
Incorporating molecular data in fungal systematics: a guide for aspiring researchers
Upload
independentCategory
view
3download
0
Botanicaljournal ofthe Linnean SocieF (1998), 127: 1 4 2 With 19 figures
Article ID: bt970164
Anatomical and molecular systematics of Asteliaceae and Hypoxidaceae
PAULA J. RUDALL*, MARK W. CHASE, DAVID F. CUTLER, JANET RUSBY AND ANETTE Y. DE BRUIJN
Jodrell Luborato?y, Royal Botanic Gardens, K e q Richmond, Surrgy 1 w 9 3DS
RecaVed October 1997; accepted f o r publication December I 9 9 7
The astelioid group of asparagoid lilies (Lilianae - Asparagales) comprises Hypoxidaceae, Asteliaceae, Blandfordia and Lamria. New information is presented on astelioid anatomy, together with a review of other systematic characters. These data are analysed in the context of recent evidence from rbcL nucleotide sequences that astelioids are related to orchids, and that astelioids and orchids @lus Alania and Bova) form a clade that is sister to all other asparagoid taxa. Hypoxidaceae and Asteliaceae differ from each other in several respects, but there are certain characters linking the two families, notably branched hairs and mucilage canals, unusual characters in Lilianae. Family diagnoses are upheld, but the precise relationships of Blandfordia and Lanuria are still poorly supported within the astelioid clade.
0 1998 The Linnran Society of London
ADDITIONAL KEY WORDS:-Asparagales - Blandfordia - cladistics - Lanana ~ Lnphiola ~ Ldianae - Orchidaceac
CONTENTS
Introduction . . . . . . . . . . . . . . . . Material and methods . . . . . . . . . . . .
Anatomical descriptions . . . . . . . . . . DNA extraction, gene amplification and sequencing . Cladistic analysis . . . . . . . . . . . . .
Results: morphological data . . . . . . . . . . . Characters for morphological analysis . . . . . .
Results of analyses . . . . . . . . . . . . . . General effects of successive approximations weighting Analysis of morphological/anatomical data . . . . DNA sequence analysis . . . . . . . . . . Combined analysis . . . , . . . . . . . .
Discussion . . . . . . . . . . . . . . . . Tree islands . . . . . . . . . . . . . . Successive approximations weighting . . . . . . Combining data . . . . . . . . . . . . . Phylogenetic considerations . . . . . . . . . Hypoxidaceae . . . . . . . . . . . . .
* Correspondence to: Dr Paula Rudall
0024-4074/98/050001+42 $25.00/0 1
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
. . . . . . .
2 2 2 3 4 5 5
15 15 17 20 20 23 23 23 27 29 29
0 1998 The Linnean Society of London

2 P. J. RUDALL ET.lL
Asteliaceae . . . . Lnnaria . . . . .
Astelioid relationships Acknowledgements . . References . . . . .
Appendix 2 . . . . .
Blandfordln . . . .
Appendix 1 . . . . .
. . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . .
30 31 31 32 33 33 35 36
INTRODUCTION
Recent DNA sequence analyses (e.g. Chase et al., 1995a) in combination with morphological studies (e.g. Chase, Rudall & Conran, 1996) have improved our understanding of family boundaries and affinities within the order Asparagales (superorder IJlianae). Asparagales comprise a ‘higher’ asparagoid clade with suc- cessive microsporogenesis and a ‘lower’ asparagoid grade, mostly with simultaneous microsporogenesis (Rudall et al., 1997). Analysis of rbcL sequence data by Chase et al. (1 995a) indicated a clade comprising Asteliaceae Dumort, Hypoxidaceae R.Br., Rlandfordiaceae Dahlgren & Clifford and Lanariaceae H. Huber ex R. Dahlgren (Table 1: the ‘astelioids’). Orchids and astelioids, together with Boryaceae M. W. Chase at al. (1996) form thc first branches of the ‘lower’ asparagoid grade. Since the systematics of this group are poorly understood, a re-assessment of both morphological and molecular data is timely. This paper presents new data on astelioid leaf anatomy and new ~bcL sequence data for the group and analyses them together with other relevant characters.
Anatomical description5
Appendix 1 lists material examined for vegetative anatomy. Living material was fixed in formalin acetic alcohol (FAA) and stored in 70% ethanol. Dried herbarium
’rum I . Astelioid genera
(;rnus No. spp Distributions Family
<d\telin Banks & Sol. 2 -5 Blnndfordia Sm. -1 Chlfospmium Skottsb. 7
(:Urculign Gaertn. r. 15 Empodium Salisb. f = Forb& Ecklon.) 10 Hvpoxidia Friedmami 1 or 2 t+po.ris L. c. 90 Innaria Aiton 1 .W//&ania Ho0k.f -1 .\fnlinerin Colla 7 .Vcva.deliu J. B. Williams 1 Paudia Ham. 2 Rhodohypoi Nel. 6 Snielln Hilliard & B. L. Rurtt 1 Spiforme Salisb (= farifhe Salisb.) 30
NZ, Austr., Parific, Mascarenes, E. Australia NZ, Fiji, Samoa southern hemisphere tropics South Africa Seychelles tropics & subtropics South Africa Tasmania Indornalaysia Australia (NSW) South Africa (SW Cape) SE Africa Lesotho South Africa
Chile Asteliaceae Blandfordiaceae Asteliaceae Hypoxidaceae Hypoxidaceae Hypoxidaceae Hypoxidaceae Lanariaceae Asteliaceae Hypoxidaccae Asteliaceae Hypoxidaceae Hypoxidaccae Hypoxidaceae Hypoxidaceae

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 3
TABLE 2. Taxa sequenced for rbcL
Genus, species Citation Voucher, herbarium Data base accession number
Alania endlichen Apostm'a s@lidioidrs Astelia alpina Astelia banksii Blandfordia punicea Blandfordia nobilis B o y a septentrionalir Collo$mmum hastatum Conanthera campanulata Curculigo capitata Doryanthes excelsa Empodium ueratrifoliam Hypoxidia rhizoptylla Hypoxis glabella Hypoxic leptocarpa Ixiolirion tartaricum Lanaria lanata Milhgania d&ora Milligania stylosa Pauridia longituba Rhodotypoxis millioides Spiloxene capasis
Chase et al., 1995a Chase et al., 1995a Rudall et al., 1997 this paper Chase et ab, 1995a this paper Rudall et aL, 1997 Chase et al., 1995a Chase et al., 1995a Chase et al., 1993 Chase et al., 1995a this paper this paper this paper Chase et al., 1993 Chase et al., 1993 Chase et aL, 1995a this paper Chase et al., 1995a this paper Chase et al., 1995a Chase et al., 1995a
3G Conran 707, A D C h e n t s 4823, CANB MW Chase 1103, K MW Chase 1072, K MW Chase 519, K MW Chase 2835, K M W Chase 2205, K Adelaide BG 875767 MW Chase 523, K MW Chase 205, NCU MW Chase 188, NCU Kirstenbosch b42-84 R. Wise s.n. OX M W C h a s e 2235,K MW Chase 108, NCU MW Chase 489, K Goldblatt 941 0, MO MW Chase 2874, K MW Chase 511, K D. Snijman 1440, WBG MW Chase 479, K UCI Arb 728
Y 14982 273705 277261 Y 14983 273694 Y 14984 Y14985 Y 14986 277312 273701 273697 Y14987 Y 14988 Y 14989 273702 273704 277313 Y 14990 273693 Y14991 277280 277281
material was revived by boiling in water. Sections were cut using a Reichert sliding microtome, stained in safranin and Alcian blue, dehydrated through an alcohol series to 100% ethanol, then placed in Histoclear, mounted into microscope slides in Euparal, and examined using bright-field optics on a Leitz Diaplan photomicroscope.
DNA extraction, gene ampl$cation and sequencing
DNA was generally extracted from either fresh silica or gel-dried leaf tissue. Details of material examined are listed in Table 2. Material newly examined here include Astelia banhii, Blandfordia nobilis, Empodium verat$lium, Hypoxidea rhizophylla, Hypoxis glabella, Millkania demjora, and Pauridia longituba.
Either 1 .O g fresh or 0.2-0.25 g silica gel-dried leaves were processed by the 2X CTAB method of Doyle & Doyle (1987). All samples were then purified on caesium chloride/ethidium bromide gradients (1.55 gml-' density). Gene am- plification of the rbcL gene was carried out using forward primers that matched the first 20 base pairs (bp) of the exon and a 20 bp reverse primer that begins at position 1370. Sequencing reactions were performed with these two primers plus two internal primers (636F GCGTTGGAGAGAGATCGTTTCT, and 724R: TCGCATGTACCYGCAGTTGC; the latter will not work on dicots) such that two overlapping segments were produced providing nearly complete coverage of both strands with only four cycle sequencing reactions. Amplified products were purified using Magic@ mini columns (Promega), following protocols provided by the manu- facturer. Standard dideoxy methods or modified dideoxy cycle sequencing with dye terminators run on an AF3I 373A automated sequencer (according to the

+ P J KLDALI, E’T.4L
manufacturer’s protocols; Applied Biosystems, Inc.) were used to sequence the amplification products directly. Approximately 1350 bp was used in the analysis for each taxon. Sequences were edited and assembled using the Sequencer Navigator and AutoAssembler software (Applied Biosystems, Inc.); rbcL sequences are aligned easily by eye because there is no internal length variation.
Cladistic anabsis
Cladistic analyses were undertaken using both heuristic and branch and bound parsimony algorithms of the soft\vare package PAUP for Macintosh (Phylogenetic Analysis using Parsimony, Version 3.1.1, Swofford, 1993) together with MacClade 3.04 (Maddison 8L hladdison, 1992) for tree manipulation and other calculations. For the morphological analysis, a branch and bound search was possible, but a similar warch on the larger matrix used in the combined analysis could not be completed, even after se\.eral days, so this was abandoned. For all analyses, heuristic searches were undertaken initially under the unordered and equal weighting criterion of Fitch parsimony (Fitch, 1971) with 1000 random sequence additions and tree liiscction-reconnection (TBR) branch swapping. Islands of equally parsimonious trtm (Xladdison, 1991) were detected if all shortest trees were not found in each replicate reaching the shortest tree length. With the morphological data two islands \\.ere discovered (see bcloiv). Successive approximations weighting (Farris, 1969) was used to ‘improve’ the matrix; in effect, this procedure optimizes the fit of the most consistent characters on the trees such that more changes are forced into the characters found to be least consistent in the initial round of analysis. The initial trees found with equal (Fitch) weights were used to re-weight all characters with a base weight of 1000 correlated to the rescaled consistency index (RC). Iterative succcssive weighting, based on trees collected at each round, was applied until the weighted length remained constant. Thc trees obtained in the final round were the ‘successive weighting trees’. We checked whether using the consistency index (CI) or retention index (RI) had any cffect on the outcome but obtained the same topologies. Several ‘fit’ options are alrailable in PAUP for use in successive weighting. It seems clear that re-weighting should be based on the ‘best fit’ of characters on any of the equally most parsimonious trees; this permits retention of characters that might be eliminated if other options, such as ‘worst fit’, were used. Bootstrapping ( 1 000 replicates with a limit of 25 trees per replicate and subtree-pruning-regrafting, SPR, swapping) was carried out with the successive weights applied. Successive approximations weighting does not ,greatly alter the patterns obtaincd with Fitch parsimony (in this case it found a subset of the trees identified with equal weighting, see below), and it simply places a penalty (by down-weighting) on characters that change frequently. We did not use Goloboffs (1 996) implied weighting procedure (PIWE) because we did not intend this paper to be an exercise in the application of dserential a posteriori weighting. Matrices are available on diskette or by email from the second author (MWC).
All astelioid genera (Hypoxidaceae, Asteliaceae, Lunaria and Blandfordia) were included in the analyses, except Neoastelia and Saniella (both monotypic), for which insufficient data and no sequences were available. Alania, Borya (Boryaceae) and Aflostczsia (Orchidaceae) were also included. Designated outgroups were Conanthera (Tccophilaeaceae), hiolirion (Ixioliriaceae) and Dolyanthes (Doryanthaceae).

SYSTEMATICS OF AS’TELIACEAE AND HYPOXIDACEAE 5
Summary of anatomical characters (data for Suniella from Hillard & Burtt, 1978; data for TABLE 3. B o y from Gaff & Churchill, 1976, and our unpublished data)
(:rIIu\ Hairs Mucilage canals Stomata Bulliform ~ c l l s
hranched absent absent branched brariched
unbranched absrnt branched branched absent hranched absent absrnt branched
branched >
sometimes present in leaves absent absent present in leaves present in rhizomrs & sometimes leaf basrs sometimes present in leaves absent from leaves present in rhizomes only absent absent absent absent from leaves present in leaves present in rhizomes only present in leaves sometimrs present in leaves & rhizomes
paracytic anomocytic paracytic paracytic paracytic/ tetracytic paracytic paracytic paracytic paracytic anomorytic paracytic paracytic paracytic paracytic paracytic parac ytic
present absent absent absent present
present present alxrnt/present present (midrib) absent ahwnt prrsent absent/prcwnt prcsrnt absent present
RESULTS: MORPHOLOGICAL DATA
Data on vegetative anatomy are presented in Appendix 2 and summarized in Table 3.
Charactersfor morphological anabsiJ
1. Mucilage canals. 0: absent, I : present Mucilage canals occur in Hypoxidaceae, Astelia and Collospemum, but not in
Blandfordia, Lanaria (Fig. 13) or Milligania (Table 3), nor in other asparagoid taxa (including orchids) or Lophiola (Fig. 14), and indeed are unusual in monocotyledons. In Hypoxidaceae mucilage canals are largely confined to the rhizomes (Fig. 8; also in the petiole of Curcul&o), whereas in Astelia and Collospemzum they are apparently absent from rhizomes but extend from the proximal to the distal ends of the leaves (Figs 9D, 10B, 11A) and are also sometimes present in flowers.
2. Trichomes. 0: absent or unbranched, 1: branched Branched (stellate) trichomes are rare in Asparagales but occur in Hypoxidaceae
(except Empodium) (Figs 1 ,2) and Astelia, Collospermum and Millkania, and also dendritic trichomes occur on inflorescences of Lunaria (Fig. 13D). This could therefore be a synapomorphy for the astelioid clade, except Bland@rdia. Branched hairs are rare in orchids, although Pridgeon (1 994) described multiseriate mycorrhizal trichomes in some species. Remarkably similar trichomes, both stellate and dendritic, occur in various genera of Haemodoraceae (Simpson, 1990), which is a homoplasy, since from other evidence Haemodoraceae belong in the commelinoid clade.
3. Bulltjim cells. 0: absent, 1: present
some other asparagoids such as some Iridaceae (Rudall, 1991, 1994). Bulliform cells are characteristic of Hypoxidaceae (Figs 4-7, although shared by

ti
Figiirc 1 . Drawings of trichomes. A, illilligania splosa. B, Astelia men&esiana. C, Empodiuni elongaturn. D, Enipodiiim plicatum. E, Curruligo uillosa. F, Curcul&o pilosa. G, H, Spiloxene .senata. J, Rhodohypoxls baurii. K, f&o.xts angurtfolia. L. Cunuligo rpcuruata. Scale har = 200 pm.
4. Tito prominent lateral veins. 0 absent, 1: present 5. Hybodermis. 0: absent, 1: present
A leaf hypodermal layer consisting of 1-3 layers of unlignified cells is present in

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 7
Figure 2. Leaf surfaces (SEW. A, Hypoxis rooperi, trichomes at margin. B, Asteliajagrans, adaxial surface covered in trichomes. C, Hypoxis urceolata, trichome base. D, Hypoxis rooperi, trichome base. E, Milligania splosa, abaxial surface with paracytic stomata. F, Astelia jagrans, stomata on abaxial surface (hairs removed). G, Hypoxis obtusa, paracytic stoma. H, Astelia3agrans, hair base. Scale bars = 200 pm in A, 500 pm in B, 100 pm in C, D, E, H, 50 pm in F, 20 pm in G.

