
Aggregation Pheromone of the Cereal Leaf Beetle: Field Evaluation and Emission from Males in the...
11
JOllmal oj Chemical Ecology, Vol. 29, No.9, September 2003 (© 2003) AGGREGATION PHEROMONE OF THE CEREAL LEAF BEETLE: FIELD EVALUATION AND EMISSION FROM MALES IN THE LABORATORY SUJAYA ALLARD A. COSSE,2 BRUCE W. ZILKOWSKI,2 and ROBERT J. BARTELT 2 1Departmenr of Enromology Oregon State University Corvallis, Oregon 97331. USA 2 USDA-ARS. National Cenrer for Agricultural Utilization Research Peoria. l/linois 61604 USA (Received February 18,2003; accepted May 9, 2003) Abstract-The previously identified, male-specific compound of the cereal leaf beetle (CLB, Chrysomelidae; Oulema melanopus), (E)-8-hydroxy-6-methyl- 6-octen-3-one, was studied further with respect to field activity and emission rate from male beetles. In a 5-week field experiment in Oregon, the compound was shown to function as an aggregation pheromone in attracting male and female CLBs migrating from overwintering sites in spring. Traps baited \vith the synthetic compound (500 f-Lg per rubber septum) caught 3.3 times more CLBs than control traps. Lower doses of the pheromone (50 and 150 f-Lg) were less attractive than the 500 f-Lg dose. One relatively abundant. volatile com- pound from the host plant (oats), (Z)-3-hexenyl acetate, that elicited responses from beetle antennae was not attractive, either by itself or as a synergist of the pheromone. Both sexes were captured about equally for all treatments. We also measured daily pheromone emission by male beetles in the laboratory. Indi- vidual males feeding on oat seedlings under greenhouse conditions emitted as much as 6 f-Lg per day, which is about 500 times higher than had been previ- ously observed under incubator conditions. The pheromone emission rate was at least five times higher during the day than at night. and in one male. emission spanned a period of 28 d. The release rate of synthetic pheromone from the 500 f-Lg septa was very similar to the maximum from single males; thus, future experiments should evaluate even higher doses. The field results indicate that the pheromone has potential as a monitoring tool for early detection of CLBs * To whom correspondence should be addressed. E-mail; [email protected] 2165 0098-0331/03/0900-2165/0 2003 Plenum Publishing Corporation
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Aggregation Pheromone of the Cereal Leaf Beetle: Field Evaluation and Emission from Males in the...

JOllmal ojChemical Ecology, Vol. 29, No.9, September 2003 (© 2003)
AGGREGATION PHEROMONE OF THE CEREAL LEAFBEETLE: FIELD EVALUATION AND EMISSION FROM
MALES IN THE LABORATORY
SUJAYA RAO,I.~ ALLARD A. COSSE,2 BRUCE W. ZILKOWSKI,2and ROBERT J. BARTELT2
1Departmenr ofEnromologyOregon State University
Corvallis, Oregon 97331. USA
2USDA-ARS. National Cenrer for Agricultural Utilization ResearchPeoria. l/linois 61604 USA
(Received February 18,2003; accepted May 9, 2003)
Abstract-The previously identified, male-specific compound of the cereal leafbeetle (CLB, Chrysomelidae; Oulema melanopus), (E)-8-hydroxy-6-methyl6-octen-3-one, was studied further with respect to field activity and emissionrate from male beetles. In a 5-week field experiment in Oregon, the compoundwas shown to function as an aggregation pheromone in attracting male andfemale CLBs migrating from overwintering sites in spring. Traps baited \viththe synthetic compound (500 f-Lg per rubber septum) caught 3.3 times moreCLBs than control traps. Lower doses of the pheromone (50 and 150 f-Lg) wereless attractive than the 500 f-Lg dose. One relatively abundant. volatile compound from the host plant (oats), (Z)-3-hexenyl acetate, that elicited responsesfrom beetle antennae was not attractive, either by itself or as a synergist of thepheromone. Both sexes were captured about equally for all treatments. We alsomeasured daily pheromone emission by male beetles in the laboratory. Individual males feeding on oat seedlings under greenhouse conditions emitted asmuch as 6 f-Lg per day, which is about 500 times higher than had been previously observed under incubator conditions. The pheromone emission rate wasat least five times higher during the day than at night. and in one male. emissionspanned a period of 28 d. The release rate of synthetic pheromone from the500 f-Lg septa was very similar to the maximum from single males; thus, futureexperiments should evaluate even higher doses. The field results indicate thatthe pheromone has potential as a monitoring tool for early detection of CLBs
* To whom correspondence should be addressed. E-mail; [email protected]
2165
0098-0331/03/0900-2165/0 (~,' 2003 Plenum Publishing Corporation

2166 RAO, COSSE, ZILKOWSKl, AND BARTELT
as they move from their overwintering sites into newly planted cereal crops inspring.
