
Differential expression and function of PACAP and VIP receptors in four human colonic adenocarcinoma...
14
Cell. Signal. Vol. 10, No. 1, pp. 13–26, 1998 ISSN 0898-6568/98 $19.00 Copyright 1998 Elsevier Science Inc. PII S0898-6568(97)00067-3 Differential Expression and Function of PACAP and VIP Receptors in Four Human Colonic Adenocarcinoma Cell Lines Vincent Lelie `vre ‡, Annie-Claire Meunier , Evelyne Caigneaux , Jack Falcon‡ and Jean-Marc Muller * Laboratoire de Biologie des Interactions Cellulaires and ‡Laboratoire de Neurobiologie Cellulaire, CNRS UMR 6558, Universite ´ de Poitiers, Faculte ´ des Sciences, 40 avenue du Recteur Pineau, 86022 Poitiers Cedex, France ABSTRACT. Human colonic adenocarcinoma cell lines have conserved several features of the native tissue. Among these is the expression of cell surface receptors for hormones and neurotransmitters that may be involved in the regulation of proliferation and differentiation processes in these cancer cells. Here, we confirm that high- affinity binding sites for the Vasoactive Intestinal Polypeptide (VIP) and for the VIP analogue Pituitary Adenyl- ate-Cyclase Activating Polypeptide (PACAP), were expressed in 4 human colonic adenocarcinoma cell lines, HT29, SW403, DLD-1 and Caco-2, that spontaneously displayed variable phenotypic properties in culture. We demonstrated that after long-term treatments, VIP and PACAP significantly reduced cell proliferation in the 4 cell lines and modulated intracellular cAMP and cGMP levels. Furthermore, conspicuous differences were ob- served from one cell type to another concerning expression of the receptor subsets or the effects of the neuropep- tides on cell growth and on cyclic nucleotides production. cell signal 10;1:13–26, 1998. 1998 Elsevier Science Inc. KEY WORDS. Human colonic cancer cell lines, VIP/PACAP receptors, Cell signalling, Cell growth and differentiation INTRODUCTION ties and on the recent cloning of the corresponding cDNAs. In the aggregate, the three VIP and PACAP receptors so far The ubiquitous 28-amino-acid neuropeptide Vasoactive In- fully characterised are now referred to as PACAP1, VIP1/ testinal polypeptide (VIP), isolated from gut, triggers im- PACAP2 and VIP2/PACAP3 [5]. The PACAP1 subtype portant processes in the gastrointestinal tract, such as stim- represents preferential receptors for both forms of PACAP ulation of smooth-muscle relaxation or of water and of 27 and 38 amino acids (deriving from differential proteol- electrolytes secretion [1]. A rich VIPergic innervation is ysis of the same precursor), with low affinity for VIP [6]. In supplied all along the gut through the autonomic ganglia of contrast, the VIP1/PACAP2 and VIP2/PACAP3 receptors the myenteric plexuses whose neuronal terminations liber- are polyvalent and share similar affinities for VIP and both ate the neuropeptide in close contact to the muscular layers forms of PACAP; they correspond to the ‘‘classical’’ VIP and to the epithelium [2]. The colon, particularly its as- receptors characterised years ago before the discovery of cending part, together with the coecum and duodenum, are PACAP, and are generally positively coupled to adenylate- the regions of the gut where the highest concentrations of cyclase [7–8]. Different versions of the PACAP1 receptor immunoreactive VIP (up to 600 pmoles/g) were detected are produced by alternative splicing of a common precursor [3]. A VIP analogue called PACAP (Pituitary Adenylate- mRNA and trigger differential stimulation of adenylate cy- Cyclase Activating Polypeptide), isolated in 1989, shares clase or phospholipase C, depending on combinations of ad- with VIP very similar distribution and biological properties ditional cassettes (called hip, hop1 and hop2) inserted into in the large intestine [4]. The VIP and PACAP actions are the third intracellular loop of the receptor [9]. This intracel- mediated through different sets of receptors which have lular loop is known to be involved in the coupling of these been subtyped on the basis of their pharmacological proper- 7 transmembrane domains receptors to the G-proteins [10]. Two polyvalent receptors, VIP1/PACAP2 and VIP2/ *Author to whom all correspondence should be addressed. E-mail: ,Jean_ PACAP3, are encoded by distinct genes. The VIP1/ [email protected]_poitiers.fr. Received 30 March 1997; and accepted 2 May 1997. PACAP2 receptor appeared likely to be the major subtype
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Differential expression and function of PACAP and VIP receptors in four human colonic adenocarcinoma...

Cell. Signal. Vol. 10, No. 1, pp. 13–26, 1998 ISSN 0898-6568/98 $19.00Copyright 1998 Elsevier Science Inc. PII S0898-6568(97)00067-3
Differential Expression and Functionof PACAP and VIP Receptors in Four
Human Colonic Adenocarcinoma Cell LinesVincent Lelievre†‡, Annie-Claire Meunier†,
Evelyne Caigneaux†, Jack Falcon‡ and Jean-Marc Muller†*†Laboratoire de Biologie des Interactions Cellulaires and
‡Laboratoire de Neurobiologie Cellulaire, CNRS UMR 6558, Universitede Poitiers, Faculte des Sciences, 40 avenue du Recteur Pineau, 86022 Poitiers Cedex, France
ABSTRACT. Human colonic adenocarcinoma cell lines have conserved several features of the native tissue.Among these is the expression of cell surface receptors for hormones and neurotransmitters that may be involvedin the regulation of proliferation and differentiation processes in these cancer cells. Here, we confirm that high-affinity binding sites for the Vasoactive Intestinal Polypeptide (VIP) and for the VIP analogue Pituitary Adenyl-ate-Cyclase Activating Polypeptide (PACAP), were expressed in 4 human colonic adenocarcinoma cell lines,HT29, SW403, DLD-1 and Caco-2, that spontaneously displayed variable phenotypic properties in culture. Wedemonstrated that after long-term treatments, VIP and PACAP significantly reduced cell proliferation in the 4cell lines and modulated intracellular cAMP and cGMP levels. Furthermore, conspicuous differences were ob-served from one cell type to another concerning expression of the receptor subsets or the effects of the neuropep-tides on cell growth and on cyclic nucleotides production. cell signal 10;1:13–26, 1998. 1998 ElsevierScience Inc.