Fi*gure 3. Crmzilgo lat$o/ra, leaf wrfaces. A, abaxial, with stomata. B, adaxial, lacking stomata. Scale bar= 100 pm.
many orchids, especially epiphytic Epidendroideae (Pridgeon, 1994), although not Apostasia (Table 3). This is an unusual character for asparagoids but is present in AJtelia, Collospmum and i\!lill&mia (Fig. 9A, D).
6. Pulh. 0: non-spinulose, 1: spinulose A~telia, Collospmum and Milligania are all recorded as having spinulose pollen,
which is lacking in the other astelioid genera and rare in asparagoids (though present in some lilioids). However, Cranwell (1953) reported that spines are indistinct or lacking in Millkania. BlQndfordia and Boya have reticulate pollen-surface sculpturing (Schulze, 1982).
7. Il/licmsporogenesis. 0: successive, 1 : simultaneous Rlicrosporogenesis is simultaneous in h.lilligania, as in most other ‘lower’ as-
paragoids, but successive in Hypoxis and Curculigo and unknown in Blandfordia, Astdza and Cu1lo.rperrnum (Rudall et al., 1997).
8. Nucellus. 0: rrassinuccllate, 1 : tcnuinucellate In the tenuinuccllatc condition, the megasporocytc is hypodermal rather than
dec$y seated; the hypodermal archespore gives rise directly to the megasporocyte without forming parietal cells. In crassinucellatc ovules (the plesiomorphic condition),
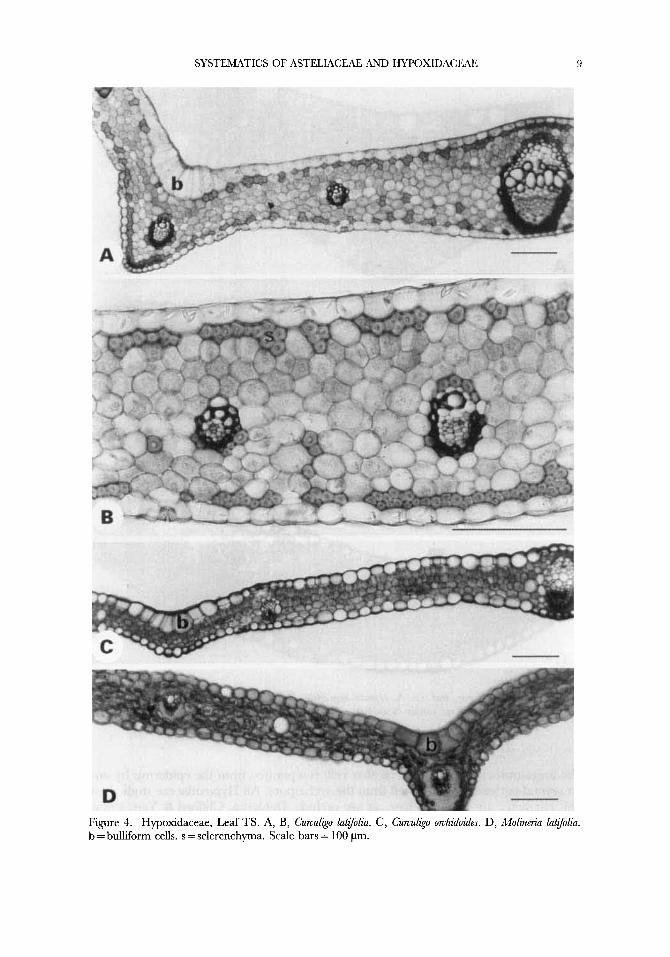
SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE
Figure 4. Hypoxidaceae, Leaf TS. A, B, Curculko lutfoliu. C, Curculip.0 orchidoides. D, Molineriu latfolia. b = bulliform cells. s = sclerenchyma. Scale bars = 100 pm.

P. J. KCDALI. ETSL.
Figure 5. bullifom cells. m = mucilage canals. Scale bars = 100 p.
Hypoxidaceae, leaf TS. A, HypoxU angust$olia. B, Spiloxm stellata. C, Spiloxene serrata. b =
the megasporocyte (megaspore mother cell) is separated from the epidermis by one or several parietal layers formed from the archespore. All Hypoxidaceae studied for this character are tenuinucellate, as are orchids (Dahlgren, Clifford & Yeo, 1985), but not Blandford& or Astelia, which are crassinucellate (unpublished data). Stevenson & Loconte (1 995) considered Hypoxidaceae close to Velloziaceae on this basis, but several molecular data sets place Velloziaceae near Stemonaceae, Pandanaceae and Cyclanthaceae (Chase et al., 1995a).
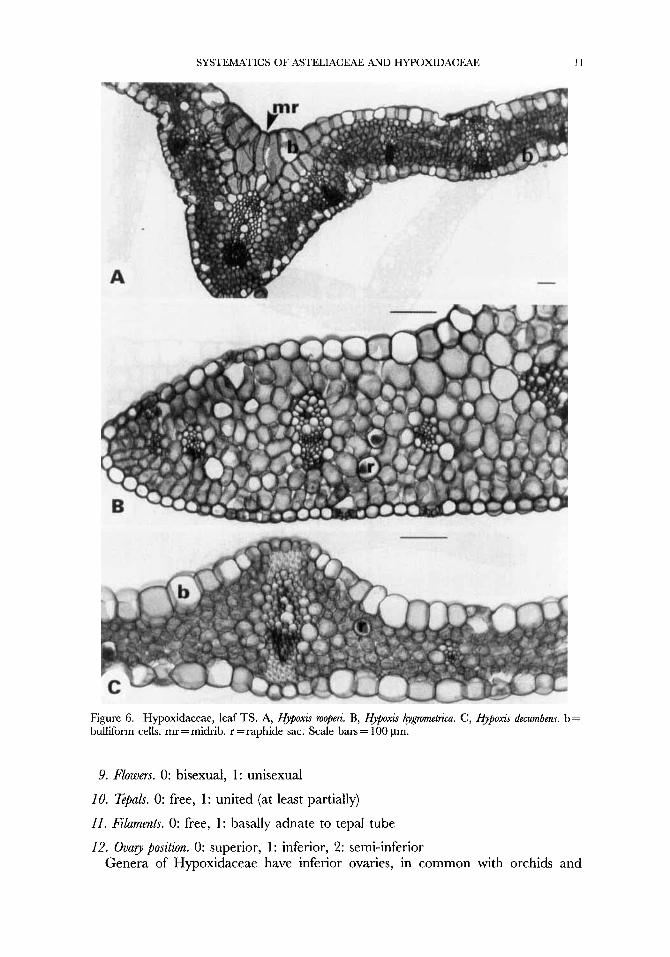
SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 11
Figure 6. Hypoxidaceae, leaf TS. A, Hypoxis mopm.. B, Hyj~oxis hygmmetriCa. C, Hypoxis decurnbens. b = bulliform cells. mr =midrib. r = raphide sac. Scale bars = 100 pm.
9. Flowers. 0: bisexual, 1: unisexual
10. Zpals. 0: free, 1: united (at least partially)
11. Filaments. 0: free, 1: basally adnate to tepal tube
12. Ovary position. 0: superior, 1 : inferior, 2: semi-inferior Genera of Hypoxidaceae have inferior ovaries, in common with orchids and
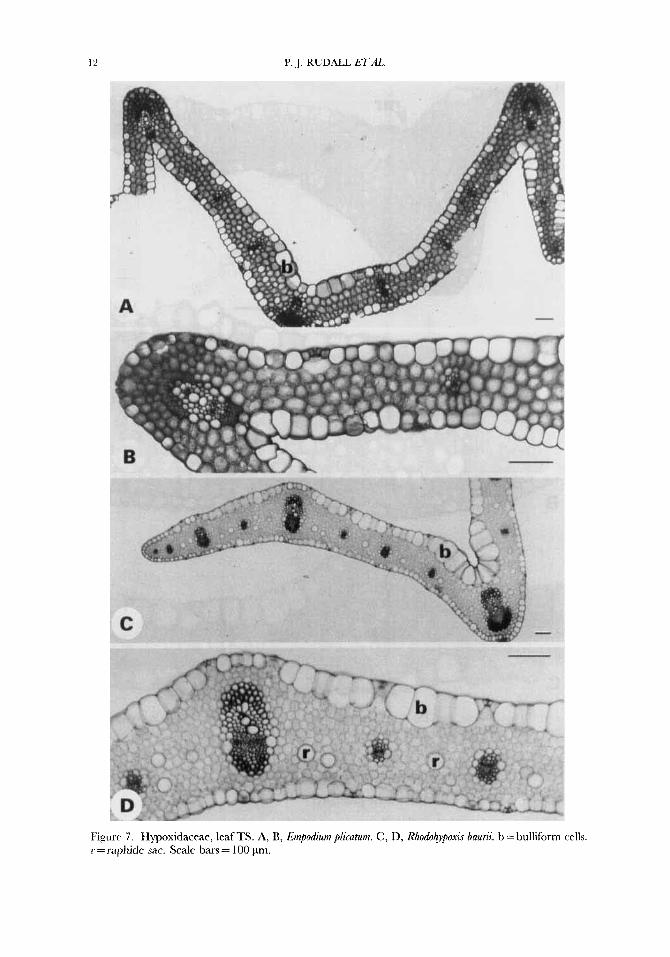
12 P. J. R U D U E T A , .
Figurr 7 . Hypoxidaceae, leaf TS. A, B, Empodium plicatum. C , D, Rhodohypoxi5 bauni. b = bulliform cells. r = raphide sac. Scale bars = 100 pm.
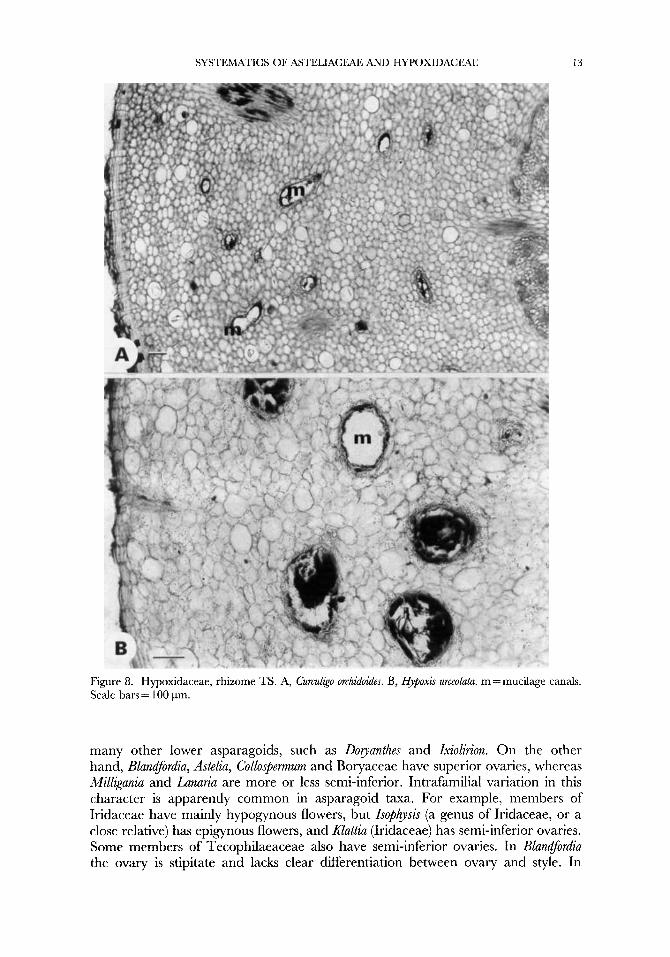
SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 13
Figure 8. Hypoxidaceae, rhizome TS. A, Curculigo orchidoides. B, Hypoxis urceolatu. m = mucilage canals. Scale bars = 100 pm.
many other lower asparagoids, such as Doryanthes and Ixiolirion. On the other hand, Blandfordia, Astelia, Collospemum and Boryaceae have superior ovaries, whereas Milligania and Lanaria are more or less semi-inferior. Intrafamilial variation in this character is apparently common in asparagoid taxa. For example, members of Iridaceae have mainly hypogynous flowers, but Isophpis (a genus of Iridaceae, or a close relative) has epigynous flowers, and Enattia (Iridaceae) has semi-inferior ovaries. Some members of Tecophilaeaceae also have semi-inferior ovaries. In Blandfordia the ovary is stipitate and lacks clear differentiation between ovary and style. In

P. J. RUDALL ETAL.
FiLpre 9. Asteliaceae, leaf sections. A, B, C, Bstelia.fiagrans. A, TS midrib, with hypodermis. B, LS blade. C , TS blade. D, Collospmurn hartatunz TS blade. h = hypodermis. r = raphide sacs. m = mucilage canals. Scale bars = 100 pm.
Lanaria there are only two large ovules per locule, with their micropyles directed downwards.
13. Placentation. 0: axile, 1: parietal Most orchids have a unilocular ovary with parietal placentation, which also occurs
in some Asteliaceae and Empodium (Hypoxidaceae). It is otherwise an unusual character in asparagoids, although also present in some Liliales such as Philesia.

