
Phytoestrogens regulate transcription and translation of vitamin D receptor in colon cancer cells
Upload
independentCategory
view
1download
0
Toxicology Letters 230 (2014) 375–381
Actions of methyl-, propyl- and butylparaben on estrogen receptor-aand -b and the progesterone receptor in MCF-7 cancer cells and non-cancerous MCF-10A cells
Anna Maria Wróbel *, Ewa Łucja GregoraszczukDepartment of Physiology and Toxicology of Reproduction, Institute of Zoology, Jagiellonian University in Kraków, Gronostajowa 9, 30-387 Krakow, Poland
H I G H L I G H T S G R A P H I C A L A B S T R A C T
� Parabens increased ESR1 mRNA andprotein expression in both MCF-7 and MCF-10A cells.
� Parabens differentially affectedESR2 and PGR expression in MCF-7 and MCF-10A cells.
� Parabens by induction of hormonereceptors may contribute to cancerinitiation/progression.
A R T I C L E I N F O
Article history:Received 7 June 2014Received in revised form 8 August 2014Accepted 11 August 2014Available online 13 August 2014
Keywords:ParabensERaERbPGRGene expressionProtein expression
A B S T R A C T
Numerous studies have shown that widely used parabens possess estrogenic properties. In the presentstudy, we examined the effects of methyl-, propyl- and butylparaben on the mRNA and proteinexpression of estrogen receptor (ER)-a (ESR1) and -b (ESR2) and the progesterone receptor (PGR).Human MCF-7 breast cancer cells and MCF-10A non-transformed breast epithelial cells were exposed toparabens at a concentration of 20 nM; 17b-estradiol at a concentration of 10 nM, was used as a positivecontrol. Both propyl- and butylparaben stimulated PGR mRNA expression in MCF-7 cells, whereasmethyl- and propylparaben PGR protein expression. In MCF-10A cells, butyl- and propylparabenincreased only PGR mRNA expression. All parabens increased ESR1 gene and protein expression inMCF-7 and with the exception of butylparaben in MCF-10A cells. All parabens significantly increasedESR2 mRNA and protein expression in MCF-7 cells, but in MCF-10A cells only ESR2 protein expression. Insummary, by virtue of their stimulatory action on the expression of ESR1, ESR2 and PGR in cancer cells,parabens can be viewed as potential contributors to breast cancer progression. Extension, the actions ofthese parabens on the expression of ERs and PGR in non-cancerous cells point to possible actions onbreast cancer initiation.
ã 2014 Elsevier Ireland Ltd. All rights reserved.
Contents lists available at ScienceDirect
Toxicology Letters
journa l homepage: www.e lsev ier .com/ locate / toxlet
* Corresponding author at: Department of Physiology and Toxicology ofReproduction, Chair of Animal Physiology, Institute of Zoology, JagiellonianUniversity, Gronostajowa 9, 30-387 Krakow, Poland.Tel.: +48 12 664 5956; fax: +48 12 664 5101.
E-mail addresses: [email protected] (A.M. Wróbel),[email protected] (E.Ł. Gregoraszczuk).
http://dx.doi.org/10.1016/j.toxlet.2014.08.0120378-4274/ã 2014 Elsevier Ireland Ltd. All rights reserved.
1. Introduction
Parabens were long considered to be harmless and have thereforebeen widely used as preservatives in thousands of consumerproducts, including cosmetics, antiperspirants and lotions(Andersen 2008; Loretz et al., 2006; Yazar et al., 2011). Many studieshave shown that these compounds are present in tissues at

376 A.M. Wróbel, E.L. Gregoraszczuk / Toxicology Letters 230 (2014) 375–381
functionally relevant concentrations—concentrations equivalent tothose that stimulate proliferation of estrogen-responsive breastcancer cells in vitro (Charles and Darbre, 2013; Wróbel andGregoraszczuk, 2013). To date, parabens have been classified as“weak estrogens”, although numerous in vitro and in vivo studieshave shown that all of the widely used parabens possess estrogenicproperties (Byford et al., 2002; Miller et al., 2001; Darbre and Harvey,2008). Owing to these properties, parabens are categorized asendocrine disrupter compounds (EDCs).
We previously reported that parabens (methyl-, propyl- andbutylparaben) stimulate the proliferation of MCF-7 breast cancercells and non-transformed MCF-10A breast epithelial cells even atlow doses (Wróbel and Gregoraszczuk, 2013). In this same study,we also showed that these parabens significantly increased17b-estradiol secretion in MCF-7 cells but decreased it inMCF-10A cells, effects that correlated with expression of thecytochrome p450 family member, CYP19A1, at gene and proteinlevels in both cell lines. In a subsequent study, we showed that thestimulation of MCF-7 cell proliferation by parabens was not due toa direct effect on the expression of genes related to the cell cycle orapoptosis. However, in non-transformed MCF-10A cells, parabensaffected the expression of major cyclins involved in cell-cycleprogression and pro-survival genes associated with apoptosis in amanner similar to that of 17b-estradiol. These results point todifferent mechanisms of action of parabens and 17b-estradiol inMCF-7 and MCF-10A cells (Wróbel and Gregoraszczuk, 2014).
