
Extra virgin olive oil potentiates the effects of aromatase inhibitors via glutathione depletion in...
8
Extra virgin olive oil potentiates the effects of aromatase inhibitors via glutathione depletion in estrogen receptor-positive human breast cancer (MCF-7) cells Amar Mohamed Ismail a,e,⇑ , Lionel L.A. In b,⇑ , Mohammad Tasyriq b , Devi Rosmy Syamsir c , Khalijah Awang c , Ayda Hussein Omer Mustafa d , Omer Fadul Idris e , Imad Fadl-Elmula a , Noor Hasima b,f a Department of Pathology, Faculty of Medicine and Health Sciences, Al-Neelain University, 12702 Khartoum, Sudan b Institute of Biological Science (Genetics & Molecular Biology), Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia c Department of Chemistry, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysia d Department of Surgery, Faculty of Medicine and Health Sciences, Al-Neelain University, 12702 Khartoum, Sudan e Department of Biochemistry, Faculty of Science and Technology, Al-Neelain University, 12702 Khartoum, Sudan f Centre for Research in Biotechnology for Agriculture (CEBAR), University of Malaya, 50603 Kuala Lumpur, Malaysia article info Article history: Received 25 January 2013 Accepted 16 October 2013 Available online 23 October 2013 Keywords: Olive oil Oleic acid Aromatase inhibitor Breast cancer Estrogen Apoptosis abstract There have been numerous evidences supporting the relationship between olive oil and cancer, with most of the attention being directed toward its fat and phenolic content. The aims of this study were to investigate whether EVOO and OA could enhance the effects of aromatase inhibitors (letrozole and anastrozole) in ER-positive MCF-7 cells, as well as to investigate its influence on cytochrome c release and GSH levels. It was observed that upon combination treatment, anti-proliferation effects and apopto- sis induction were augmented. Apoptosis was triggered via the intrinsic pathway in accordance with cytochrome c release into the cytosol based on IF–IC and ELISA observations. Intracellular GSH levels were also reduced upon EVOO/OA treatment in combination with aromatase inhibitors, and were found to be inversely correlated to cytosolic cytochrome c levels. Additionally, the estrogenic suppressive effects of letrozole and anastrozole were amplified when used in combination with EVOO/OA. Therefore, the employment of aromatase inhibitors in combination with EVOO/OA could orchestrate a reduction in intracellular estrone biosynthesis which feeds ER-positive cells, while simultaneously depleting GSH lev- els and increasing ROS generation, thus releasing cytochrome c and subsequent induction of apoptosis in MCF-7 cells. Ó 2013 Elsevier Ltd. All rights reserved. 1. Introduction Epidemiological studies have implicated that the incidence of breast cancers were found to be the lowest in Mediterranean women (La Vecchia and Bosetti, 2006). It has been suggested that this was largely due to high olive oil (OO) consumption, which con- tains a rich source of oleic omega-9 monounsaturated fatty acids (OA:x-9 MUFA) (Colomer and Menendez, 2006) ranging from 56% to 84% of total fatty acids (FA), and x-6 linoleic polyunsatu- rated fatty acids (LA PUFA;18:2n-6) ranging from 7% to 10% of total FA in the Mediterranean diet. Collectively, these fatty acids provide OO with unique characteristics such as resistance to acidification and oxidation (Perez-Jimenez et al., 2005). Beside FAs, the main phenolic component present in olive leaves, drupes and extra vir- gin olive oil (EVOO) is oleuropein which discriminates the charac- teristic differences between various types of OOs in terms of taste and stability (Andrewes et al., 2003). Previous studies have shown that olive tree (Olea eurapaea) products possess several pharmaco- logical properties including anti-oxidant, hypocholesterolemia (Bogani et al., 2007; El and Karakaya, 2009), anti-inflammatory (Bogani et al., 2007; Fabiani et al., 2008), anti-microbial (Omar, 2010), anti-viral (Lee-Huang et al., 2007) and anti-cancer proper- ties (Fu et al., 2010). In spite of these claims, the mechanism by which specific FA components in OO exert its potential protective 0278-6915/$ - see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.fct.2013.10.024 Abbreviations: Bcl-2, B-Cell Lymphoma 2; ER, estrogen receptor; E1, estrone; EVOO, extra virgin olive oil; FA, fatty acids; GSH, glutathione; MUFA, monounsat- urated fatty acid; NADPH, Nicotinamide Adenine Dinucleotide Phosphate; OA, oleic acid; OO, olive oil; PUFA, polyunsaturated fatty acid. ⇑ Corresponding authors. Address: Department of Pathology, Faculty of Medicine and Health Sciences, Al-Neelain University, 12702 Khartoum, Sudan. Tel.: +249 912144114; fax: +249 794422 (A.M. Ismail), tel.: +60 3 79675921; fax: +60 3 79675908 (L.L.A. In). E-mail addresses: [email protected] (A.M. Ismail), [email protected] (L.L.A. In). Food and Chemical Toxicology 62 (2013) 817–824 Contents lists available at ScienceDirect Food and Chemical Toxicology journal homepage: www.elsevier.com/locate/foodchemtox
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Extra virgin olive oil potentiates the effects of aromatase inhibitors via glutathione depletion in...

Food and Chemical Toxicology 62 (2013) 817–824
Contents lists available at ScienceDirect
Food and Chemical Toxicology
journal homepage: www.elsevier .com/locate / foodchemtox
Extra virgin olive oil potentiates the effects of aromatase inhibitors viaglutathione depletion in estrogen receptor-positive human breast cancer(MCF-7) cells
0278-6915/$ - see front matter � 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.fct.2013.10.024
Abbreviations: Bcl-2, B-Cell Lymphoma 2; ER, estrogen receptor; E1, estrone;EVOO, extra virgin olive oil; FA, fatty acids; GSH, glutathione; MUFA, monounsat-urated fatty acid; NADPH, Nicotinamide Adenine Dinucleotide Phosphate; OA, oleicacid; OO, olive oil; PUFA, polyunsaturated fatty acid.⇑ Corresponding authors. Address: Department of Pathology, Faculty of Medicine
and Health Sciences, Al-Neelain University, 12702 Khartoum, Sudan. Tel.: +249912144114; fax: +249 794422 (A.M. Ismail), tel.: +60 3 79675921; fax: +60 379675908 (L.L.A. In).
E-mail addresses: [email protected] (A.M. Ismail), [email protected](L.L.A. In).
