
Dynamic Granger-Geweke causality modeling with application to interictal spike propagation
Upload
independentCategory
view
1download
0
www.elsevier.com/locate/clinph
Clinical Neurophysiology 117 (2006) 2615–2626
Propagation of interictal discharges in temporal lobeepilepsy: Correlation of spatiotemporal mapping with
intracranial foramen ovale electrode recordings
Dominik Zumsteg a,*, Alon Friedman b, Heinz Gregor Wieser c, Richard A. Wennberg a
a Krembil Neuroscience Centre, Toronto Western Hospital, University of Toronto, Toronto, Canadab Departments of Physiology and Neurosurgery, Zlotowski Center for Neuroscience, Ben-Gurion University, Beer-Sheva, Israel
c Department of Neurology, University Hospital Zurich, Zurich, Switzerland
Accepted 29 July 2006Available online 6 October 2006
Abstract
Objective: We have investigated intracerebral propagation of interictal epileptiform discharges (IED) in patients with mesial temporallobe epilepsy (MTLE) by using spatiotemporal source maps based on statistical nonparametric mapping (SNPM) of low resolution elec-tromagnetic tomography (LORETA) values.Methods: We analyzed 30 patterns of IED recorded simultaneously with scalp and intracranial foramen ovale (FO) electrodes in 15 con-secutive patients with intractable MTLE. The scalp EEG signals were averaged time-locked to the peak activity in bilateral 10-contactFO electrode recordings. SNPM was applied to LORETA values and spatiotemporal source maps were created by allocating the t-valuesover time to their corresponding Brodmann areas. Propagation was defined as secondary statistically significant involvement of distinctcortical areas separated by >15 ms. The results were correlated with intracranial data obtained from FO electrode recordings and withscalp EEG recordings. All patients underwent subsequent amygdalo-hippocampectomy and outcome was assessed one year after surgery.Results: We found mesial to lateral propagation in 6/30 IED patterns (20%, four patients), lateral to mesial propagation in 4/30 IEDpatterns (13.3%, four patients) and simultaneous (within 15 ms) activation of mesial and lateral temporal areas in 6/30 IED patterns(20%, five patients). Propagation generally occurred within 30 ms and was always limited to ipsilateral cortical regions. Nine/30 IEDpatterns (30%) showed restricted activation of mesial temporal structures and no significant solutions were found in 5/30 IED patterns(16.7%). There was no clear association between the number or characteristics of IED patterns and the postsurgical outcome.Conclusions: Spatiotemporal mapping of SNPM LORETA accurately describes mesial to lateral temporal propagation of IED, and viceversa, which commonly occur in patients with MTLE.Significance: Intracerebral propagation must be considered when using non-invasive source algorithms in patients with MTLE. Spatio-temporal mapping might be useful for visualizing this propagation.� 2006 International Federation of Clinical Neurophysiology. Published by Elsevier Ireland Ltd. All rights reserved.
Keywords: Propagation; Temporal lobe epilepsy; Foramen ovale electrodes; Source localization; LORETA; SNPM
1. Introduction
Interictal epileptiform discharges (IED) are a very con-sistent finding in patients with temporal lobe epilepsy(TLE), occurring in 96–98% of patients (Marsan and Zivin,
1388-2457/$32.00 � 2006 International Federation of Clinical Neurophysiolo
doi:10.1016/j.clinph.2006.07.319
* Corresponding author. Tel.: +1 416 603 5402; fax: +1 416 603 5768.E-mail address: [email protected] (D. Zumsteg).
1970; Williamson et al., 1993). Although the recording ofseizures remains the cornerstone of presurgical evaluation,IED are considered to be important and independent indi-cators of the epileptogenic zone (So et al., 1989). The distri-bution of IED in patients with mesial temporal lobeepilepsy (MTLE) due to hippocampal sclerosis has beenreported to be smaller and more restricted to thehippocampal region compared to TLE due to otheretiologies (Hamer et al., 1999). Nonetheless, IED in MTLE
gy. Published by Elsevier Ireland Ltd. All rights reserved.

2616 D. Zumsteg et al. / Clinical Neurophysiology 117 (2006) 2615–2626
commonly involve basal or basolateral temporal corticalareas or may even occur distant to mesial temporal struc-tures independently from any involvement of the mesiallimbic structures (Marks et al., 1992). Bilateral IED occurin about 34–77%, being preponderant over the side of sei-zure onset in about 60–77% of patients (Marsan and Zivin,1970; Hughes, 1985; So et al., 1989; Hirsch et al., 1991;Williamson et al., 1993). Intracranial and surface EEGstudies as well as combined intracranial/surface EEG stud-ies have shown that mesial IED may propagate with laten-cies on the order of 10–50 ms, whether within thehippocampus in the anterior-posterior direction (Buserand Bancaud, 1983), within the temporal lobe (Alarconet al., 1997; Emerson et al., 1995; Merlet et al., 1996; Suth-erling and Barth, 1989) or even to more distant corticalareas (Alarcon et al., 1994; Emerson et al., 1995). Propaga-tion may also occur in the opposite direction, i.e., from lat-eral temporal cortical areas to mesial temporal structures(Sutherling and Barth, 1989; Stefan et al., 1991; Markset al., 1992).
The surface voltage distribution at a certain time point isdirectly related to the intracranial voltage fields generatedby the cerebral sources of the IED at that time point.IED arising from restricted and deep cortical areas suchas the hippocampus may remain undetected on scalpEEG recordings (Niedermeyer and Rocca, 1972; Alarconet al., 1994; Munari et al., 1994; Merlet et al., 1996; Fer-nandez Torre et al., 1999; Wennberg and Lozano, 2003;Gavaret et al., 2004; Nayak et al., 2004; Zumsteg et al.,2006a). Mesial temporal IED that also involve basal tobasolateral temporal cortical areas typically show a nega-tive field maximum over the ipsilateral frontotemporalscalp and a corresponding positive field maximum overthe centroparietal scalp, whereas lateral temporal discharg-es commonly show negative electrical fields extending toipsilateral mid to posterior temporal regions and a corre-sponding positive electrical field maximum over the contra-lateral temporal region (Ebersole and Wade, 1990).Furthermore, the scalp voltage distribution may show con-siderable changes during the course of the IED, clearlyindicating intracerebral propagation or extension of theepileptiform activity (Baumgartner et al., 1995; Emersonet al., 1995; Merlet et al., 1996; Merlet and Gotman,1999). This has obvious bearing on the validity of non-in-vasive source algorithms. It has been repeatedly shown thatpropagated sources may dominate at the peak of the IED,a finding that has led to the conclusion that the ascendingphase of an IED would be the most appropriate for sourcelocalization (Scherg et al., 1999; Huppertz et al., 2001;Lantz et al., 2003a). The use of distributed source modelssuch as low resolution electromagnetic tomography(LORETA) might be most suitable to study such complexactivity, since such models take into account that cerebralsources are extended areas of active cortex rather thanpoint-like dipoles. We have previously shown that the useof statistical nonparametric mapping (SNPM) of LORE-TA values applied during the ascending phase of IED
allowed for reasonable localization of mesial temporalIED, as validated by simultaneous intracranial FO elec-trode recordings (Zumsteg et al., 2005, 2006b).
The purpose of the present investigation was to evaluatethe usefulness of SNPM LORETA for the analysis of IEDwith fast changing surface voltage distributions. The use ofspatiotemporal source maps would make it possible todetermine the cortical sources underlying the fast changesof surface voltage distributions during the entire courseof the IED. Although outcome is not the principle aim ofthis study, we also assessed the relationship of SNPMLORETA source solutions to the postsurgical outcome.
2. Patients and methods
2.1. Patients
We retrospectively studied 30 IED patterns in 15 consec-utive patients (mean age 34.7 years, six women) withMTLE who had presurgical assessment with simultaneousintracranial FO electrode and scalp EEG recordings. Allpatients have subsequently undergone selective or tailoredamygdalo-hippocampectomy (AHE, Wieser et al., 2003).Postsurgical outcome was assessed using the Engelclassification one year after resective surgery (Engel et al.,1993). Table 1 shows the clinical and electrophysiologicalcharacteristics of the patients.
