
Estimating the value of point-of-care HPV testing in three low

A new multiparameter assay to assess HPV 16/18, viral loadand physical status together with gain of telomerase genes inHPV-related cancers
Wendy Theelen1, Martin Reijans2, Guus Simons2, Frans C.S. Ramaekers1, Ernst-Jan M. Speel1 and Anton H.N. Hopman1
1 Department of Molecular Cell Biology, GROW—School for Oncology and Developmental Biology, Maastricht University Medical Center, The Netherlands2 PathoFinder BV, Maastricht, The Netherlands
Oncogenic human papillomavirus (HPV) is the most important risk factor for cancer of the uterine cervix and a subgroup of
head and neck cancers. Viral load has been associated with persistence of infection, whereas integration of HPV into the host
cell genome is associated with transition to invasive disease. Viral integration is frequently correlated with loss of viral E2
and gain of the telomerase-related genes TERC and TERT. The objective of this study was to develop a rapid and sensitive
multiplex ligation-dependent probe amplification (MLPA) assay for the simultaneous analysis of viral load, integration and
copy number gain of TERC and TERT in HPV16/18-associated lesions. The performance of the assay was tested for HPV vs.
human gene copy number ratios ranging from 0.1 to 100 and for percentages of integration ranging from 0 to 100%. The
model systems used include plasmid mixtures and the HPV-positive cell lines SiHa, HeLa and CaSki described to contain a
range of 2–600 viral copies per cell. In samples with low-viral load, viral integration can be reliably determined when more
than 30% of the virus is integrated. Gain of the telomerase-related genes in the cell lines as determined by our MLPA assay
was in accordance with data reported in the literature. Our study demonstrates that within a single MLPA-reaction viral type,
load, integration and gain of TERC and TERT can be reliably determined, which will improve risk assessment for patients
suspected for HPV infection.
Human papillomavirus (HPV) plays a causal role in the de-velopment of several cancer types1 and has been identified inthe majority of uterine cervical and anal cancer cases2,3 andin approximately half of the tonsil and penile cancers.4,5
Most HPV-related cervical intraepithelial precursor lesionsare known to regress spontaneously, which implicates thatalthough HPV is a necessary cause, it is not independentlycausative for cancer. Additional factors play a role in the pro-gression and/or prognosis of these (pre)malignant lesions,including, e.g., individual susceptibility, integration of thevirus and genomic instability, as described later.
In the past years, many studies have focused on the iden-tification of markers that can predict the progression of cervi-cal intraepithelial neoplasia to cervical cancer, which maythus help to discriminate regressive from progressive HPV-related lesions. The markers include HPV type,4,6 viral load7,8
and physical status9,10 and gain of telomerase-relatedgenes.11–14 HPV16 and 18 are the most prevalent oncogenictypes in cervical cancer and are found to be responsible formore than 70% of cervical carcinomas.15,16 In tonsillar squa-mous cell carcinomas, HPV16 is predominantly found inHPV-positive cases.4 Furthermore, in cervical lesions a highHPV16 viral load is more likely to lead to a persistent infec-tion than a low-viral load,7,17,18 which in turn enhances thelikelihood of progression to cancer.
It has been found that part of the HPV genome will belost on integration into the host genome. This predomi-nantly concerns the E2 gene but may also include the E1 orL1 gene.3,19,20 Some studies have demonstrated that there isa very wide range of integration-related disruption sites inthe viral genome.20–22 Most of these deletions will causesilencing of the E2 gene, which in turn will lead to a
Key words: uterine cervical cancer, head and neck cancer, viral
integration, TERC, TERT
Additional Supporting Information may be found in the online
version of this article
Abbreviations: HPV: human papillomavirus; MLPA: multiplex
ligation-dependent probe amplification; TERC: Telomerase RNA
Component; TERT: Telomerase Reverse Transcriptase; FISH:
fluorescence in situ hybridization; CGH: comparative genomic
hybridization; MSH2: MutS homolog 2; bp: basepairs; FAM:
carboxyfluorescein; CNR: copy number ratio; qPCR: quantitative
PCR
W. Theelen, F.C.S. Ramaekers, E.J.M. Speel and A.H.N. Hopman
declare that there are no conflicts of interest
Grant sponsors: The Transnational University Limburg (Maastricht,
The Netherlands) and LIOF (Limburg Development and Investment
Company, Maastricht, The Netherlands) through PathoFinder BV
DOI: 10.1002/ijc.24844
History: Received 8 May 2009; Accepted 30 Jul 2009; Online 26
Aug 2009
Correspondence to: Wendy Theelen, Department of Molecular Cell
Biology (UNS50-box 17), Maastricht University Medical Center, P.O.
Box 616, Maastricht 6200 MD, The Netherlands,
Fax: þ31-43-388-4151, E-mail: [email protected]
Early
Detection
andDiagn
osis
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC
International Journal of Cancer
IJC

consistent overexpression of E6 and E7 gene products thatwill degrade p53 and inactivate pRb, respectively.23 It hasbeen described that integration and the size of the deletedfragment correlates with severity of the lesion and diseaseprogression.24–27 Therefore, the physical status of HPV (i.e.,integrated vs. episomal) can be an indicator for progressionto cancer.
Chromosomal aberrations found in different cancer stageshave been used to predict cancer progression. One of theaberrations frequently identified during HPV-related carcino-genesis of the cervix and the oropharyngeal mucosa is a gainof chromosome 3q, with Telomerase RNA Component(TERC) as 1 of the suggested candidate genes.12,28 Anotherfrequently found aberration involves chromosome 5p con-taining the Telomerase Reverse Transcriptase (TERT) gene.In most cases gain of this region is reported to be associatedwith progression to cancer.13,29–31
Several assays are currently available to analyze 1 ormore of the aforementioned markers. For example, PCR forHPV E2,20,21 in some cases combined with PCR for HPVE6,32 is frequently used to determine HPV physical status.When a real-time PCR analysis is performed for these tar-gets also, viral load can be determined.33 Additional testsuse HPV L1 PCR combined with reverse line blotting34 orsequencing35 for HPV typing, the detection of integratedpapillomavirus sequences36 or amplification of papillomavi-rus oncogene transcripts10 assay to determine HPV physicalstatus, fluorescence in situ hybridization (FISH) to identifyHPV type and physical status and chromosomal aberra-tions,27 and (array) comparative genomic hybridization(CGH)13,37 or molecular allelotyping11 to screen for chro-mosomal aberrations.
Aim of the underlying study was to develop a reliable andsensitive HPV multiplex ligation-dependent probe amplifica-tion (MLPA) assay for the simultaneous detection of HPVtype (16 and 18), viral load, physical status of the virus andgenomic instability as monitored by gain of TERC and TERTgene copies. The HPV assay described here combines theprinciples of 2 MLPA assays described earlier, 1 for thequantification of up to 40 human DNA sequences with ahigh specificity in a single PCR assay38 and the secondMLPA assay for viral detection.39
Material and MethodsCell lines
Human uterine cervical cancer cell lines (SiHa, HeLa andCaSki), human breast cancer cell lines (MCF7 and T47D),the human intestinal epithelial cell line CaCo-2 and thehuman immortalized nontumorigenic keratinocyte cell lineHaCaT were obtained from the American Type Culture Col-lection (ATCC, Manassas, VA). All cell lines were grownaccording to the supplier’s recommendations, and DNA wasextracted using the QIAamp DNA Mini Kit (Qiagen, Hilden,Germany) according to protocol.
Clinical samples
The following clinical samples were analyzed by MLPA andthe results verified by quantitative PCR (qPCR) and/or FISH:
1. Fresh-frozen tissue samples of 5 cases, including 1 sam-ple of normal cervix and 4 (pre)malignant samples thatwere selected to contain >60% of tumor cells. TheMLPA results for these samples were verified by bothqPCR and FISH.
2. Cytological samples of 7 cases, ranging from normal todyskaryotic and for which the presence of HPV wasknown based on GP5þ/6þ testing,34 were verified byqPCR.
3. Formalin-fixed, paraffin-embedded tissue samples of 3cases, which were stained for p16 as a surrogate markerfor HPV positivity,40 were verified by FISH.
Because of day to day experimental variation referencesamples, i.e., episomal samples and plasmid mixtures repre-senting 30% integrated HPV, were included in each experi-ment. To determine whether a clinical sample contained inte-grated HPV, the E2/E6 signal intensity ratios for the clinicalsamples were compared with the ratios for the matching ref-erence samples. Furthermore, all clinical samples are analyzedin duplicate and mixed or integrated HPV and gain forTERT and TERC are only reported when it is detected inboth duplicate measurements. This study was carried out inaccordance with local ethical guidelines.
Design and preparation of preamplification primers
and MLPA probes
For the design of the preamplification primers and thehybridizing part of the probes, sequence information avail-able from public databases was used, including the Nucleo-tide database from the National Center of Biotechnology In-formation (NCBI; http://www.ncbi.nlm.nih.gov). In additionto the Basic Local Alignment and Search Tool analysis pro-gram (NCBI), BioEdit (http://www.mbio.ncsu.edu/BioEdit/BioEdit.html) was used for sequence alignments and Primer3design software (Primer3 v 0.4.0; http://frodo.wi.mit.edu/) forprimer design.
To identify type-specific HPV16 and 18 regions for thedesign of the preamplification primers and the hybridizingpart of the MLPA probes, alignments were performed withthe sequences of all HPV types. All primers and probes wereevaluated by performing a Basic Local Alignment and SearchTool analysis against the NCBI database and were approvedwhen no mismatch was found for all known viral subtypeand variant sequences within the critical regions of the pre-amplification primers (i.e., no mismatch at the 30 end of aprimer) and MLPA probes (i.e., no mismatch within 5 nucle-otides from the ligation site). A similar approach was chosenfor the design of the preamplification primers and MLPA
Early
Detection
andDiagn
osis
960 MLPA assay for HPV-related cancers
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

probes for the human targets, i.e., b-globin, MSH2 (MutShomolog 2), TERC and TERT.
HPV probes were developed to assess HPV type, load andphysical status (Supporting Information Table 1). Because theE6 and E7 genes are nearly always present in all HPV-relatedlesions, regardless of the physical status, probes against thesegenes were designed for typing of HPV16 and 18. For thedetection of the physical status 2 E2 probe sets per HPV typewere developed, which target the sequences most frequentlydeleted on integration into the human genome. The distancebetween the 2 E2 target sequences is 600 base pairs (bp) and282 bp for HPV16 and 18, respectively. For the estimation ofviral load, 7 human probe sets were developed recognizingtarget sequences in the b-globin (2 loci), the MSH2 (1 locus),the TERT (2 loci) and the TERC gene (2 loci). The b-globingene was included in this assay because it has previouslybeen described as a reference to determine viral load in real-time PCR.41 MSH2 was selected as a reference because chro-mosome 2p numbers remain relatively stable in HPV-relatedcancers.13,29,30,42 In addition, 4 probe sets were designed totarget sequences within TERC (3q26) and TERT (5p15),which were also used to assess gain of the telomerase-associ-ated genes. An overview of the used target sequences is pro-vided in Supporting Information Table 1.