Amar Mohamed Ismail a,e,⇑, Lionel L.A. In b,⇑, Mohammad Tasyriq b, Devi Rosmy Syamsir c,Khalijah Awang c, Ayda Hussein Omer Mustafa d, Omer Fadul Idris e, Imad Fadl-Elmula a, Noor Hasima b,f
a Department of Pathology, Faculty of Medicine and Health Sciences, Al-Neelain University, 12702 Khartoum, Sudanb Institute of Biological Science (Genetics & Molecular Biology), Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysiac Department of Chemistry, Faculty of Science, University of Malaya, 50603 Kuala Lumpur, Malaysiad Department of Surgery, Faculty of Medicine and Health Sciences, Al-Neelain University, 12702 Khartoum, Sudane Department of Biochemistry, Faculty of Science and Technology, Al-Neelain University, 12702 Khartoum, Sudanf Centre for Research in Biotechnology for Agriculture (CEBAR), University of Malaya, 50603 Kuala Lumpur, Malaysia
a r t i c l e i n f o
Article history:Received 25 January 2013Accepted 16 October 2013Available online 23 October 2013
Keywords:Olive oilOleic acidAromatase inhibitorBreast cancerEstrogenApoptosis
a b s t r a c t
There have been numerous evidences supporting the relationship between olive oil and cancer, withmost of the attention being directed toward its fat and phenolic content. The aims of this study wereto investigate whether EVOO and OA could enhance the effects of aromatase inhibitors (letrozole andanastrozole) in ER-positive MCF-7 cells, as well as to investigate its influence on cytochrome c releaseand GSH levels. It was observed that upon combination treatment, anti-proliferation effects and apopto-sis induction were augmented. Apoptosis was triggered via the intrinsic pathway in accordance withcytochrome c release into the cytosol based on IF–IC and ELISA observations. Intracellular GSH levelswere also reduced upon EVOO/OA treatment in combination with aromatase inhibitors, and were foundto be inversely correlated to cytosolic cytochrome c levels. Additionally, the estrogenic suppressiveeffects of letrozole and anastrozole were amplified when used in combination with EVOO/OA. Therefore,the employment of aromatase inhibitors in combination with EVOO/OA could orchestrate a reduction inintracellular estrone biosynthesis which feeds ER-positive cells, while simultaneously depleting GSH lev-els and increasing ROS generation, thus releasing cytochrome c and subsequent induction of apoptosis inMCF-7 cells.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Epidemiological studies have implicated that the incidence ofbreast cancers were found to be the lowest in Mediterraneanwomen (La Vecchia and Bosetti, 2006). It has been suggested thatthis was largely due to high olive oil (OO) consumption, which con-tains a rich source of oleic omega-9 monounsaturated fatty acids
(OA:x-9 MUFA) (Colomer and Menendez, 2006) ranging from56% to 84% of total fatty acids (FA), and x-6 linoleic polyunsatu-rated fatty acids (LA PUFA;18:2n-6) ranging from 7% to 10% of totalFA in the Mediterranean diet. Collectively, these fatty acids provideOO with unique characteristics such as resistance to acidificationand oxidation (Perez-Jimenez et al., 2005). Beside FAs, the mainphenolic component present in olive leaves, drupes and extra vir-gin olive oil (EVOO) is oleuropein which discriminates the charac-teristic differences between various types of OOs in terms of tasteand stability (Andrewes et al., 2003). Previous studies have shownthat olive tree (Olea eurapaea) products possess several pharmaco-logical properties including anti-oxidant, hypocholesterolemia(Bogani et al., 2007; El and Karakaya, 2009), anti-inflammatory(Bogani et al., 2007; Fabiani et al., 2008), anti-microbial (Omar,2010), anti-viral (Lee-Huang et al., 2007) and anti-cancer proper-ties (Fu et al., 2010). In spite of these claims, the mechanism bywhich specific FA components in OO exert its potential protective

818 A.M. Ismail et al. / Food and Chemical Toxicology 62 (2013) 817–824
and anti-cancer effects on estrogen receptor (ER) positive humanbreast cancers remain poorly understood.
Past studies have begun to address the specific biological andmolecular mechanisms by which FAs such as OA exert its effectson cancer (Simopoulos, 2004; Trichopoulou et al., 2000). It hasbeen reported that OA, which is the main MUFA of OO specificallysuppresses Her-2/neu overexpression, which in turn interacts syn-ergistically with immunotherapies involving Herceptin� in breastcancers expressing the Her-2/neu oncogene (Menendez et al.,2005). It was also reported that in non-malignant cells, OA pro-moted an additive growth effect when used together with insu-lin-like growth factor 1 (IGF1). In contrast, OA had no growthinducing effects and inhibited IGF1-induced proliferation in malig-nant cells. This was reported in rat models where a rich diet of OOresulted in a slower progression of induced mammary tumors(Zeng et al., 2010).
Apocytochrome c is the heme-free precursor of cytochrome c,which is synthesized in the cytosol and post-translationally im-ported into the mitochondria (Wang et al., 2003). Once it entersthe intermembrane space, apocytochrome c is converted intoholocytochrome c and is attached to the inner mitochondrialmembrane where it plays a role in the production of energy (Stu-art et al., 1990). However, when cytochrome c detaches and es-capes into the cytosol, a cascade of events commences, whichultimately results in the triggering of apoptosis via the intrinsicmitochondria-mediated pathway (Fulda and Debatin, 2006; Kro-emer and Reed, 2000). In this pathway, Bcl-2 proteins regulateapoptosis by altering the mitochondrial membrane potential lead-ing to the release of cytochrome c (Newmeyer and Ferguson-Mill-er, 2003). Bcl-2 overexpression was also shown to increasecellular glutathione (GSH) levels, which is associated with in-creased resistance to apoptotic-inducing drugs (Meredith et al.,1998; Voehringer, 1999), while GSH depletion enhances apoptosisinduced by anti-cancer drugs such as cisplatin in Bcl-2 expressingcells (Rudin et al., 2003). Depletion of GSH levels have also beenreported to cause elevations in lipid peroxidase and protein car-bonylation in MCF-7 and MDA-MB-231 cells (Parihar et al.,2010). Studies have also shown that cellular levels of GSH arelinked to NADPH levels, a cofactor of glutathione reductase whichcatalyzes the reduction of glutathione disulfide to GSH. The pro-duction of GSH then reduces hyperoxides to water, maintaininga low level of oxidative stress within cells. All these indicatethe importance of GSH during oxidative stress and apoptosis inhuman mammary adenocarcinoma cells (Parihar et al., 2010;Spitz et al., 2000).
Aromatase, a member of the cytochrome P450 (CYP) enzymefamily, catalyzes the rate-limiting conversion of androstenedioneto estrone and of testosterone to estradiol, using NADPH as a cofac-tor (Ghosh et al., 2009). Increase in aromatase activity has beencorrelated to alcohol consumption, old age, obesity, insulin andgonadotropins levels (Smith and Dowsett, 2003). In post-meno-pausal women, ER-positive breast cancers often fail to respond,or develops resistance during treatment leading to disease recur-rence (Ali and Coombes, 2002). Third generation non-steroidal aro-matase inhibitors such as letrozole, vorozole and anastrozolepotentially prevent estrogen biosynthesis rather than blocking ERactivation (Santen and Harvey, 1999) and are currently being usedas adjuvant therapies for early and late stage breast cancer (Cruci-tta et al., 2001).
The present study was undertaken to investigate the effectsof EVOO and its major FA component (omega-9 OA) in combi-nation with aromatase inhibitors such as anastrozole and letroz-ole in ER-positive mammary adenocarcinoma cells. Here wehypothesized that by depleting GSH levels with EVOO and OA,the efficacy of aromatase inhibitors will be enhanced in MCF-7 cells.
2. Materials and methods
2.1. Chemicals and reagents
Dimethyl sulfoxide (DMSO) (Cat. No. 317275), 3-(4,5-dimethylthiazol-2-41)-2,5-diphenyl tetrazolium bromide (MTT) (Cat. No. 474989) and annexin V-FITCapoptosis detection kit (Cat. No. PF032) were purchased from EMD Millipore (Bille-rica, MA, USA). Roswell Park Memorial Institute (RPMI-1640) media (Cat. No. 17-702F), fetal bovine serum (FBS) (Cat. No. 14-502F), trypsin (Cat. No. 17-160E),and penicillin /streptomycin (Cat. No. 17-602F) were purchased from Lonza Inc.(Basel, Switzerland). Letrozole (Cat. No. L6545), Anastrozole (Cat. No. A2736) andoleic acid (Cat. No. O1383) were purchased from Sigma Aldrich (St. Louis, MO,USA). Commercial EVOO was purchased from Monte Verde Pte. Ltd. (KL, Malaysia).
2.2. Cell lines and culture conditions
ER-positive human breast adenocarcinoma cell lines (MCF-7) were kindly do-nated by Prof. Datuk Dr. Rohana Yusof from the Department of Molecular Medicine,Faculty of Medicine, University of Malaya, while normal human mammary epithe-lial cells (HMEC) were purchased from Lonza Inc. (Basel, Switzerland). MCF-7 cellswere cultured as monolayers in RPMI-1640 media, supplemented with L-glutamine,10.0% (v/v) FBS, 100.0 U/ml penicillin and 100.0 mg/ml streptomycin. HMEC weremaintained in mammary epithelial growth media (MEGM) with growth supple-ments provided by the manufacturer, Lonza Inc. (Basel, Switzerland). All runningcultures were maintained at 10 passages or fewer in a humidified incubator at37 �C in 5.0% CO2 and 95.0% air. All treatments in this study were conducted inFBS-free media.
