
Chrysochromulina fragaria sp. nov. (Prymnesiophyceae), a new haptophyte flagellate from Norwegian...
7
Phycologia (1999) Volume 38 (2),149-155 Published 16 July 1999 Chrysochromulina fragarin sp. noy. (Prymnesiophyceae), a new haptophyte flagellate from Norwegian waters Wsrucne Exnerra eNo BBNreEpvenoseN Sectionfor Marine Botany, Department of Biology, IJniversityof Oslo, P.O. Box 1069 Blindern,0316 Oslo, Norway W. ETKREM aNo B. Eovaeosstt. 1999. Chrysochromulina fragaria sp. nov. (Prymnesiophyceae), a new haptophyte flagellate from Norwegian waters. Phycologia 38: 149-155. Chrysochromulina fragaria sp. nov. was one of the dominant species during the 1994 and 1995 Chrysochromulina bloorns off the southem coast of Norway. It was isolated by the serial dilution culture method from a surface water sample collected during the bloom in May-June 1994. The cells are spherical, 4 to 8 pm in diametet and possess a coiling haptonema that is shorter than the two equal to subequal flagella. Each cell contains two chloroplasts, a nucleus, and one mitochondrion, which appears to be reticulate. The periplast is covered by monomorphic scales (scale faces with identical pattem) of two types arr:anged in layers. The scale faces have a pattern of radiating ribs arranged in quadrants. One type has inflexed rims, whereas the other type has erect rims. The fust intemal transcribed spacer (ITS1) rDNA sequence and scale morphology of C. fragaria are compared with that of Chrysochromulina sp- CCMP 1204. Despite the great similarities in scale morphology, their ITS1 rDNA regions are very different, indicating that they are different species- Both C- frugaia and Chrysochromulina sp. CCMP 1204 are nontoxic to A rte m i a frctn ci scana naur,lti. INTRODUCTION The number of known species in the genus Chrysochromulina Lackey has increased steadily since the description of the first marine species by Parke et al. (1955,1956), and it is now one of the marine phytoplanktonic genera with the greatest num- ber of species. More than 50 species have been described, and 38 of them have been observed in Scandinavian waters. In addition, from Scandinavian waters alone we know of 30 forms not described in the literature (Eikrem et al. 1998; Ien- sen 1998) and the number of Chrysochromulina species worldwide is expected to exceed 100 (Thomsen et al. 1994). Members of the genus Chrysochromulina are unicellular, nanoplanktonic, photosynthetic flagellates. Most species are marine; only a few occur in freshwater (Green & Jordan 1994). They possess two smooth flagella and a haptonema that is readily recognized with the light microscope. The length of the haptonema varies greatly from many times the cell di- ameter in some species of Chrysochromulina [e.9. c. 160 pm long in C. camella Leadbeater et Manton (Iradbeater & Man- ton 1969)l to less than the cell length in others [e.g. c. 5 pm in Chrysochromulina spinifera (Fournier) Pienaar et Norris (Pienaar & Norris 1979)1. Chrysochromulina cells are covered by one to several kinds of scales arranged in layers. The scales are organic and com- posed mainly of proteins and carbohydrates (Leadbeater 1994). The basic scale is a round or oval plate composed of microfibrils, but the variation is great, and many highly sculp- tured and elaborate scale forms are found. The scales of many species consist of a proximal layer of microfibrils arranged in a radial pattern, whereas the pattern of the distal microfibrillar layer is more variable. Scales may have monomorphic faces (the two scale faces having identical patterns) or dimorphic faces (the scale faces having different patterns). The cells con- tain two golden-brown chloroplasts, each with a pyrenoid that may be bulging. The nucleus is posterior or central and the outer membrane of the nuclear envelope is confluent with the endoplasmic reticulum enveloping the chloroplast (Hibberd 1980). Located immediately within the cell membrane is a layer of peripheral endoplasmic reticulum that also extends into the haptonema (Pienaar 1994). The mitochondrion prob- ably forms a network, as was shown for the coccolithophorid Pleurochrysis carterae (Braarud et Fagerland) Christensen (Beech & Wetherbee 1984). The flagellar apparatus consists of two basal bodies, the haptonematal base, microtubular and fibrous roots, and accessory and connecting fibers. The mi- crotubular root (R,), which is connected to the mature flagel- lum, may be either simple (a sheet of microtubules) or com- pound (a sheet and a crystalline bundle), and its structure varies between the species within the genus as presently de- limited. The microtubular root R, has an origin between the two basal bodies, is simple, and consists of just a few micro- tubules in the species examined so far. The roots associated with the right flagellar base contain only a few microtubules (Green & Hori 1994). With few exceptions, identification of Chrysochromulina species relies on the examination by electron microscopy of the organic scales covering the periplast (Leadbeater 1994). We present some morphological and ulffastructural details of C. fragaria sp. nov., in addition to its sequence of the flrst internal transcribed spacer (ITSI) rDNA, and compare it with that of Chrysochromulina sp. CCMP 1204, which shows a scale morphology reminiscent of that of C. fragaria sp. nov. MATERIAL AND METHODS Cultures Cultures of Chrysochromulina fragaria were established from cells isolated by the serial dilution culture method (Throndsen l97R) during a bloom dominated by species of Chrysochrom- A 283 149
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Chrysochromulina fragaria sp. nov. (Prymnesiophyceae), a new haptophyte flagellate from Norwegian...

