
Alterations in PACAP-38-like immunoreactivity in the plasma during ictal and interictal periods of...
11
Original Article Alterations in PACAP-38-like immunoreactivity in the plasma during ictal and interictal periods of migraine patients Bernadett Tuka 1,2 , Zsuzsanna Helyes 3,4 , Adrienn Markovics 3,4 , Tere ´z Bagoly 3 , Ja ´nos Szolcsa ´nyi 3 , Nikoletta Szabo ´ 1,5 , Eszter To ´ th 1 , Zsigmond Tama ´s Kincses 1,5 , La ´szlo ´ Ve ´ csei 1,2 and Ja ´nos Tajti 1 Abstract Background: Recent studies on migraineurs and our own animal experiments have revealed that pituitary adenylate cyclase-activating polypeptide-38 (PACAP-38) has an important role in activation of the trigeminovascular system. The aim of this study was to determine the PACAP-38-like immunoreactivity (LI) in the plasma of healthy subjects, and parallel with the calcitonin gene-related peptide (CGRP)-LI in migraine patients in the ictal and interictal periods. Methods: A total of 87 migraineurs and 40 healthy control volunteers were enrolled in the examination. Blood samples were collected from the cubital veins in both periods in 21 patients, and in either the ictal or the interictal period in the remaining 66 patients, and were analysed by radioimmunoassay. Results: A significantly lower PACAP-38-LI was measured in the interictal plasma of the migraineurs as compared with the healthy control group (p < 0.011). In contrast, elevated peptide levels were detected in the ictal period relative to the attack-free period in the 21 migraineurs (p PACAP-38 < 0.001; p CGRP < 0.035) and PACAP-38-LI in the overall population of migraineurs (p < 0.009). A negative correlation was observed between the interictal PACAP-38-LI and the disease duration. Conclusion: This is the first study that has provided evidence of a clear association between migraine phases (ictal and interictal) and plasma PACAP-38-LI alterations. Keywords Migraine, ictal/interictal periods, plasma PACAP-38-LI Date received: 17 October 2012; revised: 6 December 2012; 8 January 2013; accepted: 15 January 2013 Introduction It is possible to analyse the mechanisms of pain in animal models, but there are no really clinically rele- vant systems to appropriately mimic the human speci- ficity of headache diseases. Human investigations are therefore particularly important to identify the key mediators responsible for the development and progres- sion of migraine. Several hypotheses have been proposed to explain the pathological mechanisms of migraine. One of the most widely accepted theories relates to activation of the trigeminovascular system (TS), which involves vas- cular and neuronal elements (1). The TS consists of the 1 Department of Neurology, Faculty of Medicine, University of Szeged, Hungary 2 Neuroscience Research Group of the Hungarian Academy of Sciences and University of Szeged, Hungary 3 Department of Pharmacology and Pharmacotherapy, Faculty of Medicine, University of Pe ´cs, Hungary 4 Ja ´nos Szenta ´gothai Research Center, University of Pe ´cs, Hungary 5 International Clinical Research Center, St. Anne’s University Hospital Brno, Czech Republic Corresponding author: Ja ´nos Tajti, Department of Neurology, Faculty of Medicine, University of Szeged, H-6725 Szeged, Semmelweis u. 6, Hungary. Email: [email protected] Cephalalgia 33(13) 1085–1095 ! International Headache Society 2013 Reprints and permissions: sagepub.co.uk/journalsPermissions.nav DOI: 10.1177/0333102413483931 cep.sagepub.com
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of Alterations in PACAP-38-like immunoreactivity in the plasma during ictal and interictal periods of...

Original Article
Alterations in PACAP-38-likeimmunoreactivity in the plasmaduring ictal and interictal periodsof migraine patients
Bernadett Tuka1,2, Zsuzsanna Helyes3,4, Adrienn Markovics3,4,Terez Bagoly3, Janos Szolcsanyi3, Nikoletta Szabo1,5,Eszter Toth1, Zsigmond Tamas Kincses1,5, Laszlo Vecsei1,2
and Janos Tajti1
Abstract
Background: Recent studies on migraineurs and our own animal experiments have revealed that pituitary adenylate
cyclase-activating polypeptide-38 (PACAP-38) has an important role in activation of the trigeminovascular system.
The aim of this study was to determine the PACAP-38-like immunoreactivity (LI) in the plasma of healthy subjects,
and parallel with the calcitonin gene-related peptide (CGRP)-LI in migraine patients in the ictal and interictal periods.
Methods: A total of 87 migraineurs and 40 healthy control volunteers were enrolled in the examination. Blood samples
were collected from the cubital veins in both periods in 21 patients, and in either the ictal or the interictal period in the
remaining 66 patients, and were analysed by radioimmunoassay.
Results: A significantly lower PACAP-38-LI was measured in the interictal plasma of the migraineurs as compared with the
healthy control group (p< 0.011). In contrast, elevated peptide levels were detected in the ictal period relative to the
attack-free period in the 21 migraineurs (pPACAP-38< 0.001; pCGRP< 0.035) and PACAP-38-LI in the overall population of
migraineurs (p< 0.009). A negative correlation was observed between the interictal PACAP-38-LI and the disease
duration.
Conclusion: This is the first study that has provided evidence of a clear association between migraine phases (ictal and
interictal) and plasma PACAP-38-LI alterations.
Keywords
Migraine, ictal/interictal periods, plasma PACAP-38-LI
Date received: 17 October 2012; revised: 6 December 2012; 8 January 2013; accepted: 15 January 2013
Introduction
It is possible to analyse the mechanisms of pain inanimal models, but there are no really clinically rele-vant systems to appropriately mimic the human speci-ficity of headache diseases. Human investigations aretherefore particularly important to identify the keymediators responsible for the development and progres-sion of migraine.
Several hypotheses have been proposed to explainthe pathological mechanisms of migraine. One of themost widely accepted theories relates to activation ofthe trigeminovascular system (TS), which involves vas-cular and neuronal elements (1). The TS consists of the
1Department of Neurology, Faculty of Medicine, University of Szeged,
Hungary2Neuroscience Research Group of the Hungarian Academy of Sciences
and University of Szeged, Hungary3Department of Pharmacology and Pharmacotherapy, Faculty of
Medicine, University of Pecs, Hungary4Janos Szentagothai Research Center, University of Pecs, Hungary5International Clinical Research Center, St. Anne’s University Hospital
Brno, Czech Republic
Corresponding author:
Janos Tajti, Department of Neurology, Faculty of Medicine, University of
Szeged, H-6725 Szeged, Semmelweis u. 6, Hungary.
Email: [email protected]
Cephalalgia
33(13) 1085–1095
! International Headache Society 2013
Reprints and permissions:
sagepub.co.uk/journalsPermissions.nav
DOI: 10.1177/0333102413483931
cep.sagepub.com

primary sensory pseudounipolar neurons in the trigem-inal ganglion (TRIG), its terminals and the meningealvasculature. The peripheral afferents innervate thesupratentorial dura mater, the dural vasculature andthe large and small pial arteries on the surface of thebrain. The central fibres project to the area of thesecond-order neurons in the brainstem, the caudal tri-geminal nucleus (TNC). The third-order neurons arelocated in the thalamus. The activation mechanisminvolves the release of neuropeptides from the terminals(e.g. calcitonin gene-related peptide (CGRP), vaso-active intestinal peptide (VIP) and substance P (SP)(2), and functional changes such as neurogenic inflam-mation (protein extravasation and vasodilation), mastcell degranulation and sensitisation (3).