2.3. GC and GC–MS analysis
Analysis of EVOO constituents were carried out by GC and GC–MS. EVOO sam-ples were dissolved in pentane (C5H12) followed by GC analysis on the Agilent7890A gas chromatograph instrument (Agilent Technologies, Santa Clara, CA,USA) with HP-5 capillary column (30 m � 0.32 mm � 0.25 lm). The carrier gaswas nitrogen at a 1.0 ml/min constant flow and with an injection volume of 1 ll.Injector and inlet temperatures were set at 250 �C and a splitting ratio of 1:20.The oven temperature program was set as: initial temperature 90 �C hold for1 min, 90–190 �C (5 �C/min) hold for 10 min, 190–250 �C (3 �C/min) and hold for10 min, with a total run time of 61 min. GC–MS analysis was performed using theShimadzu GCMS-QP 2010 PLUS (Shimadzu Corporation, Kyoto, Japan) with DB-5capillary column (30 m � 0.25 mm � 0.25 lm) and helium as the carrier gas.The oven temperature program was set similar to the GC parameters. Mass spectrawere performed at 70 eV over a scan range of 40–600 amu. Retention indices weredetermined with C7 to C30 alkane standards as references. Relative amounts of indi-vidual compounds were based on peak areas obtained without FID response correc-tion factor. Identification of compounds was based on the GC–MS library (NIST08)while comparing Kovats retention index (KI) with literature.
2.4. MTT cell viability assay
Cytotoxic effects of EVOO, OA, letrozole and anastrozole on MCF-7 cells weredetermined using the MTT assay. Briefly, 2.0 � 104 cells were treated in triplicateswith various concentrations of letrozole (up to 50 lM) or anastrozole (up to100 lM) and/or in combination of EVOO (up to 125 lg/ml) or OA (up to 80 lg/ml) for 24 h. All EVOO and OA dosages used in this study were within or less thanphysiologically relevant concentrations. Solvent controls were conducted simulta-neously using either DMSO (for EVOO) or ethanol (for OA) with appropriate concen-trations corresponding to each treatment. MTT reagent (5.0 mg/ml) was added andcells were incubated in the dark at 37 �C. DMSO was then added to dissolve purpleformazan precipitates and a Tecan Sunrise� microwell plate reader (Tecan, Männe-dorf, Switzerland) was used to detect absorbance/reference at 570/650 nm.
2.5. Annexin V-FITC/PI apoptosis assay
Detection of apoptosis was conducted using the annexin V-FITC/PI apoptosisdetection kit according to manufacturer’s protocol. Briefly, MCF-7 cells were seededin RPMI-1640 with 10% (v/v) FBS for 24 h and replaced with FBS-free media duringtreatments. Cells were then treated for 24 h with various combinations of OA (20and 40 lg/ml) and EVOO (100 and 125 lg/ml) in combination with anastrozole(40 lM) or letrozole (20 lM). Cell pellets were washed in 1x PBS and resuspendedin 0.5 ml of media binding reagent. Annexin V-FITC (1.25 ll) was added and cellswere incubated for 15 min in the dark. After incubation, 10 ll of PI was addedand cells were analyzed immediately using a BD FACSCANTO™ II flow cytometerand the CellQuest Pro IVD software (Becton Dickenson, Mountain View, CA, USA).
A.M. Ismail et al. / Food and Chemical Toxicology 62 (2013) 817–824 819
2.6. Cytochrome c immunofluorescence–immunocytochemistry (IF–IC)
Determination of cytochrome c localization was done using the ApoTrack™cytochrome c apoptosis IF–IC antibody kit (Cat. No. MSA07) (MitoSciences Inc., Eu-gene, OR, USA) according to manufacturer’s protocol on MCF-7 cells treated withEVOO, OA and aromatase inhibitors. Briefly, monolayer cells were grown and trea-ted on coverslips in 6-well plates, using untreated cells containing treatment-equiv-alent DMSO/ethanol volumes as a solvent control for 24 h. Cells were then fixedwith 4% (v/v) paraformaldehyde dissolved in 1x PBS for 20 min at room tempera-ture. Coverslips were placed in antigen retrieval buffer (100 mM Tris, 5% w/v urea,pH 9.5), at 95oC for 10 min to improve antigen detection. Permeabilization buffer(0.5 ml of 1% Triton� X-100) was added and specimens were blocked with 10%(v/v) normal goat serum for 1 h. Cells were incubated with primary anti-cyto-chrome c IgG and anti-complex (Va) IgG for 2 h. Unbound antibodies were washedaway, followed by incubation with secondary goat anti-mouse IgG2a-FITC and goatanti-mouse IgG2b-Texas Red (TXRD) for 1 h. Imaging was processed using an in-verted fluorescence microscope, Nikon ECLIPSE TS100 with excitation/emissionwavelength at 590/620 nm for TXRD and 488/520 nm for FITC.
2.7. Cytochrome c and glutathione ELISA
In order to quantify the intracellular levels of cytochrome c and GSH, the humancytochrome c sandwich ELISA kit (Cat. No. KA0160) (Abnova, Taipei City, Taiwan)and the GSH 96-wells sandwich ELISA kit (Cat. No. 703002-96) (Cayman Chemical,Ann Arbor, MI, USA) were used according to manufacturer’s protocol. Briefly, trea-ted MCF-7 cells were collected and lysed with 500 ll of cell lysis buffer. Proteinsamples were collected and normalized using the Bradford assay to a final concen-tration of 10 lg/ml. A total of 100 ll of protein samples were added into each pre-coated well followed by the addition of biotinylated antibodies against cytochromec and GSH. Detection was done using streptavidin-HRP conjugates and TMB sub-strate. A microtiter plate reader, Tecan Sunrise� was used to detect absorbance/ref-erence at 450/650 nm and mean concentrations from triplicates were calculatedusing a standard curve.