The development ofbreast cancer is initiatedbyimbalancesin sexhormones (Katchy et al., 2011). Estrogen, a primary sex hormoneresponsible for mammary gland development, exerts its effectthrough two nuclear receptors: estrogen receptor (ER)-a (ESR1) andERb (ESR2) (Lee et al., 2012, 2013). Progesterone, also a crucial sexhormone, acts through the progesterone receptor (PGR) to playa rolenot only in normal breast development but also in the carcinogenesisof breast epithelium and progression of breast cancer (Dickson andLippman, 1995). About 40% of breast cancers are ER-negative(Dickson and Lippman, 1995); these tumors are unresponsive toanti-estrogen therapy, probably because of constitutive growthfactor expression (Dickson and Lippman,1995; Ingvarsson,1999). Ingeneral, these tumors also have a higher histologic grade and greaterproliferation rate and are associated with poor prognosis (Dicksonand Lippman, 1995). Thus, both estrogens and progesterone havebeen implicated in the development of breast cancer, and ER/PGRstatus is a strong predictive factor.
Parabens have been found to similarly induce both ESR1 andESR2, with a possibly a stronger effect on ESR1 (Gomez et al., 2005;Okubo et al., 2001; Watanabe et al., 2013). On the basis of theseobservations and our previous data, in the present study, weexamined the effect of three parabens—methyl-, propyl- andbutylparaben—on the mRNA and protein expression of ESR1,ESR2 and PGR, the most common sex hormone receptors involvedin mammary gland development and carcinogenesis.
2. Materials and methods
2.1. Reagents
Dulbecco’s modified Eagle’s medium (DMEM; with and withoutphenol red), DMEM/F12 (DMEM and Ham’s F12 medium [1:1],with and without phenol red), fulvestrant (ICI 182,780),charcoal-dextran, insulin, hydrocortisone, epidermal growth factor(EGF) and Laemmli lysis buffer were obtained from SigmaChemical Co. (St. Louis, MO, USA). Fetal bovine serum (FBS; heatinactivated), horse serum (HS; heat inactivated), penicillin,streptomycin, and trypsin-EDTA were obtained from PAA Labora-tories GmbH (Colbe, Germany). 17b-estradiol was obtained fromSteraloids, Inc. (Newport, RI, USA). Phosphate-buffered saline (PBS)
was purchased from Biomed (Lublin, Poland). Methylparaben,n-propylparaben and n-butylparaben (Sigma Chemical Co.) weredissolved in absolute ethanol. The final concentration of ethanol inthe medium for each paraben was 0.2%. At this concentration,ethanol had no effect on cell proliferation or apoptosis (data notshown).
2.2. Cell culture and treatments
MCF-7 breast cancer cells (passage number 165; American TypeCulture Collection (ATCC), Manassas, VA, USA) were routinelycultured in DMEM supplemented with 10% heat-inactivated FBS,100 IU/ml penicillin, and 100 mg streptomycin. Twenty-four hoursbefore the experiments, the medium was removed and replacedwith DMEM without phenol red supplemented with 5%dextran-coated, charcoal-treated FBS (5% DC-FBS) to excludeestrogenic effects caused by the medium. The cells were thenplated in the same medium and allowed to attach overnight.
MCF-10A human breast epithelial cells (passage number 106;ATCC) were routinely cultured in DMEM/F12 supplemented with20 ng/ml EGF, 0.01 mg/ml insulin, 500 ng/ml hydrocortisone, 5%heat-inactivated HS, 100 IU/ml penicillin, and 100 mg streptomycin.Twenty-fourhoursbeforetheexperiments,themediumwasremovedandreplacedwithDMEM/F12withoutphenolredsupplementedwith0.01 mg/ml insulin, 500 ng/ml hydrocortisone, and 5% dextran-coated charcoal-treated HS (5% DC-HS). The cells were then platedin the same medium and allowed to attach overnight.
MCF-7 cells were seeded in 96-well plates at a density of15 �103 cells/well, whereas MCF-10A cells were seeded at adensity of 12 � 103 cells/well. For evaluation of gene expression,cells were exposed to 20 nM methylparaben, n-propylparaben orn-butylparaben, or 10 nM 17b-estradiol (positive control) for 6 and24 h. Cells were treated similarly for the evaluation of proteinexpression, but exposure duration was longer (48 and 72 h) andcompounds were tested alone and in combination with the ERreceptor blocker ICI 182,780 (100 nM). The cells were cultured in ahumidified atmosphere of 5% CO2 and at 37 �C.
