
Activation of the IGF-IR system contributes to malignant growth of human and mouse medulloblastomas
12
Activation of the IGF-IR system contributes to malignant growth of human and mouse medulloblastomas Jin Ying Wang 1 , Luis Del Valle 1 , Jennifer Gordon 1 , Michele Rubini 3 , Gaetano Romano 2 , Sidney Croul 1 , Francesca Peruzzi 2 , Kamel Khalili 1 and Krzysztof Reiss* ,1 1 Center for Neurovirology and Cancer Biology, College of Science and Technology, Temple University, 1900 North 12th Street, Biology Life Science Building, Room 238 Philadelphia, Pennsylvania, PA 19122, USA; 2 Kimmel Cancer Center, Thomas Jeerson University, 233 S 10th Street, 624 BLSB Philadelphia, Pennsylvania, PA 19107, USA; 3 Universita ’Degli Studi di Ferrara Dipartimento di Medicina Sperimentale e Diagnostica, Via L. Borsari, 46-44100 Ferrara, Italy Insulin-like growth factor I receptor (IGF-IR) has been implicated in the normal and malignant growth of many cell types including cells from the central nervous system. In the cerebellar cortex IGF-IR mRNA is found in granular cells and IGF-I stimulation is mitogenic and protects cells from low-potassium-induced apoptosis. Since primitive neuroectodermal tumors/medulloblasto- mas (PNETs/medulloblastomas) are suspected to origi- nate from the external cerebellar granular layer, it is reasonable to postulate that IGF-IR and/or its signaling molecules may contribute to the transformation of these poorly dierentiated cells. To study activation of the IGF-IR system in medulloblastomas, we have utilized an antibody (anti-pY1316) that specifically recognizes the phosphorylated (active) form of the IGF-IR. Medullo- blastoma biopsy specimens were positive when examined immunohistochemically with anti-Y1316 antibody. Further analysis of the IGF-IR system was performed in three human (Daoy, TE-671, D283 Med) and four mouse (BsB8, BsB13, Bs-1b, Bs-1c) medulloblastoma cell lines. All the murine cell lines examined express IGF-IR and PI3-kinase at relatively normal levels, and grossly overexpress IRS-1, when compared with normal mouse cerebellum. Within 15 min following IGF-I stimulation both mouse and human cell lines phosphorylate the b subunit of the IGF-IR, IRS-1, Akt, and MAP kinases. They respond with cell proliferation when stimulated solely with IGF-I and are strongly inhibited when challenged with a dominant negative mutant of the IGF-IR (486/STOP), or with antisense oligonucleotides against the IGF-IR mRNA. Oncogene (2001) 20, 3857 – 3868. Keywords: IGF-IR; medulloblastoma; IRS-1; cell pro- liferation; tyrosine phosphorylation Introduction The type I insulin-like growth factor receptor (IGF-IR) is a multifunctional tyrosine kinase receptor that regulates cellular growth in at least four dierent ways: (i) it is mitogenic (Baserga, 1994; Reiss et al., 1998b); (ii) it protects normal and tumor cells from apoptosis (Gluckman et al., 1992; Sell et al., 1995); (iii) in certain circumstances it may control neuronal dierentiation (Morrione et al., 2000); and (iv) it plays an important role in transformation by several viral and cellular oncogenes, and in the maintenance of the transformed phenotype (Baserga, 1995; Baserga et al., 1997). The requirement for a functional IGF-IR in malignant transformation has been demonstrated by the use of R 7 cells, which are 3T3-like cells isolated from mouse embryos with a targeted disruption of the IGF-IR genes (Baker et al., 1993; Liu et al., 1993). Without IGF-IR, R 7 cells cannot be transformed by a number of genetic manipulations that easily transform the parental 3T3 cells. These include SV40 T antigen, activated Ha-ras oncogene (Sell et al., 1993, 1994), bovine papilloma virus E5 protein (Morrione et al., 1995), human papilloma virus (Steller et al., 1996), activated c-src (Valentinis et al., 1997), and over- expressed growth factor receptors, including EGF (Coppola et al., 1994), PDGF (DeAngelis et al., 1995) and insulin (Miura et al., 1995) receptors. In agreement with these findings, antibodies against the IGF-IR (Kalebic et al., 1994), antisense expression plasmids (Trojan et al., 1992, 1993; Resnico et al., 1994; Shapiro et al., 1994), and dominant negative mutants of the IGF-IR (Prager et al., 1994; Burgaud et al., 1995; Reiss et al., 1998a, 1999) can attenuate malignant tumor growth by inducing apoptosis, and/or by inhibiting cell proliferation. In the central nervous system (CNS), IGF-IR has been implicated in fetal and postnatal growth and development of the brain (LeRoith et al., 1993) as well as in brain tumors. The presence of an active IGF system has been shown in astrocytomas (Antoniades et al., 1992), glioblastomas (Trojan et al., 1992, 1993; Resnico et al., 1994), meningiomas (Antoniades et al., 1992), and neuroblastoma cells (Martin et al., 1992). Oncogene (2001) 20, 3857 – 3868 ª 2001 Nature Publishing Group All rights reserved 0950 – 9232/01 $15.00 www.nature.com/onc *Correspondence: K Reiss Received 13 December 2000; revised 3 April 2001; accepted 9 April 2001
Transcript of Activation of the IGF-IR system contributes to malignant growth of human and mouse medulloblastomas

Activation of the IGF-IR system contributes to malignant growth of humanand mouse medulloblastomas
Jin Ying Wang1, Luis Del Valle1, Jennifer Gordon1, Michele Rubini3, Gaetano Romano2,Sidney Croul1, Francesca Peruzzi2, Kamel Khalili1 and Krzysztof Reiss*,1
1Center for Neurovirology and Cancer Biology, College of Science and Technology, Temple University, 1900 North 12th Street,Biology Life Science Building, Room 238 Philadelphia, Pennsylvania, PA 19122, USA; 2Kimmel Cancer Center, Thomas Je�ersonUniversity, 233 S 10th Street, 624 BLSB Philadelphia, Pennsylvania, PA 19107, USA; 3Universita 'Degli Studi di FerraraDipartimento di Medicina Sperimentale e Diagnostica, Via L. Borsari, 46-44100 Ferrara, Italy
Insulin-like growth factor I receptor (IGF-IR) has beenimplicated in the normal and malignant growth of manycell types including cells from the central nervous system.In the cerebellar cortex IGF-IR mRNA is found ingranular cells and IGF-I stimulation is mitogenic andprotects cells from low-potassium-induced apoptosis.Since primitive neuroectodermal tumors/medulloblasto-mas (PNETs/medulloblastomas) are suspected to origi-nate from the external cerebellar granular layer, it isreasonable to postulate that IGF-IR and/or its signalingmolecules may contribute to the transformation of thesepoorly di�erentiated cells. To study activation of theIGF-IR system in medulloblastomas, we have utilized anantibody (anti-pY1316) that speci®cally recognizes thephosphorylated (active) form of the IGF-IR. Medullo-blastoma biopsy specimens were positive when examinedimmunohistochemically with anti-Y1316 antibody.Further analysis of the IGF-IR system was performedin three human (Daoy, TE-671, D283 Med) and fourmouse (BsB8, BsB13, Bs-1b, Bs-1c) medulloblastoma celllines. All the murine cell lines examined express IGF-IRand PI3-kinase at relatively normal levels, and grosslyoverexpress IRS-1, when compared with normal mousecerebellum. Within 15 min following IGF-I stimulationboth mouse and human cell lines phosphorylate the bsubunit of the IGF-IR, IRS-1, Akt, and MAP kinases.They respond with cell proliferation when stimulatedsolely with IGF-I and are strongly inhibited whenchallenged with a dominant negative mutant of theIGF-IR (486/STOP), or with antisense oligonucleotidesagainst the IGF-IR mRNA. Oncogene (2001) 20,3857 ± 3868.
