
Epitope-Guided Engineering of Monobody Binders forin Vivo Inhibition of Erk‑2 Signaling

CLINICAL IMMUNOLOGY AND IMMUNOPATHOLOGY
Vol. 82, No. 2, February, pp. 174–189, 1997Article No. II964290
Analysis of Autoantibodies to T-Cell Receptors among HIV-InfectedIndividuals: Epitope Analysis and Time Course
JOHN J. MARCHALONIS,* NEIL M. AMPEL,† SAMUEL F. SCHLUTER,* ANDREA GARZA,*DOUGLAS F. LAKE,* JOHN N. GALGIANI,† AND WILLIAM J. LANDSPERGER‡
*Department of Microbiology and Immunology, College of Medicine, University of Arizona, Tucson, Arizona 85724; †VeteransAdministration Medical Center, Tucson, Arizona 85723; and ‡Baxter Biotechnology, Hyland Division, Duarte, California 91010
(TCR) Vb chain (5). It was suggested that these autoan-Individuals seropositive for human immunodeficiency tibodies arise from a deregulation of the immune sys-
virus type 1 (HIV) express elevated levels of autoantibod- tem involving the production of autoantibodies againsties (AAbs) directed against recombinant T-cell receptors ‘‘public idiotopes’’ of T-cell receptor variable regions. In(TCRs) and synthetic peptide epitopes duplicating b addition, antibodies reactive with a peptide epitopechain markers. We performed longitudinal studies of from the V3 loop of gp120 of HIV were present, andanti-TCR AAbs in HIV-1-infected individuals, making these cross-reacted with T-cell receptors because ofcomparisons with uninfected sera and sera from other shared sequence and antigenic homology (antigenicindividuals infected with a nonviral agent. We deter-
mimicry) to a sequence defined by the joining segmentmined levels of autoantibodies by titration using en-of the T-cell receptor b chain (5).zyme-linked immunosorbent assay (ELISA) and devel-
Here, we analyze the autoantibody reactivities to theoped a means for characterizing ‘‘autoantibody CDR rec-recombinant TCR Va/Vb construct and peptide epi-ognition spectrotypes’’ for individual sera. Antibodytopes of the Vb domain of ten HIV-seropositive individ-levels against certain defined synthetic epitopes wereuals in a longitudinal study over a period of 2 years.substantially elevated in HIV-infected subjects relativeThe goals of the study were to characterize reactivitiesto reactivities by control groups. Individual sera showedof individual patients and determine the early timerelatively high AAb levels to a subset of CDR1 peptidecourse of anti-TCR autoantibody production in HIV in-epitopes. Two patients who subsequently developed
AIDS showed particular reactivity to Vb2.1, 8.1, 10.1, and fection. Since such autoantibodies are polyclonal (13),22.1 epitopes. Our results show that production AAbs to we devised a procedure to estimate the relative contri-TCR Vb epitopes is a general consequence of HIV infec- bution in each individual of autoantibodies againsttion. The response is individual but shows some restric- products of distinct TCR Vb gene products. To accom-tion and shifts in AAb subpopulations often occur with plish this, we derived an ‘‘autoantibody CDR1 (or FR3)time. q 1997 Academic Press recognition spectrotype’’ for each serum by quantifying
the reactivity against a set of synthetic 16-mer pep-tides, each of which corresponds to the product of a
INTRODUCTION separate Vb gene. We provide evidence that the levelsof IgG autoantibodies to the intact recombinant TCRand the characteristic peptide epitopes are elevated atOne of the major consequences of infection with hu-first analysis of the seropositive individuals, and thatman immunodeficiency virus type 1 (HIV) is the elabo-high autoantibody levels are maintained throughoutration of autoimmune reactivity similar to that foundthe 2-year period. Autoantibody recognition spectro-in graft versus host disease (1) or systemic lupus ery-type analysis disclosed that HIV-infected individualsthematosus (2). In particular, autoimmune reactivityhave individual profiles but that these have commonhas been found against MHC antigens (3, 4), T-cellcharacteristics distinguishing them from either unin-receptors (5, 6), and the Fab fragments of immunoglob-fected, asymptomatic individuals or patients infectedulin (7–9) as well to a number of other autoantigenswith a separate agent, the fungus Coccidioides immitis.(2, 10–12). We recently reported that preparations of
pooled IgG immunoglobulin isolated from the plasmasMATERIALS AND METHODSof HIV-infected individuals contain autoantibodies di-
rected against recombinant T-cell receptor Va/Vb con-structs and defined peptide epitopes corresponding to Human sera and IgG. Sera of clinically healthy in-
dividuals of both sexes in the age range of 20–60 yearsthe CDR1 and FR3 segments of the T-cell receptor
1740090-1229/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
AID Clin 4290 / a50a$$$201 12-20-96 13:57:53 clinas AP: Clin