2.2. Scalp EEG and FO electrode recordings
Twenty-three scalp electrodes were placed according tothe 10–20 System, supplemented with mastoid (M1/2)and true-temporal (T1/2) electrodes. Two 10-contact DIXIFO electrodes (each contact is 2 mm long and 0.93 mm indiameter, inter-contact distance is 2 mm) were implantedbilaterally under radiological control according to a stan-dard protocol (Wieser, 1999). The deepest contacts of theFO electrodes (contacts 1) lay bilaterally in the caudalend of the ambient cistern in proximity to posterior mesialtemporal structures (i.e., Brodmann areas (BAs) 27, 35, 26,29, and 30) and next to the lateral geniculate bodies. Theoutermost contacts of the electrodes (contacts 10) werepositioned just intracranially of the FO, adjacent to mesio-basal temporal structures (i.e., BAs 20, 36, and 28) (Wieserand Schwarz, 2001). Fig. 1 shows the position of a properlyplaced FO electrode in relation to anatomical structuresand scalp electrodes. The adequacy of the position of theFO electrodes was confirmed with post-insertion radiogra-phy. Both scalp EEG and FO electrode signals were contin-uously recorded using a Telefactor Beekeeper system with asampling rate of 200 Hz and a bandpass filter of 0.3–100 Hz. All electrodes were referenced to an FCz electrode.
2.3. Data preparation, LORETA and SNPM
Thirty IED patterns (1–4 per patient) were identified byvisual inspection of the referential FO electrode traces
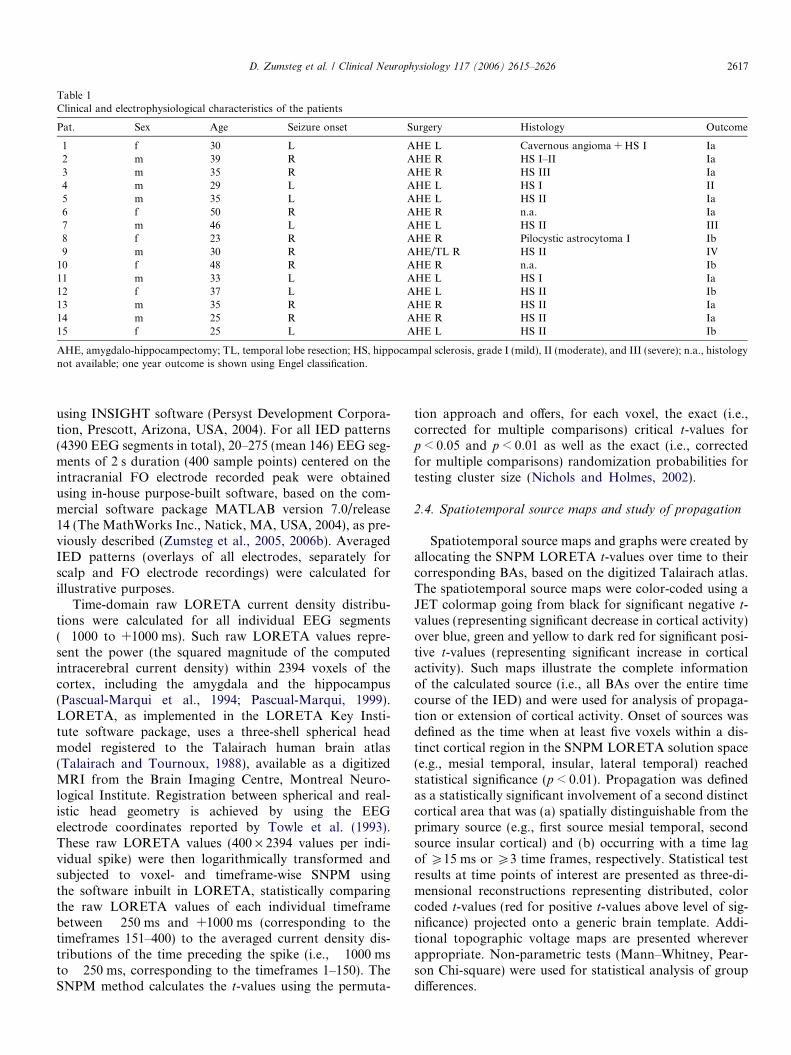
Table 1Clinical and electrophysiological characteristics of the patients
Pat. Sex Age Seizure onset Surgery Histology Outcome
1 f 30 L AHE L Cavernous angioma + HS I Ia2 m 39 R AHE R HS I–II Ia3 m 35 R AHE R HS III Ia4 m 29 L AHE L HS I II5 m 35 L AHE L HS II Ia6 f 50 R AHE R n.a. Ia7 m 46 L AHE L HS II III8 f 23 R AHE R Pilocystic astrocytoma I Ib9 m 30 R AHE/TL R HS II IV
10 f 48 R AHE R n.a. Ib11 m 33 L AHE L HS I Ia12 f 37 L AHE L HS II Ib13 m 35 R AHE R HS II Ia14 m 25 R AHE R HS II Ia15 f 25 L AHE L HS II Ib
AHE, amygdalo-hippocampectomy; TL, temporal lobe resection; HS, hippocampal sclerosis, grade I (mild), II (moderate), and III (severe); n.a., histologynot available; one year outcome is shown using Engel classification.
D. Zumsteg et al. / Clinical Neurophysiology 117 (2006) 2615–2626 2617
using INSIGHT software (Persyst Development Corpora-tion, Prescott, Arizona, USA, 2004). For all IED patterns(4390 EEG segments in total), 20–275 (mean 146) EEG seg-ments of 2 s duration (400 sample points) centered on theintracranial FO electrode recorded peak were obtainedusing in-house purpose-built software, based on the com-mercial software package MATLAB version 7.0/release14 (The MathWorks Inc., Natick, MA, USA, 2004), as pre-viously described (Zumsteg et al., 2005, 2006b). AveragedIED patterns (overlays of all electrodes, separately forscalp and FO electrode recordings) were calculated forillustrative purposes.
Time-domain raw LORETA current density distribu-tions were calculated for all individual EEG segments(�1000 to +1000 ms). Such raw LORETA values repre-sent the power (the squared magnitude of the computedintracerebral current density) within 2394 voxels of thecortex, including the amygdala and the hippocampus(Pascual-Marqui et al., 1994; Pascual-Marqui, 1999).LORETA, as implemented in the LORETA Key Insti-tute software package, uses a three-shell spherical headmodel registered to the Talairach human brain atlas(Talairach and Tournoux, 1988), available as a digitizedMRI from the Brain Imaging Centre, Montreal Neuro-logical Institute. Registration between spherical and real-istic head geometry is achieved by using the EEGelectrode coordinates reported by Towle et al. (1993).These raw LORETA values (400 · 2394 values per indi-vidual spike) were then logarithmically transformed andsubjected to voxel- and timeframe-wise SNPM usingthe software inbuilt in LORETA, statistically comparingthe raw LORETA values of each individual timeframebetween �250 ms and +1000 ms (corresponding to thetimeframes 151–400) to the averaged current density dis-tributions of the time preceding the spike (i.e., �1000 msto �250 ms, corresponding to the timeframes 1–150). TheSNPM method calculates the t-values using the permuta-
tion approach and offers, for each voxel, the exact (i.e.,corrected for multiple comparisons) critical t-values forp < 0.05 and p < 0.01 as well as the exact (i.e., correctedfor multiple comparisons) randomization probabilities fortesting cluster size (Nichols and Holmes, 2002).
2.4. Spatiotemporal source maps and study of propagation
Spatiotemporal source maps and graphs were created byallocating the SNPM LORETA t-values over time to theircorresponding BAs, based on the digitized Talairach atlas.The spatiotemporal source maps were color-coded using aJET colormap going from black for significant negative t-values (representing significant decrease in cortical activity)over blue, green and yellow to dark red for significant posi-tive t-values (representing significant increase in corticalactivity). Such maps illustrate the complete informationof the calculated source (i.e., all BAs over the entire timecourse of the IED) and were used for analysis of propaga-tion or extension of cortical activity. Onset of sources wasdefined as the time when at least five voxels within a dis-tinct cortical region in the SNPM LORETA solution space(e.g., mesial temporal, insular, lateral temporal) reachedstatistical significance (p < 0.01). Propagation was definedas a statistically significant involvement of a second distinctcortical area that was (a) spatially distinguishable from theprimary source (e.g., first source mesial temporal, secondsource insular cortical) and (b) occurring with a time lagof P15 ms or P3 time frames, respectively. Statistical testresults at time points of interest are presented as three-di-mensional reconstructions representing distributed, colorcoded t-values (red for positive t-values above level of sig-nificance) projected onto a generic brain template. Addi-tional topographic voltage maps are presented whereverappropriate. Non-parametric tests (Mann–Whitney, Pear-son Chi-square) were used for statistical analysis of groupdifferences.

