
central services - signalling projects group - dickthesignals.co.uk

Available online at www.sciencedirect.com
(2008) 748–757www.elsevier.com/locate/cellsig
Cellular Signalling 20
Activation of the cell integrity pathway is channelledthrough diverse signalling elements in fission yeast
Gregorio Barba, Teresa Soto, Marisa Madrid, Andrés Núñez,Jeronima Vicente, Mariano Gacto ⁎, José Cansado
Yeast Physiology Group
Department of Genetics and Microbiology, Facultad de Biología, University of Murcia, 30071, Campus Universitario de Espinardo, Murcia, Spain
Received 27 November 2007; accepted 14 December 2007Available online 4 January 2008
Abstract
MAPK Pmk1p is the central element of a cascade involved in the maintenance of cell integrity and other functions in Schizosaccharomycespombe. Pmk1p becomes activated by multiple stressing situations and also during cell separation. GTPase Rho2p acts upstream of the proteinkinase C homolog Pck2p to activate the Pmk1 signalling pathway through direct interaction with MAPKKK Mkh1p. In this work we analyzed thefunctional significance of both Rho2p and Pck2p in the transduction of various stress signals by the cell integrity pathway. The results indicate thatbasal Pmk1p activity can be positively regulated by alternative mechanisms which are independent on the control by Rho2p and/or Pck2p.Unexpectedly, Pck1p, another protein kinase C homolog, negatively modulates Pmk1p basal activity by an unknown mechanism. Moreover,different elements appear to regulate the stress-induced activation of Pmk1p depending on the nature of the triggering stimuli. Whereas Pmk1pactivation induced by hyper- or hypotonic stresses is channeled through Rho2p–Pck2p, other stressors, like glucose deprivation or cell walldisturbance, are transduced via other pathways in addition to that of Rho2p–Pck2p. On the contrary, Pmk1p activation observed during cellseparation or after treatment with hydrogen peroxide does not involve Rho2p–Pck2p. Finally, Pck2p function is critical to maintain a Pmk1p basalactivity that allows Pmk1p activation induced by heat stress. These data demonstrate the existence of a complex signalling network modulatingPmk1p activation in response to a variety of stresses in fission yeast.© 2008 Elsevier Inc. All rights reserved.
Keywords: Mitogen-activated protein kinase; Pmk1p; Fission yeast; Cell signalling; Cell integrity pathway; S. pombe
1. Introduction
A great challenge in modern biology is to understand howcells sense extracellular changes and produce programmedresponses for survival and adaptation. In eukaryotic organisms,the mitogen-activated protein kinase (MAPK) cascades arehighly conserved biochemical modules that transduce extra-
Abbreviations:MAPK, mitogen-activated protein kinaseMAPKK, mitogen-activated protein kinase kinaseMAPKKK, mitogen-activated protein kinasekinase kinaseSAPK, stress-activated protein kinaseGFP, green fluorescentproteinHA6H, epitope comprising hemagglutinin antigen plus six histidineresiduesHA, hemagglutininOD600, optical density at 600nm.⁎ Corresponding author. Tel.: +34 968367132; fax: +34 968363963.E-mail address: [email protected] (M. Gacto).
0898-6568/$ - see front matter © 2008 Elsevier Inc. All rights reserved.doi:10.1016/j.cellsig.2007.12.017
cellular signals and control cellular functions such as migration,proliferation and differentiation. Functional MAPK cascadescomprise MAPK kinase kinases (MAPKKKs), which phos-phorylate and activate MAPK kinases (MAPKKs), which inturn activate MAPKs by phosphorylation of two conservedthreonine and tyrosine residues [1,2]. Finally, active MAPKsphosphorylate different substrates, such as transcription factorsor other protein kinases, to promote transcriptional and post-transcriptional changes which play a critical role in theadjustment of the cell behavior to environmental conditions.
The fission yeast Schizosaccharomyces pombe has three MAPkinase cascadeswithin specific signalling pathways thatmodulateresponses to external stimuli. These are the stress activated proteinkinase (SAPK) pathway, the mating pheromone-responsive

749G. Barba et al. / Cellular Signalling 20 (2008) 748–757
pathway, and the cell integrity pathway, whose central elementsare MAPKs Sty1p/Spc1p, Spk1p, and Pmk1p/Spm1p, respec-tively [3–7]. Pmk1p/Spm1p is a structural homolog to MAPKSlt2p/Mpk1p from Saccharomyces cerevisiae [6,7], and amember of the 'classical' family of MAPKs including theextracellular signal-regulated kinases ERK1/2 (p42/p44) fromanimal cells which are activatable by phorbol esters, cytokines orosmotic stress [8]. In S. pombe, Pmk1p is involved in themaintenance of cell integrity, cytokinesis, ion homeostasis, andvacuole fusion because deletion of any gene coding forcomponents of the cascade, namelymkh1+ (encoding MAPKKKMkh1p) [9], pek1+/ skh1+ (encoding MAPKK Pek1p/Skh1p)[10,11], or pmk1+/spm1+ (encoding MAPK Pmk1p/Spm1p)[6,7], causes morphological alterations, multiseptate phenotype,cell death in the presence of potassium ions, hypersensitivity toβ-1,3 glucanases, and defective vacuole fusion [9–12]. In controland stressed cells,Mkh1p and Pek1p localize at the cytoplasm andseptum, whereas Pmk1p is found in both cytoplasm and nucleus,as well as in the mitotic spindle and septum during cytokinesis[6,7,13]. Notably, subcellular distribution of Pmk1p is unaffectedby either stress or absence of Mkh1p and Pek1p, implying thatPmk1p activation is extranuclear and that both the active andinactive forms of this kinase enter to the nucleus [13]. Theidentification of transcription factor(s) regulated by Pmk1p hasremained elusive until recently. However, observations supportthat transcription factor Atf1p, which results activated by Sty1p inresponse to several stresses [14,15], is targeted also by Pmk1p inresponse to mycafungin, an specific beta-glucan synthaseinhibitor [16].
Pmk1p is dually phosphorylated by Pek1p at threonine andtyrosine residues in positions 186 and 188, respectively [10,11].Although it was reported that Pmk1p is only activated by hightemperatures or sodium chloride [6,7], subsequent work hasshown that its activation is induced by multiple stressingsituations, including hyper- and hypotonic conditions, cell wall-damaging compounds, heat shock, glucose deprivation, andoxidative stress [13]. Moreover, Pmk1p becomes periodicallyactivated during the cell cycle, peaking at the M-G1 transition incoincidence with cell separation [17]. Interestingly, all stresssignals activating Pmk1p are exclusively funneled throughMkh1p and Pek1p, suggesting that the basic functional MAPKmodule does not have additional components [13].
There is a complex negative regulation of Pmk1p activity byseveral MAPK-phosphatases in S. pombe. Initially, the dual-specificity phosphatase Pmp1p was the only phosphatase knownto inactivate Pmk1p [18]. However, we have shown recentlythat tyrosine phosphatases Pyp1p and Pyp2p, and threoninephosphatase Ptc1p, whose expression is regulated by MAPKSty1p, are also essential for proper deactivation of Pmk1p understress [17], revealing the existence of a crosstalk between theSAPK and the cell integrity pathways in fission yeast.
