
Anatomical aspects of angiosperm root evolution
16
REVIEW: PART OF A SPECIAL ISSUE ON MATCHING ROOTS TO THEIR ENVIRONMENT Anatomical aspects of angiosperm root evolution James L. Seago Jr 1, * and Danilo D. Fernando 2 1 Department of Biological Sciences, SUNY at Oswego, Oswego, NY 13126, USA and 2 Department of Environmental and Forest Biology, SUNY College of Environmental Science and Forestry, Syracuse, NY 13210, USA * For correspondence. E-mail [email protected] Received: 29 August 2012 Revision requested: 9 October 2012 Accepted: 9 November 2012 Published electronically: 7 January 2013 † Background and Aims Anatomy had been one of the foundations in our understanding of plant evolutionary trends and, although recent evo-devo concepts are mostly based on molecular genetics, classical structural infor- mation remains useful as ever. Of the various plant organs, the roots have been the least studied, primarily because of the difficulty in obtaining materials, particularly from large woody species. Therefore, this review aims to provide an overview of the information that has accumulated on the anatomy of angiosperm roots and to present possible evolutionary trends between representatives of the major angiosperm clades. † Scope This review covers an overview of the various aspects of the evolutionary origin of the root. The results and discussion focus on angiosperm root anatomy and evolution covering representatives from basal angios- perms, magnoliids, monocots and eudicots. We use information from the literature as well as new data from our own research. † Key Findings The organization of the root apical meristem (RAM) of Nymphaeales allows for the ground meri- stem and protoderm to be derived from the same group of initials, similar to those of the monocots, whereas in Amborellales, magnoliids and eudicots, it is their protoderm and lateral rootcap which are derived from the same group of initials. Most members of Nymphaeales are similar to monocots in having ephemeral primary roots and so adventitious roots predominate, whereas Amborellales, Austrobaileyales, magnoliids and eudicots are gener- ally characterized by having primary roots that give rise to a taproot system. Nymphaeales and monocots often have polyarch (heptarch or more) steles, whereas the rest of the basal angiosperms, magnoliids and eudicots usually have diarch to hexarch steles. † Conclusions Angiosperms exhibit highly varied structural patterns in RAM organization; cortex, epidermis and rootcap origins; and stele patterns. Generally, however, Amborellales, magnoliids and, possibly, Austrobaileyales are more similar to eudicots, and the Nymphaeales are strongly structurally associated with the monocots, espe- cially the Acorales. Key words: Anatomy, angiosperms, cortex, epidermis, evolution, roots, vascular tissue. INTRODUCTION Evolutionary origin of the root A root is a highly differentiated multicellular axis found only in the sporophytes of vascular plants that typically has a rootcap, endodermis, pericycle and lateral roots. It is the main organ in vascular plants that anchors the plant body to its substrate and absorbs water and dissolved minerals to support growth and development. The free-living gameto- phytes of bryophytes, lycophytes and monilophytes grow on moist environments, and anchorage is accomplished by a system of unicellular or undifferentiated multicellular rhizoids. Evolutionarily, the root seemed to be the last of the three main vegetative organs to evolve, perhaps since early land plants grew on or near the water and so much of their early innova- tions were geared toward maximizing photosynthesis through development of stems and leaves. Vascular plants evolved at least during the Silurian based on the oldest known macro- fossil – Cooksonia (Lang, 1937; Edwards et al., 1992), but there is no record of specialized root axes in this Period (Gensel et al., 2001; Raven and Edwards, 2001). In the Early Devonian, several early land plants already had well developed stems and leaves, but only structures with consider- able similarity to roots are known, such as those of Asteroxylon mackiei (Rayner, 1984; Li and Edwards, 1995; Gensel et al., 2001; Bennett and Scheres, 2010). Although no rootcap was observed from the rooting structures of lycophytes during the Early Devonian, they had sub-terranean parenchymatous axes which performed the functions of roots and so these plants have been considered to have possessed roots (see Raven and Edwards, 2001). Bennett and Scheres (2010) suggested that these horizontal axes later evolved rootcap meristems, and so rootcap formation was a separate innovation that allowed pene- trative growth into the soil. There is no corresponding evidence for root-like structures for the euphyllophytes (monilophytes and seed plants) during the Early Devonian, and the earliest convincing evidence of root in this group is from Lorophyton goense, a Middle Devonian fern-like (cladoxylalean) plant (Fairon-Demaret and Li, 1993). Roots of extant lycophytes and euphyllophytes possess a rootcap that is derived from and functions to protect the root apical meristem (RAM), and root hairs are present on the epidermis of all major vascular plants (Jones and Dolan, 2012). Their endodermis ensures a one-way mode of water transport into the plant, while the peri- cycle is usually where the lateral roots originate. # The Author 2013. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: [email protected] Annals of Botany 112: 223–238, 2013 doi:10.1093/aob/mcs266, available online at www.aob.oxfordjournals.org Downloaded from https://academic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022
-
Upload
khangminh22 -
Category
Documents
-
view
8 -
download
0
Transcript of Anatomical aspects of angiosperm root evolution

REVIEW: PART OF A SPECIAL ISSUE ON MATCHING ROOTS TO THEIR ENVIRONMENT
Anatomical aspects of angiosperm root evolution
James L. Seago Jr1,* and Danilo D. Fernando2
1Department of Biological Sciences, SUNY at Oswego, Oswego, NY 13126, USA and 2Department of Environmentaland Forest Biology, SUNY College of Environmental Science and Forestry, Syracuse, NY 13210, USA
* For correspondence. E-mail [email protected]
Received: 29 August 2012 Revision requested: 9 October 2012 Accepted: 9 November 2012 Published electronically: 7 January 2013
† Background and Aims Anatomy had been one of the foundations in our understanding of plant evolutionarytrends and, although recent evo-devo concepts are mostly based on molecular genetics, classical structural infor-mation remains useful as ever. Of the various plant organs, the roots have been the least studied, primarilybecause of the difficulty in obtaining materials, particularly from large woody species. Therefore, this reviewaims to provide an overview of the information that has accumulated on the anatomy of angiosperm roots andto present possible evolutionary trends between representatives of the major angiosperm clades.† Scope This review covers an overview of the various aspects of the evolutionary origin of the root. The resultsand discussion focus on angiosperm root anatomy and evolution covering representatives from basal angios-perms, magnoliids, monocots and eudicots. We use information from the literature as well as new data fromour own research.† Key Findings The organization of the root apical meristem (RAM) of Nymphaeales allows for the ground meri-stem and protoderm to be derived from the same group of initials, similar to those of the monocots, whereas inAmborellales, magnoliids and eudicots, it is their protoderm and lateral rootcap which are derived from the samegroup of initials. Most members of Nymphaeales are similar to monocots in having ephemeral primary roots andso adventitious roots predominate, whereas Amborellales, Austrobaileyales, magnoliids and eudicots are gener-ally characterized by having primary roots that give rise to a taproot system. Nymphaeales and monocots oftenhave polyarch (heptarch or more) steles, whereas the rest of the basal angiosperms, magnoliids and eudicotsusually have diarch to hexarch steles.† Conclusions Angiosperms exhibit highly varied structural patterns in RAM organization; cortex, epidermis androotcap origins; and stele patterns. Generally, however, Amborellales, magnoliids and, possibly, Austrobaileyalesare more similar to eudicots, and the Nymphaeales are strongly structurally associated with the monocots, espe-cially the Acorales.
Key words: Anatomy, angiosperms, cortex, epidermis, evolution, roots, vascular tissue.
INTRODUCTION
Evolutionary origin of the root
A root is a highly differentiated multicellular axis found onlyin the sporophytes of vascular plants that typically has arootcap, endodermis, pericycle and lateral roots. It is themain organ in vascular plants that anchors the plant body toits substrate and absorbs water and dissolved minerals tosupport growth and development. The free-living gameto-phytes of bryophytes, lycophytes and monilophytes grow onmoist environments, and anchorage is accomplished by asystem of unicellular or undifferentiated multicellular rhizoids.Evolutionarily, the root seemed to be the last of the three mainvegetative organs to evolve, perhaps since early land plantsgrew on or near the water and so much of their early innova-tions were geared toward maximizing photosynthesis throughdevelopment of stems and leaves. Vascular plants evolved atleast during the Silurian based on the oldest known macro-fossil – Cooksonia (Lang, 1937; Edwards et al., 1992), butthere is no record of specialized root axes in this Period(Gensel et al., 2001; Raven and Edwards, 2001). In theEarly Devonian, several early land plants already had well
developed stems and leaves, but only structures with consider-able similarity to roots are known, such as those of Asteroxylonmackiei (Rayner, 1984; Li and Edwards, 1995; Gensel et al.,2001; Bennett and Scheres, 2010). Although no rootcap wasobserved from the rooting structures of lycophytes during theEarly Devonian, they had sub-terranean parenchymatous axeswhich performed the functions of roots and so these plantshave been considered to have possessed roots (see Raven andEdwards, 2001). Bennett and Scheres (2010) suggested thatthese horizontal axes later evolved rootcap meristems, and sorootcap formation was a separate innovation that allowed pene-trative growth into the soil. There is no corresponding evidencefor root-like structures for the euphyllophytes (monilophytesand seed plants) during the Early Devonian, and the earliestconvincing evidence of root in this group is from Lorophytongoense, a Middle Devonian fern-like (cladoxylalean) plant(Fairon-Demaret and Li, 1993). Roots of extant lycophytesand euphyllophytes possess a rootcap that is derived fromand functions to protect the root apical meristem (RAM), androot hairs are present on the epidermis of all major vascularplants (Jones and Dolan, 2012). Their endodermis ensures aone-way mode of water transport into the plant, while the peri-cycle is usually where the lateral roots originate.
# The Author 2013. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved.
For Permissions, please email: [email protected]
Annals of Botany 112: 223–238, 2013
doi:10.1093/aob/mcs266, available online at www.aob.oxfordjournals.org
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