The preamplification primers were designed to anneal asclose as possible to the 50 and the 30 end of the aforemen-tioned probe targets. For each target, 4 forward and 4 reversepreamplification primers were designed, which were tested inall possible combinations, resulting in 16 primer combina-tions per target and a total of 240 primer combinations. Foreach target the specificity of all combinations was determinedand 4 specific combinations resulting in the most optimalPCR amplification were selected for the second test. In thistest, the most sensitive primer combination was determinedthrough the use of 5 dilutions of DNA, i.e., 5 ng; 500 pg; 50pg; 5 pg and 500 fg in the 20-ll PCR reaction mixture forthe human targets and 12.5 fg; 1.25 fg; 125 ag; 12.5 ag and1.25 ag for the HPV targets. The combination with the high-est sensitivity was selected for each target. Of the 240 initiallydesigned primer combinations, the 15 combinations thatwere selected based on highest sensitivity and specificity arepresented in Supporting Information Table 2. The primerswere designed to produce preamplification fragments with amaximum of 166 (bp) in length.
Each MLPA probe set consists of 1 short synthetic oli-gonucleotide and 1 phage M13-derived, long oligonucleo-tide (Fig. 1) and gives rise to an amplification product ofunique size between 129 and 488 bp. The phage M13-derived, long oligonucleotides were prepared as previouslydescribed,38 and the short synthetic oligonucleotides andpreamplification primers were synthesized by Biolegio(Malden, The Netherlands). For storage, the oligonucleo-tides were diluted in TE-buffer (10 mM Tris-HCl pH 8.0,1 mM EDTA) to a concentration of 100 lM, which wasused as stock solution.
The specificity of the primer and probe mix for HPV16and 18 was evaluated by tests on HPV16 or 18 plasmidmodel systems, cell lines containing HPV16 or 18 and clini-cal samples containing closely related HPV types, i.e.,HPV31, 33 and 45 (data not shown), as determined with thePapilloCheck (Greiner Bio-One GmbH, Frickenhausen,Germany).
Preamplification
A multiplex preamplification PCR was performed using theQiagen Multiplex PCR kit (Qiagen, Hilden, Germany). A20-ll reaction mixture contained Qiagen mastermix (MgCl2final concentration 3 mM; dNTPs and HotstarTaq DNA po-lymerase), multiplex primer mix (final concentration 20 nMfor each forward primer and 200 nM for each reverse primer)and 10 ng of sample DNA. Amplification was performed on aBiometra T1 Thermocycler (Biometra, Gottingen, Germany)as follows: 15 min at 95�C, followed by 20 cycles of each30 sec at 94�C, 90 sec at 55�C and 90 sec at 72�C and a finalextended elongation step for 10 min at 72�C.
MLPA analysis
The MLPA analysis of the preamplified product was per-formed as described by Schouten et al.38 with minor modifi-cations. The preamplified product was diluted 5 times usingsterile water, after which 2 ll was mixed with 1.5 ll MLPA-buffer (1.5 M KCl, 300 mM Tris-HCl pH 8.5, 1 mM EDTA;MRC-Holland, Amsterdam, the Netherlands), 1.5 ll probemix (3 fmol of each synthetic probe oligonucleotide and 1.5fmol of each M13-derived oligonucleotide in TE) and 3 llsterile water. After a 5-min denaturation step at 98�C in aBiometra T1 Thermocycler with a heated lid, the mixturewas incubated for 16 hr at 60�C. For ligation, this mixturewas diluted to 40 ll with ligation buffer (2.6 mM MgCl2,5 mM Tris-HCl pH 8.5, 0.013% nonionic detergents, 0.2 mMnicotinamide adenine dinucleotide) containing 1 U heat-sta-ble Ligase-65 enzyme (MRC-Holland, Amsterdam, theNetherlands) and incubated at 54�C for 15 min, followed byligase inactivation at 98�C for 5 min. Four microliters of thismixture was added to 16 ll of PCR mixture containingdNTPs (2 mM each, Fermentas, St. Leon-Rot, Germany), 1 UTaq-polymerase (MRC-Holland, Amsterdam, the Nether-lands), 1� PCR buffer (50 mM KCl, 10 mM Tris-HCl pH8.5, 1.6 mM MgCl2) and 4 pmol of the 2 MLPA-PCR pri-mers each, with the forward primer 50-GTGGCAGGGCGC-TAC GAACAA-30 labeled with carboxyfluorescein (FAM),and the reverse primer 50-GGACGCGCCAGCAAGATC-CAATCT AGA-30. Amplification was performed on a Biome-tra T1 Thermocycler as follows: an initial cycle of 2 min at95�C, followed by 33 cycles of 30 sec at 94�C, 30 sec at 60�Cand 1 min at 72�C and a final extended elongation step for10 min at 72�C. MLPA buffers and enzymes were obtainedfrom MRC-Holland (Amsterdam, the Netherlands).
Early
Detection
andDiagn
osis
Theelen et al. 961
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

Analysis of PCR products
Amplified FAM-labeled MLPA products were analyzed byelectrophoresis on an ABI3730 genetic analyzer (Applied Bio-systems, Foster City, CA). One microliter of 20� dilutedamplified MLPA products was mixed with 8.5 ll of deionized
formamide and 0.5 ll of GeneScan-600 LIZVR size standard(Applied Biosystems, Foster City, CA) and run in GeneScanmode. All analyses were done according to the manufac-turer’s instructions. Electropherograms were analyzed byGeneMarker software (Softgenetics, State College, PA), and
Figure 1. Overview of the HPV-MLPA methodology. A preamplification step is performed with a mixture of specific primers (fwd ¼ forward
primer and rev ¼ reverse primer) for each target (Step 1). Subsequently, the MLPA reaction is initiated by hybridizing a mixture of probe
sets specific for all targets to the preamplification products (Step 2). A probe set consists of 2 oligonucleotides: 1 synthetic oligonucleotide
(left) and 1 M13-derived oligonucleotide (right). The synthetic oligonucleotide contains a universal forward priming sequence (bold probe
line) next to the target specific sequence (thin probe line). The M13-derived oligonucleotide contains a universal reverse priming sequence
(bold probe line) and a unique stuffer sequence (dotted probe line) next to the target specific sequence (thin probe line). The length of the
stuffer sequence varies between the different probes. Subsequently, the 2 oligonucleotides are joined by ligation (Step 3) and the ligated
oligonucleotides are amplified by 1 universal primer set (fwd X and rev Y; Step 4). The length of the MLPA products is the combined length
of both oligonucleotides, which is unique for each probe set because of the specific length of the stuffer sequence. The amplification
products can be analyzed by capillary electrophoresis (Step 5), for example, for HPV16 (Step5A) and HPV18 (Step5B) of HPV/human DNA
mixtures containing 3000 episomal viral copies and 3000 human genome copies (HPV/human DNA copy number ratio of 1). Each MLPA
product can be discerned due to its specific length.
Early
Detection
andDiagn
osis
962 MLPA assay for HPV-related cancers
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

peak height data were exported to Excel files for calculationof ratios, means and standard deviations.
Plasmid model systems for viral integration and viral load
To determine what percentage of integrated HPV can bedetected reliably in a background of episomal HPV and viceversa, several mixtures of HPV plasmids, mimicking eitherintegrated or episomal HPV were used. These model systemswere also used to determine the lowest concentration of HPVthat can be detected in a background of human DNA andconversely at which concentration of HPV human targets canstill be detected.
HPV16 and 18 plasmids. For each of the 2 HPV types 2plasmids were used for the model systems. One plasmid con-taining all targets and thereby mimicking episomal HPV and1 plasmid with a deletion of at least 1 of the targets resem-bling the loss of sequences as a consequence of HPV inser-tion into the human genome. This latter plasmid will be usedto mimic integrated HPV.
Two synthetic plasmids were constructed for HPV16. Asynthetic construct was designed containing the targetsequences for HPV16 in the following order: 16E2.2, 16E2.1,the reverse complementary target sequence for 16E7 and thereverse complementary target sequence for 16E6. The HPV16plasmid containing E2 mimics episomal HPV and was madeby annealing 28 overlapping oligonucleotides (Biolegio, Mal-den, The Netherlands), designed with DNA works,39 coveringthe entire construct sequence, using the Phusion High-Fidel-ity PCR Master Mix (Finnzymes, Espoo, Finland). For theHPV16 plasmid lacking E2, which mimics integrated HPV,only the 16 oligonucleotides representing the 30-half wereused. Initial annealing was performed on a Biometra T1Thermocycler as follows: an initial cycle of 30 sec at 98�C,followed by 15 cycles of 10 sec at 98�C, 30 sec at 60�C and90 sec at 72�C and a final extended elongation step for 10min at 72�C. After initial annealing the constructs wereamplified, using only the first and last oligonucleotide, as fol-lows: an initial cycle of 30 sec at 98�C, followed by 30 cyclesof 10 sec at 98�C, 30 sec at 60�C and 90 sec at 72�C and afinal extended elongation step for 10 min at 72�C. The PCRreaction was applied according to the manufacturer’s instruc-tions (Finnzymes, Espoo, Finland) in a final volume of 50 ll.The oligonucleotides were used as both primer and templateDNA and a total of 5 pmol of each oligonucleotide was usedfor each reaction. The constructs were ligated into thepGem-T Easy vector (Promega, Madison, WI) according tomanufacturer’s instructions. DNA was extracted using TheWizardV
R
Plus SV Minipreps DNA Purification System(Promega, Madison, WI) according to protocol.
The 2 HPV18 plasmids used in this study were describedpreviously.43,44 The first plasmid was described to contain a7.8 kb HPV18 construct and was found to be positive for all4 HPV18 targets. This plasmid was used to mimic episomalHPV18. The second plasmid was described to contain a
6.9 kb construct. This plasmid was shown to lack 900 bpincluding E6 and was used to mimic integrated HPV18.
Reliability of quantifying integration and copy numbers. Todetermine what percentage of integrated HPV can reliably bedetected in a background of episomal HPV and vice versa,several combinations of the aforementioned HPV plasmids,representing either integrated or episomal HPV, were madevarying from 100% episomal HPV, i.e., an E2/E6 copy num-ber ratio (CNR) being 1, to 100% integrated HPV with anE2/E6 CNR being 0. This was achieved by mixing episomalHPV and integrated HPV to obtain samples with E2/E6CNRs of 1, 0.9, 0.8–0.1 and 0. Each sample contained �3,000copies of plasmid DNA (37 fg of the HPV18 plasmids or12 fg of the HPV16 plasmids, based on the length of eachplasmid) in a background of 10 ng (3,000 copies) of normalhuman DNA (Promega, Madison). The HPV/human DNACNR in these mixtures is 1.
To determine the lowest concentration of HPV that canbe detected in a background of human DNA and converselyto determine at which concentration of HPV the human tar-gets can still be detected, variants of the above describedHPV16 integration series were designed, in that the HPV/human DNA CNR varied from 0.1 to 100. Furthermore,these series were used to assess the effect of copy numbervariations on the quantification of integration. All series weredesigned to have the same distribution in physical status withE2/E6 CNRs varying from 1.0 to 0. Three series weredesigned to contain more HPV copies than human copieswith 300,000, 150,000 or 30,000 copies of HPV, in a back-ground of 3,000 copies of human DNA (HPV/human DNACNR being 100, 50 and 10, respectively). A fourth series wasdesigned to contain less HPV copies than human DNA cop-ies, with 300 copies of HPV DNA in a background of 3,000human copies (HPV/human DNA CNR being 0.1). ForHPV18, only the series with a HPV/human DNA CNR of 0.1and 100 were prepared.
