Chromosomal microarrays testing in children with developmental disabilities and congenital anomalies
Upload
uni-frankfurtCategory
view
0download
0
[CANCER RESEARCH 59, 3357–3362, July 15, 1999]
Advances in Brief
Rapid Isolation of Chromosomal Breakpoints from Patients with t(4;11) AcuteLymphoblastic Leukemia: Implications for Basic and Clinical Research1
Martin Reichel, Esther Gillert, Ina Breitenlohner, Reinald Repp, Johann Greil, Jorn D. Beck, Georg H. Fey, andRolf Marschalek2
Chair of Genetics, University of Erlangen–Nurnberg, D-91058 Erlangen [M. R., E. G., I. B., G. H. F., R. M.], and Department of Pediatrics, University of Erlangen–Nurnberg,D-91054 Erlangen [R. R., J. G., J. D. B.], Germany
Abstract
Chromosomal translocations t(4;11)(q21;q23) are associated with agroup of acute lymphoblastic leukemias with very poor prognosis. Fromthe complete sequences of the breakpoint cluster regions of the humanMLL and AF-4 translocation partner genes, a novel set of 66 oligonucleo-tides that facilitates the rapid identification of translocation breakpointsby PCR analysis of genomic DNA was designed. For each breakpoint, apair of optimally suited primers can be assigned, which improves themonitoring of the disease during treatment. Comparison of the break-points with the corresponding parental sequences also contributes to ourbetter understanding of the illegitimate recombination events leading tothese translocations.
Introduction
Certain chromosomal translocations are known to be the initialsteps of the malignant transformation of hematopoietic cells leading tothe development of myeloid and lymphoblastic leukemias and lym-phomas (1). Translocations to theMLL3 gene (2, 3) are associatedwith an AML or ALL disease phenotype. More than 30 differentchromosomal translocations to the 11q23 region have so far beenidentified, all of which are associated with hematological malignan-cies, and many translocation partner loci have been analyzed at themolecular level (for review see Refs. 4–6). Among these transloca-tions, the chromosomal translocation t(4;11) is regularly associatedwith high-risk infant acute pro-B lymphoblastic leukemias and hasbeen proposed to initiate the development of this malignancy (4–8).Leukemic blasts with chromosomal translocation t(4;11) are highlyresistant against current treatment protocols. Although an initial re-mission can be achieved in about 80–90% of all cases, the relapse rateis exceedingly high. Thus, there is need for a better control of appliedtherapy. Monitoring of MRD allows the control of the efficacy ofinduction chemotherapy or subsequent allogeneic stem cell transplan-tation. Molecular methods for MRD-monitoring of t(4;11)-patients, sofar, were based mostly on RT-PCR analysis of breakpoint mRNAs(9). However, RT-PCR experiments using one t(4;11) break-regiontranscript as the sole molecular marker may generate ambiguousresults, as it has been demonstrated recently (10). Derivative (der) 11breakregion transcripts were identified in normal biopsy material
without any accompanying cytogenetic evidence for a chromosomalrearrangement t(4;11). In addition, the RNA may be degraded andsteady-state RNA concentration or transcriptional activity may changeduring therapy, which may negatively affect a precise analysis.
The use of genomic DNA as template for PCR experiments is veryattractive because chromosomal fusion sites are characteristic andusually stable features of t(4;11) cells (11). However, this type ofanalysis, so far, was hampered by the fact that the DNA sequence ofthe breakpoint cluster region of the humanAF-4 gene was not avail-able, and systematic analyses of t(4;11) breakpoints on the genomicDNA level could not be performed. In this study, the completebreakpoint cluster region of theAF-4 gene was subcloned and se-quenced. The sequence was used to design sets of specific oligonu-cleotides, allowing investigators to retrieve chromosomal breakpointinformation from t(4;11) patient DNA. This resulted in the assignmentof optimally suited DNA primer pairs for each individual patient. Thisnew tool can be applied in any clinical setting and may close adiagnostic gap for this group of high-risk acute leukemias.
Materials and Methods
Patient Material. Leukemic cells used in this study were collected fromchildren enrolled in treatment protocols of the German BFM-95 study group,which had been approved by institutional review boards. All patients and theirparents gave informed consent for treatment and for collection of material forbiological studies. Bone marrow biopsy material from patients P. B. (1-year-old male) and E. T. (3-year-old female) was collected at the time of diagnosis.Patient biopsy material was kindly provided by Drs. E. R. Panzer-Grumayerand O. A. Haas from the St. Anna Childrens Hospital (Vienna, Austria). Allother patient breakpoint data shown in Fig. 1 and Table 1 were obtained fromearlier studies (12).
Oligonucleotides. Breakpoint analysis was performed by amplification ofDNA fragments usingMLL- andAF-4-specific oligonucleotides (x, directed tothe telomer; y, directed to the centromer): MLLx2 (59-GGAAAAGAGTGAA-GAAGGGAATGTCTCG-39), MLLy7 (59-CAAAACTTGTGGAAGGGCT-CACAACAGACTTGG-39), AF4x2 (59-CAGTTTGTACAATGACGACA-GAAACCTGCTTCG-39), AF4x4 (59-GGCACCAGTGCATCTGCTGTCA-ACAATGGAG-39), AF4x6 (59-GGATGAACATGCGTCAGTCATGCAC-AGCTCAG-39), AF4x8 (59-AGAATTCCCTGCTGCCATGGGTGGTGC-CTG-39), AF4x10 (59-GGTAGGGTACTGTTACCATCCTGTTTTACTG-AG-39), AF4x12 (59-CTTGGTATCATCCTGCTGCTTTTTCTCCAGGTG-39), AF4x14 (59-CAGTATTTCCTGAGCTAGAGGGCTGTTGTGAG-39),AF4x16 (59-CAAAGGTCAAGTACACGACGGCTTCTTCAG-39), AF4x18(59-GTGATGAAAAGCTAATAGGCGAGGACGATAGTG-39), AF4x20 (59-GCTGTTGAGATTACTGCTGGAATAAGCCCAGG-39), AF4x22 (59-CC-CAAACTGCATTTAGGCAACTACTCTTATAGG-39), AF4x24 (59-AGA-AAAGTGTGACTTATCTAACTAGCGAGTTG-39), AF4x26 (59-GTCCTG-GTTCATCCAACATAACAAAACTCTTGG-39), AF4x28 (59-ATGCCCAT-TTATGATTTGGAAATCATAACTTGG-39), AF4x30 (59-CATGGCCGCC-TCCTTTGACAGCAATACATACG-39), AF4x32 (59-TGATTTCTTCA-ACACTTGATCTTATGGTACTGG-39), AF4y2 (59-CCTCAACTGACTGA-GAAGGCATCTTCAGTTTGG-39), AF4y4 (59-GGGTCCATGTGCATG-AGCACTCACAGAGATG-39), AF4y6 (59-ACTTGGGTCTTGGAAAT-
Received 4/28/99; accepted 6/1/99.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby markedadvertisementin accordance with18 U.S.C. Section 1734 solely to indicate this fact.
