
Systematics and biogeography of the Boana albopunctata ...
41
HAL Id: hal-03410768 https://hal.archives-ouvertes.fr/hal-03410768 Submitted on 18 Nov 2021 HAL is a multi-disciplinary open access archive for the deposit and dissemination of sci- entific research documents, whether they are pub- lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers. L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés. Systematics and biogeography of the Boana albopunctata species group (Anura, Hylidae), with the description of two new species from Amazonia Antoine Fouquet, Pedro Marinho, Alexandre Réjaud, Thiago Carvalho, Marcel Caminer, Martin Jansen, Raíssa Rainha, Miguel Rodrigues, Fernanda Werneck, Albertina Lima, et al. To cite this version: Antoine Fouquet, Pedro Marinho, Alexandre Réjaud, Thiago Carvalho, Marcel Caminer, et al.. Systematics and biogeography of the Boana albopunctata species group (Anura, Hylidae), with the description of two new species from Amazonia. Systematics and Biodiversity, Tay- lor & Francis: STM, Behavioural Science and Public Health Titles, 2021, 19 (4), pp.375-399. 10.1080/14772000.2021.1873869. hal-03410768
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Systematics and biogeography of the Boana albopunctata ...

HAL Id hal-03410768httpshalarchives-ouvertesfrhal-03410768
Submitted on 18 Nov 2021
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents whether they are pub-lished or not The documents may come fromteaching and research institutions in France orabroad or from public or private research centers
Lrsquoarchive ouverte pluridisciplinaire HAL estdestineacutee au deacutepocirct et agrave la diffusion de documentsscientifiques de niveau recherche publieacutes ou noneacutemanant des eacutetablissements drsquoenseignement et derecherche franccedilais ou eacutetrangers des laboratoirespublics ou priveacutes
Systematics and biogeography of the Boanaalbopunctata species group (Anura Hylidae) with the
description of two new species from AmazoniaAntoine Fouquet Pedro Marinho Alexandre Reacutejaud Thiago Carvalho
Marcel Caminer Martin Jansen Raiacutessa Rainha Miguel Rodrigues FernandaWerneck Albertina Lima et al
To cite this versionAntoine Fouquet Pedro Marinho Alexandre Reacutejaud Thiago Carvalho Marcel Caminer etal Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae)with the description of two new species from Amazonia Systematics and Biodiversity Tay-lor amp Francis STM Behavioural Science and Public Health Titles 2021 19 (4) pp375-3991010801477200020211873869 hal-03410768
PROOF ONLY
TSAB1873869 VOL 0 ISS 0
Systematics and biogeography of the Boana albopunctataspecies group (Anura Hylidae) with the description
of two new species from Amazonia
Antoine Fouquet Pedro Marinho Alexandre Rejaud Thiago R Carvalho Marcel A CaminerMartin Jansen Raıssa N Rainha Miguel T Rodrigues Fernanda P Werneck Albertina P Lima
Tomas Hrbek Ariovaldo A Giaretta Pablo J Venegas German Chavez and Santiago Ron
QUERY SHEET
This page lists questions we have about your paper The numbers displayed at left are hyperlinked to the location of thequery in your paper
The title and author names are listed on this sheet as they will be published both on your paper and on the Table ofContents Please review and ensure the information is correct and advise us if any changes need to be made In additionplease review your paper as a whole for typographical and essential corrections
Your PDF proof has been enabled so that you can comment on the proof directly using Adobe Acrobat For furtherinformation on marking corrections using Acrobat please visit httpjournalauthorstandfcoukproductionacrobatasphttpsauthorservicestaylorandfranciscomhow-to-correct-proofs-with-adobe
The CrossRef database (wwwcrossreforg) has been used to validate the references
AUTHOR QUERIES
Q1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak ampHo 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila-Pires et al 2010 and Avila ampKawashita -Ribeiro 2011) in the reference list or delete the citation from the text
Q2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Geurounther 1858Geurounther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please inserta citation in the text or delete the reference as appropriate
Q3 Please provide the volume number and page range
Q4 Please provide the volume number and page range
Q5 The year of publication has been changed as per Crossref details both in the list and in the text forthis reference Please check
Q6 The year of publication has been changed as per Crossref details both in the list and in the text forthis reference Please check
PROOF ONLY
Q7 If your paper introduces new zoological taxa at family-group level or below you are now required toregister your paper with ZooBank and insert the generated ZooBank ID (LSID) here Individual newtaxa need not be registered before publication this can be done subsequently should you wish Pleasego to httpwwwzoobankorgregister to complete this task You will need your article DOI to registerAfter publication you must amend your ZooBank record of your paper to reflect the date of publica-tion Please see httpwwwzoobankorghelp for further information
PROOF ONLY
Research Article
Systematics and biogeography of the Boana albopunctataspecies group (Anura Hylidae) with the descriptionof two new species from Amazonia
ANTOINE FOUQUET1 PEDRO MARINHO2 ALEXANDRE REacuteJAUD1 THIAGO R CARVALHO3 MARCEL A CAMINER45 MARTIN JANSEN6 RAIacuteSSA N RAINHA7 MIGUEL T RODRIGUES8 FERNANDA P WERNECK7 ALBERTINA P LIMA7 TOMAS HRBEK9 ARIOVALDO A GIARETTA2 PABLO J VENEGAS10 GERMAacuteN CHAacuteVEZ10 amp SANTIAGO RON4
1Laboratoire Evolution et Diversite Biologique UMR 5174 CNRS IRD Universite Paul Sabatier Batiment 4R1 31062cedex 9 118 Route de Narbonne Toulouse 31077 France2Laboratorio de Anuros Neotropicais Instituto de Ciencias Exatas e Naturais do Pontal Universidade Federal de UberlandiaItuiutaba MG Brazil3Laboratorio de Herpetologia Departamento de Biodiversidade e Centro de Aquicultura IB Universidade Estadual PaulistaRio Claro SP Brazil4Museo de Zoologıa Escuela de Biologıa Pontificia Universidad Catolica del Ecuador Quito Ecuador5Institute of Organismic and Molecular Evolution Johannes Gutenberg University Mainz Germany6Department of Terrestrial Zoology Research Institute and Nature Museum Senckenberg Frankfurt Germany7Instituto Nacional de Pesquisas da Amazonia Coordenac~ao de Biodiversidade Manaus AM Brazil8Departamento de Zoologia Universidade de S~ao Paulo Instituto de Biociencias S~ao Paulo SP Brazil9Departamento de Genetica Universidade Federal do Amazonas Manaus AM Brazil10Instituto Peruano de Herpetologıa Lima Peru
The outstanding species richness of Amazonia has fascinated biologists for centuries However the records of actualnumbers and distribution of species forming its ecosystems are so incomplete that the understanding of the historicalcauses and regional determinants of this diversity remain speculative Anuran clades have repeatedly been documentedto harbour many unnamed species in this region notably the Boana albopunctata species group Considering thedocumented distribution and the ecology of the species of that group we hypothesized that it diversified via successivetrans-riverine dispersals during the late Miocene and Pliocene after the formation of the modern Amazon watershedTo test this hypothesis we gathered an extensive dataset of 16S rDNA sequences sampled throughout Amazonia anda mitogenomic dataset representative of the diversity of the clade to (1) re-evaluate species boundaries and distributionsand (2) infer the spatio-temporal history of diversification within Amazonia We delimited 14 Operational TaxonomicUnits (OTUs) in an Amazonian clade ie 75 higher than currently recognized (14 OTUs for eight described species)Combining molecular data with morphological and acoustic data two new species Boana courtoisae sp nov fromthe eastern Guiana Shield and Boana eucharis sp nov from Southern Amazonia are described herein These speciesbelong to a clade that diversified throughout Amazonia during the last 10Ma thus more recently than co-distributedsmall terrestrial anurans but concomitantly with other more vagile vertebrates Our time-scaled phylogeny andbiogeographic analyses suggest an initial east-west divergence and confirm reciprocal trans-riverine dispersals duringthe last 5Ma The geomorphological evolution of the region and species-specific dispersal ability largely explain thesedistinct spatio-temporal patterns across anuransQ7
Key words Amphibia biodiversity Guiana Shield mitogenomics neotropics phylogenetics
IntroductionThe Neotropics harbour the most diverse ecosystems onthe planet (Jenkins et al 2013) Within the Neotropics
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253
54555657585960616263646566676869707172737475767778798081828384858687888990919293949596979899100101102103104105106
Correspondence to Antoine Fouquet E-mailfouquetantoinegmailcom Pedro Marinho E-mailpmarinho50gmailcom
ISSN 1477-2000 print 1478-0933 online The Trustees of the Natural History Museum London 2021 All Rights Reservedhttpsdxdoiorg1010801477200020211873869
Systematics and Biodiversity (2021) 0(0) 1ndash38
Author Query
AQ7 If your paper introduces new zoological taxa at family‐group level or below you are now required to register your paper with ZooBank and insert the generated ZooBank ID (LSID) here Individual new taxa need not be registered before publication this can be done subsequently should you wish Please go to httpwwwzoobankorgregister to complete this task You will need your article DOI to register After publication you must amend your ZooBank record of your paper to reflect the date of publication Please see httpwwwzoobankorghelp for further information
antoinefouquet
Note
Boana courtoisaezoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBEBoana euchariszoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
PROOF ONLY
Amazonia stands as a major biogeographic region cover-ing more than 65 million km2 including the largestcontinuous tract of tropical forest (40 of the worldrsquostropical rainforests) encompassing the most importantdrainage (Amazon River basin) and probably hostingthe highest continental biodiversity on earth (Myerset al 2000) The outstanding species richness of thisregion has raised many questions concerning its originsand diversification mechanisms While all agree on theastounding diversity of Amazonia our understanding ofthe actual number and distribution of species forming itsecosystems is in fact so incomplete (Ficetola et al2014 Meyer et al 2015 Vacher et al 2020) that ithas so far hampered investigations at the regional scaleConsequently the understanding of the historical causesand regional determinants of this outstanding diversityremain speculative (Antonelli et al 2018) Another con-sequence is that the collision between the Linnean short-fall and the biodiversity crisis is particularly evident inAmazonia (Guerra et al 2020) With almost the entireeastern and southern parts (25) of its extent beingalready deforested for agriculture an unknown propor-tion of endemic species and thus of the testimony ofAmazonian history has already vanished (Lovejoy ampNobre 2019)Anuran clades have repeatedly been documented to
harbour many unnamed species in Amazonia (Vacheret al 2020) and to display strikingly allopatric distribu-tion patterns that could provide crucial insights intoAmazoniarsquos past (Fouquet Cassini Haddad Pech ampRodrigues 2014 Rejaud et al 2020) This is notablythe case of the Boana albopunctata species group(Caminer amp Ron 2014 Funk et al 2012) This groupis only defined on the basis of molecular data(Faivovich et al 2005) and currently comprises 16valid nominal species distributed in the Caribbean andSouth America Boana albopunctata (Spix 1824)Boana alfaroi (Caminer amp Ron 2014) Boana almen-darizae (Caminer amp Ron 2014) Boana caiapo Pinheiroet al 2018 Boana calcarata (Troschel 1848) Boanadentei (Bokermann 1967) Boana fasciata (Geurounther1858) Boana heilprini (Noble 1923) Boana lancifor-mis (Cope 1871) Boana leucocheila (Caramaschi amp deNiemeyer 2003) Boana maculateralis (Caminer ampRon 2014) Boana multifasciata (Geurounther 1859)Boana paranaiba (Carvalho Giaretta amp Facure 2010)Boana steinbachi (Boulenger 1905) Boana tetete(Caminer amp Ron 2014) and Boana raniceps (Cope1862) Within this clade B heilprini is the most distinctsince it is confined to Hispaniola and is phylogeneticallydistant from a clade formed by all the other species ofthe group (Duellman Marion amp Hedges 2016) Wecan also distinguish B raniceps and the B albopunctata
clade (B lanciformis B albopunctata B multifasciataB paranaiba B caiapo B leucocheila) which occurwidely in the open habitats of the Cerrado andAmazonia (Camurugi et al 2021) from the Amazonianclade According to Caminer and Ron (2014) thisAmazonian clade is itself subdivided in two maingroups hereafter called the B calcarata (B fasciata Bcalcarata B almendarizae B maculateralis) and the Bsteinbachi (B tetete B alfaroi B steinbachi) cladesWithin this Amazonian clade Funk Caminer and
Ron (2012) identified no less than seven putativeunnamed species based on mitochondrial divergence andacoustic data These species add up to the 10 taxa thatwere valid at that time Caminer and Ron (2014) com-pleted this picture by documenting two additional line-ages describing four of the previously discoveredspecies and removing B steinbachi from its synonymywith B fasciata However sampling of both studies wascircumscribed to western Amazonia (mostly Ecuador)while the species complex occurs throughout AmazoniaPopulations from the eastern Guiana Shield lowlandsare already assumed to belong to a yet unnamed species(Caminer amp Ron 2014) but the status of numerousother populations throughout Amazonia generally iden-tified as B fasciata remain virtually unknown Morerecently Vacher et al (2020) suggested based on 16Smitochondrial DNA (mtDNA) sequences that up to 22species could exist in that group and that the distributionof most species could be circumscribed to small rangeswithin AmazoniaThese aforementioned studies improved our under-
standing of the actual diversity in the B calcaratastein-bachi clade but left virtually unexplored the temporaland spatial context of the diversification withinAmazonia The crown age of this Amazonian clade wasestimated between 16ndash11 million years ago (Ma) byFunk et al (2012) and between 14ndash9Ma by Duellmanet al (2016) Therefore the initial diversification of thatclade throughout Amazonia may have taken place dur-ing the final stage of the Pebas system a freshwaterlake occupying most of Western Amazonia from theearly Miocene (23Ma) until 10ndash9Ma (Hoorn et al2017) Meanwhile most of the diversification within theB calcaratasteinbachi clade took place during the lateNeogene (last 10Ma) From 9Ma onward this systemhas drained eastward into the Atlantic Ocean but enor-mous flooded ecosystems (Acre system) still occupied alarge portion of what we currently consider as WesternAmazonia until 7Ma (Hoorn et al 2010 Albert et al2018a) Subsequent river captures were common(Ruokolainen Moulatlet Zuquim Hoorn amp Tuomisto2018) as were climatic fluctuations that may havemodified vegetation possibly to the point of forest
107108109110111112113114115116117118119120121122123124125126127128129130131132133134135136137138139140141142143144145146147148149150151152153154155156157158159160
161162163164165166167168169170171172173174175176177178179180181182183184185186187188189190191192193194195196197198199200201202203204205206207208209210211212213214
2 A Fouquet et al
antoinefouquet
Texte inseacutereacute
mostly
PROOF ONLY
fragmentation at least in some peripheral parts ofAmazonia (Cheng et al 2013 Kirschner amp Hoorn2019) The relative roles of hydrological evolution(Albert et al 2018b Ribas Aleixo Nogueira Miyakiamp Cracraft 2012) and rainforest expansion and contrac-tion due to climate oscillations (Haffer 1969) have beenthe focus of intense debate (Leite amp Rogers 2013)Since the species of the B calcarata and B steinbachicomplexes are associated with riparian forests and thesespecies mostly breed in pools formed on the banks ofthe beds of small to medium-sized rivers very large riv-ers may act as barriers to dispersal thus reducing orcompletely impeding gene flow and inhibiting homogen-ization of differentiating populations (Naka et al 2018)However smaller and highly dynamic meandering riversprobably represent more permeable barriers Thereforewe assume that successive dispersals across majorAmazonian rivers (parapatric speciation) (Pirani et al2019) as well as hydrological changes (vicariant speci-ation) (Naka et al 2018) could have been major proc-esses of diversification in these frogsWe gathered a large mtDNA dataset for the B albo-
punctata group sampled throughout Amazonia combinedwith morphological and acoustic data for a subset ofthis group (the B steinbachi clade) to address two maingoals (1) reevaluate species boundaries (in or amongmembers) of the B albopunctata group and theirrespective distributions and (2) investigate its diversifi-cation history within Amazonia using mitogenomic datafor one terminal of each delimited species Furthermoretwo species in the B steinbachi clade that wereunnamed are described herein
Materials and methodsSpecies delimitationOur first objective was to delimit all major OperationalTaxonomic Units (OTUs) based on mtDNA main line-ages Our sampling included 55 new Boana tissue sam-ples obtained through fieldwork throughout Amazoniaand adjacent Dry Diagonal (DD Chaco CerradoCaatinga Werneck 2011) and loans from collaborators(from collections QCAZ MPEG INPA CORBIDIAAGUFU and from personal loans JMP SCF FTAAppendix 1) Samples unambiguously identified (neartype locality andor phenotypically corresponding to typematerial) as belonging to the currently recognized taxawere included except B paranaiba and B caiapo Wesequenced a 400 bp portion of the end of the 16S rDNAgene a locus commonly used for Neotropical amphibiantaxonomy and systematics (Vences Thomas Bonett ampVieites 2005) We also retrieved homologous sequences
from GenBank (389 accessions) In total we gathered444 16S sequences for this study (dataset details are pro-vided in Appendix 1 and DNA extraction and sequencingprotocols can be found in Appendix 2) These samplescover the whole distribution of the B albopunctata spe-cies group (14 included taxa) DNA sequence alignmentwas conducted on the MAFFT7 online server under theE-INS-i option with default parameters an algorithmdesigned for sequences with multiple conserved domainsand long gaps (Katoh et al 2019)We applied three DNA-based single-locus species
delimitation approaches (a) a distance-based methodthe Automated Barcode Gap Discovery (ABGDPuillandre Lambert Brouillet amp Achaz 2012) (b) amulti-rate coalescent based method the multi-ratePoisson Tree Processes model approach (mPTP Kapliet al 2017) and (c) a single-threshold coalescent-basedmethod the Generalized Mixed Yule Coalescentapproach (single threshold GMYC Monaghan et al2009 Pons et al 2006)The ABGD delimitation was performed with a prior
of intraspecific divergence P between 0001 and 01 aproxy for minimum relative gap width X of 1 and anumber of steps n equal to 30 We kept the partitionsuch that Pfrac14 0016 as it corresponds to the end of aplateau for group number and it matches thresholds ofintraspecific divergence proposed in other vertebratedelimitation studies using 16S barcodes (Puillandreet al 2012) For mPTP delimitation we first recon-structed a Maximum Likelihood (ML) tree withRAxML v824 (Stamatakis 2014) using theGTRthornGthorn I substitution model and estimated nodalsupport via 1000 parametric bootstraps We used nineoutgroups representing most other Boana species groups(Appendix 3) The mPTP delimitation was undertakenon the rooted ML tree with 5 million Markov chainMonte Carlo (MCMC) iterations sampling every10000th iteration and a 10 burn-in For the GMYCdelimitation we reconstructed a time-calibrated phyl-ogeny using BEAST 25 (Bouckaert et al 2014) Weused a birth-death population model to account forextinction processes and incomplete sampling Weincluded the same nine Boana outgroups used inRAxML reconstruction and five additional representa-tives of other Cophomantini genera (NesorohylaMyersiohyla Hyloscirtus AplastodiscusBokermannohyla) in order to include relationships thatcan be time-calibrated We used a single partition with aGTRthornGthorn I substitution model with an uncorrelatedrelaxed lognormal clock model of rate variation amongbranches (Drummond Ho Phillips amp Rambaut 2006)We used two calibration points the ages of the mostrecent common ancestor (MRCA) of Aplastodiscus and
215216217218219220221222223224225226227228229230231232233234235236237238239240241242243244245246247248249250251252253254255256257258259260261262263264265266267268
269270271272273274275276277278279280281282283284285286287288289290291292293294295296297298299300301302303304305306307308309310311312313314315316317318319320321322
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 3
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13536302v1
PROOF ONLY
Boana as well as the MRCA of a clade formed byHyloscirtus Boana Aplastodiscus andBokermannohyla which were estimated by Feng et al(2017) and consequently constrained here assuming nor-mal prior distributions of 252Ma (standard deviation[SD]frac14 28Ma) and 323Ma (SD frac14 3Ma) respectivelyWe considered these calibrations to represent the bestprior because Feng et al (2017) included a comprehen-sive dataset of nuclear loci for an extensive sampling ofanurans and used many reliable fossils as calibrationpoints Moreover Hime et al (2020) analysing an evenlarger genomic dataset found very similar time esti-mates for the TMRCA of BoanathornHyloscirtus Insteadusing fossil andor biogeographic calibrations wouldhave implied to expand the matrix to lineages distantlyrelated to Boana and would likely lead to an overesti-mation of calibration dates with our mitogenomic data-set (Molak amp Ho 2015 Papadopoulou et al 2010) Forthe MCMC parameters we used four independentchains of 100 million iterations recording every10000th iteration and a 10 burn-in We combined thelog files of the independent runs using LogCombiner25 (Drummond amp Rambaut 2007) and checked theconvergence of our parameters confirmed by allEffective Sample Size (ESS) being above 200 Then weextracted the maximum clade credibility tree using Treeannotator 25 (Drummond amp Rambaut 2007) with aburn-in of 10 After removing outgroups we per-formed a GMYC delimitation on the ultrametric treeusing the GMYC function of the splits R package v10-11 (Ezard Fujisawa amp Barraclough 2009) with athreshold interval between 0 and 10Ma and by usingthe single threshold method Operational TaxonomicUnits (OTUs) were defined using a majority-rule con-sensus from the results of the three methods ie a lin-eage is considered as being an OTU if supported by atleast two of the three methodsAll but five occurrence records were georeferenced
and used to create distribution maps with convex poly-gons under QGIS 214 (QGIS Geographic InformationSystem) OTUs were assigned to taxa based on thefieldmuseum identification and sometimes corrected inaccordance with type localities and known distribution(Frost 2019 IUCN 2020)
Morphological and acoustic variation in theBoana steinbachi cladeThe following morphometric measurements were takenon 38 males and 9 females snout-vent length (SVL)head length (HL) head width (HW) eye diameter (ED)tympanum diameter (TD) tibia length (TL) foot length(FL) thigh length (THL) and calcaneal appendage
length (CL) In addition to these we also measuredhand length (HAL) forearm length (FLL) and eyendashnos-tril distance (EN) for the morphometric characterizationof holotypes All measurements followed the definitionsand terminology of Watters Cummings Flanagan andSiler (2016) We classified the calcaneal appendage intothree-character states (i) a calcar (skin appendage withlength gt 1mm flattened or conical) a (ii) skin flap(skin appendage with length lt 1mm flattened or a(iii) tubercle a conical structure with reduced size andas such not forming an appendage coming out of theheel The specimens were measured by P Marinho(Alta Floresta Mato Grosso State and Assis BrazilAcre State) using a Mitutoyo digital calliper (to thenearest 005mm) by MT Rodrigues (Jirau and PacaasNovos Rondonia State) using a Mitutoyo digital calliper(to the nearest 001mm) by A Fouquet (French Guianaand Suriname) using a DigiMax digital calliper (nearest001mm) and by M Caminer (Ecuador Peru andBolivia) using a Mannesmann digital calliper (nearest001mm) Snout shape was assessed according to Heyeret al (1990) Digital webbing formulae followed thenotation system of Savage and Heyer (1997)Acoustic analysis of 27 recorded calling males
(Appendix 4) was conducted in Raven Pro 15 64-bit ver-sion (Bioacoustics Research Program 2012) sound figureswere produced using Seewave version 210 (SueurAubin amp Simonis 2008) and tuneR version 132(Ligges Krey Mersmann amp Schnackenberg 2014) in Rversion 350 (R Core Team 2018) Raven Pro settingswindow size frac14 512 samples window typefrac14Hann 3 dBfilter bandwidth frac14 124Hz window overlap frac14 85 hopsize frac14 77ms discrete Fourier transform (DFT) size frac141024 samples grid spacing frac14 431Hz seewave settingswindow typefrac14Hanning fast Fourier transform (FFT)size frac14 256 samples FFT overlap frac14 90 The analysedfiles are deposited in the sound collection of the lsquoMuseude Biodiversidade do Cerradorsquo (AAG-UFU) lsquoFonotecaNeotropical Jacques Vielardirsquo (FNJV) lsquoLa Sonotheque duMuseum National dHistoire Naturellersquo (MNHN) and theMacaulay library (MC) Detailed information about theanalysed sound files and accession numbers are providedin Appendix 4Call types were distinguished based on the number of
notes per call and pulsing We defined as type 1 callsthe multinote calls formed by nonpulsed notes whichconsist of the main call type emitted by males of allstudied species Type 2 calls are emitted less often thantype 1 and consist of a one-note call formed by poorlydefined pulses We only included type 1 calls in theacoustic diagnoses and the interspecific comparisonsThe acoustic traits were generally analysed manually (ifnot stated otherwise) as follows temporal traits (call
323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370371372373374375376
377378379380381382383384385386387388389390391392393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430
4 A Fouquet et al
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
1
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics 2014
antoinefouquet
Texte inseacutereacute
a
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
s
PROOF ONLY
duration note duration interval between notes numberof notes and call rise time the latter using the lsquoPeakTimersquo function) frequency traits (dominant frequencyusing the lsquoPeak Frequencyrsquo function minimum andmaximum frequency using the lsquoFrequency 5rsquo andlsquoFrequency 95rsquo functions respectively) Call traitsused in the description followed the definitions and ter-minology of Keuroohler et al (2017) using a note-cen-tred approach
Time-calibrated species phylogenyWe selected a representative for each of the 25 OTUsidentified by the species delimitation (see Results) forcomplete mitogenome sequencing to investigate interre-lationships and divergence times Mitogenomic sequen-ces were obtained using low-coverage shotgunsequencing We recovered high-quality mitochondrialgenome assemblies for 18 OTU representatives (seeAppendix 2 for details regarding mitogenome sequenc-ing assembling and annotation) For the remainingseven OTUs for which we could not obtain tissue sam-ples (Appendix 3) we gathered all the available mito-chondrial loci (12S 16S ND1 COI and Cytb) fromGenBank four of these OTUs were represented by 16Sonly We also selected 14 species as outgroups (ninewith complete mitogenomes) including representativesfor other Boana species groups and most otherCophomantini genera (Appendix 3)We extracted 12S 16S and all protein-coding
sequence regions (CDS thus removing D-Loop andtRNAs) from complete mitogenomes as well asGenBank accessions and aligned each locus independ-ently using the MAFFT7 online server For rRNAgenes we chose the E-INS-i strategy recommended forsequences with multiple conserved domains and longgaps For the protein-coding genes we chose the G-INS-i strategy designed for sequences with global hom-ology (Katoh et al 2019) Realignments of CDS con-sidering the reading frame were done and concatenatedin Geneious v918 (httpswwwgeneiouscom) Ourfinal matrix totalled 39 terminals and 13620 alignednucleotide sites Among these terminals 27 were com-plete and nine had fewer than 3000 nucleotides (20)but all were putatively closely related to terminals withcomplete dataWe selected the best-fit partition scheme and model
of evolution for each partition using PartitionFinderV211 (Lanfear et al 2016) based on the BayesianInformation Criterion (BIC) We predefined four datablocks one for rRNA genes (12S and 16S) and one foreach codon position of all CDS regions and found theGTRthorn IthornG model to best-fit all partitions We
reconstructed a time-calibrated tree using a birth-deathtree prior using BEAST 25 to account for extinctionprocesses and incomplete sampling We parameterizedunlinked substitution models according to the estimatesobtained in the PartitionFinder V211 analysis (Lanfearet al 2016) We used the same two calibration pointsas in the previous BEAST analysis Analyses wereundertaken using uncorrelated relaxed lognormal clockmodel of distribution of rate variation among branchesfor each partition (Drummond et al 2006) The Markovchain Monte Carlo (MCMC) parameters were set withfour independent chains of 100 million iterations stor-ing every 10000th iteration and a 10 burn-in Wecombined the log files of the independent runs usingLogCombiner 25 and confirmed the convergence of ourparameters as all ESS were above 200 Finally weextracted the maximum clade credibility tree using Treeannotator 25 We acknowledge that our phylogeneticreconstruction solely based on mtDNA sequences priori-tizes spatial and taxonomic completeness over genomiccoverage and can lead to overestimated divergence times(eg McCormack Heled Delaney Peterson ampKnowles 2011 Near et al 2012)
Biogeographic analysisBiogeographic inferences were undertaken on the time-calibrated phylogeny using the BioGeoBEARS R pack-age (Matzke 2013) This package reconstructs ancestralgeographic distributions and investigates the role ofeach biogeographic event with a maximum likelihoodalgorithm We compared three different models (i) alikelihood version of the Dispersal-Vicariance(DIVALIKE) model (Ronquist 1997) (ii) a likelihoodversion of the BayArea (BBM) model (Landis et al2013) and (iii) the Dispersal Extinction Cladogenesis(DEC) model (Ree amp Smith 2008) We also comparedversions of these models allowing jump dispersal asdescribed by the J parameter (Matzke 2013) Modelswere compared with the Akaike Information Criterion(AIC) We ran 50 independent BioGeoBEARS biogeo-graphic stochastic mapping to determine biogeographicevent counts for the best-fit model (Dupin et al 2017)Ree and Sanmartın (2018) voiced several criticismsagainst the use of J parameter arguing that it inflates thecontribution of cladogenetic events to the likelihoodand minimizes the contribution of anagenetic time-dependent range evolution We thus also compared theresults from the best model without jump dispersalSince this work focuses on the historical biogeog-
raphy of the Amazonian clade we discarded the twoearly diverging lineages B heilprini and B ranicepsfrom the biogeographic analyses since their distribution
431432433434435436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484
485486487488489490491492493494495496497498499500501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 5
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13560371v1)
antoinefouquet
Texte inseacutereacute
D
PROOF ONLY
ranges lie mostly outside of Amazonia We consideredthree areas within Amazonia Western Amazonia theBrazilian Shield and the Guiana Shield These areascorrespond to major geological features roughly delim-ited by modern riverine barriers the Madeira River theNegro River and the lower course of the Amazon Riverand to three large biogeographic regions known asWallacersquos districts (Hoorn et al 2010 Wallace 1854)These districts were recently confirmed as major breaksin birdsrsquo species composition (Oliveira Vasconcelos ampSantos 2017) and amphibians (Godinho amp da Silva2018 Vacher et al 2020) strengthening their status ofbiogeographic regions Because the distribution range ofthe B albopunctata group extends outside Amazoniawe included in the ancestral range reconstruction oneadditional non-Amazonian Neotropical area theDry Diagonal
ResultsSpecies delimitationOf the three tested methods of species delimitation theABGD method was found to be the most conservativedelimiting 21 OTUs while mPTP and GMYC delimited24 and 47 OTUs respectively (Appendix 1) In onecase the consensual partitioning was overly conserva-tive given that B leucocheila and B multifasciata wereconsidered as a single OTU which is contradicted bythe fact that these two taxa are phenotypically distinctTherefore we kept the different lineages delimited byGMYC in this group as distinct OTUs The final delimi-tation led to 25 OTUs (including B heilprini) in the Balbopunctata species group (Fig 1 Appendix 1)Thirteen of these OTUs could be linked to nominal spe-cies (except B caiapo and B paranaiba that were notincluded in the analysis because material was not avail-able) Conversely 11 could not be linked to any nom-inal species representing a possible 44 increase in thespecies richness of the group The Amazonian cladeitself was represented by 14 OTUs including eight nom-inal species and six putative new species (Fig 1) Theseresults also imply important changes to the geographicdistribution of the species of the group In Appendix 5we detail and justify the identification of these OTUsand their respective geographic ranges
Taxonomic accountsThe Amazonian clade of the B albopunctata group pre-viously reported as the B calcaratandashB fasciata speciescomplex by Caminer and Ron (2014) is composed oftwo major clades (the B calcarata and the B steinbachi
clades) defined by the following combination of charac-ter states (1) truncate snout in dorsal view (2) roundedhead shape and (3) presence of calcaneal appendageon heelThe B calcarata clade (Fig 1) is composed of four
species possessing a calcar and having advertisementcalls formed by a single type of call (Caminer amp Ron2014) The B steinbachi clade on the other handincludes three described and two undescribed species(Fig 1) Members of this clade lack a calcar (skin flapor tubercle present or the complete absence of calcanealappendages) and have vocal repertoires composed ofmore than one type of call except for B alfaroi forwhich a single type of call has been documented so far(Caminer amp Ron 2014)Hyla steinbachi Boulenger 1905 currently valid as
Boana steinbachi (Boulenger 1905) was describedfrom the Bolivian Province Sara Departament SantaCruz de la Sierra Bolivia We assume that Buenavistais the type locality of B steinbachi Between 1910 and1950 the Steinbach family collected many amphibiansand reptiles in Bolivia mainly at Buenavista in theDepartment of Santa Cruz (see Parker 1927) which isthe type locality of several anurans (Hamptophryneboliviana (Parker 1927) Pseudopaludicola bolivianaParker 1927 Scinax parkeri (Gaige 1929)) and thesnake Apostolepis tenuis Ruthven 1927 The materialcollected by the Steinbach family is on display in vari-ous museums such as the University of MichiganMuseum of Zoology (UMMZ see Gaige 1929) theNatural History Museum London (BMNH see Parker1927) the Zoologisches Museum Berlin (ZMB seeMeurouller 1924) and the Naturhistorisches Museum Basel(NBM pers comm Weurouest) Many decades later De laRiva (1990) synonymized H steinbachi with H fas-ciata Subsequently Jansen Bloch Schulze andPfenninger (2011) found differences in call and mtDNAdata between populations of B fasciata from Boliviaand Ecuador and suggested that their results imply a res-urrection of B steinbachi In the following yearsCaminer and Ron (2014) tentatively assigned B steinba-chi to a genetic lineage (clade J) inhabiting BolivianAmazonia near its type locality By assessing its phylo-genetic relationships in the group Caminer and Ron(2014) formally revalidated B steinbachi as a distinctspecies from nominal B fasciata The revalidation of Bsteinbachi was also supported by phenotypic dataobtained from syntypes but did not include a reexamina-tion of all diagnostic characters of the speciesNext we present an amended diagnosis of B steinba-
chi based on novel data on life colours and morphologyand describe its vocal repertoire based on topotypesfrom Bolivia and distinct OTUs from Peru and Brazil
539540541542543544545546547548549550551552553554555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592
593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644645646
6 A Fouquet et al
PROOF ONLY
647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695696697698699700
701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754
Fig 1 Subtrees of the three main clades from the chronogram obtained from the analysis of 16S sequences using BEAST2Terminals are collapsed according to the OTU recovered from the species delineation analysis The distribution of each of these OTUis depicted on the maps
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 7
PROOF ONLY
also assigned to this species in this study Specimensand calls from Acre (ldquoB aff steinbachi 1rdquo) and callsfrom the lower Madre de Dios River (ldquoB aff steinbachi2rdquo) were included in the variation of B steinbachi
Boana steinbachi (Boulenger 1905)Hyla steinbachi mdash Boulenger 1905
Hyla fasciata De la Riva 1990Hypsiboas fasciatus Jansen et al 2011
Hypsiboas sp (Clade G) Funk et al 2012Hypsiboas steinbachi Caminer amp Ron 2014
Boana steinbachi Dubois 2017Boana sp (Clade J) Meza-Joya et al 2019
Boana fasciata Vacher et al 2020
Syntypes BMNH 194721361ndash63 two adults ofunknown sex and one juvenile respectively from Saraprovince Department of Santa Cruz de La SierraBolivia Collected by Hf J Steinbach
Diagnosis Boana steinbachi is characterized by the fol-lowing combination of character states (1) skin flap onheel (2) vocal repertoire composed of more than onecall type (3) multinote call and (4) regular internoteintervals (Figs 2 3 amp 4)
Comparisons with congeners of the Amazonian cladeBoana steinbachi can be distinguished from the membersof the B calcarata clade (B almendarizae B fasciata Bcalcarata and B maculateralis) by the absence of a cal-car and by the vocal repertoire composed of more thanone type of call Among members of the B steinbachiclade it can be distinguished from B dentei B alfaroiand B tetete by the presence of a skin flap on heel (tuber-cle in B alfaroi and B tetete tubercle on one side orcompletely absent in B dentei) by its multinote call (one-note call in B dentei and B tetete) by having regularinternote intervals between call notes (notes with irregularintervals sometimes partly fused one with the next in Balfaroi) and by the vocal repertoire composed of two dis-tinct types of calls (one call type in B alfaroi) (Camineramp Ron 2014 Marinho et al 2020)
Variation Body size varies between 304 and 374mmin males and between 425 and 488mm in females(Appendix 7) Individual AAG-UFU 5918 lacks the heelskin flap on the left side topotypes SMF 88394ndash95 andSMF 88397 lack the skin flap on both sides In life(Fig 3a b) dorsal colouration is beige light brownbright yellow or orange brown in calling males withfaint brown transversal bands and dark purplish-brownfaint brown or grey longitudinal lines extending from
the snout to vent a second line extending from behindthe eye to pelvic region and a third line as a canthalstripe extending from the posterior corner of the nostrilto anterior corner of the eye and from the posterior cor-ner of the eye to midbody length on flank (the topotypesSMF 88394ndash97 lack transversal bands on dorsum indi-viduals SMF 88394 and CORBIDI 13414 lack longitu-dinal lines on dorsum and the middorsal line isextending from snout to interocular region AAG-UFU5921 lacks longitudinal lines on dorsum) Iris cream orgrey sometimes with yellow pigmentation on the upperpart of the iris Throat varying from white to yellowchest and anterior belly varying from white to creamVentral surface of hand and hind limb beige to bright
755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808
809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862
Fig 2 Adult male of Boana steinbachi (AAG-UFU 5921)from the Assis Brasil population (Acre Brazil) (A) dorsal and(B) ventral views of the body (SVL frac14 337mm) (C) dorsaland (D) lateral views of the head (HL frac14123mm HW frac14102mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 106mm) and (G) sole of thefoot (FL frac14 139mm)
8 A Fouquet et al
antoinefouquet
Texte surligneacute
we may wanna ask for full page here no
PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
TSAB1873869 VOL 0 ISS 0
Systematics and biogeography of the Boana albopunctataspecies group (Anura Hylidae) with the description
of two new species from Amazonia
Antoine Fouquet Pedro Marinho Alexandre Rejaud Thiago R Carvalho Marcel A CaminerMartin Jansen Raıssa N Rainha Miguel T Rodrigues Fernanda P Werneck Albertina P Lima
Tomas Hrbek Ariovaldo A Giaretta Pablo J Venegas German Chavez and Santiago Ron
QUERY SHEET
This page lists questions we have about your paper The numbers displayed at left are hyperlinked to the location of thequery in your paper
The title and author names are listed on this sheet as they will be published both on your paper and on the Table ofContents Please review and ensure the information is correct and advise us if any changes need to be made In additionplease review your paper as a whole for typographical and essential corrections
Your PDF proof has been enabled so that you can comment on the proof directly using Adobe Acrobat For furtherinformation on marking corrections using Acrobat please visit httpjournalauthorstandfcoukproductionacrobatasphttpsauthorservicestaylorandfranciscomhow-to-correct-proofs-with-adobe
The CrossRef database (wwwcrossreforg) has been used to validate the references
AUTHOR QUERIES
Q1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak ampHo 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila-Pires et al 2010 and Avila ampKawashita -Ribeiro 2011) in the reference list or delete the citation from the text
Q2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Geurounther 1858Geurounther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please inserta citation in the text or delete the reference as appropriate
Q3 Please provide the volume number and page range
Q4 Please provide the volume number and page range
Q5 The year of publication has been changed as per Crossref details both in the list and in the text forthis reference Please check
Q6 The year of publication has been changed as per Crossref details both in the list and in the text forthis reference Please check
PROOF ONLY
Q7 If your paper introduces new zoological taxa at family-group level or below you are now required toregister your paper with ZooBank and insert the generated ZooBank ID (LSID) here Individual newtaxa need not be registered before publication this can be done subsequently should you wish Pleasego to httpwwwzoobankorgregister to complete this task You will need your article DOI to registerAfter publication you must amend your ZooBank record of your paper to reflect the date of publica-tion Please see httpwwwzoobankorghelp for further information
PROOF ONLY
Research Article
Systematics and biogeography of the Boana albopunctataspecies group (Anura Hylidae) with the descriptionof two new species from Amazonia
ANTOINE FOUQUET1 PEDRO MARINHO2 ALEXANDRE REacuteJAUD1 THIAGO R CARVALHO3 MARCEL A CAMINER45 MARTIN JANSEN6 RAIacuteSSA N RAINHA7 MIGUEL T RODRIGUES8 FERNANDA P WERNECK7 ALBERTINA P LIMA7 TOMAS HRBEK9 ARIOVALDO A GIARETTA2 PABLO J VENEGAS10 GERMAacuteN CHAacuteVEZ10 amp SANTIAGO RON4
1Laboratoire Evolution et Diversite Biologique UMR 5174 CNRS IRD Universite Paul Sabatier Batiment 4R1 31062cedex 9 118 Route de Narbonne Toulouse 31077 France2Laboratorio de Anuros Neotropicais Instituto de Ciencias Exatas e Naturais do Pontal Universidade Federal de UberlandiaItuiutaba MG Brazil3Laboratorio de Herpetologia Departamento de Biodiversidade e Centro de Aquicultura IB Universidade Estadual PaulistaRio Claro SP Brazil4Museo de Zoologıa Escuela de Biologıa Pontificia Universidad Catolica del Ecuador Quito Ecuador5Institute of Organismic and Molecular Evolution Johannes Gutenberg University Mainz Germany6Department of Terrestrial Zoology Research Institute and Nature Museum Senckenberg Frankfurt Germany7Instituto Nacional de Pesquisas da Amazonia Coordenac~ao de Biodiversidade Manaus AM Brazil8Departamento de Zoologia Universidade de S~ao Paulo Instituto de Biociencias S~ao Paulo SP Brazil9Departamento de Genetica Universidade Federal do Amazonas Manaus AM Brazil10Instituto Peruano de Herpetologıa Lima Peru
The outstanding species richness of Amazonia has fascinated biologists for centuries However the records of actualnumbers and distribution of species forming its ecosystems are so incomplete that the understanding of the historicalcauses and regional determinants of this diversity remain speculative Anuran clades have repeatedly been documentedto harbour many unnamed species in this region notably the Boana albopunctata species group Considering thedocumented distribution and the ecology of the species of that group we hypothesized that it diversified via successivetrans-riverine dispersals during the late Miocene and Pliocene after the formation of the modern Amazon watershedTo test this hypothesis we gathered an extensive dataset of 16S rDNA sequences sampled throughout Amazonia anda mitogenomic dataset representative of the diversity of the clade to (1) re-evaluate species boundaries and distributionsand (2) infer the spatio-temporal history of diversification within Amazonia We delimited 14 Operational TaxonomicUnits (OTUs) in an Amazonian clade ie 75 higher than currently recognized (14 OTUs for eight described species)Combining molecular data with morphological and acoustic data two new species Boana courtoisae sp nov fromthe eastern Guiana Shield and Boana eucharis sp nov from Southern Amazonia are described herein These speciesbelong to a clade that diversified throughout Amazonia during the last 10Ma thus more recently than co-distributedsmall terrestrial anurans but concomitantly with other more vagile vertebrates Our time-scaled phylogeny andbiogeographic analyses suggest an initial east-west divergence and confirm reciprocal trans-riverine dispersals duringthe last 5Ma The geomorphological evolution of the region and species-specific dispersal ability largely explain thesedistinct spatio-temporal patterns across anuransQ7
Key words Amphibia biodiversity Guiana Shield mitogenomics neotropics phylogenetics
IntroductionThe Neotropics harbour the most diverse ecosystems onthe planet (Jenkins et al 2013) Within the Neotropics
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253
54555657585960616263646566676869707172737475767778798081828384858687888990919293949596979899100101102103104105106
Correspondence to Antoine Fouquet E-mailfouquetantoinegmailcom Pedro Marinho E-mailpmarinho50gmailcom
ISSN 1477-2000 print 1478-0933 online The Trustees of the Natural History Museum London 2021 All Rights Reservedhttpsdxdoiorg1010801477200020211873869
Systematics and Biodiversity (2021) 0(0) 1ndash38
Author Query
AQ7 If your paper introduces new zoological taxa at family‐group level or below you are now required to register your paper with ZooBank and insert the generated ZooBank ID (LSID) here Individual new taxa need not be registered before publication this can be done subsequently should you wish Please go to httpwwwzoobankorgregister to complete this task You will need your article DOI to register After publication you must amend your ZooBank record of your paper to reflect the date of publication Please see httpwwwzoobankorghelp for further information
antoinefouquet
Note
Boana courtoisaezoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBEBoana euchariszoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
PROOF ONLY
Amazonia stands as a major biogeographic region cover-ing more than 65 million km2 including the largestcontinuous tract of tropical forest (40 of the worldrsquostropical rainforests) encompassing the most importantdrainage (Amazon River basin) and probably hostingthe highest continental biodiversity on earth (Myerset al 2000) The outstanding species richness of thisregion has raised many questions concerning its originsand diversification mechanisms While all agree on theastounding diversity of Amazonia our understanding ofthe actual number and distribution of species forming itsecosystems is in fact so incomplete (Ficetola et al2014 Meyer et al 2015 Vacher et al 2020) that ithas so far hampered investigations at the regional scaleConsequently the understanding of the historical causesand regional determinants of this outstanding diversityremain speculative (Antonelli et al 2018) Another con-sequence is that the collision between the Linnean short-fall and the biodiversity crisis is particularly evident inAmazonia (Guerra et al 2020) With almost the entireeastern and southern parts (25) of its extent beingalready deforested for agriculture an unknown propor-tion of endemic species and thus of the testimony ofAmazonian history has already vanished (Lovejoy ampNobre 2019)Anuran clades have repeatedly been documented to
harbour many unnamed species in Amazonia (Vacheret al 2020) and to display strikingly allopatric distribu-tion patterns that could provide crucial insights intoAmazoniarsquos past (Fouquet Cassini Haddad Pech ampRodrigues 2014 Rejaud et al 2020) This is notablythe case of the Boana albopunctata species group(Caminer amp Ron 2014 Funk et al 2012) This groupis only defined on the basis of molecular data(Faivovich et al 2005) and currently comprises 16valid nominal species distributed in the Caribbean andSouth America Boana albopunctata (Spix 1824)Boana alfaroi (Caminer amp Ron 2014) Boana almen-darizae (Caminer amp Ron 2014) Boana caiapo Pinheiroet al 2018 Boana calcarata (Troschel 1848) Boanadentei (Bokermann 1967) Boana fasciata (Geurounther1858) Boana heilprini (Noble 1923) Boana lancifor-mis (Cope 1871) Boana leucocheila (Caramaschi amp deNiemeyer 2003) Boana maculateralis (Caminer ampRon 2014) Boana multifasciata (Geurounther 1859)Boana paranaiba (Carvalho Giaretta amp Facure 2010)Boana steinbachi (Boulenger 1905) Boana tetete(Caminer amp Ron 2014) and Boana raniceps (Cope1862) Within this clade B heilprini is the most distinctsince it is confined to Hispaniola and is phylogeneticallydistant from a clade formed by all the other species ofthe group (Duellman Marion amp Hedges 2016) Wecan also distinguish B raniceps and the B albopunctata