It has been hypothesized that roots evolved at least twice,once in the lycophytes and another in the euphyllophytes(Bierhorst, 1971; Kenrick and Crane, 1997; Gensel et al.,2001; Raven and Edwards, 2001; Boyce and Knoll, 2002;Jones and Dolan, 2012; Pires and Dolan, 2012). Phylogeneticanalysis that integrated key fossil taxa with extant lineagesshows that the roots of lycophytes and euphyllophytes are analo-gous (Fig. 1) (Friedman et al., 2004). This suggests that thecommon ancestors of both lycophytes and euphyllophyteslack roots, and so the possession of roots must be the result ofexhibiting the same developmental patterns in response to asimilar selective pressure (Friedman et al., 2004). Therefore,there is strong support for the independent origin of roots inland plants. However, it is still not clear how many timesroots evolved within the lycophytes and euphyllophytes. Inplants with pyramidal root apical cells [e.g. monilophytes andsome lycophytes (Selaginella)], lateral roots arise from theendodermis, whereas in plants with one or more superimposedinitials [e.g. seed plants and some lycophytes (Lycopodium)],lateral roots originate from the pericycle. The orientation andorigin of the embryonic root axis relative to the shoot axis aredifferent in monilophytes and seed plants (Gensel et al., 2001;Raven and Edwards, 2001). Specifically, the embryonic root
subtends the embryonic leaf in ferns (Cooke et al., 2004) and,although the root is formed on the basal pole of the embryoin angiosperms, it is derived from the hypophysis in eudicotsand ground meristem in monocots (Bennett and Scheres,2010). All these suggest multiple origins of the roots and there-fore warrant further investigations.
Developmental origin of the root
The early vascular plants were composed of dichotomouslybranched parenchymatous axes or telomes based from theRhynie Chert fossils. In the Zosterophyllophyta of the EarlyDevonian, most of the branches were erect and functioned asthe main organ for photosynthesis (shoot-like), whereas otherbranches were oriented horizontally and rested on the surfaceof the substrate. Some of the horizontal axes were directeddownwards and were root-like in appearance. Based on this,the roots of extant lycophytes and euphyllophytes probablyarose from an original dichotomizing axis, particularly fromthe horizontal branches that penetrated the substrate, anchoredthe plant body, and absorbed water and dissolved minerals.Thus, roots and shoots are considered homologous sincethey are both believed to have evolved from the same
Embryophytes
Polysporangiophytes
Tracheophytes
Euphyllophytes
Lignophytes
Seed plantsMoniliformopsesLycopsida
Liverworts
Cha
rale
s
Jung
enm
anni
ales
Sim
ple
thal
loid
Com
plex
thal
loid
Hap
lom
itria
les
Mos
ses
Hor
nwor
ts
Agl
aoph
yton
Rhy
niop
sida
Zos
tero
phyl
lum
Lyco
podi
ales
Sel
agin
ella
les
Isoe
tale
s
Psi
loph
yton
Fili
cale
s
Sph
enop
hyta
Mar
attia
les
Psi
lota
les
Elk
insi
a
Ang
iosp
erm
s
Cyc
adal
es
Gin
kgoa
les
Con
ifers
Gne
tale
s
Oph
iogl
ossa
les
Ane
urop
hyta
les
Arc
haeo
pter
idal
es
Lycophytes
Roots
Absent
Root-like structure
Roots
FI G. 1. Key fossil taxa are integrated into a phylogeny for analysis of root evolution which shows two evolutionary origins of roots. One origin is in the ly-copsida, perhaps from telomic systems that grow into substrate, as found in the Zosterophyllum. Another independent origin is observed in the common ancestor
of extant euphyllophytes. Illustration from Friedman et al. (2004, p. 1728), with permission from the American Journal of Botany.
Seago & Fernando — Anatomical aspects of angiosperm root evolution224
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

dichotomously branched axes (Gensel et al., 2001; Friedman,2004; Bennett and Scheres, 2010).
Molecular, developmental and genetic analyses are congru-ent with the fact that, at least in angiosperms, roots and shootsare derived from the same dichotomizing axes (Friedman,2004). In Arabidopsis thaliana, these two organs share manydevelopmental processes in common such as the: (1) formationof the endodermis which both require the expression ofSCARECROW and SHORTROOT genes (Pysh et al., 1999;Nakajima et al., 2001); (2) regulation of radial patterning ofthe ground tissues during and post-embryo formation (Fukakiet al., 1998; Wysocka-Diller et al., 2000); (3) specificationof epidermal cell fate, e.g. GLABRA2 in conjunction withWEREWOLF genes (in roots) and its functionally redundantparalogue GLABRA1 (in shoots) are required for the differen-tiation between hair- and non-hair-bearing cells in roots andshoots (Dolan and Scheres, 1998; Schiefelbein, 2003; Bruexet al., 2012); (4) provascular cell fate commitment(Scarpella et al., 2000); and (5) similar mutations affectingboth the RAM and the shoot apical meristem (SAM) (Uedaet al., 2004).
It is becoming clear that the production and maintenance ofstem cells in the RAM and SAM involve the same develop-mental patterns (Mayer et al., 1998; Haecker et al., 2004)and expression of the same genes or class of genes includingSCARECROW (Wysocka-Diller et al., 2000), WUSCHELRELATED HOMEOBOX (Haecker et al., 2004), HALTEDROOT (Ueda et al., 2004) and the CLAVATA3/ENDOSPERMSURROUNDING REGION-related gene family (Miwa et al.,2009). The stem cells in the RAM and SAM are located inregions referred to as the quiescent centre and organizingcentre, respectively, which are the signalling centres thatmake up the stem cell maintaining microenvironments inboth meristems. These two regions are considered to be func-tionally equivalent (Baurle and Laux, 2003; Byrne et al.,2003; Haecker et al., 2004; Veit, 2004; Bennett and Scheres,2010). There are many similarities in the organizations ofthe RAM and SAM, and the signalling components requiredfor stem cell initiation and maintenance seem to be relativelyconserved. This suggests that similar genes or classes ofgenes have been co-opted for use in both types of meristems(Friedman et al., 2004; Bennett and Scheres, 2010). Bennettand Scheres (2010) have proposed a mechanism for how theRAM and SAM of monilophytes and seed plants descendedfrom the ancestral generic meristem of a dichotomizing axis.
Origin of genes involved in root development
The genome sequence of the model bryophytePhyscomitrella patens has provided the opportunity to probethe genes, gene expression patterns and development asso-ciated with the gametophyte phase of the plant life cycle. Italso has allowed for the comparison of sequences and analysisof evolutionary trends across phyla and between the gameto-phyte and sporophyte generations of the plant life cycle.Molecular genetic analysis of root and shoot developmentdoes not only suggests homology of roots and shoots, butalso that genes involved in the formation of gametophyte-related structures in ancestral plants were co-opted toperform regulatory roles in the formation of sporophyte-related
structures in more evolutionarily derived plants. This is exem-plified by the ROOT HAIR DEFECTIVE 6 (RHD6) andRHD6-LIKE 1 (RSL1) genes that control the differentiationof roots hairs in A. thaliana, and two similar genes (PpRSL1and PpRSL2) that are involved in rhizoid development inP. patens (Menand et al., 2007). Rhizoids and root hairsboth elongate by tip growth and fulfil similar functions(Jones and Dolan, 2012). However, rhizoids are gametophyticfilamentous structures from the gametophytes, whereas roothairs are sporophytic tubular projections from root epidermis.It appears that genes that promoted the development of cellswith rooting functions in the gametophytes of bryophyteswere co-opted in the development of the sporophytes of vascu-lar plants or their ancestors, perhaps through gene duplicationand sub-functualization. Once expressed in the sporophyte,they promoted the development of hairs on roots. It is also pos-sible that changes in the cis-regulatory regions of RSL1 genesplayed a role in altering their expression patterns, i.e. promo-tion of their transcription in the sporophytes and repressionof their transcription in the gametophytes (Jones and Dolan,2012). The elaboration of the sporophyte generation of the vas-cular plants and particularly the large radiation of morpho-logical forms that occurred during the Devonian may havebeen achieved in part through the recruitment of genes orgenetic networks that had previously evolved and functionedin the gametophyte generation of early land plants or theirancestors. The demonstration that similar genes control the de-velopment of cells with a rooting function suggests that generecruitment is an ancient mechanism (Edwards and Feehan,1980; Menand et al., 2007). Therefore, RHD6 and RSL1genes control the development of cells with rooting functionsin bryophytes and angiosperms. However, it remains to be seenif these genes will exhibit the same function in lycophytes,monilophytes and gymnosperms. Also, it is likely that thereare other genes involved in the development of roots andother structures in the sporophyte that were recruited fromthe gametophyte. Therefore, investigations into the moleculargenetics of root structure and development in basal landplants will address the questions regarding homology ofroots and shoots and multiple origins of the roots. Our under-standing of the diversity of root anatomical structures will alsohelp in formulating or substantiating possible evolutionaryrelationships.
Angiosperm root anatomy and evolution
Much work has been done on the anatomy and developmentof roots of extant angiosperms, particularly from model herb-aceous species such as A. thaliana and Oryza sativa, and otherspecies of economic importance such as Zea mays and Glycinemax, plants that are considered to be more evolutionarilyderived. To bridge the gap in our understanding of root struc-tures, development and evolution, a review of our knowledgeof this subject that includes the major clades of the angios-perms is necessary. Therefore, this article will cover root struc-tures from plants representing the basal angiosperm lineages,such as Amborellales, Nymphaeales and Austrobaileyales(Heimsch and Seago, 2008), and representative magnoliids,monocots and eudicots. Data will be derived from the litera-ture, including our articles, and from some new research,
Seago & Fernando — Anatomical aspects of angiosperm root evolution 225
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

