
Transcriptional and post-translational regulation of neutral trehalase inSchizosaccharomyces pombe...
10
Transcriptional and post-translational regulation of mouse cation transport regulator homolog 1 Kentaro Oh-hashi • Yuki Nomura • Kiyo Shimada • Hisashi Koga • Yoko Hirata • Kazutoshi Kiuchi Received: 23 January 2013 / Accepted: 17 April 2013 Ó Springer Science+Business Media New York 2013 Abstract Recently, cation transport regulator homolog 1 (Chac1) has been identified as a novel pro-apoptotic factor in cells under endoplasmic reticulum (ER) stress. Of the three major ER stress sensors, it is suggested that ATF4 participates in the transcriptional regulation of Chac1 gene expression. The precise characterization of the Chac1 promoter, however, has not yet been elucidated. In this study, we detected the induction of Chac1 mRNA expres- sion using DNA array analysis and RT-PCR of thapsigar- gin (Tg)-inducible genes in Neuro2a cells. Chac1 mRNA expression was also induced immediately following treat- ment with tunicamycin (Tm) and brefeldin A. Character- ization of the mouse Chac1 promoter activity using a luciferase reporter assay revealed that the CREB/ATF element and amino acid response element in the mouse Chac1 promoter are functional and respond to Tm stimu- lation and ATF4 overexpression. Mutations in either ele- ment in the Chac1 promoter did not inhibit the responsiveness of this promoter to Tm and ATF4; however, mutations in both of these elements dramatically decreased the basal activity and response to ER stress stimuli. In addition to the transcriptional regulation, we found that Chac1 protein expression was only detected in the presence of MG132, a proteasome inhibitor, even though mouse Chac1 gene was transiently overexpressed in Neuro2a cells. Taken together, we are the first to demonstrate the transcriptional and post-translational regulation of Chac1 expression in a neuronal cell line. Keywords ER stress ATF4 Chac1 Abbreviations AARE Amino acid response element AP1 Activator protein 1 ATF Activating transcription factor BFA Brefeldin A bZIP Basic leucine zipper Chac1 Cation transport regulator homolog 1 C/EBP CCAAT-enhancer-binding protein CHOP C/EBP homologous protein CREB cAMP response element binding protein CRELD2 Cysteine-rich with EGF-like domains 2 eIF2a Eukaryotic translation initiation factor 2A ER Endoplasmic reticulum ERSE ER stress response element GADD153 Growth arrest and DNA-damage-inducible protein 153 GRP78 78 kDa glucose-regulated protein IRE1 Inositol-requiring enzyme 1 MANF Mesencephalic astrocyte-derived neurotrophic factor Nrf2 Nuclear factor (erythroid-derived 2)-like 2 PKA Protein kinase A PERK PRKR-like endoplasmic reticulum kinase RT-PCR Reverse transcription polymerase chain reaction Kentaro Oh-hashi, Yuki Nomura contributed equally to this study. K. Oh-hashi (&) Y. Nomura Y. Hirata K. Kiuchi Department of Biomolecular Science, Faculty of Engineering, Gifu University, 1-1 Yanagido, Gifu 501-1193, Japan e-mail: [email protected] K. Shimada H. Koga Kazusa DNA Research Institute, 2-6-7 Kazusa-Kamatari, Kisarazu, Chiba 292-0818, Japan Y. Hirata K. Kiuchi United Graduate School of Drug Discovery and Medical Information Sciences, Gifu University, 1-1 Yanagido, Gifu 501-1193, Japan 123 Mol Cell Biochem DOI 10.1007/s11010-013-1663-1
Transcript of Transcriptional and post-translational regulation of neutral trehalase inSchizosaccharomyces pombe...

Transcriptional and post-translational regulation of mouse cationtransport regulator homolog 1
Kentaro Oh-hashi • Yuki Nomura • Kiyo Shimada •
Hisashi Koga • Yoko Hirata • Kazutoshi Kiuchi
Received: 23 January 2013 / Accepted: 17 April 2013
� Springer Science+Business Media New York 2013
Abstract Recently, cation transport regulator homolog 1
(Chac1) has been identified as a novel pro-apoptotic factor
in cells under endoplasmic reticulum (ER) stress. Of the
three major ER stress sensors, it is suggested that ATF4
participates in the transcriptional regulation of Chac1 gene
expression. The precise characterization of the Chac1
promoter, however, has not yet been elucidated. In this
study, we detected the induction of Chac1 mRNA expres-
sion using DNA array analysis and RT-PCR of thapsigar-
gin (Tg)-inducible genes in Neuro2a cells. Chac1 mRNA
expression was also induced immediately following treat-
ment with tunicamycin (Tm) and brefeldin A. Character-
ization of the mouse Chac1 promoter activity using a
luciferase reporter assay revealed that the CREB/ATF
element and amino acid response element in the mouse
Chac1 promoter are functional and respond to Tm stimu-
lation and ATF4 overexpression. Mutations in either ele-
ment in the Chac1 promoter did not inhibit the
responsiveness of this promoter to Tm and ATF4; however,
mutations in both of these elements dramatically decreased
the basal activity and response to ER stress stimuli. In
addition to the transcriptional regulation, we found that
Chac1 protein expression was only detected in the presence
of MG132, a proteasome inhibitor, even though mouse
Chac1 gene was transiently overexpressed in Neuro2a
cells. Taken together, we are the first to demonstrate the
transcriptional and post-translational regulation of Chac1
expression in a neuronal cell line.
Keywords ER stress � ATF4 � Chac1
Abbreviations
AARE Amino acid response element
AP1 Activator protein 1
ATF Activating transcription factor
BFA Brefeldin A
bZIP Basic leucine zipper
Chac1 Cation transport regulator homolog 1
C/EBP CCAAT-enhancer-binding protein
CHOP C/EBP homologous protein
CREB cAMP response element binding protein
CRELD2 Cysteine-rich with EGF-like domains 2
eIF2a Eukaryotic translation initiation factor 2A
ER Endoplasmic reticulum
ERSE ER stress response element
GADD153 Growth arrest and DNA-damage-inducible
protein 153
GRP78 78 kDa glucose-regulated protein
IRE1 Inositol-requiring enzyme 1
MANF Mesencephalic astrocyte-derived
neurotrophic factor
Nrf2 Nuclear factor (erythroid-derived 2)-like 2
PKA Protein kinase A
PERK PRKR-like endoplasmic reticulum kinase
RT-PCR Reverse transcription polymerase chain
reaction
Kentaro Oh-hashi, Yuki Nomura contributed equally to this study.
