
10
Analysis of genetic similarities among species of Fragaria, Potentilla, and Duchesnea found in northwest Argentina by using morphological, anatomical, and molecular characters Marta Ontivero, Marta Arias, Juan Diaz Ricci, Judith Babot, Patricia Albornoz, and Atilio Castagnaro Abstract: Morphological, anatomical, and molecular techniques were used to characterize wild strawberry and wild strawberry-like species in northwest Argentina. Characteristics of leaves, flowers, runners, achenes, and genomic DNA polymorphisms were used to analyze similarities among Potentilla tucumanensis Castagnaro & Arias, Duchesnea indica (Andr.) Focke, and Fragaria vesca L. Comparison of phenograms obtained by using morphological and anatomical traits or genomic DNA characters revealed similar clustering of the species. Both phenograms suggest that D. indica is more closely related to P. tucumanensis than to F. vesca. Using the randomly amplified polymorphic DNA (RAPD) technique with specific primers, we detected polymorphic bands that permit the identification of P. tucumanensis, D. in- dica, and F. vesca. In addition, we report new morphological and anatomical characters that can be used as diagnostic traits for better identification of species in reproductive and vegetative states. Key words: Fragaria, Potentilla, Duchesnea, RAPD, DNA fingerprinting, morphological traits, anatomical traits. Résumé: Les auteurs ont utilisé des techniques morphologiques, anatomiques et moléculaires pour caractériser les espè- ces de fraises sauvages ainsi que les espèces qui leur ressemblent, dans le nord-ouest de l’Argentine. Ils ont utilisé les caractéristiques des feuilles, des fleurs, des stolons, des achènes et des polymorphismes de l’ADN génomique, pour analyser les similitudes entre les Potentilla tucumanensis Castagnaro & Aries, Duchesnea indica (Andr.) Focke et Fra- garia vesca L. Une comparaison des phénogrammes obtenus en utilisant les caractéres morphologiques et atomiques ou de l’ADN génomique, montrent un même regroupement pour ces espèces. Les deux phénogrammes suggèrent que le D. indica est plus étroitement apparenté au P. tucumanensis qu’au F. vesca. L’analyse d’ADN polymorphe amplifié au hasard (RAPD) avec des amorçes spécifiques montrent des bandes polymorphiques qui permettent d’identifier les P. tu- cumanensis, D. indica, et F. vesca. De plus, les auteurs rapportent de nouveaux caractères anatomo-morphologiques di- gnostiques qui peuvent être utilisés pour mieux identifier ces espèces à l’ état végétatif ou reproducteur. Mots clés : Fragaria, Potentilla, Duchesnea, RAPD, empreinte ADN, caractères morphologiques, caractères anatomiques. [Traduit par la Rédaction] Ontivero et al. 556 Introduction Genetic diversity is considered a critical factor in crop im- provement. In our laboratory, we have started a breeding program with the aim to obtain novel lines of the cultivated strawberry Fragaria × ananassa Duch. adapted to local en- vironmental conditions. Consequently, the first step of our program was to characterize wild strawberry and strawberry- like species growing in northwest Argentina, to study their genetic relationships, and to assess genetic diversity. Wild germplasm is considered a valuable resource of novel genes, including resistance to disease or environmental stresses (Harrison et al. 1993; Hancock and Luby 1993). In northwest Argentina, wild Rosaceae species related to F. × ananassa span a wide range of ecological habitats (Arias et al. 1998) and belong to the closely related genera Fragaria, Duchesnea, and Potentilla (Ellis 1961). In the ge- nus Fragaria, 18 species have been described (Hancock and Luby 1993), two of which are found in Argentina: F. chi- loensis (L.) Duch. and F. vesca L. (Zardini 1999). The genus Potentilla consists of about 500 species (Davidson 1995), with two species described in Argentina: P. anserina L. and Can. J. Bot. 78: 547–556 (2000) © 2000 NRC Canada 547 Received April 9, 1999. M. Ontivero, J.D. Ricci, J. Babot, and A. Castagnaro. 1 Instituto de Química Biológica “Dr.Bernabé Bloj,” Facultad de Bioquímica, Química y Farmacia, Instituto Superior de Investigaciones Biológicas, Departamento de Bioquímica de la Nutrición, Universidad Nacional de Tucumán, Chacabuco 461, 4000 - Tucumán, Argentina. M. Arias and P. Albornoz. Facultad de Ciencias Naturales, Instituto Miguel Lillo and Fundación Miguel Lillo, Miguel Lillo 205, 4000 - Tucumán, Argentina. 1 Author to whom all correspondence should be addressed (e-mail: [email protected]).
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Analysis of genetic similarities among species of Fragaria , Potentilla , and Duchesnea found in...

Analysis of genetic similarities among species ofFragaria, Potentilla, and Duchesnea found innorthwest Argentina by using morphological,anatomical, and molecular characters
Marta Ontivero, Marta Arias, Juan Diaz Ricci, Judith Babot, Patricia Albornoz,and Atilio Castagnaro
Abstract: Morphological, anatomical, and molecular techniques were used to characterize wild strawberry and wildstrawberry-like species in northwest Argentina. Characteristics of leaves, flowers, runners, achenes, and genomic DNApolymorphisms were used to analyze similarities amongPotentilla tucumanensisCastagnaro & Arias,Duchesnea indica(Andr.) Focke, andFragaria vescaL. Comparison of phenograms obtained by using morphological and anatomicaltraits or genomic DNA characters revealed similar clustering of the species. Both phenograms suggest thatD. indica ismore closely related toP. tucumanensisthan toF. vesca. Using the randomly amplified polymorphic DNA (RAPD)technique with specific primers, we detected polymorphic bands that permit the identification ofP. tucumanensis, D. in-dica, and F. vesca. In addition, we report new morphological and anatomical characters that can be used as diagnostictraits for better identification of species in reproductive and vegetative states.
Key words: Fragaria, Potentilla, Duchesnea, RAPD, DNA fingerprinting, morphological traits, anatomical traits.