KEY WORDS. Human colonic cancer cell lines, VIP/PACAP receptors, Cell signalling, Cell growth anddifferentiation
INTRODUCTION ties and on the recent cloning of the corresponding cDNAs.In the aggregate, the three VIP and PACAP receptors so farThe ubiquitous 28-amino-acid neuropeptide Vasoactive In-fully characterised are now referred to as PACAP1, VIP1/testinal polypeptide (VIP), isolated from gut, triggers im-PACAP2 and VIP2/PACAP3 [5]. The PACAP1 subtypeportant processes in the gastrointestinal tract, such as stim-represents preferential receptors for both forms of PACAPulation of smooth-muscle relaxation or of water andof 27 and 38 amino acids (deriving from differential proteol-electrolytes secretion [1]. A rich VIPergic innervation isysis of the same precursor), with low affinity for VIP [6]. Insupplied all along the gut through the autonomic ganglia ofcontrast, the VIP1/PACAP2 and VIP2/PACAP3 receptorsthe myenteric plexuses whose neuronal terminations liber-are polyvalent and share similar affinities for VIP and bothate the neuropeptide in close contact to the muscular layersforms of PACAP; they correspond to the ‘‘classical’’ VIPand to the epithelium [2]. The colon, particularly its as-receptors characterised years ago before the discovery ofcending part, together with the coecum and duodenum, arePACAP, and are generally positively coupled to adenylate-the regions of the gut where the highest concentrations ofcyclase [7–8]. Different versions of the PACAP1 receptorimmunoreactive VIP (up to 600 pmoles/g) were detectedare produced by alternative splicing of a common precursor[3]. A VIP analogue called PACAP (Pituitary Adenylate-mRNA and trigger differential stimulation of adenylate cy-Cyclase Activating Polypeptide), isolated in 1989, sharesclase or phospholipase C, depending on combinations of ad-with VIP very similar distribution and biological propertiesditional cassettes (called hip, hop1 and hop2) inserted intoin the large intestine [4]. The VIP and PACAP actions arethe third intracellular loop of the receptor [9]. This intracel-mediated through different sets of receptors which havelular loop is known to be involved in the coupling of thesebeen subtyped on the basis of their pharmacological proper-7 transmembrane domains receptors to the G-proteins[10]. Two polyvalent receptors, VIP1/PACAP2 and VIP2/
*Author to whom all correspondence should be addressed. E-mail: ,Jean_PACAP3, are encoded by distinct genes. The VIP1/[email protected]_poitiers.fr.
Received 30 March 1997; and accepted 2 May 1997. PACAP2 receptor appeared likely to be the major subtype

14 V. Lelievre et al.
represented in peripheral tissues, particularly in the large grade were from Sigma Chimie, France. 125I-Na (17 Ci/mg)and 125I-cAMP radioimmuno-assay kit were purchased fromintestine, with far higher levels of expression than the
PACAP1 and the VIP2/PACAP3 receptors. NEN-Du Pont, France. Microporous culture inserts (0.45mm membrane) were from Costar/polylabo, France.Since 1964, when Jorgen Fogh established the first human
colonic adenocarcinoma cell line HT29, more than 80 celllines have been derived from human colonic cancers [11]. Ac-
Cell Culture and Maintenancecording to the cell type considered, the cells displayed a varietyof phenotypes that were reminiscent of the different stages of The human colonic cells HT29, DLD-1, Caco-2 andthe malignant process, from apparently well-differentiated en- SW403 were routinely cultured in 75 cm2 culture flasks interocytes to severely dysplastic cells [12]. Furthermore, since Glutamax I DMEM (4.5 g/l glucose, without sodium py-they expressed receptors for a number of hormones and neuro- ruvate) supplemented with 10% (v/v) FCS and penicillin/transmitters, these cell lines provided precious models for in streptomycin (respectively 100 U/ml and 0.1 mg/ml), in avitro studies on regulations by different stimuli of functions of humidified atmosphere of air/CO2 (95%/5%), at 378C. Cul-the normal or malignant colonic tissue [13]. In some of these ture medium was removed and replaced by fresh mediumcell lines, enterocytelike differentiation could spontaneously every 3 d. For subcultures, cells were harvested in trypsin-take place (Caco-2, T84) or could be triggered in given culture EDTA solution (respectively 0.05/0.53mM) in PUCK’s me-conditions (HT29) [14–16]. For these reasons, they were ex- dium, for 10 min at 378C.tensively utilized for in vitro studies on the establishment of thecell polarity or on expression of specific enzymes of the colonic
Analysis of Cell Morphologyenterocytes [12]. High-affinity receptors for VIP (and PACAP,since the bivalence of the VIP receptors is now recognised), Fifty thousand cells were seeded in 1 ml culture medium, inwere demonstrated to be expressed in several colonic ade- 24-well dishes on 0.4 mm microporous polycarbonate mem-nocarcinoma cell lines [12, 17–18]. In some of them, like branes for light or electron microscopy studies. Medium wasCaco-2 or T84, these receptors triggered chloride secretion in removed and changed every 3 d until cells reached conflu-response to the stimuli, demonstrating that the cells possessed ency. The total time in culture to reach this stage was 14 d.all the necessary components to ensure this typical function of Cells on inserts were fixed for 1 h at room temperature,the normal differentiated enterocyte [19–20]. with 2.5% glutaraldehyde in sodium cacodylate buffer (0.1