2.3. Real-time polymerase chain reaction analysis
Total RNA isolation and cDNA synthesis were performed usingthe TaqMan Gene Expression Cell-to-CT Kit (Applied Biosystems,Carlsbad, CA, USA) according to the manufacturer’s protocol. Thelysis solution contains DNase I to remove genomic DNA during celllysis. The resulting pre-amplified cDNA preparations were ana-lyzed by real-time polymerase chain reaction (PCR) in aStepOnePlus Real-Time PCR System (Applied Biosystems) usingthe TaqMan Gene Expression Assay in combination with TaqManGene Expression Master Mix containing ROX (Applied Biosystems),according to the manufacturer’s instructions. The following PCRconditions were used: 2 min at 50 �C, then 10 min at 95 �C, followedby 40 cycles of 15 s at 95 �C (denaturation) and 60 s at 60 �C(annealing/elongation). Duplicate samples without cDNA for eachgene showed no DNA contamination. Expression of PGR(Hs01556702_m1), ESR1 (Hs00174860_m1) and ESR2(Hs1100353_m1), measured as cycle threshold (Ct), was normal-ized to that of glyceraldehyde-3-phosphate dehydrogenase(GAPDH; Hs99999905_m1) (DCt) to compensate for differencesin the amount of cDNA and then converted to relative expressionusing the 2�DDCt method (Livak and Schmittgen, 2001).
2.4. Western blot analysis
After incubating for 48 and 72 h, cells were washed with ice-coldPBS and lysed with Laemmli lysis buffer. The proteinconcentration incell lysates was determined using the Bradford reagent (Bio-Rad
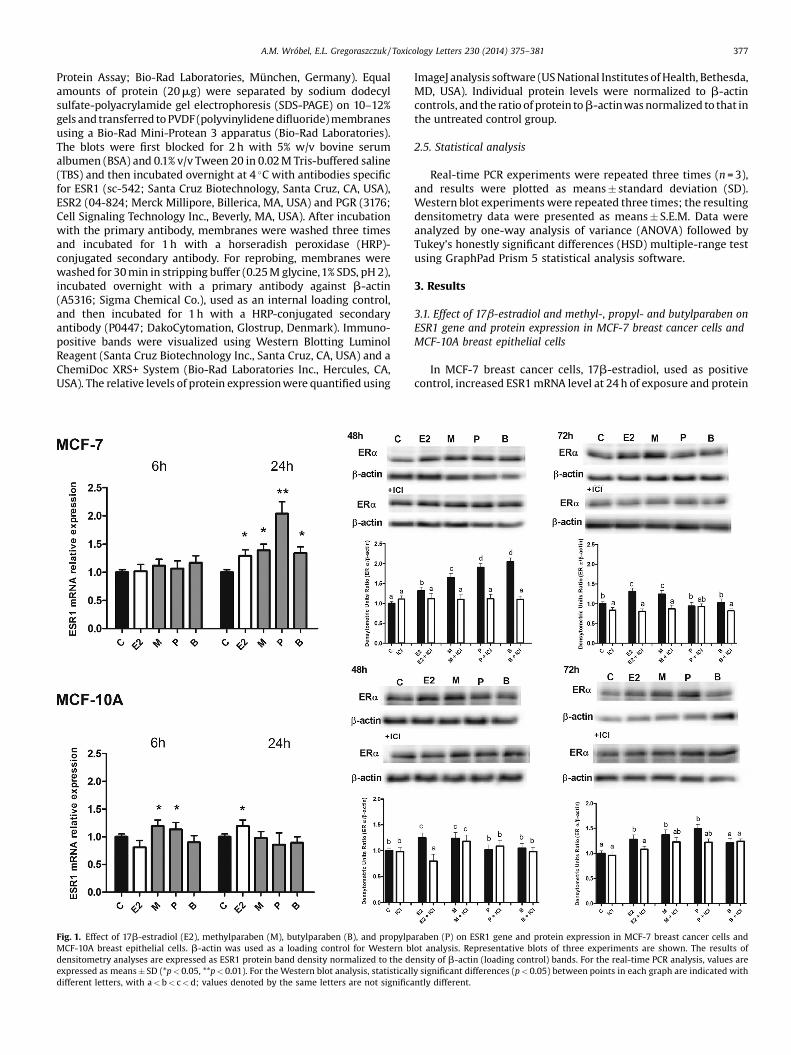
A.M. Wróbel, E.L. Gregoraszczuk / Toxicology Letters 230 (2014) 375–381 377
Protein Assay; Bio-Rad Laboratories, München, Germany). Equalamounts of protein (20mg) were separated by sodium dodecylsulfate-polyacrylamide gel electrophoresis (SDS-PAGE) on 10–12%gels and transferred to PVDF (polyvinylidene difluoride) membranesusing a Bio-Rad Mini-Protean 3 apparatus (Bio-Rad Laboratories).The blots were first blocked for 2 h with 5% w/v bovine serumalbumen (BSA) and 0.1% v/v Tween 20 in 0.02 M Tris-buffered saline(TBS) and then incubated overnight at 4 �C with antibodies specificfor ESR1 (sc-542; Santa Cruz Biotechnology, Santa Cruz, CA, USA),ESR2 (04-824; Merck Millipore, Billerica, MA, USA) and PGR (3176;Cell Signaling Technology Inc., Beverly, MA, USA). After incubationwith the primary antibody, membranes were washed three timesand incubated for 1 h with a horseradish peroxidase (HRP)-conjugated secondary antibody. For reprobing, membranes werewashed for 30 min in stripping buffer (0.25 M glycine,1% SDS, pH 2),incubated overnight with a primary antibody against b-actin(A5316; Sigma Chemical Co.), used as an internal loading control,and then incubated for 1 h with a HRP-conjugated secondaryantibody (P0447; DakoCytomation, Glostrup, Denmark). Immuno-positive bands were visualized using Western Blotting LuminolReagent (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA) and aChemiDoc XRS+ System (Bio-Rad Laboratories Inc., Hercules, CA,USA). The relative levels of protein expression were quantified using
Fig. 1. Effect of 17b-estradiol (E2), methylparaben (M), butylparaben (B), and propylpaMCF-10A breast epithelial cells. b-actin was used as a loading control for Western bdensitometry analyses are expressed as ESR1 protein band density normalized to the deexpressed as means � SD (*p < 0.05, **p < 0.01). For the Western blot analysis, statisticalldifferent letters, with a < b < c < d; values denoted by the same letters are not significa
