
Natural Product–Guided Discovery of a Fungal Chitinase Inhibitor
Upload
independentCategory
view
2download
0
ORIGINAL ARTICLE
Mee-Len Chye Æ Kai-Jun Zhao Æ Zhu-Mei He
Sathishkumar Ramalingam Æ King-Leung Fung
An agglutinating chitinase with two chitin-binding domainsconfers fungal protection in transgenic potato
Received: 29 April 2004 / Accepted: 14 August 2004 / Published online: 13 October 2004� Springer-Verlag 2004
Abstract Brassica juncea BjCHI1 is a unique chitinasewith two chitin-binding domains. Here, we show that,unlike other chitinases, potato-expressed BjCHI1 showshemagglutination ability. BjCHI1 expression in B. jun-cea seedlings is induced by Rhizoctonia solani infection,suggesting its protective role against this fungus. Toverify this, transgenic potato (Solanum tuberosum L. cv.Desiree) plants expressing BjCHI1 generated by Agro-bacterium-mediated transformation were challengedwith R. solani. We also transformed potato with acDNA encoding Hevea brasiliensis b-1,3-glucanase,designated HbGLU, and a pBI121-derivative that con-tains cDNAs encoding both BjCHI1 and HbGLU. Invitro fungal bioassays using Trichoderma viride showedthat extracts from transgenic potato lines co-expressingBjCHI1 and HbGLU inhibited fungal growth betterthan extracts from transgenic potato expressing eitherBjCHI1 or HbGLU, suggesting a synergistic effect.Consistently, in vivo fungal bioassays with soil-borneR. solani on young transgenic potato plants indicatedthat the co-expressing plants showed healthier rootdevelopment than untransformed plants or those thatexpressed either BjCHI1 or HbGLU. Light microscopyand transmission electron microscopy revealed abun-dant intact R. solani hyphae and monilioid cells in un-transformed roots and disintegrated fungus in theBjCHI1-expressing and the BjCHI1 and HbGLU co-expressing plants. Observations of collapsed epidermalcells in the co-expressing potato roots suggest that these
proteins effectively degrade the fungal cell wall, pro-ducing elicitors that initiate other defense responsescausing epidermal cell collapse that ultimately restrictsfurther fungal penetration.
Keywords Brassica Æ Chitinase Æ b-1,3-Glucanase ÆHemagglutination Æ Rhizoctonia Æ Solanum
Abbreviations BjCHI1: Brassica juncea chitinase 1 ÆCaMV: Cauliflower Mosaic Virus Æ HbGLU: Heveabrasiliensis b-1,3-glucanase Æ MS: Murashige andSkoog Æ PVDF: Polyvinylidene difluoride Æ TEM:Transmission electron microscopy Æ UDA: Urtica dioicaagglutinin
Introduction
Pathogenesis-related proteins, including chitinases andb-1,3-glucanases, are produced by plants in response tomicrobial infection. These enzymes hydrolyse chitin andglucan of fungal cell walls (Boller 1985). Schlumbaumet al. (1986) have demonstrated that purified plantchitinases demonstrate antifungal activity in vitro andthat the expression of chitinases in transgenic plants en-hances their resistance against fungal pathogens (Broglieet al. 1991; Lin et al. 1995). Chitinases act synergisticallywith b-1,3-glucanases in inhibiting fungal growth (Mauchet al. 1988; Zhu et al. 1994; Jach et al. 1995).
To understand the plant–pathogen interactions inBrassica leafy vegetables, popular in East Asia, we haveinitiated investigations on chitinases from B. juncea (In-dian mustard). We have cloned a cDNA encoding anacidic chitinase, designated BjCHI1, which is structurallydistinct frompreviously reported chitinases because of thepresence of two chitin-binding domains (Zhao and Chye1999). BjCHI1 shows closest (62%) identity to Nicotianatabacum Chia1 chitinase but, unlike Chia1 chitinases, itpossesses a second chitin-binding domain. As a result ofthe presence of two chitin-binding domains, it resembles
M.-L. Chye (&) Æ K.-J. Zhao Æ Z.-M. He Æ S. RamalingamK.-L. FungDepartment of Botany, The University of Hong Kong,Pokfulam Road, Hong Kong, ChinaE-mail: [email protected]: +852-28583477
Present address: K.-J. ZhaoInstitute of Crop Breeding and Cultivation,Chinese Academy of Agricultural Sciences, Beijing, China
Present address: Z.-M. HeBiotechnology Research Centre, Zhongshan University,Guangzhou, China
Planta (2005) 220: 717–730DOI 10.1007/s00425-004-1391-6

the precursor of Urtica dioica agglutinin (UDA). How-ever, there is only 36.9% identity between them and, un-like the UDA precursor, the chitin-binding domains ofBjCHI1 are not cleaved from the catalytic domain post-translationally (Zhao and Chye 1999). Absence of cleav-age between these two domains was confirmed by ourability to purify active tobacco-expressed BjCHI1 using aregenerated chitin column (Fung et al. 2002). Chitinaseswith single or double chitin-binding domains exist inB. juncea and the 1.3-kb BjCHI1 mRNA is induced bywounding, methyl jasmonate treatment, Aspergillus nigerinfection and caterpillar (Pieris rapae) feeding, suggestiveof its role in defense (Zhao and Chye 1999; Fung et al.2002).
Here, we verified the role of BjCHI1 in defense anddemonstrated that it is a unique chitinase with aggluti-nation ability. We show that it confers fungal protectionby its heterologous expression from the CauliflowerMosaic Virus 35S (CaMV35S) promoter in transgenicpotato. Potatowas chosen for transformation because it isa crop that is susceptible to soil-borne fungal infectionsand its tubers are produced in the soil. Since we have alsopreviously cloned a b-1,3-glucanase cDNA from Heveabrasiliensis (Chye and Cheung 1995), we have co-ex-pressed it with BjCHI1. We subsequently challengedyoung transgenic potato plants with R. solani Kuhn [te-lemorph Thanatephorus cucumeris (Frank) Donk], caus-ative agent of Rhizoctonia canker in growing potatoplants and black scurf in tubers, diseases resulting in croplosses of up to 30% (Banville et al. 1999).We present lightmicroscopy and transmission electronmicroscopy (TEM)analyses on the effect of BjCHI1 and HbGLU expressionin planta. So far, only one previous study using TEMofR.solani protection by the heterologous expression of achitinase has been reported (Benhamou et al. 1993).
Materials and methods
Infection of Brassica juncea with Rhizoctonia solani
Brassica juncea (L.) Czern &Coss seeds (gift of Prof. E.C.Pua, Department of Biological Sciences, National Uni-versity of Singapore), sown on autoclaved soil, were in-cubated in a growth chamber at 24�C with a day–nightregime of 12 h light and 12 h dark. Two-week-old seed-lings were carefully transferred to soil inoculated with aweek-old culture of R. solani. The fungus was grown atroom temperature for 1 week in potato dextrose broth(PDB) and was added to the soil 3 days before the seed-lings were planted. Leaves from seedlings were harvesteddaily from 0 to 9 days after transfer to infected soil fortotal RNA and total protein extractions for use innorthern blot and western blot analyses, respectively.
Plasmid constructs
A 1.2-kb SmaI–HindII fragment of full-length Heveabrasiliensis b-1,3-glucanase cDNA, designated HbGLU
(Chye and Cheung 1995) was cloned downstream fromthe CaMV35S promoter in the SmaI site of binaryplasmid vector pBI121 (Clontech) to generate plasmidpHEV43. A 1.3-kb SmaI fragment of full-length cDNAencoding B. juncea chitinase, BjCHI1 (Zhao and Chye1999) was similarly cloned in the SmaI site of pBI121(Clontech) to generate plasmid pBj17. Subsequentlyfrom pHEV43, a 2.04-kb HindIII fragment containingthe CaMV35S promoter and the HbGLU full-lengthcDNA was blunt-ended with Klenow and ligated to theSnaBI site within GUS of pBj17, generating plasmidpBj47. Plasmids pBj17, pBj47 and pHEV43 were used inpotato transformation.
Plant transformation
Each of the plasmids pHEV43, pBj17, pBj47 and pBI121was mobilized from Escherichia coli strain DH5a intoAgrobacterium tumefaciens strain LBA4404 by tri-parental mating using helper strain E. coli HB101(pRK2013). Potato (Solanum tuberosum L.) cultivarDesiree was obtained from Prof. Z.P. Lin, College ofLife Sciences, Peking University. Following Agrobacte-rium-mediated transformation of potato (Dietze et al.1995), transformants were selected on Murashige andSkoog (MS) medium supplemented with kanamycin(100 lg ml�1). Potato tissue cultures were maintained ina growth incubator under a 16 h light/8 h dark regime at20–25�C. Transgenic R0 potato plants were grown in soilin a growth chamber at 24�C with a day/night regime of12 h light/12 h dark.