Despite the important bibliography devoted to these M, pH 7.4). After 3 washes in PBS, cells were post-fixedfunctional aspects, much less is known on the involvement with 1% OsO4 in the same sodium cacodylate buffer. Forof VIP and PACAP in colonic tumorigenesis or on the be- light microscopy, cells were further dehydrated in ascendinghaviour of the colonic cancer cell lines after long-term acetone series, and included in araldite. Semi-thin sectionstreatments with these neuropeptides. Data from some re- were stained with toluidine blue. Observations and photo-ports, however, suggest that VIP and PACAP may modu- graphs were carried out on a Zeiss Axioplan microscopelate cell proliferation differentiation in human colonic ade- with X100 immersion lens.nocarcinoma cell lines [21–25]. Such properties have also For scanning electron microscopy (SEM), dehydratedbeen demonstrated in other epithelial cell types or in cells cells were washed with acetone before vacuum dessicationfrom the immune system or of neural or glial origin [25–26]. (in a Critical Point Dryer CPD, 020, Balzers). Then, a goldIn this context, we undertook to confirm that VIP and metallisation (metallisation device, Balzers) was achieved,PACAP could regulate cell proliferation in 4 colonic can- before observation with a JEOL JSM 35-CF scanning elec-cer cell lines: HT29, SW403, DLD-1 and Caco-2. This tron microscope.panel comprises cells commonly recognised as spontane-ously displaying enterocytelike properties (Caco-2), to be
Proliferation Assayspoorly differentiated (HT29) or to possess intermediate phe-notypes (DLD-1, SW403). With the aim of further charac- Proliferation studies were performed in extracellular matrixterising the functional mechanisms ensuring VIP and PACAP coated 24-well dishes seeded with 10.000 cells/well in 1 mleffects in these cells, we also investigated expression of the VIP culture medium. Precoating of the dishes with extracellularand PACAP receptors and effects of the neuropeptides on in- matrix was carried out according to a previously reportedtracellular cyclic nucleotides (cAMP and cGMP) production. procedure [27]. After 24 h, a daily addition (10 ml from a
100-fold concentrated solution) of specified polypeptideswas started. Culture medium was replaced every 3 d. At the
EXPERIMENTAL METHODS end of the treatments, the cells were harvested in 500 mlChemicals and Materials trypsin/EDTA and protease activity was stopped by addi-
tion of 500 ml 10% FCS in culture medium. Cell viabilityGlutamax-I Dulbecco’s Modified Eagle’s Medium (DMEM),penicillin/streptomycin, trypsin-EDTA solution, foetal calf was controlled by trypan blue exclusion. Cells from each
treatment performed in triplicate, were counted in dupli-serum (FCS) (Gibco BRL, France). VIP, PACAP-27 werefrom Neosystem, France. Bacitracin, phenylmethylsulfonyl cate using a Malassez chamber. Curve fittings of the data
were computed (GraphpadTM, ISI software/USA) accordingfluoride (PMSF) and current reagents of the highest purity

VIP- and PACAP-Mediated Inhibition of Cell Growth 15
to a sigmoid equation. The effects of different drugs tested bations, cells were placed on ice. Incubation medium wasremoved and replaced with 300 ml of 6% TCA solution.were evaluated using an analysis of variance (ANOVA).Cells were lysed by direct sonication in wells and the ex-tracts were evaporated. The resulting pellets were dissolvedin sodium acetate buffer (0.05 M, pH 5 5.9). After a shortPeptides Radioiodinationcentrifugation (in order to pellet DNA and cell debris),
For binding experiments, VIP and PACAP-27 were radioio- cAMP in the supernatants was quantified using a commer-dinated using a previously described chloramine-T tech- cially available cAMP radioimmuno-assay kit in a gamma-nique [28] with slight modifications. Briefly, 20 ml (about counter (LKB).1.8mCi) of a 125I-Na solution were mixed with 20 ml of a1024 M (about 6 mg) aqueous solution of VIP or PACAP-27. The reaction was initiated by adding Chloramine-T (2 RESULTSmg/ml) in a 0.2 M sodium phosphate buffer (pH 7.6) then Morphological Aspect of the
Colonic Adenocarcinoma Cell Lines in Culturestopped after 30 s incubation at room temperature with 20ml sodium metabisulfite (2 mg/ml) in a 0.2 M sodium phos- The morphological aspect of HT29, Caco-2, DLD-1 andphate buffer (pH 7.6). The radioiodination mix was first SW403 adenocarcinoma cells was examined at confluenteluted from a C18 SEP PAK column (Waters Associates), stages of the cultures, under light (Fig. 1) and scanning elec-with 85% acetonitrile in H2O-0.1% Trifluoroacetic acid tron (Fig. 2) microscopes. The HT29 cells appeared very(TFA) after a rinsing of the column in 10 ml H2O-0.1% poorly differentiated, growing in multilayers and presentingTFA in order to discard the free radioiodine. The sample wide intracellular spaces (Fig. 1A). Under the scanningcontaining radioiodinated peptides was separated by reverse electron microscope, their cell surface, devoid of typicalphase HPLC (Spectraphysics), using a 5 mm VYDAC C18 microvilli, displayed long irregular cellular extensionscolumn (Interchrom, France). Elution was conducted for (Fig. 2A).20 min with 0–85% linear gradient of acetonitrile in As reported by others [12], enterocytelike features wereH2O-0.1% TFA. Fractions corresponding to the monoiodi- observed for DLD-1, Caco-2 and SW403 cell lines (Fig. 1B–nated forms of VIP or PACAP-27 typically giving high spe- D). These cells were growing as monolayers, although mi-cific binding were evaporated under nitrogen and stored at crotumourlike cell foci were also observed emerging from2208C. DLD-1 and SW403 monolayers, (Fig. 1B, C). The cells also