175AUTOANTIBODIES TO TCR IN HIV INFECTION
TABLE 1
Clinical Parameters of HIV-Seropositive Individuals at Initial and 2-Year Examination
Initial 2 yearsIDNo. CD4 CD8 Status Rx CD4 CD8 Status Rx
1001 398 1030 ASX None 634 963 ASX None1014 550 1364 ASX None 378 1191 ASX None1015 499 883 ASX AZT 299 1197 ASX None1016 406 764 ASX AZT 295 870 ASX AZT1018 211 1816 ARC None 126 2672 AIDS AZT1021 429 2608 ASX None 402 2960 ASX AZT1023 148 451 ASX None 15 387 AIDS DDI1029 880 528 ASX None 740 722 ASX None2010 895 552 ASX None 1132 773 ASX None2013 915 1936 ASX None 667 2158 ASX None
Note. Rx indicates anti-retroviral therapy and status indicates HIV clinical status. ASX, asymptomatic; ARC, AIDS-related complex orsymptomatic HIV without AIDS; and AIDS, AIDS.
were obtained through the Division of Rheumatology, ogy, Medical University of South Carolina (Charleston,SC), and the V3-loop peptide was synthesized manuallyDepartment of Internal Medicine, College of Medicine,
Arizona Health Sciences Center, Tucson. Baseline sera using solid-phase peptide synthesizer (5). All peptideswere synthesized. Purity was determined by bothof untreated individuals infected with the fungus C.
immitis were obtained from the Infectious Diseases amino acid composition and sequence analysis. The pu-rity of the peptides wasú90%, and that of set Aú95%.Clinic at the Tucson VA Medical Center. Sera of 10
HIV-seropositive individuals followed routinely at 4- Intact protein antigens used were ovalbumin as an ex-ample of an irrelevant antigen unrelated to the infec-month intervals for a period of 2 years were obtained
from the HIV–AIDS Clinic at the Tucson VA Medical tion and recombinant single chain T-cell receptor con-sisting of the entire Va and Vb segments of the T-cellCenter. The health status, CD4/ and CD8/ T-cell levels
and treatment at initial presentation and at the two- JURKAT. The ovalbumin was purchased from SigmaChemical Co., and the recombinant single chain T-cellyear follow-up are listed in Table 1. In addition, plasma
pools from healthy uninfected individuals and purified receptor was prepared according to the procedures ofLake et al. (20, 21).IgG (Gammagard) were provided by Baxter Biotech
(Hyland Division, Duarte, CA). Enzyme-linked immunosorbent assay. When affin-ity-purified antibodies to peptides were used, they wereAntigens and antibodies. Sets of synthetic peptides,
including those previously shown to be reactive with prepared as previously described (13, 15) using pep-tides coupled to Sepharose 4B as the solid-phase affin-human autoantibodies (13, 14), as well as a new test set
to delineate individual autoantibody CDR1 recognition ity matrix. ELISAs were carried out essentially as pre-viously described (15, 22, 23) and performed in polysty-spectrotypes (ACRS) were used as test antigens in en-
zyme-linked immunosorbent (ELISA) assays. Table 2, rene microtiter plates. Test antigens were dissolved ata concentration of 10 mg/ml in 0.2 M carbonate buffer,using the single letter abbreviation for amino acids,
lists the set of synthetic peptides corresponding to pH 9.6, and 100 ml/well was used to coat plates. Peptideantigens were coated by drying down the solution over-CDR1 sequences of individual Vb gene products (A), a
set of peptides corresponding to the third framework night at 377C. The plates were washed three times withphosphate-buffered saline, pH 7.4, containing 0.05%of six individual Vb gene products (B), other peptides
corresponding to the joining segment of the TCR b Tween 20 (PBST), followed by quenching with PBSTcontaining 0.1% porcine gelatin (Sigma Chemicals, St.chain (13–18), and a cross-reactive peptide (5) corre-
sponding to the conserved fragment of the V3-loop of Louis, MO) for 30 min. The diluted human sera or Igs(in PBST–gelatin) were reacted with antigen for 1 hr,the IIIB strain of HIV-1 gp120 (19) (C). Set A was syn-
thesized by Chiron Mimetopes Inc. (San Diego, CA); set after which the wells were washed four times withPBST. Peroxidase-conjugated rabbit antibody to hu-B by the Biotechnology Center, University of Arizona
Research Laboratories (Tucson, AZ); the Jb by Penin- man IgM or human IgG was used as a developing re-agent (Dako Corp., Carpenteria, CA). This conjugatesula Laboratories (San Francisco, CA), the b10 peptide
by the Department of Biochemistry and Molecular Biol- was used at a dilution of 1:1000 for the anti-IgM and
AID Clin 4290 / a50a$$$202 12-20-96 13:57:53 clinas AP: Clin

176 MARCHALONIS ET AL.
TABLE 2 druplicate replicates were assayed at a serum dilutionof 1:200 in PBST–gelatin. The third quantitative ap-Synthetic Peptides Used in Studyproach was use of a ‘‘modified sum absorbance’’ which
(A) Synthetic Vb CDR1 epitopes utilized all of the data points in a titration by calculat-ing the sum of the absorbencies at each dilution andVb2.1 C R S L D F Q A T T M F W Y G Q Fcorrecting for nonspecificity by subtracting the sum ofVb3.1 C V Q D M D H E N M F W Y R Q D
Vb4.1 C Q V D S Q V T M M F W Y R Q Q the values of the absorbencies of binding of the sameVb5.1 C S P R S G D L S V Y W Y Q Q S serum or antibody preparation to uncoded PBST-Vb5.2 C S P K S G H D I V S W Y Q Q A blocked gelatin plates (24). This approach has the valueVb6.1 C D P I S G H T A L Y W Y R Q S
of using all of the data and is useful for estimatingVb6.5 C D P I S E H N R L Y W Y R Q Tvb7.1 C E Q H M G H R A M Y W Y K Q A small specific differences from background.Vb8.1 C K P I S G H N S L F W Y R Q T In order to determine the profile of autoantibody sub-Vb9.2 C E Q N L G H D T M Y W Y K Q D sets within individual sera or antibody preparations,Vb10.1 C V P I K A H S Y V Y W Y R K K we constructed sets of synthetic peptides correspondingVb11.1 C S Q T M G H D K M Y W Y Q Q D
to CDR1 (Table 2A) and FR3 (of individual Vb geneVb12.1 C H Q T E N H R Y M Y W Y R Q DVb13.1 C A Q D M N H E Y M Y W Y R Q D products). The assays were performed by two methods.Vb14.1 C S Q N M N H E Y M S W Y R Q D The first was to determine the quantitative responseVb15.1 C S Q T K G H D R M Y W Y R Q D to the individual ‘‘public idiotype.’’ This was done byVb16.1 C D P I S G H D H L Y W Y R R V
determining titer, absorbancy at a particular dilutionVb17.1 C E Q N L N H D A M Y W Y R Q Dor the ‘‘modified sum absorbance.’’ The second applica-Vb19.1 C T P E K G H T F V Y W Y Q Q N
Vb20.1 C T V E G T S N P N L Y W Y R Q A tion of ACRS was to determine the relative contribu-Vb21.1 C D P I S G H A T L Y W Y R Q I tions of the individual subsets of autoantibodies. ThisVb22.1 C V P I S G H S H V Y W Y R Q L was done in the case of the Vb CDR1 homolog set byVb23.1 C Y P I P R H D T V Y W Y Q Q G
taking the value of the autoantibody binding to theVb24.1 C S Q T L N H N V M Y W Y Q Q KVb8.1 peptide as 1.0 and normalizing the other valuesrelative to it. We chose the Vb8.1-CDR1 peptide as the(B) Synthetic Vb FR3 epitopesreferrant because it is the original one used in prior
Vb2.1 T V T S A H P E D S S F Y I C SVb5.2 N L S S L E L G D S A L Y F C AVb6.3 K I Q R T Q Q E D S A V Y L C AVb8.1 K I Q P S E P R D S A V Y F C AVb12.1 T L E S L P A P E T S V Y F C AVb17.1 T V T S A Q K N P I A F Y L C A
Other peptides
pepJb A N Y G Y T F G S G T R L T V Vpepb10 S A N Y G Y T F G S G T R L T VpepV3(HIV) R I H I Q R G P G * R A F Y T T K
1:4000 for the anti-IgG and was incubated in the wellsfor 1 hr. After five washings with PBST, 0.03% 2,2 *-azino-bis(3-ethyl-benzthioazoline-6-sulfonic acid)(Sigma Chemical Co.) in 0.1 M citrate buffer, pH 4.0,0.01% hydrogen peroxide was added. The absorbancesat 405 nm were measured in a Titertek Multiskan(Flow Laboratories), usually following 60 min of incu-bation at room temperature.
Titrations were carried out in duplicate beginningwith the human serum diluted 1:50 with PBST followed FIG. 1. Comparison of individual values for IgG autoantibodyby twofold serial dilutions. Titer was defined as the binding of 10 HIV/ patients and 22 healthy uninfected individuals
to the scTCR. Individual sera were assayed in triplicate by ELISAreciprocal dilution at which the ELISA absorbance hadat a dilution of 1/200. ‘‘Blank’’ values of binding of each serum toa value of 0.50 under standard conditions. It was ob-pig gelatin were subtracted from each value. The means of the twotained by numerical interpolation between the two di- populations (normals, 0.06 { 0.02; HIV/, 0.62 { 0.09) differ signifi-
lution values bracketing the end point. A second quan- cantly with P õ 0.01. The means of the ELISA absorbance are indi-cated by arrows.titative approach was a one-step assay in which qua-
AID Clin 4290 / a50a$$$202 12-20-96 13:57:53 clinas AP: Clin