Similar to the model described in budding yeast, it has beendemonstrated that in fission yeast the small GTPase Rho2p mayact upstream of the protein kinase C homolog Pck2p to activatethe Pmk1 MAPK signalling cascade through direct interactionwith MAPKKK Mkh1p [19]. However, the functional sig-nificance of Rho2p and Pck2p in the transduction of stress
signals activating the cell integrity pathway has not beenanalyzed. We have performed such study in this work to draw acomplex scenario. The results reveal that stresses of differentnature activate the Pmk1p MAPK cascade through distinctmechanisms which differ in the initial triggering events. Thesignal transductions may be dependent, partially-dependent orindependent on Rho2p–Pck2p function, thus implying theexistence of alternative additional elements for the cell integritypathway upstream of the MAPK module.
2. Materials and methods
2.1. Strains, plasmids and growth conditions
The S. pombe strains (Table 1) were routinely grown with shaking at 28°C inYES or EMM2 medium [20] with 3% of glucose, and supplemented withadenine, leucine, histidine or uracil (100mg/liter, Sigma Chemical) dependingon the requirements for each particular strain. Transformation of yeast strainswas performed by the lithium acetate method [20]. Mutant strains were obtainedby transformation or by mating and selecting diploids in EMM2 mediumwithout supplements. Spores were purified by glusulase treatment [21] andallowed to germinate in EMM2 plus the appropriate requirements. Correctconstruction of strains was verified by PCR, and Western blot analyses (seebelow). During experiments performed with cdc25-22 thermosensitive mutantstrains, the cells were grown in YES medium to an OD600 of 0.2 at 25°C,shifted to 37°C for 3.5h, and released from the growth arrest by transfer back to25°C.
2.2. Gene disruption and epitope tagging
The rho2+ and pck2+ null mutants were obtained by entire deletion of thecorresponding coding sequences and their replacement with the KanMX6cassette by PCR-mediated strategy using plasmid pFA6a-kanMX6 as template[22]. Strains expressing genomic versions of Pmk1p fused to HA6H epitope atits C-terminus were constructed by transformation with integrative plasmid pIH-pmk1-ura [14] previously digested with BstxI at the unique site within thepmk1+coding region. The correct construction of strains was verified by PCRand/or Western blot analyses. To obtain strains expressing a C-terminal taggedversions of Pmk1p fused to the green fluorescent protein (GFP), plasmid pIL-Pmk1-GFP [14] was digested at the unique NruI site within leu1+, and the linearfragment transformed into different disrupted strains. Transformants leu1+ wereobtained and the correct integration of the Pmk1-GFP fusion plus regulatorysequences was verified as above.
2.3. Stress treatments
Experiments designed to investigate Pmk1 activation under stress wereperformed using log-phase cell cultures (OD600 of 0.5) growing at 28°C in YESmedium and the appropriate stress treatment. In glucose deprivation experi-ments, cells were grown in YES medium with 7% glucose to an OD600 of 0.5,recovered by filtration, and resuspended in the same medium without glucosebut osmotically equilibrated with 3% glycerol [23]. Hypotonic treatment wasachieved by growing cells in YES medium plus 0.8M sorbitol and resuspensionin the same medium without polyol. In all cases 30ml of culture were harvestedby centrifugation at 4°C, the cells washed with cold PBS buffer, and the yeastpellets immediately frozen in liquid nitrogen for analysis. Neither activation ofPmkp1 or Sty1p induced by centrifugation at room temperature was observed inunstressed cells [24].
2.4. Purification and detection of activated Pmk1–Ha6H followingdifferent stresses
Cell homogenates were prepared under native conditions employing chilledacid-washed glass beads and lysis buffer (10% glycerol, 50mM Tris–HCl pH

Table 1S. pombe strains used in this study
Strain Genotype a Source/reference
MM1 h+ade6-M216 A. DuránMM2 h−ade6-M210 A. DuránMI102 h+ade6-M216 pmk1::kanMX6 Madrid et al. 2006TP319-13c h− pmk1::ura4+ T. TodaMI200 h+ade6-M216 pmk1–HA6H::ura4+ Madrid et al. 2006MI700 h+ade6-M216 rho2:: kanMX6 pmk1–HA6H:: ura4+ This workGB3 h+ade6-M216 pck2:: kanMX6 pmk1 HA6H:: ura4+ This workGB29 h+ade6-M216 rho2:: kanMX6 pck2:: kanMX6 pmk1- HA6H:: ura4+ This workMI202 h+ade6-M216 mkh1:: ura4+ pmk1–HA6H:: ura4+ Madrid et al. 2006MI203 h+ade6-M216 pek1:: kanMX6 pmk1- HA6H:: ura4+ Madrid et al. 2006MI301 h−pmk1:: ura4+ pmk1-GFP::leu1 + Madrid et al. 2006GB31 h+pmk1-GFP::leu1 + rho2:: kanMX6 pmk1–HA6H:: ura4+ This workGB32 h+pmk1-GFP::leu1 + pck2:: kanMX6 pmk1–HA6H:: ura4+ This workMI212 h+ade6-M216 pmp1::kanMX6 pmk1–HA6H:: ura4+ Madrid et al. 2007GB17 h+ade6-M216 pck2:: kanMX6 pmp1:: kanMX6 pmk1–HA6H:: ura4+ This workMI600 h−cdc25-22 pmk1–HA6H:: ura4+ Madrid et al. 2007GB4 h+ade6-M216 rho2:: kanMX6 cdc25-22 pmk1–HA6H:: ura4+ This workGB16 h+ade6-M216 pck2:: kanMX6 cdc25-22 pmk1–HA6H:: ura4+ This workJCV-7 h−ade6-M216 pck1::ura4 + Lab stockGB35 h+ade6-M216 pck1::ura4+ pmk- HA6H::ura4+ This workMI800 h−ade6-M216 pmk1::kanMX6 pck1::ura4+ This workMI705 h+ade6-M216 ptc1-13myc:: KanR pyp1::HygR pmk1–HA6H:: ura4 + Madrid et al. 2007a All strains are ura4D-18 leu1-32 except MI600, which is ura4D-18.
750 G. Barba et al. / Cellular Signalling 20 (2008) 748–757
7.5, 150mM NaCl, 0.1% Nonidet NP-40, plus specific protease andphosphatase inhibitor cocktails for fungal and yeast extracts, Sigma Chemical).The lysates were cleared by centrifugation at 13,000rpm for 15min and Pmk1–HA6H was purified by using Ni2+-NTA-agarose beads (Qiagen Inc.) [21]. Thepurified proteins were resolved in 10% SDS-PAGE gels, transferred to nitro-cellulose filters (Amersham-Pharmacia), and incubated with either monoclonalmouse anti-HA (clone 12CA5, Roche Molecular Biochemicals) or polyclonalrabbit anti-phospho-p42/44 antibodies (Cell Signalling) [13]. A polyclonalrabbit anti-Cdc2 antibody (PSTAIR; Upstate Biotechnology) was usedas loading control in some experiments. The immunoreactive bands wererevealed with anti-mouse or anti-rabbit HRP-conjugated secondary antibodies(Sigma Chemical) and the Supersignal System (Pierce). Densitometricquantification of Western blot signals was performed using Molecular AnalystSoftware (Bio-Rad).
