
Ras1 controls pheromone expression and response during mating in Cryptococcus neoformans
Upload
independentCategory
view
0download
0
RESEARCH ARTICLE
Mark Didmon Æ Kevin Davis Æ Peter Watson
Graham Ladds Æ Peter Broad Æ John Davey
Identifying regulators of pheromone signallingin the fission yeast Schizosaccharomyces pombe
Received: 23 January 2002 / Accepted: 8 May 2002 / Published online: 25 June 2002� Springer-Verlag 2002
Abstract The rate and extent of a cell’s response to anextracellular stimulus is influenced by regulators that acton the intracellular signalling machinery. Although notdirectly involved in propagating the intracellular signal,regulators control the activity of the proteins thattransmit the signals. To understand this aspect of cellsignalling, we studied the pheromone-response pathwayin the fission yeast Schizosaccharomyces pombe, a rela-tively simple signalling system in a genetically tractableorganism. Here, we describe the development of yeaststrains containing ura4 and lacZ reporter genes underthe control of the pheromone-regulated sxa2 promoterand the use of these strains to isolate mutants defective intheir ability to regulate signalling. Several different typesof mutant were identified. Some mutants were defectivein proteins already known to regulate the pheromone-signalling pathway (Rgs1, Map1, Map2). Our approachalso identified the MAP kinase phosphatase Pmp1 as aregulator of the pheromone-response pathway. Al-though previously shown to regulate other MAP kinasepathways in Sz. pombe, this is the first demonstration ofa role for Pmp1 in pheromone signalling.
Keywords Pheromone Æ Pmp1 Æ Signalling Æ Yeast
Introduction
The growth, differentiation and general metabolicactivity of most cells can be influenced by exposure to
extracellular ligands; and stimulation of the appropriatereceptors at the cell surface activates the intracellularmachinery responsible for bringing about changes in cellbehaviour. Persistent stimulation does not, however,generally bring about a continuous response, as the celladapts to the presence of the stimulus and becomes de-sensitised. Such desensitisation is a normal physiologicalprocess that both limits the extent of the response andallows cells to recover their resting state. The molecularbasis for desensitisation in eukaryotic cells is complexand involves a variety of processes that can affect thesignalling machinery at almost every level (for reviews,see Freedman and Lefkowitz 1996; Bohm et al. 1997;Keyse 2000).
To gain further insights into the desensitisation pro-cess, we studied the pheromone-response pathway in thefission yeast Schizosaccharomyces pombe (for a review,see Davey 1998). Conjugation between two haploidSz. pombe cells involves the reciprocal exchange of dif-fusible mating pheromones: M-cells (mating type minus)release M-factor and respond to the P-factor released byP-cells (mating type plus). Binding the pheromones tospecific seven-span receptors at the cell surface activatesan intracellular signalling pathway and leads to changesin cell behaviour in readiness for mating. The signaltransduction machinery contains a G protein-coupledreceptor, a mitogen-activated protein (MAP) kinasecascade and at least one transcription factor, althoughthere are many other components that influence thesignalling process. Responses induced by the phero-mones include a G1 arrest of the cell cycle, an increase inagglutination and an elongation of the cell to form ashmoo.
Desensitisation is an important feature of the pher-omone-response process. Mutations that reduce theability to recover from stimulation lead to reduced fer-tility (Fukui et al. 1986; Egel 1992; Imai and Yamamoto1992) and cells that fail to mate must recover from thepheromone and resume mitotic growth (Davey andNielsen 1994; Imai and Yamamoto 1994). Recoveryoccurs in the presence of extracellular pheromones and
Curr Genet (2002) 41: 241–253DOI 10.1007/s00294-002-0301-3
Communicated by M. YamamotoM. Didmon and K. Davis made equal contributions to this work
M. Didmon Æ K. Davis Æ P. Watson Æ G. Ladds Æ J. Davey (&)Department of Biological Sciences,University of Warwick, Coventry CV4 7AL, UKE-mail: [email protected]
P. BroadAstraZeneca Pharmaceuticals, Mereside,Alderley Park, Macclesfield SK10 4TG, UK

illustrates the fact that cells become desensitised tocontinued stimulation. The best characterised mecha-nism of desensitisation in Sz. pombe involves the carb-oxypeptidase Sxa2 (Imai and Yamamoto 1992; Laddset al. 1996; Ladds and Davey 2000). M-cells exposed toP-factor secrete Sxa2, which inactivates the extracellularpheromone by removing the C-terminal leucine residue.Sxa2 is not produced by P-cells or by non-stimulated M-cells. A second mechanism involves the Rgs1 protein(Watson et al. 1999; Pereira and Jones 2001), a memberof the regulator of G protein signalling (RGS) family ofproteins (for reviews, see Dohlman and Thorner 1997;Berman and Gilman 1998). RGS proteins act asGTPase-activating proteins and promote the inactiva-tion of Ga-GTP subunits. In a resting cell, heterotri-meric G proteins contain an a subunit that is bound toGDP, a b subunit and a c subunit. Activation of thereceptor stimulates the exchange of GDP by GTP,causes the Ga-GTP subunit to dissociate from the bcdimer and allows the separated components to regulatedownstream effectors. Signalling is terminated when theGa subunits hydrolyse GTP, return to the GDP-boundstate and reform the inactive heterotrimers. The slowintrinsic GTPase activity of the Ga subunits is increasedby the RGS proteins (for a review, see Berman andGilman 1998). It should be noted that the precise natureof the G protein associated with the pheromone-response pathway in Sz. pombe remains to be deter-mined and it is possible that a Ga-like subunit operatesindependently of any Gb or Gc subunits (Landry andHoffman 2001).
To identify and characterise other proteins involvedin the desensitisation process, we constructed yeaststrains that express reporter proteins under the controlof a pheromone-dependent promoter. Mutations thatreduce the ability of cells to recover from pheromonestimulation affect expression of the reporter protein;and the corresponding mutants can be isolated by
appropriate screening procedures. Here, we describe thebasic features of the reporter strains, their use to isolatemutants defective in their ability to regulate signallingand an initial characterisation of some of these mutants.
Materials and methods
Strains, reagents and general methods
The yeast strains used in this study are listed in Table 1. Details forconstructing the sxa2>lacZ and sxa2>ura4 reporter strains areincluded in the legend for Fig. 1. General yeast procedures wereperformed as described by Davey et al. (1995) and Ladds et al.(1996), using lithium acetate for the transformation of yeast. Cul-ture media used were: yeast extract (YE; for routine cell growth), adefined minimal medium (DMM) and the same minimal mediumsupplemented with amino acids (AA; Davey et al. 1995). Cellconcentrations were determined using a Coulter Channelyser(Beckman Coulter, Luton, UK). DNA manipulations were per-formed by standard methods. All constructs were sequenced by thedideoxynucleotide method, using double-stranded DNA as tem-plate and a series of oligonucleotide primers designed to generateoverlapping sequence data. Oligonucleotides were synthesised byAlta Bioscience (University of Birmingham, Birmingham, UK) orAstraZeneca Pharmaceuticals (Macclesfield, UK). Unless statedotherwise, amplification by polymerase chain reaction (PCR) usedPwo DNA polymerase (from Pyrococcus woesei), according to thesupplier’s instructions (Boehringer Mannheim Biochemicals,Lewes, UK). This polymerase has a 3¢–5¢ exonuclease (proof-reading) activity that reduces the introduction of errors duringamplification.
Total cellular RNA was prepared by disrupting cells with glassbeads and was used as template for semi-quantitative reversetranscriptase (RT)-PCR in the presence of [a32P]-dGTP. PCRproducts were separated by electrophoresis on a 6% acrylamide geland the dried gel was exposed to a phosphorimager screen wherethe radioactivity peaks for each product were estimated usingImageQuant (Molecular Dynamics, Pharmacia Amersham LifeSciences, Little Chalfont, UK).