ImageJ analysis software (US National Institutes of Health, Bethesda,MD, USA). Individual protein levels were normalized to b-actincontrols, and the ratio of protein to b-actin was normalized to that inthe untreated control group.
2.5. Statistical analysis
Real-time PCR experiments were repeated three times (n = 3),and results were plotted as means � standard deviation (SD).Western blot experiments were repeated three times; the resultingdensitometry data were presented as means � S.E.M. Data wereanalyzed by one-way analysis of variance (ANOVA) followed byTukey’s honestly significant differences (HSD) multiple-range testusing GraphPad Prism 5 statistical analysis software.
3. Results
3.1. Effect of 17b-estradiol and methyl-, propyl- and butylparaben onESR1 gene and protein expression in MCF-7 breast cancer cells andMCF-10A breast epithelial cells
In MCF-7 breast cancer cells, 17b-estradiol, used as positivecontrol, increased ESR1 mRNA level at 24 h of exposure and protein
raben (P) on ESR1 gene and protein expression in MCF-7 breast cancer cells andlot analysis. Representative blots of three experiments are shown. The results ofnsity of b-actin (loading control) bands. For the real-time PCR analysis, values arey significant differences (p < 0.05) between points in each graph are indicated withntly different.

378 A.M. Wróbel, E.L. Gregoraszczuk / Toxicology Letters 230 (2014) 375–381
expression at 48 and 72 h. All parabens tested significantlyincreased the level of ESR1 mRNA after 24 h and protein expressionafter 48 h, with propylparaben producing the greatest stimulatoryeffect. In addition, methylparaben, like 17b-estradiol, increasedESR1 protein expression at 72 h. Treatment of MCF-7 cells with theER receptor blocker, ICI 182,780 (100 nM), prevented the stimula-tory effects of 17b-estradiol and all parabens investigated onESR1 protein expression (Fig 1).
In MCF-10A breast epithelial cells, 17b-estradiol significantlyincreased ESR1 mRNA expression after 24 h and protein expressionafter 48 and 72 h. Methyl- and propylparaben increasedESR1 mRNA expression only at 6 h. ESR1 protein expression wasincreased after 48 h exposure to methylparaben and 72 h exposureto methyl- or propylparaben. Butylparaben had no effect onESR1 mRNA or protein expression in these cells. Interestingly,treatment with ICI 182,780 (100 nM) did not prevent the inductionof ESR1 protein expression by 17b-estradiol or any of theinvestigated parabens in non-cancerous MCF-10A cells (Fig 1).
3.2. Effect of 17b-estradiol and ethyl-, propyl- and butylparaben onESR2 gene and protein expression in MCF-7 breast cancer cells andMCF-10A breast epithelial cells
In MCF-7 cells,17b-estradiol and all tested parabens significantlyincreased ESR2 mRNA expression after 24 h of exposure and
Fig. 2. Effect of 17b-estradiol (E2), methylparaben (M), butylparaben (B), and propylpMCF-10A breast epithelial cells. b-actin was used as a loading control for Western bdensitometry analyses are expressed as ESR2 protein band density normalized to the deexpressed as means � SD (*p < 0.05, **p < 0.01, ***p < 0.001). For the Western blot analyindicated with different letters, with a < b < c; values denoted by the same letters are
ESR2proteinexpressionafter48 h.17b-estradiolandmethylparabenalso increasedESR2 proteinexpressionafter72 h,whereas butyl-andpropylparaben had no effect. As was the case for ESR1, no significantincrease in ESR2 protein was induced by parabens in cells treatedwith ICI 182,780 (100 nM) (Fig 2).
