
Cephalic salivary gland ultrastructure of worker and queen eusocial bees (Hymenoptera, Apidae)
Upload
independentCategory
view
1download
0
Regulation of Reproduction in the Primitively Eusocial WaspRopalidia marginata: on the Trail of the Queen Pheromone
Anindita Bhadra & Aniruddha Mitra & Sujata A. Deshpande &
Kannepalli Chandrasekhar & Dattatraya G. Naik & Abraham Hefetz &
Raghavendra Gadagkar
Received: 14 August 2009 /Revised: 9 February 2010 /Accepted: 25 February 2010 /Published online: 18 March 2010# Springer Science+Business Media, LLC 2010
Abstract Queens and workers are not morphologicallydifferentiated in the primitively eusocial wasp, Ropalidiamarginata. Upon removal of the queen, one of the workersbecomes extremely aggressive, but immediately drops heraggression if the queen is returned. If the queen is notreturned, this hyper-aggressive individual, the potentialqueen (PQ), will develop her ovaries, lose her hyper-aggression, and become the next colony queen. Because ofthe non-aggressive nature of the queen, and because the PQloses her aggression by the time she starts laying eggs, wehypothesized that regulation of worker reproduction in R.marginata is mediated by pheromones rather than byphysical aggression. Based on the immediate loss ofaggression by the PQ upon return of the queen, we
developed a bioassay to test whether the queen’s Dufour’sgland is, at least, one of the sources of the queenpheromone. Macerates of the queen’s Dufour’s gland, butnot that of the worker’s Dufour’s gland, mimic the queen inmaking the PQ decrease her aggression. We also correctlydistinguished queens and workers of R. marginata nests bya discriminant function analysis based on the chemicalcomposition of their respective Dufour’s glands.
Key Words Ropalidia marginata . Queen pheromone .
Potential queen . Dufour’s gland . Reproductive monopoly
Introduction
The differentiation of adult colony members into fertilequeens and functionally sterile workers is an importantfeature of insect societies of ants, bees, and wasps. In thesehighly eusocial insect species, which generally have largecolony sizes (>>100 individuals), queens and workers aremorphologically differentiated, with queens maintainingtheir reproductive monopoly with pheromones. On theother hand, in primitively eusocial species, colony sizes areusually small (<100 individuals), and queens and workersare morphologically indistinguishable, with queens be-lieved to suppress worker reproduction by physical aggres-sion (often referred to as dominance behavior) (Wilson1971). Ropalidia marginata, the subject of this study, isclassified as a primitively eusocial species, because of theabsence of morphological differentiation between queensand workers. However, the queen in R. marginata is astrikingly non-aggressive and non-interacting individualthat, nevertheless, maintains complete reproductive monop-oly. Since the queen does not use physical aggression tosuppress worker reproduction (Gadagkar 2001), we hy-
Electronic supplementary material The online version of this article(doi:10.1007/s10886-010-9770-x) contains supplementary material,which is available to authorized users.
A. Bhadra :A. Mitra : S. A. Deshpande :K. Chandrasekhar :R. Gadagkar (*)Centre for Ecological Sciences, Indian Institute of Science,Bangalore 560012, Indiae-mail: [email protected]
R. GadagkarEvolutionary and Organismal Biology Unit, Jawaharlal NehruCentre for Advanced Scientific Research,Jakkur, Bangalore, India
D. G. NaikAgharkar Research Institute,G. G. Agarkar Road,Pune, India
A. HefetzDepartment of Zoology, Tel Aviv University,Ramat Aviv,69978 Tel Aviv, Israel
J Chem Ecol (2010) 36:424–431DOI 10.1007/s10886-010-9770-x
291

pothesized that R. marginata queens may use pheromonesfor this purpose.