Among the numerous ‘‘migraine-related’’ processesand substances, the vasoactive pituitary adenylatecyclase-activating polypeptide (PACAP) has beenpotentially implicated in the pathophysiology ofmigraine (4,5). PACAP belongs to the VIP/secretin/glucagon neuropeptide superfamily, and is consideredto be a ‘‘brain-gut peptide’’ by virtue of its widespreadexpression and functions in the human body. PACAPexists in two forms: PACAP-27 and PACAP-38; thelatter is expressed predominantly in mammals.PACAP acts at one specific (PAC1) and two non-specificreceptors, which are shared between PACAP and VIP(VPAC1 and VPAC2). This can be explained by the 68%structural homology between PACAP and VIP (6–8).
This implies that PACAP-38 also has a vasodilatingeffect (9), and a broad range of data suggest that it is anintegrator of nociceptive and sensitisation processes,besides being involved in neurogenic inflammation(10–13).
PACAP is presented in the primary sensory neuronsof the TRIG (14), the parasympathetic otic and thesphenopalatine ganglia (15,16). Moreover, PACAP-38is found in the cell bodies and nerve fibres of the humanTNC and the upper regions of the cervical spinal cord,which suggests that PACAP may be closely related tothe TS (2,17). We earlier furnished evidence for thishypothesis with animal experimental results. PACAP-deficient mice displayed reduced light-aversive behav-iour (photophobia), while decreased meningeal bloodflow and c-fos expression in the TRIG and TNC weredetected relative to wild-type mice after nitroglycerol(NTG)-induced TS activation (18). Further, elevatedlevels of PACAP-27 and PACAP-38 were measured inthe plasma and the TNC 90 and 180 minutes after therespective stimulation in two animal models of TS acti-vation (19). These results suggest that PACAP may bean important mediator, and therefore a diagnosticmarker of TS activation.
The available human data point to the involvementof PACAP in the mechanisms of migraine. Schytz et al.
demonstrated that the intravenous administration ofPACAP-38 causes headache in healthy subjects, andmigraine-like attacks in migraine patients withoutaura, six hours on average after starting theinfusion (4). However, no clinical data are availableon endogenous alterations in PACAP levels in relationto migraine.
Based on the literature, we have hypothesised thatthe plasma concentration of PACAP-38 increasesduring migraine attacks. Our aims were therefore toreveal the potential relationships between thePACAP-38 levels of human plasma and the presenceof migraine headache. Radioimmunoassay (RIA) meas-urements were carried out on peripheral blood plasmasamples in order to show alterations of PACAP-38-likeimmunoreactivity (PACAP-38-LI) in migraine patientsduring the ictal and interictal periods in comparisonwith healthy control subjects. In addition, the clinicalfeatures of the disease, as well as plasma CGRP-likeimmunoreactivity (CGRP-LI) were also compared tothe PACAP-38-LI to explore possible correlations.
Materials and methods
Participants
A total of 87 migraine patients with or without auraand 40 healthy control subjects were enrolled in thisstudy. The migraineurs were selected in accordancewith the criteria of the Headache ClassificationCommittee of the International Headache Society2004 (20). The study groups were age matched. Thedemographic and clinical characteristics of the patientand control populations are summarised in Table 1.
A detailed questionnaire was used to compile ahomogeneous group of migraineurs as concerns the fea-tures of their migraine disease: the duration of themigraine, the attack frequency, allodynia (21) and theseverity of pain during attacks as measured on a visualanalogue scale (VAS). The relation of migraine attacksto the menstrual cycle (22) and to the presence of othernon-migraine, chronic pain disorders (lumbago, low-back pain, knee- and hip-joint arthrosis) was alsoassessed. Depression was not clinically diagnosed inany of the cases. Healthy volunteers serving as controlswere screened for non-reported/non-treated headaches.Subjects (patients and controls) who displayed any sig-nificant and serious non-migraine chronic disorderswere excluded from the study.
Study design and procedures
The study was approved by the Ethics Committeeof the Faculty of Medicine, University of Szeged(87/2009). All study participants gave their written
1086 Cephalalgia 33(13)

informed consent, in accordance with the Declarationof Helsinki. There were no restrictions as regardsfood and drink intake. Blood samples were drawnfrom migraineurs during an attack and/or in anattack-free period. Affected patients were asked notto start their usual attack treatment until blood sam-ples had been taken. Accordingly, 80 interictal and 28ictal samples were collected. From among the 87patients, blood samples could be collected in bothperiods from 21 migraineurs. Data of the populationof 21 migraineurs are shown in Table 2. A singleblood sample was taken from each control. Bloodsamples (6ml per subject) were taken in a sitting pos-ition during rest from the cubital vein and collectedin ice-cold glass tubes containing the anticoagulantethylenediaminetetraacetic acid (EDTA, 12mg) andthe protease inhibitor aprotinin (Trasylol, 1200 IU),and kept at 4�C until centrifugation (2000 rpm for10 minutes at 4�C). Plasma samples were stored at�80�C until the PACAP-38-LI and CGRP-LI weremeasured by RIA.
RIA measurements and data acquisition
Plasma concentrations of PACAP-38 were determinedwith specific and sensitive RIA techniques developedearlier in our laboratory (23). The PACAP-38 anti-serum 88111-3 was raised in rabbits with synthetic pep-tides conjugated to bovine serum albumin (BSA) orthyroglobulin with glutaraldehyde or carbodiimide.The high specificity and C-terminal sensitivity of thisantibody were confirmed by cross-reactivity studies,and no cross-reactivity was found with PACAP-27 orwith other related neuropeptides in either case.Following centrifugation of the plasma samples(2000 rpm at 4�C for 10 minutes), the peptide wasextracted from the plasma into three volumes of abso-lute alcohol.
After precipitation and a second centrifugation(2000 rpm at 4�C for 10 minutes), the samples weredried under a nitrogen flow and resuspended in 300mlof assay buffer before RIA determination in order toachieve a 10 times higher concentration for the RIAprocedure (10,23). The tracers were mono-125I-labelledpeptides prepared in our laboratory. Synthetic peptideswere used as RIA standards in concentrations rangingfrom 0 to 1000 fmol/ml. The assay was prepared in 1ml0.05M (pH 7.4) phosphate buffer containing 0.1Msodium chloride, 0.25% (w/v) BSA and 0.05% (w/v)sodium azide. The antiserum (100 ml, 1:10,000 dilution),the RIA tracer (100ml, 5000 cpm/tube) and the stand-ard or unknown samples (100 ml) were measured intopolypropylene tubes with the assay buffer. After incu-bation for 48–72 hours at 4�C, the antibody-boundpeptide was separated from the free peptide by the add-ition of 100 ml of separating solution (10 g charcoal, 1 gdextran and 0.5 g commercial fat-free milk powder in100ml of distilled water). Following centrifugation(3000 rpm at 4�C for 15 minutes), the contents of thetubes were gently decanted and the radioactivity of theprecipitates was measured in a gamma counter(Gamma, type NZ310). The PACAP-38 concentrationsof the unknown samples were read from calibrationcurves.