Keywords: IGF-IR; medulloblastoma; IRS-1; cell pro-liferation; tyrosine phosphorylation
Introduction
The type I insulin-like growth factor receptor (IGF-IR)is a multifunctional tyrosine kinase receptor thatregulates cellular growth in at least four di�erentways: (i) it is mitogenic (Baserga, 1994; Reiss et al.,1998b); (ii) it protects normal and tumor cells fromapoptosis (Gluckman et al., 1992; Sell et al., 1995); (iii)in certain circumstances it may control neuronaldi�erentiation (Morrione et al., 2000); and (iv) it playsan important role in transformation by several viraland cellular oncogenes, and in the maintenance of thetransformed phenotype (Baserga, 1995; Baserga et al.,1997). The requirement for a functional IGF-IR inmalignant transformation has been demonstrated bythe use of R7 cells, which are 3T3-like cells isolatedfrom mouse embryos with a targeted disruption of theIGF-IR genes (Baker et al., 1993; Liu et al., 1993).Without IGF-IR, R7 cells cannot be transformed by anumber of genetic manipulations that easily transformthe parental 3T3 cells. These include SV40 T antigen,activated Ha-ras oncogene (Sell et al., 1993, 1994),bovine papilloma virus E5 protein (Morrione et al.,1995), human papilloma virus (Steller et al., 1996),activated c-src (Valentinis et al., 1997), and over-expressed growth factor receptors, including EGF(Coppola et al., 1994), PDGF (DeAngelis et al.,1995) and insulin (Miura et al., 1995) receptors. Inagreement with these ®ndings, antibodies against theIGF-IR (Kalebic et al., 1994), antisense expressionplasmids (Trojan et al., 1992, 1993; Resnico� et al.,1994; Shapiro et al., 1994), and dominant negativemutants of the IGF-IR (Prager et al., 1994; Burgaud etal., 1995; Reiss et al., 1998a, 1999) can attenuatemalignant tumor growth by inducing apoptosis, and/orby inhibiting cell proliferation.
In the central nervous system (CNS), IGF-IR hasbeen implicated in fetal and postnatal growth anddevelopment of the brain (LeRoith et al., 1993) as wellas in brain tumors. The presence of an active IGFsystem has been shown in astrocytomas (Antoniades etal., 1992), glioblastomas (Trojan et al., 1992, 1993;Resnico� et al., 1994), meningiomas (Antoniades et al.,1992), and neuroblastoma cells (Martin et al., 1992).
Oncogene (2001) 20, 3857 ± 3868ã 2001 Nature Publishing Group All rights reserved 0950 ± 9232/01 $15.00
www.nature.com/onc
*Correspondence: K ReissReceived 13 December 2000; revised 3 April 2001; accepted 9 April2001

Little is known about the role of the IGF-IR systemduring development and progression of medulloblasto-mas, which represent about 25% of all pediatric braintumors. Recently, Patti et al. (2000) have found that anautocrine IGF-I loop activates proliferation of theDaoy human medulloblastoma cell line, suggesting thatthe IGF-IR system may be involved in the develop-ment and/or progression of these cerebellar tumors. Toaddress this issue in more detail we propose to: (i)characterize the IGF-IR system in a new transgenicmouse model of medulloblastoma; (ii) determinewhether any of the IGF-IR signaling components arealtered in mouse and human medulloblastomas incomparison with developmentally matched normalcerebellar tissue; and ®nally (iii) correlate activity ofthe IGF-IR in medulloblastoma cell lines in vitro withIGF-IR phosphorylation in medulloblastoma biopsies.
We have found that medulloblastoma biopsiesdemonstrated cells with the phosphorylated (active)form of the IGF-IR, as shown by anti-pY1316antibody staining (Rubini et al., 1999). Additionally,we examined four mouse (Bs-1b, Bs-1c, BsB8, BsB13)and three human (Daoy, TE-671, D283 Med) medullo-blastoma cell lines that are all positive for neuronalmarkers such as synaptophysin, class III b-tubulin andneuro®lament proteins, and are negative for glial®brilliary acidic protein (GFAP). The cell lines selectedpossess the major components of the IGF-IR system,and grossly overexpress IRS-1. They responded withproliferation when stimulated solely with IGF-I, andwere strongly inhibited when challenged with either adominant negative mutant of the IGF-IR or antisenseoligonucleotides against IGF-IR mRNA. Importantly,the mouse medulloblastoma cell lines characterized inthis study can be utilized in a syngenic animal model tostudy the e�cacy of anti-IGF-IR strategies directedagainst medulloblastoma cells in vivo.
Results
Activity of the IGF-IR system in biopsies from patientswith medulloblastoma
Recently an antibody against the phosphorylated(active) form of the IGF-IR has been developed andcharacterized by Rubini et al. (1999). This antibodyspeci®cally recognizes phosphotyrosine 1316 (pY1316)of the b subunit of IGF-IR, and does not cross-reactwith the corresponding moiety of the activated insulinreceptor (Rubini et al., 1999). To determine whetherIGF-IR is activated in human medulloblastomas invivo, para�n sections of medulloblastoma biopsieswere stained with the anti-pY1316 antibody. Figure1c (low magni®cation) and Figure 1e (high magni®ca-tion) shows groups of tumor cells in a representativebiopsy, which were strongly positive for anti-pY1316by immunohistochemistry. A facing section was probedwith an antibody that recognizes total IGF-IR (Figure1d,f) and cells within this section were uniformlystained. A section of normal human cerebellum, used
here as a reference tissue, was completely negative forthe anti-pY1316 antibody staining (Figure 1a) andweakly positive for the total IGF-IR immuno-staining(Figure 1c). Note that in normal human cerebellumPurkinje cells were apparently positive for the totalIGF-IR immuno-staining, cells from granular layerwere weakly positive, and cells from the molecularlayer were negative (Figure 1b). These results suggestthat while the IGF-IR protein is present in themajority of tumor cells, the phosphorylated form ofthe IGF-IR is not omnipresent since it was detectedonly in certain areas within the tumor and only someof these cells showed strong anti-pY1316 reactivity(compare Figure 1c and d). Table 1 gives thedescription of medulloblastoma biopsies examined inthis study. Six cases including one classic and ®vedesmoplastic medulloblastomas were characterized inrespect to: gender and age of the patient; location ofthe tumor; presence (+) or absence (7) of conven-tional neuronal (SY-38 and NF) and glial (GFAP)markers; and ®nally presence (+) or absence (7) ofanti-IGF-IR and anti-pY1316 antibody staining. Cellsin all six medulloblastoma cases examined werepositive for the IGF-IR, which showed uniformstaining across the sections (see also Figure 1d). Theanti-pY1316 antibody staining, although also presentin all six biopsies examined, was not as uniform asdetected for total IGF-IR antibody staining, showingareas within the tumor tissue where groups of cellswere strongly positive. It has to be emphasizedhowever, that some areas within the positive sectionwere completely negative for the anti-pY1316 antibodystaining, implying the presence of a local auto- and/orpara-ctine IGF system in these tumors. Estimatedpercentage of cells positive for the anti-pY1316 anti-body staining is given in parentheses (Table 1).