displayed an enterocytelike morphology with an overall po-larised aspect, very reduced intercellular spaces, and a dense
Binding 125I Radiotracer Peptides apical membrane reminiscent of a brush border. Indeed, thepresence of short and regular microvilli on the apical mem-Binding studies were performed on intact cells, according tobrane (with a higher density in Caco-2 and DLD-1 than inpreviously reported conditions [29]. Briefly, cells (2.5 3 105/SW403 cells) was observed under the scanning electron mi-well), were seeded in 24-well dishes and cultured for 3 d.croscope (Fig. 2B–D). In DLD-1 and SW403 cell lines, theThe cells were then incubated for 3 h at 48C, in presencemicrovilli of adjacent cells underlined very conspicuouslyof 20–25 pM 125I-VIP or 125I-PACAP and increasing concen-the cell outlines (Fig. 2B, C). In Caco-2 cells, peculiar star-trations of VIP or PACAP-27, in DMEM-150 mM HEPESshaped aggregates of microvilli were observed (Fig. 2D),(pH 7.4), containing 1% BSA, 0.1% bacitracin, and 150that have been previously described by others [30].mM PMSF. Binding reactions were stopped by cooling
dishes on ice. Cells were rinsed once with 2 ml cold PBS-0.1% BSA and lysed in 400 ml of 0.5 NaOH. Radioactivity Effects of VIP and PACAP27 on Cell Proliferationin the cell lysates was quantified in a gamma counting sys-
The proliferation data (Fig. 3), are summarized in Table 1,tem (LKB). Fitting of the data by non-linear regression waswhere two characteristic parameters have been evaluatedcomputed according to either a Hill or a two-site competi-for each experimental condition, by fitting the data to a sig-tive inhibition equation, using the GraphpadTM software.moid equation:
1. The latency which was the time (in days) to obtain thefirst doubling (20,000 cells) of the initial cell populationAnalysis of Intracellular Cyclic(10,000 cells) in a given sample.Nucleotides Levels (cAMP and cGMP)
2. The doubling time (DT, in hours), needed to obtain aCells (2.5 3 105) seeded in precoated 24-well dishes were doubling of the cell population in a given well, duringcultured for 3 d, after which time their number was deter- the period of fast growth.mined in three wells. Culture medium was then replaced by270 ml of fresh medium supplemented or not with 30 ml of Cell growth studies were carried out from seeding to con-specified drugs. Cells were incubated at 378C for 20 min, fluency. The growth curves indicated that, in control exper-with or without the specified concentrations of polypep- iments, confluency was reached after about 14 d in the 4
cell lines. At this time, about 500,000 and 800,000 cells/tides, ranging from 10212 to 1026 M. At the end of the incu-
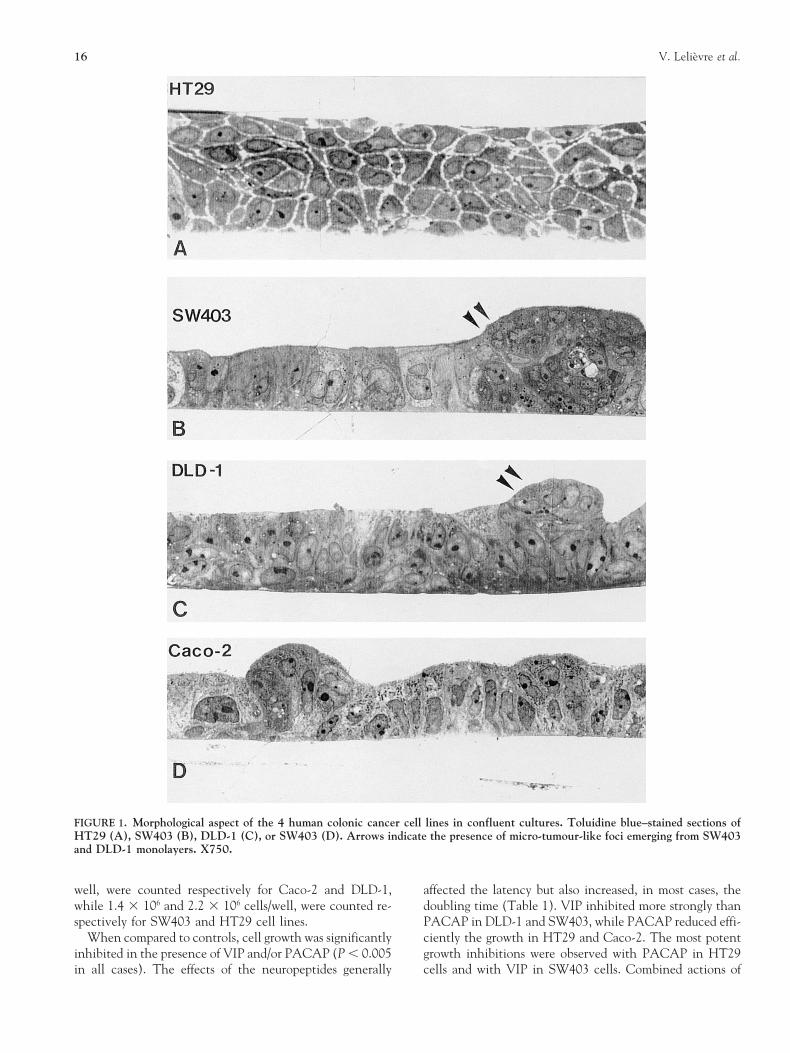
16 V. Lelievre et al.
FIGURE 1. Morphological aspect of the 4 human colonic cancer cell lines in confluent cultures. Toluidine blue–stained sections ofHT29 (A), SW403 (B), DLD-1 (C), or SW403 (D). Arrows indicate the presence of micro-tumour-like foci emerging from SW403and DLD-1 monolayers. X750.
well, were counted respectively for Caco-2 and DLD-1, affected the latency but also increased, in most cases, thedoubling time (Table 1). VIP inhibited more strongly thanwhile 1.4 3 106 and 2.2 3 106 cells/well, were counted re-
spectively for SW403 and HT29 cell lines. PACAP in DLD-1 and SW403, while PACAP reduced effi-ciently the growth in HT29 and Caco-2. The most potentWhen compared to controls, cell growth was significantly
inhibited in the presence of VIP and/or PACAP (P , 0.005 growth inhibitions were observed with PACAP in HT29cells and with VIP in SW403 cells. Combined actions ofin all cases). The effects of the neuropeptides generally

VIP- and PACAP-Mediated Inhibition of Cell Growth 17
FIGURE 2. Aspect of the cell surface of the 4 human colon cancer cell lines in confluent cultures (Scanning electron microscopy).HT29 (A) SW403 (B), DLD-1 (C) and Caco-2 (D) cells. Arrows point to the accumulated microvilli of the adjacent cells underliningthe very regular cell outlines (SW403 and DLD-1 cells). Bar: 5 mM.
Pharmacological Properties of the VIP and PACAPVIP and PACAP resulted in a clear additional effect onlyBinding Sites Expressed in Colonic Cancer Cellsin HT29 cells. In DLD-1, the combined treatment (VIP 1
PACAP) was even less powerful than the action of VIP, Data from these binding experiments (Figs. 4, 5) werecomputed according to either a Hill or a two-site compe-alone. Finally, there is no significant difference between
PACAP and (VIP 1 PACAP) treatments in Caco-2, and titive inhibition equation in order to determine two pa-rameters:between VIP and (VIP 1 PACAP) treatments in DLD-1.

18 V. Lelievre et al.
FIGURE 3. Effects of VIP, PACAP-27 or both (each at 10 nM) on the proliferation of the four colonic carcinoma cell lines: DLD-1 (A),HT29 (B), Caco-2 (C) and SW403 (D). Ten thousand cells were seeded in 24-well dishes and cultured in the absence (X) or in thepresence of VIP (n), PACAP-27 (h) or both polypeptides (s). Cell density was monitored at the indicated times in culture. Data(6SEM) are the mean of two independent triplicate experiments. Proliferation data were fitted according to a sigmoid equation.