K. Oh-hashi (&) � Y. Nomura � Y. Hirata � K. Kiuchi
Department of Biomolecular Science, Faculty of Engineering,
Gifu University, 1-1 Yanagido, Gifu 501-1193, Japan
e-mail: [email protected]
K. Shimada � H. Koga
Kazusa DNA Research Institute, 2-6-7 Kazusa-Kamatari,
Kisarazu, Chiba 292-0818, Japan
Y. Hirata � K. Kiuchi
United Graduate School of Drug Discovery and Medical
Information Sciences, Gifu University, 1-1 Yanagido,
Gifu 501-1193, Japan
123
Mol Cell Biochem
DOI 10.1007/s11010-013-1663-1

Tg Thapsigargin
Tm Tunicamycin
TRIB3 Tribbles homolog 3
Introduction
The endoplasmic reticulum (ER) is responsible for the
folding and modification of newly synthesized transmem-
brane and secretory proteins [1, 2]. Some pathophysio-
logical conditions disrupt ER function and cause the
accumulation of unfolded and/or misfolded proteins in the
ER [3, 4]. These conditions, known as ER stress, activate
various stress responses that are mediated by the three ER-
resident stress sensors PERK-ATF4 [5], IRE1 [6], and
ATF6 [7]. A variety of genes have been identified as the
downstream targets of these sensors [8–11]. Recently, we
found that the induction of CRELD2 and MANF mRNA
expression in Neuro2a cells is ER stress-dependent using
DNA array analysis and RT-PCR [12, 13]. It has been
reported that ATF6 and/or IRE1 regulate the transcription
of the CRELD2 and MANF genes via the ER stress
response element (ERSE) or ERSE-II in the 50 flanking
promoter region of each gene, respectively [12, 14]. ER-
resident chaperones, such as GRP78 [15, 16], which are
responsible for proper protein folding in the ER, are well-
known targets of ATF6 and IRE1. Furthermore, a few
factors involved in ER-associated degradation (e.g., ER
degradation-enhancing a-mannosidase-like protein and E3
ubiquitin-protein ligase HRD1) have also been reported to
be the targets of IRE1 [17, 18].
ATF4 is preferentially translated in response to various
types of environmental stress signals including ER stress
[19], which induce eIF2a phosphorylation. In addition to
ER-resident PERK [5], other kinases, such as GCN2 [20],
heme-regulated inhibitor [21] and double-stranded RNA-
dependent kinase [22] which are activated by serum starva-
tion, oxidative stress and viral infection, respectively, have
been reported to participate in various cellular processes
through the eIF2a-ATF4 pathway [23, 24]. Therefore, this
pathway has been referred to as the integrated stress response
and implicated in various diseases (e.g., neurodegenerative
diseases and inflammations) [19, 25, 26]. In our DNA array
analysis, we also found that the downstream targets of the
PERK-ATF4 pathway GADD153/CHOP [27], TRIB3 [28]
and Chac1 [29] were remarkably upregulated in Tg-treated
Neuro2a cells. The GADD153/CHOP promoter contains
multiple stress responsive elements, including an activator
protein 1 (AP1) element, amino acid response element
(AARE) and ERSE. The roles each of these elements plays in
this promoter have been characterized under various stimuli
[30–33]. The transcriptional regulation of the TRIB3 gene
via the ATF4-CHOP pathway has also been studied in detail
[28]. Chac1 was first identified as a novel ER stress-inducible
gene in endothelial cells treated with oxidized phospholipids
[29]. It has been suggested that ATF4 plays an important role
in the regulation of Chac1 gene expression [29, 34–37], but a
detailed characterization of the Chac1 gene promoter has not
yet been reported. An analysis of the 50-flanking nucleotide
sequence of the mouse Chac1 gene suggests that there are
two putative consensus sequences, a CREB/ATF element
and AARE), in this region; however, relationships between
the two and their functions in regulating Chac1 promoter
activity under ER stress have not been elucidated. Therefore,
we isolated a mouse Chac1 promoter fragment, which was
approximately 400 bp length, and characterized its promoter
activity in response to ER stress stimuli and ATF4 overex-
pression. In addition to the transcriptional regulation of the
Chac1 gene, we also investigated the involvement of the
proteasome pathway in the post-translational instability of
the Chac1 protein.
Materials and methods
Materials
Thapsigargin (Tg), tunicamycin (Tm), brefeldin A (BFA),
forskolin (F) were obtained from Sigma-Aldrich. MG132
was purchased from Peptide Institute. Antibodies against
Chac1 and actin were purchased from Abcam and Cal-
biochem, respectively.
Construction of plasmids
For generating the reporter construct of the mouse Chac1
promoter, genomic DNA from Neuro2a cells was extrac-
ted, and the mouse Chac1 promoter (-334/?112) was
amplified using polymerase chain reaction (PCR). The
PCR product was then cloned into the pGL3-Basic
(pGL3b) vector (Promega). Other constructs containing
either deleted or mutated mouse Chac1 promoters were
also prepared by PCR. The promoter region was defined
using the NCBI Reference Sequences NM_026929.4 and
NM_024111.3. For generating mouse ATF4, cDNA was
amplified from Neuro2a cell-derived mRNA by reverse
transcriptional-polymerase chain reaction (RT-PCR) and
then cloned into the pFLAG-CMV vector. Mouse Chac1
cDNA was amplified from C57/BL6 mouse brain-derived
mRNA by RT-PCR and cloned into the pcDNA3.1 vector.
Cell culture and treatment
We used Neuro2a cells for characterization of Chac1 gene
according to our previous studies about ER stress response
Mol Cell Biochem
123

[12, 13]. Neuro2a cells were maintained in Dulbecco’s
modified eagles minimum essential medium containing
8 % fetal bovine serum. The transfection of each vector
used in this study was performed using the Lipofectamine-
Plus reagent (Life Technologies) according to the manu-
facturer’s instructions. Neuro2a cells were treated with
thapsigargin (Tg) (0.1 lM), tunicamycin (Tm) (2 lg/ml),
brefeldin A (BFA) (5 lg/ml), serum starvation (serum free,
SF), forskolin (F) (10 lM) or MG132 (MG) (20 lM) for
the indicated times.