In MCF-10A cells, neither 17b-estradiol or any of the testedparabens altered ESR2 mRNA levels. However, methyl-, butyl- andpropylparaben all significantly increased ESR2 protein expressionafter 72 h, effects that were not blocked by treatment with ICI182,780 (Fig 2).
3.3. Effect of 17b-estradiol and methyl-, propyl- and butylparaben onPGR gene and protein expression in MCF-7 breast cancer cells andMCF-10A breast epithelial cells
In MCF-7 cells, 17b-estradiol increased PGR mRNA expressionafter 6 and 24 h of exposure and protein expression after 48 and72 h. Both butyl- and propylparaben stimulated PGR mRNAexpression at 24 h, whereas methylparaben did not. Parabens alsodifferentially affected PGR protein expression; both methyl- andpropylparaben increased PGR protein expression at 48 and 72 h inMCF-7 cells, whereas butylparaben did not (Fig 3).
In MCF-10A cells, propyl- and butylparaben increased PGRmRNA expression after 24 h, but none of the parabens investigatedaffected PGR protein expression at 48 or 72 h (Fig 3).
araben (P) on ESR2 gene and protein expression in MCF-7 breast cancer cells andlot analysis. Representative blots of three experiments are shown. The results ofnsity of b-actin (loading control) bands. For the real-time PCR analysis, values aresis, statistically significant differences (p < 0.05) between points in each graph arenot significantly different.

Fig. 3. Effect of 17b-estradiol (E2), methylparaben (M), butylparaben (B), and propylparaben (P) on PGR gene and protein expression in MCF-7 breast cancer cells andMCF-10A breast epithelial cells. b-actin was used as a loading control for Western blot analysis. Representative blots of three experiments are shown. The results ofdensitometry analyses are expressed as PGR protein band density normalized to the density of b-actin (loading control) bands. For the real-time PCR analysis, values areexpressed as means � SD (*p < 0.05, **p < 0.01). For the Western blot analysis, statistically significant differences (p < 0.05) between points in each graph are indicated withdifferent letters, with a < b; values denoted by the same letters are not significantly different.
A.M. Wróbel, E.L. Gregoraszczuk / Toxicology Letters 230 (2014) 375–381 379
4. Discussion
Here we show that all parabens tested significantly increasedESR1 at mRNA and protein levels in both MCF-7 breast cancer cellsand non-cancerous MCF-10A cells with the exception of butylpar-aben, which had no effect on ESR1 mRNA or protein expression inMCF-10A cells. Parabens differentially affected ESR2 mRNA andprotein expression in MCF-7 and MCF-10A cells. In MCF-7 cells, allparabens induced ESR2 mRNA and protein expression. In MCF-10Acells not a single parabens induced ESR2 mRNA expression but allsignificantly increased ESR2 protein. Interestingly, whereas block-ing ERs with ICI 182,780 prevented the stimulatory effects ofparabens on ESR1 and ESR2 protein expression in MCF-7 cells, ithad no such effect in MCF-10A cells.
Previous reports based on transient gene expression assaysdemonstrated that, unlike phytoestrogens such as coumestrol,genistein and daidzein, which have a higher affinity for ESR1 thanESR2, parabens have a similar binding affinity for ESR1 and ESR2(Kuiper et al., 1998). Okubo et al. (2001) noted that parabens canmimic the effects of the main natural estrogen, 17b-estradiol, bybinding to the ERs, stimulating ER-dependent cell growth andinfluencing the expression of estrogen-dependent genes.
On the basis of the results presented here, we suggest thatparabens have different actions on ESR1 and ESR2 proteins incancer and non-cancer cells, acting on both in cancer cells and
favoring ESR1 in non-cancerous cells. In the mammary gland,ESR1 and ESR2 play opposing roles in regulating growth anddifferentiation in response to estrogens; ESR1 promotes prolifera-tion, whereas ESR2 inhibits ESR1-mediated proliferation (Changet al., 2006; Helguero et al., 2005; Treeck et al., 2010).
The increase in ESR1 expression in response to parabensexposure is consistent with data showing that, in many cases ofbreast cancer, ESR1 expression contributes significantly to thehistory of the disease (Black et al., 1983; Thorpe et al., 1993;Hupperets et al., 1997). On the other hand, elevated levels ofESR1 in non-cancer cells following exposure to parabens suggeststhat one possible effect of parabens is an increased risk of breastcancer. A role for ESR1 in the initiation of breast cancer, as well asprogression in benign breast epithelium, have been proposed by Aliand Coombesm (2000). Furthermore estrogen-activatedESR1 regulates the expression of several key cell cycle regulatorygenes, such as c-myc (Dubik and Shiu, 1988), c-fos (van der Burget al., 1989), and cyclin D1 (Prall et al., 1997).