2.8. Quantification of estrone (E1) levels
A solid phase competitive inhibition enzyme immunoassay was used to deter-mine estrone levels in MCF-7 lysates using the Estrone ELISA kit (Cat. No. DB52051)(IBL International, Hamburg, Germany) according to manufacturer’s protocol.Briefly, rabbit anti-E1 polyclonal IgGs were pre-coated onto microplate wells fol-lowed by competitive binding between 100 ll of avidin-HRP conjugated human
0 10 25 50 75 100 125
020406080
100120
0 5 10 15 20 40 80
EVOO concentration (µg/ml)
Rel
ativ
e v
iabi
lity
(%)
OA concentration (μg/ml)
Treated with OATreated with EVOO
0
20
40
60
80
100
120
0 20 40 60 80 100 120
Rel
ativ
e v
iabi
lity
(%)
Anastrozole concentration (μM)
Anas.Anas.+EVOO (50 μg/ml)Anas.+OA (15 μg/ml)
A
C
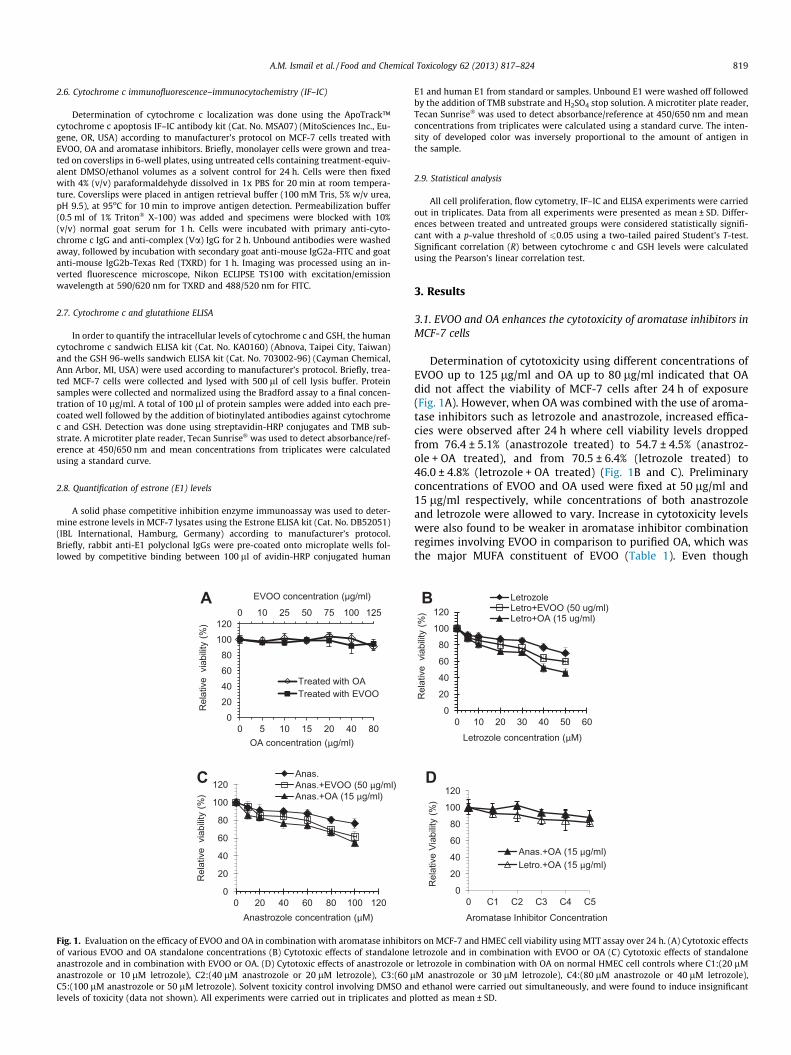
Fig. 1. Evaluation on the efficacy of EVOO and OA in combination with aromatase inhibitoof various EVOO and OA standalone concentrations (B) Cytotoxic effects of standaloneanastrozole and in combination with EVOO or OA. (D) Cytotoxic effects of anastrozole oranastrozole or 10 lM letrozole), C2:(40 lM anastrozole or 20 lM letrozole), C3:(60 lC5:(100 lM anastrozole or 50 lM letrozole). Solvent toxicity control involving DMSO anlevels of toxicity (data not shown). All experiments were carried out in triplicates and p
E1 and human E1 from standard or samples. Unbound E1 were washed off followedby the addition of TMB substrate and H2SO4 stop solution. A microtiter plate reader,Tecan Sunrise� was used to detect absorbance/reference at 450/650 nm and meanconcentrations from triplicates were calculated using a standard curve. The inten-sity of developed color was inversely proportional to the amount of antigen inthe sample.
2.9. Statistical analysis
All cell proliferation, flow cytometry, IF–IC and ELISA experiments were carriedout in triplicates. Data from all experiments were presented as mean ± SD. Differ-ences between treated and untreated groups were considered statistically signifi-cant with a p-value threshold of 60.05 using a two-tailed paired Student’s T-test.Significant correlation (R) between cytochrome c and GSH levels were calculatedusing the Pearson’s linear correlation test.
3. Results
3.1. EVOO and OA enhances the cytotoxicity of aromatase inhibitors inMCF-7 cells
Determination of cytotoxicity using different concentrations ofEVOO up to 125 lg/ml and OA up to 80 lg/ml indicated that OAdid not affect the viability of MCF-7 cells after 24 h of exposure(Fig. 1A). However, when OA was combined with the use of aroma-tase inhibitors such as letrozole and anastrozole, increased effica-cies were observed after 24 h where cell viability levels droppedfrom 76.4 ± 5.1% (anastrozole treated) to 54.7 ± 4.5% (anastroz-ole + OA treated), and from 70.5 ± 6.4% (letrozole treated) to46.0 ± 4.8% (letrozole + OA treated) (Fig. 1B and C). Preliminaryconcentrations of EVOO and OA used were fixed at 50 lg/ml and15 lg/ml respectively, while concentrations of both anastrozoleand letrozole were allowed to vary. Increase in cytotoxicity levelswere also found to be weaker in aromatase inhibitor combinationregimes involving EVOO in comparison to purified OA, which wasthe major MUFA constituent of EVOO (Table 1). Even though
0
20
40
60
80
100
120
0 10 20 30 40 50 60
Rel
ativ
e v
iabi
lity
(%)
Letrozole concentration (μM)
LetrozoleLetro+EVOO (50 ug/ml)Letro+OA (15 ug/ml)
0
20
40
60
80
100
120
0 C1 C2 C3 C4 C5
Rel
ativ
e Vi
abilit
y (%
)
Aromatase Inhibitor Concentration
Anas.+OA (15 μg/ml)Letro.+OA (15 μg/ml)
B
D
rs on MCF-7 and HMEC cell viability using MTT assay over 24 h. (A) Cytotoxic effectsletrozole and in combination with EVOO or OA (C) Cytotoxic effects of standaloneletrozole in combination with OA on normal HMEC cell controls where C1:(20 lMM anastrozole or 30 lM letrozole), C4:(80 lM anastrozole or 40 lM letrozole),d ethanol were carried out simultaneously, and were found to induce insignificantlotted as mean ± SD.

Table 1List of major compounds identified in EVOO samples using GC and GC–MS analysis.
No Retention time (min) Retention index (KI)a Compound name Molecular formula Area (%)
1 24.64 1953 Pentadecanal C15H30O 7.082 31.76 2137 Hexadecanoic acid, methyl ester (Palmitic acid – Major SFA)b C17H34O2 46.593 44.37 2472 9-Octadecanoic acid, methyl ester (Oleic acid – Major MUFA)c C19H36O2 13.234 54.18 2812 Squalene C30 H50 28.92
a KI denotes Kovats Retention Index.b SFA denotes saturated fatty acid.c MUFA denotes monounsaturated fatty acid.
820 A.M. Ismail et al. / Food and Chemical Toxicology 62 (2013) 817–824
normal HMEC controls indicated that no adverse cytotoxic effectswere observed (viability >80%) when maximum concentrations ofboth aromatase inhibitors were used in combination with OA over24 h, a slight cytostasis effect may be present (Fig. 1D). Solvent tox-icity control involving DMSO and ethanol were carried out simul-taneously, and were found to induce insignificant levels of toxicity.
