
A Holocene record of climate, vegetation change and peat bog development, east Otago, South Island,...
16
JOURNAL OF QUATERNARY SCIENCE (1999) 14 (3) 239–254 CCC 0267-8179/99/030239–16$17.50 Copyright 1999 John Wiley & Sons, Ltd. A Holocene record of climate, vegetation change and peat bog development, east Otago, South Island, New Zealand MATT S. McGLONE* and JANET M. WILMSHURST Landcare Research, PO Box 69, Lincoln, New Zealand Matt S. McGlone and Janet M. Wilmshurst, 1999. A Holocene record of climate, vegetation change and peat bog development, east Otago, South Island, New Zealand, J. Quaternary Sci., Vol. 14, pp. 239–254. ISSN 0267–8179 Received 3 August 1998; revised 26 October 1998; accepted 30 October 1998 ABSTRACT: A Holocene record of pollen, macrofossils, testate amoebae and peat humification is presented from a small montane bog. Sediment accumulation began before 9000 yr BP, but peat growth not until ca. 7000 BP. From 12 000 to 7000 yr BP, a shrub–grassland dominated under a dry climate, with increasing conifer forest and tall scrub from ca. 9600 yr BP. At 7000 yr BP a dense montane–subalpine low conifer forest established under a moist, cool climatic regime. Between 7000 and 700 yr BP the bog surface was shrubby, tending to be dry but with highly variable surface wetness. The catchment was affected by major fire at least four times between 4000 and 1000 yr BP. Both fire and bog surface wetness may have been linked to ENSO-caused variations in rainfall. Cooler, cloudier winters and disturbance by fire promoted the expansion of the broadleaf tree Nothofagus menziesii between 4000 yr BP and 1300 yr BP at the expense of the previous conifer forest–scrub vegetation. Polynesian fires (ca. 700 yr BP) reduced the vegetation to tussock grassland and bracken. Deforestation did not markedly affect the hydrology of the site. European pastoralism since ad 1860 has increased run-off and rising water tables in the bog have led to a Sphagnum-dominated cover. Copyright 1999 John Wiley & Sons, Ltd. KEYWORDS: ENSO; fire; humification; precipitation; testate amoebae. Introduction Pollen analytical studies of the late-glacial and Holocene in New Zealand have established a broad regional framework of terrestrial vegetation change and wetland development (McGlone et al., 1993). Although climatic inferences made from this framework have been used as evidence for southw- est Pacific atmospheric circulation changes (Markgraf et al., 1992; McGlone et al., 1992), the basis for them remains highly generalised and somewhat speculative. We have undertaken a detailed multiproxy study of a small wetland in the southern South Island of New Zealand in order to examine the links between terrestrial vegetation and wetland vegetation in relation to the evolution of the broader regional pattern of climatic change. A small catch- ment and wetland was chosen to enable close connections to be made between the dryland and wetland vegetation changes. The bog lies in the upland Glendhu catchment in * Correspondence to: M. S. McGlone, Landcare Research New Zealand, Canterbury Agriculture and Science Centre Ltd, P.O. Box 69, Gerald Street, Lincoln 8152, New Zealand. Contract grant sponsor: Foundation for Research, Science and Technology, New Zealand Contract grant number: C09525 a transition zone where the mild, cool, moist climates of the Otago coastline are replaced by the dry, variable, more continental climates of the interior. It is also at the northern edge of an area where precipitation is highly responsive to El Nin ˜ o–Southern Oscillation (ENSO) fluctuations (Salinger, 1980; Salinger et al., 1996). Our primary aim was to derive a record of fire and bog surface wetness variability from the bog sediments, and to link and compare this to changes in terrestrial vegetation in the surrounding catchment. Pollen, macrofossil and charcoal analyses are used to chart changes in both the dryland and the wetland vegetation. Several proxy indicators of surface wetness of the bog were utilised in this study: peat humification, testate amoebae and detailed macrofossil analyses. From these various techniques a more reliable assessment of climatic change in southern New Zealand than previously possible is presented. Glendhu study site The Glendhu drainage basin (169°459E, 45°509S) is in the headwaters of the Waipori River at the eastern edge of a schist plateau that extends inland from the east Otago coast
-
Upload
landcareresearch -
Category
Documents
-
view
0 -
download
0
Transcript of A Holocene record of climate, vegetation change and peat bog development, east Otago, South Island,...