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 15
14. Intra-ovarian trichomes. 0: absent, 1: present Intra-ovarian trichomes are present in some species of Astelia and Collospermum, in
which they are mucilage-secreting (Rudall, Prychid &Jones, in press). They are absent from other astelioids, although in orchids non-secretory trichomes, possibly homologous structures, are often present around the bases of seeds. Mucilage- secreting intra-ovarian trichomes also occur in a few monocot families, possibly to aid pollen tube growth into the micropyles (Rudall et al. in press).
15. Fruit. 0: capsule, 1: berry
capsule, rather than the more usual loculicidal asparagoid capsule.
'
A berry occurs in Astelia, Collospermum and Empodium. Blandfordia has a septicidal
16. Phytomelan (in seed coat). 0: absent, 1 : present
of Bolya (Churchill, 1987). The seed coat is hairy in Blandfordia (Huber, 1969) and papillate in some species
17. Undeyound stem. 0: rhizome, 1 : tunicated corm.
RESULTS OF ANALYSES
General efects of successive approximations wekhting
We examined the likely effects of using successive weighting by determining which characters were changing frequently. For example, among the morphological data, two characters, tepals free or united (character 10) and ovary position (character 12) changed more than six times on either the combined (Fig. 15A) or morphological trees (Fig. 15B). Several positions in rbcL also changed frequently (more than five times on the combined tree; Fig. 16), but especially relative to their numbers certain morphological characters were changing far more frequently than any of the molecular characters. This is why using weighting is appropriate, it is reasonable to ask what would be the effect of down-weighting such excessively changing characters. In this case, there was no change in overall tree length (the successively weighted trees in every case were a subset of the trees found with equal weights), but the increase in resolution was dramatic. Down-weighting frequently-changing characters resulted in a much clearer pattern from the morphological data and to a somewhat more limited extent from the rbcL data. However, if resolution were due to the effects of frequently changing characters, then successive weighting could increase tree number and decrease resolution (cf. Fay et al., in press).
Successive weighting on the morphological data eliminated one of the two islands (Fig. 17A, B) and selected an island that was more similar to the molecular tree (Fig. 17C). When successive weighting was applied to the rbcL data, then the number of trees produced was reduced from three to one, and the one tree selected was the single tree found when the morphological data were combined with the molecular. The overall effect of successive weighting was thus to improve confidence in the resolution obtained and secondly to select trees in each individual analysis that agreed with those produced when the two data sets were combined.
P. .J. KUD.\I,L E7 dl..
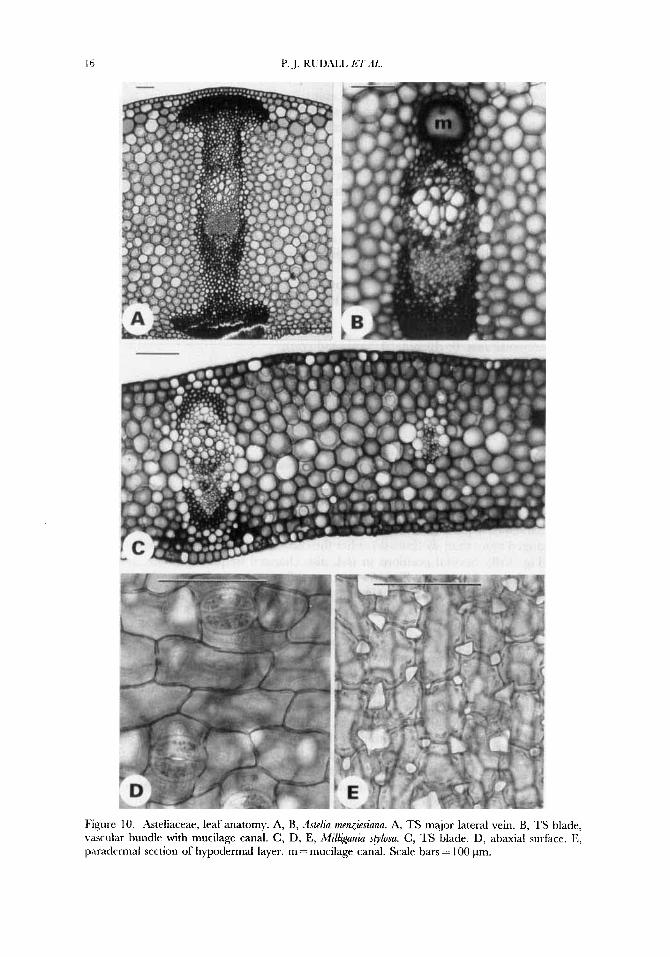
Figure 10. Asteliaceae, leaf anatomy. A, B, Bskdia menzitsinna. A, TS major lateral vein. B, TS blade, vascular bundle with mucilage canal. C, D, E, iMill~anza sglosu. C, TS blade. D, abaxial surface. E, paradermal section of hypodermal layer. m = mucilage canal. Scale bars = 100 pm.

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 17
Figure 1 1. Asteliaceae. A, Astelia banksii, LS base of leaf. B, Astelia alpina, rhizome TS. m = mucilage canal. Scale bars = 100 pm.
Anabsis of morphological/anatomical data
Using the morphological/anatomical data alone (Table 4), two ‘islands’ of trees of 32 steps were found, with a consistency index (CI) of 0.56 and a retention index (RI) of 0.7 1; in one island there were only two trees (Fig. 17A) whereas in the other there were 136 trees (Fig. 17B). Exact solutions find all trees, but the presence of
P. J. RCDAIL E T A 5
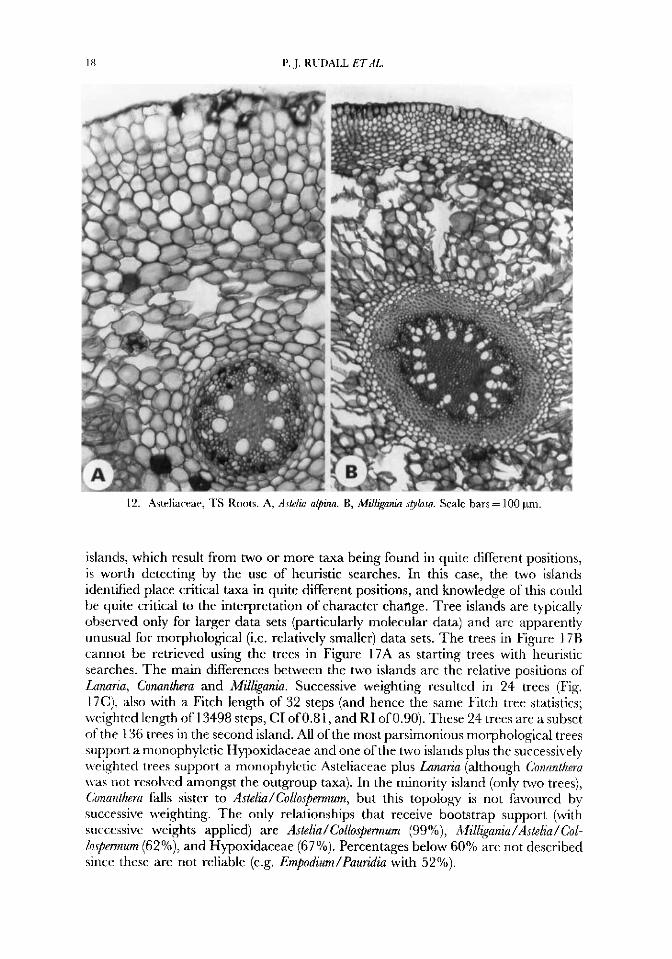
12. Asteliareae. TS Roots. A, A~telia alpirao. B, ,Mill&ania splosa. Scale bars = 100 pm
islands, which result from two or more taxa being found in quite different positions, is worth detecting by the use of heuristic searches. In this case, the two islands identified place critical taxa in quite different positions, and knowledge of this could be quite critical to the interpretation of character change. Tree islands are typically observed only for larger data sets (particularly molecular data) and are apparently unusual for morphological (i.e. relatively smaller) data sets. The trees in Figure 17B cannot be retrieved using the trees in Figure 17A as starting trees with heuristic searches. The main differences between the two islands are the relative positions of Lanaria, Conanthera and Milligania. Successive weighting resulted in 24 trees (Fig. 17C), also with a Fitch length of 32 steps (and hence the same Fitch tree statistics; weighted length of 13498 steps, CI of 0.8 1 , and RI of 0.90). These 24 trees are a subset of the 136 trees in the second island. All of the most parsimonious morphological trees support a monophyletic Hypoxidaceae and one of the two islands plus the successively iceighted trees support a monophyletic Asteliaceae plus Lanaria (although Conanthera was not resolved amongst the outgroup taxa). In the minority island (only two trees), Conanthera falls sister to Astelia/Collospmium, but this topology is not favoured by successive weighting. The only relationships that receive bootstrap support (with successive weights applied) are Astelia/Collos-ermum (99%), Mill~ania/Astelia/Col- lospennum (62%), and Hypoxidaceae (67%). Percentages below 60% are not described since these are not reliable (e.g. Empodium/Pauridia with 52%).
SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 19
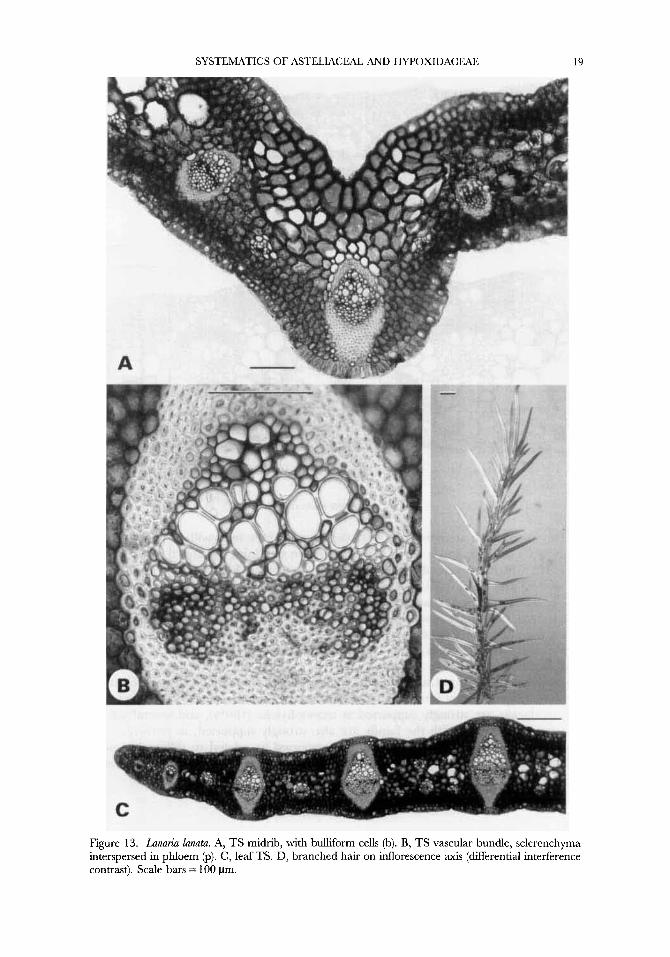
Figure 13. Lanuria lunutu. A, TS midrib, with bulliform cells (b). B, TS vascular bundle, sclerenchyma interspersed in phloem @). C, leaf TS. D, branched hair on inflorescence axis (differential interference contrast). Scale bars = 100 pm.

Figure 14. zI, B. Loplrioln arnrri~nmu, leaf TS. Scale bars = 100 pm
DJW sequence anabszi
The initial Fitch analysis using rbcL alone found three trees with a length of 446 steps, a CI of 0.70, and a RI of 0.58. Successive weighting produced a single tree (Fig. 18), which was one of the three Fitch trees (weighted length of 229 155 steps, CI of 0.92 and RI of 0.85). The main difference from the morphological trees (Fig. 17) is that amongst the astelioids, Lanaria is sister to Hypoxidaceae rather than Asteliaceae, but this position is not well supported by the bootstrap (60%). Additional differences are that Conanthera is clearly in the outLgroup (100%) and Blandfordia is 1veaMy supported (60%) as a member of the astelioids rather than related to Apostasia arid Dopznthes (which are not astelioids). The relationship of Millkaniu to Astelia/ Collospemtum (56%) is not supported by the bootstrap, although all three Fitch trees resolve Mzll@ania as sister to Astelia/Collospmum (ie. Asteliaceae are monophyletic). Hypoxidaceae are strongly supported as monophyletic (100%), and several sets of generic reiationships with the family are also strongly supported, in particular the tpvo species of Hypoxis are each strongly supported as related to different genera ( 1 0O0h bootstrap). All of the inter-familial relationships amongst the astelioids reccivc at least weak bootstrap support.
Combined anabsis
‘1’0 combine the two data sets, Molinnz’a was removed because we have no sequence data for it. Only one sequence was used for each genus in the rbcL matrix, except