If the queen is lost or removed from a colony, one of theworkers becomes highly aggressive within a few minutes; ifthe queen is not returned, this individual develops ovariesand becomes the new queen of the colony within a fewdays. We have designated this individual as the potentialqueen (PQ; Premnath et al. 1996). However, if the queen isreturned to the colony within a day of removal, the PQdecreases her aggression and swiftly returns to being atypical worker (Premnath et al. 1996; Kardile and Gadagkar2002; Sumana and Gadagkar 2003; Deshpande et al. 2006).This suggests that the PQ perceives the absence of thequeen within a few minutes. When workers are separatedfrom their queen by a wire mesh screen, one of the workersestablishes itself as the PQ and will lay eggs if the wiremesh is not removed (Sumana et al. 2007/2008). How thendo workers perceive the presence or absence of their queen?One possibility is through physical interactions with thequeen. However, we have shown that the rates ofinteraction of the PQ with the queen (both direct andindirect, via interactions with other workers) are notfrequent enough to explain the rapidity with which thequeen’s absence is perceived (Bhadra et al. 2007). Anotherpossibility is that the queen applies her pheromone to thenest surface. This possibility is supported by the observa-tion that the queen, but not the workers, frequently rubs theventral side of her abdomen on the nest surface (Bhadra etal. 2007). Because the Dufour’s gland, which opens into thetip of the abdomen, is believed to be a source of the queensignal in honeybees (Katzav-Gozansky et al. 1997; Katzav-Gozansky et al. 2002; Dor et al. 2005), we examined thepossibility that the queen pheromone of R. marginata isproduced in the Dufour’s gland.
Here, we demonstrated that a crude macerate of thequeen’s Dufour’s gland mimics her presence in a bioassay,and that queens and workers can be classified correctly by adiscriminant function analysis that uses the hydrocarbonprofiles of their respective Dufour’s glands.
Methods and Material
We collected post-emergence nests of R. marginata fromvarious localities in Bangalore (13° 00’ N and 77° 32’ E),India, and transplanted them to the Vespiary at the Centrefor Ecological Sciences, Indian Institute of Science,Bangalore. The nests were maintained in closed cagesmade of wood and fine mesh, and provided with food,water, and building material, ad libitum. All adults wereuniquely color-coded with small spots of Testors® enamelpaints (Gadagkar 2001). The queen was identified by egg-laying behavior prior to beginning the experiment.
Bioassay
We developed a bioassay for the queen pheromone basedon the observation that the PQ immediately reduces heraggression when the queen is returned to the colony. Thus,we tested whether applying crude macerate of the Dufour’sgland of the queen but not of the workers mimics the returnof the queen by a similar reduction in the aggression of thePQ. The bioassay consisted of three observation sessionslasting 36 min. each. Each session consisted of six ob-servation periods of 5 min. each, interspersed with a 1 min.break. After observing the normal queen-right colony in thefirst session (Queen-right Session), the queen and arandomly chosen worker were removed, and the queen-less colony was observed in the second session (Queen-lessSession). Before beginning observations in the third session(Treatment Session), the contents of the Dufour’s gland ofthe removed queen or a worker (each crushed in 30 µl ofRinger’s solution) or 30 µl of Ringer’s solution, wereapplied to the nest with a micropipette. The choice oftreatment was decided by drawing random numbers andwas unknown to the observer. A total of 25 nests were usedin the experiment (8 queen extract; 9 worker extract, and8 Ringer’s solution). All statistical analyses were performedusing the software package STATISTICA 7.
We used Ringer’s solution because we observed thatorganic solvents, such as pentane or acetone, had anadverse effect when applied to the nest, with the waspsbecoming agitated or even dying after contact with thesesolvents. Later, we validated the use of Ringer’s solution bychemical analysis of crude macerates of glands crushed inRinger’s solution and found a similar chemical profile tothat obtained by extracting glands in pentane.
Chemical Analysis
Preparation of Sample Seven additional colonies were usedfor chemical analysis. The queens of each colony and a totalof 18 workers (2 to 5 per colony, depending on colony size)had their Dufour’s glands dissected in distilled water under astereomicroscope, after gently pulling out the sting. Thegland was placed in a vial (chilled with dry ice and acetone),containing 5 µl pentane, and crushed with a needle.
Gas Chromatographic-Mass Spectrometric Analysis Afteradding another 5 μl pentane to the vial containing thecrushed gland, 2 μl extract was injected into an Agilent6890 gas chromatograph coupled with an Agilent 5973mass selective detector. A fused-silica capillary column,coated with 100% dimethyl polysiloxane (Agilent HP-1,60 m×250 µm×0.25 µm), was used for the gaschromatography-mass spectrometry (GC-MS) analyses.The injector port and transfer line were set at 250°C, and
J Chem Ecol (2010) 36:424–431 425
292

helium was the carrier gas. The column oven wasprogrammed from 100–275°C at 7°C min−1, held for5 min, and then to 280°C at 5°C min−1 held for 33 min.Analyses were performed in split mode (split ratio 10:1).Straight-chain and methyl-branched hydrocarbons wereidentified from their characteristic mass spectral fragmen-tation patterns, produced by electron impact ionization at70 eV. The compounds identified were either straight-chainor branched hyrdrocarbons, the spectra of which are veryclear and can be interpreted unambiguously. For example,branching at carbon 11 (11-methylHC) gives a distinct peakfragment at m/z 168 with a complementary fragmentdepending on the chain length of the compound. Likewise,14-methylHC has a pronounced peak fragment at m/z 196,etc. For dimethyl compounds, we have two branchingpoints and therefore 4 complementary peak fragments, them/z of which are characteristic to the compound. Thespectra of most of these compounds have been previouslypublished with a list of the ions characteristic to thebranching points. A blank run with 2 µl pentane confirmedthat none of the detected compounds was present asimpurities in the solvent.
