
A novel CD44 antibody identifies an epitope that is aberrantly expressed on acute lymphoblastic...
9
Immunology and Cell Biology (2003) 81, 311–319 Research Article A novel CD44 antibody identifies an epitope that is aberrantly expressed on acute lymphoblastic leukaemia cells LINDA J BENDALL, 1 ALEXANDER JAMES, 1 ANDREW ZANNETTINO, 2 PAUL J SIMMONS, 3 DAVID J GOTTLIEB 1 and KENNETH F BRADSTOCK 1 1 Westmead Institute for Cancer Research, Westmead Millennium Institute, University of Sydney, Westmead Hospital, Westmead, New South Wales, 2 Institute for Medical and Veterinary Research, Adelaide, South Australia and 3 Peter MacCallum Institute, Melbourne, Victoria, Australia Summary Previous studies have shown that the antibody 7H9D6 identifies CD44, a glycoprotein receptor for hyaluronic acid. 7H9D6 recognizes an epitope of CD44 that is not always present on CD44 molecules. The 7H9D6 antibody bound to the hyaluronic acid binding domain of CD44 and inhibited cell adhesion to immobilized hyaluronic acid. However, the expression of the 7H9D6 epitope was not sufficient for hyaluronic acid binding. Immunofluorescent staining with 7H9D6 revealed a punctate surface staining pattern, suggesting that CD44 molecules recognized by 7H9D6 are located in clusters on the cell surface. In contrast, other CD44 antibodies produced a uniform staining pattern. Early bone marrow B cells were negative for 7H9D6 but reactive with other CD44 monoclonal antibodies. In contrast, leukaemic cells from 65% of patients (28 of 43) with B lineage acute lymphoblastic leukaemia bound 7H9D6. Patients expressing the 7H9D6 epitope on their leukaemic cells had an increased risk of death (HR 3.5 95% CI 1.1–10.9, P = 0.029) and of disease relapse (HR 3.2 95% CI 1.2–8.5, P = 0.017) when corrected for white cell count. This antibody may be useful for the detection of residual disease in B lineage acute lymphoblastic leukaemia and as a prognostic indicator and for the study of CD44 function. Key words: acute lymphoblastic leukaemia, adhesion, CD44, hyaluronic acid. Introduction CD44 is a widely expressed transmembrane glycoprotein receptor for the polysaccharide, hyaluronic acid. 1 In addition CD44 binds many other extracellular matrix components including fibronectin, collagen, laminin, osteopontin, sergly- cin and the cellular ligands E and L-selectin. 2–4 CD44H has been reported to modulate a variety of cellular functions including adhesion, proliferation, presentation of growth factors, apoptosis, cell migration and metastatic behaviour. 2 Within the haematopoietic system CD44 plays a role in lymphocyte extravasation across high endothelium and homing to periph- eral lymphoid organs. 5–8 It regulates haematopoietic progeni- tor cell distribution and mobilization from the bone marrow. 9 It is also believed to mediate the adhesion of haematopoietic cells to stromal components, an interaction important for the survival and differentiation of haematopoietic progenitors in vitro. 10 Stromal contact is particularly important for normal and leukaemic B-lymphoid progenitors. 11 A distinctive feature of CD44 is that it exists in a large range of isoforms that are encoded by a single gene. Many isoforms, at least 18, are the result of alternative mRNA splicing, with 12 of the 19 exons having the potential for being alternatively spliced. 12 On haemopoietic cells, the pre- dominant form is the smallest of 80–90 kDa (CD44H) that lacks all variant exons. 13–15 However, CD44 is also subject to a range of post-translational modifications. Within the con- stant region of CD44H there are several positions for the attachment of N-linked carbohydrate, and a membrane proxi- mal serine/threonine-rich mucin-like region that provides multiple sites for O-linked carbohydrate. 12 Although there are four potential sites for the attachment of glycosaminoglycans in CD44H, it appears that the majority of heparan and chondroitin sulfate modifications occur within the alterna- tively spliced variable exon v3. 16–19 This variant can be detected in normal and malignant haemopoietic progenitors. 20 Binding to hyaluronic acid appears to be dependent on phosphorylation of Ser291 in the cytoplasmic tail and can also be modulated by the formation of dimers through the intramembrane cysteine. 21–23 CD44 is expressed by haematopoietic progenitors and its expression is temporally regulated during differentiation. 24 Expression of CD44 is also up-regulated following activation of a number of cell types. In B lymphoid progenitors, CD44 expression increases during the process of differentiation. 25 Despite expression of CD44, mature B cells do not bind hyaluronic acid unless previously activated by antigen stimu- lation. 26 Like their normal counterparts, the majority of cases of B lineage acute lymphoblastic leukaemia (ALL) express CD44. However, leukaemic cells from a significant propor- tion of cases of ALL over or under express CD44 in compar- ison to normal B cell progenitors. This has made the examination of CD44 in conjunction with other phenotypic markers useful for the detection of minimal residual disease in ALL. 27,28 Correspondence: Dr Linda J Bendall, Westmead Institute for Cancer Research, Westmead Millennium Institute, Westmead 2145, NSW, Australia. Email: [email protected] Received 3 March 2003; accepted 11 April 2003.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of A novel CD44 antibody identifies an epitope that is aberrantly expressed on acute lymphoblastic...

Immunology and Cell Biology
(2003)
81
, 311–319
Research Article
A novel CD44 antibody identifies an epitope that is aberrantly expressed on acute lymphoblastic leukaemia cells
L I N D A J B E N D A L L ,
1
A L E X A N D E R J A M E S ,
1
A N D R E W Z A N N E T T I N O ,
2
P A U L J S I M M O N S ,
3
D A V I D J G O T T L I E B
1
a n d K E N N E T H F B R A D S T O C K
1
1
Westmead Institute for Cancer Research, Westmead Millennium Institute, University of Sydney, Westmead Hospital, Westmead, New South Wales,
2
Institute for Medical and Veterinary Research, Adelaide, South Australia and
3
Peter MacCallum Institute, Melbourne, Victoria, Australia
Summary
Previous studies have shown that the antibody 7H9D6 identifies CD44, a glycoprotein receptor forhyaluronic acid. 7H9D6 recognizes an epitope of CD44 that is not always present on CD44 molecules. The 7H9D6antibody bound to the hyaluronic acid binding domain of CD44 and inhibited cell adhesion to immobilizedhyaluronic acid. However, the expression of the 7H9D6 epitope was not sufficient for hyaluronic acid binding.Immunofluorescent staining with 7H9D6 revealed a punctate surface staining pattern, suggesting that CD44molecules recognized by 7H9D6 are located in clusters on the cell surface. In contrast, other CD44 antibodiesproduced a uniform staining pattern. Early bone marrow B cells were negative for 7H9D6 but reactive with otherCD44 monoclonal antibodies. In contrast, leukaemic cells from 65% of patients (28 of 43) with B lineage acutelymphoblastic leukaemia bound 7H9D6. Patients expressing the 7H9D6 epitope on their leukaemic cells had anincreased risk of death (HR 3.5 95% CI 1.1–10.9,
P
= 0.029) and of disease relapse (HR 3.2 95% CI 1.2–8.5,
P
= 0.017) when corrected for white cell count. This antibody may be useful for the detection of residual disease inB lineage acute lymphoblastic leukaemia and as a prognostic indicator and for the study of CD44 function.
Key words
:
acute lymphoblastic leukaemia, adhesion, CD44, hyaluronic acid.