Also, the influence of the total amount of input DNA onthe capillary electrophoresis peak profile was tested. For thispurpose, the initial integration series was tested with 100 nginstead of 10 ng of total DNA.
Calculation of viral load (defined here as HPV copies perhuman gene copy) and determination of the physical status(i.e., integrated HPV, episomal HPV or mixed) are based onthe signal intensity (peak height) ratio of the probe signals inthe capillary electrophoresis profile. The viral load is esti-mated based on the average ratio of the signal intensities ofE6 and E7 to the total signal intensity of all 7 human targetsusing the following equations: L ¼ 55.9r1:11 , L ¼ 12.5r1:31 , L ¼16.8r1:31 , and L ¼ 43.2r1:41 for 16E6, 16E7, 18E6 and 18E7,respectively. With L being the viral load estimated as HPVcopies per human gene copy and r1 being the ratio betweentarget signal intensity and the total signal intensity of allhuman targets. The physical status is estimated based on the
Early
Detection
andDiagn
osis
Theelen et al. 963
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

E2/E6 signal intensity ratio for HPV16 and the E6/E2 signalintensity ratio for HPV18.
For the plasmid model system with a HPV/human DNACNR of 1, a standard integration curve was made, in whichthe known percentage of integration in the prepared mixturewas plotted against the HPV16 E2.1/E6, HPV16 E2.2/E6,HPV18 E6/E2.1 or HPV18 E6/E2.2 signal intensity ratiomeasured with the MLPA assay. For the series with aHPV16/human DNA CNR of 10, 50 and 100, these curveswere also determined.
To determine gain for TERT, the ratio between the com-bined signal intensity of TERTa and TERTc on the one handand the combined signal intensity of b-globin-a, b-globin-band MSH2 on the other hand was calculated. This signal in-tensity ratio was compared with the ratio as measured forthese targets in the normal human DNA sample. When thesignal intensity ratio was 1.5� higher than that of the refer-ence sample, TERT was classified as gained. To determinegain for TERC, the ratio between the combined signal inten-sity of TERCa and TERCb on the one hand and the com-bined signal intensity of b-globin-a, b-globin-b and MSH2on the other hand was calculated. This signal intensity ratiowas also compared with the ratio as measured for these tar-gets in the normal human DNA sample and classified as gainwhen it was 1.5� higher than that of the reference sample.
Quantitative PCR
Viral load for HPV18 in the HeLa cell culture was estimatedby qPCR using the preamplification primers for 18E6 incombination with LC-green (Idaho Technology Inc., SaltLake City, UT). A 20 ll reaction mixture contained dNTPs(2 mM each, Fermentas, St. Leon-Rot, Germany), 1 U Taq-polymerase (MRC-Holland, Amsterdam, the Netherlands),1� PCR buffer (50 mM KCl, 10 mM Tris-HCl pH 8.5,1.6 mM MgCl2), 400 fmol of each primer, 0.5 ll LC-green(Idaho Technology Inc.) and 10 ng of sample DNA. TheqPCR reactions were performed in a Rotor-Gene 6000 real-time system (Corbett Life Science, Sydney, Australia) as fol-lows: 2 min at 95�C, followed by 45 cycles of 30 sec at 94�C,90 sec at 55�C and 90 sec at 72�C and a final extended elon-gation step for 10 min at 72�C. The standard curve wasobtained by amplification of a 5-fold dilution series of 6 �106 to 1,920 copies of the HPV18 plasmid in a backgroundof 10 ng normal human DNA (Promega, Madison, USA) perreaction mixture.
Viral load and physical status for HPV16 in the SiHa andCaSki cell cultures and the fresh-frozen and cytological clini-cal samples were estimated by qPCR using the previouslydescribed primers and probes33 for 16E6 and 16E2 (Biolegio,Malden, The Netherlands) in a 20-ll PCR mixture containingdNTPs (2 mM each, Fermentas, St. Leon-Rot, Germany), 1 UTaq-polymerase (MRC-Holland, Amsterdam, the Nether-lands), 1� PCR buffer (50 mM KCl, 10 mM Tris-HCl pH8.5, 1.6 mM MgCl2), 3 pmol of each primer, 1 pmol of eachprobe and 25 ng of sample DNA. The qPCR reactions were
performed in a Rotor-Gene 6000 real-time system (CorbettLife Science, Sydney, Australia) as follows: 2 min at 95�C,followed by 50 cycles of 30 sec at 94�C, 90 sec at 55�C and90 sec at 72�C and a final extended elongation step for10 min at 72�C. The standard curves were obtained byamplification of a 10-fold dilution series of 5 � 106 to 500copies of the HPV16 plasmid.
The threshold cycle values for all standard curves wereplotted against the log of the copy number over the entirerange of dilutions and revealed a linear relationship. Thesestandard curves were used to estimate the average HPV copynumber in the individual cell lines based on duplicate meas-urements. Viral loads for the cell lines were calculated basedon the ploidy of the cell lines as estimated by FISH using ahuman chromosome 3 centromere-specific probe (see later)and a DNA mass of 7.8 pg per diploid cell.45 Viral loads forthe clinical samples were calculated based on the assumptionthat they have a diploid DNA content and a DNA mass of7.8 pg per diploid cell.45 The physical status of the virus wasdetermined on the basis of the CNR between E2 and E6.Samples were classified as integrated when the ratio was 0,mixed for ratios between 0 and 0.5 and episomal for ratiosabove 0.5 as previously described.46
Fluorescence in situ hybridization
FISH analysis on the cell lines and fresh-frozen clinical sam-ples was performed as previously described12,47 using digoxy-genin-labeled probes for HPV16 or 18 or Vysis probe sets(Vysis, Abbott Molecular, Des Plaines, IL) consisting of: (i) aDNA probe for chromosome 3 centromere labeled with thefluorescent dye Spectrum Aqua (SA), a 3q26-specific BACclone containing the TERC gene labeled with Spectrum Or-ange (SO) and a 5p15-specific BAC clone containing theTERT gene labeled with Spectrum Green (SG) or (ii) a DNAprobe for chromosome 3 centromere labeled with the fluores-cent dye Spectrum Green (SG), a 3q26-specific BAC clonecontaining the TERC gene labeled with Spectrum Orange(SO) and a probe for chromosome 7 centromere labeled withthe fluorescent dye Spectrum Aqua (SA). For the paraffin-embedded tissue sections, HPV FISH analysis was performedas described previously using the mild pretreatmentprotocol.48
Images were acquired using a Leica DMRXA microscope(Leica, Wetzlar, Germany) equipped with custom optical fil-ters for DAPI, SA, SO and SG (Chroma Technologies, Brat-tleboro, VT) with a �40 Plan Apo (NA 1.20) objective. Themicroscope was connected to a digital black and white CCDcamera (Metasystems Image Pro System, Sandhausen, Ger-many). To estimate an average copy number for the TERCand TERT targets, the FISH signals (spots) were counted in atotal of 50 interphase cells per cell line. For the fresh-frozenclinical samples, the copy number for chromosome 7 wasused as a control for the ploidy of cells. For chromosome 3and target 3q26 (TERC), the maximum number of signals
Early
Detection
andDiagn
osis
964 MLPA assay for HPV-related cancers
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC
per nucleus was determined and used as an indicator of copynumber, as described previously.12
ResultsEstimation of viral load
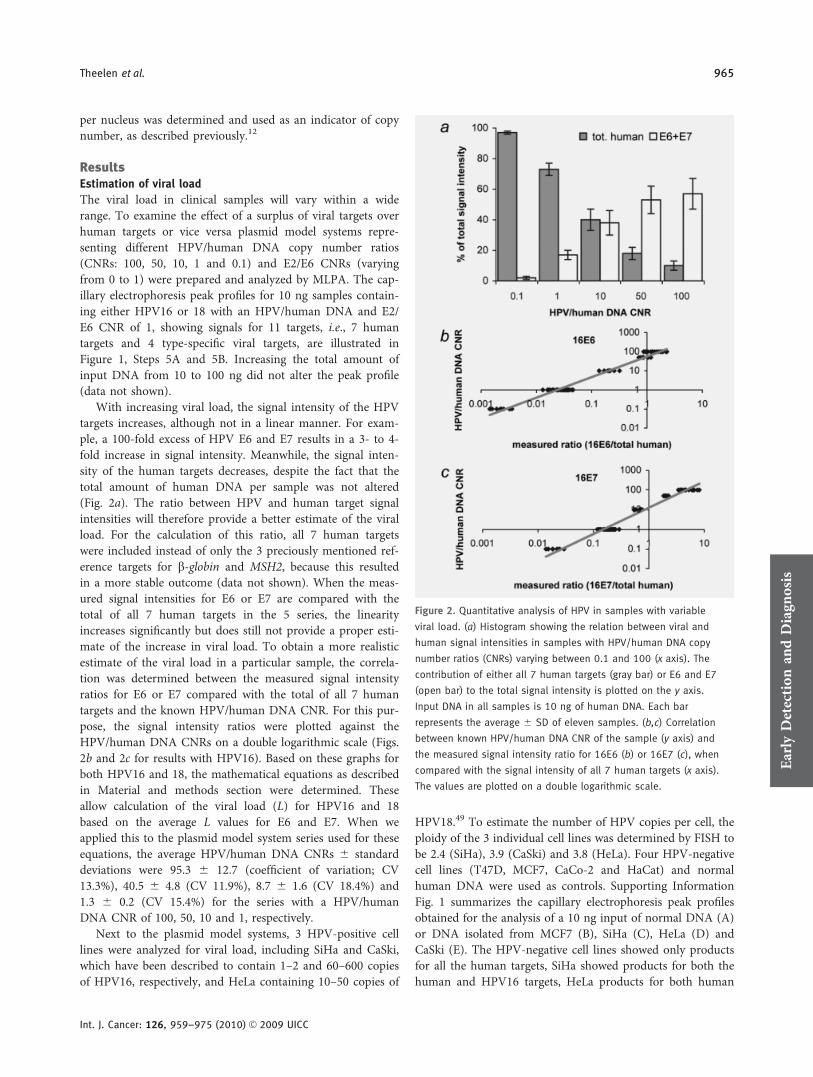
The viral load in clinical samples will vary within a widerange. To examine the effect of a surplus of viral targets overhuman targets or vice versa plasmid model systems repre-senting different HPV/human DNA copy number ratios(CNRs: 100, 50, 10, 1 and 0.1) and E2/E6 CNRs (varyingfrom 0 to 1) were prepared and analyzed by MLPA. The cap-illary electrophoresis peak profiles for 10 ng samples contain-ing either HPV16 or 18 with an HPV/human DNA and E2/E6 CNR of 1, showing signals for 11 targets, i.e., 7 humantargets and 4 type-specific viral targets, are illustrated inFigure 1, Steps 5A and 5B. Increasing the total amount ofinput DNA from 10 to 100 ng did not alter the peak profile(data not shown).