For information, 1 unit ml–1 is defined as the concentration ofP-factor that induces a response in M-cells of 4% in the cell volumeassay (Davey 1991) and is equivalent to approximately 10–9 M. Todistinguish between components from the two yeasts used in thestudy, we adopted nomenclature in which Sz. pombe proteins are
Table 1. Schizosaccharomycespombe strains Strain Genotype
JY486 mat1-M, ?mat2/3::LEU2–, leu1–, ade6-M216, ura4-D18, cyr1-D51JY522 mat1-M, ?mat2/3::LEU2–, leu1–, ade6-M216, ura4-D18, cyr1-D51, sxa2::ura4+
JY546 mat1-M, ?mat2/3::LEU2–, leu1–, ade6-M216, ura4-D18, cyr1-D51, sxa2>lacZJY744 JY546 containing pREPJY746 JY546 containing pREP3X-Rgs1JY1054 JY546 containing pREP3X-Pmp1JY603 mat1-M, ?mat2/3::LEU2–, leu1–, ade6-M216, ura4-D18, cyr1-D51, sxa2>ura4JY780 JY603 containing pREP3XJY884 JY603 containing pREP3X-Map2JY629 mat1-M, ?mat2/3::LEU2–, leu1–, ade6-M216, ura4-D18, cyr1-D51, sxa2>lacZ, Drgs1JY691 JY629 containing pREP3XJY693 JY629 containing pREP3X-Rgs1JY723 mat1-M, ?mat2/3::LEU2–, leu1–, ade6-M216, ura4-D18, cyr1-D51, sxa2>lacZ, Dpmp1JY1066 JY723 containing pREP3XJY1067 JY723 containing pREP3X-MSG5JY1068 JY723 containing pREP3X-Pmp1JY1069 JY723 containing pREP3X-Pyp1JY1070 JY723 containing pREP3X-Pyp2JY1071 JY723 containing pREP3X-CL100JY1095 mat1-M, ?mat2/3::LEU2–, leu1–, ade6-M216, ura4-D18, cyr1-D51, sxa2>ura4, Dpmp1
242

indicated as Xyz1 and genes as xyz1 and Saccharomyces cerevisiaeproteins are indicated as XYZ1 and genes as XYZ1.
Assay of b-galactosidase activity
Assays were performed using a method modified from Dohlmanet al. (1995). Sz. pombe cells were cultured to a density of�5·105 cells ml–1 in DMM and 500-ll aliquots transferred to 2-mlSafe-Lock tubes (Eppendorf, Hamburg, Germany) containing 5 llof P-factor pheromone (in HPLC-grade methanol). Tubes wereincubated at 29 �C for 16 h on a rotating wheel and 50 ll were thentransferred to 750 ll of Z-buffer containing 2.25 mM o-nitrophe-nyl-b-D-galactopyranoside. Reactions were stopped after 90 min byadding 200 ll of 2 M Na2CO3 and b-galactosidase activity wascalculated as the optical density at 420 nm (OD420) per 106 cells(determined using a Coulter Channelyser).
Mutagenesis of Sz. pombe
Ethylmethane sulfonate (EMS) mutagenesis of Sz. pombe strainswas performed as described by Guthrie and Fink (1991). Muta-genesis using the kanr cassette was based on the method of Chua etal. (2000). The kanr cassette contains the transcriptional andtranslational sequences of the TEF gene of Ashbya gossypii and isfunctional in Sz. pombe (Bahler et al. 1998). It was amplified usingthe sense primer JO1113 (N18AAGCTTGCCTCGTCCCCG-CCGGTC) and the antisense primer JO1114 (N18GTCGACACT-GGATGGCGGCGTTAG). Amplification was performed in100-ll reactions containing 60 pmol of each primer, a final MgCl2concentration of 1.5 mM and 2.5 units of Taq DNA polymerase(from Thermus aquaticus; Life Technologies, Paisley, UK). Aninitial heat-denaturation step of 95 �C for 2 min was followed by30 cycles of denaturation (94 �C for 30 s), primer annealing (55 �Cfor 60 s) and extension (72 �C for 90 s). PCR reactions were pooledto generate 200 lg of product, which was precipitated by ethanoland used to transform the Sz. pombe sxa2>ura4 reporter strain(JY603), using a modified lithium acetate method (Bahler et al.1998). Transformants were plated on rich medium containing G418sulphate (Geneticin; Life Technologies) at 100 lg ml–1 and colonieswere resuspended in 25% glycerol for storage at –80 �C. Trans-formants were screened for their ability to grow on AA-uracilsupplemented with P-factor at 0.1 units ml–1. This is too low toinduce growth of the wild-type reporter strain, but about 1 in5,000 transformants formed colonies on these plates. GenomicDNA isolated from these colonies was digested with EcoRI andligated in 100-ll reactions at concentrations of 10 lg ml–1, using3 units of T4 DNA ligase (Life Technologies) for 16 h at 12 �C.The reaction was heat-inactivated at 65 �C for 20 min and 2 ll ofthe mixture were used as template for inverse PCR with Taq DNApolymerase and oligonucleotide primers JO1117 (AT-GCTGGTCGCTATACTG) and JO1118 (GTATTCTGGGCCT-CCATG). These oligonucleotides are specific for sequences ateither end of the kanr cassette. An initial denaturation step of 95 �Cfor 2 min was followed by 30 cycles with annealing at 50 �C and a4 min extension time. The purified PCR products were cloned usingthe TOPO TA cloning vector (Invitrogen, Groningen, The Neth-erlands) and sequenced by the dideoxynucleotide method, usingprimers specific for the cloning vector. Sequences were compared tothe Sz. pombe genome using the on-line BLAST server at theSanger Centre (Cambridge, UK; http://www.sanger.ac.uk/Projects/S_pombe/blast_server.shtml).
Preparing the pREP expression constructs
The pREP series of Sz. pombe vectors allows expression of par-ticular genes to be under the control of the thiamine-repressiblenmt1 promoter (Maundrell 1993). The Sz. pombe map2 ORF wasamplified using the sense primer JO1099 (ggggatc-caccATGAAGATCACCGCTGTCATTG), which includes aBamHI site and the sequence immediately upstream of the
initiator ATG has been modified to one that is expected to bemore favourable for translation (Yun et al. 1996), and the anti-sense primer JO1100 (ggggatccTTAGCTCTCAAATTTGGCAG),which includes a BamHI site and the TTA stop anticodon formap2. The PCR product was digested with BamHI and clonedinto the BamHI site of pREP3X to generate JD1338 (pREP3X-Map2). The Sz. pombe rgs1 ORF was amplified using the senseprimer JO599 (ggggatatccaccATGCCCGCCCTTCATAACCCT-TC), which includes an EcoRV site and a modified sequenceimmediately upstream of the initiator ATG, and the antisenseprimer JO600 (ggggatatCATTAAATACCGAGCCCCCCATTC),which includes an EcoRV site and the TTA stop anticodon forrgs1. The PCR product was digested with EcoRV and cloned intothe end-filled BamHI site of pREP3X to generate JD1013(pREP3X-Rgs1). The Sz. pombe pmp1 ORF was amplified usingthe sense primer JO1143 (gatatccaccATGTCTCAAAAACTACC-TCCC), which includes an EcoRV site and a modified sequenceimmediately upstream of the initiator ATG, and the antisenseprimer JO1144 (gatatcTCAAGAAGCATCATTACTTATAC),which includes an EcoRV site and the TCA stop anticodon forpmp1. The PCR product was digested with EcoRV and clonedinto the end-filled BamHI site of pREP3X to generate JD1456(pREP3X-Pmp1). The S. cerevisiae MSG5 ORF was amplifiedusing the sense primer JO1151 (gatatccaccATGCAATTTCACT-CAGATAAG), which includes an EcoRV site and a modifiedsequence immediately upstream of the initiator ATG, and theantisense primer JO1152 (gatatcTTAAGGAAGAAACATCATC),which includes an EcoRV site and the TTA stop anticodon forMSG5. The PCR product was digested with EcoRV and clonedinto the end-filled BamHI site of pREP3X to generate JD1454(pREP3X-MSG5). The Rattus rattus CL100 ORF was amplifiedusing the sense primer JO1204 (gatatccaccATGGTGATGGA-GGTGGGC), which includes an EcoRV site and a modifiedsequence immediately upstream of the initiator ATG, and theantisense primer JO1205 (gatatcTCAGCAGCTCGGAGAGG-TTG), which includes an EcoRV site and the TCA stop anticodonfor MSG5. The PCR product was digested with EcoRV andcloned into the end-filled BamHI site of pREP3X to generateJD1489 (pREP3X-CL100). The Sz. pombe pyp1 ORF was am-plified using the sense primer JO1145 (gatatccaccATGAATTTTT-CAAACGGTTC), which includes an EcoRV site and a modifiedsequence immediately upstream of the initiator ATG, and theantisense primer JO1146 (gatatcTCATGTTAAAACCGGGA-AATG), which includes an EcoRV site and the TCA stop antic-odon for pyp1. The PCR product was digested with EcoRV andcloned into the end-filled BamHI site of pREP3X to generateJD1458 (pREP3X-Pyp1). The Sz. pombe pyp2 ORF was amplifiedusing the sense primer JO1147 (gggggatccaccATGCTCCATC-TTCTGTCTAAAG), which includes a BamHI site and a modi-fied sequence immediately upstream of the initiator ATG, and theantisense primer JO1148 (gggggatccTTAAGTCATCAAGGGCT-TGG), which includes a BamHI site and the TTA stop anticodonfor pyp1. The PCR product was digested with BamHI and clonedinto the BamHI site of pREP3X to generate JD1460 (pREP3X-Pyp2).