Key Words-Cerealleaf beetle. Oulema melanopus, Coleoptera. Chrysomelidae. aggregation pheromone. (£)-8-hydroxy-6-methyl-6-octen-3-one, field evaluation, emission rates. (Z)-3-hexenyl acetate.
INTRODUCTION
The cereal leaf beetle (CLB), Olilema melanoplls (L.) (Coleoptera: Chrysomelidae), is a new pest in cereals in the western U.S. (Rao et aI., 2002). The beetle hasbeen in eastern and midwestern regions since 1962 (Castro et aI., 1965). It wasfirst detected in the west coast states of Oregon and Washington in 1999, and itcontinues to spread. There are currently quarantine restrictions on movement ofhay and forage from infested counties in Oregon and Washington into neighboringCalifornia and Canada. Annual surveys are now required in all counties in Oregonand Washington, and these presently involve labor intensive sweep-net samplingand visual examination of plants. An effective attractant for CLBs would facilitatethe early detection of the pest in new areas during these surveys.
The CLB has one generation per year and overwinters in the adult stage.In spring, adults migrate from overwintering sites into cereal grain fields withsucculent new foliage. The adults feed, mate, and lay eggs for a period of 6 weeks.Larvae also feed on the foliage. Mature larvae drop to the ground, pupate, andemerge as adults 2-3 weeks later. These emerging adults, present in the field inlate summer, feed for 3 weeks prior to migration to overwintering sites (Royceand Simko, 2000). Adults are reproductively active only during the spring. Thoseemerging from pupation in summer feed to build up fat reserves for overwintering,but they do not mate. Hence, traps baited with an attractant would be especiallybeneficial for monitoring or managing CLBs during spring, as they move into thenewly emerging cereals.
In an earlier study, a putative male-derived aggregation pheromone (£)-8hydroxy-6-methyl-6-octen-3-one, was identified from CLB (Cosse et aI., 2002).The compound stimulated antennae (GC-EAD) of both males and females. Thetotal amount of male-specific compound emitted per male per day was observedto be 6.3 ± 7.9 ng when adults were placed in incubators and exposed to artificiallight. In addition, male and female antennae responded to four host-derived compounds present in volatiles collected from feeding beetles, namely benzaldehyde,nonanal, methyl salicylate, and (Z)-3-hexenyl acetate (HA) (Cosse et aI., 2002).Of these, HA was the most abundant in the volatile collections.
Certain chrysomelid beetles, such as rootworms (Metcalf and Metcalf, 1992)and flea beetles (Pivnick et aI., 1992) are known to be strongly attracted to specific host compounds, but such information is not known for O. melanopus. In

EVALUATION OF CEREAL LEAF BEETLE PHEROMONE 2167
beetle groups with aggregation pheromones, pheromone emission occurs at thehostlbreeding site, and some host-derived volatiles are potent synergists of thepheromone. Examples include yeast-fermentation volatiles for the Carpophilussap beetles (family Nitidulidae; Bartelt, 1999) and conifer terpenes for Dendroctonus bark beetles (family Scolytidae; Borden, 1985). Synergistic interactions ofhost compounds and pheromones have not been published for chrysomelid beetlesbut could be crucial to the development of effective attractants for monitoringcereal leaf beetle.