With increasing viral load, the signal intensity of the HPVtargets increases, although not in a linear manner. For exam-ple, a 100-fold excess of HPV E6 and E7 results in a 3- to 4-fold increase in signal intensity. Meanwhile, the signal inten-sity of the human targets decreases, despite the fact that thetotal amount of human DNA per sample was not altered(Fig. 2a). The ratio between HPV and human target signalintensities will therefore provide a better estimate of the viralload. For the calculation of this ratio, all 7 human targetswere included instead of only the 3 preciously mentioned ref-erence targets for b-globin and MSH2, because this resultedin a more stable outcome (data not shown). When the meas-ured signal intensities for E6 or E7 are compared with thetotal of all 7 human targets in the 5 series, the linearityincreases significantly but does still not provide a proper esti-mate of the increase in viral load. To obtain a more realisticestimate of the viral load in a particular sample, the correla-tion was determined between the measured signal intensityratios for E6 or E7 compared with the total of all 7 humantargets and the known HPV/human DNA CNR. For this pur-pose, the signal intensity ratios were plotted against theHPV/human DNA CNRs on a double logarithmic scale (Figs.2b and 2c for results with HPV16). Based on these graphs forboth HPV16 and 18, the mathematical equations as describedin Material and methods section were determined. Theseallow calculation of the viral load (L) for HPV16 and 18based on the average L values for E6 and E7. When weapplied this to the plasmid model system series used for theseequations, the average HPV/human DNA CNRs 6 standarddeviations were 95.3 6 12.7 (coefficient of variation; CV13.3%), 40.5 6 4.8 (CV 11.9%), 8.7 6 1.6 (CV 18.4%) and1.3 6 0.2 (CV 15.4%) for the series with a HPV/humanDNA CNR of 100, 50, 10 and 1, respectively.
Next to the plasmid model systems, 3 HPV-positive celllines were analyzed for viral load, including SiHa and CaSki,which have been described to contain 1–2 and 60–600 copiesof HPV16, respectively, and HeLa containing 10–50 copies of
HPV18.49 To estimate the number of HPV copies per cell, theploidy of the 3 individual cell lines was determined by FISH tobe 2.4 (SiHa), 3.9 (CaSki) and 3.8 (HeLa). Four HPV-negativecell lines (T47D, MCF7, CaCo-2 and HaCat) and normalhuman DNA were used as controls. Supporting InformationFig. 1 summarizes the capillary electrophoresis peak profilesobtained for the analysis of a 10 ng input of normal DNA (A)or DNA isolated from MCF7 (B), SiHa (C), HeLa (D) andCaSki (E). The HPV-negative cell lines showed only productsfor all the human targets, SiHa showed products for both thehuman and HPV16 targets, HeLa products for both human
Figure 2. Quantitative analysis of HPV in samples with variable
viral load. (a) Histogram showing the relation between viral and
human signal intensities in samples with HPV/human DNA copy
number ratios (CNRs) varying between 0.1 and 100 (x axis). The
contribution of either all 7 human targets (gray bar) or E6 and E7
(open bar) to the total signal intensity is plotted on the y axis.
Input DNA in all samples is 10 ng of human DNA. Each bar
represents the average 6 SD of eleven samples. (b,c) Correlation
between known HPV/human DNA CNR of the sample (y axis) and
the measured signal intensity ratio for 16E6 (b) or 16E7 (c), when
compared with the signal intensity of all 7 human targets (x axis).
The values are plotted on a double logarithmic scale.Early
Detection
andDiagn
osis
Theelen et al. 965
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

and HPV18 targets, whereas for CaSki only the dominantpeaks for the HPV16 targets are recognized.
For the estimation of viral load, the mathematical equa-tions, as described in Material and methods section wereused. By averaging the L values for E6 and E7 and takinginto account the ploidy of the cell, the number of HPV cop-ies per cell for SiHa was calculated to be 14, for HeLa to be22 and for CaSki to be 579. With qPCR, the viral loads weredetermined to be 29 copies per cell for SiHa, 34 copies percell for HeLa and 1,946 copies per cell for CaSki.
Estimation of viral integration
It has been described that viral integration correlates stronglywith disease progression. As a consequence of this integrationprocess, the E2 gene is (partially) deleted. Quantification ofthe E2/E6 ratio will therefore allow estimation of viral inte-gration. To test whether our MLPA assay allows an accuratedistinction of different percentages of viral integration, wehave used the plasmid samples with varying percentages ofintegrated viral copies (E2/E6 CNR varying from 0 to 1) anda constant HPV/human DNA CNR of 1. The known percent-age of integration was plotted against the measured E2.1/E6or E2.2/E6 ratio for HPV16 or the measured E6/E2.1 or E6/E2.2 ratio for HPV18 (see Material and methods section).The graphs in Figures 3a–3d clearly illustrate the linear cor-relation between these 2 parameters for HPV16 and 18.Based on the standard deviation of triplicate measurements,these curves show that integrated HPV is reliably recognizedwhen its frequency exceeds that of 30% of the total numberof viral copies. This results in a classification of episomalHPV when less than 30% of the virus in a sample showsintegration, mixed (i.e., episomal and integrated) HPV whenbetween 30% and 95% of the virus shows integration andintegrated HPV when more than 95% of the virus showsintegration. In the HPV18 plasmid model systems E6 insteadof E2 was deleted, showing that reliable quantification is in-dependent of the deleted sequence. For this purpose, the E6/E2 ratio is calculated for these samples instead of the E2/E6ratio.
Because clinical samples show large variations in viralload, the question becomes relevant to what extent this caninfluence the reliable detection of viral integration. For theseries with increasing amounts of HPV16 DNA, i.e., HPV/human DNA CNRs varying from 1 to 100, the measured sig-nal intensity ratios for E2.1/E6 and E2.2/E6 again showed alinear correlation with the known percentage of integration(Figs. 3e and 3f). However, the difference in the E2.1/E6 sig-nal intensity ratio between 90% and 10% integration sampleswas influenced by the viral load. In the series with a HPV/human DNA CNR of 1, the E2.1/E6 signal intensity ratio ofthe 10% integration sample was 12.3� larger than the E2.1/E6 signal intensity ratio of the 90% integration sample. Inthe series with a HPV/human DNA CNR of 10, 50 and 100,the difference was 6.3-, 3.7- and 3.4-fold, respectively. Similarresults were obtained for the HPV16 E2.2/E6 signal intensity
ratio, which exhibited 11.4-, 6.0-, 3.9- and 3.1-fold increasein the respective HPV/human DNA CNR series.
When the HPV/human DNA CNR was 0.1, the signalintensities for the HPV targets was too low to reliably analyzethe physical status of the virus (data not shown).
For the analysis of the physical status of HPV in the cer-vical cell lines, the E2.1/E6 and E2.2/E6 signal intensity ratioswere calculated. In HeLa, no signals were measured for the18E2.1 and 18E2.2 targets, which is interpreted as a completedeletion of these targets and thus as a 100% integration ofHPV. For SiHa and CaSki, the signal intensity ratios for16E2.1/E6 and 16E2.2/E6 were measured and compared withthe graphs in Figures 3e and 3f, based on the curve closest totheir estimated viral load (HPV/human DNA CNR 10 andHPC/human DNA CNR 100, respectively). For SiHa, it wasdetermined that �90% of the 16E2.1 target was deleted,based on a signal intensity ratio of 0.39, whereas 16E2.2 wasretained (signal intensity ratio 1.97). In CaSki, both 16E2.1(signal intensity ratio 2.52) and 16E2.2 (signal intensity ratio2.66) were retained, which could be interpreted as exclusivelyepisomal HPV but is a result of multicopy tandem repeats ofHPV in this cell line.50 Using qPCR E2/E6 CNRs of 0.04 and2.29 were found for SiHa and CaSki, respectively.
Telomerase-related genes
Gain of the telomerase-related genes is associated with pro-gression of (HPV related) cancers and can therefore be usedas a prognosticator. For this reason simultaneous detection ofTERT and TERC next to HPV increases the ability of thisMLPA assay to distinguish the more aggressive lesions. Todetermine the reliability of the MLPA assay in this respectthe copy number gains of the telomerase genes TERT andTERC as determined by MLPA were compared with theresults obtained by FISH analysis (Table 1). For the MLPAanalysis of the copy number gain for TERT and TERC, thecombined signal intensities of the 2 probes for either TERTor TERC were divided by the combined signal intensities ofthe MSH2 and both b-globin probes. These signal intensityratios were compared with the ratios as measured for thesetargets in the normal human DNA sample. When the signalintensity ratio for the cell line was 1.5� higher than that ofthe reference sample, the cell line was classified as having acopy number gain. All 3 cell lines showed a gain for TERT(6.0, 2.3 and 1.9� higher signal intensity ratio, when com-pared with normal human DNA for CaSki, SiHa and HeLa,respectively), whereas gain for TERC was only found inCaSki and SiHa (3.1� and 1.6� higher, when compared withnormal human DNA). These data were comparable with theresults of the FISH analysis for the TERC and TERT gene(Table 1), with the centromere 3 probe as a reference probefor chromosome ploidy (Fig. 4a–4c).
Application of the MLPA assay in clinical samples
To test the performance of the HPV MLPA assay in differenttypes of clinical samples, we selected a limited series of fresh-
Early
Detection
andDiagn
osis
966 MLPA assay for HPV-related cancers
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC
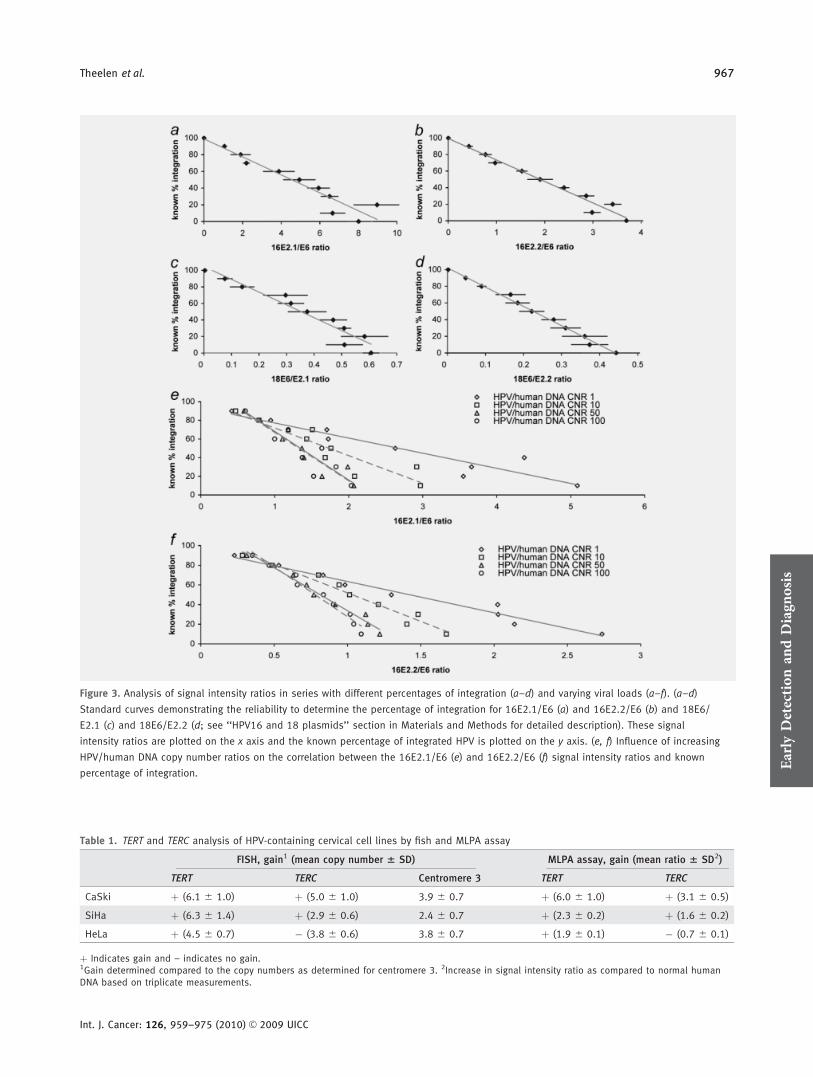
Figure 3. Analysis of signal intensity ratios in series with different percentages of integration (a–d) and varying viral loads (a–f). (a–d)
Standard curves demonstrating the reliability to determine the percentage of integration for 16E2.1/E6 (a) and 16E2.2/E6 (b) and 18E6/
E2.1 (c) and 18E6/E2.2 (d; see ‘‘HPV16 and 18 plasmids’’ section in Materials and Methods for detailed description). These signal
intensity ratios are plotted on the x axis and the known percentage of integrated HPV is plotted on the y axis. (e, f) Influence of increasing
HPV/human DNA copy number ratios on the correlation between the 16E2.1/E6 (e) and 16E2.2/E6 (f) signal intensity ratios and known
percentage of integration.