Detection of activated IGF-IR in medulloblastoma celllines
As demonstrated in Figure 1 human medulloblastomasshowed reactivity with anti-pY1316 antibody, suggest-ing continuous activation of the IGF-IR in thesetumors. This antibody was also utilized in vitro tofurther characterize its speci®city and to analysephosphorylation of the IGF-IR in medulloblastomacell lines following IGF-I stimulation. Figure 2 showsthat the immuno-reactivity is associated with IGF-Istimulation. Mouse and human medulloblastoma celllines were cultured in serum-free medium for 48 h andthe quiescent cells were subsequently stimulated withIGF-I (50 ng/ml) for 15 min (Figure 2a,c,e,g,i,k), orwere left without stimulation (Figure 2b,d,f,h,j,l). Allcell lines stimulated with IGF-I were characterized bystrong granular staining. In contrast, quiescent mouseBs-1b, Bs-1c, BsB8 cells (Figure 2b,d,f) and humanD283 Med cells (Figure 2j), tested with the anti-pY1316 antibody showed much weaker di�use stain-ing, similar to the background staining obtained whenthe primary antibody was omitted from the reaction(Figure 2i picture B). These results indicate that anti-
Activity of the IGF-IR system in medulloblastomasJY Wang et al
3858
Oncogene
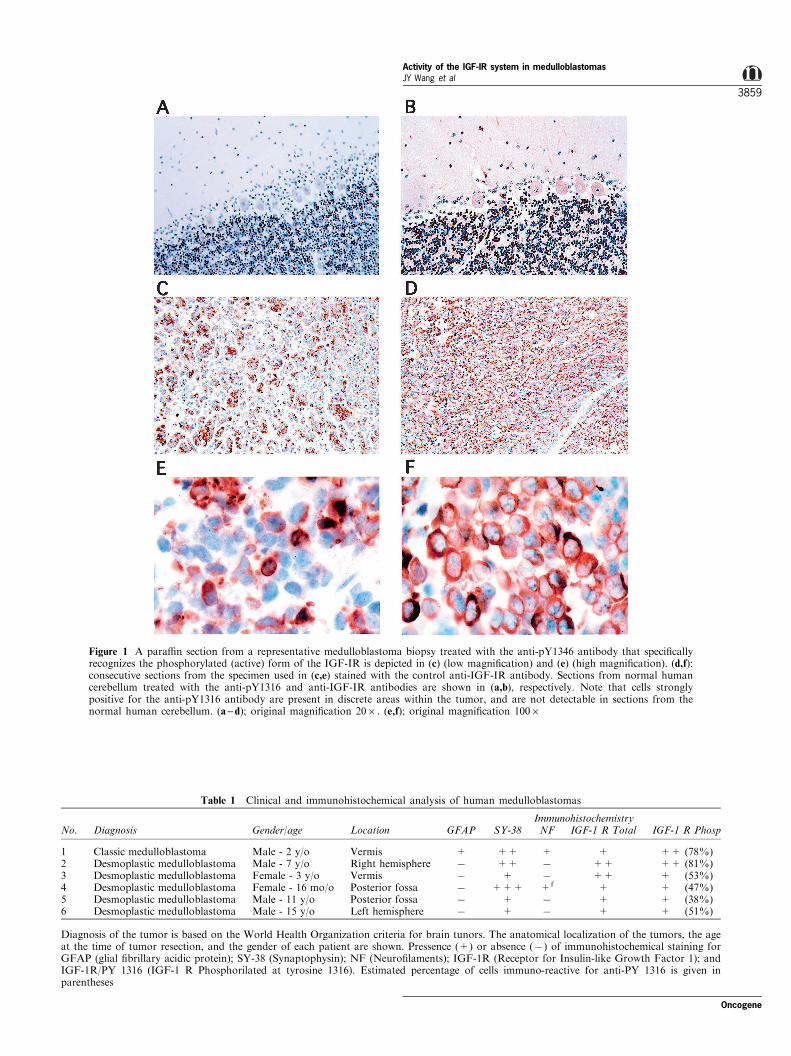
Figure 1 A para�n section from a representative medulloblastoma biopsy treated with the anti-pY1346 antibody that speci®callyrecognizes the phosphorylated (active) form of the IGF-IR is depicted in (c) (low magni®cation) and (e) (high magni®cation). (d,f):consecutive sections from the specimen used in (c,e) stained with the control anti-IGF-IR antibody. Sections from normal humancerebellum treated with the anti-pY1316 and anti-IGF-IR antibodies are shown in (a,b), respectively. Note that cells stronglypositive for the anti-pY1316 antibody are present in discrete areas within the tumor, and are not detectable in sections from thenormal human cerebellum. (a ± d); original magni®cation 206. (e,f); original magni®cation 1006
Table 1 Clinical and immunohistochemical analysis of human medulloblastomas
ImmunohistochemistryNo. Diagnosis Gender/age Location GFAP SY-38 NF IGF-1 R Total IGF-1 R Phosp
1 Classic medulloblastoma Male - 2 y/o Vermis + ++ + + ++ (78%)2 Desmoplastic medulloblastoma Male - 7 y/o Right hemisphere 7 ++ 7 ++ ++ (81%)3 Desmoplastic medulloblastoma Female - 3 y/o Vermis 7 + 7 ++ + (53%)4 Desmoplastic medulloblastoma Female - 16 mo/o Posterior fossa 7 +++ +f + + (47%)5 Desmoplastic medulloblastoma Male - 11 y/o Posterior fossa 7 + 7 + + (38%)6 Desmoplastic medulloblastoma Male - 15 y/o Left hemisphere 7 + 7 + + (51%)
Diagnosis of the tumor is based on the World Health Organization criteria for brain tunors. The anatomical localization of the tumors, the ageat the time of tumor resection, and the gender of each patient are shown. Pressence (+) or absence (7) of immunohistochemical staining forGFAP (glial ®brillary acidic protein); SY-38 (Synaptophysin); NF (Neuro®laments); IGF-1R (Receptor for Insulin-like Growth Factor 1); andIGF-1R/PY 1316 (IGF-1 R Phosphorilated at tyrosine 1316). Estimated percentage of cells immuno-reactive for anti-PY 1316 is given inparentheses
Oncogene
Activity of the IGF-IR system in medulloblastomasJY Wang et al
3859

pY1316 antibody staining is associated with IGF-Istimulation. TE-671 and Daoy cells were also positivefor the phosphorylated form of IGF-IR whenstimulated with IGF-I (Figure 2g,k). However, thedi�erence in staining intensity between cells stimulated(Figure 2g,k), or not stimulated with IGF-I (Figure2h,l) was not as apparent as in previous examples. Thismay indicate that the IGF-IR is continuously phos-phorylated in TE-671 and Daoy cells independently ofIGF-I stimulation. Insets located in the lower rightcorners show synaptophysin immuno-staining of corre-sponding cell lines (Krynska et al., 1999).
Levels of IGF-IR, IRS-1, and PI3-K in medulloblastomacell lines
Proceeding from the ®nding that IGF-IR is activated inmedulloblastoma tissues and cell lines, we looked forpotential abnormalities within the IGF-IR system.Normal cerebellar tissues isolated from mice age-matched to those from which the medulloblastomacell lines originated, were utilized as reference samples.Thick frozen sections of normal human cerebellumcollected from autopsies of 11 weeks (N906), 26 weeks(N1061) children, and 36 years old (N989) individuals,were used as reference samples for the humanmedulloblastoma cell lines. As shown in Figure 3a,b,IGF-IR proteins were present in all mouse and humanmedulloblastoma cell lines, ranging from about 26104
to 46104 IGF-IR molecules per cell. These values wereestimated on the basis of the IGF-IR protein levels in
R600 cells that are known to express 36104 IGF-IRmolecules per cell (Reiss et al., 1998b; Rubini et al.,1997), and have been repeatedly used in semi-quantitative assays of IGF-IR levels in cells and tissues(Reiss et al., 1998a,b, 1999, 2000). Levels of the IGF-IR in mouse medulloblastoma cell lines were onlyslightly altered when compared with control mousecerebellar tissue. We subsequently examined PI3-K thatlinks IGF-IR signaling with the Akt/PKB activation ±pathway/s associated with cell protection from apopto-tic death (Kulik et al., 1997; Kau�mann-Zeh et al.,1997). As shown in Figure 3 only small di�erenceswere found for PI3-K protein levels when themedulloblastoma cell lines were compared with controlmouse (Figure 3a) or human cerebellum (Figure 3b). Incontrast to IGF-IR and PI3-K, the major IGF-IRsubstrate, IRS-1 was signi®cantly increased in medullo-blastoma cell lines (Figure 3a,b). Densitometricanalysis showed that the mouse medulloblastoma celllines, Bs-1b, Bs-1c, BsB8 and BsB13 have aboutfourfold; ®vefold; 10-fold and 12-fold higher IRS-1protein levels respectively, when compared with normalmouse cerebellum. Additionally, levels of IRS-1 inmedulloblastoma cell lines were comparable to thosefound in mouse R600 ®broblasts that have previouslybeen shown to express high levels of IRS-1 (Reiss etal., 2000). Overall levels of the IGF-IR and IRS-1proteins in mouse medulloblastoma cell lines (Figure3a) were similar to those detected in human medullo-blastoma cell lines (Figure 3b) with the exception ofDaoy cells, where both IGF-IR and IRS-1 were found
Figure 2 Immunocytochemical detection of the phosphorylated IGF-IR (pY1316) in medulloblastoma cell lines. Three mouse (Bs-1b, Bs-1c, BsB8), and three human (TE-671, D283 Med, Daoy) medulloblastoma cell lines were made quiescent (48 h incubation inserum-free medium) and were stimulated with IGF-I (50 ng/ml) for 15 min (a,c,e,g,i,k), or were left without stimulation (b,d,f,h,j,l).Bs-1b cells are depicted in (a) and (b); Bs-1c cells in (c) and (d); BsB8 in (e) and (f); TE-671 cells in (g) and (h); D283 Med in (i) and(j); and ®nally Daoy in (k) and (l). Original magni®cation 406. Insets: corresponding cell lines stained with mouse anti-synaptophysin antibody (Chemicon, MAB332)
Activity of the IGF-IR system in medulloblastomasJY Wang et al
3860
Oncogene

to be present at considerably lower levels. The majordi�erence between mouse and human cells is related tothe fact that the IGF-IR and IRS-1 proteins levels were
very low (not detectable with the exposure time shownin Figure 3b) in the three control specimens (N906,N989, N1061) of human cerebellum. This comparisonbetween control tissues and tumor cell lines point to anapparent reactivation of both IGF-IR and IRS-1expression in human medulloblastoma cell lines, andto strong activation of IRS-1 in mouse medulloblasto-ma cell lines. Importantly, when the same blots werere-probed with anti-PI3-K antibody, an apparent85 kD band was detected in all protein samples,including human cerebellar tissues. To con®rm goodpreservation of cellular proteins in frozen humansamples, the blots were additionally probed with anti-Shc, anti-EGF-IR, and anti-Grb-2 antibodies respec-tively (Figure 3c).