The present study was conducted to establish whether (E)-8-hydroxy-6methyl-6-octen-3-one functions as an aggregation pheromone under field conditions. In addition, we tested whether HA was attractive to CLB or was synergisticwith the hydroxy ketone pheromone. Finally, additional information was obtainedin the laboratory on the rate and timing ofpheromone emission from male CLBs under greenhouse conditions for optimizing release rates of the synthetic pheromone.
METHODS AND MATERIALS
Chemicals, Instrumentation, and General Procedure for Collection ofVolatiles. (E)-8-Hydroxy-6-methyl-6-octen-3-one used in the field trial was synthesized as described previously (Cosse et aI., 2002). The compound was 95% pure(GC, impurities were not identified) after chromatographic purification on silicagel. The oat volatile HA was purchased from Lancaster Synthesis (Windham, NewHampshire).
Gas chromatography analyses were conducted on a Hewlett-Packard 5890 instrument equipped with flame-ionization detection. Samples were injected in splitless mode using 30 m Econocap (EC-l and EC-5) capillary columns (0.25 mm ID,0.25 fLm film thickness, Alltech Associates, Deerfield, IL). The oven temperaturewas programmed from 50 to 27SOC at 10°C per min. GC-MS was performed usinga Hewlett-Packard 5973 instrument (EL 70 eV) interfaced to an HP 6890 GC.
For volatile collections, the source material was contained in a glass tube(5 cm diam x 20 cm long) equipped with glass inlet and outlet adapters; all partshad 24/40 joints. A volatile trap was fitted to each adapter. Traps were made from5-mm diam glass tubing and contained a 5-8 mm plug of Super-Q porous polymer(80-100 mesh, Alltech). The Super-Q was held in place by fine stainless steelscreen (which was fused into the trap wall) and glass wool. A gentle vacuum wasdrawn on one of the traps so that air flowed through the assembled apparatus.Emitted volatiles were collected in the downstream trap, and the other trap servedto clean the incoming air. All collection glassware was silanized to reduce sampleadsorption. Volatiles were rinsed from the traps into vials with 300 fLl hexane(HPLC grade). Solvent desorbtion efficiency was checked by a second 300 fLlhexane rinse. Trap breakthrough was checked by placing a second Super-Q trap

2168 Rtto, COSSE, ZILKOWSKI, AND BARTELT
in series with the first The additional hexane rinse only showed trace amounts ofpheromone, and no pheromone was detected in the breakthrough trap.
Lures for Fields Study. Synthetic (E)-8-hydroxy-6-methyl-6-octen-3-onewas appropriately diluted with hexane and 10 f.LI aliquots were applied to rubbersepta (20 x 11 mm diam, red rubber, Aldrich Chemical Co., Milwaukee, WI) atrates of500, 150, and 50 f.Lg per septum followed by 300 f.LI of methylene chloride.Once the solvent had soaked into the septa, septa were aired in a fume hood for 1 hrand stored at - 70c C in tightly closed bottles until needed. Lures containing HAwere prepared in a similar manner. The synthetic ester (4 mg in 50 f.Ll hexane) wasapplied to a rubber septum, and the septum was then placed in a flat-top polypropylene microcentrifuge tube (1.5 rrJ, Bio Plas, San Francisco, CA). A single pin holewas made in the lid to slowly release the compound. Punctured tubes were storedat -70°C in tightly closed bottles until needed.
Prior to field tests, emission rates were measured from freshly prepared 500 f.L gsepta, and from 500 f.Lg septa that had been aired in a chemical hood for 1 week.The septa were placed in volatiles collectors at 2SOC with an air-flow rate of100 ml/min. Volatiles were collected for 24 hr. The amount of pheromone releasedwas quantified by GC using hexadecan-l-ol as internal standard. The releasedamounts of HA were not measured.