1 Supported by research grants SFB 466C4 (to R. M. and J. G.) and SFB 466C5 (toG. H. F. and R. M.) from the Deutsche Forschungsgemeinschaft (DFG), research grant96.047.1 (to R. M., J. G., and G. H. F.) from the Wilhelm Sander Stiftung, and support bythe J. and F. Marohn Foundation.
2 To whom requests for reprints should be addressed, at Chair of Genetics, Universityof Erlangen—Nurnberg, Staudstr. 5, D-91058 Erlangen, Germany. Phone: 49-9131-852-8530; Fax: 49-9131-852-8526; E-mail: [email protected].
3 The abbreviations used are: MLL, Mixed-Lineage Leukemia; ALL, acute lympho-blastic leukemia; AML, Acute Myeloid Leukemia; MRD, minimal residual disease;RT-PCR, reverse transcription-PCR.
3357
Research. on November 23, 2015. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

TAAATGCAAGCATCTG-39), AF4y8 (59-GCCAAAGGGATTAAGAAGA-TTAGAATGGGCTTG-39), AF4y10 (59-AGGAACAGTGTTTACCTACT-TCTGTCTCCAGAG-39), AF4y12 (59-TGGGTCTGCTCAAAGTGAGGAA-TAATCATACAG-39), AF4y14 (59-TTAACCAGTCTTTCCTCCATGATA-TGAGGACAG-39), AF4y16 (59-CACTGTCTCGTGTGCTGGCGGCTGC-AATGG-39), AF4y18 (59-AAGACTCATCTGATACACTGATTTACTGA-TCTGG-39), AF4y20 (59-CCACATGAGGCCCCATGAAGCCCATTGTG-G-39), AF4y22 (59-CCTGGCCCAGTGTTCACTTGCTAAACTTCACG-39),AF4y24 (59-TCAAGAAAATCTAGTTTGTAAGTCCTTCTCTGG-39),AF4y26 (59-CTCTTCAACACAATGGACTTCATTGGAGTAGG-39),AF4y28 (59-TGTTTACTGACTGGCCATCTGCTTTGTTCAGG-39),AF4y30 (59-GGTTTTGGGTTACAGAACTGACATGCTGAGAG-39),AF4y32 (59-CGATGACGTTCCTTGCTGAGAATTTGAGTGAG-39).
DNA Purification. Genomic DNA from 23 105 to 2 3 107 cells waspurified by ion exchange chromatography (Qiagen Genomic tips 100/G; Qia-gen Ltd., Hilden, Germany), according to the manufacturer’s recommenda-tions. The DNA was dissolved in 100ml of TE buffer (pH 8.0) and itsconcentration was determined by absorbance at 260 nm. In general, 40–60mgof high molecular weight DNA (50–100 kb) were obtained from 107 viablecells.
Amplification and Analysis of Genomic DNA Fragments SpanningTranslocation Breakpoints. Long-range PCR experiments were performedas published (13). Briefly, oligonucleotide MLLx2 was used in combinationwith one of the oligos of the AF4y2 to AF-4y32 primer set, whereas MLLy7was used in combinations with one of the AF4x2 to AF4x32 primers. All DNAamplimers presumably spanning chromosomal breakpoints were cloned andsequenced. All data were compared with the known wild-type sequences of thebreakpoint cluster regions of theMLL andAF-4 genes (3, 11, 12, 14–16) andsequences newly derived in this study. All sequence data were deposited inpublic databases under the following accession codes: breakpoint clusterregion (bcr) of theMLL gene: X83604; patient breakpoints: AJ000166 toAJ000180; Y16596 to Y16609; AJ235330 to AJ235380; Y18922 to Y18929;and breakpoint cluster region of theAF-4 gene: AJ238093.
Results
Cloning the Breakpoint Cluster Region of the AF-4 Gene.Large segments of theAF-4 gene had previously been isolated, andtwo of theAF-4contigs (contigs 2 and 3) were partly overlapping withthe AF-4 breakpoint cluster region (bcr; Ref. 16). The gap betweenthese contigs was closed by screening individual PAC clones usinggenomic probes fromAF-4contigs 2 or 3. Five PAC clones containingthe missing information (RPCI 1, 3–5, 1048b13; RPCI 1, 3–5,
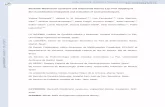
263e20; RPCI 1, 3–5, 666m14; RPCI 1, 3–5, 698g15; and RPCI 1,3–5, 583j12) were obtained from the German Resource Center inHeidelberg (RZPD). Appropriate restriction fragments were sub-cloned and finally sequenced on both strands. Where necessary, theorientation of certain genomic fragments was verified by genomicPCR experiments using oligonucleotides specifically binding to ter-minal regions of subcloned restriction fragments (data not shown).The final sequence has been deposited in public databases (Genbankaccession code AJ238093), and the structure of theAF-4 breakpointcluster region is shown in Fig. 1. The exon/intron structure (AF-4exons 2–7) and a restriction map for the enzymes BamHI (B), EcoRI(E), and HindIII (H) are indicated.