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 21
" 1 2 3 4 5 6 7 8 9
Number of steps
" 1 2 3 4 5 6 7 8 9 Number of steps
Figure 15. Numbers of morphological characters plotted against the number of times they change (as estimated from steps on one of the shortest trees). A. Morphological characters as mapped onto the combined or rbcL tree (since they are the same). B. Morphological characters mapped onto a morphological tree. Note that mapping these onto the combined tree results in a greater number of overall steps.
for two species of each of Astelia, Blandfordia, Hypoxis and Milligania, and for these, we duplicated the morphological characters, although in some cases (especially the two Astelia species) there were observable differences between them. Alternative approaches would have been either to score the morphological matrix at the species level or to combine the molecular sequences for each genus, both of which would have been difficult and inappropriate for this investigation, and would probably have yielded broadly similar topologies at the genus level.
The combined Fitch analysis, using both morphological and molecular data, found a single tree of 484 steps with a CI of 0.67 and a RI of 0.60, which was also found with successive weighting (Fig. 19; weighted length of 242093 steps with CI of 0.92 and RI of 0.87). This topology is identical to that of the rbcL tree found with successive weighting (Fig. 17), but levels of internal support are improved. For example, the monophyly of the astelioids (Asteliaceae/Blandfordiaceae/ Hypoxidaceae/Lanariaceae) rises from 60% with rbcL alone to 7o0/o, and the mono- phyly of Asteliaceae receives some bootstrap support (73 %) whereas with rbcL alone there is no support. The overall bootstrap levels are much higher in the combined analysis: morphology alone 450, rbcL done 1570, and combined 1729 (i.e. these are the sums of bootstrap values across the trees); in no case does the level of support
22
1 2 3 4 5 6 7 Number of steps
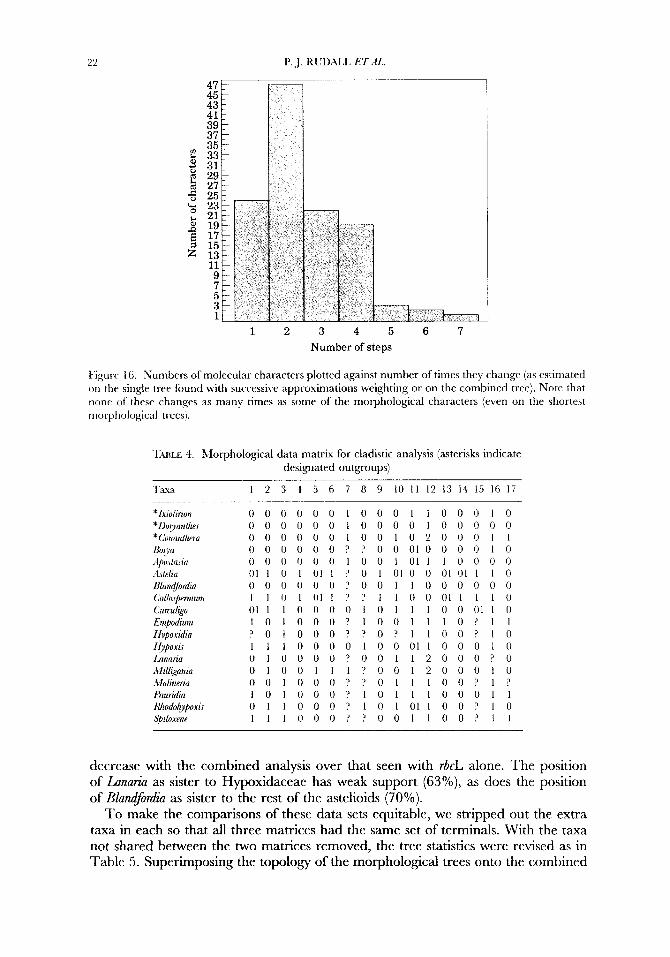
Pigurr 16. Numbers of molecular characters plotted against number of times they change (as estimated on the single tree found with successive approximations weighting or on the combined tree). Note that lion? of these changes as many times as s ~ m e of the morphological characters (even on the shortest moi-phological trees).
TULE 3. Lforphological data matrix for cladistic analysis (asterisks indicate designated outgroups)
Taus I 2 3 4 5 6 7 8 9 10 1 1 12 13 14 15 16 17
o o o o o o I o o o I I 0 0 0 I o o o o o o o I 0 0 0 0 1 o n 0 0 0 0 0 0 0 0 0 1 0 0 1 0 2 0 0 0 1 1 0 0 0 0 0 0 ? ? 0 0 0 i 0 0 0 0 1 0 0 0 0 0 0 0 1 0 0 1 0 1 1 1 0 0 0 0 011 0 1 0 1 1 ? 0 I 0 1 0 0 01011 1 0 0 0 0 0 0 0 ' 0 0 1 1 0 0 0 0 0 0 1 1 n 1 0 1 1 ? ~ i i o 0 0 1 i i i o 0 1 1 I o n n o 1 o I I 1 0 0 0 1 1 o l 0 1 0 0 0 ? i 0 0 1 1 1 0 ? 1 1 ? 0 1 O O O ? ? O ? 1 1 0 0 ' 1 0 1 1 1 0 0 0 0 1 0 0 011 0 0 0 1 0
0 1 0 0 I 1 1 ? 0 0 1 2 0 0 0 1 0 0 0 1 0 0 0 ? ? 0 1 1 1 0 0 ? 1 ?
0 1 1 0 0 0 ? 1 0 I 0 1 1 0 0 ? 1 0 1 1 1 0 0 0 ? ? 0 0 1 1 0 0 ? 1 1
o ~ o n o o ? o o i i ~ o o o ? o
I o I n o o ? I 0 1 I 1 0 0 0 1 1
decrease with the combined analysis over that seen with rbcL alone. The position of Lanuria as sister to Hypoxidaceae has weak support (63O/o), as does the position of Blandfordia as sister to the rest of the astelioids (70%).
To make the comparisons of these data sets equitable, we stripped out the extra taxa in each so that all three matrices had the same set of terminals. With the taxa not shared between the two matrices removed, the tree statistics were revised as in Table 5. Superimposing the topology of the morphological trees onto the combined

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 23
tree (or rbcL tree, they are identical trees) increased the number of steps by six (16.2%), but a similar exercise with the rbcL data on the morphological tree was only 40 steps less parsimonious (8.7%; Table 5). The molecular data are more consistent than the morphological data (0.71 versus 0.58), but the retention index of the morphological data was higher (0.72 versus 0.50). The pattern obtained with the combined data set was however identical to that produced by r6cL done.
DISCUSSION
Tree islands
It has become standard procedure to look for islands of equally parsimonious trees (cf. Maddison, 199 1) with molecular data, but this phenomenon is not restricted to such data. There is no reason apriori to believe that small morphological matrices cannot be similarly afflicted. In this instance, we subjected the morphological matrix to exactly the same methods of analyses as the rbcL data, and in undertaking random replicates of taxa entry order noted the characteristic pattern that marks islands: individual replicates found either two trees or 136 trees, but never did a single replicate find all 138 trees. The necessary correlate of this pattern is that if one tree from either island is used as a starting tree, then the other island is not found. Out of 1000 replicates, we found the small island much less frequently than the larger (78 times versus 848 times). It should also be noted that 74 times suboptimal islands were found, which means that if we had entered the taxa in one of these orders in a single heuristic search, we could have failed to find either of these shorter islands, Maddison (1991) stated that if the RI was 0.72 or less, then islands were possible, and these data produce an RI of 0.71, which makes them a candidate for the existence of islands. The r6cL data also had an RI under this limit, but we detected no islands (with 1000 replicates, it is possible that we simply did not discover a ‘rare’ island, i.e. one that was significantly less frequently found than the smaller of the two morphological islands). We wish to emphasize that such patterns must be considered possible with all classes of data.
In the end, islands exist as problems because for large matrices exact search algorithms are not feasible, and thus heuristic methods have to be used. In the case examined here, a branch and bound search could be used on the smaller morphological/anatomical matrix but not on the larger combined dataset. Even though the use of branch and bound searches avoids the problems of finding islands, it does not of course eliminate the existence of the radically different positions of the taxa involved, and knowing that these differences exist seems an important outcome of which to be aware. It involves drastically different interpretations of character evolution, which in the case of molecular data do not seem particularly significant, but in this instance it involves characters that have potential use in the taxonomy of these plants. Therefore, we investigated the nature of these two islands by the use of heuristic methods even though we found all trees with the branch and bound search as well.
Successive approximations wekhting
The use of successive weighting has been questioned by reviewers of recently submitted papers using molecular data (M. W. Chase, pers. comm.) because it was

21
A
Conanthera Lanaria Milligania Curculigo Hypoxidia Molineria
B
Empodium Pauridia Spiloxene Hypoxis Blandfordia 1p Apostasia Doryanthes Borya Ixiolirion
1 0 . Astelia
Collospermum Milligania
~onant~iera
Rhodohypoxis
O h n a r i a , I$ curculigo
Spiloxene Pauridia
Blandfordia Apostasia Doryanthes Borya Ixiolirion
Asteliaceae 0 Tecophilaeaceae Lanariaceae Asteliaceae
H ypoxidaceae
Blandfordiaceae Orchidaceae Doryanthaceae Boryaceae Ixioliriaceae
Asteliaceae
Lanariaceae [1
Tecophilaeaceae
Hypoxidaceae
Blandfordiaceae Qrchidaceae Doryanthaceae Boryaceae Ixioliriaceae
suspected of grossly affecting the patterns obtained. We have attempted in this study to examine the patterns of change in our characters such that a clear understanding of the effects of successive weighting can be obtained. The first observation is that regardless of which trees one uses (morphological, molecular, or combined) the same patterns of change are observed in most characters, and some characters do change much more frequently than others (Figs 1,2). Therefore, the effects of these frequently changing characters should be examined. In the case of the morphological data, the effect is to produce two islands of equally parsimonious trees (Fig. 17A, B) and many trees. When the overall signal in a matrix is strong, it comes through clearly, but when there is no clear signal, such high frequencies of change as in these

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 25
C
Hypoxidaceae
Asteliaceae n U
Lanariaceae Tecophilaeaceae Orchidaceae Doryanthaceae Blandfordiaceae Boryaceae Ixioliriaceae
Figure 17. Analysis of morphological data alone (Table 3). A, island one, B, island two. C, one of 24 trees found after successive approximations weighting. All trees have 32 Fitch steps with a CI of 0.56 and a RI of 0.71. The successively weighted trees have 13 498 steps with a CI of 0.81 and a RI of 0.90. Arrows indicate branches that collapse in the strict consensus tree. Numbers above branches are the numbers of steps. Numbers below branches in C are bootstrap values (based on I000 replicates with the weights from successive weighting applied).
characters only obscures the patterns observed with the more consistent characters (note that the majority of morphological characters change three or fewer times on the shortest molecular trees as well as on the shortest morphological trees). We are interested here in knowing how well the morphological patterns correspond with those found with the molecular data, but the excessive homoplasy in some characters creates two radically different patterns, one of which is quite at odds with the DNA tree. When these two patterns are condensed into a strict consensus tree, it appears as though there is no clear morphological result. Successive weighting produces quite a consistent set of relationships by down-weighting the frequently changing characters. It is not appropriate a prion' to eliminate characters, but down-weighting those that change frequently based on initial results should increase the resolution of the strict consensus tree. In this case, the use of successive weighting on the morphological data selects a topology that is more like the larger island (none of the patterns in the morphological data are strongly supported except for the sister relationship of Astelia and Collospermum; 99%). This island of trees is also more like the pattern found with the r6cL data alone (Fig. 18), as well as that of the combined analysis (Fig. 19) which is identical to that produced by r6cL alone. The DNA tree has fairly high levels of support, but the addition of the morphological data produces a large increase in support across the whole topology. Thus the use of successive weighting does three things that appear useful: (i) it discriminates between islands of equally parsimonious trees by favouring the more consistent characters and thus elevating the RI above the level at which islands are possible (note that the RI for all successive weighted trees were above the 0.72 cut-off indicated by Maddison,

26
3 - 60
~ 100
P. J. RUDALL ETAL.
~ 6 99 Astelia Asteliaalpina banksii Asteliaceae Collospermum
1 60
I ,"9 Milligania stylosa Milligania densiflora
Hypoxidia Hypoxis leptocarpa
93 Hypoxis glabella
Pauridia - Lanaria Lanariaceae 14 z
18 Blandfordia punicea 100 5 Blandfordia nobilis I Blandfordiaceae
Boryaceae 14 Alania 100 1.3 Borva
Apostasia Conanthera Doryanthes
I Zxiolirion
Orchidaceae Tecophilaeaceae Doryanthaceae Ixioliriaceae
Figure 18. Analysis of molecular data alone: single most parsimonious tree after successive ap- proximations weighting. This tree has the same Fitch length as the Fitch trees, 446 steps with a CI of 0.70 and a RI of 0.58 (weighted length 229 156 steps with a CI of 0.92 and a RI of 0.85). Numbers above branches are the numbers of Fitch substitutions (ACCTRAN optimization). Numbers below branches are bootstrap values (based on 1000 replicates with the weights from successive approximations weighting applied). 'The large arrow indicates the single branch not present in the strict consensus of the three Fitch trees.
1991); (ii) it produces a result similar to one of the two islands, which is also the one found by the molecular and combined datasets; and (iii) it permits a stronger si<gnal to be detected in the morphological data than would otherwise be the case so that the issue of data congruence can be more accurately assessed (the strict consensus of the two islands illustrated in Fig. 17A, B is so highly unresolved that it is difficult to conclude that there is any clear pattern in these morphological data). 'The trees found by successive weighting indicate that there is a pattern, and it is congruent with that produced by the molecular data. Assuming that we still would have bothered to combine such apparently signal-less data with the molecular data, the reasons for apparent synergism of two classes of information would be less obvious to us. Successive weighting revealed a much clearer pattern in the morphological data than we would have discovered had we employed only Fitch optimization. Clearly we discovered a great deal more about the value of these morphological data by using this approach.
Successive weighting at first may seem a method simply to reduce trees, but there is a geat deal more to be learned about data quality from its use. It permits identification of a few characters for which the patterns of variation obscure those produced by the majority, and it is thus a more critical method of examining data quality. It appears to work equally well for molecular and morphological data, and we recommend its use.

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 27
J
Asteliaceae
Hypoxidaceae
Lanariaceae
.-E Blandfordb Blandfordiaceae
Boryaceae
I 19 Blandfordia punicea 100
15 f Alania 100 .- 2o B o v a
Apostasia Orchidaceae Conanthera Tecophilaeaceae
Ixiolirion Ixioliriaceae 44 Doryanthes Doryanthaceae
Figure 19. Combined analysis of molecular and morphological data: single tree 484 steps with a GI of 0.67 and a RI of 0.60 (also the same single tree found with successive approximations weighting; 242 093 weighted steps with a CI of 0.92 and a RI of 0.87). Numbers above branches are the numbers of Fitch substitutions (ACCTRAN optimization). Numbers below branches are bootstrap values (based on 1000 replicates with the weights from successive approximations weighting applied).
TABLE 5. Tree data for separate and combined analyses with taxa not shared between matrices removed from all calculations (all tree lengths calculated under the Fitch criterion, rbcL tree and combined are the same tree, hence the question of how much longer the rbcL data are on the combined
tree is moot)
Tree length CI RI Percent longer
Combined tree 455 0.69 0.51 na Morphological data on morphological tree 31 0.58 0.72 na Morphological data on combined/rbcL tree 37 0.49 0.60 16.2% rbcL data on morphological tree 458 0.65 0.33 8.7% rbcL data on combined/rbcL tree 418 0.7 1 0.50 na
Combining data
Combining the two (morphological and molecular) data sets can be considered problematic because one (morphology) is scored at the genus level, including data from several species, whereas the other (rbcL) is sampled at the species (or individual)

28 P. J. RUDALL E‘TAL.
level. Scorinp the momhological matrix at the sDecies level would have resulted in
30 P. J. RUDALL ETAL.
TABLE 6. Comparison of itiilligania and other Asteliaceae
Alilligania Astelia & Collo@mum
Flowcn Filaments Style Septa1 nectaries Ovary Intra-ovarian tnchomes Fruit Pollen Mucilage canals I r a r s I raws
hermaphroditc basally adnate to tcpals tripartite short, in upper part of ovary semi-inferior absent loculicidal capsule spinulose ahsent wit11 prominent midvein with silky ‘wool’ (branched hain)
dioecious or gynodioecious free fused (sometimes very short) often extensive, branching superior sometimes present berry markedly spinulose present with two prominent lateral veins with silky ‘wool’ (branched hairs)
(Thompson, 1979), in contrast to the sulcate pollen of other astelioids (the ple- siomorphic state) and resembling the ‘operculate’ pollen of Tecophilaeaceae (Simpson, 1983, 1985). Pauridia also differs from other Hypoxidaceae in having nuclear endosperm formation (De Vos, 1948, 1949, 1963), but the presence of mucilage canals in the leaf, an unusual character in monocots, indicates its correct placement in this family.
In the rbcL analysis the two individuals sequenced for Hypoxis were not sister taxa within Hypoxidaceae. Hypoxis glabella is an unusual glabrous Australian species (Conran pers. comm.; not included in the morphological data), and may well represent a distinct taxon, although more research is necessary to assess this and the circumscription of many of the genera. For example, these data indicate that Rhodohypoxis (South Africa; Table 1 ) is surprisingly similar to a species of Hypoxis from North America, and it should be compared to other Africa species. Likewise, Spiloxene (South Africa) is not particularly divergent from H. glabella (Australia). These results indicate that a thorough examination of generic limits in Hypoxidaceae is w-arranted.
Asteliaceae
Astelia, Collospennum, Milligania and also Jveoastelia (not included in the analysis here) form a well defined family (Asteliaceae), with Milligania as sister to the other genera. The main character defining Asteliaceae is spinulose pollen (character 6, Table 4), which is absent from other astelioids. Astelia and Collospennum share several other characters which are lacking in Milligania (Table 6). Skottsberg (1934) regarded itfilligania as closely related to Astelia, but laid great emphasis on the similarities in the leaf hairs, despite “important differences in flower structure”. Our results demonstrate only weak support for the inclusion of Milligania in Asteliaceae, and it is clearly a divergent outlier (both in morphological and molecular terms) to the other genera of the family. Astelia and Collospermum, formerly congeneric, differ in our matrix (Table 4) only in some characters that are variable within Asklia. Astelia