Introduction
CD44 is a widely expressed transmembrane glycoproteinreceptor for the polysaccharide, hyaluronic acid.
1
In additionCD44 binds many other extracellular matrix componentsincluding fibronectin, collagen, laminin, osteopontin, sergly-cin and the cellular ligands E and L-selectin.
2–4
CD44H hasbeen reported to modulate a variety of cellular functionsincluding adhesion, proliferation, presentation of growth factors,apoptosis, cell migration and metastatic behaviour.
2
Withinthe haematopoietic system CD44 plays a role in lymphocyteextravasation across high endothelium and homing to periph-eral lymphoid organs.
5–8
It regulates haematopoietic progeni-tor cell distribution and mobilization from the bone marrow.
9
It is also believed to mediate the adhesion of haematopoieticcells to stromal components, an interaction important for thesurvival and differentiation of haematopoietic progenitors
in vitro
.
10
Stromal contact is particularly important for normaland leukaemic B-lymphoid progenitors.
11
A distinctive feature of CD44 is that it exists in a largerange of isoforms that are encoded by a single gene. Manyisoforms, at least 18, are the result of alternative mRNAsplicing, with 12 of the 19 exons having the potential forbeing alternatively spliced.
12
On haemopoietic cells, the pre-dominant form is the smallest of 80–90 kDa (CD44H) that
lacks all variant exons.
13–15
However, CD44 is also subject toa range of post-translational modifications. Within the con-stant region of CD44H there are several positions for theattachment of N-linked carbohydrate, and a membrane proxi-mal serine/threonine-rich mucin-like region that providesmultiple sites for O-linked carbohydrate.
12
Although there arefour potential sites for the attachment of glycosaminoglycansin CD44H, it appears that the majority of heparan andchondroitin sulfate modifications occur within the alterna-tively spliced variable exon v3.
16–19
This variant can bedetected in normal and malignant haemopoietic progenitors.
20
Binding to hyaluronic acid appears to be dependent onphosphorylation of Ser291 in the cytoplasmic tail and canalso be modulated by the formation of dimers through theintramembrane cysteine.
21–23
CD44 is expressed by haematopoietic progenitors and itsexpression is temporally regulated during differentiation.
24
Expression of CD44 is also up-regulated following activationof a number of cell types. In B lymphoid progenitors, CD44expression increases during the process of differentiation.
25
Despite expression of CD44, mature B cells do not bindhyaluronic acid unless previously activated by antigen stimu-lation.
26
Like their normal counterparts, the majority of casesof B lineage acute lymphoblastic leukaemia (ALL) expressCD44. However, leukaemic cells from a significant propor-tion of cases of ALL over or under express CD44 in compar-ison to normal B cell progenitors. This has made theexamination of CD44 in conjunction with other phenotypicmarkers useful for the detection of minimal residual diseasein ALL.
27,28
Correspondence: Dr Linda J Bendall, Westmead Institute forCancer Research, Westmead Millennium Institute, Westmead 2145,NSW, Australia. Email: [email protected]
Received 3 March 2003; accepted 11 April 2003.

312
LJ Bendall
et al
.
We have characterized a monoclonal antibody, 7H9D6,that was raised against bone marrow stromal cells. 7H9D6recognizes a subset of CD44 molecules on the cell surface. Asa result of recognizing only a fraction of CD44 molecules ithas an unusual reactivity profile on many haematopoieticcells. This is most pronounced on early B cells, which are notlabelled by 7H9D6 but are clearly CD44 positive using otherCD44 monoclonal antibodies. In addition, 7H9D6 binds allcases of adult ALL examined and 57% of childhood ALL,making it a potentially useful agent for distinguishing normalfrom leukaemic cells.
Methods
Cells and cell culture
The following cell lines were cultured in RPMI medium containing10% fetal calf serum (FCS): U937, HL60, CEM, CEMT4 and Daudiin a humidified atmosphere containing 5% CO
2
in air at 37
°
C. KG1cells were grown in RPMI containing 20% FCS. CEMT4 were kindlyprovided by Dr G Lynch (Westmead Millennium Institute, Australia)and all other cell lines were obtained from the American TissueCulture Collection (ATCC). Normal peripheral blood and bonemarrow samples were obtained from patients and normal donorsfollowing informed consent within institutional ethics committeeguidelines. Leukaemic patients were from the New Children’s Hospi-tal Westmead, and Westmead Hospital. In addition, cells from 12cases of ALL were obtained from Dr Richard Lock, the Children’sCancer Institute Australia. These samples had been expanded byxenograft in NOD/SCID mice, a procedure that has been previouslyshown not to significantly modify the immunophenotype of thesample.
29
Normal peripheral blood was collected from volunteers intoheparinized tubes. Bone marrow or peripheral blood samples werecollected from 43 patients with acute lymphoblastic leukaemia. Themononuclear layer was collected from the interface of a FicollHypaque density separation as previously described.
30
Granulocyteswere collected from the pellet and the red cells removed by hypotoniclysis in 150 mmol/L NH
4
Cl, 10 mmol/L KHCO
3
and 0.01 mmol/LEDTA pH 7.5 for 10 mins at room temperature (no loss of viability ofgranulocytes was observed as a result of hypotonic lysis of red cells).Red blood cells were also collected from the pellet after centrifuga-tion on Ficoll. All cells were washed twice in phosphate bufferedsaline (PBS) prior to use. Normal CD34 + progenitors were isolatedfrom the bone marrow mononuclear fraction by fluorescence acti-vated cell sorting after labelling with HPCA-2-PE or FITC. Whereindicated cells were cultured in the presence of PMA (10 nmol/L).
Antibodies
Purified Hermes 1 and Hermes 3 were a kind gift from Dr E Butcher(Stanford, CA, USA). 7H9D6 and H9H11 antibodies have beenpreviously described as CD44 antibodies.
31
H9H11 and 7H9D6 wereraised against human bone marrow stroma. The cDNA responsiblefor the 7H9D6 antigen was cloned from a bone marrow stromalcDNA library expressed in FDC-P1 cells.
31
Re-expression of thecDNA was performed to confirm that CD44 was responsible for the7H9D6 and H9H11 antigens. The full length cDNA has beensequenced and found to encode CD44H. The cDNA does not containcoding sequences for variant exons nor does it contain mutationswhen compared to the published sequence.
13
H9H11 has beenmapped to epitope 3 (Hermes 3 like) of CD44 by crossblockingstudies (Bendall, unpubl. obss). The following antibodies werepurchased from Becton Dickinson (Mountainview, CA, USA) –
Leu12-PE (anti-CD19), Leu4-PCP (anti-CD3), NCAM 16.2-PE (anti-CD56), Leu M3-PE (anti-CD14), HPCA-2-PE and HPCA-2-FITC(anti-CD34), from DAKO (Carpenteria, CA, USA) HD37-TRIC(anti-CD19), streptavidin-PE and streptavidin-TRIC and from Bio-genesis (Poole, England) F10-44-2 (anti-CD44). Anti-CD38 (OKT10)and anti-HLA-DR (WM2) were purified from culture supernatant orascites fluid and biotinylated in this laboratory.