RT-PCR
To estimate the expression level of each gene by RT-PCR,
total RNA was extracted from cells lysed with Trizol (Life
Technologies) [12, 13]. After incubation of total RNA for
10 min at 72 �C, cDNA was generated by reverse transcrip-
tion using total RNA (0.5 lg/10 ll), DTT (10 mM), dNTP
(0.5 mM), random ninemers (0.05 lg/10 ll), RNaseOUT (2
U/10 ll) (Life Technologies) and prime superscript III
RNase- reverse transcriptase (RT) (25 U/10 ll) (Life Tech-
nologies) for 60 min at 42 �C according to the manufacturer’s
instructions. Specific cDNAs were mixed and amplified using
a PCR reaction mixture containing the indicated primer pair
(1 lM), dNTP (0.1 mM), rTaq (0.25 U/10 ll) (Taq PCR kit,
Takara). The following RT-PCR primers were used in this
study: Chac1 sense primer, 50-CATAGGGGCAGCGACA-
AGATG-30, Chac1 antisense primer 50-CTGTGTGGCAAT-
GACCTCTTC-30, GADD153 sense primer 50-GAATAACA
GCCGGAACCTGA-30, GADD153 antisense primer 50-GGACGCAGGGTCAAGAGTAG-30, GRP78 sense primer
50-ACCAATGACCAAAACCGCCT-30, GRP78 antisense
primer 50-GAGTTTGCTGATAATTGGCTGAAC-30, GAPDH
sense primer, 50-ACCACAGTCCATGCCATCAC-30 and
GAPDH antisense primer, 50-TCCACCACCCTGTTGC
TGTA-30. The typical reaction conditions were 30 s at
96 �C, 30 s at 58 �C and 30 s at 72 �C. The results represent
20–28 cycles of amplification. The cDNA amplification
products were separated by electrophoresis on 2.0 % agarose
gels and visualized using ethidium bromide. The relative
mRNA level of Chac1 gene was calculated by comparison of
GAPDH-normalized values with the level of untreated
control cells using NIH imaging.
Reporter assay
Each reporter construct (0.1 lg) and the pGL4.70 (Rluc)
(0.04 lg) vector (Promega), an internal control, were
transfected into Neuro2a cells in a 48-well plate as previ-
ously described [12]. Twenty-four hours after transfection,
the cells were treated with Tm (2 lg/ml) or vehicle for
12 h. To determine the effect of ATF4 overexpression on
the reporter activity, the ATF4-expression vector or empty
vector (mock) (0.01 lg) was co-transfected with the
reporter construct into cells that were then cultured for
36 h. After each treatment, the cells were lysed with
1 9 passive lysis buffer (Promega) for 15 min at room
temperature and briefly centrifuged. The luciferase activity
in each lysate was measured using a Dual-Luciferase assay
system (Promega). The reporter activity in each lysate was
normalized to the co-transfected Renilla luciferase activity
and shown as the relative luciferase activity.
Western blot analysis
Cells were lysed with homogenate butter [20 mM Tris–
HCl (pH 8.0) containing 137 mM NaCl, 2 mM EDTA,
10 % glycerol, 1 % TritonX-100, 1 mM PMSF, 10 lg/ml
leupeptin and 10 lg/ml pepstatin A]. After the protein
concentration was determined, each cell lysate was dis-
solved in sodium dodecyl sulfate (SDS)-Laemmli sample
buffer [62.5 mM Tris–HCl (pH 6.8), 2 % SDS and 10 %
glycerol]. Equal amounts of cell lysate were separated on 8
or 12.5 % SDS–polyacrylamide electrophoresis gels,
immunoblotted onto polyvinylidene difluoride membrane
(GE Healthcare) and identified by enhanced chemilumi-
nescence (GE Healthcare) using antibodies against Chac1
(1:1,000) and actin (1: 5,000).
Statistical analysis
The results are expressed mean ± SD of the indicated
number. Statistical analysis was carried out by one way-
ANOVA followed by Fischer’s PLSD test. p \ 0.05 was
considered to be statistically significant.
Results
ER stress-upregulated Chac1 mRNA expression
in Neuro2a cells
The expression of Chac1, a novel ER stress-inducible gene,
has been reported to be upregulated under certain patho-
physiological conditions [29, 34–37]. Mungrue et al.
reported that Chac1 is a downstream target of ATF4 [29];
however, the precise mechanism of its transcriptional
regulation under ER stress has not yet been characterized.
Previously, we reported that the expression of the CRELD2
and MANF genes increased in Tg-treated Neuro2a cells
using DNA array analysis [12, 13]. This comprehensive
analysis also showed that Chac1 mRNA might be included
in the Tg-inducible genes. Therefore, we examined the ER
stress-induced expression of mouse Chac1 mRNA in
Neuro2a cells by RT-PCR. As shown in Fig. 1A, Tg
Mol Cell Biochem
123

treatment caused Chac1 mRNA expression, as well as
GADD153 and GRP78 mRNA expression, to increase in a
time-dependent manner. The relative mRNA level of
Chac1 gene, which was normalized with that of GAPDH
gene, was increased by 1.5-, 1.8-, and 2.3-fold after Tg
stimulation for 4, 8, and 12 h, respectively. Other ER
stress-inducing reagents, such as Tm and BFA (1.7- and
1.5-fold comparing with untreated control cells), but not
serum free (SF, 1.0-fold), also enhanced the expression
levels of Chac1 mRNA as well as those of GADD153 and
GRP78 mRNA (Fig. 1B).
Characterization of ER stress-induced mouse Chac1
promoter activity in Neuro2a cells
As shown in Fig. 2, the nucleotide sequence of the mouse
Chac1 promoter region is similar to the human Chac1
promoter region. They contain a highly homologous
domain around the CREB/ATF element and AARE (I and
II in Fig. 2), which seem to be recognized by ATF4 and its
related transcription factors.