Résumé: Les auteurs ont utilisé des techniques morphologiques, anatomiques et moléculaires pour caractériser les espè-ces de fraises sauvages ainsi que les espèces qui leur ressemblent, dans le nord-ouest de l’Argentine. Ils ont utilisé lescaractéristiques des feuilles, des fleurs, des stolons, des achènes et des polymorphismes de l’ADN génomique, pouranalyser les similitudes entre lesPotentilla tucumanensisCastagnaro & Aries,Duchesnea indica(Andr.) Focke etFra-garia vescaL. Une comparaison des phénogrammes obtenus en utilisant les caractéres morphologiques et atomiques oude l’ADN génomique, montrent un même regroupement pour ces espèces. Les deux phénogrammes suggèrent que leD. indica est plus étroitement apparenté auP. tucumanensisqu’au F. vesca. L’analyse d’ADN polymorphe amplifié auhasard (RAPD) avec des amorçes spécifiques montrent des bandes polymorphiques qui permettent d’identifier lesP. tu-cumanensis, D. indica,et F. vesca. De plus, les auteurs rapportent de nouveaux caractères anatomo-morphologiques di-gnostiques qui peuvent être utilisés pour mieux identifier ces espèces à l’ état végétatif ou reproducteur.
Mots clés: Fragaria, Potentilla, Duchesnea, RAPD, empreinte ADN, caractères morphologiques, caractères anatomiques.
[Traduit par la Rédaction] Ontivero et al. 556
Introduction
Genetic diversity is considered a critical factor in crop im-provement. In our laboratory, we have started a breedingprogram with the aim to obtain novel lines of the cultivatedstrawberryFragaria × ananassaDuch. adapted to local en-vironmental conditions. Consequently, the first step of ourprogram was to characterize wild strawberry and strawberry-like species growing in northwest Argentina, to study theirgenetic relationships, and to assess genetic diversity. Wildgermplasm is considered a valuable resource of novel genes,
including resistance to disease or environmental stresses(Harrison et al. 1993; Hancock and Luby 1993).
In northwest Argentina, wild Rosaceae species related toF. × ananassaspan a wide range of ecological habitats(Arias et al. 1998) and belong to the closely related generaFragaria, Duchesnea, andPotentilla (Ellis 1961). In the ge-nusFragaria, 18 species have been described (Hancock andLuby 1993), two of which are found in Argentina:F. chi-loensis(L.) Duch. andF. vescaL. (Zardini 1999). The genusPotentilla consists of about 500 species (Davidson 1995),with two species described in Argentina:P. anserinaL. and
Can. J. Bot.78: 547–556 (2000) © 2000 NRC Canada
547
Received April 9, 1999.
M. Ontivero, J.D. Ricci, J. Babot, and A. Castagnaro.1 Instituto de Química Biológica “Dr.Bernabé Bloj,” Facultad deBioquímica, Química y Farmacia, Instituto Superior de Investigaciones Biológicas, Departamento de Bioquímica de la Nutrición,Universidad Nacional de Tucumán, Chacabuco 461, 4000 - Tucumán, Argentina.M. Arias and P. Albornoz. Facultad de Ciencias Naturales, Instituto Miguel Lillo and Fundación Miguel Lillo, Miguel Lillo 205,4000 - Tucumán, Argentina.
1Author to whom all correspondence should be addressed (e-mail: [email protected]).
J:\cjb\cjb78\cjb-04\B00-026.vpWednesday, May 17, 2000 9:43:17 AM
Color profile: Generic CMYK printer profileComposite Default screen

P. tucumanensisCastagnaro & Arias (Zardini 1999). The ge-nus Duchesneaconsists of a single worldwide distributedspecies,D. indica (Andr.) Focke, which is also found in Ar-gentina (Zardini 1999).
In the northwest of Argentina, only one wild strawberryspecies,F. vesca, and two strawberry-like species,P. tucu-manensisand D. indica, are found. These species can bewell differentiated through classic taxonomic criteria, whichconsidered mostly floral and fruit traits (Zardini 1984;Schulze-Menz 1964). However, confusion may arise whenplants of these species are observed in a vegetative state, be-cause critical morphological differences among them havenot been described (Zardini 1984). An example of this con-fusion is the case ofD. indica, which was initially namedFragaria indica (Jackson 1895). Later, Wolf (1908) pro-posed the transfer of this species to the genusPotentilla, andKalkman (1968), following the same criteria, named thisspeciesP. indica. Currently most taxonomic authors agree,based mainly on reproductive traits, thatD. indica belongsto a separate genus (Hara and Kurosawa 1959; Schulze-Menz 1964; Zardini 1999). In the case ofP. tucumanensis, itwas initially classified asP. norvegicabut after further stud-ies it has been retained as a new species within the genusPotentilla (Castagnaro et al. 1998).
To characterize the species found in northwestern Argen-tina, we have followed the taxonomic criteria proposed byEngler (Schulze-Menz 1964) but complemented with newand more discriminating morphological and anatomical char-acters presented in this communication.
Since the aim of this study is to evaluate relationshipsamong F. vesca, P. tucumanensis, and D. indica by usingnumerical taxonomic methods, we included a large numberof morphological and anatomical characters observed in re-productive and vegetative stages. It was also our concern toanalyze whether relationships based on morphological andanatomical traits agreed with relationships based on mo-lecular characters. For this purpose, we used the randomlyamplified polymorphic DNA (RAPD) technique to evaluategenomic similarities amongF. vesca, P. tucumanensis, andD. indica. SinceF. × ananassamight be the potential recep-tor of genes coming from these wild species, it was includedin the molecular analysis.
Recently, molecular techniques have provided additionaltools for examining variation among populations and eluci-dating taxonomic positions. The RAPD technique has beenused to study genetic diversity inBrassica (Demeke et al.1992),Amaranthus(Transue et al. 1994), andOryza(Yu andNguyen 1994). Hancock and Callow (1994) used RAPDanalysis to examine variation among strawberry cultivars,and more recently, Harrison et al. (1997b) have reported theuse of this technique to assess genetic relationships amongpopulations of wild octoploids ofFragaria from variousgeographic regions of the United States. For assessing rela-tionships among populations ofFragaria, Harrison (1994)determined that RAPDs, complemented with morphologicalcharacters provided more information than restriction frag-ment length polymorphism (RFLP) alone, due to a lowRFLP variability found among these populations. There areonly two reports ofFragaria, Duchesnea, andPotentilla re-lationships based on RFLP molecular markers of the chloro-plast genome. Both show thatFragaria is more closely
related to Potentilla than to Duchesnea(Harrison et al.1993, 1997a).