clade (B lanciformis B albopunctata B multifasciataB paranaiba B caiapo B leucocheila) which occurwidely in the open habitats of the Cerrado andAmazonia (Camurugi et al 2021) from the Amazonianclade According to Caminer and Ron (2014) thisAmazonian clade is itself subdivided in two maingroups hereafter called the B calcarata (B fasciata Bcalcarata B almendarizae B maculateralis) and the Bsteinbachi (B tetete B alfaroi B steinbachi) cladesWithin this Amazonian clade Funk Caminer and
Ron (2012) identified no less than seven putativeunnamed species based on mitochondrial divergence andacoustic data These species add up to the 10 taxa thatwere valid at that time Caminer and Ron (2014) com-pleted this picture by documenting two additional line-ages describing four of the previously discoveredspecies and removing B steinbachi from its synonymywith B fasciata However sampling of both studies wascircumscribed to western Amazonia (mostly Ecuador)while the species complex occurs throughout AmazoniaPopulations from the eastern Guiana Shield lowlandsare already assumed to belong to a yet unnamed species(Caminer amp Ron 2014) but the status of numerousother populations throughout Amazonia generally iden-tified as B fasciata remain virtually unknown Morerecently Vacher et al (2020) suggested based on 16Smitochondrial DNA (mtDNA) sequences that up to 22species could exist in that group and that the distributionof most species could be circumscribed to small rangeswithin AmazoniaThese aforementioned studies improved our under-
standing of the actual diversity in the B calcaratastein-bachi clade but left virtually unexplored the temporaland spatial context of the diversification withinAmazonia The crown age of this Amazonian clade wasestimated between 16ndash11 million years ago (Ma) byFunk et al (2012) and between 14ndash9Ma by Duellmanet al (2016) Therefore the initial diversification of thatclade throughout Amazonia may have taken place dur-ing the final stage of the Pebas system a freshwaterlake occupying most of Western Amazonia from theearly Miocene (23Ma) until 10ndash9Ma (Hoorn et al2017) Meanwhile most of the diversification within theB calcaratasteinbachi clade took place during the lateNeogene (last 10Ma) From 9Ma onward this systemhas drained eastward into the Atlantic Ocean but enor-mous flooded ecosystems (Acre system) still occupied alarge portion of what we currently consider as WesternAmazonia until 7Ma (Hoorn et al 2010 Albert et al2018a) Subsequent river captures were common(Ruokolainen Moulatlet Zuquim Hoorn amp Tuomisto2018) as were climatic fluctuations that may havemodified vegetation possibly to the point of forest
107108109110111112113114115116117118119120121122123124125126127128129130131132133134135136137138139140141142143144145146147148149150151152153154155156157158159160
161162163164165166167168169170171172173174175176177178179180181182183184185186187188189190191192193194195196197198199200201202203204205206207208209210211212213214
2 A Fouquet et al
antoinefouquet
Texte inseacutereacute
mostly
PROOF ONLY
fragmentation at least in some peripheral parts ofAmazonia (Cheng et al 2013 Kirschner amp Hoorn2019) The relative roles of hydrological evolution(Albert et al 2018b Ribas Aleixo Nogueira Miyakiamp Cracraft 2012) and rainforest expansion and contrac-tion due to climate oscillations (Haffer 1969) have beenthe focus of intense debate (Leite amp Rogers 2013)Since the species of the B calcarata and B steinbachicomplexes are associated with riparian forests and thesespecies mostly breed in pools formed on the banks ofthe beds of small to medium-sized rivers very large riv-ers may act as barriers to dispersal thus reducing orcompletely impeding gene flow and inhibiting homogen-ization of differentiating populations (Naka et al 2018)However smaller and highly dynamic meandering riversprobably represent more permeable barriers Thereforewe assume that successive dispersals across majorAmazonian rivers (parapatric speciation) (Pirani et al2019) as well as hydrological changes (vicariant speci-ation) (Naka et al 2018) could have been major proc-esses of diversification in these frogsWe gathered a large mtDNA dataset for the B albo-
punctata group sampled throughout Amazonia combinedwith morphological and acoustic data for a subset ofthis group (the B steinbachi clade) to address two maingoals (1) reevaluate species boundaries (in or amongmembers) of the B albopunctata group and theirrespective distributions and (2) investigate its diversifi-cation history within Amazonia using mitogenomic datafor one terminal of each delimited species Furthermoretwo species in the B steinbachi clade that wereunnamed are described herein
Materials and methodsSpecies delimitationOur first objective was to delimit all major OperationalTaxonomic Units (OTUs) based on mtDNA main line-ages Our sampling included 55 new Boana tissue sam-ples obtained through fieldwork throughout Amazoniaand adjacent Dry Diagonal (DD Chaco CerradoCaatinga Werneck 2011) and loans from collaborators(from collections QCAZ MPEG INPA CORBIDIAAGUFU and from personal loans JMP SCF FTAAppendix 1) Samples unambiguously identified (neartype locality andor phenotypically corresponding to typematerial) as belonging to the currently recognized taxawere included except B paranaiba and B caiapo Wesequenced a 400 bp portion of the end of the 16S rDNAgene a locus commonly used for Neotropical amphibiantaxonomy and systematics (Vences Thomas Bonett ampVieites 2005) We also retrieved homologous sequences
from GenBank (389 accessions) In total we gathered444 16S sequences for this study (dataset details are pro-vided in Appendix 1 and DNA extraction and sequencingprotocols can be found in Appendix 2) These samplescover the whole distribution of the B albopunctata spe-cies group (14 included taxa) DNA sequence alignmentwas conducted on the MAFFT7 online server under theE-INS-i option with default parameters an algorithmdesigned for sequences with multiple conserved domainsand long gaps (Katoh et al 2019)We applied three DNA-based single-locus species
delimitation approaches (a) a distance-based methodthe Automated Barcode Gap Discovery (ABGDPuillandre Lambert Brouillet amp Achaz 2012) (b) amulti-rate coalescent based method the multi-ratePoisson Tree Processes model approach (mPTP Kapliet al 2017) and (c) a single-threshold coalescent-basedmethod the Generalized Mixed Yule Coalescentapproach (single threshold GMYC Monaghan et al2009 Pons et al 2006)The ABGD delimitation was performed with a prior
of intraspecific divergence P between 0001 and 01 aproxy for minimum relative gap width X of 1 and anumber of steps n equal to 30 We kept the partitionsuch that Pfrac14 0016 as it corresponds to the end of aplateau for group number and it matches thresholds ofintraspecific divergence proposed in other vertebratedelimitation studies using 16S barcodes (Puillandreet al 2012) For mPTP delimitation we first recon-structed a Maximum Likelihood (ML) tree withRAxML v824 (Stamatakis 2014) using theGTRthornGthorn I substitution model and estimated nodalsupport via 1000 parametric bootstraps We used nineoutgroups representing most other Boana species groups(Appendix 3) The mPTP delimitation was undertakenon the rooted ML tree with 5 million Markov chainMonte Carlo (MCMC) iterations sampling every10000th iteration and a 10 burn-in For the GMYCdelimitation we reconstructed a time-calibrated phyl-ogeny using BEAST 25 (Bouckaert et al 2014) Weused a birth-death population model to account forextinction processes and incomplete sampling Weincluded the same nine Boana outgroups used inRAxML reconstruction and five additional representa-tives of other Cophomantini genera (NesorohylaMyersiohyla Hyloscirtus AplastodiscusBokermannohyla) in order to include relationships thatcan be time-calibrated We used a single partition with aGTRthornGthorn I substitution model with an uncorrelatedrelaxed lognormal clock model of rate variation amongbranches (Drummond Ho Phillips amp Rambaut 2006)We used two calibration points the ages of the mostrecent common ancestor (MRCA) of Aplastodiscus and
215216217218219220221222223224225226227228229230231232233234235236237238239240241242243244245246247248249250251252253254255256257258259260261262263264265266267268
269270271272273274275276277278279280281282283284285286287288289290291292293294295296297298299300301302303304305306307308309310311312313314315316317318319320321322
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 3
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13536302v1
PROOF ONLY
Boana as well as the MRCA of a clade formed byHyloscirtus Boana Aplastodiscus andBokermannohyla which were estimated by Feng et al(2017) and consequently constrained here assuming nor-mal prior distributions of 252Ma (standard deviation[SD]frac14 28Ma) and 323Ma (SD frac14 3Ma) respectivelyWe considered these calibrations to represent the bestprior because Feng et al (2017) included a comprehen-sive dataset of nuclear loci for an extensive sampling ofanurans and used many reliable fossils as calibrationpoints Moreover Hime et al (2020) analysing an evenlarger genomic dataset found very similar time esti-mates for the TMRCA of BoanathornHyloscirtus Insteadusing fossil andor biogeographic calibrations wouldhave implied to expand the matrix to lineages distantlyrelated to Boana and would likely lead to an overesti-mation of calibration dates with our mitogenomic data-set (Molak amp Ho 2015 Papadopoulou et al 2010) Forthe MCMC parameters we used four independentchains of 100 million iterations recording every10000th iteration and a 10 burn-in We combined thelog files of the independent runs using LogCombiner25 (Drummond amp Rambaut 2007) and checked theconvergence of our parameters confirmed by allEffective Sample Size (ESS) being above 200 Then weextracted the maximum clade credibility tree using Treeannotator 25 (Drummond amp Rambaut 2007) with aburn-in of 10 After removing outgroups we per-formed a GMYC delimitation on the ultrametric treeusing the GMYC function of the splits R package v10-11 (Ezard Fujisawa amp Barraclough 2009) with athreshold interval between 0 and 10Ma and by usingthe single threshold method Operational TaxonomicUnits (OTUs) were defined using a majority-rule con-sensus from the results of the three methods ie a lin-eage is considered as being an OTU if supported by atleast two of the three methodsAll but five occurrence records were georeferenced
and used to create distribution maps with convex poly-gons under QGIS 214 (QGIS Geographic InformationSystem) OTUs were assigned to taxa based on thefieldmuseum identification and sometimes corrected inaccordance with type localities and known distribution(Frost 2019 IUCN 2020)
Morphological and acoustic variation in theBoana steinbachi cladeThe following morphometric measurements were takenon 38 males and 9 females snout-vent length (SVL)head length (HL) head width (HW) eye diameter (ED)tympanum diameter (TD) tibia length (TL) foot length(FL) thigh length (THL) and calcaneal appendage
length (CL) In addition to these we also measuredhand length (HAL) forearm length (FLL) and eyendashnos-tril distance (EN) for the morphometric characterizationof holotypes All measurements followed the definitionsand terminology of Watters Cummings Flanagan andSiler (2016) We classified the calcaneal appendage intothree-character states (i) a calcar (skin appendage withlength gt 1mm flattened or conical) a (ii) skin flap(skin appendage with length lt 1mm flattened or a(iii) tubercle a conical structure with reduced size andas such not forming an appendage coming out of theheel The specimens were measured by P Marinho(Alta Floresta Mato Grosso State and Assis BrazilAcre State) using a Mitutoyo digital calliper (to thenearest 005mm) by MT Rodrigues (Jirau and PacaasNovos Rondonia State) using a Mitutoyo digital calliper(to the nearest 001mm) by A Fouquet (French Guianaand Suriname) using a DigiMax digital calliper (nearest001mm) and by M Caminer (Ecuador Peru andBolivia) using a Mannesmann digital calliper (nearest001mm) Snout shape was assessed according to Heyeret al (1990) Digital webbing formulae followed thenotation system of Savage and Heyer (1997)Acoustic analysis of 27 recorded calling males
(Appendix 4) was conducted in Raven Pro 15 64-bit ver-sion (Bioacoustics Research Program 2012) sound figureswere produced using Seewave version 210 (SueurAubin amp Simonis 2008) and tuneR version 132(Ligges Krey Mersmann amp Schnackenberg 2014) in Rversion 350 (R Core Team 2018) Raven Pro settingswindow size frac14 512 samples window typefrac14Hann 3 dBfilter bandwidth frac14 124Hz window overlap frac14 85 hopsize frac14 77ms discrete Fourier transform (DFT) size frac141024 samples grid spacing frac14 431Hz seewave settingswindow typefrac14Hanning fast Fourier transform (FFT)size frac14 256 samples FFT overlap frac14 90 The analysedfiles are deposited in the sound collection of the lsquoMuseude Biodiversidade do Cerradorsquo (AAG-UFU) lsquoFonotecaNeotropical Jacques Vielardirsquo (FNJV) lsquoLa Sonotheque duMuseum National dHistoire Naturellersquo (MNHN) and theMacaulay library (MC) Detailed information about theanalysed sound files and accession numbers are providedin Appendix 4Call types were distinguished based on the number of
notes per call and pulsing We defined as type 1 callsthe multinote calls formed by nonpulsed notes whichconsist of the main call type emitted by males of allstudied species Type 2 calls are emitted less often thantype 1 and consist of a one-note call formed by poorlydefined pulses We only included type 1 calls in theacoustic diagnoses and the interspecific comparisonsThe acoustic traits were generally analysed manually (ifnot stated otherwise) as follows temporal traits (call
323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370371372373374375376
377378379380381382383384385386387388389390391392393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430
4 A Fouquet et al
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
1
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics 2014
antoinefouquet
Texte inseacutereacute
a
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
s
PROOF ONLY
duration note duration interval between notes numberof notes and call rise time the latter using the lsquoPeakTimersquo function) frequency traits (dominant frequencyusing the lsquoPeak Frequencyrsquo function minimum andmaximum frequency using the lsquoFrequency 5rsquo andlsquoFrequency 95rsquo functions respectively) Call traitsused in the description followed the definitions and ter-minology of Keuroohler et al (2017) using a note-cen-tred approach
Time-calibrated species phylogenyWe selected a representative for each of the 25 OTUsidentified by the species delimitation (see Results) forcomplete mitogenome sequencing to investigate interre-lationships and divergence times Mitogenomic sequen-ces were obtained using low-coverage shotgunsequencing We recovered high-quality mitochondrialgenome assemblies for 18 OTU representatives (seeAppendix 2 for details regarding mitogenome sequenc-ing assembling and annotation) For the remainingseven OTUs for which we could not obtain tissue sam-ples (Appendix 3) we gathered all the available mito-chondrial loci (12S 16S ND1 COI and Cytb) fromGenBank four of these OTUs were represented by 16Sonly We also selected 14 species as outgroups (ninewith complete mitogenomes) including representativesfor other Boana species groups and most otherCophomantini genera (Appendix 3)We extracted 12S 16S and all protein-coding
sequence regions (CDS thus removing D-Loop andtRNAs) from complete mitogenomes as well asGenBank accessions and aligned each locus independ-ently using the MAFFT7 online server For rRNAgenes we chose the E-INS-i strategy recommended forsequences with multiple conserved domains and longgaps For the protein-coding genes we chose the G-INS-i strategy designed for sequences with global hom-ology (Katoh et al 2019) Realignments of CDS con-sidering the reading frame were done and concatenatedin Geneious v918 (httpswwwgeneiouscom) Ourfinal matrix totalled 39 terminals and 13620 alignednucleotide sites Among these terminals 27 were com-plete and nine had fewer than 3000 nucleotides (20)but all were putatively closely related to terminals withcomplete dataWe selected the best-fit partition scheme and model
of evolution for each partition using PartitionFinderV211 (Lanfear et al 2016) based on the BayesianInformation Criterion (BIC) We predefined four datablocks one for rRNA genes (12S and 16S) and one foreach codon position of all CDS regions and found theGTRthorn IthornG model to best-fit all partitions We
reconstructed a time-calibrated tree using a birth-deathtree prior using BEAST 25 to account for extinctionprocesses and incomplete sampling We parameterizedunlinked substitution models according to the estimatesobtained in the PartitionFinder V211 analysis (Lanfearet al 2016) We used the same two calibration pointsas in the previous BEAST analysis Analyses wereundertaken using uncorrelated relaxed lognormal clockmodel of distribution of rate variation among branchesfor each partition (Drummond et al 2006) The Markovchain Monte Carlo (MCMC) parameters were set withfour independent chains of 100 million iterations stor-ing every 10000th iteration and a 10 burn-in Wecombined the log files of the independent runs usingLogCombiner 25 and confirmed the convergence of ourparameters as all ESS were above 200 Finally weextracted the maximum clade credibility tree using Treeannotator 25 We acknowledge that our phylogeneticreconstruction solely based on mtDNA sequences priori-tizes spatial and taxonomic completeness over genomiccoverage and can lead to overestimated divergence times(eg McCormack Heled Delaney Peterson ampKnowles 2011 Near et al 2012)
Biogeographic analysisBiogeographic inferences were undertaken on the time-calibrated phylogeny using the BioGeoBEARS R pack-age (Matzke 2013) This package reconstructs ancestralgeographic distributions and investigates the role ofeach biogeographic event with a maximum likelihoodalgorithm We compared three different models (i) alikelihood version of the Dispersal-Vicariance(DIVALIKE) model (Ronquist 1997) (ii) a likelihoodversion of the BayArea (BBM) model (Landis et al2013) and (iii) the Dispersal Extinction Cladogenesis(DEC) model (Ree amp Smith 2008) We also comparedversions of these models allowing jump dispersal asdescribed by the J parameter (Matzke 2013) Modelswere compared with the Akaike Information Criterion(AIC) We ran 50 independent BioGeoBEARS biogeo-graphic stochastic mapping to determine biogeographicevent counts for the best-fit model (Dupin et al 2017)Ree and Sanmartın (2018) voiced several criticismsagainst the use of J parameter arguing that it inflates thecontribution of cladogenetic events to the likelihoodand minimizes the contribution of anagenetic time-dependent range evolution We thus also compared theresults from the best model without jump dispersalSince this work focuses on the historical biogeog-
raphy of the Amazonian clade we discarded the twoearly diverging lineages B heilprini and B ranicepsfrom the biogeographic analyses since their distribution
431432433434435436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484
485486487488489490491492493494495496497498499500501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 5
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13560371v1)
antoinefouquet
Texte inseacutereacute
D
PROOF ONLY
ranges lie mostly outside of Amazonia We consideredthree areas within Amazonia Western Amazonia theBrazilian Shield and the Guiana Shield These areascorrespond to major geological features roughly delim-ited by modern riverine barriers the Madeira River theNegro River and the lower course of the Amazon Riverand to three large biogeographic regions known asWallacersquos districts (Hoorn et al 2010 Wallace 1854)These districts were recently confirmed as major breaksin birdsrsquo species composition (Oliveira Vasconcelos ampSantos 2017) and amphibians (Godinho amp da Silva2018 Vacher et al 2020) strengthening their status ofbiogeographic regions Because the distribution range ofthe B albopunctata group extends outside Amazoniawe included in the ancestral range reconstruction oneadditional non-Amazonian Neotropical area theDry Diagonal
ResultsSpecies delimitationOf the three tested methods of species delimitation theABGD method was found to be the most conservativedelimiting 21 OTUs while mPTP and GMYC delimited24 and 47 OTUs respectively (Appendix 1) In onecase the consensual partitioning was overly conserva-tive given that B leucocheila and B multifasciata wereconsidered as a single OTU which is contradicted bythe fact that these two taxa are phenotypically distinctTherefore we kept the different lineages delimited byGMYC in this group as distinct OTUs The final delimi-tation led to 25 OTUs (including B heilprini) in the Balbopunctata species group (Fig 1 Appendix 1)Thirteen of these OTUs could be linked to nominal spe-cies (except B caiapo and B paranaiba that were notincluded in the analysis because material was not avail-able) Conversely 11 could not be linked to any nom-inal species representing a possible 44 increase in thespecies richness of the group The Amazonian cladeitself was represented by 14 OTUs including eight nom-inal species and six putative new species (Fig 1) Theseresults also imply important changes to the geographicdistribution of the species of the group In Appendix 5we detail and justify the identification of these OTUsand their respective geographic ranges
Taxonomic accountsThe Amazonian clade of the B albopunctata group pre-viously reported as the B calcaratandashB fasciata speciescomplex by Caminer and Ron (2014) is composed oftwo major clades (the B calcarata and the B steinbachi
clades) defined by the following combination of charac-ter states (1) truncate snout in dorsal view (2) roundedhead shape and (3) presence of calcaneal appendageon heelThe B calcarata clade (Fig 1) is composed of four
species possessing a calcar and having advertisementcalls formed by a single type of call (Caminer amp Ron2014) The B steinbachi clade on the other handincludes three described and two undescribed species(Fig 1) Members of this clade lack a calcar (skin flapor tubercle present or the complete absence of calcanealappendages) and have vocal repertoires composed ofmore than one type of call except for B alfaroi forwhich a single type of call has been documented so far(Caminer amp Ron 2014)Hyla steinbachi Boulenger 1905 currently valid as
Boana steinbachi (Boulenger 1905) was describedfrom the Bolivian Province Sara Departament SantaCruz de la Sierra Bolivia We assume that Buenavistais the type locality of B steinbachi Between 1910 and1950 the Steinbach family collected many amphibiansand reptiles in Bolivia mainly at Buenavista in theDepartment of Santa Cruz (see Parker 1927) which isthe type locality of several anurans (Hamptophryneboliviana (Parker 1927) Pseudopaludicola bolivianaParker 1927 Scinax parkeri (Gaige 1929)) and thesnake Apostolepis tenuis Ruthven 1927 The materialcollected by the Steinbach family is on display in vari-ous museums such as the University of MichiganMuseum of Zoology (UMMZ see Gaige 1929) theNatural History Museum London (BMNH see Parker1927) the Zoologisches Museum Berlin (ZMB seeMeurouller 1924) and the Naturhistorisches Museum Basel(NBM pers comm Weurouest) Many decades later De laRiva (1990) synonymized H steinbachi with H fas-ciata Subsequently Jansen Bloch Schulze andPfenninger (2011) found differences in call and mtDNAdata between populations of B fasciata from Boliviaand Ecuador and suggested that their results imply a res-urrection of B steinbachi In the following yearsCaminer and Ron (2014) tentatively assigned B steinba-chi to a genetic lineage (clade J) inhabiting BolivianAmazonia near its type locality By assessing its phylo-genetic relationships in the group Caminer and Ron(2014) formally revalidated B steinbachi as a distinctspecies from nominal B fasciata The revalidation of Bsteinbachi was also supported by phenotypic dataobtained from syntypes but did not include a reexamina-tion of all diagnostic characters of the speciesNext we present an amended diagnosis of B steinba-
chi based on novel data on life colours and morphologyand describe its vocal repertoire based on topotypesfrom Bolivia and distinct OTUs from Peru and Brazil
539540541542543544545546547548549550551552553554555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592
593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644645646
6 A Fouquet et al
PROOF ONLY
647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695696697698699700
701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754
Fig 1 Subtrees of the three main clades from the chronogram obtained from the analysis of 16S sequences using BEAST2Terminals are collapsed according to the OTU recovered from the species delineation analysis The distribution of each of these OTUis depicted on the maps
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 7
PROOF ONLY
also assigned to this species in this study Specimensand calls from Acre (ldquoB aff steinbachi 1rdquo) and callsfrom the lower Madre de Dios River (ldquoB aff steinbachi2rdquo) were included in the variation of B steinbachi
Boana steinbachi (Boulenger 1905)Hyla steinbachi mdash Boulenger 1905
Hyla fasciata De la Riva 1990Hypsiboas fasciatus Jansen et al 2011
Hypsiboas sp (Clade G) Funk et al 2012Hypsiboas steinbachi Caminer amp Ron 2014
Boana steinbachi Dubois 2017Boana sp (Clade J) Meza-Joya et al 2019
Boana fasciata Vacher et al 2020
Syntypes BMNH 194721361ndash63 two adults ofunknown sex and one juvenile respectively from Saraprovince Department of Santa Cruz de La SierraBolivia Collected by Hf J Steinbach
Diagnosis Boana steinbachi is characterized by the fol-lowing combination of character states (1) skin flap onheel (2) vocal repertoire composed of more than onecall type (3) multinote call and (4) regular internoteintervals (Figs 2 3 amp 4)
Comparisons with congeners of the Amazonian cladeBoana steinbachi can be distinguished from the membersof the B calcarata clade (B almendarizae B fasciata Bcalcarata and B maculateralis) by the absence of a cal-car and by the vocal repertoire composed of more thanone type of call Among members of the B steinbachiclade it can be distinguished from B dentei B alfaroiand B tetete by the presence of a skin flap on heel (tuber-cle in B alfaroi and B tetete tubercle on one side orcompletely absent in B dentei) by its multinote call (one-note call in B dentei and B tetete) by having regularinternote intervals between call notes (notes with irregularintervals sometimes partly fused one with the next in Balfaroi) and by the vocal repertoire composed of two dis-tinct types of calls (one call type in B alfaroi) (Camineramp Ron 2014 Marinho et al 2020)
Variation Body size varies between 304 and 374mmin males and between 425 and 488mm in females(Appendix 7) Individual AAG-UFU 5918 lacks the heelskin flap on the left side topotypes SMF 88394ndash95 andSMF 88397 lack the skin flap on both sides In life(Fig 3a b) dorsal colouration is beige light brownbright yellow or orange brown in calling males withfaint brown transversal bands and dark purplish-brownfaint brown or grey longitudinal lines extending from
the snout to vent a second line extending from behindthe eye to pelvic region and a third line as a canthalstripe extending from the posterior corner of the nostrilto anterior corner of the eye and from the posterior cor-ner of the eye to midbody length on flank (the topotypesSMF 88394ndash97 lack transversal bands on dorsum indi-viduals SMF 88394 and CORBIDI 13414 lack longitu-dinal lines on dorsum and the middorsal line isextending from snout to interocular region AAG-UFU5921 lacks longitudinal lines on dorsum) Iris cream orgrey sometimes with yellow pigmentation on the upperpart of the iris Throat varying from white to yellowchest and anterior belly varying from white to creamVentral surface of hand and hind limb beige to bright
755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808
809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862
Fig 2 Adult male of Boana steinbachi (AAG-UFU 5921)from the Assis Brasil population (Acre Brazil) (A) dorsal and(B) ventral views of the body (SVL frac14 337mm) (C) dorsaland (D) lateral views of the head (HL frac14123mm HW frac14102mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 106mm) and (G) sole of thefoot (FL frac14 139mm)
8 A Fouquet et al
antoinefouquet
Texte surligneacute
we may wanna ask for full page here no
PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Q7 If your paper introduces new zoological taxa at family-group level or below you are now required toregister your paper with ZooBank and insert the generated ZooBank ID (LSID) here Individual newtaxa need not be registered before publication this can be done subsequently should you wish Pleasego to httpwwwzoobankorgregister to complete this task You will need your article DOI to registerAfter publication you must amend your ZooBank record of your paper to reflect the date of publica-tion Please see httpwwwzoobankorghelp for further information
PROOF ONLY
Research Article
Systematics and biogeography of the Boana albopunctataspecies group (Anura Hylidae) with the descriptionof two new species from Amazonia
ANTOINE FOUQUET1 PEDRO MARINHO2 ALEXANDRE REacuteJAUD1 THIAGO R CARVALHO3 MARCEL A CAMINER45 MARTIN JANSEN6 RAIacuteSSA N RAINHA7 MIGUEL T RODRIGUES8 FERNANDA P WERNECK7 ALBERTINA P LIMA7 TOMAS HRBEK9 ARIOVALDO A GIARETTA2 PABLO J VENEGAS10 GERMAacuteN CHAacuteVEZ10 amp SANTIAGO RON4
1Laboratoire Evolution et Diversite Biologique UMR 5174 CNRS IRD Universite Paul Sabatier Batiment 4R1 31062cedex 9 118 Route de Narbonne Toulouse 31077 France2Laboratorio de Anuros Neotropicais Instituto de Ciencias Exatas e Naturais do Pontal Universidade Federal de UberlandiaItuiutaba MG Brazil3Laboratorio de Herpetologia Departamento de Biodiversidade e Centro de Aquicultura IB Universidade Estadual PaulistaRio Claro SP Brazil4Museo de Zoologıa Escuela de Biologıa Pontificia Universidad Catolica del Ecuador Quito Ecuador5Institute of Organismic and Molecular Evolution Johannes Gutenberg University Mainz Germany6Department of Terrestrial Zoology Research Institute and Nature Museum Senckenberg Frankfurt Germany7Instituto Nacional de Pesquisas da Amazonia Coordenac~ao de Biodiversidade Manaus AM Brazil8Departamento de Zoologia Universidade de S~ao Paulo Instituto de Biociencias S~ao Paulo SP Brazil9Departamento de Genetica Universidade Federal do Amazonas Manaus AM Brazil10Instituto Peruano de Herpetologıa Lima Peru
The outstanding species richness of Amazonia has fascinated biologists for centuries However the records of actualnumbers and distribution of species forming its ecosystems are so incomplete that the understanding of the historicalcauses and regional determinants of this diversity remain speculative Anuran clades have repeatedly been documentedto harbour many unnamed species in this region notably the Boana albopunctata species group Considering thedocumented distribution and the ecology of the species of that group we hypothesized that it diversified via successivetrans-riverine dispersals during the late Miocene and Pliocene after the formation of the modern Amazon watershedTo test this hypothesis we gathered an extensive dataset of 16S rDNA sequences sampled throughout Amazonia anda mitogenomic dataset representative of the diversity of the clade to (1) re-evaluate species boundaries and distributionsand (2) infer the spatio-temporal history of diversification within Amazonia We delimited 14 Operational TaxonomicUnits (OTUs) in an Amazonian clade ie 75 higher than currently recognized (14 OTUs for eight described species)Combining molecular data with morphological and acoustic data two new species Boana courtoisae sp nov fromthe eastern Guiana Shield and Boana eucharis sp nov from Southern Amazonia are described herein These speciesbelong to a clade that diversified throughout Amazonia during the last 10Ma thus more recently than co-distributedsmall terrestrial anurans but concomitantly with other more vagile vertebrates Our time-scaled phylogeny andbiogeographic analyses suggest an initial east-west divergence and confirm reciprocal trans-riverine dispersals duringthe last 5Ma The geomorphological evolution of the region and species-specific dispersal ability largely explain thesedistinct spatio-temporal patterns across anuransQ7
Key words Amphibia biodiversity Guiana Shield mitogenomics neotropics phylogenetics
IntroductionThe Neotropics harbour the most diverse ecosystems onthe planet (Jenkins et al 2013) Within the Neotropics
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253
54555657585960616263646566676869707172737475767778798081828384858687888990919293949596979899100101102103104105106
Correspondence to Antoine Fouquet E-mailfouquetantoinegmailcom Pedro Marinho E-mailpmarinho50gmailcom
ISSN 1477-2000 print 1478-0933 online The Trustees of the Natural History Museum London 2021 All Rights Reservedhttpsdxdoiorg1010801477200020211873869
Systematics and Biodiversity (2021) 0(0) 1ndash38
Author Query
AQ7 If your paper introduces new zoological taxa at family‐group level or below you are now required to register your paper with ZooBank and insert the generated ZooBank ID (LSID) here Individual new taxa need not be registered before publication this can be done subsequently should you wish Please go to httpwwwzoobankorgregister to complete this task You will need your article DOI to register After publication you must amend your ZooBank record of your paper to reflect the date of publication Please see httpwwwzoobankorghelp for further information
antoinefouquet
Note
Boana courtoisaezoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBEBoana euchariszoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
PROOF ONLY
Amazonia stands as a major biogeographic region cover-ing more than 65 million km2 including the largestcontinuous tract of tropical forest (40 of the worldrsquostropical rainforests) encompassing the most importantdrainage (Amazon River basin) and probably hostingthe highest continental biodiversity on earth (Myerset al 2000) The outstanding species richness of thisregion has raised many questions concerning its originsand diversification mechanisms While all agree on theastounding diversity of Amazonia our understanding ofthe actual number and distribution of species forming itsecosystems is in fact so incomplete (Ficetola et al2014 Meyer et al 2015 Vacher et al 2020) that ithas so far hampered investigations at the regional scaleConsequently the understanding of the historical causesand regional determinants of this outstanding diversityremain speculative (Antonelli et al 2018) Another con-sequence is that the collision between the Linnean short-fall and the biodiversity crisis is particularly evident inAmazonia (Guerra et al 2020) With almost the entireeastern and southern parts (25) of its extent beingalready deforested for agriculture an unknown propor-tion of endemic species and thus of the testimony ofAmazonian history has already vanished (Lovejoy ampNobre 2019)Anuran clades have repeatedly been documented to
harbour many unnamed species in Amazonia (Vacheret al 2020) and to display strikingly allopatric distribu-tion patterns that could provide crucial insights intoAmazoniarsquos past (Fouquet Cassini Haddad Pech ampRodrigues 2014 Rejaud et al 2020) This is notablythe case of the Boana albopunctata species group(Caminer amp Ron 2014 Funk et al 2012) This groupis only defined on the basis of molecular data(Faivovich et al 2005) and currently comprises 16valid nominal species distributed in the Caribbean andSouth America Boana albopunctata (Spix 1824)Boana alfaroi (Caminer amp Ron 2014) Boana almen-darizae (Caminer amp Ron 2014) Boana caiapo Pinheiroet al 2018 Boana calcarata (Troschel 1848) Boanadentei (Bokermann 1967) Boana fasciata (Geurounther1858) Boana heilprini (Noble 1923) Boana lancifor-mis (Cope 1871) Boana leucocheila (Caramaschi amp deNiemeyer 2003) Boana maculateralis (Caminer ampRon 2014) Boana multifasciata (Geurounther 1859)Boana paranaiba (Carvalho Giaretta amp Facure 2010)Boana steinbachi (Boulenger 1905) Boana tetete(Caminer amp Ron 2014) and Boana raniceps (Cope1862) Within this clade B heilprini is the most distinctsince it is confined to Hispaniola and is phylogeneticallydistant from a clade formed by all the other species ofthe group (Duellman Marion amp Hedges 2016) Wecan also distinguish B raniceps and the B albopunctata
clade (B lanciformis B albopunctata B multifasciataB paranaiba B caiapo B leucocheila) which occurwidely in the open habitats of the Cerrado andAmazonia (Camurugi et al 2021) from the Amazonianclade According to Caminer and Ron (2014) thisAmazonian clade is itself subdivided in two maingroups hereafter called the B calcarata (B fasciata Bcalcarata B almendarizae B maculateralis) and the Bsteinbachi (B tetete B alfaroi B steinbachi) cladesWithin this Amazonian clade Funk Caminer and
Ron (2012) identified no less than seven putativeunnamed species based on mitochondrial divergence andacoustic data These species add up to the 10 taxa thatwere valid at that time Caminer and Ron (2014) com-pleted this picture by documenting two additional line-ages describing four of the previously discoveredspecies and removing B steinbachi from its synonymywith B fasciata However sampling of both studies wascircumscribed to western Amazonia (mostly Ecuador)while the species complex occurs throughout AmazoniaPopulations from the eastern Guiana Shield lowlandsare already assumed to belong to a yet unnamed species(Caminer amp Ron 2014) but the status of numerousother populations throughout Amazonia generally iden-tified as B fasciata remain virtually unknown Morerecently Vacher et al (2020) suggested based on 16Smitochondrial DNA (mtDNA) sequences that up to 22species could exist in that group and that the distributionof most species could be circumscribed to small rangeswithin AmazoniaThese aforementioned studies improved our under-
standing of the actual diversity in the B calcaratastein-bachi clade but left virtually unexplored the temporaland spatial context of the diversification withinAmazonia The crown age of this Amazonian clade wasestimated between 16ndash11 million years ago (Ma) byFunk et al (2012) and between 14ndash9Ma by Duellmanet al (2016) Therefore the initial diversification of thatclade throughout Amazonia may have taken place dur-ing the final stage of the Pebas system a freshwaterlake occupying most of Western Amazonia from theearly Miocene (23Ma) until 10ndash9Ma (Hoorn et al2017) Meanwhile most of the diversification within theB calcaratasteinbachi clade took place during the lateNeogene (last 10Ma) From 9Ma onward this systemhas drained eastward into the Atlantic Ocean but enor-mous flooded ecosystems (Acre system) still occupied alarge portion of what we currently consider as WesternAmazonia until 7Ma (Hoorn et al 2010 Albert et al2018a) Subsequent river captures were common(Ruokolainen Moulatlet Zuquim Hoorn amp Tuomisto2018) as were climatic fluctuations that may havemodified vegetation possibly to the point of forest
107108109110111112113114115116117118119120121122123124125126127128129130131132133134135136137138139140141142143144145146147148149150151152153154155156157158159160
161162163164165166167168169170171172173174175176177178179180181182183184185186187188189190191192193194195196197198199200201202203204205206207208209210211212213214
2 A Fouquet et al
antoinefouquet
Texte inseacutereacute
mostly
PROOF ONLY
fragmentation at least in some peripheral parts ofAmazonia (Cheng et al 2013 Kirschner amp Hoorn2019) The relative roles of hydrological evolution(Albert et al 2018b Ribas Aleixo Nogueira Miyakiamp Cracraft 2012) and rainforest expansion and contrac-tion due to climate oscillations (Haffer 1969) have beenthe focus of intense debate (Leite amp Rogers 2013)Since the species of the B calcarata and B steinbachicomplexes are associated with riparian forests and thesespecies mostly breed in pools formed on the banks ofthe beds of small to medium-sized rivers very large riv-ers may act as barriers to dispersal thus reducing orcompletely impeding gene flow and inhibiting homogen-ization of differentiating populations (Naka et al 2018)However smaller and highly dynamic meandering riversprobably represent more permeable barriers Thereforewe assume that successive dispersals across majorAmazonian rivers (parapatric speciation) (Pirani et al2019) as well as hydrological changes (vicariant speci-ation) (Naka et al 2018) could have been major proc-esses of diversification in these frogsWe gathered a large mtDNA dataset for the B albo-
punctata group sampled throughout Amazonia combinedwith morphological and acoustic data for a subset ofthis group (the B steinbachi clade) to address two maingoals (1) reevaluate species boundaries (in or amongmembers) of the B albopunctata group and theirrespective distributions and (2) investigate its diversifi-cation history within Amazonia using mitogenomic datafor one terminal of each delimited species Furthermoretwo species in the B steinbachi clade that wereunnamed are described herein
Materials and methodsSpecies delimitationOur first objective was to delimit all major OperationalTaxonomic Units (OTUs) based on mtDNA main line-ages Our sampling included 55 new Boana tissue sam-ples obtained through fieldwork throughout Amazoniaand adjacent Dry Diagonal (DD Chaco CerradoCaatinga Werneck 2011) and loans from collaborators(from collections QCAZ MPEG INPA CORBIDIAAGUFU and from personal loans JMP SCF FTAAppendix 1) Samples unambiguously identified (neartype locality andor phenotypically corresponding to typematerial) as belonging to the currently recognized taxawere included except B paranaiba and B caiapo Wesequenced a 400 bp portion of the end of the 16S rDNAgene a locus commonly used for Neotropical amphibiantaxonomy and systematics (Vences Thomas Bonett ampVieites 2005) We also retrieved homologous sequences
from GenBank (389 accessions) In total we gathered444 16S sequences for this study (dataset details are pro-vided in Appendix 1 and DNA extraction and sequencingprotocols can be found in Appendix 2) These samplescover the whole distribution of the B albopunctata spe-cies group (14 included taxa) DNA sequence alignmentwas conducted on the MAFFT7 online server under theE-INS-i option with default parameters an algorithmdesigned for sequences with multiple conserved domainsand long gaps (Katoh et al 2019)We applied three DNA-based single-locus species
delimitation approaches (a) a distance-based methodthe Automated Barcode Gap Discovery (ABGDPuillandre Lambert Brouillet amp Achaz 2012) (b) amulti-rate coalescent based method the multi-ratePoisson Tree Processes model approach (mPTP Kapliet al 2017) and (c) a single-threshold coalescent-basedmethod the Generalized Mixed Yule Coalescentapproach (single threshold GMYC Monaghan et al2009 Pons et al 2006)The ABGD delimitation was performed with a prior
of intraspecific divergence P between 0001 and 01 aproxy for minimum relative gap width X of 1 and anumber of steps n equal to 30 We kept the partitionsuch that Pfrac14 0016 as it corresponds to the end of aplateau for group number and it matches thresholds ofintraspecific divergence proposed in other vertebratedelimitation studies using 16S barcodes (Puillandreet al 2012) For mPTP delimitation we first recon-structed a Maximum Likelihood (ML) tree withRAxML v824 (Stamatakis 2014) using theGTRthornGthorn I substitution model and estimated nodalsupport via 1000 parametric bootstraps We used nineoutgroups representing most other Boana species groups(Appendix 3) The mPTP delimitation was undertakenon the rooted ML tree with 5 million Markov chainMonte Carlo (MCMC) iterations sampling every10000th iteration and a 10 burn-in For the GMYCdelimitation we reconstructed a time-calibrated phyl-ogeny using BEAST 25 (Bouckaert et al 2014) Weused a birth-death population model to account forextinction processes and incomplete sampling Weincluded the same nine Boana outgroups used inRAxML reconstruction and five additional representa-tives of other Cophomantini genera (NesorohylaMyersiohyla Hyloscirtus AplastodiscusBokermannohyla) in order to include relationships thatcan be time-calibrated We used a single partition with aGTRthornGthorn I substitution model with an uncorrelatedrelaxed lognormal clock model of rate variation amongbranches (Drummond Ho Phillips amp Rambaut 2006)We used two calibration points the ages of the mostrecent common ancestor (MRCA) of Aplastodiscus and
215216217218219220221222223224225226227228229230231232233234235236237238239240241242243244245246247248249250251252253254255256257258259260261262263264265266267268
269270271272273274275276277278279280281282283284285286287288289290291292293294295296297298299300301302303304305306307308309310311312313314315316317318319320321322
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 3
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13536302v1
PROOF ONLY
Boana as well as the MRCA of a clade formed byHyloscirtus Boana Aplastodiscus andBokermannohyla which were estimated by Feng et al(2017) and consequently constrained here assuming nor-mal prior distributions of 252Ma (standard deviation[SD]frac14 28Ma) and 323Ma (SD frac14 3Ma) respectivelyWe considered these calibrations to represent the bestprior because Feng et al (2017) included a comprehen-sive dataset of nuclear loci for an extensive sampling ofanurans and used many reliable fossils as calibrationpoints Moreover Hime et al (2020) analysing an evenlarger genomic dataset found very similar time esti-mates for the TMRCA of BoanathornHyloscirtus Insteadusing fossil andor biogeographic calibrations wouldhave implied to expand the matrix to lineages distantlyrelated to Boana and would likely lead to an overesti-mation of calibration dates with our mitogenomic data-set (Molak amp Ho 2015 Papadopoulou et al 2010) Forthe MCMC parameters we used four independentchains of 100 million iterations recording every10000th iteration and a 10 burn-in We combined thelog files of the independent runs using LogCombiner25 (Drummond amp Rambaut 2007) and checked theconvergence of our parameters confirmed by allEffective Sample Size (ESS) being above 200 Then weextracted the maximum clade credibility tree using Treeannotator 25 (Drummond amp Rambaut 2007) with aburn-in of 10 After removing outgroups we per-formed a GMYC delimitation on the ultrametric treeusing the GMYC function of the splits R package v10-11 (Ezard Fujisawa amp Barraclough 2009) with athreshold interval between 0 and 10Ma and by usingthe single threshold method Operational TaxonomicUnits (OTUs) were defined using a majority-rule con-sensus from the results of the three methods ie a lin-eage is considered as being an OTU if supported by atleast two of the three methodsAll but five occurrence records were georeferenced
and used to create distribution maps with convex poly-gons under QGIS 214 (QGIS Geographic InformationSystem) OTUs were assigned to taxa based on thefieldmuseum identification and sometimes corrected inaccordance with type localities and known distribution(Frost 2019 IUCN 2020)
Morphological and acoustic variation in theBoana steinbachi cladeThe following morphometric measurements were takenon 38 males and 9 females snout-vent length (SVL)head length (HL) head width (HW) eye diameter (ED)tympanum diameter (TD) tibia length (TL) foot length(FL) thigh length (THL) and calcaneal appendage
length (CL) In addition to these we also measuredhand length (HAL) forearm length (FLL) and eyendashnos-tril distance (EN) for the morphometric characterizationof holotypes All measurements followed the definitionsand terminology of Watters Cummings Flanagan andSiler (2016) We classified the calcaneal appendage intothree-character states (i) a calcar (skin appendage withlength gt 1mm flattened or conical) a (ii) skin flap(skin appendage with length lt 1mm flattened or a(iii) tubercle a conical structure with reduced size andas such not forming an appendage coming out of theheel The specimens were measured by P Marinho(Alta Floresta Mato Grosso State and Assis BrazilAcre State) using a Mitutoyo digital calliper (to thenearest 005mm) by MT Rodrigues (Jirau and PacaasNovos Rondonia State) using a Mitutoyo digital calliper(to the nearest 001mm) by A Fouquet (French Guianaand Suriname) using a DigiMax digital calliper (nearest001mm) and by M Caminer (Ecuador Peru andBolivia) using a Mannesmann digital calliper (nearest001mm) Snout shape was assessed according to Heyeret al (1990) Digital webbing formulae followed thenotation system of Savage and Heyer (1997)Acoustic analysis of 27 recorded calling males
(Appendix 4) was conducted in Raven Pro 15 64-bit ver-sion (Bioacoustics Research Program 2012) sound figureswere produced using Seewave version 210 (SueurAubin amp Simonis 2008) and tuneR version 132(Ligges Krey Mersmann amp Schnackenberg 2014) in Rversion 350 (R Core Team 2018) Raven Pro settingswindow size frac14 512 samples window typefrac14Hann 3 dBfilter bandwidth frac14 124Hz window overlap frac14 85 hopsize frac14 77ms discrete Fourier transform (DFT) size frac141024 samples grid spacing frac14 431Hz seewave settingswindow typefrac14Hanning fast Fourier transform (FFT)size frac14 256 samples FFT overlap frac14 90 The analysedfiles are deposited in the sound collection of the lsquoMuseude Biodiversidade do Cerradorsquo (AAG-UFU) lsquoFonotecaNeotropical Jacques Vielardirsquo (FNJV) lsquoLa Sonotheque duMuseum National dHistoire Naturellersquo (MNHN) and theMacaulay library (MC) Detailed information about theanalysed sound files and accession numbers are providedin Appendix 4Call types were distinguished based on the number of
notes per call and pulsing We defined as type 1 callsthe multinote calls formed by nonpulsed notes whichconsist of the main call type emitted by males of allstudied species Type 2 calls are emitted less often thantype 1 and consist of a one-note call formed by poorlydefined pulses We only included type 1 calls in theacoustic diagnoses and the interspecific comparisonsThe acoustic traits were generally analysed manually (ifnot stated otherwise) as follows temporal traits (call
323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370371372373374375376
377378379380381382383384385386387388389390391392393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430
4 A Fouquet et al
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
1
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics 2014
antoinefouquet
Texte inseacutereacute
a
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
s
PROOF ONLY