with methods, including brightfield and epifluorescence photo-microscopy, from Seago et al. (1999b, 2005), Seago (2002),Soukup et al. (2005) and Heimsch and Seago (2008).
Formation of below-ground axes from rhizomes into rootshas been considered with regard to various ecological andfunctional aspects of evolutionary phenomena (e.g. Cairney,2000; Raven and Edwards, 2001; Brundrett, 2002; Sperry,2003; Jackson et al., 2009; Pires and Dolan, 2012).However, more generally with regard to angiosperms,researchers have examined structural features such as the tran-sition from tracheids to vessel elements and specific xylemcells associated with water/mineral conduction (Carlquist,1975; Carlquist and Schneider, 2001, 2002, 2009; Schneiderand Carlquist, 2002), RAMs (Philipson, 1990; Barlow, 1994,2002; Clowes, 2000; Groot and Rost, 2001; Groot et al.,2004; Heimsch and Seago, 2008) and, to a lesser degree, thecortical derivatives of the RAM (Shishkoff, 1987; Seagoet al., 2005).
There are generally two very different developmental andstructural aspects to angiosperm root systems. The primaryroot system is derived from the radicle and tends to be domin-ant in eudicots, and gives rise to lateral roots with variousdegrees of branching. In monocots, the primary root is oftenephemeral, and so adventitious roots (derived usually fromstems and leaves) and seminal roots (derived from mesocotyl)comprise their root systems where they also produce lateralroots (Bell and Bryan, 2008, p. 126; for nature of seminalroots, cf. Weaver, 1926; Hayward, 1938). The ephemeralnature of primary roots and dominance of adventitious rootsin monocots present a difficulty in comparisons, especiallysince adventitious roots (even from secondary growth) alsooccur in many eudicots. However, it seems quite likely thatat least some groups of basal angiosperms were derived fromancestors whose plant structure was mainly rhizomatousprior to flower evolution. Nevertheless, besides the differencein the origin of primary and adventitious roots, there is gener-ally no difference in the anatomy of primary and adventitiousroots, primary roots and their lateral roots, and adventitiousroots and their lateral roots in angiosperms. However, as willbe realized in this review, the differences lie in the types orgroups of plants and their environments.
DATA FROM THE LITERATUREAND NEW RESULTS
Amborellales
This order is represented by Amborella trichopoda, the onlymember of the family Amborellaceae. It is an understoreyshrub or small tree on moist soils or near streams, and isendemic to New Caledonia. Adventitious and lateral rootsare characterized by a diarch stele, but triarch steles are alsopresent; secondary xylem has tracheids but no vessels(Carlquist and Schneider, 2001). The cortex is delimited tothe inside by an endodermis with distinct Casparian bands(CBs) and suberin lamellae (SL; Fig. 2A, B) with passagecells (Fig. 2C). The middle cortex is occupied by non-radiallyarranged parenchyma cells (Fig. 2B–D; Heimsch and Seago,2008), and there is no aerenchyma. Secondary xylem is ac-companied by more cells of the endodermis developing SL
(cf. Fig. 2A–C); endodermal cells dilatate or increase innumber as some cells of the adjacent cortex layer thickenthrough addition of wall material that is fluorescent (cf.Fig. 2C, D), but the endodermis does not undergo this kindof dilatation as in some eudicots (Lux and Luxova, 2003;Sottnıkova and Lux, 2003). An irregular, multilayered hypo-dermis with an exodermis with CBs and SL underlies the epi-dermis (Fig. 2B–D). Both endodermis and exodermis remainbefore pericyclic derivatives initiate periderm production.Amborella RAMs are eudicot-like (Heimsch and Seago, 2008).
Nymphaeales
The Nymphaeales are currently and historically aquatic (Lesand Schneider, 1995; Friis et al., 1999, 2001, 2009). Roots ofNymphaeaceae and Cabombaceae are always adventitious inwater and have been previously characterized (Seago, 2002;Seago et al., 2000b, 2005; Carlquist and Schneider, 2002,2009). The salient traits are pentarch to polyarch stele inNymphaeaceae (Fig. 3D, E) and often monarch stele in the ex-tremely small roots of the Cabombaceae (Fig. 3C, two xylemcells) and Hydatellaceae (Fig. 3A; cf. Arabidopsis of eudicots,Baum et al., 2002). However, Conard (1905) described diarchyin Nymphaea species, but such a condition has not been con-firmed because of the ephemeral nature of the primary roots.Distinctive expansigenous aerenchyma is present in theNymphaeaceae and Cabombaceae. An endodermis with CBsand sometimes a small amount of SL, prominent astrosclereidsof the Nymphaeaceae and their absence in other Nymphaeales,and uniseriate exodermis with CBs and SL (Fig. 3D, E; Seagoet al., 2005) are also salient features of the Nymphaeales.
Trithuria filamentosa, a representative of the familyHydatellaceae, usually has one central tracheid surroundedby phloem (Fig. 3A) and pericycle (our results; Rudall et al.,2007), but we have also observed two tracheids in the stele;xylem of T. filamentosa lacks vessels, unlike theNymphaeaceae and Cabombaceae (Carlquist and Schneider,2009). We show here clearly that T. filamentosa has a cortexwith an endodermis with CBs in anticlinal walls, and, likeCabomba, the SL are more prominent on the outer tangentialwalls (note: root endodermis is continuous with the stem endo-dermis). In the Hydatellaceae, the cortical aerenchyma isderived through schizogeny, followed by expansigeny(Fig. 3B, right insert). There is an exodermis with CBs andSL under the epidermis (Fig. 3A).
A brief depiction of the RAM and root tip of T. filamentosais necessary; there is a closed, three-tiered meristem with a dis-tinct, distal tier for the very small rootcap, a separate tier ofground meristem/protoderm for cortex and epidermis, andthe proximal tier is for the procambium (Fig. 3B; Heimschand Seago, 2008). Its RAM appears slightly more monocotlike (tiered monocot in Heimsch and Seago, 2008) thanCabomba-like (tiered basal angiosperm in Heimsch andSeago, 2008). The root tip of T. filamentosa does not have acleft separating the lateral rootcap from the epidermis, andthe rootcap and its portion of the RAM overlie the proto-derm/ground meristem tier (Fig. 3B). This is very similar tothe closed or tiered RAM of the monocots under theHeimsch and Seago (2008) characterization (and alsoClowes, 2000). In T. filamentosa, the tier for ground
Seago & Fernando — Anatomical aspects of angiosperm root evolution226
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

meristem/protoderm is most clearly aligned with the epider-mis, and there is a very clear distinction between the rootcapinitials and the ground meristem/protoderm. The images ofnewly germinated T. submersa primary roots in Rudall et al.(2009) and in Friedman et al. (2012; their fig. 6A) appear tobe very similar to our image in Fig. 3B. Further, Fig 3B(left insert) shows a radial arrangement of cortical cell filesthat are like monocots derived from tiered monocot RAMs(Heimsch and Seago, 2008); this distinctive radial cortex isnot usually found in the Cabombaceae (Seago, 2002; Seagoet al., 2005; Heimsch and Seago, 2008). Strangely, the RAMand root tip of some Lemnaceae (Araceae, Alismatales) arevery similar to the Cabomba-tiered basal angiosperm-typeRAM with a cleft between the lateral rootcap and epidermis(see Landolt, 1998), except that its RAM has more pronouncedtiered monocot cell lineages like Trithuria.
Austrobaileyales
Illicium floridanum is used here as a representative of theorder. Like other members of this clade, it is terrestrial(Thomas, 1914; Metcalfe, 1987); its lateral roots are oftensmall and diarch, with early-maturing phloem fibres and anendodermis with CBs and SL delimiting a very small cortexwith some radial cell patterns and without an exodermis(Fig. 2E). Its adventitious roots are mostly diarch (Metcalfe,1987), but tetrarch patterns are also found, and early secondarygrowth produces secondary xylem with vessel elements, anarrow phloem region without noticeable pericyclic develop-ment into phellogen, and a redivided endodermis, fully com-plete with CBs and even thick SL (Fig. 2F); and anextensive hypodermis which is probably exodermal. Theseroots show a much more extensive cortex with irregular cellpatterns, typical of an open RAM origin for cortex, although
A
F
ED
B C
FI G. 2. Basal angiosperms, Amborellaceae and Austrobaileyales: Amborella trichopoda (A) Diarch. Early exodermis. Scale bar ¼ 110 mm. (B) Diarch withxylem continuous across poles, endodermis with early suberin lamellae (SL) development. Complex exodermis. Mid-cortex non-radial. Scale bar ¼ 120 mm.(C) Secondary growth, endodermis with SL and passage cells. Scale bar ¼ 120 mm. (D) Redivided endodermis, and some evidence of redivided mid-cortexand exodermis cells; secondary xylem without vessels, phloem surrounding secondary xylem. Scale bar ¼ 80 mm. Illicium floridanum (E) Diarch root withCasparian bands and early suberin formation. Scale bar ¼ 140 mm. (F) Secondary xylem with vessels and redivided endodermis with SL, and some evidenceof redivided exodermal cells. Scale bar ¼ 130 mm. Abbreviations (used both here and on other figures): c, cortex; e, epidermis; en, endodermis; ex, exodermis,
l, lacuna; p, passage cell of endodermis; s, secondary xylem; v, velamen; x, primary xylem.
Seago & Fernando — Anatomical aspects of angiosperm root evolution 227
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

lateral roots have monocot-like RAMs (Heimsch and Seago,2008).
Magnoliids
This clade is represented by four orders, i.e. Canellales,Laurales, Magnoliales and Piperales (APG, 2009). However,
since there is no information on the roots of the Canellales, wewill deal only with the representative families from the otherthree orders: Calycanthaceae, Lauraceae, Magnoliaceae,Annonaceae, Aristolochiaceae, Piperaceae and Saururaceae.The magnoliids are terrestrial and their primary and lateralroots vary from diarch to hexarch (present study; Thomas,1914; Metcalfe and Chalk, 1950a, b; Metcalfe, 1987); they are
A C
D
E
B
FI G. 3. Basal angiosperms, Nymphaeales: (A) Trithuria filamentosa – monarch stele, nine-celled endodermis with Casparian bands (CBs) and suberin lamellae(SL), mid-cortex with aerenchyma and collapsed radiating cells, exodermis and epidermis. Scale bar ¼ 20 mm. (B) Median longisection of root tip with tieredbasal angiosperm (TB) apical organization (arrow). Scale bar ¼ 20 mm. Insets: left – radial cortical cell files, scale bar ¼ 40 mm; right – aerenchyma lacunaeexpanded by cell expansion, scale bar ¼ 35 mm. (C) Cabomba caroliniana – monarch stele, nine-celled endodermis, schizogenous–expansigenous aerenchyma,exodermis with double CBs; photograph from Seago (2002), with permission from the Journal of the Torrey Botanical Society. Scale bar ¼ 65 mm. (D) Nupharlutea – polyarch stele, astrosclereids in expansigenous aerenchyma, exodermis with CBs and SL; photograph from Seago (2002), with permission from the TorreyBotanical Society. Scale bar ¼ 200 mm. (E) Nymphaea odorata – polyarch stele (seven xylem poles), astrosclereids, expansigenous aerenchyma. Scale bar ¼
60 mm. See Fig. 2 for list of abbreviations.
Seago & Fernando — Anatomical aspects of angiosperm root evolution228
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