Statistical analysis
Data are expressed as median� SD if not stated other-wise. The normality of the data was tested with theShapiro-Wilk test. Group comparisons were carriedout with Student’s unpaired, paired t tests and theWilcoxon-test with SPSS 17.0. Data were analysedwith a multivariate test (repeated-measure analysis ofvariance (ANOVA)) in the case of menstruation cycleand chronic pain condition related to PACAP-38 level.Statistical significance was accepted at p< 0.05.
Table 1. Mean data on healthy control volunteers and 87 migraineurs (age, gender, type of migraine (with or without aura), disease
duration (years), attack frequency, allodynia and VAS score are shown.
Healthy control subjects n¼ 40
Mean age (years) Gender
36.60� 11.84
, n¼ 26
< n¼ 14
Migraine
patients
n¼ 87
Mean age
(years) Gender
Type of
migraine
Mean duration
of disease
(years)
Mean attack
frequency/
year
Allodynia
n¼ 23
Mean
VAS-score
37.91� 10.17 , n¼ 79
< n¼ 8
MA n¼ 18
MO n¼ 69
12.99� 9.61 36.80� 28.44 0. level n¼ 64
1. level n¼ 9
2. level n¼ 9
3. level n¼ 5
7.94� 1.75
MA: migraine with aura; MO: migraine without aura; allodynia: 0: absence of allodynia; 1: mild allodynia; 2: moderate allodynia; 3: serious allodynia;
VAS: visual analogue score.
Tuka et al. 1087

Results
Differences in plasma PACAP-38-LI betweenmigraineurs and healthy controls
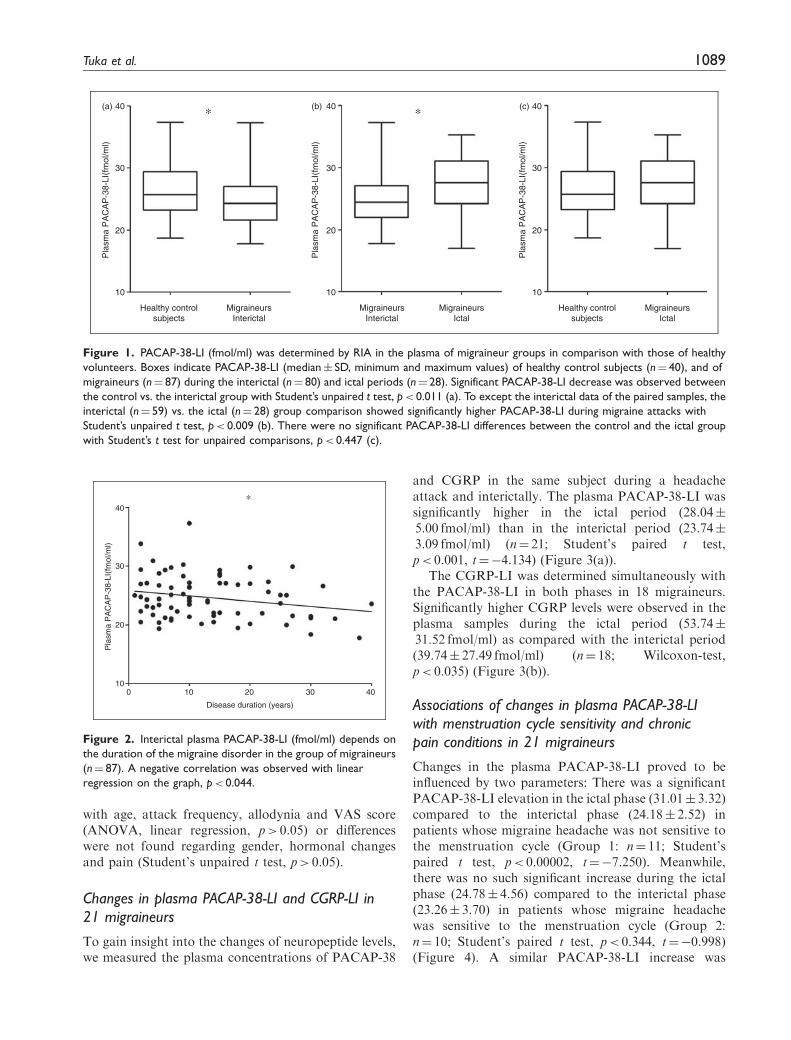
As concerns the total of 87 migraine patients (n¼ 87), asignificantly lower PACAP-38-LI was determined in theinterictal plasma of the migraineurs (n¼ 80; 24.60�3.59 fmol/ml) than in that of the healthy volunteers(n¼ 40; 26.54� 4.43 fmol/ml; Student’s unpaired t test,p< 0.011, t¼ 2.578) (Figure 1(a)). However, the plasmasamples from the patients during their migraine attacks(n¼ 28) exhibited a significantly higher PACAP-38 con-centration (27.39� 4.67 fmol/ml) as compared withthe interictal samples (n¼ 59; 24.91� 3.73 fmol/ml;Student’s unpaired t test, p< 0.009, t¼�2.676)
(the interictal data of those 21 migraineurs whoseplasma samples were collected from both periodswere excluded from this analysis to avoid the statisticalproblems caused by the paired samples) (Figure 1(b)).No difference was found when the ictal samples werecompared with those of the controls (Student’s t testfor unpaired comparisons, p< 0.447, t¼�0.765)(Figure 1(c)).
Association between plasma PACAP-38-LI andduration of migraine
A negative correlation was found between the durationof the migraine and the interictal PACAP-38-LI(n¼ 87; linear regression, p< 0.044, R¼ 0.231)(Figure 2). Plasma PACAP-38-LI did not correlate
Table 2. Demographic data on 21 migraineurs, whose samples from both the ictal and the interictal periods were analysed. The age,
gender, type of migraine (with or without aura), disease duration (years), attack frequency, allodynia, VAS score, PACAP-38-LI in
plasma samples (fmol/ml) originating from interictal and ictal phases, the time of the previous attack before interictal blood sampling
(days) and the duration of the present headache (hours) are shown for each patient.