Results and discussion
Constructing the reporter strains
We previously reported the construction ofSz. pombe mat1-Pm>lacZ reporter strains and their useto demonstrate the role of Rgs1 in the pheromone-response process (Watson et al. 1999). These strains canalso be used to screen for mutants defective in signalregulation, but such screens are limited by the need touse M-factor, which is difficult to synthesise and onlyavailable in small quantities (Wang et al. 1994); and they
243

are further complicated by the fact that the mat1-Pmgene is expressed at low level in the absence of phero-mone stimulation (Nielsen et al. 1992; Aono et al. 1994;Davey and Nielsen 1994; Watson et al. 1999). Wetherefore sought a pheromone-dependent promoter thatwas induced by exposure to P-factor. Unlike M-factor,which is both carboxyl-methylated and farnesylated(Davey 1992), P-factor is an unmodified peptide (Imaiand Yamamoto 1994) that can be synthesised in largeamounts.
The sxa2 gene encodes a carboxypeptidase that playsan important role in the pheromone response (Imai andYamamoto 1992; Ladds et al. 1996; Ladds and Davey2000). M-cells exposed to P-factor secrete Sxa2, whichinactivates the extracellular pheromone by removing theC-terminal leucine residue. Northern blot analysis (Imaiand Yamamoto 1992) and enzymatic studies (Laddset al. 1996; Ladds and Davey 2000) indicate that Sxa2is not produced by P-cells or non-stimulated M-cells. Wetherefore constructed sxa2-based reporter strains using atwo-step strategy to replace the sxa2 ORF with eitherlacZ or ura4 (Fig. 1). Integrating the reporter constructsinto the chromosomal sxa2 locus avoids the variabilitythat can complicate the use of plasmid-borne reporters(see, for example, Aono et al. 1994). The Escherichia colilacZ gene encodes b-galactosidase and is a commonchoice of reporter because of the availability of chrom-ogenic substrates. The Sz. pombe ura4 gene encodesorotidine 5¢-monophosphate decarboxylase (Grimmet al. 1988), an enzyme involved in the biosynthesis ofuracil. Depending upon the growth conditions, thesxa2>ura4 strain can be used to select strains that eitherexpress or do not express the reporter construct. Ura4 isrequired for growth in the absence of uracil, but itconverts 5-fluoro-orotic acid (FOA) into a toxic product(Boeke et al. 1984), leading to cell death.
Two other modifications were made to the reporterstrains. Removing the information at the mat2 and mat3loci locks the P-type information at mat1 and ensuresthe strains are unable to switch mating type (Klar andMiglio 1986). The strains also lack the cyr1 gene thatencodes adenylate cyclase (Yamawaki-Kataoka et al.1989; Young et al. 1989). Nutritional regulation ofthe pheromone-response pathway complicates mating-related studies in Sz. pombe, as cells are repressed formating during mitotic growth. The mechanism by whichcells detect nutritional status is unclear, but matingfunctions are inhibited by high intracellular levels ofcAMP and mutants lacking cyr1 respond to pheromonesduring mitotic growth (Davey and Nielsen 1994; Imaiand Yamamoto 1994).
Exposing the sxa2>lacZ reporter strain (JY546) toP-factor resulted in the time- and pheromone-dependentproduction of b-galactosidase (Fig. 2a). The sxa2 tran-script is normally induced within 1 h of exposure to P-factor, reaches a peak at 2–3 h and is undetectable by5 h (Imai and Yamamoto 1992). Sxa2 production mir-rors gene expression, such that the protein appears after1–2 h and reaches a plateau after 4–5 h (Ladds et al.
1996), the stable protein remaining long after the dis-appearance of the transcript. The activity of b-galact-osidase was detectable at times consistent withexpression of sxa2, but took longer to reach a plateau.This discrepancy could partly reflect differences in thetime taken for Sxa2 and b-galactosidase to assemble intotheir active conformations, but reporter expression isalso likely to be influenced by the absence of a functionalSxa2 in the sxa2>lacZ reporter strains. Sxa2 is a majorfactor in the ability of cells to recover from stimulationby P-factor and cells lacking sxa2 show a prolongedpheromone response (Imai and Yamamoto 1992; Laddset al. 1996). The low-level production of b-galactosidasein the absence of P-factor is a consequence of the ab-sence of a functional cyr1 gene (adenylate cyclase) in thesxa2>lacZ reporter strain. Wild-type cells do not
Fig. 1. Constructing the sxa2 reporter strains. The sxa2 openreading frame (ORF) was first replaced with a 1.8-kb Schizosac-charomyces pombe ura4+ cassette (Grimm et al. 1988). Thecomplete sxa2 locus was amplified by PCR, using the sense primerJO760 [ggggggtacCATGGCTAGAAATCCGCCATTGTGTG;lower-case letters are not complementary to sxa2 but theoligonucleotide includes a KpnI site (ggtac*C) and an NcoI site(c*CATGG), where digestion with NcoI leaves ends that are fullyhomologous to the chromosomal sequence] and the antisenseprimer JO683 (CTTCTCGTAAAGGCACATTGACGG; comple-mentary to a region immediately downstream of the BamHI site atposition 2,043). The resulting PCR product was cloned into theKpnI and BamHI sites of pSP72 (Promega, Southampton, UK) togenerate JD808 (pSP72 containing the sxa2 locus). This was used astemplate for PCR with JO746 (TGAAAAGAGAGACAATG;antisense primer complementary to a region immediately upstreamof the ATG initiator codon for sxa2) and JO745 (TA-AAAGTTTAATATC; sense primer complementary to a regionthat includes the TAA stop codon for sxa2) and the product wasligated either to the ura4+ cassette (to generate JD857, pSP72containing a construct suitable for disruption of sxa2) or to PCRproducts corresponding to either the lacZ ORF (to generate JD954,pSP72 containing the sxa2>lacZ reporter construct) or the ura4ORF (to generate JD929, pSP72 containing the sxa2>ura4reporter construct). The lacZ ORF was prepared by amplificationusing the sense primer JO660 (ATGCAGCTGGCACGAC-AGGTTTCCCGAC; includes the ATG initiator codon and next25 bases of the lacZ ORF) and the antisense primer JO661(TTTTTGACACCAGACCAACTGGTAATGGTAGC; comple-mentary to the 3¢ end of the lacZ ORF but lacks the stopanticodon). The ura4 ORF was prepared by amplification usingthe sense primer JO828 (ATGGATGCTAGAGTATTTC; includesthe ATG initiator codon and next 16 bases of the ura4 ORF)and the antisense primer JO759 (ATGCTGAGAAAGTCTTTGC;complementary to the 3¢ end of the ura4 ORF but lacks the stopanticodon). JY486 (a mating stable M-cell lacking cyr1) wastransformed with a NcoI-BamHI fragment corresponding to thesxa2::ura4+ construct (isolated from JD857); and stable ura+
transformants were initially screened by PCR and replacement ofthe sxa2 locus was confirmed by Southern blot. A correctsxa2::ura4+ disruptant (JY522) was then transformed with theNcoI–BamHI fragments corresponding to the sxa2>lacZ reporter(isolated from JD954) or the sxa2>ura4 reporter (isolated fromJD929). Stable ura– transformants were selected by their ability togrow in the presence of 5¢ fluoro-orotic acid (FOA; Boeke et al.1984) and homologous integration of the reporter constructs at thesxa2 locus was confirmed by Southern blot for JY546 (sxa2>lacZ)and JY603 (sxa2>ura4). Southern blot analysis was performed ongenomic DNA digested with PvuII and HindIII and a probecorresponding to the 5¢ untranslated region of sxa2
c
244

express sxa2 during normal mitotic growth (Imai andYamamoto 1992; Ladds et al. 1996), but disruption ofcyr1 relieves nutrient repression and causes low-levelexpression, even in the absence of pheromone (Yabanaand Yamamoto 1996).
Exposing the sxa2>ura4 reporter strain (JY603) toP-factor resulted in the pheromone-dependent produc-tion of orotidine 5¢-monophosphate decarboxylase(Fig. 2b). Cells require the expression of ura4 to grow inthe absence of uracil and the reporter strain did notgrow on plates lacking P-factor, or on plates containingP-factor at 1 unit ml–1. The addition of P-factor to atleast 10 units ml–1 induced sufficient expression of thesxa2>ura4 reporter to support colony formation. Thecomplementary situation was observed on plates con-taining uracil and FOA, the sxa2>ura4 reporter beingunable to form colonies on plates containing P-factor ator above 10 units ml–1.