JOURNAL OF QUATERNARY SCIENCE (1999) 14 (3) 239–254 CCC 0267-8179/99/030239–16$17.50Copyright 1999 John Wiley & Sons, Ltd.
A Holocene record of climate, vegetationchange and peat bog development, eastOtago, South Island, New ZealandMATT S. McGLONE* and JANET M. WILMSHURSTLandcare Research, PO Box 69, Lincoln, New Zealand
Matt S. McGlone and Janet M. Wilmshurst, 1999. A Holocene record of climate, vegetation change and peat bog development, east Otago, South Island, NewZealand, J. Quaternary Sci., Vol. 14, pp. 239–254. ISSN 0267–8179
Received 3 August 1998; revised 26 October 1998; accepted 30 October 1998
ABSTRACT: A Holocene record of pollen, macrofossils, testate amoebae and peat humificationis presented from a small montane bog. Sediment accumulation began before 9000 yr BP, butpeat growth not until ca. 7000 BP. From 12 000 to 7000 yr BP, a shrub–grassland dominatedunder a dry climate, with increasing conifer forest and tall scrub from ca. 9600 yr BP. At 7000yr BP a dense montane–subalpine low conifer forest established under a moist, cool climaticregime. Between 7000 and 700 yr BP the bog surface was shrubby, tending to be dry but withhighly variable surface wetness. The catchment was affected by major fire at least four timesbetween 4000 and 1000 yr BP. Both fire and bog surface wetness may have been linked toENSO-caused variations in rainfall. Cooler, cloudier winters and disturbance by fire promotedthe expansion of the broadleaf tree Nothofagus menziesii between 4000 yr BP and 1300 yr BPat the expense of the previous conifer forest–scrub vegetation. Polynesian fires (ca. 700 yr BP)reduced the vegetation to tussock grassland and bracken. Deforestation did not markedly affectthe hydrology of the site. European pastoralism since ad 1860 has increased run-off and risingwater tables in the bog have led to a Sphagnum-dominated cover. Copyright 1999 JohnWiley & Sons, Ltd.
KEYWORDS: ENSO; fire; humification; precipitation; testate amoebae.
Introduction
Pollen analytical studies of the late-glacial and Holocene inNew Zealand have established a broad regional frameworkof terrestrial vegetation change and wetland development(McGlone et al., 1993). Although climatic inferences madefrom this framework have been used as evidence for southw-est Pacific atmospheric circulation changes (Markgraf et al.,1992; McGlone et al., 1992), the basis for them remainshighly generalised and somewhat speculative.
We have undertaken a detailed multiproxy study of asmall wetland in the southern South Island of New Zealandin order to examine the links between terrestrial vegetationand wetland vegetation in relation to the evolution of thebroader regional pattern of climatic change. A small catch-ment and wetland was chosen to enable close connectionsto be made between the dryland and wetland vegetationchanges. The bog lies in the upland Glendhu catchment in
* Correspondence to: M. S. McGlone, Landcare Research New Zealand,Canterbury Agriculture and Science Centre Ltd, P.O. Box 69, Gerald Street,Lincoln 8152, New Zealand.Contract grant sponsor: Foundation for Research, Science and Technology,New ZealandContract grant number: C09525
a transition zone where the mild, cool, moist climates ofthe Otago coastline are replaced by the dry, variable, morecontinental climates of the interior. It is also at the northernedge of an area where precipitation is highly responsive toEl Nino–Southern Oscillation (ENSO) fluctuations (Salinger,1980; Salinger et al., 1996). Our primary aim was to derivea record of fire and bog surface wetness variability from thebog sediments, and to link and compare this to changes interrestrial vegetation in the surrounding catchment. Pollen,macrofossil and charcoal analyses are used to chart changesin both the dryland and the wetland vegetation. Severalproxy indicators of surface wetness of the bog were utilisedin this study: peat humification, testate amoebae and detailedmacrofossil analyses. From these various techniques a morereliable assessment of climatic change in southern NewZealand than previously possible is presented.