To evaluate the roles these elements in regulating the
Chac1 promoter activity, various luciferase reporter
BA
Con Tg Tm BFA SF
GAPDH
Chac1
0 4 8 12
GRP78GRP78
GADD153GADD153
GAPDH
Chac1
Time (h)
Fig. 1 ER stress-induced
Chac1 mRNA expression in
Neuro2a cells. A Neuro2a cells
were treated with 0.1 lM Tg for
the indicated times. B Neuro2a
cells were treated with Tg
(0.1 lM), Tm (2 lg/ml), BFA
(5 lg/ml), serum free (SF) or
vehicle (Con) for 8 h. The total
RNA isolated from each sample
was subjected to RT-PCR as
described in the ‘‘Materials and
methods’’ section
ACCGTTTAGACCACTGAGAGGGCCTTCCCGGGAGA----CGGCGGGTGCCCGCGCCCATT **** *** * * ****** ** * ******* ****AGGGGCCCAGGTCATGAG-GGGGCGGCTGGGGAGACGCCCGCCCGGTGCCC--GCCCTGG
CTAAGGCCCGTTCCCGGTGCCAGCAGCCTGACGCAATCTGACTCGCCCTTGCTGCATGCC *** ** *** **** ** ************* ** *****************GCCCGGCGGGTGCCCCCTGCCCGCCGCCTGACGCAATCCGAGGCGCCCTTGCTGCATGCC
AGTTCTGGGGAAACCTTCCCTGCCGTTGCATCACTCCTGCCAGGTCTGCCACGGGAGGAG * **** ******* **************************** ****** **** *GGCGCTGGAGAAACCTCCCCTGCCGTTGCATCACTCCTGCCAGGTTTGCCAC-AGAGGTG
GAGCCGAACCAATCAGCACGCGCGTTGGCCCTGGGGCGGGCTGCTCTGGTACCCACTCGG**** ************ ***** ** **** **GAGCTGAACCAATCAGCGGGCGCG------CT-----GGGC-----------CC------
AGGGCGGCGCGGTTAACCGAGAAGCAAAATCTAAAGGTGATTGGCCAGAAAGAGATTTGC ************* ******** **** *** ********** ** * *******---GCGGCGCGGTTAAGTGAGAAGCAGAATCCGAAGTTGATTGGCCAAAAGGGGATTTGC
CCCGCCCCAATCATCGGGACAGCTGGAGGCAGTCTGAGGGCCTTCCTGGCCGGAGGGTTT*** ********* ** * ******* ** ** * * * *** *** ** *****CCCACCCCAATCACCGCGGCAGCTGGCCGCGGTGGGCGTGGTCTCCCGGCAAGAAGGTTT
AAAGAGCGCCTCTGGAGGGACCACTCAGCTGGAACGACCGATCGGTGCCAGGCCAGGTGT**** ******* *** * ********** * ***** ******************AAAGGGCGCCTCCATGGGGGGCGCTCAGCTGGAGCTACCGAGCGGTGCCAGGCCAGGTGT
ACGCGTCCGTCGGTCCTTCCGTGCCCGTGCCGGAGACCAGTCTCGGAGGCCACCCGGGTC ************* ********************* * ******* ** *** *GTGCGTCCGTCGGTCTTTCCGTGCCCACGCCGGAGACCAGCCCCGGAGGCCGCCTGGGCC
mouse
human
+142
-334
+127
-318I
II
Fig. 2 Nucleotide sequences of
the 50-flanking region of the
mouse and human Chac1 gene.
Nucleotide sequences conserved
between the 50-flanking
promoter region of the mouse
and human Chac1 genes are
indicated with asterisks. The
conserved CREB/ATF element
(I) and AARE (II) are shown in
a box. The transcriptional
direction and putative
transcriptional start sites of the
mouse and human Chac1 gene
are shown with arrows and
arrowheads, respectively
Mol Cell Biochem
123
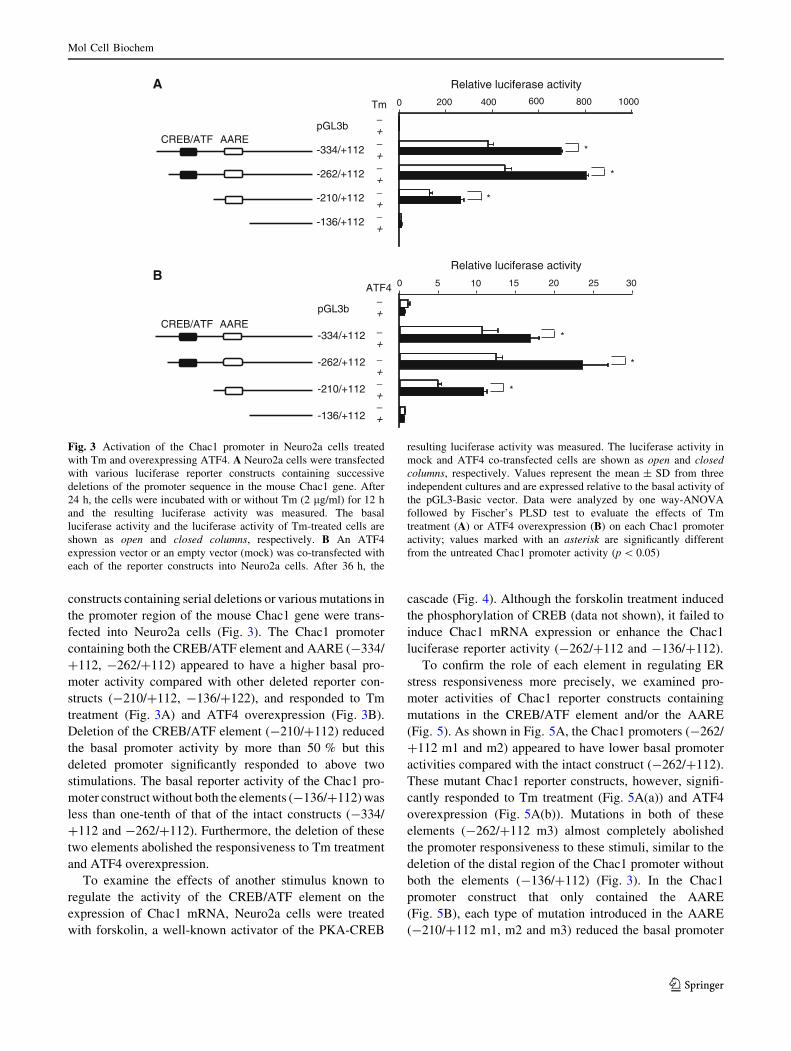
constructs containing serial deletions or various mutations in
the promoter region of the mouse Chac1 gene were trans-
fected into Neuro2a cells (Fig. 3). The Chac1 promoter
containing both the CREB/ATF element and AARE (-334/
?112, -262/?112) appeared to have a higher basal pro-
moter activity compared with other deleted reporter con-
structs (-210/?112, -136/?122), and responded to Tm
treatment (Fig. 3A) and ATF4 overexpression (Fig. 3B).
Deletion of the CREB/ATF element (-210/?112) reduced
the basal promoter activity by more than 50 % but this
deleted promoter significantly responded to above two
stimulations. The basal reporter activity of the Chac1 pro-
moter construct without both the elements (-136/?112) was
less than one-tenth of that of the intact constructs (-334/
?112 and -262/?112). Furthermore, the deletion of these
two elements abolished the responsiveness to Tm treatment
and ATF4 overexpression.
To examine the effects of another stimulus known to
regulate the activity of the CREB/ATF element on the
expression of Chac1 mRNA, Neuro2a cells were treated
with forskolin, a well-known activator of the PKA-CREB
cascade (Fig. 4). Although the forskolin treatment induced
the phosphorylation of CREB (data not shown), it failed to
induce Chac1 mRNA expression or enhance the Chac1
luciferase reporter activity (-262/?112 and -136/?112).