Fig. 1. Lateral (top) and mesial (bottom) view of the left hemisphereshowing the location of a left FO electrode and scalp electrodes in relationto anatomical structures. Note the intimate relation of the most posteriorFO electrode contact 1 with the posterior aspects of the hippocampus andthe parahippocampal gyrus as well as the proximity of the outermostcontacts 5–10 with the uncus.
2618 D. Zumsteg et al. / Clinical Neurophysiology 117 (2006) 2615–2626
3. Results
3.1. Source solutions and intracerebral propagation
Nineteen of 30 IED patterns (63.3%) showed a restrict-ed intracranial electrical field in FO electrode recordingsand were accompanied by a simultaneous low-amplitudescalp EEG discharge showing a scalp voltage distributionthat is considered to be typical for mesial temporal dis-charges, i.e., a negative field maximum over the ipsilateralsubtemporal scalp and a corresponding positive field max-imum over the centroparietal scalp. As previously report-ed, SNPM LORETA correctly localized the sources in 14of these 19 IED patterns (73.7%), whereas no significantsolution was found for the remaining five IED patterns(Zumsteg et al., 2005, 2006b). These 19 IED patterns
are henceforth referred to as ‘‘mesial’’ IED patterns.The onset times of these single sources with respect tothe IED peak activity in FO electrode recordings aregiven in Table 2.
Eleven of 30 IED patterns (36.7%) in the same collec-tive of patients were considered to be of basolateral orlateral neocortical temporal origin, based on their electro-physiological characteristics (henceforth termed ‘‘lateral’’IED patterns). They consistently showed high amplitudescalp EEG discharges with a scalp topography that hasbeen commonly associated with temporal IED involvingbasal or basolateral temporal cortical areas, i.e., a nega-tive field maximum over the ipsilateral frontotemporalor lateral temporal scalp and a corresponding positivefield maximum over the contralateral temporal scalp.With respect to FO electrode recordings, nine of the ‘‘lat-eral’’ IED patterns (81.8%) revealed a wide anterior orposterior electrical field, with either no phase reversal ora broad positive phase reversal within the scope of theFO electrode, thus suggesting a wide intracranial fieldwith involvement of basolateral or lateral temporal corti-cal areas. Two of the ‘‘lateral’’ IED patterns (18.2%), incontrast, showed a negative phase reversal within thescope of the FO electrode, suggesting a restricted intracra-nial electrical field comparable to ‘‘mesial’’ IED patterns.These IED patterns were nevertheless considered to reflectdischarges with maximal involvement of basolateral orlateral temporal cortical structures because of their highamplitude (>200 lV) and typical lateral voltage distribu-tion in scalp EEG recordings (it is of note that they werealso considered as such during presurgical clinical assess-ment of the patients).
We found mesial to lateral propagation in 6/30 (20%)IED patterns (Fig. 2A). Two of the 19 ‘‘mesial’’ IED pat-terns showed clear propagation from mesial to ipsilateralbasolateral or lateral temporal cortical areas within about30 ms (Fig. 3 depicts a representative example), and 4/11‘‘lateral’’ IED patterns (all showing a wide posterior fieldin FO electrode recordings) revealed a significant increaseof ipsilateral mesial temporal activity preceding the lateralactivation by about 15–25 ms (raising the possibility thatthese four ‘‘lateral’’ IED patterns actually reflect ‘‘mesial’’IED patterns with fast propagation to basolateral or lateraltemporal cortical areas). The onset times of the primarymesial and secondary propagated lateral temporal sourcesare given in Table 2.
Lateral to mesial propagation within about 15–25 mswas found in 4/30 IED patterns (13.3%; all classified as‘‘lateral’’ IED patterns, Fig. 2B and Table 2). They consis-tently revealed a wide anterior intracranial field in FO elec-trode recordings, thus suggesting a primary source inbasolateral to lateral temporal cortical areas with subse-quent propagation into ipsilateral mesial temporal struc-tures. A representative example is shown in Fig. 4.
Simultaneous activation of mesial and lateral temporalareas was found in 6/30 IED patterns (20%). Three of thosewere classified as ‘‘mesial’’ IED patterns (i.e., they were
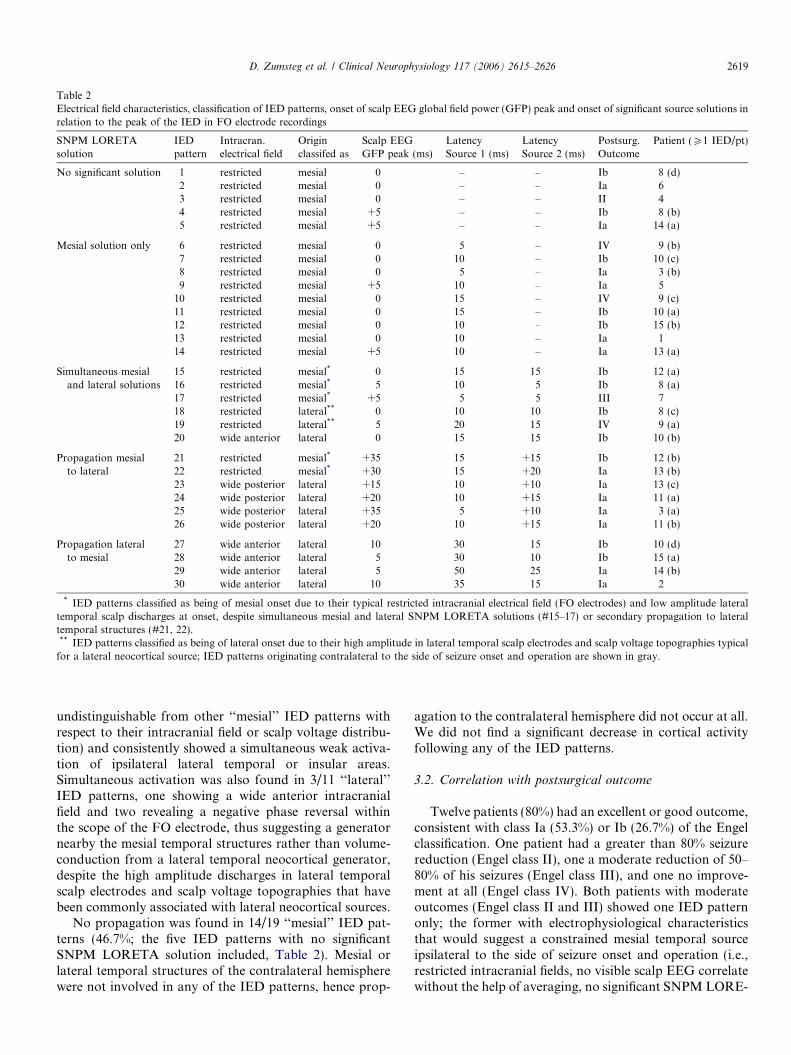
Table 2Electrical field characteristics, classification of IED patterns, onset of scalp EEG global field power (GFP) peak and onset of significant source solutions inrelation to the peak of the IED in FO electrode recordings
SNPM LORETAsolution
IEDpattern
Intracran.electrical field
Originclassifed as
Scalp EEGGFP peak (ms)
LatencySource 1 (ms)
LatencySource 2 (ms)
Postsurg.Outcome
Patient (P1 IED/pt)
No significant solution 1 restricted mesial 0 – – Ib 8 (d)2 restricted mesial 0 – – Ia 63 restricted mesial 0 – – II 44 restricted mesial +5 – – Ib 8 (b)5 restricted mesial +5 – – Ia 14 (a)
Mesial solution only 6 restricted mesial 0 � 5 – IV 9 (b)7 restricted mesial 0 �10 – Ib 10 (c)8 restricted mesial 0 �5 – Ia 3 (b)9 restricted mesial +5 �10 – Ia 5
10 restricted mesial 0 �15 – IV 9 (c)11 restricted mesial 0 �15 – Ib 10 (a)12 restricted mesial 0 �10 – Ib 15 (b)13 restricted mesial 0 �10 – Ia 114 restricted mesial +5 �10 – Ia 13 (a)
Simultaneous mesialand lateral solutions
15 restricted mesial* 0 �15 �15 Ib 12 (a)16 restricted mesial* �5 �10 �5 Ib 8 (a)17 restricted mesial* +5 �5 �5 III 718 restricted lateral** 0 �10 �10 Ib 8 (c)19 restricted lateral** �5 �20 �15 IV 9 (a)20 wide anterior lateral 0 �15 �15 Ib 10 (b)
Propagation mesialto lateral
21 restricted mesial* +35 �15 +15 Ib 12 (b)22 restricted mesial* +30 �15 +20 Ia 13 (b)23 wide posterior lateral +15 �10 +10 Ia 13 (c)24 wide posterior lateral +20 �10 +15 Ia 11 (a)25 wide posterior lateral +35 �5 +10 Ia 3 (a)26 wide posterior lateral +20 �10 +15 Ia 11 (b)
Propagation lateralto mesial
27 wide anterior lateral �10 �30 �15 Ib 10 (d)28 wide anterior lateral �5 �30 �10 Ib 15 (a)29 wide anterior lateral �5 �50 �25 Ia 14 (b)30 wide anterior lateral �10 �35 �15 Ia 2
* IED patterns classified as being of mesial onset due to their typical restricted intracranial electrical field (FO electrodes) and low amplitude lateraltemporal scalp discharges at onset, despite simultaneous mesial and lateral SNPM LORETA solutions (#15–17) or secondary propagation to lateraltemporal structures (#21, 22).** IED patterns classified as being of lateral onset due to their high amplitude in lateral temporal scalp electrodes and scalp voltage topographies typical
for a lateral neocortical source; IED patterns originating contralateral to the side of seizure onset and operation are shown in gray.