Table 1. TERT and TERC analysis of HPV-containing cervical cell lines by fish and MLPA assay
FISH, gain1 (mean copy number 6 SD) MLPA assay, gain (mean ratio 6 SD2)
TERT TERC Centromere 3 TERT TERC
CaSki þ (6.1 6 1.0) þ (5.0 6 1.0) 3.9 6 0.7 þ (6.0 6 1.0) þ (3.1 6 0.5)
SiHa þ (6.3 6 1.4) þ (2.9 6 0.6) 2.4 6 0.7 þ (2.3 6 0.2) þ (1.6 6 0.2)
HeLa þ (4.5 6 0.7) � (3.8 6 0.6) 3.8 6 0.7 þ (1.9 6 0.1) � (0.7 6 0.1)
þ Indicates gain and – indicates no gain.1Gain determined compared to the copy numbers as determined for centromere 3. 2Increase in signal intensity ratio as compared to normal humanDNA based on triplicate measurements.
Early
Detection
andDiagn
osis
Theelen et al. 967
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC
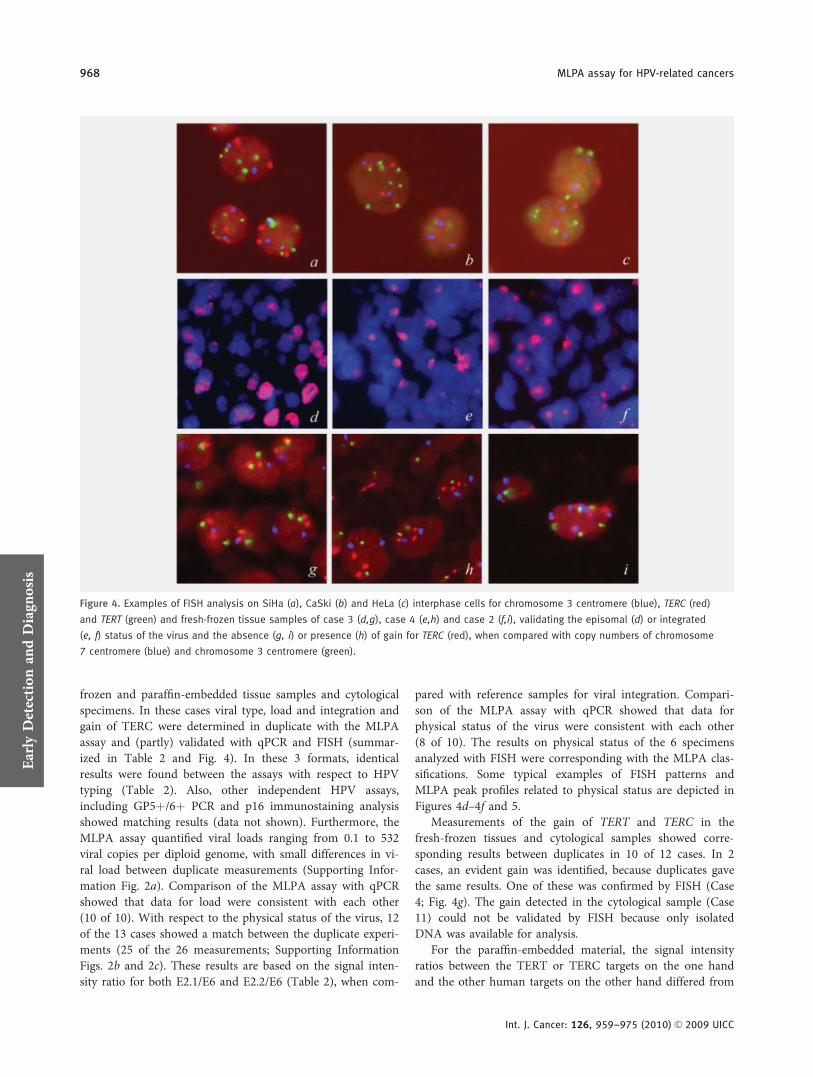
frozen and paraffin-embedded tissue samples and cytologicalspecimens. In these cases viral type, load and integration andgain of TERC were determined in duplicate with the MLPAassay and (partly) validated with qPCR and FISH (summar-ized in Table 2 and Fig. 4). In these 3 formats, identicalresults were found between the assays with respect to HPVtyping (Table 2). Also, other independent HPV assays,including GP5þ/6þ PCR and p16 immunostaining analysisshowed matching results (data not shown). Furthermore, theMLPA assay quantified viral loads ranging from 0.1 to 532viral copies per diploid genome, with small differences in vi-ral load between duplicate measurements (Supporting Infor-mation Fig. 2a). Comparison of the MLPA assay with qPCRshowed that data for load were consistent with each other(10 of 10). With respect to the physical status of the virus, 12of the 13 cases showed a match between the duplicate experi-ments (25 of the 26 measurements; Supporting InformationFigs. 2b and 2c). These results are based on the signal inten-sity ratio for both E2.1/E6 and E2.2/E6 (Table 2), when com-
pared with reference samples for viral integration. Compari-son of the MLPA assay with qPCR showed that data forphysical status of the virus were consistent with each other(8 of 10). The results on physical status of the 6 specimensanalyzed with FISH were corresponding with the MLPA clas-sifications. Some typical examples of FISH patterns andMLPA peak profiles related to physical status are depicted inFigures 4d–4f and 5.
Measurements of the gain of TERT and TERC in thefresh-frozen tissues and cytological samples showed corre-sponding results between duplicates in 10 of 12 cases. In 2cases, an evident gain was identified, because duplicates gavethe same results. One of these was confirmed by FISH (Case4; Fig. 4g). The gain detected in the cytological sample (Case11) could not be validated by FISH because only isolatedDNA was available for analysis.
For the paraffin-embedded material, the signal intensityratios between the TERT or TERC targets on the one handand the other human targets on the other hand differed from
Figure 4. Examples of FISH analysis on SiHa (a), CaSki (b) and HeLa (c) interphase cells for chromosome 3 centromere (blue), TERC (red)
and TERT (green) and fresh-frozen tissue samples of case 3 (d,g), case 4 (e,h) and case 2 (f,i), validating the episomal (d) or integrated
(e, f) status of the virus and the absence (g, i) or presence (h) of gain for TERC (red), when compared with copy numbers of chromosome
7 centromere (blue) and chromosome 3 centromere (green).
Early
Detection
andDiagn
osis
968 MLPA assay for HPV-related cancers
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

Table
2.Resu
ltsoftheHPVMLPAassayin
routinely
obtainedclinicalsamplesofpatients
withlesionsoftheuterinecervix,whichwere
alsovalidatedbyqPCRand/orfish
Material
Case
Clinicalinform
ation
MLP
AHPV16/18
qPCRHPV16
FISH
MLP
AFISH
Diagnosis
AgeFollow-
up
Exp
TypeLoad
E2.1/E6
Sign
ratio
E2.2/E6
Sign
ratio
E2.1
1
TargetE2.2
1
TargetPhysical
Status
Load
E2/E6
Ratio
Physical
Status2
TypePhysical
Status
TERT
ratio3
TERC
ratio3
TERT
gain
TERC
gain
Telo.
gain
TERC
gain
Fresh
-frozen
1Norm
al
65
n.a.
1�
��
��
��
��
�1.04
0.78
��
�tissue
2�
��
��
��
��
�0.60
0.63
��
2CIN2/3
43
n.a.
116
1.2
2.9
1.4
RPD
Mix
0.7
0Int
16
Int
0.85
1.17
��
��
216
1.5
3.6
1.6
RPD
Mix
0.7
0.11
Mix
0.72
0.78
��
3CIN2/3
27
n.a.
116
241
1.6
1.6
RR
Epi
129
0.73
Epi
16
Epi
1.07
1.44
��
��
216
256
1.3
1.5
RR
Epi
95
0.84
Epi
0.72
0.93
��
4SCC
n.a
n.a.
116
157
1.1
1.3
PD
PD
Mix
245
0.46
Mix
16
Int
0.93
1.79
�þ
þþ
216
140
1.2
1.5
PD
PD
Mix
153
0.36
Mix
0.78
1.64
�þ
5SCC
52
n.a.
116
8.1
0.03
2.3
DR
Int
3.5
0Int
1.03
0.83
��
�2
16
10.8
0.03
1.8
DR
Int
3.8
0Int
0.70
0.48
��
Cytological
6Norm
al
60
n.a.
1�
��
��
��
��
�0.85
0.93
��
�samples
2�
��
��
��
��
�2.58
2.07
þþ
7Norm
al
29
CIN24
116
3.3
5.2
3.0
RR
Epi
3.6
0.45
Mix
1.06
0.91
��
�2
16
3.8
12.0
4.0
RR
Epi
2.6
0.50
Mix
1.24
1.09
��
8Atypical
30
n.a.
116
9.0
1.3
2.0
PD
REpi
15.2
0.69
Epi
0.52
0.61
��
�2
16
9.4
7.9
2.4
RR
Epi
7.5
0.51
Epi
0.84
1.16
��
9Dyskaryotic56
Norm
al5
116
45
1.6
1.2
PD
PD
Mix
40
0.44
Mix
1.38
1.43
��
�2
16
31
2.4
1.4
PD
PD
Mix
42
0.26
Mix
1.56
2.39
þþ
10
Dyskaryotic31
CIN25
116
93
1.2
0.9
PD
PD
Mix
86
0.50
Mix
0.91
0.81
��
�2
16
99
2.8
1.2
PD
PD
Mix
54
0.001
Mix
1.10
1.45
��
11
Dyskaryotic30
CIN15
116
283
1.9
1.1
RR
Epi
185
0.61
Epi
2.13
2.08
þþ
þ2
16
320
2.8
1.1
RR
Epi
328
0.55
Epi
1.81
1.63
þþ
12
Dyskaryotic30
CIN15
116
532
1.2
1.3
PD
RMix
724
0.67
Epi
0.47
0.53
��
�2
16
460
1.8
1.3
PD
RMix
710
0.55
Epi
0.36
0.62
��
Early
Detection
andDiagn
osis
Theelen et al. 969
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

that of the plasmid model systems, which implies that forthis type of material optimization is needed to determinegain of TERT and TERC (Table 2).