Analysis of the AF-4 Breakpoint Cluster Region for the Pres-ence of Repetitive DNA Sequences and Potential TopoisomeraseII or Nonamer/Heptamer Sequences.The complete DNA sequenceof the AF-4 bcr (52.909 bp) was analyzed for the presence of repet-itive DNA elements by using the Blast2 server from the NationalCenter for Biotechnology Information.4 Repetitive DNA element se-quences (Alu J: U14567, LINE-1: U93562, and Kpn-1: X03145) wereused to identify homologous DNA sequences within the breakpointcluster region of theAF-4 gene. A total of 26 Alu repetitive elements(19 of 26) or Alu remnants (,250 bp: 7 of 26), but no repetitiveLINE-1 or Kpn-1 elements were identified. A search for potentialtopoisomerase II consensus sequences or nonamer/heptamer signalsequences was performed by using the computer program DNAInspector II. Due to the highly degenerated character, potential topoi-somerase II consensus sequences were only accepted with deviationsfrom the known consensus sequence (59-RNYNNCNNGYNGGT-NYNY-39; Ref. 17) of not more than one mismatch. Two perfect sitesand 30 sites with one mismatch were identified during the analysis ofboth DNA strands (Fig. 1). Nonamer/Heptamer signal sequences wereaccepted with up to five mismatches from the known consensussequences of the canonical 7-23-9, 7-12-9, 9-23-7, or 9-12-7 signalsequences (18). Ten mismatched DNA sequences were identified(three sites with four mismatches; seven sites with five mismatches),which are also shown in Fig. 1. The precise locations of all Alu
4 http://www.ncbi.nlm.nih.gov/gorf/bl2.html.
Fig. 1. Breakpoint cluster region of the humanAF-4 gene.Top, size scale.Clusters 1–3, clusters of translocation breakpoints.Vertical barsrepresent breakpoints from derivative11 (brx der11) and derivative 4 (brx der4) chromosomes, mapped as described in the text.Black line, introns;black boxes, exons. Numbering of exons of theAF-4 gene accordingto Ref. 16. A restriction map is shown for the breakpoint cluster regions.B, E, andH, restriction cleavage sites for the enzymes BamHI, EcoRI, and HindIII. Numbers represent fragmentsizes in bp. Alu elements (cw, clockwise;ccw, counter clockwise) are indicated astriangles in the exon-intron map.Topo, topoisomerase II consensus sequences (long vertical line,zero mismatches;short vertical line, one mismatch) analyzed for both the coding and the non-coding DNA strands.VDJ, potential nonamer/heptamer signal sequences for canonical7-23-9, 7-12-9, 9-23-7, and 9-12-7 motifs (long or short vertical lines, four or five mismatches).
3358
CHROMOSOMAL TRANSLOCATIONS T(4;11)
Research. on November 23, 2015. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
elements and consensus sequences in the breakpoint cluster region ofthe humanAF-4 gene are summarized in Table 2.
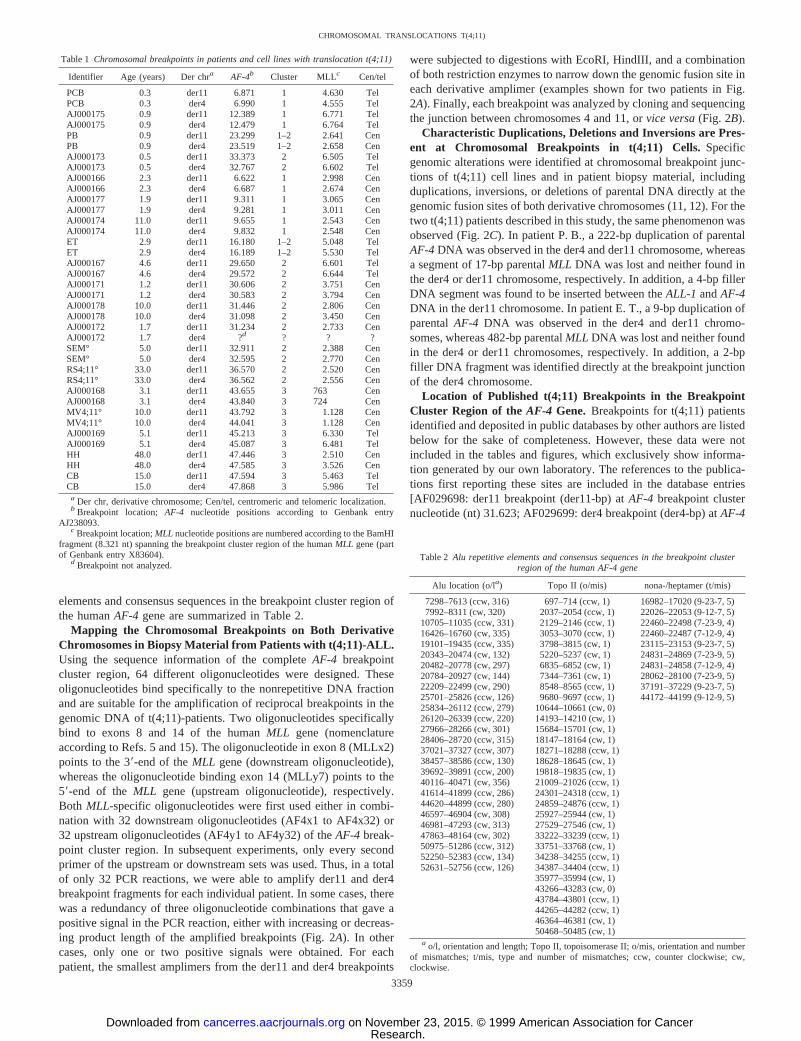
Mapping the Chromosomal Breakpoints on Both DerivativeChromosomes in Biopsy Material from Patients with t(4;11)-ALL.Using the sequence information of the completeAF-4 breakpointcluster region, 64 different oligonucleotides were designed. Theseoligonucleotides bind specifically to the nonrepetitive DNA fractionand are suitable for the amplification of reciprocal breakpoints in thegenomic DNA of t(4;11)-patients. Two oligonucleotides specificallybind to exons 8 and 14 of the humanMLL gene (nomenclatureaccording to Refs. 5 and 15). The oligonucleotide in exon 8 (MLLx2)points to the 39-end of theMLL gene (downstream oligonucleotide),whereas the oligonucleotide binding exon 14 (MLLy7) points to the59-end of theMLL gene (upstream oligonucleotide), respectively.Both MLL-specific oligonucleotides were first used either in combi-nation with 32 downstream oligonucleotides (AF4x1 to AF4x32) or32 upstream oligonucleotides (AF4y1 to AF4y32) of theAF-4 break-point cluster region. In subsequent experiments, only every secondprimer of the upstream or downstream sets was used. Thus, in a totalof only 32 PCR reactions, we were able to amplify der11 and der4breakpoint fragments for each individual patient. In some cases, therewas a redundancy of three oligonucleotide combinations that gave apositive signal in the PCR reaction, either with increasing or decreas-ing product length of the amplified breakpoints (Fig. 2A). In othercases, only one or two positive signals were obtained. For eachpatient, the smallest amplimers from the der11 and der4 breakpoints