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 29
Phylogenetic considerations
Hypoxidaceae and Asteliaceae differ from each other in gynoecium position and pollen sculpturing, but they are similar in other respects, notably branched hairs and mucilage canals, both unusual or unique characters in Lilianae. According to Skottsberg (1 934), Brongniart also put Asteliaceae close to Hypoxidaceae. There is now much evidence that the astelioids (as listed in Table 1) form a clade, mainly from the southern hemisphere, although the position of the isolated genus Blandfordia is still unresolved. However, to regard the astelioids as a single large family is untenable in view of the large differences between them and the relative distinctness of Hypoxidaceae. Hypoxidaceae (nine genera), Asteliaceae (three or four genera), Lanariaceae (monogeneric) and Blandfordiaceae (monogeneric) are here upheld as separate and distinct families. Lanuria is most likely the sister taxon to both Hy- poxidaceae (Fig. 18) and Asteliaceae (Fig. 17).
Hypoxidaceae
Both morphological and molecular analyses consistently supported Hypoxidaceae as a monophyletic clade. In the morphological analysis (Fig. 17), the following characters were apomorphies for Hypoxidaceae: (3) the presence of bulliform cells, (7) successive microsporogenesis, (8) tenuinucellate ovules, ( 1) mucilage canals and (2) branched trichomes, the latter two characters shared with Asteliaceae. Hilliard & Burtt ( 1978) considered Rhodohypoxis close to Hypoxis, and Empodium not closely related to Molineria, the latter supported by the morphological data, although there is a lack of sequence data for Molinm’u. Hilliard & Burtt (1978) noted several characters that they considered homoplasious within Hypoxidaceae, such as the presence of an ovary ‘beak‘, variation in fruit, seed and underground stem mor- phology, and also variation in stamen morphology: in Empodium, Molineria, Rhodo~poxis and Saniella the filament joins the anther low down on the outer face, whereas in Curculigo, Hypoxis, Pauridia and Spiloxene the filament joins the anthers medianly in the sinus between its basal lobes. They suggested that the presence of mucilage canals in the leaves may be indicative of a close relationship, but since this character is variable within some genera (Table 3), it requires further research.
Both Empodium and Pauridia are on long branches in the rbcL analysis (Fig. 18), and this perhaps reflects their equally great morphological differences from the rest of the family. Empodium differs from other Hypoxidaceae in several respects: it has a unilocular ovary filled with mucilage, parietal placentation (as in orchids) and the fruit is a berry rather than a capsule (Markotter, 1936; Thompson, 1978), all characters in common with some Asteliaceae rather than Hypoxidaceae. Empodium also differs from most Hypoxidaceae in having a several-layered micropylar nucellus (although still tenuinucellate, as the archesporial cell is hypodermal), Allium-type of embryo sac development, and a chalaza1 haustorium from the basal endosperm chamber (De Vos, 1949). However, based on the tenuinucellate condition and other characters, Empodium is correctly placed in Hypoxidaceae. Dahlgren, Clifford & Yeo (1 985) were uncertain about including the Cape genus Pauridia in Hypoxidaceae because it has only three stamens. However, from a study of floral vasculature, Thompson (1 978) convincingly demonstrated that the unusual six-lobed stigma in Paudia represents three staminodes fused to the style. Pollen of Pauridia is bisulcate

30 P. J. RUDALL E T A .
TABLE 6. Comparison of Milligania and other Asteliaceae
Milligania Astelia & Collospennum
Flowers Filaments Style Septal nectaries
Intra-ovarian trichomes Fruit Pollen Mucilage canals Leaves Leaves
Ovary
hermaphrodite basally adnate to tepals tripartite short, in upper part of ovary semi-inferior absent loculicidal capsule spinulose absent with prominent midvein with silky ‘wool’ (branched hairs)
dioecious or gynodioecious free fused (sometimes very short) often extensive, branching superior sometimes present berry markedly spinulose present with two prominent lateral veins with silky ‘wool’ @ranched hairs)
(Thompson, 1979), in contrast to the sulcate pollen of other astelioids (the ple- siomorphic state) and resembling the ‘operculate’ pollen of Tecophilaeaceae (Simpson, 1983, 1985). Pauridia also differs from other Hypoxidaceae in having nuclear endosperm formation (De Vos, 1948, 1949, 1963), but the presence of mucilage canals in the leaf, an unusual character in monocots, indicates its correct placement in this family.
In the rbcL analysis the two individuals sequenced for Hypoxis were not sister taxa within Hypoxidaceae. Hypoxis glabella is an unusual glabrous Australian species (Conran pers. comm.; not included in the morphological data), and may well represent a distinct taxon, although more research is necessary to assess this and the circumscription of many of the genera. For example, these data indicate that Rhoddypoxis (South Africa; Table 1) is surprisingly similar to a species of Hypoxis from North America, and it should be compared to other Africa species. Likewise, Spiloxene (South Afi-ica) is not particularly divergent from H. glabella (Australia). These results indicate that a thorough examination of generic limits in Hypoxidaceae is warranted.
Asteliaceae
Astelia, Collospmum, Milligania and also Neoastelia (not included in the analysis here) form a well defined family (Asteliaceae), with Milligania as sister to the other genera. The main character defining Asteliaceae is spinulose pollen (character 6, Table 4), which is absent from other astelioids. Astelia and Collospmum share several other characters which are lacking in Milligania (Table 6). Skottsberg (1 934) regarded Millkania as closely related to Astelia, but laid great emphasis on the similarities in the leaf hairs, despite “important differences in flower structure”. Our results demonstrate only weak support for the inclusion of Milligania in Asteliaceae, and it is clearly a divergent outlier (both in morphological and molecular terms) to the other genera of the family. Astelia and Collospmum, formerly congeneric, differ in our matrix (Table 4) only in some characters that are variable within Astelia. Astelia is a widespread genus with several distinct sections (Skottsberg, 1934), and a more detailed (species-level) morphological matrix, together with greater sampling for rbcL, is required to resolve the systematics of Asteliaceae (including Neoastelia). Like Hypoxidaceae, generic delimitation is in need of critical review.

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 31
Lunaria
The results of the morphological and molecular analyses differed only with respect to the position of Lunaria, which was sister to Asteliaceae in the morphological trees (Fig. 17B, C), and to Hypoxidaceae in the molecular (Fig. 18) and combined (Fig. 19) trees. In neither case is there any internal support, and the exact position of Lunaria must still be considered enigmatic.
Lunaria is a taxonomically isolated genus, formerly included in Haemodoraceae (e.g. by Hutchinson, 1959; Geerinck, 1969), and more recently (Dahlgren et al., 1985) in Tecophilaeaceae, although Dahlgren & Van Wyk (1 988) and G. Dahlgren (1989) put it in Lanariaceae, which is supported here. Lunaria resembles Haemo- doraceae in inflorescence morphology and tomentose flowers, but differs from them in many other respects (Simpson, 1990). It has thick leaves, as in Asteliaceae, with a clearly defined midrib tissue, as in both Asteliaceae and Hypoxidaceae, and also has paracytic stomata, as in most lower asparagoids except Blandfordia. It lacks hairs on the leaves, but has copious highly branched hairs on the flowers and inflorescences. It lacks mucilage canals in the leaves. Dora & Edwards (1 99 1) recorded the presence of a novel biflavone in Lunaria lanata and observed that biflavonoids have been found in few other monocotyledons: Patersonia and Isophysis (Iridaceae), Xemphyta (Velloziaceae) and Lophiola.
The monotypic north American genus Lophiola has often been associated with Lunaria. Since Simpson & Dickison (1981), Zavada, Xu & Edwards (1983) and Simpson (1 990) demonstrated that Lophiola is not close to Haemodoraceae, with which it was long associated (on the basis of inflorescence morphology, tomentose flowers and unifacial leaves), it was placed by default with Lunaria. However, Lophiola and Lunaria differ in leaf morphology and anatomy and are not closely related from rbcL data (Chase et al., unpubl.), which indicate that Lophiola belongs within Narthecieae (Melanthiaceae s.1.; JVarthecium, Metanarthecium and Aletris).
Blandfordia
Although the rbcL tree indicates a placement for Blandfordia as sister to the other astelioids (Fig. 18), its position here is not supported in the morphological analysis (Fig. 17). In some individual trees, Blandfordia and Apostasia were a sister pair on the basis of lack of phytomelan in the seed coat, perhaps an unreliable character in view of the marked differences in seed morphology (large hairy seeds in Blandfordia, ‘dust’ seeds in orchids).
Morphologically, Blandfordia does not fit well into the astelioid group, or more importantly anywhere else. Stevenson & Loconte (1 995) also noted that Blandfordia shows a lack of homology with other Lilianae. Blandfordia has previously been associated with Hemerocallis, Hesperocallis, Hosta, Lucocrinum and Phormium (e.g. Krause, 1930) and Kniphofia (Hutchinson, 1959). Fulvio & Cave (1964) noted that although it has characters in common with these genera, it differs from all of them, for example in having a septicidal (rather than loculicidal) capsule. They could find little from embryology that would help in assessing relationships, although they concluded that Blandfordia is not closely related to either Hememcallis or Kniphoja. It has an unusual hairy seed coat (Huber, 1969), which differs from that of Eriospmum in that the latter has long pointed silky hairs, whereas Blandfordia has much shorter

32 P. J. RCDALL E‘TAL
rounded hairs. Some species of Bova (Boryaceae: Chase, Rudall & Conran, 1996) have papillate testas (Churchill, 1987), possibly homologous with the hairy testa of Blandfordia, although there are no other obvious similarities between the two genera. The gynoecium of Blandfordia is stipitate, and the ovary is not distinct from the style. Skottsberg (1 934) also noted that in some species ofdsklia the pistil is “bottle-shaped, with a gradual transition between ovary and style”, although in other Astelia species the style is short or absent. Blandfordia lacks branched hairs and resin canals and has anomocytic stomata (Table 3), in contrast to the tetracytic or paracytic stomata of other astelioids and many other ‘lower’ asparagoids including the apostasioid orchids (Stem, Cheadle & Thorsch, 1993). In orchids, however, both anomocytic and tetracytic/paracytic stomata1 types occur. In spite of clear molecular patterns for Blandfordia (Chase et al., 1995a and this paper) that indicate a relationship to the lower asparagoids, it remains difficult to defend this position based on morphological or anatomical patterns. At the same time, an argument for a better position elsewhere is lacking, leaving us with the impression that more research is required.
Astelioid relationships
Astelioids are amongst the early-branching asparagoid taxa (the paraphyletic ‘lower asparagoids’) from both molecular and morphological evidence (e.g. mi- crosporogenesis: RudaIl et al., 1997). Analysis of rbcL placed them in the first- branching clade of Asparagales, together with Orchidaceae and Boryaceae (Chase et al., 1995a). Most of these genera (Table 1) have long been of uncertain affinity. Previously, Asteliaceae were linked with various Asparagales, especially Dracaena (Bentham & Hooker, 1883), which is now placed close to Convallariaceae in the ‘higher’ asparagoids (Chase et al., 1995a). Cordyline (including Cohnia) was formerly put in Asteliaceae (Dahlgren et al., 1985), but correctly belongs among the ‘higher’ asparagoids in Lomandraceae (Chase et al., 1996). Hypoxidaceae have been allied with a wide range of families, including several members of Asparagales (Geerinck, 1968) and Haemodoraceae. However, as several authors have demonstrated, a close relationship between Hypoxidaceae and Haemodoraceae can be ruled out. Haemodoraceae resemble Hypoxidaceae in several respects, but clearly belong in the commelinoid clade from analysis of both rbcL sequence data and morphological data (e.g. Chase et al., 1995a, b).
Hutchinson (1 959) considered the apostasioid orchids to be derived from Hy- poxidaceae, especially Curculigo, and noted that “the diandrous orchids, the most primitive”, have monosulcate grains with “little pattern”, as in Hypoxidaceae. Stern et al. (1 993) discussed the relationships of Hypoxidaceae with apostasioid orchids, but found few similarities. Orchids have many characters that are unique or unusual amongst asparagoids (e.g. silica bodies, velamen, mesophyll idioblasts and microspermy) or difficult to assess (e.g. mycorrhizal association). Silica bodies occur in leaves of some terrestrial and epiphytic orchids (Dahl Moller & Rasmussen, 1984), but amongst monocotyledons they are otherwise present only in the commelinoid clade, not in astelioids or other Asparagales. Some epiphytic orchids have a several- layered epidermis (velamen) in their aerial roots, which is not recorded in astelioids, even among epiphytic species of Astelia and Collospmum. Mesophyll idioblasts with helical thickenings are fairly common in orchids (Pridgeon, 1994), but apparently absent from astelioids (and other Asparagales). Microspermy (numerous, minute