3.2. Induction of apoptosis by EVOO and OA in combination witharomatase inhibitors
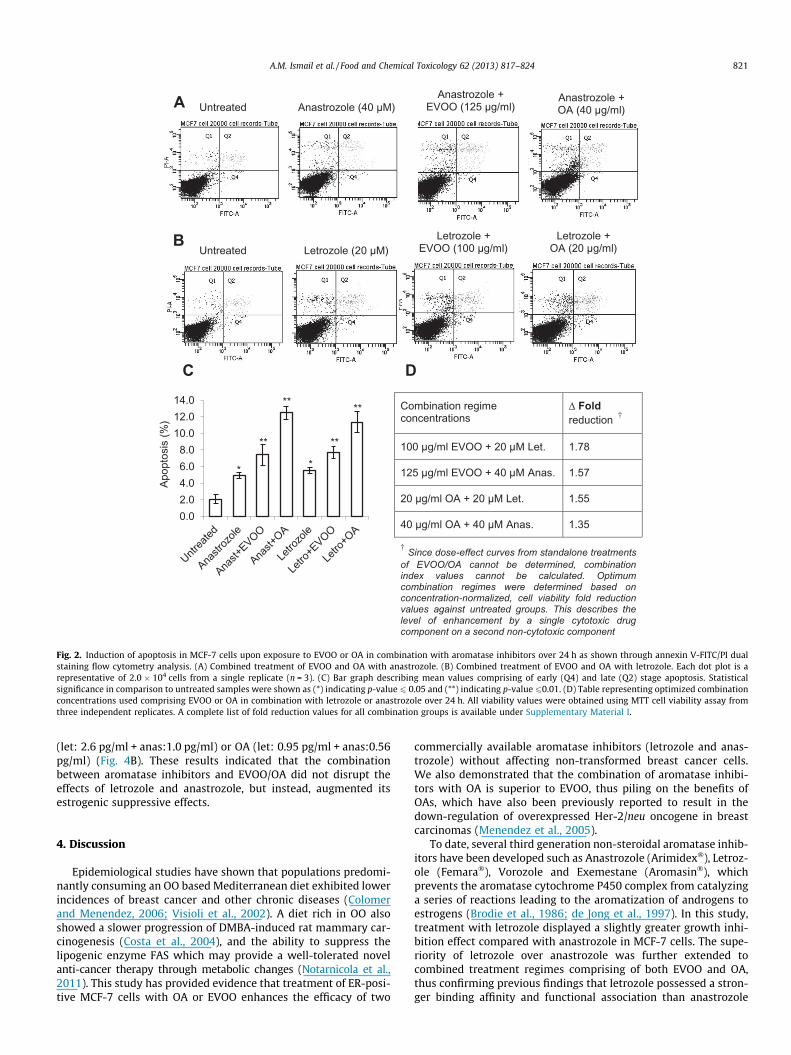
In order to justify the optimum combination ratio betweenEVOO/OA with letrozole and anastrozole, a series of combinationtreatments were performed using the MTT cell viability assay.Since EVOO and OA were non-cytotoxic components in a dual drugcombination regime, combination index values as well as the pres-ence of synergistic relationships cannot be determined. Therefore,optimal combinations were justified based on normalized cell via-bility fold reduction values against untreated groups. This de-scribes the level of enhancement by a single cytotoxic drugcomponent on a second non-cytotoxic component (Chou, 2006).Optimum combination concentrations were obtained and usedfor subsequent assays (Fig. 2D). Annexin V-FITC/PI flow cytometryresults indicated that the population of cells shifted from viablecells to early and late stage apoptosis, followed by secondarynecrosis after 24 h of treatment (Fig. 2A and B). As with previousMTT cytotoxicity analysis, the extent of apoptosis was greatest inletrozole + OA and anastrozole + OA combination treatment groupsat 11.32% and 12.48% respectively, followed by combined treat-ment groups containing EVOO and lastly standalone aromataseinhibitors (Fig. 2C). This indicated that both EVOO and OA playeda role towards aromatase inhibitor’s augmentation of apoptosis,and that OA was indeed the major FA constituent in EVOO respon-sible for this observation.
3.3. EVOO/OA in combination with aromatase inhibitors modulates therelease of cytochrome c from the mitochondria
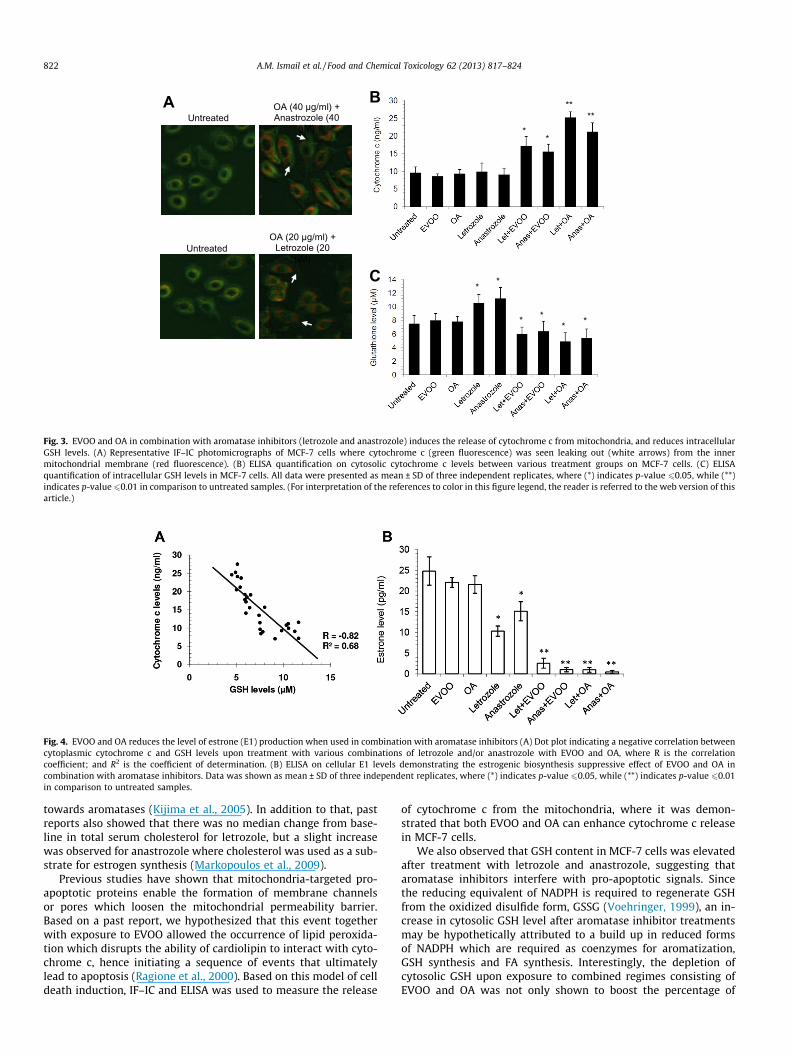
Since a combined treatment of EVOO and/or OA with aromataseinhibitors (letrozole and anastrozole) successfully induced apopto-sis in MCF-7 cells, we then investigated the facilitation of cyto-chrome c release from inner mitochondria membrane to thecytosol. IF–IC results showed that untreated cells indicated anoverlapping presence of cytochrome c within the mitochondrialmembrane with minimal signs of cytochrome c release. However,when both aromatase inhibitors were used in combination withOA, a release of cytochrome c into the cytosol was observed(Fig. 3A). ELISA on cytosolic cytochrome c levels confirmed ourIF–IC observations, where both standalone letrozole and anastroz-ole treated cells did not show any significant difference in cytosoliccytochrome c levels compared to untreated cells (Fig. 3B). In con-trast, combination treatment involving EVOO or OA showed a1.5–2.5 fold increase in cytosolic cytochrome c levels, thus affirm-ing the involvement of the intrinsic pathway. The effect of OA-combined with letrozole in comparison to anastrozole was also
shown to be superior in terms of cytochrome c release by19.2 ± 2.1% (Fig. 3B). In reference to annexin V-FITC data demon-strated opposing observations on the superiority of letrozole overanastrozole compared to cytochrome c release and initial MTTdata, its findings were statistically insignificant (1.16% variationbetween anastrozole and letrozole with a p-value of 0.28) andwas therefore disregarded.
3.4. EVOO/OA nullifies the increase of GSH levels by aromataseinhibitors
Cellular levels of GSH are positively correlated to NADPH levels,which are in turn increased through the inhibition of aromatiza-tion reactions by aromatase inhibitors. Therefore, we investigatedwhether EVOO and OA could counteract the increasing levels ofGSH following exposure to letrozole and anastrozole in MCF-7 cellsthrough ELISA. Our observations show that upon letrozole andanastrozole treatment, levels of GSH were increased to10.5 ± 1.3 lM and 11.2 ± 1.6 lM respectively, compared to un-treated cells (7.5 ± 1.2 lM) presumably from an increase in NADPHlevels (Fig. 3C). However, when both aromatase inhibitors wereused in combination with EVOO or OA, this increase in GSH levelswere nullified and reduced to 6.0 ± 1.0 lM by EVOO and4.9 ± 1.3 lM by OA, which was below untreated control levels(Fig. 3C). These findings revealed that treatment with aromataseinhibitors alone increases GSH levels, while combination of aroma-tase inhibitors with EVOO or OA depletes cellular GSH levels.