Northern blot analysis
Twenty lg of RNA, extracted from whole plants (Nagyet al. 1988), was denatured at 50�C for 30 min in glyoxal,separated by electrophoresis in a 1.5% agarose gel andtransferred to Hybond-N membrane (Amersham). Blots,pre-hybridized for 4–6 h, were hybridized with random-primed 32P-labelled BjCHI1 cDNA or 32P-labelledHbGLU cDNA, in a solution containing 50% formamide,1· Denhardt¢s, 6· SSPE (1· SSPE: 0.15 MNaCl, 0.01 MNaH2PO4, 0.001 M EDTA, pH 7.4), 0.1% SDS,100 lg ml�1 denatured salmon sperm DNA and 10%dextran sulphate at 42�C overnight. Blots were washed in0.1· standard saline citrate (1· SSC: 150 mM NaCl,15 mM sodium citrate, pH 7.0) containing 0.1% SDS at65�C.
Southern blot analysis
For Southern blot analysis, 20 lg genomic DNA iso-lated according to Dellaporta et al. (1983), was digestedwith restriction endonucleases, separated by electro-phoresis in a 0.7% agarose gel and blotted onto Hy-bond-N (Amersham) membranes (Sambrook et al.
718

1989). Membranes were prehybridized in a solutioncontaining 50% deionized formamide, 6· SSC, 5·Denhardt¢s, 1% SDS, 100 lg ml�1 denatured salmonsperm DNA at 42�C for 4–6 h. Random-primed 32P-labelled BjCHI1 cDNA or 32P-labelled HbGLU cDNAwas added and hybridized overnight. Membranes werewashed in 0.1· SSC, 0.1% SDS at 65�C.
Preparation of polyclonal antibodies againstHbGLU and BjCHI1
A synthetic peptide (SDLQSLTNPSNAKS) corre-sponding to amino acids 94–107 of HbGLU (Chye andCheung 1995) was purchased from Chiron Technologies(Clayton, Australia), conjugated with keyhole limpethemocyanin (Sigma), mixed with Freund’s completeadjuvant and subsequently injected into rabbit for rais-ing polyclonal antibodies according to Sambrook et al.(1989). A synthetic peptide (YKEEIDKSDPHC) corre-sponding to amino acids 231–242 of BjCHI1 (Zhao andChye 1999) was similarly treated for immunization ofrabbit to raise polyclonal antibodies; these anti-BjCHI1-specific antibodies were purchased from Chiron Tech-nologies. Blood was collected and antibodies againstHbGLU were purified using Protein A–SepharoseCL-4B (Pharmacia) followed by a column consisting ofthe HbGLU peptide coupled to cyanogen bromide-activated Sepharose 4B (Pharmacia) while antibodiesagainst BjCHI1 were purified using a column of theBjCHI1 peptide coupled to Thiopropyl-Sepharose 6B(Pharmacia).
Western blot analysis
Total plant protein was prepared from whole plantsaccording to Kush et al. (1990). Twenty lg of totalprotein was separated by SDS–PAGE and transferredonto Hybond-C (Amersham) membrane as described bySambrook et al. (1989). In western blot analysis, cross-reacting bands were detected using polyclonal antibodiesagainst BjCHI1 or polyclonal antibodies againstHbGLU, following the procedures described in theAmplified Alkaline Phosphatase Goat Anti-Rabbit Im-mun-Blot Assay Kit (BioRad).
Purification of BjCHI1 from transgenic potatousing affinity chromatography
Regenerated colloidal chitin was prepared for affinitychromatography according to the procedure describedby Boller (1992). BjCHI1 was purified using 20-mlregenerated colloidal chitin columns following Iseli et al.(1993). Extraction and all subsequent steps were per-formed at 4�C. Transgenic potato leaves (100 g) fromline pBj17-P6 were homogenized in 100 mM sodiumacetate buffer (pH 4). The homogenate was centrifuged
at 18,000 g for 10 min and ammonium sulfate was ad-ded to 65% saturation. Following overnight incubation,the pellet was collected by centrifugation at 10,000 g for10 min. The spun pellet was re-dissolved in 20 mM Tris(HCl), pH 8, and was loaded onto the column equili-brated in the same solution. Subsequently, the columnwas eluted successively with 20 mM Tris (HCl), pH 8,followed by 20 mM sodium acetate buffer (pH 5.5), and100 mM acetic acid (pH 3). Eluates were collected in300-ll fractions. A tenth of each portion (30 ll) wasloaded for analysis using SDS–PAGE. Eluates with theexpected protein band were dialyzed against 10 mMsodium phosphate buffer (pH 7.2) overnight at 4�C,lyophilized and stored at �70�C. BjCHI1 purified usingaffinity chromatography was used for SDS–PAGE fol-lowed by N-terminal sequencing and for hemagglutina-tion assays.
N-terminal sequencing of purified BjCHI1
BjCHI1 purified from transgenic potato line pBj17-P6 byaffinity chromatography using regenerated colloidalchitin, was electrophoresed on an SDS–polyacrylamidegel and transferred to a polyvinylidene difluoride(PVDF) membrane filter (Invitrogen) according toinstructions of the manufacturer. The membrane wasstained with Coomassie Brilliant Blue and de-stained ina solution of 50% (v/v) methanol and 10% (v/v) aceticacid. The band of apparent molecular mass 52 kDa wasexcised and used in N-terminal sequencing, which wasrepeated to confirm results. Edman degradation wasperformed using an HP Protein Sequencer (ModelG1000A) according to the manufacturer’s instructions.
Estimation of total carbohydrate contentof purified BjCHI1
Total carbohydrate content of potato-expressedBjCHI1, purified by affinity chromatography usingregenerated colloidal chitin, was carried out using phe-nol–sulfuric acid assays following the procedure ofTaylor (1995). Assays were carried out in duplicates andabsorbance was read at 490 nm against a set of glucosestandards (ranging from 0 lg to 100 lg) from which astandard curve was prepared. Bovine fetuin (Sigma) wasused as a positive control.
Hemagglutination assays
Hemagglutination assays were carried out using potato-expressed BjCHI1 that was purified by affinity chroma-tography using regenerated colloidal chitin. These assayswere carried out according to the procedure of Doeset al. (1999) in a microtiter tray. Final protein concen-trations tested were at 0 (control), 4.2, 8.3, 33.3 and66.7 lg of purified BjCHI1 protein per ml. In each
719

protein-containing well, trypsin-treated rabbit erythro-cytes (30 ll) were added and 10 mM sodium phosphatebuffer (pH 7.2) was used to make up the volume to60 ll. The reaction was left for 1 h at room temperature,before documentation by photography.
Preparation of plant protein extractsfor enzyme assays
Plant protein extracts were prepared according to themethod of Boller et al. (1983). Plants were ground to finepowder in liquid nitrogen, transferred to 0.1 M sodiumcitrate (pH 5.0) buffer containing 1% (v/v) b-mercap-toethanol and vortexed. Following incubation at 50�Cfor 10 min and cooling on ice for 10 min, the sample wascentrifuged (20,900 g for 5 min at 4�C), and the super-natant was used as the crude protein in assays forchitinase and b-1,3-glucanase activities. Protein con-centration was determined according to Bradford(1976).
b-1, 3-Glucanase assays
Colorimetric assays for b-1,3-glucanase were carried outfollowing the method of Abeles and Forrence (1970).Each reaction mixture consisting of 100 ll (25 lg) plantprotein extract and 100 ll of 2% (w/v) laminarin (Sig-ma) was incubated at 50�C for 2 h. The reaction wasterminated by addition of 600 ll of dinitrosalicylic re-agent and heating for 5 min at 100�C. Following coolingto room temperature, the contents were diluted 1:20 inwater and the absorbance was measured at 500 nm.