IGF-IR signaling in medulloblastoma cell lines
The immunocytochemical data depicted in Figures 1and 2 demonstrate the initial activation steps of theIGF-IR system, i.e., autophosphorylation of the b-subunit of IGF-IR. By immuno-precipitating the IGF-IR and probing corresponding blots with the anti-phospho-tyrosine antibody, we were able to con®rmIGF-IR phosphorylation in selected medulloblastomacell lines (Figure 4a). Only cells stimulated with theIGF-I (+) had an apparent 97 kD (IGF-IRb) band.Interestingly, TE-671 cells have an additional strongband at the level of about 80 ± 85 kD that is tyrosinephosphorylated and co-precipitates with the IGF-IRindependently from the IGF-I stimulation. Loadingconditions were monitored by re-probing the sameblots with the antibody directed against the b subunitof IGF-IR (lower part of Figure 4a).
Activated IGF-IR recruits and phosphorylates itsmajor substrate, IRS-1. Figure 4b shows that selectedmouse and human medulloblastoma cell lines phos-phorylated IRS-1 on tyrosine residues within 15 minafter IGF-I stimulation. Forty-eight hours followingserum starvation IRS-1 tyrosine phosphorylation wasnot detectable in Bs-1b, BsB8 and Daoy cells. Incontrast, serum starved TE-671 cells possessed detect-able levels of the tyrosine phosphorylated IRS-1,suggesting the possibility of an autocrine activation inthese human cells. The lower part of Figure 4b showsloading conditions in this experiment.
Activated IGF-IR leads to the activation of Ras/MAP kinase pathway via SHC proteins or partially byrecruiting IRS-1 (Myers et al., 1993; Waters et al.,1993). Although this is a routine procedure, we haveexamined Erk1/Erk2 phosphorylation following IGF-Itreatment (Figure 4c). In all medulloblastoma cell linesexamined, short term IGF-I stimulation resulted inErk1/Erk2 phosphorylation on threonine 202 andtyrosine 204, the event that triggers their kinase activity(Payne et al., 1991). In contrast to residual phosphor-ylation of the IRS-1 detected in TE-671 cells culturedin serum-free medium (Figure 4b), MAP kinases werenot activated without additional stimulation, suggest-ing that exogenous IGF-I is still required to activateMAP kinases in this experimental setting. To monitor
Figure 3 IGF-IR, PI3-K, and IRS-1 protein levels in medullo-blastomas. (a) Proteins detected by Western blots developed forthe IGF-IR (b subunit), IRS-1, and PI3-K in R600 mouse®broblasts, mouse medulloblastoma cell lines (Bs-1b, Bs-1c, BsB8,BsB13), and control mouse cerebellum. (b): Protein levels of theIGF-IR, PI3-K and IRS-1 detected in R600 ®broblasts, humancerebellum (N906, N989, N1061), and human medulloblastomacell lines (TE-671, Daoy, D283 Med). Total proteins (50 mg perlane) were isolated from the cell lines cultured under standardcondition and from mouse and human cerebellar tissues (seeMaterial and methods for details). (c): To determine goodpreservation of the human samples blots from (b) were `stripped'and re-probed with anti-Shc, anti-EGF-R and anti-Grb-2antibodies. Only ®rst four lanes are shown including proteinextract from R600 cells as a positive control
Oncogene
Activity of the IGF-IR system in medulloblastomasJY Wang et al
3861

loading conditions, the same blots were re-probed withthe antibody that recognizes Erk1 and Erk2 indepen-dent of their phosphorylation status, (lower part ofFigure 4c).
The Western blot depicted in Figure 4d is the sameone shown in Figure 4c after re-probing with the anti-phospho-Akt antibody. This was done to monitor Aktphosphorylation in the same experimental samples asdescribed for MAP kinases. Samples that had demon-strated activated MAP kinases following IGF-Istimulation were also characterized by an apparentAkt/PKB phosphorylation (Figure 4d). A clearabnormality in Akt activation was detected in TE-671cells. In contrast to all other cell lines, serum starved
TE-671 was able to keep Akt phosphorylated, evenwhen the process of serum starvation was prolongedup to 72 h (not shown).
IGF-I-dependent growth and survival of medulloblastomacell lines
Our results (Figure 3) suggest that the IGF-IR andIRS-1 proteins detected in mouse and human medullo-blastoma cell lines are expressed at levels (Reiss et al.,1998b; Rubini et al., 1997) that ought to support cellproliferation and survival. In fact, Figure 5 shows thatmedulloblastoma cell lines are indeed able to prolifer-ate when stimulated solely with IGF-I. Cell prolifera-tion was evaluated by determining cell number prior tostimulation (T0), and at two di�erent time pointsfollowing IGF-I stimulation, as described in ourprevious papers (Reiss et al., 1998b; Morrione et al.,2000). All cell lines examined also showed restrictedabilities to increase cell number when cultured inserum-free medium without stimulation. However,these responses were marginal when compared withthe magnitude of cell proliferation in the presence ofIGF-I. There was again one exception, TE-671 humancells proliferated in serum-free medium almost ase�ciently as in the presence of IGF-I (Figure 5). Inall other cell lines an average of about threefoldincrease in cell number was observed over a period of 4days following IGF-I stimulation (Figure 5).
When JCV T-antigen positive (BsB8, BsB13) andnegative (Bs-1b, Bs-1c) mouse cell lines were analysedin monolayer cultures their growth responses to IGF-Istimulation (Figure 5), or to serum stimulation (notshown) were similar. Di�erences began to emerge whenthe cells were analysed in anchorage-independentconditions. To deny attachment, cells were culturedon 35 mm dishes covered with polyHema as previouslydescribed (Reiss et al., 1998b; Valentinis et al., 1999a).In suspension, T-positive cell lines (BsB8 and BsB13)survive well in serum-free medium (Figure 6a). Incontrast, T-negative cell lines (Bs-1b and Bs-1c) die insuspension when cultured in serum-free medium,showing 57% decrease and 32% decrease in cellnumber respectively. The cell loss was completelyprevented by the IGF-I (Figure 6a), or partiallyattenuated by the presence of 10% FBS (Figure 6bcompare open bars). The T-positive cells BsB8 andBsB13 responded to IGF-I treatment with cellproliferation showing 227 and 128% increases in cellnumber respectively. On the other hand, T-negative Bs-1b and Bs-1c cells grew much slower showing only 8%increase and 31% increase in cell number whenstimulated solely with the IGF-I (Figure 6a).