Field Study. The experiment was conducted at the Oregon State UniversityAgricultural Experiment Station at Union in a newly-seeded 2.4-hectare field ofoats, May 9 to June 13, 2002. Normal cereal production practices were adoptedat the experimental site, but no insecticides were applied to the crop. The studycommenced about 10 d before seedling oats emerged from the soil.
There were six experimental treatments. Four of these tested for individualand synergistic activity of the 500 f.Lg dose of the pheromone (P500) and HA; thesetreatments were P500, HA, P500 + HA, and unbaited control. It was consideredlikely, a priori, that a synergistic response to the combination would occur and,therefore, a sensitive way to test for a dose response to the pheromone would bein combination with the synergist Thus, the final two treatments were the 50 and150 f.Lg doses of synthetic pheromone in combination with HA (P50 +HA andP150 +HA, respectively). Resources did not allow testing all possible combinations.
The experiment was set up as a randomized block design with 10 replicates.In each block, traps were set up in a line, 15 m apart. P and HA lures were attachedwith separate clips on the downwind side of yellow sticky traps (Pherocon® AM,Trece Inc.) attached to bamboo stakes for trapping CLB adults. The trap heightwas initially 60 em and later was raised to 90 em above ground level.
Traps were checked, and lures and sticky traps were replaced weekly for 5weeks. Captured CLB adults were counted, removed from the sticky traps, andpreserved in 70% alcohol. Samples of adults collected from P500 traps on May18 (190 CLBs) and May 30 (144 CLBs) were sexed using characters described

EVALUATION OF CEREAL LEAF BEETLE PHEROMONE 2169
by Myser and Schultz (1967) to estimate the sex ratio of adults trapped in theexperiment.
Statistical Analyses. Counts of CLB adults on traps were transformed usinglog(X + 1) to stabilize variance and then submitted to a two-way analysis ofvariance, using Statistix for Windows software (Analytical Software, 1998). In thisanalysis, the blocking factor was the replicate-by-week interaction (a total of 50levels); thus, any week-to-week shifts in CLB population density in I or more ofthe 10 replicate areas would not inflate residual error and obscure treatment difference. Such shifts would occur, for example, if the field margins were infestedfirst, and the central areas became infested later. Treatment effects were assessedwith three linear contrasts involving the four means, P500 + HA, P500, HA, andcontrol: (1) effect ofP500, (2) effect ofHA, and (3) interaction ofP500 and HA. Inaddition, a linear contrast was fitted for P500 + HA, P150 + HA, and P50 +HA,using equal spacing of levels (which is approximately true for the logarithms ofthe doses). Finally, individual means were compared to the control, using the leastsignificant difference (LSD) test at a = 0.05.
Laboratory Study ofPheromone Emission from Male CLB. The CLB adultsused in this study were field collected during late summer and early fall of2001 from CLB-infested oat fields at Lake County, MT, Spokane Co., WY, andMultonomah Co., OR, and shipped overnight to Peoria, IL. Upon arrival, beetleswere placed in cages containing oat seedlings and moist cardboard rolls for shelter,and cooled from 15 to 4°C over a lO-d period for winter storage in total darkness(Ruthann Berry, USDA!APHISIPPQ, Niles, MI, unpublished 2000). Beetles werekept under these conditions for at least 4 months. After this period, beetles were removed from the cardboard rolls and sexed using characters described in Myser andSchultz (1967). Beetles were placed individually in 35-ml plastic cups with freshpieces of oat seedlings as food sources in the greenhouse (21-30°C, 14L:10D).Food was replaced every morning.
A previous study indicated a positive correlation between pheromone production and visible feeding damage (Cosse et aI., 2002). Therefore, only those malesthat showed feeding activity after spending several days in the cups were chosenfor the subsequent pheromone emission study.