Statistical Analyses Both multivariate and univariate anal-yses were carried out. To reduce the number of peaksused in the multivariate analyses, only those present in atleast 70% of all individuals were considered. Beforeapplying any multivariate test, the areas under each peakwere transformed by √(X=+0.5) to eliminate zero values(Zar 1999). Before applying the transformation, weconfirmed that group variances were proportional to themeans. The transformed data were subjected to the furthertransformation:
Zp:j ¼ lnAp:j
g Aj
� �
" #
where, Ap.j is the area of the peak p for individual j, g(Aj) isthe geometric mean of all peaks considered for analysis inindividual j, and Zp.j is the transformed area of peak p forindividual j (Reyment 1989). These transformed areas thenwere subjected to principal components (PC) analysis,followed by stepwise discriminant function analysis. Allpeaks had communality <0.8 on PC1 and were consideredsubsequently for discriminant analysis. The significance ofWilk’s λ (for canonical discriminant function), and thepercentage of correct assignments (for classification dis-criminant functions) were used to evaluate the validity ofthe discriminant functions. For univariate analysis, the areasunder each peak were transformed into percentages of totalarea under all peaks for each individual, and Mann-Whitney U tests were carried out to determine whetherqueens and workers differed for each compound.
An index of chemical diversity was calculated for eachindividual, using the Shannon Weiner Index of speciesdiversity (Shannon 1948; Krebs 1989), by the followingformula:
H0 ¼ �
XS
i¼1
pi ln pi
!
where H’ is the index of chemical diversity, S is the totalnumber of peaks in the respective individual, pi is therelative abundance of each peak (i.e., the area under therespective peak divided by total area under all peaks in thatindividual). A Mann-Whitney U test was carried out to testwhether queens and workers differed with respect to indicesof chemical diversity.
Squared Euclidean distances were calculated to estimatethe chemical distances between all possible pairs ofindividuals, using the standardized percentages of areaunder each peak (percentages calculated out of total areaunder all peaks as mentioned earlier). Z scores for thematrices were used to standardize the percentages, and thedifferences between groups were analyzed by Mann-Whitney U tests. Statistical analyses used the softwarepackages StatistiXL, version 1.7 and Mystat 12.
Validation of Bioassay
Because we carried out the bioassay using Ringer’ssolution, and the chemicals we identified from the Dufour’sgland are insoluble in Ringer’s, we determined whether theRinger’s solution macerates used in the bioassays actuallycontained the Dufour’s gland compounds or not. Eachgland was crushed in 30 µl Ringer’s solution, as in thebioassay, and the contents transferred to a clean glass vial.The Ringer’s solution was evaporated from the vial in anoven at 30°C, the dry vial chilled, and 10 µl pentane addedto it. Two microliters of this extract were analyzed by GCusing an Agilent 6890 gas chromatograph equipped with aflame ionization detector and a fused silica capillary columncoated with 5% phenyl methyl siloxane (Agilent HP-5, 30 m ×320 μm × 0.25 μm). General GC conditions were the same asthose in the GC-MS analyses. The Ringer’s solution wasanalyzed similarly to rule out any organic compounds thatmight be present as impurities. Peaks were identified byanalyzing a sample of 21 glands in 20 µl pentane by both GC-MS and GC.
Further, to rule out the possible involvement of othercompounds in the Dufour’s gland-Ringer’s solution that areinsoluble in pentane, we analyzed glands crushed inacetone. GC conditions were identical to those used in thepentane extracts, and blank runs were also carried out toexclude solvent-based impurities.