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 33
‘dust’ seeds) or miniaturization of ovules is an orchid character. Many astelioids have ovaries with numerous small ovules, the plesiomorphic condition for Asparagales, but in orchids there are hundreds of ovules on branching placentas per ovary. Mi- crospermy is otherwise rare in monocotyledons, although it occurs elsewhere in the angiosperms; for example, Endress (1 990) reported at least two separate developments in Magnoliidae. Orchids are also well known for their obligate mycorrhizal as- sociations. Aluniu and B o y also have mycorrhizal roots (Keighery, 1984), and Laughton (1964) reported mycorrhizal roots not only in Hypoxis, but also in several other South African asparagoids, such as Anthericum, Scilla and Gladiolus, indicating that this character may be widespread and requires further assessment.
ACKNOWLEDGEMENTS
We are greatful to the following for supplying material for molecular analysis: Peter Goldblatt (Missouri Botanical Garden), Rosemary Wise (Dept of Plant Sciences, Oxford University), Mark Whitten (Florida Museum of Natural History) and Tony Hall (Alpine Section, RBG, Kew).
REFERENCES
Arber A. 1922. On the nature of the ‘blade’ in certain monocotyledonous leaves. Annals ofBotany 36:
Bentham G Hooker JD. 1883. Genera Plantarum 3. London. Betts W. 1920. Notes on the autecology of certain plants of the peridotit belt, Nelson. 1: Structure
of some of the plants (No. 3). Transactions ofthe New zealand Institute 52. Chase MW, Soltis DE, Olmstead RG, Morgan D, Les, DH, Mishler BD, Duvall MR, Price
RAY Hills HG, Qiu Y-L, Kron KAY Rettig JH, Conti E, Palmer JD, Manhart JR, Sytsma KJ, Michaels HJ, Kress WJ, Donoghue MJ, Clark WD, Hedren My Gaut BS, Jansen RK, Kim K;T, Wimpee CF, Smith JF, Furnier GR, Straus SH, Xiang Q-Y, Plunkett GM, Soltis PS, Swenson S, Eguiarte LEY Learn GH, Barrett SCH, Graham S, Dayanandan S, Albert VA. 1993. Phylogenetics of seed plants: an analysis of nucleotide sequences from the plastid gene rcbL. Annals ofthe Missouri Botanical Garden 8 0 528-580.
Chase MW, Duvall MR, Hills HG, Conran JG, Cox AV, Eguiarte LEY Hartwell J, Fay MF, Caddick LR, Cameron KM, Hoot S. 1995a. Molecular systematics of Lilianae. In: Rudall, PJ, Cribb PJ, Cutler DF, Humphries CJ, eds. Monocotyledons: Systematics and Evolution. London: Royal Botanic Gardens, Kew, 109-137.
Chase My Stevenson DW, Wilkin P, Rudall P. 199513. Monocot systematics: a combined analysis. In: Rudall PJ, Cribb PJ, Cutler DF, Humphries CJ, eds. Monocotyledons: Systematics and Evolution. London: Royal Botanic Gardens, Kew, 685-730.
Chase My R u d d P, Conran J. 1996. New circumscriptions and a new family of asparagoid lilies: genera formerly included in Anthericaceae. Kern Bulletin 51: 667-680.
Chase M, Rudall P, Conran J. 1997. Validation of the family name Boryaceae. K m Bulletin 52: 416.
Churchill DM. 1987. Bolya. Flora ofAustralia 45: 268-279. Cranwell LM. 1953. New Zealand pollen studies. The monocotyledons. Bulletin ofthe Auckland Institute
Dahl Meller JD, Rasmussen H. 1984. Stegmata in Orchidales: character state distribution and
Dahlgren RMT, Clifford HT, Ye0 PF. 1985. 7 h e families ofthe monocotyledons. Berlin: Springer-Verlag. Dahlgren RMT, Van Wyk AE. 1988. Structure and relationships of families endemic to or centered
in southern Africa. In: Goldblatt P, Lowry PP, eds. Modem systematic studies in Ajkcan botany. Proceedings
329-35 1.
and Museum 3: 1-91,
polarity. BotunicalJoumal of the Linnean Society 89: 53-76.

34 P. J. RUDALL E'TAL.
ofthe Ekventh Plenary Meeting ofthe Association for the Taronomic study ofthe Flora o f Tmpical A&ca. Missouri: Missouri Botanical Garden, 1-94.
Dahlgren G. 1989. An updated angiosperm classification. Botanical Journal ofthe Iknean Sociep 100:
De Vos MP. 1948. The development of the ovule and the seed in the Hypoxideae. I. Zanthe Salisb.
De Vos MP. 1949. The development of the ovule and the seed in the Hypoxidae. 11. The genera
De Vos MP. 1963. Studies on the embryology and relationships of South African genera of the
Dora G, EdwardsJM. 1991. Taxonomic status of Lanuria hnata and isolation of a novel biflavone.
DoyleJJ, DoyleJL. 1987. A rapid DNA isolation procedure from small quantities of fresh leaf tissue.
Endress PK. 1990. Evolution of reproductive structures and functions in primitive angiosperms
197-203.
Journal of South Ajhcan Botany 14: 159-169.
Paunrlia Haw. and Forbesia Ecklon. Journal of South Afncan Botany 15: 13-22.
Haemodoraceae: Lanaria At. Journal of South A h a n Botany 29: 79-90.
Journal of M u r a l Products 54: 79&80 1.
Phvtochical Bulletin 19: 11-15.
(hfa,gnoliidae). Memoirs of the Nmo 25rk Botanual Garden 55: 5-34. Farris JS. 1969. A successive approximation approach to character weighting. $sternatic <oolagy 18:
37G-385. Fay MF, Olmstead RG, RichardsonJE, Santiago E, Prance GT, Chase MW. 1998. Molecular
Fitch WM. 1971. Towards defining the course of evolution: minimum change for a specific tree
Fulvio TE, Cave MS. 1964. Embryolog): of Blandfordia nobilis Smith (Ldiaceae), with special reference
Gaff DF, Churchill DM. 1976. Bova nitida Labill. - an Australian species in the Liliaceae with
Geerinck D. 1968. Considerations taxonomiques au sujet des Haemodoraceae et de Hypoxidaceae
Geerinck D. 1969. Genera des Haemodoraceae et des Hypoxidaceae. Bulletin of the Jardin Botanique
Goloboff PA. 1996. PIWE Phszon 2.5. Program and Documentation. Published privately by the
Heidemau ME. 1983. Studies of diagnostic features in the genus Hypoxis L. (Hypoxidaceae R.Br.)
Hilliard OM, Burtt BL. 1978. Notes on some plants from southern Africa chiefly from Natal: VII.
Huber H. 1969. Die Samenmerkmale und Venvandtschaftsverhaltnisse der Liliifloren. Mitt Bot.
Hutchinson J. 1959. 'The families ofjowm'ng plants. I f . Monoco$ledonr. 2nd edition. Oxford Oxford
Keighery GJ. 1984. The Johnsonieae (Liliaceae); biology and classification. Flora 175: 103-108. Krause K. 1930. Liliaceae. In: Engler A, Prantl K, eds. Die .Matiirlichen gYanzenfamilien. ed. 2. 15a:
LLaughton EM. 1964. Occurence of fungal hyphae in young roots of South African indigenous
Lov L. 1926. Zur Kenntnis der Entfaltungszellen monokotyler Blatter. Flora 120 283-343. Maddison DR. 1991. Discoveq and importance of multiple islands of most-parsimonious trees.
Maddison WP, Maddison DR. 1992. MacClade verswn 3.02. Sunderland, Mass. Sinauer Associates. Markotter EL. 1936. Die lewensgeskiedenis van sekere geslagte van die Ameryllidaceae. Bnna1.c ofthe
[Jniveisip of Stpllmbosch XIV, A2. Morton CM, Mori SA, Prance GT, Karol KG, Chase MW. 1997. Phylogenetic relationships of
Iarythidaceae: a cladistic analysis using rbcL sequences and morphological data. American Journal o j Bohy 84: 530-540.
Nel G. 1914a. Studien uber Amaryllidaceae-Hypoxideae, unter besonderer Reriicksichtigung der afrikanischen Arten. Botanische Jahrbiiclw 51: 234-286.
Nel G. 1914b. Die afrikanischen Arten der Amerayllidaceae-Hypoxideae. Botanische Jahrbuch~r 51:
data support the inclusion of DuckPodendmn cestroidt-5 in Solanaceae. k m Bulletin: in press.
topology. Syskmatic ~ o o l o g ~ 20: 406-4 16.
to its taxonomical position. Phytomorphologp 14: 487-499.
desiccation tolerant leaves. Australian Journal of Botany 24: 209-224.
(hlonocotyltdones). Bulletin de la Sock% Royale a5 Botanique de Bekque 101: 265-278.
.!Vational de Behue 39: 167-1 77.
author. New York.
on the Witwatersrand. Bothalia 14: 889-893.
.hbtesfim the Ryal Botanic Garden, Minbuyh 36: 43-76.
Statssamni. ; M u c h 8 2 19-538.
University Press.
227--386.
plants. Botanical Gazettt 125 38-40.
Systematic ~ o o h g y 40: 3 15-328.
28 7-~~340.

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 35
Pridgeon AM. 1994. Systematic anatomy of Orchidaceae: Resource or anachronism? In: Pridgeon
Rudall PJ. 1991. Leaf anatomy in Tigridieae (Iridaceae). Plant &stmatics and Evolution 175: 1-10, Rudall PJ. 1994. Anatomy and systematics of Iridaceae. Botanical Journal of the Linnean Sociep 114:
Rudall PJ, Furness CAY Chase MW, Fay MF. 1997. Microsporogenesis and pollen sulcus type in
Rudall PJ, Prychid C, Jones C. In press. Intra-ovarian trichomes in monocotyledons. In: SJ
Schmidt C. 1891. Uber den Blatten einiger xerophilen Miifloren. Botanische <entralblatt 47: 1 6 , 33-42,
Schnarf W. 1892. Beitrage zur Anatomie der Hypoxideen und einiger verwandter Pflanzen. Botanische i&tralblatt 52: 145-153, 177-184, 209-217, 241-249, 289-296, 321-327.
Schulze R. 1893. Beitrage zur vergleichenden Anatomie der Ldiaceen, Haemodoraceen, Hy- poxidoideen und Velloziaceen. Botunische Jahrbuchr 17: 295-394.
Schulze W. 1982. Beitrage zur Taxonomie der Wiifloren. IX. Antericaceae. Wzcsenrcha$m <ehchrzz der Friedrich-Schiller- Univer~~t , Mathmatisch-Natumissmchajlliche 31: 29 1-307,
Shah GL, Gopal BV. 1970. Structure and development of stomata on the vegetative and Roral organs of some Amaryllidaceae. Annals $Botany 34: 737-749.
Sheahan MC, Chase MW. 1997. A phylogenetic analysis of Zygophyllaceae R. Br. based on morphological, anatomical, and rbcL DNA sequence data. BotunicalJournal ofthe Linnean Sociep 122: 279-300.
Simpson MG. 1983. Pollen ultrastructure of the Haemodoraceae and its taxonomic significance. Grana 22: 79-103.
Simpson MG. 1985. Pollen ultrastructure of the Tecophilaeaceae. Gana 2 4 77-92. Simpson MG. 1990. Phylogeny and classification of the Haemodoraceae. Annals ofthe Missouri Botanical
Garden 77: 722-784. Simpson MG, Dickison WC. 1981. Comparative anatomy of Lochnanthes and Lophiola (Haemo-
doraceae). Flora 171: 95-133. Skottsberg C. 1934. Studies in the genus Astelia Banks et Solander. Kungl. Svmka Wtenskaps&mim
Handlingar 14: 1-106. Stem WL, Cheadle VI, Thorsch J. 1993. Apostasiads, systematic anatomy, and the origins of
Orchidaceae. BotanicalJournal ofthe Linnean Sociep 111: 41 1-455. Stevenson DW, Loconte H. 1995. Cladistic analysis of monocot families. In: Rudall PJ, Cribb PJ,
Cutler DF, Humphries CJ, eds. Monocopledom: &stmatics and Evolution. London: Royal Botanic Gardens, Kew, 543-578.
Swofford DL. 1993. PAUP: Phylogenetic Anabsis Using Parsimony, Version 3.1.1. Computer Program distributed by the Illinois Natural History Survey, Champaign, Illinois.
Thompson MF. 1976. Studies in the Hypoxidaceae. I. Vegetative morphology and anatomy. Bothalia
Thompson MF. 1978. Studies in the Hypoxidaceae. 11. Floral morphology and anatomy. Bothalia
Thompson MF. 1979. Studies in the Hypoxidaceae. 111. The genus Paudia. Bothalia 12: 621-625. Zavada My Xu XL, Edwards JM. 1983. On the taxonomic status of Lojhiola aurea Ker-Gawler.
AM, ed. Proceedings ofthe 14th World Orchid Confmmce. HMSO: Edinburgh, 84-91.
1-21.
Asparagales (Lilianae). Canadian Journal ofBotany 75: 408430.
Owens, PJ Rudall, eds. Repmductive BioloQ. London: Royal Botanic Gardens, Kew.
97-107, 164-170.
12: 11 1-1 17.
12: 429-435.
Rhodora 85: 73-8 1.
APPENDIX 1
Material examined Hortus K e w m i (HK) refers to specimens in the Kew living collections, followed by the accession
number. Dried herbarium material from the Kew Herbarium is indicated by ‘K, followed by the collector’s name and number. Asterisks refer to microscope slides already in the Jodrell slide collection, sometimes with little or no accession information. No material of Saniella or jl’eoastelia was available, although Hiuiard & Burtt (1978) described the leaf anatomy of Sanielh. This list refers mainly to leaf material, although for some genera rhizomes and roots were also available (species given in descriptions).

