
A portable system for measuring carbon dioxide and water vapour exchange of leaves
9
plant. Cell atxd Ettvironment (1982) 5, 179-186, TECHNICAL REPORT portable system for measuring carbon dioxide and xA/ater vapour exchange of leaves C, FIELD*, J, A, BERRYt & H, A, MOONEY* *Department of Biological Sciences, Stanford University, Stanford, CA 94305 U,S,A, and fDepartment of Plant Biology, Carnegie Institution of Washington, Stanfot-d, CA 94305 U,S,A, CIW-DPB Publieation No, 735 Received 5 May 1981; aeeepted for publication 8 July 1981 Abstract. The apparatus deseribed here is a fully portable, steady-state gas exchange systet-t-i for simultaneous t-neasurements of the CO, exchatige and transpiration of single, attaehed leaves. The leaf cuvette provides teti-ipetature, humidity, and CO,- concenttation control. The system is suitable for either surveys or detailed studies of photosynthetic and stomatal responses to environmental variables, Representative data demonstrate the response time characteristics of the system and constitute the first field evidence of stotnatal behaviour consistent with a recent hypothesis eoncerning the optimum pattern of stomatal conductance for the maximization of water- use-eflieieney, li^ev-words: Lepechinia calycina (Benth,) Epl.; Lamiceae; pitcher sage; gas exchange system; photosytithesis; ttanspiiatioti; humidity lesponse; waler-use-efficiency. Introduction Gas exchange systems for measuring photosynthesis and transpitation in the field have tended to emphasize either aecutacy and environmental control at the eost of portability, or portability and convenienee, saerifieing environt-nental eonttol and accuraey. The appat-atus described here incorpotates recent technical and conceptual advanees to bring accuracy and control to a compact, fully portable system. The system pt-ovides sufficient control of the leaf envitonment to allow investigations of photo- synthetic and stotnatal responses to tempetature, light intensity, and vapour pressure deficit. With minor modifieations it can be used iti studies of the CO, coneentt ation dependence of photosynthesis. Carbon dioxide and water vapour exehange of single, attaehed leaves ean be measut-ed with an accut-acy comparable to that attained in the sophisticated rnobile laboratory described by Mooney et al (1971), Though untested in extreme habitats sueh as Death Correspondetice: Dr Christopher Field, Department of Biology, University of Utah, Salt Lake City, UT 84112, U,S.A. 0140-7791/82/0400-0179$02,()0 © 1982 Blackwell Scientific Publications Valley in the summer, the portable system has beefi used to study photosynthetie properties of plants growing under natut-al eonditions in sites throughout California, including Death Valley (Seemann, Field & Berry, 1980), Bodega Head (H, A, Mooney, unpublished), and the Sierra Nevada (C, Field, unpublished). The most aeeurate laboratory gas exchange systems are open, diffet-ential systems (cf, Ehleringer & Bjorkman, 1977), With these systems, both photo- synthesis and tt-anspit-ation at-e ealeulated from tneasured flow rates and the CO, and watet--vapour concentration differences between the air entering and that leaving the cuvette, A major feature limiting the portability of open, differential systems is the requirement for temperature-eontroUed hutnidi- fieation devices and heated gas How lines (Blootn et al., 1980), Consider an open system with a well-stirred cuvette. If the entering air stream is dry instead of moist, the cuvette equilibtates at some relative humidity determined by the leaf transpiration rate and the air flow rate through the euvette. Decreasing the air flow tate increases the steady-state telative hutnidity, and increasing the air flow rate deereases it. While the systetn is still fully differential, the iticoming humidity is zero, Beardsell, Jarvis & Davidson (1972) developed a portable porometer for tt-anspiration measutei-tients that is ati open, diffeiential system using only dry air as the inlet gas. Their steady-state porotneter operates with no tempetature-controlled components and with a single humidity sensor, iti the leaf euvette. Transpiration is calculated as the simple product of the air flow rate and the vapour density of the air in the euvette, Porometers of the type described by Beardsell et al, (1972) are tiot suitable for sitnultaneous measute- ments of photosynthesis atid transpiration. At flow rates of dry air low enough to balance tt-anspiration at tnoderate or high euvette humidities, photosynthesis causes large depletions in ambient CO, concentrations. Such CO, depletions affeet photo- synthetic performance, complicating studies of photo- 179
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A portable system for measuring carbon dioxide and water vapour exchange of leaves

plant. Cell atxd Ettvironment (1982) 5, 179-186,
TECHNICAL REPORT
portable system for measuring carbon dioxide andxA/ater vapour exchange of leaves
C, FIELD*, J, A, BERRYt & H, A, MOONEY* *Department of Biological Sciences, StanfordUniversity, Stanford, CA 94305 U,S,A, and fDepartment of Plant Biology, Carnegie Institution ofWashington, Stanfot-d, CA 94305 U,S,A,CIW-DPB Publieation No, 735
Received 5 May 1981; aeeepted for publication 8 July 1981
Abstract. The apparatus deseribed here is a fullyportable, steady-state gas exchange systet-t-i forsimultaneous t-neasurements of the CO, exchatige andtranspiration of single, attaehed leaves. The leafcuvette provides teti-ipetature, humidity, and CO,-concenttation control. The system is suitable foreither surveys or detailed studies of photosyntheticand stomatal responses to environmental variables,Representative data demonstrate the response timecharacteristics of the system and constitute the firstfield evidence of stotnatal behaviour consistent with arecent hypothesis eoncerning the optimum pattern ofstomatal conductance for the maximization of water-use-eflieieney,
li^ev-words: Lepechinia calycina (Benth,) Epl.; Lamiceae; pitchersage; gas exchange system; photosytithesis; ttanspiiatioti;humidity lesponse; waler-use-efficiency.