duration note duration interval between notes numberof notes and call rise time the latter using the lsquoPeakTimersquo function) frequency traits (dominant frequencyusing the lsquoPeak Frequencyrsquo function minimum andmaximum frequency using the lsquoFrequency 5rsquo andlsquoFrequency 95rsquo functions respectively) Call traitsused in the description followed the definitions and ter-minology of Keuroohler et al (2017) using a note-cen-tred approach
Time-calibrated species phylogenyWe selected a representative for each of the 25 OTUsidentified by the species delimitation (see Results) forcomplete mitogenome sequencing to investigate interre-lationships and divergence times Mitogenomic sequen-ces were obtained using low-coverage shotgunsequencing We recovered high-quality mitochondrialgenome assemblies for 18 OTU representatives (seeAppendix 2 for details regarding mitogenome sequenc-ing assembling and annotation) For the remainingseven OTUs for which we could not obtain tissue sam-ples (Appendix 3) we gathered all the available mito-chondrial loci (12S 16S ND1 COI and Cytb) fromGenBank four of these OTUs were represented by 16Sonly We also selected 14 species as outgroups (ninewith complete mitogenomes) including representativesfor other Boana species groups and most otherCophomantini genera (Appendix 3)We extracted 12S 16S and all protein-coding
sequence regions (CDS thus removing D-Loop andtRNAs) from complete mitogenomes as well asGenBank accessions and aligned each locus independ-ently using the MAFFT7 online server For rRNAgenes we chose the E-INS-i strategy recommended forsequences with multiple conserved domains and longgaps For the protein-coding genes we chose the G-INS-i strategy designed for sequences with global hom-ology (Katoh et al 2019) Realignments of CDS con-sidering the reading frame were done and concatenatedin Geneious v918 (httpswwwgeneiouscom) Ourfinal matrix totalled 39 terminals and 13620 alignednucleotide sites Among these terminals 27 were com-plete and nine had fewer than 3000 nucleotides (20)but all were putatively closely related to terminals withcomplete dataWe selected the best-fit partition scheme and model
of evolution for each partition using PartitionFinderV211 (Lanfear et al 2016) based on the BayesianInformation Criterion (BIC) We predefined four datablocks one for rRNA genes (12S and 16S) and one foreach codon position of all CDS regions and found theGTRthorn IthornG model to best-fit all partitions We
reconstructed a time-calibrated tree using a birth-deathtree prior using BEAST 25 to account for extinctionprocesses and incomplete sampling We parameterizedunlinked substitution models according to the estimatesobtained in the PartitionFinder V211 analysis (Lanfearet al 2016) We used the same two calibration pointsas in the previous BEAST analysis Analyses wereundertaken using uncorrelated relaxed lognormal clockmodel of distribution of rate variation among branchesfor each partition (Drummond et al 2006) The Markovchain Monte Carlo (MCMC) parameters were set withfour independent chains of 100 million iterations stor-ing every 10000th iteration and a 10 burn-in Wecombined the log files of the independent runs usingLogCombiner 25 and confirmed the convergence of ourparameters as all ESS were above 200 Finally weextracted the maximum clade credibility tree using Treeannotator 25 We acknowledge that our phylogeneticreconstruction solely based on mtDNA sequences priori-tizes spatial and taxonomic completeness over genomiccoverage and can lead to overestimated divergence times(eg McCormack Heled Delaney Peterson ampKnowles 2011 Near et al 2012)
Biogeographic analysisBiogeographic inferences were undertaken on the time-calibrated phylogeny using the BioGeoBEARS R pack-age (Matzke 2013) This package reconstructs ancestralgeographic distributions and investigates the role ofeach biogeographic event with a maximum likelihoodalgorithm We compared three different models (i) alikelihood version of the Dispersal-Vicariance(DIVALIKE) model (Ronquist 1997) (ii) a likelihoodversion of the BayArea (BBM) model (Landis et al2013) and (iii) the Dispersal Extinction Cladogenesis(DEC) model (Ree amp Smith 2008) We also comparedversions of these models allowing jump dispersal asdescribed by the J parameter (Matzke 2013) Modelswere compared with the Akaike Information Criterion(AIC) We ran 50 independent BioGeoBEARS biogeo-graphic stochastic mapping to determine biogeographicevent counts for the best-fit model (Dupin et al 2017)Ree and Sanmartın (2018) voiced several criticismsagainst the use of J parameter arguing that it inflates thecontribution of cladogenetic events to the likelihoodand minimizes the contribution of anagenetic time-dependent range evolution We thus also compared theresults from the best model without jump dispersalSince this work focuses on the historical biogeog-
raphy of the Amazonian clade we discarded the twoearly diverging lineages B heilprini and B ranicepsfrom the biogeographic analyses since their distribution
431432433434435436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484
485486487488489490491492493494495496497498499500501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 5
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13560371v1)
antoinefouquet
Texte inseacutereacute
D
PROOF ONLY
ranges lie mostly outside of Amazonia We consideredthree areas within Amazonia Western Amazonia theBrazilian Shield and the Guiana Shield These areascorrespond to major geological features roughly delim-ited by modern riverine barriers the Madeira River theNegro River and the lower course of the Amazon Riverand to three large biogeographic regions known asWallacersquos districts (Hoorn et al 2010 Wallace 1854)These districts were recently confirmed as major breaksin birdsrsquo species composition (Oliveira Vasconcelos ampSantos 2017) and amphibians (Godinho amp da Silva2018 Vacher et al 2020) strengthening their status ofbiogeographic regions Because the distribution range ofthe B albopunctata group extends outside Amazoniawe included in the ancestral range reconstruction oneadditional non-Amazonian Neotropical area theDry Diagonal
ResultsSpecies delimitationOf the three tested methods of species delimitation theABGD method was found to be the most conservativedelimiting 21 OTUs while mPTP and GMYC delimited24 and 47 OTUs respectively (Appendix 1) In onecase the consensual partitioning was overly conserva-tive given that B leucocheila and B multifasciata wereconsidered as a single OTU which is contradicted bythe fact that these two taxa are phenotypically distinctTherefore we kept the different lineages delimited byGMYC in this group as distinct OTUs The final delimi-tation led to 25 OTUs (including B heilprini) in the Balbopunctata species group (Fig 1 Appendix 1)Thirteen of these OTUs could be linked to nominal spe-cies (except B caiapo and B paranaiba that were notincluded in the analysis because material was not avail-able) Conversely 11 could not be linked to any nom-inal species representing a possible 44 increase in thespecies richness of the group The Amazonian cladeitself was represented by 14 OTUs including eight nom-inal species and six putative new species (Fig 1) Theseresults also imply important changes to the geographicdistribution of the species of the group In Appendix 5we detail and justify the identification of these OTUsand their respective geographic ranges
Taxonomic accountsThe Amazonian clade of the B albopunctata group pre-viously reported as the B calcaratandashB fasciata speciescomplex by Caminer and Ron (2014) is composed oftwo major clades (the B calcarata and the B steinbachi
clades) defined by the following combination of charac-ter states (1) truncate snout in dorsal view (2) roundedhead shape and (3) presence of calcaneal appendageon heelThe B calcarata clade (Fig 1) is composed of four
species possessing a calcar and having advertisementcalls formed by a single type of call (Caminer amp Ron2014) The B steinbachi clade on the other handincludes three described and two undescribed species(Fig 1) Members of this clade lack a calcar (skin flapor tubercle present or the complete absence of calcanealappendages) and have vocal repertoires composed ofmore than one type of call except for B alfaroi forwhich a single type of call has been documented so far(Caminer amp Ron 2014)Hyla steinbachi Boulenger 1905 currently valid as
Boana steinbachi (Boulenger 1905) was describedfrom the Bolivian Province Sara Departament SantaCruz de la Sierra Bolivia We assume that Buenavistais the type locality of B steinbachi Between 1910 and1950 the Steinbach family collected many amphibiansand reptiles in Bolivia mainly at Buenavista in theDepartment of Santa Cruz (see Parker 1927) which isthe type locality of several anurans (Hamptophryneboliviana (Parker 1927) Pseudopaludicola bolivianaParker 1927 Scinax parkeri (Gaige 1929)) and thesnake Apostolepis tenuis Ruthven 1927 The materialcollected by the Steinbach family is on display in vari-ous museums such as the University of MichiganMuseum of Zoology (UMMZ see Gaige 1929) theNatural History Museum London (BMNH see Parker1927) the Zoologisches Museum Berlin (ZMB seeMeurouller 1924) and the Naturhistorisches Museum Basel(NBM pers comm Weurouest) Many decades later De laRiva (1990) synonymized H steinbachi with H fas-ciata Subsequently Jansen Bloch Schulze andPfenninger (2011) found differences in call and mtDNAdata between populations of B fasciata from Boliviaand Ecuador and suggested that their results imply a res-urrection of B steinbachi In the following yearsCaminer and Ron (2014) tentatively assigned B steinba-chi to a genetic lineage (clade J) inhabiting BolivianAmazonia near its type locality By assessing its phylo-genetic relationships in the group Caminer and Ron(2014) formally revalidated B steinbachi as a distinctspecies from nominal B fasciata The revalidation of Bsteinbachi was also supported by phenotypic dataobtained from syntypes but did not include a reexamina-tion of all diagnostic characters of the speciesNext we present an amended diagnosis of B steinba-
chi based on novel data on life colours and morphologyand describe its vocal repertoire based on topotypesfrom Bolivia and distinct OTUs from Peru and Brazil
539540541542543544545546547548549550551552553554555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592
593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644645646
6 A Fouquet et al
PROOF ONLY
647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695696697698699700
701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754
Fig 1 Subtrees of the three main clades from the chronogram obtained from the analysis of 16S sequences using BEAST2Terminals are collapsed according to the OTU recovered from the species delineation analysis The distribution of each of these OTUis depicted on the maps
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 7
PROOF ONLY
also assigned to this species in this study Specimensand calls from Acre (ldquoB aff steinbachi 1rdquo) and callsfrom the lower Madre de Dios River (ldquoB aff steinbachi2rdquo) were included in the variation of B steinbachi
Boana steinbachi (Boulenger 1905)Hyla steinbachi mdash Boulenger 1905
Hyla fasciata De la Riva 1990Hypsiboas fasciatus Jansen et al 2011
Hypsiboas sp (Clade G) Funk et al 2012Hypsiboas steinbachi Caminer amp Ron 2014
Boana steinbachi Dubois 2017Boana sp (Clade J) Meza-Joya et al 2019
Boana fasciata Vacher et al 2020
Syntypes BMNH 194721361ndash63 two adults ofunknown sex and one juvenile respectively from Saraprovince Department of Santa Cruz de La SierraBolivia Collected by Hf J Steinbach
Diagnosis Boana steinbachi is characterized by the fol-lowing combination of character states (1) skin flap onheel (2) vocal repertoire composed of more than onecall type (3) multinote call and (4) regular internoteintervals (Figs 2 3 amp 4)
Comparisons with congeners of the Amazonian cladeBoana steinbachi can be distinguished from the membersof the B calcarata clade (B almendarizae B fasciata Bcalcarata and B maculateralis) by the absence of a cal-car and by the vocal repertoire composed of more thanone type of call Among members of the B steinbachiclade it can be distinguished from B dentei B alfaroiand B tetete by the presence of a skin flap on heel (tuber-cle in B alfaroi and B tetete tubercle on one side orcompletely absent in B dentei) by its multinote call (one-note call in B dentei and B tetete) by having regularinternote intervals between call notes (notes with irregularintervals sometimes partly fused one with the next in Balfaroi) and by the vocal repertoire composed of two dis-tinct types of calls (one call type in B alfaroi) (Camineramp Ron 2014 Marinho et al 2020)
Variation Body size varies between 304 and 374mmin males and between 425 and 488mm in females(Appendix 7) Individual AAG-UFU 5918 lacks the heelskin flap on the left side topotypes SMF 88394ndash95 andSMF 88397 lack the skin flap on both sides In life(Fig 3a b) dorsal colouration is beige light brownbright yellow or orange brown in calling males withfaint brown transversal bands and dark purplish-brownfaint brown or grey longitudinal lines extending from
the snout to vent a second line extending from behindthe eye to pelvic region and a third line as a canthalstripe extending from the posterior corner of the nostrilto anterior corner of the eye and from the posterior cor-ner of the eye to midbody length on flank (the topotypesSMF 88394ndash97 lack transversal bands on dorsum indi-viduals SMF 88394 and CORBIDI 13414 lack longitu-dinal lines on dorsum and the middorsal line isextending from snout to interocular region AAG-UFU5921 lacks longitudinal lines on dorsum) Iris cream orgrey sometimes with yellow pigmentation on the upperpart of the iris Throat varying from white to yellowchest and anterior belly varying from white to creamVentral surface of hand and hind limb beige to bright
755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808
809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862
Fig 2 Adult male of Boana steinbachi (AAG-UFU 5921)from the Assis Brasil population (Acre Brazil) (A) dorsal and(B) ventral views of the body (SVL frac14 337mm) (C) dorsaland (D) lateral views of the head (HL frac14123mm HW frac14102mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 106mm) and (G) sole of thefoot (FL frac14 139mm)
8 A Fouquet et al
antoinefouquet
Texte surligneacute
we may wanna ask for full page here no
PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Research Article
Systematics and biogeography of the Boana albopunctataspecies group (Anura Hylidae) with the descriptionof two new species from Amazonia
ANTOINE FOUQUET1 PEDRO MARINHO2 ALEXANDRE REacuteJAUD1 THIAGO R CARVALHO3 MARCEL A CAMINER45 MARTIN JANSEN6 RAIacuteSSA N RAINHA7 MIGUEL T RODRIGUES8 FERNANDA P WERNECK7 ALBERTINA P LIMA7 TOMAS HRBEK9 ARIOVALDO A GIARETTA2 PABLO J VENEGAS10 GERMAacuteN CHAacuteVEZ10 amp SANTIAGO RON4
1Laboratoire Evolution et Diversite Biologique UMR 5174 CNRS IRD Universite Paul Sabatier Batiment 4R1 31062cedex 9 118 Route de Narbonne Toulouse 31077 France2Laboratorio de Anuros Neotropicais Instituto de Ciencias Exatas e Naturais do Pontal Universidade Federal de UberlandiaItuiutaba MG Brazil3Laboratorio de Herpetologia Departamento de Biodiversidade e Centro de Aquicultura IB Universidade Estadual PaulistaRio Claro SP Brazil4Museo de Zoologıa Escuela de Biologıa Pontificia Universidad Catolica del Ecuador Quito Ecuador5Institute of Organismic and Molecular Evolution Johannes Gutenberg University Mainz Germany6Department of Terrestrial Zoology Research Institute and Nature Museum Senckenberg Frankfurt Germany7Instituto Nacional de Pesquisas da Amazonia Coordenac~ao de Biodiversidade Manaus AM Brazil8Departamento de Zoologia Universidade de S~ao Paulo Instituto de Biociencias S~ao Paulo SP Brazil9Departamento de Genetica Universidade Federal do Amazonas Manaus AM Brazil10Instituto Peruano de Herpetologıa Lima Peru
The outstanding species richness of Amazonia has fascinated biologists for centuries However the records of actualnumbers and distribution of species forming its ecosystems are so incomplete that the understanding of the historicalcauses and regional determinants of this diversity remain speculative Anuran clades have repeatedly been documentedto harbour many unnamed species in this region notably the Boana albopunctata species group Considering thedocumented distribution and the ecology of the species of that group we hypothesized that it diversified via successivetrans-riverine dispersals during the late Miocene and Pliocene after the formation of the modern Amazon watershedTo test this hypothesis we gathered an extensive dataset of 16S rDNA sequences sampled throughout Amazonia anda mitogenomic dataset representative of the diversity of the clade to (1) re-evaluate species boundaries and distributionsand (2) infer the spatio-temporal history of diversification within Amazonia We delimited 14 Operational TaxonomicUnits (OTUs) in an Amazonian clade ie 75 higher than currently recognized (14 OTUs for eight described species)Combining molecular data with morphological and acoustic data two new species Boana courtoisae sp nov fromthe eastern Guiana Shield and Boana eucharis sp nov from Southern Amazonia are described herein These speciesbelong to a clade that diversified throughout Amazonia during the last 10Ma thus more recently than co-distributedsmall terrestrial anurans but concomitantly with other more vagile vertebrates Our time-scaled phylogeny andbiogeographic analyses suggest an initial east-west divergence and confirm reciprocal trans-riverine dispersals duringthe last 5Ma The geomorphological evolution of the region and species-specific dispersal ability largely explain thesedistinct spatio-temporal patterns across anuransQ7
Key words Amphibia biodiversity Guiana Shield mitogenomics neotropics phylogenetics
IntroductionThe Neotropics harbour the most diverse ecosystems onthe planet (Jenkins et al 2013) Within the Neotropics
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253
54555657585960616263646566676869707172737475767778798081828384858687888990919293949596979899100101102103104105106
Correspondence to Antoine Fouquet E-mailfouquetantoinegmailcom Pedro Marinho E-mailpmarinho50gmailcom
ISSN 1477-2000 print 1478-0933 online The Trustees of the Natural History Museum London 2021 All Rights Reservedhttpsdxdoiorg1010801477200020211873869
Systematics and Biodiversity (2021) 0(0) 1ndash38
Author Query
AQ7 If your paper introduces new zoological taxa at family‐group level or below you are now required to register your paper with ZooBank and insert the generated ZooBank ID (LSID) here Individual new taxa need not be registered before publication this can be done subsequently should you wish Please go to httpwwwzoobankorgregister to complete this task You will need your article DOI to register After publication you must amend your ZooBank record of your paper to reflect the date of publication Please see httpwwwzoobankorghelp for further information
antoinefouquet
Note
Boana courtoisaezoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBEBoana euchariszoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
PROOF ONLY
Amazonia stands as a major biogeographic region cover-ing more than 65 million km2 including the largestcontinuous tract of tropical forest (40 of the worldrsquostropical rainforests) encompassing the most importantdrainage (Amazon River basin) and probably hostingthe highest continental biodiversity on earth (Myerset al 2000) The outstanding species richness of thisregion has raised many questions concerning its originsand diversification mechanisms While all agree on theastounding diversity of Amazonia our understanding ofthe actual number and distribution of species forming itsecosystems is in fact so incomplete (Ficetola et al2014 Meyer et al 2015 Vacher et al 2020) that ithas so far hampered investigations at the regional scaleConsequently the understanding of the historical causesand regional determinants of this outstanding diversityremain speculative (Antonelli et al 2018) Another con-sequence is that the collision between the Linnean short-fall and the biodiversity crisis is particularly evident inAmazonia (Guerra et al 2020) With almost the entireeastern and southern parts (25) of its extent beingalready deforested for agriculture an unknown propor-tion of endemic species and thus of the testimony ofAmazonian history has already vanished (Lovejoy ampNobre 2019)Anuran clades have repeatedly been documented to
harbour many unnamed species in Amazonia (Vacheret al 2020) and to display strikingly allopatric distribu-tion patterns that could provide crucial insights intoAmazoniarsquos past (Fouquet Cassini Haddad Pech ampRodrigues 2014 Rejaud et al 2020) This is notablythe case of the Boana albopunctata species group(Caminer amp Ron 2014 Funk et al 2012) This groupis only defined on the basis of molecular data(Faivovich et al 2005) and currently comprises 16valid nominal species distributed in the Caribbean andSouth America Boana albopunctata (Spix 1824)Boana alfaroi (Caminer amp Ron 2014) Boana almen-darizae (Caminer amp Ron 2014) Boana caiapo Pinheiroet al 2018 Boana calcarata (Troschel 1848) Boanadentei (Bokermann 1967) Boana fasciata (Geurounther1858) Boana heilprini (Noble 1923) Boana lancifor-mis (Cope 1871) Boana leucocheila (Caramaschi amp deNiemeyer 2003) Boana maculateralis (Caminer ampRon 2014) Boana multifasciata (Geurounther 1859)Boana paranaiba (Carvalho Giaretta amp Facure 2010)Boana steinbachi (Boulenger 1905) Boana tetete(Caminer amp Ron 2014) and Boana raniceps (Cope1862) Within this clade B heilprini is the most distinctsince it is confined to Hispaniola and is phylogeneticallydistant from a clade formed by all the other species ofthe group (Duellman Marion amp Hedges 2016) Wecan also distinguish B raniceps and the B albopunctata
clade (B lanciformis B albopunctata B multifasciataB paranaiba B caiapo B leucocheila) which occurwidely in the open habitats of the Cerrado andAmazonia (Camurugi et al 2021) from the Amazonianclade According to Caminer and Ron (2014) thisAmazonian clade is itself subdivided in two maingroups hereafter called the B calcarata (B fasciata Bcalcarata B almendarizae B maculateralis) and the Bsteinbachi (B tetete B alfaroi B steinbachi) cladesWithin this Amazonian clade Funk Caminer and
Ron (2012) identified no less than seven putativeunnamed species based on mitochondrial divergence andacoustic data These species add up to the 10 taxa thatwere valid at that time Caminer and Ron (2014) com-pleted this picture by documenting two additional line-ages describing four of the previously discoveredspecies and removing B steinbachi from its synonymywith B fasciata However sampling of both studies wascircumscribed to western Amazonia (mostly Ecuador)while the species complex occurs throughout AmazoniaPopulations from the eastern Guiana Shield lowlandsare already assumed to belong to a yet unnamed species(Caminer amp Ron 2014) but the status of numerousother populations throughout Amazonia generally iden-tified as B fasciata remain virtually unknown Morerecently Vacher et al (2020) suggested based on 16Smitochondrial DNA (mtDNA) sequences that up to 22species could exist in that group and that the distributionof most species could be circumscribed to small rangeswithin AmazoniaThese aforementioned studies improved our under-
standing of the actual diversity in the B calcaratastein-bachi clade but left virtually unexplored the temporaland spatial context of the diversification withinAmazonia The crown age of this Amazonian clade wasestimated between 16ndash11 million years ago (Ma) byFunk et al (2012) and between 14ndash9Ma by Duellmanet al (2016) Therefore the initial diversification of thatclade throughout Amazonia may have taken place dur-ing the final stage of the Pebas system a freshwaterlake occupying most of Western Amazonia from theearly Miocene (23Ma) until 10ndash9Ma (Hoorn et al2017) Meanwhile most of the diversification within theB calcaratasteinbachi clade took place during the lateNeogene (last 10Ma) From 9Ma onward this systemhas drained eastward into the Atlantic Ocean but enor-mous flooded ecosystems (Acre system) still occupied alarge portion of what we currently consider as WesternAmazonia until 7Ma (Hoorn et al 2010 Albert et al2018a) Subsequent river captures were common(Ruokolainen Moulatlet Zuquim Hoorn amp Tuomisto2018) as were climatic fluctuations that may havemodified vegetation possibly to the point of forest
107108109110111112113114115116117118119120121122123124125126127128129130131132133134135136137138139140141142143144145146147148149150151152153154155156157158159160
161162163164165166167168169170171172173174175176177178179180181182183184185186187188189190191192193194195196197198199200201202203204205206207208209210211212213214
2 A Fouquet et al
antoinefouquet
Texte inseacutereacute
mostly
PROOF ONLY
fragmentation at least in some peripheral parts ofAmazonia (Cheng et al 2013 Kirschner amp Hoorn2019) The relative roles of hydrological evolution(Albert et al 2018b Ribas Aleixo Nogueira Miyakiamp Cracraft 2012) and rainforest expansion and contrac-tion due to climate oscillations (Haffer 1969) have beenthe focus of intense debate (Leite amp Rogers 2013)Since the species of the B calcarata and B steinbachicomplexes are associated with riparian forests and thesespecies mostly breed in pools formed on the banks ofthe beds of small to medium-sized rivers very large riv-ers may act as barriers to dispersal thus reducing orcompletely impeding gene flow and inhibiting homogen-ization of differentiating populations (Naka et al 2018)However smaller and highly dynamic meandering riversprobably represent more permeable barriers Thereforewe assume that successive dispersals across majorAmazonian rivers (parapatric speciation) (Pirani et al2019) as well as hydrological changes (vicariant speci-ation) (Naka et al 2018) could have been major proc-esses of diversification in these frogsWe gathered a large mtDNA dataset for the B albo-
punctata group sampled throughout Amazonia combinedwith morphological and acoustic data for a subset ofthis group (the B steinbachi clade) to address two maingoals (1) reevaluate species boundaries (in or amongmembers) of the B albopunctata group and theirrespective distributions and (2) investigate its diversifi-cation history within Amazonia using mitogenomic datafor one terminal of each delimited species Furthermoretwo species in the B steinbachi clade that wereunnamed are described herein
Materials and methodsSpecies delimitationOur first objective was to delimit all major OperationalTaxonomic Units (OTUs) based on mtDNA main line-ages Our sampling included 55 new Boana tissue sam-ples obtained through fieldwork throughout Amazoniaand adjacent Dry Diagonal (DD Chaco CerradoCaatinga Werneck 2011) and loans from collaborators(from collections QCAZ MPEG INPA CORBIDIAAGUFU and from personal loans JMP SCF FTAAppendix 1) Samples unambiguously identified (neartype locality andor phenotypically corresponding to typematerial) as belonging to the currently recognized taxawere included except B paranaiba and B caiapo Wesequenced a 400 bp portion of the end of the 16S rDNAgene a locus commonly used for Neotropical amphibiantaxonomy and systematics (Vences Thomas Bonett ampVieites 2005) We also retrieved homologous sequences
from GenBank (389 accessions) In total we gathered444 16S sequences for this study (dataset details are pro-vided in Appendix 1 and DNA extraction and sequencingprotocols can be found in Appendix 2) These samplescover the whole distribution of the B albopunctata spe-cies group (14 included taxa) DNA sequence alignmentwas conducted on the MAFFT7 online server under theE-INS-i option with default parameters an algorithmdesigned for sequences with multiple conserved domainsand long gaps (Katoh et al 2019)We applied three DNA-based single-locus species
delimitation approaches (a) a distance-based methodthe Automated Barcode Gap Discovery (ABGDPuillandre Lambert Brouillet amp Achaz 2012) (b) amulti-rate coalescent based method the multi-ratePoisson Tree Processes model approach (mPTP Kapliet al 2017) and (c) a single-threshold coalescent-basedmethod the Generalized Mixed Yule Coalescentapproach (single threshold GMYC Monaghan et al2009 Pons et al 2006)The ABGD delimitation was performed with a prior
of intraspecific divergence P between 0001 and 01 aproxy for minimum relative gap width X of 1 and anumber of steps n equal to 30 We kept the partitionsuch that Pfrac14 0016 as it corresponds to the end of aplateau for group number and it matches thresholds ofintraspecific divergence proposed in other vertebratedelimitation studies using 16S barcodes (Puillandreet al 2012) For mPTP delimitation we first recon-structed a Maximum Likelihood (ML) tree withRAxML v824 (Stamatakis 2014) using theGTRthornGthorn I substitution model and estimated nodalsupport via 1000 parametric bootstraps We used nineoutgroups representing most other Boana species groups(Appendix 3) The mPTP delimitation was undertakenon the rooted ML tree with 5 million Markov chainMonte Carlo (MCMC) iterations sampling every10000th iteration and a 10 burn-in For the GMYCdelimitation we reconstructed a time-calibrated phyl-ogeny using BEAST 25 (Bouckaert et al 2014) Weused a birth-death population model to account forextinction processes and incomplete sampling Weincluded the same nine Boana outgroups used inRAxML reconstruction and five additional representa-tives of other Cophomantini genera (NesorohylaMyersiohyla Hyloscirtus AplastodiscusBokermannohyla) in order to include relationships thatcan be time-calibrated We used a single partition with aGTRthornGthorn I substitution model with an uncorrelatedrelaxed lognormal clock model of rate variation amongbranches (Drummond Ho Phillips amp Rambaut 2006)We used two calibration points the ages of the mostrecent common ancestor (MRCA) of Aplastodiscus and
215216217218219220221222223224225226227228229230231232233234235236237238239240241242243244245246247248249250251252253254255256257258259260261262263264265266267268
269270271272273274275276277278279280281282283284285286287288289290291292293294295296297298299300301302303304305306307308309310311312313314315316317318319320321322
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 3
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13536302v1
PROOF ONLY
Boana as well as the MRCA of a clade formed byHyloscirtus Boana Aplastodiscus andBokermannohyla which were estimated by Feng et al(2017) and consequently constrained here assuming nor-mal prior distributions of 252Ma (standard deviation[SD]frac14 28Ma) and 323Ma (SD frac14 3Ma) respectivelyWe considered these calibrations to represent the bestprior because Feng et al (2017) included a comprehen-sive dataset of nuclear loci for an extensive sampling ofanurans and used many reliable fossils as calibrationpoints Moreover Hime et al (2020) analysing an evenlarger genomic dataset found very similar time esti-mates for the TMRCA of BoanathornHyloscirtus Insteadusing fossil andor biogeographic calibrations wouldhave implied to expand the matrix to lineages distantlyrelated to Boana and would likely lead to an overesti-mation of calibration dates with our mitogenomic data-set (Molak amp Ho 2015 Papadopoulou et al 2010) Forthe MCMC parameters we used four independentchains of 100 million iterations recording every10000th iteration and a 10 burn-in We combined thelog files of the independent runs using LogCombiner25 (Drummond amp Rambaut 2007) and checked theconvergence of our parameters confirmed by allEffective Sample Size (ESS) being above 200 Then weextracted the maximum clade credibility tree using Treeannotator 25 (Drummond amp Rambaut 2007) with aburn-in of 10 After removing outgroups we per-formed a GMYC delimitation on the ultrametric treeusing the GMYC function of the splits R package v10-11 (Ezard Fujisawa amp Barraclough 2009) with athreshold interval between 0 and 10Ma and by usingthe single threshold method Operational TaxonomicUnits (OTUs) were defined using a majority-rule con-sensus from the results of the three methods ie a lin-eage is considered as being an OTU if supported by atleast two of the three methodsAll but five occurrence records were georeferenced
and used to create distribution maps with convex poly-gons under QGIS 214 (QGIS Geographic InformationSystem) OTUs were assigned to taxa based on thefieldmuseum identification and sometimes corrected inaccordance with type localities and known distribution(Frost 2019 IUCN 2020)
Morphological and acoustic variation in theBoana steinbachi cladeThe following morphometric measurements were takenon 38 males and 9 females snout-vent length (SVL)head length (HL) head width (HW) eye diameter (ED)tympanum diameter (TD) tibia length (TL) foot length(FL) thigh length (THL) and calcaneal appendage
length (CL) In addition to these we also measuredhand length (HAL) forearm length (FLL) and eyendashnos-tril distance (EN) for the morphometric characterizationof holotypes All measurements followed the definitionsand terminology of Watters Cummings Flanagan andSiler (2016) We classified the calcaneal appendage intothree-character states (i) a calcar (skin appendage withlength gt 1mm flattened or conical) a (ii) skin flap(skin appendage with length lt 1mm flattened or a(iii) tubercle a conical structure with reduced size andas such not forming an appendage coming out of theheel The specimens were measured by P Marinho(Alta Floresta Mato Grosso State and Assis BrazilAcre State) using a Mitutoyo digital calliper (to thenearest 005mm) by MT Rodrigues (Jirau and PacaasNovos Rondonia State) using a Mitutoyo digital calliper(to the nearest 001mm) by A Fouquet (French Guianaand Suriname) using a DigiMax digital calliper (nearest001mm) and by M Caminer (Ecuador Peru andBolivia) using a Mannesmann digital calliper (nearest001mm) Snout shape was assessed according to Heyeret al (1990) Digital webbing formulae followed thenotation system of Savage and Heyer (1997)Acoustic analysis of 27 recorded calling males
(Appendix 4) was conducted in Raven Pro 15 64-bit ver-sion (Bioacoustics Research Program 2012) sound figureswere produced using Seewave version 210 (SueurAubin amp Simonis 2008) and tuneR version 132(Ligges Krey Mersmann amp Schnackenberg 2014) in Rversion 350 (R Core Team 2018) Raven Pro settingswindow size frac14 512 samples window typefrac14Hann 3 dBfilter bandwidth frac14 124Hz window overlap frac14 85 hopsize frac14 77ms discrete Fourier transform (DFT) size frac141024 samples grid spacing frac14 431Hz seewave settingswindow typefrac14Hanning fast Fourier transform (FFT)size frac14 256 samples FFT overlap frac14 90 The analysedfiles are deposited in the sound collection of the lsquoMuseude Biodiversidade do Cerradorsquo (AAG-UFU) lsquoFonotecaNeotropical Jacques Vielardirsquo (FNJV) lsquoLa Sonotheque duMuseum National dHistoire Naturellersquo (MNHN) and theMacaulay library (MC) Detailed information about theanalysed sound files and accession numbers are providedin Appendix 4Call types were distinguished based on the number of
notes per call and pulsing We defined as type 1 callsthe multinote calls formed by nonpulsed notes whichconsist of the main call type emitted by males of allstudied species Type 2 calls are emitted less often thantype 1 and consist of a one-note call formed by poorlydefined pulses We only included type 1 calls in theacoustic diagnoses and the interspecific comparisonsThe acoustic traits were generally analysed manually (ifnot stated otherwise) as follows temporal traits (call
323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370371372373374375376
377378379380381382383384385386387388389390391392393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430
4 A Fouquet et al
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
1
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics 2014
antoinefouquet
Texte inseacutereacute
a
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
s
PROOF ONLY
duration note duration interval between notes numberof notes and call rise time the latter using the lsquoPeakTimersquo function) frequency traits (dominant frequencyusing the lsquoPeak Frequencyrsquo function minimum andmaximum frequency using the lsquoFrequency 5rsquo andlsquoFrequency 95rsquo functions respectively) Call traitsused in the description followed the definitions and ter-minology of Keuroohler et al (2017) using a note-cen-tred approach
Time-calibrated species phylogenyWe selected a representative for each of the 25 OTUsidentified by the species delimitation (see Results) forcomplete mitogenome sequencing to investigate interre-lationships and divergence times Mitogenomic sequen-ces were obtained using low-coverage shotgunsequencing We recovered high-quality mitochondrialgenome assemblies for 18 OTU representatives (seeAppendix 2 for details regarding mitogenome sequenc-ing assembling and annotation) For the remainingseven OTUs for which we could not obtain tissue sam-ples (Appendix 3) we gathered all the available mito-chondrial loci (12S 16S ND1 COI and Cytb) fromGenBank four of these OTUs were represented by 16Sonly We also selected 14 species as outgroups (ninewith complete mitogenomes) including representativesfor other Boana species groups and most otherCophomantini genera (Appendix 3)We extracted 12S 16S and all protein-coding
sequence regions (CDS thus removing D-Loop andtRNAs) from complete mitogenomes as well asGenBank accessions and aligned each locus independ-ently using the MAFFT7 online server For rRNAgenes we chose the E-INS-i strategy recommended forsequences with multiple conserved domains and longgaps For the protein-coding genes we chose the G-INS-i strategy designed for sequences with global hom-ology (Katoh et al 2019) Realignments of CDS con-sidering the reading frame were done and concatenatedin Geneious v918 (httpswwwgeneiouscom) Ourfinal matrix totalled 39 terminals and 13620 alignednucleotide sites Among these terminals 27 were com-plete and nine had fewer than 3000 nucleotides (20)but all were putatively closely related to terminals withcomplete dataWe selected the best-fit partition scheme and model
of evolution for each partition using PartitionFinderV211 (Lanfear et al 2016) based on the BayesianInformation Criterion (BIC) We predefined four datablocks one for rRNA genes (12S and 16S) and one foreach codon position of all CDS regions and found theGTRthorn IthornG model to best-fit all partitions We
reconstructed a time-calibrated tree using a birth-deathtree prior using BEAST 25 to account for extinctionprocesses and incomplete sampling We parameterizedunlinked substitution models according to the estimatesobtained in the PartitionFinder V211 analysis (Lanfearet al 2016) We used the same two calibration pointsas in the previous BEAST analysis Analyses wereundertaken using uncorrelated relaxed lognormal clockmodel of distribution of rate variation among branchesfor each partition (Drummond et al 2006) The Markovchain Monte Carlo (MCMC) parameters were set withfour independent chains of 100 million iterations stor-ing every 10000th iteration and a 10 burn-in Wecombined the log files of the independent runs usingLogCombiner 25 and confirmed the convergence of ourparameters as all ESS were above 200 Finally weextracted the maximum clade credibility tree using Treeannotator 25 We acknowledge that our phylogeneticreconstruction solely based on mtDNA sequences priori-tizes spatial and taxonomic completeness over genomiccoverage and can lead to overestimated divergence times(eg McCormack Heled Delaney Peterson ampKnowles 2011 Near et al 2012)
Biogeographic analysisBiogeographic inferences were undertaken on the time-calibrated phylogeny using the BioGeoBEARS R pack-age (Matzke 2013) This package reconstructs ancestralgeographic distributions and investigates the role ofeach biogeographic event with a maximum likelihoodalgorithm We compared three different models (i) alikelihood version of the Dispersal-Vicariance(DIVALIKE) model (Ronquist 1997) (ii) a likelihoodversion of the BayArea (BBM) model (Landis et al2013) and (iii) the Dispersal Extinction Cladogenesis(DEC) model (Ree amp Smith 2008) We also comparedversions of these models allowing jump dispersal asdescribed by the J parameter (Matzke 2013) Modelswere compared with the Akaike Information Criterion(AIC) We ran 50 independent BioGeoBEARS biogeo-graphic stochastic mapping to determine biogeographicevent counts for the best-fit model (Dupin et al 2017)Ree and Sanmartın (2018) voiced several criticismsagainst the use of J parameter arguing that it inflates thecontribution of cladogenetic events to the likelihoodand minimizes the contribution of anagenetic time-dependent range evolution We thus also compared theresults from the best model without jump dispersalSince this work focuses on the historical biogeog-
raphy of the Amazonian clade we discarded the twoearly diverging lineages B heilprini and B ranicepsfrom the biogeographic analyses since their distribution
431432433434435436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484
485486487488489490491492493494495496497498499500501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 5
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13560371v1)
antoinefouquet
Texte inseacutereacute
D
PROOF ONLY
ranges lie mostly outside of Amazonia We consideredthree areas within Amazonia Western Amazonia theBrazilian Shield and the Guiana Shield These areascorrespond to major geological features roughly delim-ited by modern riverine barriers the Madeira River theNegro River and the lower course of the Amazon Riverand to three large biogeographic regions known asWallacersquos districts (Hoorn et al 2010 Wallace 1854)These districts were recently confirmed as major breaksin birdsrsquo species composition (Oliveira Vasconcelos ampSantos 2017) and amphibians (Godinho amp da Silva2018 Vacher et al 2020) strengthening their status ofbiogeographic regions Because the distribution range ofthe B albopunctata group extends outside Amazoniawe included in the ancestral range reconstruction oneadditional non-Amazonian Neotropical area theDry Diagonal
ResultsSpecies delimitationOf the three tested methods of species delimitation theABGD method was found to be the most conservativedelimiting 21 OTUs while mPTP and GMYC delimited24 and 47 OTUs respectively (Appendix 1) In onecase the consensual partitioning was overly conserva-tive given that B leucocheila and B multifasciata wereconsidered as a single OTU which is contradicted bythe fact that these two taxa are phenotypically distinctTherefore we kept the different lineages delimited byGMYC in this group as distinct OTUs The final delimi-tation led to 25 OTUs (including B heilprini) in the Balbopunctata species group (Fig 1 Appendix 1)Thirteen of these OTUs could be linked to nominal spe-cies (except B caiapo and B paranaiba that were notincluded in the analysis because material was not avail-able) Conversely 11 could not be linked to any nom-inal species representing a possible 44 increase in thespecies richness of the group The Amazonian cladeitself was represented by 14 OTUs including eight nom-inal species and six putative new species (Fig 1) Theseresults also imply important changes to the geographicdistribution of the species of the group In Appendix 5we detail and justify the identification of these OTUsand their respective geographic ranges
Taxonomic accountsThe Amazonian clade of the B albopunctata group pre-viously reported as the B calcaratandashB fasciata speciescomplex by Caminer and Ron (2014) is composed oftwo major clades (the B calcarata and the B steinbachi
clades) defined by the following combination of charac-ter states (1) truncate snout in dorsal view (2) roundedhead shape and (3) presence of calcaneal appendageon heelThe B calcarata clade (Fig 1) is composed of four
species possessing a calcar and having advertisementcalls formed by a single type of call (Caminer amp Ron2014) The B steinbachi clade on the other handincludes three described and two undescribed species(Fig 1) Members of this clade lack a calcar (skin flapor tubercle present or the complete absence of calcanealappendages) and have vocal repertoires composed ofmore than one type of call except for B alfaroi forwhich a single type of call has been documented so far(Caminer amp Ron 2014)Hyla steinbachi Boulenger 1905 currently valid as
Boana steinbachi (Boulenger 1905) was describedfrom the Bolivian Province Sara Departament SantaCruz de la Sierra Bolivia We assume that Buenavistais the type locality of B steinbachi Between 1910 and1950 the Steinbach family collected many amphibiansand reptiles in Bolivia mainly at Buenavista in theDepartment of Santa Cruz (see Parker 1927) which isthe type locality of several anurans (Hamptophryneboliviana (Parker 1927) Pseudopaludicola bolivianaParker 1927 Scinax parkeri (Gaige 1929)) and thesnake Apostolepis tenuis Ruthven 1927 The materialcollected by the Steinbach family is on display in vari-ous museums such as the University of MichiganMuseum of Zoology (UMMZ see Gaige 1929) theNatural History Museum London (BMNH see Parker1927) the Zoologisches Museum Berlin (ZMB seeMeurouller 1924) and the Naturhistorisches Museum Basel(NBM pers comm Weurouest) Many decades later De laRiva (1990) synonymized H steinbachi with H fas-ciata Subsequently Jansen Bloch Schulze andPfenninger (2011) found differences in call and mtDNAdata between populations of B fasciata from Boliviaand Ecuador and suggested that their results imply a res-urrection of B steinbachi In the following yearsCaminer and Ron (2014) tentatively assigned B steinba-chi to a genetic lineage (clade J) inhabiting BolivianAmazonia near its type locality By assessing its phylo-genetic relationships in the group Caminer and Ron(2014) formally revalidated B steinbachi as a distinctspecies from nominal B fasciata The revalidation of Bsteinbachi was also supported by phenotypic dataobtained from syntypes but did not include a reexamina-tion of all diagnostic characters of the speciesNext we present an amended diagnosis of B steinba-
chi based on novel data on life colours and morphologyand describe its vocal repertoire based on topotypesfrom Bolivia and distinct OTUs from Peru and Brazil
539540541542543544545546547548549550551552553554555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592
593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644645646
6 A Fouquet et al
PROOF ONLY
647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695696697698699700
701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754
Fig 1 Subtrees of the three main clades from the chronogram obtained from the analysis of 16S sequences using BEAST2Terminals are collapsed according to the OTU recovered from the species delineation analysis The distribution of each of these OTUis depicted on the maps
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 7
PROOF ONLY
also assigned to this species in this study Specimensand calls from Acre (ldquoB aff steinbachi 1rdquo) and callsfrom the lower Madre de Dios River (ldquoB aff steinbachi2rdquo) were included in the variation of B steinbachi
Boana steinbachi (Boulenger 1905)Hyla steinbachi mdash Boulenger 1905
Hyla fasciata De la Riva 1990Hypsiboas fasciatus Jansen et al 2011
Hypsiboas sp (Clade G) Funk et al 2012Hypsiboas steinbachi Caminer amp Ron 2014
Boana steinbachi Dubois 2017Boana sp (Clade J) Meza-Joya et al 2019
Boana fasciata Vacher et al 2020
Syntypes BMNH 194721361ndash63 two adults ofunknown sex and one juvenile respectively from Saraprovince Department of Santa Cruz de La SierraBolivia Collected by Hf J Steinbach
Diagnosis Boana steinbachi is characterized by the fol-lowing combination of character states (1) skin flap onheel (2) vocal repertoire composed of more than onecall type (3) multinote call and (4) regular internoteintervals (Figs 2 3 amp 4)
Comparisons with congeners of the Amazonian cladeBoana steinbachi can be distinguished from the membersof the B calcarata clade (B almendarizae B fasciata Bcalcarata and B maculateralis) by the absence of a cal-car and by the vocal repertoire composed of more thanone type of call Among members of the B steinbachiclade it can be distinguished from B dentei B alfaroiand B tetete by the presence of a skin flap on heel (tuber-cle in B alfaroi and B tetete tubercle on one side orcompletely absent in B dentei) by its multinote call (one-note call in B dentei and B tetete) by having regularinternote intervals between call notes (notes with irregularintervals sometimes partly fused one with the next in Balfaroi) and by the vocal repertoire composed of two dis-tinct types of calls (one call type in B alfaroi) (Camineramp Ron 2014 Marinho et al 2020)
Variation Body size varies between 304 and 374mmin males and between 425 and 488mm in females(Appendix 7) Individual AAG-UFU 5918 lacks the heelskin flap on the left side topotypes SMF 88394ndash95 andSMF 88397 lack the skin flap on both sides In life(Fig 3a b) dorsal colouration is beige light brownbright yellow or orange brown in calling males withfaint brown transversal bands and dark purplish-brownfaint brown or grey longitudinal lines extending from
the snout to vent a second line extending from behindthe eye to pelvic region and a third line as a canthalstripe extending from the posterior corner of the nostrilto anterior corner of the eye and from the posterior cor-ner of the eye to midbody length on flank (the topotypesSMF 88394ndash97 lack transversal bands on dorsum indi-viduals SMF 88394 and CORBIDI 13414 lack longitu-dinal lines on dorsum and the middorsal line isextending from snout to interocular region AAG-UFU5921 lacks longitudinal lines on dorsum) Iris cream orgrey sometimes with yellow pigmentation on the upperpart of the iris Throat varying from white to yellowchest and anterior belly varying from white to creamVentral surface of hand and hind limb beige to bright
755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808
809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862
Fig 2 Adult male of Boana steinbachi (AAG-UFU 5921)from the Assis Brasil population (Acre Brazil) (A) dorsal and(B) ventral views of the body (SVL frac14 337mm) (C) dorsaland (D) lateral views of the head (HL frac14123mm HW frac14102mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 106mm) and (G) sole of thefoot (FL frac14 139mm)
8 A Fouquet et al
antoinefouquet
Texte surligneacute
we may wanna ask for full page here no
PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Amazonia stands as a major biogeographic region cover-ing more than 65 million km2 including the largestcontinuous tract of tropical forest (40 of the worldrsquostropical rainforests) encompassing the most importantdrainage (Amazon River basin) and probably hostingthe highest continental biodiversity on earth (Myerset al 2000) The outstanding species richness of thisregion has raised many questions concerning its originsand diversification mechanisms While all agree on theastounding diversity of Amazonia our understanding ofthe actual number and distribution of species forming itsecosystems is in fact so incomplete (Ficetola et al2014 Meyer et al 2015 Vacher et al 2020) that ithas so far hampered investigations at the regional scaleConsequently the understanding of the historical causesand regional determinants of this outstanding diversityremain speculative (Antonelli et al 2018) Another con-sequence is that the collision between the Linnean short-fall and the biodiversity crisis is particularly evident inAmazonia (Guerra et al 2020) With almost the entireeastern and southern parts (25) of its extent beingalready deforested for agriculture an unknown propor-tion of endemic species and thus of the testimony ofAmazonian history has already vanished (Lovejoy ampNobre 2019)Anuran clades have repeatedly been documented to
harbour many unnamed species in Amazonia (Vacheret al 2020) and to display strikingly allopatric distribu-tion patterns that could provide crucial insights intoAmazoniarsquos past (Fouquet Cassini Haddad Pech ampRodrigues 2014 Rejaud et al 2020) This is notablythe case of the Boana albopunctata species group(Caminer amp Ron 2014 Funk et al 2012) This groupis only defined on the basis of molecular data(Faivovich et al 2005) and currently comprises 16valid nominal species distributed in the Caribbean andSouth America Boana albopunctata (Spix 1824)Boana alfaroi (Caminer amp Ron 2014) Boana almen-darizae (Caminer amp Ron 2014) Boana caiapo Pinheiroet al 2018 Boana calcarata (Troschel 1848) Boanadentei (Bokermann 1967) Boana fasciata (Geurounther1858) Boana heilprini (Noble 1923) Boana lancifor-mis (Cope 1871) Boana leucocheila (Caramaschi amp deNiemeyer 2003) Boana maculateralis (Caminer ampRon 2014) Boana multifasciata (Geurounther 1859)Boana paranaiba (Carvalho Giaretta amp Facure 2010)Boana steinbachi (Boulenger 1905) Boana tetete(Caminer amp Ron 2014) and Boana raniceps (Cope1862) Within this clade B heilprini is the most distinctsince it is confined to Hispaniola and is phylogeneticallydistant from a clade formed by all the other species ofthe group (Duellman Marion amp Hedges 2016) Wecan also distinguish B raniceps and the B albopunctata
clade (B lanciformis B albopunctata B multifasciataB paranaiba B caiapo B leucocheila) which occurwidely in the open habitats of the Cerrado andAmazonia (Camurugi et al 2021) from the Amazonianclade According to Caminer and Ron (2014) thisAmazonian clade is itself subdivided in two maingroups hereafter called the B calcarata (B fasciata Bcalcarata B almendarizae B maculateralis) and the Bsteinbachi (B tetete B alfaroi B steinbachi) cladesWithin this Amazonian clade Funk Caminer and
Ron (2012) identified no less than seven putativeunnamed species based on mitochondrial divergence andacoustic data These species add up to the 10 taxa thatwere valid at that time Caminer and Ron (2014) com-pleted this picture by documenting two additional line-ages describing four of the previously discoveredspecies and removing B steinbachi from its synonymywith B fasciata However sampling of both studies wascircumscribed to western Amazonia (mostly Ecuador)while the species complex occurs throughout AmazoniaPopulations from the eastern Guiana Shield lowlandsare already assumed to belong to a yet unnamed species(Caminer amp Ron 2014) but the status of numerousother populations throughout Amazonia generally iden-tified as B fasciata remain virtually unknown Morerecently Vacher et al (2020) suggested based on 16Smitochondrial DNA (mtDNA) sequences that up to 22species could exist in that group and that the distributionof most species could be circumscribed to small rangeswithin AmazoniaThese aforementioned studies improved our under-
standing of the actual diversity in the B calcaratastein-bachi clade but left virtually unexplored the temporaland spatial context of the diversification withinAmazonia The crown age of this Amazonian clade wasestimated between 16ndash11 million years ago (Ma) byFunk et al (2012) and between 14ndash9Ma by Duellmanet al (2016) Therefore the initial diversification of thatclade throughout Amazonia may have taken place dur-ing the final stage of the Pebas system a freshwaterlake occupying most of Western Amazonia from theearly Miocene (23Ma) until 10ndash9Ma (Hoorn et al2017) Meanwhile most of the diversification within theB calcaratasteinbachi clade took place during the lateNeogene (last 10Ma) From 9Ma onward this systemhas drained eastward into the Atlantic Ocean but enor-mous flooded ecosystems (Acre system) still occupied alarge portion of what we currently consider as WesternAmazonia until 7Ma (Hoorn et al 2010 Albert et al2018a) Subsequent river captures were common(Ruokolainen Moulatlet Zuquim Hoorn amp Tuomisto2018) as were climatic fluctuations that may havemodified vegetation possibly to the point of forest
107108109110111112113114115116117118119120121122123124125126127128129130131132133134135136137138139140141142143144145146147148149150151152153154155156157158159160
161162163164165166167168169170171172173174175176177178179180181182183184185186187188189190191192193194195196197198199200201202203204205206207208209210211212213214
2 A Fouquet et al
antoinefouquet
Texte inseacutereacute
mostly
PROOF ONLY
fragmentation at least in some peripheral parts ofAmazonia (Cheng et al 2013 Kirschner amp Hoorn2019) The relative roles of hydrological evolution(Albert et al 2018b Ribas Aleixo Nogueira Miyakiamp Cracraft 2012) and rainforest expansion and contrac-tion due to climate oscillations (Haffer 1969) have beenthe focus of intense debate (Leite amp Rogers 2013)Since the species of the B calcarata and B steinbachicomplexes are associated with riparian forests and thesespecies mostly breed in pools formed on the banks ofthe beds of small to medium-sized rivers very large riv-ers may act as barriers to dispersal thus reducing orcompletely impeding gene flow and inhibiting homogen-ization of differentiating populations (Naka et al 2018)However smaller and highly dynamic meandering riversprobably represent more permeable barriers Thereforewe assume that successive dispersals across majorAmazonian rivers (parapatric speciation) (Pirani et al2019) as well as hydrological changes (vicariant speci-ation) (Naka et al 2018) could have been major proc-esses of diversification in these frogsWe gathered a large mtDNA dataset for the B albo-
punctata group sampled throughout Amazonia combinedwith morphological and acoustic data for a subset ofthis group (the B steinbachi clade) to address two maingoals (1) reevaluate species boundaries (in or amongmembers) of the B albopunctata group and theirrespective distributions and (2) investigate its diversifi-cation history within Amazonia using mitogenomic datafor one terminal of each delimited species Furthermoretwo species in the B steinbachi clade that wereunnamed are described herein
Materials and methodsSpecies delimitationOur first objective was to delimit all major OperationalTaxonomic Units (OTUs) based on mtDNA main line-ages Our sampling included 55 new Boana tissue sam-ples obtained through fieldwork throughout Amazoniaand adjacent Dry Diagonal (DD Chaco CerradoCaatinga Werneck 2011) and loans from collaborators(from collections QCAZ MPEG INPA CORBIDIAAGUFU and from personal loans JMP SCF FTAAppendix 1) Samples unambiguously identified (neartype locality andor phenotypically corresponding to typematerial) as belonging to the currently recognized taxawere included except B paranaiba and B caiapo Wesequenced a 400 bp portion of the end of the 16S rDNAgene a locus commonly used for Neotropical amphibiantaxonomy and systematics (Vences Thomas Bonett ampVieites 2005) We also retrieved homologous sequences
from GenBank (389 accessions) In total we gathered444 16S sequences for this study (dataset details are pro-vided in Appendix 1 and DNA extraction and sequencingprotocols can be found in Appendix 2) These samplescover the whole distribution of the B albopunctata spe-cies group (14 included taxa) DNA sequence alignmentwas conducted on the MAFFT7 online server under theE-INS-i option with default parameters an algorithmdesigned for sequences with multiple conserved domainsand long gaps (Katoh et al 2019)We applied three DNA-based single-locus species
delimitation approaches (a) a distance-based methodthe Automated Barcode Gap Discovery (ABGDPuillandre Lambert Brouillet amp Achaz 2012) (b) amulti-rate coalescent based method the multi-ratePoisson Tree Processes model approach (mPTP Kapliet al 2017) and (c) a single-threshold coalescent-basedmethod the Generalized Mixed Yule Coalescentapproach (single threshold GMYC Monaghan et al2009 Pons et al 2006)The ABGD delimitation was performed with a prior
of intraspecific divergence P between 0001 and 01 aproxy for minimum relative gap width X of 1 and anumber of steps n equal to 30 We kept the partitionsuch that Pfrac14 0016 as it corresponds to the end of aplateau for group number and it matches thresholds ofintraspecific divergence proposed in other vertebratedelimitation studies using 16S barcodes (Puillandreet al 2012) For mPTP delimitation we first recon-structed a Maximum Likelihood (ML) tree withRAxML v824 (Stamatakis 2014) using theGTRthornGthorn I substitution model and estimated nodalsupport via 1000 parametric bootstraps We used nineoutgroups representing most other Boana species groups(Appendix 3) The mPTP delimitation was undertakenon the rooted ML tree with 5 million Markov chainMonte Carlo (MCMC) iterations sampling every10000th iteration and a 10 burn-in For the GMYCdelimitation we reconstructed a time-calibrated phyl-ogeny using BEAST 25 (Bouckaert et al 2014) Weused a birth-death population model to account forextinction processes and incomplete sampling Weincluded the same nine Boana outgroups used inRAxML reconstruction and five additional representa-tives of other Cophomantini genera (NesorohylaMyersiohyla Hyloscirtus AplastodiscusBokermannohyla) in order to include relationships thatcan be time-calibrated We used a single partition with aGTRthornGthorn I substitution model with an uncorrelatedrelaxed lognormal clock model of rate variation amongbranches (Drummond Ho Phillips amp Rambaut 2006)We used two calibration points the ages of the mostrecent common ancestor (MRCA) of Aplastodiscus and
215216217218219220221222223224225226227228229230231232233234235236237238239240241242243244245246247248249250251252253254255256257258259260261262263264265266267268
269270271272273274275276277278279280281282283284285286287288289290291292293294295296297298299300301302303304305306307308309310311312313314315316317318319320321322
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 3
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13536302v1
PROOF ONLY
Boana as well as the MRCA of a clade formed byHyloscirtus Boana Aplastodiscus andBokermannohyla which were estimated by Feng et al(2017) and consequently constrained here assuming nor-mal prior distributions of 252Ma (standard deviation[SD]frac14 28Ma) and 323Ma (SD frac14 3Ma) respectivelyWe considered these calibrations to represent the bestprior because Feng et al (2017) included a comprehen-sive dataset of nuclear loci for an extensive sampling ofanurans and used many reliable fossils as calibrationpoints Moreover Hime et al (2020) analysing an evenlarger genomic dataset found very similar time esti-mates for the TMRCA of BoanathornHyloscirtus Insteadusing fossil andor biogeographic calibrations wouldhave implied to expand the matrix to lineages distantlyrelated to Boana and would likely lead to an overesti-mation of calibration dates with our mitogenomic data-set (Molak amp Ho 2015 Papadopoulou et al 2010) Forthe MCMC parameters we used four independentchains of 100 million iterations recording every10000th iteration and a 10 burn-in We combined thelog files of the independent runs using LogCombiner25 (Drummond amp Rambaut 2007) and checked theconvergence of our parameters confirmed by allEffective Sample Size (ESS) being above 200 Then weextracted the maximum clade credibility tree using Treeannotator 25 (Drummond amp Rambaut 2007) with aburn-in of 10 After removing outgroups we per-formed a GMYC delimitation on the ultrametric treeusing the GMYC function of the splits R package v10-11 (Ezard Fujisawa amp Barraclough 2009) with athreshold interval between 0 and 10Ma and by usingthe single threshold method Operational TaxonomicUnits (OTUs) were defined using a majority-rule con-sensus from the results of the three methods ie a lin-eage is considered as being an OTU if supported by atleast two of the three methodsAll but five occurrence records were georeferenced
and used to create distribution maps with convex poly-gons under QGIS 214 (QGIS Geographic InformationSystem) OTUs were assigned to taxa based on thefieldmuseum identification and sometimes corrected inaccordance with type localities and known distribution(Frost 2019 IUCN 2020)
Morphological and acoustic variation in theBoana steinbachi cladeThe following morphometric measurements were takenon 38 males and 9 females snout-vent length (SVL)head length (HL) head width (HW) eye diameter (ED)tympanum diameter (TD) tibia length (TL) foot length(FL) thigh length (THL) and calcaneal appendage
length (CL) In addition to these we also measuredhand length (HAL) forearm length (FLL) and eyendashnos-tril distance (EN) for the morphometric characterizationof holotypes All measurements followed the definitionsand terminology of Watters Cummings Flanagan andSiler (2016) We classified the calcaneal appendage intothree-character states (i) a calcar (skin appendage withlength gt 1mm flattened or conical) a (ii) skin flap(skin appendage with length lt 1mm flattened or a(iii) tubercle a conical structure with reduced size andas such not forming an appendage coming out of theheel The specimens were measured by P Marinho(Alta Floresta Mato Grosso State and Assis BrazilAcre State) using a Mitutoyo digital calliper (to thenearest 005mm) by MT Rodrigues (Jirau and PacaasNovos Rondonia State) using a Mitutoyo digital calliper(to the nearest 001mm) by A Fouquet (French Guianaand Suriname) using a DigiMax digital calliper (nearest001mm) and by M Caminer (Ecuador Peru andBolivia) using a Mannesmann digital calliper (nearest001mm) Snout shape was assessed according to Heyeret al (1990) Digital webbing formulae followed thenotation system of Savage and Heyer (1997)Acoustic analysis of 27 recorded calling males
(Appendix 4) was conducted in Raven Pro 15 64-bit ver-sion (Bioacoustics Research Program 2012) sound figureswere produced using Seewave version 210 (SueurAubin amp Simonis 2008) and tuneR version 132(Ligges Krey Mersmann amp Schnackenberg 2014) in Rversion 350 (R Core Team 2018) Raven Pro settingswindow size frac14 512 samples window typefrac14Hann 3 dBfilter bandwidth frac14 124Hz window overlap frac14 85 hopsize frac14 77ms discrete Fourier transform (DFT) size frac141024 samples grid spacing frac14 431Hz seewave settingswindow typefrac14Hanning fast Fourier transform (FFT)size frac14 256 samples FFT overlap frac14 90 The analysedfiles are deposited in the sound collection of the lsquoMuseude Biodiversidade do Cerradorsquo (AAG-UFU) lsquoFonotecaNeotropical Jacques Vielardirsquo (FNJV) lsquoLa Sonotheque duMuseum National dHistoire Naturellersquo (MNHN) and theMacaulay library (MC) Detailed information about theanalysed sound files and accession numbers are providedin Appendix 4Call types were distinguished based on the number of
notes per call and pulsing We defined as type 1 callsthe multinote calls formed by nonpulsed notes whichconsist of the main call type emitted by males of allstudied species Type 2 calls are emitted less often thantype 1 and consist of a one-note call formed by poorlydefined pulses We only included type 1 calls in theacoustic diagnoses and the interspecific comparisonsThe acoustic traits were generally analysed manually (ifnot stated otherwise) as follows temporal traits (call
323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370371372373374375376
377378379380381382383384385386387388389390391392393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430
4 A Fouquet et al
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
1
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics 2014
antoinefouquet
Texte inseacutereacute
a
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
s
PROOF ONLY
duration note duration interval between notes numberof notes and call rise time the latter using the lsquoPeakTimersquo function) frequency traits (dominant frequencyusing the lsquoPeak Frequencyrsquo function minimum andmaximum frequency using the lsquoFrequency 5rsquo andlsquoFrequency 95rsquo functions respectively) Call traitsused in the description followed the definitions and ter-minology of Keuroohler et al (2017) using a note-cen-tred approach
Time-calibrated species phylogenyWe selected a representative for each of the 25 OTUsidentified by the species delimitation (see Results) forcomplete mitogenome sequencing to investigate interre-lationships and divergence times Mitogenomic sequen-ces were obtained using low-coverage shotgunsequencing We recovered high-quality mitochondrialgenome assemblies for 18 OTU representatives (seeAppendix 2 for details regarding mitogenome sequenc-ing assembling and annotation) For the remainingseven OTUs for which we could not obtain tissue sam-ples (Appendix 3) we gathered all the available mito-chondrial loci (12S 16S ND1 COI and Cytb) fromGenBank four of these OTUs were represented by 16Sonly We also selected 14 species as outgroups (ninewith complete mitogenomes) including representativesfor other Boana species groups and most otherCophomantini genera (Appendix 3)We extracted 12S 16S and all protein-coding
sequence regions (CDS thus removing D-Loop andtRNAs) from complete mitogenomes as well asGenBank accessions and aligned each locus independ-ently using the MAFFT7 online server For rRNAgenes we chose the E-INS-i strategy recommended forsequences with multiple conserved domains and longgaps For the protein-coding genes we chose the G-INS-i strategy designed for sequences with global hom-ology (Katoh et al 2019) Realignments of CDS con-sidering the reading frame were done and concatenatedin Geneious v918 (httpswwwgeneiouscom) Ourfinal matrix totalled 39 terminals and 13620 alignednucleotide sites Among these terminals 27 were com-plete and nine had fewer than 3000 nucleotides (20)but all were putatively closely related to terminals withcomplete dataWe selected the best-fit partition scheme and model
of evolution for each partition using PartitionFinderV211 (Lanfear et al 2016) based on the BayesianInformation Criterion (BIC) We predefined four datablocks one for rRNA genes (12S and 16S) and one foreach codon position of all CDS regions and found theGTRthorn IthornG model to best-fit all partitions We
reconstructed a time-calibrated tree using a birth-deathtree prior using BEAST 25 to account for extinctionprocesses and incomplete sampling We parameterizedunlinked substitution models according to the estimatesobtained in the PartitionFinder V211 analysis (Lanfearet al 2016) We used the same two calibration pointsas in the previous BEAST analysis Analyses wereundertaken using uncorrelated relaxed lognormal clockmodel of distribution of rate variation among branchesfor each partition (Drummond et al 2006) The Markovchain Monte Carlo (MCMC) parameters were set withfour independent chains of 100 million iterations stor-ing every 10000th iteration and a 10 burn-in Wecombined the log files of the independent runs usingLogCombiner 25 and confirmed the convergence of ourparameters as all ESS were above 200 Finally weextracted the maximum clade credibility tree using Treeannotator 25 We acknowledge that our phylogeneticreconstruction solely based on mtDNA sequences priori-tizes spatial and taxonomic completeness over genomiccoverage and can lead to overestimated divergence times(eg McCormack Heled Delaney Peterson ampKnowles 2011 Near et al 2012)
Biogeographic analysisBiogeographic inferences were undertaken on the time-calibrated phylogeny using the BioGeoBEARS R pack-age (Matzke 2013) This package reconstructs ancestralgeographic distributions and investigates the role ofeach biogeographic event with a maximum likelihoodalgorithm We compared three different models (i) alikelihood version of the Dispersal-Vicariance(DIVALIKE) model (Ronquist 1997) (ii) a likelihoodversion of the BayArea (BBM) model (Landis et al2013) and (iii) the Dispersal Extinction Cladogenesis(DEC) model (Ree amp Smith 2008) We also comparedversions of these models allowing jump dispersal asdescribed by the J parameter (Matzke 2013) Modelswere compared with the Akaike Information Criterion(AIC) We ran 50 independent BioGeoBEARS biogeo-graphic stochastic mapping to determine biogeographicevent counts for the best-fit model (Dupin et al 2017)Ree and Sanmartın (2018) voiced several criticismsagainst the use of J parameter arguing that it inflates thecontribution of cladogenetic events to the likelihoodand minimizes the contribution of anagenetic time-dependent range evolution We thus also compared theresults from the best model without jump dispersalSince this work focuses on the historical biogeog-
raphy of the Amazonian clade we discarded the twoearly diverging lineages B heilprini and B ranicepsfrom the biogeographic analyses since their distribution
431432433434435436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484
485486487488489490491492493494495496497498499500501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 5
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13560371v1)
antoinefouquet
Texte inseacutereacute
D
PROOF ONLY
ranges lie mostly outside of Amazonia We consideredthree areas within Amazonia Western Amazonia theBrazilian Shield and the Guiana Shield These areascorrespond to major geological features roughly delim-ited by modern riverine barriers the Madeira River theNegro River and the lower course of the Amazon Riverand to three large biogeographic regions known asWallacersquos districts (Hoorn et al 2010 Wallace 1854)These districts were recently confirmed as major breaksin birdsrsquo species composition (Oliveira Vasconcelos ampSantos 2017) and amphibians (Godinho amp da Silva2018 Vacher et al 2020) strengthening their status ofbiogeographic regions Because the distribution range ofthe B albopunctata group extends outside Amazoniawe included in the ancestral range reconstruction oneadditional non-Amazonian Neotropical area theDry Diagonal
ResultsSpecies delimitationOf the three tested methods of species delimitation theABGD method was found to be the most conservativedelimiting 21 OTUs while mPTP and GMYC delimited24 and 47 OTUs respectively (Appendix 1) In onecase the consensual partitioning was overly conserva-tive given that B leucocheila and B multifasciata wereconsidered as a single OTU which is contradicted bythe fact that these two taxa are phenotypically distinctTherefore we kept the different lineages delimited byGMYC in this group as distinct OTUs The final delimi-tation led to 25 OTUs (including B heilprini) in the Balbopunctata species group (Fig 1 Appendix 1)Thirteen of these OTUs could be linked to nominal spe-cies (except B caiapo and B paranaiba that were notincluded in the analysis because material was not avail-able) Conversely 11 could not be linked to any nom-inal species representing a possible 44 increase in thespecies richness of the group The Amazonian cladeitself was represented by 14 OTUs including eight nom-inal species and six putative new species (Fig 1) Theseresults also imply important changes to the geographicdistribution of the species of the group In Appendix 5we detail and justify the identification of these OTUsand their respective geographic ranges
Taxonomic accountsThe Amazonian clade of the B albopunctata group pre-viously reported as the B calcaratandashB fasciata speciescomplex by Caminer and Ron (2014) is composed oftwo major clades (the B calcarata and the B steinbachi
clades) defined by the following combination of charac-ter states (1) truncate snout in dorsal view (2) roundedhead shape and (3) presence of calcaneal appendageon heelThe B calcarata clade (Fig 1) is composed of four
species possessing a calcar and having advertisementcalls formed by a single type of call (Caminer amp Ron2014) The B steinbachi clade on the other handincludes three described and two undescribed species(Fig 1) Members of this clade lack a calcar (skin flapor tubercle present or the complete absence of calcanealappendages) and have vocal repertoires composed ofmore than one type of call except for B alfaroi forwhich a single type of call has been documented so far(Caminer amp Ron 2014)Hyla steinbachi Boulenger 1905 currently valid as
Boana steinbachi (Boulenger 1905) was describedfrom the Bolivian Province Sara Departament SantaCruz de la Sierra Bolivia We assume that Buenavistais the type locality of B steinbachi Between 1910 and1950 the Steinbach family collected many amphibiansand reptiles in Bolivia mainly at Buenavista in theDepartment of Santa Cruz (see Parker 1927) which isthe type locality of several anurans (Hamptophryneboliviana (Parker 1927) Pseudopaludicola bolivianaParker 1927 Scinax parkeri (Gaige 1929)) and thesnake Apostolepis tenuis Ruthven 1927 The materialcollected by the Steinbach family is on display in vari-ous museums such as the University of MichiganMuseum of Zoology (UMMZ see Gaige 1929) theNatural History Museum London (BMNH see Parker1927) the Zoologisches Museum Berlin (ZMB seeMeurouller 1924) and the Naturhistorisches Museum Basel(NBM pers comm Weurouest) Many decades later De laRiva (1990) synonymized H steinbachi with H fas-ciata Subsequently Jansen Bloch Schulze andPfenninger (2011) found differences in call and mtDNAdata between populations of B fasciata from Boliviaand Ecuador and suggested that their results imply a res-urrection of B steinbachi In the following yearsCaminer and Ron (2014) tentatively assigned B steinba-chi to a genetic lineage (clade J) inhabiting BolivianAmazonia near its type locality By assessing its phylo-genetic relationships in the group Caminer and Ron(2014) formally revalidated B steinbachi as a distinctspecies from nominal B fasciata The revalidation of Bsteinbachi was also supported by phenotypic dataobtained from syntypes but did not include a reexamina-tion of all diagnostic characters of the speciesNext we present an amended diagnosis of B steinba-
chi based on novel data on life colours and morphologyand describe its vocal repertoire based on topotypesfrom Bolivia and distinct OTUs from Peru and Brazil
539540541542543544545546547548549550551552553554555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592
593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644645646
6 A Fouquet et al
PROOF ONLY
647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695696697698699700
701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754
Fig 1 Subtrees of the three main clades from the chronogram obtained from the analysis of 16S sequences using BEAST2Terminals are collapsed according to the OTU recovered from the species delineation analysis The distribution of each of these OTUis depicted on the maps
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 7
PROOF ONLY
also assigned to this species in this study Specimensand calls from Acre (ldquoB aff steinbachi 1rdquo) and callsfrom the lower Madre de Dios River (ldquoB aff steinbachi2rdquo) were included in the variation of B steinbachi
Boana steinbachi (Boulenger 1905)Hyla steinbachi mdash Boulenger 1905
Hyla fasciata De la Riva 1990Hypsiboas fasciatus Jansen et al 2011
Hypsiboas sp (Clade G) Funk et al 2012Hypsiboas steinbachi Caminer amp Ron 2014
Boana steinbachi Dubois 2017Boana sp (Clade J) Meza-Joya et al 2019
Boana fasciata Vacher et al 2020
Syntypes BMNH 194721361ndash63 two adults ofunknown sex and one juvenile respectively from Saraprovince Department of Santa Cruz de La SierraBolivia Collected by Hf J Steinbach
Diagnosis Boana steinbachi is characterized by the fol-lowing combination of character states (1) skin flap onheel (2) vocal repertoire composed of more than onecall type (3) multinote call and (4) regular internoteintervals (Figs 2 3 amp 4)
Comparisons with congeners of the Amazonian cladeBoana steinbachi can be distinguished from the membersof the B calcarata clade (B almendarizae B fasciata Bcalcarata and B maculateralis) by the absence of a cal-car and by the vocal repertoire composed of more thanone type of call Among members of the B steinbachiclade it can be distinguished from B dentei B alfaroiand B tetete by the presence of a skin flap on heel (tuber-cle in B alfaroi and B tetete tubercle on one side orcompletely absent in B dentei) by its multinote call (one-note call in B dentei and B tetete) by having regularinternote intervals between call notes (notes with irregularintervals sometimes partly fused one with the next in Balfaroi) and by the vocal repertoire composed of two dis-tinct types of calls (one call type in B alfaroi) (Camineramp Ron 2014 Marinho et al 2020)
Variation Body size varies between 304 and 374mmin males and between 425 and 488mm in females(Appendix 7) Individual AAG-UFU 5918 lacks the heelskin flap on the left side topotypes SMF 88394ndash95 andSMF 88397 lack the skin flap on both sides In life(Fig 3a b) dorsal colouration is beige light brownbright yellow or orange brown in calling males withfaint brown transversal bands and dark purplish-brownfaint brown or grey longitudinal lines extending from
the snout to vent a second line extending from behindthe eye to pelvic region and a third line as a canthalstripe extending from the posterior corner of the nostrilto anterior corner of the eye and from the posterior cor-ner of the eye to midbody length on flank (the topotypesSMF 88394ndash97 lack transversal bands on dorsum indi-viduals SMF 88394 and CORBIDI 13414 lack longitu-dinal lines on dorsum and the middorsal line isextending from snout to interocular region AAG-UFU5921 lacks longitudinal lines on dorsum) Iris cream orgrey sometimes with yellow pigmentation on the upperpart of the iris Throat varying from white to yellowchest and anterior belly varying from white to creamVentral surface of hand and hind limb beige to bright
755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808
809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862
Fig 2 Adult male of Boana steinbachi (AAG-UFU 5921)from the Assis Brasil population (Acre Brazil) (A) dorsal and(B) ventral views of the body (SVL frac14 337mm) (C) dorsaland (D) lateral views of the head (HL frac14123mm HW frac14102mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 106mm) and (G) sole of thefoot (FL frac14 139mm)
8 A Fouquet et al
antoinefouquet
Texte surligneacute
we may wanna ask for full page here no
PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
fragmentation at least in some peripheral parts ofAmazonia (Cheng et al 2013 Kirschner amp Hoorn2019) The relative roles of hydrological evolution(Albert et al 2018b Ribas Aleixo Nogueira Miyakiamp Cracraft 2012) and rainforest expansion and contrac-tion due to climate oscillations (Haffer 1969) have beenthe focus of intense debate (Leite amp Rogers 2013)Since the species of the B calcarata and B steinbachicomplexes are associated with riparian forests and thesespecies mostly breed in pools formed on the banks ofthe beds of small to medium-sized rivers very large riv-ers may act as barriers to dispersal thus reducing orcompletely impeding gene flow and inhibiting homogen-ization of differentiating populations (Naka et al 2018)However smaller and highly dynamic meandering riversprobably represent more permeable barriers Thereforewe assume that successive dispersals across majorAmazonian rivers (parapatric speciation) (Pirani et al2019) as well as hydrological changes (vicariant speci-ation) (Naka et al 2018) could have been major proc-esses of diversification in these frogsWe gathered a large mtDNA dataset for the B albo-
punctata group sampled throughout Amazonia combinedwith morphological and acoustic data for a subset ofthis group (the B steinbachi clade) to address two maingoals (1) reevaluate species boundaries (in or amongmembers) of the B albopunctata group and theirrespective distributions and (2) investigate its diversifi-cation history within Amazonia using mitogenomic datafor one terminal of each delimited species Furthermoretwo species in the B steinbachi clade that wereunnamed are described herein
Materials and methodsSpecies delimitationOur first objective was to delimit all major OperationalTaxonomic Units (OTUs) based on mtDNA main line-ages Our sampling included 55 new Boana tissue sam-ples obtained through fieldwork throughout Amazoniaand adjacent Dry Diagonal (DD Chaco CerradoCaatinga Werneck 2011) and loans from collaborators(from collections QCAZ MPEG INPA CORBIDIAAGUFU and from personal loans JMP SCF FTAAppendix 1) Samples unambiguously identified (neartype locality andor phenotypically corresponding to typematerial) as belonging to the currently recognized taxawere included except B paranaiba and B caiapo Wesequenced a 400 bp portion of the end of the 16S rDNAgene a locus commonly used for Neotropical amphibiantaxonomy and systematics (Vences Thomas Bonett ampVieites 2005) We also retrieved homologous sequences
from GenBank (389 accessions) In total we gathered444 16S sequences for this study (dataset details are pro-vided in Appendix 1 and DNA extraction and sequencingprotocols can be found in Appendix 2) These samplescover the whole distribution of the B albopunctata spe-cies group (14 included taxa) DNA sequence alignmentwas conducted on the MAFFT7 online server under theE-INS-i option with default parameters an algorithmdesigned for sequences with multiple conserved domainsand long gaps (Katoh et al 2019)We applied three DNA-based single-locus species
delimitation approaches (a) a distance-based methodthe Automated Barcode Gap Discovery (ABGDPuillandre Lambert Brouillet amp Achaz 2012) (b) amulti-rate coalescent based method the multi-ratePoisson Tree Processes model approach (mPTP Kapliet al 2017) and (c) a single-threshold coalescent-basedmethod the Generalized Mixed Yule Coalescentapproach (single threshold GMYC Monaghan et al2009 Pons et al 2006)The ABGD delimitation was performed with a prior
of intraspecific divergence P between 0001 and 01 aproxy for minimum relative gap width X of 1 and anumber of steps n equal to 30 We kept the partitionsuch that Pfrac14 0016 as it corresponds to the end of aplateau for group number and it matches thresholds ofintraspecific divergence proposed in other vertebratedelimitation studies using 16S barcodes (Puillandreet al 2012) For mPTP delimitation we first recon-structed a Maximum Likelihood (ML) tree withRAxML v824 (Stamatakis 2014) using theGTRthornGthorn I substitution model and estimated nodalsupport via 1000 parametric bootstraps We used nineoutgroups representing most other Boana species groups(Appendix 3) The mPTP delimitation was undertakenon the rooted ML tree with 5 million Markov chainMonte Carlo (MCMC) iterations sampling every10000th iteration and a 10 burn-in For the GMYCdelimitation we reconstructed a time-calibrated phyl-ogeny using BEAST 25 (Bouckaert et al 2014) Weused a birth-death population model to account forextinction processes and incomplete sampling Weincluded the same nine Boana outgroups used inRAxML reconstruction and five additional representa-tives of other Cophomantini genera (NesorohylaMyersiohyla Hyloscirtus AplastodiscusBokermannohyla) in order to include relationships thatcan be time-calibrated We used a single partition with aGTRthornGthorn I substitution model with an uncorrelatedrelaxed lognormal clock model of rate variation amongbranches (Drummond Ho Phillips amp Rambaut 2006)We used two calibration points the ages of the mostrecent common ancestor (MRCA) of Aplastodiscus and
215216217218219220221222223224225226227228229230231232233234235236237238239240241242243244245246247248249250251252253254255256257258259260261262263264265266267268
269270271272273274275276277278279280281282283284285286287288289290291292293294295296297298299300301302303304305306307308309310311312313314315316317318319320321322
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 3
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
amp Brumfield 2018
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13536302v1
PROOF ONLY
Boana as well as the MRCA of a clade formed byHyloscirtus Boana Aplastodiscus andBokermannohyla which were estimated by Feng et al(2017) and consequently constrained here assuming nor-mal prior distributions of 252Ma (standard deviation[SD]frac14 28Ma) and 323Ma (SD frac14 3Ma) respectivelyWe considered these calibrations to represent the bestprior because Feng et al (2017) included a comprehen-sive dataset of nuclear loci for an extensive sampling ofanurans and used many reliable fossils as calibrationpoints Moreover Hime et al (2020) analysing an evenlarger genomic dataset found very similar time esti-mates for the TMRCA of BoanathornHyloscirtus Insteadusing fossil andor biogeographic calibrations wouldhave implied to expand the matrix to lineages distantlyrelated to Boana and would likely lead to an overesti-mation of calibration dates with our mitogenomic data-set (Molak amp Ho 2015 Papadopoulou et al 2010) Forthe MCMC parameters we used four independentchains of 100 million iterations recording every10000th iteration and a 10 burn-in We combined thelog files of the independent runs using LogCombiner25 (Drummond amp Rambaut 2007) and checked theconvergence of our parameters confirmed by allEffective Sample Size (ESS) being above 200 Then weextracted the maximum clade credibility tree using Treeannotator 25 (Drummond amp Rambaut 2007) with aburn-in of 10 After removing outgroups we per-formed a GMYC delimitation on the ultrametric treeusing the GMYC function of the splits R package v10-11 (Ezard Fujisawa amp Barraclough 2009) with athreshold interval between 0 and 10Ma and by usingthe single threshold method Operational TaxonomicUnits (OTUs) were defined using a majority-rule con-sensus from the results of the three methods ie a lin-eage is considered as being an OTU if supported by atleast two of the three methodsAll but five occurrence records were georeferenced
and used to create distribution maps with convex poly-gons under QGIS 214 (QGIS Geographic InformationSystem) OTUs were assigned to taxa based on thefieldmuseum identification and sometimes corrected inaccordance with type localities and known distribution(Frost 2019 IUCN 2020)
Morphological and acoustic variation in theBoana steinbachi cladeThe following morphometric measurements were takenon 38 males and 9 females snout-vent length (SVL)head length (HL) head width (HW) eye diameter (ED)tympanum diameter (TD) tibia length (TL) foot length(FL) thigh length (THL) and calcaneal appendage
length (CL) In addition to these we also measuredhand length (HAL) forearm length (FLL) and eyendashnos-tril distance (EN) for the morphometric characterizationof holotypes All measurements followed the definitionsand terminology of Watters Cummings Flanagan andSiler (2016) We classified the calcaneal appendage intothree-character states (i) a calcar (skin appendage withlength gt 1mm flattened or conical) a (ii) skin flap(skin appendage with length lt 1mm flattened or a(iii) tubercle a conical structure with reduced size andas such not forming an appendage coming out of theheel The specimens were measured by P Marinho(Alta Floresta Mato Grosso State and Assis BrazilAcre State) using a Mitutoyo digital calliper (to thenearest 005mm) by MT Rodrigues (Jirau and PacaasNovos Rondonia State) using a Mitutoyo digital calliper(to the nearest 001mm) by A Fouquet (French Guianaand Suriname) using a DigiMax digital calliper (nearest001mm) and by M Caminer (Ecuador Peru andBolivia) using a Mannesmann digital calliper (nearest001mm) Snout shape was assessed according to Heyeret al (1990) Digital webbing formulae followed thenotation system of Savage and Heyer (1997)Acoustic analysis of 27 recorded calling males
(Appendix 4) was conducted in Raven Pro 15 64-bit ver-sion (Bioacoustics Research Program 2012) sound figureswere produced using Seewave version 210 (SueurAubin amp Simonis 2008) and tuneR version 132(Ligges Krey Mersmann amp Schnackenberg 2014) in Rversion 350 (R Core Team 2018) Raven Pro settingswindow size frac14 512 samples window typefrac14Hann 3 dBfilter bandwidth frac14 124Hz window overlap frac14 85 hopsize frac14 77ms discrete Fourier transform (DFT) size frac141024 samples grid spacing frac14 431Hz seewave settingswindow typefrac14Hanning fast Fourier transform (FFT)size frac14 256 samples FFT overlap frac14 90 The analysedfiles are deposited in the sound collection of the lsquoMuseude Biodiversidade do Cerradorsquo (AAG-UFU) lsquoFonotecaNeotropical Jacques Vielardirsquo (FNJV) lsquoLa Sonotheque duMuseum National dHistoire Naturellersquo (MNHN) and theMacaulay library (MC) Detailed information about theanalysed sound files and accession numbers are providedin Appendix 4Call types were distinguished based on the number of
notes per call and pulsing We defined as type 1 callsthe multinote calls formed by nonpulsed notes whichconsist of the main call type emitted by males of allstudied species Type 2 calls are emitted less often thantype 1 and consist of a one-note call formed by poorlydefined pulses We only included type 1 calls in theacoustic diagnoses and the interspecific comparisonsThe acoustic traits were generally analysed manually (ifnot stated otherwise) as follows temporal traits (call
323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370371372373374375376
377378379380381382383384385386387388389390391392393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430
4 A Fouquet et al
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
1
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics 2014
antoinefouquet
Texte inseacutereacute
a
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
s
PROOF ONLY
duration note duration interval between notes numberof notes and call rise time the latter using the lsquoPeakTimersquo function) frequency traits (dominant frequencyusing the lsquoPeak Frequencyrsquo function minimum andmaximum frequency using the lsquoFrequency 5rsquo andlsquoFrequency 95rsquo functions respectively) Call traitsused in the description followed the definitions and ter-minology of Keuroohler et al (2017) using a note-cen-tred approach
Time-calibrated species phylogenyWe selected a representative for each of the 25 OTUsidentified by the species delimitation (see Results) forcomplete mitogenome sequencing to investigate interre-lationships and divergence times Mitogenomic sequen-ces were obtained using low-coverage shotgunsequencing We recovered high-quality mitochondrialgenome assemblies for 18 OTU representatives (seeAppendix 2 for details regarding mitogenome sequenc-ing assembling and annotation) For the remainingseven OTUs for which we could not obtain tissue sam-ples (Appendix 3) we gathered all the available mito-chondrial loci (12S 16S ND1 COI and Cytb) fromGenBank four of these OTUs were represented by 16Sonly We also selected 14 species as outgroups (ninewith complete mitogenomes) including representativesfor other Boana species groups and most otherCophomantini genera (Appendix 3)We extracted 12S 16S and all protein-coding
sequence regions (CDS thus removing D-Loop andtRNAs) from complete mitogenomes as well asGenBank accessions and aligned each locus independ-ently using the MAFFT7 online server For rRNAgenes we chose the E-INS-i strategy recommended forsequences with multiple conserved domains and longgaps For the protein-coding genes we chose the G-INS-i strategy designed for sequences with global hom-ology (Katoh et al 2019) Realignments of CDS con-sidering the reading frame were done and concatenatedin Geneious v918 (httpswwwgeneiouscom) Ourfinal matrix totalled 39 terminals and 13620 alignednucleotide sites Among these terminals 27 were com-plete and nine had fewer than 3000 nucleotides (20)but all were putatively closely related to terminals withcomplete dataWe selected the best-fit partition scheme and model
of evolution for each partition using PartitionFinderV211 (Lanfear et al 2016) based on the BayesianInformation Criterion (BIC) We predefined four datablocks one for rRNA genes (12S and 16S) and one foreach codon position of all CDS regions and found theGTRthorn IthornG model to best-fit all partitions We
reconstructed a time-calibrated tree using a birth-deathtree prior using BEAST 25 to account for extinctionprocesses and incomplete sampling We parameterizedunlinked substitution models according to the estimatesobtained in the PartitionFinder V211 analysis (Lanfearet al 2016) We used the same two calibration pointsas in the previous BEAST analysis Analyses wereundertaken using uncorrelated relaxed lognormal clockmodel of distribution of rate variation among branchesfor each partition (Drummond et al 2006) The Markovchain Monte Carlo (MCMC) parameters were set withfour independent chains of 100 million iterations stor-ing every 10000th iteration and a 10 burn-in Wecombined the log files of the independent runs usingLogCombiner 25 and confirmed the convergence of ourparameters as all ESS were above 200 Finally weextracted the maximum clade credibility tree using Treeannotator 25 We acknowledge that our phylogeneticreconstruction solely based on mtDNA sequences priori-tizes spatial and taxonomic completeness over genomiccoverage and can lead to overestimated divergence times(eg McCormack Heled Delaney Peterson ampKnowles 2011 Near et al 2012)
Biogeographic analysisBiogeographic inferences were undertaken on the time-calibrated phylogeny using the BioGeoBEARS R pack-age (Matzke 2013) This package reconstructs ancestralgeographic distributions and investigates the role ofeach biogeographic event with a maximum likelihoodalgorithm We compared three different models (i) alikelihood version of the Dispersal-Vicariance(DIVALIKE) model (Ronquist 1997) (ii) a likelihoodversion of the BayArea (BBM) model (Landis et al2013) and (iii) the Dispersal Extinction Cladogenesis(DEC) model (Ree amp Smith 2008) We also comparedversions of these models allowing jump dispersal asdescribed by the J parameter (Matzke 2013) Modelswere compared with the Akaike Information Criterion(AIC) We ran 50 independent BioGeoBEARS biogeo-graphic stochastic mapping to determine biogeographicevent counts for the best-fit model (Dupin et al 2017)Ree and Sanmartın (2018) voiced several criticismsagainst the use of J parameter arguing that it inflates thecontribution of cladogenetic events to the likelihoodand minimizes the contribution of anagenetic time-dependent range evolution We thus also compared theresults from the best model without jump dispersalSince this work focuses on the historical biogeog-
raphy of the Amazonian clade we discarded the twoearly diverging lineages B heilprini and B ranicepsfrom the biogeographic analyses since their distribution
431432433434435436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484
485486487488489490491492493494495496497498499500501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 5
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13560371v1)
antoinefouquet
Texte inseacutereacute
D
PROOF ONLY
ranges lie mostly outside of Amazonia We consideredthree areas within Amazonia Western Amazonia theBrazilian Shield and the Guiana Shield These areascorrespond to major geological features roughly delim-ited by modern riverine barriers the Madeira River theNegro River and the lower course of the Amazon Riverand to three large biogeographic regions known asWallacersquos districts (Hoorn et al 2010 Wallace 1854)These districts were recently confirmed as major breaksin birdsrsquo species composition (Oliveira Vasconcelos ampSantos 2017) and amphibians (Godinho amp da Silva2018 Vacher et al 2020) strengthening their status ofbiogeographic regions Because the distribution range ofthe B albopunctata group extends outside Amazoniawe included in the ancestral range reconstruction oneadditional non-Amazonian Neotropical area theDry Diagonal
ResultsSpecies delimitationOf the three tested methods of species delimitation theABGD method was found to be the most conservativedelimiting 21 OTUs while mPTP and GMYC delimited24 and 47 OTUs respectively (Appendix 1) In onecase the consensual partitioning was overly conserva-tive given that B leucocheila and B multifasciata wereconsidered as a single OTU which is contradicted bythe fact that these two taxa are phenotypically distinctTherefore we kept the different lineages delimited byGMYC in this group as distinct OTUs The final delimi-tation led to 25 OTUs (including B heilprini) in the Balbopunctata species group (Fig 1 Appendix 1)Thirteen of these OTUs could be linked to nominal spe-cies (except B caiapo and B paranaiba that were notincluded in the analysis because material was not avail-able) Conversely 11 could not be linked to any nom-inal species representing a possible 44 increase in thespecies richness of the group The Amazonian cladeitself was represented by 14 OTUs including eight nom-inal species and six putative new species (Fig 1) Theseresults also imply important changes to the geographicdistribution of the species of the group In Appendix 5we detail and justify the identification of these OTUsand their respective geographic ranges
Taxonomic accountsThe Amazonian clade of the B albopunctata group pre-viously reported as the B calcaratandashB fasciata speciescomplex by Caminer and Ron (2014) is composed oftwo major clades (the B calcarata and the B steinbachi
clades) defined by the following combination of charac-ter states (1) truncate snout in dorsal view (2) roundedhead shape and (3) presence of calcaneal appendageon heelThe B calcarata clade (Fig 1) is composed of four
species possessing a calcar and having advertisementcalls formed by a single type of call (Caminer amp Ron2014) The B steinbachi clade on the other handincludes three described and two undescribed species(Fig 1) Members of this clade lack a calcar (skin flapor tubercle present or the complete absence of calcanealappendages) and have vocal repertoires composed ofmore than one type of call except for B alfaroi forwhich a single type of call has been documented so far(Caminer amp Ron 2014)Hyla steinbachi Boulenger 1905 currently valid as