D. Zumsteg et al. / Clinical Neurophysiology 117 (2006) 2615–2626 2619
undistinguishable from other ‘‘mesial’’ IED patterns withrespect to their intracranial field or scalp voltage distribu-tion) and consistently showed a simultaneous weak activa-tion of ipsilateral lateral temporal or insular areas.Simultaneous activation was also found in 3/11 ‘‘lateral’’IED patterns, one showing a wide anterior intracranialfield and two revealing a negative phase reversal withinthe scope of the FO electrode, thus suggesting a generatornearby the mesial temporal structures rather than volume-conduction from a lateral temporal neocortical generator,despite the high amplitude discharges in lateral temporalscalp electrodes and scalp voltage topographies that havebeen commonly associated with lateral neocortical sources.
No propagation was found in 14/19 ‘‘mesial’’ IED pat-terns (46.7%; the five IED patterns with no significantSNPM LORETA solution included, Table 2). Mesial orlateral temporal structures of the contralateral hemispherewere not involved in any of the IED patterns, hence prop-
agation to the contralateral hemisphere did not occur at all.We did not find a significant decrease in cortical activityfollowing any of the IED patterns.
3.2. Correlation with postsurgical outcome
Twelve patients (80%) had an excellent or good outcome,consistent with class Ia (53.3%) or Ib (26.7%) of the Engelclassification. One patient had a greater than 80% seizurereduction (Engel class II), one a moderate reduction of 50–80% of his seizures (Engel class III), and one no improve-ment at all (Engel class IV). Both patients with moderateoutcomes (Engel class II and III) showed one IED patternonly; the former with electrophysiological characteristicsthat would suggest a constrained mesial temporal sourceipsilateral to the side of seizure onset and operation (i.e.,restricted intracranial fields, no visible scalp EEG correlatewithout the help of averaging, no significant SNPM LORE-

Fig. 2. Source solutions and corresponding topographical voltage maps of the six IED patterns with mesial to lateral propagation (A, patterns 21–26) andthe four IED patterns with lateral to mesial propagation (B, patterns 27–30). For each IED pattern, the left panels refer to the source localization of theprimary source, the right panels to the source localization of the secondary propagated source. The numbers below the voltage maps denote to the timepoints with reference to the IED peak in FO electrode recordings. The source solutions of the nine IED patterns with exclusive mesial sources, and thesix IED patterns with simultaneous mesial and lateral source solutions have been previously reported (Zumsteg et al., 2006a,b) and are not shown (seeTable 2).
2620 D. Zumsteg et al. / Clinical Neurophysiology 117 (2006) 2615–2626
TA solution), the latter with a restricted intracranial field butwith simultaneous mesial and lateral temporal SNPMLORETA solutions. The patient with bad outcome (Engelclass IV) showed three IED patterns, all of which revealeda restricted intracranial field, two originating from mesialtemporal structures contralateral to the side of seizure onsetand operation (Table 2).
The eight patients with excellent outcome (Engel class Ia)had significantly fewer IED patterns (1.6 ± 0.7) when com-pared to patients with good outcome (Engel class Ib;3 ± 1.2; Mann–Whitney, p < 0.05). The excellent outcomegroup also had a higher rate of restricted IED patterns only(50% vs 0%) and a higher percentage of patients with mesialonset IED patterns (37.5% vs 25%) or no contralateral IEDpatterns (37.5% vs 25%), but none of these differencesreached statistical significance (Pearson Chi-Square; p-val-ues 0.083, 0.665, 0.665). No significant differences werefound with respect to propagation of IED patterns, occur-ring in 75% of patients with Engel class Ia and in all patientswith Engel class Ib outcome (Pearson Chi-Square; p-values:mesial to lateral 0.665; lateral to mesial 0.386; simultaneous0.14; all 0.157).
4. Discussion
Propagation or extension of epileptiform activity fromthe epileptogenic region to distant areas is well known tooccur in patients with MTLE. It is mediated by fiber tractsconnecting the mesial temporal structures with ipsilaterallateral temporal and even more distant cortical areas suchas the orbitofrontal region, via the subicular cortex, theenthorhinal cortex and perirhinal/parahippocampal corti-ces (Gloor, 1997). The electrophysiology of the efferentconnections of the amygdala and the hippocampal systemto other cortical areas has been extensively studied inpatients with intractable epilepsy during stereotaxic explo-rations of mesial temporal structures (Feindel and Penfield,1954; Wieser, 1983; Pandya and Yeterian, 1987; Wilsonet al., 1990; Engel, 1993). Previous studies have also report-ed on the usefulness of non-invasive source models for theinvestigation of propagation of interictal epileptiformactivity. Studies using spatiotemporal dipole modeling(either moving or multiple dipole algorithms) have demon-strated sequential activation of different dipole sources,typically occurring at an early phase of the IED, usually
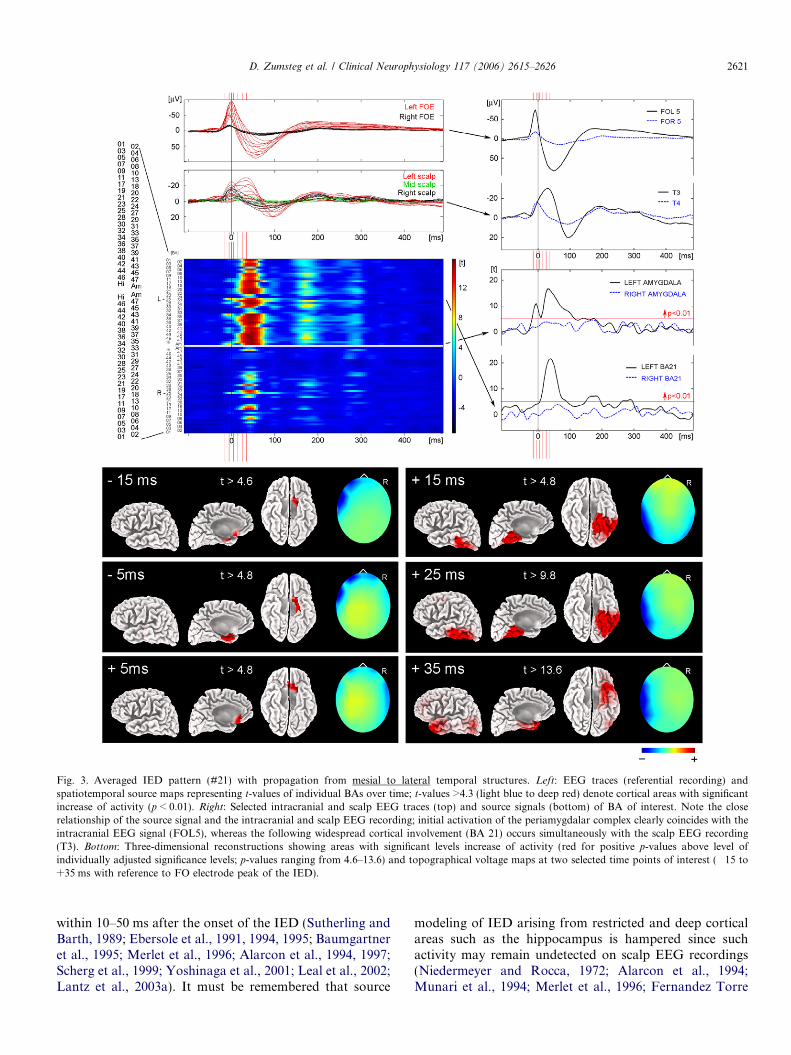
Fig. 3. Averaged IED pattern (#21) with propagation from mesial to lateral temporal structures. Left: EEG traces (referential recording) andspatiotemporal source maps representing t-values of individual BAs over time; t-values >4.3 (light blue to deep red) denote cortical areas with significantincrease of activity (p < 0.01). Right: Selected intracranial and scalp EEG traces (top) and source signals (bottom) of BA of interest. Note the closerelationship of the source signal and the intracranial and scalp EEG recording; initial activation of the periamygdalar complex clearly coincides with theintracranial EEG signal (FOL5), whereas the following widespread cortical involvement (BA 21) occurs simultaneously with the scalp EEG recording(T3). Bottom: Three-dimensional reconstructions showing areas with significant levels increase of activity (red for positive p-values above level ofindividually adjusted significance levels; p-values ranging from 4.6–13.6) and topographical voltage maps at two selected time points of interest (�15 to+35 ms with reference to FO electrode peak of the IED).