1. IC50, the concentration (in nM) of unlabelled polypep- By definition, 125I-PACAP-27 binds to both bivalentVIP/PACAP and PACAP preferring receptors (PACAP1),tide giving half-maximal inhibition of the radiotracer
binding. while 125I-VIP only interacts with the bivalent receptors.Hence the Bmax value obtained using the 125I-PACAP-27 as2. Bmax, the total number of sites/cell interacting with a
given radiotracer. a radiotracer represents the total number of (PACAP1 1

VIP- and PACAP-Mediated Inhibition of Cell Growth 19
TABLE 1. Effects of VIP and/or PACAP-27 (P27) on the proliferationprocesses of four human colonic adenocarcinoma cell lines
Latency Doubling timeCell lines Treatments (hours) (hours)
DLD-1 Control 72 41VIP 10 nM 84** 48**P27 10 nM 84** 43*VIP 1 P27 (10 nM each) 80 48**
HT29 Control 26 25VIP 10 nM 47 27*P27 10 nM 54** 35VIP 1 P27 (10 nM each) 52** 41
Caco-2 Control 96 44VIP 10 nM 108** 48P27 10 nM 108** 56**VIP 1 P27 (10 nM each) 112** 58**
SW403 Control 43 36VIP 10 nM 67** 50**P27 10 nM 60 43VIP 1 P27 (10 nM each) 65** 49**
Latency and Doubling time were obtained from Figure 3. Statistical analysis of the data(ANOVA) indicated the significant differences between the treatments and controls(P , 0.005), except between data pointed by an asterisk and also the significantdifferences between two different treatments (P , 0.005), except between data pointedby a double asterisk.
VIP/PACAP) receptors. The Bmax value obtained using 125I- in HT29 cells, demonstrated that 10 nM VIP or PACAP-27VIP corresponds to the total number of VIP/PACAP recep- caused approximately a 10–15-fold elevation of basal cAMPtors. The Bmax of the PACAP1 receptors is the difference: levels, with a transient peak from 5 to 25 min incubation,(Bmax
125I-PACAP-27 2 Bmax125I-VIP). followed by a slow decay. For this reason, the dose-response
Our results, reported in Tables 2 and 3, indicate that experiments presented here were conducted at the time ofHT29, DLD-1 and SW403 cells preferentially expressed bi- 20 min incubation.valent VIP/PACAP receptors (88, 90 and 94%, respec- In the aggregate, VIP and PACAP were much more effi-tively). In contrast, Caco-2 cells preferentially expressed cient at increasing basal cAMP levels (up to 15-fold inthe PACAP preferring subtype (60%). Another noticeable HT29 cells) than in cGMP (a maximal 1.5-fold increase byobservation was the higher affinity of the 125I-VIP binding VIP was observed in DLD-1 cells). Concerning the effectssites in the HT29 cells (IC50 0.6 nM), when compared with of VIP and PACAP on intracellular cAMP levels, the mostthe other cell lines (IC50 ranging from 2 to 7 nM according sensitive (with EC50 values of about 25 pM) and most po-to the Hill equation). Several displacement curves, how- tent (up to 15-fold stimulation) responses by far to eitherever, strongly diverged from a simple sigmoid equation (Ta- polypeptide were observed in HT29 cells (Fig. 6C). In theseble 2). This was the case for example in SW403 cells (Figs. cells, the (VIP 1 PACAP) treatment shifted the dose-5C, D), where the experimental data were more accurately response curve to the left. Similar results were observed infitted using a double sigmoid equation (describing interac- DLD-1 cells except that they were more sensitive to VIPtion of the ligand with two binding sites), than a single sig- than to PACAP (Fig. 6A). Potent effects of VIP and PACAPmoid equation (Hill equation). Hence, as indicated in Table were also seen in Caco-2 cells, but only for quite elevated2, for 125I-VIP binding in SW403 cells, very high-affinity bind- concentrations of the neuropeptides (EC50 $ 25 nM) (Fig.ing sites (IC50 5 0.4 pM), were distinguished from lower affin- 7A). Finally, in SW403 cells, only PACAP induced anity sites (IC50 5 4 nM) (Fig. 5C, Table 2). In other cases, like increase in cAMP levels, whereas no effect was seen withfor example 125I-PACAP binding to HT29 cells, the data accu- either VIP or (VIP 1 PACAP) (Fig. 7C).rately fitted a single-site sigmoid equation (Fig. 4D) with, how- Cyclic GMP levels were measured in parallel, in the sameever, a Hill number value much lower than 1.
experimental samples as for cAMP analysis. In DLD-1,HT29 and Caco-2 cells, a slight dose-dependent increase
Effects of VIP and PACAP-27 (about 1.5-fold for the maximal response) of intracellularOn Intracellular cAMP and cGMP Levels cGMP was observed in response to VIP but not in response
to PACAP (Figs. 6B, 6D, and 7B). However, the combinedThe effects of VIP, PACAP-27 or of (VIP 1 PACAP-27)use of (PACAP 1 VIP) resulted in a stronger effect thanon intracellular cAMP or cGMP productions were assayedVIP, alone (Figs. 6B, 6D and 7B). Radically opposite effectsfor a time of incubation of 20 min, in dose-response experi-
ments (Figs. 6 and 7). Preliminary time-course experiments were observed in SW403 cells, since either VIP or PACAP

20 V. Lelievre et al.
FIGURE 4. Specific binding of 125I-VIP or 125I-PACAP on DLD-1 (A, B) and HT29 (C, D) cells. Cells were seeded at 2.5 3 105 cells/well in 24-well dishes, and cultured for 3 d. Incubations with radioligands (20–25 pM): 125I-VIP (left panel) or 125I-PACAP (rightpanel) and increasing concentrations of VIP (n) or PACAP-27 (h), were performed at 48C during 180 min. Nonspecific binding wassubtracted for each value. Data are the means (6SEM) of two independent triplicate experiments and were fitted according to eithera Hill equation or a two-site competitive inhibition equation.
decreased (by about 2.5-fold, for the maximal response) including colonic adenocarcinoma cell lines [5, 25]. Fur-thermore, it has been suggested that VIP might be actingbasal cGMP levels (Fig. 7D). These effects were seen forthrough an autocrine/paracrine mechanism in human ma-concentrations as low as 10 pM for VIP. In these experi-lignant cell lines, such as neuroblastoma, pancreatic carci-ments, VIP or (VIP 1 PACAP) had similar efficiency butnoma or non-small cell lung cancer [31–36]. A peculiar sys-were clearly less potent that PACAP, alone.tem is represented by the human colonic adenocarcinomacells Lo-Vo: at concentrations below 100 nM, VIP stimu-
DISCUSSION lates both intracellular cAMP production and proliferation;Cell Morphology and Growth at concentrations over 100 nM, no further increase in theRecent reviews suggested that VIP or PACAP may modu- cAMP levels is observed, whereas VIP appeared to inhibit
cell growth [37]. Some years ago, the group of Laburthelate in vitro, cell growth and differentiation in cancer cells,

VIP- and PACAP-Mediated Inhibition of Cell Growth 21
FIGURE 5. Specific binding of 125I-VIP or 125I-PACAP on Caco-2 (A, B) and SW403 (C, D) cells. Cells were seeded at 2.5 3 105 cells/well in 24-well dishes, and cultured for 3 d. Incubations with radioligands (20–25 pM): 125I-VIP (left panel) or 125I-PACAP (rightpanel) and increasing concentrations of VIP (n) or PACAP-27 (h), were performed at 48C during 180 min. Nonspecific binding wassubtracted for each value. Data are the means (6SEM) of two independent triplicate experiments and were fitted according to eithera Hill equation or a two-site competitive inhibition equation.
demonstrated an increased sensitivity to VIP upon differen- cells [12]. Hence, beside the regulations that they exert ontypical functions of the differentiated colonic epithelium,tiation of the colonic adenocarcinoma cells Caco-2 [38].