Phycologia (1999) Volume 38 (2),149-155 Published 16 July 1999
Chrysochromulina fragarin sp. noy. (Prymnesiophyceae), a new haptophyteflagellate from Norwegian waters
I Wsrucne Exnerra eNo BBNre EpvenoseN
1 Sectionfor Marine Botany, Department of Biology, IJniversity of Oslo, P.O. Box 1069 Blindern,0316 Oslo, NorwayI
W. ETKREM aNo B. Eovaeosstt. 1999. Chrysochromulina fragaria sp. nov. (Prymnesiophyceae), a new haptophyte flagellate
from Norwegian waters. Phycologia 38: 149-155.
Chrysochromulina fragaria sp. nov. was one of the dominant species during the 1994 and 1995 Chrysochromulina bloorns offthe southem coast of Norway. It was isolated by the serial dilution culture method from a surface water sample collected duringthe bloom in May-June 1994. The cells are spherical, 4 to 8 pm in diametet and possess a coiling haptonema that is shorter thanthe two equal to subequal flagella. Each cell contains two chloroplasts, a nucleus, and one mitochondrion, which appears to bereticulate. The periplast is covered by monomorphic scales (scale faces with identical pattem) of two types arr:anged in layers.The scale faces have a pattern of radiating ribs arranged in quadrants. One type has inflexed rims, whereas the other type haserect rims. The fust intemal transcribed spacer (ITS1) rDNA sequence and scale morphology of C. fragaria are compared withthat of Chrysochromulina sp- CCMP 1204. Despite the great similarities in scale morphology, their ITS1 rDNA regions are verydifferent, indicating that they are different species- Both C- frugaia and Chrysochromulina sp. CCMP 1204 are nontoxic toA rte m i a frctn c i s c a na naur,lti.
INTRODUCTION
The number of known species in the genus ChrysochromulinaLackey has increased steadily since the description of the firstmarine species by Parke et al. (1955,1956), and it is now oneof the marine phytoplanktonic genera with the greatest num-ber of species. More than 50 species have been described, and38 of them have been observed in Scandinavian waters. Inaddition, from Scandinavian waters alone we know of 30forms not described in the literature (Eikrem et al. 1998; Ien-sen 1998) and the number of Chrysochromulina speciesworldwide is expected to exceed 100 (Thomsen et al. 1994).
Members of the genus Chrysochromulina are unicellular,nanoplanktonic, photosynthetic flagellates. Most species aremarine; only a few occur in freshwater (Green & Jordan1994). They possess two smooth flagella and a haptonema thatis readily recognized with the light microscope. The length ofthe haptonema varies greatly from many times the cell di-ameter in some species of Chrysochromulina [e.9. c. 160 pm
long in C. camella Leadbeater et Manton (Iradbeater & Man-ton 1969)l to less than the cell length in others [e.g. c. 5 pmin Chrysochromulina spinifera (Fournier) Pienaar et Norris(Pienaar & Norris 1979)1.
Chrysochromulina cells are covered by one to several kindsof scales arranged in layers. The scales are organic and com-posed mainly of proteins and carbohydrates (Leadbeater
1994). The basic scale is a round or oval plate composed ofmicrofibrils, but the variation is great, and many highly sculp-tured and elaborate scale forms are found. The scales of manyspecies consist of a proximal layer of microfibrils arranged ina radial pattern, whereas the pattern of the distal microfibrillarlayer is more variable. Scales may have monomorphic faces(the two scale faces having identical patterns) or dimorphicfaces (the scale faces having different patterns). The cells con-tain two golden-brown chloroplasts, each with a pyrenoid thatmay be bulging. The nucleus is posterior or central and the
outer membrane of the nuclear envelope is confluent with theendoplasmic reticulum enveloping the chloroplast (Hibberd
1980). Located immediately within the cell membrane is alayer of peripheral endoplasmic reticulum that also extendsinto the haptonema (Pienaar 1994). The mitochondrion prob-ably forms a network, as was shown for the coccolithophoridPleurochrysis carterae (Braarud et Fagerland) Christensen(Beech & Wetherbee 1984). The flagellar apparatus consistsof two basal bodies, the haptonematal base, microtubular andfibrous roots, and accessory and connecting fibers. The mi-crotubular root (R,), which is connected to the mature flagel-lum, may be either simple (a sheet of microtubules) or com-pound (a sheet and a crystalline bundle), and its structurevaries between the species within the genus as presently de-limited. The microtubular root R, has an origin between thetwo basal bodies, is simple, and consists of just a few micro-tubules in the species examined so far. The roots associatedwith the right flagellar base contain only a few microtubules(Green & Hori 1994).
With few exceptions, identification of Chrysochromulinaspecies relies on the examination by electron microscopy ofthe organic scales covering the periplast (Leadbeater 1994).We present some morphological and ulffastructural details ofC. fragaria sp. nov., in addition to its sequence of the flrstinternal transcribed spacer (ITSI) rDNA, and compare it withthat of Chrysochromulina sp. CCMP 1204, which shows ascale morphology reminiscent of that of C. fragaria sp. nov.
MATERIAL AND METHODS
Cultures
Cultures of Chrysochromulina fragaria were established fromcells isolated by the serial dilution culture method (Throndsen
l97R) during a bloom dominated by species of Chrysochrom-
A 283149

150 Phycologia, Vol. 38 (2), 1999
ulina off the southern coast of Norway in May-June 1994.Strain Sl9 originated from a surface water sample collectedon 20 May 1994 off Flodevigen (58"25'N, 8"46'E), and strainU21 was from a surface water sample collected on 2 June1994 off Grimstad (58'20'N, 8'36'E).