The assays used to monitor the activity of the dif-ferent reporters are not directly comparable and wesought an alternative assay to demonstrate that the twostrains display similar responses to P-factor. Exposureof a cyr1– strain to mating pheromones induces a G1arrest of the cell cycle and a consequent increase in cellvolume, leading to the formation of a shmoo (Daveyand Nielsen 1994; Imai and Yamamoto 1994). Thesxa2>lacZ (JY546) and sxa2>ura4 (JY603) reporterstrains were therefore exposed to P-factor and cell
number and median cell volume determined, using aCoulter Channelyser (Fig. 2c). A non-synchronous cul-ture contains cells of various sizes and the median cellvolume is a useful measure of cell size (this is the volumethat divides the distribution into two equal groups, suchthat 50% of the cells are larger than the median vol-ume). In the absence of P-factor, each reporter strainhad a generation time of about 5 h and a median cellvolume of about 72 lm3. The two strains also behavedsimilarly when exposed to P-factor; undergoing com-parable cell-cycle arrests and very similar increases incell volume. It is difficult to precisely compare the twostrains using such a non-quantitative assay, but the re-sults suggest that the two reporter strains have similarsensitivities to P-factor.
Isolating signalling mutants
The sxa2>ura4 reporter strain provides a convenientway of isolating mutants with altered sensitivity topheromone stimulation and could help identify proteinsthat regulate the signalling machinery. For example, astrain with a reduced ability to negatively regulate theresponse machinery might require lower levels ofP-factor to induce functional expression of thesxa2>ura4 reporter than the 10 units ml–1 needed bywild-type cells (see Fig. 2b).
245

Mutagenesis with EMS generated many potentialsignalling mutants, but the subsequent analysis of theseisolates proved difficult. Genetic analysis was hamperedby the greatly reduced fertility of strains lacking afunctional sxa2 gene (Imai and Yamamoto 1992) andour attempts to rescue the defects by complementationwith Sz. pombe libraries was complicated by the isola-tion of extragenic suppressors, most of which did notdirectly affect the signalling machinery. We thereforesought an alternative method of generating the signal-ling mutants.
Efficient joining of non-homologous DNA sequencesin Sz. pombe (Goedecke et al. 1994) leads to a rate ofillegitimate recombination that is high enough to beexploited for insertional mutagenesis (Chua et al. 2000).In this approach, a cassette containing a selectablemarker possessing termini of degenerate sequence be-comes integrated randomly into the Sz. pombe genomeand provides an efficient method of mutagenesis. The
cassette is then recovered with its associated genomicsequence and the availability of the Sz. pombe genomesequence enables identification of the integration site.We chose the kanr gene from E. coli as the marker be-cause it is functional in Sz. pombe (Bahler et al. 1998)and the lack of sequence homology with the yeast ge-nome ensures illegitimate integration. Fully degeneratetails of 18 bp were added to each end of the cassette byPCR and the linear product was used to transform thesxa2>ura4 reporter strain (JY603). Transformants werescreened for their ability to grow on AA-uracil supple-mented with P-factor at 0.1 units ml–1. This is too low toinduce growth of the wild type reporter strain but about1 in 5,000 transformants formed colonies on theseplates. About half of these isolates could also grow inthe absence of P-factor, suggesting that the sxa2>ura4was constitutively expressed in these cells. Isolates wereclassified as either group A (reporter produced inresponse to lower than normal levels of P-factor) or group
246

B (reporter produced in the absence of P-factor;Fig. 3a).
Genomic DNA was prepared from each strain, di-gested with EcoRI and analysed by Southern blot, usinga probe specific for the kanr gene (Fig. 3b). Each isolateappeared to possess a single insertion event and the kanr
cassette was integrated at a different site in most, if notall, transformants. The EcoRI-digested fragments ofgenomic DNA were circularised and used as templatefor inverse PCR to amplify the regions flanking each endof the kanr cassette. The PCR products were cloned andtheir sequences compared to the Sz. pombe genome us-ing the BLAST server at the Sanger Centre (Cambridge,UK). Sequence analysis identified the locations for mostof the integration events. As reported by Chua et al.(2000), many of the integration events involved deletionswithin the genome and at the ends of the cassette.
Several of the genes affected by the integration eventsencode proteins previously shown to regulate intracel-lular signalling (although not necessarily the phero-mone-signalling pathway) and demonstrate the ability ofthe approach to identify such proteins. Some of theother genes affected in our strains do not encode signalregulators, but we can use their known functions topropose explanations as to why they were isolated byour screen. We have so far analysed 74 mutants but herelimit our discussion to five isolates that illustrate thesedifferent types of mutants. Many other isolates are stillbeing characterised and will be described elsewhere.
Integrations that affect Map1, Map2and the Sxa2 reporter
Integration of the kanr cassette 296 bp upstream of themap1 gene produced a strain that responded to very low
levels of P-factor (one of the group A mutants inFig. 3a). The map1 gene was originally defined by amutation that caused P-specific sterility (Egel 1973), butwas subsequently shown to encode a transcription factorthat regulates expression of mating-type genes in bothM-cells and P-cells (Yabana and Yamamoto 1996;Nielsen et al. 1996). Its mode of action in M-cells is notclearly defined, but overexpression of map1 in a mating-competent cell greatly increases expression of mating-related genes, including sxa2 (Yabana and Yamamoto1996). Semi-quantitative RT-PCR (using cdc2 as theinternal standard) revealed a 5-fold increase in map1mRNA in the mutant containing the integrated kanr
cassette, compared with the normal reporter strain(Fig. 4a). Although the relevant levels of Map1 proteinin the two strains is not known, our observed increase inmap1 expression would be expected to increase thepheromone sensitivity of the sxa2>ura4 reporter strain,leading to Ura4 production at lower than normal levelsof P-factor. The need for Map1 to interact with proteinsonly produced following pheromone stimulation wouldexplain why the map1 mutant fails to grow in the ab-sence of P-factor. Consistent with the earlier observationof Yabana and Yamamoto (1996), overexpression of aplasmid-borne map1 from the thiamine-repressible nmt1promoter greatly increased the pheromone sensitivity ofthe sxa2 reporter strains (data not shown).
Integration of the kanr cassette 830 bp upstream of themap2 gene produced a strain that induced expression ofthe sxa2>ura4 reporter in the absence of P-factor (one ofthe group B mutants in Fig. 3a). A second group B mu-tant had the kanr cassette integrated very close to thisposition. The map2 gene encodes the P-factor precursorand is not normally expressed in M-cells (Imai andYamamoto 1994). Previous studies have demonstratedthat ectopic expression of the ‘‘wrong’’ mating phero-mone in yeast cells leads to an autocrine stimulation of theresponse pathway as the cell responds to its own phero-mone (Manfredi et al. 1996; Kitamura et al. 1996). Todemonstrate this also occurs in the sxa2>ura4 reporter,we introduced a plasmid in which expression of the map2gene was under the control of the thiamine-repressiblenmt1 promoter (JY884; JY603 containing pREP3X-Map2). An equivalent strain was transformed with thevector alone (JY780; JY603 containing pREP3X). Asexpected, both strains formed colonies on AA-uracilplates supplemented with P-factor at 10 units ml–1
(Fig. 4b). The strain lacking Map2 (JY780) could notform colonies in the absence of P-factor and the ability ofthe pREP3X-Map2 transformant to grow was dependenton the thiamine content of the medium, forming coloniesin the absence of thiamine (nmt1 promoter on) but not inthe presence of thiamine (nmt1 promoter off). Integrationof the kanr cassette upstream of the map2 gene is likely tobring about the same effect by interfering with themechanism that normally prevents expression of thisgene, thereby leading to the production of P-factor.