In this work, we evaluate the relationships amongF. vesca,P. tucumanensis, andD. indica by using morphological, ana-tomical, and genomic characters, and provide a more com-plete description of these species.
Materials and methods
Plant materialTo include a wide range of ecological habitats of wild strawber-
ries and strawberry-like species growing in Tucumán, specimens ofF. vesca, D. indica, and P. tucumanensiswere collected from dif-ferent sites of the region and cultivated in a greenhouse. The culti-vars of F. × ananassawere obtained from Estación ExperimentalObispo Colombres, Tucumán (EEAOC). Table 1 shows the list ofaccession numbers, provenance information, and the number of in-dividuals used in this study.
Brief description of speciesThe four species under study are herbaceous, 5–30 cm in height,
with pubescent three-foliolate leaves and hermaphroditic flowerswith five petals.Fragaria vesca: perennial herb, runnering, whiteflowers in an inflorescence, fruiting receptacle fleshy red, achenessuperficial (Staudt 1962).Duchesnea indica: perennial herb, run-nering, solitary yellow flowers, fruiting receptacle fleshy red,achenes prominent (Focke 1888).Potentilla tucumanensis: annualherb, runners absent, solitary yellow flowers, fruiting receptacledry brown, achenes sunken (Castagnaro et al. 1998).Fragaria ×ananassa: is a hybrid derived from the wild American speciesF. chiloensis (L.) Duch., andF. virginiana Duch.; annual herb,runnering, white flowers in an inflorescence, fruiting receptaclefleshy red, achenes superficial (Duchesne 1766; Darow 1966).
Morphological and anatomical studiesThe morphological and anatomical characters evaluated are
listed in Table 2. Morphological characters were observed macro-scopically on fresh and dried material grown in the field and in thegreenhouse. For microscopic examination, freshly collected leavesobtained from 5 or 10 individuals of each of the accessions listedin Table 1 were fixed in formalin – acetic acid – alcohol (FAA) for2 or 3 days and then preserved in 70% ethyl alcohol. Fixed mate-rial was processed according to Strittmather (1986) for light mi-croscopy and scanning electron microscopy (SEM). Samples formicrotome sectioning were dehydrated through a tertiary-butanolseries, infiltrated with liquid paraffin, sectioned at 10–15µm, andstained with Safranin – fast green. Sections were observed by nor-mal- and polarized-light microscopy. Samples for SEM examina-tion were stepwise dehydrated with increasing concentrations ofethanol, stored in 100% acetone, critical-point dried, and gold–pal-ladium sputter-coated.
Molecular studies
DNA extractionTotal DNA was extracted from young leaves of each individual
of all accessions studied, following the CTAB (hexadecyltrimethyl-ammonium bromide) procedure described by Rogers and Bendich(1988). DNA concentration was determined at 260 and 280 nmwith a Gilson spectrophotometer. Quality of the DNA was evalu-ated by electrophoresis in 0.7% agarose gel stained with ethidiumbromide (0.5µg/mL). Buffer and running conditions were accord-ing to Sambrook et al. (1989).
© 2000 NRC Canada
548 Can. J. Bot. Vol. 78, 2000
J:\cjb\cjb78\cjb-04\B00-026.vpWednesday, May 17, 2000 9:43:17 AM
Color profile: Generic CMYK printer profileComposite Default screen

Polymerase chain reactionDNA from D. indica, F. vesca, F. × ananassa, andP. tucuman-
ensis were analyzed with a set of 10 random-sequence 10-merprimers (OPJ series from Operon Technologies). Samples of DNAwere obtained by pooling equimolar quantities of independent ex-tractions of DNA from each of the accessions under study. The setof 10 primers used in experiments (Table 3) was chosen based onthe clarity and reproducibility of banding patterns in preliminaryscreening. Reaction mixtures were prepared in 20µL containing:1.5 mM MgCl2, 0.1 mM of each dNTP (deoxyribonuleotides),0 2. µM primer, 20 ng DNA template, 0.75 U of Taq polymerase(Promega), and 2µL of Taq buffer (×10). The program used was asfollows: denaturation, 1 min at 94°C; annealing, 1 min at 40°C; ex-tension, 2 min at 72°C; number of cycles: 40. The program startedwith a thermal treatment of 5 min at 94°C and finished with an ex-tension of 5 min at 72°C. The reaction was carried out inan Ericomp Delta cycler I (Ericomp Inc., San Diego, Calif.). Am-plification products were separated by electrophoresis in a 1.5%agarose gel in tris–borate–EDTA buffer (TBE) running buffer(Sambrook et al. 1989). Electrophoresis was carried out by apply-ing 2.8 V/cm during the first 20 min and 4 V/cm for 1 h 20 min.Gels were stained with ethidium bromide (0.5µg/mL) for 30 min,washed three times with distilled water, and photographed with aPolaroid camera (Polaroid film 667). RAPD experiments were re-peated three times to avoid false results and to assure reproduc-ibility. Negative controls, without DNA, were included in eachexperiment. Molecular weights were estimated usingEcoRI/HindIII-digested lambda DNA, and PCR markers from Promega.
Data analysisTo build a similarity matrix for numerical analysis, data from
morphological and anatomical observations were scored as present(1) or absent (0) in each of the three species considered. RAPDdata were also scored by analyzing PCR-amplified bands. Photo-graphs of agarose gel electrophoresis were digitized by using aVideo Scope International Ltd. camera, and image processing wascarried out with the Image 1.45 software.
Only bands showing in all replicates were considered and scoredas present (1) in each sample. Minor bands that were diagnostic ofthe rare alleles but could not be scored with certainty in pooledDNA were not included. The similarity matrix was built by consid-ering presence (1) or absence (0) of bands in each sample.