Chrysochromulina sp. strain CCMP 12M, obtained fromthe Provasoli-Guillard National Center for Culture of MarinePhytoplankton (CCMP), was isolated by R. Selvin from a wa-ter sample collected at Baffin Bay, Canada (76"25'N,
82'55'W) on 3 June 1986.Stock cultures of C. fragaria were grown in filtered sea-
water diluted to 30 psu, enriched with nutrients, vitamins, andtrace metals as in IMR l/2 medium (Eppley et al. 1967), andsupplemented with 1O nM selenite. The cultures were grownat 15"C under white fluorescent light with a photon fluencerate (PFR) of about 50 pmol photons m-2 s-r and a l2:l2hLD cycle. Stock cultures of strain CCMP l2O4 were grownin K-medium (Keller et al. 1987) at 3oC under similar lightconditions as above. Stock cultures were transferred to newmedium every 14 d.
Light microscopy
Living cells of the strains S19 and CCMP l2O4 were studiedwith a Nikon Microphot FX fitted with phase contrast anddifferential interference contrast optics and electronic flash.Strain S19 was photographed using phase contrast and flash.
Electron microscopy
Positively stained whole mounts of strains Sl9 and CCMPl2O4 were prepared according to Moestrup (1984). Some ofthe preparations were shadowed with gold-palladium using anEdward's Speedivac 12 E6 coating unit, c. 30" angle of shad-owing.
Thin sections of Sl9 were prepared according to the fol-lowing protocol. The cells were fixed in 2Vo glutaraldehyde inmedium for 2 h and rinsed 3 times at 30-min intervals inmedium followed by 2 times at l0-min intervals in 0.1 M Nacacodylate buffer (pH 8). Postfixation was carried out in l%oosmium tetroxide and lvo ferricyanide in 0.1 M Na cacodylatebuffer for 3 h. The cells were then rinsed 3 times at lS-minintervals in Na cacodylate buffer and 2 times at lo-min in-tervals in distilled water. The samples were left overnight in2Vo aqteous uranyl acetate. Subsequently the cells were rinsedin distilled water and dehydrated in an ethanol series startingat 30Vo and gradually rising to 96Vo. The dehydration wasconcluded with changes in IOOVo ethanol 4 times at l0-minintervals and changes in propylene oxide 2 times at lo-minintervals. The pellets were left overnight in a l:1 mixture ofpropylene oxide and Epon embedding resin (Burke & Giesel-man l97l). Finally, the cells were changed in Epon 3 timesat l-h intervals before they were polymerized at 50'C for 12h. The thin sections were stained with lead citrate. Thin sec-tions and whole mounts were viewed in a Jeol 120oex trans-mission electron microscope.
Artemia bioassay
Toxicity tests were performed using a strain of the crustaceanArtemia franciscana (Creasel, Deinze, Belgium) as previouslydescribed (Edvardsen 1993). Sixty nauplii were exposed to C.
fragaria strain U21 (9 X 108 cells I r) and Chrysochromulinasp. strain CCMP I2O4 (3 x lO8 cells I r), and 30 nauplii wereexposed to C. fragaria strain Sl9 for 24 h and 48 h at 25'Cin darkness. Seawater was used as control.
DNA extraction, amplification, and sequencing
DNA was extracted using a 3Vo CTAB (hexadecyltrimethylam-
monium bromide) procedure (Doyle & Doyle 1990). Biotiny-lated and nonbiotinylated primers ITSI and ITS2 (White er a/.1990) were used to amplify the internal transcribed spacer I(ITSl) region. Polymerase chain reaction (PCR) amplificationswere performed in a Techne Genius using the following am-plification profile: 95'C for 5 min, 30 cycles at 50'C for I min,72"C for I min and 95'C for I min, and a final extension at72"C for 7 min. The PCR reaction mixtures (50 p,l) containedI U Taq DNA polymerase (Advanced Biotechnologies, UK),50 ng DNA, 0.5 pM of each primer, 200 pM dNTR 2.5 mMMgClr, 5 pl lOX reaction buffer, and 5Vo acetamide. Single-stranded DNA, obtained from the PCR products by using M-280 streptavidine-coated magnetic Dynabeads (Dynal AS, Nor-way), were sequenced directly with the use of the Sequenaceversion 2.0 DNA Sequencing Kit (United States Biochemical)according to the manufacturer's protocol and were run manuallyon 6Eo acrylanide gels (Sambrook et al. 1989). DNA sequenc-es have been submitted to GenBank: C. fragaria Sl9,AJ2387O8, Chrysochromulina sp. CCMP 1204, AJ2387O9.
The alignment was performed automatically using ClustalW in the GCG package. The distance value was estimated byKimura two-parameter analysis (Kimura l98O) using PHYLIP(Felsenstein 1995).
OBSERVATIONS
Description of the new species
Chrysochromulina fragaria Eikrem et Edvardsen sp. nov.
Figs l-16
DIAGNosIS: Cellulae sphaericae-subsphaericae 4-8 pm, appendicesin tenuem depressionem apicaliter insertae. Haptonema (3-9 p"m)flagellis plerumque aequalibus (10-16 u,m) brevius. Periplastussquamis duarum formarum tectus. Squamae aut marginibus inflexis(50-100 nm) ovales vel orbiculares 0.35-0.75 x 0.45-0.8 lrm, autmarginibus erectis (150-200 nm) orbicularis vel ovales 0.5-0.8 x0.5-0.8 pm. Facies proximales et distales utriusque formae squa-marum striis radiantibus in quattuor quadrantes dispositis instructae.
Cells spherical to subspherical measuring 4-8 pm, append-ages inserted apically in a slight depression. Haptonema (3-9
pm) shorter than the usually equal flagella (10-16 pm). Peri-plast covered with scales of two types. Scales with inflexedrims (50-100 nm) oval to round 0.35-{.75 by 0.45-{.8 pm
and scales with erect rims (150-2OO nm) round to oval 0.5-0.8 by 0.5{).8 pm. Proximal and distal faces of both scaletypes with radiating ribs arranged in four quadrants.