DiscussionSeveral factors have been reported to play a role in the clinicalbehavior of HPV-related (pre)malignant lesions. Simultaneousassessment of the most important parameters, i.e., viral type,load and physical status and aberrations in the host genome,is of great diagnostic and prognostic importance. In theunderlying study, we show that a multiparameter MLPA assay,based on 15 selected targets for which new probes were devel-oped, can reliably assess HPV 16/18, viral load and integrationof the virus, together with copy number gain of the telomer-ase-related genes TERT and TERC. Our MLPA HPV E2, E6and E7 probes demonstrated to be type specific and alloweddetection of a single copy of HPV per cell. Our study alsodemonstrates the feasibility to detect even lower viral loads, asoften encountered in clinical samples with admixtures of vari-able amounts of noninfected cells. Furthermore, viral integra-tion can reliably be detected when at least 30% of the HPV isincorporated into the human genome. Combined with theseparameters for HPV, gain for TERT and TERC could be deter-mined in cell samples with varying viral loads.
The HPV MLPA assay
The quantitative MLPA assays described previously are onlybeing used for the analysis of small range gene copy numbervariations.38 For the determination of viral load and viralTa
ble
2.Resu
ltsoftheHPVMLPAassayin
routinely
obtainedclinicalsamplesofpatients
withlesionsoftheuterinecervix,whichwere
alsovalidatedbyqPCRand/orfish
(Continued)
Material
Case
Clinicalinform
ation
MLP
AHPV16/18
qPCRHPV16
FISH
MLP
AFISH
Diagnosis
AgeFollow-
up
Exp
TypeLoad
E2.1/E6
Sign
ratio
E2.2/E6
Sign
ratio
E2.1
1
TargetE2.2
1
TargetPhysical
Status
Load
E2/E6
Ratio
Physical
Status2
TypePhysical
Status
TERT
ratio3
TERC
ratio3
TERT
gain
TERC
gain
Telo.
gain
TERC
gain
Paraffin�
13
CIN2/3
35
SCC
118
0.3
01.0
DR
Int
18
Int
Inconcl.Inconcl.Inconcl.Inconcl.
�embedded
218
0.3
00.9
DR
Int
tissue
14
CIN2/3
38
SCC
116
82.2
1.2
0.7
PD
PD
Mix
16
Int
Inconcl.Inconcl.Inconcl.Inconcl.
þ2
16
82.6
0.9
0.6
PD
PD
Mix
15
AdC
44
n.a.
118
1.5
0.2
0.4
PD
PD
Mix
18
Int
Inconcl.Inconcl.Inconcl.Inconcl.
�2
18
1.2
0.1
0.2
PD
PD
Mix
þindicatesdetected;–indicatesnotdetectedandopencellsindicate
thattheexp
erimentwasnotperform
edforthatsample.
MLPA¼
Multiplexligation-dependentprobeamplification;qPCR¼
quantitative
PCR;FISH¼
fluorescence
insitu
hyb
ridization;CIN
¼cervicalintraepithelialneoplasia;SCC¼
squamouscell
carcinoma;AdC¼
adenocarcinoma;n.a.¼
notavailable;exp
¼exp
erimentalnumber;signratio¼
signalintensity
ratio;R¼
retained;PD¼
partly
deleted;D¼
deleted;epi¼
episomal;
mix
¼mixed;int¼
integrated;inconcl.¼
inconclusive;telo.¼
telomerase
gene.
1Targetstatusis
determ
inedbasedoncompariso
nofthesignalintensity
ratiooftheclinicalsample
tothesignalintensity
ratioofthematchingexp
erimentalreference
samples.
2Statusis
determ
inedbasedonthecriteriareportedbyRuutu
etal51;E2/E6ratio>0.5
¼episomal;E2/E6ratio>0upto
0.5
¼mixed;E2/E6ratio0¼
integrated.3Increase
insignalintensity
ratioas
comparedto
norm
alhumanDNA.4Histologicalfollow-upafter14months.
5Histologicalfollow-upafter2–3months.
Figure 5. Examples of MLPA capillary electrophoresis peak profiles
for DNA isolated from fresh-frozen tissue sample Case 4 with
mixed physical status for HPV16 (a), cytological sample Case 8
with episomal HPV16 (b) and paraffin-embedded tissue sample
case 13 with fully integrated HPV18 as concluded from the
complete deletion of 18E2.1 (c).
Early
Detection
andDiagn
osis
970 MLPA assay for HPV-related cancers
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

integration, often a qPCR assay is used.51 In this study, wewanted to investigate whether it is possible to use the MLPAassay to simultaneously quantify all 3 aspects. Similar toqPCR this MLPA assay also uses the E2/E6 ratio for thedetermination of viral load.33,51,52 However, our MLPA assayshowed that it is possible to distinguish a lower percentage ofintegration (30% of integration with the MLPA assay vs. 50%of integration with the qPCR) and thus identify a largergroup of mixed cases where the qPCR would label part ofthese as episomal.51
In this MLPA assay also, a preamplification step is incor-porated to increase sensitivity and to facilitate detection ofvery small amounts of HPV. Based on previous reports, thelower threshold value for viral load detection is 0.2 copiesper cell, with the majority of clinical cases, however, harbor-ing at least 1 copy per cell.33,52 The incorporation of the pre-amplification step is needed in clinical samples, such as biop-sies or cytological specimens, from which only small amountsof DNA can be obtained. Because of the competition effectbetween the multiple primers in the preamplification step,the accuracy of the assay for detecting all 4 parameters islimited for the more extreme situations, e.g., samples with avery high-viral load or with a very low-viral load. Althoughviral load measurements proved to be feasible for 0.1 copiesper cell, the assay is developed to optimally analyze the 4 pa-rameters in samples with a viral load range between 1 and500 copies per diploid genome. We anticipate to improve theaccuracy of the assay for samples with higher and lower viralloads by varying the experimental conditions.
HPV16 and 18 typing
The identification of HPV16 and 18 infections has an addedvalue in the screening for cervical (pre)neoplasia in that thesensitivity of detection is increased, when compared withcytological analysis only.53 In this screening approach false-positive results are highly unwanted. The specificity of theprobes for HPV16 or 18 is warranted in that their selectionis based on the alignment of the sequences of the 30 mostprevalent genital HPV types. Furthermore, our studies showthat there is no crosshybridization of these HPV16 or 18probes with each others genome or with the closely relatedHPV types 31, 33 and 45 or with human sequences. Withour MLPA assay a false-positive outcome is furthermore pre-vented through the use of 4 probes per HPV type. An HPVinfection is only scored as being positive when a product isdetected for both the E6 and E7 probe, whereas in a largegroup of cases also products will be detected for 1 or both ofthe E2 probes, providing additional proof for the reliability ofthe outcome. When dealing with samples with extreme low-viral loads (<1 copy per 10 cells), the signals for the HPVtargets will be too low for reliable analysis. For these samples,the reaction can be repeated with HPV probes alone.Although this approach does not provide exact informationon viral load, the sensitivity of typing will be improved.
Viral load
The presence of a high-viral load in premalignant lesions ofthe uterine cervix is described to be an indicator for progres-sion to malignancy,54 whereas increased viral load correlateswith improved survival in patients with tonsil carcinomas.8
The underlying study shows that viral load can reliably bedetermined based on the signal intensity of E6 and E7, whencompared with the human targets. A detection range of 1copy of HPV per 10 cells in a sample up to several hundredsof copies per cell is feasible. However, in case of extremelyhigh-viral loads (>500 copies per cell), it can no longer beaccurately quantified because of the low or absent signals forthe human targets, which is illustrated, for example, by thepeak profile for CaSki. An important conclusion from ourstudy is, however, that this HPV MLPA assay can reliablydistinguish samples with high-viral load. A comparison withqPCR not only confirmed the high-viral load for CaSki butalso showed that qPCR analysis results in a 4-fold highercopy number estimation. When comparing our results toreports in the literature49,50,55 using, for example, qPCR andSouthern blotting to quantify viral load in CaSki, it becomesobvious that also these approaches result in a highly variableHPV copy number estimation, ranging from �60 to �1,700copies per cell. Furthermore, comparison between MLPA andqPCR in the fresh-frozen and cytological samples demon-strated a good correlation for the estimation of viral load inclinical settings.
For calculating viral load, the signal intensities for all 7human targets are used as a reference. Among these are alsothe 4 targets for TERT and TERC, which are expected to begained in many cases. Because we could, however, showthat the use of 7 reference targets results in more stable esti-mations than 3 reference targets and moreover that copynumber changes for these genes are relatively minor com-pared with the changes in viral load, it is both save and nec-essary to include these targets for the estimation of viralload.
HPV integration
As can be deduced from the literature 70–80% of cervicalhigh-grade preneoplasia, carcinoma in situ and carcinomacases show disruption of E2 expression on integration of viralDNA into the human genome.20,56 In majority of cases, thisis a result of deletion of E2, but deletions of the E1 gene arealso described.20–22 For HPV16, E2 deletions are found in60–80% of the cases,21,52,57 whereas for HPV18 E2 this isfound in 90–100% of the cases.58,59 In head and neck lesions,the findings concerning these E2 deletions are similar to cer-vical lesions. Nearly all HPV16-infected head and necklesions show a disruption of this gene.60,61 Deletions in thesegenes are not limited to special regions.20,21 In our study, 2probes directed against different positions in the E2 genewere selected to determine viral integration. For HPV16,these probes were located within the 2 regions that were
Early
Detection
andDiagn
osis
Theelen et al. 971
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

described to be deleted in �90% of high-grade cervicallesions, when taken together.20,21 For HPV18, this detailedinformation is not available and the 2 probes used in theunderlying study were therefore randomly selected. By using2 probe sets for the detection of integration instead of 1, thenumber of cases with detected integration will increase, but itis still possible that some cases with integration will bemissed. The probability to detect viral integration with ourMLPA assay can be increased by adding additional probesdispersed over the E1 and E2 genes. In cervical invasive can-cers, it has been described that multiple copies of HPV canintegrate in tandem repeats,62 and it may be obvious thatsuch integrations will be missed when analyzing E1 or E2deletions.
A recently described mismatch at position 3425 in theAfrican-1 HPV16 variant63 is located within the criticalregion of our HPV16E2.1 forward primer, which will prob-ably result in an unjustified E2 deletion and therefore anunjustified classification of integration for this variant. How-ever, the incidence of this variant has been found to be sig-nificant only in some African countries, whereas it was notdetected in patients with cervical carcinomas in Asia, Europeand Northern America.64
From studies with the plasmid model systems, it hasbecome clear that integration can be reliably detected when itconcerns at least 30% of the virus in a given sample. Further-more, the detection of integration was shown to be hamperedby a high-viral load, and it is therefore suggested that dilu-tion of high-viral load samples with human DNA will resultin a better detection of integration.