Chitinase assays
Colorimetric chitinase assays were carried out accordingto Wirth and Wolf (1990). The reaction mixture of150 ll of carboxymethyl/chitin/Remazol Brilliant Violet(Loewe Biochemica; 2 mg ml�1 stock solution), 150 llof 0.2 M sodium acetate (pH 5.0) and 300 ll (15 lg)protein extract was incubated at 37�C for 0.5 h. Thereaction was terminated with 150 ll of 1 N HCl, fol-lowed by incubation on ice for 5–10 min, before cen-trifugation (20,900 g for 3 min at 4�C). The supernatantwas used to read absorbance at 550 nm. For the blank,300 ll of 0.1 M sodium citrate buffer (pH 5.0) replacedthe protein sample.
In vitro bioassays using Trichoderma viride
T. viride Persoon (American Type Culture Collection12582) was cultured on a plate of potato dextrose(Difco) agar (PDA). It was used for in vitro hyphalinhibition tests following the method of Schlumbaumet al. (1986). A plug of growing T. viride culture on PDAwas transferred to the centre of a fresh PDA plate.
Following incubation at 25�C for 24 h, during which thehyphae grew outwards from the centre, wells were boredon the outer surface of the PDA, equidistant from theplug. Plant protein extract (50 lg) was added to eachwell and the plate was further incubated in the dark at25�C after which growth inhibition of T. viride was ob-served. Photographs were taken at 16 h and 24 h.
In vivo bioassays using R. solani in MS
R. solani grown on a PDA plate was used to inoculate500 ml of MS liquid medium. The culture was incubatedwith shaking (80 rpm) at 28�C for 1 week. Twenty-fi-ve ml of this R. solani culture was transferred to a 250-ml sterile culture flask. Each tissue-cultured plant on MSagar medium, of potato lines, pBj17-P6, pBj47-P10 andwild type, was transferred to a flask containing the 25 mlof R. solani. The cultures were then incubated with rootssubmerged at 28�C under a 12 h light/12 h dark cycle.After 5 days, the plant roots were harvested and fixedfor light microscopy.
In vivo bioassays using R. solani in soil
In vivo bioassays were carried out on young potatoplants according to the method of Jach et al. (1995). R.solani incubated on solid PDA medium at 25�C for 5–6 days was inoculated into 100 ml of liquid PDB andincubated with shaking (100 rpm) at room temperaturefor 1–2 weeks. Subsequently, the culture was transferredto a conical flask containing 500 ml of fresh PDB andfurther incubated for 3–4 days. This culture was thenthoroughly mixed with 6 l of sterilized soil (Bio-MixSuper; Schoonebeek, The Netherlands) and added to 8 lsterilized soil in a plant growth tray. After 10 days, thesoil was uniformly mixed and potato plants, originallyfrom tissue culture and previously grown for 2 weeks insterilized soil, were transplanted into the infected soiland further incubated in a growth chamber at 24�Cunder a 12 h light/12 h dark regime. Photographs weretaken of potato plants at 2 weeks following transplan-tation to infected soil. Plants grown in sterilized soilwithout fungus were used as controls. Roots fromtransgenic potato line pBj47-P10 and untransformedpotato were harvested for examination by light micros-copy and TEM.
Light microscopy and TEM of root sectionsfrom R. solani-infected potato
Lesion-containing roots were excised and fixed in asolution of 2% (v/v) glutaraldehyde in 0.1 mM sodiumphosphate buffer (pH 7.2) for 24 h at room temperature,followed by a solution of 1% (w/v) osmium tetroxide in0.1 mM sodium phosphate buffer (pH 7.2), for 4 h atroom temperature. They were dehydrated in a graded
720

ethanol series (30, 50, 70, 80, 90, 100 and 100%), eachfor 2 h at room temperature, and embedded in SpurrLow Viscosity Embedding Kit (Electron MicroscopySciences). For light microscopy, 1-lm sections were cutusing a glass knife, collected on microscope slides,stained with 1% (w/v) aqueous Toluidine Blue andphotographed using a Leica DMRXA microscope. ForTEM, 0.6- to 0.7-lm sections were cut with a diamondknife, collected on Formar-coated copper grids, stainedwith 2% uranyl acetate for 15 min, and 2% lead citratefor 10 min, followed by examination under a PhilipsEM-208S electron microscope.
Results
BjCHI1 expression is induced by R. solani infection
To investigate the effect of R. solani infection on BjCHI1expression in B. juncea, seedlings were grown in soil pre-inoculated with this soil fungus and RNA was extractedfrom leaves for northern blot analysis. Our resultsdemonstrate that the expression of the 1.3-kb BjCHI1mRNA increases 1 day following growth in infected soil(Fig. 1a). A cross-reacting band of apparent molecularmass of 37 kDa (arrow, Fig. 1b), which is expected ofthe mature BjCHI1 protein, accumulates from day 3 ofgrowth in infected soil. An approx. 40-kDa faint band(arrowhead, Fig. 1b) above the 37-kDa band, could bethe pre-protein. In B. juncea, the BjCHI1 pre-protein ofcalculated molecular mass 42.77 kDa, undergoes post-translational cleavage whereby an N-terminal signalpeptide and a C-terminal vacuolar targeting peptide areremoved to yield mature BjCHI1 of calculated molecu-lar mass 39.67 kDa (Fung et al. 2002).
Expression of BjCHI1 and HbGLU mRNAin transgenic potato
The observation that BjCHI1 expression is induced byR. solani infection in B. juncea seedlings led us toinvestigate its potential use against R. solani by heter-ologous expression in transgenic potato. To this end,BjCHI1 cDNA-containing plasmids, pBj17 and pBj47,were introduced into potato by A. tumefaciens-mediatedtransformation. Plasmid pBj47 also contains theHbGLU cDNA, enabling us to investigate any syner-gistic effect of HbGLU on BjCHI1 action. PlasmidpHEV43 containing the HbGLU cDNA alone and vec-tor control, pBI121, were also used in potato transfor-mation. About 20 independent kanamycin-resistantputative transgenic plants from each transformationwere examined by northern, Southern and western blotanalyses. Results of analyses of representative lines fromeach transformation that were eventually chosen forexperiments on R. solani infection are shown in Figs. 2and 3. A 1.2-kb hybridizing HbGLU mRNA (arrow-head, Fig. 2a) was detected using a 32P-labelled HbGLU
cDNA probe in a northern blot analysis of transgenicpotato lines co-expressing HbGLU and BjCHI1, pBj47-P10 (lane 2), pBj47-P8 (lane 3), pBj47-P7 (lane 4), and atransgenic potato line expressing HbGLU alone,pHEV43-P14 (lane 5). This band is absent in untrans-formed potato (Fig. 2a, lane 1). A 1.3-kb BjCHI1hybridizing mRNA (arrow, Fig. 2b) was detected with32P-labelled BjCHI1 cDNA in transgenic potato linespBj47-P10 (lane 2), pBj47-P8 (lane 3), pBj47-P7 (lane 4)and pBj17-P6 (lane 6). This band is absent in untrans-formed potato (Fig. 2b, lane 1) and pBI121-transformedpotato (Fig. 2b, lane 5).
Southern blot analysis of transgenic potato lines
Subsequently, DNA from transgenic potato lines wasused in Southern blot analysis with 32P-labelled BjCHI1
Fig. 1a–c BjCHI1 expression in Brassica juncea seedlings follow-ing Rhizoctonia solani infection. a Northern blot analysis with32P-labelled BjCHI1 cDNA of total RNA from B. juncea leavesharvested 0 days (lane 1), 1 day (lane 2), 2 days (lane 3), 3 days(lane 4), 4 days (lane 5), 5 days (lane 6), 6 days (lane 7), 7 days(lane 8), 8 days (lane 9) and 9 days (lane 10) after growth in R.solani-infected soil. The same blot re-probed with 32P-labelled 18SrDNA is shown below. Arrow 1.3-kb BjCHI1 mRNA, arrowhead18S rRNA. b Western blot analysis using anti-BjCHI1 antibodies.Total protein from B. juncea leaves was harvested 0 days (lane 1),1 day (lane 2), 2 days (lane 3), 3 days (lane 4), 4 days (lane 5),5 days (lane 6), 6 days (lane 7), 7 days (lane 8), 8 days (lane 9) and9 days (lane 10) after growth in R. solani-infected soil. ArrowCross-reacting 37-kDa BjCHI1, arrowhead pre-protein. c Coomas-sie Blue-stained protein gel loaded as in b demonstrates equalamounts of protein in each well
721

or HbGLU cDNA (data not shown). The results ofSouthern blot analysis using EcoRI-digested DNAprobed with the 32P-labelled HbGLU cDNA showed thepresence of a 1.2-kb HbGLU EcoRI-hybridizing band intransgenic potato lines pHEV43-P14, pBj47-P7, pBj47-P8
and pBj47-P10. This band was absent in both untrans-formed potato and in pBI121-transformed potato. The1.2-kb EcoRI-hybridizing band corresponds to the 1.2-kb HbGLU full-length cDNA in pHEV43. Using32P-labelled BjCHI1 cDNA and HindIII-digested DNAin Southern blot analysis, 0.9-kb HindIII-hybridizingbands were detected in transgenic potato lines pBj47-P7,pBj47-P8, pBj47-P10 and pBj17-P6. This band corre-sponds to the fragment between the second and thirdinternal HindIII sites within the BjCHI1 cDNA (Zhaoand Chye 1999) in pBj17. This band was absent in un-transformed potato and in pBI121-transformed potato.