Integration of the kanr cassette at 900–1,200 bp up-stream of the sxa2>ura4 reporter gene produced three
Fig. 2a–c. Pheromone-dependent expression of the sxa2>reporterconstructs. a The sxa2>lacZ reporter strain (JY546) was exposedto P-factor and b-galactosidase production assayed using o-nitrophenyl-b-D-galactosidase as substrate. Activity is expressedas OD420 units per 106 cells (see Materials and methods). In the leftpanel, cells were exposed to P-factor at either 0 or 1,000 units ml–1
and assayed at hourly intervals. In the right panel, cells wereexposed to various concentrations of P-factor for 16 h before beingassayed. The values shown are the means of triplicate determina-tions from a typical experiment. b The sxa2>ura4 reporter strain(JY603) was exposed to P-factor and orotidine 5¢-monophosphatedecarboxylase determined either by cell growth in the absence ofuracil (upper series of plates) or by cell death in the presence of 5-FOA (lower series of plates). Plates were supplemented withincreasing concentrations of P-factor (0–100 units ml–1) andapproximately 2,000 cells were spread on each plate. Plates wereincubated at 29 �C for 3 days. AA Defined minimal medium withamino acids c The sxa2>lacZ (JY546) and sxa2>ura4 (JY603)reporter strains were grown in defined minimal medium at 29 �C toa concentration of 106 cells ml–1. Each sample was divided in twoand P-factor was added to one half to a final concentration of1,000 units ml–1. Cell numbers and median cell volumes weredetermined using a Coulter Channelyser. There were no differencesbetween the strains in the absence of P-factor and, for clarity, onlythe results for the sxa2>lacZ strain are shown
b
247

strains that induced reporter expression in the absence ofP-factor (another group B mutant in Fig. 3a). The sxa2gene is normally subject to tight transcriptional regula-tion (Imai and Yamamoto 1992, 1994; Ladds et al. 1996;this study) and it is not a surprise that this integrationcould affect elements that are required for repressingexpression of sxa2 in the absence of P-factor.
Identification of Rgs1 as a regulatorof pheromone signalling
Disruption of the rgs1 gene produced 12 strains (16% ofthe mutants analysed) that expressed the sxa2>ura4
reporter in the absence of P-factor. As described in theIntroduction, Rgs1 was first identified by the Sz. pombesequencing project as a hypothetical protein with simi-larity to members of the RGS family. We previously re-ported that Sz. pombe P-cells lacking rgs1 (Drgs1) arehypersensitive to stimulation by the M-factor pheromone(Watson et al. 1999) and therefore expected this to be oneof the genes identified in our screens. To quantitate therole of Rgs1 in regulating signalling in response toP-factor, we disrupted the rgs1 gene in the sxa2>lacZ(JY546) strain and assayed b-galactosidase activity fol-lowing exposure to P-factor (Fig. 5). Loss of rgs1 in-creased production of b-galactosidase in the absence ofP-factor and also made the cells more sensitive to pher-omone stimulation. These changes were due to the loss ofrgs1, as they were overcome by expression of a plasmid-borne rgs1 gene. These results are similar to those wereported for Rgs1 in P-cells stimulated with M-factor(Watson et al. 1999) and demonstrates that Rgs1 playsthe same role in both cell types (Pereira and Jones 2001).
Identification of Pmp1 as a regulatorof pheromone signalling
Ten of the mutants (14% of those analysed) with in-creased sensitivity to mating pheromone (group A-typemutants in Fig. 3a) had mutations in the pmp1 gene.Pmp1 (which is also called Dsp1) is a dual-specificity
Fig. 3a, b. Isolation of signalling mutants. a JY603 (sxa2>ura4)was transformed with a construct in which the kanr gene fromEscherichia coli was flanked by fully degenerate tails of 18 bp.Transformants were screened for their ability to grow on definedminimal medium with amino acids and uracil, supplemented withP-factor at 0.1 units ml–1. This is too low to induce growth of thewild-type reporter strain, but about 1 in 5,000 transformantsformed colonies on these plates. Isolates were classified as eithergroup A (reporter produced in response to lower than normal levelsof P-factor, including mutations affecting the map1 or pmp1 genes)or group B (reporter produced in the absence of P-factor, includingmutations affecting the map2, sxa2 or rgs1 genes). b Genomic DNAwas prepared from seven randomly selected signalling mutants,digested with EcoRI and analysed by Southern blot, using a probespecific for the kanr gene. The EcoRI-digested fragments ofgenomic DNA were circularised and used as template for inversePCR to identify the regions flanking each end of the kanr cassette
248

phosphatase that was isolated as a regulator of the MAPkinase Pmk1 (Sugiura et al. 1998). Pmk1 is part of thesignalling pathway that maintains ion homeostasis andcell integrity (Toda et al. 1996), but it was also isolated(and called Spm1) as a stress-activated regulator of cellmorphogenesis (Zaitsevskaya-Carter and Cooper 1997).Results suggest that Pmp1 negatively regulates Pmk1 bydirect dephosphorylation (Sugiura et al. 1998) . Pmp1
had not previously been implicated in regulating thepheromone response, but this signalling pathway alsoinvolves a MAP kinase (Spk1) and this suggested anobvious role for Pmp1. A one-step gene-replacementstrategy was therefore used to prepare sxa2>lacZ re-porter strains lacking pmp1 to further investigate thispossibility.
Strains with and without pmp1 were assayed for b-galactosidase activity following exposure to P-factor(Fig. 6a). Disruption of pmp1 caused a greater responseto stimulation above 10 units ml–1 and this was over-come by expression of a plasmid-borne pmp1 gene(pREP3X-Pmp1). Given that pmp1 was identified in ascreen at 0.1 units ml–1, it was initially surprising thatdisruption of the gene had no noticeable affect on b-galactosidase activity at low levels of P-factor. We havesince observed this with a number of other genes andbelieve it is a consequence of the time-frames over whichthe different reporters are measured.
Complementation of Dpmp1 by dual-specificityphosphatases
Pmp1 is closely related to the budding yeast MSG5 dual-specificity phosphatase. This was isolated as a