The following nomenclature was used: NA, number of charac-ters (bands) present in samples A and B; NB, number of characters
present in sample A and absent in B; NC, number of characterspresent in sample B and absent in A. Similarities were evaluatedby estimating the Dice coefficient according to: SD = 2NA/(2NA +NB + NC). We have used the Dice (equal to Nei and Li) coeffi-cient because it excludes shared absence as a criterion of similarityand gives more weight to shared than to unshared characters. Fur-thermore, this coefficient has been recommended for the evaluationof genetic similarities when using RAPD data (Lamboy 1994). Wealso verified that similar results were obtained with any coefficient(e.g., simple matching coefficient or Jaccard), when more than 50characters were included in the analysis (data not shown).
Cluster analysis was made by utilizing the unweighted pairgroup method with arithmetic mean (UPGMA) because this is themost widely used method in ecology and systematics (Crisci andLopez-Armengol 1983; van Tongeren 1995). The evaluation ofsimilarity coefficients and cluster analysis were carried out withthe NTSYS program (Rohlf 1993).
Results
A list of all morphological and anatomical characters in-cluded in our analysis is summarized in Table 2. These char-acters permit recognition of each species not only whenplants are in a reproductive but also in a vegetative state.However, characters common to the three species, namely:five petals and hermaphroditic flowers, numerous stamensand styles and ranunculaceous stomata (not shown in Ta-ble 2) were included in the numerical analysis, to keep thesame statistical criteria used for molecular analysis, wherewe considered all molecular markers (see Materials andmethods).
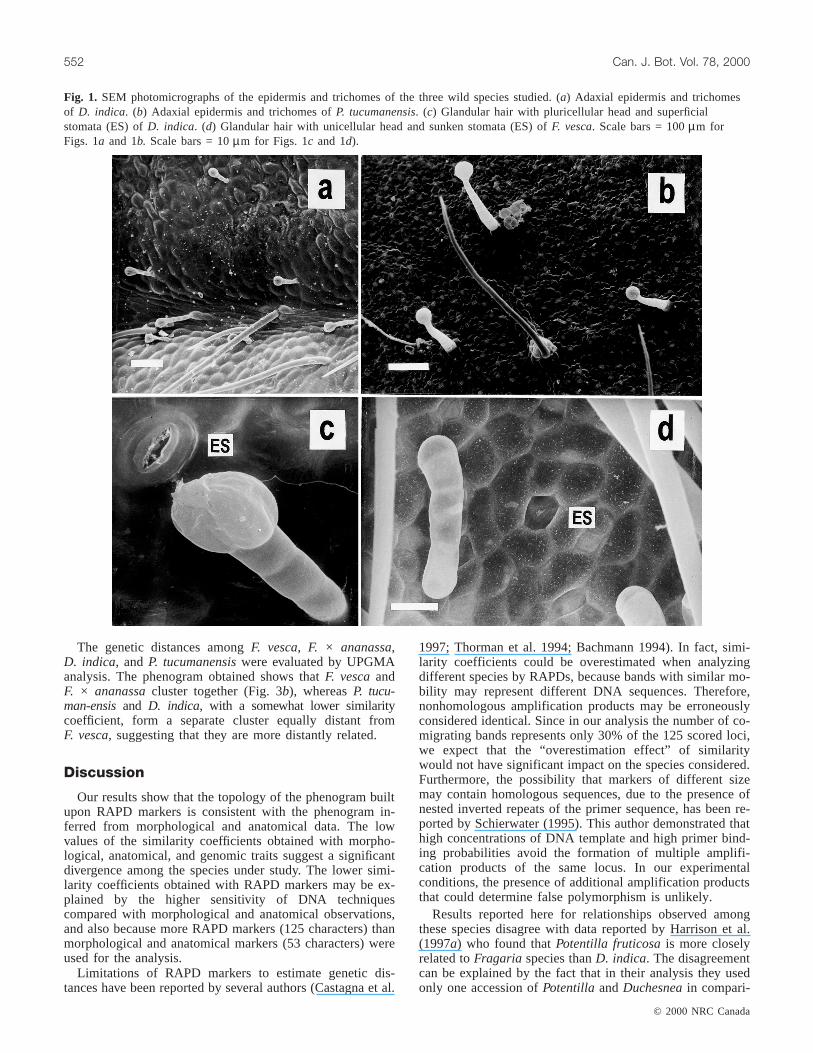
The type of trichomes and stomata observed inD. indica,P. tucumanensis, and F. vescaleaves are shown in Fig. 1.The analysis revealed thatD. indica has glandular hairs withpluricellular heads on the adaxial and abaxial epidermis(Figs. 1a-1c), F. vescahas glandular hairs with unicellularheads on both epidermal surfaces (Fig. 1d), andP. tucuman-ensis has glandular hairs with unicellular and pluricellularheads on the adaxial epidermis (Fig. 1b) and only unicellularheads on the abaxial epidermis (Table 2). We also, observeddifferences among stomata of these species, whereasD. in-dica has superficial (not prominent) stomata (Fig. 1c),
© 2000 NRC Canada
Ontivero et al. 549
Species Accession No. Locality Habitat Elevation (m) Sample size
F. vesca 1 Taficillo Highlands 1600 10F. vesca 2 Villa Nougés Highlands 1400 10P. tucumanensis 1 Taficillo Semiarid 1500 10P. tucumanensis 2 Potrero de Trancas Semiarid 1600 10D. indica 1 Taficillo Subtropical rain forest 1850 5D. indica 2 Tafí del Valle Highland 2000 5D. indica 3 Trancas Semiarid 800 5D. indica 4 Lules Subtropical rain forest 900 5D. indica 5 Potrero de las Tablas Highland 1000 5F. × ananassa
var. Chandler 1 INTA Cultured 600 5var. Sweet Charlie 2 INTA Cultured 600 5var. Pájaro 3 EEAOC Cultured 600 5var. Milsei Tudla 4 INTA Cultured 600 5var. Oso Grande 5 EEAOC Cultured 600 5
Note: INTA, Instituto Nacional de Tecnología Agropecuaria, EEA-Famaillá; EEAOC, Estación Experimental Agro-industrial “ObispoColombres.” Taficillo, Trancas, Tafí del Valle, and Lules are localities in northwest Argentina that have different ecological environments.