426 J Chem Ecol (2010) 36:424–431
293

Results
The Bioassay
In all nests, a PQ was obvious after queen removal, due to asignificant increase in its aggression from the Queen-rightSession to the Queen-less Session (Wilcoxon matchedpairs signed-ranks test: P=0.012 for queen-gland andRinger’s-solution treatments, and P=0.007 for worker-gland treatment). There was a significant reduction in thePQ’s aggression from the Queen-less Session to theTreatment Session (Fig. 1) in nests treated with the queen’sDufour’s gland macerate (P=0.012). However, in neststreated with worker-gland macerate or Ringer’s solution,the PQ did not show any change in aggression from theQueen-less Session to the Treatment Session (P=0.593 forworker and P=0.401 for Ringer’s).
Chemical Analysis
Of 18 workers, two were eliminated from statisticalanalyses because they had poorly developed glands, andmost compounds in their glands were below the detectablelimit of the GC-MS analysis. All queens had welldeveloped glands. The Dufour’s gland contained a seriesof linear, monomethyl and dimethyl branched alkanes with21 to 33 carbon atoms in the main chains. We did not findany unsaturated compounds. Considering all individuals,we found about 30 different compounds (Fig. 2 and
Table 1). We did not find any compound or set ofcompounds present exclusively in queens or in workers.
We found that: 1) queens had a significantly greater per-centage peak area than workers for three compounds (peaks#16, #24, and #29; Mann-Whitney U test, U=96, P=0.003for peak #16; U=91, P=0.009 for peak #24; and U=96.5,P=0.002 for peak #29); 2) a significantly lower percentagepeak area than workers for one compound (peak #13; U=83,P=0.038; and 3) they were indistinguishable from workers inpercentage peak areas for the remaining compounds (P>0.05). When the queens and workers of each colony wereexamined separately, the differences based on these com-pounds were not consistent. There was no compound thatwas consistently greater or lower in any queen compared toall the workers tested from her colony. The differencesbetween queens and workers became significant only whenall queens were pooled together and compared with allworkers pooled together. However, when we added up thepercentage areas under peaks 16, 24, and 29, for whichqueens had higher percentage areas than workers, queens ofeach colony were greater than workers from their respectivecolonies, except for one colony, which was a small colonywith only three workers, out of which two were newlyeclosed. Queens and workers did not differ with respect tototal area under all peaks (U=59, P=0.871) or total numberof peaks present (U=64.5, P=0.579).
We differentiated all queens and workers by a stepwisediscriminant analysis (Wilk’s λ=0.214, P<0.001, classifica-tion analysis: 100% correct classification; Fig. 3). Peak
0
50
100
150
200
250
300
350
a
a
b
bb
b
a
b
c
QR QL T QR QL T QR QL T
Frq
/hr
of
Do
min
ance
Beh
avio
ur
of
the
PQ
Queen gland Worker gland Ringer’s solution
Fig. 1 Mean and standard deviation of the frequency per hour ofdominance behavior exhibited by the potential queen from Ropalidiamarginata nests in the three sessions of the bioassay (Queen-right,Queen-less and Treatment; N=8, 9, and 8, respectively) when the nestwas exposed to queen Dufour’s gland macerate, worker Dufour’s