Volatiles collections were first made from seven individual males under greenhouse conditions using glass collectors as described earlier. To accurately detect(>0.5 nglhr) and analyze the daily pheromone emission patterns, only those beetles that showed relatively high and consistent amounts of pheromone (relativeabundance estimated by GCfMS) during collections preceding the day and nightstudy were selected (3 out of 7 beetles). The day-night collection apparatus wasessentially the same as the standard collector, and each contained one male CLBand an oat seeding (10-15 cm tall) with the roots in a vial of water. The difference between the two systems was that the upper adaptor on the day-nightapparatus had two ports, and a trap and vacuum line were attached to each. A
2170 RAo, COSSE, ZILKOWSKl, AND BARTELT
switching system using a solenoid valve on an electric timer controlled whichof the two vacuum lines and traps was in use at any moment. Daily pheromoneemissions were collected for 30 d in two portions: a 16-hr collection that included the entire photophase and the 2 hr of the scotophase during which morningand evening twilight occurred, and a second 8-hr collection that included the remainder of the scotophase. The air-flow rate was 120 ml/min. Collectors weregenerally changed early each morning, and the oat seedlings were replaced every other day. Because of the laborious nature of the experiments and the limited amount of apparatus, volatile collections were limited to three individualbeetles.
RESULTS
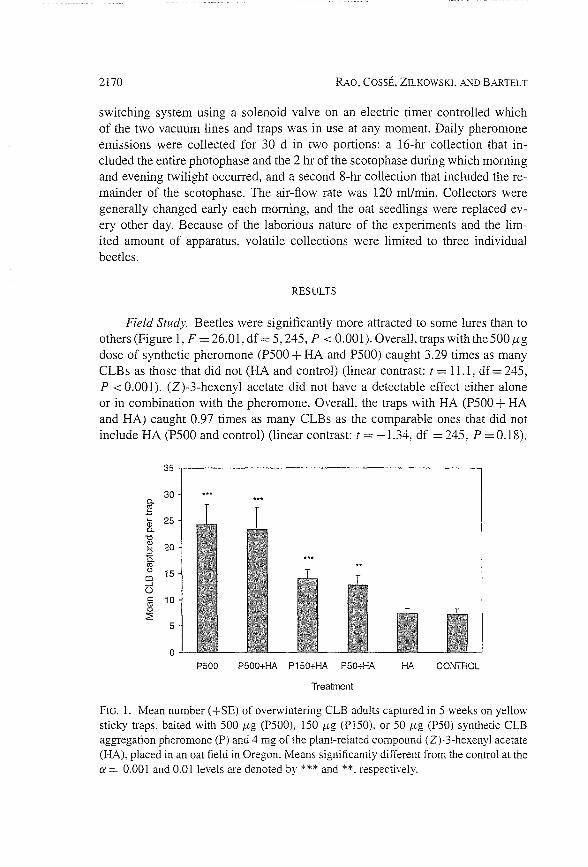
Field Study. Beetles were significantly more attracted to some lures than toothers (Figure I, F = 26.01, df = 5, 245, P < 0.001). Overall, traps with the 500 Jlg
dose of synthetic pheromone (P500 + HA and P500) caught 3.29 times as manyCLBs as those that did not (HA and control) (linear contrast: t = ILl, df = 245,P < 0.001). (Z)-3-hexenyl acetate did not have a detectable effect either aloneor in combination with the pheromone. Overall, the traps with HA (P500 + HAand HA) caught 0.97 times as many CLBs as the comparable ones that did notinclude HA (P500 and control) (linear contrast: t = -1.34, df =245, P =0.18).
35
30a.~a; 25a.'0~ 20::li5.C1l()
15co...J0c: 10C1l(lJ
:25
0
P500 P500+HA P150+HA P50+HA
Treatment
HA CONTROL
FIG. 1. Mean number (+SE) of overwintering CLB adults captured in 5 weeks on yellowsticky traps, baited with 500 J.Lg (P500), 150 J.Lg (P150), or 50 J.Lg (P50) synthetic CLBaggregation pheromone (P) and 4 mg of the plant-related compound (Z)-3-hexenyl acetate(HA), placed in an oat field in Oregon. Means significantly different from the control at thea = 0.001 and 0.01 levels are denoted by *,,* and **, respectively.