Patient Age Gender
Type of
migraine
Duration
of disease
(years)
Attack
frequency/
year Allodynia
VAS
score
Interictal
sample
PACAP-38-LI
(fmol/ml)
Previous
attack
(days ago)
Ictal sample
PACAP-38-LI
(fmol/ml)
Duration
of headache
before
sampling
(hours)
1 24 , MO 3 17 0 10 24.30 9 26.50 4
2 39 , MO 26 30 0 9 20.00 2 29.16 96
3 46 , MO 18 24 0 9 19.50 7 23.90 4
4 44 , MO 30 47 3 7 18.40 9 20.90 12
5 45 , MO 2 12 0 9 26.94 6 20.40 5
6 27 , MO 14 30 2 8 20.50 6 17.00 24
7 35 , MO 20 12 0 8 27.00 6 31.20 7
8 39 , MO 30 109 2 9 21.20 10 25.90 7
9 31 < MA 10 52 0 7 24.11 7 33.21 8
10 21 , MO 5 24 0 7 20.41 7 25.18 3
11 38 , MO 2 30 0 7 22.34 5 31.81 24
12 46 , MO 10 36 0 8 26.36 7 28.05 10
13 17 < MO 10 12 0 8 23.51 10 35.28 2
14 30 , MO 10 12 1 5 26.43 7 23.30 24
15 39 < MO 10 66 0 8 27.15 1 35.28 2
16 37 , MO 9 30 0 8 25.23 7 33.70 48
17 50 , MO 14 12 0 8 21.59 21 26.92 5
18 52 , MO 14 26 3 8 22.08 5 30.71 60
19 35 , MO 25 52 0 10 25.05 1 31.69 8
20 58 , MO 15 21 0 8 27.09 7 29.18 14
21 35 , MA 6 20 2 10 29.33 20 29.67 20
Mean 37.52 13.48 32.10 8.14 23.74 7.62 28.04 18.43
SD 10.34 8.61 23.23 1.20 3.09 4.95 5.00 23.33
MA: migraine with aura; MO: migraine without aura; allodynia: 0: absence of allodynia; 1: mild allodynia; 2: moderate allodynia; 3: serious allodynia; VAS:
visual analogue score; PACAP-38-LI: pituitary adenylate cyclase-activating polypeptide-38 like immunoreactivity.
1088 Cephalalgia 33(13)
with age, attack frequency, allodynia and VAS score(ANOVA, linear regression, p> 0.05) or differenceswere not found regarding gender, hormonal changesand pain (Student’s unpaired t test, p> 0.05).
Changes in plasma PACAP-38-LI and CGRP-LI in21 migraineurs
To gain insight into the changes of neuropeptide levels,we measured the plasma concentrations of PACAP-38
and CGRP in the same subject during a headacheattack and interictally. The plasma PACAP-38-LI wassignificantly higher in the ictal period (28.04�5.00 fmol/ml) than in the interictal period (23.74�3.09 fmol/ml) (n¼ 21; Student’s paired t test,p< 0.001, t¼�4.134) (Figure 3(a)).
The CGRP-LI was determined simultaneously withthe PACAP-38-LI in both phases in 18 migraineurs.Significantly higher CGRP levels were observed in theplasma samples during the ictal period (53.74�31.52 fmol/ml) as compared with the interictal period(39.74� 27.49 fmol/ml) (n¼ 18; Wilcoxon-test,p< 0.035) (Figure 3(b)).
Associations of changes in plasma PACAP-38-LIwith menstruation cycle sensitivity and chronicpain conditions in 21 migraineurs
Changes in the plasma PACAP-38-LI proved to beinfluenced by two parameters: There was a significantPACAP-38-LI elevation in the ictal phase (31.01� 3.32)compared to the interictal phase (24.18� 2.52) inpatients whose migraine headache was not sensitive tothe menstruation cycle (Group 1: n¼ 11; Student’spaired t test, p< 0.00002, t¼�7.250). Meanwhile,there was no such significant increase during the ictalphase (24.78� 4.56) compared to the interictal phase(23.26� 3.70) in patients whose migraine headachewas sensitive to the menstruation cycle (Group 2:n¼ 10; Student’s paired t test, p< 0.344, t¼�0.998)(Figure 4). A similar PACAP-38-LI increase was
10
Healthy controlsubjects
Healthy controlsubjects
MigraineursInterictal
MigraineursIctal
MigraineursInterictal
MigraineursIctal
20
Pla
sma
PA
CA
P-3
8-LI
(fm
ol/m
l)
30
40(a)
Pla
sma
PA
CA
P-3
8-LI
(fm
ol/m
l)
(b)
10
20
30
40
Pla
sma
PA
CA
P-3
8-LI
(fm
ol/m
l)
(c)
10
20
30
40∗ ∗
Figure 1. PACAP-38-LI (fmol/ml) was determined by RIA in the plasma of migraineur groups in comparison with those of healthy
volunteers. Boxes indicate PACAP-38-LI (median� SD, minimum and maximum values) of healthy control subjects (n¼ 40), and of
migraineurs (n¼ 87) during the interictal (n¼ 80) and ictal periods (n¼ 28). Significant PACAP-38-LI decrease was observed between
the control vs. the interictal group with Student’s unpaired t test, p< 0.011 (a). To except the interictal data of the paired samples, the
interictal (n¼ 59) vs. the ictal (n¼ 28) group comparison showed significantly higher PACAP-38-LI during migraine attacks with
Student’s unpaired t test, p< 0.009 (b). There were no significant PACAP-38-LI differences between the control and the ictal group
with Student’s t test for unpaired comparisons, p< 0.447 (c).
10
Disease duration (years)
20
Pla
sma
PA
CA
P-3
8-LI
(fm
ol/m
l)
30
∗40
0 10 20 30 40
Figure 2. Interictal plasma PACAP-38-LI (fmol/ml) depends on
the duration of the migraine disorder in the group of migraineurs
(n¼ 87). A negative correlation was observed with linear
regression on the graph, p< 0.044.
Tuka et al. 1089

detected in the ictal phase (30.01� 3.69) vs. the interictalphase (24.15� 2.47) of patients, who did not havechronic pain-related conditions (Group 1: n¼ 15;Student’s paired t test, p< 0.00005, t¼�5.716).However, there was no difference in the ictal phase(23.13� 4.64) compared to the interictal phase(22.71� 4.40) in patients, who had other non-migraine,chronic pain disorders (Group 2: n¼ six; Student’spaired t test, p< 0.833, t¼�0.222) (Figure 5).
To reveal the relationship between ictal-interictalPACAP-38 levels and menstruation and chronic pain,a repeated-measure ANOVA was used. A multivariatetest showed a significant main effect of PACAP-38measurements in different phases of the disease(F(1,19)¼ 22.579, p< 0.0001) and a main effect of head-ache related to menstruation cycle (F(1,19)¼ 22.257,
p< 0.0001). The interaction of the two factors wasalso significant (F(1,19)¼ 9.096, p< 0.007). A similarANOVA was carried out for identifying if ictal/interictal PACAP-38 level changes are influenced byconcomitant chronic pain conditions. While significantmain effects were identified by analysis (ictal/interictal:F(1,19)¼ 11.392, p< 0.003, chronic pain condition:F(1,19)¼ 0.553, p< 0.446), the interaction was not sig-nificant in this case (p> 0.05).
No correlations were found between the level of pep-tides and age, gender, attack frequency, allodynia orVAS score.