Introduction
G a s exchange systems for measuring photosynthesisa n d transpitation in the field have tended toemphasize either aecutacy and environmental controla t the eost of portability, or portability andconvenienee, saerifieing environt-nental eonttol andaccuraey. The appat-atus described here incorpotatesrecent technical and conceptual advanees to bringaccuracy and control to a compact, fully portablesystem. The system pt-ovides sufficient control of theleaf envitonment to allow investigations of photo-synthetic and stotnatal responses to tempetature, lightintensity, and vapour pressure deficit. With minormodifieations it can be used iti studies of the CO,coneentt ation dependence of photosynthesis. Carbondioxide and water vapour exehange of single,attaehed leaves ean be measut-ed with an accut-acycomparable to that attained in the sophisticatedrnobile laboratory described by Mooney et al (1971),Though untested in extreme habitats sueh as Death
Correspondetice: Dr Christopher Field, Department of Biology,University of Utah, Salt Lake City, UT 84112, U,S.A.0140-7791/82/0400-0179$02,()0© 1982 Blackwell Scientific Publications
Valley in the summer, the portable system has beefiused to study photosynthetie properties of plantsgrowing under natut-al eonditions in sites throughoutCalifornia, including Death Valley (Seemann, Field &Berry, 1980), Bodega Head (H, A, Mooney,unpublished), and the Sierra Nevada (C, Field,unpublished).
The most aeeurate laboratory gas exchange systemsare open, diffet-ential systems (cf, Ehleringer &Bjorkman, 1977), With these systems, both photo-synthesis and tt-anspit-ation at-e ealeulated fromtneasured flow rates and the CO, and watet--vapourconcentration differences between the air enteringand that leaving the cuvette, A major feature limitingthe portability of open, differential systems isthe requirement for temperature-eontroUed hutnidi-fieation devices and heated gas How lines (Blootn etal., 1980),
Consider an open system with a well-stirredcuvette. If the entering air stream is dry instead ofmoist, the cuvette equilibtates at some relativehumidity determined by the leaf transpiration rateand the air flow rate through the euvette. Decreasingthe air flow tate increases the steady-state telativehutnidity, and increasing the air flow rate deereases it.While the systetn is still fully differential, theiticoming humidity is zero, Beardsell, Jarvis &Davidson (1972) developed a portable porometer fortt-anspiration measutei-tients that is ati open,diffeiential system using only dry air as the inlet gas.Their steady-state porotneter operates with notempetature-controlled components and with a singlehumidity sensor, iti the leaf euvette. Transpiration iscalculated as the simple product of the air flow rateand the vapour density of the air in the euvette,
Porometers of the type described by Beardsell et al,(1972) are tiot suitable for sitnultaneous measute-ments of photosynthesis atid transpiration. At flowrates of dry air low enough to balance tt-anspiration attnoderate or high euvette humidities, photosynthesiscauses large depletions in ambient CO,concentrations. Such CO, depletions affeet photo-synthetic performance, complicating studies of photo-
179
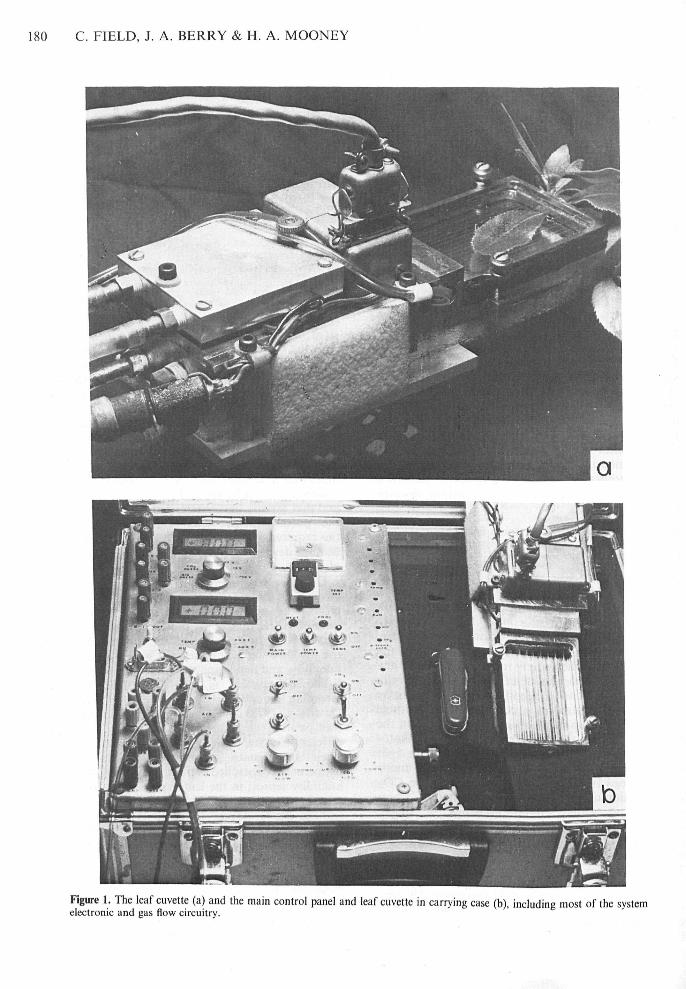
180 C. FIELD, J. A. BERRY & H. A. MOONEY
Figure 1. The leaf cuvette (a) atid the tnain control patiel and leaf cuvette in carrying case (b), including most of the systemelectronic and gas flow circuitry.