gland macerate or Ringer’s solution (control). Comparisons are byWilcoxon matched pairs signed-ranks test among the three sessionswithin each treatment. Different letters denote significant differenceamong bars (P<0.05)
J Chem Ecol (2010) 36:424–431 427
294
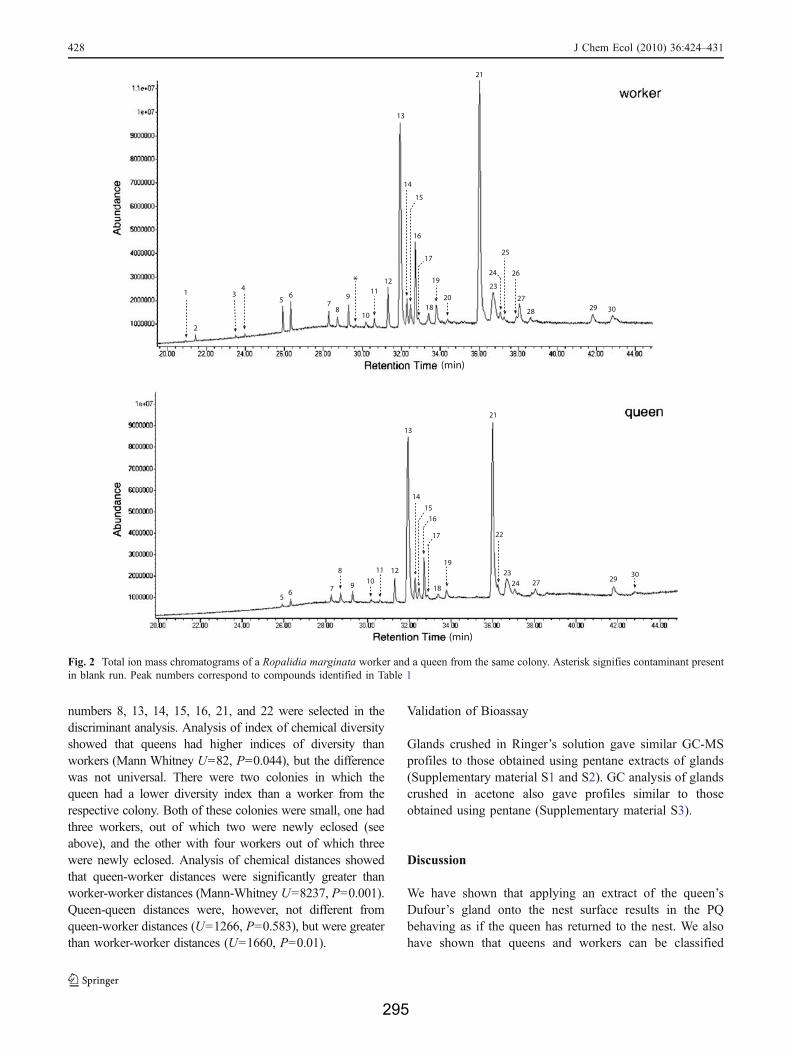
numbers 8, 13, 14, 15, 16, 21, and 22 were selected in thediscriminant analysis. Analysis of index of chemical diversityshowed that queens had higher indices of diversity thanworkers (Mann Whitney U=82, P=0.044), but the differencewas not universal. There were two colonies in which thequeen had a lower diversity index than a worker from therespective colony. Both of these colonies were small, one hadthree workers, out of which two were newly eclosed (seeabove), and the other with four workers out of which threewere newly eclosed. Analysis of chemical distances showedthat queen-worker distances were significantly greater thanworker-worker distances (Mann-Whitney U=8237, P=0.001).Queen-queen distances were, however, not different fromqueen-worker distances (U=1266, P=0.583), but were greaterthan worker-worker distances (U=1660, P=0.01).
Validation of Bioassay
Glands crushed in Ringer’s solution gave similar GC-MSprofiles to those obtained using pentane extracts of glands(Supplementary material S1 and S2). GC analysis of glandscrushed in acetone also gave profiles similar to thoseobtained using pentane (Supplementary material S3).
Discussion
We have shown that applying an extract of the queen’sDufour’s gland onto the nest surface results in the PQbehaving as if the queen has returned to the nest. We alsohave shown that queens and workers can be classified
1
2
34
56
78
9
*
10
1112
13
14
15
16
17
18
19
20
21
23
24
25
26
27
28 29 30
56 7
8
910
11 12
13
14
15
16
17
18
19
21
22
23
24 27 2930
(min)
(min)
Fig. 2 Total ion mass chromatograms of a Ropalidia marginata worker and a queen from the same colony. Asterisk signifies contaminant presentin blank run. Peak numbers correspond to compounds identified in Table 1
428 J Chem Ecol (2010) 36:424–431
295

correctly based on the contents of their Dufour’s glands.These results are consistent with the hypothesis that, in R.marginata, the workers (at least the PQ) perceive thepresence of their queen through contents of the Dufour’sgland that she applies to the nest surface. It should be notedthat our ability to classify queens and workers correctlyrequired that we consider the effect of several compoundssimultaneously with no single compound being adequatefor this purpose. We do not rule out the role of othercompounds or indeed of other mechanisms contributing toqueen recognition by colony members. We do not have directevidence that the compounds tested here are responsible forthe lack of ovarian development of the workers in thepresence of the queen. However, if workers use an honest
signal of their queen’s fertility to refrain from reproduction, assuggested by Keller and Nonacs (1993), the compounds weidentified also may be involved in reproductive regulation inR. marginata, thus permitting the queen to maintainreproductive monopoly in spite of her docility.