D. Zumsteg et al. / Clinical Neurophysiology 117 (2006) 2615–2626 2621
within 10–50 ms after the onset of the IED (Sutherling andBarth, 1989; Ebersole et al., 1991, 1994, 1995; Baumgartneret al., 1995; Merlet et al., 1996; Alarcon et al., 1994, 1997;Scherg et al., 1999; Yoshinaga et al., 2001; Leal et al., 2002;Lantz et al., 2003a). It must be remembered that source
modeling of IED arising from restricted and deep corticalareas such as the hippocampus is hampered since suchactivity may remain undetected on scalp EEG recordings(Niedermeyer and Rocca, 1972; Alarcon et al., 1994;Munari et al., 1994; Merlet et al., 1996; Fernandez Torre
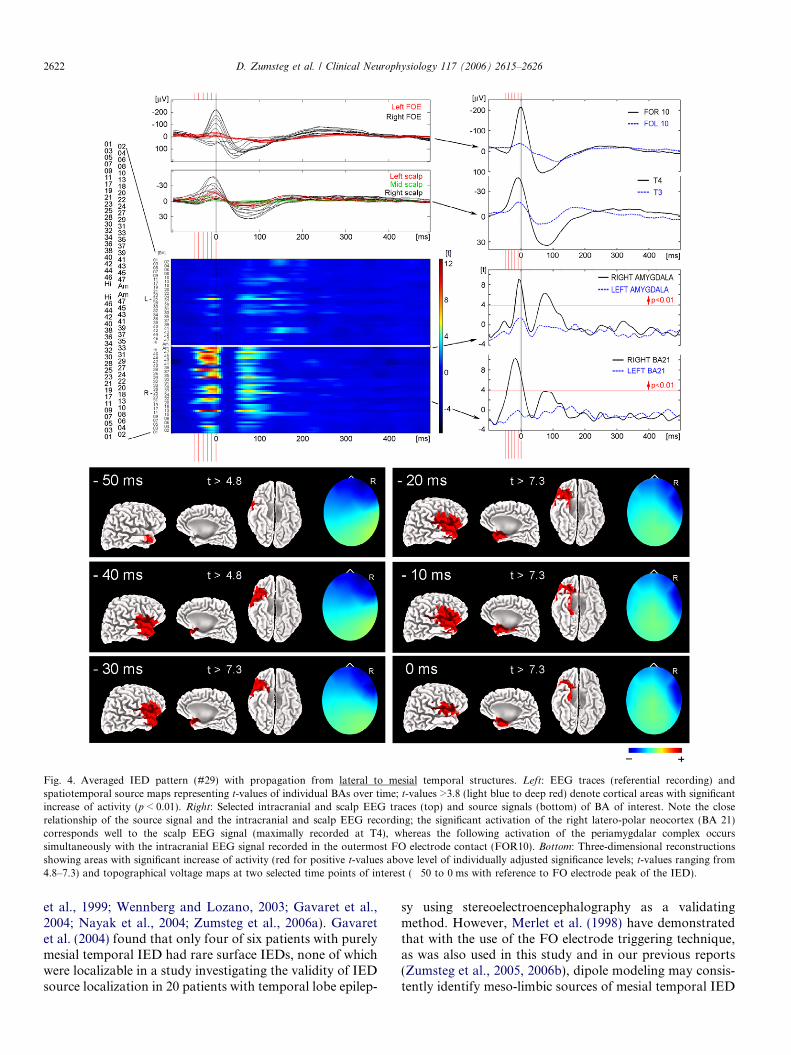
Fig. 4. Averaged IED pattern (#29) with propagation from lateral to mesial temporal structures. Left: EEG traces (referential recording) andspatiotemporal source maps representing t-values of individual BAs over time; t-values >3.8 (light blue to deep red) denote cortical areas with significantincrease of activity (p < 0.01). Right: Selected intracranial and scalp EEG traces (top) and source signals (bottom) of BA of interest. Note the closerelationship of the source signal and the intracranial and scalp EEG recording; the significant activation of the right latero-polar neocortex (BA 21)corresponds well to the scalp EEG signal (maximally recorded at T4), whereas the following activation of the periamygdalar complex occurssimultaneously with the intracranial EEG signal recorded in the outermost FO electrode contact (FOR10). Bottom: Three-dimensional reconstructionsshowing areas with significant increase of activity (red for positive t-values above level of individually adjusted significance levels; t-values ranging from4.8–7.3) and topographical voltage maps at two selected time points of interest (�50 to 0 ms with reference to FO electrode peak of the IED).
2622 D. Zumsteg et al. / Clinical Neurophysiology 117 (2006) 2615–2626
et al., 1999; Wennberg and Lozano, 2003; Gavaret et al.,2004; Nayak et al., 2004; Zumsteg et al., 2006a). Gavaretet al. (2004) found that only four of six patients with purelymesial temporal IED had rare surface IEDs, none of whichwere localizable in a study investigating the validity of IEDsource localization in 20 patients with temporal lobe epilep-
sy using stereoelectroencephalography as a validatingmethod. However, Merlet et al. (1998) have demonstratedthat with the use of the FO electrode triggering technique,as was also used in this study and in our previous reports(Zumsteg et al., 2005, 2006b), dipole modeling may consis-tently identify meso-limbic sources of mesial temporal IED

D. Zumsteg et al. / Clinical Neurophysiology 117 (2006) 2615–2626 2623
patterns: the source localizations were in good accordancewith the FO electrode contact recording the intracranialspike with maximal amplitude. Bast et al. (2004, 2005)investigated interictal activity using a sequential analyisdipole modeling technique (Scherg et al., 1999) in patientswith neocortical epilepsy due to cortical malformations. Infocal cortical dysplasia, IED patterns were complex in fiveof nine patients, thus requiring two dipole sources forappropriate modeling of propagation during the earlyphase of the IED pattern (Bast et al., 2004). In polymicro-gyria, a single dipole was appropriate in two of ninepatients, while two (four patients) or four (one patient)equivalent dipoles were required in five patients, and theremaining two patients revealed even more complex, rotat-ing activity during the early phase of the IED patterns(Bast et al., 2005).