3.5. Cytochrome c release is inversely correlated to GSH levels
In order to investigate whether the occurrence of apoptosisthrough the intrinsic pathway was associated to GSH levels, a Pear-son correlation analysis was performed. The correlation dot plotindicated that the release of cytochrome c from the mitochondriainner membrane was negatively correlated towards the levels ofGSH with a correlation coefficient, R of �0.82 (Fig. 4A). Thestrength of the relationship as represented by the coefficient ofdetermination, R2, indicates that at least 68% (R2 = 0.68) of the totalvariation in cytosolic cytochrome c release is inversely related toGSH levels, which is in turn hypothetically affected by levels of ex-cess NADPH within the cell.
3.6. EVOO/OA enhances the estrogenic suppressive effects of aromataseinhibitors
To determine whether EVOO/OA disrupted the efficacy of aro-matase inhibitors owing to reduced GSH levels and the under uti-lization of NADPH, an ELISA was performed on estrone (E1) levels,which is the predominant estrogen in post-menopausal women. Itwas observed that intracellular E1 levels were reduced as expectedwhen exposed to standalone letrozole (10.3 pg/ml) and standaloneanastrozole (15.1 pg/ml) from untreated cells (24.8 pg/ml). E1levels were further significantly reduced when cells wereexposed to both aromatase inhibitors in combination with EVOO
0.02.04.06.08.0
10.012.014.0
Apop
tosi
s (%
)
Combination regime concentrations
Δ Fold reduction †
100 µg/ml EVOO + 20 µM Let. 1.78
125 µg/ml EVOO + 40 µM Anas. 1.57
20 µg/ml OA + 20 µM Let. 1.55
40 µg/ml OA + 40 µM Anas. 1.35
† Since dose-effect curves from standalone treatments of EVOO/OA cannot be determined, combination index values cannot be calculated. Optimum combination regimes were determined based on concentration-normalized, cell viability fold reduction values against untreated groups. This describes the level of enhancement by a single cytotoxic drug component on a second non-cytotoxic component
C D
A Untreated Anastrozole (40 µM) Anastrozole +
EVOO (125 µg/ml)Anastrozole +OA (40 µg/ml)
B Letrozole +EVOO (100 µg/ml)
Letrozole +OA (20 µg/ml)Untreated Letrozole (20 µM)
** **
****
**
Fig. 2. Induction of apoptosis in MCF-7 cells upon exposure to EVOO or OA in combination with aromatase inhibitors over 24 h as shown through annexin V-FITC/PI dualstaining flow cytometry analysis. (A) Combined treatment of EVOO and OA with anastrozole. (B) Combined treatment of EVOO and OA with letrozole. Each dot plot is arepresentative of 2.0 � 104 cells from a single replicate (n = 3). (C) Bar graph describing mean values comprising of early (Q4) and late (Q2) stage apoptosis. Statisticalsignificance in comparison to untreated samples were shown as (*) indicating p-value 6 0.05 and (**) indicating p-value 60.01. (D) Table representing optimized combinationconcentrations used comprising EVOO or OA in combination with letrozole or anastrozole over 24 h. All viability values were obtained using MTT cell viability assay fromthree independent replicates. A complete list of fold reduction values for all combination groups is available under Supplementary Material I.
A.M. Ismail et al. / Food and Chemical Toxicology 62 (2013) 817–824 821
(let: 2.6 pg/ml + anas:1.0 pg/ml) or OA (let: 0.95 pg/ml + anas:0.56pg/ml) (Fig. 4B). These results indicated that the combinationbetween aromatase inhibitors and EVOO/OA did not disrupt theeffects of letrozole and anastrozole, but instead, augmented itsestrogenic suppressive effects.
4. Discussion
Epidemiological studies have shown that populations predomi-nantly consuming an OO based Mediterranean diet exhibited lowerincidences of breast cancer and other chronic diseases (Colomerand Menendez, 2006; Visioli et al., 2002). A diet rich in OO alsoshowed a slower progression of DMBA-induced rat mammary car-cinogenesis (Costa et al., 2004), and the ability to suppress thelipogenic enzyme FAS which may provide a well-tolerated novelanti-cancer therapy through metabolic changes (Notarnicola et al.,2011). This study has provided evidence that treatment of ER-posi-tive MCF-7 cells with OA or EVOO enhances the efficacy of two
commercially available aromatase inhibitors (letrozole and anas-trozole) without affecting non-transformed breast cancer cells.We also demonstrated that the combination of aromatase inhibi-tors with OA is superior to EVOO, thus piling on the benefits ofOAs, which have also been previously reported to result in thedown-regulation of overexpressed Her-2/neu oncogene in breastcarcinomas (Menendez et al., 2005).
To date, several third generation non-steroidal aromatase inhib-itors have been developed such as Anastrozole (Arimidex�), Letroz-ole (Femara�), Vorozole and Exemestane (Aromasin�), whichprevents the aromatase cytochrome P450 complex from catalyzinga series of reactions leading to the aromatization of androgens toestrogens (Brodie et al., 1986; de Jong et al., 1997). In this study,treatment with letrozole displayed a slightly greater growth inhi-bition effect compared with anastrozole in MCF-7 cells. The supe-riority of letrozole over anastrozole was further extended tocombined treatment regimes comprising of both EVOO and OA,thus confirming previous findings that letrozole possessed a stron-ger binding affinity and functional association than anastrozole
A B
C
UntreatedOA (20 µg/ml) +
Letrozole (20 µM)
UntreatedOA (40 µg/ml) + Anastrozole (40
µM)
** **
* *
**
****
Fig. 3. EVOO and OA in combination with aromatase inhibitors (letrozole and anastrozole) induces the release of cytochrome c from mitochondria, and reduces intracellularGSH levels. (A) Representative IF–IC photomicrographs of MCF-7 cells where cytochrome c (green fluorescence) was seen leaking out (white arrows) from the innermitochondrial membrane (red fluorescence). (B) ELISA quantification on cytosolic cytochrome c levels between various treatment groups on MCF-7 cells. (C) ELISAquantification of intracellular GSH levels in MCF-7 cells. All data were presented as mean ± SD of three independent replicates, where (*) indicates p-value 60.05, while (**)indicates p-value60.01 in comparison to untreated samples. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of thisarticle.)
Fig. 4. EVOO and OA reduces the level of estrone (E1) production when used in combination with aromatase inhibitors (A) Dot plot indicating a negative correlation betweencytoplasmic cytochrome c and GSH levels upon treatment with various combinations of letrozole and/or anastrozole with EVOO and OA, where R is the correlationcoefficient; and R2 is the coefficient of determination. (B) ELISA on cellular E1 levels demonstrating the estrogenic biosynthesis suppressive effect of EVOO and OA incombination with aromatase inhibitors. Data was shown as mean ± SD of three independent replicates, where (*) indicates p-value 60.05, while (**) indicates p-value 60.01in comparison to untreated samples.
822 A.M. Ismail et al. / Food and Chemical Toxicology 62 (2013) 817–824
towards aromatases (Kijima et al., 2005). In addition to that, pastreports also showed that there was no median change from base-line in total serum cholesterol for letrozole, but a slight increasewas observed for anastrozole where cholesterol was used as a sub-strate for estrogen synthesis (Markopoulos et al., 2009).
Previous studies have shown that mitochondria-targeted pro-apoptotic proteins enable the formation of membrane channelsor pores which loosen the mitochondrial permeability barrier.Based on a past report, we hypothesized that this event togetherwith exposure to EVOO allowed the occurrence of lipid peroxida-tion which disrupts the ability of cardiolipin to interact with cyto-chrome c, hence initiating a sequence of events that ultimatelylead to apoptosis (Ragione et al., 2000). Based on this model of celldeath induction, IF–IC and ELISA was used to measure the release
of cytochrome c from the mitochondria, where it was demon-strated that both EVOO and OA can enhance cytochrome c releasein MCF-7 cells.