The next set of experiments analysed the degree towhich impairments of IGF-IR function a�ect growthand survival of medulloblastoma cell lines in culture.These experiments were also conducted in anchorage-independent conditions, i.e. when cells are deniedattachment to the extracellular matrix. Figure 6bshows the results from one human and two mousemedulloblastoma cell lines. The cells were plated in
Figure 4 IGF-I ±mediated phosphorylation of the IGF-IR andits major signaling molecules. Indicated above quiescent cells werestimulated (+) or not stimulated (7) with IGF-I (50 ng/ml) for15 min. (a): IGF-IR immuno-precipitated from 500 mg of totalproteins with anti-IGF-IR antibody (Calbiochem), and with anti-mouse agarose-conjugated IgG (Sigma). Corresponding blotswere probed ®rst with anti-phosphotyrosine (PY) antibody(Transduction Laboratories, PY20: HRP) (upper blot), and re-probed with anti-IGF-IRb antibody (Santa Cruz) (lower blot).(b): IRS-1 immuno-precipitated from 500 mg of total proteins withrabbit anti-IRS-1 antibody (Upstate Biotechnology), and withprotein A-agarose (Calbiochem). Corresponding blots wereprobed ®rst with anti-phosphotyrosine (PY) antibody (Transduc-tion Laboratories, PY20: HRP) (upper blot), and re-probed withanti-IRS-1 antibody (Upstate Biotechnology) (lower blot). (c):Western blot for the phosphorylated ERKs (p-Erk1/Erk2) (upperblot) and for total Erk2/Erk2 (lower blot) in medulloblastoma celllines. (d): The same blot as in (c) re-probed with the antibodydirected against phosphorylated form of the Akt/PKB (p-Akt)
Activity of the IGF-IR system in medulloblastomasJY Wang et al
3862
Oncogene

suspension at 16105/ml in the presence of 10% FBS,and the cell number was evaluated 48 h later. Althoughnot observed in monolayer cultures (Figure 5), therewas again a dramatic di�erence in cell proliferationwhen selected cell lines were cultured in suspension.For instance, mouse BsB8 cells and human TE-671cells are characterized by 96 and 93% increase in cellnumber respectively, over the period of 2 days in thepresence of 10% FBS. Conversely, a 7% decrease inthe number of Bs-1b cells was observed under the sameconditions (Figure 6b, open bars). Subsequently, cellgrowth and survival were analysed following challengewith antisense oligonucleotides against the IGF-IRmRNA (AS), or after treatment with the conditionedmedium containing a dominant negative mutant of theIGF-IR (486/CM) (Reiss et al., 1998a). As illustratedin Figure 6b all cell lines responded strongly to thetreatments. In comparison to the cell number deter-mined before the treatment (T0), the number of Bs-1bcells decreased by 69 and 79% following AS treatmentor 486/CM treatment respectively. The number of Bs-B8 cells was reduced by 66% following AS treatmentand by 36% in the presence of 486/CM. Finally, cellloss observed in TE-671 was 24% for AS treatment,and 43% for 486/CM treatment. Importantly, cells incontrol conditions treated either with equal concentra-tions of the mismatched oligonucleotides (MM), orwith control-conditioned medium (C/CM) (see Materi-al and methods), were much less a�ected. When the Bs-1b cells were treated with MM oligonucleotides or withC/CM, 24 and 27% decreases in cell number wereobserved respectively. BsB8 cells cultured in the
presence of MM or C/CM were still able to proliferate,showing a 51 and 73% increase in cell number. Thesame was observed for TE-671 cells that showed a 73%increase in cell number after the treatment with MMoligonucleotides, and a 74% increase following C/CMtreatment. The inset in Figure 6b shows the speci®cityand e�ciency of the antisense treatment. Whencompared with untreated cultures (lane 5), BsB8 cellstreated with AS showed dose and time dependentdownregulation of the IGF-IR protein. IGF-IR wasbarely detectable by Western blotting when BsB8 cellswere exposed to 150 nM AS for 36 h (Figure 6b, lane4).
Discussion
The ®nding that the phosphorylated form of the IGF-IR is present in medulloblastoma biopsies (Figure 1)emphasizes the potential involvement of the IGF-IRsystem in the development and progression of thesetumors. These results clearly indicate that the IGF-IRsystem is not only functional in vitro when cells arestimulated with IGF-I (Patti et al., 2000), but is alsocontinuously active in several medulloblastomas exam-ined in this study.
We have selected three human and four mousemedulloblastoma cell lines to explore further theinvolvement of IGF-IR in the growth of medulloblas-tomas. Our ®ndings can be summarized as follows: (i)mouse and human medulloblastoma cell lines allexpress functional IGF-IR that is tyrosine phosphory-
Figure 5 IGF-I induced proliferation of mouse and human medulloblastoma cell lines. All cell lines examined were plated at 16104
cells/35 mm dish in the presence of 10% FBS. Following 24 h incubation attached cells were washed 36with HBSS, and serum-containing medium was replaced with serum-free medium (SFM). Cells were cultured in SFM for an additional 48 h, and cells werestimulated with 50 ng/ml of IGF-I (IGF), or were left without stimulation (SF). Cell number was determined at the time (T0),before stimulation, and at 48 and 96 h following IGF-I stimulation. Ordinate: cell number6104. Abscissa: Time after stimulation.The results are representative of three experiments and are shown with standard deviation
Oncogene
Activity of the IGF-IR system in medulloblastomasJY Wang et al
3863

lated following IGF-I stimulation; (ii) levels of theIGF-IR protein in mouse and human medulloblastomacell lines examined range from about 26104 to 46104
IGF-IR molecules per cell which were not signi®cantlydi�erent from those found in control mouse andhuman cerebellum; (iii) PI3-K, one of the IGF-IRsignaling molecules, was not signi®cantly di�erent incell lines from control tissues; (iv) in contrast, levels ofIRS-1 protein, the major substrate for both IGF-Ireceptor and insulin receptor, were found to be at leastseveral folds greater in mouse and human medullo-blastoma cell lines when compared with correspondingnormal mouse and human cerebellum (Figure 3);®nally, (v) both mouse and human medulloblastomacell lines die in suspension cultures when the functionof the IGF-IR was challenged by a dominant negativemutant of the IGF-IR (486/STOP), or by antisenseoligonucleotides aimed against the IGF-IR mRNA.
Certain di�erences between mouse and humanmedulloblastoma cell lines emerged, particularly whenhuman tumor cell were compared with the normalhuman cerebellum. It appears that the IGF-IR andIRS-1 protein levels are extremely low in humanpediatric cerebellum (Figure 3b). Therefore, in compar-ison to the low levels in the control human tissues,both IGF-IR and IRS-1 are apparently reactivated inall three human medulloblastoma cell lines examined.This is distinct from mouse medulloblastoma cell linesin which only IRS-1 protein is strongly increased, andthe IGF-IR protein level is not signi®cantly di�erentfrom normal cerebellar controls (compare Figure 3aand b). There is of course concern related to thecontrol human materials used in this study. Thesections of human cerebellar tissues were acquiredbetween 13.5 ± 36 h post mortem. Thus the possibilityexists that the proteins isolated from these sectionswere not well preserved. Microscopic examination ofthe sections, and the abundant presence of the PI3-Kband (85 kD) detected in all protein samples, includingnormal human cerebellar tissues (Figure 3), partiallyexcludes this possibility. Additionally, blots containingproteins from normal human cerebellum were re-probed with other antibodies (anti-EGF-R, anti-SHCand anti-Grb-2), demonstrating in all cases thepresence of the corresponding proteins in this humanmaterial (Figure 3c).
The ®nding that IRS-1 protein is strongly up-regulated in all medulloblastoma cell lines, in combina-tion with the presence of the functional IGF-IR mayhave implications as to the pathways which control thegrowth of primitive neuroectodermal tumors. Addi-tionally, low levels of IRS-1 protein in normaldi�erentiated cerebellar tissues may have also otherimplications. As previously reported, signaling fromthe activated IGF-IR can trigger cellular di�erentiationbut only in the absence of IRS-1 protein (Valentinis etal., 1999b). Therefore the possibility exists thatdevelopmental down-regulation of IRS-1 protein maybe partially responsible for IGF-I-mediated cerebellardi�erentiation. Conversely, reactivation of IRS-1expression in cells from the external granular layer
Figure 6 Growth and survival of medulloblastoma cell lines inanchorage-independence. (a) JCV T- antigen positive (BsB8,BsB13) and negative (Bs-1b, Bs-1c) cell lines were cultured inanchorage-independence in serum-free medium (SF) or werestimulated with IGF-I (IGF). Cell lines used in the experimentare shown in the table located below the histogram. Cell numberwas determined at the time of treatment (T0), and 48 h followingthe treatment. Results are presented as a percent increase ordecrease in cell number over T0, and are shown with standarddeviation. (b) Impairment of the IGF-IR function in selectedmedulloblastoma cell lines. Two di�erent treatments wereemployed: antisense oligonucleotides against IGF-IR mRNA(Reiss et al., 1995) and the conditioned medium containing adominant negative mutant of the IGF-IR designed 486/STOP (seeMaterials and methods for details). Cells were cultured inanchorage-independence in the presence of 10% FBS. Treatmentsand cell lines used in the experiment are shown in the tablelocated below the histogram. Cell number was determined at thetime of treatment (T0), and 48 h following the treatment. Resultsare presented as a per cent increase or decrease in cell numberover T0, and are shown with standard deviation. Individualvalues are indicated below the histogram. Inset: Western blotshowing IGF-IR protein levels in BsB8 cells treated with antisenseoligonucleotides against the IGF-IR mRNA (AS). Cells weretreated with 100 or 150 nM of AS for 16 and 36 h. Note apparentdownregulation of the IGF-IR protein by AS in a dose- and time-dependent manner
Activity of the IGF-IR system in medulloblastomasJY Wang et al
3864
Oncogene

may contribute to the development and expansion ofmedulloblastomas. Mechanistically, IRS-1 binds to theactivated IGF-IR at Tyr 950, amplifying the mitogenicsignal via coupling the receptor with the PI3-Kpathways and partially with the Ras/MAP kinasepathways (Kulik et al., 1997; Reiss et al., 2000).Additionally, the IRS-1 protein has transformingproperties but only when overexpressed in cells witha functional IGF-IR (Fei et al., 1995). In addition, theIRS-1 protein has been found to co-precipitate with theSV40 T antigen. Simultaneous ectopic expression ofboth IRS-1 and SV40 T is able to sustain thetransformed phenotype of the cell, even in the absenceof the IGF-IR (Fei et al., 1995). We have selected twomouse medulloblastoma cell lines (Bs-1b and Bs-1c)that are JCV T-antigen negative and two cell lines(BcB8 and BsB13) that are JCV T-antigen positive(Krynska et al., 2000). When the cells were analysed inmonolayer cultures their growth responses followingIGF-I stimulation were similar (Figure 5). Di�erencesbegan to emerge when the cells were analysed inanchorage-independent conditions. In suspension, T-positive cell lines (BsB8 and BsB13) survived well inserum-free medium and proliferated following IGF-Istimulation (Figure 6a). Conversely, T-negative celllines (Bs-1b and Bs-1c) died in suspension whencultured in serum-free medium (Figure 6a). The cellloss was completely prevented by IGF-I (Figure 6a), orpartially attenuated by 10% FBS (Figure 6b compareopen bars). Finally, T-positive cells responded to amuch greater degree than T-negative cells whenstimulated solely with IGF-I. At present we can onlyspeculate about the mechanism/s responsible for thedi�erences in cell behavior in anchorage-independence.Considering that both T-positive and T-negative cellshave similar levels of the IGF-IR and P13-K proteins,and both grossly overexpress IRS-1 (Figure 3), it isfeasible that the IGF-IR signaling pathway and JCV T-antigen may cooperate by amplifying the signaltowards transformation in this cellular system. Thereare several reports that support this notion: JCV T-antigen has high homology to SV40 large T-antigen(Gallia et al., 1998); both JCV T-antigen and SV40 T-antigen have transforming properties (Gallia et al.,1998); SV40 T-antigen is not able to transform cells inthe absence of IGF-IR (Sell et al., 1993); cellsexpressing functional IGF-IR can be transformedeither by ectopic expression of SV40 T-antigen or byIRS-1 overexpression (Fei et al., 1995); IRS-1 co-precipitates with the SV40 T-antigen (Fei et al., 1995),and SV40 T-antigen enhances expression from theIGF-1 promoter (Porcu et al., 1994). Whether similarinteractions exist between the IGF-IR system and theJCV T-antigen, and whether these interaction/s con-tribute to the development and progression ofmedulloblastoma remains to be determined.