Several studies have reported that ESR2 is expressed at highlevels in normal breast tissue and is frequently lost duringmultistage carcinogenesis (Park et al., 2003; Roger et al., 2001;Shaaban et al., 2003; Skliris et al., 2003), suggesting a role forESR2 as a tumor suppressor (Skliris et al., 2003). In the currentreport, we show a lack of parabens action on ESR2 mRNAexpression but stimulatory on protein expression in

380 A.M. Wróbel, E.L. Gregoraszczuk / Toxicology Letters 230 (2014) 375–381
non-cancerous MCF-10A cells. These findings are not in accordwith data showing that ESR2 is involved in inhibiting human breastcancer cell proliferation by repressing the transcription of c-myc,cyclin D1 and cyclin A genes, and increasing the expression of thecyclin-dependent kinase inhibitors, p21Waf1/Cip1 and p27Kip1,effects that collectively lead to cell cycle arrest in the G2 phase(Chang et al., 2006; Paruthiyil et al., 2004; Frasor et al., 2006).However, supports data of Kuiper et al. (1998) showing parabenssimilar binding affinity for ESR1 and ESR2.
In the presence of ICI 182,780, a blocker of both ERs, thestimulatory effects of parabens on ESR1 and ESR2 proteinexpression were absent in MCF-7 cells but were retained inMCF-10A cells. This suggests that the actions of parabens in breastcancer cells are mediated by ERs, but another mechanism mediatesthe effects of parabens in non-cancer cells. In non-cancer cells, it islikely that the most important determinant of the action ofparabens is the ratio of ESR1 to ESR2 protein expression. Speirset al. (1999) reported that the ratio of ESR1 to ESR2 changes duringbreast cancer progression, with increased expression of ESR2 inrelapsed patients exhibiting tamoxifen resistance. Testing tumorsfor both ERs and PGRs is a standard part of breast cancer diagnosticregimens.
Our data also showed that propyl- and butylparaben increasedPGR mRNA expression in MCF-7 breast cancer cells andmethyl- and propylparaben increased PGR protein expression inthese cells. In non-cancer cells, propyl- and butylparaben increasedPGR mRNA level but had no effect on protein expression. Wesuggest that parabens that possess estrogenic properties mayinduce, in addition to PGR mRNA, another factor, such as aco-repressor (Horwitz et al., 1996), whose recruitment is requiredfor blocking transcriptional activation of target genes by theantagonist-occupied PGR.
Apart from ERs, PGR is the most widely studied marker in breasttissue. Of the few studies on PGR expression in premalignant andpreinvasive lesions, some have suggested that expression of thePGR also increases with increasing atypia (Allred et al., 2001).Experimental and clinical evidence suggests that progesteroneand its nuclear receptor, PGR, are critical in controlling mammarygland tumorigenesis by regulating the expression of key genesinvolved in breast cancer growth and activating signal transduc-tion pathways through a rapid or non-genomic mechanism(Elizalde and Proietti, 2012).
PGR protein expression is associated with a favorable prognosisin patients with primary breast cancer. In particular, the frequencyof metastases was found to be reduced among patients withPGR-positive tumors compared to patients with PGR-negativetumors. Moreover, the survival of patients whose tumors lose PGRexpression is significantly reduced compared with those whoretain it (William et al., 1986). Thus, PGRs appear to be animportant factor in the prognosis and management of breastcancer. By comparison, measurement of ER was found to be lesseffective in guiding the prognosis of early breast cancer (Pichonet al., 1980).
In summary, by virtue of their stimulatory action on theexpression of ESR1, ESR2 and PGR in MCF-7 cancer cells, parabenscan be viewed as potential contributors to breast cancerprogression. Extension, the actions of these parabens on theexpression of ERs and PGR in non-cancerous MCF-10A cells, pointto possible actions of these compounds on breast cancer initiation.The fact that the ER blocker, ICI 182,780, prevented the action ofparabens in MCF-7 cells but not in MCF-10A cells suggests adifferent mechanism of paraben action in these two cell lines.
Conflict of interest
The authors declare that there are no conflicts of interest.
Transparency document
The Transparency document associated with this article can befound in the online version.
Acknowledgments
This study was supported by the National Sciences Centre from2011 to 2014 as project DEC-2011/01/N/NZ7/00015. This paperincludes some of the data presented in Anna Wróbel’s PhDdissertation.
References
Ali, S., Coombesm, R.C., 2000. Estrogen receptor alpha in human breast cancer:occurrence and significance. J. Mammary Gland Biol. Neoplasia 5 (3), 271–281.
Allred, D.C., Mohsin, S.K., Fuqua, S.A., 2001. Histological and biological evolution ofhuman premalignant breast disease. Endocr. Relat. Cancer 8 (1), 47–61.
Andersen, F.A., 2008. Final amended report on the safety assessment ofmethylparaben, ethylparaben, propylparaben, isopropylparaben, butylparaben,isobutylpraben and benzylparaben as used in cosmetic products. Inte. J. Toxicol.27 (4), 1–82.