were subjected to digestions with EcoRI, HindIII, and a combinationof both restriction enzymes to narrow down the genomic fusion site ineach derivative amplimer (examples shown for two patients in Fig.2A). Finally, each breakpoint was analyzed by cloning and sequencingthe junction between chromosomes 4 and 11, orvice versa(Fig. 2B).
Characteristic Duplications, Deletions and Inversions are Pres-ent at Chromosomal Breakpoints in t(4;11) Cells.Specificgenomic alterations were identified at chromosomal breakpoint junc-tions of t(4;11) cell lines and in patient biopsy material, includingduplications, inversions, or deletions of parental DNA directly at thegenomic fusion sites of both derivative chromosomes (11, 12). For thetwo t(4;11) patients described in this study, the same phenomenon wasobserved (Fig. 2C). In patient P. B., a 222-bp duplication of parentalAF-4DNA was observed in the der4 and der11 chromosome, whereasa segment of 17-bp parentalMLL DNA was lost and neither found inthe der4 or der11 chromosome, respectively. In addition, a 4-bp fillerDNA segment was found to be inserted between theALL-1 andAF-4DNA in the der11 chromosome. In patient E. T., a 9-bp duplication ofparentalAF-4 DNA was observed in the der4 and der11 chromo-somes, whereas 482-bp parentalMLL DNA was lost and neither foundin the der4 or der11 chromosomes, respectively. In addition, a 2-bpfiller DNA fragment was identified directly at the breakpoint junctionof the der4 chromosome.
Location of Published t(4;11) Breakpoints in the BreakpointCluster Region of theAF-4 Gene. Breakpoints for t(4;11) patientsidentified and deposited in public databases by other authors are listedbelow for the sake of completeness. However, these data were notincluded in the tables and figures, which exclusively show informa-tion generated by our own laboratory. The references to the publica-tions first reporting these sites are included in the database entries[AF029698: der11 breakpoint (der11-bp) atAF-4 breakpoint clusternucleotide (nt) 31.623; AF029699: der4 breakpoint (der4-bp) atAF-4
Table 1 Chromosomal breakpoints in patients and cell lines with translocation t(4;11)
Identifier Age (years) Der chra AF-4b Cluster MLLc Cen/tel
PCB 0.3 der11 6.871 1 4.630 TelPCB 0.3 der4 6.990 1 4.555 TelAJ000175 0.9 der11 12.389 1 6.771 TelAJ000175 0.9 der4 12.479 1 6.764 TelPB 0.9 der11 23.299 1–2 2.641 CenPB 0.9 der4 23.519 1–2 2.658 CenAJ000173 0.5 der11 33.373 2 6.505 TelAJ000173 0.5 der4 32.767 2 6.602 TelAJ000166 2.3 der11 6.622 1 2.998 CenAJ000166 2.3 der4 6.687 1 2.674 CenAJ000177 1.9 der11 9.311 1 3.065 CenAJ000177 1.9 der4 9.281 1 3.011 CenAJ000174 11.0 der11 9.655 1 2.543 CenAJ000174 11.0 der4 9.832 1 2.548 CenET 2.9 der11 16.180 1–2 5.048 TelET 2.9 der4 16.189 1–2 5.530 TelAJ000167 4.6 der11 29.650 2 6.601 TelAJ000167 4.6 der4 29.572 2 6.644 TelAJ000171 1.2 der11 30.606 2 3.751 CenAJ000171 1.2 der4 30.583 2 3.794 CenAJ000178 10.0 der11 31.446 2 2.806 CenAJ000178 10.0 der4 31.098 2 3.450 CenAJ000172 1.7 der11 31.234 2 2.733 CenAJ000172 1.7 der4 ?d ? ? ?SEM° 5.0 der11 32.911 2 2.388 CenSEM° 5.0 der4 32.595 2 2.770 CenRS4;11° 33.0 der11 36.570 2 2.520 CenRS4;11° 33.0 der4 36.562 2 2.556 CenAJ000168 3.1 der11 43.655 3 763 CenAJ000168 3.1 der4 43.840 3 724 CenMV4;11° 10.0 der11 43.792 3 1.128 CenMV4;11° 10.0 der4 44.041 3 1.128 CenAJ000169 5.1 der11 45.213 3 6.330 TelAJ000169 5.1 der4 45.087 3 6.481 TelHH 48.0 der11 47.446 3 2.510 CenHH 48.0 der4 47.585 3 3.526 CenCB 15.0 der11 47.594 3 5.463 TelCB 15.0 der4 47.868 3 5.986 Tela Der chr, derivative chromosome; Cen/tel, centromeric and telomeric localization.b Breakpoint location; AF-4 nucleotide positions according to Genbank entry
AJ238093.c Breakpoint location;MLL nucleotide positions are numbered according to the BamHI
fragment (8.321 nt) spanning the breakpoint cluster region of the humanMLL gene (partof Genbank entry X83604).