Analysis of chemical diversity showed that queens, ingeneral, had higher indices than workers, implying thatqueens have either greater numbers of compounds, or a moreeven distribution (i.e., less variable ratios, within individu-als) of compounds. Since we did not find any differencebetween queens and workers with respect to numbers ofcompounds present, it is likely that the distribution ofrelative proportions of compounds is more even in the caseof queens and less even in the case of workers. The two
Table 1 Peak numbers, retention times, identity and average percent areas and standard deviations of dufour’s gland compounds of queens andworkers in Ropalidia marginata
Peak # Retention Time (min) Identity of compound % area of each compound mean±standarddeviation
workers queens
1 20.83 Heneicosane 0.16±0.46 0.0008±0.002
2 21.431 11-Methylheneicosane 0.6±0.99 0.04±0.08
3 23.504 Tricosane 0.54±1.51 0.03±0.07
4 23.963 11-Methyltricosane 0.37±0.774 0.007±0.012
5 25.914 Pentacosane 1.85±3.06 1.12±1.96
6 26.323 Mixture of 11- and 13-methylpentacosane 3.99±3.58 3.28±4.40
7 28.267 Heptacosane 0.63±1.14 1.63±2.78
8 28.714 Mixture of 11- and 13-methylheptacosane 1.77±3.08 1.90±1.99
9 29.288 3-Methylheptacosane 2.19±2.754 1.062±2.069
10 30.187 Mixture of 12-, 14-, 16- and 18-methyloctacosane 0.341±0.59 0.25±0.33
11 30.614 4-Methyloctacosane 0.56±1.16 0.27±0.53
12 31.29 Nonacosane 1.63±3.10 6.74±10.40
13 31.915 Mixture of 11-, 13- and 15-methylnonacosane 41.87±30.06 16.95±14.97
14 32.279 5-Methylnonacosane 3.24±4.81 7.30±9.29
15 32.463 11-, 15-Dimethylnonacosane 7.814±9.753 4.54±4.09
16 32.712 3-Methylnonacosane 2.13±2.24 16.43±8.95
17 32.859 2-Methylnonacosane 0.25±0.91 0.10±0.20
18 33.395 8-Methyltriacontane 2.70±4.23 0.73±1.20
19 33.79 14-, 16-Dimethyltriacontane 1.92±2.26 1.02±1.95
20 34.377 2-Methyltriacontane 0.02±0.06 0.05±0.09
21 35.984 Mixture of 11-, 13- and 15-methylhentriacontane 12.03±17.53 14.98±17.40
22 36.2 Mixture of 7- and 9-methylhentriacontane 0.60±1.68 1.58±2.15
23 36.698 Mixture of 11-, 17- and 13-, 17-dimethylhentriacontane 11.10±11.09 9.99±8.40
24 37.029 3-Methylhentriacontane 0.02±0.07 1.39±2.37
25 37.253 Mixture of 5-, 21-, 5-,19- and 5-, 17-dimethyldotriacontane 0.02±0.07 1.11±2.04
26 37.896 14-Methyldotriacontane 0.17±0.32 0.66±0.76
27 38.056 8-Methyldotriacontane 1.84±3.52 0.38±0.7
28 38.643 14-, 18-Dimethyldotriacontane 0.04±0.12 0.63±1.45
29 41.8 Mixture of 13-, 15- and 17-methyltritriacontane 0.43±1.42 5.31±7.22
30 42.813 13-, 19-Dimethyltritriacontane 0.08±0.15 0.55±1.03
J Chem Ecol (2010) 36:424–431 429
296

colonies in which the queen did not have the highest indexvalue were small colonies with only one mature worker, withthe rest of the workers being newly eclosed. Chemicalprofiles of old, well established queens could be differentfrom those of newly established queens, as queens areknown to be behaviorally dominant during the nest initiationperiod or pre-emergence phase (Gadagkar 2001). Also, inthis species, the old queen sometimes gets overthrown witha worker becoming the PQ and ultimately taking over as thenew queen of the colony, a phenomenon known as queenturnover in natural colonies (Gadagkar 2001). It is possiblethat queens that are on the verge of being overthrown havea declining queen signal and are chemically different fromnormal queens. Since we do not have any information onthe history of each colony, and also did not measure theovarian development of the queens used in our GC-MSanalyses, we cannot make any conclusive statement as towhy these colonies were different from the others. Chemicaldistances show that queen-worker distances are higher thanworker-worker distances, indicating that queens and work-ers are more different from each other than workers arefrom other workers. However, queen-queen distances aredifferent from worker-worker, but not queen-worker dis-tances, implying that more variability in peak area (relativeamount) occurs among queens (between individuals).