When EcoRI-digested DNA was probed with 32P-labelled BjCHI1 cDNA in Southern blot analysis,hybridizing bands of various sizes were seen withtransgenic potato lines pBj47-P10, pBj47-P8 and pBj47-P7, suggesting that they are independent lines. Suchhybridizing bands were absent in untransformed potatoand in pBI121-transformed potato.
Analysis of BjCHI1 and HbGLU expressedin transgenic potato
Following northern blot analysis and Southern blotanalysis, transgenic potato plants were further analyzedby western blot analysis using anti-HbGLU antibodiesand anti-BjCHI1 antibodies. Western blot analysis usinganti-HbGLU antibodies (Fig. 3a) on crude protein fromtransgenic potato lines pBj47-P7 (lane 1), pBj47-P8
(lane 2), pBj47-P10 (lane 3) and pHEV43-P14 (lane 4)showed a cross-reacting band with an apparent molec-
Fig. 2a, b Northern blot analysis of R0 transgenic potato (Solanumtuberosum) plants. a Expression of the 1.2-kb hybridizing HbGLUmRNA (arrowhead) as detected with 32P-labelled HbGLU cDNA inuntransformed potato (lane 1) and transgenic potato lines pBj47-P10 (lane 2), pBj47-P8 (lane 3), pBj47-P7 (lane 4) and pHEV43-P14
(lane 5). b Expression of the 1.3-kb BjCHI1-hybridizing mRNA(arrow) as detected with 32P-labelled BjCHI1 cDNA in untrans-formed potato (lane 1) and transgenic potato lines pBj47-P10
(lane 2), pBj47-P8 (lane 3), pBj47-P7 (lane 4), pBI121-transformedpotato (lane 5) and pBj17-P6 (lane 6)
Fig. 3a–c Analysis of HbGLU and BjCHI1 expressed fromtransgenic potato. a Western blot analysis using anti-HbGLUantibodies on crude protein from transgenic potato R0 lines pBj47-P7 (lane 1), pBj47-P8 (lane 2), pBj47-P10 (lane 3), pHEV43-P14
(lane 4), pBI121-transformed potato (lane 5) and untransformedpotato (lane 6). Arrowhead 35-kDa HbGLU. b Western blotanalysis using anti-BjCHI1 antibodies on crude protein fromtransgenic potato R0 lines pBj47-P7 (lane 1), pBj47-P8 (lane 2),pBj47-P10 (lane 3), pHEV43-P14 (lane 4), pBI121-transformedpotato (lane 5), untransformed potato (lane 6) and transgenicpotato line pBj17-P6 (lane 7). Arrow 52-kDa BjCHI1. c Hemag-glutination assays on potato-expressed BjCHI1 purified by affinitychromatography using regenerated colloidal chitin. Each wellcontains purified BjCHI1 (in lg ml�1 amounts as indicated abovewell), 30 ll trypsin-treated rabbit erythrocytes and 10 mM sodiumphosphate buffer (pH 7.2) in a final volume of 60 ll. Thephotograph was taken after 1 h reaction at room temperature.The control (con) lacking BjCHI1 is on the right
722

ular mass of 35 kDa corresponding to HbGLU. Thisband was absent in pBI121-transformed potato (Fig. 3a,lane 5) and untransformed potato (lane 6).
Western blot analysis using anti-BjCHI1-specificantibodies (Fig. 3b) on crude protein from transgenicpotato lines pBj47-P7 (lane 1), pBj47-P8 (lane 2), pBj47-P10 (lane 3), pHEV43-P14 (lane 4), pBI121-transformedpotato (lane 5), untransformed potato (lane 6) andtransgenic potato line pBj17-P6 (lane 7) showed a cross-reacting BjCHI1 band, with an apparent molecular massof 52 kDa in lines pBj47-P7 (lane 1), pBj47-P8 (lane 2),pBj47-P10 (lane 3) and pBj17-P6 (lane 7). This band wasabsent in the transgenic line expressing HbGLU alone,pHEV43-P14 (Fig. 3b, lane 4), in pBI121-transformedpotato (Fig. 3b, lane 5) and in untransformed potato(Fig. 3b, lane 6).
The apparent molecular mass (52 kDa) of potato-expressed BjCHI1 observed in Fig. 3b resembles that oftobacco-expressed BjCHI1 (Fung et al. 2002), but islarger than that of native BjCHI1 which has a calculatedmass of 39.67 kDa (Fung et al. 2002). Since a BjCHI1-specific peptide that has no homology to any other re-ported chitinases (Zhao and Chye 1999) was used inraising antibodies against BjCHI1, it is unlikely that thiscross-reacting band, absent in the controls, does notcorrespond to BjCHI1. Nonetheless, we carried out N-terminal sequencing to further confirm the identity ofthe 52-kDa band. To this end, affinity chromatographyusing a regenerated colloidal chitin column was used topurify BjCHI1 from potato line pBj17-P6. The yieldfrom affinity column chromatography was 3 lg affinity-purified protein per g fresh leaf sample. The chitinaseactivity of purified potato-expressed BjCHI1 was0.023 U lg�1 min�1. Following SDS–PAGE analysisand blotting onto PVDF membrane, the CoomassieBlue-stained protein band was eluted for N-terminalsequencing. The results show that the identity of the firstten amino acids corresponded to amino acids 21–30(EQCGSQSIPE) of BjCHI1. These results further con-firmed that the N-terminal signal peptide of heterolo-gously expressed BjCHI1 is cleaved, as predicted, even inpotato. An estimation of the carbohydrate content ofpotato-expressed BjCHI1 revealed that it consists of20% carbohydrate. The reduced mobility of BjCHI1 inSDS–PAGE, which has been attributed to O-glycosyla-tion, resembles that of potato lectin (Van Damme et al.2004).
To investigate whether BjCHI1 resembles plant lec-tins that hemagglutinate, including bean phytohemag-glutinin (Chrispeels and Raikhel 1991), UDA (Doeset al. 1999) and the mannose-binding lectin from blue-bell (Wright et al. 1999), hemagglutination assays wereconducted using affinity-purified BjCHI1 from potatoR0 line pBj17-P6. Figure 3c shows that potato-expressedBjCHI1 agglutinates rabbit erythrocytes at a concen-tration of 8.3 lg ml�1 while agglutination is absent inthe control lacking BjCHI1. Agglutination was moresignificant when BjCHI1 concentration was increased to33.3 and 66.7 lg ml�1 (Fig. 3c).
Chitinase and b-1,3-glucanase activity assays
Transgenic potato lines were tested for chitinase and b-1,3-glucanase activity assays and compared to pBI121-transformed potato. Glucanase assays (Fig. 4a) usingcrude protein from the pBI121 transformant andtransgenic potato lines pBj47-P7, pBj47-P8 and pBj47-P10 indicated higher levels of b-1,3-glucanase activity inthe pBj47 transgenic lines. Chitinase assays (Fig. 4b)using crude protein show that transgenic potato linespBj47-P7, pBj47-P8 and pBj47-P10 had higher levels ofchitinase activities than the pBI121 transformant.Activities in the pBI121 transformant are due to thepresence of endogenous potato b-1,3-glucanases andchitinases.
In vitro bioassays using T. viride
Subsequently, crude extracts from transgenic potatolines were tested in vitro for T. viride growth inhibition.Extracts from untransformed potato and a transgenicline transformed with pBI121 were used as controls inthese bioassays. Photographs taken at 16 h (Fig. 5a) and24 h (Fig. 5b) following addition of protein extracts towells show greatest inhibition of T. viride growth usingprotein from the BjCHI1 and HbGLU co-expressingtransgenic potato line pBj47-P10 (well 1). Inhibition wasabsent in the buffer-only control (well 2) and in extractsfrom untransformed potato (well 3) and pBI121-trans-formed potato (well 4). Transgenic potato lines pBj17-P6, expressing BjCHI1 only (well 5), and pHEV43-P14,expressing HbGLU only (well 6), show weaker inhibi-tion than pBj47-P10 (well 1).