d Breakpoint not analyzed.Table 2 Alu repetitive elements and consensus sequences in the breakpoint cluster
region of the human AF-4 gene
Alu location (o/la) Topo II (o/mis) nona-/heptamer (t/mis)
7298–7613 (ccw, 316) 697–714 (ccw, 1) 16982–17020 (9-23-7, 5)7992–8311 (cw, 320) 2037–2054 (ccw, 1) 22026–22053 (9-12-7, 5)
10705–11035 (ccw, 331) 2129–2146 (ccw, 1) 22460–22498 (7-23-9, 4)16426–16760 (cw, 335) 3053–3070 (ccw, 1) 22460–22487 (7-12-9, 4)19101–19435 (ccw, 335) 3798–3815 (cw, 1) 23115–23153 (9-23-7, 5)20343–20474 (cw, 132) 5220–5237 (cw, 1) 24831–24869 (7-23-9, 5)20482–20778 (cw, 297) 6835–6852 (cw, 1) 24831–24858 (7-12-9, 4)20784–20927 (cw, 144) 7344–7361 (cw, 1) 28062–28100 (7-23-9, 5)22209–22499 (cw, 290) 8548–8565 (ccw, 1) 37191–37229 (9-23-7, 5)25701–25826 (ccw, 126) 9680–9697 (ccw, 1) 44172–44199 (9-12-9, 5)25834–26112 (ccw, 279) 10644–10661 (cw, 0)26120–26339 (ccw, 220) 14193–14210 (cw, 1)27966–28266 (cw, 301) 15684–15701 (cw, 1)28406–28720 (ccw, 315) 18147–18164 (cw, 1)37021–37327 (ccw, 307) 18271–18288 (ccw, 1)38457–38586 (ccw, 130) 18628–18645 (cw, 1)39692–39891 (ccw, 200) 19818–19835 (cw, 1)40116–40471 (cw, 356) 21009–21026 (ccw, 1)41614–41899 (ccw, 286) 24301–24318 (ccw, 1)44620–44899 (ccw, 280) 24859–24876 (ccw, 1)46597–46904 (cw, 308) 25927–25944 (cw, 1)46981–47293 (cw, 313) 27529–27546 (cw, 1)47863–48164 (cw, 302) 33222–33239 (ccw, 1)50975–51286 (ccw, 312) 33751–33768 (cw, 1)52250–52383 (ccw, 134) 34238–34255 (ccw, 1)52631–52756 (ccw, 126) 34387–34404 (ccw, 1)
35977–35994 (cw, 1)43266–43283 (cw, 0)43784–43801 (ccw, 1)44265–44282 (ccw, 1)46364–46381 (cw, 1)50468–50485 (cw, 1)
a o/l, orientation and length; Topo II, topoisomerase II; o/mis, orientation and numberof mismatches; t/mis, type and number of mismatches; ccw, counter clockwise; cw,clockwise.
3359
CHROMOSOMAL TRANSLOCATIONS T(4;11)
Research. on November 23, 2015. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
breakpoint cluster nt 10.779; AF029700: der4-bp at nt 4.500;AF024540: der11-bp at nt 19.487; AF024543: der11-bp at nt 10.853;S79285: der11-bp at nt 33.950; S79287: der4-bp at nt 33.947;AJ000170: der11-bp at nt 28.033; AJ000179: der11-bp at nt 35.611;AJ000180: der11-bp at nt 36.691, fused to the inverted region of nt36.691 to 36.734].
Discussion
Two major advances follow from the results of this study: (a) animproved method allowing investigators to diagnose and monitorpatients with t(4;11)-ALL more rapidly and more reliably than it waspossible so far; and (b) an improved ability to carry out the retrospec-tive analysis of breakpoints in biopsy material and leukemia-derivedcell lines. The results of this study provide investigators with anincreased probability of actually finding the breakpoint in each indi-vidual sample and of correctly reconstructing the detailed recombi-nation events that may have led to a particular translocation. Bothadvances are achieved by using an optimally suited pair of PCRprimers and the PCR amplification of genomic DNA for the analysisof each individual breakpoint. A method for the rapid assignment ofthe optimally suited primer pair for individual breakpoints is pre-sented in this study.