IGF-IR protein levels between 26104 to 46104
IGF-IR molecules per cell seem to be high enough(Reiss et al., 1998b) to transduce both proliferative andsurvival signals in medulloblastoma cell lines. Indeed,15 min following IGF-I stimulation all medulloblasto-
ma cell lines responded with auto-phosphorylation ofthe IGF-IR (Figures 2 and 4a), and tyrosinephosphorylation of IRS-1 (Figure 4b). There are,however, some di�erences in the activation of signalingmolecules between the medulloblastoma cell linesanalysed. For instance, when IGF-IR was immuno-precipitated from TE-671 cells, a strong additionalband of about 80 ± 85 kD was detected when blots wereprobed with an anti-phosphotyrosine antibody (Figure3a). The presence of this additional phosphoproteinexclusively in TE-671 cells did not depend on IGF-Istimulation. In addition, this band was not detectedwhen the same blots were re-probed with anti-IGF-IR(Figure 3a lower panel) or with anti-PI3-K (p85)antibodies (not shown), excluding the possibility of acontinuous association between the IGF-IR and PI3-Kin these cells. It is possible also that this unidenti®edphosphoprotein is responsible for false positive im-muno-staining of TE-671 cells in serum-free medium(Figure 2k), since tyrosine phosphorylation of the IGF-IR is not detected in these cells cultured in serum-freemedium (Figure 4a).
Another abnormality of IGF-IR signaling involvesresidual phosphorylation of IRS-1 (Figure 3b) andAkt/PKB (Figure 3d) repeatedly detected in TE-671cells, but not in other medulloblastoma cell linescultured in serum-free medium. This constitutivephosphorylation of the signaling molecules may beresponsible for the ability of this cell line to growwithout exogenous stimulation (Figure 5). One ex-planation may be that TE-671 cells are stimulated in anautocrine manner via production and secretion of IGF-I. Such a possibility has already been proposed forDaoy cells (Patti et al., 2000). In our hands however,and in contrast to TE-671, Daoy cells still requireexogenous IGF-I to proliferate (Figure 5). It must beemphasized that although TE-671 were isolated from abiopsy fragment from a cerebellar medulloblastoma,and were classi®ed once as a medulloblastoma cell line,several later articles criticized this classi®cation (forinstance: Stratton et al., 1989; Pappas et al., 1996).Accordingly, TE-671 cells possess features that cate-gorize them as rhabdomyosarcomas rather thanmedulloblastomas (Stratton et al., 1989). This includesmutation within N-Ras gene (Fults et al., 1989),expression of the muscle-type nicotinic acetylcholinereceptor (Pappas et al., 1996), and the presence ofintermediate ®lament protein, desmin (Pappas et al.,1996). Since our present data show a strong discre-pancy in the pattern of IGF-IR activation between TE-671 cells, and other mouse and human medulloblasto-ma cell lines examined, this extends even further thedi�erent origin of TE-671 cells.
Strong evidence supporting the role of the IGF-IRsystem in growth and survival of medulloblastoma celllines comes from experiments in which the IGF-IR waschallenged either with antisense oligonucleotidesagainst the IGF-IR mRNA, or with a dominantnegative mutant of the IGF-IR, designated as 486/STOP (Figure 6b). The 486/STOP product has beenshown in our previous work to compete with the IGF-
Oncogene
Activity of the IGF-IR system in medulloblastomasJY Wang et al
3865

IR function, inhibiting growth and survival of cancercells in both in vitro and in vivo experiments (Reiss etal., 1998a; D'Ambrosio et al., 1996; Dunn et al., 1998).Importantly, our results show that medulloblastomacells die in suspension cultures when challenged withthe AS oligonucleotides or with the 486/STOP product(Figure 6b). It has been previously reported thatdi�erent manipulations aimed against the IGF-IRfunction have a much more dramatic e�ect on cancercells forced to grow in anchorage-independence thanon cells growing in monolayer cultures (Baserga et al.,1997). This may suggest the possibility of targeting theIGF-IR function when tumor cells in®ltrate adjacenttissues. In this stage medulloblastoma cells may beforced temporarily to grow and survive anchorage-independence, a condition in which anti-apoptoticfunction of the IGF-IR is critical (Baserga et al.,1997). Importantly, these treatments could be lessharmful to normal anchorage-dependent host braincells.
In summary, the medulloblastoma cell lines exam-ined express major components of the IGF-IR systemand grossly overexpress IRS-1 protein. Within 15 minafter IGF-I stimulation mouse and human cell linesphosphorylate the b subunit of the IGF-IR, IRS-I,Akt/PKB, and MAP kinases; respond with prolifera-tion when stimulated solely with the IGF-I; and arestrongly inhibited when challenged with a dominantnegative mutant of the IGF-IR (486/STOP), or withantisense oligonucleotides against the IGF-IR mRNA.The signi®cance of the fact that the IGF-IR maycontribute to the growth of medulloblastoma cells wasvalidated by a positive staining of biopsies frommedulloblastoma patients with an antibody thatrecognizes the phosphorylated (active) form of theIGF-IR.
Materials and methods
Clinical samples
A total of six human medulloblastoma samples were obtainedfrom the pathology archives of the following institutions:MCP-Hahnemann University (Philadelphia, PA, USA), andSt. Christopher's Hospital for Children (Philadelphia, PA,USA). Formalin-®xed, para�n embedded surgical resectionswere histologically classi®ed according to the World HealthOrganization Classi®cation of Tumors in the NervousSystem. Samples of frozen human cerebellum (N906, N989and N1061) were kindly provided by Dr Mary Herman andDr Joel E Kleinman from the Neuropathology Section-CBDB NIMH (Bethesda, MD, USA).