Boana steinbachi (Boulenger 1905) was describedfrom the Bolivian Province Sara Departament SantaCruz de la Sierra Bolivia We assume that Buenavistais the type locality of B steinbachi Between 1910 and1950 the Steinbach family collected many amphibiansand reptiles in Bolivia mainly at Buenavista in theDepartment of Santa Cruz (see Parker 1927) which isthe type locality of several anurans (Hamptophryneboliviana (Parker 1927) Pseudopaludicola bolivianaParker 1927 Scinax parkeri (Gaige 1929)) and thesnake Apostolepis tenuis Ruthven 1927 The materialcollected by the Steinbach family is on display in vari-ous museums such as the University of MichiganMuseum of Zoology (UMMZ see Gaige 1929) theNatural History Museum London (BMNH see Parker1927) the Zoologisches Museum Berlin (ZMB seeMeurouller 1924) and the Naturhistorisches Museum Basel(NBM pers comm Weurouest) Many decades later De laRiva (1990) synonymized H steinbachi with H fas-ciata Subsequently Jansen Bloch Schulze andPfenninger (2011) found differences in call and mtDNAdata between populations of B fasciata from Boliviaand Ecuador and suggested that their results imply a res-urrection of B steinbachi In the following yearsCaminer and Ron (2014) tentatively assigned B steinba-chi to a genetic lineage (clade J) inhabiting BolivianAmazonia near its type locality By assessing its phylo-genetic relationships in the group Caminer and Ron(2014) formally revalidated B steinbachi as a distinctspecies from nominal B fasciata The revalidation of Bsteinbachi was also supported by phenotypic dataobtained from syntypes but did not include a reexamina-tion of all diagnostic characters of the speciesNext we present an amended diagnosis of B steinba-
chi based on novel data on life colours and morphologyand describe its vocal repertoire based on topotypesfrom Bolivia and distinct OTUs from Peru and Brazil
539540541542543544545546547548549550551552553554555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592
593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644645646
6 A Fouquet et al
PROOF ONLY
647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695696697698699700
701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754
Fig 1 Subtrees of the three main clades from the chronogram obtained from the analysis of 16S sequences using BEAST2Terminals are collapsed according to the OTU recovered from the species delineation analysis The distribution of each of these OTUis depicted on the maps
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 7
PROOF ONLY
also assigned to this species in this study Specimensand calls from Acre (ldquoB aff steinbachi 1rdquo) and callsfrom the lower Madre de Dios River (ldquoB aff steinbachi2rdquo) were included in the variation of B steinbachi
Boana steinbachi (Boulenger 1905)Hyla steinbachi mdash Boulenger 1905
Hyla fasciata De la Riva 1990Hypsiboas fasciatus Jansen et al 2011
Hypsiboas sp (Clade G) Funk et al 2012Hypsiboas steinbachi Caminer amp Ron 2014
Boana steinbachi Dubois 2017Boana sp (Clade J) Meza-Joya et al 2019
Boana fasciata Vacher et al 2020
Syntypes BMNH 194721361ndash63 two adults ofunknown sex and one juvenile respectively from Saraprovince Department of Santa Cruz de La SierraBolivia Collected by Hf J Steinbach
Diagnosis Boana steinbachi is characterized by the fol-lowing combination of character states (1) skin flap onheel (2) vocal repertoire composed of more than onecall type (3) multinote call and (4) regular internoteintervals (Figs 2 3 amp 4)
Comparisons with congeners of the Amazonian cladeBoana steinbachi can be distinguished from the membersof the B calcarata clade (B almendarizae B fasciata Bcalcarata and B maculateralis) by the absence of a cal-car and by the vocal repertoire composed of more thanone type of call Among members of the B steinbachiclade it can be distinguished from B dentei B alfaroiand B tetete by the presence of a skin flap on heel (tuber-cle in B alfaroi and B tetete tubercle on one side orcompletely absent in B dentei) by its multinote call (one-note call in B dentei and B tetete) by having regularinternote intervals between call notes (notes with irregularintervals sometimes partly fused one with the next in Balfaroi) and by the vocal repertoire composed of two dis-tinct types of calls (one call type in B alfaroi) (Camineramp Ron 2014 Marinho et al 2020)
Variation Body size varies between 304 and 374mmin males and between 425 and 488mm in females(Appendix 7) Individual AAG-UFU 5918 lacks the heelskin flap on the left side topotypes SMF 88394ndash95 andSMF 88397 lack the skin flap on both sides In life(Fig 3a b) dorsal colouration is beige light brownbright yellow or orange brown in calling males withfaint brown transversal bands and dark purplish-brownfaint brown or grey longitudinal lines extending from
the snout to vent a second line extending from behindthe eye to pelvic region and a third line as a canthalstripe extending from the posterior corner of the nostrilto anterior corner of the eye and from the posterior cor-ner of the eye to midbody length on flank (the topotypesSMF 88394ndash97 lack transversal bands on dorsum indi-viduals SMF 88394 and CORBIDI 13414 lack longitu-dinal lines on dorsum and the middorsal line isextending from snout to interocular region AAG-UFU5921 lacks longitudinal lines on dorsum) Iris cream orgrey sometimes with yellow pigmentation on the upperpart of the iris Throat varying from white to yellowchest and anterior belly varying from white to creamVentral surface of hand and hind limb beige to bright
755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808
809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862
Fig 2 Adult male of Boana steinbachi (AAG-UFU 5921)from the Assis Brasil population (Acre Brazil) (A) dorsal and(B) ventral views of the body (SVL frac14 337mm) (C) dorsaland (D) lateral views of the head (HL frac14123mm HW frac14102mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 106mm) and (G) sole of thefoot (FL frac14 139mm)
8 A Fouquet et al
antoinefouquet
Texte surligneacute
we may wanna ask for full page here no
PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Boana as well as the MRCA of a clade formed byHyloscirtus Boana Aplastodiscus andBokermannohyla which were estimated by Feng et al(2017) and consequently constrained here assuming nor-mal prior distributions of 252Ma (standard deviation[SD]frac14 28Ma) and 323Ma (SD frac14 3Ma) respectivelyWe considered these calibrations to represent the bestprior because Feng et al (2017) included a comprehen-sive dataset of nuclear loci for an extensive sampling ofanurans and used many reliable fossils as calibrationpoints Moreover Hime et al (2020) analysing an evenlarger genomic dataset found very similar time esti-mates for the TMRCA of BoanathornHyloscirtus Insteadusing fossil andor biogeographic calibrations wouldhave implied to expand the matrix to lineages distantlyrelated to Boana and would likely lead to an overesti-mation of calibration dates with our mitogenomic data-set (Molak amp Ho 2015 Papadopoulou et al 2010) Forthe MCMC parameters we used four independentchains of 100 million iterations recording every10000th iteration and a 10 burn-in We combined thelog files of the independent runs using LogCombiner25 (Drummond amp Rambaut 2007) and checked theconvergence of our parameters confirmed by allEffective Sample Size (ESS) being above 200 Then weextracted the maximum clade credibility tree using Treeannotator 25 (Drummond amp Rambaut 2007) with aburn-in of 10 After removing outgroups we per-formed a GMYC delimitation on the ultrametric treeusing the GMYC function of the splits R package v10-11 (Ezard Fujisawa amp Barraclough 2009) with athreshold interval between 0 and 10Ma and by usingthe single threshold method Operational TaxonomicUnits (OTUs) were defined using a majority-rule con-sensus from the results of the three methods ie a lin-eage is considered as being an OTU if supported by atleast two of the three methodsAll but five occurrence records were georeferenced
and used to create distribution maps with convex poly-gons under QGIS 214 (QGIS Geographic InformationSystem) OTUs were assigned to taxa based on thefieldmuseum identification and sometimes corrected inaccordance with type localities and known distribution(Frost 2019 IUCN 2020)
Morphological and acoustic variation in theBoana steinbachi cladeThe following morphometric measurements were takenon 38 males and 9 females snout-vent length (SVL)head length (HL) head width (HW) eye diameter (ED)tympanum diameter (TD) tibia length (TL) foot length(FL) thigh length (THL) and calcaneal appendage
length (CL) In addition to these we also measuredhand length (HAL) forearm length (FLL) and eyendashnos-tril distance (EN) for the morphometric characterizationof holotypes All measurements followed the definitionsand terminology of Watters Cummings Flanagan andSiler (2016) We classified the calcaneal appendage intothree-character states (i) a calcar (skin appendage withlength gt 1mm flattened or conical) a (ii) skin flap(skin appendage with length lt 1mm flattened or a(iii) tubercle a conical structure with reduced size andas such not forming an appendage coming out of theheel The specimens were measured by P Marinho(Alta Floresta Mato Grosso State and Assis BrazilAcre State) using a Mitutoyo digital calliper (to thenearest 005mm) by MT Rodrigues (Jirau and PacaasNovos Rondonia State) using a Mitutoyo digital calliper(to the nearest 001mm) by A Fouquet (French Guianaand Suriname) using a DigiMax digital calliper (nearest001mm) and by M Caminer (Ecuador Peru andBolivia) using a Mannesmann digital calliper (nearest001mm) Snout shape was assessed according to Heyeret al (1990) Digital webbing formulae followed thenotation system of Savage and Heyer (1997)Acoustic analysis of 27 recorded calling males
(Appendix 4) was conducted in Raven Pro 15 64-bit ver-sion (Bioacoustics Research Program 2012) sound figureswere produced using Seewave version 210 (SueurAubin amp Simonis 2008) and tuneR version 132(Ligges Krey Mersmann amp Schnackenberg 2014) in Rversion 350 (R Core Team 2018) Raven Pro settingswindow size frac14 512 samples window typefrac14Hann 3 dBfilter bandwidth frac14 124Hz window overlap frac14 85 hopsize frac14 77ms discrete Fourier transform (DFT) size frac141024 samples grid spacing frac14 431Hz seewave settingswindow typefrac14Hanning fast Fourier transform (FFT)size frac14 256 samples FFT overlap frac14 90 The analysedfiles are deposited in the sound collection of the lsquoMuseude Biodiversidade do Cerradorsquo (AAG-UFU) lsquoFonotecaNeotropical Jacques Vielardirsquo (FNJV) lsquoLa Sonotheque duMuseum National dHistoire Naturellersquo (MNHN) and theMacaulay library (MC) Detailed information about theanalysed sound files and accession numbers are providedin Appendix 4Call types were distinguished based on the number of
notes per call and pulsing We defined as type 1 callsthe multinote calls formed by nonpulsed notes whichconsist of the main call type emitted by males of allstudied species Type 2 calls are emitted less often thantype 1 and consist of a one-note call formed by poorlydefined pulses We only included type 1 calls in theacoustic diagnoses and the interspecific comparisonsThe acoustic traits were generally analysed manually (ifnot stated otherwise) as follows temporal traits (call
323324325326327328329330331332333334335336337338339340341342343344345346347348349350351352353354355356357358359360361362363364365366367368369370371372373374375376
377378379380381382383384385386387388389390391392393394395396397398399400401402403404405406407408409410411412413414415416417418419420421422423424425426427428429430
4 A Fouquet et al
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
1
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics 2014
antoinefouquet
Texte inseacutereacute
a
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
s
PROOF ONLY
duration note duration interval between notes numberof notes and call rise time the latter using the lsquoPeakTimersquo function) frequency traits (dominant frequencyusing the lsquoPeak Frequencyrsquo function minimum andmaximum frequency using the lsquoFrequency 5rsquo andlsquoFrequency 95rsquo functions respectively) Call traitsused in the description followed the definitions and ter-minology of Keuroohler et al (2017) using a note-cen-tred approach
Time-calibrated species phylogenyWe selected a representative for each of the 25 OTUsidentified by the species delimitation (see Results) forcomplete mitogenome sequencing to investigate interre-lationships and divergence times Mitogenomic sequen-ces were obtained using low-coverage shotgunsequencing We recovered high-quality mitochondrialgenome assemblies for 18 OTU representatives (seeAppendix 2 for details regarding mitogenome sequenc-ing assembling and annotation) For the remainingseven OTUs for which we could not obtain tissue sam-ples (Appendix 3) we gathered all the available mito-chondrial loci (12S 16S ND1 COI and Cytb) fromGenBank four of these OTUs were represented by 16Sonly We also selected 14 species as outgroups (ninewith complete mitogenomes) including representativesfor other Boana species groups and most otherCophomantini genera (Appendix 3)We extracted 12S 16S and all protein-coding
sequence regions (CDS thus removing D-Loop andtRNAs) from complete mitogenomes as well asGenBank accessions and aligned each locus independ-ently using the MAFFT7 online server For rRNAgenes we chose the E-INS-i strategy recommended forsequences with multiple conserved domains and longgaps For the protein-coding genes we chose the G-INS-i strategy designed for sequences with global hom-ology (Katoh et al 2019) Realignments of CDS con-sidering the reading frame were done and concatenatedin Geneious v918 (httpswwwgeneiouscom) Ourfinal matrix totalled 39 terminals and 13620 alignednucleotide sites Among these terminals 27 were com-plete and nine had fewer than 3000 nucleotides (20)but all were putatively closely related to terminals withcomplete dataWe selected the best-fit partition scheme and model
of evolution for each partition using PartitionFinderV211 (Lanfear et al 2016) based on the BayesianInformation Criterion (BIC) We predefined four datablocks one for rRNA genes (12S and 16S) and one foreach codon position of all CDS regions and found theGTRthorn IthornG model to best-fit all partitions We
reconstructed a time-calibrated tree using a birth-deathtree prior using BEAST 25 to account for extinctionprocesses and incomplete sampling We parameterizedunlinked substitution models according to the estimatesobtained in the PartitionFinder V211 analysis (Lanfearet al 2016) We used the same two calibration pointsas in the previous BEAST analysis Analyses wereundertaken using uncorrelated relaxed lognormal clockmodel of distribution of rate variation among branchesfor each partition (Drummond et al 2006) The Markovchain Monte Carlo (MCMC) parameters were set withfour independent chains of 100 million iterations stor-ing every 10000th iteration and a 10 burn-in Wecombined the log files of the independent runs usingLogCombiner 25 and confirmed the convergence of ourparameters as all ESS were above 200 Finally weextracted the maximum clade credibility tree using Treeannotator 25 We acknowledge that our phylogeneticreconstruction solely based on mtDNA sequences priori-tizes spatial and taxonomic completeness over genomiccoverage and can lead to overestimated divergence times(eg McCormack Heled Delaney Peterson ampKnowles 2011 Near et al 2012)
Biogeographic analysisBiogeographic inferences were undertaken on the time-calibrated phylogeny using the BioGeoBEARS R pack-age (Matzke 2013) This package reconstructs ancestralgeographic distributions and investigates the role ofeach biogeographic event with a maximum likelihoodalgorithm We compared three different models (i) alikelihood version of the Dispersal-Vicariance(DIVALIKE) model (Ronquist 1997) (ii) a likelihoodversion of the BayArea (BBM) model (Landis et al2013) and (iii) the Dispersal Extinction Cladogenesis(DEC) model (Ree amp Smith 2008) We also comparedversions of these models allowing jump dispersal asdescribed by the J parameter (Matzke 2013) Modelswere compared with the Akaike Information Criterion(AIC) We ran 50 independent BioGeoBEARS biogeo-graphic stochastic mapping to determine biogeographicevent counts for the best-fit model (Dupin et al 2017)Ree and Sanmartın (2018) voiced several criticismsagainst the use of J parameter arguing that it inflates thecontribution of cladogenetic events to the likelihoodand minimizes the contribution of anagenetic time-dependent range evolution We thus also compared theresults from the best model without jump dispersalSince this work focuses on the historical biogeog-
raphy of the Amazonian clade we discarded the twoearly diverging lineages B heilprini and B ranicepsfrom the biogeographic analyses since their distribution
431432433434435436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484
485486487488489490491492493494495496497498499500501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 5
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13560371v1)
antoinefouquet
Texte inseacutereacute
D
PROOF ONLY
ranges lie mostly outside of Amazonia We consideredthree areas within Amazonia Western Amazonia theBrazilian Shield and the Guiana Shield These areascorrespond to major geological features roughly delim-ited by modern riverine barriers the Madeira River theNegro River and the lower course of the Amazon Riverand to three large biogeographic regions known asWallacersquos districts (Hoorn et al 2010 Wallace 1854)These districts were recently confirmed as major breaksin birdsrsquo species composition (Oliveira Vasconcelos ampSantos 2017) and amphibians (Godinho amp da Silva2018 Vacher et al 2020) strengthening their status ofbiogeographic regions Because the distribution range ofthe B albopunctata group extends outside Amazoniawe included in the ancestral range reconstruction oneadditional non-Amazonian Neotropical area theDry Diagonal
ResultsSpecies delimitationOf the three tested methods of species delimitation theABGD method was found to be the most conservativedelimiting 21 OTUs while mPTP and GMYC delimited24 and 47 OTUs respectively (Appendix 1) In onecase the consensual partitioning was overly conserva-tive given that B leucocheila and B multifasciata wereconsidered as a single OTU which is contradicted bythe fact that these two taxa are phenotypically distinctTherefore we kept the different lineages delimited byGMYC in this group as distinct OTUs The final delimi-tation led to 25 OTUs (including B heilprini) in the Balbopunctata species group (Fig 1 Appendix 1)Thirteen of these OTUs could be linked to nominal spe-cies (except B caiapo and B paranaiba that were notincluded in the analysis because material was not avail-able) Conversely 11 could not be linked to any nom-inal species representing a possible 44 increase in thespecies richness of the group The Amazonian cladeitself was represented by 14 OTUs including eight nom-inal species and six putative new species (Fig 1) Theseresults also imply important changes to the geographicdistribution of the species of the group In Appendix 5we detail and justify the identification of these OTUsand their respective geographic ranges
Taxonomic accountsThe Amazonian clade of the B albopunctata group pre-viously reported as the B calcaratandashB fasciata speciescomplex by Caminer and Ron (2014) is composed oftwo major clades (the B calcarata and the B steinbachi
clades) defined by the following combination of charac-ter states (1) truncate snout in dorsal view (2) roundedhead shape and (3) presence of calcaneal appendageon heelThe B calcarata clade (Fig 1) is composed of four
species possessing a calcar and having advertisementcalls formed by a single type of call (Caminer amp Ron2014) The B steinbachi clade on the other handincludes three described and two undescribed species(Fig 1) Members of this clade lack a calcar (skin flapor tubercle present or the complete absence of calcanealappendages) and have vocal repertoires composed ofmore than one type of call except for B alfaroi forwhich a single type of call has been documented so far(Caminer amp Ron 2014)Hyla steinbachi Boulenger 1905 currently valid as
Boana steinbachi (Boulenger 1905) was describedfrom the Bolivian Province Sara Departament SantaCruz de la Sierra Bolivia We assume that Buenavistais the type locality of B steinbachi Between 1910 and1950 the Steinbach family collected many amphibiansand reptiles in Bolivia mainly at Buenavista in theDepartment of Santa Cruz (see Parker 1927) which isthe type locality of several anurans (Hamptophryneboliviana (Parker 1927) Pseudopaludicola bolivianaParker 1927 Scinax parkeri (Gaige 1929)) and thesnake Apostolepis tenuis Ruthven 1927 The materialcollected by the Steinbach family is on display in vari-ous museums such as the University of MichiganMuseum of Zoology (UMMZ see Gaige 1929) theNatural History Museum London (BMNH see Parker1927) the Zoologisches Museum Berlin (ZMB seeMeurouller 1924) and the Naturhistorisches Museum Basel(NBM pers comm Weurouest) Many decades later De laRiva (1990) synonymized H steinbachi with H fas-ciata Subsequently Jansen Bloch Schulze andPfenninger (2011) found differences in call and mtDNAdata between populations of B fasciata from Boliviaand Ecuador and suggested that their results imply a res-urrection of B steinbachi In the following yearsCaminer and Ron (2014) tentatively assigned B steinba-chi to a genetic lineage (clade J) inhabiting BolivianAmazonia near its type locality By assessing its phylo-genetic relationships in the group Caminer and Ron(2014) formally revalidated B steinbachi as a distinctspecies from nominal B fasciata The revalidation of Bsteinbachi was also supported by phenotypic dataobtained from syntypes but did not include a reexamina-tion of all diagnostic characters of the speciesNext we present an amended diagnosis of B steinba-
chi based on novel data on life colours and morphologyand describe its vocal repertoire based on topotypesfrom Bolivia and distinct OTUs from Peru and Brazil
539540541542543544545546547548549550551552553554555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592
593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644645646
6 A Fouquet et al
PROOF ONLY
647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695696697698699700
701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754
Fig 1 Subtrees of the three main clades from the chronogram obtained from the analysis of 16S sequences using BEAST2Terminals are collapsed according to the OTU recovered from the species delineation analysis The distribution of each of these OTUis depicted on the maps
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 7
PROOF ONLY
also assigned to this species in this study Specimensand calls from Acre (ldquoB aff steinbachi 1rdquo) and callsfrom the lower Madre de Dios River (ldquoB aff steinbachi2rdquo) were included in the variation of B steinbachi
Boana steinbachi (Boulenger 1905)Hyla steinbachi mdash Boulenger 1905
Hyla fasciata De la Riva 1990Hypsiboas fasciatus Jansen et al 2011
Hypsiboas sp (Clade G) Funk et al 2012Hypsiboas steinbachi Caminer amp Ron 2014
Boana steinbachi Dubois 2017Boana sp (Clade J) Meza-Joya et al 2019
Boana fasciata Vacher et al 2020
Syntypes BMNH 194721361ndash63 two adults ofunknown sex and one juvenile respectively from Saraprovince Department of Santa Cruz de La SierraBolivia Collected by Hf J Steinbach
Diagnosis Boana steinbachi is characterized by the fol-lowing combination of character states (1) skin flap onheel (2) vocal repertoire composed of more than onecall type (3) multinote call and (4) regular internoteintervals (Figs 2 3 amp 4)
Comparisons with congeners of the Amazonian cladeBoana steinbachi can be distinguished from the membersof the B calcarata clade (B almendarizae B fasciata Bcalcarata and B maculateralis) by the absence of a cal-car and by the vocal repertoire composed of more thanone type of call Among members of the B steinbachiclade it can be distinguished from B dentei B alfaroiand B tetete by the presence of a skin flap on heel (tuber-cle in B alfaroi and B tetete tubercle on one side orcompletely absent in B dentei) by its multinote call (one-note call in B dentei and B tetete) by having regularinternote intervals between call notes (notes with irregularintervals sometimes partly fused one with the next in Balfaroi) and by the vocal repertoire composed of two dis-tinct types of calls (one call type in B alfaroi) (Camineramp Ron 2014 Marinho et al 2020)
Variation Body size varies between 304 and 374mmin males and between 425 and 488mm in females(Appendix 7) Individual AAG-UFU 5918 lacks the heelskin flap on the left side topotypes SMF 88394ndash95 andSMF 88397 lack the skin flap on both sides In life(Fig 3a b) dorsal colouration is beige light brownbright yellow or orange brown in calling males withfaint brown transversal bands and dark purplish-brownfaint brown or grey longitudinal lines extending from
the snout to vent a second line extending from behindthe eye to pelvic region and a third line as a canthalstripe extending from the posterior corner of the nostrilto anterior corner of the eye and from the posterior cor-ner of the eye to midbody length on flank (the topotypesSMF 88394ndash97 lack transversal bands on dorsum indi-viduals SMF 88394 and CORBIDI 13414 lack longitu-dinal lines on dorsum and the middorsal line isextending from snout to interocular region AAG-UFU5921 lacks longitudinal lines on dorsum) Iris cream orgrey sometimes with yellow pigmentation on the upperpart of the iris Throat varying from white to yellowchest and anterior belly varying from white to creamVentral surface of hand and hind limb beige to bright
755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808
809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862
Fig 2 Adult male of Boana steinbachi (AAG-UFU 5921)from the Assis Brasil population (Acre Brazil) (A) dorsal and(B) ventral views of the body (SVL frac14 337mm) (C) dorsaland (D) lateral views of the head (HL frac14123mm HW frac14102mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 106mm) and (G) sole of thefoot (FL frac14 139mm)
8 A Fouquet et al
antoinefouquet
Texte surligneacute
we may wanna ask for full page here no
PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
duration note duration interval between notes numberof notes and call rise time the latter using the lsquoPeakTimersquo function) frequency traits (dominant frequencyusing the lsquoPeak Frequencyrsquo function minimum andmaximum frequency using the lsquoFrequency 5rsquo andlsquoFrequency 95rsquo functions respectively) Call traitsused in the description followed the definitions and ter-minology of Keuroohler et al (2017) using a note-cen-tred approach
Time-calibrated species phylogenyWe selected a representative for each of the 25 OTUsidentified by the species delimitation (see Results) forcomplete mitogenome sequencing to investigate interre-lationships and divergence times Mitogenomic sequen-ces were obtained using low-coverage shotgunsequencing We recovered high-quality mitochondrialgenome assemblies for 18 OTU representatives (seeAppendix 2 for details regarding mitogenome sequenc-ing assembling and annotation) For the remainingseven OTUs for which we could not obtain tissue sam-ples (Appendix 3) we gathered all the available mito-chondrial loci (12S 16S ND1 COI and Cytb) fromGenBank four of these OTUs were represented by 16Sonly We also selected 14 species as outgroups (ninewith complete mitogenomes) including representativesfor other Boana species groups and most otherCophomantini genera (Appendix 3)We extracted 12S 16S and all protein-coding
sequence regions (CDS thus removing D-Loop andtRNAs) from complete mitogenomes as well asGenBank accessions and aligned each locus independ-ently using the MAFFT7 online server For rRNAgenes we chose the E-INS-i strategy recommended forsequences with multiple conserved domains and longgaps For the protein-coding genes we chose the G-INS-i strategy designed for sequences with global hom-ology (Katoh et al 2019) Realignments of CDS con-sidering the reading frame were done and concatenatedin Geneious v918 (httpswwwgeneiouscom) Ourfinal matrix totalled 39 terminals and 13620 alignednucleotide sites Among these terminals 27 were com-plete and nine had fewer than 3000 nucleotides (20)but all were putatively closely related to terminals withcomplete dataWe selected the best-fit partition scheme and model
of evolution for each partition using PartitionFinderV211 (Lanfear et al 2016) based on the BayesianInformation Criterion (BIC) We predefined four datablocks one for rRNA genes (12S and 16S) and one foreach codon position of all CDS regions and found theGTRthorn IthornG model to best-fit all partitions We
reconstructed a time-calibrated tree using a birth-deathtree prior using BEAST 25 to account for extinctionprocesses and incomplete sampling We parameterizedunlinked substitution models according to the estimatesobtained in the PartitionFinder V211 analysis (Lanfearet al 2016) We used the same two calibration pointsas in the previous BEAST analysis Analyses wereundertaken using uncorrelated relaxed lognormal clockmodel of distribution of rate variation among branchesfor each partition (Drummond et al 2006) The Markovchain Monte Carlo (MCMC) parameters were set withfour independent chains of 100 million iterations stor-ing every 10000th iteration and a 10 burn-in Wecombined the log files of the independent runs usingLogCombiner 25 and confirmed the convergence of ourparameters as all ESS were above 200 Finally weextracted the maximum clade credibility tree using Treeannotator 25 We acknowledge that our phylogeneticreconstruction solely based on mtDNA sequences priori-tizes spatial and taxonomic completeness over genomiccoverage and can lead to overestimated divergence times(eg McCormack Heled Delaney Peterson ampKnowles 2011 Near et al 2012)
Biogeographic analysisBiogeographic inferences were undertaken on the time-calibrated phylogeny using the BioGeoBEARS R pack-age (Matzke 2013) This package reconstructs ancestralgeographic distributions and investigates the role ofeach biogeographic event with a maximum likelihoodalgorithm We compared three different models (i) alikelihood version of the Dispersal-Vicariance(DIVALIKE) model (Ronquist 1997) (ii) a likelihoodversion of the BayArea (BBM) model (Landis et al2013) and (iii) the Dispersal Extinction Cladogenesis(DEC) model (Ree amp Smith 2008) We also comparedversions of these models allowing jump dispersal asdescribed by the J parameter (Matzke 2013) Modelswere compared with the Akaike Information Criterion(AIC) We ran 50 independent BioGeoBEARS biogeo-graphic stochastic mapping to determine biogeographicevent counts for the best-fit model (Dupin et al 2017)Ree and Sanmartın (2018) voiced several criticismsagainst the use of J parameter arguing that it inflates thecontribution of cladogenetic events to the likelihoodand minimizes the contribution of anagenetic time-dependent range evolution We thus also compared theresults from the best model without jump dispersalSince this work focuses on the historical biogeog-
raphy of the Amazonian clade we discarded the twoearly diverging lineages B heilprini and B ranicepsfrom the biogeographic analyses since their distribution
431432433434435436437438439440441442443444445446447448449450451452453454455456457458459460461462463464465466467468469470471472473474475476477478479480481482483484
485486487488489490491492493494495496497498499500501502503504505506507508509510511512513514515516517518519520521522523524525526527528529530531532533534535536537538
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 5
antoinefouquet
Texte inseacutereacute
(alignment available from figsharecom at httpsdoiorg106084m9figshare13560371v1)
antoinefouquet
Texte inseacutereacute
D
PROOF ONLY
ranges lie mostly outside of Amazonia We consideredthree areas within Amazonia Western Amazonia theBrazilian Shield and the Guiana Shield These areascorrespond to major geological features roughly delim-ited by modern riverine barriers the Madeira River theNegro River and the lower course of the Amazon Riverand to three large biogeographic regions known asWallacersquos districts (Hoorn et al 2010 Wallace 1854)These districts were recently confirmed as major breaksin birdsrsquo species composition (Oliveira Vasconcelos ampSantos 2017) and amphibians (Godinho amp da Silva2018 Vacher et al 2020) strengthening their status ofbiogeographic regions Because the distribution range ofthe B albopunctata group extends outside Amazoniawe included in the ancestral range reconstruction oneadditional non-Amazonian Neotropical area theDry Diagonal
ResultsSpecies delimitationOf the three tested methods of species delimitation theABGD method was found to be the most conservativedelimiting 21 OTUs while mPTP and GMYC delimited24 and 47 OTUs respectively (Appendix 1) In onecase the consensual partitioning was overly conserva-tive given that B leucocheila and B multifasciata wereconsidered as a single OTU which is contradicted bythe fact that these two taxa are phenotypically distinctTherefore we kept the different lineages delimited byGMYC in this group as distinct OTUs The final delimi-tation led to 25 OTUs (including B heilprini) in the Balbopunctata species group (Fig 1 Appendix 1)Thirteen of these OTUs could be linked to nominal spe-cies (except B caiapo and B paranaiba that were notincluded in the analysis because material was not avail-able) Conversely 11 could not be linked to any nom-inal species representing a possible 44 increase in thespecies richness of the group The Amazonian cladeitself was represented by 14 OTUs including eight nom-inal species and six putative new species (Fig 1) Theseresults also imply important changes to the geographicdistribution of the species of the group In Appendix 5we detail and justify the identification of these OTUsand their respective geographic ranges
Taxonomic accountsThe Amazonian clade of the B albopunctata group pre-viously reported as the B calcaratandashB fasciata speciescomplex by Caminer and Ron (2014) is composed oftwo major clades (the B calcarata and the B steinbachi
clades) defined by the following combination of charac-ter states (1) truncate snout in dorsal view (2) roundedhead shape and (3) presence of calcaneal appendageon heelThe B calcarata clade (Fig 1) is composed of four
species possessing a calcar and having advertisementcalls formed by a single type of call (Caminer amp Ron2014) The B steinbachi clade on the other handincludes three described and two undescribed species(Fig 1) Members of this clade lack a calcar (skin flapor tubercle present or the complete absence of calcanealappendages) and have vocal repertoires composed ofmore than one type of call except for B alfaroi forwhich a single type of call has been documented so far(Caminer amp Ron 2014)Hyla steinbachi Boulenger 1905 currently valid as
Boana steinbachi (Boulenger 1905) was describedfrom the Bolivian Province Sara Departament SantaCruz de la Sierra Bolivia We assume that Buenavistais the type locality of B steinbachi Between 1910 and1950 the Steinbach family collected many amphibiansand reptiles in Bolivia mainly at Buenavista in theDepartment of Santa Cruz (see Parker 1927) which isthe type locality of several anurans (Hamptophryneboliviana (Parker 1927) Pseudopaludicola bolivianaParker 1927 Scinax parkeri (Gaige 1929)) and thesnake Apostolepis tenuis Ruthven 1927 The materialcollected by the Steinbach family is on display in vari-ous museums such as the University of MichiganMuseum of Zoology (UMMZ see Gaige 1929) theNatural History Museum London (BMNH see Parker1927) the Zoologisches Museum Berlin (ZMB seeMeurouller 1924) and the Naturhistorisches Museum Basel(NBM pers comm Weurouest) Many decades later De laRiva (1990) synonymized H steinbachi with H fas-ciata Subsequently Jansen Bloch Schulze andPfenninger (2011) found differences in call and mtDNAdata between populations of B fasciata from Boliviaand Ecuador and suggested that their results imply a res-urrection of B steinbachi In the following yearsCaminer and Ron (2014) tentatively assigned B steinba-chi to a genetic lineage (clade J) inhabiting BolivianAmazonia near its type locality By assessing its phylo-genetic relationships in the group Caminer and Ron(2014) formally revalidated B steinbachi as a distinctspecies from nominal B fasciata The revalidation of Bsteinbachi was also supported by phenotypic dataobtained from syntypes but did not include a reexamina-tion of all diagnostic characters of the speciesNext we present an amended diagnosis of B steinba-
chi based on novel data on life colours and morphologyand describe its vocal repertoire based on topotypesfrom Bolivia and distinct OTUs from Peru and Brazil
539540541542543544545546547548549550551552553554555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592
593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644645646
6 A Fouquet et al
PROOF ONLY
647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695696697698699700
701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754
Fig 1 Subtrees of the three main clades from the chronogram obtained from the analysis of 16S sequences using BEAST2Terminals are collapsed according to the OTU recovered from the species delineation analysis The distribution of each of these OTUis depicted on the maps
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 7
PROOF ONLY
also assigned to this species in this study Specimensand calls from Acre (ldquoB aff steinbachi 1rdquo) and callsfrom the lower Madre de Dios River (ldquoB aff steinbachi2rdquo) were included in the variation of B steinbachi
Boana steinbachi (Boulenger 1905)Hyla steinbachi mdash Boulenger 1905
Hyla fasciata De la Riva 1990Hypsiboas fasciatus Jansen et al 2011
Hypsiboas sp (Clade G) Funk et al 2012Hypsiboas steinbachi Caminer amp Ron 2014
Boana steinbachi Dubois 2017Boana sp (Clade J) Meza-Joya et al 2019
Boana fasciata Vacher et al 2020
Syntypes BMNH 194721361ndash63 two adults ofunknown sex and one juvenile respectively from Saraprovince Department of Santa Cruz de La SierraBolivia Collected by Hf J Steinbach
Diagnosis Boana steinbachi is characterized by the fol-lowing combination of character states (1) skin flap onheel (2) vocal repertoire composed of more than onecall type (3) multinote call and (4) regular internoteintervals (Figs 2 3 amp 4)
Comparisons with congeners of the Amazonian cladeBoana steinbachi can be distinguished from the membersof the B calcarata clade (B almendarizae B fasciata Bcalcarata and B maculateralis) by the absence of a cal-car and by the vocal repertoire composed of more thanone type of call Among members of the B steinbachiclade it can be distinguished from B dentei B alfaroiand B tetete by the presence of a skin flap on heel (tuber-cle in B alfaroi and B tetete tubercle on one side orcompletely absent in B dentei) by its multinote call (one-note call in B dentei and B tetete) by having regularinternote intervals between call notes (notes with irregularintervals sometimes partly fused one with the next in Balfaroi) and by the vocal repertoire composed of two dis-tinct types of calls (one call type in B alfaroi) (Camineramp Ron 2014 Marinho et al 2020)
Variation Body size varies between 304 and 374mmin males and between 425 and 488mm in females(Appendix 7) Individual AAG-UFU 5918 lacks the heelskin flap on the left side topotypes SMF 88394ndash95 andSMF 88397 lack the skin flap on both sides In life(Fig 3a b) dorsal colouration is beige light brownbright yellow or orange brown in calling males withfaint brown transversal bands and dark purplish-brownfaint brown or grey longitudinal lines extending from
the snout to vent a second line extending from behindthe eye to pelvic region and a third line as a canthalstripe extending from the posterior corner of the nostrilto anterior corner of the eye and from the posterior cor-ner of the eye to midbody length on flank (the topotypesSMF 88394ndash97 lack transversal bands on dorsum indi-viduals SMF 88394 and CORBIDI 13414 lack longitu-dinal lines on dorsum and the middorsal line isextending from snout to interocular region AAG-UFU5921 lacks longitudinal lines on dorsum) Iris cream orgrey sometimes with yellow pigmentation on the upperpart of the iris Throat varying from white to yellowchest and anterior belly varying from white to creamVentral surface of hand and hind limb beige to bright
755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808
809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862
Fig 2 Adult male of Boana steinbachi (AAG-UFU 5921)from the Assis Brasil population (Acre Brazil) (A) dorsal and(B) ventral views of the body (SVL frac14 337mm) (C) dorsaland (D) lateral views of the head (HL frac14123mm HW frac14102mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 106mm) and (G) sole of thefoot (FL frac14 139mm)
8 A Fouquet et al
antoinefouquet
Texte surligneacute
we may wanna ask for full page here no
PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
ranges lie mostly outside of Amazonia We consideredthree areas within Amazonia Western Amazonia theBrazilian Shield and the Guiana Shield These areascorrespond to major geological features roughly delim-ited by modern riverine barriers the Madeira River theNegro River and the lower course of the Amazon Riverand to three large biogeographic regions known asWallacersquos districts (Hoorn et al 2010 Wallace 1854)These districts were recently confirmed as major breaksin birdsrsquo species composition (Oliveira Vasconcelos ampSantos 2017) and amphibians (Godinho amp da Silva2018 Vacher et al 2020) strengthening their status ofbiogeographic regions Because the distribution range ofthe B albopunctata group extends outside Amazoniawe included in the ancestral range reconstruction oneadditional non-Amazonian Neotropical area theDry Diagonal
ResultsSpecies delimitationOf the three tested methods of species delimitation theABGD method was found to be the most conservativedelimiting 21 OTUs while mPTP and GMYC delimited24 and 47 OTUs respectively (Appendix 1) In onecase the consensual partitioning was overly conserva-tive given that B leucocheila and B multifasciata wereconsidered as a single OTU which is contradicted bythe fact that these two taxa are phenotypically distinctTherefore we kept the different lineages delimited byGMYC in this group as distinct OTUs The final delimi-tation led to 25 OTUs (including B heilprini) in the Balbopunctata species group (Fig 1 Appendix 1)Thirteen of these OTUs could be linked to nominal spe-cies (except B caiapo and B paranaiba that were notincluded in the analysis because material was not avail-able) Conversely 11 could not be linked to any nom-inal species representing a possible 44 increase in thespecies richness of the group The Amazonian cladeitself was represented by 14 OTUs including eight nom-inal species and six putative new species (Fig 1) Theseresults also imply important changes to the geographicdistribution of the species of the group In Appendix 5we detail and justify the identification of these OTUsand their respective geographic ranges
Taxonomic accountsThe Amazonian clade of the B albopunctata group pre-viously reported as the B calcaratandashB fasciata speciescomplex by Caminer and Ron (2014) is composed oftwo major clades (the B calcarata and the B steinbachi
clades) defined by the following combination of charac-ter states (1) truncate snout in dorsal view (2) roundedhead shape and (3) presence of calcaneal appendageon heelThe B calcarata clade (Fig 1) is composed of four
species possessing a calcar and having advertisementcalls formed by a single type of call (Caminer amp Ron2014) The B steinbachi clade on the other handincludes three described and two undescribed species(Fig 1) Members of this clade lack a calcar (skin flapor tubercle present or the complete absence of calcanealappendages) and have vocal repertoires composed ofmore than one type of call except for B alfaroi forwhich a single type of call has been documented so far(Caminer amp Ron 2014)Hyla steinbachi Boulenger 1905 currently valid as
Boana steinbachi (Boulenger 1905) was describedfrom the Bolivian Province Sara Departament SantaCruz de la Sierra Bolivia We assume that Buenavistais the type locality of B steinbachi Between 1910 and1950 the Steinbach family collected many amphibiansand reptiles in Bolivia mainly at Buenavista in theDepartment of Santa Cruz (see Parker 1927) which isthe type locality of several anurans (Hamptophryneboliviana (Parker 1927) Pseudopaludicola bolivianaParker 1927 Scinax parkeri (Gaige 1929)) and thesnake Apostolepis tenuis Ruthven 1927 The materialcollected by the Steinbach family is on display in vari-ous museums such as the University of MichiganMuseum of Zoology (UMMZ see Gaige 1929) theNatural History Museum London (BMNH see Parker1927) the Zoologisches Museum Berlin (ZMB seeMeurouller 1924) and the Naturhistorisches Museum Basel(NBM pers comm Weurouest) Many decades later De laRiva (1990) synonymized H steinbachi with H fas-ciata Subsequently Jansen Bloch Schulze andPfenninger (2011) found differences in call and mtDNAdata between populations of B fasciata from Boliviaand Ecuador and suggested that their results imply a res-urrection of B steinbachi In the following yearsCaminer and Ron (2014) tentatively assigned B steinba-chi to a genetic lineage (clade J) inhabiting BolivianAmazonia near its type locality By assessing its phylo-genetic relationships in the group Caminer and Ron(2014) formally revalidated B steinbachi as a distinctspecies from nominal B fasciata The revalidation of Bsteinbachi was also supported by phenotypic dataobtained from syntypes but did not include a reexamina-tion of all diagnostic characters of the speciesNext we present an amended diagnosis of B steinba-
chi based on novel data on life colours and morphologyand describe its vocal repertoire based on topotypesfrom Bolivia and distinct OTUs from Peru and Brazil
539540541542543544545546547548549550551552553554555556557558559560561562563564565566567568569570571572573574575576577578579580581582583584585586587588589590591592
593594595596597598599600601602603604605606607608609610611612613614615616617618619620621622623624625626627628629630631632633634635636637638639640641642643644645646
6 A Fouquet et al
PROOF ONLY
647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695696697698699700
701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754
Fig 1 Subtrees of the three main clades from the chronogram obtained from the analysis of 16S sequences using BEAST2Terminals are collapsed according to the OTU recovered from the species delineation analysis The distribution of each of these OTUis depicted on the maps
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 7
PROOF ONLY
also assigned to this species in this study Specimensand calls from Acre (ldquoB aff steinbachi 1rdquo) and callsfrom the lower Madre de Dios River (ldquoB aff steinbachi2rdquo) were included in the variation of B steinbachi
Boana steinbachi (Boulenger 1905)Hyla steinbachi mdash Boulenger 1905
Hyla fasciata De la Riva 1990Hypsiboas fasciatus Jansen et al 2011
Hypsiboas sp (Clade G) Funk et al 2012Hypsiboas steinbachi Caminer amp Ron 2014
Boana steinbachi Dubois 2017Boana sp (Clade J) Meza-Joya et al 2019
Boana fasciata Vacher et al 2020
Syntypes BMNH 194721361ndash63 two adults ofunknown sex and one juvenile respectively from Saraprovince Department of Santa Cruz de La SierraBolivia Collected by Hf J Steinbach
Diagnosis Boana steinbachi is characterized by the fol-lowing combination of character states (1) skin flap onheel (2) vocal repertoire composed of more than onecall type (3) multinote call and (4) regular internoteintervals (Figs 2 3 amp 4)
Comparisons with congeners of the Amazonian cladeBoana steinbachi can be distinguished from the membersof the B calcarata clade (B almendarizae B fasciata Bcalcarata and B maculateralis) by the absence of a cal-car and by the vocal repertoire composed of more thanone type of call Among members of the B steinbachiclade it can be distinguished from B dentei B alfaroiand B tetete by the presence of a skin flap on heel (tuber-cle in B alfaroi and B tetete tubercle on one side orcompletely absent in B dentei) by its multinote call (one-note call in B dentei and B tetete) by having regularinternote intervals between call notes (notes with irregularintervals sometimes partly fused one with the next in Balfaroi) and by the vocal repertoire composed of two dis-tinct types of calls (one call type in B alfaroi) (Camineramp Ron 2014 Marinho et al 2020)
Variation Body size varies between 304 and 374mmin males and between 425 and 488mm in females(Appendix 7) Individual AAG-UFU 5918 lacks the heelskin flap on the left side topotypes SMF 88394ndash95 andSMF 88397 lack the skin flap on both sides In life(Fig 3a b) dorsal colouration is beige light brownbright yellow or orange brown in calling males withfaint brown transversal bands and dark purplish-brownfaint brown or grey longitudinal lines extending from
the snout to vent a second line extending from behindthe eye to pelvic region and a third line as a canthalstripe extending from the posterior corner of the nostrilto anterior corner of the eye and from the posterior cor-ner of the eye to midbody length on flank (the topotypesSMF 88394ndash97 lack transversal bands on dorsum indi-viduals SMF 88394 and CORBIDI 13414 lack longitu-dinal lines on dorsum and the middorsal line isextending from snout to interocular region AAG-UFU5921 lacks longitudinal lines on dorsum) Iris cream orgrey sometimes with yellow pigmentation on the upperpart of the iris Throat varying from white to yellowchest and anterior belly varying from white to creamVentral surface of hand and hind limb beige to bright
755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808
809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862
Fig 2 Adult male of Boana steinbachi (AAG-UFU 5921)from the Assis Brasil population (Acre Brazil) (A) dorsal and(B) ventral views of the body (SVL frac14 337mm) (C) dorsaland (D) lateral views of the head (HL frac14123mm HW frac14102mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 106mm) and (G) sole of thefoot (FL frac14 139mm)
8 A Fouquet et al
antoinefouquet
Texte surligneacute
we may wanna ask for full page here no
PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
647648649650651652653654655656657658659660661662663664665666667668669670671672673674675676677678679680681682683684685686687688689690691692693694695696697698699700
701702703704705706707708709710711712713714715716717718719720721722723724725726727728729730731732733734735736737738739740741742743744745746747748749750751752753754
Fig 1 Subtrees of the three main clades from the chronogram obtained from the analysis of 16S sequences using BEAST2Terminals are collapsed according to the OTU recovered from the species delineation analysis The distribution of each of these OTUis depicted on the maps
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 7
PROOF ONLY
also assigned to this species in this study Specimensand calls from Acre (ldquoB aff steinbachi 1rdquo) and callsfrom the lower Madre de Dios River (ldquoB aff steinbachi2rdquo) were included in the variation of B steinbachi
Boana steinbachi (Boulenger 1905)Hyla steinbachi mdash Boulenger 1905
Hyla fasciata De la Riva 1990Hypsiboas fasciatus Jansen et al 2011
Hypsiboas sp (Clade G) Funk et al 2012Hypsiboas steinbachi Caminer amp Ron 2014
Boana steinbachi Dubois 2017Boana sp (Clade J) Meza-Joya et al 2019