Studies using distributed source models, which take intoaccount that IED may originate from a complex pattern ofcortical sources due to sequential activation of differentmesiobasal, lateral temporal or even distant neocorticalareas, have also shown evidence for epileptic source prop-agation (Fuchs et al., 1999; Huppertz et al., 2001; Lantzet al., 2001, 2003a). Huppertz et al. (2001), for example,demonstrated early propagation (i.e., before the peak ofthe IED) of epileptiform activity to anterior, posterior,and partly contralateral temporal regions in two of threepatients with lateral temporal lesions using non-linear min-imum L1-norm current density reconstructions. Fuchs et al.(1999) compared the localization accuracy of several linearand non-linear current density reconstructions (as well asdipole models) in two IED patterns and found that rawLORETA analysis resulted in the most blurred and toodeeply localized solutions, whereas the non-linear methodsyielded more focal results and the L1-norm results seemedto be overfocused (the inclusion of orientational con-straints, however, resulted in much more focused LORE-TA solutions). Lantz et al. (2003a) investigated IEDpatterns of 15 patients with focal epilepsy using a spatio-temporal segmentation procedure (Pascual-Marqui et al.,1995) and EPIFOCUS source localization. The authorsfound one to five different voltage maps during the risingphase of the IEDs, clearly suggesting early propagationof the IED in most of their patients (only three of 16patients showed a single map with source solutions withinthe epileptogenic lesion during the entire rising phase of theIED, whereas the remaining revealed several maps withsource solutions either within (five patients) or outside(eight patients) the epileptogenic lesion). These findingsclearly indicate the presence of several consecutively acti-vated generators during the course of surface recordedIEDs.
The main aim of the present study was to investigate theusefulness of SNPM LORETA source maps for the analy-sis of intracerebral propagation or extension of interictalepileptiform activity in IED patterns by using clinicalEEG data of simultaneous scalp and intracranial FO elec-trode recordings. Hence, the methods of this study differ
from previous investigations in several ways. First, unlikeprevious studies using distributed source models, we per-formed timeframe-wise comparison of the source signalon a statistical basis. We have previously established thevalidity of our SNPM LORETA approach for the localiza-tion of mesial temporal IED. We found that SNPMLORETA, but not a raw LORETA analysis of averagedIED patterns (as commonly performed in previous LORE-TA studies), accurately localized the source generators ofmesial temporal IED (Zumsteg et al., 2005, 2006b). Second,our approach allowed for direct correlation of SNPMLORETA source solutions with the findings of simulta-neous intracranial EEG recordings (see Figs. 3 and 4). Inprevious distributed source model studies, the localizationsof model-defined sources have usually been correlated tosites of structural MRI lesions and/or ‘‘confirmed’’ by apositive postsurgical outcome (Huppertz et al., 2001; Lantzet al., 2003a). This approach is based on the assumptionthat the site of the epileptogenic lesion, whether definedby MRI or by positive postsurgical outcome, necessarilyreflects the source(s) of the interictal IED. This assumptionmay be correct in many cases, but it is nevertheless an over-simplification and may actually be misleading, in particularwith respect to propagating activity and with respect topatients with MTLE. In these patients, interictal epilepti-form activity is well known to occur (or propagate) distantto or even contralateral to the seizure onset zone and neo-cortical spikes may frequently be recorded independentlyfrom any involvement of the mesial limbic structures(and are not of prognostic significance with respect to post-surgical seizure control after AHE (Wennberg et al.,1997a,b)). Hence, intracranial EEG recordings remain thegold standard for the accurate delineation of IED. We ofcourse acknowledge that some of the above mentionedstudies have also used intracranial EEG recordings for val-idation but, to our knowledge, invasive EEG recordingshave not been performed simultaneously with the scalpEEG recordings in any of these studies using distributedsource models. Given the difficulties in the assignment ofscalp-recorded IED to match with results from precedingor subsequent invasive EEG recordings, however, simulta-neous recording is clearly preferable. This is especially truein patients with more than one pattern of IED (as was thecase in more than half of the patients in our collective).Furthermore, the use of FO electrodes for simultaneousrecording has clear advantages in comparison to otherintracranial electrodes since FO electrodes do not needtrepanation, thus avoiding alterations of the ‘‘conductiveproperties’’ of the skull and resultant inappropriatenessof the head models used by the model. Third, we have usedspatiotemporal SNPM LORETA source maps for theinvestigation of IED propagation. Such maps illustratethe complete information of the calculated source (i.e., allBAs over the entire course of the IED) and are thus idealfor the determination of fast changing cortical sources.Considering that intracerebral propagation most likelyrepresents a very fast and probably slightly asynchronous

2624 D. Zumsteg et al. / Clinical Neurophysiology 117 (2006) 2615–2626
process approximating a continuous phenomenon, ratherthan taking place in discrete measureable stages, we wouldsuggest that such source maps are superior to spatiotempo-ral segmentation procedures as used in the study of Lantzet al. (2003a), for example.
Overall, we found a high concordance between FO elec-trode recordings and model-defined mesial temporal sourc-es. The statistical correlate of the source signal (t-values)within mesial temporal structures generally paralleled theintracranial EEG signal, whereas the t-values correspond-ing to basolateral to lateral temporal cortical areas typical-ly showed a close temporal relationship with the scalp EEGsignal recorded from the surface electrode nearest to thesecortical structures. We found mesial to lateral propagationin 20%, lateral to mesial propagation in 13.3% and simul-taneous (within 15 ms) activation of mesial and lateral tem-poral structures in 20% of IED patterns. This propagationgenerally occurred within 35 ms and was always limited toipsilateral cortical regions. In about half of the IED pat-terns (46.7%), however, the SNPM source solutionsremained restricted to mesial temporal structures duringthe entire course of the IED (those five IED patterns withno significant source solution included). It is interesting tonote that propagation between mesial and lateral temporalareas (and vice versa) frequently involved the lateral tem-poral pole and the temporobasal cortex, occurring in halfof the IED patterns with mesial to lateral propagation(IED patterns 24–26), and in three of four IED patternswith lateral to mesial propagation (28–30) in our study.This ‘‘mediating’’ role of the temporal pole and temporo-basal cortical areas is in keeping with the anatomy of thisregion (it is anatomically connected to both mesial and lat-eral temporal structures). Moreover, it is also in goodaccordance with the findings of a previous study by Bouri-en et al. (2005). These authors found the internal temporalpole and the temporobasal cortex to be conjointly involvedwith IED originating in mesial and lateral temporal struc-tures, as evidenced by intracerebral recordings in 15patients with MTLE. With respect to intracranial electricalfield characteristics, IED patterns with propagation orextension to extramesial areas tended to have a wide andanterior intracranial electrical field, suggesting a morewidespread and more superficial cortical activation. Thestrictly mesial IED patterns, in contrast, generally revealeda restricted intracranial field in FO electrode recordingsand, without the help of averaging, no clear counterpartin scalp EEG recordings. However, it is important to notethat this does not necessarily mean that IED patterns withrestricted intracranial fields would not show propagationor extension into ipsilateral basolateral or lateral temporalareas. We actually found mesial to lateral propagation intwo, and simultaneous activation of mesial and lateral tem-poral areas in three of those IED patterns.