Labelling for flow cytometry
Cells were labelled with directly conjugated, biotinylated or unconju-gated antibodies in the presence of 10% heat inactivated humanserum at room temperature (RT) for 10 min and washed once withPBS containing 0.1% bovine serum albumin (BSA) and 0.1% sodiumazide. If required, the second layer, either streptavidin conjugated tophycoerythrin or tricolor or sheep antimouse fluorescein (SAMF)(Amrad, Melbourne, Australia) was then added and the cells incu-bated for a further 10 min at RT and washed again. 7H9D6 andH9H11 were purified from mouse ascites or hybridoma culturesupernatant and conjugated to fluorescein isothiocyanate by estab-lished methods.
32
Purified H9H11, 7H9D6 and Hermes 1 wereconjugated to biotin. Briefly, 40
µ
L of 0.1 M biotinamidocaproateN-hydroxysuccinimide ester (Sigma, Sydney Australia) in dimethylformamide was added to 1 mL of purified antibody (1 mg/mL) inPBS and incubated at RT for 1 h. Uncoupled biotinamidocaproateN-hydroxysuccinimide ester was separated from labelled antibody ona PD10 column (Pharmacia Biotech, Sydney, Australia).
Quantification of antigen sites
The number of binding sites for 7H9D6 and H9H11 was determinedby flow cytometry. Cells were labelled with saturating concentrationsof FITC-conjugated antibodies. Simply Cellular goat antimousebeads (Becton Dickinson) were similarly labelled with saturatingconcentrations of antibodies. Standard fluorescence beads (BectonDickinson) were used to calculate the effective fluorescein/proteinratio for each antibody from the molecules of equivalent solublefluorochrome (MESF) of the labelled beads. This value was then usedto calculate the number of antibody binding sites on cells accordingto the manufacturer’s instructions. Control cells labelled with anirrelevant control antibody or cells negative for CD44 labelled withCD44 antibodies were analysed to allow for correction due to non-specific binding of antibody or autofluorescence.
Immunofluorescent microscopy
Cells were washed in PBS, cytospun onto slides, air dried and fixedin 3.7% formaldehyde at 4
°
C for 15 min. Following washing, theslides were blocked with 10% FCS in PBS overnight at 4
°
C andlabelled with antibodies diluted in 10% FCS in PBS for 1 h at RT.Following washes in PBS, cells were incubated for 1 h at RT withsheep antimouse FITC (1 in 10 dilution) or avidin Texas Red (1 in250 dilution) as second layers for unconjugated or biotinylatedantibodies, respectively. Sheep antimouse FITC (SAMF) was dilutedin PBS containing 10% FCS and avidin Texas Red in 150 mmol/LNaCl, 10 mmol/L NaHCO
3
and 0.5% BSA pH 8.2. Labelled cellswere viewed under an Olympus fluorescent microscope.
In capping experiments, cells were labelled with 7H9D6 orcontrol culture supernatant at 4
°
C for 30 min and washed twice inPBS. Sheep anti-mouse fluorescein (SAMF) was added at a 1 in 100dilution and cells incubated at 4
°
C for 10 min, then at 37
°
C for 1–3 h.Cells were washed once in PBS and fixed in 3.7% formaldehydeat 4
°
C for 15 min. Cells were incubated with H9H11-biotin,

CD44 antibody recognizes malignant pre-B ALL
313
7H9D6-biotin or control antibody at 4
°
C for 30 min, washed in PBSand labelled with Texas Red avidin as described above. Cells werecytospun onto slides and viewed under an Olympus fluorescentmicroscope.
Adhesion assays
Adhesion assays were performed as previously described.
30
Briefly,hyaluronic acid was coated onto 96-well plates at 100
µ
g/mL over-night at 4
°
C. After four washes in PBS the wells were blocked with1% BSA in PBS at 37
°
C for 2 h. Plates were washed twice with PBSprior to use. Cells were labelled with sodium
51
chromate and plated at5
×
10
4
cells/mL in RPMI medium containing 1% BSA. Plates wereincubated at 37
°
C in a humidified incubator for the indicated timeperiods and nonadherent cells removed by washing twice with RPMImedium. Adherent cells were lysed in 2% Triton-X 100 and lysatescounted in a gamma counter. Where indicated F10-44-2, a CD44activating antibody, was added at a final concentration of 1
µ
g/mL,20 min prior to plating cells in adhesion assays.
Statistics
Kaplan-Meier curves were used to illustrate the event free and overallsurvival distributions. Cox proportional hazard models were used totest for the association between 7H9D6 expression and event free oroverall survival both unadjusted and adjusted for white cell count(WCC). The hazards ratios (HR) and their 95% confidence intervals(CI) were used to quantify the degree of association.
Results
7H9D6 recognizes the hyaluronic acid binding domain of CD44
CD44 antibodies bind to one of three well-defined physicallydistinct regions on CD44. Some but not all antibodies thatrecognize region 1 (Hermes 2-like) inhibit T cell rosetting,antibodies that recognize region 2 (Hermes 1-like) blockadhesion to hyaluronic acid and antibodies that recognizeregion 3 (Hermes 3-like) block lymphocyte binding tomucosal high endothelial venules.
33
Excess unlabelled anti-bodies directed against the hyaluronic acid binding domain(Hermes 1, 3F12), but not other CD44 antibodies, blocked thebinding of FITC conjugated 7H9D6, demonstrating that7H9D6 recognizes region 2 (Fig. 1a). Interestingly, excessunlabelled 7H9D6 only partially blocked the binding ofHermes 1, suggesting that some CD44 molecules that bindHermes-1 do not bind 7H9D6 or that 7H9D6 has a loweraffinity for CD44 than Hermes-1 (Fig. 1b). Since 7H9D6binding can be cross blocked by antibodies to region 2, thehyaluronic acid binding domain of CD44, we investigated theability of 7H9D6 to inhibit the adhesion of the haematopoieticprogenitor cell line KG1 to hyaluronic acid. 7H9D6 was justas effective as the Hermes 1 antibody at inhibiting theadhesion of KG1 cells to immobilized hyaluronic acid(Fig. 1c). In contrast, Hermes 3 and H9H11 had little effecton binding of KG1 to hyaluronic acid as expected (data notshown).