Table 1. Locations, habitats, and populations ofFragaria vesca, Potentilla tucumanensis, Duchesnea indica, andFragaria × ananassaused in this study.
J:\cjb\cjb78\cjb-04\B00-026.vpWednesday, May 17, 2000 9:43:18 AM
Color profile: Generic CMYK printer profileComposite Default screen

© 2000 NRC Canada
550 Can. J. Bot. Vol. 78, 2000
Species
Traits F. vesca P. tucumanensis D. indica
RunnersRunnering + – +Runnering with rooting nodes – – +Runner color green – – +Runner color pinkish + – –Stipules tripartite caulinar + – +LeavesStipules bipartite – + –Color dark green – + +Color bright green + – –Venation type semicraspedodromous + – +Venation type brochidodromous – + –Very branched secondary venation – + –Scarcely branched secondary venation – – +Not branched secondary venation + – –Leaf pubescence density: few – – +Leaf pubescence density: intermediate – + –Leaf pubescence density: many hairs + – –Petiole green – + +Petiole pink + – –Petiole pubescence density: few – – +Petiole pubescence density: intermediate – + –Petiole pubescence density: many hairs + – –Petiole pubescence: hairs appressed – + +Petiole pubescence: hairs appressed and ascending + – –Glandular hair with unicellular head in upper epidermis + + –Glandular hair with unicellular head in abaxial epidermis + + –Glandular hair with pluricellular head in upper epidermis – + +Glandular hair with pluricellular head in abaxial epidermis – – +One layer of palisade – + +Three layers of palisade + – –Druse crystals in idioblasts and clustered sandy crystals. + – –Druse crystals in idioblasts – – +Crystals prismatic and small (not in idioblasts) – + –Stomata sunken + – –Stomata superficial – – +Stomata prominent – + –FlowersFlower yellow – + +Flower white + – –Flower, 4–9 mm diameter – + –Flower, 15–25 mm diameter + – +Flowers hermaphroditic + + +Petals lanceolate – + –Petals orbiculate + – +Fruiting receptacle fleshy red + – +Fruiting receptacle dry brown – + –AchenesAchenes color red – – +Achenes color light brown + + –Position of achenes: sunken – + –Position of achenes: superficial + – –Position of achenes: prominent – – +
Note: +, presence of the character state; –, absence of the character state.
Table 2. Morphological and anatomical characters, grouped by plant part, inFragaria vesca, Potentillatucumanensis, and Duchesnea indica.
J:\cjb\cjb78\cjb-04\B00-026.vpWednesday, May 17, 2000 9:43:18 AM
Color profile: Generic CMYK printer profileComposite Default screen

F. vescahas sunken stomata (Fig. 1d) and P. tucumanensishas prominent stomata (Table 2).
Foliar morphology and vasculature and leaf anatomy ofthe three species are illustrated in Fig. 2. Differences in leafvenation (Figs.2a, 2c, and 2e) and anatomy, including num-ber of palisade layers, stomata position, location and type ofcrystals (Figs. 2b, 2d, 2f; Table 2) are evident.
The data obtained from morphological and anatomicalanalysis were used to evaluate relationships among the threewild species. The phenogram shown in Fig. 3a reveals thatF. vescais more distantly related to the other two species,which cluster together.
Preliminary RAPD experiments carried out with templateDNA coming from each accession listed in Table 1 showedsimilar amplification profiles within each taxon for eachprimer. However some faint bands were present in particulargenotypes and absent in others, suggesting the existence ofintraspecific polymorphism (Fig. 4). Since the purpose ofthis study was not to evaluate intraspecific diversity, but the
genetic differences among wild and cultivated species, wepooled equimolar quantities of DNA from all accessions ofeach species, to strengthen common traits and minimize in-traspecific polymorphism. The latter means that each poolcontains DNA of the same species coming from differentecological habitats. Therefore, we assumed that each poolrepresents the genetic diversity of wild germplasm of north-west Argentina.
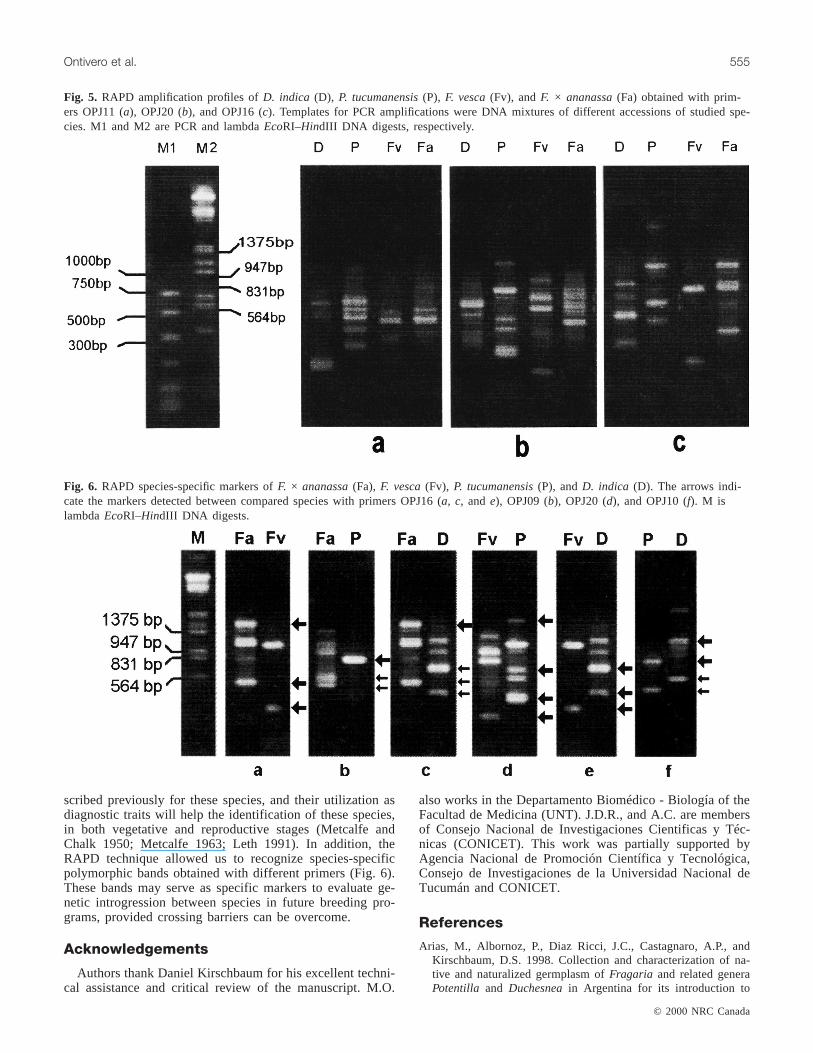
The number and type of RAPD bands obtained with the10 selected primers and for the pooled DNA samples ofD. indica, F. vesca, F. × ananassa, andP. tucumanensisarepresented in Table 3. Results show abundant genomic poly-morphisms among species with all primers used. We de-tected 125 fragments, 87 of which are polymorphic (70%),and the other 38 bands shared (i.e., comigrating or same mo-lecular weight) by two or three of the studied species. Nobands were shared by all four species (Fig. 5). This outcomereveals that a high degree of genetic diversity exists amongthe four species studied.