For routine clinical diagnosis and monitoring of t(4;11) break-points, the preferred method, so far, was the RT-PCR method basedon the amplification of breakregion mRNAs and corresponding first-strand cDNAs by the use of mixed primer pairs with one primer eachderived from exon sequences of theMLL and AF-4 translocationpartner genes (9). This method has the advantage of generating rapidinformation about the location of the breakpoints to the nearest exon,irrespective of their precise location within the large introns in thebreakpoint cluster region of theAF-4 gene (15). The disadvantages ofthis method are that it requires a significant amount of cells (105 to107) and that these cells must be sufficiently fresh and well preservedto guarantee the integrity of the breakregion mRNA. Due to therelatively large amounts of cells required, this method was oftendifficult to apply for the purpose of monitoring MRD during thecourse of a therapy. RT-PCR methods and, in particular, nestedRT-PCR methods also sometimes generated results that were difficultto interpret, pointing to the existence of breakpoint RNA species incases in which no cytogenetic evidence for a t(4;11) translocation wasobserved (10). The method presented here allows the assignment ofbreakpoints with comparable speed as RT-PCR methods (i.e., withina couple of hours). It requires fewer cells, and the quality of the biopsymaterial is less critical than for RT-PCR analysis because genomic
Fig. 2. Patient breakpoint junctions identified by genomic PCR. A,left, PCR amplimers for patient ET:Lanes 1and6, l phage size standard;Lane 2, MLLx23 AF4y13; Lane3, MLLx2 3 AF4y15; Lane 4, MLLx23 AF4y19; Lanes 5and10, negative control;Lane 7, AF4x103 MLLy7; Lane 8, AF4x113 MLLy7; Lane 9, AF4x123 MLLy7. Secondpanel from left, DNA digests for patient ET:Lane a:undigested der11 amplimer;Lanes b–d:der11 amplimer digested with EcoRI, HindIII, EcoRI/HindIII;Lane e,l phage sizestandard;Lane f, undigested der4 amplimer;Lanes g–i:der4 amplimer digested with EcoRI, HindIII, EcoRI/HindIII.Third panel from left, PCR amplimers for patient PB:Lanes 1and6: l phage size standard;Lane 2:MLLx2 3 AF4y17; Lane 3:MLLx2 3 AF4y18; Lane 4:MLLx2 3 AF4y19; Lanes 5and10: negative control;Lane 7:AF4x14 3 MLLy7;Lane 8:AF4x15 3 MLLy7; Lane 9:AF4x16 3 MLLy7. Right, DNA digests for patient PB:Lane a, undigested der11 amplimer;Lanes b–d:der11 amplimer digested with EcoRI,HindIII, EcoRI/HindIII; Lane e,l phage size standard;Lane f, undigested der4 amplimer;Lanes g–i:der4 amplimer digested with EcoRI, HindIII, EcoRI/HindIII. Primers used forthe amplification process are specified in “Materials and Methods.”B, DNA sequences of reciprocal breakpoints in the der11 and der4 chromosomes, respectively.Dots, breakpoints.Filler DNA is underlined.MLL sequences are shown in upper case letters;AF-4sequences are shown in lower case letters.C, model of recombinedMLL andAF-4alleles.Top, parentalMLL allele (white boxes);bottom, parental AF-4 allele (grey boxes);middle, der11 and der4 alleles after recombination;■, presence of filler DNA;letters, wild-type DNA fragments.For patients E. T. and P. B., deletion of fragments B. DuplicatedAF-4 DNA is present in both derivative alleles of both patients.
3360
CHROMOSOMAL TRANSLOCATIONS T(4;11)
Research. on November 23, 2015. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

DNA is degraded less rapidly than breakregion RNA. Therefore,poorly preserved samples that may no longer be suitable for aRT-PCR analysis may still give useful results with the methodpresented here. Because it requires fewer cells, this method mayalso allow MRD monitoring in a greater number of cases thanconventional RT-PCR methods. Continuous complete remission ofpatients with t(4;11)-ALL so far has only been observed in cases inwhich RT-PCR-based MRD monitoring did not detect any breakre-gion transcripts. Patients scoring positive by RT-PCR did notremain in complete remission (9). The method presented here canbe used to obtain an independent verification of RT-PCR resultswith equal speed and sensitivity. In some cases this is likely torepresent a valuable advantage. This may particularly apply tocases where RT-PCR methods indicated a rearrangement in theabsence of cytogenetic evidence for a t(4;11) translocation (10). Insuch cases, the demonstration of a rearrangement on the genomicDNA level by the methods presented here may add decisiveadditional evidence.
The application of the method described here to a sizeable cohort oft(4;11) leukemias generated useful new information. First, the distri-bution of chromosomal breakpoints (Fig. 1 and Table 1) suggests anonrandom occurrence of translocation-breaks within theAF-4 gene.Preferred sites (clusters) of breakpoints were detected within the48-kb breakpoint cluster region of theAF-4 gene, which is locatedbetween exons 3 and 7 (Fig. 1). A total of 37 breakpoints weremapped in this region, including 33 assigned in previous studies (11,12). Ten of these breakpoints fell into cluster 1 (;6 kb), 13 intocluster 2 (;9 kb), and 10 constituted cluster 3 (;5 kb). Only thenewly assigned breakpoints for patients P. B. and E. T. fell outside ofthese clusters (Fig. 1). Together, these three clusters covered 22 of the48 kb of the breakpoint region. The DNA in these clusters was foundto contain significantly fewer repetitive Alu elements than the remain-der of the breakpoint region. Only nine Alu elements (.250 bp) werefound within or overlapping these three clusters, whereas eight Aluelements plus nine Alu remnants (,250 bp) were found in theremainder of the breakpoint region (Fig. 1 and Table 2). Only thebreakpoint of patient C. B. on the derivative 4 chromosome (nt 47.869in theAF-4breakpoint cluster region) fell directly into an Alu element(nt 47.863–48.164); all other 36 breakpoints were located in nonre-petitive DNA sequences (Tables 1 and 2). Therefore, these new dataconfirm and extend the conclusion previously drawn, that recombi-nation between Alu elements is unlikely to be the mechanism respon-sible for a major fraction of t(4;11) translocations (11, 12). Recom-bination between Alu elements is important, however, in thegeneration of internal duplications within theMLL gene (19) anddeletions in the humanBRCA1andhMLH1 genes (20, 21).
Only one breakpoint was found to coincide with a consensusrecognition sequence for topoisomerase II (17). This was thebreakpoint on the derivative 11 chromosome of the cell lineMV4;11 (nt 43.793). None of the remaining 36 breakpoints (Fig. 1and Table 1) coincided with topoisomerase II binding sites orsignal sequences by V(D)J recombinases (Table 2). Thus, the newdata obtained here confirmed and extended previously publishedconclusions, that the actions of topoisomerase II and V(D)J recom-binases are unlikely to be a major component in the events gen-erating t(4;11) translocations (12).