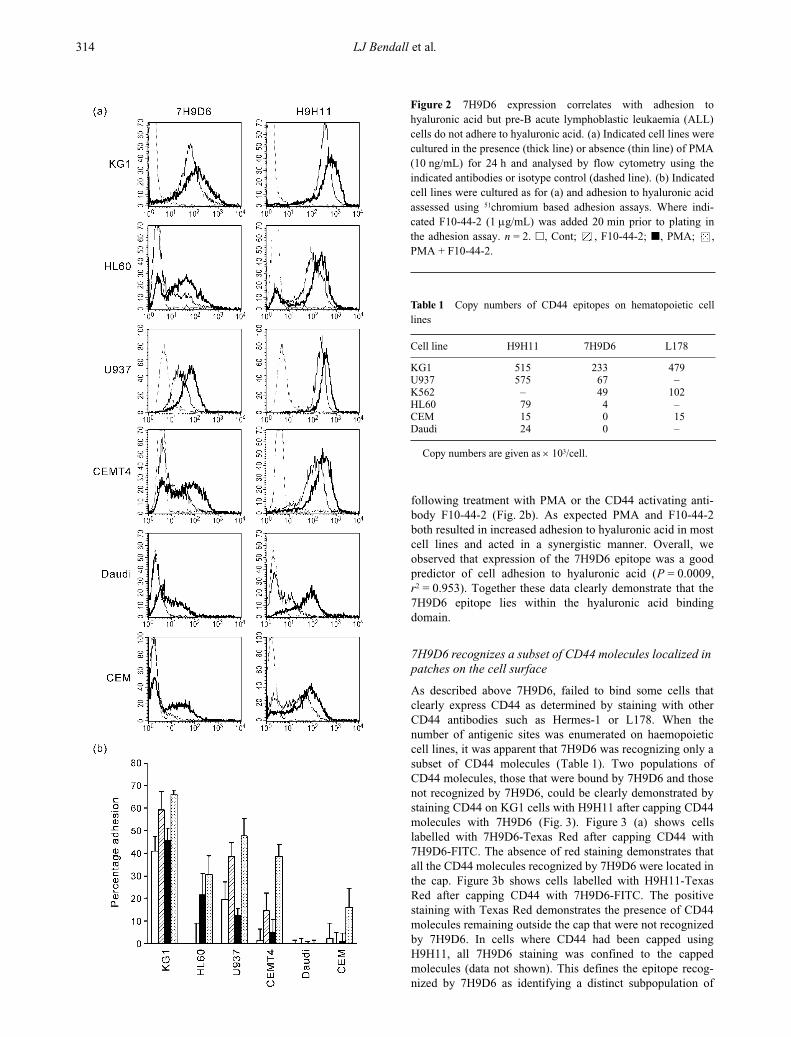
We next examined a range of haemopoietic cell linesincluding HL60, KG1, U937, CEM, Daudi and CEMT4 fortheir expression of the 7H9D6 epitope when unstimulated andafter PMA activation for 24 h. All cell lines expressed CD44
as determined by H9H11 binding although Daudi onlyexpressed very low levels (Fig. 2a, thin line). In contrast,CEM and Daudi did not bind 7H9D6 and HL60 and CEMT4expressed only very low levels of the 7H9D6 epitope. Follow-ing PMA activation, total CD44 was up-regulated on all cellsexamined. The 7H9D6 epitope was also up-regulated on allcells with CEM and Daudi becoming positive for this antigen(Fig. 2a, dark line). Phorbyl myristate acetate induced up-regulation of the 7H9D6 antigen could be blocked by cyclo-phosphamide indicating that it was dependent on new proteinsynthesis (data not shown). We also examined the adhesion ofthese cell lines to hyaluronic acid, when unstimulated, and
Figure 1
7H9D6 binds to the Hermes-1-like epitope 2 andblocks binding to hyaluronic acid. KG1 cells were incubated withthe indicated unlabelled antibody (L178, Hermes-1, 3F12 orHermes-3) for 10 min prior to the addition of FITC conjugated7H9D6 (a)
or biotinylated Hermes 1 (b).
After an additional10 min incubation, cells were washed. Those labelled with bio-tinylated Hermes 1 were then incubated with streptavidin PE for10 mins and washed again prior to analysis. (c)
KG1 cells wereallowed to adhere to hyaluronic acid or BSA in the presence orabsence of 7H9D6 or Hermes-1. Bars indicate standard error.*
P
< 0.05.
n
= 3.
314
LJ Bendall
et al
.
following treatment with PMA or the CD44 activating anti-body F10-44-2 (Fig. 2b). As expected PMA and F10-44-2both resulted in increased adhesion to hyaluronic acid in mostcell lines and acted in a synergistic manner. Overall, weobserved that expression of the 7H9D6 epitope was a goodpredictor of cell adhesion to hyaluronic acid (
P
= 0.0009,
r
2
= 0.953). Together these data clearly demonstrate that the7H9D6 epitope lies within the hyaluronic acid bindingdomain.
7H9D6 recognizes a subset of CD44 molecules localized in patches on the cell surface
As described above 7H9D6, failed to bind some cells thatclearly express CD44 as determined by staining with otherCD44 antibodies such as Hermes-1 or L178. When thenumber of antigenic sites was enumerated on haemopoieticcell lines, it was apparent that 7H9D6 was recognizing only asubset of CD44 molecules (Table 1). Two populations ofCD44 molecules, those that were bound by 7H9D6 and thosenot recognized by 7H9D6, could be clearly demonstrated bystaining CD44 on KG1 cells with H9H11 after capping CD44molecules with 7H9D6 (Fig. 3). Figure 3 (a) shows cellslabelled with 7H9D6-Texas Red after capping CD44 with7H9D6-FITC. The absence of red staining demonstrates thatall the CD44 molecules recognized by 7H9D6 were located inthe cap. Figure 3b shows cells labelled with H9H11-TexasRed after capping CD44 with 7H9D6-FITC. The positivestaining with Texas Red demonstrates the presence of CD44molecules remaining outside the cap that were not recognizedby 7H9D6. In cells where CD44 had been capped usingH9H11, all 7H9D6 staining was confined to the cappedmolecules (data not shown). This defines the epitope recog-nized by 7H9D6 as identifying a distinct subpopulation of
Figure 2
7H9D6 expression correlates with adhesion tohyaluronic acid but pre-B acute lymphoblastic leukaemia (ALL)cells do not adhere to hyaluronic acid. (a) Indicated cell lines werecultured in the presence (thick line) or absence (thin line) of PMA(10 ng/mL) for 24 h and analysed by flow cytometry using theindicated antibodies or isotype control (dashed line). (b) Indicatedcell lines were cultured as for (a) and adhesion to hyaluronic acidassessed using
51
chromium based adhesion assays. Where indi-cated F10-44-2 (1
µ
g/mL) was added 20 min prior to plating inthe adhesion assay.
n
= 2.
�
, Cont; , F10-44-2;
�
, PMA; ,PMA + F10-44-2.
Table 1
Copy numbers of CD44 epitopes on hematopoietic celllines
Cell line H9H11 7H9D6 L178
KG1 515 233 479U937 575 67 –K562 – 49 102HL60 79 4 –CEM 15 0 15Daudi 24 0 –
Copy numbers are given as
×
10
3
/cell.

CD44 antibody recognizes malignant pre-B ALL
315
CD44 molecules on the cell surface. This is in keeping withthe inability of 7H9D6 to completely crossblock Hermes-1binding (Fig. 1b).
Staining of haematopoietic cells including KG1, U937 andT cells with a standard CD44 antibody such as H9H11 orHermes 1 produced a uniform surface labelling as illustratedin Fig. 3c (and data not shown). In contrast 7H9D6 produceda punctate staining pattern on each of these cells (Fig. 3d anddata not shown). When cells stained with 7H9D6 wereexamined by confocal microscopy it was clear that thesepatches were located on the cell surface (Fig. 3e). Clearly7H9D6 recognizes a subpopulation of CD44 molecules thatdemonstrate a distinctive localization in patches on the cellsurface.