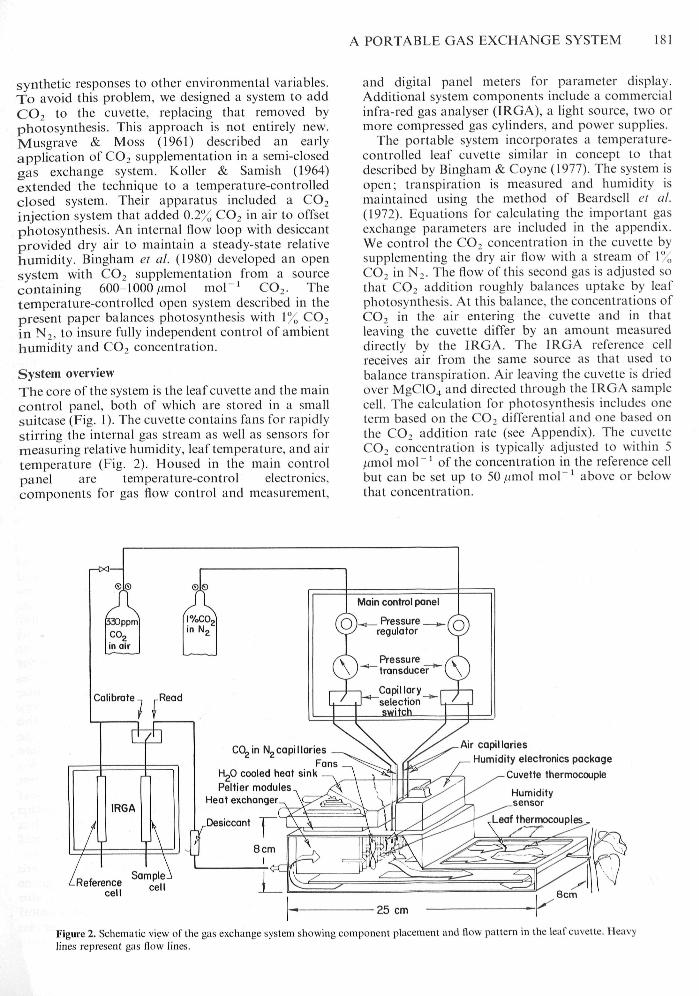
A PORTABLE GAS EXCHANGE SYSTEM 181
synthetic responses to other environmental variables.To avoid this problem, we designed a system to addCO", to the euvette, replaeing that retnoved byphotosynthesis. This appt-oach is not entit-ely new,Musgrave & Moss (1961) described an earlyapplication of CO, supplementation in a semi-closedgas exehange system, Koller & Samish (1964)extended the teehnique to a temperature-controlledclosed system. Their appat-atus included a CO,injection system that added 0,2% CO, in air to offsetphotosynthesis. An internal flow loop with desiccantprovided dry air to tnaintain a steady-state telativehutnidity. Bingham et al, (1980) developed an opensystem with CO-, supplementation from a sourcecontaining 600-1000/nnol mol"' CO,, Thetemperature-eontrolled open system described in thepresent paper balatices photosynthesis with 1% COTin N2, to insure fully itidependent eontrol of ambienthumidity and CO, concentration.
System overviewThe core of the system is the leaf cuvette and the maincontrol panel, both of whieh are stored in a smallsuitcase (̂ Fig, 1), The cuvette eontains fans for rapidlystirring the internal gas streatn as well as sensors formeasuring telative hutnidity, leaf tetnpet-atute, and airtempetature (Fig, 2), Housed in the main eontrolpanel are tempetature-eontrol electronics,components for gas flow control and measurement.
and digital panel meters for parameter display.Additional system eomponents inelude a cotnmetcialinfra-t-ed gas analyser (IRGA), a light souree, two ormore cotnpressed gas eylinders, and power supplies.
The portable system incorporates a temperatut-e-controlled leaf euvette similar in concept to thatdescribed by Bingham & Coyne (1977), The systetn isopen; tt-anspiration is measuted and hutnidity ismaintained using the method of Beardsell et al.(1972), Equations for calculating the impot-tant gasexchange patameters are ineluded in the appendix.We control the CO, eoticentt-ation in the cuvette bysuppletnenting the dt-y air flow with a streatn of 1",,CO, in N,, The flow of this seeond gas is adjusted sothat CO, addition roughly balatices uptake by leafphotosynthesis. At this balance, the eoneentrations ofCO, in the air entering the euvette and in thatleaving the euvette differ by an amount measuredditectly by the IRGA, The IRGA referenee eellreeeives air from the same souree as that used tobalanee transpiration. Air leaving the cuvette is driedover MgClO^ and diteeted through the IRGA satnpleeell. The calculation for photosynthesis ineludes oneterm based on the CO, differential and one based onthe CO, addition rate (see Appendix), The cuvetteCO, concentration is typieally adjusted to within 5/(mol mol" ' of the eoneentration in the referenee cellbut can be set up to 50/nnol mol"' above or belowthat concentration.