Analysis of the cervical cell lines showed that the selectedprobes enabled detection of viral integration in HeLa(HPV18) and SiHa (HPV16). For SiHa, a deletion of E2.1(position 3460–3512) and retention of E2.2 was expectedbased on the published data.49 We clearly detected a 90%reduction of the copy numbers for E2.1. This partial retain-ment of the E2.1 sequence was confirmed by our qPCRresults, and Roberts et al.55 also showed recently, by usingqPCR, that a small fraction of E2 could still be detected inSiHa. It is most likely that in �10% of these cells, the E2.1sequence is retained despite integration, rather than a pres-ence of episomal copies of the virus. In CaSki, integrationcould not be detected because it has been described thatthis cell line contains several tandem repeats of integratedHPV16 genomes, with retained E2 targets.50 Moreover, theE2/E6 ratio above 1 seen with qPCR even implies the pres-ence of more E2 copies compared with E6, which is inagreement with previous restriction and qPCR analysis forthis cell line.50,55 In the clinical samples, a similar correla-tion between MLPA and qPCR for the detection of viralintegration was seen. In those cases where FISH could beperformed, an episomal or integrated status of the viruscould be confirmed, whereas the MLPA assay is able todetect a mixed status of the virus in cases where FISH onlydetects integrated virus.
Gain of telomerase-related genes
Gain of the telomerase-related genes has been correlated withprogression of cervical neoplasia12,28 and prognosis of headand neck squamous cell carcinomas.13,31 It was shown thatTERT and TERC amplifications are seen in �30–40% of cer-vical carcinomas.65 In oropharyngeal carcinomas gain ofTERC was even found in �70% of the cases.13 Our MLPAassay showed a gain for TERT in all 3 cervical cell lines andalso for TERC in CaSki and SiHa, which was confirmed byFISH and the high resolution array CGH data of Lockwoodet al.37
Gain for TERT and TERC was demonstrated in 1 fresh-frozen and 1 cytological sample. In the fresh-frozen samples,we were able to confirm this by FISH analysis. However, forthe cytological samples, no cells were available for confirma-tion. In the formalin-fixed and paraffin-embedded samples sofar unexplained, aberrant signal intensity ratios were seenbetween the TERT or TERC targets on the one hand and theother human targets on the other hand, indicating that opti-mization for these parameters in paraffin-embedded samplesis needed.
In a number of clinical samples ranging from normal tocarcinoma, the MLPA assay was able to discriminate betweenhigh- and low-grade lesions based on the presence of viralintegration and/or a high-viral load, as verified by independ-ent assays. Larger numbers of samples need to be analyzed tofurther determine the clinical significance of this association.Furthermore, these results show that so far the future formatsfor the assay will be fresh-frozen tissue and cytological sam-ples. Although gain for TERT and TERC is detected in 1cytological sample, it should be taken into account that mix-ing of the tumor cells with normal cells in these samples willstrongly influence the sensitivity to detect gain of the telo-merase genes.
We can conclude that the new multiparameter HPV MLPAassay described here allows a reliable detection and quantifica-tion of HPV16 and 18. Furthermore, the procedure enables thedetection of viral integration in samples with different viralloads and can also be combined with analysis of genomic insta-bility based on gain of telomerase-related genes. Application ofthe assay in clinical samples will pave the way to improve riskassessment for patients suspected for HPV infection.
AcknowledgementsM. Reijans and G. Simons are employee and CEO of PathoFinder BV (Maas-tricht, The Netherlands), respectively. The authors acknowledge Dr. Phil-ippe Delvenne (Department of Pathology, University of Liege, CHU Sart-Tilman, Liege, Belgium), Dr. Els Meulemans (Department of Pathology,Maastricht University Medical Centre; MUMC, The Netherlands) andDr. Frank Smedts (Department of Pathology, Erasmus University MedicalCenter, Rotterdam, The Netherlands) for providing us with some clinicalsamples, Ms. Brit Mulder (PathoFinder BV) for support with the develop-ment of the MLPA probes, Ms. Annick Haesevoets and Ms. Mieke Henfling(Department of Molecular Cell Biology, MUMC) for FISH analysis and cellculturing, respectively, Dr. Will Lockwood and Dr. Wan Lam (BritishColumbia Cancer Research Centre, Vancouver, Canada) for the latest
Early
Detection
andDiagn
osis
972 MLPA assay for HPV-related cancers
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

update of the TERC and TERT array CGH data, Dr. Adriaan van den Brule(Catharina Hospital, PAMM Laboratories, Eindhoven, The Netherlands),Ms. Sandra Claessen (Department of Molecular Cell Biology, MUMC), Dr.Andrea Romano (Department of Gynaecology, MUMC) and Mr. Jos Dree-
sen (Department of Population Genetics, Genomics & Bioinformatics,MUMC) for fruitful discussions and Dr. Jan Schouten (MRC Holland, Am-sterdam, The Netherlands) for fruitful discussions and for providing us withtheMSH2 and TERT-c probes.
References
1. Scheurer ME, Tortolero-Luna G, Adler-Storthz K. Human papillomavirusinfection: biology, epidemiology, andprevention. Int J Gynecol Cancer 2005;15:727–46.
2. Hoots BE, Palefsky JM, Pimenta JM, SmithJS. Human papillomavirus type distributionin anal cancer and anal intraepitheliallesions. Int J Cancer 2009;124:2375–83.
3. Walboomers JM, Jacobs MV, Manos MM,Bosch FX, Kummer JA, Shah KV, SnijdersPJ, Peto J, Meijer CJ, Munoz N. Humanpapillomavirus is a necessary cause ofinvasive cervical cancer worldwide. J Pathol1999;189:12–19.
4. Klussmann JP, Weissenborn SJ, WielandU, Dries V, Kolligs J, Jungehuelsing M,Eckel HE, Dienes HP, Pfister HJ, FuchsPG. Prevalence, distribution, and viral loadof human papillomavirus 16 DNA intonsillar carcinomas. Cancer 2001;92:2875–84.
5. Kalantari M, Villa LL, Calleja-Macias IE,Bernard HU. Human papillomavirus-16and -18 in penile carcinomas: DNAmethylation, chromosomal recombinationand genomic variation. Int J Cancer 2008;123:1832–40.
6. Munoz N, Bosch FX, de Sanjose S, HerreroR, Castellsague X, Shah KV, Snijders PJ,Meijer CJ. Epidemiologic classification ofhuman papillomavirus types associatedwith cervical cancer. N Engl J Med 2003;348:518–27.
7. Dalstein V, Riethmuller D, Pretet JL, LeBail Carval K, Sautiere JL, Carbillet JP,Kantelip B, Schaal JP, Mougin C.Persistence and load of high-risk HPV arepredictors for development of high-gradecervical lesions: a longitudinal Frenchcohort study. Int J Cancer 2003;106:396–403.
8. Cohen MA, Basha SR, Reichenbach DK,Robertson E, Sewell DA. Increased viralload correlates with improved survival inHPV-16-associated tonsil carcinomapatients. Acta Otolaryngol 2008;128:583–9.
9. Begum S, Cao D, Gillison M, Zahurak M,Westra WH. Tissue distribution of humanpapillomavirus 16 DNA integration inpatients with tonsillar carcinoma. ClinCancer Res 2005;11:5694–9.
10. Klaes R, Woerner SM, Ridder R,Wentzensen N, Duerst M, Schneider A,Lotz B, Melsheimer P, von KnebelDoeberitz M. Detection of high-riskcervical intraepithelial neoplasia andcervical cancer by amplification of
transcripts derived from integratedpapillomavirus oncogenes. Cancer Res1999;59:6132–6.
11. Arias-Pulido H, Narayan G, Vargas H,Mansukhani M, Murty VV. Mappingcommon deleted regions on 5p15 incervical carcinoma and their occurrence inprecancerous lesions. Mol Cancer 2002;1:3.
12. Hopman AHN, Theelen W, HommelbergPP, Kamps MA, Herrington CS, MorrisonLE, Speel EJM, Smedts F, Ramaekers FCS.Genomic integration of oncogenic HPVand gain of the human telomerase geneTERC at 3q26 are strongly associatedevents in the progression of uterinecervical dysplasia to invasive cancer.J Pathol 2006;210:412–19.
13. Klussmann JP, Mooren JJ, Lehnen M,Claessen SM, Stenner M, Huebbers CU,Weissenborn SJ, Wedemeyer I, Preuss SF,Straetmans JM, Manni JJ, Hopman AHN,et al. Genetic signatures of HPV-relatedand unrelated oropharyngeal carcinomaand their prognostic implications. ClinCancer Res 2009;15:1779–86.
14. Wentzensen N, von Knebel Doeberitz M.Biomarkers in cervical cancer screening.Dis Markers 2007;23:315–30.
15. Bosch FX, Lorincz A, Munoz N, Meijer CJ,Shah KV. The causal relation betweenhuman papillomavirus and cervical cancer.J Clin Pathol 2002;55:244–65.
16. Clifford G, Franceschi S, Diaz M, MunozN, Villa LL. Chapter 3: HPV type-distribution in women with and withoutcervical neoplastic diseases. Vaccine 2006;24(suppl 3):S26–34.
17. Ho GY, Bierman R, Beardsley L, Chang CJ,Burk RD. Natural history of cervicovaginalpapillomavirus infection in young women.N Engl J Med 1998;338:423–8.
18. Gravitt PE, Kovacic MB, Herrero R,Schiffman M, Bratti C, Hildesheim A,Morales J, Alfaro M, Sherman ME,Wacholder S, Rodriguez AC, Burk RD.High load for most high risk humanpapillomavirus genotypes is associated withprevalent cervical cancer precursors butonly HPV16 load predicts the developmentof incident disease. Int J Cancer 2007;121:2787–93.
19. Chen CM, Shyu MP, Au LC, Chu HW,Cheng WT, Choo KB. Analysis of deletionof the integrated human papillomavirus 16sequence in cervical cancer: a rapidmultiplex polymerase chain reactionapproach. J Med Virol 1994;44:206–11.
20. Arias-Pulido H, Peyton CL, Joste NE,Vargas H, Wheeler CM. Humanpapillomavirus type 16 integration incervical carcinoma in situ and in invasivecervical cancer. J Clin Microbiol 2006;44:1755–62.
21. Vernon SD, Unger ER, Miller DL, Lee DR,Reeves WC. Association of humanpapillomavirus type 16 integration in theE2 gene with poordisease-free survivalfrom cervical cancer. Int J Cancer 1997;74:50–6.
22. Ziegert C, Wentzensen N, Vinokurova S,Kisseljov F, Einenkel J, Hoeckel M, vonKnebel Doeberitz M. A comprehensiveanalysis of HPV integration loci inanogenital lesions combining transcriptand genome-based amplificationtechniques. Oncogene 2003;22: 3977–84.
23. zur Hausen H. Papillomaviruses andcancer: from basic studies to clinicalapplication. Nat Rev Cancer 2002;2:342–50.
24. Cheung JL, Lo KW, Cheung TH, Tang JW,Chan PK. Viral load. E2 gene disruptionstatus, and lineage of humanpapillomavirus type 16 infection in cervicalneoplasia. J Infect Dis 2006;194:1706–12.
25. Badaracco G, Venuti A, Sedati A, MarcanteML. HPV16 and HPV18 in genital tumors:significantly different levels of viralintegration and correlation to tumorinvasiveness. J Med Virol 2002;67:574–82.