For one patient (AJ000172; Table 1), only the breakpoint on thederivative 11, but not the breakpoint on the derivative 4, chromosomewas obtained by applying the new methods described above. Thisresult was unexpected because a derivative 4 breakregion transcriptwas identified by RT-PCR analysis of biopsy material from the same
patient.5 A similar result was obtained for patient L. L. M. and the cellstrains derived from this patient (11). The breakpoint on the derivative11 chromosome of this patient was found to reside outside of thepresently known breakpoint cluster region of theAF-4gene. The mostplausible explanation of these findings is that in a few rare cases thetranslocation breakpoints cannot be definitively located by the meth-ods presented here, presumably due to incomplete characterized par-tial or total duplications of sequences from the wild-typeAF-4 gene.5
Another interesting set of observations made using the methodsdescribed above relates to the distribution of translocation breakpointswithin the MLL and AF-4 genes and the correlation between thelocation of breakpoints and the age of the patient at the time ofdiagnosis. It has been suggested recently that early infant leukemiashave a preferred breakpoint distribution in the 39-terminal portion ofthe MLL breakpoint cluster region (22, 23). When we analyzed thecurrent set of data, there was a difference between the distribution ofbreakpoints of patients below or above 1 year of age, at least for thebreakpoints in theMLL gene. Three of four patients below or around1 year of age have breakpoints in the telomeric part of theMLLbreakpoint cluster region and have their breakpoints predominantly inAF-4 cluster 1 (two of three) or cluster 2 (one of three). Eight of 12older patients have their breakpoints in the centromeric portion ofMLL and their corresponding breakpoints distributed in theAF-4clusters 1 (3 of 8), 2 (3 of 8), and 3 (2 of 8). Older patients withbreakpoints in the telomeric portion of theMLL breakpoint clusterregion (4 of 12) have their breakpoints inAF-4 cluster 1–2 (1 of 4),2 (1 of 4), or 3 (2 of 4). This may reflect a specific tendency; however,more data are necessary to validate this observation as a real signif-icant difference.
Finally, the breakpoints of both patients that were reported here forthe first time (patients E. T. and P. B.) showed similar characteristicfeatures as the previously published breakpoints (11, 12). Both werenot balanced at the fine-structure level and showed deletions andduplications of sequences at the breakpoint. For patients E. T. andP. B.,MLL sequences were deleted (482 bp and 17 bp, respectively),andAF-4 sequences were duplicated (9 bp and 222 bp, respectively)at the breakpoints (Figure 2,B andC). Filler DNA segments of 2 bpand 4 bp were also detected at the breakpoints on the derivative 4 andderivative 11 chromosomes, respectively, for both patients. The pres-ence of duplications, inversions, deletions, filler DNA and mini-directrepeat sequences at the translocation breakpoints has previously beenreported and was interpreted to indicate that these translocations werelikely to result from DNA damage and the subsequent action of theerror-prone DNA repair pathway (12). The error-prone pathway isonly one of several known DNA-repair mechanisms, and it is not themost frequently used pathway, but a default option that is mainly usedwhen the major pathways are inoperative or impaired. The finding oftell-tale signs for the action of the error-prone pathway rather than themajor repair pathways at t(4;11) translocation breakpoints indicatesthat t(4;11) translocations may preferentially occur in cells in whichthe major DNA repair pathway is impaired. This is an attractive novelhypothesis, and the new methods presented here provide improvedtools for testing its predictions in a large number of leukemia cases.
Acknowledgments
Pieter de Jong and P. Ioannou are gratefully acknowledged for constructionof the RPCI 1, 3-5 Human PAC library at Roswell Park Cancer Institute(Buffalo, NY). We thank Drs. Panzer-Grumayer and Haas from the ChildrensHospital for providing patient biopsy material.
5 Unpublished data.
3361
CHROMOSOMAL TRANSLOCATIONS T(4;11)
Research. on November 23, 2015. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

References
1. Rabbitts, T. H. Chromosomal translocations in human cancer. Nature (Lond.),372:143–149, 1994.
2. Ziemin van der Poel, S., McCabe, N. R., Gill, H. J., Espinosa, R., III, Patel, Y.,Harden, A., Rubinelli, P., Smith, S. D., LeBeau, M. M., Rowley, J. D., and Diaz,M. O. Identification of a gene,MLL, that spans the breakpoint in 11q23 translocationsassociated with human leukemias. Proc. Natl. Acad. Sci. USA,88: 10735–10739,1991.
3. Gu, Y., Nakamura, T., Alder, H., Prasad, R., Canaani, O., Cimino, G., Croce, C. M.,and Canaani, E. The t(4;11) chromosome translocation of human acute leukemiasfuses theALL-1 gene, related to Drosophila trithorax, to theAF-4 gene. Cell,71:701–708, 1992.
4. Bernard, O. A., and Berger, R. Molecular basis of 11q23 rearrangements in hema-topoietic malignant proliferations. Genes Chromosomes Cancer,13: 75–85, 1995.
5. Marschalek, R., Nilson, I., Lochner, K., Greim, R., Siegler, G., Greil, J., Beck, J. D.,and Fey, G. H. The structure of the humanALL-1/MLL/HRXgene. Leuk. Lymphoma,27: 417–428, 1997.
6. Waring, P. M., and Cleary, M. L. Disruption of a homolog of trithorax by 11q23translocations: leukemogenic and transcriptional implications. Curr. Top. Microbiol.Immunol.,220: 1–23, 1997.
7. Ford, A. M., Ridge, S. A., Cabrera, M. E., Mahmoud, H., Steel, C. M., Chan, L. C.,and Greaves, M. In utero rearrangements in the trithorax-related oncogene in infantleukemias. Nature (Lond.),363: 358–360, 1993.
8. Canaani, E., Nowell, P. C., and Croce, C. M. Molecular genetics of 11q23 chromo-some translocations. Adv. Cancer Res.,66: 213–234, 1995.