HoLoTYPE: Figures l-9.ETyMoLocy: Latin fragaria (strawberry). The organism
gives the impression of the shape of a squat strawberry in thelight microscope.

Eikrem & Edvardsen'A new species of Chrysochromulina t 5 l
Figs 1-7. Chrysochromulina fragaria sp. nov.Figs 1, 2. Light micrographs (phase contrast) of living cells.Figs 3-5. Shadow-cast whole mounts of scales. Distal scale faces (arrows), proximal scale faces (arrowheads). Outer layer scales (white arrow,white arrowhead), inner layer scales (black arrows, black arrowheads).Figs 6, 7. Thin sections of scales.
Fig. 6. Cross section showing scales with inflexed rims (arrow) and scales with upright rims (arrowhead).Fig. 7. Tangential section of scales.
Microscopic observations of Chrysochromulina fragariasp. nov. strain S19
The spherical to subspherical cells (4-8 pm) bear a hapto-nema (3-8 pm) that is shorter than the two equal or subequalflagella (10-14 pm) and is capable ofcoiling. The appendagesare inserted in a slight apical depression (Figs l,2). Two typesof scales cover the periplast. We have not been able to producesections of cells with intact scale investments. but we assume
that the scales are ilTanged in two layers, with the scales hav-ing the tall, erect rims forming the outer layer. We also assumethat the rims are on the distal face of the scales, which isusually the case in Chrysochromulina species (Leadbeater1994). The inner layer consists of round to oval plate scales(0.35--0.75 by 0.45-{.8 pm) with an inflexed rim (50-100nm) with concentric fibers. Both scale faces have radiatingribs (c. 65-90) extending from center to edge (proximal face)or rim (distal face). The round scales of the outer layer (0.5-
r52 Phycologia, Vol. 38 (2), 1999
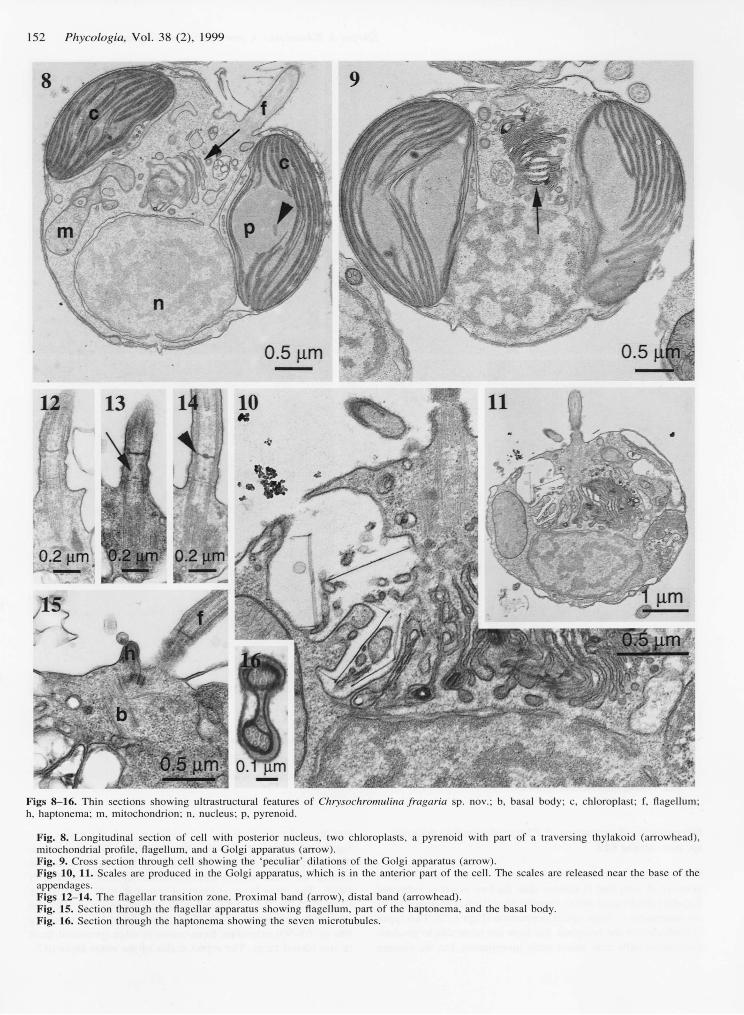
Figs 8-16. Thin sections showing ultrastructural features of Chrysochromulina fragaria sp. nov.; b, basal body; c, chloroplast; f, flagellum;h, haptonema; m, mitochondrion; n, nucleus; p, pyrenoid.
Fig. 8. Longitudinal section of cell with posterior nucleus, two chloroplasts, a pyrenoid with part of a traversing thylakoid (arrowhead),mitochondrial profile, flagellum, and a Golgi apparatus (arrow).Fig. 9. Cross section through cell showing the 'peculiar' dilations of the Golgi apparatus (arrow).Figs 10, 11. Scales are produced in the Golgi apparatus, which is in the anterior part of the cell. The scales are released near the base of theappendages.Figs 12-14. The flagellar transition zone. Proximal band (arrow), distal band (arrowhead).Fig. 15. Section through the flagellar apparatus showing flagellum, part of the haptonema, and the basal body.Fig. 16. Section through the haptonema showing the seven microtubules.
$

Figs 17, 18. Whole mounts of Chrysochromulina sp. CCMP 12O4.