7H9D6 binds acute lymphoblastic leukaemia cells but not normal B cell progenitors
Unlike most CD44 antibodies, 7H9D6 failed to stain allperipheral blood mononuclear cells. Although 7H9D6 recog-nized all T cells (CD3 positive), monocytes and granulocytes,it only bound to a subset of peripheral B cells (52
±
19%,
n
= 3) and NK cells (62
±
23%,
n
= 3) (Fig. 4a). Although redblood cells were positive for CD44, they did not bind 7H9D6(data not shown). Of note, the CD44 epitope identified by7H9D6 was expressed at a significantly reduced level onnormal bone marrow CD34
+
cells than the H9H11 epitope(Fig. 4b). Most interestingly, CD19
+
CD34
+
B-lymphoid
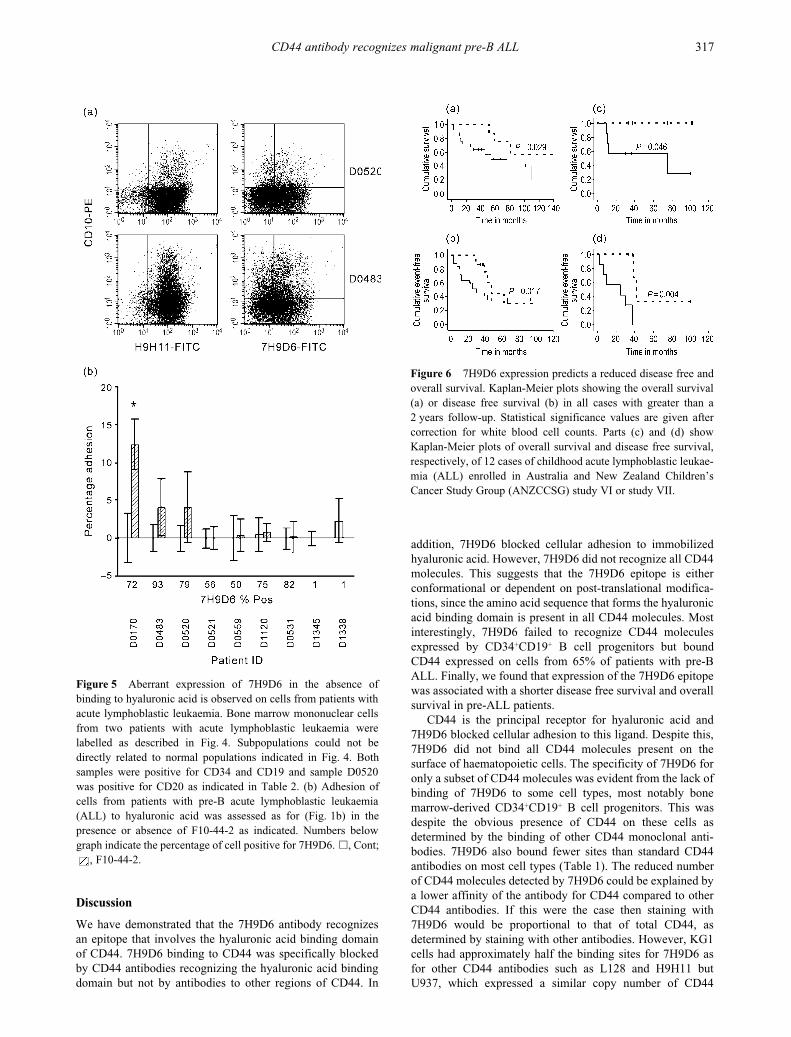
progenitors, although CD44 positive, failed to bind 7H9D6 ata detectable level (Fig. 4b). 7H9D6 expression appeared onbone marrow CD19 positive cells after the loss of CD34expression and with increasing expression of CD20 (Fig. 4c).In contrast, 7H9D6 bound leukaemic cells from 65% (28 of43) of cases of ALL (Table 2). Unlike normal B cell progeni-tors, 7H9D6 binding was not related to the absence of CD34expression or the presence of CD20 (Table 2 and Fig. 5a).CD44 was detected on 87% (all but 4 of 31) ALL cases testedusing other CD44 antibodies such as H9H11 and L178(Table 2 and data not shown). However, regardless of theirexpression of CD44 or the 7H9D6 epitope, cells from ALLpatients failed to bind hyaluronic acid, and cells from onlyone patient bound hyaluronic acid following treatment withthe CD44 activating antibody F10-44-2 (Fig. 5b). This isconsistent with the failure of normal B cells to adhere tohyaluronic acid, even in the presence of cell activators such asstimulating antibodies.
26
The unusual reactivity profile of7H9D6 on haematopoietic cells observed here is consistentwith this antibody recognizing only some CD44 molecules.The lack of expression of the 7H9D6 epitope on normal bonemarrow pre-B cells contrasts with its presence on the majorityof pre-B ALL cells.
Figure 3
7H9D6 recognizes a subset of CD44 molecules locatedin patches on the cell surface. KG1 cells were labelled with 7H9D6and the bound CD44 molecules capped using sheep antimousefluorescein (SAMF) for 3 h at 37
°
C. Cells were fixed then labelledwith (A) 7H9D6-biotin or (B) H9H11-biotin and visualized withavidin Texas Red. Original magnification 40
×
. Immunofluorescentstaining of KG1 cells using (C) H9H11-biotin and avidin Texas Redand (D) 7H9D6-FITC. (E) Confocal analysis of cells labelled with7H9D6 biotin and avidin Texas Red. A section through the centre ofthe cell is shown. Original magnification 40
×
.
Figure 4
7H9D6 has an unusual reactivity profile for CD44. (a)Peripheral blood mononuclear cells were labelled with the indi-cated antibodies and analysed by flow cytometry. All antibodiesexcept H9H11 were directly conjugated. H9H11 was biotinylatedand detected with streptavidin-tricolour. Quadrants were set togive less than 1% positive events on negative controls. (b)
CD34
+
cells were sorted from normal bone marrow and labelled with theindicated directly conjugated antibodies and analysed by flowcytometry. Quadrants were set to give less than 1% positiveevents on negative controls. (c)
Bone marrow mononuclear cellswere labelled with CD10-PE, CD19-PE-Cy5 and CD34-APC, and7H9D6-FITC, H9H11-FITC or CD20-FITC. Data was collectedon CD19 + cells with low side scatter. Populations of maturingB cell progenitors were gated using CD34 and CD10.CD10
+
CD34
+
cells are shown in pink, CD10 bright CD34
–
cells ingreen and CD10 negative to dull, CD34
–
in blue.
316
LJ Bendall
et al
.