Fig. 4a, b Glucanase and chitinase assays on transgenic potatolines. a Glucanase assays using crude protein from the pBI121transformant and transgenic R0 lines transformed with pBj47.b Chitinase assays using crude protein from the pBI121 transfor-mant and transgenic R0 lines transformed with pBj47. Error barsrepresent standard deviations of three separate experiments
723
In vivo bioassays using R. solani
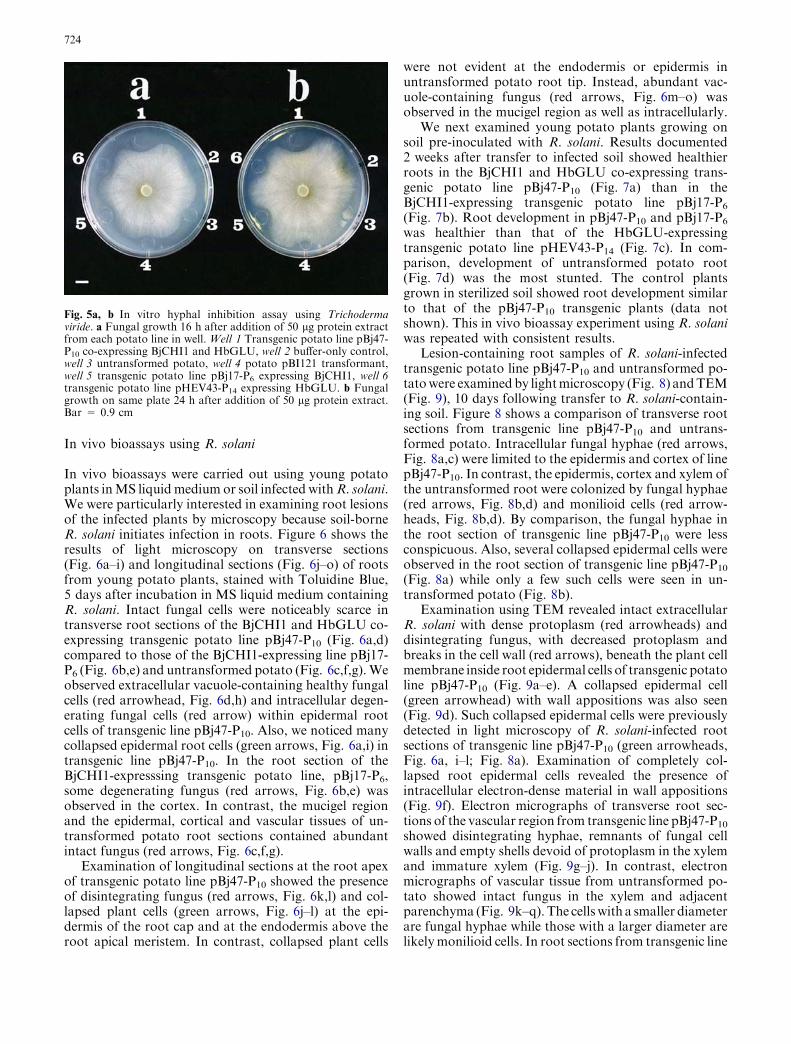
In vivo bioassays were carried out using young potatoplants inMS liquidmedium or soil infected withR. solani.We were particularly interested in examining root lesionsof the infected plants by microscopy because soil-borneR. solani initiates infection in roots. Figure 6 shows theresults of light microscopy on transverse sections(Fig. 6a–i) and longitudinal sections (Fig. 6j–o) of rootsfrom young potato plants, stained with Toluidine Blue,5 days after incubation in MS liquid medium containingR. solani. Intact fungal cells were noticeably scarce intransverse root sections of the BjCHI1 and HbGLU co-expressing transgenic potato line pBj47-P10 (Fig. 6a,d)compared to those of the BjCHI1-expressing line pBj17-P6 (Fig. 6b,e) and untransformed potato (Fig. 6c,f,g).Weobserved extracellular vacuole-containing healthy fungalcells (red arrowhead, Fig. 6d,h) and intracellular degen-erating fungal cells (red arrow) within epidermal rootcells of transgenic line pBj47-P10. Also, we noticed manycollapsed epidermal root cells (green arrows, Fig. 6a,i) intransgenic line pBj47-P10. In the root section of theBjCHI1-expresssing transgenic potato line, pBj17-P6,some degenerating fungus (red arrows, Fig. 6b,e) wasobserved in the cortex. In contrast, the mucigel regionand the epidermal, cortical and vascular tissues of un-transformed potato root sections contained abundantintact fungus (red arrows, Fig. 6c,f,g).
Examination of longitudinal sections at the root apexof transgenic potato line pBj47-P10 showed the presenceof disintegrating fungus (red arrows, Fig. 6k,l) and col-lapsed plant cells (green arrows, Fig. 6j–l) at the epi-dermis of the root cap and at the endodermis above theroot apical meristem. In contrast, collapsed plant cells
were not evident at the endodermis or epidermis inuntransformed potato root tip. Instead, abundant vac-uole-containing fungus (red arrows, Fig. 6m–o) wasobserved in the mucigel region as well as intracellularly.
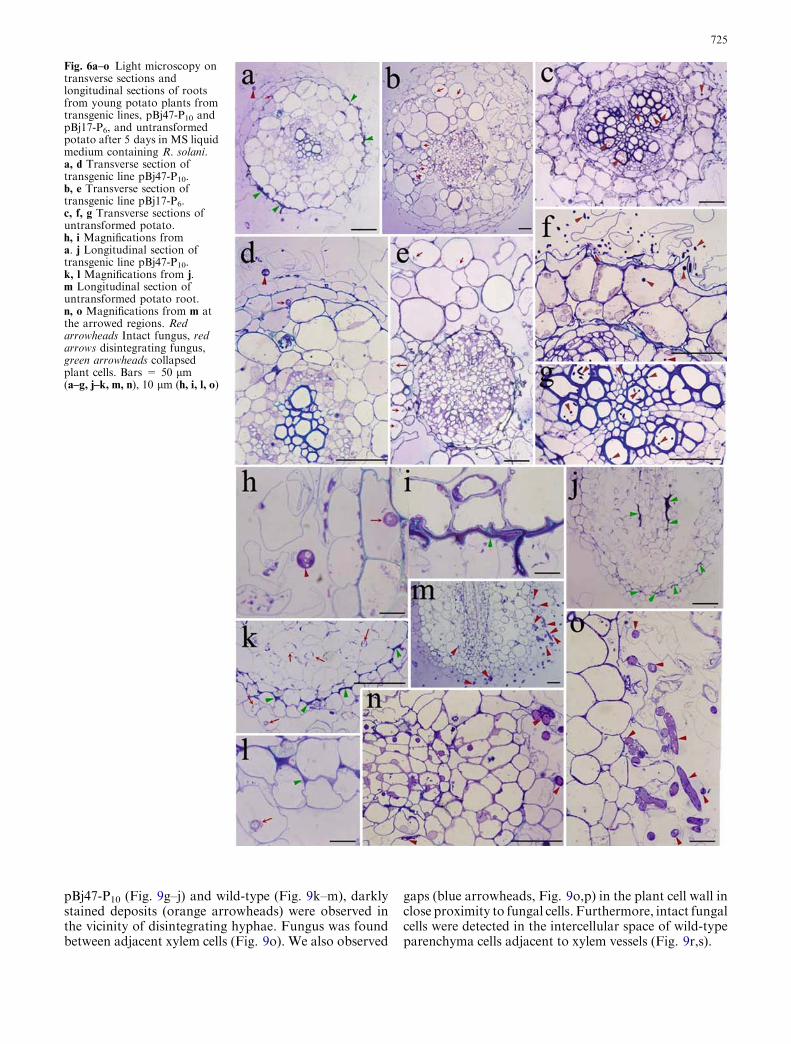
We next examined young potato plants growing onsoil pre-inoculated with R. solani. Results documented2 weeks after transfer to infected soil showed healthierroots in the BjCHI1 and HbGLU co-expressing trans-genic potato line pBj47-P10 (Fig. 7a) than in theBjCHI1-expressing transgenic potato line pBj17-P6
(Fig. 7b). Root development in pBj47-P10 and pBj17-P6
was healthier than that of the HbGLU-expressingtransgenic potato line pHEV43-P14 (Fig. 7c). In com-parison, development of untransformed potato root(Fig. 7d) was the most stunted. The control plantsgrown in sterilized soil showed root development similarto that of the pBj47-P10 transgenic plants (data notshown). This in vivo bioassay experiment using R. solaniwas repeated with consistent results.