Fig. 4a, b. Modifying the expression levels of map1 and map2. aSemi-quantitative RT-PCR was used to investigate expression ofthe map1 gene in the normal reporter strain (map1+) and in amutant containing the kanr cassette integrated 296 bp upstream ofthe map1 ORF (kanR-map1). Total RNA was prepared from bothstrains and used as template for amplification in the presence of[a32P]-GTP, using oligonucleotide primers specific for map1 andcdc2. The cdc2 transcript was not affected by the developmentalstate of the cells and demonstrated that approximately equalamounts of RNA were used as template in each reaction. The PCRproducts were separated by electrophoresis on a 6% acrylamide geland the dried gel exposed to a phosphorimager screen. b Thesxa2>ura4 reporter (JY603) was transformed with a plasmidcontaining the map2 gene (encodes P-factor) under the control ofthe thiamine-repressible nmt1 promoter (to generate strain JY884).Transformation with the vector alone provided a control strain(JY780)
Fig. 5. The influence of Rgs1 on pheromone-dependent transcrip-tion. Pheromone-dependent transcription of a sxa2>lacZ reporterconstruct was assayed by monitoring b-galactosidase activity incells with different combinations of Rgs1. The sxa2>lacZ reporter(JY546, rgs1+) was transformed with a plasmid containing the rgs1gene under the control of the thiamine-repressible nmt1 promoter(to generate JY746). An equivalent sample was transformed withthe vector alone to provide a control strain (JY744). A sxa2>lacZreporter strain lacking rgs1 (JY629, Drgs1) was transformed withthe same plasmids to generate JY691 (Drgs1 with pREP3X) andJY693 (Drgs1 with pREP3X-Rgs1). Cells were grown to mid-exponential phase and treated with P-factor for 16 h before beingharvested. The b-galactosidase activity was determined colorimet-rically (see Materials and methods). The values shown are themeans of triplicate determinations. There was no apparentdifference between the rgs1+ and Drgs1 strains in the presence ofpREP3X-Rgs1 and, for clarity, only the results for the Drgs1 strain(JY693) are shown
249
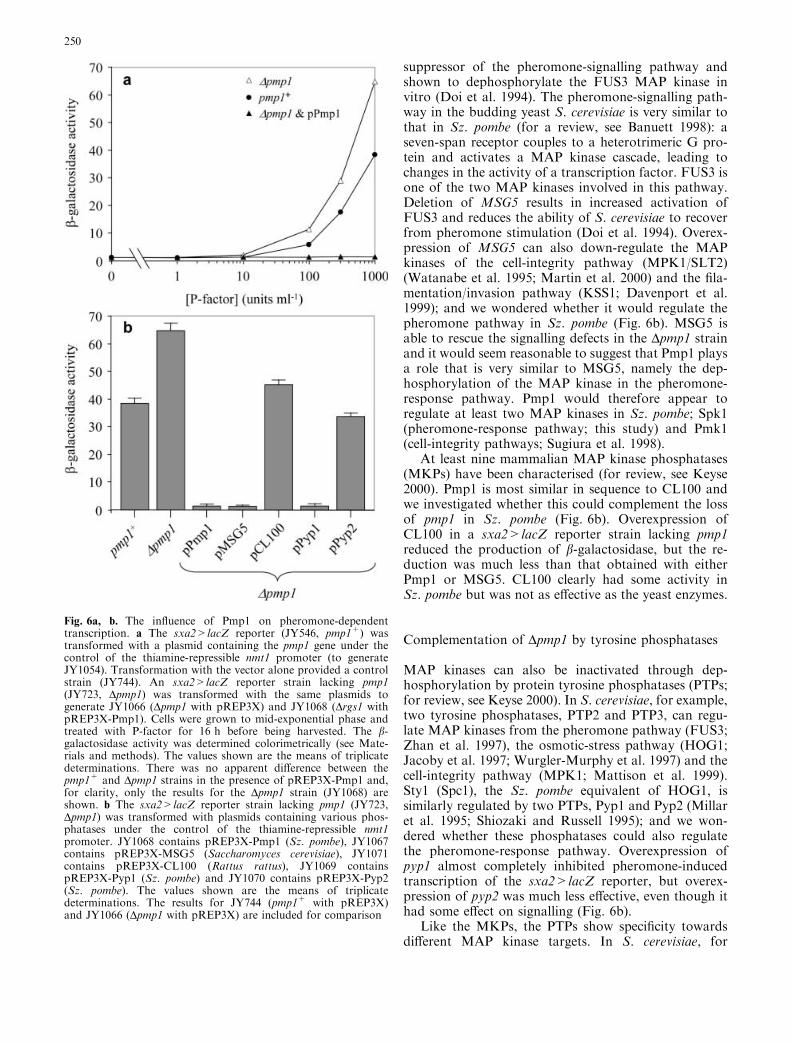
suppressor of the pheromone-signalling pathway andshown to dephosphorylate the FUS3 MAP kinase invitro (Doi et al. 1994). The pheromone-signalling path-way in the budding yeast S. cerevisiae is very similar tothat in Sz. pombe (for a review, see Banuett 1998): aseven-span receptor couples to a heterotrimeric G pro-tein and activates a MAP kinase cascade, leading tochanges in the activity of a transcription factor. FUS3 isone of the two MAP kinases involved in this pathway.Deletion of MSG5 results in increased activation ofFUS3 and reduces the ability of S. cerevisiae to recoverfrom pheromone stimulation (Doi et al. 1994). Overex-pression of MSG5 can also down-regulate the MAPkinases of the cell-integrity pathway (MPK1/SLT2)(Watanabe et al. 1995; Martin et al. 2000) and the fila-mentation/invasion pathway (KSS1; Davenport et al.1999); and we wondered whether it would regulate thepheromone pathway in Sz. pombe (Fig. 6b). MSG5 isable to rescue the signalling defects in the Dpmp1 strainand it would seem reasonable to suggest that Pmp1 playsa role that is very similar to MSG5, namely the dep-hosphorylation of the MAP kinase in the pheromone-response pathway. Pmp1 would therefore appear toregulate at least two MAP kinases in Sz. pombe; Spk1(pheromone-response pathway; this study) and Pmk1(cell-integrity pathways; Sugiura et al. 1998).
At least nine mammalian MAP kinase phosphatases(MKPs) have been characterised (for review, see Keyse2000). Pmp1 is most similar in sequence to CL100 andwe investigated whether this could complement the lossof pmp1 in Sz. pombe (Fig. 6b). Overexpression ofCL100 in a sxa2>lacZ reporter strain lacking pmp1reduced the production of b-galactosidase, but the re-duction was much less than that obtained with eitherPmp1 or MSG5. CL100 clearly had some activity inSz. pombe but was not as effective as the yeast enzymes.
Complementation of Dpmp1 by tyrosine phosphatases
MAP kinases can also be inactivated through dep-hosphorylation by protein tyrosine phosphatases (PTPs;for review, see Keyse 2000). In S. cerevisiae, for example,two tyrosine phosphatases, PTP2 and PTP3, can regu-late MAP kinases from the pheromone pathway (FUS3;Zhan et al. 1997), the osmotic-stress pathway (HOG1;Jacoby et al. 1997; Wurgler-Murphy et al. 1997) and thecell-integrity pathway (MPK1; Mattison et al. 1999).Sty1 (Spc1), the Sz. pombe equivalent of HOG1, issimilarly regulated by two PTPs, Pyp1 and Pyp2 (Millaret al. 1995; Shiozaki and Russell 1995); and we won-dered whether these phosphatases could also regulatethe pheromone-response pathway. Overexpression ofpyp1 almost completely inhibited pheromone-inducedtranscription of the sxa2>lacZ reporter, but overex-pression of pyp2 was much less effective, even though ithad some effect on signalling (Fig. 6b).
Like the MKPs, the PTPs show specificity towardsdifferent MAP kinase targets. In S. cerevisiae, for
Fig. 6a, b. The influence of Pmp1 on pheromone-dependenttranscription. a The sxa2>lacZ reporter (JY546, pmp1+) wastransformed with a plasmid containing the pmp1 gene under thecontrol of the thiamine-repressible nmt1 promoter (to generateJY1054). Transformation with the vector alone provided a controlstrain (JY744). An sxa2>lacZ reporter strain lacking pmp1(JY723, Dpmp1) was transformed with the same plasmids togenerate JY1066 (Dpmp1 with pREP3X) and JY1068 (Drgs1 withpREP3X-Pmp1). Cells were grown to mid-exponential phase andtreated with P-factor for 16 h before being harvested. The b-galactosidase activity was determined colorimetrically (see Mate-rials and methods). The values shown are the means of triplicatedeterminations. There was no apparent difference between thepmp1+ and Dpmp1 strains in the presence of pREP3X-Pmp1 and,for clarity, only the results for the Dpmp1 strain (JY1068) areshown. b The sxa2>lacZ reporter strain lacking pmp1 (JY723,Dpmp1) was transformed with plasmids containing various phos-phatases under the control of the thiamine-repressible nmt1promoter. JY1068 contains pREP3X-Pmp1 (Sz. pombe), JY1067contains pREP3X-MSG5 (Saccharomyces cerevisiae), JY1071contains pREP3X-CL100 (Rattus rattus), JY1069 containspREP3X-Pyp1 (Sz. pombe) and JY1070 contains pREP3X-Pyp2(Sz. pombe). The values shown are the means of triplicatedeterminations. The results for JY744 (pmp1+ with pREP3X)and JY1066 (Dpmp1 with pREP3X) are included for comparison
250
example, although both PTP2 and PTP3 regulate bothHOG1 and FUS3, PTP2 is the main regulator of HOG1,while PTP3 is the main regulator of FUS3 (Jacoby et al.1997; Wurgler-Murphy et al. 1997; Zhan et al. 1997).Our results suggest that a similar situation exists inSz. pombe. Pyp1 and Pyp2 can both regulate Sty1 andSpk1 but, whereas Pyp2 is the better regulator of Sty1(Millar et al. 1995; Gaits and Russell 1999), our resultssuggest that Pyp1 is the more effective against Spk1(Fig. 6b).