There are a number of limitations in our methods thatneed to be acknowledged with respect to the study of prop-agated activity. First, it is difficult to estimate the extent ofpropagated activity outside of the restricted region from
which the FO electrodes are recording. Whereas FO elec-trodes are well positioned to record from basomedial tem-poral cortex, they are relatively far from the temporal tip,basolateral and lateral temporal cortices (Wieser and Sch-warz, 2001). Notwithstanding, there is indirect evidencethat one can seemingly trust those SNPM LORETA solu-tions. Numerous studies have shown that IED patternswith wide intracranial electrical fields (in particular thosewith wide anterior fields) represent more widespread prop-agated activity involving mesial, basal and lateral areas ofthe temporal cortex (Alarcon et al., 1994). Furthermore, itis in good accordance with our own experience and under-standing derived from FO electrode recordings and subse-quent tailored amygdalo-hippocampectomy under ECoGguidance in over 400 patients with MTLE (we usually per-form intraoperative ECoG recordings with simultaneous 4contact strip electrodes recording from the hippocampusand 32 contact grid electrodes placed over the ipsilateralbasal and lateral region). Second, the spatial resolution ofLORETA is admittedly limited. This spatial smearing effectof LORETA is particularly evident in cases where IED pat-terns reveal a high signal to noise ratio (e.g., towards thepeak of the IED). In such situations, SNPM LORETAmay offer very extended and widespread source solutions,raising the question of whether in such cases propagationmight reflect simple smearing-related ‘‘extension’’ of sourcesolutions rather than true sequential activation of distinctcortical areas. Although we cannot exclude this possibilitywith certainty in all situations, there are nevertheless sever-al arguments making us believe that SNPM LORETA isindeed capable of accurately localizing sources of propa-gated activity. Most importantly, simple smearing-related‘‘extension’’ is very unlikely to occur in cases where the sec-ondary source is actually emerging in an area that is ana-tomically distant to the primary source, while theprimary source is lost at the same time (see Fig. 3, forexample). With simple extension caused by smearing onewould expect the ‘‘second’’ source to be simply ‘‘addedto’’ and adjacent to the primary source. Furthermore,propagation was commonly accompanied by obviouschanges in the topographical voltage maps during thecourse of the IED, clearly suggesting propagation. Howev-er, the spatial smearing effect of LORETA is also criticalwith respect to the localization of activation to BA, as per-formed for the creation of spatiotemporal source maps.Moreover, there is no exact functional delineation of theseareas and most individuals will not have a brain thatmatches the ‘‘generic brain’’ of the Talairach atlas. Third,the low number (23) of scalp electrodes and the low sam-pling frequency of 200 Hz used in this study are other lim-itations with respect to investigation of the propagation ofIED. This is of course due to the fact that these are clinicaldata, analyzed retrospectively. The localization accuracyfor LORETA and other distributed source models has beenshown to increase substantially up to 64 electrodes (Lantzet al., 2003b). However, some of the most critical addition-al electrodes in the basal temporal regions were at least par-

D. Zumsteg et al. / Clinical Neurophysiology 117 (2006) 2615–2626 2625
tially represented (mastoids, T1/2). Fourth, it must also beacknowledged that LORETA is using a three shell spheri-cal head model with assumed conductivities of scalp, skull,and brain, which may lead to inaccurate localization ofsources, irrespective of the number of sensors used. Lastly,analysis of propagation was clearly limited by the low sam-pling frequency of 200 Hz. We considered activity to bepropagating only if the propagation (from mesial to lateraland vice versa) extended over at least three sample points(15 ms). It can be assumed that with a higher samplingfrequency at least some of the six IED patterns with simul-taneous activation of mesial and lateral temporal structureswould actually have reflected IED patterns showing fastmesial to lateral propagation, or vice versa.
Statistical correlation of the electrophysiological IEDcharacteristics and postsurgical outcome was limited a pri-ori by the small sample size of this study (only one patientin each of the unfavorable outcome classes II–IV). Never-theless, the lack of a clear association between the postsur-gical outcome and the number, laterality or propagationcharacteristics of IED patterns is consistent with previousreports showing neocortical spikes to be of no prognosticsignificance for outcome after selective AHE in patientswith MTLE (Polkey et al., 1989; Cendes et al., 1993; Wenn-berg et al., 1997b, 1998).
In conclusion, the findings of this study confirm the com-mon occurrence of mesial to lateral temporal propagation ofIED, and vice versa, and reemphasize that such propagationmust always be taken into consideration when using non-in-vasive source algorithms in patients with MTLE. Spatiotem-poral mapping of SNPM LORETA allows for reliabledescription of such propagation or extension. However, itmust be emphasized again that IED arising from deep mesialtemporal structures usually remain undetected in scalp EEGrecordings and, therefore, from a clinical point of view, ourtechnique might not be helpful for the identification of prop-agated interictal activity of mesial temporal onset. If futureresearch in this area using combined intracranial and scalpEEG might eventually facilitate the development of somenoninvasive recording technique that could, with a reason-ably high degree of sensitivity and specificity, identify focaltemporal limbic epileptiform activity, this would representa true advance, though the means of achieving this technicalgoal are not immediately obvious.
Acknowledgment
This project has been supported by the Swiss NationalScience Foundation (Grant PA00A-101502).
References
Alarcon G, Guy CN, Binnie CD, Walker SR, Elwes RD, Polkey CE.Intracerebral propagation of interictal activity in partial epilepsy:implications for source localisation. J Neurol Neurosurg Psychiatry1994;57:435–49.
Alarcon G, Garcia Seoane JJ, Binnie CD, Martin Miguel MC, Juler J,Polkey CE, et al. Origin and propagation of interictal discharges in
the acute electrocorticogram. Implications for pathophysiology andsurgical treatment of temporal lobe epilepsy. Brain 1997;120:2259–82.
Bast T, Oezkan O, Rona S, Stippich C, Seitz A, Rupp A, et al. EEG andMEG source analysis of single and averaged interictal spikes revealsintrinsic epileptogenicity in focal cortical dysplasia. Epilepsia2004;45:621–31.
Bast T, Ramantani G, Boppel T, Metzke T, Ozkan O, Stippich C, et al.Source analysis of interictal spikes in polymicrogyria: loss of relevantcortical fissures requires simultaneous EEG to avoid MEG misinter-pretation. Neuroimage 2005;25:1232–41.
Baumgartner C, Lindinger G, Ebner A, Aull S, Serles W, Olbrich A, et al.Propagation of interictal epileptic activity in temporal lobe epilepsy.Neurology 1995;45:118–22.
Bourien J, Bartolomei F, Bellanger JJ, Gavaret M, Chauvel P, WendlingF. A method to identify reproducible subsets of co-activated structuresduring interictal spikes. Application to intracerebral EEG in temporallobe epilepsy. Clin Neurophysiol 2005;116:443–55.
Buser P, Bancaud J. Unilateral connections between amygdala andhippocampus in man: a study of epileptic patients with depthelectrodes. Electroencephalogr Clin Neurophysiol 1983;55:1–12.
Cendes F, Dubeau F, Olivier A, Cukiert A, Andermann E, Quesney F,et al. Increased neocortical spiking and surgical outcome afterselective amygdalo-hippocampectomy. Epilepsy Res 1993;16:195–206.
Ebersole JS. EEG dipole modelling in complex partial epilepsy. BrainTopogr 1991;4:113–23.
Ebersole JS. Non-invasive localization of the epileptogenic focus by EEGdipole modeling. Acta Neurol Scand Suppl 1994;152:20–8.
Ebersole JS, Wade PB. Spike voltage topography and equivalentdipole localization in complex partial epilepsy. Brain Topogr1990;3:21–34.
Ebersole JS, Hawes S, Scherg M. Intracranial EEG validation of spikepropagation predicted by dipole models. Electroencephalogr ClinNeurophysiol 1995;95:18P–9P.
Emerson RG, Turner CA, Pedley TA, Walczak TS, Forgione M.Propagation patterns of temporal spikes. Electroencephalogr ClinNeurophysiol 1995;94:338–48.
Engel Jr J. Intracerebral recordings: organization of the human epilep-togenic region. J Clin Neurophysiol 1993;10:90–8.
Engel Jr J, Van Ness PC, Rasmussen TB, Ojemann LM. Outcome withrespect to epileptic seizures. In: Engel Jr J, editor. Surgical treatment ofthe epilepsies. NewYork: Raven Press; 1993. p. 09–621.
Feindel W, Penfield W. Localization of discharge in temporal lobeautomatism. AMA Arch Neurol Psychiatry 1954;72:603–30.
Fernandez Torre JL, Alarcon G, Binnie CD, Polkey CE. Compar-ison of sphenoidal, foramen ovale and anterior temporalplacements for detecting interictal epileptiform discharges inpresurgical assessment for temporal lobe epilepsy. Clin Neuro-physiol 1999;110:895–904.
Fuchs M, Wagner M, Kohler T, Wischmann HA. Linear and nonlinearcurrent density reconstructions. J Clin Neurophysiol 1999;16:267–95.
Gavaret M, Badier JM, Marquis P, Bartolomei F, Chauvel P. Electricsource imaging in temporal lobe epilepsy. J Clin Neurophysiol2004;21:267–82.
Gloor P. The temporal lobe and the limbic system. New York,Oxford: Oxford University Press; 1997.
Hamer HM, Najm I, Mohamed A, Wyllie E. Interictal epileptiformdischarges in temporal lobe epilepsy due to hippocampal sclerosisversus medial temporal lobe tumors. Epilepsia 1999;40:1261–8.
Hirsch LJ, Spencer SS, Williamson PD, Spencer DD, Mattson RH.Comparison of bitemporal and unitemporal epilepsy defined by depthelectroencephalography. Ann Neurol 1991;30:340–6.
Hughes JR. Long-term clinical and EEG changes in patients with epilepsy.Arch Neurol 1985;42:213–23.
Huppertz HJ, Hoegg S, Sick C, Lucking CH, Zentner J, Schulze-Bonhage A, et al. Cortical current density reconstruction ofinterictal epileptiform activity in temporal lobe epilepsy. ClinNeurophysiol 2001;112:1761–72.