177AUTOANTIBODIES TO TCR IN HIV INFECTION
studies (13, 14) and is frequently the major bindingspecificity both in human and murine (34) sera. Thesame procedure was used with the Vb FR3 homologset with the binding to the Vb8.1 set defined as equalto 1.0.
RESULTS
Autoantibodies to recombinant scTCR in sera of indi-vidual HIV-infected patients. The sera from HIV-in-fected patients at presentation were tested for IgG au-toantibodies to the recombinant single chain T-cell re-ceptor (scTCR) and the results were compared withthose of a group of uninfected, asymptomatic controlindividuals (Fig. 1). Individual sera were assayed byELISA at a dilution of 1/200. ‘‘Blank’’ values of bindingof each serum to pig gelatin were subtracted from eachdetermination. The means of the two populations (con-trols, 0.06 { 0.02; HIV-infected, 0.62 { 0.09) differedsignificantly (P õ 0.01). These results supports previ-ous findings comparing purified IgG immunoglobulinpools from HIV-infected and uninfected individuals (5).
Since sera were available from six time points at 4-month intervals for each patient, it was possible todetermine a time course of autoantibody binding to therecombinant single chain T-cell receptor (Fig. 2). Tomonitor status of antibodies against HIV, we deter-mined the time course for appearance of antibodies tothe synthetic V3 loop peptide of HIV gp120. As amarker for an irrelevant antibody specificity, we as-sayed binding to ovalbumin. The kinetic analysis of
FIG. 3. Titrations of sera of coccidiomycosis patients, HIV/ indi-viduals, and asymptomatic individuals to the CDR1 peptide (pepb3,upper figure) and to the FR3 peptide (pepb8, lower figure). Datagiven are means { SE. Data for the HIV/ individuals were obtainedat the original examination.
the average of all HIV-infected patients shows that theantibody level to the viral peptide stays high throughthe course of this study. The autoantibodies to thescTCR are substantial at the beginning of the responseand have a tendency to move upwards toward the end.The bulk average response to ovalbumin is lower than
FIG. 2. Time course of response of HIV seropositive individuals the other two and tends to drift downward. It is appar-to the irrelevant antigen ovalbumin, to the V3 loop peptide of gp120 ent that the antibodies directed against the viral V3of HIV-1, and to the recombinant single chain TCR. Data given are loop peptide are the highest and show little variationmeans { SE of the combined ELISA absorbance of the patients at a
during this time course. Considerable variations aredilution of 1/200 at each time point. (j) Binding to ovalbumin; (l)binding to pepV3; (m) binding to recombinant single chain TCR. shown with respect to the irrelevant antigen which do
AID Clin 4290 / a50a$$$202 12-20-96 13:57:53 clinas AP: Clin
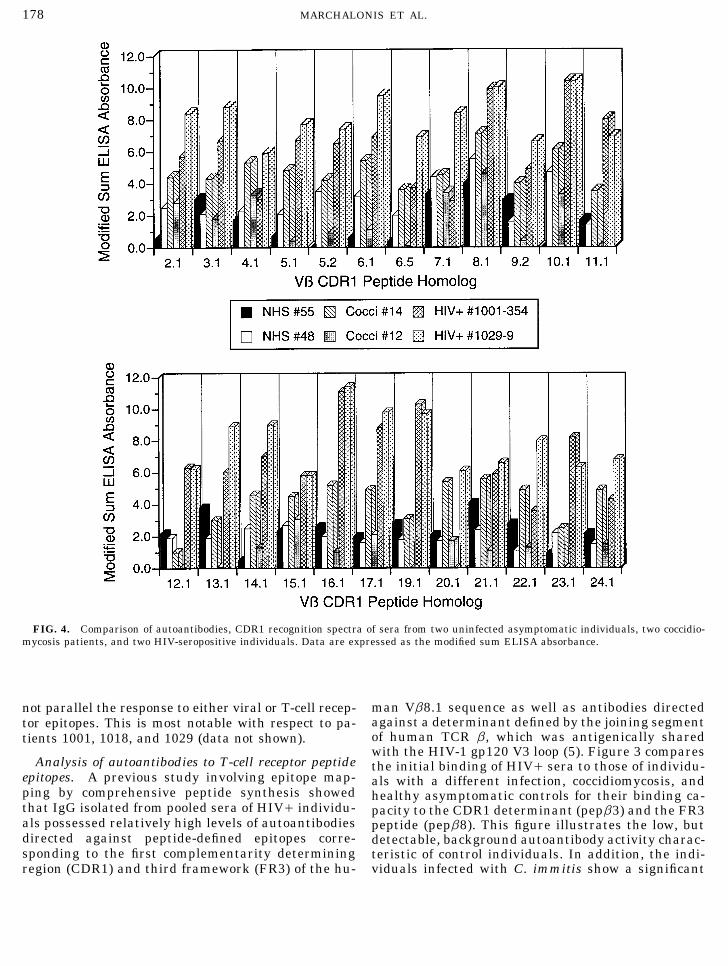
178 MARCHALONIS ET AL.
FIG. 4. Comparison of autoantibodies, CDR1 recognition spectra of sera from two uninfected asymptomatic individuals, two coccidio-mycosis patients, and two HIV-seropositive individuals. Data are expressed as the modified sum ELISA absorbance.
man Vb8.1 sequence as well as antibodies directednot parallel the response to either viral or T-cell recep-against a determinant defined by the joining segmenttor epitopes. This is most notable with respect to pa-of human TCR b, which was antigenically sharedtients 1001, 1018, and 1029 (data not shown).with the HIV-1 gp120 V3 loop (5). Figure 3 compares
Analysis of autoantibodies to T-cell receptor peptide the initial binding of HIV/ sera to those of individu-epitopes. A previous study involving epitope map- als with a different infection, coccidiomycosis, andping by comprehensive peptide synthesis showed healthy asymptomatic controls for their binding ca-that IgG isolated from pooled sera of HIV/ individu- pacity to the CDR1 determinant (pepb3) and the FR3als possessed relatively high levels of autoantibodies peptide (pepb8). This figure illustrates the low, butdirected against peptide-defined epitopes corre- detectable, background autoantibody activity charac-sponding to the first complementarity determining teristic of control individuals. In addition, the indi-
viduals infected with C. immitis show a significantregion (CDR1) and third framework (FR3) of the hu-
AID Clin 4290 / a50a$$$202 12-20-96 13:57:53 clinas AP: Clin