usually tetrarch or pentarch, but hexarch is also common (e.g.Magnolia, Liriodendron, Asimina, Laurus, Aristolochia,Saururus; Fig. 4A–D). For primary roots, Calycanthus andAsimina (see Thomas, 1914; Hayat and Canright, 1965;Metcalfe, 1987) are initially diarch and Saururus is tetrarch(present study; Calycanthus may also be tetrarch, Thomas,1914). Adventitious and lateral roots in Laurus (Lauraceae)and Aristolochia (Aristolochiaceae) can also be tetrarch ordiarch (with two distinct and widely separate protophloem ele-ments at each pole). In the Piperaceae, Metcalfe (1987) reporteddiarchy and tetrarchy in Peperomia and polyarchy in Piper. Inmost magnoliids, the centre of the stele often is occupied bymetaxylem (Fig. 4B, C) or sclerenchyma (Fig. 4D). A pericycledelimits the stele, and the cortex is delimited on its interior by an
endodermis which forms CBs and later SL; there is usually a uni-seriate exodermis (Fig. 4B, D); depending on the state of earlysecondary growth, an epidermis may or may not be present inolder roots after exodermis maturation. Endodermis and exoder-mis increase in cell number during secondary growth. All mag-noliids so far reported have common initials in their RAMs(Heimsch and Seago, 2008).
Monocots
Most monocot roots are adventitious and arise endogenouslyusually from stems or leaves (Tomlinson, 1961, 1969, 1982;Metcalfe, 1971; Ayensu, 1972; Keating, 2002; Bell andBryan, 2008). In mature roots, the stele is often polyarch (six
A B
C D
FI G. 4. Magnoliids, Magnoliales. (A) Magnolia soulangeana – young root, hexarch stele, endodermis. Scale bar ¼ 65 mm. (B) Secondary xylem with vessels,redivided endodermis with suberin lamellae (SL) and passage cells, exodermis. Scale bar ¼ 30 mm. (C) Aristolochia sp. – tetrarch stele with early secondaryxylem, endodermis with Casparian bands (CBs) and early suberin deposition, early stage of exodermis wall deposition. Scale bar ¼ 55 mm. (D) Saururuscernuus – tetrarch root with early secondary growth, endodermal CBs, exodermis with CBs and partial SL. Scale bar ¼ 115 mm. See Fig. 2 for list of
abbreviations.
Seago & Fernando — Anatomical aspects of angiosperm root evolution 229
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022
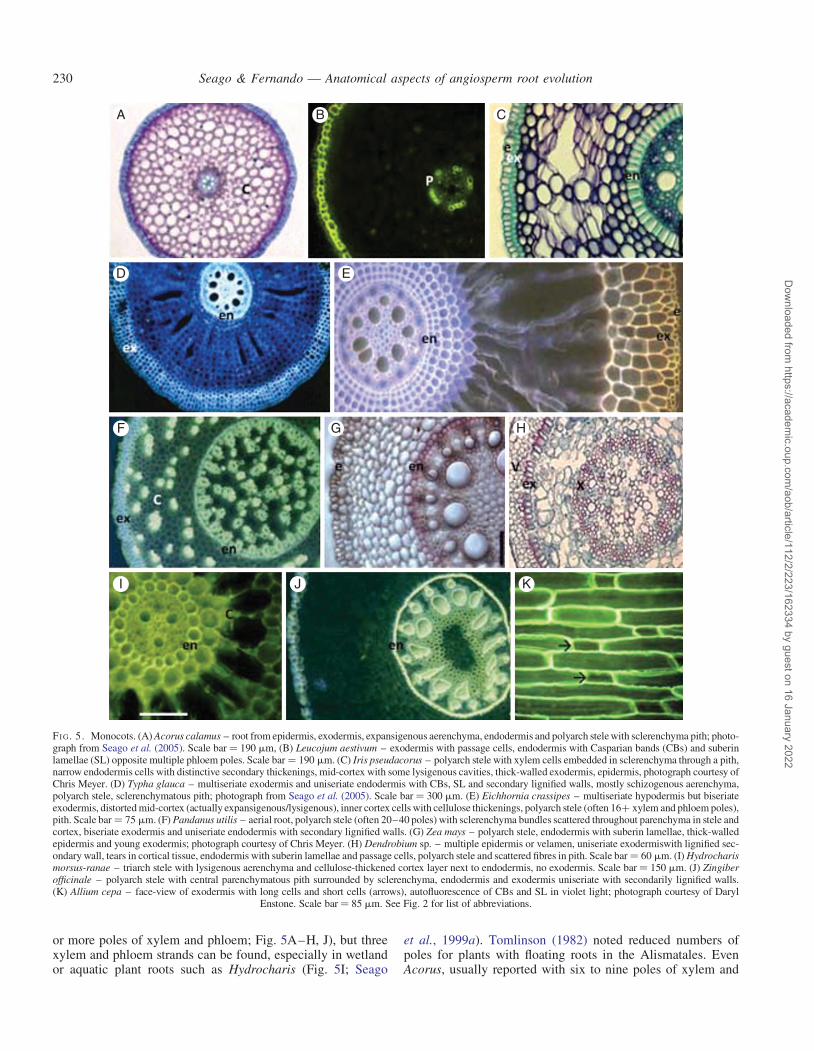
or more poles of xylem and phloem; Fig. 5A–H, J), but threexylem and phloem strands can be found, especially in wetlandor aquatic plant roots such as Hydrocharis (Fig. 5I; Seago
et al., 1999a). Tomlinson (1982) noted reduced numbers ofpoles for plants with floating roots in the Alismatales. EvenAcorus, usually reported with six to nine poles of xylem and
A
D E
B C
F G H
I J K
FI G. 5. Monocots. (A) Acorus calamus – root from epidermis, exodermis, expansigenous aerenchyma, endodermis and polyarch stele with sclerenchyma pith; photo-graph from Seago et al. (2005). Scale bar ¼ 190 mm, (B) Leucojum aestivum – exodermis with passage cells, endodermis with Casparian bands (CBs) and suberinlamellae (SL) opposite multiple phloem poles. Scale bar ¼ 190 mm. (C) Iris pseudacorus – polyarch stele with xylem cells embedded in sclerenchyma through a pith,narrow endodermis cells with distinctive secondary thickenings, mid-cortex with some lysigenous cavities, thick-walled exodermis, epidermis, photograph courtesy ofChris Meyer. (D) Typha glauca – multiseriate exodermis and uniseriate endodermis with CBs, SL and secondary lignified walls, mostly schizogenous aerenchyma,polyarch stele, sclerenchymatous pith; photograph from Seago et al. (2005). Scale bar ¼ 300 mm. (E) Eichhornia crassipes – multiseriate hypodermis but biseriateexodermis, distorted mid-cortex (actually expansigenous/lysigenous), inner cortex cells with cellulose thickenings, polyarch stele (often 16+ xylem and phloem poles),pith. Scale bar ¼ 75 mm. (F) Pandanus utilis – aerial root, polyarch stele (often 20–40 poles) with sclerenchyma bundles scattered throughout parenchyma in stele andcortex, biseriate exodermis and uniseriate endodermis with secondary lignified walls. (G) Zea mays – polyarch stele, endodermis with suberin lamellae, thick-walledepidermis and young exodermis; photograph courtesy of Chris Meyer. (H) Dendrobium sp. – multiple epidermis or velamen, uniseriate exodermiswith lignified sec-ondary wall, tears in cortical tissue, endodermis with suberin lamellae and passage cells, polyarch stele and scattered fibres in pith. Scale bar ¼ 60 mm. (I) Hydrocharismorsus-ranae – triarch stele with lysigenous aerenchyma and cellulose-thickened cortex layer next to endodermis, no exodermis. Scale bar ¼ 150 mm. (J) Zingiberofficinale – polyarch stele with central parenchymatous pith surrounded by sclerenchyma, endodermis and exodermis uniseriate with secondarily lignified walls.(K) Allium cepa – face-view of exodermis with long cells and short cells (arrows), autofluorescence of CBs and SL in violet light; photograph courtesy of Daryl
Enstone. Scale bar ¼ 85 mm. See Fig. 2 for list of abbreviations.
Seago & Fernando — Anatomical aspects of angiosperm root evolution230
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