To confirm the role of each element in regulating ER
stress responsiveness more precisely, we examined pro-
moter activities of Chac1 reporter constructs containing
mutations in the CREB/ATF element and/or the AARE
(Fig. 5). As shown in Fig. 5A, the Chac1 promoters (-262/
?112 m1 and m2) appeared to have lower basal promoter
activities compared with the intact construct (-262/?112).
These mutant Chac1 reporter constructs, however, signifi-
cantly responded to Tm treatment (Fig. 5A(a)) and ATF4
overexpression (Fig. 5A(b)). Mutations in both of these
elements (-262/?112 m3) almost completely abolished
the promoter responsiveness to these stimuli, similar to the
deletion of the distal region of the Chac1 promoter without
both the elements (-136/?112) (Fig. 3). In the Chac1
promoter construct that only contained the AARE
(Fig. 5B), each type of mutation introduced in the AARE
(-210/?112 m1, m2 and m3) reduced the basal promoter
Relative luciferase activity
0 5 10 15 20 25 30
-136/+112
-210/+112
-262/+112
-334/+112
pGL3b
CREB/ATF AARE
pGL3b
-334/+112
-262/+112
-210/+112
-136/+112
CREB/ATF AARE
0 200 400 600 800 1000
Relative luciferase activity
Tm−+−+−+−+−+
−+
−+
−+−+−+
ATF4
*
*
*
*
*
*
B
A
Fig. 3 Activation of the Chac1 promoter in Neuro2a cells treated
with Tm and overexpressing ATF4. A Neuro2a cells were transfected
with various luciferase reporter constructs containing successive
deletions of the promoter sequence in the mouse Chac1 gene. After
24 h, the cells were incubated with or without Tm (2 lg/ml) for 12 h
and the resulting luciferase activity was measured. The basal
luciferase activity and the luciferase activity of Tm-treated cells are
shown as open and closed columns, respectively. B An ATF4
expression vector or an empty vector (mock) was co-transfected with
each of the reporter constructs into Neuro2a cells. After 36 h, the
resulting luciferase activity was measured. The luciferase activity in
mock and ATF4 co-transfected cells are shown as open and closedcolumns, respectively. Values represent the mean ± SD from three
independent cultures and are expressed relative to the basal activity of
the pGL3-Basic vector. Data were analyzed by one way-ANOVA
followed by Fischer’s PLSD test to evaluate the effects of Tm
treatment (A) or ATF4 overexpression (B) on each Chac1 promoter
activity; values marked with an asterisk are significantly different
from the untreated Chac1 promoter activity (p \ 0.05)
Mol Cell Biochem
123

activity and rendered the mutant construct unresponsive to
Tm treatment (Fig. 5B(a)) and ATF4 overexpression
(Fig. 5B(b)).
Post-translational regulation of Chac1
via the proteasome pathway in Neuro2a cells
In addition to the transcriptional regulation of the mouse
Chac1 gene expression, we also investigated the expression
of the mouse Chac1 protein in Neuro2a cells transiently
transfected with the mouse Chac1 gene because the
intrinsic Chac1 protein expression level could not be
detected in cells cultured in the presence or absence of ER
stress stimuli (data not shown). The expression of the
Chac1 protein could only be detected 36 h after transfec-
tion in cells treated with MG132, a proteasome inhibitor
(Fig. 6). Furthermore, the expression of the intrinsic Chac1
mRNA was hardly affected by MG132 treatment, which is
unlike the effects seen with the Tg and Tm treatments.
Discussion
The induction of Chac1 mRNA expression in several
models [34–37] has been reported since Chac1 was first
identified as a novel ER stress-inducible gene in endothe-
lial cells treated with oxidized phospholipids [29]. Our
study using DNA array analysis and RT-PCR shows that
Tg-triggered ER stress also increases the expression level
of Chac1 mRNA in a neuronal cell line, Neuro2a cells. We
also discovered that Chac1 expression is regulated by both
transcriptional and post-translational mechanisms. The
expression pattern of Chac1 mRNA in Neuro2a cells is
similar to other typical ER stress-inducible genes, such as
GADD153/CHOP [27] and GRP78 [15, 16]. In the 50
upstream region of the mouse Chac1 gene, there are two
consensus sequences, a CREB/ATF element and an AARE,
that are well conserved between the mouse and human
Chac1 promoter region. Typically, the three ER-resident
stress sensors PERK [5], IRE1 [6] and ATF6 [7] are
simultaneously activated in response to various stimuli that
trigger ER dysfunction and induce the expression of sev-
eral downstream target genes [8–11]. The active form of
PERK phosphorylates eIF2a, which induces ATF4 trans-
lation. Furthermore, phospho-eIF2a inhibits global protein
synthesis [23]. It has been demonstrated that the increased
level of ATF4 generated in response to ER stress specifi-
cally recognizes the AARE/NSRE and induces the tran-
scription of target genes, such as GADD153/CHOP [27,
33], TRIB3 [28] and asparagines synthetase [38]. Regard-
ing the regulation of the Chac1 promoter, Romanski et al.
[35] recently reported that the CREB/ATF element, just
upstream of the AARE, in the human Chac1 promoter
specifically responds to ATF4 overexpression by enhanc-
ing the promoter activity in human aortic endothelial cells.
In our study, we show that while the CREB/ATF element
in the mouse Chac1 promoter, which is well conserved
with the human Chac1 promoter, is important for the basal
promoter activity, Chac1 reporter constructs lacking the
CREB/ATF element (-210/?112) or containing a muta-
tion in the CREB/ATF element (-262/?112 m1) still
respond to Tm treatment and ATF4 overexpression. Fur-
thermore, mutations in the AARE of the mouse Chac1
Con Tm F
GAPDH
Chac1
A B0 250 500 750 1000
Relative luciferase activityTm F
pGL3b
-262/+112
-136/+112
+−
− −
+ −
+−
− −
+ −
+−
− −
+ −
*
*
Fig. 4 Forskolin failed to induce Chac1 mRNA expression or
enhance its promoter activity in Neuro2a cells. A Neuro2a cells were
treated with 10 lM forskolin (F), 2 lg/ml Tm or vehicle (Con) for
12 h. The total RNA isolated from each sample was subjected to RT-
PCR as described in the ‘‘Materials and methods’’ section. B Neuro2a
cells were transfected with the indicated Chac1 reporter construct.