Black, R., Prescott, R., Bers, K., Hawkins, A., Stewart, H., Forrest, P., 1983. Tumourcellularity, oestrogen receptors and prognosis in breast cancer. Clin. Oncol.9311–9318.
Byford, J.R., Shaw, L.E., Drew, M.G.B., Pope, G.S., Sauer, M.J., Darbre, P.D., 2002.Oestrogenic activity of parabens in MCF7 human breast cancer cells. J. SteroidBiochem. Mol. Biol. 80, 49–60.
Chang, E.C., Frasor, J., Komm, B., Katzenellenbogen, B.S., 2006. Impact of estrogenreceptor beta on gene networks regulated by estrogen receptor alpha in breastcancer cells. Endocrinology 147, 4831–4842.
Charles, A.K., Darbre, P.D., 2013. Combinations of parabens at concentrationsmeasured in human breast tissue can increase proliferation of MCF-7 humanbreast cancer cells. J. Appl. Toxicol. 33, 390–398.
Darbre, P.D., Harvey, P.W., 2008. Paraben esters: review of recent studies ofendocrine toxicity, absorption, esterase and human exposure, and discussion ofpotential human health risks. J. Appl. Toxicol. 28, 561–578.
Dickson, R.B., Lippman, M.E., 1995. Growth factors in breast cancer. Endocr. Revi. 16,559–589.
Dubik, D., Shiu, R.P.C., 1988. Transcriptional regulation of c-myc oncogeneexpression by estrogen in hormone-responsive human breast cancer cells.J. Biol. Chem. 263, 12705–12708.
Elizalde, P.V., Proietti, J., 2012. The molecular basis of progesterone receptor actionin breast carcinogenesis. Horm. Mol. Biol. Clin. Invest. 9 (2), 105–117.
Frasor, J., Chang, E.C., Komm, B., Lin, C.Y., Vega, VB, Liu, E.T., Miller, L.D., Smeds, J.,Bergh, J., Katzenellenbogen, B.S., 2006. Gene expression preferentially regulatedby tamoxifen in breast cancer cells and correlations with clinical outcome.Cancer Res. 66, 7334–7340.
Gomez, E., Pillon, A., Fenet, H., Rosain, D., Duchesne, M.J., Nicolas, J.C., 2005.Estrogenic activity of cosmetic components in reporter cell lines: parabens, UVscreens, and musks. J. Toxicol. Environ. Health A 68, 239–251.
Helguero, L.A., Faulds, M.H., Gustafsson, J.A., Haldosen, L.A., 2005. Estrogenreceptors alpha (ERa) and beta (ERb) differentially regulate proliferationand apoptosis of the normal murine mammary epithelial cell line HC11.Oncogene 24, 6605–6616.
Horwitz, K.B., Jackson, T.A., Bain, D.L., Richer, J.K., Takimoto, G.S., Tung, L.,1996. Nuclearreceptor coactivators and corepressors. Mol. Endocrinol. 10 (10), 1167–1177.
Hupperets,P.S., Volovics, L., Schouten, L.J., Jager, J.J., Schouten, H.C.,Hillen, H.F., Blijham,G.H.,1997. The prognostic significance of steroid receptor activity in tumor tissuesof patients with primary breast cancer. Am. J. Clin. Oncol. 20, 546–551.
Ingvarsson, S., 1999. Molecular genetics of breast cancer progression. Semin. CancerBiol. 9, 277–288.
Katchy, A., Edvardsson, K., Aydogdu, E., Williams, C., 2011. Estradiol-activatedestrogen receptor alpha does not regulate mature microRNAs in T47D breastcancer cells. J. Steroid Biochem. Mol. Biol. 128, 145–153.
Kuiper, G.G.J.M., Lemmen, J.G., Carlsson, B., Corton, J.C., Safe, S.H., Saag, P.T., Burg, B.,Gustafsson, J.Å., 1998. Interaction of estrogenic chemicals and phytoestrogenswith estrogen receptor b. Endocrinology 138, 4252–4263.
Lee, H.R., Jeung, E.B., Cho, M.H., Kim, T.H., Leung, P.C., Choi, K.C., 2013. Molecularmechanism(s) of endocrine-disrupting chemicals and their potent oestroge-nicity in diverse cells and tissues that express oestrogen receptors. J. Cell. Mol.Med. 17, 1–11.
Lee, H.R., Kim, T.H., Choi, K.C., 2012. Functions and physiological roles of two types ofestrogen receptors, ERa and ERb, identified by estrogen receptor knockoutmouse. Lab. Anim. Res. 28, 71–76.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2�DDCT Method. Methods 25, 402–408.
Loretz, L., Api, A.M., Barraj, L., Burdick, J., Davis de, A., Dressler, W., Gilberti, E., Jarrett,G., Mann, S., Laurie Pan, Y.H., Re, T., Renskers, K., Scrafford, C., Vater, S., 2006.Exposure data for personal care products: hairspray spray perfume, liquidfoundation, shampoo, body wash, and solid antiperspirant. Food Chem. Toxicol.44, 2008–2018.