© 2000 NRC Canada
Ontivero et al. 551
Primers Shared bands
No. Sequence (5′–3′)Total no.of bands
Polymorphic bandsamong Fv, Fa, P, D No.
Speciescompared Size (bp)
OPJ-04 CCGAACACGG 17 14 1 P, D 18001 Fa, D 7501 Fa, D 415
OPJ-05 CTCCATGGGG 5 3 1 Fa, P 10851 Fa, Fv 565
OPJ-08 CATACCGTGG 13 4 1 Fa, Fv, D 12001 Fa, P, D 6902 Fa, D 2100, 14002 Fa, Fv 950, 3752 P, D 1580, 4601 Fv, D 565
OPJ-09 TGAGCCTCAC 10 10 0 — —OPJ-10 AAGCCCGAGG 13 10 1 Fa, D 1735
1 P, D 6551 Fv, D 565
OPJ-11 ACTCCTGCGA 10 7 1 P, D 8501 Fv, D 7001 Fa, Fv 650
OPJ-13 CCACACTACC 15 10 2 P, D 520, 4501 Fa, P 17351 Fv, P 10001 Fa, Fv 830
OPJ-15 TGTAGCAGGG 11 8 1 Fa, Fv, P 6551 P, D 9101 Fa, Fv 395
OPJ-16 CTGCTTAGGG 14 11 1 Fa, Fv, P 10001 Fa, P 14701 Fa, D 1120
OPJ-20 AAGCGGCCTC 17 10 1 Fa, Fv, P 9502 Fa, Fv 1200, 8301 Fa, P 10452 Fa, D 750, 4501 P, D 620
Total characters 125 87 38
Table 3. List of primers, numbers of polymorphic bands, and numbers of bands shared byFragaria vesca(Fv),Potentilla tucumanensis(P), Duchesnea indica(D), and Fragaria × ananassa(Fa).
J:\cjb\cjb78\cjb-04\B00-026.vpWednesday, May 17, 2000 9:43:19 AM
Color profile: Generic CMYK printer profileComposite Default screen
The genetic distances amongF. vesca, F. × ananassa,D. indica, and P. tucumanensiswere evaluated by UPGMAanalysis. The phenogram obtained shows thatF. vescaandF. × ananassacluster together (Fig. 3b), whereasP. tucu-man-ensisand D. indica, with a somewhat lower similaritycoefficient, form a separate cluster equally distant fromF. vesca, suggesting that they are more distantly related.
Discussion
Our results show that the topology of the phenogram builtupon RAPD markers is consistent with the phenogram in-ferred from morphological and anatomical data. The lowvalues of the similarity coefficients obtained with morpho-logical, anatomical, and genomic traits suggest a significantdivergence among the species under study. The lower simi-larity coefficients obtained with RAPD markers may be ex-plained by the higher sensitivity of DNA techniquescompared with morphological and anatomical observations,and also because more RAPD markers (125 characters) thanmorphological and anatomical markers (53 characters) wereused for the analysis.
Limitations of RAPD markers to estimate genetic dis-tances have been reported by several authors (Castagna et al.
1997; Thorman et al. 1994; Bachmann 1994). In fact, simi-larity coefficients could be overestimated when analyzingdifferent species by RAPDs, because bands with similar mo-bility may represent different DNA sequences. Therefore,nonhomologous amplification products may be erroneouslyconsidered identical. Since in our analysis the number of co-migrating bands represents only 30% of the 125 scored loci,we expect that the “overestimation effect” of similaritywould not have significant impact on the species considered.Furthermore, the possibility that markers of different sizemay contain homologous sequences, due to the presence ofnested inverted repeats of the primer sequence, has been re-ported by Schierwater (1995). This author demonstrated thathigh concentrations of DNA template and high primer bind-ing probabilities avoid the formation of multiple amplifi-cation products of the same locus. In our experimentalconditions, the presence of additional amplification productsthat could determine false polymorphism is unlikely.
Results reported here for relationships observed amongthese species disagree with data reported by Harrison et al.(1997a) who found thatPotentilla fruticosais more closelyrelated toFragaria species thanD. indica. The disagreementcan be explained by the fact that in their analysis they usedonly one accession ofPotentilla andDuchesneain compari-
© 2000 NRC Canada
552 Can. J. Bot. Vol. 78, 2000
Fig. 1. SEM photomicrographs of the epidermis and trichomes of the three wild species studied. (a) Adaxial epidermis and trichomesof D. indica. (b) Adaxial epidermis and trichomes ofP. tucumanensis. (c) Glandular hair with pluricellular head and superficialstomata (ES) ofD. indica. (d) Glandular hair with unicellular head and sunken stomata (ES) ofF. vesca. Scale bars = 100µm forFigs. 1a and 1b. Scale bars = 10µm for Figs. 1c and 1d).