Pelfier modulesHeat exchanger
Air capillariesHumidify electronics package
Cuvette thermocouple
25 cm
Figure 2, Schematic view of the gas exchange systetn showing component placei-nent and tlow pattern in the leaf cuvette. Heavylities represent gas flow lines.

182 C, FIELD, J, A, BERRY & H. A, MOONEY
Elow eontrol
To eontrol flow rates, we regulate the pressure behindstainless steel capillaries mounted in the cuvette andattached to the control panel with 1,58 mm LD,polyurethane tubing (Fig, 2), Miniature single-stageregulators (Clippard Instrument Laboratory,Cincinnati, Ohio) allow us to adjust the gas pressuresfrom 0,06 to 0,5 MPa (10 to 80 psi). Pressures aremeasured with electronic pressure transducers(LX1720G, National Semiconductor Corp,, SantaClara, California). Two capillaries (short segments ofstainless steel tubing 0.127 mm to 0,254 mm LD.)calibrated in overlapping flow ranges provide air flowrates from 2 to 40cm-' s ' ' over the above pressurerange. Similar but erimped capillaries yield flows of1% CO2 in N2 between 10 and 200 mm^ s ' withpressures from 0,06 to 0,5 MPa, Because flow ratesfor the COj-enriched gas are always less than 1% ofthe air flow rates, changes in the flow of CO^-enrichedgas exert negligible effects on cuvette humidity.
The compressed air used to balanee transpiration isobtained commercially (Liquid Carbonic Corp,, SanCarlos, California) and is not dried further(maximum vapour pressure = 0,03 kPa), Carbondioxide concentrations in the compressed air aretypically close to 360 /tmol mol ~', One per cent COj inN2 is mixed manometrically from compressed CO2and N-,, The pressure-versus-flow cafibrations for thecapillaries were determined over tetnperatures from10 to 35 C and fitted to regression equations with amaximum error of ±5%, With the capillariesmounted in the cuvette, their temperatures areinsensitive to fluctuations in ambient temperature.The calibrations have proved stable even afterextensive field use.
The euvetteSeveral considerations influenced the shape of theeuvette (Fig, 2), For access to leaves near the groundor elosely spaeed on stems, we required minitnum top-to-bottom depth. To minimize the leaf boundarylayer and eliminate the possibility of stagnant eddies,we chose a continuous-loop internal circulation path.Because many of the system transducers andcontrollers are mounted in the cuvette, we wereforced to include a gas conditioning and monitoringcompartment as an integral part of the leaf chamber.
The cuvette, whieh is approximately 25 x 8 x 8 cmexternally, includes a leaf ehamber with the internaldimensions 6,25 x 8,75 x 1 cm. The internal volume ofthe cuvette is approximately 175 cm^. Except for theglass witidow and the plastic fan bodies, the cuvette isconstructed entirely of nickel-plated brass, providinga large thermal mass with low water absorptivity.
The two cuvette fans (V241L, Micronel U,S,,Hudson, Massachusetts) produce no measurable CO2or ozone (Bingham & Coyne, 1977), As determined
from measured transpiration, the boundary layerresistance for a 3 x 3 cm square of filter paper is0,015stnm ^
The humidity sensor, tnounted just downstreatii ofthe fans, is a Vaisala Humicap (WeathermeasureCorp,, Sacramento, California), a thin-film polymersensor with a linearity of 1% and a tetnperatutesensitivity of only 0,05% RH C~', The electronicspackage mounted above the Humicap is described byBingham et al (1980), Temperature sensitivity of theHumicap electronics package has been minimized bythe use of ultra-low drift resistors and capacitors,Humicap temperature is tneasured with apermanently mounted thermocouple a fewmillimeters from the setisor.
Carbon dioxide in N2 and air enter the cuvette justdownstream of the fans at a point of tnaxitnumturbulenee (Fig, 2), Because air addition ratesrepresent only a small percentage of the totalcirculation around the cuvette, humidity gradientscaused by the progressive hutnidifieation of airflowing across the leaf are negligible. After passingover the leaf, the humidified gas stream passes alongthe bottom of the cuvette. At the back of the cuvette,the gas-flow out of the chamber passes through aMgClO4 drying tube to the IRGA, The internalcirculation is drawn across the temperature-eontrolled heat exchanger to start another loop.
Temperature eontrolThe cuvette, which is the only tetnperature-controlledcomponent of the gas exehange systetn, is heated andcooled by a pair of solid state thermoelectric (Peltier-effect) modules (CPI,4-71-06L, Melcor, Trenton,New Jersey) mounted on top of the cuvette. Waterpumped through an automobile heater-radiator andthen through the heat exchanger above the Peltiermodules provides a heat sink near ambienttemperature. The temperature-control circuit ismodified from Bingham et al (1980), Using the samedeviee to heat and eool minitnizes the energyconsumption for temperature control. While themaximum power consumption for rapid temperaturechanges is approximately 100 W, the steady-statetemperature-control power ranges from 15 to 50 W,
Leaf temperature is measured with two 0,25 mmcoppet-constantan thermocouples. Usually, the leaftemperature is within 1 C of the Hutnicaptemperature, but at vety high light, especially with achamber temperature well below ambient, the leaftnay be up to 4" wanner than the humidity sensor,Thermoeouples on different parts of the leaf aregenerally within 0,3" of each other and neverseparated by tnore than 1,2 C, Under full sunirradiance (with a water filter) at an environmentaltemperature of 25 C, leaf tetnperature can bemaintained within CTC from 15" to 40C,

A PORTABLE GAS EXCHANGE SYSTEM 183
IRGAThe design of the gas exchange system does notrequire the incorpotation of any particular infra-redgas analyser. The instrutnent we have used extensivelyin the field is a standat-d laboratory device, an ADCSeries 225, Mat-k II (Analytical Development Co,Ltd,, Hoddesdon, England), Ease of calibration(Parkinson & Legg, 1978) and independence of linefrequency have made this IRGA a standatd fieldinstrutnent (Oechel & Lawtence, 1979). Accut-atemeasut-ements can be obtained with as little as 20 minof warm-up, and the IRGA is fully stable after onehour.