[1t: tt. Whole cell with detached scales. Stained with uranyl ace-
Fig. 18. Details of scales. Shadow-cast preparation showing the twoscale types (arrow, arrowhead).
0.8 by 0.5-{.8 pm) have tall erect rims (150-200 nm) withno obvious pattern, and the radiating ribs (c. 75-90) extendfrom the center to the rim on the distal face and from centerto edge on the proximal face (Figs 3-7). The pattern createdby the radiating ribs divides the scales into four distinct quad-rants (Fig. 7) in both scale types.
The cells contain two golden-brown, parietal chloroplasts,each with an immersed pyrenoid that may be bulging and maybe traversed by thylakoids, a posterior nucleus, a mitochon-drion, and a Golgi body where the scales are produced (Figs
8-l I ) .The flagellar apparatus (Fig. 15) has not been studied in
detail, but some features have been revealed. The flagella havea proximal band consisting of a tripartite plate and a distalband with a transitional plate (Figs 12-14). The extended partof the haptonema contains seven microtubules (Fig. 16).
To allow a comparison between the scales of C. fragariasp. nov. and those of Chrysochromulina sp. CCMP 1204, wehave included electron micrographs (Figs 17, 18) of a wholecell and detached scales. Under the light microscope the cellsappear spherical to pear shaped (8-14 by G-10 pm). The cellsmay be strongly compressed dorsoventrally. The haptonema(12-20 pm) is slightly shorter than the flagella (14-24 p"m).
The scales are of two types: one type has an erect rim (85-
100 nm) and measures 0.'74.9 by 0.8-1.0 pm; the rim of theother type appears inflexed (35-50 nm) and it measures 0.5-0.8 by 0.6-0.9 pm. All scale faces have radiating ribs (c. 75-8s).
Artemia bioassay
Strains of C. fragaria (S19 and U21) and Chrysochromulinasp. CCMP l2O4 that were grown in complete medium were
Eikrem & Edvardsen' A new species of Chrysochromulina 153
nontoxic to Artemia nauplii (all 60/30 nauplii survived in eachtest) when exposed for 24 and 48 h under the conditions used.
ITSI nucleotide sequences
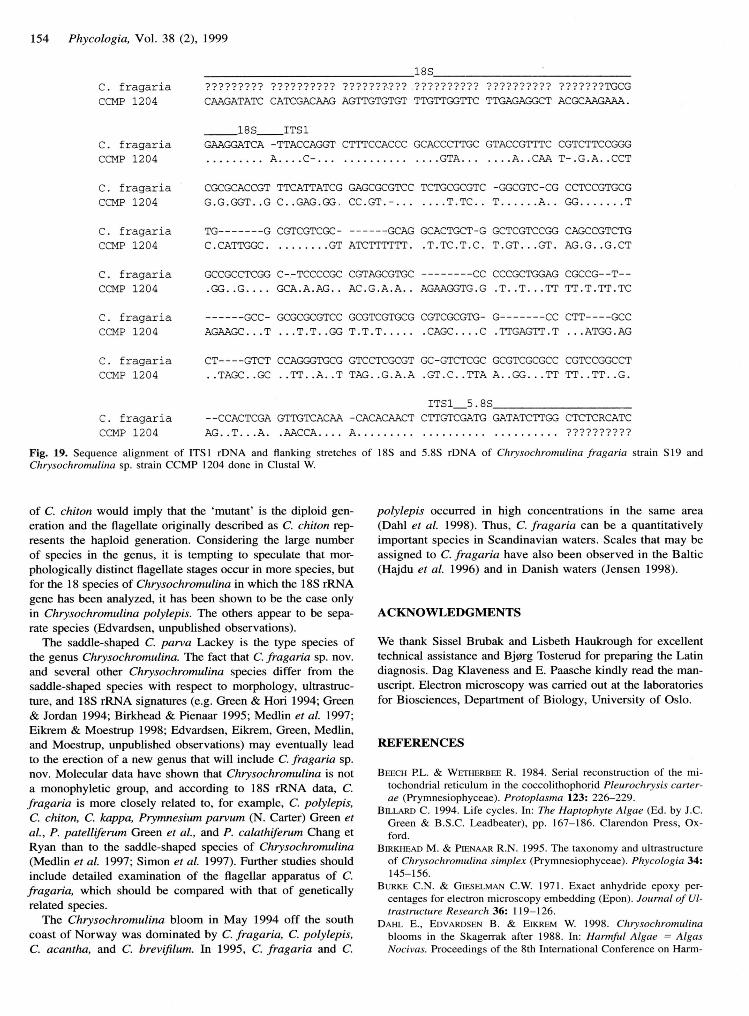
Nucleotide sequences of ITS I rDNA and flanking stretches ofl8S and 5.8S rDNA regions were determined for C. fragariastrain S19 and Chrysochromulina sp. strain CCMP l2O4 (Fig.l9). Terminals of the l8S rRNA coding region and startingpositions of the 5.8S region were determined by comparisonwith other haptophytes (Larsen & Medlin 1997; Edvardsen &Medlin 1998). The length of the ITSI region was 326 basepairs (bp) in C. fragaria and 381 bp in Chrysochromulina sp.strain CCMP l2O4.The G/C content of ITS1 was much higherin C. fragaria (1l%o) than in CCMP l2O4 (58Vo). Pairwisecomparisons, including gaps of strain Sl9 and CCMP l2M,revealed a distance value of approximately O.6l (Kimura two-parameter) for the ITSI region.