Suppression of the PQ’s dominance is not complete afterapplying the queen’s Dufour’s gland extract to the nest,although the level of dominance is significantly higher thanthat observed in the queen-right condition. It should benoted that in earlier studies (Sumana and Gadagkar 2003)we saw that, after keeping the queen away from her colonyfor a day followed by queen reintroduction on the next day,the dominance of the PQ remained significantly higher thanin the normal queen-right condition, suggesting that the PQ
takes more than a day to lower her dominance to the normallevel. Also, in the present study, we observed that the effectof the queen’s Dufour’s gland macerate wanes with timeand the PQ again starts to increase her aggression. Overall,we conclude that the queen’s Dufour’s gland chemicals areat least one of the components of the queen signal and canact as a proxy for the queen herself.
Our results add to the growing evidence for chemicalcommunication between queens and workers in primitivelyeusocial wasps. For example, Downing (1991) demonstrat-ed that the contents of the Dufour’s gland in Polistesfuscatus act as an egg-marking pheromone. Sledge et al.(2001) showed that queens and workers in Polistesdominulus differ in their cuticular hydrocarbon signatures.Dapporto et al. (2007) suggested that the contents of theVan der Vecht’s gland may function as a queen pheromonein Polistes gallicus. Unlike Polistes queens, that are alsoaggressive in addition to producing a chemical cuesignaling their presence, R. marginata queens appear torely more heavily on pheromones for communication withworkers, as they are non-aggressive and interact directlywith other members of their colonies relatively infrequently.Our results are similar to the situation in honeybees inwhich queens are non-aggressive and where there is goodevidence for the contents of Dufour’s glands as signals offertility (Katzav-Gozansky et al. 2002).
Acknowledgements We thank the Department of Science andTechnology, the Department of Biotechnology, the Council forScientific and Industrial Research and the Ministry of Environmentand Forests, Government of India for financial assistance, and RobinCrewe for helpful comments and encouragement. We also thank AnjaliRajasekharan, Pooja Muralidharan and Gautam Pramanik, Centre forEcological Sciences, Indian Institute of Science, Bangalore, fortechnical assistance with the GC-MS analyses and, Prasanta Das,Department of Inorganic and Physical Chemistry, Indian Institute ofScience, Bangalore, for providing useful suggestions regardinginterpretation of mass spectra. AB carried out the behavioral observa-tions. SAD, KC, and AM did the dissections and prepared the extracts.AM carried out the chemical analysis, with help and guidance fromDGN and AH. The paper was co-written by AB, AM, and RG, and RGsupervised the overall work. All experiments reported here comply withthe current laws of the country in which they were performed.
References
BHADRA, A., IYER, P. L., SUMANA, A., DESHPANDE, S. A., GHOSH, S.,and GADAGKAR, R. 2007. How do workers of the primitivelyeusocial wasp Ropalidia marginata detect the presence of theirqueens? J. Theor. Biol. 246: 574–582.
DAPPORTO, L., SANTINI, A., DANI, F. R., and TURILLAZZI, S. 2007.Workers of a Polistes paper wasp detect the presence of their queenby chemical cues. Chem. Senses 32: 795–802. doi:10.1093/chemse/bjm047.
DESHPANDE, S. A., SUMANA, A., SURBECK, M., and GADAGKAR, R.2006. Wasp who would be queen: A comparative study of twoprimitively eusocial species. Curr. Sci. 91: 332–336.
-4
-3
-2
-1
0
1
2
3
4
5
Sco
res
on d
iscr
imin
ant f
unct
ion
Workers Queens
Fig. 3 Scores on discriminant function analysis for gas chromato-graphic analyses of Dufour’s glands of Ropalidia marginata queensand workers (nqueens=7, nworkers=16, Wilk’s λ=0.214, P<0.001,classification analysis: 100% correct classification)
430 J Chem Ecol (2010) 36:424–431
297

DOR, R., KATZAV-GOZANSKY, T., and HEFETZ, A. 2005. Dufour’sgland pheromone as a reliable fertility signal among honeybee(Apis mellifera) workers. Behav. Ecol. 49: 401–409.
DOWNING, H. A. 1991. A role of the Dufour's gland in the dominanceinteractions of the paper wasp, Polistes fuscatus (Hymenoptera:Vespidae). J. Insect Behav. 4: 557–565.
GADAGKAR, R. 2001. The Social Biology of Ropalidia marginata:Toward Understanding the Evolution of Eusociality. HarvardUniversity Press, Cambridge.
KARDILE, S. P., and GADAGKAR, R. 2002. Docile sitters and activefighters in paper wasps: a tale of two queens. Naturwissenschaften89: 302–304. doi:10.1007/s00114-002-0306-2.
KATZAV-GOZANSKY, T., SOROKER, V., and HEFETZ, A. 1997. Plasticityof caste specific Dufour’s gland secretion in the honey bee (Apismellifera L.). Naturwissenschaften 84: 238–241.