179AUTOANTIBODIES TO TCR IN HIV INFECTION
elevation in binding to both peptides relative to that epitopes duplicating the CDR1 segments of 24 individ-shown by the normal controls. The autoantibody lev- ual human Vb genes. All of these analyses were per-els characteristic of the HIV-infected individuals are formed in the same trial to ensure accurate compari-substantially higher, however, than those of either son. The comparison confirms the previous quantita-group. tive differences and shows that sera of HIV patients
contain autoantibodies against a number of the individ-Analysis of autoantibody CDR1 recognition spectro-ual epitopes. Taking the modified sum ELISA ab-types. The preceding data establish that HIV-infectedsorbance equal to 5 as a point of reference, 19 individ-individuals have high levels of autoantibodies directedual epitopes showed binding of at least of this level.against Vb-CDR1 and FR3 peptide-defined epitopes atThe highest activities found in these HIV/ sera werepresentation, and that these levels remain highdirected against Vb8.1, Vb10.1, Vb16.1, and Vb19.1.throughout the time course of this study. To determineThe only autoantibody reactivities in the normal serawhether shifts in populations of autoantibodies di-to reach the level of 5.0 was antiVb8.1, although activ-rected against particular Vb region epitopes variedity was noted against Vb8.1, Vb13.1, and Vb21.1. Thewith time, we devised a means to determine autoanti-levels of autoantibodies in the coccidiomycosis-infectedbody CDR1 or FR3 recognition spectrotypes. In thisindividuals were lower than those of the HIV/ pa-procedure, we synthesized peptides corresponding totients, but reactivity §5.0 was observed to the Vb8.1the CDR1 segments of 24 individual human Vb germ-peptide and to Vb4.1, 5.1, 6.1, 10.1, and 15.1.line genes (see Table 1) and measured at the same
In order to diminish the contribution of differencestime, under the same conditions, the binding spectrain quantitative levels and different bleeds and obtainof autoantibodies of individual patients. Figure 4 com-a spectrotype reflecting subpopulations of antibodies ofpares autoantibody binding of two asymptomatic indi-different specificities, we normalized the binding toviduals (NHS48 and 55), two coccidiomycosis patientsthat of the Vb8.1 peptide as shown in Table 3. Since(patients 12 and 14) and two HIV/ individuals (pa-
tients 1001 and 1029) to the panel of synthetic peptide the reactivity to this peptide is generally the highest
TABLE 3
Normalized Autoantibody CDR1 Recognition Spectrotype Analysis
HIV/
Vb Epitope Normals (12) Cocci (19) Initial (8) 2 years (8)
2.1 0.48 { 0.09 0.67 { 0.17 0.69 { 0.18 0.81 { 0.213.1 0.56 { 0.10 0.67 { 0.05 0.54 { 0.18 0.82 { 0.274.1 0.39 { 0.07 0.61 { 0.09 0.54 { 0.09 0.47 { 0.135.1 0.38 { 0.07 0.34 { 0.07 0.36 { 0.10 0.57 { 0.215.2 0.43 { 0.10 0.51 { 0.07 0.40 { 0.14 0.60 { 0.206.1 0.47 { 0.11 0.36 { 0.06 0.49 { 0.15 0.69 { 0.246.5 0.40 { 0.10 0.30 { 0.05 0.34 { 0.14 0.47 { 0.227.1 0.70 { 0.06 1.01 { 0.21 0.42 { 0.16 0.60 { 0.208.1 1.00 1.00 1.00 1.009.2 0.47 { 0.09 0.41 { 0.07 0.51 { 0.17 0.66 { 0.26
10.1 0.91 { 0.22 1.19 { 0.25 0.85 { 0.14 1.32 { 0.2811.1 0.31 { 0.06 0.28 { 0.06 0.44 { 0.17 0.90 { 0.2912.1 0.59 { 0.06 0.24 { 0.06 0.36 { 0.12 0.82 { 0.2813.1 0.64 { 0.09 0.35 { 0.08 0.43 { 0.15 0.82 { 0.2714.1 0.36 { 0.06 0.54 { 0.09 0.59 { 0.15 0.85 { 0.2915.1 0.58 { 0.06 0.68 { 0.12 1.20 { 0.32 1.36 { 0.2916.1 0.68 { 0.06 0.51 { 0.07 0.91 { 0.15 1.41 { 0.3517.1 0.53 { 0.07 0.46 { 0.06 0.62 { 0.16 0.96 { 0.3519.1 0.66 { 0.08 0.35 { 0.05 0.65 { 0.16 1.07 { 0.3820.1 0.43 { 0.05 0.36 { 0.06 0.42 { 0.14 0.51 { 0.2321.1 0.67 { 0.09 0.50 { 0.07 0.49 { 0.20 0.87 { 0.3222.1 0.55 { 0.08 0.39 { 0.07 0.49 { 0.17 0.82 { 0.2523.1 0.45 { 0.09 0.25 { 0.05 0.38 { 0.12 0.82 { 0.3324.1 0.43 { 0.09 0.31 { 0.07 0.47 { 0.15 0.84 { 0.20
Note. Shaded values differ significantly from corresponding values for the normal population at the P õ 0.05 level by Student’s t test.
AID Clin 4290 / a50a$$$202 12-20-96 13:57:53 clinas AP: Clin

180 MARCHALONIS ET AL.
FIG. 5. Comparison of ‘‘bulk’’ (average values) of normalized autoantibodies CDR1 reactivity spectrotypes for uninfected asymptomaticindividuals, coccidiomycosis patients, and HIV-seropositive individuals at first presentation and after 2 years. A comparison is made withaffinity purified antibody (AP GG anti-b) to the Vb8.1 CDR1 peptide (pepb3) affinity-purified from intravenous immunoglobulin.
irrespective of absolute titer or modified sum ELISA asymptomatic and HIV-infected sets of individuals byshowing reactivity against the Vb7.1 epitope equal toabsorbance, it was appropriate to express binding to
the other individual CDR1 epitopes relative to this that directed against the Vb8.1 standard. The coccidio-mycosis patients, in addition, have decreased relativestandard for comparison. This table presents the popu-
lation means for normal individuals and coccidiomy- binding to Vb12.1 and 19.1 epitopes by comparisonwith the normal population. The coccidiomycosis- andcosis- and HIV-infected patients, giving means { stan-
dard error. Consistent with previous studies (5, 6), re- HIV-infected individuals showed elevated binding tothe Vb10.1 epitope, and the HIV-infected individualsactivity to the Vb8.1 CDR1 peptide (known as b3 or
pepb3 in previous publications) was higher in control had particular elevations to Vb2.1, 11.1, 14.1, 15.1,16.1, 17.1, and 19.1. It is of particular interest thatsubjects than that to any of the other epitopes. Some
reactivity, however, was observed to all members of the certain of these showed major increases at 2 years rela-tive to the initial value, e.g., autoantibodies to CDR1set. The mean values of this data set are plotted in Fig.
5. The coccidiomycosis patients differ from both the of Vb11.1, 16.1, 17.1, 19.1, 21.1, 22.1, 23.1, and 24.1.
AID Clin 4290 / a50a$$$202 12-20-96 13:57:53 clinas AP: Clin