EVALUATION OF CEREAL LEAF BEETLE PHEROMONE 2171
70
-'" 60<ll<ll;;:Q; 500.."D 40~::J15.. 30C1lu
CD--J 20ut:C1l 10<D:2
02 3
Weeks
4 5
FIG. 2. Comparison of mean numbers (+5E) of overwintering CLB adults captured eachweek on yellow sticky traps baited with 500 f-Lg synthetic CLB aggregation pheromone(PSOO) and unbaited control traps placed in an oat field in Oregon.
Furthermore, the effect of P500 in the presence of HA was not different from theeffect of P500 in the absence of HA (i.e., there was no interaction; linear contrast: t = 1.01, df = 245, P = 0.32). There was a significant increase in trap catchwith increasing dose of pheromone (for the linear contrast including P50 +HA,P150 + HA, and P500 + HA, t =3.95, df =245, P < 0.001). Considering individual means, those for all treatments except HA were significantly higher than thecontrol (P < 0.003).
Numbers of beetles captured on a single trap in a week ranged from 0 to 140.Random samples from P500 traps showed that males and females were capturedabout equally; on May 18, the female:male ratio was 1.0 (N = 190), and on May30, it was 1.1 (N = 114). The numbers of adults trapped across all treatmentsincreased from an average of 12.5 beetles per trap in the first week to 27.5 pertrap in the third week. Subsequently there was a decline and the weekly averagedropped to 1.08 per trap in the fifth week. The pattern of change over weeks wassimilar for all treatments (Figure 2).
The release rate test conducted prior to the field experiment indicated that thesynthetic pheromone was emitted at 5.6 ± 0.9 f..(g/d (N = 3) from freshly preparedrubber septa (P500), and 3.6 ± 1.5 f..(g/d (N = 3) from septa that had been airedfor 1 week in a fumehood.
Pheromone Emission from Males. Traces of pheromone (GeIMS) were detected 3-10 d after removal of the beetles from cold storage (7 out of7 beetles). Thedaily mean pheromone release rates (± SD) from individual male beetles (prior togreenhouse equipment failure) was 3.4 (±1.7) f..(g/d (N = 27), and the maximumwas 6.0 f..(g/d. The periodicity of daily pheromone release by three individual

2172 RA.O, COSSE, ZILKOWSKI, AND BARTELT
male cereal leaf beetles is presented in Figure 3. For each male, pheromonerelease rate was at least five times higher during the photophase than duringthe scotophase. Pheromone secretion ceased completely when the greenhousetemperature reached 45°C because of equipment failure, but it started again whenthe normal temperature range (2 I-30°C) was reestablished.
DISCUSSION
In an earlier study, Cosse et al. (2002) demonstrated that the compound (E)
8-hydroxy-6-methyl-6-octen-3-one isolated from male CLB adults elicited strongresponses from antennae of both male and female CLBs in the laboratory. In thepresent study. we demonstrated that the compound attracted both sexes under fieldconditions, functioning as an aggregation pheromone.
Aggregation pheromones are well known in the Coleoptera (Borden, 1985),but they were not known in the Chrysomelidae until recently. The first chrysomelidpheromone to be identified was the female-produced sex pheromone of the westerncom rootworm (Guss et aI., 1982), which attracts only males. Chemically relatedpheromones were subsequently defined for seven other Diabrotica species andsubspecies (Krysan et aI., 1989, and references therein). It is now known, however,that males in other chrysomelid species produce long-range pheromones to whichboth sexes respond. Examples include the Colorado potato beetle, Leptinotarsadecemlineata (Say) (Dickens et aI., 2002), and the crucifer flea beetle, Phyllotretacruciferae (Goeze) (Peng et aI., 1999; Bartelt et aI., 2001). The present studyprovides further evidence for the latter pattern.