LightThe systetn can be used with either natur-al or artificialillumination. In situations wher-e constant lightintensity is crucial, we use a 300 W cool-beammedium fioodlight (General Electric Co., Cleveland,Ohio), Intensity is adjusted with neutr-al densityscreens and measut-ed with a Li-Cor Ouantum Pr-obe(Li-Cor Inc., Lincoln, Nebraska),
Physieat and te.sponse eharacteristies
The whole system, except the IRGA and a lamp forartificial illutnination, is power-ed by an 80 amp-hour,marine-type lead-acid battery. Depending on ambientand cuvette tetnper-atur-es, the battery provides 1-2days of set-vice between techargings. In r-emotelocations, a portable generator (EM400, HondaMotor Co., Tokyo) power-s the IRGA and the latnp.The generator can be used to charge the batter-ywhenever the lamp is not in oper-ation. Maxitnutnpower consumption for the system without artificialillutnitiation is 300 W. With artificial illutnination, itis 600 W, or about one-tenth the power tequirernentof the mobile laboratory of Mooney et at, (1971).Wher-e por-tability is a prime eoncern, we use size Ecompressed gas cylinders, keeping the weight of eachcomponent under 30 kg. The entir-e system, completewith tools, cotnpt-essed gas, and fuel for 2 days ofopet-ation, fits easily in a compact station wagon. Itcan be carried by one person in four loads. The timerequir-ed to unpack and assemble the system isapproxitnately 1 hour. Often the first tneasut-etnentsare calculated within an hour of arriving at theexperimental site. All gas exchange pat-atnetet-s at-ecalculated itntnediately in the field with apt-ogt-ammable calculator (HP41C, Hewlett-Paekar-dCorp., Corvallis, Oregon).
The aecut-acy of the system is r-educed vet-y little bythe large fluctuations in atnbient tetnpetatureencountet-ed in the field, Tetnperatur-e dr-ift in nosubsystem is greater than 0,1% per C, keeping driftfrom 10" to 40'C under 5%, At very hightemperatures atid high radiation loads, the system isusually shaded to prevent damage.
Time lags between control action and effect at-egenet-ally short. The Peltier tnodules heat or eool thecuvette at gteater than 2 Ctnin^'. Hutnidity can beadjusted to a steady state in less than 15 s, but thetime required to produce a given change in cuvettehutnidity is limited by the leafs tr-anspiration t-ate.The titne lag in photosynthesis tneasut-etnents andCOj-concentt-ation control depends on the air flowtate through the systetn. Equilibtation titnes rangefr-otn less than 1 min to a maximum of 5-7 tnin.Typically, leaf tetnpet-ature is controlled to +0.1 C,vapour pressure deficit to ± 0,05 kPa, and atnbientCO. to ±5/(mol rnol"'. The option of measuringphotosynthesis with different cotnbinations of nullbalance and open system cotnponents provides auseful check on the calibration of the system.
Representative results
Compari.son wittt an open laboratory system
To check absolute accur-acy and to establish that thefield system does not interfer-e with leaf futiction overextended periods, we simultaneously tneasut-ed gasexchange on two visually-matched leaves on a singlepotted shrub with two eompletely independentsystetns, the portable system and the open labot-atot-ysystetn desct-ibed by Winner & Mooney (1980). Over2 h of observations with utichanging cuvetteeotiditions, the average diffetence between the tesultsfrotn the two systems was 6.3% for net photosynthesisand 7,7% for stomatal conductance. With the fieldsystem, variation in both pat-atnetet-s was less than 2%dut-ing the entite interval.
Leaf responses to relative humidity
By manipulating air flow t-ates, we can maintain theleaf cuvette at any t-elative hutnidity. This capabilityallows us to study leaf responses to the water--vapourcoticentt-ation gt-adient between leaf and air (Arr),Data frotn a r-epresentative experiment highlight thepor-table system's capabilities and the robustness ofthe results to rigot-ous analysis.