2.5. Fluorescence microscopy
A Leica DM 4000B fluorescence microscope with a 100× objective wasemployed, and images were captured with a cooled Leica DC 300F camera andIM50 software, and then imported into Adobe PhotoShop 8.0 (Adobe Systems).Small aliquots (10μl) of cells growing in YES medium to an OD600 of 0.5were loaded onto poly L-lysine coated slides or fixed with formaldehyde asdescribed [25]. Calcofluor white was employed for specific cell wall/septumstaining [25].
2.6. Plate assay of stress sensitivity for growth
Wild type and mutant strains of S. pombe were grown in YES liquidmedium to log-phase. Appropriate dilutions were spotted per duplicate on YESor EMM2 solid media containing 2% (w/v) bacto-agar (Difco) and sup-plemented with the indicated compounds. Incubation was either at 28°C for3days or at 37°C for 2–3days.
2.7. Reproducibility of results
All experiments were repeated at least three times with similar results.Representative results are shown.
3. Results
3.1. Basal Pmk1p phosphorylation is not completely abolishedin rho2Δ or pck2Δ cells
We have previously reported that in all stressing conditionsboth the basal and the stress-induced activation of Pmk1p iscompletely dependent on Mkh1p and Pek1p function, suggest-ing the existence of a linear, non-branched MAPK cascade [13].Recent work [19] has shown that GTPase Rho2p acts upstreamof protein kinase C homolog Pck2p as a regulator of the fissionyeast Pmk1p MAPK pathway. We thus analyzed the basalPmk1p activity during growth of control, rho2Δ, pck2Δ,rho2Δ pck2Δ, mkh1Δ, and pek1Δ cells expressing achromosomal HA6H-tagged version of Pmk1p by employinga p42/44 antibody [13]. As expected, MAPK activity wasundetectable in growing cells of mkh1Δ, and pek1Δ mutants(Fig. 1A). Interestingly, however, a low but reproducible basallevel of Pmk1 phosphorylation was detected in rho2Δ cells ascompared with control cells. An even lower but still recogniz-able activation was also observed in pck2Δ and rho2Δ pck2Δmutants (Fig. 1A). These results indicate that there arealternative signalling pathways independent on the control byRho2p and/or Pck2p to maintain Pmk1p basal activity.
Contrary to wild type cells, deletion of members of theRho2p–Pck2p–Pmk1p MAPK cascade enables cells to grow inthe presence of 0.2M MgCl2 plus 0.5μg/ml of the specificinhibitor of calcineurin FK506 (Fig. 1B) [19]. Because growthof fission yeast in this medium is determined by the absence ofPmk1p activity, we reasoned that an increase in the concentra-tion of either MgCl2 or FK506 would permit to distinguishbetween the components essential for MAPK activity (Mkh1p,
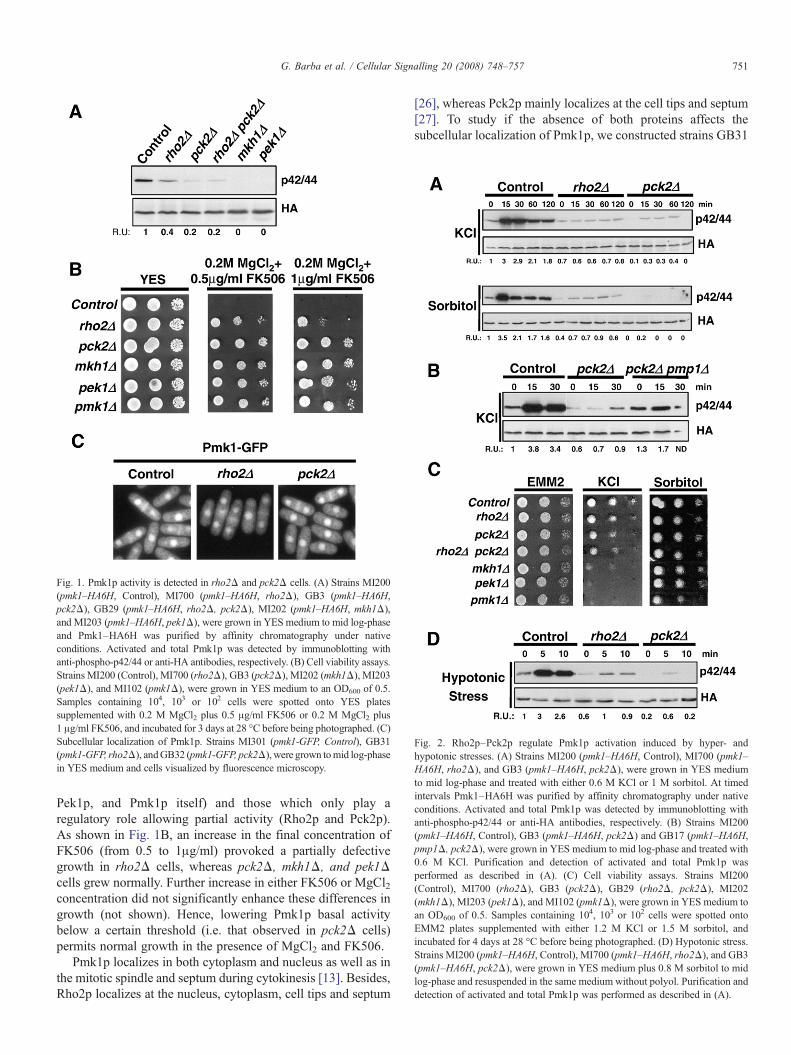
Fig. 2. Rho2p–Pck2p regulate Pmk1p activation induced by hyper- andhypotonic stresses. (A) Strains MI200 (pmk1–HA6H, Control), MI700 (pmk1–HA6H, rho2Δ), and GB3 (pmk1–HA6H, pck2Δ), were grown in YES mediumto mid log-phase and treated with either 0.6 M KCl or 1 M sorbitol. At timedintervals Pmk1–HA6H was purified by affinity chromatography under nativeconditions. Activated and total Pmk1p was detected by immunoblotting withanti-phospho-p42/44 or anti-HA antibodies, respectively. (B) Strains MI200(pmk1–HA6H, Control), GB3 (pmk1–HA6H, pck2Δ) and GB17 (pmk1–HA6H,pmp1Δ, pck2Δ), were grown in YES medium to mid log-phase and treated with0.6 M KCl. Purification and detection of activated and total Pmk1p wasperformed as described in (A). (C) Cell viability assays. Strains MI200(Control), MI700 (rho2Δ), GB3 (pck2Δ), GB29 (rho2Δ, pck2Δ), MI202(mkh1Δ), MI203 (pek1Δ), and MI102 (pmk1Δ), were grown in YES medium toan OD600 of 0.5. Samples containing 104, 103 or 102 cells were spotted ontoEMM2 plates supplemented with either 1.2 M KCl or 1.5 M sorbitol, andincubated for 4 days at 28 °C before being photographed. (D) Hypotonic stress.Strains MI200 (pmk1–HA6H, Control), MI700 (pmk1–HA6H, rho2Δ), and GB3(pmk1–HA6H, pck2Δ), were grown in YES medium plus 0.8 M sorbitol to midlog-phase and resuspended in the same medium without polyol. Purification anddetection of activated and total Pmk1p was performed as described in (A).