181AUTOANTIBODIES TO TCR IN HIV INFECTION
FIG. 6. Autoantibody CDR1 recognition spectrotype of patient 1016 comparing values at initial presentation and at 2 years. (Top) Dataas binding titer. (Bottom) Data normalized with the reactivity with Vb8.1 CDR1 defined as 1.0.
Overall, it is possible to recognize distinctive trends of antibody to Vb5.1 which increases substantially. Thenormalized spectrotype does not indicate a substantialin spectrotype among the distinct pools of individuals.
Affinity-purified IgG autoantibody to the b3 peptide population shift. Sera of patient 1029 (Fig. 7) showstrong activities against Vb2.1, 3.1, 6.1, 8.1, 9.1, 10.1,(13, 25) from intravenous immunoglobulin is shown for
comparison. The major activity is against the Vb8.1 11.1, 13.1, 14.1, 16.1, 17.1, 19.1, 21.1, and 22.1. Bindingto Vb epitopes 3.1, 6.1, 11.1, and 21.1 increase withpeptide with detectable cross-reactivity against Vb5.2,
Vb6.5, Vb9.2, and Vb11.1. time, while binding to 10.1 shows a relative decrease.The autoanti TCR profile for patient 2010 (Fig. 8).The anti-CDR1 spectrotype analysis of sera of four
individual HIV/ patients comparing values at initial shows major binding to Vb peptide epitopes 2.1, 3.1,6.1, 7.1, 8.1, 10.1, 11.1, 16.1, 19.1, 21.1, and 22.1. Thepresentation and at the 2-year follow-up are illustrated
in Figs. 6 through 9. The upper figure in each gives independence of particular constituent autoantibodiesis indicated by the fact that relative levels of autoanti-titers for the binding to particular Vb CDR1 synthetic
epitopes and the lower figure is normalized with reac- bodies to Vb2.1, 3.1, 6.1, 11.1, 12.1, 16.1, and 17.1 in-crease by the end of the 2-year period. By contrast,tivity to the Vb8.1 CDR1 epitope defined as 1.0. The
major binding specificities in the serum of patient 1016 binding to Vb8.1 and 21.1 is essentially unchanged andthat to Vb10.1 has decreased. Autoantibodies in the(Fig. 6) are directed against Vb3.1, 5.1, 8.1, 9.1, 10.1,
13.1, 16.1, 19.1, 21.1, 22.1, and 23.1. There is a ten- two bleeds of patient 1018 (Fig. 9) show a relativelysimplified pattern with extremely high titers directeddency for titers against the majority of these to drop
during the 2-year period follow-up with the exception against Vb8.1, 10.1, and 22.1 at the second bleed. By
AID Clin 4290 / a50a$$$202 12-20-96 13:57:53 clinas AP: Clin

182 MARCHALONIS ET AL.
FIG. 7. Autoantibody CDR1 recognition spectratype of patient 1029 comparing values at initial presentation and at 2 years. (Top) Dataas binding titer. (Bottom) Data normalized with the reactivity with Vb8.1 CDR1 defined as 1.0.
comparison with the other three sera, substantial titers seropositive individuals, likewise, produce autoanti-bodies directed against the entire set, but the titers areare present against Vb2.1, 3.1, 4.1, and 24.1. The nor-
malized spectrotype analysis shows that the activities much higher. We constructed a normalized spectrotypeanalysis (Fig. 12) by comparing absorbance values foragainst Vb8.1, 10.1, and 22.1 retain their relative con-
tributions, but that relative binding activity to Vb2.1, the normal plasma at a dilution of 1:200 and ab-sorbance values for the coccidiomycosis patients at a3.1, 4.1, 13.1, 14.1, 16.1, 17.1, 21.1, and 24.1 show sub-
stant relative decreases. dilution of 1/3200 and HIV/ patients at a dilution of1:25,600. Consistent with our above studies of the VbAnalysis of binding to theVbFR3 homolog set. Hexa- CDR1 homolog set, there is individual variation as indi-decamers corresponding to the original framework 3 cated by the standard errors, but the HIV/ individualspeptide recognized by human autoantibodies (‘‘b8’’; 13, have relatively elevated levels of autoantibody to the14) were synthesized, and the autoantibody spectrum Vb2.1, 5.2, and 17.1 peptides by comparison with bothto the set determined for normal plasma and pooled the normal plasma and the coccidiomycosis patients.immunoglobulin, sera of patients with coccidiomycosis,
and sera of HIV/ individuals was determined (Figs.DISCUSSION10 and 11). As illustrated in Fig. 10, the normal prepa-
rations contain low levels of autoantibodies to the en-tire set, and these are increased in the patients infected Clonospecific autoantibodies to human TCRs have
been found in allograft transplantation (26). Autoanti-with coccidiomycosis. As shown in Fig. 11, the HIV-
AID Clin 4290 / a50a$$$202 12-20-96 13:57:53 clinas AP: Clin

183AUTOANTIBODIES TO TCR IN HIV INFECTION
FIG. 8. Autoantibody CDR1 recognition spectratype of patient 2010 comparing values at initial presentation and at 2 years. (Top) Dataas binding titer. (Bottom) Data normalized with the reactivity with Vb8.1 CDR1 defined as 1.0.
bodies to recombinant Va/Vb constructs (14, 27) and Vb-FR3 epitopes and to peptides expressing a mimicrydeterminant shared between the V3 loop of HIV-1Vb-peptide-defined epitopes occur in asymptomatic in-
dividuals (13, 14) and in normal physiological processes gp120 and the joining (Jb) segment of human TCR. Wefollowed binding of naturally occurring IgG antibodiesincluding aging (22) and pregnancy (28). Elevated lev-
els of such autoantibodies appear to vary in autoim- to ovalbumin as an irrelevant antigen because approxi-mately 70% of our Tucson population expresses suchmune diseases (14, 22, 25) and in retroviral infection
(5, 6). Analyses involving immune affinity purification antibodies (14). Although it might be expected that lev-els of antibodies to this common dietary antigen mightand inhibition with peptides (13, 14) as well as isolation
of monoclonal anti-TCR autoantibodies from EBV- reflect the state of health of the patients immune sys-tem, no correlation was found with the levels of anti-transformed human B cell lines (29) have established
that these represent populations of specific IgM and bodies to the HIV peptide or with autoantibodies toTCR. The anti V3/Jb epitope is an example of trueIgG antibodies. Here, we show that elevated levels of
IgG autoantibodies to TCR are a characteristic serum antigenic mimicry between an infectious agent and ahost protein because binding of affinity-purified anti-component of HIV-infected humans.
We report that HIV-infected individuals have high Jb and anti-V3 to their respective ligands is cross-in-hibited by either peptide and the actual immunogen inlevels of anti-TCR autoantibodies at first examination
and that elevated levels persist through 2 years of clini- humans is the viral peptide (5). It is of interest thatwe have not been able to induce antibodies to this pep-cal follow-up. This conclusion held for autoantibodies
to the rscTCR, to peptides duplicating Vb-CDR1 and tide by immunization of mice (30), and antibodies of
AID Clin 4290 / a50a$$$202 12-20-96 13:57:53 clinas AP: Clin