Boana fasciata Vacher et al 2020
Syntypes BMNH 194721361ndash63 two adults ofunknown sex and one juvenile respectively from Saraprovince Department of Santa Cruz de La SierraBolivia Collected by Hf J Steinbach
Diagnosis Boana steinbachi is characterized by the fol-lowing combination of character states (1) skin flap onheel (2) vocal repertoire composed of more than onecall type (3) multinote call and (4) regular internoteintervals (Figs 2 3 amp 4)
Comparisons with congeners of the Amazonian cladeBoana steinbachi can be distinguished from the membersof the B calcarata clade (B almendarizae B fasciata Bcalcarata and B maculateralis) by the absence of a cal-car and by the vocal repertoire composed of more thanone type of call Among members of the B steinbachiclade it can be distinguished from B dentei B alfaroiand B tetete by the presence of a skin flap on heel (tuber-cle in B alfaroi and B tetete tubercle on one side orcompletely absent in B dentei) by its multinote call (one-note call in B dentei and B tetete) by having regularinternote intervals between call notes (notes with irregularintervals sometimes partly fused one with the next in Balfaroi) and by the vocal repertoire composed of two dis-tinct types of calls (one call type in B alfaroi) (Camineramp Ron 2014 Marinho et al 2020)
Variation Body size varies between 304 and 374mmin males and between 425 and 488mm in females(Appendix 7) Individual AAG-UFU 5918 lacks the heelskin flap on the left side topotypes SMF 88394ndash95 andSMF 88397 lack the skin flap on both sides In life(Fig 3a b) dorsal colouration is beige light brownbright yellow or orange brown in calling males withfaint brown transversal bands and dark purplish-brownfaint brown or grey longitudinal lines extending from
the snout to vent a second line extending from behindthe eye to pelvic region and a third line as a canthalstripe extending from the posterior corner of the nostrilto anterior corner of the eye and from the posterior cor-ner of the eye to midbody length on flank (the topotypesSMF 88394ndash97 lack transversal bands on dorsum indi-viduals SMF 88394 and CORBIDI 13414 lack longitu-dinal lines on dorsum and the middorsal line isextending from snout to interocular region AAG-UFU5921 lacks longitudinal lines on dorsum) Iris cream orgrey sometimes with yellow pigmentation on the upperpart of the iris Throat varying from white to yellowchest and anterior belly varying from white to creamVentral surface of hand and hind limb beige to bright
755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808
809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862
Fig 2 Adult male of Boana steinbachi (AAG-UFU 5921)from the Assis Brasil population (Acre Brazil) (A) dorsal and(B) ventral views of the body (SVL frac14 337mm) (C) dorsaland (D) lateral views of the head (HL frac14123mm HW frac14102mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 106mm) and (G) sole of thefoot (FL frac14 139mm)
8 A Fouquet et al
antoinefouquet
Texte surligneacute
we may wanna ask for full page here no
PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
also assigned to this species in this study Specimensand calls from Acre (ldquoB aff steinbachi 1rdquo) and callsfrom the lower Madre de Dios River (ldquoB aff steinbachi2rdquo) were included in the variation of B steinbachi
Boana steinbachi (Boulenger 1905)Hyla steinbachi mdash Boulenger 1905
Hyla fasciata De la Riva 1990Hypsiboas fasciatus Jansen et al 2011
Hypsiboas sp (Clade G) Funk et al 2012Hypsiboas steinbachi Caminer amp Ron 2014
Boana steinbachi Dubois 2017Boana sp (Clade J) Meza-Joya et al 2019
Boana fasciata Vacher et al 2020
Syntypes BMNH 194721361ndash63 two adults ofunknown sex and one juvenile respectively from Saraprovince Department of Santa Cruz de La SierraBolivia Collected by Hf J Steinbach
Diagnosis Boana steinbachi is characterized by the fol-lowing combination of character states (1) skin flap onheel (2) vocal repertoire composed of more than onecall type (3) multinote call and (4) regular internoteintervals (Figs 2 3 amp 4)
Comparisons with congeners of the Amazonian cladeBoana steinbachi can be distinguished from the membersof the B calcarata clade (B almendarizae B fasciata Bcalcarata and B maculateralis) by the absence of a cal-car and by the vocal repertoire composed of more thanone type of call Among members of the B steinbachiclade it can be distinguished from B dentei B alfaroiand B tetete by the presence of a skin flap on heel (tuber-cle in B alfaroi and B tetete tubercle on one side orcompletely absent in B dentei) by its multinote call (one-note call in B dentei and B tetete) by having regularinternote intervals between call notes (notes with irregularintervals sometimes partly fused one with the next in Balfaroi) and by the vocal repertoire composed of two dis-tinct types of calls (one call type in B alfaroi) (Camineramp Ron 2014 Marinho et al 2020)
Variation Body size varies between 304 and 374mmin males and between 425 and 488mm in females(Appendix 7) Individual AAG-UFU 5918 lacks the heelskin flap on the left side topotypes SMF 88394ndash95 andSMF 88397 lack the skin flap on both sides In life(Fig 3a b) dorsal colouration is beige light brownbright yellow or orange brown in calling males withfaint brown transversal bands and dark purplish-brownfaint brown or grey longitudinal lines extending from
the snout to vent a second line extending from behindthe eye to pelvic region and a third line as a canthalstripe extending from the posterior corner of the nostrilto anterior corner of the eye and from the posterior cor-ner of the eye to midbody length on flank (the topotypesSMF 88394ndash97 lack transversal bands on dorsum indi-viduals SMF 88394 and CORBIDI 13414 lack longitu-dinal lines on dorsum and the middorsal line isextending from snout to interocular region AAG-UFU5921 lacks longitudinal lines on dorsum) Iris cream orgrey sometimes with yellow pigmentation on the upperpart of the iris Throat varying from white to yellowchest and anterior belly varying from white to creamVentral surface of hand and hind limb beige to bright
755756757758759760761762763764765766767768769770771772773774775776777778779780781782783784785786787788789790791792793794795796797798799800801802803804805806807808
809810811812813814815816817818819820821822823824825826827828829830831832833834835836837838839840841842843844845846847848849850851852853854855856857858859860861862
Fig 2 Adult male of Boana steinbachi (AAG-UFU 5921)from the Assis Brasil population (Acre Brazil) (A) dorsal and(B) ventral views of the body (SVL frac14 337mm) (C) dorsaland (D) lateral views of the head (HL frac14123mm HW frac14102mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 106mm) and (G) sole of thefoot (FL frac14 139mm)
8 A Fouquet et al
antoinefouquet
Texte surligneacute
we may wanna ask for full page here no
PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
yellow Posterior surface of thigh groin and posteriorhalf of flank spotted in black with a bright yellow orwhite background colour Brown dots scattered on dor-sum dorsal surface of limbs bordering the lower lipmental region and chest In preservative the coloursfade the bright yellow colour sometimes present on dor-sum ventral surfaces of limbs groin and flank arebeige or pale cream Iris grey Flank and groin macula-tion are denser and more extensive in females
Vocal repertoire (Fig 4) We analysed calls of 13males (see Appendix 4 for information about soundrecordings and Appendix 6 for the complete descriptivestatistics) The vocal repertoire of Boana steinbachi iscomposed of two distinct types of calls (type 1 nfrac14 149calls of 13 males type 2 nfrac14 43 calls of five males)
that are emitted sporadically at irregular intervals Thetype 1 call of B steinbachi lasts 130ndash430ms and iscomposed of 3ndash8 nonpulsed notes lasting 4ndash52ms sepa-rated by intervals of 1ndash67ms The rise time is at 2ndash98of call duration The minimum frequency ranges from1335ndash1981Hz the maximum frequency from3336ndash4479Hz and the dominant frequency from1688ndash3402Hz The type 2 call is composed of one notewith poorly defined pulses Notes last 47ndash87ms Therise time is at 18ndash60 of note duration The minimumfrequency ranges from 1453ndash2015Hz the maximum fre-quency from 2713ndash4522Hz and the dominant frequencyfrom 1688ndash2813Hz The limited sample size for eachpopulation with recorded calls prevented us from evalu-ating the geographic variation of acoustic traits acrosspopulations of B steinbachi
863864865866867868869870871872873874875876877878879880881882883884885886887888889890891892893894895896897898899900901902903904905906907908909910911912913914915916
917918919920921922923924925926927928929930931932933934935936937938939940941942943944945946947948949950951952953954955956957958959960961962963964965966967968969970
Fig 3 Life colours in dorsolateral view and detail of the patterns on flank and groin (A B) Boana steinbachi (topotype SMF88394and non-topotype AAG-UFU 5918 respectively) (C D) B eucharis sp nov (AAG-UFU 6503 holotype) and (E F) B courtoisaesp nov (holotype MNHN-RA-20200001 and an unvouchered specimen respectively)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 9
antoinefouquet
Texte surligneacute
pedro thiago these ranges are a bit strange 4-52 or 40-52 1-67 really 2-98
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-
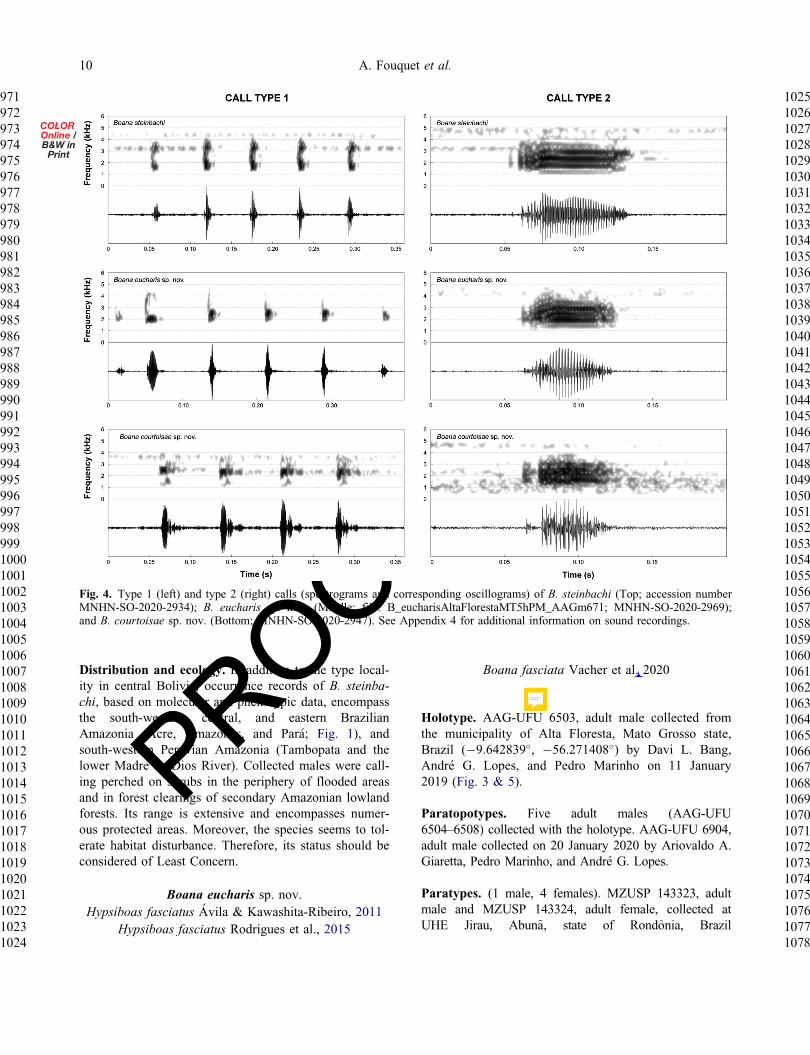
PROOF ONLY
Distribution and ecology In addition to the type local-ity in central Bolivia occurrence records of B steinba-chi based on molecular and phenotypic data encompassthe south-western central and eastern BrazilianAmazonia (Acre Amazonas and Para Fig 1) andsouth-western Peruvian Amazonia (Tambopata and thelower Madre de Dios River) Collected males were call-ing perched on shrubs in the periphery of flooded areasand in forest clearings of secondary Amazonian lowlandforests Its range is extensive and encompasses numer-ous protected areas Moreover the species seems to tol-erate habitat disturbance Therefore its status should beconsidered of Least Concern
Boana eucharis sp novHypsiboas fasciatus Avila amp Kawashita-Ribeiro 2011
Hypsiboas fasciatus Rodrigues et al 2015
Boana fasciata Vacher et al 2020
Holotype AAG-UFU 6503 adult male collected fromthe municipality of Alta Floresta Mato Grosso stateBrazil (9642839 56271408) by Davi L BangAndre G Lopes and Pedro Marinho on 11 January2019 (Fig 3 amp 5)
Paratopotypes Five adult males (AAG-UFU6504ndash6508) collected with the holotype AAG-UFU 6904adult male collected on 20 January 2020 by Ariovaldo AGiaretta Pedro Marinho and Andre G Lopes
Paratypes (1 male 4 females) MZUSP 143323 adultmale and MZUSP 143324 adult female collected atUHE Jirau Abun~a state of Rondonia Brazil
9719729739749759769779789799809819829839849859869879889899909919929939949959969979989991000100110021003100410051006100710081009101010111012101310141015101610171018101910201021102210231024
102510261027102810291030103110321033103410351036103710381039104010411042104310441045104610471048104910501051105210531054105510561057105810591060106110621063106410651066106710681069107010711072107310741075107610771078
Fig 4 Type 1 (left) and type 2 (right) calls (spectrograms and corresponding oscillograms) of B steinbachi (Top accession numberMNHN-SO-2020-2934) B eucharis sp nov (Middle file B_eucharisAltaFlorestaMT5hPM_AAGm671 MNHN-SO-2020-2969)and B courtoisae sp nov (Bottom MNHN-SO-2020-2947) See Appendix 4 for additional information on sound recordings
10 A Fouquet et al
antoinefouquet
Note
zoobankorgact51CC7B40-2D6B-4A9E-AF50-AB34D4CE1042
antoinefouquet
Texte inseacutereacute
PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
(9699410 65358875) MZUSP 143238 adultfemale collected at UHE Jirau Porto Velho state ofRondonia Brazil (8748483 63903465) MZUSP159228 (field No MTR25896) adult female collected atPARNA Pacaas Novos state of Rondonia Brazil(10786979 63627305) MZUSP 159227 (field NoMTR25798) adult female collected at Pacaas Novos stateof Rondonia Brazil (10786979 63627305)
Diagnosis Boana eucharis sp nov is characterized bythe following combination of character states (1) tuber-cle on heel (2) vocal repertoire composed of more than
one call type (3) multinote call and (4) regular inter-note intervals (Figs 3 4 amp 5)
Comparisons with congeners of the Amazonian cladeBoana eucharis sp nov is distinguished from the mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B eucharis sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroiby having regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type Caminer amp Ron 2014 Marinho et al 2020)Boana eucharis sp nov can be distinguished in almostall cases from B steinbachi by the presence of tubercleon heel (skin flap in B steinbachi but see Variation)In addition B eucharis sp nov is sister of all remain-ing species of the B steinbachi clade (except the allo-patric B dentei) The phylogenetic relationships withinthis clade and spatial distribution strongly support thedistinct specific status of B eucharis sp nov relative toits closest relatives (Fig 1)
Description of holotype (Figs 3 amp 5) Adult male SVL308mm FL 120mm ED 38mm TD 20mm TL178mm THL 158mm CL 02mm HAL 104mmFLL 59mm EN 33mm head slightly longer (HL119mm) than wide (HW 103mm) and wider thanbody snout rounded in lateral view truncate in dorsalview EN shorter than ED canthus rostralis indistinctrounded loreal region concave internarial area convexnostril slightly protuberant directed laterally interorbitalarea slightly convex eye large strongly protuberant ED19 times TD tympanic membrane undifferentiatedtympanic annulus evident rounded concealed poster-iorly by the supratympanic fold running from the pos-terior corner of the eye to arm insertion Tongue ovoidwidely attached to mouth floor six vomerine teeth oneach vomer vomers barely separated posteromedial tochoanae choanae ovoid Arm slender axillary mem-brane absent ill-defined low tubercles present alongventrolateral edge of forearm relative length of fingersIlt IIlt IVlt III fingers bearing large oval discs subar-ticular tubercles prominent ovoid to conical singlesupernumerary tubercles present palmar tubercle smallelongated prepollical tubercle large flat elliptical pre-pollex enlarged covered by skin nuptial excrescencesabsent webbing absent between fingers Tubercle ontibiotarsal articulation scattered tubercles along the
107910801081108210831084108510861087108810891090109110921093109410951096109710981099110011011102110311041105110611071108110911101111111211131114111511161117111811191120112111221123112411251126112711281129113011311132
113311341135113611371138113911401141114211431144114511461147114811491150115111521153115411551156115711581159116011611162116311641165116611671168116911701171117211731174117511761177117811791180118111821183118411851186
Fig 5 Holotype of Boana eucharis sp nov (AAG-UFU6503) from Alta Floresta (Mato Grosso Brazil) (A) dorsal and(B) views of the body (SVL frac14 308mm) (C) dorsal and (D)lateral views of the head (HL frac14 119mm HW frac14 103mm)(E) detail in dorsal view of the tubercle on heels (F) palm ofthe hand (HAL frac14 87mm) and (G) and sole of the foot (FLfrac14 131mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 11
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
t
antoinefouquet
Texte surligneacute
same here it looks too small
PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
external edge of tarsus and foot toes bearing discs slightlywider than long smaller than those of fingers relativelength of toes Ilt IIltVltIIIlt IV outer metatarsal tuber-cle ill-defined small rounded inner metatarsal tuberclelarge ovoid subarticular tubercles single low roundedsupernumerary tubercles restricted to the sole of foot web-bing formula of toes I 2ndash212 II 1thornndash212 III 112ndash212 IV3ndash112 V Skin on dorsum head and dorsal surfaces oflimbs and flank mostly smooth skin on belly and thighcoarsely granular skin on throat and chest finely granulararm forearm and shank smooth Cloacal opening directedposteriorly at upper level of thigh short simple cloacalsheath covering cloacal opening round tubercles belowand on the sides of the opening
Colours of holotype In preservative dorsum greyishbrown with scattered minute black dots faint brownmiddorsal line extending from the tip of the snout topelvic region fragmented in interorbital region a secondline extending from behind the eye to pelvic regionfragmented and faint at midbody length and a third lineas a canthal stripe extending from the posterior cornerof the nostril to anterior corner of the eye and from theposterior corner of the eye to midbody length on flankdorsal surface of limbs greyish brown with transversalfaint brown bars flank beige with dark irregular spotsposterior surface of thigh beige with dark irregularspots venter cream white with brown spots on the men-tal region and chest ventral surface of limbs cream witha narrow brown stripe on the outer edge of the handforearm thigh tarsal fold and foot limb bones partiallyvisible through skin white In life (Fig 3c d) dorsumbeige with a purplish brown middorsal line extendingfrom the tip of snout to pelvic region a second lineextending from behind the eye to pelvic region frag-mented and faint at midbody length and a third as can-thal stripe from the posterior border of nostril toanterior corner of the eye and from the posterior cornerof the eye to midbody length on flank dorsal surface ofhindlimbs with faint brown transversal bands minutedark brown dots scattered on the dorsal surface of limbsand dorsum flank bright yellow with dark brown irregu-lar blotches on groin posterior surface of thigh paleyellowish with dark brown blotches
Variation Body size varies between 308 and 347mmin males (Appendix 7) The paratopotype AAG-UFU6504 lacks the heel tubercle on the right side andMZUSP 80790 (from Rondonia) does not possess thetubercle on either side In life (Fig 3c d) dorsal colour-ation varies from beige to brown orange brown in call-ing males with dark purplish-brown longitudinal linesand faint brown transversal bands (individuals MZUSP
159227 and MZUSP 159228 do not have longitudinallines extending from snout to pelvic region or a canthalstripe) Throat varies from white to yellow chest andanterior belly varying from white to cream Iris creamor grey sometimes with yellow pigmentation on theupper iris Ventral surface of hand and hind limb mostlybright yellow Posterior surface of thigh groin and pos-terior half of flank spotted in black with a bright yellowor white background colour Brown dots scattered ondorsum dorsal surface of limbs bordering the lower lipmental region and chest We did not observe dichro-matic patterns between male and female specimens Inpreservative the colours become paler the bright yellowtone occasionally on ventral surfaces of limbs groin andflank is beige or pale cream Iris grey
Vocal repertoire Calls of seven males were recorded atthe type locality in southern Brazilian Amazonia(Appendix 6) The vocal repertoire of Boana eucharis iscomposed of two distinct calls (type 1 nfrac14 133 calls fromseven males type 2 nfrac14 62 calls from seven males) thatare emitted sporadically at irregular intervals The type 1call (Fig 4) lasts 290ndash420ms and consists of 3ndash7 non-pulsed notes that last 3ndash60ms separated by intervals of27ndash82ms The rise time is at 2ndash97 of call duration Theminimum frequency ranges from 1864ndash2250Hz the max-imum frequency from 2196ndash4220Hz and the dominantfrequency from 2147ndash2462Hz The type 2 call consists ofone note with poorly defined pulses Notes last 29ndash64msThe rise time is at 24ndash78 of note duration The min-imum frequency ranges from 1593ndash2067Hz the maximumfrequency from 2712ndash3491Hz and the dominant fre-quency from 1938ndash3143Hz
Distribution and ecology Boana eucharis sp nov isknown from southern Amazonia in the Brazilian statesof Mato Grosso and Rondonia (Fig 1) Males callperched on shrubs in flooded areas associated with theborder of secondary-growth or disturbed forestsSympatric anuran species at the type locality of Beucharis sp nov are Boana leuchocheila B albopunc-tata Dendropsophus cruzi Engystomops freibergiLeptodactylus vastus L petersii Pithecopus hypochon-drialis Scinax garbei and S nebulosus The species isnot abundant It is possible that the species range ismore extensive than the five populations reported in thisstudy which suggests a Data Deficient conservation cat-egory for B eucharis sp nov However it is importantto highlight that the southern limits of Amazonia areoverall highly impacted by habitat conversion and thisspecies could be classified at least as VulnerableNevertheless the known occurrence recordings compriseat least two protected conservation units Pacaas Novos
118711881189119011911192119311941195119611971198119912001201120212031204120512061207120812091210121112121213121412151216121712181219122012211222122312241225122612271228122912301231123212331234123512361237123812391240
124112421243124412451246124712481249125012511252125312541255125612571258125912601261126212631264126512661267126812691270127112721273127412751276127712781279128012811282128312841285128612871288128912901291129212931294
12 A Fouquet et al
PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
National Park and possibly the Cristalino State Parkand the species seems to tolerate a certain extent ofhuman disturbance to forest habitats since calling maleswere sampled at forest borders and clearings
Etymology The specific epithet is derived from theGreek word eukharis which means gracious or charis-matic as a reference to the delicate and gracious aspectof the species
Boana courtoisae sp novHyla fasciata Lescure amp Marty 2000Hyla fasciata Faivovich et al 2005
Hypsiboas fasciatus Fouquet et al 2007Hyla fasciata Avila-Pires et al 2010
Hypsiboas sp (Clade H) Funk et al 2012Hypsiboas fasciatus Ouboter amp Jairam 2012
Hypsiboas fasciatus Cole et al 2013Hypsiboas sp (Clade H) Caminer amp Ron 2014
Boana cf fasciata Fouquet et al 2019Boana fasciata Vacher et al 2020
Holotype MNHN-RA-20200001 adult male collected atAlikene French Guiana (320906 52402000) by JPVacher and S Cally on 12 February 2015 (Figs 3 amp 6)
Paratopotypes An adult female (MNHN-RA-20200002) collected with the holotype
Paratypes (12 males 3 females) MNHN-RA-20200004 adult male collected at Saut Taconet FrenchGuiana (403249 52526188) MNHN-RA-20200005 adult female collected at Saut GrandMachicou French Guiana (3897416 52583565)MNHN-RA-20200006ndash7 two adult males collected atSaul French Guiana (3615576 53227093)MNHN-RA-20200008 adult male collected at Flat dela Waki French Guiana (3089500 0398460)MNHN-RA-20200009ndash12 four adult males collected atSipaliwini Suriname (2097530 56147200)MNHN-RA-20200013 an adult male collected at EkiniFrench Guiana (4050000 52466700) MNHN-RA-20200014ndash16 three adult males collected at MitarakaFrench Guiana (2235770 54449280) MNHN-RA-20200003 an adult female collected at Inini TolengaFrench Guiana (3663159 53928308)
Other material Twelve additional males and threefemales (Appendix 8) from French Guiana and Surinamewere also assigned to B courtoisae They were examinedand included in the analysis of the variation but were notdeposited and not included in the type series
Diagnosis Boana courtoisae sp nov is characterizedby the following combination of character states (1)skin flap on heel (2) vocal repertoire composed ofmore than one call type (3) multinote call and (4)regular internote intervals (5) multi-blotched pattern ongroin and flank of males (Figs 3 4 amp 6)
Comparisons with congeners of the Amazonian cladeBoana courtoisae sp nov is distinguished from mem-bers of the B calcarata clade (B almendarizae B fas-ciata B calcarata and B maculateralis) by theabsence of a calcar and by the vocal repertoire com-posed of more than one type of call Within the B stein-bachi clade B courtoisae sp nov can be distinguishedfrom B dentei and B tetete by its multinote call (one-note calls in B dentei and B tetete) and from B alfaroi
129512961297129812991300130113021303130413051306130713081309131013111312131313141315131613171318131913201321132213231324132513261327132813291330133113321333133413351336133713381339134013411342134313441345134613471348
134913501351135213531354135513561357135813591360136113621363136413651366136713681369137013711372137313741375137613771378137913801381138213831384138513861387138813891390139113921393139413951396139713981399140014011402
Fig 6 Holotype of Boana courtoisae sp nov (MNHN-RA-20200001) from Alikene (French Guiana) (A) dorsal and (B)ventral views of the body (SVL frac14 310mm adult male) (C)dorsal and (D) lateral views of the head (HL frac14 115mm HWfrac14 108mm) (E) detail in dorsal view of the skin flap on heels(F) palm of the hand (HAL frac14 93mm) and (G) sole of thefoot (FL frac14 120mm)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 13
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Texte inseacutereacute
antoinefouquet
Note
zoobankorgact4F8ACA9F-F6F1-4605-BD6C-6D4650AACCBE13
PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
by its regular internote intervals and vocal repertoirecomposed of two distinctive types of calls (in B alfaroicall notes having irregular intervals sometimes partlyfused one with the next and repertoire formed by onecall type) (Caminer amp Ron 2014 Marinho et al 2020)Boana courtoisae sp nov can be distinguished inalmost all cases from B eucharis sp nov by the pres-ence of skin flap on heel (tubercle in B eucharis spnov but see Variation) In addition B courtoisae spnov can be distinguished from B eucharis sp nov andB steinbachi by a multi-blotched pattern on flank andgroin of males (fewer blotches or transversal bands andspots in males of the other two species Fig 3)Although B courtoisae sp nov and B eucharis spnov are distinguished by subtle differences in morph-ology and colouration the two species are not directlyphylogenetically related to each other since B courtoi-sae sp nov is sister of the remaining species that formthe B steinbachi clade (B steinbachi B alfaroi Btetete and B eucharis sp nov) which strongly sup-ports the distinct specific status relative to its closest rel-atives (Fig 6)
Description of holotype Adult male SVL 310mm FL120mm ED 44mm TD 18mm TL 189mm THL158mm CL 04mm HAL frac14 93mm FLL 62mm EN42mm head slightly longer (HL 115mm) than wide(HW 109mm) and wider than body snout rounded inlateral view truncate in dorsal view (Fig 5) EN shorterthan ED canthus rostralis indistinct rounded lorealregion concave internarial area convex nostrils slightlyprotuberant directed laterally interorbital area slightlyconvex eye large strongly protuberant ED 25 timesTD tympanum membrane undifferentiated tympanicannulus evident rounded concealed posteriorly bysupratympanic fold running from the posterior corner ofthe eye to arm insertion Tongue ovoid widely attachedto mouth floor vomerine odontophores triangular witharched base barely separated posteromedial to choanaebearing eight vomerine teeth on each side choanaeovoid Arm slender axillary membrane absent indistinctlow tubercles present along ventrolateral edge of fore-arm relative length of fingers Ilt IIlt IVlt III fingersbearing large oval discs subarticular tubercles promin-ent ovoid to conical single supernumerary tuberclespresent palmar tubercle small elongated prepollicaltubercle large flat elliptical prepollex enlarged clawshaped nuptial excrescences absent webbing absentbetween fingers Skin flap on tibiotarsal articulationscattered tubercles on tarsus and along ventrolateraledge of foot toes bearing discs slightly wider than longsmaller than those of fingers relative length of toesIlt IIltVltIIIlt IV outer metatarsal tubercle ill defined
small round inner metatarsal tubercle large elongatedand elliptical subarticular tubercles single lowrounded supernumerary tubercles restricted to the soleof foot webbing formula of toes I2mdash212II1thorn-212III112-212IV3-112V Skin on dorsum head and dorsal surfa-ces of limbs smooth skin on flanks smooth with weaklongitudinal wrinkles posterior to the arm skin on ven-ter coarsely granular skin on ventral surfaces of headand thighs granular those of shanks smooth Cloacalopening directed posteriorly at upper level of thighsshort simple cloacal sheath covering cloacal openinground tubercles below and on the sides of the opening
Colour of holotype In preservative dorsum beige withscattered minute black dots and spots (Fig 6) faintbrown narrow middorsal line extends from the tip of thesnout to the vent faint brown transversal bands on dor-sum dorsal surface of limbs beige with transversal faintbrown bars flank white with dark irregular spots pos-terior surface of thigh white with dark irregularblotches venter creamy white with brown spots on thethroat and chest ventral surface of limbs whitish creamwith scattered dots on the forearm ventral surface ofshank cream a discontinuous brown stripe varying inwidth along its length on the outer edge of the handand forearm limb bones (visible through skin) white Inlife dorsum beige with a faint brown narrow longitu-dinal line from snout to vent dorsal surface of limbsbeige with faint brown transversal bands scatteredminute black dots on the dorsal surfaces of limbs anddorsum flank white to light yellow with dark irregularblotches venter white to cream scattered brown fleckson the throat and chest and bordering the lower lipventral surface of limbs pale yellow with bright yellowgranules discs and webbing yellow iris cream with anundefined upper yellow band limb bones (visiblethrough skin) white
Variation Body size varies between 308 and 359mmin males and between 430 and 459mm in females(Appendix 7) In life (Fig 4endashf) dorsal colourationvaries from beige to bright yellow or orange brownwith faint brown transversal bands markings purplish-brown to dark brown longitudinal lines and many scat-tered brown spots covering all over the dorsum Throatchest and anterior belly varying from white to creamwith scattered spots covering the entire ventral surfacein both sexes Flank and groin with dark brown to blackblotches on a bright yellow bluish or white backgroundVentral surface of hand foot and legs varying frombright yellow to dark grey covered with black spotsPosterior surface of thigh striped or spotted in blackwith a bright yellow or white background colour
140314041405140614071408140914101411141214131414141514161417141814191420142114221423142414251426142714281429143014311432143314341435143614371438143914401441144214431444144514461447144814491450145114521453145414551456
145714581459146014611462146314641465146614671468146914701471147214731474147514761477147814791480148114821483148414851486148714881489149014911492149314941495149614971498149915001501150215031504150515061507150815091510
14 A Fouquet et al
PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
sometimes with tints of blue particularly pronounced infemales (individual MNHN-RA-20200006 has noblotches or dots on the posterior surface of thigh orspots on belly) Iris grey or cream sometimes with ayellow upper band (the individual MNHN-RA-20200015 has also black markings surrounding theiris) Our collected individuals exhibit a dimorphic col-ouration between males and females The ventral surfaceof thigh in males varies from bright yellow to greenishyellow In females the colouration of the ventral surfaceof thigh is blue bluish grey or dark grey The groinregion flank and posterior surface of thigh have moretints of blue in females In addition flank and groinmaculation is denser and more extensive in females Inpreservative the colours fade becoming pale the brightyellow tone sometimes present on dorsum is beige orbrown with brown transversal bands and lines Ventralsurfaces of body limbs groin and flank are beige orpale cream The blue tints on groin flank and posteriorsurface of thigh completely vanish as well as the darkblue or grey colours on ventral surfaces of femalersquosthigh Iris grey
Vocal repertoire Calls of five males were recordedfrom French Guiana and Suriname The vocal repertoireof B courtoisae is composed of two types of call (type1 nfrac14 13 calls from five males type 2 nfrac14 5 calls fromthree males) that are emitted sporadically at irregularintervals The type 1 call (Fig 4) lasts 140ndash240ms con-sisting of 3ndash4 nonpulsed notes that last 5ndash36ms sepa-rated by intervals of 27ndash82ms The rise time is at2ndash97 of call duration The minimum frequency rangesfrom 1335ndash1875 the maximum frequency from2627ndash3101Hz and the dominant frequency from1981ndash2972Hz The type 2 call consists of one note withpoorly defined pulses Notes last 39ndash79ms The risetime is at 39ndash82 of note duration The minimum fre-quency ranges from 1464ndash1723Hz the maximum fre-quency from 2713ndash3144Hz and the dominantfrequency from 1680ndash2412Hz
Distribution and ecology Boana courtoisae sp nov isdistributed throughout the eastern Guiana Shield inFrench Guiana Suriname Guyana and adjacentBrazilian Amazonia in the states of Amapa Para (Avila-Pires et al 2010) and possibly Roraima (pending con-firmation) The species could possibly occur in the stateof Amazonas as well The species is not abundant andfound in scattered populations Nevertheless its range isextensive and encompasses numerous protected areasMoreover the species seems to tolerate habitat disturb-ance indicated by the use of forest borders and clear-ings Therefore the conservation status of B courtoisae
sp nov might be classified as Least Concern (pendinga formal evaluation by IUCN team) It is a nocturnalspecies found in primary and secondary forest associatedwith the flooded zones of slow streams and medium-sized rivers The males call perched at low height onthe adjacent vegetation or even overlooking the waterforming small groups of 2ndash10 individuals separated by afew metres from each other A single clutch wasobserved at Mitaraka French Guiana and containedapproximately 1100 beige eggs deposited directly in thewater and forming a film on the surface
Etymology This species is dedicated to our friendElodie Courtois in honour of her invaluable contribu-tion to field herpetology in French Guiana notably themonitoring of populations of threatened species and dis-covery of previously undocumented species and manynatural history observations
Mitogenomic phylogenyThe mitogenomic phylogeny is well resolved with theexception of the position of the clade formed by B cine-rascens thorn B punctata and the position of B pellucenswithin the genus (Fig 7) Within the focal species grouponly the relationships among the different OTUs of theB albopunctata clade remain ambiguous Boana heil-prini is strongly supported as the sister species of allother members of the B albopunctata species groupwhose crown age is estimated to date back to 173Ma(144ndash202) Boana raniceps forms a clade with all theother species of the group whose crown age is estimatedto date back to 132Ma (111ndash155) The remainder ofspecies forms three main clades The B albopunctataclade diverges from the Amazonian species 109Ma(91ndash127) The two Amazonian clades diverged99Ma (83ndash116)
Biogeographic inferencesModel comparisons identified DECthorn J as the best-fitmodel (Appendix 9) According to both DEC andDECthorn J models the ancestral range of the B albopunc-tata species group remains largely ambiguous sinceeach of the major lineages is widely distributed inAmazonia and even further for the B albopunctataclade However the ancestral range of the B steinbachiclade is supported to be located in the Guiana Shield bythe DECthorn J model (Fig 8A) and while it remainsambiguous for the DEC model all states with high like-lihood encompass the Guiana Shield (Fig 8B) Thisgroup probably dispersed southward to the BrazilianShield 5Ma as suggested by the phylogenetic position
151115121513151415151516151715181519152015211522152315241525152615271528152915301531153215331534153515361537153815391540154115421543154415451546154715481549155015511552155315541555155615571558155915601561156215631564
156515661567156815691570157115721573157415751576157715781579158015811582158315841585158615871588158915901591159215931594159515961597159815991600160116021603160416051606160716081609161016111612161316141615161616171618
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 15
antoinefouquet
Texte inseacutereacute
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-
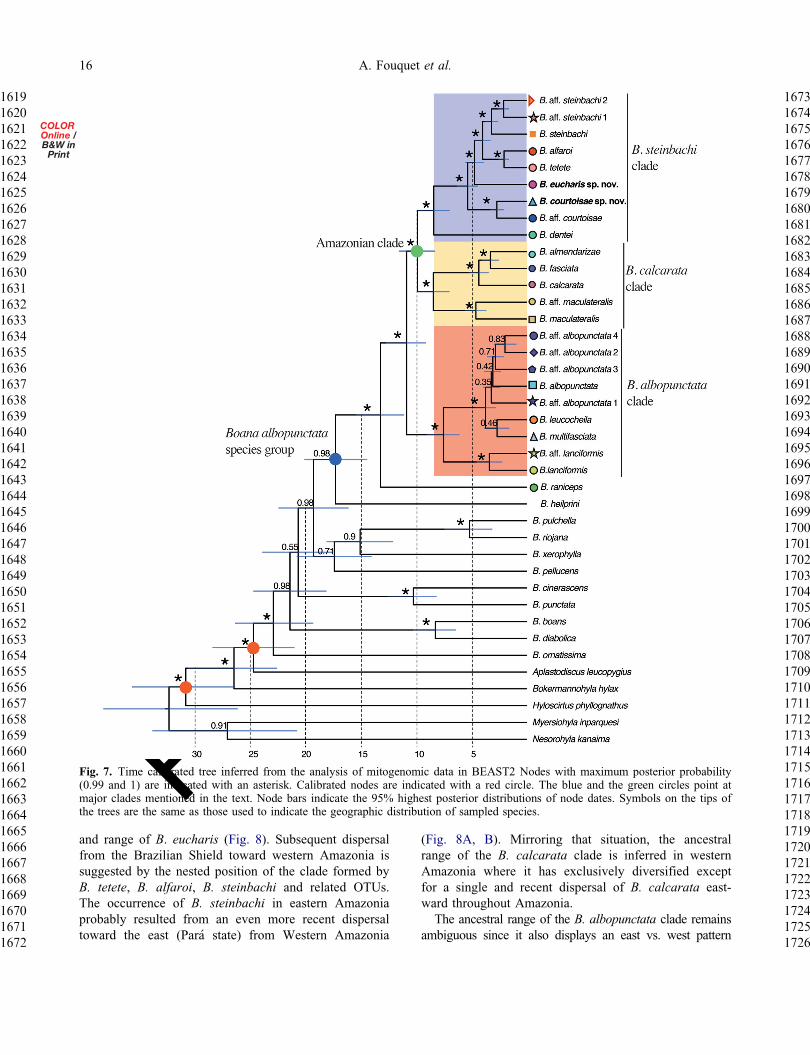
PROOF ONLY
and range of B eucharis (Fig 8) Subsequent dispersalfrom the Brazilian Shield toward western Amazonia issuggested by the nested position of the clade formed byB tetete B alfaroi B steinbachi and related OTUsThe occurrence of B steinbachi in eastern Amazoniaprobably resulted from an even more recent dispersaltoward the east (Para state) from Western Amazonia
(Fig 8A B) Mirroring that situation the ancestralrange of the B calcarata clade is inferred in westernAmazonia where it has exclusively diversified exceptfor a single and recent dispersal of B calcarata east-ward throughout AmazoniaThe ancestral range of the B albopunctata clade remains
ambiguous since it also displays an east vs west pattern
161916201621162216231624162516261627162816291630163116321633163416351636163716381639164016411642164316441645164616471648164916501651165216531654165516561657165816591660166116621663166416651666166716681669167016711672
167316741675167616771678167916801681168216831684168516861687168816891690169116921693169416951696169716981699170017011702170317041705170617071708170917101711171217131714171517161717171817191720172117221723172417251726
Fig 7 Time calibrated tree inferred from the analysis of mitogenomic data in BEAST2 Nodes with maximum posterior probability(099 and 1) are indicated with an asterisk Calibrated nodes are indicated with a red circle The blue and the green circles point atmajor clades mentioned in the text Node bars indicate the 95 highest posterior distributions of node dates Symbols on the tips ofthe trees are the same as those used to indicate the geographic distribution of sampled species
16 A Fouquet et al
PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
with B lanciformis occurring only in western Amazoniaand the rest of the species of the clade occurring in theCerrado and eastern Amazonia However this east-westdivergence seems more recent in that group (75Ma) thanthe B calcarata vs B steinbachi divergence (99Ma)
DiscussionSpecies richness and distributionWith 25 putative species and two taxa (B caiapo and Bparanaiba) that could not be included in this study the
172717281729173017311732173317341735173617371738173917401741174217431744174517461747174817491750175117521753175417551756175717581759176017611762176317641765176617671768176917701771177217731774177517761777177817791780
178117821783178417851786178717881789179017911792179317941795179617971798179918001801180218031804180518061807180818091810181118121813181418151816181718181819182018211822182318241825182618271828182918301831183218331834
Fig 8 Ancestral area reconstruction for the Boana albopunctata species group (B heilprini and B raniceps excluded) using BioGeoBearsassuming (A) a DECthorn J model and (B) a DEC model (Appendix 5) most likely biogeographic scenarios plotted on the chronogramobtained with BEAST 25 (Numbers on branches are posterior clade probabilities those 095 are indicated with an asterisk Node barsindicate the 95 highest posterior distributions of node dates Squares on the tips of the trees indicate the geographic distribution of extantspecies sampled in the phylogeny Pie charts on nodes show the most likely reconstructions of ancestral areas the size of each sliceproportional to the maximum likelihood Colours corresponding to the different geographic distributions are depicted on the left
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 17
antoinefouquet
Texte inseacutereacute
gt
PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
species richness of the B albopunctata group may actu-ally reach 27 species ie 70 higher than currentlyrecognized (16 valid species) Within the Amazonianclade the actual number of species may be 44 higherthan currently recognized (14 OTU for 10 describedvalid species including the two species describedherein) We could not gather sufficient phenotypic datafor the OTU identified as B aff maculateralis but weare confident that such data will contribute to its taxo-nomic resolution Moreover our genetic samplingremains limited specifically in Colombia Venezuelaand south-eastern Amazonia and more species in thegroup probably remain undocumentedThe discovery of yet undescribed species in
Amazonia is not surprising since new species descrip-tions of squamates and anurans keep accumulating at afast pace (eg Carvalho et al 2020 Kok et al 2018)In fact almost all systematic investigation of broadlydistributed groups in Amazonia led to the understandingthat they actually represent species complexes oftenhiding narrowly distributed and remotely diverging spe-cies within Amazonia (eg Fouquet et al 2014 Vacheret al 2020) This recurrent pattern is also illustratedherein in the B albopunctata species group notably byB fasciata a taxon that was used to design populationsthroughout Amazonia until the conclusions of Caminerand Ron (2014) The extent of the actual diversity ofanurans in Amazonia remains so speculative that itcould be three to four times higher than the current600 species occurring in that region according to theIUCN (Vacher et al 2020)Relationships and distribution of the species we found
within the Amazonian clade strikingly mirror thosefound in other groups of anurans notably B semilineatagroup (Caminer amp Ron 2020 Fouquet et al 2016Peloso et al 2018) Osteocephalus (Jungfer et al2013) Dendropsophus minutus (Gehara et al 2014)Allobates (Rejaud et al 2020) Amazophrynella (Rojaset al 2018) Adenomera (Fouquet et al 2014) andmore broadly matches a pattern of allopatry throughoutAmazonia in which communities are spatially structuredforming distinct bioregions (Vacher et al 2020)However the history of Pan-Amazonian diversificationis relatively recent in the case of the Amazonian cladeof Boana studied herein compared with other taxa suchas Allobates (Rejaud et al 2020) or Amazophrynella(Rojas et al 2018) A combination of historical andcontemporary climatic heterogeneity as well as species-specific dispersal ability and niche breadth (Sheu et al2020) is probably responsible for these common anddistinct spatio-temporal patterns across taxa Our time-scaled phylogeny and biogeographic analyses providesome insights into the historical processes responsible
for the diversification of the B albopunctata speciesgroup in Amazonia
BiogeographyWith nine OTUs the species richness in westernAmazonia is confirmed to be higher than in the GuianaShield (4 OTUs) and the Brazilian Shield (3 OTUs)suggesting that the climatic conditions and historicalgeomorphological dynamism in this region notablyhydrological changes may have played a major role inB gr albopunctata diversificationThe initial diversification within the Amazonian clade
between the B calcarata clade in the west and the Bsteinbachi in the east of Amazonia dates back to about10Ma This estimate is relatively younger than those ofFunk et al (2012) and Duellman et al (2016) partlybecause the divergences estimated by Feng et al (2017)which were used as calibrations herein are overall morerecent than the nodes of those previous studies that werebased on a lower amount of genomic data Other eastwest divergences putatively simultaneous to the onefound herein within the Amazonian clade of the B albo-punctata group are documented in the Adenomera heyericlade (Carvalho et al 2020 Fouquet et al 2014) in twoinstances within the Allobates trilineatus clade (Rejaudet al 2020) in Ameerega (Guillory et al 2020) andprobably in many other lineages for which sampling anddating are still missing This 10 Myr old node coincideswith the end of the Pebas system and the transition froma western watershed drained to the north to a Pan-Amazonian system drained to the east (Hoorn et al2017) Available data indicate that the rise of the VaupesArch around 10Ma completely separated the WesternAmazon and Llanos basins (Hoorn et al 2010 Jaramilloet al 2017) We assume that this new configuration mayhave permitted the dispersal between the Guiana Shieldand Western Amazonia and could be responsible for this10 Myr old divergence in the Boana of the Amazonianclade and the other mentioned groupsSubsequently both groups have apparently diversified
in situ ie within Western Amazonia and within theGuiana Shield until some 5Ma This date coincides withthe divergence between B courtoisae sp nov and theother species of the B steinbachi clade and suggests adispersal from the Guiana to the Brazilian Shield acrossthe transcontinental configuration of the Amazon Riverwhich contradicts our expectation that such divergencewould precede this configuration (9Ma Hoorn et al2017) Temporally concordant northsouth divergencesare documented within Allobates tapajos and betweenA bacurau and A sumtuosus (Rejaud et al 2020) inChiasmocleis (de Sa et al 2019) and most likely other
183518361837183818391840184118421843184418451846184718481849185018511852185318541855185618571858185918601861186218631864186518661867186818691870187118721873187418751876187718781879188018811882188318841885188618871888
188918901891189218931894189518961897189818991900190119021903190419051906190719081909191019111912191319141915191619171918191919201921192219231924192519261927192819291930193119321933193419351936193719381939194019411942
18 A Fouquet et al
PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
lineages of terrestrial vertebrates for which the historiesof diversification remain undocumented The processesthat may have fostered multiple trans-Amazon dispersalsaround 5Ma remain highly speculative Considerableuncertainty remains about the timing and amplitude ofhistorical topographic hydrological and vegetationalchanges in Amazonia (Albert et al 2018a BicudoSacek de Almeida Bates amp Ribas 2019 CampbellFrailey amp Romero-Pittman 2006 Hoorn et al 2017Latrubesse et al 2010) The sediment discharge in theAmazon fan was relatively modest until 5Ma and vastlyincreased in the Pliocene-Pleistocene (Albert et al2018a Hoorn et al 2017) The lower course of theAmazon River may have become an impassable barrierfor these taxa only from the Miocene-Pliocene boundaryonward Moreover this period also coincides with vege-tational and climatic changes notably the expansion ofgrasslands not only in the Andes and the Cerrado butalso within Amazonia (Kirschner amp Hoorn 2019)Subsequently ie the last 5 My both lineages diver-
sified extensively in Western Amazonia notably alongthe foothills of the Andes This diversification has prob-ably been fostered by the combination of increasingavailability of suitable terra-firme habitat due to theretreat of the lacustrine ecosystem and dynamic rivercapture (Albert et al 2018a) The ancestors of the Bcalcarata clade have presumably diversified in the west-ern Acre system ie between the Andes and theAmazon rainforest in the east (Latrubesse et al 2010)Only recently dispersal towards the east seems to haveoccurred notably in B calcarata This last species dis-plays the largest range in the clade which may beexplained by the large diversity of rivers used by thatspecies and the fact that large rivers do not representefficient barriers for its dispersalThe diversification of the B albopunctata clade has
taken place both in Amazonia and in the Cerrado andappears overall more recent than within the Amazonianclade These lineages probably originated from transi-tional ecosystems between the Dry Diagonal andAmazonia This diversification may be partly related tochanges in climate and probable forest retreat in theeastern part of Amazonia during the Late Pliocene andPleistocene (Cheng et al 2013 Ledru et al 2000Pennington et al 2000 Van der Hammen ampHooghiemstra 2000)
AcknowledgementsThis study benefited from an Investissement drsquoAvenirgrant managed by the Agence Nationale de laRecherche (CEBA ref ANR-10-LABX-25-01 TULIPref ANR-10-LABX-0041 ANAEE-France ANR-11-
INBS-0001) AF and FPW acknowledge the FrenchBrazilian GUYAMAZON program action (IRD CNRSCTG CIRAD and Brazilian Fundac~ao de Amparo aPesquisa do Estado do Amazonas-FAPEAM1013039501100004916 062009622018) MTR thanks ConselhoNacional de Desenvolvimento Cientıfico e Tecnologico(CNPq)1013039501100003593 Fundac~ao de Amparo aPesquisa do Estado de S~ao Paulo [FAPESP1013039501100001807 grant numbers 200310335-8 201150146-6] and NSF-FAPESP Dimensions of BiodiversityProgram [grant numbers BIOTA 201350297-0 NSF-DEB 1343578] and NASA1013039100000104 SRRacknowledges a grant from SENESCYT (Arca de NoeInitiative) FPW thanks CNPq (Productivity Fellowship)FAPEAM Coordenac~ao de Aperfeicoamento de Pessoalde Nıvel Superior-CAPES (Visiting ProfessorFellowship) and the LrsquoOreal-UNESCO For Women InScience Program TH thanks CNPq (ProductivityFellowship) and CNPqSISBIOTA [5633482010-0]Financial support was received through a research grantfrom the National Council for Scientific andTechnological Development (CNPq 4469352014-0)TRC is a recipient of a postdoctoral fellowship fromFAPESP (201708489-0) PM is a recipient of aMasterrsquos fellowship from Coordenac~ao deAperfeicoamento de Pessoal de Nıvel Superior (CAPES888872013562018-00) AAG receives financialsupport and grants from CNPq (4469352014-0 3009032015-4 and 3051692019-0) We thank the assistance ofAG Lopes BFV Teixeira and DL Bang as fieldcompanions We are grateful to CFB Haddad MLLyra and Centro de Estudos de Insetos Sociais (CEIS)for providing financial and logistic support and trainingin DNA sequencing partly funded by a research grantfrom FAPESP (201350741-7 CFBH) The MacaulayLibrary (ML) at the Cornell Lab of OrnithologyFonoteca Neotropical Jacques Vielliard (FNJV) enabledaccess to sound files The Cornell Lab of Ornithologyprovided a free license of Raven Pro Software We alsowarmly thank members of MTR lab and Arcadis-Logosfor help in fieldwork as well as J Lima M Blanc J-P Vacher E Courtois B Villette M Dewynter QMartinez R Jairam and P Ouboter for theircontribution with material used in this work
Supplemental dataSupplemental data for this article can be accessed herehttpsdoiorg1010801477200020211873869
194319441945194619471948194919501951195219531954195519561957195819591960196119621963196419651966196719681969197019711972197319741975197619771978197919801981198219831984198519861987198819891990199119921993199419951996
199719981999200020012002200320042005200620072008200920102011201220132014201520162017201820192020202120222023202420252026202720282029203020312032203320342035203620372038203920402041204220432044204520462047204820492050
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 19
antoinefouquet
Texte inseacutereacute
We finally thank Peter Olson (EiC) Mark Wilkinson (AE) and two anonymous reviewers for their comments and suggestions on the manuscript
antoinefouquet
Texte inseacutereacute
S Castroviejo-Fisher J M Padial
PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