Discussion
This study provided the first evidence that PACAP-38concentration in the plasma of migraineurs is signifi-cantly lower in the interictal period as compared withthat of healthy volunteers, but increases during migraineattacks relative to the attack-free period. These resultssuggest that PACAP-38 might be an important medi-ator of the pathophysiology of migraine. The differencein the plasma levels of PACAP-38 in migraineurs in thetwo periods indicate that this peptide is involved in thedevelopment of the attacks.
Effects of PACAP-38 related to migraine
PACAP-38 can be released from both the central andthe peripheral terminals of the pseudounipolar neurons
0
50
100
150
Pla
sma
CG
RP
-LI(
fmol
/ml)
10
20
30
40(a)
(b)
Pla
sma
PA
CA
P-3
8-LI
(fm
ol/m
l)
∗
∗
Interictal Ictal
Interictal Ictal
Figure 3. PACAP-38-LI (fmol/ml) was determined by RIA on
plasma samples from migraineurs in both interictal and ictal
periods. (a). Plots of individual data for each patient (n¼ 21). A
significant difference was observed between the levels in the two
phases with Student’s paired t test, p< 0.001. CGRP-LI (fmol/ml)
was determined by RIA on plasma samples from migraineurs in
both interictal and ictal periods. (b). Plots of individual data for
each patient (n¼ 18). A significant difference was observed
between the levels in the two phases with Wilcoxon-test,
p< 0.035. The thick grey lines represent the mean values.
10
Group 1: Menstruation cycle-independent migraineurs
Group 2: Menstruation cycle-sensitive migraineurs
Interictal Ictal Interictal Ictal
20
30
40
Pla
sma
PA
CA
P-3
8-LI
(fm
ol/m
l)
∗
Figure 4. Plasma PACAP-38-LI (fmol/ml) in the interictal and
ictal periods of two subpopulations of 21 migraineurs charac-
terised on the basis of menstruation cycle dependence. Group 1:
menstruation cycle-independent migraineurs (n¼ 11); group 2:
migraine patients whose headache was sensitive to their men-
struation (n¼ 10). Each box represents the median� SD, min-
imum and maximum values of the results. A significant difference
in PACAP-38-LI was observed between the interictal and ictal
phases in group 1 with Student’s paired t test, p< 0.00002.
1090 Cephalalgia 33(13)

of the TRIG (24) in response to specific triggers. Itenters the systemic circulation, and binds to its recep-tors (PAC1, VPAC1 and VPAC2) on the meningeal ves-sels and second-order sensory neurons. Zagami andcolleagues published (25) that the stimulation of thesuperior sagittal sinus causes extracranial release ofPACAP. PACAP is known to induce neurogenicinflammation, mast cell degranulation, neuronal activa-tion and sensitisation (4,26). These effects can contrib-ute to the development of severe headache and possiblyto the aggravation of the attacks.
A magnetic resonance imaging (MRI) angiographicstudy revealed that PACAP-38 infusion-induced head-ache is related to significant dilation of the middle men-ingeal arteries (MMAs), in contrast with the middlecerebral arteries (27). The intracarotid infusion ofPACAP-38 produces significant dilation of the duralartery in the rat, and this administration route hasproved to be more effective than the intravenousmode (28). PACAP-38 also exerts potent vasodilatingaction on pressurised MMAs, which can be influencedby a PAC1 receptor antagonist. Multiple variants of thePAC1 receptor have been found in addition to theVPAC2 receptors in rat MMAs (29).
PACAP and VIP receptors in the human meningealand coronary arteries were recently characterised withpharmacological modulators, and these peptides werefound to have lower potency and efficacy in meningeal
vasculature than in coronary arteries. This study con-cluded that processes of PACAP-38-induced migraine-like headache may not involve meningeal vasodilationrather than sensitisation of peripheral and central sen-sory trigeminal fibres (9). The reported effects of aPACAP infusion regarding neurogenic inflammationand mast cell degranulation in relation to migraineare controversial (26). It was concluded from a studyof the mast cell degranulating effects of different trun-cated PACAP and VIP fragments on rat peritoneal anddural mast cells that phospholipase C-mediated mastcell degranulation is implicated in PACAP-inducedmigraine (30).
Possible mechanisms related to the elevatedPACAP-38-LI during migraine attacks
There are some anatomical and physiological factorswhich might be involved in the migraine-like head-ache-inducing effect of PACAP-38.
(a) PACAP-38 is a sensory (14), sympathetic (15)and parasympathetic (16) neuropeptide (31), which isreleased from the nerve endings (2,14,17,32) at thedural or other cranial compartments. It can modulatevessels and nerve fibres through its receptors leading toelevated intracellular cyclic adenosine monophosphate(cAMP) levels (33). There are several lines of evidencethat the increased cAMP levels cause sensitisation andactivation of the trigeminal neurons (34) and meningealnociceptors (35,36), therefore inducing delayed head-ache (37).
The headache-inducing action of PACAP-38 wasfirst described in 2009 by Schytz and colleagues, whodemonstrated that PACAP-38 has a simple headache-evoking effect in healthy volunteers but provokes severemigraine-like attacks in susceptible subjects. Adecreased mean blood flow velocity in the middle cere-bral artery and an increase in diameter of the superficialtemporal artery were observed 20 minutes following theinfusion (4). It is possible that the PACAP-38 has apositive-feedback, self-amplifying effect, which cancontribute to the maintenance and aggravation ofheadache.
Based on similar features and receptors of PACAPand VIP, as well as the supposed complex mechanismof migraine, it is likely that migraine cannot be relatedonly to PACAP-38. Its causative role is equivocal, butits involvement is unquestioned.
This peptide can contribute to the evolution ofmigraine attacks in cooperation with other regulatoryneuropeptides, molecules and enzymes, like VIP,CGRP, 5-HT, SP and nitric oxide (NO). When theheadache starts in response to specific triggers (vaso-dilation, plasma protein extravasation, mast celldegranulation, neuropeptide release, neurogenic
10Interictal Ictal Interictal Ictal
20
30
40P
lasm
a P
AC
AP
-38-
LI(f
mol
/ml)
∗
Group 1: Migrainerus withoutchronic pain conditions
Group 2: Migraineurs withchronic pain conditions
Figure 5. Plasma PACAP-38-LI (fmol/ml) in the interictal and
ictal periods of two subpopulations of 21 migraineurs charac-
terised on the basis of chronic pain conditions. Group 1: patients
without chronic low back pain (n¼ 15); group 2: patients with
simultaneous low back pain (n¼ 6). Each box represents the
median� SD, minimum and maximum values of the results. A
significant difference in PACAP-38-LI was observed between the
interictal and ictal phases in group 1 with Student’s paired t test,
p< 0.00005.
Tuka et al. 1091

inflammation), the presence and aggravation of painmay lead to the release of PACAP-38 from the nerveterminals as a self-triggering process. This can be anexplanation for the finding that the increase ofplasma PACAP-38-LI in the ictal phases of migraineurscompared to the interictal one is moderate butsignificant.