184 MARCHALONIS ET AL.
FIG. 9. Autoantibody CDR1 recognition spectratype of patient 1018 comparing values at initial presentation and at 2 years. (Top) Dataas binding titer. (Bottom) Data normalized with the reactivity with Vb8.1 CDR1 defined as 1.0.
this specificity do not occur in nonretrovirally infected tide to illustrate the relative contributions of subsetsof autoantibodies of distinct Vb-CDR1 peptide resultshumans (14). Rabbits by contrast, produce high titer
antibodies to conjugates of the Jb peptide (31) which from an unusual process of immunoregulation becauseall mammals tested including humans (14), mice (34),bind to human T-cells (32) and can block in vitro infec-
tion of T-cells by live HIV (J. J. Marchalonis, S. F. rabbits, and lower primates (J. J. Marchalonis and A.Garza, unpublished observations) have naturally oc-Schluter, and R. T. Kennedy, unpublished observa-
tions). curring autoantibodies. These peptides lack ostensivehomology to viral peptides. Although rabbits are excel-The Vb-CDR1 epitopes are defined completely by the
individual Vb gene and can, thus, be considered ‘‘public lent antibody producers to most peptides in our compre-hensive peptide epitope mapping set (14, 20), immuni-idiotopes’’ (14, 33). We have used this property to define
autoantibody CDR1 recognition spectrotypes by con- zation with the Vb8.1 CDR1 peptide does not generateincreased antibody, but can cause diminution in titerstructing a set of homologous peptides, each of which
corresponds to the product of an individual Vb gene, (25). Consequently, it is noteworthy that retroviral in-fections in humans (5, 6, this paper) and mice (34)and determining the autoantibody profiles of individual
and sets of patients. The assays were performed by brings about major increases in autoantibodies toVbCDR1 epitopes.ELISA and incorporated both quantitative levels based
upon titers or modified sum absorbancy and normal- The ACRS approach enables us to construct charac-teristic spectrotypes for populations of ASX individualsized comparisons with the reactivity to the Vb8.1 pep-
AID Clin 4290 / a50a$$$202 12-20-96 13:57:53 clinas AP: Clin

185AUTOANTIBODIES TO TCR IN HIV INFECTION
FIG. 10. Titrations of purified normal intravenous immunoglobulin, pooled plasma from asymptomatic individuals, and three coccidio-mycosis patients to the panel of peptides representing the FR3 epitope of Vb8.1, 2.1, 5.2, 6.3, 12.1, and 17.1.
and patients infected either with coccidiomycosis or differences were found with respect to relative bindingto distinct CDR1 epitopes; for example, the normalizedHIV infection. Furthermore, we obtained spectra for
individuals and compare the same HIV-infected pa- profile of coccidiomycosis patients was comparable tothat of normals with the exception of relatively de-tients over a 2-year period. Quantitatively, both groups
of infected patients showed an increase in the level of creased levels of autoantibodies to Vb12.1 and Vb19.1.By contrast, the HIV-infected populations initiallyautoantibodies relative to the normal set, but the HIV-
infected group had considerably higher levels than did showed relative elevations to Vb14.1 and 15.1, and,after 2 years, Vb2.1, 11.1, 14.1, 15.1, 16.1, 17.1, 19.1,the fungally infected group. In addition, characteristic
AID Clin 4290 / a50a$$$203 12-20-96 13:57:53 clinas AP: Clin

186 MARCHALONIS ET AL.
FIG. 11. Titrations showing the binding of individual HIV-seropositive patients taken at the 2-year bleed to the set of FR3 homologs.The arrows in Figs. 10 and 11 indicate the dilutions at which absorbancies were used to calculate normalized autoantibody spectrotypes.
23.1, and 24.1. The population data and individual pa- fered in the two distinct infections. In terms of ACRScomparison, the HIV-infected group showed decreasedtient analyses were not consistent with a simple pro-
cess of polyclonal activation by the virus, because there levels of autoantibodies to the Vb7.1 epitope, but mark-edly raised levels to the Vb11.1, 12.1, 16.1, 19.1, andwas a selectivity in the response in which some autoan-
tibodies declined whereas others increased. The com- 23.1 epitopes. The distinctions between the two in-fected groups were most prominent in the comparisonparison between the coccidiomycosis- and HIV-infected
groups showed that the quantitative and qualitative with the 2-year HIV-infected patient assays. Our ACRSapproach to evaluating an overall profile of autoanti-features of autoantibodies to the CDR1 epitopes dif-
AID Clin 4290 / a50a$$$203 12-20-96 13:57:53 clinas AP: Clin