phloem (Keating, 2002; Soukup et al., 2005), may have onlyfive poles (Soukup et al., 2005). Very high numbers ofstrands (≥20) occur in plants with large and/or aerial rootssuch as in Pandanaceae and Arecaceae (Fig. 5F, G, J;Tomlinson, 1961, 1982; French, 1987a, b).
Figure 5 shows other features of some diverse monocotsfrom the Acorales to the Zingiberales. The endodermis is uni-seriate and most often has cell wall stages I, II and III [e.g.Fig. 5C, D, F, H, J; see Meyer et al. (2009) for endodermalcell wall stages]. The middle cortex has great variability, butone of the most unique features is the occurrence of scleren-chyma bundles in roots of many aerial plants (Fig. 5F;Tomlinson, 1961; Keating, 2002). Aerenchyma is a commonfeature in the many families with aquatic or wetland species(Fig. 5A, B, D). In basal monocots (Acorus), aerenchyma isexpansigenous, and in derived monocots it is often ofvarying schizo-lysigenous to lysigenous types (Fig. 5I; Seagoet al., 2005; Jung et al., 2008). However, aerenchyma patternsin most angiosperms are still not well represented (Jacksonet al., 2009; Van der Valk, 2012), except that the membersof Cyperaceae are well known for their unusual tangential lysi-geny (Seago et al., 2005; see also Metcalfe, 1971). Often, inplants with aerial roots, especially large roots, the air spacesin the cortex are described as cavities (Tomlinson, 1961)because they are not developed or organized like typical aer-enchyma. A hypodermis ranges from uniseriate to multiseri-ate exodermis (Fig. 5A, C–H, J), and in many plants isdimorphic with long and short cells (Fig. 5K; Shishkoff,1987). A velamen occurs in some aerial roots, especially inorchids (Fig. 5H; Ayensu, 1962; Zankowski, 1987; Keating,2002).
The single most distinctive feature of monocots, as reportedextensively by Clowes (2000) and Heimsch and Seago (2008),is the developmental association between ground meristem andprotoderm in the RAM.
Eudicots
As early as 1914, Thomas compared the vasculature ofmany seedling roots that are now classified as either magno-liids or eudicots; he found them to vary from diarch tooctarch, but in Metcalfe and Chalk (1950a, b) and Metcalfe(1987) very few species’ roots are heptarch and octarch. Inthe basal eudicot Ranunculales, there is diarchy (Aquilegia,some Ranunculus; Thomas, 1914), tetrarchy (otherRanunculus, Berberis; Thomas, 1914) and pentarchy(Podophyllum). Thus, while vascular tissues of the stele inmost eudicots have two to six poles or strands of alternatingxylem and phloem (Fig. 6A, E–I; Metcalfe and Chalk,1950a, b), importantly, some groups near the base of the eudi-cots (Proteales) and core eudicots (Gunnerales) often havepolyarch steles. Nelumbo lutea (Proteales, Nelumbonaceae;Fig. 6B) is aquatic, but Gunnera (Gunnerales, Gunneraceae)grows both in aquatic and terrestrial habitats, and its speciesoften have large differences in numbers of xylem poles evenin terrestrial plants, as we have collected (cf. G. perpensa,Fig. 6C, and G. killipiana, Fig. 6D), even though the rootsmay be similar or dissimilar in size; tetrarch steles are appar-ently characteristic of these basal species (J.L. Seago, pers.obs.; A. Soukup and E. Tylova, pers. comm.; Wilkinson,
2000). Roots of species across the eudicot spectrum mayhave diarchy (Fig. 6I; e.g. Thomas, 1914; Hayward, 1938;Baum et al., 2002), especially when those roots are small, asin secondary root stages (Seago, 1973; Byrne et al., 1977).In many Fabaceae, primary roots are often triarch to hexarch(Fig. 6E), although in Glycine lateral roots originate in thediarch condition (Ambler et al., 1971) and then develop tet-rarchy (Byrne et al., 1977); tetrarchy is very common in eudi-cots (Metcalfe and Chalk, 1950a, b; Seago, 1971).
The cortex is delimited internally by the endodermis whichvaries as much in eudicots as it does in monocots, and oftenpassage cells with CBs are opposite protoxylem and SL cellsare opposite the protophloem (Fig. 6A, B, G); stage III cellwalls appear to be less common in eudicots. Air spaces in theform of aerenchyma are found most commonly in aquatic eudi-cots (e.g. Fig. 6G, H). There is an exodermis in many eudicots(e.g. Fig. 6A–C, H), but it is typically lacking in nodule-producing roots with an open transversal RAM such aslegumes (Heimsch and Seago, 2008). Many eudicots, especiallythe many trees and shrubs, have secondary root growth, even ifvery limited as in small herbaceous plants (e.g. Fig. 6F;Metcalfe and Chalk, 1950a, b). Secondary root growth is prob-ably accompanied by a dilatated endodermis and exodermis inmany species, as in Gentiana (Fig. 6J; Sottnikova and Lux,2003) and Medicago (our observations).
Resin canals in roots are known but relatively little studied(French, 1987a). Laticifers are more a feature of eudicots (e.g.Ipomoea purpurea; Seago, 1971; and Lactuca sativa, J.L.Seago, pers. obs.; see also Metcalfe and Chalk, 1950a, b;Metcalfe, 1967) than of monocots where they are rare(Metcalf, 1967). Root laticifer development has been studied(e.g. Seago, 1971), and crystalliferous and tanniniferous cells, es-pecially in rootcap or cortex, are also well known (e.g. Seago andMarsh, 1989).
All eudicots have a RAM with the protoderm/epidermisassociated developmentally with the lateral rootcap (Clowes,2000; Groot et al., 2004; Heimsch and Seago, 2008).
Air spaces in roots
Aerenchyma types have been presented by several research-ers (e.g. Justin and Armstrong, 1987; Evans, 2004), but theexplanations for the development of intercellular spaces intoaerenchymatous lacunae by Seago et al. (2005) are the onlyadequate explanations for the roles of cell division, cell expan-sion, cell separations and cell deaths that can account for thetypes of root cortical aerenchyma: expansigeny, schizogenyand lysigeny. Based on this feature, Seago et al. (2005) andJung et al. (2008) best provide the possible evolutionarypath from basal angiosperms to monocots or eudicots.Clearly, the earliest root aerenchyma in angiosperms wasmost probably by expansigeny (Fig. 7A – Acorus;Nymphaeales of basal angiosperms and Acorales of monocots;Seago et al., 2005; Soukup et al., 2005), the lacunae arising bycell division and cell expansion, not by schizogeny (Fig. 7B)or lysigeny (Fig. 7C, as noted by Jackson et al., 2009).Particularly in monocots, various kinds of lysigeny arose inmore derived families of several orders. The occurrence of dia-phragms across aerenchymatous lacunae has been notedand even studied in detail (e.g. in Cabombaceae and
Seago & Fernando — Anatomical aspects of angiosperm root evolution 231
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

A B C
D E F
G H I J
FI G. 6. Eudicots. (A) Ranunculus repens – tetrarch stele, endodermis with passage cells, cortex non-aligned, exodermis Casparian bands (CBs) and suberinlamellae (SL); brightly fluorescing epidermis. Scale bar ¼ 85 mm. (B) Nelumbo lutea – polyarch stele with sclerified pith, endodermis with passage cellsand suberized cells, exodermis suberized. Scale bar ¼ 115 mm. (C) Gunnera perpensa – polyarch stele (seven poles), endodermis, mid-cortex with expansigen-ous spaces, endodermis with CBs and SL. Scale bar ¼ 190 mm. (D) Gunnera killipiana – polyarch stele with 18 xylem strands, bundles within pith and paren-chyma. Scale bar ¼ 200 mm. (E) Medicago sativa – typical legume triarch stele, non-radial cortex, no hypodermis/exodermis. Scale bar ¼ 95 mm; inset:somewhat unusual tetrarch stele observed in very few primary roots. (F) Fraxinus americana – secondary xylem, but pentarch primary xylem visible. Scalebar ¼ 140 mm. (G) Rumex crispus – pentarch stele with partial sixth pole and central metaxylem, endodermis with CBs, expansigenous aerenchyma, no exo-dermis. Scale bar ¼ 135 mm. (H) Nymphoides crenata – pentarch with pith, endodermis with CBs only and exodermis with CBs and SL, astrosclereids in mid-cortex with aerenchyma. (I) Artemisia lavandulaefolia – diarch primary root, no pith, endodermis with CBs only, faint CB staining in hypodermis; photographcourtesy of Chaodong Yang. Scale bar ¼ 80 mm. (J) Gentiana asclepiadea – root with dilatated endodermis and exodermis in early secondary growth; photo-
graph courtesy of Alexander Lux. Scale bar ¼ 45 mm. See Fig. 2 for list of abbreviations.
Seago & Fernando — Anatomical aspects of angiosperm root evolution232
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

Nymphaeaceae of the Nymphaeales, Seago et al., 2005; theHydatellaceae do not appear to have diaphragms, possibly aconsequence of having only cell expansion and no furthercell divisions contributing to the lacunae). The presence orabsence of diaphragms has not been widely studied acrossmonocots and eudicots (Seago et al., 2005).
In legumes, vascular cavities can be found in the pith ofsome triarch Pisum roots (Fig. 7D) under flooded conditions(Niki and Gladish, 2001). Legumes do not have the cortex de-velopment or structures that allow easy formation of aeren-chyma (Seago et al., 2005) or exodermis (Heimsch andSeago, 2008). Such cavities are not considered aerenchyma.Secondary aerenchyma, aerenchymatous phellem derivedfrom phellogen, can also occur in wetland plants (Lythrum sal-icaria; Stevens et al., 1997) and may be found in the fossilrecord in related species (see Little and Stockey, 2003).
DISCUSSION ON SELECTED ASPECTSOF ROOT ANATOMY
Root apical meristem
From the concepts of Barlow (1994, 2002) on increasing com-plexity and quiescence, to Clowes (1994) on epidermis origins,the possible evolutionary path of RAM organization has beenpresented in three major studies by Clowes (2000), Groot et al.(2004) and Heimsch and Seago (2008). The latter authors pre-sented an analysis of RAMs with several manifestations ofclosed and open types and reported that some specimens ofAmborella trichopoda and the magnoliids contain commoninitials for most meristematic tissues of the root. As statedabove, Heimsch and Seago (2008) further related the openand closed RAMs (with cortex and epidermis association) inthe nymphaealean families (Cabombaceae, Nymphaeaceaeand now the Hydatellaceae) to the monocots. In Friedmanet al. (2012), fig. 6 corroborates our findings herein forT. filamentosa that its primary, adventitious and lateral rootshave a tiered monocot-type RAM (sensu Clowes, 1994,2000; Heimsch and Seago, 2008), and the pattern of corticaldevelopment from a tiered RAM further illustrates a monocot-like root.
In overcoming some of the questions which Les andSchneider (1995) posed about the lack of solid evidence fornymphaealean and monocot phylogenetic connections, weargue that there is no stronger anatomical evidence for aNymphaealean–monocot connection than the RAM and
cortex, because such a type of anatomy is not found inAmborellaceae, magnoliids and eudicots. These patternsclearly arose in the ancestors of monocots, i.e. various ances-tral, early basal angiosperms, as the patterns of epidermis andlateral rootcap connections characteristic of some basal angios-perms and all eudicots must have separately so arisen.
Further, there appears to be a clear association betweenRAM organization and the patterns of lateral rootcap cellsand their sloughing (Hamamoto et al., 2006). Open RAM pro-duces more cells and releases individual living border cells,whereas closed RAM releases sheets or groups of dead cells.The fate of lateral rootcap cells in the tiered or closed RAMsof Cabombaceae and Hydatellaceae, as well as the open trans-versal RAMs of Nymphaeaceae, need to be examined to deter-mine if the same relationships holds for RAMs of these basalangiosperms. The differentiation of epidermal cells, especiallyin simple tiered RAMs, has received enormous attention in justa select few species (Bruex et al., 2012; Jones and Dolan,2012), and this needs to be expanded.
Cortex: endodermis and hypodermis
The endodermis is a well-defined structural feature of angio-sperm roots (Kroemer, 1903; Van Fleet, 1950; Wilson andPeterson, 1983; Seago and Marsh, 1989; Seago et al., 1999b;Soukup et al., 2005; Meyer et al., 2009), except possibly inholoparasites (Kuijt and Bruns, 1987). A hypodermis is theoutermost cell layers of the cortex derived by periclinal divi-sions in the outer ground meristem (Seago and Marsh,1989). When CBs are present and SL are also alwayspresent, the cell layer(s) is termed exodermis (Kroemer,1903; Wilson and Peterson, 1983; Perumalla et al., 1990;Peterson and Perumalla, 1990; Seago et al., 1999b).Multiseriate exodermis is much more common in monocotsthan in eudicots (Seago et al., 1999a, b; Peterson andPerumalla, 1990). Two different cell types can occur in exo-dermis – long cells and short cells; Shishkoff (1987) reportedno dimorphic hypodermis in Nymphaeales (see also Seagoet al., 2000b) and Laurales, but found them in Magnolialesand in basal eudicot Ranunculales (not in the Papaveraceae,however). Dimorphic hypodermis as seen in Allium cepa(Fig. 5K) is fairly common in monocots. The exodermis andits passage cells can have major effects on root–fungus asso-ciations (e.g. Baylis, 1972; Brundrett, 2002).
There have been analyses of root structures with regard totheir application to systematics (e.g. French, 1987a, b;
A B C D
FI G. 7. Aerenchyma, air cavities. (A) Expansigeny – cell division and cell expansion, in Acorus calamus; photograph courtesy of Ales Soukup. Scale bar ¼60 mm. (B) Schizogeny – cell wall separations, in Typha glauca; photograph from Seago et al. (2005). Scale bar ¼ 90 mm. (C) Lysigeny – cell deaths, in Oryzasativa; photograph courtesy of Ales Soukup. Scale bar ¼ 140 mm. (D) Vascular or stelar air cavity (asterisk) – cell/tissue death, in Pisum sativum; photograph
courtesy of Daniel Gladish and Suma Sreekanta. Scale bar ¼ 150 mm. See Fig. 2 for list of abbreviations.
Seago & Fernando — Anatomical aspects of angiosperm root evolution 233
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