3 6 P. J. RUDALL E T A L
We assume that the astelioid group includes the genera of Hypoxidaceae and Asteliaceae that have not yet been cvaluated with rbcL.
Astelia: A. alpina R.Br. var. alpinu (HK: 1992484); A. banhii A. Cunn. (HK: 1986-3620); A. jaguns Colenso (HK: 1989-2690); ,4. gundis Ho0k.f. (HK: 1972-3488); A . mieiesiuna Sm. (HK 1975-1250); A. m o s a Banks & Soland ex Ho0k.f (HK 1958-27302). ~ l a ~ ~ ~ d t a : *B.ftammeu Lindl. (HK: 1957), *B. mag-inata Herb. (HK), *B. nobilis Sm. (RBG, Sydney; HK I954), B. punkra Sweet (HK). Collospennum: C. hastaturn (Colenso) Skottsb. (HK: 1986-3592). Cu.-.l@: C. capitulata Kuntze (HK 1973-13968); *C. latijbliu Dryand. (P.B. Tomlinson, 1959); *C. orchoidoides Gaertn (HK 1956); *C. pilosa (Schum. & Thonn.) Engl. (P.B. Tomlinson, Ghana, 30.3.57); *O'. rrcuntztu Dryand. (HK 1956); *C. zillasa W d . (P.B. Tomlinson, Singapore, 1956). Empodium: E. rlongatum (Nel) B.L. Burtt (K. Hilliard & Burtt 951 1); E. plicatum (K: Salter 502; Goldblatt & Manning s.n., v. 95, Cape). Hypowidin: H. rtiizophyllu (Baker) F. Friedmann (K: Friedmann s.n., Seychelles, 26.i. 1876). H3;bo~iC: H. angustifoha Lam. (HK: 1970-937; H K 1970-2698); H decurnbens Lam. (HK: 1969-6477, 1976-6038); H. h_l,PTometrica Labill. (HK: 1968-57408; & * W.M.C., Tasmania, 1951); H. obtusa Burch (HK: 1973-1989); H. mopen' T. Moore ( H K 1976-406; H. uneolata Nel (HK: 1975-280; & * P.B. Tomlinson, Cameroon, 12.3.57); *H. Lillosa L.f. (HK 1956); H. sp. ( H K 1969-4283). lanuria: L. lunata Druce ( K Salter 4158; MO: Goldblatt 9110). laphzola: L. americuna C o d e (W.M. Whitten, Florida, 29-05-95). ,2iill&znta: M. denszj4ora Ho0k.f. (HK: 1970- 132); X% sylosa F.Muell. ex Benth (HK. 1969-5 1859). ,Wolinmu ,I.% latfoliu Herb ( K de Vogel & Vermeulen 6907). Paudia: F? longitubu M E . Thompson (K: Thompson 95). Rhodo/ypoxix *R. baurii Nel (HK: 1960); R. milloides (Baker) Hilliard & B.L.Burtt (HK: 1980-1 744); *R. 'Stella' (HK: 1960). Spi'pilo.wze: S. aquatzcu (L.f.) Fourc. (K: Garside 4029); S. schlechteri (Bolus) Garside ( K Garside 4855); S. svnatu (Thumb.) Garside (K: Garside 4821); S. stellah Salisb. (K: Garside 4070; Garside s.n. 12.ix.32).
APPENDIX 2
Genus d e s c ~ p ~ ~ o ~ f o ~ lruf unatomp
or root description is gven, no material Has available. Information from the literature is incorporated into the descriptions as indicated. Where no rhizome
LEAF ANATOW: C. capitulata, C. latlfolia, C. orchidoides, C. pilosa, C. recuwatu, C. villosa; other data from Arber (1 922), LQV (1926), Shah & Gopal (1 970). LEAF SURFACE (Figs IE,F,L, 3) EPIDERMIS: cells mainly h ided , not axially elongated, although slightly axially elongated in C. orchidoides, C. pilosu; containing tiny crystals in C. latiiolia (Fig. 3). HAIRS (Fig. lE,F,L): sparse, mainly abaxial; stellate, usually with 1 or 2 unicellular unbranched 'arms' from a multicellular base, except in C. CUpZiUhhZ, in Lvhich hairs with 1- 4 unicellular unbranched arms and C. ziillosu, in which hairs more common (especially abaxially), and with up to c. 8 unicellular unbranched arms radiating from multicellular base. One or more arms often thicker than the others. Arms sometimes spiralling in dried material, e.g. in C. rapitulata. STOMATA: present on abaxial surface only in C. capitulata, C. recuruatn, on both surfaces in C. latfolia, C. orchidaides, C. pilosa, C. uillosa; usually paracytic (although reported as 60% tetracytic in C. orchidoidps by Shah & Gopal, 1970). I.WF .rs (Fig. 4A-C) O I ~ . I U E : more or less plicate, often with a prominent midrib. EPIDERMIS: hulliform cells present on both surfaces in sinuses opposite ridges. r m s o P m L : of varying thickness, from e.g. 14 cell layers in C. pilorn to c. 5 cell layers in C. capitulnta, 3 4 cell layers in C. onhidoides, not differentiated into palisade; enlaraed axially rlongated raphide sacs interspersed (raphide sacs large in e.g. C. latifblia). In some

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 37
species (e.g. C. latijilia, C. recuroata) 1-2 layers of sclerenchyma cells present beneath epidermis at ridges (opposite sinuses). In C. lat@lia, C. remruata, C. villosa small hypodermal strands of sclerenchyma present beneath both surfaces (Fig. 4A,B). MUCILAGE CANALS: absent from leaf blades (though present in petioles and rhizomes of some species). MARGINS: usually with 2-3 layers of hypodermal sclerenchyma, or margin bundles in C. pilosa. VASCULAR BUNDLES: in a single row, with phloem abaxially and xylem adaxially. Main lateral bundles opposite dorsal sinuses; ventral sinuses occur between lateral bundles. BUNDLE SHEATH: outer sheath a single layer of thin-walled parenchymatous cells. Inner sheath: sclerenchyma present at phloem and xylem poles of most bundles, often extending to epidermis at larger bundles. CRYSTALS: raphides present in idioblasts in mesophyll; styloids absent. In C. capitulata, C. latfolia, C. recuruata, small crystals present in epidermal cells. LEAF BASE (PETIOLE)
TS OUTLINE: thick, V-shaped. EPIDERMIS: bulliform cells absent. MESOPHYLL: in C. I.t@lia, C. recuraata, C. villosa small hypodermal strands of sclerenchyma present beneath both surfaces. MUCILAGE CANALS:
present in C. uillosa, interspersed in ground tissue. VASCULAR BUNDLES: in one row in C. orchidoides, otherwise in two or more rows, each with phloem abaxially and xylem adaxially. In larger petioles, i.e. of C. latijilia, C. recuruata, C. uillosa, peripheral row of small bundles present beneath both abaxial and adaxial epidermises, consisting of sclerenchyma and phloem, or sometimes small amount of xylem, often with reverse orientation. RHIZOME: C. latfolia, C. orchidoides, C. pilosa, C. villosa Mucilage canals interspersed throughout parenchyma (Fig. 8A). ROOT: C. latfolia, C. villosa OUTLINE: in cross section usually round or oval. EPIDERMIS: cells even (not alternately long and short). Root hairs sometimes branched. CORTEX: 1-2 (-3) subepidermal cell layers thick walled. Inner cortex consisting of large loosely-packed cells, mainly circular in cross section. ENDODERMIS: 1 cell layer thick.
Empodium
LEAF ANATOMY: E. elongaturn, E. plicatum. Data for E. plicatum also from Thompson (1976).
EPIDERMIS: cells 4-sided, axially elongated. HAIRS: unicellular, unbranched pointed hairs present on leaf ridges (Fig. IC,D). STOMATA: present on both surfaces in intercostal regions; paracytic. LEAF TS (Fig. 7A,B) OUTLINE: plicate. EPIDERMIS: bulliform cells present in E. plicatum, within the folds of the plicate leaf. MESOPHYLL: in 5-7 layers, not differentiated into palisade and spongy tissue. MUCILAGE CANALS: occasional sparse mucilage canals present in E. plicatum, not associated with vascular bundles; not seen in E. elongaturn. MARGINS: lacking sclerenchyma. VASCULAR BUNDLES: in one row, with similar orientation (xylem adaxial), with a large vascular bundle at adaxial ridges. Main lateral bundles opposite dorsal sinuses; ventral sinuses occur between lateral bundles. BUNDLE SHEATH: outer sheath a single layer of thin-walled parenchymatous cells, often incomplete. Inner sheath: sclerenchyma cap present at xylem pole only, or sometimes at both poles of larger bundles, not extending to epidermises as girders. CRYSTALS: raphides present in enlarged idioblasts interspersed in mesophyll.
LEAF SURFACE
Hypoxidia
LEAF ANATOMY: H. rhizophylla
EPIDERMIS: cells slightly axially elongated. HAIRS: present in material examined as broken multicellular hair bases (i.e. probably stellate). STOMATA: mainly abaxial, paracytic.
OUTLINE: plicate, with a prominent midrib (as in Curculigo). EPIDERMIS: bulliform cells present, alternating on abaxial and adaxial surfaces. MESOPHYLL: interspersed with enlarged raphides sacs. MUCILAGE CANALS:
present, scattered in mesophyll. MARGINS: not seen. VASCULAR BUNDLES: in a single row, with phloem abaxially and xylem adaxially. Midrib with a ring of vascular bundles. BUNDLE SHEATH: outer sheath a single layer of thin-walled parenchymatous cells. Inner sheath: sclerenchyma cap at both poles of
LEAF SURFACE
LEAF TS

38 P. J. RUDALL ETilL.
larger bundles. extending to epidermis at larger bundles. Sclerenchyma completely surrounding midrib bundles. CRYSTALS: raphides present in large idioblasts in mesophyll; styloids absent.
Hypoxis
LEAF AMTOMY: H. atgwt@lia, H. akcumbens, H. tygmrnebica, H. obtwa, H. moper', H. urceolata, H uillosa, H. sp. Leaf anatomical data also in Heideman (1 983), Lov (l926), Nel (1 9 l4a, b) LEAF SURFACE (Figs IK, 2A, C, D, G) EPIDERMIS: cells polygonal or slightly axially elongated. HAIRS (Figs IK, 2A, C, D): stellate, consisting of one or more unicellular unbranched 'arms' radiating from a multicellular base; central arm often thicker than others. Usually 1-2 arms present in hairs of H. angustfolia, H. decumbens, 2--3 arms in hairs of H. Tooperi, H. urceolata, H. obtusa, 4-8 arms in hairs of H. uillosa. Unicellular arms varying in length, to long (up to 2-3mm) in H. iillosa; and hairs varying in density from sparse in H. tygmmetrica to relatively dense in H. uillosa; present mainly at margins in H. decumbens. Arms often spiralling in dried material. STOMATA: present on both surfaces, although often slightly more common on abaxial; paracytic (Fig. 2G). Slight striations radiating from stomata on subsidiary cells in H. obtusa. LEAF 1 s (Figs 5-4, 6&C) O~TI.INE: \'-shaped, with prominent midrib and sometimes with prominent main lateral veins making leaf slightly plicate, e.g. in H. angustfaha, H. cillosa. EPIDERMIS: bulliform cells present in H. anpustfolia, mainly on adaxial surface; othenvise generally absent, except on adaxial side of midrib. In H. obtusu epidermal cells relatively thick (I-. 5-14 pm thick, as opposed to c. 1.5-8 pm thick in most species, or c. 3 13 pni thick in H. mopm). YESOPHYLL: of varying thickness, from e.g. 8-10 cell layers in H. roopeti, H. urwolata to c. 4 cell layers in H. aiigustfolia, not differentiated into palisade (except outer 2-3 cell layers slightly palisade in H. obtusa). Enlarged axially elongated raphide sacs interspersed throughout centre of mesophyll, between vascular bundles. MUCILAGE (:ANALS: absent (though present in rhizomes of some species). % i m G I x s : without sclerenchyma. VASCULAR BLNDLES: in one row, with phloem ahaxially and xylem adaxially. BUNDLE SHEATH: outer sheath a single layer of thin-walled parenchymatous cells. Inner sheath: sclerenchyma usually extending to epidermises as girders at larger bundles (except in H. hygromttrica). In H. obtusa, sclerenchyma girders widening in hypodermal region immediately beneath epidermises. CRYSTALS: raphides present in raphide sacs in mesophyll; styloids absent. RHIZOME: H. decumbtns, H. typmetrica, H. sessiljhora, H. urceohta, H. uillosa Mucilage canals interspersed throughout parenchyma (Fig. 8B). ROOT: H. cierunibens, H. hygiometrica, H. urceolata, H. iillosa OUTLINE: in cross section usually round or oval. EPIDERMIS: cells even (not alternately long and short), occasionally containing fungal hyphae. Root hairs sometimes branched. CORTEX: subepidermal cell layer thick walled, but most of outer cortical region usually collapsed (in contractile roots). Inner cortex consisting of large loosely-packed cells, mainly circular in cross section, hut axially elongated. oval in LS; cells becoming smaller towards central vascular region. ENDODERMIS:
1 cell layer thick.
I .E.~F ~LUATOMY: 'W. latfolia
E P I I m a i I s : cell slightly axially elongated. HAIRS: very sparse or absent. STOMATA: present on abaxial surface only, paracytic. LEAF 'rs (Fig. +D) ovi-ux: more or less plicate, with abaxial ridges more prominent than adaxial. EPIDERMIS: bulliform cells present on both surfaces in sinuses opposite ridges. MESOPHYLL: 5-7 cell layers thick; enlarged axially elongated raphide sacs interespersed. At main lateral bundles 1-2 layers of sclerenchyma cells present beneath epidermis at ridges (opposite sinuses). MUCILAGE CANALS: absent. MARGINS: with occasional scattered subepidermal fibres, and thick walled epidermal cells. VASCUIAR BUNDLES: in a single row, with phloem ahaxially and xylem adaxially. Main lateral bundles opposite dorsal sinuses; smaller laterals opposite ventral sinuses. BLNDLE SHEATH: outer sheath a single layer of thin-walled parenchymatous cells. Inner sheath: sclerenchyma present at phloem and xylem poles of most bundles,
LEAF SCTRFACE

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 39
often extending to epidermis at larger bundles. CRYSTALS: raphides present in idioblasts in mesophyll; styloids absent.
Pauridia
LEAF ANATOMY: P longituba. Other data for P longtuba and P minuta from Thompson (1976, 1979).
EPIDERMIS consisting of axially elongated cells. HAIRS: absent. STOMATA; present on both surfaces, paracytic.
OUTLINE: Narrow, V-shaped (with a midrib), or sometimes semi-terete in P minuta. EPIDERMIS: bulliform cells present on the adaxid surface of midrib. MESOPHYLL: sometimes with an adaxial palisade layer. MUCILAGE CANALS: present, associated with vascular bundles at adaxial (xylem) poles. MARGINS: without sclerenchyma. VASCULAR BUNDLES: in a single arc, with phloem abaxially and xylem adaxially. BUNDLE SHEATH: outer sheath a single layer of thin-walled parenchymatous cells. Inner sheath: sclerenchyma generally absent. CRYSTALS: raphides present in idioblasts in mesophyl; styloids absent.
LEAF SURFACE
LEAF TS
LEAF ANATOMY: R. baurii, R. milloides, R. ‘Stella’. Other leaf anatomical data in Hilliard & Burtt (1978). LEAF SURFACE (Fig. 1J EPIDERMIS consisting of slightly axially elongated cells. HAIRS: sparse or absent in R. milloides (except over midrib); in R. baurii more frequent on adaxial surface, especially near margins; stellate, i.e. consisting of one or more arms radiating from multicellular base, central arm thicker than rest. STOMATA: present on abaxial surface only in R. milloides, or on both surfaces in R. baurii, R. ‘Stella’, although more frequent on abaxial surface, paracytic. LEAF TS (Fig. 7C,D) O U T L I ~ : V-shaped. EPIDERMIS: bulliform cells present along almost entire adaxid surface (except over main lateral veins). MESOPHYLL: c. 8 cell layers thick; largely undifferentiated; occasional enlarged, axially elongated raphid sacs present. MUCILAGE CANALS: absent (although present in rhizomes). MARGINS:
without thick-wded cells. VASCULAR BUNDLES: in a single row, with phloem abaxidy and xylem adaxially. BUNDLE SHEATH: outer sheath a single layer of thin-walled parenchymatous cells. Inner sheath: sclerenchyma present at both xylem and phloem poles of larger bundles; at main lateral bundles sclerenchyma extending to abaxial epidermis as girders in R. milloides (not in R. bauriz). CRYSTALS:
raphides present in idioblasts in mesophyll; styloids absent. mizom: R. baurii Mucilage canals interspersed throughout parenchyma.
Spiloxene
LEAF ANATOMY: S. aquatica, S. schlechten’; S. serrata, S. stellata. Data for S. aquatica, S. capensis, S.Jlaccida, S. minuta, S. ouata, S. schlechteri, S. serrata, also from Thompson (1976). LEAF SURFACE (Fig. 1H) EPIDERMIS consisting of axially elongated cells. HAIRS: often absent, but if present (in S. capensis, S. serrata), multicellular and often branched (stellate), either regularly or irregularly (Fig. 1 H). STOMATA:
present on both surfaces, scattered; paracytic. LEAF TS (Fig. 5B,C) OUTLINE: U- or V-shaped (with a midrib) in S. capensis, S. flaccida, S. serrata, S. minutu, S. stellatu; U- shaped (without a midrib) in S. ovata; terete in S. aquatica, S. schlechteri. EPIDERMIS: bulliform cells present in S. minuta, S. serrata. MESOPHYLL: sometimes with an adaxial palisade layer. Aerenchyma (large air spaces) present in S. aquatica, S. serrata, S. stellata. MUCILAGE CANALS: present