Fig. 1. Pmk1p activity is detected in rho2Δ and pck2Δ cells. (A) Strains MI200(pmk1–HA6H, Control), MI700 (pmk1–HA6H, rho2Δ), GB3 (pmk1–HA6H,pck2Δ), GB29 (pmk1–HA6H, rho2Δ, pck2Δ), MI202 (pmk1–HA6H, mkh1Δ),and MI203 (pmk1–HA6H, pek1Δ), were grown in YES medium to mid log-phaseand Pmk1–HA6H was purified by affinity chromatography under nativeconditions. Activated and total Pmk1p was detected by immunoblotting withanti-phospho-p42/44 or anti-HA antibodies, respectively. (B) Cell viability assays.Strains MI200 (Control), MI700 (rho2Δ), GB3 (pck2Δ), MI202 (mkh1Δ), MI203(pek1Δ), and MI102 (pmk1Δ), were grown in YES medium to an OD600 of 0.5.Samples containing 104, 103 or 102 cells were spotted onto YES platessupplemented with 0.2 M MgCl2 plus 0.5 μg/ml FK506 or 0.2 M MgCl2 plus1 μg/ml FK506, and incubated for 3 days at 28 °C before being photographed. (C)Subcellular localization of Pmk1p. Strains MI301 (pmk1-GFP, Control), GB31(pmk1-GFP, rho2Δ), andGB32 (pmk1-GFP, pck2Δ), were grown tomid log-phasein YES medium and cells visualized by fluorescence microscopy.
751G. Barba et al. / Cellular Signalling 20 (2008) 748–757
Pek1p, and Pmk1p itself) and those which only play aregulatory role allowing partial activity (Rho2p and Pck2p).As shown in Fig. 1B, an increase in the final concentration ofFK506 (from 0.5 to 1μg/ml) provoked a partially defectivegrowth in rho2Δ cells, whereas pck2Δ, mkh1Δ, and pek1Δcells grew normally. Further increase in either FK506 or MgCl2concentration did not significantly enhance these differences ingrowth (not shown). Hence, lowering Pmk1p basal activitybelow a certain threshold (i.e. that observed in pck2Δ cells)permits normal growth in the presence of MgCl2 and FK506.
Pmk1p localizes in both cytoplasm and nucleus as well as inthe mitotic spindle and septum during cytokinesis [13]. Besides,Rho2p localizes at the nucleus, cytoplasm, cell tips and septum
[26], whereas Pck2p mainly localizes at the cell tips and septum[27]. To study if the absence of both proteins affects thesubcellular localization of Pmk1p, we constructed strains GB31

Fig. 3. Partial role of Pck2p–Rho2p function in Pmk1p activation induced bycell wall stress or glucose deprivation. (A) Strains MI200 (pmk1–HA6H,Control), MI700 (pmk1–HA6H, rho2Δ), and GB3 (pmk1–HA6H, pck2Δ), weregrown in YES medium to mid log-phase and treated with either 15 mMCaffeineor 1 mg/ml Calcofluor. At timed intervals Pmk1–HA6H was purified by affinitychromatography under native conditions. Activated and total Pmk1p wasdetected by immunoblotting with anti-phospho-p42/44 or anti-HA antibodies,respectively. (B) Strains MI200 (pmk1–HA6H, Control), and GB29 (pmk1–HA6H, rho2Δ, pck2Δ), were grown in YES medium to mid log-phase andtreated with 15 mM Caffeine. Purification and detection of activated and totalPmk1p was performed as described in (A). (C) Cell viability assays. StrainsMI200 (Control), MI700 (rho2Δ), GB3 (pck2Δ), GB29 (rho2Δ, pck2Δ),MI202 (mkh1Δ), MI203 (pek1Δ), and MI102 (pmk1Δ), were grown in YESmedium to an OD600 of 0.5. Samples containing 104, 103 or 102 cells werespotted onto YES plates supplemented with either 5 mM Caffeine or 1 mg/mlCalcofluor, and incubated for 3 days at 28 °C before being photographed. (D)Glucose deprivation. Strains MI200 (pmk1–HA6H, Control), MI700 (pmk1–HA6H, rho2Δ), and GB3 (pmk1–HA6H, pck2Δ), were grown in YES mediumwith 7% glucose to an OD600 of 0.5, and resuspended in the same mediumwithout glucose and osmotically equilibrated with 3% glycerol. Purification anddetection of activated and total Pmk1p was performed as described in (A).
752 G. Barba et al. / Cellular Signalling 20 (2008) 748–757
and GB32, expressing a C-terminal tagged version of Pmk1fused to GFP in a rho2Δ or pck2Δ background, respectively.Results in Fig. 1C show that the absence of either protein didnot alter the distribution pattern of Pmk1-GFP.
3.2. Pmk1p activation by hypertonic and hypotonic stresses isregulated through Rho2p–Pck2p
The existence of a low but measurable basal MAPK activityin rho2Δ and pck2Δ cells prompted us to identify the signallingnetworks that modulate Pmk1p activation in response to avariety of stresses and their dependence on Rho2p and/or Pck2pfor signalling. The level of Pmk1 phosphorylation increasesquickly and transiently when S. pombe cells are subjected to asalt-induced osmostress (NaCl or KCl) or shifted to hypertonicmedium (sorbitol) [13]. As shown in Fig. 2A, the presence ofRho2p and/or Pck2p was critical for Pmk1p activation duringtreatment with 0.6M KCl or 1M sorbitol, suggesting that bothproteins play an integral role in the signalling cascade thatmodulates MAPK activation under osmotic stress. Moreover,the increased MAPK activity in pck2Δ or rho2Δ cells byadditional deletion of pmp1+, which encodes a dual specificityphosphatase downregulating Pmk1p basal activity [18], was notsufficient to significantly enhance Pmk1p phosphorylationunder osmostress (Fig. 2B). These results confirm that anactivation signal transduced from Rho2p–Pck2p to the MAPKmodule is needed when S. pombe is subjected to a hypertonicstress. We also determined whether the absence of any of themembers of Pmk1p cascade would affect cell viability underhypertonic stress. As indicated in Fig. 2C, only mkh1Δ, pek1Δand pmk1Δ cells were clearly hypersensitive to growth in thepresence of 1M KCl, whereas all mutants tested grew similar towild type cells in medium containing 1M sorbitol. We alsoanalyzed Pmk1p activation in control, rho2Δ and pck2Δ cellssubjected to a hypotonic stress, which induces a very quick andtransient phosphorylation of the MAPK in control cells [13](Fig. 2D). Like under osmostress, Rho2p and/or Pck2p deletionstrongly decreased Pmk1p activation during hypotonic condi-tions (Fig. 2D).