(b) Numerous studies have approached the effects ofPACAP-38 involved in headache from vasodilatingaspects. There are controversial results regarding therelaxing actions of PACAP-38 on meningeal or cerebralarteries (9,29,38,39). The vasodilating properties to asmall extent or indirectly are likely to contribute tothe development of headache, e. g. PACAP inducesvascular effects mediated via activation of perivascularnerves (40). The vascular effects cannot be excluded,but based on the literature it seems that these may beless relevant in migraine (9). PACAP-38 is more potentin sensitising trigeminal peripheral sensory fibres dir-ectly and also through mast cell degranulation(4,26,30). These mechanisms can be related to the phe-nomenon of throbbing headache, which can correlatewith the increased PACAP-38-LI during the ictal phaseof migraineurs found in our study. Subsequently, itwould be plausible that the activation of the second-order trigeminal neurons by PACAP-38 can result in adirect central sensitisation (4).
Complementary mechanisms behind reducedPACAP-38-LI during interictal phase of migrainepatients
The lower concentration of PACAP-38 observed in theinterictal period, and especially the correlation of theinterictal plasma PACAP-38-LI with disease duration,might be explained indirectly in terms of brain energydeficit (an impairment of the cerebral and striatedmuscle energy metabolism (41,42), elevated lactatelevels (43,44), abnormalities of mitochondrial compart-ments (45) and imbalanced Mg2þ concentration in theneurons (46,47)).
It may additionally be hypothesised that the lowinterictal peptide concentration might be associatedwith degenerative changes affecting the PACAP-releasing circuitries (cortical atrophy (48), iron depos-ition in the periaqueductal grey matter (49) andincreased levels of markers indicative of neuronal andglial damage (50)).
The possible role of the blood-brain barrier (BBB)
The ability of PACAP to pass through the intact BBB islimited; it needs specific transporters in specific brainareas (51,52). However, the enhanced BBB permeabil-ity in migraine may allow PACAP to penetrate into the
cerebrospinal fluid and exert its central effects. Fromthe opposite aspect, the PACAP released in the brain(6) can also penetrate through the BBB, and hence maybe detected in the plasma. There are assumptions thatthe integrity of the BBB is disrupted in migraineurs(53,54). A transient brain dysfunction, vasogenic cere-bral oedema and a damaged BBB can develop in lipo-polysaccharide-induced brain injury in rats in responseto the over-expression of matrix metalloproteinase 9(55). This enzyme can degrade the basal membrane,resulting in structural impairment of the BBB, alteredplasma levels of which have been observed in patientswith migraine (56–59).
Correlation of PACAP-38-LI with the menstruationcycle and other chronic pain conditions inmigraineurs
Migraine attacks are often closely related to femalehormonal changes (22). Hence, it is interesting thatPACAP-38-LI increases significantly during the ictalperiod only in those women whose headache is notrelated to the menstrual cycle. Although the influenceof the menstruation cycle in the PACAP-38-LI changescannot be excluded, there is evidence that the plasmaconcentration of PACAP-38 is relatively stable andindependent of gender, age, food intake or female hor-monal cycles in healthy subjects (60).
It is well established that PACAP-38 plays animportant role in a variety of other pain conditions(61–63). In opposition, there was no increasedPACAP-38-LI in the ictal phase in patients with otherchronic pain conditions. Moreover, we did not find anystatistical proof of the influence of the chronic pain andalterations of the PACAP-38 level.
Possible relations between the PACAP-38 andCGRP in migraineurs
More than 20 years ago, the release of CGRP wasdemonstrated in the extracerebral circulation ofhumans and cats in response to TS activation (64,65).CGRP-containing neurons have been detected in thehuman TRIG (14,66), and elevated plasma CGRPlevels have been described in migraine (67,68) andother types of primary headache (69). Moreover, anintravenous infusion of human alpha-CGRP causesmigraine headache (37,70), and there is a significantpositive correlation between plasma levels of CGRPand headache severity scores in NTG-induced migraineattacks (71). However, one controversial study ques-tions the importance of CGRP in migraine (72). Infact, CGRP-receptor antagonists are effective in thetreatment of migraine disorder (73), but their side-effect profile, with special emphasis on hepatotoxicity,
1092 Cephalalgia 33(13)

currently makes this drug development direction prob-lematic (74–76). Our results draw attention to a pos-sible influence between the PACAP and CGRP systemsin migraine pathogenesis.
Conclusions
It is concluded that PACAP-38 might be implicated inthe development of migraine headache. There is asso-ciation between migraine periods (ictal and interictal)
and alterations in PACAP-38 levels in the plasma. Thequantitative changes in this peptide in the plasma arerelated to the disease duration, the menstruation cycleand the presence of other pain-related disorders. Thecausative role of PACAP-38 in migraine headachedemands further studies. Our results indicate the needfor investigations of the role of plasma PACAP-38 as aputative biomarker of migraine, which might providenew perspectives as concerns the identification of a newtarget in the therapy of migraine.
Clinical implications
. The PACAP-38 has an important role in the mechanisms of migraine attack.
. A significantly lower PACAP-38 concentration in the interictal plasma of migraineurs as compared withhealthy controls is associated with the duration of migraine disease.
. The elevated plasma PACAP-38 level in the ictal phase relative to the attack-free period is attributable to thereleasing peptide from the PACAP-containing neurons, which can contribute to the development and/or theaggravation of migraine attacks.
Funding
This work was supported by Hungarian grants: DevelopingCompetitiveness of Universities in the South TransdanubianRegion (SROP-4.2.1.B-10/2/KONV-2010-0002, SROP-
4.2.2.A-11/1/KONV-2012-0024 and SROP-4.2.2/B-10/1-2010-0029), the Neuroscience Research Group of theHungarian Academy of Sciences and the University ofSzeged, SROP-4.2.1.B-09/1/KONV-2010-0005, K-2012-
TAMOP-4.2.2.A-11/1/KONV.-0443534/130, OTKA (PD104715) and FNUSA-ICRC (CZ.1.05/1.1.00/02.0123).B. Tuka is a Gedeon Richter Plc.-sponsored PhD candidate.
Dr Kincses was supported by the Bolyai ScholarshipProgramme of the Hungarian Academy of Sciences.
Conflict of interest
None declared.
References
1. Tajti J, Pardutz A, Vamos E, et al. Migraine is a neuronaldisease. J Neural Transm 2011; 118: 511–524.
2. Tajti J, Uddman R and Edvinsson L. Neuropeptide local-
ization in the ‘‘migraine generator’’ region of the humanbrainstem. Cephalalgia 2001; 21: 96–101.
3. Edvinsson L and Uddman R. Neurobiology in primary
headaches. Brain Res Brain Res Rev 2005; 48: 438–456.4. Schytz HW, Birk S, Wienecke T, et al. PACAP38 induces
migraine-like attacks in patients with migraine withoutaura. Brain 2009; 132: 16–25.
5. Schytz HW, Olesen J and Ashina M. The PACAPreceptor: A novel target for migraine treatment.Neurotherapeutics 2010; 7: 191–196.
6. Vaudry D, Falluel-Morel A, Bourgault S, et al. Pituitaryadenylate cyclase-activating polypeptide and its receptors:20 years after the discovery. Pharmacol Rev 2009; 61:
283–357.