KATZAV-GOZANSKY, T., SOROKER, V., and HEFETZ, A. 2002. Evolu-tion of worker sterility in honey bees: egg-laying workers expressqueen-like secretion in Dufour’s gland. Behav. Ecol. Sociobiol.51: 588–589.
KELLER, L., and NONACS, P. 1993. The role of queen pheromones in socialinsects: queen control or queen signal? Anim. Behav. 45: 787–794.
KREBS, C. J. 1989. Ecological Methodology. Harper & Row, New York.
PREMNATH, S., SINHA, A., and GADAGKAR, R. 1996. Dominancerelationships in the establishment of reproductive division oflabour in a primitively eusocial wasp (Ropalidia marginata). Behav.Ecol. Sociobiol. 39: 125–132, doi:10.1007/s002650050274.
REYMENT, R. A. 1989. Compositional data analysis. Terra Nova. 1:29–34.
SHANNON, C. E. 1948. A mathematical theory of communication. TheBell Syst. Tech. J. 27: 379–423 & 623–656.
SLEDGE, M. F., BOSCARO, F., and TURILLAZZI, S. 2001. Cuticularhydrocarbons and reproductive status in the social wasp Polistesdominulus. Behav. Ecol. Sociobiol. 58: 270–276.
SUMANA, A., and GADAGKAR, R. 2003. Ropalidia marginata—aprimitively eusocial wasp society headed by behaviourally non-dominant queens. Curr. Sci. 84: 1464–1468.
SUMANA, A., DESHPANDE, S. A., BHADRA, A., and GADAGKAR, R.2007/2008. Workers of the primitively eusocial wasp Ropalidiamarginata do not perceive their queen across a wire meshpartition. J. Ethol. 26: 207–212. doi:10.1007/s10164-007-0049-9.
WILSON, E. O. 1971. The Insect Societies. The Belknap Press ofHarvard University Press, Cambridge.
ZAR, J. H. 1999. Biostatistical Analysis. Pearson Education Inc., TanPrints Private Ltd., India.
J Chem Ecol (2010) 36:424–431 431
298

Journal of Chemical Ecology© Springer Science+Business Media, LLC 201010.1007/s10886-010-9770-x
Below is the link to the electronic supplementary material.
Supplementary material S1 Flame ionization detection gas chromatogram of a Ropalidiamarginata Dufour’s gland crushed in Ringer’s solution, evaporated to dryness andresuspended in pentane. Asterisk signifies contaminant from Ringer’s solution. Identities ofpeak numbers are given in Table 1. Compare the chromatogram of this extraction method withthat obtained in S2 (PDF 34 kb)
Supplementary material S2 Comparison of gas chromatograms of the same sample of aRopalidia marginata Dufour’s gland extracted in pentane and analyzed by flame ionizationdetection (FID) and mass spectrometry (total ion chromatogram; TIC). Identities of peaknumbers are given in Table 1 (PDF 188 kb)
Supplementary material S3 Flame ionization detection gas chromatogram of a Ropalidiamarginata Dufour’s gland crushed and extracted in acetone. Identities of peak numbers aregiven in Table 1. Compare the chromatogram of this extraction method with that obtained in S2(PDF 19 kb)
Electronic Only Material: Regulation of Reproduction in the Primiti... http://www.springerlink.com/content/44u6714475108137/10886...
1 of 1 07-Apr-10 10:52 AM
299

13
12
78 9
10
11
56
*
14
15
16
17
1819
20
21
22
23
24
26
27
28 2930
Retention Time (min)
Supplementary Material
Click here to download Supplementary Material: 1952S1.ps
300

5
6
7
8
9
12
10
11
13
14
15
16
17
1819
20
21
22
23
24 26
27
28
29 30
25
432
1
Retention Time (min)
FID, GC analysis
Retention Time (min)
TIC, GC-MS analysis
1
324
5
67
8
9
10
11
12
13
14
16
15
17
18 19
20
21
22
23
24
25
27
2628
29 30
Supplementary Material
Click here to download Supplementary Material: 1952S2.ps
301

13
21
Retention Time (min)
89 10
6
4
2
1
11
15
14
16
17
18 19
20
23
24
26
27
28
2930
Acetone
Supplementary Material
Click here to download Supplementary Material: 1952S3.ps
302
Copyright © 2022 FDOKUMEN




