Long term effects of Dpmp1
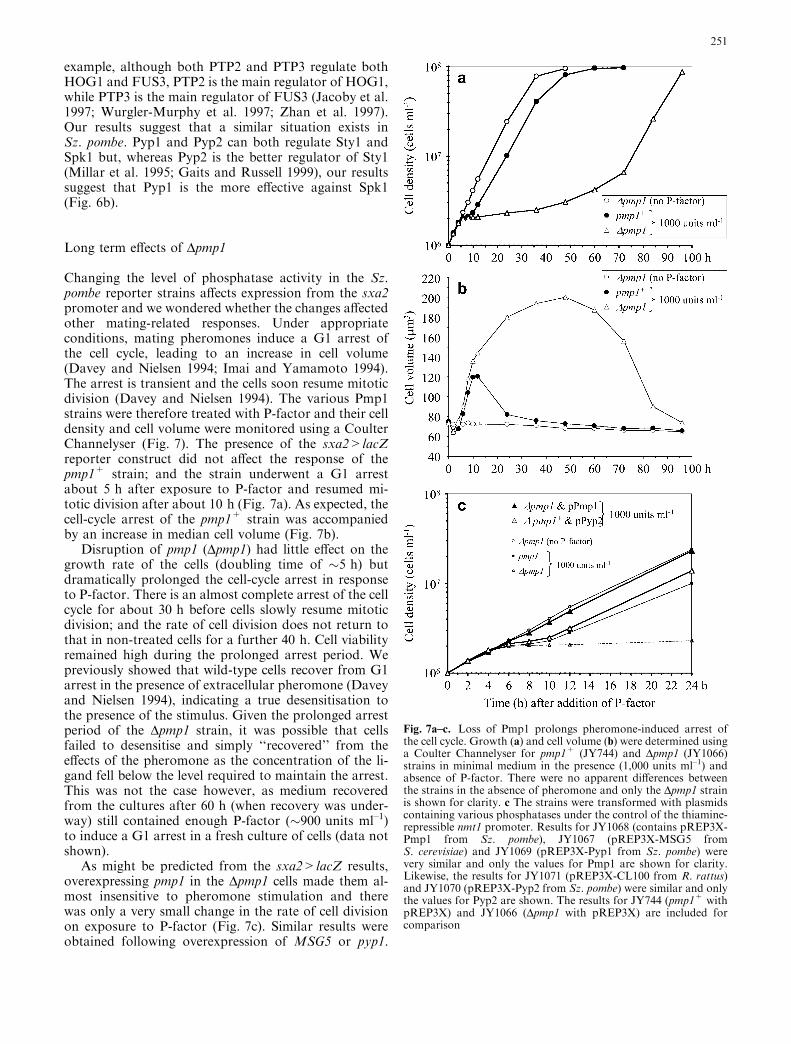
Changing the level of phosphatase activity in the Sz.pombe reporter strains affects expression from the sxa2promoter and we wondered whether the changes affectedother mating-related responses. Under appropriateconditions, mating pheromones induce a G1 arrest ofthe cell cycle, leading to an increase in cell volume(Davey and Nielsen 1994; Imai and Yamamoto 1994).The arrest is transient and the cells soon resume mitoticdivision (Davey and Nielsen 1994). The various Pmp1strains were therefore treated with P-factor and their celldensity and cell volume were monitored using a CoulterChannelyser (Fig. 7). The presence of the sxa2>lacZreporter construct did not affect the response of thepmp1+ strain; and the strain underwent a G1 arrestabout 5 h after exposure to P-factor and resumed mi-totic division after about 10 h (Fig. 7a). As expected, thecell-cycle arrest of the pmp1+ strain was accompaniedby an increase in median cell volume (Fig. 7b).
Disruption of pmp1 (Dpmp1) had little effect on thegrowth rate of the cells (doubling time of �5 h) butdramatically prolonged the cell-cycle arrest in responseto P-factor. There is an almost complete arrest of the cellcycle for about 30 h before cells slowly resume mitoticdivision; and the rate of cell division does not return tothat in non-treated cells for a further 40 h. Cell viabilityremained high during the prolonged arrest period. Wepreviously showed that wild-type cells recover from G1arrest in the presence of extracellular pheromone (Daveyand Nielsen 1994), indicating a true desensitisation tothe presence of the stimulus. Given the prolonged arrestperiod of the Dpmp1 strain, it was possible that cellsfailed to desensitise and simply ‘‘recovered’’ from theeffects of the pheromone as the concentration of the li-gand fell below the level required to maintain the arrest.This was not the case however, as medium recoveredfrom the cultures after 60 h (when recovery was under-way) still contained enough P-factor (�900 units ml–1)to induce a G1 arrest in a fresh culture of cells (data notshown).
As might be predicted from the sxa2>lacZ results,overexpressing pmp1 in the Dpmp1 cells made them al-most insensitive to pheromone stimulation and therewas only a very small change in the rate of cell divisionon exposure to P-factor (Fig. 7c). Similar results wereobtained following overexpression of MSG5 or pyp1.
Fig. 7a–c. Loss of Pmp1 prolongs pheromone-induced arrest ofthe cell cycle. Growth (a) and cell volume (b) were determined usinga Coulter Channelyser for pmp1+ (JY744) and Dpmp1 (JY1066)strains in minimal medium in the presence (1,000 units ml–1) andabsence of P-factor. There were no apparent differences betweenthe strains in the absence of pheromone and only the Dpmp1 strainis shown for clarity. c The strains were transformed with plasmidscontaining various phosphatases under the control of the thiamine-repressible nmt1 promoter. Results for JY1068 (contains pREP3X-Pmp1 from Sz. pombe), JY1067 (pREP3X-MSG5 fromS. cerevisiae) and JY1069 (pREP3X-Pyp1 from Sz. pombe) werevery similar and only the values for Pmp1 are shown for clarity.Likewise, the results for JY1071 (pREP3X-CL100 from R. rattus)and JY1070 (pREP3X-Pyp2 from Sz. pombe) were similar and onlythe values for Pyp2 are shown. The results for JY744 (pmp1+ withpREP3X) and JY1066 (Dpmp1 with pREP3X) are included forcomparison
251

Overexpressing pyp2 in the Dpmp1 strain reduced but didnot abolish the G1 arrest; and the strains gave responsessimilar to pmp1+ cells (Fig. 7c). A very similar resultwas obtained following overexpression of CL100.
In conclusion, we developed Sz. pombe strains con-taining reporter genes under the control of a phero-mone-regulated promoter that allowed signalling to bemonitored during normal mitotic growth. A lacZ re-porter provides a quantitative measure of signallingthrough the response pathway, while a ura4 reporterallows a growth-based screen to identify mutants withdefects in proteins that regulate the signalling machin-ery. As expected, we isolated mutations in proteinsinvolved in the signalling pathway (the ligand, the re-ceptor, the reporter construct) and known, or suspected,to regulate the signalling machinery (Rgs1, Pmp1). Weare currently characterising other mutants isolated bythis approach.
Acknowledgements This work was supported by a Co-operativeAward in Science and Engineering from the Biotechnology andBiological Sciences Research Council and AstraZeneca Pharma-ceuticals (97/A2/C/03552, MD), the Cancer Research Campaign(SP1972; K.D., P.W.) and the Wellcome Trust (049472; G.L.).Some of this work was performed while J.D. was a Lister InstituteResearch Fellow. We thank Jurg Bahler (ICRF, London), OlafNielsen (University of Copenhagen), Bob Old (University ofWarwick) and Masayuki Yamamoto (University of Tokyo) forplasmids and strains.
References
Aono T, Yanai H, Miki F, Davey J, Shimoda C (1994) Matingpheromone-induced expression of the mat1-Pm gene ofSchizosaccharomyces pombe: identification of signalling com-ponents and characterisation of upstream controlling elements.Yeast 10:757–770
Bahler J, Wu J-Q, Longtim MS, Shaah NV, McKenzie A,Steever AB, Wach A, Philippsen P, Pringle JR (1998) Heter-ologous modules for efficient and versatile PCR-based genetargeting in Schizosaccharomyces pombe. Yeast 14:943–951
Banuett F (1998) Signalling in the yeasts: an informational cascadewith links to the filamentous fungi. Microbiol Mol Biol Rev62:249–274
Berman DM, Gilman AG (1998) Mammalian RGS proteins: bar-barians at the gate. J. Biol. Chem. 273:1269–1272
Boeke JD, LaCroute F, Fink GR (1984) A positive selection formutants lacking orotidine-5¢-phosphate decarboxylase activityin yeast: 5-fluoro-orotic acid resistance. Mol Gen Genet197:345–346
Bohm SK, Grady EF, Bunnett NW (1997) Regulatory mechanismsthat modulate signalling by G protein-coupled receptors. Bio-chem J 322:1–18
Chua G, Taricani L, Stangle W, Young PG (2000) Insertionalmutagenesis based on illegitimate recombination inSchizosaccharomyces pombe. Nucleic Acids Res 28:E53
Davenport KD, Williams KE, Ullmann BD, Gustin MC (1999)Activation of the Saccharomyces cerevisiae filamentation/inva-sion pathway by osmotic stress in high-osmolarity glycogenpathway mutants. Genetics 153:1091–1103
Davey J (1991) Isolation and quantitation of M-factor, a diffusiblemating factor from the fission yeast Schizosaccharomycespombe. Yeast 7:357–366
Davey J (1992) Mating pheromones of the fission yeastSchizosaccharomyces pombe: purification and structural char-acterisation of M-factor and isolation and structural charac-
terisation of two genes encoding the pheromone. EMBO J11:951–960
Davey J (1998) Fusion of a fission yeast. Yeast 14:1529–1566Davey J, Nielsen O (1994) Mutations in cyr1 and pat1 reveal
pheromone-induced G1 arrest in the fission yeastSchizosaccharomyces pombe. Curr Genet 26:105–112
Davey J, Egel R, Nielsen O (1995) Pheromone procedures in fissionyeast. In: Adolph KW (ed) Microbial gene techniques; methodsin molecular genetics, vol 6B. Academic Press, San Diego,pp 247–263