40 P.,J. RUDAI,I, E 7 , i L .
in S. capensis, S.Jlaccida, S. ouata, S. schlechteri. S. stnata, absent from leaves of S. aquatica, S. minuta, though present in rhizomes. If present, always associated with vascular bundles at adaxial (xylem) poles, often extending to adaxial epidermis, except in terete leaves (S. aquatica, S. schlechterz). MARGINS: without sclerenchyma, sometimes with thick walled epidermal cells (in S. stellata). VASCULAR BUNDLES: all collateral in a single arc, with phloem adaxially and xylem abaxially (i.e. leaves biracial), except in terete leaves, where in a ring, with xylem towards centre. BVXDLE SHEATH: outer sheath a single layer of thin-walled parenchymatous cells. Inner sheath: small amount of sclerenchyma sometimes present at xylem and phloem poles (not extending to epidermises) in S. stellata, otherwise absent. CRYSTALS: raphides present in idioblasts in mesophyll; styloids absent.
Asklia
LEAF ~ A T O M Y : A. alpina var. alpina, d. bailksii, A . j q a n s , A. grandis, A. menziesiana, A. neniosa. Skottsberg’s 1 1934) comprehensive monogfaph on Astelia and Collospennurn includes some descriptions of the trichomes (following Betts. 1920) and leaf morphology, as well as flower and inflorescence structure. LEAP SI’RU(;TTIRE (Figs IB, 2B, F, H) F . P i r m m i s consisting of polygonal (not elongated) cells, with 4-6 sides. HAIRS: present on both surfaces; xAc-like or stellate, i.e. arising from a multicellular base, long, muticellular (Figs l K , 2H) with marginal cells peeling off to form a matted, untidy ‘wool’ over surface (Fig. 2B). STOMATA: present on abaxial surface only; parac).tic; sometimes slightly sunken (e.g. in A . grandis). Subsidiary cells sometimes with cuticular striations radiating from stomata in -4. j a p a n s , A . menzesiana; in A. fraprans these present only roirrid abaxial stomata, not adaxial (Fig. 2F). IEAF TS (Figs Y&C, 10A,B) OITL.INE: V-shaped, but thickest at middle of hvo sides of lamina. In A. menziesiana, A. neruosa, ridged over tWo prominent lateral bundles (Fig. lO.4j. EPIDERMIS: bulliform cells absent. MESOPHYLL: 6- 16 cell layers thick, not differentiated into palisade; occasional enlarged, axially elongated raphide sacs interspersed in leaf centre (Fig. 9B,C). Adaxial hypodermal layer of 1 --2 cells usually present; hypodermal cells identifiable as slightly larger, thicker-walled or more lignified than others (Fig. 9A). MUCILAGE CUM.S: absent from ‘-1. alpina, d. banhii, d.Jiagrans, A. nenoJa; present in A. grandis, A . menzjesiana (Fig. 1 OB), associatcd with adaxial side (i.e. xylem pole) of middle-range bundles, or occasionally large lateral bundles, sometimes within outer bundle sheath. MARGINS: without sclerenchyma. VASCULAR BUNDLES:
in a single row, lvith phloem abaxially and ?c);lem adaxially. BUNDLE SHEATH: outer sheath a single Iaycr of thin-walled parench!matous cells. Innrr sheath: sclerenchyna present at xylem poles of most bundles, sometimes extending round to phloem pole, and present at both poles of largest bundles; at smallrr bundles sometimes extending to abaxial epidermis; at two main lateral bundles extending to both epidermises as girders (not in A . aijina); in A j agram, A. neruosa, A . menziesiana girders wider immediately within epidermis. C:RYST.ALS: raphides present in idioblasts in mesophyll; styloids usually absent. but present in A. bankrii. in which both bundles of raphides and solitary styloids and also transitional typcs (e.g. a pair of large needle-like crystals) present in enlarged idioblasts in mesophyll. R t m o m : ‘4. a@ina, 11. banksii, .-I. ‘grandis 1,ackiiig mucilagr canals (Fig. 1 IB)? except in leaf bases (Fig. 1 1A). Raphide sacs present, sometimes cnlargcd and numerous (in .4. gandi.7). rm(w: A. akin0 (Fig. I+l3), .i. grand& ’-1. banksii orTL.ISE: in cross section usually round, hut aerial roots of ‘4. gmrfdi,r flattened, oval. EPIDERMIS sometimes thick ~vallecl (in 8. grandis, d. banhi& especially on inner periclinal walls, e.g. in :I. bnnkii. Epidermal cells even (not alternately long and short). Subepidermal layer sometimes with alternately long and short cells, in -4. alpina. ..1. banksii. CORTEX: Groups of cortical cells containing furig-al h!phae present in material of d. alpina. Tannin cells present in inner cortex flattened aerial roo1s of .A. grand& ESDODEILMIS 1- 2 cell layers thick.
LEAF .LU~TOMY: (L hastaturn
E w D E w m consisting of slightly axially elongated cells. IWIKS: present on both surfaces, stcllate, i.e. LLW SC’RPA(X

SYSTEMATICS OF ASTELIACEAE AND HYPOXIDACEAE 41
consisting of several unicellular ‘arms’ from a hi- or multi-cellular base, forming a dense ‘felt’ over both surfaces, as in Astelia. STOMATA: present on both surfaces, although more frequent on abaxial surface; paracytic. LEAF TS (Fig. 9D) OUTLINE: thick, V-shaped. EPIDERMIS: bulliform cells absent, except on adaxial surface of midrib. MESOPHYLL: 9-12 cell layers thick, not differentiated into palisade; 1-2 cell layers of larger hypodermal cells present adaxially, at least in some parts of leaf (Fig. 9D). Occasional enlarged, axially elongated raphide sacs interspersed in leaf centre. MUCILAGE CANALS: present (Fig. 9D), associated with adaxial pole of larger vascular bundles or occasionally large lateral bundles (not midrib). MARGINS: without sclerenchyma. VASCULAR BUNDLES: in a single row, with phloem abaxially and xylem adaxially. BUNDLE
SHEATH: outer sheath a single layer of thin-walled parenchymatous cells. Inner sheath: sclerenchyma present at xylem poles of most bundles, sometimes extending round to phloem pole, and present at both poles of largest bundles; at smaller bundles sometimes extending to abaxial epidermis; at 2 main lateral bundles extending to both epidermises as girders. CRYSTALS: raphides present in idioblasts in mesophyll; styloids absent.
Milligania
LEAF ANATOMY: M. densgora, M. splasa
EPIDERMIS: cells slightly axially elongated or 4-6 sided. Surface covered with fine, mesh-like striations. HAIRS (Fig. 1A): present on abaxial surface and at margins; stellate, i.e. consisting of several long unbranched multicellular ‘arms’ radiating from a multicellular base; forming a dense layer over entire surface. STOMATA: present on both surfaces; paracytic (Fig. IOD). LEAF TS (Fig IOC,E). OUTLINE: V-shaped, with midrib. EPIDERMIS: bulliform cells absent. MESOPHYLL: 10-1 3 cell layers; cells oval or rounded in TS and LS (not differentiated into palisade and spongy), slightly larger in leaf centre (Fig. 1 OC). Occasional enlarged, axially elongated cells (raphide sacs) present in centre. Thick- walled, non-lignified mesophyll cells forming girders between larger vascular bundles and epidermis, and forming a single-layered abaxial hypodermis (Fig. 1 OC,E). Tannin cells interspersed throughout mesophyll. MUCILAGE CANALS: absent. MARGINS: with thick-walled hypodermal cells. VASCULAR BUNDLES:
in a single row, with phloem abaxially and xylem adaxially. BUNDLE SHEATH: outer sheath a single layer of thin-walled parenchymatous cells. Inner sheath: sclerenchyma cap present at both xylem and phloem poles of larger bundles. CRYSTALS: raphides present in large sacs in mesophyll; styloids absent. RHIZOME: M. densgara Lacking mucilage canals. ROOT: M. densgora, M. sglosa (Fig. 14B) OUTLINE: oval in cross section. EPIDERMIS: cells even (not alternatively long and short), containing fungal hyphae. CORTEX: groups of central cortical cells with numerous tiny surface projections. ENDODERMIS
c. 5-6 cell layers thick.
LEAF SURFACE
Blandfordia
LEAF ANATOMY: B. Jammta, B. malginata, B. nobilis. Other data for Blandfardia leaf anatomy in Schulze (1 893). LEAF SURFACE
EPIDERMIS consisting of axially elongated cells, sometimes with thickened rounded end walls. HAIRS: ahsent. STOMATA: present on both surfaces; more or less paracytic; sometimes anomocytic.
OUTLINE: thick, V-shaped. EPIDERMIS: bulliform cells absent. MESOPHYLL: c. 13 cell layers thick; not differentiated into palisade. MUCILAGE CANALS: absent. MARGINS: without sclerenchyma. VASCULAR
BUNDLES: in a single row, with phloem abaxially and xylem adaxially. BUNDLE SHEATH: outer sheath a single layer of thin-walled parenchymatous cells. Inner sheath: sclerenchyma caps at both phloem and xylem poles of all bundles, not extending to epidermises as girders, although outer sheaths sometimes adjacent to adaxial epidermis (especially in B. nobilis), and
LEAF T.S.

+2 P..J. KUDALL E 7 . K
midrib bundle cap sometimes \c-ithin onc cell layer of al~axial epidermis. (:RYSTAI.S: both raphides and styloids abrcnt. RHIZOME: I?. puii icw~
Lacking rrrucilagc canals.
OLTLINE: ov;d in C1'CJS section. EPIDERMIS: crlls ciwi (not alterately lo hlphac. ( :orrm: root contractile, kl-ith regions of collapsed tissue in indi\klual large thicker-\\-allcd cells). ENDODERMIS 1-2 cell layers thick.
ROO?': B. /W/ifPn short), containing fungal cortcx, interspersed with
LEAF .LY.VIOMY: L. kmntcr. Othcr data I'or 1 ~ 1 7 ( i k I d anatomy in Gecrinck (1968), Schulzc (1 893), Schmidt (189 I 1, Schnarf i 1892).
t m m n i i s consisting of pulygonal (not elongated\ c d s with 4-6 siclcs. HAIRS: ahsent from leaves, althoirgh thick ridges present at marpis. and copious dcnsc. highl)- branched (dcndritic) hairs presmt
LEAF S1'RFr\(:b.
ncn and flo\\-ers (Fig. I ?Dj. si-o\i;\T.-\: prcsent on both surfaces; paracytic.
O~TI,ISF,: \7-shapecl. thick (Fig. 124). EPIUEKWS: bulliforni cells present over midrib. Iliick walled, lignificd cclls preseiit o \ r r girders at large-ct ImiNes, in small groups between largest hundlcs, and at margins. ME,SOPHYI.I.: corisistiiig of 3 5 la) of rounded photosynthetic cells beneath lioth surfaces, and larger ~~~irc~icliymatous cells I>et\\ern theni. Xlso groups of largr cells with dark contents (tannins)
('cntrc of Icaf. M ~ ~ : I I A ( ; E (:As..u.s: abscnt. XIARGISS: with thick mdled, lignified epidermal I A R B ~ S D I . E S : largest bundles (ivith girders) alternating with smallcr bundles (without girdcrs),
in ;i hinglc rm\ . l\-ith phloem a1)asiall)- and xylem adasially; two rcws of tiny bundles (randomly orit-ntcd, mainly lacking sclel-rnchymai also present in spaces lict\vccn larger bundles (Fig. 12C:). BI-SDLE s I u T i x outer sheath a single la)-cr of thin-\vallcd parenchymatous cells, extending to both epidermises at larger bunclles. Innrr sheath: sclcrrnch)nia cstending to both epidermises as girders at larger bundles sclcrcnchpa encircling all cscept smallest bundles. i\t largest bundles, sclercnchyma dividing phloem regions into t\\-o strands? or intcrspersed with central part of phloem tissue (Fig. 121)). (:RYSTALS: raphides abscnt; small cuboidal st).loids prescnt in outer bundle sheath cells surrounding sclerench)ina at hrgcr vascular tiundles, and occasionally also just mithin lignified epidermal cells.
laphiah
I.. (iioil(l. Othcr data for Lo/hio/n leaf anatomy in Simpson and Dickison (1981), Schulze i 1892). Since the monot?pic north Xnierican genus Lophiola has often bccn associated leaf anatomy description is included here, although much evidence indicates that it
elong in ;\sparagalcs (see discussioni. and it is therefore not included in the cladistic analysis.
w i i m w i s consisting of asially elongated cells. HAIRS: ahsent. STOMATA: prcsrnt on both surfaces in cquitant distal I Lipper) part o f leaf (ix. adasial surface), lackiiig kom abaxial (sheathing) surface at leaf 1x1s~: ,inoniocytic. LEAF 1's (Fig. 13j OVTLINE: ellipsoidal (leaves cquitant, with bifacial sheathing base). EPIDERMIS: without bulliform cells. \tEsoPHm.L: consisting of 1-2 layers of' photosyiithetic, mainly palisade cells immediately within cpidcrmis, and central region of largcr parenchymatous tissue, cells becoming larger towards centre of Icat: M~Y:IL.-\GE (XVALS: absent. i t . -e iss: lacking sclerenchynia, but marginal epidermal cells thick \valled. VAS(:UL\R BI%~L.ES: in tivo rows in equitant distal (upper) part of leaf, with xylem poles towards centre. B~TDLE SHEATH: outer sheath a single layer of large thin-\vallcd parenchymatous cells, extending to epidermises as girders. Inner sheath: sclerenchynia present at phloem poles, also interspersed in phloem, and dividing phloem region into two or more strands. CRYSTALS: raphides present, though sparse and small; st)-loids absent. RHIZOME: L. nuna rhizome (data from Simpson & Dickison, 198 1). Lacking mucilage canals.
Copyright © 2022 FDOKUMEN




