Cell lines
The cell lines used include D283 Med human cells (ATCC#HTB-185) that originate from peritoneal metastases from aboy with medulloblastoma; Daoy human cell line (ATCC#HTB-186) isolated from a posterior fossa tumor of a 4-year-old boy; TE-671 human cells (ATCC# CRL-8805) isolatedfrom a biopsy of a cerebellar medulloblastoma; and fourmouse cell lines (Bs-1b, Bs-1c, BsB8, BsB13) derived from
cerebellar medulloblastomas that developed spontaneously intransgenic mice expressing the early genome of the archetypeform of the JC virus (Krynska et al., 2000). All mouse celllines included are positive for the major neuronal markers. Inaddition, Bs-1b and Bs-1c cells are JCV T-antigen negative,and BsB8 and BsB13 are JCV T-antigen positive (Krynskaet al., 2000). R600 mouse embryo ®broblasts that express36104 IGF-IR molecules per cell (Rubini et al., 1997; Reisset al., 1998b) were included to estimate levels of the IGF-IRexpression in medulloblastomas. R7/486/HSP/Flag cells wereengineered to secrete large quantities of a dominant negativemutant of IGF-IR designated as 486/STOP. R7/486/HSP/Flag cells are based on R7 cells derived from IGF-IR geneknockout mice, and do not express IGF-IR (Rubini et al.,1997). These cells were stably transduced with a retroviralexpression vector containing 486/STOP cDNA cloned inframe with the Flag tag. Expression of the fusion protein486/STOP-Flag is under control of Drosophila heat shockpromoter (HSP) that ensures e�cient production of the 486/STOP product at 398C. Control R7 GR219 cells weretransduced with an `empty' retroviral vector.
Growth in monolayer
All cell lines were cultured in Dulbecco's Modi®ed EagleMedium (DMEM) (GIBCO±BRL, Grand Island, NY, USA)containing 10% fetal bovine serum (FBS), at 378C in a 9.6%CO2 atmosphere. To make cells quiescent, they were seededat a density of 2.56103/cm2 in the presence of serum. After24 h, the cells were washed with Hank's balanced saltsolution (HBSS) and incubated in serum-free medium(SFM) [DMEM supplemented with 0.1% bovine serumalbumin (SIGMA)]. The cells became quiescent after 48 hin SFM, which was renewed with fresh medium every 24 h.Growth response to IGF-I was tested by stimulatingquiescent cells with IGF-I (GIBCO±BRL, Grand Island,NY, USA) at 50 ng/ml.
Preparation of the 486/STOP conditioned medium
To prepare conditioned medium (CM) containing the 486/STOP product, 70% con¯uent R7/486/HSP/Flag cells grownin DMEM plus 10% FBS and 1 mg/ml of Puromycin werewashed three times with PBS, and cultured in serum-freemedium at 398C for an additional 24 h. The CM wascollected and aliquots frozen at 7808C. Control CM (C/CM)was prepared in the same way from the R7GR219 cells thatcontain the empty retroviral vector. Collected conditionedmedia were concentrated about 10-fold using Centriprep 50centrifugal ®lters (Amicon, Bedford, MA, USA), analysed byWestern blotting for the presence of the 486/STOP productand utilized to compete with the IGF-IR function by mixingone volume of the 486/CM with one volume of the culturemedium (DMEM+10% FBS). Control cultures were treatedthe same way with the exception that 486/CM was replacedwith the C/CM.
Cell growth and survival in suspension
To determine this parameter, quiescent cells were detachedfrom a culture dish with 0.02% EDTA (disodium ethylene-diamine tetraacetate), and seeded on dishes coated withpoly(2-hydroxyethyle methacrylate) [poly(HEMA)] (Aldrich,Milwaukee, WI, USA), prepared according to the methodol-ogy previously described (Reiss et al., 1998b). Cells wereseeded in 10% FBS at concentration of 16105 cells/35 mmdish, and were treated either with 5 mM antisense oligonucleo-
Activity of the IGF-IR system in medulloblastomasJY Wang et al
3866
Oncogene

tides (Reiss et al., 1995), or with the 486/CM, or were leftuntreated. Forty-eight hours later cell suspensions werecollected, dissociated with 0.25% trypsin, and cells werecounted in a Bright-line Hemocytometer. Results represent anaverage of three experiments, and are expressed as per centincrease or decrease in cell number over time zero (T0).Sequences of phosphorothioate antisense (AS) and mis-matched (MM) oligonucleotides to the IGF-IR mRNA werepreviously described (Reiss et al., 1995).
Western blots and immuno-precipitation
To determine total IGF-IR, IRS-1 and PI3-K levels,monolayer cultures or small fragments of cerebellar tissuewere lysed on ice with 400 ml of lysis bu�er [(50 mM HEPES);pH 7.5; 150 mM NaCl; 1.5 mM MgCl2; 1 mM EGTA; 10%glycerol; 1% Triton X-100; 1 mM phenylmethylsulfonyl¯uoride (PMSF); 0.2 mM Na-orthovanadate and 10 mg/mlaprotinin]. Protein concentration was determined by a Bio-Rad Protein Assay (BioRad, Hercules, CA, USA), and 50 mgof total proteins separated on a 4 ± 15% gradient SDS ±PAGE (BioRad) and transferred into nitrocellulose mem-branes. Blots were blocked with 5% non-fat dry milk inTBST (10 mM Tris-HCl, pH 7.5, 150 mM NaCl, 0.1% Tween20), probed with rabbit anti-IGF-IRb antibody (Santa CruzInc. Santa Cruz, CA, USA); rabbit anti-IRS-1 antibody(Upstate Biotechnology Inc., Lake Placid, NY, USA), rabbitanti-PI3-K (p85) antibody (Upstate Biotechnology), and withanti-rabbit-HRP conjugated secondary antibody (Calbio-chem, San Diego, CA, USA). Blots were developed withECL detection reagents (Amersham, Arlington, IL, USA). Todetermine phosphorylated forms of IGF-IR, IRS-1, Akt/PKBand MAP kinases, quiescent cells were stimulated with IGF-I(50 ng/ml) and total proteins were extracted 15 min afterstimulation. Total and phosphorylated Akt/PKB levels weredetermined by PhosphoPlus Akt(Ser473) antibody kit (NewEngland BioLabs, Beverly, MA, USA), and total andphosphorylated ERKs levels were determined by Phospho-Plus Phospho-p44/42 MAP kinase (Thr202/Tyr204) antibodykit (New England BioLabs, Beverly, MA, USA). Todetermine IGF-IR tyrosine phosphorylation IGF-IR wasimmuno-precipitated from 500 mg protein extracts withmouse anti-IGF-IR antibody (Calbiochem, San Diego, CA,USA) and agarose conjugated anti-mouse IgG (Sigma, St.Louis, MO, USA), and corresponding blots developed ®rstwith anti-phosphotyrosine (PY) antibody (TransductionLaboratories, Lexington, KY, USA) (PY20: HRP), and re-probed with anti-IGF-IR antibody (Santa Cruz). Todetermine IRS-1 tyrosine phosphorylation IRS-1 was im-muno-precipitated from 500 mg protein extracts with rabbitanti-IRS-1 antibody (Transduction Laboratories) and agarose
conjugated protein A (Calbiochem). Corresponding blotswere developed ®rst with anti-phosphotyrosine (PY20) anti-body, and re-probed with N-terminal anti-IRS-1 antibody(Santa Cruz).
Immunohistochemistry
Formalin ®xed, para�n embedded samples were sectioned at4 microns thickness and stained with Hematoxylin-Eosin forroutine histological analysis. Immunohistochemistry wasperformed by using the avidin-biotin-peroxidase complexsystem, according to the manufacturer's instructions (Vectas-tain Elite ABC Peroxidase Kit; Vector Laboratories). Brie¯y,sections were depara�nized in xylene and re-hydratedthrough alcohols up to water. Sections were heated in0.01 M sodium citrate bu�er (pH 6.0) to 958C under vacuumfor 40 min and allowed to cool for 20 min at roomtemperature for non-enzymatic antigen retrieval. The slideswere rinsed with PBS and incubated in MeOH/3% H2O2 for20 min to quench endogenous peroxidase. Sections were thenwashed with PBS and blocked in PBS/0.1% BSA containing5% normal horse or goat serum for 2 h at room temperature,and incubated overnight at room temperature with primaryantibodies. The primary antibodies utilized were rabbit anti-pY1316 IGF-IR (1 : 500 dilution); rabbit anti-IGF-IR (1 : 500dilution, Santa Cruz); anti-GFAP antibody (1 : 100 dilution,DACO); anti-neuro®laments antibody (1 : 500 dilution, Stern-berger Monoclonals); and anti-synaptophysin antibody(1 : 500 dilution, clone SY38; Roche Molecular, Indianapolis,IN, USA). Secondary antibodies used were biotin-conjugatedanti-mouse and anti-rabbit IgGs (Vector). Following second-ary antibodies, avidin-biotin peroxidase complexes (Vector)were incubated for 1 h at room temperature, sections weredeveloped with diaminobenzidine substrate (Sigma), counter-stained with hematoxylin and mounted with Permont. Forimmunocytochemistry, cells cultured on poly-D-lysine coatedglass chamber slides (Nunc) were washed in cold PBS, ®xedwith cold acetone and stained as described for immunohis-tochemistry.