Lesion-containing root samples of R. solani-infectedtransgenic potato line pBj47-P10 and untransformed po-tatowere examined by lightmicroscopy (Fig. 8) andTEM(Fig. 9), 10 days following transfer to R. solani-contain-ing soil. Figure 8 shows a comparison of transverse rootsections from transgenic line pBj47-P10 and untrans-formed potato. Intracellular fungal hyphae (red arrows,Fig. 8a,c) were limited to the epidermis and cortex of linepBj47-P10. In contrast, the epidermis, cortex and xylem ofthe untransformed root were colonized by fungal hyphae(red arrows, Fig. 8b,d) and monilioid cells (red arrow-heads, Fig. 8b,d). By comparison, the fungal hyphae inthe root section of transgenic line pBj47-P10 were lessconspicuous. Also, several collapsed epidermal cells wereobserved in the root section of transgenic line pBj47-P10
(Fig. 8a) while only a few such cells were seen in un-transformed potato (Fig. 8b).
Examination using TEM revealed intact extracellularR. solani with dense protoplasm (red arrowheads) anddisintegrating fungus, with decreased protoplasm andbreaks in the cell wall (red arrows), beneath the plant cellmembrane inside root epidermal cells of transgenic potatoline pBj47-P10 (Fig. 9a–e). A collapsed epidermal cell(green arrowhead) with wall appositions was also seen(Fig. 9d). Such collapsed epidermal cells were previouslydetected in light microscopy of R. solani-infected rootsections of transgenic line pBj47-P10 (green arrowheads,Fig. 6a, i–l; Fig. 8a). Examination of completely col-lapsed root epidermal cells revealed the presence ofintracellular electron-dense material in wall appositions(Fig. 9f). Electron micrographs of transverse root sec-tions of the vascular region from transgenic line pBj47-P10
showed disintegrating hyphae, remnants of fungal cellwalls and empty shells devoid of protoplasm in the xylemand immature xylem (Fig. 9g–j). In contrast, electronmicrographs of vascular tissue from untransformed po-tato showed intact fungus in the xylem and adjacentparenchyma (Fig. 9k–q). The cellswith a smaller diameterare fungal hyphae while those with a larger diameter arelikelymonilioid cells. In root sections from transgenic line
Fig. 5a, b In vitro hyphal inhibition assay using Trichodermaviride. a Fungal growth 16 h after addition of 50 lg protein extractfrom each potato line in well. Well 1 Transgenic potato line pBj47-P10 co-expressing BjCHI1 and HbGLU, well 2 buffer-only control,well 3 untransformed potato, well 4 potato pBI121 transformant,well 5 transgenic potato line pBj17-P6 expressing BjCHI1, well 6transgenic potato line pHEV43-P14 expressing HbGLU. b Fungalgrowth on same plate 24 h after addition of 50 lg protein extract.Bar = 0.9 cm
724
pBj47-P10 (Fig. 9g–j) and wild-type (Fig. 9k–m), darklystained deposits (orange arrowheads) were observed inthe vicinity of disintegrating hyphae. Fungus was foundbetween adjacent xylem cells (Fig. 9o). We also observed
gaps (blue arrowheads, Fig. 9o,p) in the plant cell wall inclose proximity to fungal cells. Furthermore, intact fungalcells were detected in the intercellular space of wild-typeparenchyma cells adjacent to xylem vessels (Fig. 9r,s).
Fig. 6a–o Light microscopy ontransverse sections andlongitudinal sections of rootsfrom young potato plants fromtransgenic lines, pBj47-P10 andpBj17-P6, and untransformedpotato after 5 days in MS liquidmedium containing R. solani.a, d Transverse section oftransgenic line pBj47-P10.b, e Transverse section oftransgenic line pBj17-P6.c, f, g Transverse sections ofuntransformed potato.h, i Magnifications froma. j Longitudinal section oftransgenic line pBj47-P10.k, l Magnifications from j.m Longitudinal section ofuntransformed potato root.n, o Magnifications from m atthe arrowed regions. Redarrowheads Intact fungus, redarrows disintegrating fungus,green arrowheads collapsedplant cells. Bars = 50 lm(a–g, j–k, m, n), 10 lm (h, i, l, o)
725
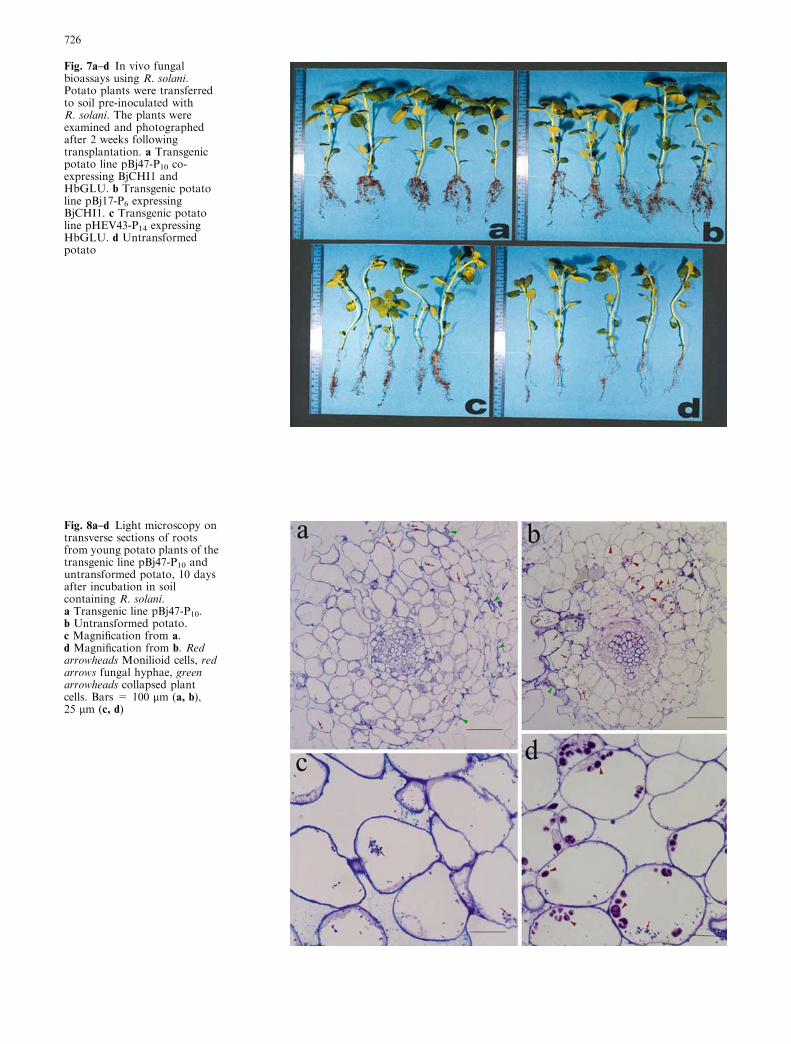
Fig. 7a–d In vivo fungalbioassays using R. solani.Potato plants were transferredto soil pre-inoculated withR. solani. The plants wereexamined and photographedafter 2 weeks followingtransplantation. a Transgenicpotato line pBj47-P10 co-expressing BjCHI1 andHbGLU. b Transgenic potatoline pBj17-P6 expressingBjCHI1. c Transgenic potatoline pHEV43-P14 expressingHbGLU. d Untransformedpotato
Fig. 8a–d Light microscopy ontransverse sections of rootsfrom young potato plants of thetransgenic line pBj47-P10 anduntransformed potato, 10 daysafter incubation in soilcontaining R. solani.a Transgenic line pBj47-P10.b Untransformed potato.c Magnification from a.d Magnification from b. Redarrowheads Monilioid cells, redarrows fungal hyphae, greenarrowheads collapsed plantcells. Bars = 100 lm (a, b),25 lm (c, d)
726

Discussion
Using northern blot analysis, we show here that themRNA encoding BjCHI1, a chitinase distinct from pre-viously reported chitinases because of the presence oftwo-chitin binding domains, is induced by R. solaniinfection. We also detected a corresponding increase inBjCHI1 protein on western blot analysis of leaf tissuefrom the infected B. juncea seedlings. Our results suggestthat the BjCHI1 mRNA and its corresponding proteinaccumulate following R. solani infection, implicating arole in fungal defense. In a resistant cultivar of B. napus,the mRNA of ChB4, a chitinase with one chitin-bindingprotein, was induced by Phoma lingam infection and wasdetected within a day on northern blot analysis (Ras-mussen et al. 1992), as in the case for BjCHI1 mRNAfollowing R. solani infection.