This phenomenon was due to an increase in the number of VIP and PACAP may take part in the establishment andmaintenance of the enterocytic phenotype [20]. The aim ofVIP receptors and, consequently, of adenylate cyclase re-
sponse to the peptide. These data were consistent with their the present study was to know whether a correlation couldbe established between the state of differentiation of co-previous demonstration that, in the human colon and rat
small intestine, the concentration of VIP receptors was lonic cancer cells and their response to these neuropeptides,in terms of proliferation, receptor expression, and coupling.more elevated in the mature epithelial cells than in root

22 V. Lelievre et al.
TABLE 2. IC50 and Hill number values of 125I-VIP and 125I-PACAP binding sites in 4 humancolonic adenocarcinoma cell lines
IC50 (nM) IC50 (nM)125I-VIP 125I-PACAPCell Competitor Hill Hill
lines ligands Site 1 Site 2 number Site 1 Site 2 number
DLD-1 VIP 0.24 28 0.55 0.33 25 0.57PACAP-27 19 130 0.71 10 none 0.94
HT29 VIP 0.6 none 0.72 1.4 none 0.47PACAP-27 0.8 none 0.78 2.0 none 0.64
Caco-2 VIP 2.0 none 0.96 8.5 none 0.95PACAP-27 0.47 none 0.81 0.021 4.3 0.42
SW403 VIP 0.0004 4.0 0.78 0.0038 6.0 0.99PACAP-27 0.00045 9 0.93 0.0005 2.7 0.88
Data were obtained from Figures 4 and 5, according to one-site or two-site competition inhibition equations.Computed Hill number corresponded either to a mean value for the overall displacement curves (DLD-1 orCaco-2 cells), or was not taking into consideration the part of the curves corresponding to the very high-affinityinteraction (SW403).
125I-VIP and 125I-PACAP Binding SitesFrom our rough morphological examination of the 4 celllines, it appears that confluent Caco-2, DLD-1 and SW403 The binding experiments revealed that in DLD-1, HT29cells displayed several features of enterocytes (growth in and SW403 cells, 88 to 94% of the radiolabelled VIP ormonolayer, polarised aspect, very limited intercellular PACAP binding sites were represented by polyvalent VIP/spaces, apical microvilli). Without any doubt, Caco-2 PACAP receptors. In contrast, the well-differentiatedmonolayers were those that resembled the more normal co- Caco-2 cells expressed a majority (60%) of PACAP1 recep-lonic mucosa. Microtumourlike foci, in which cells were tors. This raises the interesting issue of a relationship be-growing quite anarchically, were conspicuous in DLD-1 and tween the expression and function of these receptors andSW403 cells. In comparison, HT29 cells appeared poorly the state of differentiation of Caco-2 cells, which closely re-differentiated. lated to the normal colonic epithelial cells (when compared
Daily addition of VIP or PACAP to the culture medium to the others herein studied). When 125I-VIP was used as ainhibited cell proliferation in the 4 cell lines. However, the radiotracer, the displacement curves diverged from a simplemost obvious effects were observed in HT29 and SW403 sigmoid equation, except in Caco-2 cells. This reflects ei-cells which had, by far, the fastest growth rates, yielding to ther a complex interaction on one category of binding sites,the highest saturation densities. In all cell lines, the con- or the existence of at least two categories of binding sites.centration used for each peptide was not sufficient to induce This later possibility gained support from the observationthe maximal effect (data not shown). However, clear addi- that the data fitted well a two-sites model—one of very hightive effects of VIP and PACAP were seen only in HT29 affinity (IC50) and one of lower affinity—in the DLD-1cells, suggesting that these neuropeptides might influence and SW403 cells. The usual IC50 for 125I-VIP is the nano-proliferation through at least partly distinct mechanisms in molar range. It would be interesting to investigate thethis cell line. In contrast, the absence of additive effect in nature of the putative very high-affinity VIP/PACAP bind-the other cell lines might reflect reciprocal antagonistic ing sites exhibiting IC50 values in the picomolar range. It isproperties of VIP and PACAP on cell growth inhibition. noteworthy that subnanomolar concentrations of VIP can
trigger mitogenic effects, neuronal survival and release ofThis is particularly obvious in DLD-1 cells.
TABLE 3. Bmax values and representativeness of VIP and PACAP bind-ing sites in 4 human colonic adenocarcinoma cell lines
BMAX (sites per cell) Representativity (%)
Cell lines PACAP VIP/PACAP PACAP VIP/PACAP
DLD-1 1900 21000 10 90HT29 1820 22000 12 88Caco-2 13000 9000 60 40SW403 1350 17000 6 94
Data were obtained from Figures 4 and 5. PACAP: PACAP-preferring receptors.VIP/PACAP: bivalent receptors with high affinity for VIP and PACAP.

VIP- and PACAP-Mediated Inhibition of Cell Growth 23
FIGURE 6. Dose-dependent effects of VIP or PACAP-27 on intracellular cAMP or cGMP levels in DLD-1 (A, B) and HT29 (C, D)cells. Cells monolayers in 24-well dishes were incubated for 20 min at 378C, in the presence of the indicated concentrations of VIP(n) or PACAP-27 (h), or both polypeptides (s). Intracellular cAMP (left panel) and cGMP (right panel) contents were determinedusing commercially available RIA kits. Data are the means (6SEM) of two independent triplicate experiments.
glial neurotrophic substances in cultured sympathic neuro- 1. DLD-1 and HT29 cells were very sensitive to VIP andPACAP. The fact that VIP and PACAP had more thanblasts [39].additive effects when used together at submaximal con-centrations suggests that VIP/PACAP and PACAP1 re-
Modulation of Cyclic AMP Levels by VIP and PACAP ceptors were present (as suggested from the binding ex-The cell lines could be classified into three different types periments; see above) and were acting in a cooperative
manner.as a function of their response to the neuropeptides:

24 V. Lelievre et al.
FIGURE 7. Dose-dependent effects of VIP or PACAP-27 on intracellular cAMP or cGMP levels in Caco-2 (A, B) and SW403 (C,D) cells. Cells monolayers in 24-well dishes were incubated for 20 min at 378C, in the presence of the indicated concentrations of VIP(n) or PACAP-27 (h), or both polypeptides (s). Intracellular cAMP (left panel) and cGMP (right panel) contents were determinedusing commercially available RIA kits. Data are the mean (6SEM) of two independent triplicate experiments.