The release rate from freshly prepared rubber septa with the highest doseof synthetic pheromone (P500) used in the field study was similar to the maximum daily rate released by a single male under greenhouse conditions, and therate for I-week-old septa was similar to the average rate for single males under greenhouse conditions. Furthermore, if the daytime release from live males(Figure 3) is concentrated in just a few hours, then the peak natural emission ratefrom males may be higher than that from the rubber septa. It is possible that alllures in the field experiment emitted the pheromone at a lower rate than the naturalmaximum from males. Using incubators and artificial light, Cosse et al. (2002)showed a pheromone release rate of 6 ng/beetle/d. In the current study, the dailypheromone release rates from individual male beetles were about 500 times higherin the greenhouse than they were in the incubator with fluorescent lighting. Furthermore, pheromone collections in the previous study were made from groups of up tofive beetles. The grouping of beetles may have negatively affected the pheromonerelease rate. However, the original intent was to deploy lures that released in therange of 10-100 male equivalents, based on the initial emission data (Cosse et aI.,2002). The present information on emissions from males indicates that the initial

EVALUATION OF CEREAL LEAF BEETLE PHEROMONE 2173
*
, ,I_..... __ 8111~~~~~-o------------,
300
200
..... 100::J0::I: 0.....(])c.."C 100(])
13.9:? 300'0<.)(]) 200c:0E0 100.....(]).c:c..
0"-0(/)
E 100~0)0 300c:rnZ
200
100
0
100
0 5 10 15 20 25 30 35
Days after removal from overwintering storage
40
A
B
DayNight
c
DayNight
45
FIG. 3. Periodicity of pheromone emissions from three individual male Oulema melanopus adults (A. B, C) feeding on oat seedlings in a greenhouse environment. Gray barsrepresent 24-h collections. white bars represent daytime collections. and black bars represent nighttime collections. * Denotes the first collection. ** denotes the first day/nightcollection, €I denotes the final collection, and ,} denotes overheating of the green house(45'C).
target was too low and suggests that considerably higher doses of the compoundshould be tested in the field, to determine if stronger responses are possible.
The results of the present study differ from those of a preliminary study conducted in late summer 2001 (Rao et aI., 2002). During the earlier study, comparedwith control traps, there were apparently more beetles captured on traps baited

2174 RAo, COSSE, ZILKOWSKl, AN"]) BARTELT
with CLB aggregation pheromone, but the differences were not significant In theearlier study, the beetles were newly emerged, nonreproductive adults that werefeeding prior to entering overwintering diapause, In the present study, the captured adult CLBs had already overwintered and were moving into the oat field tobegin reproduction, It is likely that the effect of the pheromone on CLB behaviordepends on their physiological state (reproductive or diapause) with diapausingbeetles being less likely to respond, In this study, numbers of trapped CLB adultsincreased for the first 3 weeks and then declined. By the fifth week there were fewcaptures. This pattern likely reflects, first, the immigration of beetles into the field,and, eventually, the cessation of reproductive activity.
Many studies have documented that host volatiles enhance responses of insects to aggregation pheromones (Borden, 1985; Bartelt, 1999). Cosse et al. (2002)observed that a number of volatiles from damaged oat seedlings elicited strongantennal responses from male and female CLBs. However, the most abundant ofthese, HA, was neither an attractant nor a pheromone synergist Pheromone synergism by host volatiles may still occur in the CLB, using blends of host compoundsor adjusting the emission rate.
Finally, further attention must be given to trap design. In an earlier attempt todevelop a survey technique for CLB, sticky boards of various colors were tested(Wilson and Shade, 1967). Of the various colors tested, yellow attracted the greatestnumbers of adults. Whereas the yellow stick')' traps used in the present study wereeffective in trapping CLBs, our observations indicated that after capture, somebeetles were dislodged from the trap.
The present study suggests that the CLB aggregation pheromone may beuseful for monitoring and managingCLB, especially in the spring when newfields are infested by overwintered beetles. One particular benefit with the CLBpheromone is that it attracts both sexes.
Acknowledgments-We thank Bryon Quebbeman and Damn Walenta for their assistance withthe field study.