The data discussed her-e are based on a singleexperiment in which we tneasur-ed the tnagnitude andtime cout-se of humidity effects on net photosynthesisand stomatal cotiductatice, A leaf of a field-growndrought-deciduous shrub {Lepeehinia catycitta(Benth.) Epl.) was held at light satutation atid 20.5 Cthrough the experiment. Ambient COj concentrationwas 355/(tnol mol"'. After the leaf equilibt-ated atone vapour eoncentration gradient, cuvettehumidity was deer-eased by 10",, over an intet-val ofabout 5 s. Each hutnidity step represented approxi-mately a 2,5 mtnol tiiol"' incr-ease in vapourcoticentration gt-adient. Net photosynthesis andstomatal conductance wer-e measured every minuteafter eaeh humidity step until these par-atnetersStabilized at new values. Between steps, cuvette

184 C. FIELD, J. A. BERRY & H. A. MOONEY
- 600
-550
-500
-450
4 0 0
-300
-290
- 280
- 270
800 -
- 260oo
1 9 5-10 -5 0 25 30
J25O5 10 15
Time ( min)Figure 3. Time eourse of changes in stomatal eonductance, netphotosynthesis and CO2 internal following a step change in watervapour concentration gradient (Aiv) at time 0 for E. calycitta. Thestep in Aw represents a humidity ehange from 87% to 76% at20.5 C.
humidity was held to within 1% of the desired values.The units used here for vapour concentration gradientand stomatal conductance are those proposed byCowan (1977).
Both stomatal conductance and net photosynthesisresponded quickly to a decrease in humidity (Fig. 3).With a vapour concentration gradient step from 5 to7.5 mmol m o l ' , conductance began to decrease inless than 1 min. The initial increase in photosynthesisis an artifact due to a flushing time lag between theleaf chamber and the IRGA. Because the humiditysensor is in the cuvette, the time lag for conductancemeasurements is very small. With vapourconcentration gradient steps of 2.5 mmol mol" '(0.25 kPa, 10% RH at 20.5X), stomatal conductanceand net photosynthesis stabilized in about 15 min.The pattern of changes in net photosynthesisparalleled changes in CO2 internal.
Using only equilibrium values measured after netphotosynthesis and stomatal conductance hadstabilized, the results of this experiment (Fig. 4)qualitatively parallel those from studies of otherVPD-sensitive species (Davies & Kozlowski, 1974;Warrit, Landsberg & Thorpe, 1980). Over a vapourconcentration gradient range from 4 to 18 mmolm o P ' , stomatal conductance decreased by 71%, from790 to 230 mmol m~^ s" ' . Over this range, CO2internal declined by 8%, and net photosynthesisdecreased by 30%.
Assuming a COj-compensation point of 80
600-
400 -
200 -
{i 20 -
•r 16 -" E
"o
• I l 2 -
QJ
"a.
<\l
tJ /I _
Net
-1
0
1 1
1 i
-300
280 !_o
-260
240 -
10( mmoi moi"
i5_ 22020
Figure 4. Stomatal conductance, net photosynthesis, and CO,intemat versus water-vapour eoncentration gradient (Aw), for E.calycltia. The maximum stomatat conductance is equivatent to19.6 mm s '.
mol ' and a constant mesophyll conductance, themodel of Gaastra (1959) as modified by Ehleringer &Miller (1975) pt edicts net photosynthetic tates thatclosely parallel observed values (Fig. 5). The observedrelationship is similar to the tnobile laboratory resultsof Schulze et at. (1972) and serves as a functionalcheck on the portable system calibrations. Theobserved pattern demonsttates that over a wide ratigeof flow rates and humidities, the system operateswithout the introduction of artifact.
Using the calculation procedute outlined byFarquhar, Schulze & Kuppers (1980), the hutnidityresponse data can be teinterpreted in the context ofoptimizing stomatal conductatice for themaxitnization of water-use-efficiency. This highly sim-plified calculation procedure is appropriate only withcuvette conditions where the boundary layer resistanceis small and the leaf can be held at the temperatureoptimum for photosynthesis. Through the hutnidity
Figure 5. Predieted (-
200 400 600 800Conductance (mmol m"^ s"')
. ) and measured (• • •) net photosynthesisversus stomatal conductatice for L. calycitta. Predicted values arebased on the model of Gaastra (1959) as modified by Ehleringer &Miller (1975). assuming a COj compensation point of 80/imolmol" ' and a mesophyll conductance of 99.3 mmol m " ^ s ' ' .
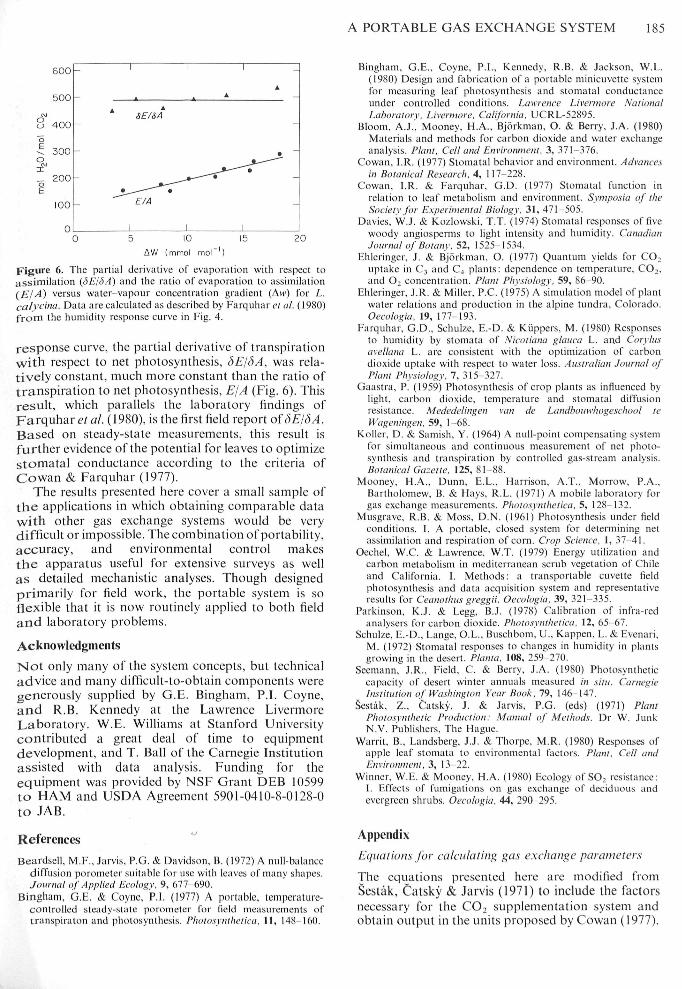
A PORTABLE GAS EXCHANGE SYSTEM 185
6 0 0 -
5 0 0 -
4 0 0 -
o:£•_ 200 -
100 -
300 -
AW (mmol mol" )
Figure 6, The partial derivative of evaporation with respect toassimilation (<yE/<)A) and the ratio of evaporation to assimilation(ElA) versus water-vapour eoneentration gradient (Aw) for L.calvcitia. Data are calculated as described by Farquhar et al. (1980)from the humidity response curve in Fig. 4.