We also observed that GSH content in MCF-7 cells was elevatedafter treatment with letrozole and anastrozole, suggesting thataromatase inhibitors interfere with pro-apoptotic signals. Sincethe reducing equivalent of NADPH is required to regenerate GSHfrom the oxidized disulfide form, GSSG (Voehringer, 1999), an in-crease in cytosolic GSH level after aromatase inhibitor treatmentsmay be hypothetically attributed to a build up in reduced formsof NADPH which are required as coenzymes for aromatization,GSH synthesis and FA synthesis. Interestingly, the depletion ofcytosolic GSH upon exposure to combined regimes consisting ofEVOO and OA was not only shown to boost the percentage of

Fig. 5. A simplified mechanism depicting an association between fatty acids (OA from EVOO) and aromatase inhibitors on estrone biosynthesis, NADPH levels, GSH depletion,lipid peroxidation, cytochrome c release, and the induction of apoptosis.
A.M. Ismail et al. / Food and Chemical Toxicology 62 (2013) 817–824 823
apoptotic cells, but also enhanced the efficiency of aromataseinhibitors as shown by reduced E1 biosynthetic levels. Throughmechanisms that have yet to be elucidated, we concluded thatOAs in EVOO play a significant role in GSH synthesis, lipid peroxi-dation and ROS generation, which have been shown to be involvedin the induction of apoptosis as well as other cell death-mediatingfactors originating from severe oxidative stress (Buttke and Sand-strom, 1994; Toborek et al., 2002).
A negative correlation between GSH levels and cytosolic cyto-chrome c also reinforced the hypothesis that depletion in GSH isconnected to lipid peroxidation, the release of cytochrome c andsubsequently, apoptosis. GSH requires constant replenishmentfrom GSSG, and NADPH is known to be essential for its regenera-tion in cellular defence against oxidative stress to promote cancercell growth and survival (Kil et al., 2010). The fact that NADPH uti-lization occurs as a result of FA oxidation inhibition even in thepresence of glutamine and glucose suggests that NADPH produc-tion powered by these nutrients may either be insufficient or im-poses a minimal impact on maintaining the anti-oxidant system.These suggest that there is a link between FA, GSH, ROS and aroma-tase inhibitors, where NADPH is shared and provided for regener-ation of the GSH anti-oxidant system to protect against oxidativedamage (Yuneva et al., 2007). It is also proposed that the inhibitionof aromatization by letrozole or anastrozole on its own increasescellular GSH levels, resulting in higher drugs resistance occurrence.The administration of FA, specifically natural OA from EVOO cantherefore be employed in combination with aromatase inhibitorsto reduce GSH levels, which has previously been shown to be a ma-jor culprit for drug resistance (Chen and Kuo, 2010).
This study shows that treatment of MCF-7 cells with aromataseinhibitors presumably increases reduced forms of NADPH, leadingto the provision of reducing equivalents to regenerate intracellularGSH. The declination of GSH levels in the presence of aromataseinhibitors and EVOO/OA provides direct evidence that EVOO playsa role in GSH depletion through disruption of membrane potentialand lipid peroxidation which enhances the release of cytochrome cfrom the mitochondria, inducing apoptosis in MCF-7 cells (Fig. 5).Even though EVOO contains a wide variety of other metabolitesincluding polyphenols, iridoids and triterpenes which could possi-bly influence the overall activity of EVOO, the consistent paralleleffects observed between EVOO and purified OA in all experimentsserved as a strong indicator that OA remains the major constituentand key player responsible in augmenting the anti-cancer effects ofaromatase inhibitors.
5. Conclusion
In conclusion, the employment of aromatase inhibitors in com-bination with EVOO/OA could orchestrate a reduction in
intracellular estrogen biosynthesis which feeds ER-positive cells,while simultaneously depleting GSH levels and the subsequentinduction of apoptosis.
Conflict of Interest
The authors declare that there are no conflicts of interest.
Acknowledgements
This study was supported by the Al-Neelain University ResearchGrant (57-02-03-1020), the University of Malaya Postgraduate Re-search Grants (PPP) (PV043-2011A and PV050/2012A), the HighImpact Research Grant (HIR) (UM-MOHE UM.C/625/1/HIR/MOHE/SC/37) and the University of Malaya Research Grant (UMRG)(RG037-10BIO).
Appendix A. Supplementary material
Supplementary data associated with this article can be found, inthe online version, at http://dx.doi.org/10.1016/j.fct.2013.10.024.
References
Ali, S., Coombes, R.C., 2002. Endocrine-responsive breast cancer and strategies forcombating resistance. Nature Reviews. Cancer 2, 101–112.
Andrewes, P., Busch, J.L., de Joode, T., Groenewegen, A., Alexandre, H., 2003. Sensoryproperties of virgin olive oil polyphenols: identification of deacetoxy-ligstrosideaglycon as a key contributor to pungency. Journal of Agricultural and FoodChemistry 51, 1415–1420.
Bogani, P., Galli, C., Villa, M., Visioli, F., 2007. Postprandial anti-inflammatory andantioxidant effects of extra virgin olive oil. Atherosclerosis 190, 181–186.
Brodie, A.M., Wing, L.Y., Goss, P., Dowsett, M., Coombes, R.C., 1986. Aromataseinhibitors and the treatment of breast cancer. Journal of Steroid Biochemistry24, 91–97.
Buttke, T.M., Sandstrom, P.A., 1994. Oxidative stress as a mediator of apoptosis.Immunology Today 15, 7–10.
Chen, H.H., Kuo, M.T., 2010. Role of glutathione in the regulation of Cisplatinresistance in cancer chemotherapy. Metal-based Drugs.
Chou, T.C., 2006. Theoretical basis, experimental design, and computerizedsimulation of synergism and antagonism in drug combination studies.Pharmacological Reviews 58, 621–681.
Colomer, R., Menendez, J.A., 2006. Mediterranean diet, olive oil and cancer. Clinical& Translational Oncology : Official Publication of the Federation of SpanishOncology Societies and of the National Cancer Institute of Mexico 8, 15–21.
Costa, I., Moral, R., Solanas, M., Escrich, E., 2004. High-fat corn oil diet promotes thedevelopment of high histologic grade rat DMBA-induced mammaryadenocarcinomas, while high olive oil diet does not. Breast Cancer Researchand Treatment 86, 225–235.
Crucitta, E., Locopo, N., Silvestris, N., De Lena, M., Lorusso, V., 2001. The role ofletrozole (Femara(R)) in breast cancer therapy: a clinical review. Drugs of Today37, 639–644.
de Jong, P.C., van de Ven, J., Nortier, H.W., Maitimu-Smeele, I., Donker, T.H., Thijssen,J.H., Slee, P.H., Blankenstein, R.A., 1997. Inhibition of breast cancer tissuearomatase activity and estrogen concentrations by the third-generationaromatase inhibitor vorozole. Cancer Research 57, 2109–2111.

824 A.M. Ismail et al. / Food and Chemical Toxicology 62 (2013) 817–824
El, S.N., Karakaya, S., 2009. Olive tree (Olea europaea) leaves: potential beneficialeffects on human health. Nutrition Reviews 67, 632–638.
Fabiani, R., Rosignoli, P., De Bartolomeo, A., Fuccelli, R., Servili, M., Montedoro, G.F.,Morozzi, G., 2008. Oxidative DNA damage is prevented by extracts of olive oil,hydroxytyrosol, and other olive phenolic compounds in human bloodmononuclear cells and HL60 cells. The Journal of Nutrition 138, 1411–1416.
Fu, S., Arraez-Roman, D., Segura-Carretero, A., Menendez, J.A., Menendez-Gutierrez,M.P., Micol, V., Fernandez-Gutierrez, A., 2010. Qualitative screening of phenoliccompounds in olive leaf extracts by hyphenated liquid chromatography andpreliminary evaluation of cytotoxic activity against human breast cancer cells.Analytical and Bioanalytical Chemistry 397, 643–654.
Fulda, S., Debatin, K.M., 2006. Extrinsic versus intrinsic apoptosis pathways inanticancer chemotherapy. Oncogene 25, 4798–4811.
Ghosh, D., Griswold, J., Erman, M., Pangborn, W., 2009. Structural basis for androgenspecificity and oestrogen synthesis in human aromatase. Nature 457, 219–223.