DISCUSSION
Of the species of Chrysochromulina already described, thereare several that resemble C. fragaria when examined by lightmicroscopy. For instance, C. minor Parke et Manton, C. kappaParke et Manton, C. brevffilum Parke et Manton, and C. ad-riatica Leadbeater have the same general appearance as C.
fragaria and are in the same size range. Also, species that aretypically oblong or pear shaped may produce cells that areround (e.g. C. polylepis Parke et Manton) and may then beconfused with C. fragaria at the light microscope level.
Despite the similarities in cell and scale morphology, ITS Idata show that Chrysochromulina sp. CCMP 1204 should beconsidered a separate species from C. fragaria. The scales ofC. fragaria are in the same size range as the scales of Chrys-ochromulina sp. CCMP l2M and the number of radiating ribsoverlap. The height of the upright rims and the width of theinflexed rims in C. fragaria are greater than the correspondingrims in Chrysochromulina sp. CCMP 1204. Length of hap-tonema and flagella and cell size are somewhat overlappingin the two species, but Chrysochromulina sp. CCMP 1204tend to be larger with longer appendages. Another distinguish-ing character at the light microscope level is the pronounceddorsoventral compression of many cells in the Chrys-ochromulina sp. CCMP 1204 cultures; this has not been ob-served in cultures of C. fragaria sp. nov.
The scales of both C. fragaria and Chrysochromulina sp.CCMP 1204 closely resemble the scales of what once was be-lieved to be a mutant of C. chiton Parke et Manton (Manton
1966, 1967a, 1967b). In the light of recent research on C. pol-ylepis (Edvardsen & Paasche 1992; Edvardsen & Medlin 1998)one may speculate that C. chiton has two distinct flagellateforms. In the original culture, C. chiton (Parke et al. 1958)possessed two types of scales with dimorphic scale faces. Aftersome time in the history of the culture a flagellate covered withtwo types of scales with monomorphic scale faces turned up(Manton 1966, 1967a, 1967b). According to a hypothesis basedon the morphology of underlayer scales in coccolithophorids,scales having dimorphic scale faces represent the haploid gen-eration and scales with monomorphic scale faces the diploidgeneration (Billard 1994). Applying this hypothesis to the case
154 Phycologia, Vol. 38 (2), 1999
a € r a a a r i a
ccMP 1204
C F r a a a r i a
CCI.,IP 1204
C f r a n a r i a
ccI4P 1204
t ' - f raaari a
ccMP 1204
. € r a a a r i a
ccMP 1204
. F r a a a r i a
ccMP 1204
C f r ^ d ^ r ; ^
ccMP 1204
Lf tUAIAIU LAILUALIuU AGTTGTGTGT' TTGTTGGTTC? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? T @ GI ruaunsr 6vwM.
GTACCGTTTC CGTCTTCCGGG
. . . . 4 . . L l s I - . U . 4 . . L L I
- s u t L - L u L L r u u u l u L g
T . . . . . . A . . G G . . . . . . . 7
GCTCGTCCGG CAGCCGTCTGf t ^ r i 4 i ^ ^ i i t
L8s_rTs1u M t u A - t l A g l n | , s f v r I r L u n u L v g u d u g L r l w
. . . A . . . . C - . . . . . G T A . . .
CCCGCACCGT TTCAT'TATCG GAGCC'CGTCC TCTGCGCGTC
G . G . G G T . . G C . . G A G . G G , C C . G T . - . . . . . . . T . T C . .
TG---- - - -G CGTCGTCGC- -- -_--GCAG GCACTGCT-G
C.CATTC€C. . .GT ATCTTTTTT . .T .TC .T .C .
GCCGCCTCGG C--TCCCCGC CGTAGCGTGC^ ^ ^ r n r ^ a ^ ^ t i
- - - - - - w L - w w \ * g t L u u L g t L u r w u
A G A A C C . . . T . . . T . T . . C ' G T . T . T . . . . .
. . 1 6 w . . 9 u . . f l . . n . . t r n u . . 9 . n . 6
AGAAGGTG.G .T . .T . . .TT TT.T .TT.TC
l 4 - u t u l u \ & g u u r u u u g u L L 9 r L L \ . & L l
. G T . C . . T T A A . . G G . . . T T T T , . T T . . G .
ITS1-5 .85-C. fragaria --CCACTCGA GTTGTCACAA -CACACAACT CTTGTCGATG GATATCTTC'G CTCTCRCATC
c c ] { P 1 2 0 4 A G . . T . . . A . . A A C C A . . . . A . ? ? ? ? ? ? ? ? ? ?
Fig. 19. Sequence alignment of ITSI rDNA and flanking stretches of 18S and 5.8S rDNA of Chrysochromulina fragaria strain Sl9 andChrysochromulina sp. strain CCMP 1204 done in Clustal W
of C. chiton would imply that the 'mutant' is the diploid gen-eration and the flagellate originally described as C. chiton rcp-resents the haploid generation. Considering the large numberof species in the genus, it is tempting to speculate that mor-phologically distinct flagellate stages occur in more species, butfor the 18 species of Chrysochromulina in which the l8S rRNAgene has been analyzed, it has been shown to be the case onlyin Chrysochromulina polylepis. The others appear to be sepa-rate species (Edvardsen, unpublished observations).
The saddle-shaped C. parva Lackey is the type species ofthe genus Chrysochromulina. TIte fact that C. fragaia sp. nov.and several other Chrysochromulina species differ from thesaddle-shaped species with respect to morphology, ultrastruc-ture, and l8S rRNA signatures (e.g. Green & Hori 1994; Green& Jordan 1994: Birkhead & Pienaar 1995: Medlin et al. 1997:Eikrem & Moestrup 1998; Edvardsen, Eikrem, Green, Medlin,and Moestrup, unpublished observations) may eventually leadto the erection of a new genus that will include C. fragaria sp.nov. Molecular data have shown that Chrysochromulina is nota monophyletic group, and according to l8S rRNA data, C.
fragaria is more closely related to, for example, C. polylepis,
C. chiton, C. kappa, Prymnesium parvum (N. Carter) Green etal., P. patelliferum Green et al., and P. calathiferum Chang etRyan than to the saddle-shaped species of Chrysochromulina(Medlin et al. 1997; Simon et al. 1997). Further studies shouldinclude detailed examination of the flagellar apparatus of C.
fragaria, which should be compared with that of geneticallyrelated species.