After 24 h, the cells were incubated with forskolin, Tm, or vehicle for
12 h and the resulting luciferase activity was measured. Values
represent the mean ± SD from three independent cultures and are
expressed relative to the basal activity of the pGL3-Basic vector. Data
were analyzed by one way-ANOVA followed by Fischer’s PLSD test
to evaluate the effect of each treatment on the Chac1 promoter
activity; values marked with an asterisk are significantly different
from the Tm-treated Chac1 promoter activity (p \ 0.05)
Mol Cell Biochem
123

promoter (-262/?112 m2) also appeared to have no effect
on the response of the promoter to these stimuli. Taken
together, the results from our mutant and deletion Chac1
reporter constructs suggest that while the CREB/ATF ele-
ment and AARE in the mouse Chac1 promoter respond to
ER stress stimuli, they do not act in a synergistic manner.
These two elements, however, may work together to
maintain the basal activity of the Chac1 promoter because
mutations in both of these elements (-262/?112 m3)
significantly decrease the reporter activity compared with
the single mutation Chac1 reporter constructs (-262/?112
m1 and m2).
A
tcgc
tcgc
-262 +112
CREB/ATF AARE
cagcag
-262/+112-262/+112 m1-262/+112 m2-262/+112 m3
0 100 200 300 400ATF4
pGL3b
-262/+112 m1
-262/+112 m2
-262/+112 m3
-262/+112CREB/ATF AARE
Relative luciferase activity
pGL3b
-262/+112 m1
-262/+112 m2
-262/+112 m3
-262/+112CREB/ATF AARE
Relative luciferase activity
0 100 200 300 400 500 600Tm−+−+−+−+−+
−+−+−+−+−+
*
*
*
*
*
*
(b)
(a)
-210 +112AARE
caggac
gcga
-210/+112-210/+112 m1-210/+112 m2-210/+112 m3
0 20 40 60 80 100 120
Relative luciferase activity
-210/+112 m1
-210/+112 m2
-210/+112 m3
-136/+112
-210/+112
pGL3bAARE
0 100 200 300 400
Relative luciferase activity
Tm
pGL3b
-136/+112
-210/+112
-210/+112 m1
-210/+112 m2
-210/+112 m3
AARE
B
−+−+−+−+−+−+
ATF4
−+−+−+−+−+−+
*
*
(b)
(a)
Fig. 5 Mutagenesis analysis of
the mouse Chac1 promoter. The
mutated sequences of the
CREB/ATF element and AARE
in the mouse Chac1 promoter
are shown in small letters. A,
B Neuro2a cells were
transfected with the indicated
luciferase reporter construct.
After 24 h, the cells were
incubated with or without Tm
(2 lg/ml) for 12 h, and the
resulting luciferase activity was
measured. The basal luciferase
activity and the luciferase
activity of the Tm-treated cells
are shown as open and closedcolumns, respectively (a). The
ATF4 expression vector or an
empty vector (mock) was co-
transfected with each of the
reporter constructs into Neuro2a
cells. After 36 h, the resulting
luciferase activity was measured
(b). The luciferase activity in
mock and ATF4 co-transfected
cells are shown as open and
closed columns, respectively.
Values represent the
mean ± SD from three
independent cultures and are
expressed relative to the basal
activity of the pGL3-Basic
vector. Data were analyzed by
one way-ANOVA followed by
Fischer’s PLSD test to evaluate
the effect of Tm treatment (a) or
ATF4 overexpression (b) on
each Chac1 promoter activity;
values marked with an asteriskare significantly different from
the untreated Chac1 promoter
activity (p \ 0.05)
Mol Cell Biochem
123

ATF4, also known as CREB2, is one of the CREB/ATF
family transcription factors [39]. It is well known that
CREB is phosphorylated by several kinases, including
PKA and MAP kinases, to regulate the expression of var-
ious different genes [40]. In this study, CREB was well
phosphorylated when cells were treated with forskolin
(data not shown). The Chac1 mRNA expression pattern
and the results from our luciferase reporter assays after
each of these treatments (Fig. 4) indicate that the PKA-
dependent signaling pathway does not induce the expres-
sion of the Chac1 gene through the CREB/ATF element
and AARE. We also examined the effect of PMA, a potent
PKC activator, on Chac1 transcription, but it neither
induced Chac1 mRNA expression nor enhanced the lucif-
erase reporter activity (data not shown). Therefore, our
results suggest that the expression of the mouse Chac1
gene is specifically upregulated by ER stress-triggering
stimuli.
It has been reported that Chac1 mRNA expression
increases in several pathological conditions [29, 34–37], but
the mechanism regulating Chac1 protein expression has not
been well characterized. Mungre et al. [29] demonstrated
that the overexpression of a GFP-tagged Chac1 caused
HEK293 and human aortic endothelial cells to undergo
apoptotic cell death though the activation of the apoptosis-
promoting factors caspase-3 and AIF and the suppression of
the apoptosis-inhibitory factor TNFRSF6B. The Chac1
protein expression pattern, however, has not yet been
studied. In this study, we transfected Neuro2a cells with a
mouse Chac1 gene construct that did not contain any epi-
tope-tags, but Chac1 protein could only be detected in cells
treated MG132. We also examined the intrinsic expression
of Chac1 protein in the presence or absence of MG132 and
other ER stress stimuli, but no specific signal could be
detected under our experimental conditions (data not
shown). As Chac1 protein is rapidly degraded in the pro-
teasome pathway and extremely unstable in cells, the
amount of intrinsic protein would be below the detection
level of our experimental techniques. Protein degradation
through the proteasome followed by ubiquitination is an
important mechanism that regulates various cellular signals
and processes, including the cell cycle, differentiation and
apoptosis [41]. Similarly, the expression of many factors
involved in the ER stress response has been reported to be
regulated in a transcriptional and translational manner by a
proteasome-dependent pathway. In the case of ATF4
expression, Jiang and Wek reported that proteasome inhi-
bition leads to the accumulation of ATF4 protein without
affecting the expression level of ATF4 mRNA [24]. They
also showed that the expression levels of GADD153 mRNA
and protein increased as a result of MG132-induced ATF4
stabilization and phospho-eIF2a-required ATF4 accumu-
lation. In this study, we also found that the amount of ATF4
protein increased in Neuro2a cells treated with MG132.
This ATF4 protein increase was more apparent with
MG132 treatment than with Tg or Tm treatment (data not
shown). Furthermore, the accumulation of ATF4 protein in
MG132-treated Neuro2a cells was independent of the
expression level of Chac1 mRNA. We also observed that
there was only a marginal increase in GADD153 mRNA
expression during proteasome inhibition. Even though the
precise mechanism preventing MG132 from significantly
inducing the expression of Chac1 and GADD153 mRNA in
Neuro2a cells despite the increase in ATF4 protein remains
unknown, differences in the MG132-stabilizing factors
between mouse embryonic fibroblasts and Neuro2a cells
may influence ATF4-mediated gene expression following
proteasome inhibition. It has been demonstrated that tran-
scription factors consisting of a basic leucine zipper domain
(bZIP) similar to ATF4 are capable of interacting with other
bZIP factors [42]. It has also been reported that ATF4
cooperates with C/EBPb [38], GADD153 [43], and Nrf2
[44] to enhance the expression of target genes. Therefore,
further studies characterizing these factors in several cell
types may be required to elucidate the transcriptional and
post-transcriptional regulation of Chac1 expression.