187AUTOANTIBODIES TO TCR IN HIV INFECTION
creases were observed in reactions to Vb5.1, 9.1, 11.,16.1, and 21.1 in some individuals. Furthermore, pa-tient 2010 showed decreases in titer to Vb8.1 and 10.1coupled with marked increases to Vb2.1, 3.1, 11.1, 12.1,13.1, 16.1, and 17.1.
It will be necessary to obtain sera from individualsprior to infection and over time during the variousstages of disease in order to obtain a complete under-standing of the correlation of anti-TCR autoantibodyproduction with retroviral infection and immunopatho-genesis of HIV infection. The present study shows thatgeneration of autoantibodies to TCR public idiotopes isa major consequence of infection with HIV, and thatthere is a selection for autoantibodies directed againstsubsets of Vb gene products. Whether this is a conse-quence of autoimmunization to Vb epitopes on T-cellsubsets amplified by putative HIV superantigens (37–
FIG. 12. Comparison of normal plasma pool with the individual 39) or of other mechanisms for generating autoantibod-coccidiomycosis and HIV-seropositive patients showing normalizedies to autologous TCR remains to be determined. With-spectrotypes to the Vb FR3 epitopes. (h) Normal plasma pool; (l)out preinfection baseline data for individuals, we can-coccidiomycosis; (l) HIV-seropositive patient. In the two patient
groups, the mean value is indicated by the R. not determine whether a population shift in specifici-ties occurred consequent to the infections. However,the amount of antibody as indicated by titers increasessubstantially relative to those of uninfected individualsbody reactivity is based upon binding to sets of homolo-
gous synthetic peptides corresponding to individual Vb and internal studies of serial bleeds of infected individ-uals indicates that relative increases and decreasesgene products. Recent studies by others have used
quantitative immunoblotting to analyze the multiple and population shifts can occur following infection.V-region interactions among normal human IgG (35)
ACKNOWLEDGMENTSand its repertoire for autologous protein tissue anti-gens (36). These approaches parallel the present one
This research was supported in part by grants from Arizona Dis-in demonstrating the capacity to recognize distinct pro-ease Control Research Commission, Baxter Biotechnology/Hylandfiles of polyclonal autoantibodies to Ig V-region deter- Division to J.J.M., and research supported in part by the U.S. Office
minants (35). of Veterans Affairs to N.A. and J.G.Although 8 of the 10 HIV-seropositive individuals
followed were healthy during the full time course, one REFERENCESpatient presented with ARC and progressed to AIDS
1. Shearer, G. M., Allogeneic leukocytes as a possible factor in in-by the last examination. The individual spectrotypeduction of aids in homosexual men. N. Engl. J. Med. 308, 223–comparison shows an overall increase in titer for most224, 1983.autoantibody specificities, but a simplification of the
2. Silvestris, F., Williams, R. C., Jr., and Dammacco, F., Autoreac-profile because of exceptional increases in binding of tivity in HIV-1 infection: The role of molecular mimicry. Clin.Vb8.1, 10.1, and 22.1. Thus, certain autoantibody lev- Immunol. Immunopathol. 75, 197–205, 1995.els such as that to Vb16.1 have increased in titer from 3. Golding, H., Shearer, G. M., Hillman, K., Lucas, P., Manischew-6393 to 7162 but decreased in relative contribution itz, P., Zajac, R. A., Clerici, M., Gress, R. E., Boswell, R. N., and
Golding, B., Common epitope in human immunodeficiency virusfrom 0.92 to 0.36. Moreover, although the titer to(HIV)1-gp41 and HLA Class II elicits immunosuppressive auto-Vb11.1 more than doubled from 2340 to 5144, the rela-antibodies capable of contributing to immune dysfunction intive values declined from 0.34 to 0.26. By contrast, au- HIV-I-infected individuals. J. Clin. Invest. 83, 1430–1432, 1989.
toantibody titers to Vb8.1, Vb10.1, and Vb22.1 epi- 4. Golding, H., Robey, F. A., Gates, F. T., Linder, W., Beining, P. R.,topes each increased threefold and thus maintained Hoffman, T., and Golding, B., Identification of homologous re-their relative positions. A second patient was asymp- gions in human immunodeficiency virus I gp41 and human MHC
class II b1 domain. I. Monoclonal antibodies against the gp41-tomatic at presentation but developed AIDS at 2 years.derived peptide and patients’ sera react with native HLA ClassThe autoantibody CDR1 spectrotype of this individual,II antigens, suggesting a role for autoimmunity in the pathogen-likewise, showed major reactivities for pepVb8.1, 10.1, esis of acquired immune deficiency syndrome. J. Exp. Med. 167,
and 22.1 as well as relatively high binding to pepVb2.1. 914, 1988.In general, autoantibodies to Vb8.1 and 10.1 made 5. Lake, D. F., Schluter, S. F., Wang, E., Bernstein, R. M., Edmund-
son, A. B., and Marchalonis, J. J., Autoantibodies to the a/b T-the largest relative contributions, but substantial in-
AID Clin 4290 / a50a$$$203 12-20-96 13:57:53 clinas AP: Clin