9. Cimino, G., Elia L., Rivolta, A., Rapanotti, M. C., Rossi, V., Alimena, G., Annino, L.,Canaani, E., LoCoco, F., and Biondi, A. Clinical relevance of residual diseasemonitoring by polymerase chain reaction in patients with ALL-1/AF-4 positive-acutelymphoblastic leukemia. Br. J. Haematol.,92: 659–664, 1996.
10. Uckun, F. M., Herman-Hatten, K., Crotty, M. L., Sensel, M. G., Sather, H. N.,Tuel-Ahlgren, L., Sarquis, M. B., Bostrom, B., Nachman, J. B., Steinherz, P. G.,Gaynon, P. S., and Heerema, N. Clinical significance of MLL-AF4 fusion transcriptexpression in the absence of a cytogenetically detectable t(4;11)(q21;q23) chromo-somal translocation. Blood,92: 810–821, 1998.
11. Reichel, M., Gillert, E., Nilson, I., Siegler, G., Greil, J., Fey, G. H., and Marschalek,R. Fine structure of translocation breakpoints in leukemic blasts with chromosomaltranslocation t(4;11): the DNA damage-repair model of translocation. Oncogene,17:3035–3044, 1998.
12. Gillert, E., Leis, T., Repp, R., Reichel, M., Hosch, A., Breitenlohner, I., Angermuller, S.,Borkhardt, A., Harbott, J., Lampert, F., Griesinger, F., Greil, J., Fey, G. H., andMarschalek, R. A DNA damage repair mechanism is involved in the origin of chromo-somal translocations t(4;11) in primary leukemic cells. Oncogene, in press, 1999.
13. Leis, T., Repp, R., Borkhardt, A., Metzler, M., Schlager, F., Harbott, J., and Lampert,F. A new fingerprint method for sequence analysis of chromosomal translocations atthe genomic DNA level. Leukemia (Baltimore),12: 758–763, 1998.
14. Marschalek, R., Greil, J., Lochner, K., Nilson, I., Siegler, G., Zweckbronner, I., Beck,J. D., and Fey, G. H. Molecular analysis of the chromosomal breakpoint and fusiontranscripts in the acute lymphoblastic SEM cell line with chromosomal translocationt(4;11). Br. J. Haematol.,90: 308–320, 1995.
15. Nilson, I., Lochner, K., Siegler, G., Greil, J., Beck, J. D., Fey, G. H., and Marschalek,R. The exon/intron structure of the human ALL-1 (MLL) gene involved in translo-cations to chromosomal region 11q23 and acute leukemias. Br. J. Haemotol.,93:966–972, 1996.
16. Nilson, I., Reichel, M., Ennas, M. G., Greim, R., Knorr, C., Siegler, G., Greil, J., Fey,G. H., and Marschalek, R. The exon/intron structure of the humanAF-4 gene, amember of theAF-4/LAF-4/FMR-2gene family coding for a nuclear protein withstructural alterations in acute leukemia. Br. J. Haematol.,98: 157–169, 1997.
17. Spitzner, J. R., and Muller, M. T. A consensus sequence for cleavage by vertebrateDNA topoisomerase II. Nucleic Acids Res.,16: 5533–5556, 1988.
18. Sakano, H., Huppi, K., Heinrich, G., and Tonegawa, S. Sequences at the somaticrecombination sites of immunoglobulin light-chain genes. Nature (Lond.),280:288–294, 1979.
19. Strout, M. P., Marcucci, G., Bloomfield, C. D., and Caligiuri, M. A. The partialtandem duplication of ALL1 (MLL) is consistently generated by Alu-mediatedhomologous recombination in acute myeloid leukemia. Proc. Natl. Acad. Sci. USA,95: 2390–2395, 1998.
20. Puget, N., Torchard, D., Serova-Sinilnikova, O. M., Lynch, H. T., Feunteun, J.,Lenoir, G. M., and Mazoyer, S. A 1-kb Alu-mediated germ-line deletion removingBRCA1 exon 17. Cancer Res.,57: 828–831, 1997.
21. Mauillon, J. L., Michel, P., Limacher, J. M., Latouche, J. B., Dechelotte, P.,Charbonnier, F., Martin, C., Moreau, V., Metayer, J., Paillot, B., and Frebourg, T.Identification of novel germline hMLH1 mutations including a 22 kb Alu-mediateddeletion in patients with familial colorectal cancer. Cancer Res.,56: 5728–5733,1996.
22. Strissel-Broeker, P. L., Super, H. G., Thirman, M. J., Pomykala, H., Yonebayashi, Y.,Tanabe, S., Zeleznik-Le, N., and Rowley, J. D. Distribution of 11q23 breakpointswithin the MLL breakpoint cluster region in de novo acute leukemia and in treatment-related acute myeloid leukemia: correlation with scaffold attachment regions andtopoisomerase II consensus binding sites. Blood,87: 1912–1922, 1996.
23. Cimino, G., Rapanotti, M. C., Biondi, A., Elia, L., Lo Coco, F., Price, C., Rossi, V.,Rivolta, A., Canaani, E., Croce, C. M., Mandelli, F., and Greaves, M. Infant acuteleukemias show the same biased distribution ofALL1 gene breaks as topoisomeraseII-related secondary acute leukemias. Cancer Res.,57: 2879–2883, 1997.
3362
CHROMOSOMAL TRANSLOCATIONS T(4;11)
Research. on November 23, 2015. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from

1999;59:3357-3362. Cancer Res Martin Reichel, Esther Gillert, Ina Breitenlohner, et al. Basic and Clinical Researchwith t(4;11) Acute Lymphoblastic Leukemia: Implications for Rapid Isolation of Chromosomal Breakpoints from Patients
Updated version
http://cancerres.aacrjournals.org/content/59/14/3357
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/59/14/3357.full.html#ref-list-1
This article cites 22 articles, 8 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/59/14/3357.full.html#related-urls
This article has been cited by 3 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
Research. on November 23, 2015. © 1999 American Association for Cancercancerres.aacrjournals.org Downloaded from
Copyright © 2022 FDOKUMEN