The Chrysochromulina bloom in May 1994 off the southcoast of Norway was dominated by C. fragaria, C. polylepis,
C. acantha, and C. brevifilum. In 1995, C. fragaria and C.
polylepis occurred in high concentrations in the same area(Dahl et al. 1998). Thus, C. fragaria can be a quantitativelyimportant species in Scandinavian waters. Scales that may beassigned to C. fragaria have also been observed in the Baltic(Hajdu et al. 1996) and in Danish waters (Jensen 1998).
ACKNOWLEDGMENTS
We thank Sissel Brubak and Lisbeth Haukrough for excellenttechnical assistance and Bj0rg Tosterud for preparing the Latindiagnosis. Dag Klaveness and E. Paasche kindly read the man-uscript. Electron microscopy was carried out at the laboratoriesfor Biosciences, Department of Biology, University of Oslo.
REFERENCES
BEECH PL. & Wrrnrnnrr R. 1984. Serial reconstruction of the mi-tochondrial reticulum in the coccolithophorid Pleurochrysis carter-ae (Prymnesiophyceae). Protoplasma 123: 226-229.
Bu-rann C. 1994. Life cycles. ln: The Haptophyte Algae (Ed. by J.C.Green & B.S.C. Leadbeater), pp. 167-186. Clarendon Press, Ox-ford.
BrnrHe.co M. & PTENAAR R.N. 1995. The taxonomy and ultrastructureof Chrysochromulina s implex (Prymnesiophyceae). P hycologia 342145-156.
BURKE C.N. & GmsrlveN C.W. 1971. Exact anhydride epoxy per-centages for electron microscopy embedding (Epon). Joumal of Ul-trastructure Research 362 l19-126.
DAHL E'., EovenoseN B. & Exneu W. 1998. Chrysochromulinablooms in the Skagerrak after 1988. In: Harmful Algae : AlgasNocivas. Proceedings of the Sth International Conference on Harm-

ful Afgae. Vigo, Spain, 1997 (Ed. by B. Reguera, J. Blanco, M.-L'
Fern6ndez & T. Wyatt), pp. 104-105. International Oceanographic
Commission, UNESCO, Vigo, Spain.Dovlr J.J & DoYLE J.L. 1990. Isolation of plant DNA from fresh
tissue. Focas l2z 13-15.EovnnosrN B. 1993. Toxicity of Chrysochromulina species (Prym-
nesiophyceae) to the brine shrimp, Artemia salina. ln: Toxic Phy-
toplankton Blooms in the Sea (Ed. by T.J. Smayda & Y. Shimizu),pp. 681-686. Elsevier, Amsterdam.
EDVARDsEN B. & Mru-IN L. 1998. Genetic analyses of authentic and
alternate forms of Chrysochromulina polylepis (Haptophyta). P/ty-
cologia 37: 275-283.EDvARDsEN B. & PAAscHE E. 1992. Two motile stages of Chryso-
chromulina polylepis (Prymnesiophyceae): morphology, growth,
and toxicity. Journal of Phycology 282 lO4-114.
ETKREM W.. JENSEN M.@.. Moesrnup @. & TunoNoseN J. 1998. An
illustrated key to the unmineralized prymnesiophyceaen flagellates
of Scandinavian marine waters with special reference to the genus
Chrysochromulina. In: The Genus Chrysochromulina (Prymnesio-
phyceae) in Scandinavian Coastal Waters-Diversity, Abundance
and Ecology, pt. 5 (By M.O. Jensen), pp. l-36. PhD Thesis, Uni-
versity of Copenhagen.EIKREM W. & Mopsrnup Q. 1998. Structural analysis of the flagellar
apparatus and the scaly periplast in Chrysochromulina scutellum sp.
nov. (Prymnesiophyceae, Haptophyta) from the Skagerrak and the
Baltic. Phycologia 372 132-153.EppLEy R.W., HoLMEs R.W. & Srnlcxrexn J.D.H. 1967. Sinking rates
of marine phytoplankton measured with a fluorometer. Journal of
Experimental Marine Biology and Ecology. 1: 191-208.
Fer-srNsrnIr J. 1995. PHYLIP (Phylogeny Inference Package) version
3.57c. Distributed by the author. Department of Genetics, University
of Washington, Seattle.GnreN J.C. & Honr T. 1994. Flagella and flagellar roots. In: The Hap'
tophyte Algae (Ed. by J.C. Green & B.S.C. Leadbeater), pp.47-71.