2626 D. Zumsteg et al. / Clinical Neurophysiology 117 (2006) 2615–2626
Lantz G, Grave de Peralta R, Spinelli L, Seeck M, Michel CM. Epilepticsource localization with high density EEG: how many electrodes areneeded? Clin Neurophysiol 2003b;114:63–9.
Lantz G, Spinelli L, Seeck M, Peralta Menendez RG, Sottas CC, MichelCM. Propagation of interictal epileptiform activity can lead toerroneous source localizations: a 128-channel EEG mapping study. JClin Neurophysiol 2003a;20:311–9.
Leal AJ, Passao V, Calado E, Vieira JP, Silva Cunha JP. Interictal spikeEEG source analysis in hypothalamic hamartoma epilepsy. ClinNeurophysiol 2002;113:1961–9.
Marks DA, Katz A, Booke J, Spencer DD, Spencer SS. Comparison andcorrelation of surface and sphenoidal electrodes with simultaneousintracranial recording: an interictal study. Electroencephalogr ClinNeurophysiol 1992;82:23–9.
Marsan CA, Zivin LS. Factors related to the occurrence of typicalparoxysmal abnormalities in the EEG records of epileptic patients.Epilepsia 1970;11:361–81.
Merlet I, Garcia-Larrea L, Gregoire MC, Lavenne F, Mauguiere F.Source propagation of interictal spikes in temporal lobe epilepsy.Correlations between spike dipole modelling and [18F]fluorodeoxy-glucose PET data. Brain 1996;119:377–92.
Merlet I, Garcia-Larrea L, Ryvlin P, Isnard J, Sindou M, Mauguiere F.Topographical reliability of mesio-temporal sources of interictal spikesin temporal lobe epilepsy. Electroencephalogr Clin Neurophysiol1998;107:206–12.
Merlet I, Gotman J. Reliability of dipole models of epileptic spikes. ClinNeurophysiol 1999;110:1013–28.
Munari C, Hoffmann D, Francione S, Kahane P, Tassi L, Lo RG, et al.Stereo-electroencephalography methodology: advantages and limits.Acta Neurol Scand Suppl 1994;152:56–67.
Nayak D, Valentin A, Alarcon G, Garcia Seoane JJ, Brunnhuber F, JulerJ, et al. Characteristics of scalp electrical fields associated with deepmedial temporal epileptiform discharges. Clin Neurophysiol2004;115:1423–35.
Nichols TE, Holmes AP. Nonparametric permutation tests for functionalneuroimaging: a primer with examples. Hum Brain Mapp2002;15:1–25.
Niedermeyer E, Rocca U. The diagnostic significance of sleep electroen-cephalograms in temporal lobe epilepsy. A comparison of scalp anddepth tracings. Eur Neurol 1972;7:119–29.
Pandya DN, Yeterian EH. Hodology of limbic and related structures:cortical and commissural connections. In: Wieser HG, Elger CE,editors. Presurgical evaluation of epileptics. Berlin: Springer-Verlag;1987. p. 3–14.
Pascual-Marqui RD. Review of methods solving the EEG inverseproblem. Int J Bioelectromagnetism 1999;1:75–86.
Pascual-Marqui RD, Michel CM, Lehmann D. Low resolution electro-magnetic tomography: a new method for localizing electrical activity inthe brain. Int J Psychophysiol 1994;18:49–65.
Pascual-Marqui RD, Michel CM, Lehmann D. Segmentation of brainelectrical activity into microstates: model estimation and validation.IEEE Trans Biomed Eng 1995;42:658–65.
Polkey C, Binnie C, Janota I. Acute hippocampal recording andpathology at temporal lobe resection and amygdalo-hippocampectomyfor epilepsy. J Neurol Neurosurg Psychiatry 1989;52:1050–7.
Scherg M, Bast T, Berg P. Multiple source analysis of interictal spikes:goals, requirements, and clinical value. J Clin Neurophysiol1999;16:214–24.
So N, Gloor P, Quesney LF, Jones-Gotman M, Olivier A, Andermann F.Depth electrode investigations in patients with bitemporal epileptiformabnormalities. Ann Neurol 1989;25:423–31.
Stefan H, Schneider S, Abraham-Fuchs K, Pawlik G, Feistel H, Bauer J,et al. The neocortico to mesio-basal limbic propagation of focal
epileptic activity during the spike-wave complex. ElectroencephalogrClin Neurophysiol 1991;79:1–10.
Sutherling WW, Barth DS. Neocortical propagation in temporal lobespike foci on magnetoencephalography and electroencephalography.Ann Neurol 1989;25:373–81.
Talairach J, Tournoux P. Co-planar stereotaxic atlas of the humanbrain. Stuttgart: Thieme; 1988.
Towle VL, Bolanos J, Suarez D, Tan K, Grzeszczuk R, Levin DN, et al.The spatial location of EEG electrodes: locating the best-fitting sphererelative to cortical anatomy. Electroencephalogr Clin Neurophysiol1993;86:1–6.
Wennberg R, Quesney F, Olivier A, Dubeau F. Mesial temporal versuslateral temporal interictal epileptiform activity: comparison of chronicand acute intracranial recordings. Electroencephalogr Clin Neuro-physiol 1997a;102:486–94.
Wennberg R, Quesney F, Olivier A, Dubeau F. Induction of burst-suppression and activation of epileptiform activity after methohexitaland selective amygdalo-hippocampectomy. Electroencephalogr ClinNeurophysiol 1997b;102:443–51.
Wennberg RA, Lozano AM. Intracranial volume conduction of corticalspikes and sleep potentials recorded with deep brain stimulatingelectrodes. Clin Neurophysiol 2003;114:1403–18.
Wennberg RA, Quesney LF, Dubeau F, Olivier A. Increasedneocortical spiking and surgical out-come after selective amygd-alo-hippocampectomy. In: Quesney LF, Binnie CD, Chatrian G-E, editors. Electrocorticography: current trends and futureperspectives. Electroenceph Clin Neurophysiol Suppl, vol.8. NEw York: WIley; 1998. p. 105–11.
Wieser HG, Ortega M, Friedman A, Yonekawa Y. Long-term seizureoutcomes following amygdalo-hippocampectomy. J Neurosurg2003;98:751–63.
Wieser HG, Schwarz U. Topography of foramen ovale electrodes by 3Dimage reconstruction. Clin Neurophysiol 2001;112:2053–6.
Wieser HG. Electroclinical features of the psychomotor seizure. Lon-don: Butterworths; 1983.
Wieser HG. Stereoelectroencephalography and foramen ovale electroderecording. In: Niedermeyer E, Lopes da Silva F, editors. Electroen-cephalography: basic principles, clinical applications, and relatedfields. Baltimore: Williams & Wilkins; 1999. p. 725–40.
Williamson PD, French JA, Thadani VM, Kim JH, Novelly RA, SpencerSS, et al. Characteristics of medial temporal lobe epilepsy: II.Interictal and ictal scalp electroencephalography, neuropsychologicaltesting, neuroimaging, surgical results, and pathology. Ann Neurol1993;34:781–7.
Wilson CL, Isokawa M, Babb TL, Crandall PH. Functional connectionsin the human temporal lobe: I. Analysis of limbic system pathwaysusing neuronal responses evoked by electrical stimulation. Exp BrainRes 1990;82:279–92.
Yoshinaga H, Hattori J, Nakahori T, Ohtsuka Y, Oka E, Tomita S, et al.Combined use of sphenoidal electrodes and the dipole localizationmethod for the identification of the mesial temporal focus. Eur JNeurol 2001;8:149–56.
Zumsteg D, Andrade DM, Wennberg RA. Clinical electrophysiologyfactors indicative of intractability in patients with temporal lobeepilepsy. Adv Neurol 2006a;97:45–62.
Zumsteg D, Friedman A, Wennberg RA, Wieser HG. Source localizationof mesial temporal interictal epileptiform discharges: correlation withintracranial foramen ovale electrode recordings. Clin Neurophysiol2005;116:2810–8.
Zumsteg D, Friedman A, Wieser HG, Wennberg RA. Source localizationof interictal epileptiform discharges: comparison of three differenttechniques to improve signal to noise ratio. Clin Neurophysiol2006b;117:562–71.
Copyright © 2022 FDOKUMEN




