ORCIDAntoine Fouquet httporcidorg0000-0003-4060-0281Pedro Marinho httporcidorg0000-0001-6432-5312Thiago R Carvalho httporcidorg0000-0003-0910-2583Marcel A Caminer httporcidorg0000-0002-5827-7462Miguel T Rodrigues httporcidorg0000-0003-3958-9919Ariovaldo A Giaretta httporcidorg0000-0001-7054-129XSantiago Ron httporcidorg0000-0001-6300-9350
ReferencesAcosta-Galvis A R Lasso C A amp Morales-Betancourt
M A (2018) First record of Boana maculateralis (Camineramp Ron 2014) and Boana tetete (Caminer amp Ron
Q1 2014)(Anura Hylidae) in Colombia Check List 14(3)549ndash554 httpsdoiorg1015560143549Q2
Albert J S Val P amp Hoorn C (2018a) The changingcourse of the Amazon River in the Neogene center stagefor Neotropical diversification Neotropical Ichthyology16(3) e180033 httpsdoiorg1015901982-0224-20180033
Albert J S Craig J M Tagliacollo V A amp Petry P(2018b) Upland and lowland fishes A test of the RiverCapture Hypothesis In C M Hoorn A Perrigo amp AAntonelli (Eds) Mountains Climate and Biodiversity (pp273ndash294) Wiley
Antonelli A Ariza M Albert J Andermann T AzevedoJ Bacon C Faurby S Guedes T Hoorn C LohmannL G Matos-Maravı P Ritter C D Sanmartın ISilvestro D Tejedor M Ter Steege H Tuomisto HWerneck F P Zizka A amp Edwards S V (2018)Conceptual and empirical advances in Neotropicalbiodiversity research PeerJ 6 e5644 httpsdoiorg107717peerj5644
Avila R W amp Kawashita-Ribeiro R A (2011)Herpetofauna of S~ao Jo~ao da Barra Hydroelectric Plantstate of Mato Grosso Check List 7(6) 750ndash755 httpsdoiorg101556011014
Avila-Pires T C S D Hoogmoed M S amp RochaW A D (2010) Notes on the Vertebrates of northern ParaBrazil a forgotten part of the Guianan Region IHerpetofauna Boletim Do Museu Paraense Emılio Goeldi5(1) 13ndash122
Bicudo T C Sacek V de Almeida R P Bates J M ampRibas C C (2019) Andean tectonics and MantleDynamics as a Pervasive Influence on Amazonianecosystem Scientific Reports 9(1) 1ndash11 httpsdoiorg101038s41598-019-53465-y
Bokermann W C A (1967) Nova espece de Hyla do Amapa(Amphibia Hylidae) Revista Brasileira de Biologia 27109ndash112
Bouckaert R Heled J Keurouhnert D Vaughan T WuC H Xie D Suchard M A Rambaut A ampDrummond A J (2014) BEAST 2 A Software Platformfor Bayesian Evolutionary Analysis PLoS Comput Biol
10(4) e1003537 httpsdoiorg101371journalpcbi1003537
Boulenger G A (1905) Descriptions of new taillessbatrachians in the collection of the British Museum Annalsand Magazine of Natural History 16(92) 180ndash184 httpsdoiorg10108003745480509443666
Caminer M A amp Ron S R (2014) Systematics of treefrogsof the Hypsiboas calcaratus and Hypsiboas fasciatusspecies complex (Anura Hylidae) with the description offour new species Zookeys 370 1
Caminer M A amp Ron S R (2020) Molecular phylogenyand morphology of Ecuadorian frogs of the genus Boana(Anura Hylidae) with the description of two new speciesZoological Journal of the Linnean Society 190(1)149ndash180 httpsdoiorg101093zoolinneanzlaa002
Campbell K E Jr Frailey C D amp Romero-Pittman L(2006) The Pan-Amazonian Ucayali Peneplain lateNeogene sedimentation in Amazonia and the birth of themodern Amazon River system PalaeogeographyPalaeoclimatology Palaeoecology 239(1-2) 166ndash219httpsdoiorg101016jpalaeo200601020
Camurugi F Gehara M Fonseca E M Zamudio K RHaddad C F B Colli G R Thome M T C PradoC P A Napoli M F amp Garda A A (2021) Isolationby environment and recurrent gene flow shaped theevolutionary history of a continentally distributedNeotropical treefrog Journal of Biogeography in press Q3
Caramaschi U amp de Niemeyer H (2003) New species ofthe Hyla albopunctata group from central Brazil (AmphibiaAnura Hylidae) Boletim do Museu Nacional Nova SerieZoologia 504 1ndash8
Carvalho T R Giaretta A A amp Facure K G (2010) Anew species of Hypsiboas Wagler (Anura Hylidae) closelyrelated to H multifasciatus Geurounther from southeasternBrazil Zootaxa 2521 37ndash52
Carvalho T R Bang D L Teixeira B F V amp GiarettaA A (2017) First record of Boana alfaroi (Caminer ampRon 2014) (Anura Hylidae) in Brazil Check List 13135ndash139
Carvalho T R Moraes L C J L Lima A P Fouquet APeloso P L V Pavan D Drummond L O RodriguesM T Giaretta A A Gordo M Neckel-Oliveira S ampHaddad C F B (2020) Systematics and historicalbiogeography of Neotropical foam-nesting frogs of theAdenomera heyeri clade (Leptodactylidae) with thedescription of six new Amazonian species ZoologicalJournal of the Linnean Society in press httpsdoiorg101093zoolinneanzlaa051 Q4
Ceballos G Ehrlich P R Barnosky A D Garcıa APringle R M amp Palmer T M (2015) Acceleratedmodern human-induced species losses Entering the sixthmass extinction Science Advances 1(5) e1400253 httpsdoiorg101126sciadv1400253
Cheng H Sinha A Cruz F W Wang X Edwards R LdrsquoHorta F M Ribas C C Vuille M Stott L D ampAuler A S (2013) Climate change patterns in Amazoniaand biodiversity Nature Communications 4(1) 1ndash6 httpsdoiorg101038ncomms2415
Cole C J Townsend C R Reynolds R P MacCullochR D amp Lathrop A (2013) Amphibians and reptiles ofGuyana South America illustrated keys annotated speciesaccounts and a biogeographic synopsis Proceedings of theBiological Society of Washington 125(4) 317ndash578 httpsdoiorg1029880006-324X-1254317
205120522053205420552056205720582059206020612062206320642065206620672068206920702071207220732074207520762077207820792080208120822083208420852086208720882089209020912092209320942095209620972098209921002101210221032104
210521062107210821092110211121122113211421152116211721182119212021212122212321242125212621272128212921302131213221332134213521362137213821392140214121422143214421452146214721482149215021512152215321542155215621572158
20 A Fouquet et al
Author Query
AQ4 Please provide the volume number and page range
Author Query
AQ3 Please provide the volume number and page range
Author Query
AQ2 There is no mention of (Ceballos et al 2015 Fouquet et al 2007a Fouquet et al 2012 Guumlnther 1858 Guumlnther 1859 Klaus amp Matzke 2020 Silva et al 2020 and Sturaro et al 2020) in the text Please insert a citation in the text or delete the reference as appropriate
Author Query
AQ1 Please provide complete details for (Naka et al 2018 Naka et al 2018 Hime et al 2020 Molak amp Ho 2015 Papadopoulou et al 2010 Drummond amp Rambaut 2007 Bioacoustics Research Program 2012 Ronquist 1997 Landis et al 2013 Ree amp Smith 2008 Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011) in the reference list or delete the citation from the text
antoinefouquet
Texte inseacutereacute
Center for Conservation Bioacoustics (2014) Raven Pro Interactive Sound Analysis Software (Version 15) Computer software available at httpravensoundsoftwarecom The Cornell Lab of Ornithology Ithaca New York USA
antoinefouquet
Note
We addedmodified these references in the list except Avila‐Pires et al 2010 and Avila amp Kawashita ‐Ribeiro 2011 which were already listed (the issue being the Aacute of Aacutevila was sometimes not properly formatted in the text)
antoinefouquet
Note
We deleted the references from the list except Guumlnther 1858 Guumlnther 1859 that are actually cited in the text L148 and 152
antoinefouquet
Note
this article is still in early view httpsdoiorg101111jbi14035
antoinefouquet
Barrer
antoinefouquet
Texte inseacutereacute
91(2) 395ndash433 httpsdoiorg101093zoolinneanzlaa051
antoinefouquet
Note
we added the missing information
PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Cope E D (1862 Catalogues of the reptiles obtained duringthe explorations of the Parana Paraguay Vermejo andUruguay Rivers by Capt Thos J Page USN and ofthose procured by Lieut N Michler US Top EngCommander of the expedition conducting the survey of theAtrato River [Paper presentation] Proceedings of theAcademy of Natural Sciences of Philadelphia 14 346ndash359
Cope E D (1871) Eighth contribution to the herpetology oftropical America Proceedings of the AmericanPhilosophical Society 11 553ndash559 1870
De la Riva I (1990) Lista preliminar comentada de losanfibios de Bolivia con datos sobre su distribucionBollettino Museo Regionale di Scienze Naturali Tori 8261ndash319
de Sa R O Tonini J F R van Huss H Long A CuddyT Forlani M C Peloso P L Zaher H amp HaddadC F (2019) Multiple connections between Amazonia andAtlantic Forest shaped the phylogenetic and morphologicaldiversity of Chiasmocleis Mehely 1904 (AnuraMicrohylidae Gastrophryninae) Molecular Phylogeneticsand Evolution 130 198ndash210 httpsdoiorg101016jympev201810021
Dewynter M Marty C Blanc M Gaucher P Vidal NFretey T de Massary J C Fouquet A (2008) Liste desAmphibiens et des Reptiles de Guyane Available onlinehttpwwwchelidaecompdfdewynter2008pdf
Drummond A J Ho S Y W Phillips M J amp RambautA (2006) Relaxed phylogenetics and dating withconfidence PLoS Biology 4(5) e88 httpsdoiorg101371journalpbio0040088
Dubois A (2017) The nomenclatural status of HysaplesiaHylaplesia Dendrobates and related nomina (AmphibiaAnura) with general comments on zoological nomenclatureand its governance as well as on taxonomic databases andwebsites Bionomina 11(1) 1ndash48 httpsdoiorg1011646bionomina1111
Duellman W E Marion A B amp Hedges S B (2016)Phylogenetics classification and biogeography of thetreefrogs (Amphibia Anura Arboranae) Zootaxa 4104(1)1ndash109 httpsdoiorg1011646zootaxa410411
Dupin J Matzke N J Seuroarkinen T Knapp S OlmsteadR G Bohs L amp Smith S D (2017) Bayesianestimation of the global biogeographical history of theSolanaceae Journal of Biogeography 44(4) 887ndash899httpsdoiorg101111jbi12898
Ezard T Fujisawa T amp Barraclough T (2009) splitsSPeciesrsquo LImits by Threshold Statistics R package version10-11r29
Faivovich J Haddad C F Garcia P C Frost D RCampbell J A amp Wheeler W C (2005) Systematicreview of the frog family Hylidae with special reference toHylinae phylogenetic analysis and taxonomic revisionBulletin of the American Museum of Natural History294(1) 1ndash240 httpsdoiorg1012060003-0090(2005)294[0001SROTFF]20CO2
Feng Y J Blackburn D C Liang D Hillis D M WakeD B Cannatella D C amp Zhang P (2017)Phylogenomics reveals rapid simultaneous diversificationof three major clades of Gondwanan frogs at theCretaceous-Paleogene boundary Proceedings of theNational Academy of Sciences of the United States ofAmerica 114(29) E5864ndashE5870 httpsdoiorg101073pnas1704632114
Ficetola G F Rondinini C Bonardi A Katariya VPadoa-Schioppa E amp Angulo A (2014) An evaluation ofthe robustness of global amphibian range maps Journal ofBiogeography 41(2) 211ndash221 httpsdoiorg101111jbi12206
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007) Underestimation of species richnessin Neotropical frogs revealed by mtDNA analyses PLoSOne 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Cassini C S Haddad C F B Pech N ampRodrigues M T (2014) Species delimitation patterns ofdiversification and historical biogeography of theNeotropical frog genus Adenomera (AnuraLeptodactylidae) Journal of Biogeography 41(5) 855ndash870httpsdoiorg101111jbi12250
Fouquet A Martinez Q Zeidler L Courtois E AGaucher P Blanc M Lima J D Souza S MRodrigues M T Lima J D Souza S M RodriguesM T amp Kok P J R (2016) Cryptic diversity in theHypsiboas semilineatus species group (Amphibia Anura)with the description of a new species from the easternGuiana Shield Zootaxa 4084(1) 79ndash104 httpsdoiorg1011646zootaxa408413
Fouquet A Gilles A Vences M Marty C Blanc M ampGemmell N J (2007a) Underestimation of speciesrichness in Neotropical frogs revealed by mtDNA analysesPLoS One 2(10) e1109 httpsdoiorg101371journalpone0001109
Fouquet A Loebmann D Castroviejo-Fisher S PadialJ M Orrico V G Lyra M L Roberto I J Kok P JHaddad C F amp Rodrigues M T (2012) From Amazoniato the Atlantic forest Molecular phylogeny ofPhyzelaphryninae frogs reveals unexpected diversity and astriking biogeographic pattern emphasizing conservationchallenges Molecular Phylogenetics and Evolution 65(2)547ndash561 httpsdoiorg101016jympev201207012
Fouquet A Vidal N amp Dewynter M (2019) TheAmphibians of the Mitaraka massif French GuianaZoosystema 41(sp1) 359ndash374 httpsdoiorg105252zoosystema2019v41a19
Frost D R (2019) Amphibian Species of the World anOnline Reference Version 61 (Date of access) ElectronicDatabase accessible at httpsamphibiansoftheworldamnhorgindexphp American Museum of Natural History NewYork USA
Funk W C Caminer M amp Ron S R (2012) High levelsof cryptic species diversity uncovered in Amazonian frogsProceedings of the Royal Society B Biological Sciences279(1734) 1806ndash1814 httpsdoiorg101098rspb20111653 Q5
Gaige H T (1929) Three new tree-frogs from Panama andBolivia Occasional Papers of the Museum of ZoologyUniversity of Michigan 207 1ndash6
Gehara M Crawford A J Orrico V G D Rodrıguez ALeurootters S Fouquet A Barrientos L S Brusquetti FDe la Riva I Ernst R Urrutia G G Glaw FGuayasamin J M Heuroolting M Jansen M Kok P J RKwet A Lingnau R Lyra M hellip Keuroohler J (2014)High levels of diversity uncovered in a widespread nominaltaxon continental phylogeography of the Neotropical treefrog Dendropsophus minutus PloS One 9(9) e103958httpsdoiorg101371journalpone0103958
215921602161216221632164216521662167216821692170217121722173217421752176217721782179218021812182218321842185218621872188218921902191219221932194219521962197219821992200220122022203220422052206220722082209221022112212
221322142215221622172218221922202221222222232224222522262227222822292230223122322233223422352236223722382239224022412242224322442245224622472248224922502251225222532254225522562257225822592260226122622263226422652266
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 21
Author Query
AQ5 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Drummond A J amp Rambaut A (2007) BEAST Bayesian evolutionary analysis by sampling trees BMC evolutionary biology 7(1) 1ndash8
antoinefouquet
Barrer
PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Godinho M B d C amp Da Silva F R (2018) The influenceof riverine barriers climate and topography on thebiogeographic regionalization of Amazonian anuransScientific Reports 8(1) 1ndash11 httpsdoiorg101038s41598-018-21879-9
Guerra V Jardim L Llusia D Marquez R amp BastosR P (2020) Knowledge status and trends in description ofamphibian species in Brazil Ecological Indicators 118106754 httpsdoiorg101016jecolind2020106754
Guillory W X French C M Twomey E M Chavez GPrates I von May R De la Riva I Leurootters S ReichleS Serrano-Rojas S J Whitworth A amp Brown J L(2020) Phylogenetic relationships and systematics of theAmazonian poison frog genus Ameerega usingultraconserved genomic elements Mol Phylogenet Evol142 106638 httpsdoiorg101016jympev2019106638
Geurounther A C L G (1858) Neue Batrachier in derSammlung des britischen Museums Archiv FeurourNaturgeschichte 24 319ndash328 httpsdoiorg105962bhlpart5288
Geurounther A C L G (1859) 1858 Catalogue of theBatrachia Salientia in the Collection of the British MuseumTaylor and Francis
Haffer J (1969) Speciation in Amazonian forest birdsScience (New York NY)) 165(3889) 131ndash137 httpsdoiorg101126science1653889131
Heyer W R Rand A S Cruz C A G Peixoto O L ampNelson C E (1990) Frogs of Boraceia Arquivos deZoologia 31 231ndash410
Hoorn C Wesselingh F P ter Steege H BermudezM A Mora A Sevink J Sanmartin I Sanchez-Meseguer A Anderson C L Figueiredo J P JaramilloC Riff D Negri F R Hooghiemstra H Lundberg JStadler T Seuroarkinen T amp Antonelli A (2010) Amazoniathrough time Andean uplift climate change landscapeevolution and biodiversity Science (New York NY))330(6006) 927ndash931 httpsdoiorg101126science1194585
Hoorn C Bogota-A G R Romero-Baez M LammertsmaE I Flantua S G Dantas E L Dino R do CarmoD A amp Chemale F Jr (2017) The Amazon at seaOnset and stages of the Amazon River from a marinerecord with special reference to Neogene plant turnover inthe drainage basin Global and Planetary Change 15351ndash65 httpsdoiorg101016jgloplacha201702005
Jansen M Bloch R Schulze A amp Pfenninger M (2011)Integrative inventory of Boliviarsquos lowland anurans revealshidden diversity Zoologica Scripta 40(6) 567ndash583 httpsdoiorg101111j1463-6409201100498x
Jaramillo C Romero I DApolito C Bayona G DuarteE Louwye S Escobar J Luque J Carrillo-Brice~noJ D Zapata V Mora A Schouten S Zavada MHarrington G Ortiz J amp Wesselingh F P (2017)Miocene flooding events of western Amazonia ScienceAdvances 3(5) e1601693 httpsdoiorg101126sciadv1601693
Jenkins C N Pimm S L amp Joppa L N (2013) Globalpatterns of terrestrial vertebrate diversity and conservationProceedings of the National Academy of Sciences 110(28)E2602ndashE2610 httpsdoiorg101073pnas1302251110
Jungfer K-H Faivovich J Padial J M Castroviejo-FisherS Lyra M M V M Berneck B Iglesias P P KokP J R MacCulloch R D Rodrigues M T VerdadeV K Torres Gastello C P Chaparro J C Valdujo
P H Reichle S Moravec J Gvozdık V Gagliardi-Urrutia G Ernst R hellip F B Haddad C (2013)Systematics of spiny-backed treefrogs (HylidaeOsteocephalus) an Amazonian puzzle Zoologica Scripta42(4) 351ndash380 httpsdoiorg101111zsc12015
Kapli P Lutteropp S Zhang J Kobert K Pavlidis PStamatakis A amp Flouri T (2017) Multi-rate Poisson treeprocesses for single-locus species delimitation undermaximum likelihood and Markov chain Monte CarloBioinformatics (Oxford England)) 33(11) 1630ndash1638httpsdoiorg101093bioinformaticsbtx025
Katoh K Rozewicki J amp Yamada K D (2019) MAFFTonline service multiple sequence alignment interactivesequence choice and visualization Briefings inBioinformatics 20(4) 1160ndash1166 httpsdoiorg101093bibbbx108
Kirschner J A amp Hoorn C (2019) The onset of grasses inthe Amazon drainage basin evidence from the fossil recordFrontiers of Biogeography 12 e44827
Klaus K V amp Matzke N J (2020) Statistical Comparisonof Trait-dependent Biogeographical Models indicates thatPodocarpaceae Dispersal is influenced by both Seed ConeTraits and Geographical Distance Systematic Biology69(1) 61ndash75 httpsdoiorg101093sysbiosyz034
Keuroohler J Jansen M Rodrıguez A Kok P J R ToledoL F Emmrich M Glaw F Haddad C F B Reuroodel M-O amp Vences M (2017) The use of bioacoustics in anurantaxonomy theory terminology methods andrecommendations for best practice Zootaxa 4251(1)1ndash124 httpsdoiorg1011646zootaxa425111
Kok P J Bittenbinder M A van den Berg J K Marques-Souza S Sales Nunes P M Laking A E Teixeira MJr Fouquet A Means D B MacCulloch R D ampRodrigues M T (2018) Integrative taxonomy of thegymnophthalmid lizard Neusticurus rudis Boulenger 1900identifies a new species in the eastern Pantepui regionnorth-eastern South America Journal of Natural History52(13-16) 1029ndash1066 httpsdoiorg1010800022293320181439541
Lanfear R Frandsen P B Wright A M Senfeld T ampCalcott B (2016) PartitionFinder 2 New methods forselecting partitioned models of evolution for molecular andmorphological phylogenetic analyses Molecular Biologyand Evolution 34(3) 772ndash773
Latrubesse E M Cozzuol M da Silva-Caminha S ARigsby C A Absy M L amp Jaramillo C (2010) TheLate Miocene paleogeography of the Amazon Basin and theevolution of the Amazon River system Earth-ScienceReviews 99(3-4) 99ndash124 httpsdoiorg101016jearscirev201002005
Ledru M-P Blanc P Charles-Dominique P Fournier MMartin L Riera B amp Tardy C (2000) Reconstitution delrsquoecosysteme forestier guyanais au cours de lrsquoHolocenesuperieur apport de la palynologie In M Servan amp SServant Vildar (Eds) Dynamique a long terme desecosystemes forestiers intertropicaux (pp 199ndash204)UNESCO
Leite R N amp Rogers D S (2013) Revisiting Amazonianphylogeography Insights into diversification hypothesesand novel perspectives Organisms Diversity amp Evolution13(4) 639ndash664 httpsdoiorg101007s13127-013-0140-8
Lescure J amp Marty C (2000) Atlas des amphibiens deGuyane Collection patrimoines naturels
226722682269227022712272227322742275227622772278227922802281228222832284228522862287228822892290229122922293229422952296229722982299230023012302230323042305230623072308230923102311231223132314231523162317231823192320
232123222323232423252326232723282329233023312332233323342335233623372338233923402341234223432344234523462347234823492350235123522353235423552356235723582359236023612362236323642365236623672368236923702371237223732374
22 A Fouquet et al
antoinefouquet
Texte inseacutereacute
Hime P M Lemmon A R Lemmon E C M Prendini E Brown J M Thomson R C Kratovil J D Noonan B P Pyron R A Peloso P L Kortyna M L Keogh J S Donnellan S C Mueller R L Raxworthy C J Kunte K Ron S Das S Gaitonde N Green D M Labisko J Che J amp Weisrock D W (2021) Phylogenomics reveals ancient gene tree discordance in the amphibian tree of life Systematic biology 70(1) 49ndash66
antoinefouquet
Texte inseacutereacute
Landis M J Matzke N J Moore B R amp Huelsenbeck J P (2013) Bayesian analysis of biogeography when the number of areas is large Systematic biology 62(6) 789ndash804
antoinefouquet
Texte surligneacute
this ref is cited L152
antoinefouquet
Texte surligneacute
this ref is cited L 148
antoinefouquet
Barrer
PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Ligges U Krey S Mersmann O Schnackenberg S(2014) TuneR Analysis of music Accessed May 8 2017Available at httpr-forger-projectorgprojectstuner
Lima A P Keller C amp Rebelo G H (2017) Estudosambientais no Rio Madeira trecho Cachoeira de SantoAntonio-Abun~a (Rondonia) Herpetofauna Relatorioelaborado para Furnas Centrais Eletricas SA como parte doEstudo de Viabilidade dos AHEs Santo Antonio e Jiraupara o Aproveitamento Hidreletrico do Rio Madeira INPAManaus
Lovejoy T E amp Nobre C (2019) Amazon tipping pointLast chance for action Science Advances 5(12) eaba2949httpsdoiorg101126sciadvaba2949Q6
Marinho P Costa-Campos C E Pezzuti T L Magalh~aesR F Souza M R D Haddad C F B Giaretta A Aamp De Carvalho T R (2020) The Amapa treefrog Boanadentei (Bokermann 1967) diagnosis and redescriptionJournal of Natural History 54(15-16) 971ndash990 httpsdoiorg1010800022293320201777336
Matzke N J (2013) BioGeoBEARS biogeography withBayesian (and likelihood) evolutionary analysis in R scriptsR Package Version 02 1 2013
McCormack J E Heled J Delaney K S Peterson A Tamp Knowles L L (2011) Calibrating divergence times onspecies trees versus gene trees implications for speciationhistory of Aphelocoma jays Evolution InternationalJournal of Organic Evolution 65(1) 184ndash202 httpsdoiorg101111j1558-5646201001097x
Meyer C Kreft H Guralnick R amp Jetz W (2015) Globalpriorities for an effective information basis of biodiversitydistributions Nature Communications 6(1) 1ndash8 httpsdoiorg101038ncomms9221
Medina-Rangel G F Mendez-Galeano M A amp Calderon-Espinosa M L (2019) Herpetofauna of San Jose delGuaviare Guaviare Colombia Biota Colombiana 20(1)75ndash90 httpsdoiorg1021068c2019v20n01a05
Medina-Rangel G F Thompson M E Ruiz-ValderramaD H Fajardo Mu~noz W Lombana Lugo J Londo~no-Guarnizo C A Moquena Carbajal C Rıos Rosero H DSanchez Pamo J E amp Sanchez E (2019) Anfibios yreptiles In N Pitman A Salazar Molano F SamperSamper C Vriesendorp A Vasquez Ceron A del CampoT L Miller E A Matapi Yucuna M E Thompson L deSouza D Alvira Reyes D F Stotz N Kotlinski TWachter E Woodward amp R Botero Garcıa (Eds)Colombia Bajo CaguanndashCaqueta Rapid Biological andSocial Inventories report 30 (pp 111ndash454) Field Museum
Meza-Joya F L Ramos-Pallares E amp Hernandez-Jaimes C(2019) Hidden diversity in frogs within Boanacalcaratandashfasciata and Boana geographica speciescomplexes from Colombia Herpetology Notes 12391ndash400
Monaghan M T Wild R Elliot M Fujisawa T BalkeM Inward D J G Lees D C Ranaivosolo REggleton P Barraclough T G amp Vogler A P (2009)Accelerated species inventory on Madagascar usingcoalescent-based models of species delineation SystematicBiology 58(3) 298ndash311 httpsdoiorg101093sysbiosyp027
Meurouller L (1924) Neue Batrachier aus Ost-BrasilienSenckenbergiana Biologica 6 169ndash177
Myers N Mittermeier R A Mittermeier C G da FonsecaG A amp Kent J (2000) Biodiversity hotspots forconservation priorities Nature 403(6772) 853ndash858
Near T J Eytan R I Dornburg A Kuhn K L MooreJ A Davis M P Wainwright P C Friedman M ampSmith W L (2012) Resolution of ray-finned fishphylogeny and timing of diversification Proceedings of theNational Academy of Sciences 109(34) 13698ndash13703httpsdoiorg101073pnas1206625109
Noble G K (1923) Six new batrachians from the DominicanRepublic American Museum Novitates 61 1ndash6
Oliveira U Vasconcelos M F amp Santos A J (2017)Biogeography of Amazon birds rivers limit speciescomposition but not areas of endemism Scientific Reports7(1) 1ndash11 httpsdoiorg101038s41598-017-03098-w
Ouboter P E amp Jairam R (2012) Amphibians of SurinameBrill 376 pp
Pansonato A Avila R W Kawashita-Ribeiro R A ampMorais D H (2011) Advertisement call and newdistribution records of Hypsiboas leucocheilus (AnuraHylidae) Salamandra 47 55ndash58
Parker H W ( (1927) ) The brevicipitid frogs allied to thegenus Hypopachus Occasional Papers of the Museum ofZoology University of Michigan 187 1ndash6
Pennington R T Prado D E amp Pendry C A (2000)Neotropical seasonally dry forests and Quaternaryvegetation changes Journal of Biogeography 27(2)261ndash273 httpsdoiorg101046j1365-2699200000397x
Peloso P L De Oliveira R M Sturaro M J RodriguesM T Lima-Filho G R Bitar Y O Wheeler W C ampAleixo A (2018) Phylogeny of map tree frogs Boanasemilineata species Group with a new amazonian species(Anura Hylidae) South American Journal of Herpetology13(2) 150ndash169 httpsdoiorg102994SAJH-D-17-000371
Pinheiro P D P Cintra C E D Valdujo P H da SilvaH L R Martins I A da Silva N J Jr amp GarciaP C A (2018) A new species of the Boana albopunctatagroup (Anura Hylidae) from the Cerrado of Brazil SouthAmerican Journal of Herpetology 13(2) 170ndash182 httpsdoiorg102994SAJH-D-17-000401
Pirani R M Werneck F P Thomaz A T Kenney M LSturaro M J Avila-Pires T C S Peloso P L VRodrigues M T amp Knowles L L (2019) Testing mainAmazonian rivers as barriers across time and space withinwidespread taxa Journal of Biogeography 46(11)2444ndash2456 httpsdoiorg101111jbi13676
Pons J Barraclough T G Gomez-Zurita J Cardoso ADuran D P Hazell S Kamoun S Sumlin W D ampVogler A P (2006) Sequence-based species delimitationfor the DNA taxonomy of undescribed insects SystematicBiology 55(4) 595ndash609 httpsdoiorg10108010635150600852011
Prado C P Haddad C F B amp Zamudio K R (2012)Cryptic lineages and Pleistocene population expansion in aBrazilian Cerrado frog Molecular Ecology 21(4) 921ndash941httpsdoiorg101111j1365-294X201105409x
Puillandre N Lambert A Brouillet S amp Achaz G (2012)ABGD Automatic Barcode Gap Discovery for primaryspecies delimitation Molecular Ecology 21(8) 1864ndash1877httpsdoiorg101111j1365-294X201105239x
R Core Team (2018) R a language and environment forstatistical computing R Foundation for StatisticalComputing Version 350 Accessed July 29 2020Available at httpswwwR-projectorg
Ree R H amp Sanmartın I (2018) Conceptual and statisticalproblems with the DECthorn J model of founder-eventspeciation and its comparison with DEC via model
237523762377237823792380238123822383238423852386238723882389239023912392239323942395239623972398239924002401240224032404240524062407240824092410241124122413241424152416241724182419242024212422242324242425242624272428
242924302431243224332434243524362437243824392440244124422443244424452446244724482449245024512452245324542455245624572458245924602461246224632464246524662467246824692470247124722473247424752476247724782479248024812482
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 23
Author Query
AQ6 The year of publication has been changed as per Crossref details both in the list and in the text for this reference Please check
antoinefouquet
Texte inseacutereacute
Naka L N amp Brumfield R T (2018) The dual role of Amazonian rivers in the generation and maintenance of avian diversity Science advances 4(8) eaar8575
antoinefouquet
Texte inseacutereacute
Molak M amp Ho S Y (2015) Prolonged decay of molecular rate estimates for metazoan mitochondrial DNA PeerJ 3 e821
antoinefouquet
Texte inseacutereacute
Papadopoulou A Anastasiou I amp Vogler A P (2010) Revisiting the insect mitochondrial molecular clock the mid-Aegean trench calibration Molecular biology and evolution 27(7) 1659ndash1672
PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
selection Journal of Biogeography 45(4) 741ndash749 httpsdoiorg101111jbi13173
Rejaud A Rodrigues M T Crawford A J Castroviejo-Fisher S Jaramillo A F Chaparro J C Glaw FGagliardi-Urrutia G Moravec J De la Riva I J PerezP Lima A P Werneck F P Hrbek T Ron S RErnst R Kok P J R Driskell A Chave J amp FouquetA (2020) Historical biogeography identifies a possible roleof the Pebas system in the diversification of the Amazonianrocket frogs (Aromobatidae Allobates) Journal ofBiogeography 47(11) 2472ndash2482 httpsdoiorg101111jbi13937
Ribas C C Aleixo A Nogueira A C R Miyaki C Y ampCracraft J (2012) A palaeobiogeographic model for bioticdiversification within Amazonia over the past three millionyears Proceedings Biological Sciences 279(1729)681ndash689 httpsdoiorg101098rspb20111120
Rodrigues D J Noronha J C Vindica V F amp BarbosaF R (2015) Biodiversidade do Parque Estadual doCristalino Attema Editorial
Rojas-Zamora R R de Carvalho V T Avila R W deAlmeida A P de Oliveira E A Menin M amp GordoM (2017) Hypsiboas maculateralis Caminer amp Ron 2014new to Brazil Herpetozoa 30 108ndash114
Rojas R R Fouquet A Ron S R Hernandez-Ruz E JMelo-Sampaio P R Chaparro J C Vogt R C deCarvalho V T Pinheiro L C Avila R W Farias I PGordo M amp Hrbek T (2018) A Pan-Amazonian speciesdelimitation high species diversity within the genusAmazophrynella (Anura Bufonidae) PeerJ 6 e4941httpsdoiorg107717peerj4941
Ruokolainen K Moulatlet G M Zuquim G Hoorn C ampTuomisto H (2018) River network rearrangements inAmazonia shake biogeography and civil security Preprints2018090168
Ruthven A G (1927) Description of an apparently newspecies of Apostolepis from Bolivia Occasional Papers ofthe Museum of Zoology University of Michigan 188 1ndash2
Savage J M amp Heyer W R (1997) Digital webbingformulae for anurans a refinement Herpetological Review28 131
Sheu Y Zurano J P Ribeiro-Junior M A Avila-PiresT C S Rodrigue M Ts Colli G R amp Werneck F P(2020) The combined role of dispersal and niche evolutionin the diversification of Neotropical lizards Ecology andEvolution 10(5) 2608ndash2625 httpsdoiorg101002ece36091
Silva L A Magalh~aes F M Thomassen H Leite F S FGarda A A Brand~ao R A Haddad C F B GiarettaA A amp Carvalho T R (2020) Unraveling the speciesdiversity and relationships in the Leptodactylus mystaceuscomplex (Anura Leptodactylidae) with the description ofthree new Brazilian species Zootaxa 4779 151ndash189
Spix J B v (1824) Animalia nova sive Species novaeTestudinum et Ranarum quas in itinere per Brasiliam annisMDCCCXVIIndashMDCCCXX jussu et auspiciis MaximilianiJosephi I Bavariae Regis F S Heuroubschmann
Stamatakis A (2014) RAxML version 8 a tool forphylogenetic analysis and post-analysis of large
phylogenies Bioinformatics (Oxford England)) 30(9)1312ndash1313 httpsdoiorg101093bioinformaticsbtu033
Sturaro M J Costa J C L Maciel A O Lima-FilhoG R Rojas-Runjaic F J Mejia D P Ron S R ampPeloso P L (2020) Resolving the taxonomic puzzle ofBoana cinerascens (Spix 1824) with resurrection of Hylagranosa gracilis Melin 1941 (Anura Hylidae) Zootaxa4750(1) 1ndash30 httpsdoiorg1011646zootaxa475011
Sueur J Aubin T amp Simonis C (2008) Seewave a freemodular tool for sound analysis and synthesis Bioacoustics18(2) 213ndash226 httpsdoiorg1010800952462220089753600
Troschel F H (1848) Theil 3 Versuch einerZusammenstellung der Fauna und Flora von Britisch-Guiana Schomburgk R ed Reisen in Britisch-Guiana inden Jahren 1840ndash44 Im Auftrage Sr Majesteuroat des Keuroonigsvon Preussen ausgefeurouhrt 645ndash661J J Weber
IUCN (2020) The IUCN Red List of Threatened SpeciesVersion 2020-1 httpswwwiucnredlistorg Downloadedon 19 March 2020
Vacher J-P Chave J Ficetola F Sommeria-Klein GTao S Thebaud C Blanc M Camacho A CassimiroJ Colston T J Dewynter M Ernst R Gaucher PGome s J O Jairam R Kok P J R Dias Lima JMartinez Q Marty C hellip Fouquet A (2020) Largescale DNA-based survey of Amazonian frogs suggest a vastunderestimation of species richness and endemism Journalof Biogeography 47(8) 1781ndash1791 httpsdoiorg101111jbi13847
Van der Hammen T amp Hooghiemstra H (2000) Neogeneand Quaternary history of vegetation climate and plantdiversity in Amazonia Quaternary Science Reviews 19(8)725ndash742 httpsdoiorg101016S0277-3791(99)00024-4
Vences M Thomas M Bonett R M amp Vieites D R(2005) Deciphering amphibian diversity through DNAbarcoding chances and challenges PhilosophicalTransactions of the Royal Society B Biological Sciences360(1462) 1859ndash1868 httpsdoiorg101098rstb20051717
Wallace A R (1854) On the monkeys of the AmazonAnnals and Magazine of Natural History 14(84) 451ndash454httpsdoiorg101080037454809494374
Watters J L Cummings S T Flanagan R L amp SilerC D (2016) Review of morphometric measurements usedin anuran species descriptions and recommendations for astandardized approach Zootaxa 4072(4) 477ndash495 httpsdoiorg1011646zootaxa407246
Werneck F P (2011) The diversification of eastern SouthAmerican open vegetation biomes historical biogeographyand perspectives Quaternary Science Reviews 30(13-14)1630ndash1648 httpsdoiorg101016jquascirev201103009
Associate Editor Dr Mark Wilkinson
248324842485248624872488248924902491249224932494249524962497249824992500250125022503250425052506250725082509251025112512251325142515251625172518251925202521252225232524252525262527252825292530253125322533253425352536
253725382539254025412542254325442545254625472548254925502551255225532554255525562557255825592560256125622563256425652566256725682569257025712572257325742575257625772578257925802581258225832584258525862587258825892590
24 A Fouquet et al
antoinefouquet
Texte inseacutereacute
13Ronquist F (1997) Dispersal-vicariance analysis a new approach to the quantification of historical biogeography Systematic Biology 46(1) 195ndash203
antoinefouquet
Texte inseacutereacute
Ree R H amp Smith S A (2008) Maximum likelihood inference of geographic range evolution by dispersal local extinction and cladogenesis Systematic biology 57(1) 4ndash14
antoinefouquet
Texte inseacutereacute
Aacute
antoinefouquet
Barrer
antoinefouquet
Barrer
PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
259125922593259425952596259725982599260026012602260326042605260626072608260926102611261226132614261526162617261826192620262126222623262426252626262726282629263026312632263326342635263626372638263926402641264226432644
264526462647264826492650265126522653265426552656265726582659266026612662266326642665266626672668266926702671267226732674267526762677267826792680268126822683268426852686268726882689269026912692269326942695269626972698
Appendix 1 Samples included in molecular analyses including results from DNA-basedspecies delineation Some of this newly analysed material was obtained through QCAZMPEG INPA (H) CORBIDI AAGUFU collections and from personal loans from JoseManuel Padial (JMP) Santiago Castroviejo Fisher (SCF) Jucivaldo Dias Lima (FTA)Quentin Martinez (QM) Michel Blanc (AG)
GenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtAF467269 CM015 B_raniceps 1 13 1 1 1 Crique YiYi French_Guiana 5483333 5315gtAY843657 MACN37795 B_raniceps 1 13 1 2 1 Santa Fe Vera Argentina 29459031 60208386gtJF790127 AS0169 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtJF790128 AS0218 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790129 AS0303 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790130 AS0304 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790132 AS0306 B_raniceps 1 13 1 2 1 Velasco Caparu Bolivia 14912117 61082467gtJF790133 AS0468 B_raniceps 1 13 1 1 1 Yucuma Los Lagos Bolivia 12772 658109gtJF790134 AS0570 B_raniceps 1 13 1 2 1 nuflo de Chavez San Sebastian Bolivia 16359633 62000050gtKDQF01002617 H3905 B_raniceps 1 13 1 1 1 UHE Jirau Mutum RO 9601348 650574150gtKDQF01002739 LPT38 B_raniceps 1 13 1 1 1 Laranjal do Jari AP 0600855 5238887gtKDQF01002991 MPEG36750 B_raniceps 1 13 1 2 1 Canaa dos Carajas PA 6075562 50243245gtKDQF01004033 PNA105 B_raniceps 1 13 1 2 1 Parque Nacional do Araguaia TO 108166667 501833333gtKDQF01004063 QM052 B_raniceps 1 13 1 1 1 Guatemala French_Guiana 515 52633333gtKF723064 MNKA9349 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723065 SMF94093 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723066 SMF94094 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723067 SMF94263 B_raniceps 1 13 1 2 1 Estancia Bechler Bolivia 1751720 6328998gtKF723068 MNKA10459 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723069 SMF94241 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKF723070 SMF94246 B_raniceps 1 13 1 2 1 San Sebastian Bolivia 163596 620001gtKU495281 CFBHT00940 B_raniceps 1 13 1 2 1 Santa Fe do Sul SP 20228 50944gtKU495282 CFBHT02353 B_raniceps 1 13 1 2 1 Cafelandia e Cafesopolis SP 21838 497gtKU495283 CFBHT13473 B_raniceps 1 13 1 2 1 campus de Veterinaria da UFBA Bahia 11935 38112gtKU495284 CFBHT07327 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495285 TG146 B_raniceps 1 13 1 2 1 Estreito MA 6548 47445gtKU495286 TG265 B_raniceps 1 13 1 2 1 Paranaita MT 9671 56482gtKU495287 CFBHT07328 CFBHT07328 B_raniceps 1 13 1 2 1 Pariquera-Acu SP 2466 47796gtKU495288 TG158 B_raniceps 1 13 1 2 1 Porto Franco MA 635 47406gtMW370296 AG234 B_raniceps 1 13 1 1 1 Comte Terrain michel French_Guiana 4679471 52338531gtMW370317 MTR36896 B_raniceps 1 13 1 1 1 Tefe AM 336396 6469653gtKDQF01003728 MTR33845 B_afflanciformis 2 5 2 13 2 Tefe Estrada do Abial AM 345944009 647509004gtMW370318 MTR36907 B_afflanciformis 2 5 2 13 2 Tefe AM 334554 6471122gtAY326054 WED54081 B_lanciformis 4 6 4 3 4 Puyo Pastaza Ecuador 1375502 78004787gtAY843636 MJH564 B_lanciformis 4 6 4 4 4 Loreto Alpahuayo Peru 394403 73606866gtJN970644 QCAZ23803 B_lanciformis 4 6 4 3 4 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970645 QCAZ23809 B_lanciformis 4 6 4 3 4 Mendez River N of Ecuador 27291 782849gtJN970646 QCAZ30936 B_lanciformis 4 6 4 3 4 Huino Ecuador 0606 77173gtJN970647 QCAZ18175 B_lanciformis 4 6 4 3 4 Estacion Biologica Jatun Sacha Ecuador 10675 776152778gtJN970648 QCAZ20641 B_lanciformis 4 6 4 3 4 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970649 QCAZ18237 B_lanciformis 4 6 4 3 4 Nangaritza Ecuador 4335192 7867974gtJN970650 QCAZ31017 B_lanciformis 4 6 4 3 4 Centro Shuar Yawi Ecuador 44301 78631608gtKF157594 isolate7768 B_lanciformis 4 6 4 5 4 Venezuela NA NAgtKP149371 AJC3975 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149451 AJC3373 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149455 AJC3971 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtKP149465 AJC3973 B_lanciformis 4 6 4 5 4 Casanare Sabanalarga Colombia 4773 73037gtMW370316 MTR36402 B_lanciformis 4 6 4 4 4 S~ao Pedro Rio Ica AM 303339 6888413gtAY843613 NMP6V71250 B_affmaculateralis 5 11 5 45 5 Anguilla Peru Peru 3662914 73924860gtKDQF01003645 MTR28015 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 744309497 736576660gtKDQF01003658 MTR28077 B_affmaculateralis 5 11 5 36 5 Serra do Divisor AC 743399298 736609450gtMW370306 CORBIDI6182 B_affmaculateralis 5 11 5 36 5 Jenaro Herrera Requena Peru 4903812 73663577gtMW370314 MTR35995 B_affmaculateralis 5 11 5 46 5 Comunidade Cuiaua Rio Ica AM 288619 6837169gtMW370312 JMP2306 B_affmaculateralis 5 11 5 36 5 quebradas_Pinsha_y_Yanayacu Loreto 512 738gtJN970541 QCAZ40082 B_maculateralis 6 12 6 37 6 Comunidad Santa Rosa Ecuador 102337 7748359gtJN970545 QCAZ43710 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970546 QCAZ43712 B_maculateralis 6 12 6 37 6 Coca Ecuador 047782 7698982gtJN970552 QCAZ44452 B_maculateralis 6 12 6 37 6 Chiroisla north bank Ecuador 057564999 758998299gtJN970555 QCAZ44184 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970556 QCAZ44185 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970559 QCAZ44248 B_maculateralis 6 12 6 37 6 Eden Ecuador 049833 7607112gtJN970562 QCAZ44636 B_maculateralis 6 12 6 37 6 Huiririma Ecuador 071163 7562395gtJN970563 QCAZ44651 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970564 QCAZ44673 B_maculateralis 6 12 6 37 6 Santa Teresita Ecuador 090087 7541357gtJN970568 QCAZ43825 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970570 QCAZ43827 B_maculateralis 6 12 6 37 6 La Primavera south bank Ecuador 044427 7678685gtJN970572 QCAZ43897 B_maculateralis 6 12 6 37 6 La Primavera north bank Ecuador 0431 7678648gtJN970576 QCAZ44531 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970577 QCAZ44532 B_maculateralis 6 12 6 37 6 San Vicente Ecuador 067901 7565112gtJN970595 QCAZ28401 B_maculateralis 6 12 6 37 6 Cuyabeno beach Ecuador 0265433 75891733gtJN970598 QCAZ44020 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970599 QCAZ44021 B_maculateralis 6 12 6 37 6 La Selva Lodge Ecuador 050868 7636493gtJN970611 QCAZ27940 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 25
PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
269927002701270227032704270527062707270827092710271127122713271427152716271727182719272027212722272327242725272627272728272927302731273227332734273527362737273827392740274127422743274427452746274727482749275027512752
275327542755275627572758275927602761276227632764276527662767276827692770277127722773277427752776277727782779278027812782278327842785278627872788278927902791279227932794279527962797279827992800280128022803280428052806
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970612 QCAZ28004 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970613 QCAZ28024 B_maculateralis 6 12 6 37 6 Zabalo Ecuador 0318133 7576625gtJN970523 QCAZ18271 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970524 QCAZ48584 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970525 QCAZ48585 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970526 QCAZ48593 B_fasciata 7 20 7 35 7 La Pradera Via Gualaquiza Ecuador 2285 78094gtJN970531 QCAZ26497 B_fasciata 7 20 7 35 7 Limon-Macas road km 8 Ecuador 28858 7839708gtJN970534 QCAZ17016 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970535 QCAZ17030 B_fasciata 7 20 7 35 7 Tiink Ecuador 3333 78451806gtJN970619 QCAZ41488 B_fasciata 7 20 7 35 7 Las Orquidias Ecuador 422903 7865775gtJN970620 QCAZ41575 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970621 QCAZ41576 B_fasciata 7 20 7 35 7 Miazi Alto Ecuador 425044 7861356gtJN970622 QCAZ24866 B_fasciata 7 20 7 35 7 Romerillos Alto Ecuador 422725 7893875gtJN970623 QCAZ31037 B_fasciata 7 20 7 35 7 Centro Shuar Yawi Ecuador 44301 78631608gtJN970624 QCAZ23147 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970625 QCAZ23148 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970626 QCAZ48583 B_fasciata 7 20 7 35 7 Zamora Ecuador 406695 78950817gtJN970520 QCAZ32638 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970521 QCAZ32639 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970522 QCAZ32645 B_almendarizae 8 21 8 33 8 Guamote-Macas road Ecuador 221635 7828986gtJN970527 QCAZ25958 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970528 QCAZ26300 B_almendarizae 8 21 8 33 8 Limon Ecuador 29046 78386931gtJN970529 QCAZ39647 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970530 QCAZ39650 B_almendarizae 8 21 8 33 8 Limon via Gualaceo Ecuador 29796 7844147gtJN970532 QCAZ23810 B_almendarizae 8 21 8 33 8 Mendez River N of Ecuador 27291 782849gtJN970533 QCAZ26438 B_almendarizae 8 21 8 33 8 San Juan Bosco Ecuador 202 7792gtJN970538 QCAZ17944 B_almendarizae 8 9 8 34 8 Rio Hollin Ecuador 069547 7772958gtJN970614 QCAZ24392 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970615 QCAZ24394 B_almendarizae 8 9 8 34 8 Rio Lagarto Ecuador 0678184 76396556gtJN970616 QCAZ31449 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970617 QCAZ31450 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtJN970618 QCAZ31452 B_almendarizae 8 9 8 34 8 Rio Pastaza Ecuador 141283 7826878gtAY326056 KU202911 B_calcarata 9 10 9 29 9 Misahualli Ecuador 1033333 77670278gtJN970536 QCAZ40085 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970537 QCAZ40084 B_calcarata 9 10 9 29 9 8 km S of Tena Ecuador 10449 7776951gtJN970542 QCAZ43935 B_calcarata 9 10 9 29 9 anangu Ecuador 052492 7638445gtJN970547 QCAZ43713 B_calcarata 9 10 9 29 9 Coca Ecuador 047782 7698982gtJN970548 QCAZ43789 B_calcarata 9 10 9 29 9 9 km S of Coca Ecuador 049986 7700828gtJN970550 QCAZ44422 B_calcarata 9 10 9 29 9 Chiroisla Ecuador 057564999 758998299gtJN970553 QCAZ44177 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970558 QCAZ44247 B_calcarata 9 10 9 29 9 Eden Ecuador 049833 7607112gtJN970567 QCAZ43824 B_calcarata 9 10 9 29 9 La Primavera south bank Ecuador 044427 7678685gtJN970575 QCAZ44530 B_calcarata 9 10 9 29 9 San Vicente Ecuador 067901 7565112gtJN970578 QCAZ16798 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970580 QCAZ43256 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970581 QCAZ43259 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970584 QCAZ20290 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970585 QCAZ20305 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970586 QCAZ17825 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 0678194 763965555gtJN970587 QCAZ31446 B_calcarata 9 10 9 29 9 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970588 QCAZ14956 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970589 QCAZ14958 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970590 QCAZ14957 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970591 QCAZ14965 B_calcarata 9 10 9 29 9 Canelos Ecuador 159 7775gtJN970597 QCAZ25434 B_calcarata 9 10 9 29 9 La Selva Lodge Ecuador 0498167 76373833gtJN970606 QCAZ28181 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970607 QCAZ28185 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970608 QCAZ28197 B_calcarata 9 10 9 29 9 Cuyabeno Ecuador 00886 761419gtJN970628 WED57649 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970630 WED58712 B_calcarata 9 10 9 30 9 Cuzco Amazonico Peru 12583 69083gtJN970631 WED59271 B_calcarata 9 10 9 42 9 Cuzco Amazonico Peru 12583 69083gtJN970633 CM131 B_calcarata 9 10 9 32 9 crique Margot French_Guiana 5443190 53972344gtJN970635 PG105 B_calcarata 9 10 9 32 9 Kaw route French_Guiana 4516105 5210053gtJN970636 PG123 B_calcarata 9 10 9 32 9 Toponowini French_Guiana 30521 52705gtJN970639 AG217 B_calcarata 9 10 9 32 9 Trinite Crique Grand Leblond French_Guiana 46709 532843gtKDQF01000164 AF0605 B_calcarata 9 10 9 32 9 Trou Poisson mare French_Guiana 5373475 53097575gtKDQF01000277 AF0848 B_calcarata 9 10 9 32 9 Chutes Voltaire camp French_Guiana 5052639 54088444gtKDQF01000291 AF0867 B_calcarata 9 10 9 32 9 Chutes gregoire French_Guiana 5097472 53050667gtKDQF01000338 AF0941 B_calcarata 9 10 9 32 9 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01000408 AF1106 B_calcarata 9 10 9 32 9 Mana Ilet Lezard rive G French_Guiana 5054481 538007gtKDQF01000537 AF1441 B_calcarata 9 10 9 32 9 Belizon Mare French_Guiana 4368663 52322931gtKDQF01000583 AF1543 B_calcarata 9 10 9 32 9 Saul roche bateau French_Guiana 360654 5317625gtKDQF01000587 AF1549 B_calcarata 9 10 9 32 9 Matoury French_Guiana 485708 52339175gtKDQF01000646 AF1726 B_calcarata 9 10 9 32 9 Chutes Voltaire French_Guiana 503255 5408689gtKDQF01000760 AF1952 B_calcarata 9 10 9 31 9 Sipaliwini Suriname 202439 5612508gtKDQF01000908 AF2341 B_calcarata 9 10 9 32 9 Ekini French_Guiana 405 524667gtKDQF01000986 AF2562 B_calcarata 9 10 9 32 9 RN2 PK65 mare French_Guiana 4487368 52347811gtKDQF01001120 AF2849 B_calcarata 9 10 9 31 9 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001145 AF2931 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001158 AF2954 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001159 AF2955 B_calcarata 9 10 9 32 9 Memora G French_Guiana 331774 5219329gtKDQF01001184 AF3000 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001185 AF3001 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001193 AF3022 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001194 AF3023 B_calcarata 9 10 9 32 9 Memora D AP 331294 5218036gtKDQF01001212 AF3060 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001213 AF3061 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404
(continued)
26 A Fouquet et al
PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
280728082809281028112812281328142815281628172818281928202821282228232824282528262827282828292830283128322833283428352836283728382839284028412842284328442845284628472848284928502851285228532854285528562857285828592860
286128622863286428652866286728682869287028712872287328742875287628772878287928802881288228832884288528862887288828892890289128922893289428952896289728982899290029012902290329042905290629072908290929102911291229132914