Kijima, I., Itoh, T., Chen, S., 2005. Growth inhibition of estrogen receptor-positiveand aromatase-positive human breast cancer cells in monolayer and spheroidcultures by letrozole, anastrozole, and tamoxifen. The Journal of SteroidBiochemistry and Molecular Biology 97, 360–368.
Kil, I.S., Chung, K.H., Park, J.W., 2010. Silencing of mitochondrial NADP+-dependentisocitrate dehydrogenase gene enhances selenite-induced apoptosis. FreeRadical Research 44, 332–339.
Kroemer, G., Reed, J.C., 2000. Mitochondrial control of cell death. Nature Medicine 6,513–519.
La Vecchia, C., Bosetti, C., 2006. Diet and cancer risk in Mediterranean countries:open issues. Public Health Nutrition 9, 1077–1082.
Lee-Huang, S., Huang, P.L., Zhang, D., Lee, J.W., Bao, J., Sun, Y., Chang, Y.T., Zhang, J.,Huang, P.L., 2007. Discovery of small-molecule HIV-1 fusion and integraseinhibitors oleuropein and hydroxytyrosol: Part I. fusion [corrected] inhibition.Biochemical and Biophysical Research Communications 354, 872–878.
Markopoulos, C., Polychronis, A., Dafni, U., Koukouras, D., Zobolas, V.,Tzorakoleftherakis, E., Xepapadakis, G., Gogas, H., 2009. Lipid changes inbreast cancer patients on exemestane treatment: final results of the TEAMGreek substudy. Annals of Oncology: Official Journal of the European Society forMedical Oncology/ESMO 20, 49–55.
Menendez, J.A., Vellon, L., Colomer, R., Lupu, R., 2005. Oleic acid, the mainmonounsaturated fatty acid of olive oil, suppresses Her-2/neu (erbB-2)expression and synergistically enhances the growth inhibitory effects oftrastuzumab (Herceptin) in breast cancer cells with Her-2/neu oncogeneamplification. Annals of Oncology: Official Journal of the European Society forMedical Oncology/ESMO 16, 359–371.
Meredith, M.J., Cusick, C.L., Soltaninassab, S., Sekhar, K.S., Lu, S., Freeman, M.L., 1998.Expression of Bcl-2 increases intracellular glutathione by inhibitingmethionine-dependent GSH efflux. Biochemical and Biophysical ResearchCommunications 248, 458–463.
Newmeyer, D.D., Ferguson-Miller, S., 2003. Mitochondria: releasing power for lifeand unleashing the machineries of death. Cell 112, 481–490.
Notarnicola, M., Pisanti, S., Tutino, V., Bocale, D., Rotelli, M.T., Gentile, A., Memeo, V.,Bifulco, M., Perri, E., Caruso, M.G., 2011. Effects of olive oil polyphenols on fattyacid synthase gene expression and activity in human colorectal cancer cells.Genes & Nutrition 6, 63–69.
Omar, S.H., 2010. Oleuropein in olive and its pharmacological effects. ScientiaPharmaceutica 78, 133–154.
Parihar, A., Parihar, M.S., Nazarewicz, R., Ghafourifar, P., 2010. Importance ofcytochrome c redox state for ceramide-induced apoptosis of human mammaryadenocarcinoma cells. Biochimica et Biophysica Acta 1800, 646–654.
Perez-Jimenez, F., Alvarez de Cienfuegos, G., Badimon, L., Barja, G., Battino, M.,Blanco, A., Bonanome, A., Colomer, R., Corella-Piquer, D., Covas, I., Chamorro-Quiros, J., Escrich, E., Gaforio, J.J., Garcia Luna, P.P., Hidalgo, L., Kafatos, A., Kris-Etherton, P.M., Lairon, D., Lamuela-Raventos, R., Lopez-Miranda, J., Lopez-Segura, F., Martinez-Gonzalez, M.A., Mata, P., Mataix, J., Ordovas, J., Osada, J.,Pacheco-Reyes, R., Perucho, M., Pineda-Priego, M., Quiles, J.L., Ramirez-Tortosa,M.C., Ruiz-Gutierrez, V., Sanchez-Rovira, P., Solfrizzi, V., Soriguer-Escofet, F., dela Torre-Fornell, R., Trichopoulos, A., Villalba-Montoro, J.M., Villar-Ortiz, J.R.,Visioli, F., 2005. International conference on the healthy effect of virgin olive oil.European Journal of Clinical Investigation 35, 421–424.
Ragione, F.D., Cucciolla, V., Borriello, A., Pietra, V.D., Pontoni, G., Racioppi, L., Manna,C., Galletti, P., Zappia, V., 2000. Hydroxytyrosol, a natural molecule occurring inolive oil, induces cytochrome c-dependent apoptosis. Biochemical andBiophysical Research Communications 278, 733–739.
Rudin, C.M., Yang, Z., Schumaker, L.M., VanderWeele, D.J., Newkirk, K., Egorin, M.J.,Zuhowski, E.G., Cullen, K.J., 2003. Inhibition of glutathione synthesis reversesBcl-2-mediated cisplatin resistance. Cancer Research 63, 312–318.
Santen, R.J., Harvey, H.A., 1999. Use of aromatase inhibitors in breast carcinoma.Endocrine-Related Cancer 6, 75–92.
Simopoulos, A.P., 2004. The traditional diet of Greece and cancer. European Journalof Cancer Prevention : The Official Journal of the European Cancer PreventionOrganisation 13, 219–230.
Smith, I.E., Dowsett, M., 2003. Aromatase inhibitors in breast cancer. The NewEngland Journal of Medicine 348, 2431–2442.
Spitz, D.R., Sim, J.E., Ridnour, L.A., Galoforo, S.S., Lee, Y.J., 2000. Glucose deprivation-induced oxidative stress in human tumor cells. A fundamental defect inmetabolism? Annals of the New York Academy of Sciences 899, 349–362.
Stuart, R.A., Nicholson, D.W., Wienhues, U., Neupert, W., 1990. Import ofapocytochrome c into the mitochondrial intermembrane space along acytochrome c1 sorting pathway. The Journal of Biological Chemistry 265,20210–20219.
Toborek, M., Lee, Y.W., Garrido, R., Kaiser, S., Hennig, B., 2002. Unsaturated fattyacids selectively induce an inflammatory environment in human endothelialcells. The American Journal of Clinical Nutrition 75, 119–125.
Trichopoulou, A., Lagiou, P., Kuper, H., Trichopoulos, D., 2000. Cancer andMediterranean dietary traditions. Cancer Epidemiology, Biomarkers &Prevention: A Publication of the American Association for Cancer Research,Cosponsored by the American Society of Preventive Oncology 9, 869–873.
Visioli, F., Poli, A., Gall, C., 2002. Antioxidant and other biological activities ofphenols from olives and olive oil. Medicinal Research Reviews 22, 65–75.
Voehringer, D.W., 1999. BCL-2 and glutathione: alterations in cellular redox statethat regulate apoptosis sensitivity. Free Radical Biology & Medicine 27, 945–950.
Wang, X., Han, X., Yang, F., 2003. Peptide 68–88 of apocytochrome c plays a crucialrole in its insertion into membrane and binding to mitochondria. Science inChina Series C, Life sciences/Chinese Academy of Sciences 46, 18–27.
Yuneva, M., Zamboni, N., Oefner, P., Sachidanandam, R., Lazebnik, Y., 2007.Deficiency in glutamine but not glucose induces MYC-dependent apoptosis inhuman cells. The Journal of Cell Biology 178, 93–105.
Zeng, L., Biernacka, K.M., Holly, J.M., Jarrett, C., Morrison, A.A., Morgan, A., Winters,Z.E., Foulstone, E.J., Shield, J.P., Perks, C.M., 2010. Hyperglycaemia confersresistance to chemotherapy on breast cancer cells: the role of fatty acidsynthase. Endocrine-Related Cancer 17, 539–551.




