3.3. Activation of Pmk1p induced by cell wall stress or glucosearrest is partially dependent on Pck2p–Rho2p function
Pmk1p undergoes a clear and delayed activation by treatmentwith caffeine or calcofluor white, whose effects have beenrelated to changes in the architecture and biosynthesis of theyeast cell wall [13,28,29]. A comparative analysis of Pmk1pphosphorylation in control, pck2Δ, or rho2Δ cells treated withthe above compounds, showed that Pmk1p activation was stillevident in the absence of Rho2p or Pck2p, although with lowerintensity than in control cells (Fig. 3A). The double deletion ofrho2+ and pck2+ did not enhance the low Pmk1p activationobserved in the corresponding single disruptants after treatmentwith caffeine (Fig. 3B), calcofluor or sodium vanadate (notshown). Similar to control cells, all these mutants displayed asimilar ability for growth in the presence of caffeine, whereaspek1Δ and pmk1Δ cells showed a partially affected growth in
the presence of calcofluor (Fig. 3C). Glucose depletion induces adelayed increase in Pmk1p activation in control cells maintainedin osmotically stabilized medium [13]. However, as Fig. 3D

753G. Barba et al. / Cellular Signalling 20 (2008) 748–757
indicates, glucose withdrawal from the medium induced a partialPmk1p activation in pck2Δ or rho2Δ cells as compared withcontrol cells. As a whole, these results suggest that the activationof Pmk1p induced by cell wall alteration or by glucose depletionis transduced by at least two different pathways, one of theminvolving Rho2p and Pck2p.
3.4. Diverse pathways regulate the oxidative stress dependentactivation of Pmk1p
Pmk1p is moderately activated by high concentrations(N1mM) of hydrogen peroxide in a rather rapid and maintainedmanner [14] (Fig. 4A). Interestingly, deletion of rho2+ or pck2+
genes did not affect significantly Pmk1p activation by hydrogenperoxide, as compared to the MAPK activation in control cells(Fig. 4A). Treatment with different pro-oxidants, like DEM orParaquat, also induces a clear Pmk1p activation [13]. However,in clear contrast to the activation elicited by hydrogen peroxide,
Fig. 4. Activation of Pmk1p induced by oxidative stress is regulated through diversrho2Δ), and GB3 (pmk1–HA6H, pck2Δ), were grown in YES medium to mid log-pParaquat. At timed intervals Pmk1–HA6H was purified by affinity chromatogrimmunoblotting with anti-phospho-p42/44 or anti-HA antibodies, respectively. (B)GB29 (rho2Δ, pck2Δ), MI202 (mkh1Δ), MI203 (pek1Δ), and MI102 (pmk1Δ), wercells were spotted onto YES plates supplemented with either 2 mM hydrogen peroxidbeing photographed.
Pmk1p activation by DEM was strongly dependent on Rho2pand Pck2p (Fig. 4A), and that induced by Paraquat was partiallysuppressed by rho2+ deletion and absent in pck2Δ cells(Fig. 4A). Moreover, a plate sensitivity assay for growth in thepresence of the above compounds revealed that pck2Δ cellsdisplayed growth sensitivity in the presence of DEM or Paraquatthat was similar to that of mkh1Δ, pek1Δ and pmk1Δ cellswhereas, on the contrary, cell viability was not modified in thepresence of hydrogen peroxide (Fig. 4B). Hence, the oxidativestress-induced activation of Pmk1p in fission yeast is transduceddifferently depending on the nature of the triggering stressor.
3.5. Role of Pck2p in Pmk1p activation at high temperature
Thermal stress promotes a delayed activation of Pmk1p infission yeast [13]. The kinetics and intensity of Pmk1pphosphorylation after transfer of yeast cultures from 30 to 40°Cwas not significantly affected by deletion of rho2+ gene, whereas
e pathways. (A) Strains MI200 (pmk1–HA6H, Control), MI700 (pmk1–HA6H,hase and treated with either 5 mM hydrogen peroxide, 4 mM DEM, or 4 mMaphy under native conditions. Activated and total Pmk1p was detected byCell viability assays. Strains MI200 (Control), MI700 (rho2Δ), GB3 (pck2Δ),e grown in YES medium to an OD600 of 0.5. Samples containing 104, 103 or 102
e, 0.5 mM DEM, or 0.5 mM Paraquat, and incubated for 3 days at 28 °C before
754 G. Barba et al. / Cellular Signalling 20 (2008) 748–757
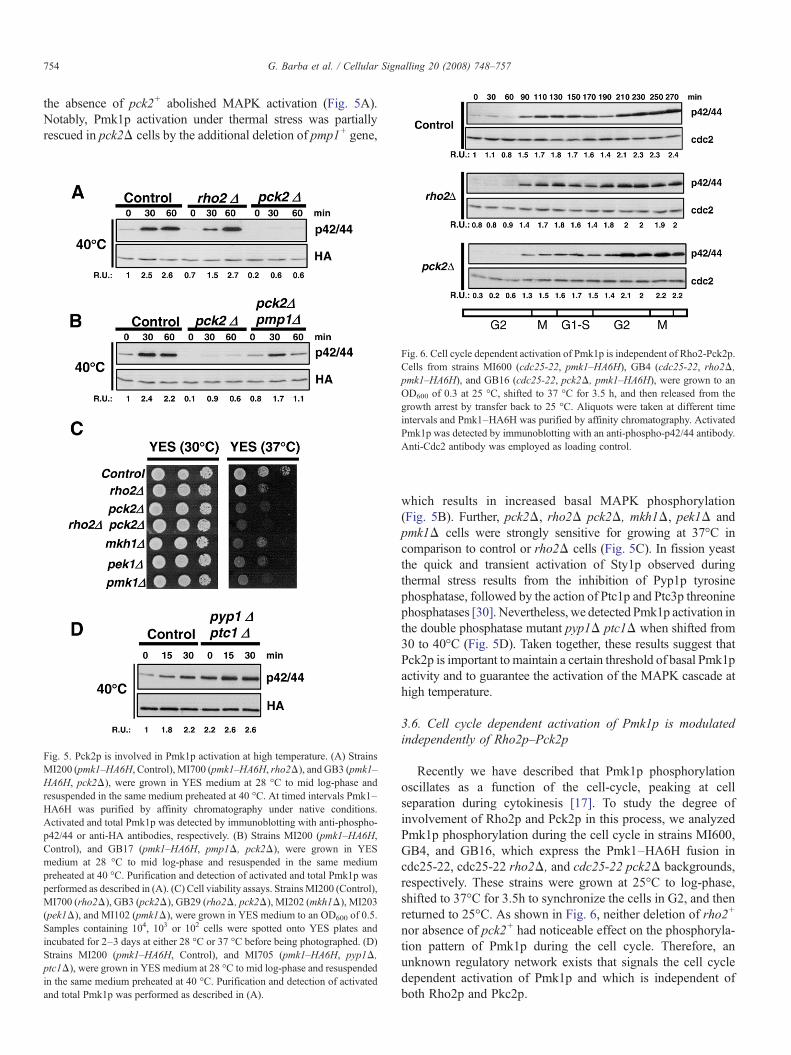
the absence of pck2+ abolished MAPK activation (Fig. 5A).Notably, Pmk1p activation under thermal stress was partiallyrescued in pck2Δ cells by the additional deletion of pmp1+ gene,
Fig. 5. Pck2p is involved in Pmk1p activation at high temperature. (A) StrainsMI200 (pmk1–HA6H, Control), MI700 (pmk1–HA6H, rho2Δ), and GB3 (pmk1–HA6H, pck2Δ), were grown in YES medium at 28 °C to mid log-phase andresuspended in the same medium preheated at 40 °C. At timed intervals Pmk1–HA6H was purified by affinity chromatography under native conditions.Activated and total Pmk1p was detected by immunoblotting with anti-phospho-p42/44 or anti-HA antibodies, respectively. (B) Strains MI200 (pmk1–HA6H,Control), and GB17 (pmk1–HA6H, pmp1Δ, pck2Δ), were grown in YESmedium at 28 °C to mid log-phase and resuspended in the same mediumpreheated at 40 °C. Purification and detection of activated and total Pmk1p wasperformed as described in (A). (C) Cell viability assays. StrainsMI200 (Control),MI700 (rho2Δ), GB3 (pck2Δ), GB29 (rho2Δ, pck2Δ), MI202 (mkh1Δ), MI203(pek1Δ), and MI102 (pmk1Δ), were grown in YES medium to an OD600 of 0.5.Samples containing 104, 103 or 102 cells were spotted onto YES plates andincubated for 2–3 days at either 28 °C or 37 °C before being photographed. (D)Strains MI200 (pmk1–HA6H, Control), and MI705 (pmk1–HA6H, pyp1Δ,ptc1Δ), were grown in YES medium at 28 °C to mid log-phase and resuspendedin the same medium preheated at 40 °C. Purification and detection of activatedand total Pmk1p was performed as described in (A).