In conclusion, we first characterized two ATF4
responsive elements, CREB/ATF and AARE, in mouse
Con Tm
GAPDH
Chac1
Tg MG
GADD153
Chac1
Con MGTmTg
Actin
A BkDa
25
50
37
37
Fig. 6 Proteasome inhibition increased the Chac1 protein expression
level in Neuro2a cells. A Neuro2a cells were treated with 0.1 lM Tg,
2 lg/ml Tm, 20 lM MG132 (MG), or vehicle (Con) for 12 h. The
total RNA isolated from each sample was subjected to RT-PCR as
described in the ‘‘Materials and methods’’ section. B Twenty-four
hours after transfection of Chac1 expression construct, the cells were
incubated with Tg, Tm, MG132 (MG) or vehicle (Con) for 12 h. The
expression of Chac1 and actin was detected using the western blot
analysis described in the ‘‘Materials and methods’’ section
Mol Cell Biochem
123

Chac1 promoter using luciferase reporter assay. As it has
been demonstrated that a variety of signals triggered by ER
stress, oxidative stress and viral infection are integrated
into ATF4 pathways via eIF2a phosphorylation [19–22],
further characterization of Chac1 promoter in different
types of cells might contribute to understand various cel-
lular processes and diseases (e.g., neurodegenerative dis-
eases and inflammations) [25, 26]. In this study, we also
demonstrated that Chac1 protein is unstable within the cells
and its intracellular amount might be definitely regulated
via a proteasome system. Very recently, Kumar et al. [45]
reported that the pro-apoptotic feature of Chac1 is attrib-
uted to the specific degradation of glutathione through its
own c-glutamyl-cyclotransferase activity. Therefore, our
findings about the Chac1 promoter would support the
phenomena that the perturbation of cellular redox status
occurs in response to ER stress [46]. Further character-
ization of Chac1 may give new insight into preventing the
onset and progression of the various diseases caused by the
stress responses via the eIF2a-ATF4 pathway.
References
1. Gething MJ, Sambrook J (1992) Protein folding in the cell.
Nature 355:33–45
2. Helenius A, Marquardt T, Braakman I (1992) The endoplasmic
reticulum as a protein-folding compartment. Trends Cell Biol
2:227–231
3. Kim I, Xu W, Reed JC (2008) Cell death and endoplasmic
reticulum stress: disease relevance and therapeutic opportunities.
Nat Rev Drug Discov 7:1013–1030
4. Lindholm D, Wootz H, Korhonen L (2006) ER stress and neu-
rodegenerative diseases. Cell Death Differ 13:385–392
5. Harding HP, Zhang Y, Ron D (1999) Protein translation and
folding are coupled by an endoplasmic-reticulum-resident kinase.
Nature 397:271–274
6. Calfon M, Zeng H, Urano F, Till JH, Hubbard SR, Harding HP,
Clark SG, Ron D (2002) IRE1 couples endoplasmic reticulum
load to secretory capacity by processing the XBP-1 mRNA.
Nature 415:92–96
7. Zhu C, Johansen FE, Prywes R (1997) Interaction of ATF6 and
serum response factor. Mol Cell Biol 17:4957–4966
8. Okada T, Yoshida H, Akazawa R, Negishi M, Mori K (2002)
Distinct roles of activating transcription factor 6 (ATF6) and
double-stranded RNA-activated protein kinase-like endoplasmic
reticulum kinase (PERK) in transcription during the mammalian
unfolded protein response. Biochem J 366:585–594
9. Rutkowski DT, Kaufman RJ (2003) All roads lead to ATF4. Dev
Cell 4:442–444
10. Yoshida H, Matsui T, Yamamoto A, Okada T, Mori K (2001)
XBP1 mRNA is induced by ATF6 and spliced by IRE1 in
response to ER stress to produce a highly active transcription
factor. Cell 107:881–891
11. Lee AH, Iwakoshi NN, Glimcher LH (2003) XBP-1 regulates a
subset of endoplasmic reticulum resident chaperone genes in the
unfolded protein response. Mol Cell Biol 23:7448–7459
12. Oh-hashi K, Koga H, Ikeda S, Shimada K, Hirata Y, Kiuchi K
(2009) CRELD2 is a novel endoplasmic reticulum stress-induc-
ible gene. Biochem Biophys Res Commun 387:504–510
13. Oh-hashi K, Tanaka K, Koga H, Hirata Y, Kiuchi K (2012) Intra-
cellular trafficking and secretion of mouse mesencephalic astro-
cyte-derived neurotrophic factor. Mol Cell Biochem 363:35–41
14. Mizobuchi N, Hoseki J, Kubota H, Toyokuni S, Nozaki J, Naitoh
M, Koizumi A, Nagata K (2007) ARMET is a soluble ER protein
induced by the unfolded protein response via ERSE-II element.
Cell Struct Funct 32:41–50
15. Yamamoto K, Sato T, Matsui T, Sato M, Okada T, Yoshida H,
Harada A, Mori K (2007) Transcriptional induction of mamma-
lian ER quality control proteins is mediated by single or com-
bined action of ATF6a and XBP1. Dev Cell 13:365–376
16. Wang M, Ye R, Barron E, Baumeister P, Mao C, Luo S, Fu Y,
Luo B, Dubeau L, Hinton DR, Lee AS (2010) Essential role of
the unfolded protein response regulator GRP78/BiP in protection
from neuronal apoptosis. Cell Death Differ 17:488–498
17. Yoshida H, Matsui T, Hosokawa N, Kaufman RJ, Nagata K, Mori
K (2003) A time-dependent phase shift in the mammalian
unfolded protein response. Dev Cell 4:265–271
18. Yamamoto K, Suzuki N, Wada T, Okada T, Yoshida H, Kaufman
RJ, Mori K (2008) Human HRD1 promoter carries a functional
unfolded protein response element to which XBP1 but not ATF6
directly binds. J Biochem 144:477–486
19. Harding HP, Novoa I, Zhang Y, Zeng H, Wek R, Schapira M, Ron
D (2000) Regulated translation initiation controls stress-induced
gene expression in mammalian cells. Mol Cell 6:1099–1108
20. Berlanga JJ, Santoyo J, De Haro C (1999) Characterization of a
mammalian homolog of the GCN2 eukaryotic initiation factor 2akinase. Eur J Biochem 265:754–762
21. Chen JJ, London IM (1995) Regulation of protein synthesis by
heme-regulated eIF-2a kinase. Trends Biochem Sci 20:105–108
22. Clemens MJ, Elia A (1997) The double-stranded RNA-dependent
protein kinase PKR: structure and function. J Interferon Cytokine
Res 17:503–524
23. Vattem KM, Wek RC (2004) Reinitiation involving upstream
ORFs regulates ATF4 mRNA translation in mammalian cells.