REFERENCES
ANALIlfCAL SOFTWARE. 1998. Statistix for Windows User's Manual. Analytical Software. Tallahassee. Florida.
BARTELT, R. J. 1999. Sap beetles. pp. 69-89, in J. Hardie and A. K. Minks (eds.). Pheromones of
Non-Lepidopteran Insects Associated with Agricultural Plants. CABI Publishing, Wallingford,
United Kingdom.
BARTELT, R. 1., COSSE, A. A., ZILKOWSKL B. w., WEISLEDER, D., and MOMANY', F. A. 2001. Male
specific sesguiterpenes from Plzyl/otreta and Aplztlzona flea beetles. J. Clzem. Ecol. 27:2397-2423.BORDEN, J. H. 1985. Aggregation pheromones, pp. 257-285, in G. A. Kerkut and L. 1. Gilbert (eds.).
Comprehensive Insect Physiology, Biochemistry, and Pharmacology. Pergamon, Oxford, UnitedKingdom.

EVALUATION OF CEREAL LEAF BEETLE PHEROMONE 2175
CASTRO, R. R., RUPPEL, R. E. and GOMULINSKI, M. L. 1965. Natural history of the cereal leaf beetlein Michigan. Mich. Agric. Exp. Sm. Q. Bull. 47:623-653.
COSSE, A. A., BARTELT, R. 1.. and ZILKOWSKI, B. W. 2002. Identification and electrophysiologicalactivity of a novel hydroxy ketone emitted by male cereal leaf beetle. 1. Nat. Prod. 65: 1156-1160.
DICKENS, J. c., OLIVER, J. E., HOLLISTER, B., DAVIS, J. c., and KLUN. J. A. 2002. Breaking a paradigm:Male-produced aggregation pheromone for the Colorado potato beetle. J. Exp. Bioi. 205:1925
1933.Guss. P. L.. TuMLINSON. J. H.. SONNET. P. E.. and PROVEAUX, A. T. 1982. Identification of a female
produced sex pheromone of the western corn rootworrn Diabrolica virgifera virgifera. J. Chem.
Ecol. 8:545-556.KRYSAN, J. L., MCDONALD, 1. c., and TUMLINSON, J. H. 1989. Phenogram based on allozymes and
its relationship to classical biosystematics and pheromone structure among eleven Diabroticites(Coleoptera: Chrysomelidae). Ann. Emomol. Soc. Am. 82:574--581.
METCALF, R. L. and METCALF, E. R. 1992. Plant Kairomones in Insect Ecology and Control. Chapman
and Hall. New York.MYSER, W. C. and SCHULTZ. W. B. 1967. Sexing the adult cereal leaf beetle, Oulema melanopus
(Coleoptera: Chrysomelidae). AnI/, Emomol. Soc. Am. 60:1329.PENO, C., BARTELT, R. 1.. and WEISS, M. 1999. Male crucifer flea beetles produce an aggregation
pheromone. J. Physiol. Emomol. 24:98-99.PIVNICK, K. A., LAMB, R. J., and REED, D. 1992. Response of flea beetles, Phyllolrera spp.. to mustard
oils and nitriles in field trapping experiments. 1. Chem. Ecol. 18:863-873.RAO, S., QUEBBEMAN, B., and WALENTA, D. 2002. Field evaluation of cereal leaf beetle pheromone
in Oregon, pp. 84--85, in W. C. Young (ed.). Seed Production Research. Oregon State University
Publication (ExtiCrs 121).ROYCE, L. A., and SIMKO, B. 2000. Cereal Leaf Beetle: Identification, Control, and California Quar
antine Act. EM 8762, Extension Service, Oregon State University, Oregon.
WILSON. M. and SHADE. R. E. 1967. Relative attractiveness of various luminescent colors to the cerealleaf beetle and the meadow spittle bug. 1. Econ. Emomol. 60:578-580.
Supplied by theU.S. Dept. of Agriculture
National Center for AgriculturalUtilization Research Peoria, at




