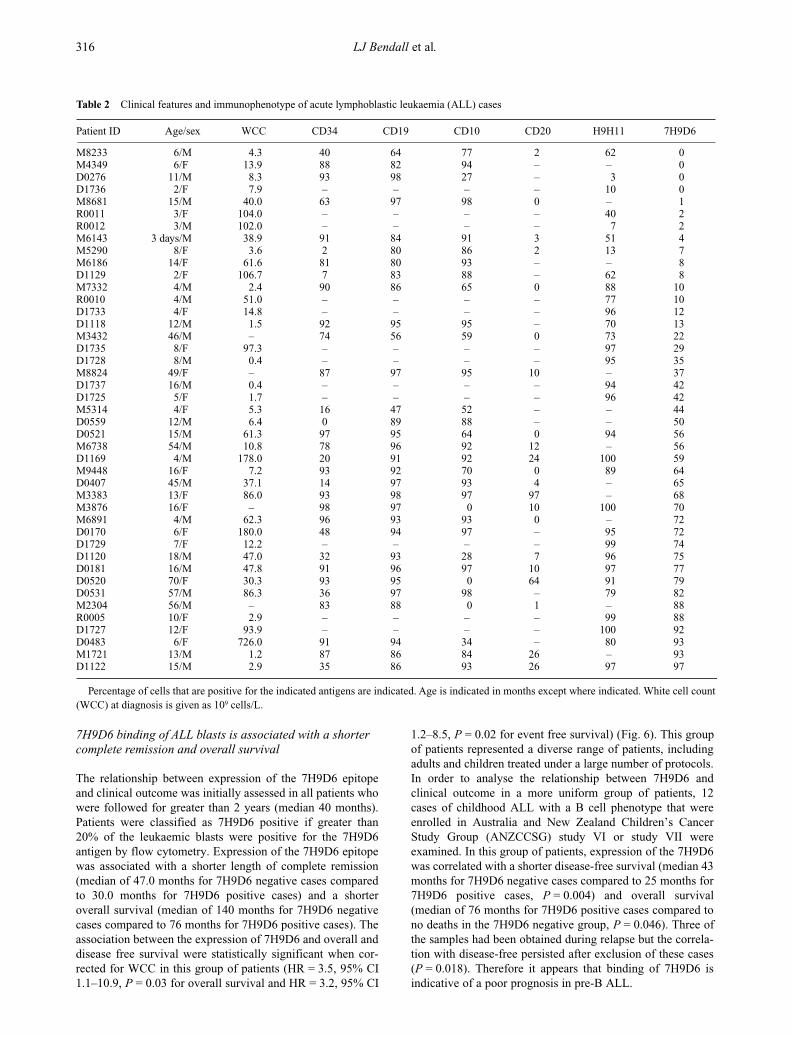
7H9D6 binding of ALL blasts is associated with a shorter complete remission and overall survival
The relationship between expression of the 7H9D6 epitopeand clinical outcome was initially assessed in all patients whowere followed for greater than 2 years (median 40 months).Patients were classified as 7H9D6 positive if greater than20% of the leukaemic blasts were positive for the 7H9D6antigen by flow cytometry. Expression of the 7H9D6 epitopewas associated with a shorter length of complete remission(median of 47.0 months for 7H9D6 negative cases comparedto 30.0 months for 7H9D6 positive cases) and a shorteroverall survival (median of 140 months for 7H9D6 negativecases compared to 76 months for 7H9D6 positive cases). Theassociation between the expression of 7H9D6 and overall anddisease free survival were statistically significant when cor-rected for WCC in this group of patients (HR = 3.5, 95% CI1.1–10.9,
P
= 0.03 for overall survival and HR = 3.2, 95% CI
1.2–8.5,
P
= 0.02 for event free survival) (Fig. 6). This groupof patients represented a diverse range of patients, includingadults and children treated under a large number of protocols.In order to analyse the relationship between 7H9D6 andclinical outcome in a more uniform group of patients, 12cases of childhood ALL with a B cell phenotype that wereenrolled in Australia and New Zealand Children’s CancerStudy Group (ANZCCSG) study VI or study VII wereexamined. In this group of patients, expression of the 7H9D6was correlated with a shorter disease-free survival (median 43months for 7H9D6 negative cases compared to 25 months for7H9D6 positive cases,
P
= 0.004) and overall survival(median of 76 months for 7H9D6 positive cases compared tono deaths in the 7H9D6 negative group,
P
= 0.046). Three ofthe samples had been obtained during relapse but the correla-tion with disease-free persisted after exclusion of these cases(
P
= 0.018). Therefore it appears that binding of 7H9D6 isindicative of a poor prognosis in pre-B ALL.
Table 2
Clinical features and immunophenotype of acute lymphoblastic leukaemia (ALL) cases
Patient ID Age/sex WCC CD34 CD19 CD10 CD20 H9H11 7H9D6
M8233 6/M 4.3 40 64 77 2 62 0M4349 6/F 13.9 88 82 94 – – 0D0276 11/M 8.3 93 98 27 – 3 0D1736 2/F 7.9 – – – – 10 0M8681 15/M 40.0 63 97 98 0 – 1R0011 3/F 104.0 – – – – 40 2R0012 3/M 102.0 – – – – 7 2M6143 3 days/M 38.9 91 84 91 3 51 4M5290 8/F 3.6 2 80 86 2 13 7M6186 14/F 61.6 81 80 93 – – 8D1129 2/F 106.7 7 83 88 – 62 8M7332 4/M 2.4 90 86 65 0 88 10R0010 4/M 51.0 – – – – 77 10D1733 4/F 14.8 – – – – 96 12D1118 12/M 1.5 92 95 95 – 70 13M3432 46/M – 74 56 59 0 73 22D1735 8/F 97.3 – – – – 97 29D1728 8/M 0.4 – – – – 95 35M8824 49/F – 87 97 95 10 – 37D1737 16/M 0.4 – – – – 94 42D1725 5/F 1.7 – – – – 96 42M5314 4/F 5.3 16 47 52 – – 44D0559 12/M 6.4 0 89 88 – – 50D0521 15/M 61.3 97 95 64 0 94 56M6738 54/M 10.8 78 96 92 12 – 56D1169 4/M 178.0 20 91 92 24 100 59M9448 16/F 7.2 93 92 70 0 89 64D0407 45/M 37.1 14 97 93 4 – 65M3383 13/F 86.0 93 98 97 97 – 68M3876 16/F – 98 97 0 10 100 70M6891 4/M 62.3 96 93 93 0 – 72D0170 6/F 180.0 48 94 97 – 95 72D1729 7/F 12.2 – – – – 99 74D1120 18/M 47.0 32 93 28 7 96 75D0181 16/M 47.8 91 96 97 10 97 77D0520 70/F 30.3 93 95 0 64 91 79D0531 57/M 86.3 36 97 98 – 79 82M2304 56/M – 83 88 0 1 – 88R0005 10/F 2.9 – – – – 99 88D1727 12/F 93.9 – – – – 100 92D0483 6/F 726.0 91 94 34 – 80 93M1721 13/M 1.2 87 86 84 26 – 93D1122 15/M 2.9 35 86 93 26 97 97
Percentage of cells that are positive for the indicated antigens are indicated. Age is indicated in months except where indicated. White cell count(WCC) at diagnosis is given as 10
9
cells/L.
CD44 antibody recognizes malignant pre-B ALL
317
Discussion
We have demonstrated that the 7H9D6 antibody recognizesan epitope that involves the hyaluronic acid binding domainof CD44. 7H9D6 binding to CD44 was specifically blockedby CD44 antibodies recognizing the hyaluronic acid bindingdomain but not by antibodies to other regions of CD44. In
addition, 7H9D6 blocked cellular adhesion to immobilizedhyaluronic acid. However, 7H9D6 did not recognize all CD44molecules. This suggests that the 7H9D6 epitope is eitherconformational or dependent on post-translational modifica-tions, since the amino acid sequence that forms the hyaluronicacid binding domain is present in all CD44 molecules. Mostinterestingly, 7H9D6 failed to recognize CD44 moleculesexpressed by CD34
+
CD19
+
B cell progenitors but boundCD44 expressed on cells from 65% of patients with pre-BALL. Finally, we found that expression of the 7H9D6 epitopewas associated with a shorter disease free survival and overallsurvival in pre-ALL patients.