2. Caco-2 cells were sensitive only to high concentrations the pre-confluent stage of the culture, as well as efficientreceptor/adenylate-cyclase coupling only in differenti-of VIP or PACAP. Because the total number of binding
sites for 125I-VIP and 125I-PACAP was similar in all cell ated Caco-2 cells (i.e., post-confluent culture) [38, 40].3. SW403 responded only to PACAP while a combinationlines, it is suggested that, under our experimental condi-
tions—i.e., pre-confluent cultures—(i) phosphodiester- of VIP and PACAP triggered no elevation of the cAMPcontents. There are at least two possible interpretationsase activity was high and/or, (ii) the receptors involved
were poorly coupled to the adenylate cyclase. This corre- of such data: (i) VIP and PACAP may interact with a bi-valent VIP/PACAP receptor, PACAP behaving like anlates well with previous data obtained in this cell line,
reporting a surge in phosphodiesterase activity during agonist and VIP like an antagonist towards this binding

VIP- and PACAP-Mediated Inhibition of Cell Growth 25
site; (ii) the PACAP stimulation of cAMP production growth with equal efficiency in two fast growing cell lines:HT29 and SW403. In the HT29 cell line, which appearswas triggered through PACAP-preferring receptors, this
response being counteracted through binding of VIP to poorly differentiated, VIP triggered very potent and sensi-tive stimulation of cAMP production and a discrete in-a VIP/PACAP receptor. This latter hypothesis appears,
however, quite unlikely considering the very low repre- crease of cGMP levels. In the SW403 cells that display mor-phological properties of enterocytes, VIP did not elevatesentativeness (6%) of the PACAP1 receptors in this cell
line. cAMP levels but induced a significant decrease of basalcGMP concentration. These unexpected findings could beinterpreted if one considers that an as yet unidentified sub-
Modulation of Cyclic GMP Levels by VIP and PACAP class of VIP/PACAP receptors, which may not be classicallycoupled to G proteins, is required for the regulation of cellConcurrently with cAMP analysis, cGMP levels were deter-growth. Expression of these receptors in the cell types con-mined in the same experimental samples. The data on regu-sidered here may also explain the multiple sites displace-lation of intracellular cGMP in response to VIP or PACAP,ment curves, observed in our binding experiments, usingrevealed two types of cell behavior:125I-VIP radiotracer. In this respect, the observation that
1. In Caco-2, DLD-1 and HT29 cells, VIP—but not PA- high-affinity receptors that are not coupled to G proteinsCAP—caused a slight dose-dependent increase of intra- might mediate the mitogenic actions of VIP during braincellular cGMP. In a combined use (VIP 1 PACAP), a development, deserves to be cited [42]. These binding sitespotentialisation of the effects of VIP and PACAP was may or may not represent a distinct molecular species, sincenoticed, that may be mediated through PACAP1 recep- they could represent differential states of functional cou-tors. As a matter of fact, this potentialisation was partic- pling of the same receptor to various transduction mecha-ularly conspicuous in Caco-2 cells where an important nisms. In a recent report, we demonstrated that expressionrepresentativeness (60%) of the PACAP1 receptors was of the mRNAs of VIP and PACAP receptor subtypes wasdemonstrated. reduced, while VIP and PACAP binding increased, during
2. In SW403 cells, both VIP or PACAP, triggered a de- retinoic acid-induced differentiation of IMR-32 humancrease in basal cGMP levels, PACAP being more effi- neuroblastoma cells. Therefore, we concluded that such dis-cient. A combination of both had the same effect as VIP, crepancy could be due either to expression of an as yet notalone. Such a phenomenon suggests that VIP may antag- cloned VIP/PACAP receptor or to the coupling of receptorsonize PACAP, possibly through a process of reciprocal to different G proteins, resulting in modified binding kinet-exclusion from a common VIP/PACAP receptor. Fur- ics [43]. To investigate these issues further, analysis of thethermore, this interpretation correlates the above pro- expression of G protein components in these cell linesposed hypothesis of an antagonistic effect of VIP on the could be helpful. Another attractive approach should be toPACAP-induced stimulation of cAMP production in check the proliferative effects of VIP and PACAP in cellsSW403 cells. transfected with dominant negative mutant genes encoding
various components of different signal transduction pathways.
CONCLUSIONS We thank F. Chevalier for her very helpful technical assistance. Part of thiswork was supported by a grant from ‘Fondation Langlois, France’.
In the aggregate, analysis of the effects of VIP and PACAPon cyclic nucleotide levels reveal that differential coupling
Referencesof the PACAP1 or of the polyvalent VIP/PACAP recep-1. Said S. I. (1991) Trends Endocrinol. Metab. 3, 107–112.tors, may occur from a colonic adenocarcinoma cell line to 2. Hakanson R., Sundler F. and Uddman R. (1982) Distribution
another. Hence the complex binding kinetics obtained in and topography of peripheral VIP nerve fibers: Functional im-these cell lines, using 125I-VIP as a radiotracer, may reflect plications. In: Vasoactive intestinal peptide, Advances in Peptide
Hormone Research Series (Said S. I. ed.), pp. 121–144. Raventhe different functional states of the VIP/PACAP receptors.Press, New York.Such data can be brought together with those of a recent re-
3. Chayvialle J. A., Miyata M., Descos F., Rayford P. L. andport from our group, where switches in the expression and Thompson J. C. (1982) Physiological observations on vaso-function of PACAP and VIP receptors were observed, along active intestinal peptide in digestive tract. In: Vasoactive intes-
tinal peptide, Advances in Peptide Hormone Research Series (Saidthe phenotypic interconversion of human neuroblastomaS. I. ed.). pp. 447–456. Raven Press, New York.cells. Fibroblastlike cells expressing mainly PACAP-prefer-
4. Miyata A., Arimura A., Dahl D. H., Minamino N., Uehararing receptors that triggered stimulation of intracellularA., Jiang L., Culler M. D. and Coy D. H. (1989) Biochem. Bio-
cGMP production, spontaneously converted into neuro- phys. Res. Com. 164, 567–574.blastlike cells that mostly possessed VIP/PACAP receptors 5. Rawlings S. R. and Hezareh M. (1996) Endocrine Rev. 17,
4–29.positively coupled to adenylate-cyclase [41]. No such obvi-6. Christophe J. (1993) Biochim. Biophys. Acta 1154, 183–199.ous relationship between the functional coupling of the re-7. Ishihara T., Shigemoto R., Mori K., Takahashi K. and Nagataceptors and the growth or differentiation properties of the S. (1992) Neuron 8, 811–819.