respotise curve, the partial derivative of transpirationwith respect to net photosynthesis, dEj^A, was rela-tively constant, tnuch tnore constant than the tatio oftranspiration to net photosynthesis, EjA (Fig, 6), Thisresult, whieh parallels the laboratory findings ofFarquhar c'/ al (1980), is the fitst field teport of 6EI6A.Based on steady-state tneasutetnents, this result isfurther evidetice of the potential for leaves to optitiiizestomatal conductance aecotding to the criteria ofCowan & Farquhar (1977),
The tesults ptesented hete cover a stnall sample ofthe applieations in whieh obtaining cotnparable datawith other gas exchange systems would be verydifficult or impossible. The eotnbination of portability,accuracy, and environmental conttol makesthe apparatus useful for extensive sutveys as wellas detailed mechanistic analyses. Though designedprimarily for field work, the portable systetn is soflexible that it is now routinely appfied to both fieldand laboratory probletns.
Acknowledgments
Not only tnany of the systetn concepts, but technicalad'vice and many difficult-to-obtain cotnponents wetegenerously supplied by G,E, Bingham, P,I, Coyne,and R,B. Kennedy at the Lawtence LivertnoreLaboratoty. W.E. Williatns at Stanford Universitycontributed a great deal of titne to equiptnentdevelopment, and T. Ball of the Cartiegie Institutionassisted with data analysis. Funding for theequiptnent was provided by NSF Grant DEB 10599to HAM and USDA Agreement 5901-0410-8-0128-0to JAB.
References
Beardsell, M.F., Jarvis, P.G. & Davidson, B. (1972) A null-balancediffusion porometer suitable for use with leaves of many shapes.Journal of Applied Eeology. 9, 677-690.
Bingham, G,E. & Coyne, P.I. (1977) A portable, temperature-controlled steady-state porometer for field measurements oftranspiraton and photosynthesis. Photosynthetiea. II , 148 160.
Bingham, G.E., Coyne, P.L, Kentiedy, R.B. & Jaekson, W.L.(1980) Design and fabrication of a portable minicuvette systemfor measuring leaf photosynthesis and stomatal conductanceunder controlled conditions. Eawrenee Livertnore NatiottalLaboratory. Livettiiore. California, UCRL-52895.
Bloom, A.J., Mooney, H.A., Bjorkman, O. & Berry, J.A. (1980)Materials and methods for earbon dioxide and water exehangeanalysis. Plant. Cell and Environtnetu. 3, 371-376.
Cowan, I.R. (1977) Stomatal behavior and environment. Adyaneesin Botanieai Researeh, 4, 117-228.
Cowan, I.R. & Farquhar, G.D. (1977) Stomatal function inrelation to leaf metabolism and environment. Symposia of theSoeiety for Experimental Biology. 31, 471-505.
Davies. W.J. & Kozlowski, T.T. (1974) Stomatal responses of tivewoody angiospemis to light intensity and humidity. CatiadianJournal of Botany. 52, 1525-1534.
Ehleringer, J. & Bjorkman, O. (1977) Quantum yields for CO.uptake in C3 and C4 plants: dependence on temperature, CO,,and O; concentration. Platit Physiology. 59, 86-90.
Ehleringer, J.R. & Miller, P.C. (1975) A simulation model of plantwater relations and production in the alpine tundra, Colorado.Oeeologia. 19, 177 193.
Farquhar, G.D., Schulze, E.-D. & Kuppeis, M. (1980) Responsesto humidity by stomata of Nieotiana glauea L. and Corylusavettana L. are consistent with the optimization of earbondioxide uptake with respeet to water loss. Australian Jourtial ofPlatU Physiology. 7, 315 327.
Gaastra, P. (1959) Photosynthesis of erop plants as influenced bylight, carbon dioxide, temperature and stomatal diffusionresistanee. Mededelingen van de Eandbouwhogesehool teWagetiingen, 59, 1-68.
Roller, D. & Samish, Y. (1964) A null-point eoiiipensating systemfor simultaneous and continuous measurement of net photo-synthesis and transpiration by controlled gas-stream analysis.Botanieai Gazette. 125,81 88.