J:\cjb\cjb78\cjb-04\B00-026.vpWednesday, May 17, 2000 9:43:21 AM
Color profile: Generic CMYK printer profileComposite Default screen

son with 26 accessions ofFragaria and also because theyused chloroplastic DNA which shows only maternally inher-ited mutations.
The data presented in this paper show differences amongP. tucumanensis, F. vesca, and D. indica, as revealed bythe examination of morphological and anatomical characters,
© 2000 NRC Canada
Ontivero et al. 553
Fig. 2. Foliar morphology and leaf cross-sections of the three species studied. (a and b) Leaf of P. tucumanensisshowingbrochidodromous venation and very branched secondary venation (a) and prismatic crystals (CR), prominent stomata (ES), and a singlemesophyl palisade (MS) (b). (c and d) Leaf of D. indica showing semicraspedodromous venation and scarcely branched secondary ve-nation (c) and druse crystals in special cells (CR), superficial stomata (ES), and a single mesophyl palisade (MS) (d). (e and f) Leaf ofF. vescashowing semicraspedodromous venation with no branched secondary venation (e) and druse crystals in special cells (CR),clustered sandy crystals (CC), hidden stomata (ES), and triple palisade mesophyl (MS) (f). Scale bar = 2.5 mm for Fig. 2a; Scalebars = 5 mm for Figs. 2c and 2e. Scale bars = 31.6µm for Figs. 2b, 2d, and 2f.
J:\cjb\cjb78\cjb-04\B00-026.vpWednesday, May 17, 2000 9:43:24 AM
Color profile: Generic CMYK printer profileComposite Default screen

and confirmed by analysis of DNA polymorphism. Theseresults extend the current information about the identity ofthese species and provide useful data to recognize wildstrawberries and related species of northwestern Argentina.
Furthermore, some morphological and anatomical charactersincluded in this work, such as runner type and color, leafvenation, type of trichomes, stomata position, crystal inclu-sions, and number of palisade layers, have not been de-
© 2000 NRC Canada
554 Can. J. Bot. Vol. 78, 2000
Fig. 3. Phenograms generated with morphological and anatomical characters (a) and RAPD characters (b). D, D. indica; P,P. tucumanensis; Fv, F. vesca; Fa, F. × ananassa.
Fig. 4. RAPD amplification profiles of different genotypes ofF. × ananassa(a), P. tucumanensis(b), and D. indica using the primerOPJ08 (c). Arrows indicate faint polymorphic bands observed within each species. M is lambdaEcoRI–HindIII DNA digests.
J:\cjb\cjb78\cjb-04\B00-026.vpWednesday, May 17, 2000 9:43:27 AM
Color profile: Generic CMYK printer profileComposite Default screen
scribed previously for these species, and their utilization asdiagnostic traits will help the identification of these species,in both vegetative and reproductive stages (Metcalfe andChalk 1950; Metcalfe 1963; Leth 1991). In addition, theRAPD technique allowed us to recognize species-specificpolymorphic bands obtained with different primers (Fig. 6).These bands may serve as specific markers to evaluate ge-netic introgression between species in future breeding pro-grams, provided crossing barriers can be overcome.
Acknowledgements
Authors thank Daniel Kirschbaum for his excellent techni-cal assistance and critical review of the manuscript. M.O.
also works in the Departamento Biomédico - Biología of theFacultad de Medicina (UNT). J.D.R., and A.C. are membersof Consejo Nacional de Investigaciones Cientificas y Téc-nicas (CONICET). This work was partially supported byAgencia Nacional de Promoción Científica y Tecnológica,Consejo de Investigaciones de la Universidad Nacional deTucumán and CONICET.
References
Arias, M., Albornoz, P., Diaz Ricci, J.C., Castagnaro, A.P., andKirschbaum, D.S. 1998. Collection and characterization of na-tive and naturalized germplasm ofFragaria and related generaPotentilla and Duchesneain Argentina for its introduction to
© 2000 NRC Canada
Ontivero et al. 555
Fig. 5. RAPD amplification profiles ofD. indica (D), P. tucumanensis(P), F. vesca(Fv), andF. × ananassa(Fa) obtained with prim-ers OPJ11 (a), OPJ20 (b), and OPJ16 (c). Templates for PCR amplifications were DNA mixtures of different accessions of studied spe-cies. M1 and M2 are PCR and lambdaEcoRI–HindIII DNA digests, respectively.
Fig. 6. RAPD species-specific markers ofF. × ananassa(Fa), F. vesca(Fv), P. tucumanensis(P), andD. indica (D). The arrows indi-cate the markers detected between compared species with primers OPJ16 (a, c, and e), OPJ09 (b), OPJ20 (d), and OPJ10 (f). M islambdaEcoRI–HindIII DNA digests.
J:\cjb\cjb78\cjb-04\B00-026.vpWednesday, May 17, 2000 9:43:35 AM
Color profile: Generic CMYK printer profileComposite Default screen

© 2000 NRC Canada
556 Can. J. Bot. Vol. 78, 2000
strawberry breeding programs.In Proceedings of the NaturalResources Forum 98, June 9 and 10, Gainesville, Fla. Center forNatural Resources. Institute of Food and Agriculture, Universityof Florida, Gainesville, Fla.
Bachmann, K. 1994. Molecular markers in plant ecology. NewPhytol. 126: 403–418.
Castagna, R., Gnocchi, S., Perenzin, S., and Heun, M. 1997. Ge-netic variability of the wild diploid wheatTriticum urartu re-vealed by RFLP and RAPD markres. Theor. Appl. Genet.94:424–430.
Castagnaro, A., Diaz Ricci, J., Arias, M., and Albornoz, P. 1998.A new shouthern hemisphere species ofPotentilla (Rosaceae).Novon, 8: 333–336.
Crisci, J.V., and López-Armengol, M.F. 1983. Introducción a lateoría y práctica de la taxonomía numérica. The General secre-tariat of the Organization of American States, Washington, D.C.
Darow, G.M. 1966. The strawberry. The New England Institute forMedical Research, N.Y. pp.45–65.