Clarendon Press, Oxford.GREEN J.C. & JoRDAN R.W. 1994. Systematic history and taxonomy.
lil The Haptophyte Algae (Ed. by J.C. Green & B.S.C. Leadbeater),
pp. l-21. Clarendon Press, Oxford.Hnrou S., LARssoN U. & Morsrnup @. 1996. Seasonal dynamics of
Chrysochromulina species (Prymnesiophyceae) in a coastal area
and a nutrient-enriched inlet of the Northern Baltic proper. Botanica
Marina 39: 281-295.Hrnseno D.J. 198O. Prymnesiophytes (: Haptophytes). In: Phytofa-
gellates (Ed. by E.R. Cox), pp. 273-317. Elsevier/North Holland,
New York.JrNsrN M.@. 1998. Seasonal dynamics of Chrysochromulina species
(Prymnesiophyceae, Haptophyta) in Danish coastal waters: diver-
sity, abundance and ecology. ln: The Gezus Chrysochromulina( Prymnesiophyceae) in Scandinavian Coastal Waters-Diversity,
Abundance and Ecology, pt. 4 (By M.@. Jensen), pp. 1-38' PhD
Thesis, University of Copenhagen.Kruune M. 1980. A simple method for estimating evolutionary rates
of base substitutions through comp.uative studies of nucleotide se-
quences. Journal of Molecular Evolution 16: I I l-120.
K-ellrn M.D., SrlvIN R.C., Cr-nus W. & GuIllnnD R.R.L. 1987. Me-
dia for the culture of oceanic ultraphytoplankton. Joumal of Phy-
cology 232 633-638.LARsEN A. & MEoI-n L.K. 1997. Inter- and intraspecific genetic var-
iation in twelve Prymnesizz (Haptophyceae) clones. Journal of
Phycology 33: 1007-1015.LEADBEA'IER B.S.C. 1994. Cell coverings.ln: The Haptophyte Algae
Eikrem & Edvardsen' A new species of Chrysochromulina 155
(Ed. by J.C. Green & B.S.C. Leadbeater), pp. 23-46. ClarendonPress, Oxford.
LEADBEATER B.S.C. & MANToN I. 1969. Chrysochromulina camellasp. nov. and C. cymbium sp. nov., two new relatives of C. strobilus
Parke and Manton. Archiv fiir Mikrobiologie 68'. 116-132.MAr.rroN l. 1966. Further observations on the fine structure of Chrys-
ochromulina chiton, with special reference to the pyrenoid. Journalof Cell Science l: 187-192.
Mrvroru l. 196'7a. Further observations on scale formation in Chrys-
ochromulina chiton. Joumal of Cell Science 2t 411-418.
MANToN l. 1967b. Further observations on the fine structure of Chrys'ochromulina chiton with special reference to the haptonema, "pe-
culiar" Golgi structure and scale production. Journal of Cell Sci'ence 2'. 265-272.
MEor-rN L.K., KooIsrRA W.H.C.E, PorrBn D., SauNoens G.W. & AN-nEnsEN R.A. 1997. Phylogenetic relationships ofthe "golden algae"(haptophytes, heterokont chromophytes) and their plastids. ln: TheOrigins of Algae and Their Plastids (Ed. by D. Bhattacharya), pp.
1 87 -219. Springer-Verlag, Vienna.MoEsrRUp O. 1984. Further studies on Nephroselmis and its allies
(Prasinophyceae). ll. Mamiella gen. nov. Mamiellaceae fam. nov.
Mamiellales ord. nov. Nordic Journal of Botany 4z lO9-121.Pnnrr M., MANToN I. & Cunxr B. 1955. Studies on marine flagel-
lates II. Three new species of Chrysochromulina. Journal of theMarine Biological Association of the United Kingdom 342 579-6O9.
Panxr M., MeNroN I. & Clenxr B. 1956. Studies on marine flagel-
lates III. Three further species of Chrysochromulina. Journal oftheMarine Biological Association of the United Kingdom 35'.387-414.
Penxs M., MANToN I. & Clnnrc B. 1958. Studies on marine flagel-lates IV. Morphology and microanatomy of a new species of Chrys-ochromulina. Journal of the Marine Biological Association of the
United Kingdom 37.. 2O9-228.PTENAAR R.N. 1994. Ultrastructure and calcification of coccolitho-
phores. In: Coccolithophores (Ed. by A. Winter & W.G. Siesser),pp. 13-37. Cambridge University Press, Cambridge.
PTENAAR R.N. & NoRRrs R.E. 1979. The ultrastructure of the flagellate
Chrysochromulina spinifera (Fournier) comb. nov. (Prymnesiophy-
ceae) with special reference to scale production. Phycologia 18:99-
108.SaNrsnoor J., Fnlrscs E.E & MeNInrIs T. 1989. Molecular Cloning:
A Inboratory Manual, 2nd ed.,3 vols. Cold Spring Harbor Labo-
ratory Press, New York.SrvoN N.. BnENNen J., EDVARDSEN B. & MEDLIN L.K. 199'7. The iden-
tification of Chrysochromulina and Prymnesium species (Haptophy-
ta, Prymnesiophyceae) using fluorescent or chemiluminescent oli-gonucleotide probes: a means for improving studies on toxic algae.European Journal of Phycology 32:393-401.
THoMSEN H.A., BucK K.R. & CrnvrzF.P. 1994. Haptophytes as com-ponents of marine phytoplankton. ln: The Haptophyte Algae (Ed.
by J.C. Green & B.S.C. Leadbeater), pp. 187-208. Clarendon Press,
Oxford.TunoNnsEN J. 1978. The dilution culture method. ln: Phytoplankton
Manual (Ed. by A. Sournia), pp.218-224. Monographs on Ocean-
ographic Methodology 6, UNESCO, Paris.Wsrrr TJ., BnuNs T., LrE S. & Tlvlon J. 1990. Amplification and
direct sequencing of fungal ribosomal RNA genes for phylogenet-
ics. In: PCR Protocols: A Guide to Methods and Applications (Ed.
by M.A. Innis, D.H. Gelfand, J.J. Sninsky & T.J. White), pp. 315-
322. Academic Press, San Diego.
Accepted 24 March 1999




