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01001214 AF3062 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001220 AF3069 B_calcarata 9 10 9 32 9 Mitan G French_Guiana 26284 5255404gtKDQF01001277 AF3162 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001278 AF3164 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001279 AF3165 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001283 AF3170 B_calcarata 9 10 9 32 9 Mitan D AP 262764 5254195gtKDQF01001323 AF3248 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001331 AF3257 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001341 AF3270 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001342 AF3271 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001343 AF3272 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001344 AF3273 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001345 AF3274 B_calcarata 9 10 9 32 9 Taparabo AP 4016066 51693802gtKDQF01001353 AF3287 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001354 AF3288 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001355 AF3289 B_calcarata 9 10 9 32 9 Trois paletuviers French_Guiana 4054582 51677007gtKDQF01001384 AF3357 B_calcarata 9 10 9 32 9 Bakhuis Creek 10k Suriname 465616 5678649gtKDQF01001425 AF3454 B_calcarata 9 10 9 32 9 Spari Creek Suriname 523285 5580462gtKDQF01001842 BM335 B_calcarata 9 10 9 41 9 UHE Belo Monte PA 2950584 51936607gtKDQF01001921 BPN0802 B_calcarata 9 10 9 31 9 Kayser Suriname 3048297 56578653gtKDQF01002023 BPN2893 B_calcarata 9 10 9 32 9 RAP Basecamp Suriname 246554 5562955gtKDQF01002048 BPN3456 B_calcarata 9 10 9 31 9 Mapari Guyana 3348083 59275833gtKDQF01002149 CAAM38 B_calcarata 9 10 9 32 9 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002510 FL45 B_calcarata 9 10 9 32 9 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002600 H2487 B_calcarata 9 10 9 30 9 UHE Jirau RO 959492 650650814gtKDQF01002916 MPEG30468 B_calcarata 9 10 9 31 9 Paru FLOTA PA 09439694 532363000gtKDQF01003034 MTD47753 B_calcarata 9 10 9 32 9 Kabo forestry consession Suriname 5378132 55621491gtKDQF01003057 MTD47926 B_calcarata 9 10 9 32 9 Iwokrama Guyana 46713889 58685gtKDQF01003166 MTR10021 B_calcarata 9 10 9 44 9 Lago Cipotuba AM 58013889 602211111gtKDQF01003568 MTR24073 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKDQF01003575 MTR24122 B_calcarata 9 10 9 32 9 Oiapoque AP 38794167 517709722gtKDQF01003750 MTR968364 B_calcarata 9 10 9 43 9 Apiacas MT 9650846 57393665gtKDQF01003993 PK1504 B_calcarata 9 10 9 32 9 Kaieteur NP Guyana 514976 59483128gtKDQF01004113 QM442 B_calcarata 9 10 9 32 9 Montagne de fer French_Guiana 54074 535548gtKDQF01004357 TJC1246 B_calcarata 9 10 9 32 9 Black Water Creek Guyana 507311 5924635gtKR811158 MTR24072 B_calcarata 9 10 9 32 9 Oiapoque AP 38355278 518333333gtKR811159 DAN012 B_calcarata 9 10 9 32 9 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKU221856 B_calcarata 9 10 9 29 9 San Jacinto Peru 4674982 73958995gtMF583738 LEO711 B_calcarata 9 10 9 29 9 Putumayo Valle del Guamuez 04161 768752gtMW370322 AF3769 B_calcarata 9 10 9 32 9 Voltzberg Suriname 468169 5618568gtMW370325 FTA182 B_calcarata 9 10 9 32 9 Rio Amapa Grande AP 2127111 51191333gtMW370328 JMP1569 B_calcarata 9 10 9 29 9 Confluencia Supai_y_Sabalo Peru 3065 72168gtAF467270 CM B_dentei 10 14 10 14 10 Kaw French_Guiana 4516105 5210053gtKDQF01000097 AF0233 B_dentei 10 14 10 14 10 Montagne des singes French_Guiana 5066667 52716667gtKDQF01000135 AF0540 B_dentei 10 14 10 14 10 Savane virginie layon French_Guiana 41959 52149gtKDQF01000227 AF0727 B_dentei 10 14 10 14 10 Paracou French_Guiana 52754 529236gtKDQF01000319 AF0913 B_dentei 10 14 10 14 10 Pic Coudreau du Sud French_Guiana 22534 543534gtKDQF01000377 AF1029 B_dentei 10 14 10 14 10 Saul Galbao French_Guiana 361125 53288548gtKDQF01000450 AF1212 B_dentei 10 14 10 14 10 Trinite French_Guiana 46025 534143gtKDQF01000495 AF1330 B_dentei 10 14 10 14 10 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000501 AF1337 B_dentei 10 14 10 14 10 Machicou French_Guiana 3890031 52573535gtKDQF01000578 AF1529 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 41959 52149gtKDQF01000637 AF1695 B_dentei 10 14 10 14 10 Saul Limonade French_Guiana 356311 5319083gtKDQF01000846 AF2107 B_dentei 10 14 10 14 10 RN2 corridor 5 French_Guiana 403265 5199093gtKDQF01001036 AF2645 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 357236 5396731gtKDQF01001065 AF2697 B_dentei 10 14 10 14 10 Alikene camp polissoir French_Guiana 321792 5239747gtKDQF01001109 AF2797 B_dentei 10 14 10 14 10 Mitaraka layon C French_Guiana 223577 5444928gtKDQF01001202 AF3041 B_dentei 10 14 10 14 10 Memora G French_Guiana 331774 5219329gtKDQF01001218 AF3066 B_dentei 10 14 10 14 10 Mitan G French_Guiana 26284 5255404gtKDQF01001264 AF3146 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001289 AF3180 B_dentei 10 14 10 14 10 Mitan D AP 262764 5254195gtKDQF01001763 AP024 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002148 CAAM37 B_dentei 10 14 10 14 10 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002205 CM013 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01002219 CM090 B_dentei 10 14 10 14 10 Tibourou French_Guiana 4416667 523gtKDQF01002394 DAN015 B_dentei 10 14 10 14 10 St Georges RN 2 PK168 French_Guiana 400126 51931057gtKDQF01002403 DAN037 B_dentei 10 14 10 14 10 St Georges piste Saut Maripa French_Guiana 3862884 51863422gtKDQF01002418 DSM064 B_dentei 10 14 10 14 10 Savane Virginie French_Guiana 4199878 52136443gtKDQF01002513 FL90 B_dentei 10 14 10 14 10 Plote PPBIO Porto Grande AP 0979331 51614895gtKDQF01002531 FTA170 B_dentei 10 14 10 14 10 Rio Amapa Grande AP 2127111 51191333gtKDQF01003258 MTR13753 B_dentei 10 14 10 14 10 Serra do Navio AP 09180556 520027778gtKDQF01003277 MTR13850 B_dentei 10 14 10 14 10 Lourenco AP 23236111 516452778gtKDQF01003811 PG032 B_dentei 10 14 10 14 10 Kaw2 French_Guiana 4516105 5210053gtKDQF01003812 PG035 B_dentei 10 14 10 14 10 Matecho French_Guiana 375 53033333gtKDQF01003843 PG234 B_dentei 10 14 10 14 10 Haute Wanapi French_Guiana 25134 538211gtMW370338 AG428 B_dentei 10 14 10 14 10 Atachi Bakka French_Guiana 36553 53844gtMW370343 AG509 B_dentei 10 14 10 14 10 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370344 AG095BM029 B_dentei 10 14 10 14 10 Kaw2 CD6PK 47 French_Guiana 4571 522206gtMW370346 AG216 B_dentei 10 14 10 14 10 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370320 AF2984 B_dentei 10 14 10 14 10 Memora D AP 328215 5220263gtMW370321 AF3628 B_dentei 10 14 10 14 10 Itoupe_400 French_Guiana 302208 5311046gtMW370323 AM009 B_dentei 10 14 10 14 10 Inini_Tolenga French_Guiana 3663159 53928308gtMW370324 BOAM034 B_dentei 10 14 10 14 10 Borne 4 French_Guiana 23709 537728gtMW370326 FTA91 B_dentei 10 14 10 14 10 Rio Vila Nova AP 0449335 52022137gtJN970539 QCAZ40080 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970540 QCAZ40081 B_tetete 13 16 13 17 13 Comunidad Santa Rosa Ecuador 102137 7747819gtJN970627 WED60015 B_tetete 13 17 13 18 13 San Jacinto Ecuador 23125 758628gtKU221865 B_tetete 13 17 13 18 13 San Jacinto Peru 4674982 73958995
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 27
PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
291529162917291829192920292129222923292429252926292729282929293029312932293329342935293629372938293929402941294229432944294529462947294829492950295129522953295429552956295729582959296029612962296329642965296629672968
296929702971297229732974297529762977297829792980298129822983298429852986298729882989299029912992299329942995299629972998299930003001300230033004300530063007300830093010301130123013301430153016301730183019302030213022
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtJN970543 QCAZ43977 B_alfaroi 14 2 14 19 14 anangu Ecuador 052492 7638445gtJN970544 QCAZ43682 B_alfaroi 14 2 14 19 14 Coca Ecuador 047782 7698982gtJN970549 QCAZ44351 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057997 7591769gtJN970551 QCAZ44425 B_alfaroi 14 2 14 19 14 Chiroisla Ecuador 057564999 758998299gtJN970554 QCAZ44180 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970557 QCAZ44191 B_alfaroi 14 2 14 19 14 Eden Ecuador 049833 7607112gtJN970560 QCAZ44634 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970561 QCAZ44635 B_alfaroi 14 2 14 19 14 Huiririma Ecuador 071163 7562395gtJN970565 QCAZ44788 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970566 QCAZ44790 B_alfaroi 14 2 14 19 14 Nuevo Roacafuerte Ecuador 091927 7540103gtJN970569 QCAZ43826 B_alfaroi 14 2 14 19 14 La Primavera south bank Ecuador 044427 7678685gtJN970571 QCAZ43894 B_alfaroi 14 2 14 19 14 La Primavera north bank Ecuador 0431 7678648gtJN970573 QCAZ44527 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970574 QCAZ44528 B_alfaroi 14 2 14 19 14 San Vicente Ecuador 067901 7565112gtJN970579 QCAZ19328 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970582 QCAZ43262 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970583 QCAZ43263 B_alfaroi 14 2 14 19 14 Estacion Cientifica Yasuni Ecuador 068931 7642903gtJN970592 QCAZ28272 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970593 QCAZ28278 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0265433 75891733gtJN970594 QCAZ28398 B_alfaroi 14 2 14 19 14 Cuyabeno beach Ecuador 0241517 75930533gtJN970596 QCAZ25410 B_alfaroi 14 2 14 19 14 La Selva Lodge Ecuador 0498167 76373833gtJN970600 QCAZ44025 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970601 QCAZ44027 B_alfaroi 14 2 14 19 14 La Selva Lodge entrance Ecuador 050868 7636493gtJN970602 QCAZ44851 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970603 QCAZ44853 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970604 QCAZ44856 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970605 QCAZ44858 B_alfaroi 14 2 14 19 14 Panacocha 25 km S of Ecuador 04712 766667gtJN970609 QCAZ28240 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtJN970610 QCAZ28315 B_alfaroi 14 2 14 19 14 Cuyabeno Ecuador 00886 761419gtKF955305 QCAZ50785 B_alfaroi 14 2 14 19 14 Panacocha Ecuador 04712 766667gtMF583737 LEO720 B_alfaroi 14 2 14 19 14 Putumayo Valle del Guamuez Colombia 04161 768752gtJN970634 CM168 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtJN970637 CM118 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4884444 52258611gtKDQF01000183 AF0641 B_aff courtoisae 15 4 15 16 15 Cayenne Rorota French_Guiana 4879124 52257589gtKDQF01000335 AF0937 B_aff courtoisae 15 4 15 16 15 Cayenne Vidal French_Guiana 4884433 52288819gtKDQF01000337 AF0940 B_aff courtoisae 15 4 15 16 15 Matoury Lac des Americains French_Guiana 4852891 52352092gtKDQF01002239 CM149 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002246 CM174 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002266 CM262 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002267 CM263 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002271 CM273 B_aff courtoisae 15 4 15 16 15 Guatemala French_Guiana 515 52633333gtKDQF01002424 DSM100 B_aff courtoisae 15 4 15 16 15 Monsinery French_Guiana 480338 5247466gtKDQF01004062 QM051 B_aff courtoisae 15 4 15 16 15 Matiti French_Guiana 5035425 52565573gtKDQF01004066 QM076 B_aff courtoisae 15 4 15 16 15 Tonnegrande grenouillere French_Guiana 4837603 52444249gtAY549335 AMNHA164081 AMNHA164081 B_courtoisae 16 18 16 15 16 Iwokrama Cowfly camp Guyana 46713889 58685gtEU201108 CM229 B_courtoisae 16 18 16 15 16 Saul French_Guiana 3625556 53207222gtJN970638 AG213 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970640 AG221 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46709 532843gtJN970641 AF0097 B_courtoisae 16 18 16 15 16 Road to Apura Suriname 5183333 55616667gtKDQF01000489 MNHN-RA-20200004 AF1322 B_courtoisae 16 18 16 15 16 Saut Taconet French_Guiana 403249 52526188gtKDQF01000494 MNHN-RA-20200005 AF1328 B_courtoisae 16 18 16 15 16 Saut Grand Machicou French_Guiana 3897416 52583565gtKDQF01000584 MNHN-RA-20200007 AF1546 B_courtoisae 16 18 16 15 16 Saul gros arbre French_Guiana 3615576 53227093gtKDQF01000636 AF1694 B_courtoisae 16 18 16 15 16 Saul Limonade French_Guiana 356311 5319083gtKDQF01000753 AF1934 B_courtoisae 16 18 16 15 16 Flat de la Waki French_Guiana 30895 5339846gtKDQF01000829 AF2090 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000830 AF2091 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000847 MNHN-RA-20200009 AF2114 B_courtoisae 16 18 16 15 16 Sipaliwini Suriname 209753 561472gtKDQF01000898 AF2319 B_courtoisae 16 18 16 15 16 Trinite French_Guiana 46025 534143gtKDQF01000909 MNHN-RA-20200013 AF2342 B_courtoisae 16 18 16 15 16 Ekini French_Guiana 405 524667gtKDQF01001002 AF2584 B_courtoisae 16 18 16 15 16 Sinnamary French_Guiana 5372835 52960608gtKDQF01001038 MNHN-RA-20200002 AF2654 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001059 MNHN-RA-20200001 AF2686 B_courtoisae 16 18 16 15 16 Alikene camp 1 French_Guiana 320906 52402gtKDQF01001118 MNHN-RA-20200014 AF2839 B_courtoisae 16 18 16 15 16 Mitaraka layon B French_Guiana 223577 5444928gtKDQF01001211 AF3059 B_courtoisae 16 18 16 15 16 Mitan G French_Guiana 26284 5255404gtKDQF01001424 AF3452 B_courtoisae 16 18 16 15 16 Spari Creek Suriname 523285 5580462gtKDQF01001456 AF3695 B_courtoisae 16 18 16 15 16 Itoupe French_Guiana 301498 5313286gtKDQF01001686 MNHN-RA-20200003 AM007 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001687 AM008 B_courtoisae 16 18 16 15 16 Inini Tolenga French_Guiana 3663159 53928308gtKDQF01001733 AMS394 B_courtoisae 16 18 16 15 16 Ants Creek Guyana 3305817 59324067gtKDQF01001735 AMS420 B_courtoisae 16 18 16 15 16 Land Turtle Landing Swamp Guyana 3180717 59421967gtKDQF01001918 BPN0794 B_courtoisae 16 18 16 15 16 Ralleighvallen Suriname 47166667 562166667gtKDQF01002020 BPN2861 B_courtoisae 16 18 16 15 16 RAP Basecamp 1 Suriname 2477 5562941gtKDQF01002044 BPN3448 B_courtoisae 16 18 16 15 16 Mapari Guyana 3348083 59275833gtKDQF01002140 CAAM28 B_courtoisae 16 18 16 15 16 Montagne cacao French_Guiana 2349128 53215985gtKDQF01002908 MPEG30336 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002909 MPEG30341 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01002910 MPEG30342 B_courtoisae 16 18 16 15 16 REBIO Maicuru PA 08286194 539312000gtKDQF01003773 NZCS45 B_courtoisae 16 18 16 15 16 rosebel Suriname 5124392 55249254gtKDQF01004011 PK3520 B_courtoisae 16 18 16 15 16 Turtle Camp Iwokrama Guyana 46718 586847gtMW370337 AG422 B_courtoisae 16 18 16 15 16 Atachi Bakka French_Guiana 36553 53844gtMW370339 AG444 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370340 AG446 B_courtoisae 16 18 16 15 16 Alikene French_Guiana 3220044 5237855gtMW370342 AG485 B_courtoisae 16 18 16 15 16 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370345 AG072 B_courtoisae 16 18 16 15 16 Trinite Crique Grand Leblond French_Guiana 46709 532843gtMW370333 AF3775 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtMW370334 AF3791 B_courtoisae 16 18 16 15 16 Voltzberg_CI camp Suriname 468169 5618568gtKDQF01002632 HJ071 B_eucharis 17 1 17 40 17 UHE Jirau RO 8748483 63903465
(continued)
28 A Fouquet et al
PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
302330243025302630273028302930303031303230333034303530363037303830393040304130423043304430453046304730483049305030513052305330543055305630573058305930603061306230633064306530663067306830693070307130723073307430753076
307730783079308030813082308330843085308630873088308930903091309230933094309530963097309830993100310131023103310431053106310731083109311031113112311331143115311631173118311931203121312231233124312531263127312831293130
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01002656 HJ443 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01002657 HJ447 B_eucharis 17 1 17 28 17 UHE Jirau RO 969941 65358875gtKDQF01003636 MTR25798 B_eucharis 17 1 17 27 17 Pacaas Novos PARNA RO 107869789 636273050gtKDQF01003752 MTR968510 B_eucharis 17 1 17 27 17 Apiacas MT 9650846 57393665gtKDQF01003759 MTR977049 B_eucharis 17 1 17 27 17 Juruena MT 103236111 584927778gtMW370327 H2472 B_eucharis 17 1 17 28 17 UHE Jirau RO 960283365 650583638gtMW370329 MZUSP 159228 MTR25896 B_eucharis 17 1 17 27 17 Pacaas Novos_PARNA RO 107815800 636267350gtAY843632 AMNHA168405 B_heilprini 18 15 18 47 25 pet_trade NAgtJN970629 WED57865 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtJN970632 WED59350 B_affsteinbachi(2) 12 3 19 26 18 Cuzco Amazonico Peru 12583 69083gtKU205472 WED55554 B_affsteinbachi(2) 12 3 19 26 18 Puerto Maldonado Peru 12618187 69167000gtMW370330 AAGUFU5917 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370331 AAGUFU5918 B_affsteinbachi(2) 12 3 19 26 18 Assis AC 10933098 69594418gtMW370350 SCF395 B_affsteinbachi(2) 12 3 19 26 18 Cocama Peru 10399 71165gtKDQF01003399 MTR18819 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003402 MTR18844 B_affsteinbachi(1) 11 3 11 22 11 Lago Chaviana AM 418314 6148801gtKDQF01003646 MTR28018 B_affsteinbachi(1) 11 3 11 21 11 Serra do Divisor AC 743707300 736571960gtKDQF01003722 MTR33822 B_affsteinbachi(1) 11 3 11 23 11 Japura Trilha da Anta AM 177207062 663725554gtMW370297 CORBIDI11192 B_affsteinbachi(1) 11 3 11 20 11 Saniri Malvinas Peru 11846765 7294gtMW370298 CORBIDI12128 B_affsteinbachi(1) 11 3 11 20 11 Rio Tapiche Peru 7448717 73940944gtMW370299 CORBIDI13035 B_affsteinbachi(1) 11 3 11 21 11 Coronel Pontillo Peru 8662599 74059897gtMW370307 CORBIDI8542 B_affsteinbachi(1) 11 3 11 20 11 Poyentimari Peru 123175 728922gtMW370308 CORBIDI8698 B_affsteinbachi(1) 11 3 11 39 11 Datem del Maranon Peru 4171196 76932370gtMW370309 CORBIDI8974 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370310 CORBIDI8975 B_affsteinbachi(1) 11 3 11 20 11 Rio Corrientes Peru 2735647 76031778gtMW370313 MTR35970 B_affsteinbachi(1) 11 3 11 23 11 Comunidade Cachoeirinha AM 287148 6834707gtMW370315 MTR36143 B_affsteinbachi(1) 11 3 11 23 11 Acaı Rio Ica AM 290452 6833869gtMW370319 MTR36968 B_affsteinbachi(1) 11 3 11 22 11 Coari AM 408864 6309844gtJF790135 MJ1300 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790136 MJ1301 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790137 MJ1302 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtJF790138 MJ1310 B_steinbachi 12 3 12 24 12 Sara Buenavista Bolivia 17481683 63684867gtKDQF01001813 BM048 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01001848 BM355 B_steinbachi 12 3 12 25 12 UHE Belo Monte PA 2950584 51936607gtKDQF01002994 MPEG36753 MPEG36753 B_steinbachi 12 3 12 25 12 Canaa dos Carajas PA 6075562 50243245gtMW370300 CORBIDI13415 CORBIDI13415 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370301 CORBIDI13417 CORBIDI13417 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370302 CORBIDI13453 CORBIDI13453 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370303 CORBIDI13454 CORBIDI13454 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370304 CORBIDI13482 CORBIDI13482 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370305 CORBIDI15138 CORBIDI15138 B_steinbachi 12 3 12 24 12 Tambopata Peru 11962370 70213704gtMW370311 JMP088 B_steinbachi 12 3 12 24 12 Chalalan_Franz_Tamayo Bolivia 14428168 67922330gtAY549317 ZUEC12053 B_albopunctata 3 7 3 6 19 Campinas SP 22926527 47002675gtJQ627203 10 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtJQ627205 114 B_albopunctata 3 7 3 6 19 Sao Carlos SP 2198 4787gtKY002929 MCNAM19674 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002930 MCNAM19670 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY002955 MCNAM19624 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKY007134 MCNAM17920 B_albopunctata 3 7 3 6 19 MG 191694 437148gtKY911220 MCNAM19707 B_albopunctata 3 7 3 6 19 MG 19815346 43661281gtKDQF01002967 MPEG33422 B_spgralbopunctata4 3 7 21 10 20 Projeto Tocantizinho PA 6061700 56296620gtKDQF01002983 MPEG36699 B_spgralbopunctata4 3 7 21 10 20 Itaituba PA 631731705 557986444gtJF790116 AS0470 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12772 658109gtJF790117 MJ1326 B_leucocheila 3 7 20 11 21A Yucuma Los Lagos Bolivia 12771967 65810933gtKDQF01003124 MTR00212 B_leucocheila 3 7 20 11 21A Claudia MT 11585 551355556gtKDQF01003634 MTR25723 B_leucocheila 3 7 20 11 21A Pacaas Novos RO 108158520 636260410gtKDQF01002443 ESTR00081 B_multifasciata 3 7 20 12 21B Carolina MA 722941 4726147gtKDQF01002465 ESTR0652 B_multifasciata 3 7 20 12 21B Estreito MA 656055555 474511111gtMW370347 H02 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtMW370348 H13 B_multifasciata 3 7 20 12 21B Belem PA 1439224 48417892gtKDQF01001810 BM044 B_spgralbopunctata2 3 7 22 25 22 UHE Belo Monte PA 2950584 51936607gtKDQF01003631 MTR25676 B_spgralbopunctata1 3 8 23 38 23 Pacaas Novos RO 107805610 636266239gtAY843648 AMCC101446 B_spgralbopunctata3 3 19 24 7 24 Demerara Guyana 6505246 58108209gtEU201110 AF0047 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtEU201111 CM241 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtJN970642 ROM22756 B_spgralbopunctata3 3 19 24 7 24 Baramita Guyana 73666 604833gtJN970643 ROM28435 B_spgralbopunctata3 3 19 24 8 24 Paramakatoi Guyana 469722 5978666gtKDQF01000029 AF0048 B_spgralbopunctata3 3 19 24 7 24 Petit Saut French_Guiana 5066667 5305gtKDQF01000041 AF0079 B_spgralbopunctata3 3 19 24 9 24 St Georges French_Guiana 3888232 51807804gtKDQF01000163 AF0603 B_spgralbopunctata3 3 19 24 9 24 St elie pist PK8 French_Guiana 5357068 53035859gtKDQF01000340 AF0943 B_spgralbopunctata3 3 19 24 9 24 Matoury French_Guiana 4852891 52352092gtKDQF01000479 AF1272 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573gtKDQF01000592 AF1570 B_spgralbopunctata3 3 19 24 7 24 St elie piste French_Guiana 5357068 53035859gtKDQF01000700 AF1832 B_spgralbopunctata3 3 19 24 9 24 Montagne des Singes French_Guiana 507148 526927gtKDQF01000870 AF2173 B_spgralbopunctata3 3 19 24 9 24 Sipaliwini air strip Suriname 202682 5612564gtKDQF01001448 AF3666 B_spgralbopunctata3 3 19 24 9 24 Itoupe French_Guiana 302208 5311046gtKDQF01002087 BPN3687 B_spgralbopunctata3 3 19 24 7 24 Parabara BC Guyana 2181833 59337017gtKDQF01002218 CM088 B_spgralbopunctata3 3 19 24 9 24 Tibourou French_Guiana 4416667 523gtKDQF01002220 CM093 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002262 CM242 B_spgralbopunctata3 3 19 24 9 24 Saul French_Guiana 3625556 53207222gtKDQF01002301 CM338 B_spgralbopunctata3 3 19 24 9 24 Cayenne Montjoly French_Guiana 4916667 52266667gtKDQF01002415 DSM048 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01002423 DSM097 B_spgralbopunctata3 3 19 24 9 24 Monsinery French_Guiana 480338 5247466gtKDQF01003251 MTR13743 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003256 MTR13750 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003257 MTR13751 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003259 MTR13769 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778
(continued)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 29
PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Appendix 2 Molecular data acquisitionWe extracted DNA from liver or muscle tissue (thigh ortoe-clip) of the 307 samples using the Wizard Genomicextraction protocol (Promega Madison WI USA) Wetargeted a 400bp fragment of the 16S rDNA We usedprimers N16R and N16F (Salducci et al 2005) to whichwe added NNN thorn 8-nucleotide labels (hereafter designatedas lsquotagsrsquo) for sample identification as all resulting PCRproducts were mixed into single libraries 32 tags forforward primer (N16R) and 36 tags for reverse primer(N16F) The final volume of PCRs was 20ml andcontained 2ml of DNA extract diluted 10-fold 10mlof Amplitaq GoldV
R
360 Master Mix (Life TechnologiesCarlsbad CA USA) 584ml of Nuclease Free WaterAmbion (Thermo Fisher Scientific MA USA) 025mMof each primer and 32mg of Bovine Serum Albumin(Roche Diagnostic Basel Switzerland) We ran the PCRin duplicate for each sample and we included blankPCR controls in the analysis using nuclease-free wateras template First the mixture was denaturated at 95 C for10min followed by 40 cycles of 30 s at 95 C 30 s at55 C and 1min at 72 C followed by a final step of 7minat 72 C Libraries of mixed PCR products were sequencedusing 2 250 paired-end reads sequencing technologythrough MiSeq high throughput sequencing (Illumina) atthe Genopole (Toulouse France) The resulting outputswere analysed with the OBITOOLS software suite (Boyeret al 2016a) Paired-end reads were assembled andmerged and we used the tag attached to the primer toassign each reads to its label Then we removed lowquality reads (alignment scores lt50 containing Ns orshorter than 50bp) The resulting batch of reads wasdereplicated while keeping the coverage information(number of reads merged) All sequences lt100bp werediscarded Eventually all the sequences that we included inour dataset were gt380bp long
Mitogenome sequencing assemblingand annotationFor complete mitochondrial sequencing we used 200ng ofgenomic DNA per sample to build libraries at theGenotoul-GeT-PlaGe core facility (Toulouse France)Genomic DNA was fragmented by sonication fragmentswere size-selected (50ndash400bp) adenylated and ligated toindexed sequencing adapters Eight cycles of PolymeraseChain Reaction (PCR) were applied to amplify librariesbefore library quantification and validation A total of 48libraries were multiplexed to be sequenced on one lane onan Illumina HiSeq 3000 flow cell (Illumina Inc SanDiego CA) Read assembly was performed using theORGanelle ASseMbler (Boyer et al 2016b) a pythonprogram developed especially for organelle and ribosomalDNA reconstructions from a genome skimming datasetThe first step is to sort and index the reads with the oaindex command Then with the oa buildgraph commandthe program uses a seed (reference sequence) to assemblethe reads and creates a graph with all the fragmentsassembled Finally the oa unfold command finds anoptimal path in the assembly graph and reconstructs theconsensus sequence (Boyer et al 2016b)For the oa index command we used two options (i)
ndashestimate-length 09 which estimates the readsrsquo lengthusing 90 of the reads and (ii) ndashbypass-filtering whenthe read indexing failed due to an over-filtering thatresulted in an empty index For the oa buildgraphcommand we used default settings in most cases exceptif the assembly graph was empty due to errors incoverage estimation In these cases we modified thendashcoverage value to be slightly lower than the estimateFor the oa unfold command we used the default optionsas long as it was able to find the optimal assembly pathIf not we checked the assembling graph with the yEd
313131323133313431353136313731383139314031413142314331443145314631473148314931503151315231533154315531563157315831593160316131623163316431653166316731683169317031713172317331743175317631773178317931803181318231833184
318531863187318831893190319131923193319431953196319731983199320032013202320332043205320632073208320932103211321232133214321532163217321832193220322132223223322432253226322732283229323032313232323332343235323632373238
ContinuedGenBank accession Voucher Field Number Species_ID mPTP ABGD mPTP GMYC Cons Locality State Lat Long
gtKDQF01003260 MTR13776 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003264 MTR13793 B_spgralbopunctata3 3 19 24 9 24 Serra do Navio AP 09180556 520027778gtKDQF01003299 MTR13953 B_spgralbopunctata3 3 19 24 9 24 Laranjal do Jari AP 07166667 523833333gtKDQF01003454 MTR20411 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 336977 6144177gtKDQF01003472 MTR20525 B_spgralbopunctata3 3 19 24 7 24 EE Maraca RR 33326 613824gtKDQF01003486 MTR20658 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 446905 6113607gtKDQF01003491 MTR20680 B_spgralbopunctata3 3 19 24 8 24 Pacaraima RR 4473034897 611352092gtKDQF01003809 PG026 B_spgralbopunctata3 3 19 24 9 24 Cayenne Rorota French_Guiana 4874784 52261119gtKDQF01003813 PG036 B_spgralbopunctata3 3 19 24 9 24 Kaw2 French_Guiana 4516105 5210053gtKDQF01003845 PG248 B_spgralbopunctata3 3 19 24 9 24 Haute Wanapi French_Guiana 25134 538211gtKDQF01004238 SMNS12093 B_spgralbopunctata3 3 19 24 7 24 Bamboo landing Guyana 5325 580455556gtKDQF01004356 TJC1243 B_spgralbopunctata3 3 19 24 7 24 Black Water Creek Guyana 507297 5924899gtKDQF01004364 TJC1268 B_spgralbopunctata3 3 19 24 7 24 NARIL Guyana 512324 5911266gtKR811154 PG779 B_spgralbopunctata3 3 19 24 9 24 St Georges Savane 14 juillet French_Guiana 3967639 5187225gtKR811155 MTR24173 B_spgralbopunctata3 3 19 24 9 24 Oiapoque AP 37960278 518629167gtKR811156 MTR24194 B_spgralbopunctata3 3 19 24 9 24 Lourenco AP 23526389 516152222gtKR811157 AF0749 B_spgralbopunctata3 3 19 24 9 24 St Georges savane French_Guiana 3925482 51788964gtMW370336 AG334 B_spgralbopunctata3 3 19 24 9 24 RN2 PK103 French_Guiana 436387 5227747gtMW370341 AG480 B_spgralbopunctata3 3 19 24 9 24 Saul Monts Belvedere French_Guiana 3719347 53412809gtMW370332 AF3704 B_spgralbopunctata3 3 19 24 9 24 Itoupe_200 French_Guiana 301498 5313286gtMW370335 AF3816 B_spgralbopunctata3 3 19 24 7 24 Voltzberg_CI camp Suriname 468169 5618568gtMW370349 QM108 B_spgralbopunctata3 3 19 24 9 24 Matiti French_Guiana 5035425 52565573
30 A Fouquet et al
PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Graph Editor (organic layout default parameters) andselected the green fragments (that correspond to similarseed fragments) and extracted them with the ndashpathndashoption in the oa unfold command The fragments werethen mapped onto a seed with the Geneious (R917Biomatters Ltd Auckland New Zealand) map toreference function with medium-low sensitivity and oneiteration The mapping consensus was extracted andconsidered as a mitochondrial genome assemblyFor annotation sequences were aligned on the
MAFFT7 online server under default parameters exceptthe use of E-INS-i strategy for rDNA which is designedfor sequences with multiple conserved domains andlong gaps (Katoh et al 2019) Then using theGeneious R917 transfer annotation function weannotated all the alignment sequences from a referencesequence (Anomaloglossus baeobatrachus NC_030054)Finally we checked that each coding sequence (CDS)
began with a methionine (ATA or ATG codon forvertebrate mitochondrial genomes) and ended with astop codon For the final alignment we kept the CDS12Sndash16S loci
Supplementary referencesBoyer F Coissac E Viari A (2016b) The ORGanelle
ASseMbler Retrieved from httpspypiorgprojectORGasm
Boyer F Mercier C Bonin A Le Bras Y Taberlet P ampCoissac E (2016a) obitools A unix-inspired softwarepackage for DNA metabarcoding Mol Ecol Resour 16(1)176ndash182 httpsdoiorg1011111755-099812428
Salducci M-D Marty C Fouquet A amp Gilles A (2005)Phylogenetic relationships and biodiversity in Hylids(Anura Hylidae) from French Guiana Comptes RendusBiologies 328(10-11) 1009ndash1024 httpsdoiorg101016jcrvi200507005
323932403241324232433244324532463247324832493250325132523253325432553256325732583259326032613262326332643265326632673268326932703271327232733274327532763277327832793280328132823283328432853286328732883289329032913292
329332943295329632973298329933003301330233033304330533063307330833093310331133123313331433153316331733183319332033213322332333243325332633273328332933303331333233333334333533363337333833393340334133423343334433453346
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 31
PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Appendix 3 Completion for themitogenomic matrix
334733483349335033513352335333543355335633573358335933603361336233633364336533663367336833693370337133723373337433753376337733783379338033813382338333843385338633873388338933903391339233933394339533963397339833993400
340134023403340434053406340734083409341034113412341334143415341634173418341934203421342234233424342534263427342834293430343134323433343434353436343734383439344034413442344334443445344634473448344934503451345234533454
mtDNA
Terminals Cons voucher 12S 16S ND1 COI CytB
Boana_eucharis 1 MZUSP 159227 MTR25798Boana aff lanciformis 2 MTR33845 MTR33845Boana aff maculateralis 16 MTR28015 MTR28015Boana aff steinbachi(1) 22 MTR33822 MTR33822Boana_aff steinbachi(2) 24 SCF395 SCF395Boana_albopunctata 5 ZUEC12053 AY549317 AY549370Boana_alfaroi 9 QCAZ44528 QCAZ44528Boana_almendarizae 10 QCAZ31449 QCAZ31449Boana_calcarata 11 H2487 H2487Boana_dentei 12 AF2797 AF2797Boana_fasciata 13 QCAZ24866 QCAZ24866Boana_heilprini 14 AMNHA168405 AY843632 KF794126 AY843864Boana_lanciformis 15 MJH564 AY843636 AY843870Boana_leucocheila 6 MTR25723 MTR25723Boana_maculateralis 17 QCAZ43827 QCAZ43827Boana_multifasciata 8 ESTR00081 ESTR00081Boana_raniceps 19 MPEG36750 MPEG36750Boana_affcourtoisae 20 AF0937 AF0937Boana_courtoisae 21 AF2584 AF2584Boana_sp gr albopunctata 3 18 AF2173 AF2173Boana_sp gr albopunctata 1 7 MTR25676 MTR25676Boana_sp gr albopunctata 2 3 BM044 BM044 BM044Boana_sp gr albopunctata 4 4 MPEG33422 MPEG33422Boana_steinbachi 23 MJ1301 MJ1301Boana_tetete 25 QCAZ40080 QCAZ40080Aplastodiscus_sp Cophomantinae_OG MTR26425 MTR26425Boana_boans Boana_OG AF1981 AF1981Boana_cinerascens Boana_OG TJC1100 TJC1100Boana_diabolica Boana_OG AF1827 AF1827Boana_ornatissima Boana_OG AF1912 AF1912Boana_pellucens Boana_OG WED53621 AY326058Boana_pulchella Boana_OG MACN37788 AY549352 KF794138 AY549405Boana_punctata Boana_OG AF3280 AF3280Boana_riojana Boana_OG MACN37507 AY549356Boana_xerophylla Boana_OG AF1797 AF1797Bokermannohyla_sp Cophomantinae_OG MTR26423 MTR26423Hyloscirtus_sp Cophomantinae_OG AF4400 AF4400Myersiohyla_inparquesi Cophomantinae_OG RWM17688 AY843672Nesorohyla_kanaima Cophomantinae_OG ROM39582 AY843634 GQ366307 AY843868
32 A Fouquet et al
PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Appendix 4 Detailed information onsound files analyzed for this studyLetters following numbers ofrecordings in AAG-UFU collection(eg 1andashe) indicate the number ofsound files for each recorded maleBoana courtoisaeSound file ndash MNHN-SO-2020-2943 Unvoucheredrecording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2944 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2946 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2945 Unvouchered
recording Recorded from Rorota French GuianaSound file ndash MNHN-SO-2020-2947 Unvouchered
recording Recorded from Sipaliwini SurinameSound file ndash MNHN-SO-2020-2948 Unvouchered
recording Recorded from Paletuviers French Guiana
Boana eucharisSound files ndash Boana_eucharisAltaFlorestaMT1andasheDLB_AAGm671 MNHN-SO-2020-2949ndash53 recordedon 11 January 2019 at 744 pm air 245 C voucherAAG-UFU 6503 (holotype) Recorded from AltaFloresta Mato Grosso BrazilSound files ndash Boana_eucharisAltaFlorestaMT2andash
cDLB_AAGm671 MNHN-SO-2020-2954ndash56 recordedon 11 January 2019 at 804 pm air 245 C VoucherAAG-UFU 6504 (paratype) Recorded from AltaFloresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT3andash
cDLB_AAGm671 MNHN-SO-2020-2957ndash59 recordedon 11 January 2019 at 810 pm air 245 CUnvouchered recording Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT4andashbDLB_
AAGm671 MNHN-SO-2020-2960ndash61 recorded on 11January 2019 at 905pm air 245 C Unvoucheredrecording Recorded from Alta Floresta Mato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT5andashiPM_
AAGm671 MNHN-SO-2020-2962ndash70 recorded on 20January 2020 at 802 pm air 251 C Voucher AAG-UFU 6901 (paratype) Recorded from Alta FlorestaMato Grosso BrazilSound file ndash Boana_eucharisAltaFlorestaMT6andashbAA
Gm671 MNHN-SO-2020-2971ndash72 recorded on 20January 2020 at 918 pm air 250 C Unvoucheredrecording Recorded from Alta Floresta MatoGrosso Brazil
Sound file ndash Boana_eucharisAltaFlorestaMT7andashfAAGm671 MNHN-SO-2020-2973ndash78 and MNHN-SO-2020-2980ndash81 recorded on 20 January 2020 at948 pm air 250 C Unvouchered recording Recordedfrom Alta Floresta Mato Grosso Brazil
Boana steinbachiSound file ndash MNHN-SO-2020-2934 Voucher SMF88394Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2935 Voucher SMF88397
Recorded from Buenavista Sara province BoliviaSound file ndash MNHN-SO-2020-2936 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2937 Unvouchered
recording Recorded from Rio Matos Beni BoliviaSound file ndash MNHN-SO-2020-2938 Unvouchered
recording Recorded from Tambopata Madre de Dios PeruSound file ndash 14-Hyla fasciata Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198328 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198676 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML198677 Unvouchered recording
Recorded from Tambopata Madre de Dios PeruSound file ndash ML222268 Unvouchered recording
Recorded from Cuzco Amazonico PeruSound file ndash Boana_steinbachiAssisBrasilAC1andashd
TRC_AAGm671 MNHN-SO-2020-2939ndash42 recorded on12 February 2017 at 800pm air 262 C Voucher AAG-UFU 5917 Recorded from Assis Brasil Acre BrazilSound file ndash FNJV12840 recorded on 4 December
1986 at 800 pm air 235 C Unvouchered recordingRecorded from Altamira Para BrazilSound file ndash FNJV12841 recorded on 7 December
1986 at 1000 pm air 235 C Unvouchered recordingRecorded from Altamira Para Brazil
Appendix 5 Details and justificationsfor the identification of these OTUsand their respectivegeographic rangesBoana raniceps (1 OTU)Boana raniceps is recovered as a single OTU distantlyrelated from the rest of the species and widelydistributed latitudinally from Argentina to FrenchGuiana and longitudinally from Bahia to centralAmazonia throughout the Cerrado and Chaco Thisdistribution encompasses the type locality Paraguay
345534563457345834593460346134623463346434653466346734683469347034713472347334743475347634773478347934803481348234833484348534863487348834893490349134923493349434953496349734983499350035013502350335043505350635073508
350935103511351235133514351535163517351835193520352135223523352435253526352735283529353035313532353335343535353635373538353935403541354235433544354535463547354835493550355135523553355435553556355735583559356035613562
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 33
PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
The Boana albopunctata clade (9 OTUs)Boana albopunctata is restricted to a small range in thestates of Sao Paulo and Minas Gerais encompassing itstype locality However additional data (Prado Haddadamp Zamudio 2012) indicate that the actual range of thisOTU extends at least to the states of Goias MatoGrosso do Sul and Parana However the otherpopulations previously identified as B albopunctatafurther north in the Cerrado and in Amazonia probablybelong to a distinct species as already suggested byPrado et al (2012)Boana leucocheila is tentatively assigned to one of
the OTU distributed in Mato Grosso Rondonia andBolivia This distribution encompasses the type locality(Caramaschi amp de Niemeyer 2003) and otherpopulations also reported in Mato Grosso (PansonatoAvila Kawashita-Ribeiro amp Morais 2011) Theexamination of specimens (MTR) corroborated thisidentificationBoana multifasciata was sampled at the type locality
in Belem Para state and in two localities (Carolina andEstreito) at the border between the states of Maranh~aoand Tocantins This species may be circumscribed to thenorthern part of the Cerrado and adjacent parts ofAmazonia The range of this species probably extendsfurther southward at least in northern CerradoHowever the Amazonian populations previouslyidentified as B multifasciata are suggested to belong toa distinct species (see hereafter)The populations previously identified as Boana
multifasciata (B aff albopunctata 3) from the GuianaShield and as Boana albopunctata from Belo MontePara (B aff albopunctata 2) Itaituba Para (B affalbopunctata 4) and Rondonia (B aff albopunctata 1)are suggested to belong to yet undescribed species Thetwo taxa (B paranaiba and B caiapo) that could not beincluded in our analysis are described from localities inthe Cerrado (see Carvalho Giaretta amp Facure 2010Pinheiro et al 2018) and are thus unlikely tocorrespond to these Amazonian OTUs Given thephenotypic differences observed among B leucocheilaB albopunctata and B multifasciata these OTUs mayalso be phenotypically distinct from each other(Caramaschi amp de Niemeyer 2003 Carvalho et al2010) Nevertheless we did not examine variationacross these populations because it was outside thescope of our study Therefore these OTUs should beconsidered as Unconfirmed Candidate Species (UCS)Boana lanciformis has a wide range in Western
Amazonia (Peru Ecuador Colombia and AmazonasState in Brazil) encompassing its type locality in theDistrict Pebas Region of Loreto Peru (incorrectlyascribed to Ecuador by Cope) However one population
from Tefe (Amazonas Brazil) in central Amazonia isfound genetically distinct and identified as a separateOTU We did not examine variation across thesepopulations thus this OTU will be considered as UCSInterestingly the range of these two OTUs does notoverlap with neither Boana raniceps nor any of theother OTU related to B albopunctata
The Boana calcarata clade (5 OTUs)Boana calcarata is recovered as a single OTU largelydistributed throughout Amazonia thus encompassing itstype locality in GuyanaBoana almendarizae and B fasciata display ranges
circumscribed to the slopes of the Andes of Ecuador(type localities in Morona Santiago and Zamora-Chinchipe provinces respectively) and northern Peru(Amazonas province) as already established by Caminerand Ron (2014) Similarly B maculateralis displays asmall range in Ecuador but in the lowlands Its actualrange extends at least to Meta (Acosta-Galvis et al2018) and Guaviare (Medina-Rangel Mendez-Galeanoamp Calderon-Espinosa 2019) in Colombia and probablyto adjacent Peru However the populations from Loretoin northern Peru previously documented by Caminerand Ron (2014) as distinct from B maculateraliscluster with other populations from Loreto in northernPeru and from Rio Ica (Amazonas) and Serra doDivisor (Acre) in north-western Brazil forming adistinct OTU (B aff maculateralis) Populations fromLeticia in south-eastern Colombia (Acosta-Galvis et al2018) Japura (Amazonas) in north-western Brazil(Rojas-Zamora et al 2017) and from Manu NationalPark in southern Peru (Caminer amp Ron 2014) probablydisplay this lineage as well We did not examinevariation across these populations thus this OTU willbe considered as UCS
The Boana steinbachi clade (9 OTUs)Boana dentei displays a small range in the easternmostpart of the Guiana Shield lowlands (Amapa and FrenchGuiana) encompassing its type locality (Serrado Navio)Boana alfaroi and B tetete both display small ranges
in Amazonian lowlands of Ecuador as documented byCaminer and Ron (2014) Moreover the range of Balfaroi extends further north to Putumayo (Meza-JoyaRamos-Pallares amp Hernandez-Jaimes 2019) andCaqueta (Medina-Rangel et al 2019) in ColombiaHowever the record from Acre provided by Carvalhoet al (2017) corresponds to B aff steinbachi 1 (seebelow) The range of B tetete extends to Loreto Peru
356335643565356635673568356935703571357235733574357535763577357835793580358135823583358435853586358735883589359035913592359335943595359635973598359936003601360236033604360536063607360836093610361136123613361436153616
361736183619362036213622362336243625362636273628362936303631363236333634363536363637363836393640364136423643364436453646364736483649365036513652365336543655365636573658365936603661366236633664366536663667366836693670
34 A Fouquet et al
PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
confirming records from Acosta-Galvis et al (2018)Nevertheless their record from Leticia most likelycorresponds to either B alfaroi or B aff steinbachi 1Boana steinbachi occurs from Bolivia (type locality)
to south-eastern Peru (Tambopata Madre de Dios) andmore surprisingly in eastern Brazilian Amazonia (Para)This extensive range with a wide gap between Boliviaand Para and the fact that acoustic data associated withvoucher specimens were not available from the Parapopulations did not allow us to confirm the taxonomicidentity of these populations as conspecific withnominal B steinbachi (see below) Another group ofrelated populations extends widely in WesternAmazonia of Peru Colombia and Brazil forming adistinct OTU (B aff steinbachi 1) The populationspreviously identified as Boana alfaroi (Carvalho BangTeixeira amp Giaretta 2017) belong to this OTU butsince we did not find any phenotypic diagnosis amongthe OTUs within this clade we treated them asconspecific with B steinbachi Additional populationsfrom southern and eastern Peru (lower Madre de DiosRiver Madre de Dios and Cocama Ucayali) areidentified as a distinct small-range OTU (B affsteinbachi 2) We examined the type series of Boanasteinbachi examined the morphology and analysed callsof a series of topotypes and provided an amendeddiagnosis of the nominal species
Populations from Rondonia (Pacaas Novos and Jirau)and Mato Grosso (Apiacas Juruena and Alta Floresta)form a distinct small-range lineage identified as adistinct OTU (Boana sp 1) These populations were sofar confused with B fasciata (Avila amp Kawashita-Ribeiro 2011 Lima Keller amp Rebelo 2017) Weassessed the phylogenetic relationships andmorphological and acoustic characters of Boana sp 1which led us to describe it herein as a new species(see below)Populations from the Eastern Guiana Shield form two
additional OTUs Boana sp 2 which extends throughoutthe region and Boana sp 3 which is circumscribed tocoastal French Guiana These populations werepreviously identified as Boana fasciata (Fouquet et al2007 Lescure amp Marty 2000 Dewynter et al 2008)and subsequently suggested to belong to an unnamedspecies (Funk et al 2012 Caminer amp Ron 2014 ColeTownsend Reynolds MacCulloch amp Lathrop 2013Fouquet Vidal amp Dewynter 2019) We examinedmorphological and acoustic characters of specimens ofBoana sp 2 and described it herein as a new species(see below) However we tentatively consider Bana sp3 as conspecific with Boana sp 2 since no clearphenotypic differentiation was found across GuianaShield populations
367136723673367436753676367736783679368036813682368336843685368636873688368936903691369236933694369536963697369836993700370137023703370437053706370737083709371037113712371337143715371637173718371937203721372237233724
372537263727372837293730373137323733373437353736373737383739374037413742374337443745374637473748374937503751375237533754375537563757375837593760376137623763376437653766376737683769377037713772377337743775377637773778
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 35
antoinefouquet
Texte inseacutereacute
Aacute
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-
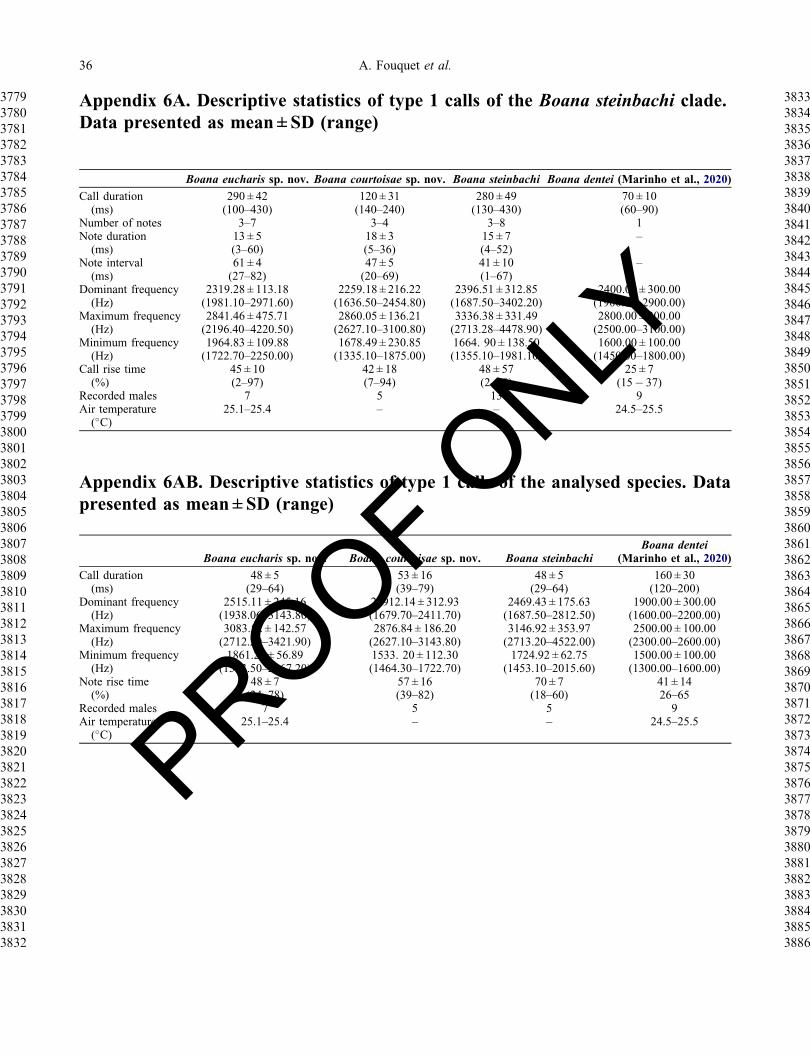
PROOF ONLY
377937803781378237833784378537863787378837893790379137923793379437953796379737983799380038013802380338043805380638073808380938103811381238133814381538163817381838193820382138223823382438253826382738283829383038313832
383338343835383638373838383938403841384238433844384538463847384838493850385138523853385438553856385738583859386038613862386338643865386638673868386938703871387238733874387538763877387838793880388138823883388438853886
Appendix 6A Descriptive statistics of type 1 calls of the Boana steinbachi cladeData presented as meanplusmn SD (range)
Appendix 6AB Descriptive statistics of type 1 calls of the analysed species Datapresented as meanplusmn SD (range)
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachi Boana dentei (Marinho et al 2020)
Call duration(ms)
290 plusmn 42(100ndash430)
120 plusmn 31(140ndash240)
280 plusmn 49(130ndash430)
70 plusmn 10(60ndash90)
Number of notes 3ndash7 3ndash4 3ndash8 1Note duration
(ms)13 plusmn 5(3ndash60)
18 plusmn 3(5ndash36)
15 plusmn 7(4ndash52)
ndash
Note interval(ms)
61 plusmn 4(27ndash82)
47 plusmn 5(20ndash69)
41 plusmn 10(1ndash67)
ndash
Dominant frequency(Hz)
231928 plusmn 11318(198110ndash297160)
225918 plusmn 21622(163650ndash245480)
239651 plusmn 31285(168750ndash340220)
240000 plusmn 30000(190000ndash290000)
Maximum frequency(Hz)
284146 plusmn 47571(219640ndash422050)
286005 plusmn 13621(262710ndash310080)
333638 plusmn 33149(271328ndash447890)
280000 plusmn 20000(250000ndash310000)
Minimum frequency(Hz)
196483 plusmn 10988(172270ndash225000)
167849 plusmn 23085(133510ndash187500)
1664 90 plusmn 13850(135510ndash198110)
160000 plusmn 10000(145000ndash180000)
Call rise time()
45 plusmn 10(2ndash97)
42 plusmn 18(7ndash94)
48 plusmn 57(2ndash98)
25 plusmn 7(15 37)
Recorded males 7 5 13 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
Boana eucharis sp nov Boana courtoisae sp nov Boana steinbachiBoana dentei
(Marinho et al 2020)
Call duration(ms)
48 plusmn 5(29ndash64)
53 plusmn 16(39ndash79)
48 plusmn 5(29ndash64)
160 plusmn 30(120ndash200)
Dominant frequency(Hz)
251511 plusmn 24516(193806ndash314380)
2191214 plusmn 31293(167970ndash241170)
246943 plusmn 17563(168750ndash281250)
190000 plusmn 30000(160000ndash220000)
Maximum frequency(Hz)
308352 plusmn 14257(271220ndash342190)
287684 plusmn 18620(262710ndash314380)
314692 plusmn 35397(271320ndash452200)
250000 plusmn 10000(230000ndash260000)
Minimum frequency(Hz)
186125 plusmn 5689(159350ndash206720)
1533 20 plusmn 11230(146430ndash172270)
172492 plusmn 6275(145310ndash201560)
150000 plusmn 10000(130000ndash160000)
Note rise time()
48 plusmn 7(24ndash78)
57 plusmn 16(39ndash82)
70 plusmn 7(18ndash60)
41 plusmn 1426ndash65
Recorded males 7 5 5 9Air temperature
(C)251ndash254 ndash ndash 245ndash255
36 A Fouquet et al
antoinefouquet
Barrer
PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
388738883889389038913892389338943895389638973898389939003901390239033904390539063907390839093910391139123913391439153916391739183919392039213922392339243925392639273928392939303931393239333934393539363937393839393940
394139423943394439453946394739483949395039513952395339543955395639573958395939603961396239633964396539663967396839693970397139723973397439753976397739783979398039813982398339843985398639873988398939903991399239933994
Appendix 7 Descriptive statistics of morphometric traits (in mm) fromspecimens of the Boana steinbachi clade topotypes and additional specimens ofB steinbachi and the type series of B eucharis sp nov and B courtoisae spnov Data presented as meanplusmn SD (range) Abbreviations are SVL5 snout-ventlength FOOT5 foot length HL5 head length HW5 head width ED5 eyediameter TD5 tympanum diameter TL5 tibia length FL5 femur lengthCL5 calcaneal appendage length sample sizes for CL are as follows Bsteinbachi (N 5 6 males) B eucharis sp nov (N 5 7 males) and B courtoisaesp nov (all specimens were measured)
Appendix 8 Additional Boana courtoisae sp nov specimensMalesAF1925 AF1934 French Guiana Flat de la WakiAF2319 French Guiana TriniteAF3059 AF3070 French Guiana MitanAF3694 French Guiana ItoupeAM008 French Guiana Inini TolengaAF2115 AF2195 Suriname SipaliwiniAF3452 Suriname Spari CreekAF3775 AF3784 Suriname Voltzberg CI camp
FemalesAF2584 French Guiana SinnamaryAF1694 French Guiana Saul ndash LimonadeAF1544 French Guiana Saul ndash Gros arbre
Boana steinbachiBoana eucharis sp nov
Boana courtoisae sp nov
MalesN5 18
FemalesN5 6
MalesN5 7
MalesN5 13
FemalesN5 3
SVL 336 plusmn 16(304ndash374)
449 plusmn 21(425ndash488)
327 plusmn 12(308ndash347)
331 plusmn 19(308ndash359)
442 plusmn 15(430ndash459)
FL 140 plusmn 14(114ndash166)
180 plusmn 08(166ndash188)
133 plusmn 04(127ndash137)
136 plusmn 11(118ndash155)
190 plusmn 05(186ndash195)
HL 115 plusmn 21(79ndash134)
109 plusmn 03(105ndash114)
121 plusmn 07(107-129)
121 plusmn 08(108ndash140)
155 plusmn 06(150ndash161)
HW 108 plusmn 08(92ndash121)
150 plusmn 04(144ndash155)
107 plusmn 05(101ndash118)
113 plusmn 10(100ndash129)
145 plusmn 10(136ndash156)
ED 40 plusmn 04(32ndash49)
43 plusmn 04(39ndash51)
42 plusmn 02(38ndash47)
47 plusmn 04(40ndash54)
51 plusmn 06(44ndash55)
TD 21 plusmn 03(13ndash25)
25 plusmn 05(19ndash29)
20 plusmn 02(16ndash22)
21 plusmn 02(18ndash23)
24 plusmn 000
TL 187 plusmn 21(142ndash217)
226 plusmn 14(205ndash243)
184 plusmn 07(171ndash196)
191 plusmn 11(177ndash209)
261 plusmn 06(256ndash267)
THL 188 plusmn 19(148ndash211)
254 plusmn 09(244ndash271)
167 plusmn 14(149ndash186)
175 plusmn 12(158ndash195)
237 plusmn 15(221ndash251)
CAL 05 plusmn 01(03ndash06)
mdash 02 plusmn 00(01ndash02)
05 plusmn 01(03ndash06)
07 plusmn 01(06ndash07)
Systematics and biogeography of the Boana albopunctata species group (Anura Hylidae) 37
PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-

PROOF ONLY
Appendix 9 BioGeoBears models399539963997399839994000400140024003400440054006400740084009401040114012401340144015401640174018401940204021402240234024402540264027402840294030403140324033403440354036403740384039404040414042404340444045404640474048
404940504051405240534054405540564057405840594060406140624063406440654066406740684069407040714072407340744075407640774078407940804081408240834084408540864087408840894090409140924093409440954096409740984099410041014102
Models LnL d e j AIC DAIC
DEC1 J -46445 689E-03 100E-12 0040 9889 0DIVAlikethorn J 46798 959E-03 200E-09 0033 996 071BAYAREAthorn J 50002 696E-3 100E-12 0047 106 711DEC 5278 0015 100E-12 0 1096 1071DIVAlike 55922 0014 100E-12 0 1158 1691BAYAREA 61097 0014 0147 0 1262 2731
38 A Fouquet et al
- Abstract
-
- Introduction
- Materials and methods
-
- Species delimitation
- Morphological and acoustic variation in the Boana steinbachi clade
- Time-calibrated species phylogeny
- Biogeographic analysis
-
- Results
-
- Species delimitation
- Taxonomic accounts
-
- mkchapTSAB_S0003-S2002-S3001_sec
-
- Syntypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Variation
- Vocal repertoire
- Distribution and ecology
- Holotype
- Paratopotypes
- Paratypes
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colours of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
- Holotype
- Paratopotypes
- Paratypes
- Other material
- Diagnosis
- Comparisons with congeners of the Amazonian clade
- Description of holotype
- Colour of holotype
- Variation
- Vocal repertoire
- Distribution and ecology
- Etymology
-
- Mitogenomic phylogeny
- Biogeographic inferences
-
- Discussion
-
- Species richness and distribution
- Biogeography
-
- Acknowledgements
- Supplemental data
- Orcid
- References
- thinsp
-
- Appendix 2 Molecular data acquisition
- Mitogenome sequencing assembling and annotation
-
- Supplementary references
-
- Boana courtoisae
- Boana eucharis
- Boana steinbachi
- Boana raniceps (1 OTU)
- The Boana albopunctata clade (9 OTUs)
- The Boana calcarata clade (5 OTUs)
- The Boana steinbachi clade (9 OTUs)
-




