A.M. Wróbel, E.L. Gregoraszczuk / Toxicology Letters 230 (2014) 375–381 381
Miller, D., Wheals, B.B., Beresford, N., Sumpter, J.P., 2001. Estrogenic activity ofphenolic additives determined by an in vitro yeast bioassay. Environ. HealthPerspect. 109, 133–138.
Okubo, T., Yokoyama, Y., Kano, K., Kano, I., 2001. ER-dependent estrogenic activity ofparabens assessed by proliferation of human breast cancer MCF-7 cells andexpression of ERa and PR. Food Chem. Toxicol. 39, 1225–1232.
Park, B.W., Kim, K.S., Heo, M.K., et al., 2003. Expression of estrogen receptor-beta innormal mammary and tumor tissues: is it protective in breast carcinogenesis?Breast Cancer Res. Treat. 80 (1), 79–85.
Paruthiyil, S., Parmar, H., Kerekatte, V., Cunha, G.R., Firestone, G.L., Leitman, D.C.,2004. Estrogen receptor beta inhibits human breast cancer cell proliferation andtumor formation by causing a G2 cell cycle arrest. Cancer Res. 64, 423–428.
Pichon, M.F., Pallud, C., Brunet, M., Milgrom, E., 1980. Relationship of presence ofprogesterone receptors to prognosis in early breast cancer. Cancer Res. 40 (9),3357–3360.
Prall, O.W.J., Sarcevic, B., Musgrove, E.A., Watts, C.K.W., Sutherland, R.L., 1997.Estrogen-induced activation of cdk4 and cdk2 during G1-S phase progression isaccompanied by increased cyclin D1 expression and decreased cyclin-dependent kinase inhibitor association with cyclin E-cdk2. J. Biol. Chem. 272,10882–10894.
Roger, P., Sahla, M.E., Makela, S., et al., 2001. Decreased expression of estrogenreceptor beta protein in proliferative preinvasive mammary tumors. Cancer Res.61 (6), 2537–2541.
Shaaban, A.M., O'Neill, P.A., Davies, M.P., et al., 2003. Declining estrogen receptor-beta expression defines malignant progression of human breast neoplasia. Am.J. Surg. Pathol. 27 (12), 1502–1512.
Skliris, G.P., Munot, K., Bell, S.M., et al., 2003. Reduced expression of oestrogenreceptor beta in invasive breast cancer and its re-expression using DNA methyltransferase inhibitors in a cell line model. J. Pathol. 201 (2), 213–220.
Speirs, V., Parkes, A.T., Kerin, M.J., et al.,1999. Coexpression of estrogen receptor a andb: poor prognostic factors in human breast cancer? Cancer Res. 59, 525–528.
Thorpe, S.M., Christensen, I.J., Rasmussen, B.B., Rose, C., 1993. Short recurrence-freesurvival associated with high oestrogen receptor levels in the natural history ofpostmenopausal, primary breast cancer. Eur. J. Cancer 29 A, 971–977.
Treeck, O., Lattrich, C., Springwald, A., Ortmann, O., 2010. Estrogen receptor betaexerts growth-inhibitory effects on human mammary epithelial cells. BreastCancer Res. Treat. 120, 557–565.
van der Burg, B., van Selm-Miltenburg, A.J.P., de Laat, S.W., van Zoelen, E.J.J., 1989.Direct effects of estrogen on c-fos and c-myc protooncogene expression andcellular proliferation in human breast cancer cells. Mol. Cell. Endocrinol. 64,223–228.
Watanabe, Y., Kojima, H., Takeuchi, S., Uramaru, N., Ohta, S., Kitamura, S., 2013.Comparative study on transcriptional activity of 17 parabens mediated byestrogen receptor a and b and androgen receptor. Food Chem. Toxicol. 57,227–234.
William, L., McGuire, M.D., Clark, G.M., 1986. Role of progesterone receptors inbreast cancer. Cancer J. Clin. 36 (5), 302–309.
Wróbel, A., Gregoraszczuk, E.Ł., 2014. Differential effect of methyl-, butyl- andpropylparaben and 17-b estradiol on selected cell cycle and apoptosis gene andprotein expression in MCF-7 breast cancer cells and MCF-10A non-malignantcells. J. Appl. Toxicol doi:http://dx.doi.org/10.1002/jat.2978.
Wróbel, A., Gregoraszczuk, E.Ł., 2013. The effects of single and repeated in vitroexposure of three forms of parabens, methyl-, butyl- and propylparabens on theproliferation and estradiol secretion in MCF-7 and MCF-10A cells. Pharmacol.Rep. 65, 484–493.
Yazar, K., Johnsson, S., Lind, M.L., Boman, A., Liden, C., 2011. Preservatives andfragrances in selected consumer-available cosmetics and detergents. ContactDermatitis 64, 265–272.
Copyright © 2022 FDOKUMEN




