Mooney, H.A., Dunn, E.L., Harrison, A.T., Morrow, P,A.,Bartholomew, B. & Hays, R.L. (1971) A mobile laboratory forgas exehange measurements. Photosynthetiea. 5, 128 132.
Musgrave, R.B. & Moss, D.N. (1961) Photosynthesis under fieldconditions. I. A portable, closed system lor determining netassimilation and respiration of corn. Crop Seienee. 1, 37 41.
Oechel, W.C. & Lawrenee, W.T. (1979) Energy utilization andcarbon metabolism in mediterranean serub vegetation of Chileand California. I. Methods: a transportable euvette fieldphotosynthesis and data acquisition system and representativeresults for Ceanothus greggii. Oeeologia. 39, 321 335.
Parkinson, K.J. & Legg, B.J. (1978) Calibration of infra-redanalysers for carbon dioxide. Photosynthetiea. 12, 65-67.
Schulze, E.-D., Lange, O.L., Busehbom, U., Kappen, L. & Evenari,M. (1972) Stomatal responses to ehanges in humidity in plantsgrowing in the desert. Planta. 108, 259 270.
Seemann, J.R., Field, C. & Berry, J.A. (1980) Photosynthetieeapaeity of desert winter annuals measured iti situ. CarnegieInstitution of Washington Year Book. 19, 146-147.
Sestak, Z., Catsky, J. & Jarvis, P.G. (eds) (1971) PlatilPhotosynthetic Production: Manual of Methods. Dr W. JunkN.V. Publishers, The Hague.
Warrit, B., Landsberg, J.J. & Thorpe, M.R. (1980) Responses ofapple leaf stomata to environmental factors. Plant. Cell andEtivironnient. 3, 13-22.
Winner, W.E. & Mooney, H.A. (1980) Eeology of SO, resistance:I. Effects of fumigations on gas exchange of deciduous andevergreen shrubs. Oeeologia. 44, 290-295.
Appendix
Eqttations for calculating gas exehatige parattieters
T h e e q u a t i o n s p t e s e n t e d h e t e a t e tnodified f totnSestak, Catsky & Jarvis (1971) to include the factorsnecessaty for the CO2 suppletnentation systetn andobtain output in the units proposed by Cowan (1977),

186 C. FIELD, J. A. BERRY & H. A. MOONEY
The rate of transpiration,given by
£ = (w • E •
I m
tn) (1)where w (mmol H2O mol ' air) is the mole fraction ofwater vapour in the ambient air, F (cm-* s"') is thetemperature- and pressure-corrected flow tate of dryair into the cuvette, .v (cm') is the leaf surface area andm (1) is the ideal gas tnolar volume at the cuvettetemperature and pressure. The air flow calculationmay require two types of temperature correction, oneto account for physical changes in capillary propertiesand one to account for Charles' law. Cortections forthe latter temperature effect and the effect of pressurecancel the temperature and pressute corrections forthe molar volume. The factor of 10 corrects the units.IV is calculated from the tetnperature, the humidityand the atmospheric pressure as follows:
VV = (r • Vh • 10^)//7 (2)
where r is the relative humidity as a fraction of 1, v,,(kPa) is the saturated vapour pressure at the Humicaptemperature, and p (kPa) is the attnospheric pressure.The factor of 10̂ corrects the units.
Leaf conductance, the sum of stomatal andcuticular conductance, g (mtnol m "^ s" '), is given by
g = {E•W)|^w (3)
where E is transpiration and Aw (mmol H2O mol~'air) is the water-vapour concentration gradientbetween the leaf intercellular spaces and the cuvetteair. Aw is given by
where v, (kPa) is the saturated vapour pressure atleaf temperature; p and M' have already been defined.Except in applications requiring very high precision(e.g. the calculation of ("lEj^A), the boundary layer
resistance is ignored in the calculation of g.Correcting for the boundary layer resistance results innegligible changes in g unless g is very large. Thecorrection is not routinely included because itdepends on leaf size and shape.
Photosynthesis, A (/(mol CO, m" ^ s~'), is given by
A=[{E • ^c)|W /))] m) (5)
where Ac (/(tnol mol ') is the differential reading onthe IRGA, J (mtn^ s"') is the pressure- andtemperatut e-corrected flow tate of COi-enriched gasinto the cuvette, and /; is the concentration of the CO2in the CO2-enriched gas as a ftaction of \. E, s, and tnhave already been defined. Again, most of thepressure and temperatute corrections for the fiowrates and the molar volume cancel each other. Thefactors of lO"* and lO"*̂ cortect the units.
The carbon dioxide concenttation in the leafintercellular spaces, q (/(tnol tnol"') is given by
where c.^ (/(mol moP ' ) is the CO2 concentration inthe air leaving the cuvette. The factor of lO'̂ correctsthe units, and 1.56 is the tatio of the tnoleculardiffusivities of water vapour atid CO2 in air. For thiscalculation, we assume that cuticular conductance isnegligible and that g represents only stomatalconductance, c, is given by
where c, (/(mol mol" ') is the CO2 concentration in thedry air cylinder and all other cariables have alreadybeen defined. The effect of the calculation for c^ is tocorrect the CO2 concentration measured at the IRGAfor the removal of the water vapour by the desiccant.In the absence of a desiccant,
c.^ = c.+Ac. (8)





