Keating, 2002), but differences in interpretations present someproblems. Kauff et al. (2000) found a dimorphic rhizodermisin Hydrocharitaceae and Pontederiaceae, but Seago et al.(1999a, 2000a) showed developmentally that only the outerlayer is the epidermis. Hydrocharis (Fig. 5I) does not havean exodermis, and Pontederia has a uniseriate exodermis asthe outer layer of a dimorphic or trimorphic hypodermiswhose inner layers are only suberized (see related Eichhornia,Fig. 5E).
Vascular tissues
The similarities between members of the Nymphaeales andthe Acorales have been noted with regard to xylem cell structure(Schneider and Carlquist, 1995, 2002; Carlquist and Schneider,1997) as well as cortex structure (Seago et al., 2005). Monocotshave far more aquatic/wetland species and families (Les andSchneider, 1995; Van der Valk, 2012) than do eudicots, andtheir roots, mostly adventitious, are often polyarch. In thebasal angiosperms, two of the families, Cabombaceae andHydatellaceae, have predominantly monarch roots, while theNymphaeaceae are dominated by species with mainly polyarchroots, as are Acorus (Acoraceae), sister to the rest of the mono-cots, and the Araceae (Keating, 2002). Most of the remainder ofthe monocots are polyarch, except for aquatic families such asHydrocharitaceae (Seago et al., 1999a) and Lemnaceae(Landolt, 1998), plants with tiny roots which have triarch ormonarch steles, respectively.
According to Metcalfe and Chalk (1950a, b), Popham(1966), Esau (1977) and Metcalf (1987), eudicots are generallydepicted as having two to six poles or strands of primaryxylem and phloem (often, apparently, diarch in young lateralroots; Byrne and Heimsch, 1968; Byrne, 1973). It seems thatdiarchy is more common in primary roots (derived from radi-cles), at least in the basal angiosperms. That some wetlandeudicots at the base of the core eudicots (Gunnera) and nearthe base of the basal eudicots (Nelumbo) are strikingly poly-arch, such as Nymphaeaceae and the vast majority of mono-cots, raises interesting questions. Possibly, plants evolving inaquatic/wetland conditions retain and express the genes neces-sary for adventitious root production and polyarchy morefrequently than non-aquatic plants. The greater the numberof poles or strands of xylem and phloem (heptarchy andabove), the less likely it is that secondary growth may occur,whereas diarch to hexarch patterns can lead more easily to sec-ondary growth.
Primary and adventitious roots
Since so many species, especially among basal angiosperms(including Nymphaeales, e.g. Friedman et al., 2012) and eudi-cots, have two cotyledons with a diarch vascular pattern inprimary and other roots, except in the nymphaealeanTrithuria, leading to the possibility that diarchy is stronglyrelated to the dicotyledonary condition, then one mightexpect that monocot primary roots might have a monarchprimary or seminal root. Such is clearly not the case; and,too many basal angiosperms and magnoliids have patternsother than diarchy.
Another aspect of development and structure which shouldbe examined more closely is the state of embryo developmentand structure at the time of maturation and germination.Amborellales, Nymphaeales and Austrobaileyales have verysmall embryos with little differentiation (Martin, 1946; Tobeet al., 2000; Friedman et al., 2012; see also Taylor et al.,2006, for fossil Nymphaeaceae). This might be important tothe balance between a primary root system and adventitiousroot systems, to the relative state of development in primaryroots vs. adventitious roots and to differences in origin ofmonocots and eudicots from basal angiosperm ancestors.
Most eudicots, when producing adventitious roots, formthem from more or less typical eudicot vascular patterns instems, bundles in one ring with a remnant procambial strandor incipient vascular cambium. Most monocots, on the otherhand, form adventitious roots from stems with scattered vascu-lar bundles or two or more rings of vascular bundles, so thatone could argue that it is the number of available vascularbundles that produces the greater number of xylem andphloem poles in monocot roots. Thus, vascular patterns inembryonically produced roots might reflect vascular bundledistributions of their stems. The contributions of moleculargenetics will have a major impact on our understanding of evo-lution of vascular patterns in roots (see Scarpella and Meijer,2004).
Root symbioses, mycorrhizae and nodules
On the matter of mycorrhizae, after Baylis (1972), Simonet al. (1993) and Cairney (2000), Brundrett’s (2002) thesisof rhizomes evolving into roots related fungal inhabitationsto root evolution. Brundrett offered possible root structuralfeatures needed to accompany the evolutionary pathways.Mycorrhizal roots can sometimes be extremely modified(e.g. Imhof, 1997, 2001). For nodules, the study of Soltiset al. (1995) confirmed that there are only two eudicot families(Fabaceae, Ulmaceae) with Rhizobium nodules and eight fam-ilies (Betulaceae, Casuarinaceae, Elaeagnaceae, Myricaceae,Rhamnaceae, Rosaceae, Datiscaceae and Coriariaceae) withFrankia actinomycetes. A recent study by Markmann et al.(2008) suggests how the evolutionary path may have involvedthe same genes in both nodulating bacteria, but the root struc-tural paths have not been well addressed. Heimsch and Seago(2008) and Seago et al. (2005) briefly discussed the ramifica-tions of RAM and cortex structure, respectively, in relation tonodule formation. It should be noted that these families withnodulating roots are not closely associated with basal angios-perms or basal eudicots, and none is found in the monocotswhere epidermal origin is associated with cortex, not lateralrootcap; roots with bacterial symbioses seem likely to repre-sent a derived condition in angiosperms.
In summary, root anatomy offers many interesting perspec-tives on developmental patterns, systematics and evolutionaryrelationships but, since their structure can vary depending onthe type of experimental conditions, their importance isoften less appreciated. However, when roots are examinedbased on their typical habitats, they can be useful when com-paring groups of plants. Therefore, based on the informationpresented in this overview, there appears to be a generaltrend in angiosperm root structure (see summarized
Seago & Fernando — Anatomical aspects of angiosperm root evolution234
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

TABLE 1. Summary of selected, but typical anatomical features of angiosperm roots (including RAMs from Heimsch and Seago, 2008)
Taxa HabitatTypes of roots
examinedTypes of
steleStage ofgrowth Endodermis
Mid-cortexpattern Aerenchyma Hypodermis exodermis RAM
Amborellales Terrestrial Adventitious;lateral
Diarch;triarch
Primary;secondary
All with CBs and SL Non-radial None All with CBs and SL Epidermis-lateral rootcap;common initials, irregularepidermis
Nymphaeales Aquatic Adventitious Monarchpolyarch
Primaryonly
Some CBs; some CBsand SL
Radial;non-radial
Expansigenous All with CBs and SL Epidermis-cortex
Austrobaileyales Terrestrial Adventitious;lateral
Diarchtetrarch
Primary;secondary
All with CBs SL;some dilatated
Non-radial None All with CBs and SL Epidermis-cortex;common initials, irregularepidermis
Magnoliids Terrestrial Primary;adventitious;lateral
Diarch tohexarch
Primary;secondary
All with CBs and SL;some dilatated
Non-radial None All with CBs and SL;some dilatated
Common initials, irregularepidermis
Monocots Aquatic Adventitious Polyarch Primaryonly
Some only CBs; someCBs and SL; manyCBs, SL andsecondary walls
Radial;non-radial
Expansigenous;schizogenous;lysigenous
None; some CBs andSL; many CBs, SL andsecondary walls
Epidermis-cortexTerrestrial Primary; lateral Triarch in a
few
Eudicots Terrestrial Primary; lateral Diarch tohexarch
Primary;mostlysecondary
Many CBs; some CBsand SL; some CBs,SL and secondarywalls
Radian;non-radial
Schizogenous;lysigenous;expansigenous
None; some CBs andSL; some CBs, SL andecondary walls; somedilatated
Epidermis-lateral rootcap
Aquatic Adventitious Polyarch insome basalaquatics
Sea
go
&F
erna
nd
o—
An
ato
mica
la
spects
of
an
gio
sperm
roo
tevo
lutio
n2
35
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