The induction of BjCHI1 by R. solani infection led usto further investigate its potential for crop protection byits heterologous expression. Protection against soil-borne R. solani by the co-expression of BjCHI1 andHbGLU would be particularly pertinent in applicationsfor transgenic potato because root-derived tubers areharvested from the soil. The stunted root developmentseen in untransformed potato and the HbGLU-expressing plants are likely due to fungal invasion,resulting in destruction of the epidermal, cortical andvascular tissues. This reduces the capacity of the rootsystem to transport water and other solutes, critical forthe development not only of the roots, but the wholeplant. Hence it was not surprising that stunted rootdevelopment was correlated with reduced foliage inuntransformed potato and HbGLU transformants.
With chitinases possessing one chitin-binding domain,Mauch et al. (1988) have shown that the in vitro com-bination of such a chitinase with b-1,3-glucanase stronglyinhibits fungal growth. Broglie et al. (1991) demonstratedthat the constitutive expression of a bean chitinase intransgenic tobacco confers fungal protection against R.solani. Later Zhu et al. (1994) showed that the constitu-tive expression of a rice chitinase with an alfalfa b-1,3-glucanase resulted in further enhanced fungal protectionof transgenic tobacco against Cercospora nicotiane whencompared to transgenic lines that expressed eitherchitinase or glucanase per se. However, in these previousstudies, investigations using light microscopy and TEMwere not carried out. Benhamou et al. (1993) used TEMto examine the effect of the expression of a bean chitinasein transgenic canola and correlated resistance with anti-microbial activity of the bean chitinase in planta.
Using light microscopy and TEM, we here report theeffect in planta of the expression of both b-1,3-glucanaseand a unique hemagglutinating chitinase with two chi-tin-binding domains. Examination of transgenic roots ofline pBj47-P10, following R. solani infection in MSsolution or in soil, showed the presence of collapsedplant cells, primarily at the epidermis. These epidermalcells form the first line of defense against pathogens and
their degeneration and subsequent lignification wouldenhance protection. Similar collapsed cells at the endo-dermis seen in the longitudinal root sections of linepBj47-P10 would limit penetration of fungus into thevascular stele. The electron-dense material seen at wallappositions of epidermal cells is due to the impregnationof anti-microbial phenolics (Foster et al. 1983). In TEM,we also observed accumulation of darkly stained elec-tron-dense deposits in the vicinity of disintegrating (inline pBj47-P10) and intact (in untransformed potato)fungal hyphae, and conclude that they are produced bythe plant in defense. Observations of gaps in xylem wallsin untransformed potato roots, in the vicinity of fungalcells, suggest that the fungus secretes extracellular cell-wall-digesting enzymes, likely cutinases, pectinases andcellulases, which are important in host penetration(Weinhold and Sinclair 1999). As in the case of theexpression of a bean endochitinase in transgenic canola(Benhamou et al. 1993), we too, observed degradation ofthe plant cell wall at some distance from the precedingfungus, reinforcing the view that the fungal extracellularhydrolytic enzymes are diffusible. It appears that inuntransformed potato, R. solani penetrates the host viathese gaps, as well as through the pits in the xylem,which we observed were ramified with fungus.
Disintegrating fungus, consistently seen in lightmicroscopy and in TEM of root sections from transgenicline pBj47-P10, was presumably the result of the actionof constitutively expressed BjCHI1 and HbGLU. Thesedisintegrating cells showed decreased protoplasmiccontent and/or degenerated fungal cell walls. It appearsthat the co-expression of BjCHI1 and HbGLU actseffectively in degrading the fungal cell wall leading toprotoplasm leakage. The presence of more hyphae in theBjCHI1-expressing line than the BjCHI1 and HbGLUco-expressing line, as detected by light microscopy,suggests that BjCHI1 and HbGLU work synergisticallyin planta. This is consistent with observations of theirsynergistic effect in T. viride bioassays. Two forms(Roberts 1999) of the fungus, hyphae from mycelia andlarger monilioid cells, were observed in untransformedpotato root by TEM, consistent with their detection insuch root sections by light microscopy.
In transgenic line pBj47-P10, fungal-derived elicitorsresulting from the action of BjCHI1 and HbGLU on theinvading fungus, as well as, plant-derived elicitors fromthe collapsing root epidermal cells, could trigger furtherplant defense responses. It has been reported thatchitosan oligomers, derived from fungal cell walls, notonly inhibit fungal growth but also induce plant defenseresponses (Leuba and Stossel 1986; Hadwinger et al.1988; Benhamou and Theriault 1992; Benhamou et al.1994). Furthermore, these elicitors could trigger lignifi-cation (Isaac 1992) at the collapsed cells, reinforcing theepidermis to form a structural barrier that limits furtherinvasion. Observation of more collapsed root epidermalcells in transgenic line pBj47-P10 was correlated with lessfungus in its epidermis, cortex and the vascular tissue. Incontrast, fewer collapsed epidermal cells resulted in
727

fungal ramification in the epidermis, cortex and thevascular tissue of wild-type root sections.
The use of BjCHI1 in crop protection is promisingbecause of its chimeric combination of two chitin-bind-
ing proteins at its N-terminus and a chitinase catalyticdomain at its C-terminus. It appears well prepared forplant defense, with the combined protective propertiesof chitinases and lectins with multiple chitin-binding
728

domains. Other than its anti-fungal action, it remains tobe determined if BjCHI1 shows any anti-bacterial andanti-insect action. By the duplication of its chitin-bind-ing domain, it resembles UDA, which is known topossess insect anti-nutrient activity (Chrispeels andRaikhel 1991). The extra chitin-binding domain inBjCHI1 seems to have given it the advantage of agglu-tination ability over other known chitinases that have atmost only one such domain. The significance of suchagglutination ability in enhancing its anti-microbial ac-tion will be further investigated.
Acknowledgments We thank Prof. Z.P. Lin (Peking University) forwild-type potato Desiree, Prof. E.C. Pua (National University ofSingapore) for B. juncea seeds, X.Z. Ouyang for technical assis-tance in microscopy, A. Mak for N-terminal sequencing, and W.Ho and P. Tam for helpful discussions. This project was supportedby the Department of Botany (University of Hong Kong), theBiotechnology Research Institute (Hong Kong University of Sci-ence and Technology) and the Research Grants Council of HongKong (Plant and Fungal Biotechnology Area of Excellence). K.J.Z.(Postdoctoral Fellowship) and K.L.F. (Postgraduate Studentship)were supported by the University of Hong Kong.