CD44 is the principal receptor for hyaluronic acid and7H9D6 blocked cellular adhesion to this ligand. Despite this,7H9D6 did not bind all CD44 molecules present on thesurface of haematopoietic cells. The specificity of 7H9D6 foronly a subset of CD44 molecules was evident from the lack ofbinding of 7H9D6 to some cell types, most notably bonemarrow-derived CD34
+
CD19
+
B cell progenitors. This wasdespite the obvious presence of CD44 on these cells asdetermined by the binding of other CD44 monoclonal anti-bodies. 7H9D6 also bound fewer sites than standard CD44antibodies on most cell types (Table 1). The reduced numberof CD44 molecules detected by 7H9D6 could be explained bya lower affinity of the antibody for CD44 compared to otherCD44 antibodies. If this were the case then staining with7H9D6 would be proportional to that of total CD44, asdetermined by staining with other antibodies. However, KG1cells had approximately half the binding sites for 7H9D6 asfor other CD44 antibodies such as L128 and H9H11 butU937, which expressed a similar copy number of CD44
Figure 5
Aberrant expression of 7H9D6 in the absence ofbinding to hyaluronic acid is observed on cells from patients withacute lymphoblastic leukaemia. Bone marrow mononuclear cellsfrom two patients with acute lymphoblastic leukaemia werelabelled as described in Fig. 4. Subpopulations could not bedirectly related to normal populations indicated in Fig. 4. Bothsamples were positive for CD34 and CD19 and sample D0520was positive for CD20 as indicated in Table 2. (b)
Adhesion ofcells from patients with pre-B acute lymphoblastic leukaemia(ALL) to hyaluronic acid was assessed as for (Fig. 1b) in thepresence or absence of F10-44-2 as indicated. Numbers belowgraph indicate the percentage of cell positive for 7H9D6.
�
, Cont;, F10-44-2.
Figure 6
7H9D6 expression predicts a reduced disease free andoverall survival. Kaplan-Meier plots showing the overall survival(a) or disease free survival (b) in all cases with greater than a2 years follow-up. Statistical significance values are given aftercorrection for white blood cell counts. Parts (c) and (d) showKaplan-Meier plots of overall survival and disease free survival,respectively, of 12 cases of childhood acute lymphoblastic leukae-mia (ALL) enrolled in Australia and New Zealand Children’sCancer Study Group (ANZCCSG) study VI or study VII.

318
LJ Bendall
et al.
molecules when examined using H9H11, had only one tenththe number of sites for 7H9D6. Most importantly, two poolsof CD44, one recognized by 7H9D6 and the other not, wereclearly demonstrated in antibody capping experiments. Asignificant proportion of CD44 molecules remained outsidethe capped region following crosslinking of 7H9D6 (Fig. 3).However, the CD44 molecules that were not capped by7H9D6 could be stained by other CD44 antibodies, such asH9H11, but were not recognized by 7H9D6. Therefore7H9D6 recognizes a subpopulation of CD44 molecules on thecell surface. Interestingly, 7H9D6 staining resulted in apunctate pattern on the cells surface that was not clearlyevident when the same cells were stained with standard CD44antibodies. This demonstrates a distinct distribution of7H9D6 recognized-CD44 molecules on the cell surface.
The precise nature of the epitope recognized by 7H9D6remains to be determined. CD44 has been reported to existin many different forms resulting from alternative mRNAsplicing and post-translational modifications including theattachment of carbohydrate, dimerization, sulfation and phos-phorylation.2 A number of attempts were made to identify thedistinguishing features of the 7H9D6 epitope. Removal ofsialic acid with neuraminidase or N-linked carbohydrate withN-glycosidase F did not alter staining with 7H9D6, suggest-ing that N-linked carbohydrate is not involved in the 7H9D6epitope (data not shown). CD44 is known to associate withthe actin cytoskeleton, an association that could result inclustering, thereby explaining the patchy staining pattern.However, disruption of the cytoskeleton with a number ofagents did not diminish the punctate staining seen with7H9D6 (data not shown). It has been reported that function-ally active CD44 partitions into Triton X-100 insoluble lipidrafts within the plasma membrane, however, co-localizationof 7H9D6 staining with that of known raft associating anti-gens could not be demonstrated (data not shown). Despitethese data, it is still possible that clustering of CD44 isnecessary for 7H9D6 binding. Factors such as O-linkedcarbohydrate, sulfation, phosphorylation or association withother proteins could also be important and have yet to betested. However, it seems that alternative mRNA splicing isnot involved since expression of cDNA lacking any variantexons can encode the 7H9D6 epitope.
CD44 can be over or under expressed on ALL cells andthis altered expression can be useful for the detection ofminimal residual disease.27,28 Most interestingly, 7H9D6 com-pletely failed to recognize CD44 expressed by normal bonemarrow B cell progenitors. Following loss of CD34, the Bcell progenitors initially remain negative for 7H9D6 stainingbut acquire the 7H9D6 epitope at the time of CD10 downregulation and acquisition of CD20, an immature B cellphenotype (Fig. 4). In contrast to the absence of staining onnormal early B cell progenitors, 7H9D6 recognized CD44expressed by the majority of cases of B lineage ALL, regard-less of the relative stage of maturity of the leukaemic blasts(Table 2). Dworzak et al.27,28 have demonstrated that leukae-mic populations of B lineage cells in the marrow can beidentified by the aberrant expression of surface markersincluding that of CD44. However, CD44 is only aberrantlyexpressed in approximately one third of cases of B lineageALL. In contrast the epitope recognized 7H9D6 is aberrantlyexpressed on two thirds of B lineage ALL cases, making this
antibody a better candidate for distinguishing normal fromleukaemic pre-B cells in the bone marrow.
How the presence of the 7H9D6 epitope alters CD44function is not known. Therefore, the reasons for the absenceof the 7H9D6 epitope on B cell progenitors and the conse-quences of its aberrant expression in B cell leukaemias are notobvious. CD44 has long been implicated in the regulation ofhaematopoietic cell production by the bone marrow.10,34,35 Themild phenotype of CD44 knockout mice demonstrates thatCD44 is not essential for normal haemopoiesis and is morelikely to play a role under situations of stress.9 However, theprecise role for CD44 in haematopoiesis still remains elusive.The association between 7H9D6 expression and pooroutcome suggests that the expression of the 7H9D6 epitopemay impact on the biology of pre-B ALL. A larger study on amore uniform group of patients is needed to confirm andquantify the prognostic value of 7H9D6 epitope expression.The failure of cells from patients with pre-B ALL to adhere tohyaluronic acid also suggests that the function of CD44recognized by the 7H9D6 antibody may not relate tohyaluronic acid binding but to interactions with other CD44ligands. Further work regarding the nature of the CD44epitope recognized by 7H9D6 is likely to provide insightsinto CD44 function, normal B cell maturation and the biologyof pre-B ALL.
Acknowledgements
This work was supported by the National Health and MedicalResearch Council of Australia, the University of SydneyCancer Fund and the Cure Cancer Australia Foundation. Wewould like to thank Mary Sartor for cell sorting, ChristineSmythe for assistance with confocal microscopy, Karen Bythfor statistical analysis of data, Dr Richard Lock for theprovision of xenograft samples and Dr Peter Shaw, Dr MarieBleakley, Dr Sukesh Nair and Ms Penny Stavros for assist-ance in obtaining patient information.
References
1 Underhill C. CD44: The hyaluronan receptor. J. Cell Sci. 1992;103: 293–8.
2 Bendall L, Gottlieb D. CD44 and adhesion of normal and leu-kaemic CD34+ cells to bone marrow stroma. Leuk. Lymphoma1999; 32: 427–39.
3 Dimitroff CJ, Lee JY, Fuhlbrigge RC, Sackstein R. A distinctglycoform of CD44 is an 1-selectin ligand on human hemato-poietic cells. Proc. Natl Acad. Sci. USA 2000; 97: 13 841–6.
4 Dimitroff CJ, Lee JY, Rafii S, Fuhlbrigge RC, Sackstein R.CD44 is a major E-selectin ligand on human hematopoietic pro-genitor cells. J. Cell Biol. 2001; 153: 1277–86.