Dohlman HG, Thorner J (1997) RGS proteins and signaling byheterotrimeric G proteins. J Biol Chem 272:3871–3874
Dohlman HG, Apaniesk D, Chen Y, Song J, Nusskern D (1995)Inhibition of G protein signalling by dominant gain-of-functionmutations in Sst2p, a pheromone desensitisation factor inSaccharomyces cerevisiae. Mol Cell Biol 15:3635–3643
Doi K, Gartner A, Ammerer G, Errede B, Shinkawa H, SugimotoK, Matsumoto K (1994) MSG5, a novel protein phosphatasepromotes adaptation to pheromone response in S. cerevisiae.EMBO J 13:61–70
Egel R (1973) Genes involved in mating-type expression in fissionyeast. Mol Gen Genet 122:339–343
Egel R (1992) Pheromone detection and pheromone hypersensitivemutants in the fission yeast Schizosaccharomyces pombe. Yeast8S:S363
Freedman NJ, Lefkowitz RJ (1996) Desensitisation of G protein-coupled receptors. Recent Prog Horm Res 51:319–353
Fukui Y, Kozasa T, Kaziro Y, Takeda T, Yamamoto M (1986)Role of a ras homolog in the life cycle of Schizosaccharomycespombe. Cell 44:329–336
Gaits F, Russell P (1999) Active nucleocytoplasmic shuttling re-quired for function and regulation of stress-activated kinaseSpc1/Sty1 in fission yeast. Mol Biol Cell 10:1395–1407
Goedecke W, Pfeiffer P, Vielmetter W (1994) Non-homologousDNA ends joining in Schizosaccharomyces pombe efficientlyeliminates double-strand breaks from haploid sequences. Nu-cleic Acids Res 22:2094–2101
Grimm C, Kohli J, Murray J, Maundrell K (1988) Genetic engi-neering of Schizosaccharomyces pombe: a system for gene dis-ruption and replacement using the ura4 gene as a selectablemarker. Mol Gen Genet 215:81–86
Guthrie E, Fink GR (1991) Guide to yeast genetics and molecularbiology. Methods Enzymol 174
Imai Y Yamamoto M (1992) Schizosaccharomyces pombe sxa1+
and sxa2+ encode putative proteases involved in the matingresponse. Mol Cell Biol 12:1827–1834
Imai Y, Yamamoto M (1994) The fission yeast mating pheromoneP-factor: its molecular structure, gene structure and physio-logical activities to induce gene expression and G1 arrest in themating partner. Genes Dev 8:328–338
Jacoby T, Flanagan H, Faykin A, Seto AG, Mattison C, Ota I(1997) Two protein-tyrosine phosphatases inactivate the os-motic stress response pathway in yeast by targeting the mitogen-activated protein kinase, HOG1. J Biol Chem 272:17749–17755
Keyse SM (2000) Protein phosphatases and the regulation of mi-togen-activated protein kinase signalling. Curr Opin Cell Biol12:186–192
Kitamura K, Nakamura T, Miki F, Shimoda C (1996) Autocrineresponse of Schizosaccharomyces pombe haploid cells to matingpheromones. FEMS Microbiol Lett 143:41–45
Klar AJS, Miglio LM (1986) Initiation of meiotic recombination bydouble-strand DNA breaks in Sz. pombe. Cell 46:725–731
Ladds G, Davey J (2000) Sxa2 is a serine carboxypeptidase thatdegrades extracellular P-factor in the fission yeastSchizosaccharomyces pombe. Mol Microbiol 36:377–390
Ladds G, Rasmussen EM, Young T, Nielsen O, Davey J (1996)The sxa2-dependent inactivation of the P-factor mating pher-omone in the fission yeast Schizosaccharomyces pombe. MolMicrobiol 20:35–42
Landry S, Hoffman CS (2001) The git5 Gb and git11 Gc form anatypical Gbc dimer acting in the fission yeast glucose/cAMPpathway. Genetics 157:1159–1168
252

Manfredi JP, Klein C, Herrero JJ, Byrd DR, Trueheart J,Wiesler WT, Fowlkes DM, Broach JR (1996) Yeast a-matingfactor structure–activity relationship derived from geneticallyselected peptide agonists and antagonists of STE2p. Mol CellBiol 16:4700–4709
Martin H, Rodriquez-Pachon JM, Ruiz C, Nombela C, Molina M(2000) Regulatory mechanisms for modulation of signalingthrough the cell integrity Slt2-mediated pathway inSaccharomyces cerevisiae. J Biol Chem 275:1511–1519
Mattison CP, Spencer SS, Kresgee KA, Lee J, Ota IM (1999)Differential regulation of the cell wall integrity mitogen-acti-vated protein kinase pathway in budding yeast by the proteintyrosine phosphatases Ptp2 and Ptp3. Mol Cell Biol 19:7651–7660
Maundrell K (1993) Thiamine-repressible expression vectors pREPand pRIP for fission yeast. Gene 123:127–130
Millar JBA, Buck V, Wilkinson MG (1995) Pyp1 and Pyp2PTPases dephosphorylate an osmosensing MAP kinase con-trolling cell size at division in fission yeast. Genes Dev 9:2117–2130
Nielsen O, Davey J, Egel R (1992) The ras1 function of Schizo-saccharomyces pombe mediates pheromone-induced transcrip-tion. EMBO J 11:1391–1395
Nielsen O, Friis T, Kjaerulff S (1996) The Schizosaccharomycespombe map1 gene encodes an SRF/MCM1-related proteinrequired for P-cell specific gene expression. Mol Gen Genet253:387–392
Pereira PS, Jones NC (2001) The RGS domain-containing fissionyeast protein, Rgs1p, regulates pheromone signalling and isrequired for mating. Genes Cells 6:789–802
Shiozaki K, Russell P (1995) Cell-cycle control linked to extracel-lular environment by MAP kinase pathway in fission yeast.Nature 378:739–743
Sugiura R, Toda T, Shuntoh H, Yanagida M, Kuno T (1998)pmp1+, a suppressor of calcineurin deficiency, encodes a novelMAP kinase phosphatase in fission yeast. EMBO J 17:140–148
Toda T, Dhut S, Supertifurga G, Gotoh Y, Nishida E, Sugiura R(1996) The fission yeast pmk1+ gene encodes a novel mitogen-
activated protein kinase homolog which regulates cell integrityand functions co-ordinately with the protein kinase C pathway.Mol Cell Biol 16:6752–6764
Wang S-H, Xue C-B, Nielsen O, Davey J, Naider F (1994)Chemical synthesis of the M-factor mating pheromone fromSchizosaccharomyces pombe. Yeast 10:595–601
Watanabe Y, Irie K, Matsumoto K (1995) Yeast RLM1 encodes aserum response factor-like protein that may function down-stream of the Mpk1 (Slt2) mitogen-activated protein kinasepathway. Mol Cell Biol 15:5740–5749
Watson P, Davis K, Didmon M, Broad P, Davey J (1999) An RGSprotein regulates the pheromone response in the fission yeastSchizosaccharomyces pombe. Mol Microbiol 33:623–634
Wurgler-Murphy SM, Maeda T, Witten EA, Saito H (1997) Reg-ulation of the Saccharomyces cerevisiae HOG1 mitogen-activated protein kinase by the PTP2 and PTP3 protein tyrosinephosphatases. Mol Cell Biol 17:1289–1297
Yabana N, Yamamoto M (1996) Schizosaccharomyces pombemap1+ encodes a MADS-box family protein required for celltype-specific gene expression. Mol Cell Biol 16:3420–3428
Yamawaki-Kataoka Y, Tamaoki T, Choe H-R, Tanaka H,Kataoka T (1989) Adenylate cyclases in yeast: a comparison ofthe genes from Schizosaccharomyces pombe and Saccharomycescerevisiae. Proc Natl Acad Sci USA 86:5693–5697
Young D, Riggs M, Field J, Vojtek A, Broek D, Wigler M (1989)The adenylyl cyclase gene from Schizosaccharomyces pombe.Proc Natl Acad Sci USA 86:7989–7993
Yun D-F, Laz TM, Clements JM, Sherman F (1996) mRNA se-quences influencing translation and the selection of AUG ini-tiator codons in the yeast Saccharomyces cerevisiae. MolMicrobiol 19:1225–1239
Zaitsevskaya-Carter T, Cooper JA (1997) Spm1, a stress-activatedMAP kinase that regulates morphogenesis in Sz. pombe. EMBOJ 16:1318–1331
Zhan X-L, Deschenes RJ, Guan K-L (1997) Differential regulationof FUS3 MAP kinase by tyrosine-specific phosphatases PTP2/PTP3 and dual-specificity phosphatase MSG5 in Saccharomy-ces cerevisiae. Genes Dev 11:1690–1702
253
Copyright © 2022 FDOKUMEN




