Fig. 6. Cell cycle dependent activation of Pmk1p is independent of Rho2-Pck2p.Cells from strains MI600 (cdc25-22, pmk1–HA6H), GB4 (cdc25-22, rho2Δ,pmk1–HA6H), and GB16 (cdc25-22, pck2Δ, pmk1–HA6H), were grown to anOD600 of 0.3 at 25 °C, shifted to 37 °C for 3.5 h, and then released from thegrowth arrest by transfer back to 25 °C. Aliquots were taken at different timeintervals and Pmk1–HA6H was purified by affinity chromatography. ActivatedPmk1p was detected by immunoblotting with an anti-phospho-p42/44 antibody.Anti-Cdc2 antibody was employed as loading control.
which results in increased basal MAPK phosphorylation(Fig. 5B). Further, pck2Δ, rho2Δ pck2Δ, mkh1Δ, pek1Δ andpmk1Δ cells were strongly sensitive for growing at 37°C incomparison to control or rho2Δ cells (Fig. 5C). In fission yeastthe quick and transient activation of Sty1p observed duringthermal stress results from the inhibition of Pyp1p tyrosinephosphatase, followed by the action of Ptc1p and Ptc3p threoninephosphatases [30]. Nevertheless, we detected Pmk1p activation inthe double phosphatase mutant pyp1Δ ptc1Δ when shifted from30 to 40°C (Fig. 5D). Taken together, these results suggest thatPck2p is important to maintain a certain threshold of basal Pmk1pactivity and to guarantee the activation of the MAPK cascade athigh temperature.
3.6. Cell cycle dependent activation of Pmk1p is modulatedindependently of Rho2p–Pck2p
Recently we have described that Pmk1p phosphorylationoscillates as a function of the cell-cycle, peaking at cellseparation during cytokinesis [17]. To study the degree ofinvolvement of Rho2p and Pck2p in this process, we analyzedPmk1p phosphorylation during the cell cycle in strains MI600,GB4, and GB16, which express the Pmk1–HA6H fusion incdc25-22, cdc25-22 rho2Δ, and cdc25-22 pck2Δ backgrounds,respectively. These strains were grown at 25°C to log-phase,shifted to 37°C for 3.5h to synchronize the cells in G2, and thenreturned to 25°C. As shown in Fig. 6, neither deletion of rho2+
nor absence of pck2+ had noticeable effect on the phosphoryla-tion pattern of Pmk1p during the cell cycle. Therefore, anunknown regulatory network exists that signals the cell cycledependent activation of Pmk1p and which is independent ofboth Rho2p and Pkc2p.

Fig. 7. Pck1p is a negative regulator of Pmk1p activity. (A) Left panel; strainsMI200 (pmk1–HA6H, Control) and GB35 (pmk1–HA6H, pck1Δ) were grown inYES medium to mid log-phase and treated with 0.6 M KCl. Right panel; strainsMI200 and GB35 were grown in YES medium to mid log-phase. In both casesPmk1–HA6H was purified by affinity chromatography under native conditions,and activated and total Pmk1p was detected by immunoblotting with anti-phospho-p42/44 or anti-HA antibodies, respectively. (B) Chloride sensitivityassays. Strains MI200 (Control), MI102 (pmk1Δ), GB35 (pck1Δ), and MI800(pmk1Δ, pck1Δ), were grown in YES medium to an OD600 of 0.5. Samplescontaining 105, 104, 103 or 102 cells were spotted onto YES plates supplementedwith 0.2 MMgCl2 and incubated for 3 days at 28 °C before being photographed.
Fig. 8. Proposedmodel showing different stress signals involved in the activationof the Pmk1p cell integrity pathway in S. pombe and their dependence/independence of Rho2p and Pck2p for transduction. Question marks indicateunknown elements involved in the regulation of Pmk1p basal activity in additionto Rho2p and Pck2p. Solid arrows indicate direct interactions.
755G. Barba et al. / Cellular Signalling 20 (2008) 748–757
3.7. Pck1p negatively regulates Pmk1p basal activity
After determining the role of Pck2p in Pmk1p activationunder different environmental situations, we focused ourattention onto Pck1p, the other PKC ortholog in fission yeastand potential candidate to positively modulate MAPK activity.However, contrary to our prediction, pck1+ deletion induced aclear increase in both the basal and the osmostress-inducedactivation of Pmk1p as compared to control cells (Fig. 7A). Infission yeast the phosphorylation state of Pmk1p has a dramaticeffect on chloride homeostasis, and Pmk1p hyperactivationelicits a strong sensitivity to this anion [18]. As demonstrated inFig. 7B, cells lacking pck1+ did not grow in the presence of0.2M MgCl2, but the chloride sensitivity of this mutant wasrescued by additional deletion of pmk1+, suggesting thatPmk1p hyperactivation is responsible for such behavior(Fig. 7B). Taken together, these results indicate that, contraryto Pck2p, Pck1p acts as a negative regulator of the cell integritypathway in fission yeast.
4. Discussion
The finding that various pathways regulate Pmk1p phos-phorylation in S. pombe depending on the nature of the stimulusis somehow surprising (Fig. 8). Maintenance of the integrity ofthe MAPK cascade is essential to allow Pmk1p activation underall stressing circumstances, but we found that Pmk1pphosphorylation is still detected to some extent in the absenceof either Rho2p GTPase or the PKC-ortholog Pck2p, which are
the only known upstream activators of the cascade [19]. Thistype of multiple regulation is in clear contrast with the control ofthe cell integrity pathway in the budding yeast S. cerevisiae,where Pkc1p (Pck2p ortholog) is absolutely required for activityof Mpk1p/Slt2p (Pmk1p ortholog) [31]. Therefore, our resultsdisclose that at least one more protein kinase in fission yeast isinvolved in the activation of the Pmk1p MAPK cascade, inaddition to Pck2p. The identity of this alternative component isat present unknown. Its role as a regulator of the MAPK cascadeappears less relevant than Pck2p, since basal Pmk1p activity israther low in pck2Δ cells as compared to control cells. Also,there must exist at least another element channeling Pmk1pactivation through Pck2p alternatively to Rho2p, because thedeficit in basal MAPK kinase activity observed in the absenceof this GTPase was less evident than in pck2Δ cells. Oneattractive candidate to perform such a role is Rho1p GTPase,whose active form (GTP bound) interacts with both Pck1p andPck2p [32].