information in Table 1) and, in general, we note that theAmborellales and magnoliids have many root structural fea-tures like those of eudicots, whereas the Nymphaeales rootsare strikingly similar to those of the monocots, especiallybasal monocots such as Acorales. The Austrobaileyales are en-igmatic and have root structural features which do not aligneasily to either monocots or eudicots. Clearly, the basalangiosperms require far more anatomical examination to cor-roborate the findings of molecular phylogenetic analyses.
ACKNOWLEDGEMENTS
For their assistance in various ways, we thank Arnold Salazar,Ales Soukup, Daryl Enstone, Christopher Meyer, ChaodongYang, Carol Peterson, Marilyn Seago, Edita Tylova,Alexander Lux, Daniel Gladish, Olga Votrubova and two an-onymous reviewers. J.L.S. received a travel grant fromSUNY Oswego Office of International Education to supportthe ISRR trip. The late Charles Heimsch is specially notedbecause he instilled in J.L.S. the idea that root evolution inangiosperms is very important.
LITERATURE CITED
Ambler JE, Brown JC, Gauch HC. 1971. Sites of iron reduction in soybeanplants. Agronomy Journal 63: 95–97.
APG. 2009. An update of the Angiosperm Phylogeny Group classification forthe orders and families of flowering plants: APG III. Botanical Journal ofthe Linnean Society 161: 105–121.
Ayensu ES. 1972. Anatomy of the monocotyledons. VI. Dioscoreales. Oxford:Clarendon Press.
Barlow PW. 1994. Structure and function at the root apex – phylogenetic andontogenetic perspectives on apical cells and quiescent centres. Plant andSoil 167: 1–16.
Barlow PW. 2002. Cellular patterning in root meristems: its origins and sig-nificance. In: Waisel Y, Eshel A, Kafkafi U. eds. Plant roots: thehidden half. New York: Marcel Dekker, 49–82.
Baum SF, Dubrovsky JG, Rost TL. 2002. Apical organization and matur-ation of the cortex and vascular cylinder in Arabidopsis thaliana(Brassicaceae) roots. American Journal of Botany 89: 908–920
Baurle I, Laux T. 2003. Apical meristems: the plant’s fountain of youth.Bioessays 25: 961–970.
Baylis GTS. 1972. Fungi, phosporus, and the evolution of root systems. Search3: 257–258.
Bell AD, Bryan A. 2008. Plant form an illustrated guide to flowering plantmorphology. Portland OR: Timber Press.
Bennett T, Scheres B. 2010. Root development – two meristems for the priceof one? Current Topics in Developmental Biology 91: 67–102.
Bierhorst DW. 1971. Morphology of vascular plants. New York: MacMillan.Boyce CK, Knoll AH. 2002. Evolution of developmental potential and the
multiple independent origin of leaves in Paleozoic vascular plants.Paleobiology 28: 70–100.
Bruex A, Kainkaryam RM, Wieckowski Y, et al. 2012. A gene regulatorynetwork for root epidermis cell differentiation in Arabidopsis. PLoSGenetics 8: e1002446. http://dx.doi.org/10.1371/journal.pgen.1002446.
Brundrett MC. 2002. Coevolution of roots and mycorrhizas of land plants.New Phytologist 154: 275–304.
Byrne JM. 1973. The root apex of Malva sylvestris. III. Lateral root develop-ment and the quiescent center. American Journal of Botany 60: 657–662.
Byrne JM, Heimsch C. 1968. The root apex of Linum. American Journal ofBotany 55: 1011–1019.
Byrne JM, Pesacreta TC, Fox JA. 1977. Development and structure of thevascular connection between the primary and secondary root of Glycinemax (L.) Merr. American Journal of Botany 64: 946–959.
Byrne ME, Kidner CA, Martiennsen RA. 2003. Plant stem cells: divergentpathways and common themes in shoots and roots. Current Opinion inGenetics and Development 13: 551–557.
Cairney JWG. 2000. Evolution of mycorrhiza systems. Naturwissenschaften87: 467–475.
Carlquist S. 1975. Ecological strategies of xylem evolution. Berkeley, CA:University of California Press.
Carlquist S, Schneider EL. 1997. Origins and nature of vessels inMonocotyledons. I. Acorus. International Journal of Plant Sciences158: 52–56.
Carlquist S, Schneider EL. 2001. Vegetative anatomy of the New Caledonianendemic Amborella trichopoda: relationships with the Illiciales andimplications for vessel origin. Pacific Science 55: 305–312.
Carlquist S, Schneider SL. 2002. The tracheid–vessel element transition inangiosperms involves multiple independent features: cladistic conse-quences. American Journal of Botany 89: 185–195.
Carlquist S, Schneider SL. 2009. Do tracheid microstructure and the presenceof minute crystals link Nymphaeaceae, Cabombaceae and Hydatellaceae?Botanical Journal of the Linnean Society 159: 572–582.
Clowes FAL. 1994. Origin of the epidermis in root meristems. NewPhytologist 127: 335–347.
Clowes FAL. 2000. Patterns of root meristem development in angiosperms.New Phytologist 146: 83–94.
Conard HS. 1905. The waterlilies: a monograph of the genus Nymphaea.Washington, DC: The Carnegie Institute of Washington.
Cooke TJ, Poli D, Cohen JD. 2004. Did auxin play a crucial role in the evo-lution of novel body plans during the Late Silurian-Early Devonian radi-ation in land plants? In: Hemsley A, Poole I. eds. The evolution of plantphysiology. London: Academic, 85–107.
Dolan L, Scheres B. 1998. Root pattern: shooting in the dark? Cell andDevelopmental Biology 9: 201–206.
Edwards D, Feehan J. 1980. Records of Cooksonia-type sporonagia from lateWenlock strata in Ireland. Nature 287: 41–42.
Edwards D, Davies KL, Axe L. 1992. A vascular conducting strand in theearly land plant Cooksonia. Nature 357: 683–685.
Esau K. 1977. Plant anatomy. New York: John Wiley & Son.Evans DE. 2004. Aerenchyma formation. New Phytologist 161: 35–49.Fairon-Demart M, Li CS. 1993. Lorophyton goense gen. et sp. Nov. from the
Lower Givetian of Belgium and a discussion of Lower DevonianCladoxylopsida. Review of Paleobiology and Palynology 77: 1–22.
French JC. 1987a. Systematic survey of resin canals in roots of Araceae.Botanical Gazette 148: 360–371.
French JC. 1987b. Systematic occurrence of a sclerotic hypodermis in roots ofAraceae. American Journal of Botany 74: 891–903.
Friedman WE, Moore RC, Purugganan MD. 2004. The evolution of plantdevelopment. American Journal of Botany 91: 1726–1741.
Friedman WE, Bachelier JB, Hormaza JI. 2012. Embryology in Trithuriasubmerse (Hydatellaceae) and relationships between embryo, endosperm,and perisperm in early-diverging flowering plants. American Journal ofBotany 99: 1083–1095.
Friis EM, Pedersen KR, Crane PR. 1999. Early angiosperm diversification:the diversity of pollen associated with the Early Cretaceous (Barremian orAptian) of Western Portugal. International Journal of Plant Sciences 161(suppl): S169–S182.
Friis EM, Pedersen KR, Crane PR. 2001. Fossil evidence of water lilies(Nymphaeales) in the Early Cretaceous. Nature 410: 357–360.
Friis EM, Pedersen KR, Von Balthazar M, Grimm GW, Crane PR. 2009.Monetianthus mirus gen. et sp. nov., a nymphaealean flower from theEarly Cretaceous of Portugal. International Journal of Plant Sciences170: 1086–1101.
Fukaki H, Wysocka-Diller J, Kato T, Fugisawa H, Benfey PN, Tasaka M.1998. Genetic evidence that the endodermis is essential for shoot gravi-tropism in Arabidopsis thaliana. The Plant Journal 14: 425–430.
Gensel PG, Kotyk ME, Basinger JF. 2001. Morphology of above- andbelow-ground structures in early Devonian (Pragian-Emsian) plants. In:Gensel PG, Edwards D. eds. Plants invade the land: evolutionary and en-vironmental perspectives. New York: Columbia University Press,83–102.
Groot EP, Rost TL. 2001. Patterns of apical organization in roots of floweringplants. In: Proceedings of the 6th Symposium of the International Societyof Root Research. Nagoya, Japan, Japanese Society for Root Research,8–9, Bagoya, Japan.
Groot EP, Doyle JA, Nichol SA, Rost TL. 2004. Phylogenetic distributionand evolution of root apical meristem organization in dicotyledonousangiosperms. International Journal of Plant Sciences 165: 97–105.
Seago & Fernando — Anatomical aspects of angiosperm root evolution236
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