26. Tonon SA, Picconi MA, Bos PD, ZinovichJB, Galuppo J, Alonio LV, Teyssie AR.Physical status of the E2 human papillomavirus 16 viral gene in cervical preneoplasticand neoplastic lesions. J Clin Virol 2001;21:129–34.
27. Hopman AHN, Smedts F, Dignef W,Ummelen M, Sonke G, Mravunac M,Vooijs GP, Speel EJM, Ramaekers FCS.Transition of high-grade cervicalintraepithelial neoplasia to micro-invasivecarcinoma is characterized by integrationof HPV 16/18 and numerical chromosomeabnormalities. J Pathol 2004;202:23–33.
28. Heselmeyer K, Schrock E, du Manoir S,Blegen H, Shah K, Steinbeck R, Auer G,Ried T. Gain of chromosome 3q definesthe transition from severe dysplasia toinvasive carcinoma of the uterine cervix.Proc Natl Acad Sci USA 1996;93:479–84.
29. Huang FY, Kwok YK, Lau ET, Tang MH,Ng TY, Ngan HY. Genetic abnormalitiesand HPV status in cervical and vulvarsquamous cell carcinomas. Cancer GenetCytogenet 2005;157:42–8.
Early
Detection
andDiagn
osis
Theelen et al. 973
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

30. Matthews CP, Shera KA, McDougall JK.Genomic changes and HPV type incervical carcinoma. Proc Soc Exp Biol Med2000;223:316–21.
31. Singh B, Stoffel A, Gogineni S, Poluri A,Pfister DG, Shaha AR, Pathak A, Bosl G,Cordon-Cardo C, Shah JP, Rao PH.Amplification of the 3q26.3 locus isassociated with progression to invasivecancer and is a negative prognostic factorin head and neck squamous cellcarcinomas. Am J Pathol 2002;161:365–71.
32. Badaracco G, Venuti A. Physical status ofHPV types 16 and 18 in topographicallydifferent areas of genital tumours and inpaired tumour-free mucosa. Int J Oncol2005;27:161–7.
33. Peitsaro P, Johansson B, Syrjanen S.Integrated human papillomavirus type 16is frequently found in cervical cancerprecursors as demonstrated by a novelquantitative real-time PCR technique. JClin Microbiol 2002;40:886–91.
34. van den Brule AJ, Pol R, Fransen-Daalmeijer N, Schouls LM, Meijer CJ,Snijders PJ. GP5þ/6þ PCR followed byreverse line blot analysis enables rapid andhigh-throughput identification of humanpapillomavirus genotypes. J Clin Microbiol2002;40:779–87.
35. Huang SL, Chao A, Hsueh S, Chao FY,Huang CC, Yang JE, Lin CY, Yan CC,Chou HH, Huang KG, Huang HJ, Wu TI,et al. Comparison between the HybridCapture II Test and an SPF1/GP6þ PCR-based assay for detection of humanpapillomavirus DNA in cervical swabsamples. J Clin Microbiol 2006;44:1733–9.
36. Luft F, Klaes R, Nees M, Durst M,Heilmann V, Melsheimer P, von KnebelDoeberitz M. Detection of integratedpapillomavirus sequences by ligation-mediated PCR (DIPS-PCR) and molecularcharacterization in cervical cancer cells. IntJ Cancer 2001;92:9–17.
37. Lockwood WW, Coe BP, Williams AC,MacAulay C, Lam WL. Whole genometiling path array CGH analysis ofsegmental copy number alterations incervical cancer cell lines. Int J Cancer 2007;120:436–43.
38. Schouten JP, McElgunn CJ, Waaijer R,Zwijnenburg D, Diepvens F, Pals GRelative quantification of 40 nucleic acidsequences by multiplex ligation-dependentprobe amplification. Nucleic Acids Res2002;30:e57.
39. Reijans M, Dingemans G, Klaassen CH,Meis JF, Keijdener J, Mulders B, Eadie K,van Leeuwen W, van Belkum A,Horrevorts AM, Simons G. RespiFinder: anew multiparameter test to differentiallyidentify fifteen respiratory viruses. J ClinMicrobiol 2008;46:1232–40.
40. Redman R, Rufforny I, Liu C, WilkinsonEJ, Massoll NA. The utility of p16(Ink4a)in discriminating between cervicalintraepithelial neoplasia 1 andnonneoplastic equivocal lesions of thecervix. Arch Pathol Lab Med 2008;132:795–9.
41. Klussmann JP, Weissenborn SJ, WielandU, Dries V, Eckel HE, Pfister HJ, FuchsPG. Human papillomavirus-positivetonsillar carcinomas: a different tumorentity? Med Microbiol Immunol 2003;192:129–32.
42. Wilting SM, Snijders PJ, Meijer GA, YlstraB, van den Ijssel PR, Snijders AM,Albertson DG, Coffa J, Schouten JP, van deWiel MA, Meijer CJ, Steenbergen RD.Increased gene copy numbers atchromosome 20q are frequent in bothsquamous cell carcinomas andadenocarcinomas of the cervix. J Pathol2006;209:220–30.
43. Boshart M, Gissmann L, Ikenberg H,Kleinheinz A, Scheurlen W, zur Hausen H.A new type of papillomavirus DNA, itspresence in genital cancer biopsies and incell lines derived from cervical cancer.EMBO J 1984;3:1151–7.
44. Seedorf K, Krammer G, Durst M, Suhai S,Rowekamp WG. Human papillomavirustype 16 DNA sequence. Virology 1985;145:181–5.
45. Lee GM, Thornthwaite JT, Rasch EM.Picogram per cell determination of DNAby flow cytofluorometry. Anal Biochem1984;137:221–6.
46. Kulmala SM, Shabalova IP, Petrovitchev N,Syrjanen KJ, Gyllensten UB, Johansson BC,Syrjanen SM. Type-specific persistence ofhigh-risk human papillomavirus infectionsin the New Independent States of theformer Soviet Union Cohort Study CancerEpidemiol Biomarkers Prev 2007;16:17–22.
47. Hopman AH, Ramaekers FC. Processingand staining of cell and tissue material forinterphase cytogenetics. Curr ProtocCytom 2001; Chapter 8:Unit 8 5.
48. Hopman AHn, Kamps MA, Smedts F,Speel EJM, Herrington CS, Ramaekers FCS.HPV in situ hybridization: impact ofdifferent protocols on the detection ofintegrated HPV. Int J Cancer 2005;115:419–28.
49. Meissner JD. Nucleotide sequences andfurther characterization of humanpapillomavirus DNA present in the CaSkiSi Ha and. He La cervical carcinoma celllines. J Gen Virol 1999;80:1725–33.
50. Baker CC, Phelps WC, Lindgren V, BraunMJ, Gonda MA, Howley PM. Structuraland transcriptional analysis of humanpapillomavirus type 16 sequences incervical carcinoma cell lines. J Virol 1987;61:962–71.
51. Ruutu MP, Kulmala SM, Peitsaro P,Syrjanen SM. The performance of theHPV16 real-time PCR integration assay.Clin Biochem 2008;41:423–8.
52. Cricca M, Morselli-Labate AM, VenturoliS, Ambretti S, Gentilomi GA, Gallinella G,Costa S, Musiani M, Zerbini M. ViralDNA load, physical status and E2/E6 ratioas markers to grade HPV16 positivewomen for high-grade cervical lesions.Gynecol Oncol 2007;106: 549–57.
53. Lorincz AT, Richart RM. Humanpapillomavirus DNA testing as an adjunctto cytology in cervical screening programs.Arch Pathol Lab Med 2003;127:959–68.
54. Saunier M, Monnier-Benoit S, Mauny F,Dalstein V, Briolat J, Riethmuller D,Kantelip B, Schwarz E, Mougin C, PretetJL. Analysis of human papillomavirus type16 (HPV16) DNA load and physical statefor identification of HPV16-infectedwomen with high-grade lesions or cervicalcarcinoma. J Clin Microbiol 2008;46:3678–85.
55. Roberts I, Ng G, Foster N, Stanley M,Herdman MT, Pett MR, Teschendorff A,Coleman N. Critical evaluation of HPV16gene copy number quantification by SYBRgreen PCR. BMC Biotechnol 2008;8:57.
56. Vinokurova S, Wentzensen N, Kraus I,Klaes R, Driesch C, Melsheimer P,Kisseljov F, Durst M, Schneider A, vonKnebel Doeberitz M. Type-dependentintegration frequency of humanpapillomavirus genomes in cervical lesions.Cancer Res 2008;68:307–13.
57. Rousseau MN, Costes V, Konate I, NagotN, Foulongne V, Ouedraogo A, Van dePerre P, Mayaud P, Segondy M. Viral loadand genomic integration of HPV 16 incervical samples from HIV-1-infected anduninfected women in Burkina Faso. J MedVirol 2007;79:766–70.
58. Corden SA, Sant-Cassia LJ, Easton AJ,Morris AG. The integration of HPV-18DNA in cervical carcinoma. Mol Pathol1999;52:275–82.
59. Hudelist G, Manavi M, Pischinger KI,Watkins-Riedel T, Singer CF, Kubista E,Czerwenka KF. Physical state andexpression of HPV DNA in benign anddysplastic cervical tissue: different levels ofviral integration are correlated with lesiongrade. Gynecol Oncol 2004;92:873–80.
60. Badaracco G, Rizzo C, Mafera B, Pichi B,Giannarelli D, Rahimi SS, Vigili MG,Venuti A. Molecular analyses andprognostic relevance of HPV in head andneck tumours. Oncol Rep 2007;17:931–9.
61. Kim SH, Koo BS, Kang S, Park K, Kim H,Lee KR, Lee MJ, Kim JM, Choi EC, ChoNH. HPV integration begins in thetonsillar crypt and leads to the alteration
Early
Detection
andDiagn
osis
974 MLPA assay for HPV-related cancers
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC

of p16. EGFR and c-myc during tumorformation. Int J Cancer 2007;120:1418–25.
62. Wagatsuma M, Hashimoto K, MatsukuraT. Analysis of integrated humanpapillomavirus type 16 DNA in cervicalcancers: amplification of viral sequencestogether with cellular flanking sequences.J Virol 1990;64:813–21.
63. Jiang M, Baseman JG, Koutsky LA, FengQ, Mao C, Kiviat NB, Xi LF. Sequencevariation of human papillomavirus type 16and measurement of viral integration byquantitative PCR. J Clin Microbiol 2009;47:521–6.
64. Yamada T, Manos MM, Peto J, Greer CE,Munoz N, Bosch FX, Wheeler CM. Humanpapillomavirus type 16 sequence variation
in cervical cancers: a worldwideperspective. J Virol 1997;71:2463–72.
65. Zhang A, Maner S, Betz R, Angstrom T,Stendahl U, Bergman F, Zetterberg A,Wallin KL. Genetic alterations in cervicalcarcinomas: frequent low-levelamplifications of oncogenes are associatedwith human papillomavirus infection. Int JCancer 2002;101:427–33.
Early
Detection
andDiagn
osis
Theelen et al. 975
Int. J. Cancer: 126, 959–975 (2010) VC 2009 UICC
Copyright © 2022 FDOKUMEN




