188 MARCHALONIS ET AL.
cell receptors in human immunodeficiency virus (HIV) infection: 20. Lake, D. F., Bernstein, R. M., Hersh, E. M., Kaymaz, H.,Schluter, S. F., and Marchalonis, J. J., Construction and serolog-Dysregulation and mimicry. Proc. Natl. Acad. Sci. USA 91,
10849–10853. ical characterization of a recombinant human single chain T-cellreceptor (scTCR). Biochem. Biophys. Res. Commun. 201, 1502–6. Marchalonis, J. J., Lake, D. F., Schluter, S. F., Dehghanpisheh,1509, 1994.K., Watson, R. R., Ampel, N. M., and Galgiani, J. N., In ‘‘Immu-
21. Lake, D. F., Bernstein, R. M., Schluter, S. F., and Marchalonis,nobiology of Proteins and Peptides’’ (M. Z. Atassi, Ed.), Vol. VIII,J. J., A method for generating diverse single chain proteins usingp. 211, Plenum, New York, 1995.a universal (Gly4-ser)3 encoding oligonucleotide. Biotechniques7. Susal, C., Daniel, V., Oberg, H. H., Terness, P., Huth-Kuhne, A.,19, 700–702, 1995.Zimmermann, R., and Opelz, G., Striking inverse association of
22. Marchalonis, J. J., Schluter, S. F., Wilson, L., Yocum, D. E.,IgG anti-Faby antibodies and CD4 counts in patients with ac-Boyer, J. R., and Kay, M. M. B., Natural human antibodies toquired immunodeficiency syndrome (AIDS)/AIDS-related com-synthetic peptide autoantigens: Correlations with age and auto-plex. Blood 79, 954–957, 1992.immune disease. Gerontology 39, 65–79, 1993.8. Silvestris, F., Di Loreto, M., Romito, A., Grizzuti, M. A., and
23. Marchalonis, J. J., Dedeo,glu. F., Kaymaz, H., Schluter, S. F.,Dammacco, F., Distribution and antigenic analysis of circulatingand Edmundson, A. B., Antigenic mapping of a human l lightF(ab*)2-reactive IgG in patients with HIV-1 infection. Clin. Im-chain: Correlation with 3-dimensional structure. J. Prot. Chem.munol. Immunopathol. 73, 229–234, 1994.11, 129–137, 1992.9. Susal, C., Daniel, V., and Opelz, G., Does AIDS emerge from a
24. Rosenshein, I. L., and Marchalonis, J. J., Broad phylogenetic ex-disequilibrium between two complementary groups of moleculespression of Heavy-chain determinants detected by rabbit anti-that mimic MHC? Immunol. Today 17, 114–119, 1996.sera to VHa allotypes. Mol. Immunol. 22, 1177–1185, 1985.10. Ditzel, H. J., Barbas, S. M., Barbas, C. F. III, and Burton, D. R.,
25. Marchalonis, J. J., Kaymaz, H., Schluter, S. F., and Yocum,The nature of the autoimmune antibody repertoire in humanD. E., Human autoantibodies to a synthetic putative T-cell recep-immunodeficiency virus type 1 infection. Proc. Natl. Acad. Sci.tor b-chain regulatory idiotype: Expression in autoimmunity andUSA 91, 3710–3714, 1994.aging. Exp. Clin. Immunogenet. 10, 1–15, 1993.11. Najean, Y., and Rain, J. D., The mechanism of thrombocytopenia
26. Duffy, B. F., Mathew, J. M., Flye, M. W., and Mohanakumar, T.,in patients with HIV infection. J. Lab. Clin. Med. 123, 415–420,Development of autoantibodies to T-cell clonotypic structures1994.in a liver–kidney allograft recipient. Transplant 56, 212–216,12. Winfield, J. B., Mimura, T., and Fernstein, P. D., Autoantibodies1993.to CD45 in systemic lupus erythematosus. Int. J. Clin. Lab. Res.
27. Lake, D. F., Landsperger, W. J., Bernstein, R. M., Schluter, S. F.,21, 292–295, 1992.and Marchalonis, J. J., In ‘‘Immunobiology of Proteins and Pep-13. Marchalonis, J. J., Kaymaz, H., Dedeoglu, F., Schluter, S. F.,tides’’ (M. Z. Atassi, Ed.), Vol. VIII, Plenum, New York, 1994.Yocum, D. E., and Edmundson, A. B., Human autoantibodies re-
28. Wang, E., Lake, D., Winfield, J. B., and Marchalonis, J. J., IgGactive with synthetic autoantigens from T-cell receptor b chain.autoantibodies to ‘‘switch peptide’’ determinants of TCR a/b inProc. Natl. Acad. Sci. USA 89, 3325–3329, 1992.human pregnancy. Clin. Immunol. Immunopathol. 73, 224–228,14. Marchalonis, J. J., Schluter, S. F., Wang, E., Dehghanpisheh, K.,1994.Lake, D., Yocum, D. E., Edmundson, A. B., and Winfield, J. B.,
29. Dedeoglu, F., Kaymaz, H., Klein, G., and Marchalonis, J. J.,Synthetic autoantigens of immunoglobulins and T-cell receptors:Light and heavy chains specifying a human IgM k autoantibodyTheir recognition in aging, infection and autoimmunity. Proc.to a T-cell receptor Vb-antigen. Immunol. Lett. 38, 223–227,Soc. Expt. Biol. 207, 129–147, 1994.1993.15. Kaymaz, H., Dedeoglu, F., Schluter, S. F., Edmundson, A. B.,
30. Marchalonis, J. J., Schluter, S. F., Hubbard, R. A., McCabe, C.,and Marchalonis, J. J., Reactions of anti-immunoglobulin seraand Allen, R. C., Immunoglobulin epitopes defined by syntheticwith synthetic T-cell receptor peptides: Implications for thepeptides corresponding to joining-region sequence: Conservationthree-dimensional structure and function of the TCR b chain.of determinants and dependence upon the presence of an arginylInt. Immunol. 5, 491–502, 1993.or lysyl residue for cross-reaction between light chains and T16. Champagne, E., Huchenq, A., Sevin, J., Casteran, N., and Rubin,cell receptor chains. Mol. Immunol. 25, 771–778, 1988.B., An alternative method for T-cell receptor reportoire analysis:
31. Schluter, S. F., and Marchalonis, J. J., Antibodies to syntheticClustering of human Vb subfamilies selected in responses tojoining segment peptide of the T-cell receptor b-chain: Serologi-staphylococcal enterotoxins B and E. Mol. Immunol. 30, 877–cal cross-reaction between products of T-cell receptor genes, anti-886, 1993.gen binding T-cell receptors and immunoglobulins. Proc. Natl.17. Toyonaga, B., and Mak, T. W., Genes of the T-cell antigen recep-Acad. Sci. USA 83, 1872–1876, 1986.tor in normal and malignant T-cells. Annu. Rev. Immunol. 5,
32. Shankey, T. V., Schluter, S. F., and Marchalonis, J. J., Flow cyto-585–620, 1987.metric analysis of human lymphocytes using affinity purified18. Marchalonis, J. J., Schluter, S. F., Hubbard, R. A., Diamanduros,antibody to T cell receptors synthetic J region. Cell Immunol.A., Barker, W. C., and Pumphrey, R. S. H., Conservation of im-118, 526–531, 1989.munoglobulin variable and joining region structure and the de-
33. Zaghouani, H., Bonella, F. A., Meek, K., and Bona, C., Molecularsign of universal anti-immunoglobulin antibodies reactive withbasis for expression of the A48 regulatory idiotype on antibodiesantigen-binding T cell receptors. Int. Rev. Immunol. 3, 241–273,encoded by immunoglobulin variable region genes from various1988.families. Proc. Natl. Acad. Sci. USA 86, 2341–2345, 1989.19. LaRosa, G. J., Davide, J. P., Weinhold, K., Waterbury, J. A.,
34. Dehghanpisheh, K., Huang, D., Schluter, S. F., Watson, R. R.,Profy, A. T., Lewis, J. A., Langlois, A. J., Dreesman, G. R., Bos-and Marchalonis, J. J., Production of IgG autoantibodies to T-well, N., Shadduck, P., Holley, L. H., Karplus, M., Bolognesi,cell receptors in mice infected with the retrovirus LP-BM5. Int.D., Matthews, T. J., Emini, E. A., and Putney, S. D., ConservedImmunol. 7, 31–36, 1995.sequence and structural elements in the HIV-1 principal neu-
tralizing determinant. Science 249, 932–935, 1990. 35. Ayouba, A., Peltre, G., and Coutinho, A., Quantitative analysis
AID Clin 4290 / a50a$$$203 12-20-96 13:57:53 clinas AP: Clin

189AUTOANTIBODIES TO TCR IN HIV INFECTION
of multiple V-region interactions among normal human IgG. 38. Scott-Algara, D., Lafon, M., Vuillier, F., Pialoux, G., Dauguet,Eur. J. Immunol. 26, 710–716, 1996. C., and Dighiero, G., Viral superantigen-induced hyporespon-
siveness of T cells and polyclonal B cell activation in HIV-1 infec-36. Mouthon, L., Haury, M., La Croix-Desmazes, S., Barreau, C.,Coutinho, A., and Kazatchkine, M. D., Analysis of the normal tion. Eur. J. Immunol. 24, 2595–2601, 1994.human IgG antibody repertoire: Evidence that IgG autoantibod- 39. Imberti, L., Sottini, A., Bettinard, A., Pouti, M., and Primi, D.,ies of healthy adults recognize a limited and conserved set of Selective depletion in HIV infection of T cells that bear specificprotein antigens in homologous tissues. J. Immunol. 154, 5769–
T cell receptor V b sequences. Science 254, 860–862, 1991.5778, 1995.40. Susal, C., Karopelin, M., Daniel, V., and Opelz, G., Molecular37. Akolkar, P. N., Gulwani-Akolkar, B., and Silver, J., The influ-
mimicry between HIV-1 and antigen receptor molecules: A clueence of non-HLA genes on the human T-cell receptor repertoire.Scand. J. Immunol. 42, 598–256, 1995. to the pathogenesis of AIDS. Vox Sang, 65, 10–17, 1993.
Received August 1, 1996; accepted with revision October 22, 1996
AID Clin 4290 / a50a$$$203 12-20-96 13:57:53 clinas AP: Clin
Copyright © 2022 FDOKUMEN




