References
Abeles FB, Forrence LE (1970) Temporal and hormonal control ofb-1,3-glucanase in Phaseolus vulgaris L. Plant Physiol 45:395–400
Banville GJ, Carling DE, Otrysko BE (1999) Rhizoctonia diseaseon potato. In: Sneh B, Jabaji-Hare S, Neate S, Dijst G(eds) Rhizoctonia species: taxonomy, molecular biology, ecol-ogy, pathology and disease control. Kluwer, Dordrecht,pp 321–330
Benhamou N, Theriault G (1992) Treatment with chitosan en-hances resistance of tomato plants to the crown and root rotpathogen, Fusarium oxysporum f. sp. radicis-lycopersici. PhysiolMol Plant Pathol 41:33–52
Benhamou N, Broglie K, Chet I, Broglie R (1993) Cytology ofinfection of 35S-bean chitinase transgenic canola plants byRhizoctonia solani: cytochemical aspects of chitin breakdown invivo. Plant J 4:295–305
Benhamou N, Lafontaine PJ, Nicole M (1994) Seed treatment withchitosan induces systemic resistance to Fusarium crown androot rot in tomato plants. Phytopathology 84:1432–1444
Boller T (1985) Induction of hydrolases as a defense reactionagainst pathogens. In: Key JL, Kosuge T (eds) Cellular andmolecular biology of plant stress. Liss, New York, pp 247–262
Boller T (1992) Biochemical analysis of chitinase and b-1,3-glu-canses. In: Gurr SJ, McPherson MJ, Bowles DJ (eds) Molecularplant pathology: a practical approach, vol II. IRL Press, Ox-ford, pp 23–30
Boller T, Gehri A, Mauch F, Vogeli U (1983) Chitinase in beanleaves: induction by ethylene, purification, properties, andpossible function. Planta 157:22–31
Bradford MM (1976) A rapid and sensitive method for the quan-titation of microgram quantities of protein utilizing the prin-ciple of protein–dye binding. Anal Biochem 72:248–254
Broglie K, Chet I, Holliday M, Cressman R, Biddle P, Knowlton S,Mauvais CJ, Broglie R (1991) Transgenic plants with enhancedresistance to the fungal pathogen Rhizoctonia solani. Science254:1194–1197
Chrispeels MJ, Raikhel NV (1991) Lectins, lectin genes, and theirrole in plant defense. Plant Cell 3:1–9
Chye ML, Cheung KY (1995) b-1,3-glucanase is highly-expressedin laticifers of Hevea brasiliensis. Plant Mol Biol 29:397–402
Dellaporta SL, Wood J, Hicks JB (1983) A plant DNA mini-preparation: version II. Plant Mol Biol Rep 1:19–21
Dietze J, Blau A, Willmitzer L (1995) Agrobacterium-mediatedtransformation of potato (Solanum tuberosum). In: Potrykus I,Spangenberg G (eds) Gene transfer to plants. Cold SpringHarbor Laboratory Press, New York, pp 24–29
Does M, Houterman PM, Dekker HL, Cornelissen BJ (1999)Processing, targeting and antifungal activity of stinging nettleagglutinin in transgenic tobacco. Plant Physiol 120:421–431
Foster RC, Rovira AD, Cock TW (1983) Ultrastructure of theroot-soil interface. The American Phytopathogical Society, StPaul, MN
Fung KL, Zhao KJ, He ZM, Chye ML (2002) Brassica junceachitinase BjCHI1 binds chitin and shows antifungal activity invitro. Plant Mol Biol 50:283–294
Hadwinger LA, Chiang C, Victory S, Horovitz D (1988) Themolecular biology of chitosan in plant–pathogen interactionsand its application to agriculture. In: Skjak G, Anthonsen B,Sandford P (eds) Chitin and chitosan: sources, chemistry, bio-chemistry, physical properties and applications. Elsevier,Amsterdam, pp 119–138
Isaac S (1992) Fungal–plant interactions. Chapman and Hall,London
Iseli B, Boller T, Neuhaus J-M (1993) The N-terminal cysteine-richdomain of tobacco class I chitinase is essential for chitinbinding but not for catalytic or antifungal activity. PlantPhysiol 103:221–226
Jach G, Gornhardt B, Mundy J, Logemann J, Pinsdorf E, Leah R,Schell J, Maas C (1995) Enhanced quantitative resistanceagainst fungal disease by combinatorial expression of differentbarley antifungal proteins in transgenic tobacco. Plant J 8:97–109
Kush A, Goyvaerts E, Chye ML, Chua NH (1990) Laticifer specificgene expression of Hevea brasiliensis (rubber tree). Proc NatlAcad Sci USA 87:1787–1790
Fig. 9a–s Electron micrographs of transverse sections of roots fromyoung potato plants of transgenic line pBj47-P10 (a–j) anduntransformed potato (k–s), 10 days after incubation in soilcontaining R. solani. a Extracellular intact fungus (red arrows)compared to intracellular disintegrating fungus (red arrowheads)in a root epidermal cell of transgenic potato line pBj47-P10.b, c Magnifications of a show intracellular disintegrating fungus.d Collapsed root epidermal cell (green arrowhead) with wallappositions (WA), extracellular intact fungus and intracellulardisintegrating fungus in a root epidermal cell of transgenic potatoline pBj47-P10. e Magnification of d shows intracellular disintegrat-ing fungus below plant cell membrane. f Collapsed root epidermalcells show wall appositions. g Transverse section of root vasculartissue from transgenic potato line pBj47-P10. h, i Magnifications ofxylem (X) in g show intracellular disintegrating fungal hyphae anddarkly-stained deposits (orange arrowhead). j Magnification ofimmature xylem (IX) in g shows disintegrating fungus and darklystained deposits. k Transverse section of root vascular tissue fromuntransformed potato shows fungus in xylem and parenchyma (P).l Magnification of xylem in k shows fungal hyphae and darklystained deposits. m Magnification of l shows darkly staineddeposits. n Magnification of a parenchyma cell in k shows intactfungal cells. o Transverse section of root vascular tissue fromuntransformed potato shows fungus and gaps (blue arrowheads) inthe plant cell wall. p Magnification of top parenchyma cell in oshows gap in cell wall and intact fungal cells. q Magnification ofbottom xylem vessel in o shows an intact fungal cell. r Transversesection of intercellular space (IS) neighbouring parenchyma cells inuntransformed potato root. s Magnification of fungal cell from r.Red arrowhead Intact fungus, red arrow degenerating fungus, greenarrowhead collapsed epidermal cell, orange arrowhead darklystained deposits, CW plant cell wall, CM plant cell membrane, ISintercellular space, IX immature xylem, LB lipid body, N plantnucleus, P parenchyma cell, WA wall appositions, X xylem. Bars =600 nm (a, i, n, p, q, s), 200 nm (b, c, e, h, j, m), 2 lm (d, f, g, k, l, o, r)
b
729

Leuba JL, Stossel P (1986) Chitosan and other polyamines: anti-fungal activity and interaction with biological membranes. In:Muzzarelli R, Jeauniaux C, Gooday GW (eds) Chitin in natureand technology. Plenum, New York, pp 215–222
Lin W, Anuratha CS, Datta K, Potrykus I, Muthukrishnan S,Datta SK (1995) Genetic engineering of rice for resistance tosheath blight. Bio/Technology 13:686–691
Mauch F, Mauch-Mani B, Boller T (1988) Antifungal hydrolases inpea tissue II. Inhibition of fungal growth by combinations ofchitinase and b-1,3-glucanase. Plant Physiol 88:936–942
Nagy F, Kay SA, Chua NH (1988) Analysis of gene expression intrangenic plants. In: Gelvin SV, Schilperoort RA (eds) Plantmolecular biology manual. Kluwer, Dordrecht, pp B4:1–29
Rasmussen U, Bojsen K, Collinge DB (1992) Cloning and char-acterization of a pathogen-induced chitinase in Brassica napus.Plant Mol Biol 20:277–287
Roberts P (1999) Rhizoctonia-forming fungi. A taxonomic guide.Royal Botanic Gardens, Kew
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular cloning: alaboratory manual. Cold Spring Harbor Laboratory Press,New York
Schlumbaum A, Mauch F, Vogeli U, Boller T (1986) Plantchitinases are potent inhibitors of fungal growth. Nature324:365–367
Taylor KACC (1995) A modification of the phenol–sulfuric acidmethod of total sugar determination. Appl Biochem Biotechnol53:207–214
Van Damme EJM, Barre A, Rouge P, Peumans WJ (2004) Potatolectin: an updated model of a unique chimeric plant protein.Plant J 37:34–45
Weinhold AR, Sinclair JB (1999) Rhizoctonia solani: penetration,colonization and host response. In: Sneh B, Jabaji-Hare S,Neate S, Dijst G (eds) Rhizoctonia species: taxonomy, molec-ular biology, ecology, pathology and disease control. Kluwer,Dordrecht, pp 163–174
Wirth SJ, Wolf GA (1990) Dye-labelled substrates for the assayand detection of chitinase and lysozyme activity. J MicrobiolMethods 12:197–205
Wright LM, Van Damme EJM, Barre A, Allens AK, Van Leuven,Reynolds CD, Rouge P, Peumans W (1999) Isolation, charac-terization, molecular cloning and molecular modeling of twolectins of different specificities from bluebell (Scilla campanu-lata) bulbs. Biochem J 340:299–308
Zhao KJ, Chye ML (1999) Methyl jasmonate induces expression ofa novel Brassica juncea chitinase with two chitin-binding do-mains. Plant Mol Biol 40:1009–1018
Zhu Q, Maher EA, Masoud S, Dixon RA, Lamb CJ (1994) En-hanced protection against fungal attack by constitutive co-expression of chitinase and glucanase genes in trangenic to-bacco. Bio/Technology 12:807–812
730
Copyright © 2022 FDOKUMEN




