Davidson, C.G. 1995.Potentilla fruticosa, taxonomy and interna-tional registration. Acta Hortic.413: 157–160.
Demeke, T., Adams, R.P., and Chibbar, R. 1992. Potential taxo-nomic use of random amplified polymorphic DNA (RAPD): acase study in Brassica. Theor. Appl. Genet.84: 990–994.
Duchesne, A.N. 1766. Histoire naturelle des Fraisiers. Paris.Ellis, J.R. 1961.Fragaria–Potentillaintergeneric hybridization and
evolution in Fragaria. Symposium of Experimental Taxonomy.Proc. Linn. Soc. Lond.173: 99–106.
Focke, W.O. 1888. Rosaceae.In Engler & Prantl. Die NatüralischePflanzenfamilie III,3: 1–61.
Hancock, J.F., and Callow, P.A. 1994. Randomly amplified poly-morphic DNAs in the cultivated strawberry,Fragaria × anan-assa. J. Am. Soc. Hortic. Sci.119: 862–864.
Hancock, J.F., and Luby, J.J. 1993. Genetic resources at our door-step: the wild strawberries. BioScience,43: 141–147.
Hara, H., and Kurosawa, S. 1959. On theDuschesnea indicagroup.J. Jpn. Bot.34: 161–166.
Harrison, R.E. 1994. Genetic relationships and geographical differ-entiation of native strawberries (Fragaria spp.). Ph.D. thesis,University of Minnesota, St. Paul, Min.
Harrison, R.E., Luby, J.J., and Furnier, G. 1993. Molecular investi-gation of the chloroplast genome inFragaria spp. Acta Hortic.348: 395–402.
Harrison, R.E., Luby, J.J., and Furnier, G.R. 1997a. ChloroplastDNA restriction fragment variation among strawberry (Fragariaspp.) taxa. J. Am. Soc. Hortic. Sci.122: 63–68.
Harrison, R.E., Luby, J.J., Furnier, G., and Hancock, J.F. 1997b.Morphological and molecular variation among populations ofoctoploidFragaria virginiana andF. chiloensis(Rosaceae) fromNorth America. Am. J. Bot.84: 612–620.
Jackson, D.B. 1895. Index Kewensis. An enumeration of the gen-era and species of flowers and plants. Vol I. Clarendon Press,Oxford. p. 974.
Kalkman, C. 1968.Potentilla, DuchesneaandFragaria in Malesia(Rosaceae). Blumea,16: 325–354.
Lamboy, W.F. 1994. Computing genetic similarity coefficientsfrom RAPD data: the effects of PCR artifacts. PCR methods andapplications. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, N.Y. pp. 31–37.
Leth, M. 1991. Micromorphological characters in the systematicsof Baltic cinquefoils (Potentilla). Botaanalised uurimused.VI(Scripta Botanica). Tallinn. pp. 150–164.
Metcalfe, C.R. 1963. Comparative anatomy as a modern botanicaldiscipline. Adv. Bot. Res. (London),1: 101–147.
Metcalfe, C.R., and Chalk, L. 1950.In Anatomy of the Dicotyle-dons. Vols. I–II. Clarendon Press, Oxford.
Rogers, S.O., and Bendich, A.J. 1988. Plant molecular manual A6.Kluwer Academic Publishers, Dordrecht, Belgium. pp. 1–10.
Rohlf, F.J. 1993. NTSYS-PC. Numerical taxonomy and multivar-iate analysis for the IBM PC and compatible. Version 1.8. Ap-plied biostatistics Inc., Exeter Software, New York.
Sambrook, J., Fritsch, E.F., and Maniatis, T. 1989. Molecular Clon-ing. A laboratory manual. Cold Spring Harbor Laboratory Press,Cold Spring Harbor, N.Y.
Schierwater, B. 1995. Arbitrarily amplified DNA in systematicsand phylogenetics. Electrophoresis,16: 1643–1647.
Schulze-Menz, G.K. 1964. Rosales.In A. Engler’s Syllabus derPflanzenfamilien. 12th ed. f.88. Gebr. Borntraeger, Berlin.pp. 213–216.
Staudt, G. 1962. Taxonomic studies in the genusFragaria. Typi-fication of Fragaria species known at the time of Lineaeus. Can.J. Bot. 40: 869–886.
Strittmather, D. 1986. Manual de técnicas en histología vegetal.Edited by Ana D’Ambrogio de Argüeso. Hemisferio Sur. 1°Edición, Chap. II, pp. 52. Editorial Hemisferio Sur. Bs. As., Ar-gentina.
Thorman, C.E., Ferreira, M.E., Camargo, L.E.A., Tivang, J.G., andOsborn, T.C. 1994. Comparison of RFLP and RAPD markersfor estimating genetic relationships within and among cruci-ferous species. Theor. Appl. Genet.88: 973–980.
Transue, D.K., Fairbanks, D.J., Roeson, L.R., and Andersen, W.R.1994. Species identification by RAPD analysis of grain ama-ranth genetic resources. Crop Sci.34: 1385–1389.
van Tongeren, O.F.R. 1995. Cluster analysis. Data analysis in com-munity and landscape ecology. Cambridge University Press,New York. pp. 175–203.
Wolf, T. 1908. Monographie der GattungPotentilla. Bibl. Bot.71: 664.
Yu, L.X., and Nguyen, H.T. 1994. Genetic variation detected withRAPD markers among upland and lowland rice cultivars (Oryzasativa L.). Theor. Appl. Genet.87: 668–672.
Zardini, E.M. 1984.In Los géneros de las Fanerógamas de Ar-gentina.Edited byA.T. Hunzinker. Bol. Soc. Argent. Bot.23:218–220.
Zardini, E.M. 1999. Catálogo de las plantas vasculares en la Repú-blica Argentina.In Rosaceae.Edited by F.O. Zuloaga and O.Monrrone. Vol 2. Missouri Botanical Garden Press, St. Louis,Mo. pp. 981–992.
J:\cjb\cjb78\cjb-04\B00-026.vpWednesday, May 17, 2000 9:43:35 AM
Color profile: Generic CMYK printer profileComposite Default screen




