colonic malignant cell lines considered here, could be ob- 8. Lutz E. M., Sheward W. J., West K. M., Morrow J. A., FinkG. and Harmer A. J. (1993) FEBS Lett. 334, 3–8.served in the present study. For example, VIP inhibited cell

26 V. Lelievre et al.
9. Spengler D., Waeber C., Pantaloni C., Holsboer F., Bockaert 25. Muller J. M., Lelievre V., Becq-Giraudon L. and MeunierA. C. (1995) Mol. Neurobiol. 10, 115–134.J., Seeburg P. H. and Journot L. (1993) Nature 365, 170–175.
26. Gozes I. and Brenneman D. E. (1993) J. Mol. Neurosci. 4, 1–9.10. Okamoto T., Murayama T., Hayashi Y., Inagaki M., Ogata E.27. Bellot F., Pichon J., Luis J., El Battari A., Secchi J., Cau P. andand Nishimoto I. (1991) Cell 67, 723–730.
Marvaldi J. (1985) Int. J. Cancer. 36, 609–615.11. Fogh J. and Trempe G. (1975) Human tumor cells in vitro. In:28. Martin J. L., Rose K., Hughes G. J. and Magistretti P. J. (1986)Human Tumor Cells ‘‘in Vitro’’ (Fogh J. ed.) pp. 115–141. Ple-
J. Biol. Chem. 261, 5320–5327.num Publishing Corp., New York.29. Muller J.-M., Luis J., Fantini J., Abadie B., Giannellini F.,12. Zweibaum A., Laburthe M., Grasset E. and Louvard D. (1991)
Marvaldi J. and Pinchon J. (1985) Eur. J. Biochem. 151, 411–In: Use of cultured cell lines of intestinal cell differentiation and417.function (Field M. and Frizzel R. A. eds.). Handbook of Physi-
30. Pinto M., Robine-Leon S., Appay M.-D., Kedinger M., Tri-ology, The Gastrointestinal System, Sec. 6, Chapt. 7, pp. adou N., Dussaulx E., Lacroix B., Simon-Assman P., Haffen223–255. Am. Physiol. Soc., Washington, DC. K., Fogh J. and Zweibaum A. (1983) Biol. Cell 47, 323–330.13. Laburthe M. and Amiranoff B. (1989) In: Peptide receptors in 31. Muller J. M., Lolait S. J., Yu V. C., Sadee W. and Waschek
the intestinal epithelium. (Makhlouf G. ed.), Handbook of Phys- J. A. (1989) J. Biol. Chem. 264, 3647–3650.iology, the Gastrointestinal System, Neural and Endocrine 32. Buscail L., Cambillau C., Seva C., Scehama J.-C., de Neef P.,Biology, Sect. 6, Vol. II, Chapt. 11, pp. 215–244. Am Physiol. Robberecht P., Christophe J., Susini C. and Vaysse N. (1992)Soc., Washington, DC. Gastroenterol. 103, 1002–1008.
14. Augeron C. and Laboisse C. (1984) Cancer Res. 44, 3961– 33. O’Dorisio M. S., Fleshman D. J., Qualman S. J. and O’Dorisio3969. T. M. (1992) Regul. Pept. 37, 213–226.
15. Pinto M., Robine-Leon S., Appay M. D., Kedinger M., Tri- 34. Moody T. W., Zia F., Draoui M., Brenneman D. E., Fridkinadou N., Dussaulx E., Lacroix B., Simon-Assmann P., Haffen M., Davidson A. and Gozes I. (1993) Proc. Natl. Acad. Sci.
USA 90, 4345–4349.K., Fogh J. and Zweibaum A. (1983) Biol. Cell 47, 323–330.35. Pence J. C. and Shorter N. A. (1993) Arch. Surg. 128, 591–16. Dharmsathaphorn K., McRoberts J. A., Mandel K. G., Tisdale
595.L. D. and Masui H. (1984) Am. J. Physiol. 246, G204–G208.36. Suzuki N., Harada M., Hosoya M. and Fujino M. (1994) Eur.17. Laburthe M., Rousset M., Chevalier G., Boissard C., Dupont
J. Biochem. 223, 147–153.A., Zweibaum A. and Rosselin G. (1980) Cancer Res. 40,37. Yu D., Seitz P. K., Selvanayagam P., Rajaraman S., Townsend2529–2533.
C. M. Jr. and Cooper C. W. (1992) Endocrinol. 131, 1188–18. Waschek J. A. (1995) Cancer Lett. 92, 143–149.1194.19. Grasset E., Bernabeu J. and Pinto M. (1984) Am. J. Physiol.
38. Laburthe M., Rousset M., Rouyer-Fessard A., Couvineau A.,248, C410–418.Chevalier G. and Zweibaum A. (1987) J. Biol. Chem. 262,20. Nguyen T. D., Heintz G. G. and Cohn J. A. (1992) Gastroent- 10180–10184.
erol. 103, 539–544. 39. Pincus D. W., DiCicco-Bloom E. M. and Black I. B. Nature21. Hoosein N. M., Black B. E. and Brattain D. E. (1989) Neuron (1990) 343, 564–567.
8, 811–819. 40. Savini F., Brardi S., Tatone D. and Spoto G. (1995) Life Sci.22. Chelbi-Alix M. K., Boissard C., Sripati C. E., Rosselin G. and 56, 421–425.
Thang M. N. (1992) Peptides 12, 1085–1093. 41. Lelievre V., Becq-Giraudon L., Meunier A.-C. and Muller23. Gamet L., Murat J-C., Remaury A., Remesy C., Valet P., Paris J.-M. (1996) Neuropeptides 30, 313–322.
H. and Denis-Pouxviel C. (1992) J. Cell. Physiol. 150, 501– 42. Gressens P., Hill J. M., Gozes I., Fridkin M. and Brenneman509. D. E. (1993) Nature 362, 155–158.
24. Bjork J., Nilsson J., Hultcrantz R. and Johansson C. (1993) 43. Waschek J. A., Lelievre V., Bravo D. T., Nguyen T. andMuller J.-M. (1997) Peptides, in press.Scand. J. Gastroenterol. 28, 879–884.




