7. Arimura A. PACAP: The road to discovery. Peptides
2007; 28: 1617–1619.8. Laburthe M and Couvineau A. Molecular pharmacology
and structure of VPAC receptors for VIP and PACAP.
Regul Pept 2002; 108: 165–173.9. Chan KY, Baun M, de Vries R, et al. Pharmacological
characterization of VIP and PACAP receptors in the
human meningeal and coronary artery. Cephalalgia2011; 31: 181–189.
10. Nemeth J, Reglodi D, Pozsgai G, et al. Effect of pituitaryadenylate cyclase activating polypeptide-38 on sensory
neuropeptide release and neurogenic inflammation inrats and mice. Neuroscience 2006; 143: 223–230.
11. Helyes Z, Pozsgai G, Borzsei R, et al. Inhibitory effect of
PACAP-38 on acute neurogenic and non-neurogenicinflammatory processes in the rat. Peptides 2007; 28:1847–1855.
12. Mabuchi T, Shintani N, Matsumura S, et al. Pituitaryadenylate cyclase-activating polypeptide is required forthe development of spinal sensitization and induction of
neuropathic pain. J Neurosci 2004; 24: 7283–7291.13. Zhang Y, Malmberg AB, Sjolund B, et al. The effect of
pituitary adenylate cyclase activating peptide (PACAP)on the nociceptive formalin test. Neurosci Lett 1996;
207: 187–190.14. Tajti J, Uddman R, Moller S, et al. Messenger molecules
and receptor mRNA in the human trigeminal ganglion.
J Auton Nerv Syst 1999; 76: 176–183.15. Uddman R, Tajti J, Moller S, et al. Neuronal messengers
and peptide receptors in the human sphenopalatine and
otic ganglia. Brain Res 1999; 826: 193–199.16. Csati A, Tajti J, Kuris A, et al. Distribution of vasoactive
intestinal peptide, pituitary adenylate cyclase-activatingpeptide, nitric oxide synthase, and their receptors in
human and rat sphenopalatine ganglion. Neuroscience2012; 202: 158–168.
Tuka et al. 1093

17. Uddman R, Tajti J, Hou M, et al. Neuropeptide expres-sion in the human trigeminal nucleus caudalis and in thecervical spinal cord C1 and C2. Cephalalgia 2002; 22:
112–116.18. Markovics A, Kormos V, Gaszner B, et al. Pituitary
adenylate cyclase-activating polypeptide plays a key rolein nitroglycerol-induced trigeminovascular activation in
mice. Neurobiol Dis 2012; 45: 633–644.19. Tuka B, Helyes Z, Markovics A, et al. Peripheral and
central alterations of pituitary adenylate cyclase activat-
ing polypeptide-like immunoreactivity in the rat inresponse to activation of the trigeminovascular system.Peptides 2012; 33: 307–316.
20. Lipton RB, Bigal ME, Steiner TJ, et al. Classification ofprimary headaches. Neurology 2004; 63: 427–435.
21. Aguggia M. Allodynia and migraine. Neurol Sci 2012;
33(Suppl 1): S9–S11.22. Karli N, Baykan B, Ertas M, et al. Impact of sex hormo-
nal changes on tension-type headache and migraine: Across-sectional population-based survey in 2,600
women. J Headache Pain 2012; 13: 557–565.23. Jakab B, Reglodi D, Jozsa R, et al. Distribution of
PACAP-38 in the central nervous system of various spe-
cies determined by a novel radioimmunoassay. J BiochemBiophys Methods 2004; 61: 189–198.
24. Baeres FM and Moller M. Origin of PACAP-immuno-
reactive nerve fibers innervating the subarachnoidalblood vessels of the rat brain. J Cereb Blood FlowMetab 2004; 24: 628–635.
25. Zagami AS, Edvinsson L, Hoskin KL, et al. Stimulation
of the superior sagittal sinus causes extracranial release ofPACAP. Cephalalgia 1995; 15: 109.
26. Schytz HW. Investigation of carbachol and PACAP38 in
a human model of migraine. Dan Med Bull 2010; 57:B4223.
27. Amin FM, Asghar MS, Guo S, et al. Headache and pro-
longed dilatation of the middle meningeal artery byPACAP38 in healthy volunteers. Cephalalgia 2012; 32:140–149.
28. Gupta S, Bhatt DK, Boni LJ, et al. Improvement of theclosed cranial window model in rats by intracarotid infu-sion of signalling molecules implicated in migraine.Cephalalgia 2010; 30: 27–36.
29. Syed AU, Koide M, Braas KM, et al. Pituitary adenylatecyclase-activating polypeptide (PACAP) potently dilatesmiddle meningeal arteries: Implications for migraine.
J Mol Neurosci 2012; 48: 574–583.30. Baun M, Pedersen MH, Olesen J, et al. Dural mast cell
degranulation is a putative mechanism for headache
induced by PACAP-38. Cephalalgia 2012; 32: 337–345.31. Knutsson M and Edvinsson L. Distribution of mRNA
for VIP and PACAP receptors in human cerebral arteriesand cranial ganglia. Neuroreport 2002; 13: 507–509.
32. Klimaschewski L, Hauser C and Heym C.PACAP immunoreactivity in the rat superior cervicalganglion in comparison to VIP. Neuroreport 1996; 7:
2797–2801.33. Dickson L, Aramori I, McCulloch J, et al. A systematic
comparison of intracellular cyclic AMP and calcium sig-
nalling highlights complexities in human VPAC/PAC
receptor pharmacology. Neuropharmacology 2006; 51:1086–1098.
34. Ingram SL and Williams JT. Modulation of the hyperpo-
larization-activated current (Ih) by cyclic nucleotides inguinea-pig primary afferent neurons. J Physiol 1996;492(Pt 1): 97–106.
35. Burstein R, Yamamura H, Malick A, et al. Chemical
stimulation of the intracranial dura induces enhancedresponses to facial stimulation in brain stem trigeminalneurons. J Neurophysiol 1998; 79: 964–982.
36. Strassman AM, Raymond SA and Burstein R. Sensitiza-tion of meningeal sensory neurons and the origin of head-aches. Nature 1996; 384: 560–564.
37. Lassen LH, Haderslev PA, Jacobsen VB, et al. CGRPmay play a causative role in migraine. Cephalalgia 2002;22: 54–61.
38. Jansen-Olesen I, Gulbenkian S, Engel U, et al. Peptider-gic and non-peptidergic innervation and vasomotorresponses of human lenticulostriate and posterior cere-bral arteries. Peptides 2004; 25: 2105–2114.
39. Goadsby PJ. The vascular theory of migraine – a greatstory wrecked by the facts. Brain 2009; 132: 6–7.
40. Uddman R, Goadsby PJ, Jansen I, et al. PACAP, a VIP-
like peptide: Immunohistochemical localization andeffect upon cat pial arteries and cerebral blood flow.J Cereb Blood Flow Metab 1993; 13: 291–297.
41. Barbiroli B, Montagna P, Cortelli P, et al. Abnormalbrain and muscle energy metabolism shown by 31P mag-netic resonance spectroscopy in patients affected bymigraine with aura. Neurology 1992; 42: 1209–1214.