Haecker A, Grob-Hardt R, Geiges B, et al. 2003. Expression dynamics ofWOX genes mark cell fate decisions during early embryonic patterningin Arabidopsis thaliana. Development 131: 657–668.
Hamamoto L, Hawes MC, Rost TL. 2006. The production and release ofliving rot cap border cells is a function of root apical meristem type indicotyledonous angiosperm plants. Annals of Botany 97: 917–923.
Hayat MA, Canright JE. 1965. The developmental anatomy of theAnnonaceae. I. Embryo and early seedling structure. American Journalof Botany 52: 228–237.
Hayward HE. 1938. The structure of economic plants. New York: Macmillan.Heimsch C, Seago JL Jr. 2008. Organization of the root apical meristem in
angiosperms. American Journal of Botany 95: 1–21.Imhof S. 1997. Root anatomy and mycotrophy of the achlorophyllous Voyria
tenella Hook. (Gentianaceae). Botanica Acta 110: 298–305.Imhof S. 2001. Subterranean structures and mycotrophy of the achlorophyl-
lous Dictyostega orobanchoides (Burmanniaceae). Revista de BiologiaTropical 49: 239–247.
Jackson MB, Ishizawa K, Ito O. 2009. Evolution and mechanisms of planttolerance to flooding stress. Annals of Botany 103: 137–142.
Jones VAS, Dolan L. 2012. The evolution of root hairs and rhizoids. Annals ofBotany 110: 205–212.
Jung J, Lee SC, Cho HK. 2008. Anatomical patterns of aerenchyma inaquatic and wetland plants. Journal of Plant Biology 51: 428–439.
Justin SHFW, Armstrong W. 1987. The anatomical characteristics of rootsand plant response to soil flooding. New Phytologist 105: 465–495.
Kauff F, Rudall PJ, Conran JG. 2000. Systematic root anatomy ofAsparagales and other monocotyledons. Plant Systematics andEvolution 223: 139–154.
Keating RC. 2002. Anatomy of the monocotyledons. IX. Acoraceae andAraceae. Oxford: Clarendon Press.
Kenrick P, Crane PR. 1997. The origin and early evolution of plants on land.Nature 389: 33–39.
Kroemer K. 1903. Wurzelhaut Hypodermis und Endodermis derAngiospermwurzel. Bibliotheca Botanica 59: 1–151.
Kuijt J, Bruns D. 1987. Roots in Corynaea (Balanophoraceae). NordicJournal of Botany 7: 539–542.
Landolt W. 1998. Anatomy of the Lemnaceae (duckweeds). In: Landolt E,Jager-Zurn I, Schnell RAA. eds. Extreme adaptations in angiospermoushydrophytes, I. Berlin: Gebruder Borntraeger, 1–127.
Lang WH. 1937. On the plantremains from the Downtonian of England andWales. Philosophical Transactions of the Royal Society B: BiologicalSciences 227: 245–291.
Les DH, Schneider EL. 1995. The Nymphaeales, Alismatidae, and the theoryof an aquatic monocotyledon origin. In: Rudall PJ, Cribb PJ, Cutler DF,Humphries CJ. eds. Monocotyledons: systematics and evolution. Kew:Royal Botanic Gardens, 23–42.
Li CS, Edwards D. 1995. The reinvestigation of Halle Drepanophycus spinae-formis Gopp, from the lower Devonian of Yunnan province, southernChina. Botanical Journal of the Linnean Society 118: 163–192.
Little SA, Stockey RA. 2003. Vegetative growth of Decodon allenbyensis(Lythraceae) from the Middle Eocene Princeton Chert with anatomicalcomparisons to Decodon verticillatus. International Journal of PlantSciences 164: 453–469.
Lux A, Luxova M. 2003. Growth and differentiation of root endodermis inPrimula acaulis Jacq. Biologia Plantarum 47: 91–97.
Markman K, Giczey G, Parniske M. 2008. Functional adaptation of a plantreceptor-kinase paved the way for the evolution of intracellular root sym-bioses with bacteria. PLOS Biology 6: 0497–0506.
Martin AC. 1946. The comparative internal morphology of seeds. AmericanMidland Naturalist 36: 513–560.
Mayer KFX, Schoof H, Haecker A, Lenhard M, Jurgemns G, Laux T.1998. The role of WUSHEL in regulating stem cell fate in Arabidopsisshoot meristem. Cell 95: 805–815.
Menand B, Yi K, Youannic S, et al. 2007. An ancient mechanism controls thedevelopment of cells with a rooting function in land plants. Science 316:1477–1480.
Metcalfe CR. 1967. Distribution of latex in the plant kingdom. EconomicBotany 21: 115–127.
Metcalfe CR. 1971. Anatomy of the monocotyledons. V. Cyperaceae. Oxford:Clarendon Press.
Metcalfe CR. 1987. Anatomy of the dicotyledons. III. Magnoliales, Illiciales,and Laurales. Oxford: Clarendon Press.
Metcalfe CR, Chalk L. 1950a. Anatomy of the dicotyledons. Volume I.Oxford: Clarendon Press.
Metcalfe CR, Chalk L. 1950b. Anatomy of the Dicotyledons. Volume II.Oxford: Clarendon Press.
Meyer CJ, Seago JL Jr, Peterson CA. 2009. Environmental effects on thematuration of the endodermis and multiseriate exodermis of Iris germa-nica roots. Annals of Botany 103: 687–702.
Miwa H, Kinoshita A, Fukuda H, Sawa S. 2009. Plant meristems:CLAVATA3/ESR-related signaling in the shoot apical meristem and theroot apical meristem. Journal of Plant Research 122: 31–39.
Nakajima K, Sena G, Nawy T, Benfey PN. 2001. Intercellular movement ofthe putative transcription factor SHR in root patterning. Nature 413:307–311.
Niki T, Gladish DK. 2001. Changes in growth and structure of pea primaryroots (Pisum sativum L. cv Alaska) as a result of sudden flooding.Plant and Cell Physiology 42: 694–702.
Perumalla CJ, Peterson CA, Enstone DE. 1990. A survey of angiospermspecies to detect hypodermal Casparian bands. I. Roots with a uniseriatehypodermis and epidermis. Botanical Journal of the Linnean Society 103:93–112.
Peterson CA, Perumalla CJ. 1990. A survey of angiosperm species to detecthypodermal Casparian bands. II. Roots with a multiseriate hypodermis orepidermis. Botanical Journal of the Linnean Society 103: 113–125.
Philipson WR. 1990. The significance of apical meristems in the phylogeny ofland plants. Plant Systematics and Evolution 173: 17–38.
Pires ND, Dolan L. 2012. Morphological evolution in land plants: newdesigns with old genes. Philosophical Transactions of the Royal SocietyB: Biological Sciences 367: 508–518.
Popham RA. 1966. Laboratory manual for plant anatomy. St. Louis, MO:C. V. Mosby.
Pysh LD, Wysocka-Diller JW, Camillen C, Bouchez D, Benfey PN. 1999.The GRAS gene family in Arabidopsis: sequence characterization andbasic expression analysis of the SCARECROW-LIKE genes. The PlantJournal 18: 111–119.
Raven JA, Edwards D. 2001. Roots: evolutionary origins and biogeochemicalsignificance. Journal of Experimental Botany 52: 381–401.
Rayner RJ. 1984. New finds of Drepanophycus spinaeformis Goppert fromthe lower Devonian of Scotland. Transactions of the Royal Society ofEdinburgh, Earth Sciences 75: 333–363.
Rudall PJ, Eldridge T, Tratt J, et al. 2009. Seed fertilization, development,and germination in Hydatellaceae (Nymphaeales): implications for endo-sperm evolution in early angiosperms. American Journal of Botany 96:1581–1593.
Rudall PJ, Sokoloff DD, Remizowa MV, et al. 2007. Morphology ofHydatellaceae, an anomalous aquatic family recently recognized as anearly-divergent angiosperm lineage. American Journal of Botany 94:1073–1092.
Scarpella E, Meijer AH. 2004. Pattern formation in the vascular system ofmonocot and dicot plant species. New Phytologist 164: 209–242.
Scarpella E, Rueb S, Boot KSM, Hoge JHC, Meijer AH. 2000. A role forthe rice homeobox gene Oshox1 in provascular cell fate commitment.Development 127: 3655–3669.
Schiefelbein J. 2003. Cell-fate specification in the epidermis: a common pat-terning mechanism in the root and shoot. Current Opinion in PlantBiology 6: 74–78.
Schneider EL, Carlquist S. 1995. Vessel origins in Nymphaeaceae: Euryaleand Victoria. Botanical Journal of the Linnean Society 119: 185–193.
Schneider SL, Carlquist S. 2002. Vessels in Brasenia (Cabombaceae): newperspectives on vessel origin in primary xylem of angiosperms.American Journal of Botany 83: 1236–1240.
Seago JL. 1971. Developmental anatomy in roots of Ipomoea purpurea.I. Radicle and primary root. American Journal of Botany 58: 604–615.
Seago JL. 1973. Developmental anatomy in roots of Ipomoea purpurea. II.Initiation and development of secondary roots. American Journal ofBotany 60: 607–618.
Seago JL Jr 2002. The root cortex of Nymphaeaceae, Cabombaceae andNelumbonaceae. Journal of the Torrey Botanical Society 129: 1–9.
Seago JL Jr, Marsh LC. 1989. Adventitious root development in Typhaglauca, with emphasis on the cortex. American Journal of Botany 76:909–923.
Seago JL Jr, Peterson CA, Enstone DE. 1999a. Cortical ontogeny in roots ofthe aquatic plant, Hydrocharis morsus-ranae L. Canadian Journal ofBotany 77: 113–121.
Seago & Fernando — Anatomical aspects of angiosperm root evolution 237
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022

Seago JL Jr, Peterson CA, Enstone DE, Scholey CA. 1999b. Developmentof the endodermis and hypodermis of Typha glauca Godr. and Typhaangustifolia L. roots. Canadian Journal of Botany 77: 122–134.
Seago JL Jr, Peterson CA, Enstone DE. 2000a. Cortical development inroots of the aquatic plant, Pontederia cordata L. American Journal ofBotany 87: 1116–1127.
Seago JL Jr, Peterson CA, Kinsley LJ, Broderick J. 2000b. Developmentand structure of the root cortex in Caltha palustris L. and Nymphaeaodorata Ait. Annals of Botany 86: 631–640.
Seago JL Jr, Marsh LC, Stevens KJ, Soukup A, Votrubova O, EnstoneDE. 2005. A re-examination of the root cortex in wetlandflowering plants with respect to aerenchyma. Annals of Botany 96:565–579.
Shishkoff N. 1987. Distribution of the dimorphic hypodermis of roots inangiosperm families. Annals of Botany 60: 1–15.
Simon L, Bousquet J, Levesque, LaLonde M. 1993. Origin and diversifica-tion of endomycorrhizal fungi and coincidence with vascular land plants.Nature 363: 67–69.
Soltis DE, Soltis PS, Morgan DR, et al. 1995. Chloroplast gene sequence datasuggest a single origin of the predisposition for symbiotic nitrogen fix-ation in angiosperms. Proceedings of the National Academy ofSciences, USA 92: 2647–2651.
Sottnıkova A, Lux A. 2003. Development, dilation and subdivision of corticallayers of gentian (Gentiana asclepiadea) root. New Phytologist 160:135–143.
Soukup A, Seago JL Jr, Votrubova. 2005. Developmental anatomy of theroot cortex of the basal monocotyledon, Acorus calamus (Acorales,Acoraceae). Annals of Botany 96: 379–385.
Sperry JS. 2003. Evolution of water transport and xylem structure.International Journal of Plant Sciences 164: S115–S127.
Stevens KJ, Peterson RL, Stephenson GR. 1997. Morphological and ana-tomical responses of Lythrum salicaria L. (purple loosestrife) to animposed water gradient. International Journal of Plant Sciences 158:172–183.
Taylor W, DeVore ML, Pigg KB. 2006. Susiea newsalemae gen. et sp. nov.(Nymphaeaceae): Euryale-like seeds from the Late Paleocene Almont
flora, North Dakota, USA. International Journal of Plant Sciences 167:1271–1278.
Thomas EN. 1914. Seedling anatomy of Ranales, Rhoeadales, and Rosales.Annals of Botany 28: 695–733.
Tobe H, Jaffre T, Raven PH. 2000. Embryology of Amborella(Amborellaceae), descriptions and polarity of character states. Journalof Plant Research 113: 271–280.
Tomlinson PB. 1961. Anatomy of the monocotyledons. II. Palmae. Oxford:Clarendon Press.
Tomlinson PB. 1969. Anatomy of the Monocotyledons. III. Commelinales-Zingiberales. Oxford: Clarendon Press.
Tomlinson PB. 1982. Anatomy of the Monocotyledons. VII. Helobiae(Alismatidae). Oxford: Clarendon Press.
Ueda M, Matsiu K, Ishiguro S, et al. 2004. The HALTED ROOT gene encod-ing the 26S proteasome subunit RPT2a is essential for the maintenance ofArabidopsis meristems. Development 131: 2101–2111.
Van der Valk AG. 2012. The biology of freshwater wetlands. Oxford: OxfordUniversity Press.
Van Fleet DS. 1950. A comparison of histochemical and anatomical charac-teristics of the hypodermis with the endodermis in vascular plants.American Journal of Botany 37: 721–725.
Veit B. 2004. Determination of cell fate in apical meristems. Current Opinionin Plant Biology 7: 57–64.
Weaver JE. 1926. Root development of field crops. New York: McGraw-Hill.Wilkinson HP. 2000. A revision of the anatomy of Gunneraceae. Botanical
Journal of the Linnean Society 134: 233–266.Wilson CA, Peterson CA. 1983. Chemical composition of the epidermal,
hypodermal, endodermal and intervening cortical cell walls of variousplant roots. Annals of Botany 51: 759–769.
Wysocka-Diller JW, Helariutta Y, Fukaki H, Malamy JE, Benfey PN.2000. Molecular analysis of SCARECROW function reveals a radial pat-terning mechanism common in root and shoot. Development 127:595–603.
Zankowski PM, Fraser D, Rost TL, Reynolds TL. 1987. The developmentalanatomy of velamen and exodermis in aerial roots of Epidendrum iba-guense. Lindleyana 2: 1–7.
Seago & Fernando — Anatomical aspects of angiosperm root evolution238
Dow
nloaded from https://academ
ic.oup.com/aob/article/112/2/223/162334 by guest on 16 January 2022




