AcknowledgmentsSamples of frozen human cerebellum were kindly providedby Dr Mary Herman and Dr Joel E Kleinman fromNeuropathology Section-CBDB NIMH, Bethesda, MD,USA. This work is supported by grant NIH PO-1 NS36466. Reiss K, Tu X, Romano G, Wang J, Baserga R.Mechanism of action of a dominant-negative mutant of theIGF-I receptor (submitted for publication).
References
Antoniades HN, Galanopoulos T, Neville-Golden J andMaxwell M. (1992). Int. J. Cancer, 50, 215 ± 222.
Baker J, Liu JP, Robertson EJ and Efstratiadis A. (1993).Cell, 75, 73 ± 82.
Baserga R. (1994). Cell, 79, 927 ± 930.Baserga R. (1995). Cancer Res., 55, 240 ± 252.Baserga R, Hongo A, Rubini M, Prisco M and Valentinis B.
(1997). Biochim. Biophys. Act., 1332, F105 ± F126.Burgaud JL, Resnico� M and Baserga R. (1995). Biophys.
Biochem. Res. Comm., 214, 475 ± 481.
Coppola D, Ferber A, Miura M, Sell C, D'Ambrosio C,Rubin R and Baserga R. (1994). Mol. Cell. Biol., 14,4588 ± 4595.
D'Ambrosio C, Ferber A, Resnico� M and Baserga R.(1996). Cancer Res., 56, 4013 ± 4020.
DeAngelis T, Ferber A and Baserga RJ. (1995). J. CellPhysiol., 164, 214 ± 221.
Dunn SE, Ehrlich M, Sharp NJH, Reiss K, Solomon G,Hawkins R, Baserga R and Barrett JC. (1998). CancerRes., 58, 3353 ± 3361.
Oncogene
Activity of the IGF-IR system in medulloblastomasJY Wang et al
3867

Fei ZL, D'Ambrosio C, Li S, Surmacz E and Baserga R.(1995). Mol. Cell. Biol., 15, 4232 ± 4239.
Fults D, Maness PF, Nakamura Y and White R. (1989).Brain Res., 503, 281 ± 287.
Gallia GL, Gordon J and Khalili K. (1998). J. Neurovirol., 4,175 ± 181.
Gluckman P, Klempt N, Guan J, Mallard C, Sirimanne E,Dragumow M, Klempt M, Singh K, Williams C andNikolics K. (1992). Biochem. Biophys. Res. Comm., 182,593 ± 599.
Kalebic T, Tsokos M and Helman LJ. (1994). Cancer Res.,54, 5531 ± 5534.
Kau�mann-Zeh A, Rodriguez-Viciana P, Ulrich E, GilbertC, Co�er P, Downward J and Evan G. (1997).Nature, 385,544 ± 548.
Krynska B, Del Valle L, Gordon J, Otte J, Croul S andKhalili K. (2000). Virology, 274, 65 ± 74.
Krynska B, Del Valle L, Croul S, Gordon J, Katsetos CD,Carbone M, Giordano A and Khalili K. (1999). Proc.Natl. Acad. Sci. USA, 96, 11519 ± 11524.
Kulik G., Klippel A and Weber MJ. (1997). Mol. Cell. Biol.,17, 1595 ± 1606.
LeRoith D, Werner H, Faria TN, Kato H, Adamo M andRoberts Jr CT. (1993). Ann. NY Acad. Sci., 692, 22 ± 32.
Liu JP, Baker J, Perkins AS, Robertson EJ and EfstratiadisA. (1993). Cell, 75, 59 ± 72.
Martin DM, Yee D, Carlson RO and Feldman EL. (1992).Mol. Brain Res., 15, 241 ± 246.
Miura M, Li S and Baserga R. (1995). Cancer Res., 55, 663 ±667.
Morrione A, DeAngelis T and Baserga R. (1995). J. Virol.,69, 5300 ± 5303.
Morrione A, Romano G, Navarro M, Reiss K, Valentinis B,Devs M, Rosen MR, Eves E and Baserga R. (2000).Cancer Res., 60, 2263 ± 2272.
Myers MJ, Sun XJ, Cheatham B, Jachna BR, Glasheen EM,Backer JM and White MF. (1993). Endocrinol., 132,1421 ± 1430.
Pappas IS, Sophianos D, Tzartos S and Tsiftsoglou AS.(1996). Cell Growth Di�er., 7, 797 ± 809.
Patti R, Reddy CD, Geoerger B, Grotzer MA, RaghunathM, Sutton LN and Phillips PC. (2000). Int. J. Oncol., 3,577 ± 584.
Payne DM, Rossomando AJ, Martino P, Erickson AK, HerJH, Shabanowitz J, Hunt DF, Weber MJ and Sturgill TW.(1991). EMBO J., 10, 885 ± 892.
Porcu P, Grana X, Li S, Swantek J, De Luca A, Giordano Aand Baserga R. (1994). Oncogene, 9, 2125 ± 2134.
Prager D, Li HL, Asa S and Melmed S. (1994). Proc. Natl.Acad. Sci. USA, 91, 2181 ± 2185.
Reiss K, Cheng W, Kajstura J, Sonnenblick EH, Meggs LGand Anversa P. (1995). Am. J. Physiol., 269, H943 ±H951.
Reiss K, D'Ambrosio C, Tu X and Tu C. (1998a). Clin.Cancer Res., 4, 2647 ± 2655.
Reiss K, Valentinis B, Tu X and Baserga R. (1998b). Exp.Cell Res., 242, 361 ± 372.
Reiss K, Wang JY, Romano G, Furnari FB, Cavenee WK,Morrione A, Tu X and Baserga R. (2000). Oncogene., 19,2687 ± 2694.
Reiss K, Yumet G, Shan S, Huang Z, Alnemri E, SrinivasulaSM, Wang JY, Morrione A and Baserga R. (1999). J. Cell.Physiol., 181, 124 ± 135.
Resnico� M, Sell CH, Rubini M, Coppola D, D'AmbrosioC, Baserga R and Rubin R. (1994). Cancer Res., 54, 2218 ±2222.
Rubini M, D'Ambrosio C, Carturan S, Yumet G, CatalanoE, Shan S, Huang Z, Criscuolo M, Pi�eri M and BasergaR. (1999). Exp. Cell. Res., 251, 22 ± 32.
Rubini M, Hongo A, D'Ambrosio C and Baserga R. (1997).Exp. Cell Res., 230, 284 ± 292.
Sell C, Baserga R and Rubin R. (1995). Cancer Res., 55,303 ± 306.
Sell C, Dumenil G, Deveaud C, Miura M, Coppola D,DeAngelis T, Rubin R, Efstratiadis A and Baserga R.(1994). Mol. Cell. Biol., 14, 3604 ± 3612.
Sell C, Rubini M, Rubin R, Liu JP, Efstratiadis A andBaserga R. (1993). Proc. Natl. Acad. Sci. USA, 90, 11217 ±11221.
Shapiro DN, Jones BG, Shapiro LH, Dias P and HoughtonPJ. (1994). J. Clin. Invest., 94, 1235 ± 1242.
Steller MA, Zou Z, Schiller JT and Baserga R. (1996). CancerRes., 56, 5087 ± 5091.
Stratton MR, Darling J, Pilkington GJ, Lantos PL, ReevesBR and Cooper CS. (1989). Carcinogenesis, 10, 899 ± 905.
Trojan J, Blossey BK, Johnson TR, Rudin SD, TykocinskiM, Ilan J and Ilan J. (1992). Proc. Natl. Acad. Sci. USA,89, 4874 ± 4878.
Trojan J, Johnson TR, Rudin SD, Ilan J, Tykocinski ML andIlan I. (1993). Science, 256, 94 ± 97.
Valentinis B, Morrione A, Peruzzi F, Reiss K and Baserga R.(1999a). Oncogene, 18, 1827 ± 1836.
Valentinis B, Morrione A, Taylor SJ and Baserga R. (1997).Mol. Cell. Biol., 17, 3744 ± 3754.
Valentinis B, Romano G, Peruzzi F, Morrione A, Prisco M,Soddu S, Cristofanelli B, Sacchi A and Baserga R.(1999b). J. Biol. Chem., 274, 12423 ± 12430.
Waters SB, Yamauchi K and Pessin JE. (1993). J. Biol.Chem., 268, 22231 ± 22234.
Activity of the IGF-IR system in medulloblastomasJY Wang et al
3868
Oncogene




