42. Montagna P, Cortelli P and Barbiroli B. Magnetic reson-ance spectroscopy studies in migraine. Cephalalgia 1994;14: 184–193.
43. Watanabe H, Kuwabara T, Ohkubo M, et al. Elevationof cerebral lactate detected by localized 1H-magnetic res-onance spectroscopy in migraine during the interictal
period. Neurology 1996; 47: 1093–1095.44. Kaufmann P, Shungu DC, Sano MC, et al. Cerebral
lactic acidosis correlates with neurological impairment
in MELAS. Neurology 2004; 62: 1297–1302.45. Sangiorgi S, Mochi M, Riva R, et al. Abnormal platelet
mitochondrial function in patients affected by migrainewith and without aura. Cephalalgia 1994; 14: 21–23.
46. Welch KM and Ramadan NM. Mitochondria, magne-sium and migraine. J Neurol Sci 1995; 134: 9–14.
47. Peikert A, Wilimzig C and Kohne-Volland R. Prophy-
laxis of migraine with oral magnesium: Results from aprospective, multi-center, placebo-controlled anddouble-blind randomized study. Cephalalgia 1996; 16:
257–263.48. May A. New insights into headache: An update on func-
tional and structural imaging findings. Nat Rev Neurol2009; 5: 199–209.
49. Welch KM, Nagesh V, Aurora SK, et al. Periaqueductalgray matter dysfunction in migraine: Cause or the burdenof illness? Headache 2001; 41: 629–637.
50. Yilmaz N, Karaali K, Ozdem S, et al. Elevated S100Band neuron specific enolase levels in patients withmigraine-without aura: Evidence for neurodegeneration?
Cell Mol Neurobiol 2011; 31: 579–585.
1094 Cephalalgia 33(13)

51. Nonaka N, Banks WA, Mizushima H, et al. Regionaldifferences in PACAP transport across the blood-brainbarrier in mice: A possible influence of strain, amyloid
beta protein, and age. Peptides 2002; 23: 2197–2202.52. Nonaka N, Farr SA, Nakamachi T, et al. Intranasal
administration of PACAP: Uptake by brain and regionalbrain targeting with cyclodextrins. Peptides 2012; 36:
168–175.53. Gao HM, Li L, Zhang KL, et al. Impact of migraine
attacks on the blood-brain barrier. Chin Med J (Engl)
2010; 123: 2559–2561.54. Dreier JP, Jurkat-Rott K, Petzold GC, et al. Opening of
the blood-brain barrier preceding cortical edema in a
severe attack of FHM type II. Neurology 2005; 64:2145–2147.
55. Mun-Bryce S and Rosenberg GA. Gelatinase B modu-
lates selective opening of the blood-brain barrier duringinflammation. Am J Physiol 1998; 274: R1203–R1211.
56. Ashina M, Tvedskov JF, Lipka K, et al. Matrix metallo-proteinases during and outside of migraine attacks with-
out aura. Cephalalgia 2010; 30: 303–310.57. Leira R, Sobrino T, Rodrıguez-Yanez M, et al. Mmp-9
immunoreactivity in acute migraine. Headache 2007; 47:
698–702.58. Gursoy-Ozdemir Y, Qiu J, Matsuoka N, et al. Cortical
spreading depression activates and upregulates MMP-9.
J Clin Invest 2004; 113: 1447–1455.59. Martins-Oliveira A, Goncalves FM, Speciali JG, et al.
Specific matrix metalloproteinase 9 (MMP-9) haplotypeaffect the circulating MMP-9 levels in women with
migraine. J Neuroimmunol 2012; 252: 89–94.60. Borzsei R, Mark L, Tamas A, et al. Presence of pituitary
adenylate cyclase activating polypeptide-38 in human
plasma and milk. Eur J Endocrinol 2009; 160: 561–565.61. Sandor K, Bolcskei K, McDougall JJ, et al. Divergent
peripheral effects of pituitary adenylate cyclase-activating
polypeptide-38 on nociception in rats and mice. Pain2009; 141: 143–150.
62. Schytz HW, Holst H, Arendt-Nielsen L, et al.
Cutaneous nociception and neurogenic inflammationevoked by PACAP38 and VIP. J Headache Pain 2010;11: 309–316.
63. Dickinson T and Fleetwood-Walker SM. Neuropeptides
and nociception: Recent advances and therapeutic impli-cations. Trends Pharmacol Sci 1998; 19: 346–348.
64. Goadsby PJ, Edvinsson L and Ekman R. Release ofvasoactive peptides in the extracerebral circulation ofhumans and the cat during activation of the trigemino-
vascular system. Ann Neurol 1988; 23: 193–196.65. Zagami AS, Goadsby PJ and Edvinsson L. Stimulation
of the superior sagittal sinus in the cat causes release ofvasoactive peptides. Neuropeptides 1990; 16: 69–75.
66. Eftekhari S, Salvatore CA, Calamari A, et al. Differentialdistribution of calcitonin gene-related peptide and itsreceptor components in the human trigeminal ganglion.
Neuroscience 2010; 169: 683–696.67. Goadsby PJ, Edvinsson L and Ekman R. Vasoactive pep-
tide release in the extracerebral circulation of humans
during migraine headache. Ann Neurol 1990; 28: 183–187.68. Ashina M, Bendtsen L, Jensen R, et al. Evidence for
increased plasma levels of calcitonin gene-related peptide
in migraine outside of attacks. Pain 2000; 86: 133–138.69. Goadsby PJ. Calcitonin gene-related peptide antagonists
as treatments of migraine and other primary headaches.Drugs 2005; 65: 2557–2567.
70. Hansen JM, Hauge AW, Olesen J, et al. Calcitonin gene-related peptide triggers migraine-like attacks in patientswith migraine with aura. Cephalalgia 2010; 30:
1179–1186.71. Juhasz G, Zsombok T, Modos EA, et al. NO-induced
migraine attack: Strong increase in plasma calcitonin
gene-related peptide (CGRP) concentration and negativecorrelation with platelet serotonin release. Pain 2003; 106:461–470.
72. Hansen JM, Thomsen LL, Olesen J, et al. Calcitonin
gene-related peptide does not cause migraine attacks inpatients with familial hemiplegic migraine. Headache2011; 51: 544–553.
73. Edvinsson L and Linde M. New drugs in migraine treat-ment and prophylaxis: Telcagepant and topiramate.Lancet 2010; 376: 645–655.
74. Ho TW, Edvinsson L and Goadsby PJ. CGRP and itsreceptors provide new insights into migraine pathophysi-ology. Nat Rev Neurol 2010; 6: 573–582.
75. Edvinsson L and Petersen KA. CGRP-receptor antagon-ism in migraine treatment. CNS Neurol Disord DrugTargets 2007; 6: 240–246.
76. Tfelt-Hansen P and Olesen J. Possible site of action of
CGRP antagonists in migraine. Cephalalgia 2011; 31:748–750.
Tuka et al. 1095




