Proc Natl Acad Sci USA 101:11269–11274
24. Jiang HY, Wek RC (2005) Phosphorylation of the a-subunit of
the eukaryotic initiation factor-2 (eIF2a) reduces protein syn-
thesis and enhances apoptosis in response to proteasome inhibi-
tion. J Biol Chem 280:14189–14202
25. Lange PS, Chavez JC, Pinto JT, Coppola G, Sun CW, Townes
TM, Geschwind DH, Ratan RR (2008) ATF4 is an oxidative
stress-inducible, prodeath transcription factor in neurons in vitro
and in vivo. J Exp Med 205:1227–1242
26. Woo CW, Cui D, Arellano J, Dorweiler B, Harding H, Fitzgerald
KA, Ron D, Taba I (2009) Adaptive suppression of the ATF4-
CHOP branch of the unfolded protein response by toll-like
receptor signalling. Nat Cell Biol 11:1473–1480
27. Luethy JD, Fargnoli J, Park JS, Fornace AJ Jr, Holbrook NJ
(1990) Isolation and characterization of the hamster gadd153
gene. Activation of promoter activity by agents that damage
DNA. J Biol Chem 265:16521–16526
28. Ohoka N, Yoshii S, Hattori T, Onozaki K, Hayashi H (2005)
TRB3, a novel ER stress-inducible gene, is induced via ATF4-
CHOP pathway and is involved in cell death. EMBO J
24:1243–1255
29. Mungrue IN, Pagnon J, Kohannim O, Gargalovic PS, Lusis AJ
(2009) CHAC1/MGC4504 is a novel proapoptotic component of
the unfolded protein response, downstream of the ATF4-ATF3-
CHOP cascade. J Immunol 182:466–476
30. Guyton KZ, Xu Q, Holbrook NJ (1996) Induction of the mam-
malian stress response gene GADD153 by oxidative stress: role
of AP-1 element. Biochem J 314:547–554
31. Oh-hashi K, Maehara K, Isobe K (2004) Hydrogen peroxide
induces GADD153 in Jurkat cells through the protein kinase
C-dependent pathway. Redox Rep 9:173–178
Mol Cell Biochem
123

32. Ma Y, Brewer JW, Diehl JA, Hendershot LM (2002) Two distinct
stress signaling pathways converge upon the CHOP promoter
during the mammalian unfolded protein response. J Mol Biol
318:1351–1365
33. Bruhat A, Jousse C, Carraro V, Reimold AM, Ferrara M, Fa-
fournoux P (2000) Amino acids control mammalian gene tran-
scription: activating transcription factor 2 is essential for the
amino acid responsiveness of the CHOP promoter. Mol Cell Biol
20:7192–7204
34. Magne L, Blanc E, Legrand B, Lucas D, Barouki R, Rouach H,
Garlatti M (2011) ATF4 and the integrated stress response are
induced by ethanol and cytochrome P450 2E1 in human hepa-
tocytes. J Hepatol 54:729–737
35. Romanoski CE, Che N, Yin F, Mai N, Pouldar D, Civelek M, Pan
C, Lee S, Vakili L, Yang WP, Kayne P, Mungrue IN, Araujo JA,
Berliner JA, Lusis AJ (2011) Network for activation of human
endothelial cells by oxidized phospholipids: a critical role of
heme oxygenase 1. Circ Res 109:e27–e41
36. Goebel G, Berger R, Strasak AM, Egle D, Muller-Holzner E,
Schmidt S, Rainer J, Presul E, Parson W, Lang S, Jones A,
Widschwendter M, Fiegl H (2012) Elevated mRNA expression of
CHAC1 splicing variants is associated with poor outcome for
breast and ovarian cancer patients. Br J Cancer 106:189–198
37. Galluzzi L, De Santi M, Crinelli R, De Marco C, Zaffaroni N,
Duranti A, Brandi G, Magnani M (2012) Induction of endo-
plasmic reticulum stress response by the indole-3-carbinol cyclic
tetrameric derivative CTet in human breast cancer cell lines.
PLoS One 7:e43249
38. Siu F, Bain PJ, LeBlanc-Chaffin R, Chen H, Kilberg MS (2002)
ATF4 is a mediator of the nutrient-sensing response pathway that
activates the human asparagine synthetase gene. J Biol Chem
277:24120–24127
39. Hai T, Hartman MG (2001) The molecular biology and nomen-
clature of the activating transcription factor/cAMP responsive
element binding family of transcription factors: activating tran-
scription factor proteins and homeostasis. Gene 273:1–11
40. Johannessen M, Delghandi MP, Moens U (2004) What turns
CREB on? Cell Signal 16:1211–1227
41. Navon A, Ciechanover A (2009) The 26 S proteasome: from basic
mechanisms to drug targeting. J Biol Chem 284:33713–33718
42. Newman JR, Keating AE (2003) Comprehensive identification of
human bZIP interactions with coiled-coil arrays. Science
300:2097–2101
43. Su N, Kilberg MS (2008) C/EBP homology protein (CHOP)
interacts with activating transcription factor 4 (ATF4) and neg-
atively regulates the stress-dependent induction of the asparagine
synthetase gene. J Biol Chem 283:35106–35117
44. He CH, Gong P, Hu B, Stewart D, Choi ME, Choi AM, Alam J
(2001) Identification of activating transcription factor 4 (ATF4)
as an Nrf2-interacting protein. Implication for heme oxygenase-1
gene regulation. J Biol Chem 276:20858–20865
45. Kumar A, Tikoo S, Maity S, Sengupta S, Sengupta S, Kaur A, Kumar
A, Bachhawat AK (2012) Mammalian proapoptotic factor ChaC1
and its homologues function as c-glutamyl cyclotransferases acting
specifically on glutathione. EMBO Rep 13(12):1095–1101
46. McCullough KD, Martindale JL, Klotz LO, Aw TY, Holbrook NJ
(2001) Gadd153 sensitizes cells to endoplasmic reticulum stress
by down-regulating Bcl2 and perturbing the cellular redox state.
Mol Cell Biol 21:1249–1259
Mol Cell Biochem
123




