The observation of a detectable MAPK phosphorylation inthe absence of Rho2p and Pck2p allowed us to analyze theactive role of both proteins in Pmk1p activation under a varietyof stresses. The results clearly indicate that Pmk1p activationsinduced by hypo- or hypertonic stresses, and that achieved aftertreatment with the oxidant agent DEM are fully dependent onthe signalling provided by Rho2p–Pck2p (Fig. 8). In S.cerevisiae hypotonic stress modulates Slt2p phosphorylationdownstream of Rho1p-Pkc1p [33], which suggests that buddingand fission yeasts sense this particular stress through verydifferent mechanisms. Contrariwise, activation of Slt2p underhyperosmotic stress is also dependent on Rho1p and Pkc1p, theRho2p and Pck2p orthologs, and shows a very slow kinetics ascompared to Pmk1p. Importantly, although Pmk1p activation

756 G. Barba et al. / Cellular Signalling 20 (2008) 748–757
induced by either sorbitol or KCl was very similar, disruption ofthe MAPK cascade prompted a severe growth defect only in thepresence of potassium ions. Hence, Pmk1p activation inducedby hypertonic stress appears related to an increased externalosmolarity, independently on the nature of the stimulus, where-as basal MAPK activity is involved in the maintenance ofpotassium homeostasis. Also, it has been shown that alkalyni-zation of the growth medium induces a quick and transientactivation of MAPK Slt2p in S. cerevisiae [34]. We havesimilarly observed that cells of S. pombe growing in richmedium (pH 5.4) show a clear increase in Pmk1p phosphoryla-tion after transfer to the same medium buffered at pH 7 orabove. Like activation by hypertonic or hypotonic stresses, thePmk1p activation under alkaline pH appears totally dependenton Rho2p–Pck2p function (unpublished results).
In the budding yeast S. cerevisiae, a family of five cellsurface sensors (Wsc1-3p, Mid2p and Mtl1p) are involved inthe signalling to Rho1p of the cell wall status in order to activatethe cell integrity pathway [35]. In S. pombe, however, plasmamembrane receptors activating the Pmk1p MAPK cascade viaRho2p have not yet been characterized. One potential candidateto fulfill such a role would be Wsc1p, a serine-rich membraneprotein featuring a signal peptide and a WSC domain which isO-mannosylated in vivo [36]. However, we have found thatneither the kinetics nor the intensity of Pmk1p activation undercell wall stress is significantly affected by deletion of wsc1+
(unpublished results). It is tempting to speculate that Wsc1p isnot a member of the cell integrity pathway in fission yeast orthat other putative plasma membrane receptors perform asimilar role in a redundant fashion.
Together with the occurrence of stress-induced activations ofPmk1p that are fully dependent on Rho2p–Pck2p function, ourresults reveal other MAPK activations which only partially relyupon both proteins. Such is the case of the Pmk1p activationinduced by cell wall perturbations with caffeine or calcofluor, orthat triggered by glucose depletion. Again, this outcome sharplycontrasts with the critical role played in S. cerevisiae by Pck1pin the cell wall-mediated stress activation of Slt2p [28]. In thefission yeast, glucose withdrawal promotes a cellular arrest atG2 phase and profound morphological changes similar to thoseobserved during the transition to stationary phase of growth[37]. Because cell wall stress and glucose depletion cause asimilar Pmk1p activation, it seems likely that the subjacenttriggering stimulus after glucose withdrawal be related to cellwall remodeling during arrest.
We also demonstrate that the activation of Pmk1p induced byhydrogen peroxide, high temperature, or that occurring duringcell cycle is not transduced through Rho2p–Pck2p. In the firstcase, our findings are somewhat intriguing since Rho2p and/orPck2p mediate Pmp1p activation induced by other oxidants likeDEM or Paraquat. Moreover, in budding yeast, Pkc1p isrequired for Slt2p activation in response to hydrogen peroxideand oxidants like diamide, and these upper elements of the cellintegrity pathway are also required for survival and adaptationto the oxidative stress provoked by such compounds [38]. Onthe contrary, in fission yeast, the cell viability of MAPK kinasecascade mutants was not significantly affected in presence of
hydrogen peroxide. As a whole, our data provide evidence forthe existence of a specific mechanism to modulate Pmk1pactivity in response to this oxidant whose functional andbiological significance remains unknown.
Several evidences anticipated that Pmk1p activation inducedby thermal stress might be due to the inhibition of MAPKphosphatases: (i) Pyp1p and Ptc1p downregulate both Sty1pand Pmk1p [17]; (ii) Sty1p activation during thermal stressresults from the inhibition of Pyp1p tyrosine phosphatase [30],and (iii) the activation of Pmk1p by high temperatures takesplace downstream of Pck2p. However, the kinetics of Pmk1pactivation is clearly slower than in the case of Sty1p [13,30],and the heat stress also induced MAPK activation in the doublephosphatase mutant pyp1Δ ptc1Δ. These results stronglysuggest that heat stress promote Pmk1p activation by actingdirectly on the MAPK cascade itself, as has been proposed forthermal activation of Slt2p in budding yeast [33]. Nevertheless,Pck2p function is critical to maintain a sufficient Pmk1p basalactivity and to allow MAPK activation at this condition.
While Pck2p has been recently established as an upstreamregulator of the MAPK Pmk1p cascade, Pck1p appearsinvolved in cell integrity signalling by acting independentlyand in parallel to Pmk1p [19]. Unexpectedly, we have foundthat Pck1p not only does not positively regulate Pmk1pactivation but, on the contrary, it behaves as a negative regulatorof the cascade, since basal Pmk1p activity is increased inpmk1Δ cells. Moreover, this negative regulation is biologicallysignificant, because the chloride sensitivity of pck1Δ cells isdue to Pmk1p hyperactivation. Hence, in fission yeast Pck1pand Pck2p antagonistically regulate Pmk1p activity, eitherpositively (Pck2p) or negatively (Pck1p). Pull-down assayshave shown that, contrary to Pck2p, Pck1p and Mkh1p do notassociate in vivo [19]. Therefore, it seems very likely thatPck1p-mediated downregulation of the Pmk1p MAPK cascademight be an indirect event (i.e. not exerted by direct associationwith components of the cascade), and that the cell wall integritydefects related to the absence of Pck1p might in turn beresponsible for Pmk1p hyperactivation. In any case, theseresults highlight the complex interactions established in fissionyeast between the Pmk1p cell integrity pathway and membersof the PKC family of proteins.