5 Jalkanen S, Saari S, Kalimo H et al. Lymphocyte migration intothe skin: the role of lymphocyte homing receptor (CD44) andendothelial cell antigen (HECA-452). J. Invest. Dermatol. 1990;94: 786–92.
6 Salmi M, Jalkanen S. Regulation of lymphocyte traffic tomucosa-associated lymphatic tissues. Gastroenterol. Clin. NorthAm. 1991; 20: 495–510.
7 Picker LJ, Terstappen LW, Rott LS, Streeter PR, Stein H,Butcher EC. Differential expression of homing-associated adhe-sion molecules by T cell subsets in man. J. Immunol. 1990; 145:3247–55.

CD44 antibody recognizes malignant pre-B ALL 319
8 Toyama-Sorimachi N, Miyake K, Miyasaka M. Activation ofCD44 induces ICAM-1/LFA-1-independent, Ca2+, Mg (2+) –independent adhesion pathway in lymphocyte–endothelial cellinteraction. Eur. J. Immunol. 1993; 23: 439–46.
9 Schmits R, Filmus J, Gerwin N et al. CD44 regulates hematopoi-etic progenitor distribution, granuloma formation, and tumori-genicity. Blood 1997; 90: 2217–33.
10 Miyake K, Medina KL, Hayashi S-I, Ono S, Hamaoka T,Kincade PW. Monoclonal antibodies to Pgp-1/CD44 blocklymphohemopoiesis in long-term bone marrow cultures. J. Exp.Med. 1990; 171: 477–88.
11 Manabe A, Coustan-Smith E, Behm FG, Raimondi SC,Campana D. Bone marrow-derived stromal cells prevent apop-totic cell death in B-lineage acute lymphoblastic leukemia. Blood1992; 79: 2370–7.
12 Screaton G, Bell M, Jackson D, Cornelis F, Gerth U, Bell J.Genomic structure of DNA encoding the lymphocyte homingreceptor CD44 reveals at least 12 alternatively spliced exons.Proc. Natl Acad. Sci. USA 1992; 89: 12 160–4.
13 Stamenkovic I, Amiot M, Pesando J, Seed B. A lymphocytemolecule implicated in lymph node homing is a member of thecartilage link protein family. Cell 1989; 56: 1057–62.
14 Nottenburg C, Rees G, St John T. Isolation of mouse CD44cDNA structural features are distinct from the primate cDNA.Proc. Natl Acad. Sci. USA 1989; 86: 8521–5.
15 Goldstein LA, Zhou DF, Picker LJ et al. A human lymphocytehoming receptor, the hermes antigen, is related to cartilageproteoglycan core and link proteins. Cell 1989; 56: 1063–72.
16 Jackson D, Bell J, Timans J, Shields J, Whittle N. Proteoglycanforms of the lymphocyte homing receptor CD44 are alternativelyspliced variants containing the v3 exon. J. Cell Biol. 1995; 128:673–85.
17 Bennett KL, Jackson DG, Simon JC et al. CD44 isoformscontaining exon V3 are responsible for the presentation ofheparin-binding growth factor. J. Cell Biol. 1995; 128: 687–98.
18 Faassen AE, Schrager JA, Klein DJ, Oegema TR, Couchman JR,McCarthy JB. A cell surface chondroitin sulfate proteoglycan,immunologically related to CD44, is involved in type I collagen-mediated melanoma cell motility and invasion. J. Cell Biol.1992; 116: 521–31.
19 Faassen AE, Mooradian DL, Tranquillo RT et al. Cell surfaceCD44-related chondroitin sulfate proteoglycan is required fortransforming growth factor-beta-stimulated mouse melanomacell motility and invasive behavior on type I collagen. J. Cell Sci.1993; 105: 501–11.
20 Bendall LJ, Gottlieb DJ, Bradstock KF. The expression of CD44variants in normal haemopoietic progenitors and acute myeloidleukaemia cells. Leukemia 2000; 14: 1239–46.
21 Pure E, Camp R, Peritt D, Panettieri RJ, Lazaar A, Nayak S.Defective phosphorylation and hyaluronate binding of CD44with point mutations in the cytoplasmic domain. J. Exp. Med.1995; 181: 55–62.
22 Legg JW, Lewis CA, Parsons M, Ng T, Isacke CM. A novelPKC-regulated mechanism controls CD44 ezrin association anddirectional cell motility. Nat. Cell Biol. 2002; 4: 399–407.
23 Liu D, Sy M. Phorbol myristate acetate stimulates the dimeriza-tion of CD44 involving a cysteine in the transmembrane domain.J. Immunol. 1997; 159: 2702–11.
24 Kansas GS, Muirhead MJ, Dailey MO. Expression of the CD11/CD18, leukocyte adhesion molecule 1, and CD44 adhesion mol-ecules during normal myeloid and erythroid differentiation inhumans. Blood 1990; 76: 2483–92.
25 Ryan DH, Nuccie BL, Abboud CN, Liesveld JL. Maturation-dependent adhesion of human B cell precursors to the bonemarrow microenvironment. J. Immunol. 1990; 145: 477–84.
26 Hathcock KS, Hirano H, Murakami S, Hodes RJ. CD44 expres-sion on activated B cells. Differential capacity for CD44-dependent binding to hyaluronic acid. J. Immunol. 1993; 151:6712–22.
27 Dworzak MN, Fritsch G, Fleischer C et al. Comparative pheno-type mapping of normal vs. malignant pediatric B-lymphopoiesisunveils leukemia-associated aberrations. Exp. Hematol. 1998;26: 305–13.
28 Dworzak MN, Stolz F, Froschl G et al. Detection of residualdisease in pediatric B-cell precursor acute lymphoblastic leuke-mia by comparative phenotype mapping: a study of five casescontrolled by genetic methods. Exp. Hematol. 1999; 27: 673–81.
29 Lock R, Liem N, Farnsworth M et al. The nonobese diabetic/severe combined immunodeficient (NOD/SCID) mouse modelof childhood acute lymphoblastic leukemia reveals intrinsicdifferences in biologic characteristics at diagnosis and relapse.Blood 2002; 99: 4100–8.
30 Kortlepel K, Bendall LJ, Gottlieb DJ. Human acute myeloidleukemia cells express adhesion proteins and bind to bonemarrow fibroblast monolayers and extracellular matrix proteins.Leukemia 1993; 7: 1174–9.
31 Zannettino A, Rayner J, Ashman L, Gonda T, Simmons P. Apowerful new technique for isolating genes encoding cell surfaceantigens using retroviral expression cloning. J. Immunol. 1996;156: 611–20.
32 The T, Feltkamp T. Conjugation of fluorescein isothiocyanate toantibodies. I. Experiments on the conditions of conjugation.Immunology 1970; 18: 865–73.
33 Anstee D, Gardner B, Spring F et al. New monoclonal antibodiesin CD44 and CD58: their use to quantify CD44 and CD58 onnormal human erythrocytes and to compare the distribution ofCD44 and CD58 in human tissues. Immunology 1991; 74:197–205.
34 Rossbach H-C, Krizanac-Bengez L, Santos E, Gooley T,Sandmaier B. An Antibody to CD44 enhances hematopoiesis inlong-term marrow cultures. Exp. Hematol. 1996; 24: 221–7.
35 Sandmaier B, Storb R, Appelbaum F, Gallatin W. An antibodythat facilitates hematopoietic engraftment recognizes CD44.Blood 1990; 76: 630–5.




