In conclusion, our data provide evidence for the existence inS. pombe of a complex regulatory network modulating both thebasal level and the activation of the Pmk1p MAPK cascade inresponse to a variety of stressful stimuli. In this organism,hypertonic, hypotonic, and alkaline stresses are transduced byRho2p GTPase via Pck2p. However, other stimuli like glucosearrest, or the presence of cell wall perturbing agents arechannelled to Pmk1p either through Rho2p–Pck2p or analternative pathway. Finally, Pmk1p activation during cellseparation or treatment with hydrogen peroxide is fullyindependent on Rho2p–Pck2p function, whereas activation byheat stress relies upon the basal MAPK activity provided byPck2p function. Identification of these alternative regulatoryelements deserves further characterization to improve ourknowledge on the architecture of MAPK cascades in eukaryotecells.

757G. Barba et al. / Cellular Signalling 20 (2008) 748–757
Acknowledgements
We thank A. Durán, P. Pérez and T. Toda for kind supply ofyeast strains, and to F. Garro and V. Hernandis for excellenttechnical assistance.M.M. andA.N. are predoctoral fellows fromMinisterio de Educación, Cultura y Deporte (FPU) and fromFundación Séneca (Spain), respectively. This workwas supportedby grants BFU2005-01401/BMC from MEC to J.C., and 00475/PI/04 from Fundación Séneca (Región de Murcia), Spain.
References
[1] C.J. Marshall, Cell 80 (1995) 179.[2] A.J. Waskiewicz, J.A. Cooper, Curr. Opin. Cell Biol. 7 (1995) 798.[3] T. Toda, M. Shimanuki, M. Yanagida, Genes Dev. 5 (1991) 60.[4] S. Hohmann, Microbiol. Mol. Biol. Rev. 66 (2002) 300.[5] K. Shiozaki, P. Russell, Nature 378 (1995) 739.[6] T. Toda, S. Dhut, G. Superti-Furga, Y. Gotoh, Y.E. Nishida, R. Sugiura,
T. Kuno, Mol. Cell. Biol. 16 (1996) 6752.[7] T. Zaitsevskaya-Carter, J.A. Cooper, EMBO J. 16 (1997) 1318.[8] P.P. Roux, J. Blenis, Microbiol. Mol. Biol. Rev. 68 (2004) 320.[9] A.S. Sengar, N.A.Markley, N.J.Marini, D.Young,Mol. Cell. Biol. 17 (1997)
3508.[10] R. Sugiura, T. Toda, S. Dhut, H. Shuntoh, T. Kuno, Nature 399 (1999) 479.[11] R. Loewith, A. Hubberstey, D. Young, J. Cell Sci. 113 (2000) 153.[12] N. Bone, J.B. Millar, T. Toda, J. Armstrong, Curr. Biol. 29 (1998) 135.[13] M.Madrid, T. Soto, H.K. Khong, A. Franco, J. Vicente, P. Pérez, M. Gacto,
J. Cansado, J. Biol. Chem. 281 (2006) 2033.[14] K. Shiozaki, P. Russell, Genes Dev. 10 (1996) 2276.[15] M.G. Wilkinson, M. Samuels, T. Takeda, M.W. Toone, J. Shieh, T. Toda,
J.B.A. Millar, N. Jones, Genes Dev. 10 (1996) 2289.[16] H. Takada, M. Nishimura, Y. Asayama, Y. Mannse, S. Ishiwata, A. Kita,
A. Doi, A. Nishida, N. Kai, S. Moriuchi, H. Tohda, Y. Giga-Hama, T. Kuno,R. Sugiura, Mol. Biol. Cell (2007) [Electronic publication ahead of print].
[17] M. Madrid, A. Nuñez, T. Soto, J. Vicente-Soler, M. Gacto, J. Cansado,Mol. Biol. Cell 18 (2007) 4405.
[18] R. Sugiura, T. Toda, H. Shuntoh, M. Yanagida, T. Kuno, EMBO J. 17 (1998)140.
[19] Y.Ma, T. Kuno, A. Kita, Y. Asayama, R. Sugiura, Mol. Biol. Cell 17 (2006)5028.
[20] S. Moreno, A. Klar, P. Nurse, Methods Enzymol. 194 (1991) 795.[21] T. Soto, F.F. Beltrán, V. Paredes, M. Madrid, J.B.A. Millar, J. Vicente-
Soler, J. Cansado, M. Gacto, Eur. J. Biochem. 269 (2002) 1.[22] J. Bahler, J.Q.Wu,M.S. Longtine,N.G. Shah, A.McKenzie III, A.B. Steever,
A. Wach, P. Philippsen, J.R. Pringle, Yeast 14 (1998) 943.[23] M. Madrid, T. Soto, A. Franco, V. Paredes, J. Vicente, E. Hidalgo, M. Gacto,
J. Cansado, J. Biol. Chem. 279 (2004) 41594.[24] T. Soto, A. Nuñez, M. Madrid, J. Vicente, M. Gacto, J. Cansado,
Microbiology 153 (2007) 1519.[25] C. Alfa, P. Fantes, J. Hyams, M. Mcleod, E. Warbrick (Eds.), Experiments
with Fission Yeast, Cold Spring Harbor Laboratory Press, Cold SpringHarbor, New York, 1993, p. 16.
[26] D. Hirata, K. Nakano, M. Fukui, H. Takenaka, T. Miyakawa, I. Mabuchi,J. Cell Sci. 111 (1998) 149.
[27] A. Matsuyama, R. Arai, Y. Yashiroda, A. Shirai, A. Kamata, S. Sekido,Y. Kobayashi, A. Hashimoto, M. Hamamoto, Y. Hiraoka, S. Horinouchi,M. Yoshida, Nat. Biotechnol. 24 (2006) 841.
[28] H. Martín, J.M. Rodriguez-Pachon, C. Ruiz, C. Nombela, M. Molina,J. Biol. Chem. 275 (2000) 1511.
[29] P. Pérez, J.C. Ribas, Methods 33 (2004) 245.[30] A.N. Nguyen, K. Shiozaki, Genes Dev. 13 (1999) 1653.[31] Y. Kamada, U.S. Jung, J. Piotrowski, D.E. Levin, Genes Dev. 9 (1995)
1559.[32] M. Arellano, M.H. Valdivieso, T.M. Calonge, P.M. Coll, A. Duran,
P. Perez, J. Cell Sci. 112 (1999) 3569.[33] J.C. Harrison, T.R. Zyla, E.S. Bardes, D.J. Lew, J. Biol. Chem. 279 (2004)
2616.[34] R. Serrano, H. Martín, A. Casamayor, J. Ariño, J. Biol. Chem. 281 (2006)
39785.[35] D.E. Levin, Microbiol. Mol. Biol. Rev. 69 (2005) 262.[36] T. Willer, M. Brandl, M. Sipiczki, S. Strahl, Mol. Microbiol. 57 (2005)
156.[37] G. Costello, L. Rodgers, D. Beach, Curr. Genet. 11 (1986) 119.[38] F.Vilella, E.Herrero, J. Torres,M.A.De laTorre-Ruiz, J.Biol.Chem. 280 (2005)
9149.
Copyright © 2022 FDOKUMEN




