Glendhu study site
The Glendhu drainage basin (169°459E, 45°509S) is in theheadwaters of the Waipori River at the eastern edge of aschist plateau that extends inland from the east Otago coast

240 JOURNAL OF QUATERNARY SCIENCE
(Fig. 1). The drainage basin is approximately 220 ha in areaand ranges in elevation from 460 to 680 m. The surroundinguplands are characterised by broad, undulating mountaincrests between 400 m and 1000 m in altitude, cut by steep-sided deeply incised river valleys. The flanks of the rangesare thickly clad with loess and solifluction deposits (Wood,1969; Bruce et al., 1973). The climate of the region tendsto be cool and moist. The broad-topped uplands interceptthe prevailing southwesterly to southerly rain-bearing winds.Low cloud and fog are common, especially between altitudesof 600 and 800 m (Carter, 1994). At the study site the meanannual temperature is 7.4°C (1980–1990; Fahey and Watson,1991). Mean annual rainfall in the region ranges between1600 mm on the coastal mountains declining to 600 mminland. An annual rainfall at the site of ca. 1350 mm (range958–1572 mm) is distributed relatively evenly throughout theyear, typically as small events of long duration and lowintensity (Fahey and Watson, 1991). Infrequent snow showersoccur from May through to November (late autumn to earlysummer). Snow lies at the site for no more than 20 days ayear, and usually for less than a week at a time (AlexWatson, pers. comm.).
Before human settlement, dense forest and scrub coveredthe region up to the treeline at around 1000–1100 m (Blissand Mark, 1974). Fire and pastoralism have reduced bothforest and scrub to small remnant patches in an unbrokentract of grassland. Chionochloa rigida (narrow-leaved snowtussock) is abundant throughout, as it tolerates a wide range
Figure 1 Glendhu study area.
Copyright 1999 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 14(3) 239–254 (1999)
of soils and climatic conditions (Carter, 1994). Forest rem-nants are largely of Nothofagus menziesii (silver beech), withmuch rarer stands of Podocarpus hallii (mountain totara). Inmontane and subalpine areas, scrub species Hebe odora,and Ozothamnus leptophyllus are common, invading snowtussock in many areas. The Glendhu study site is in aChionochloa rigida tussock grassland, with the introducedgrasses Agrostis capillaris (browntop) and Anthoxanthumodoratum (sweet vernal) a prominent secondary component.Stunted shrubs of Leptospermum scoparium (manuka) andOzothamnus leptophyllus, low growing small-leaved Copro-sma spp., Muehlenbeckia complexa, and dwarf shrubs Per-nettya macrostigma and Cyathodes fraseri are common onopen ground, while Pteridium esculentum (bracken), Lyco-podium fastigiatum, Anisotome aromatica, Wahlenbergiaalbomarginata, Gnaphalium traversii and Hieracium spp. arefrequent between the tussocks.
Small valley head, riparian, cushion and Sphagnum bogsare present throughout the east Otago uplands from about400 m altitude, becoming ubiquitous above 700 m (Johnson,1988). The Glendhu bog is at the head of a small (3.6 ha)first-order catchment (Fig. 1), and forms part of a long-term experimental hydrological study begun in 1979 of theheadwaters of the Waipori River (O’Loughlin et al., 1984;Fahey and Watson, 1991; Fahey and Jackson, 1997). Thecentre of the 0.39 ha bog is dominated by mosaics of Empod-isma minus (wire rush), and Sphagnum cristatum and S.falcatulum, forming an even surface. The better drained

241HOLOCENE VEGETATION AND CLIMATE IN NEW ZEALAND
edges are largely in Chionochloa rigida tussocks, togetherwith Carex and Juncus spp., and a number of herbaceousspecies (Forstera tenella, Pratia angulata, Gunnera monoica),prostrate shrubs (Pernettya macrostigma, Coprosmaperpusilla), small shrubs (Hebe odora and Ozothamnusleptophyllus) and a fern (Polystichum vestitum).
Field and laboratory methods
A core was taken from the deepest, central part of the bog(Fig. 1). The upper 50 cm was too fibrous to sample cleanlyby coring, so peat blocks were dug out . A D-section corerwas used to sample from 50 cm to 190 cm. The basal siltsand silty peats were sampled with a 5-cm chamber Hillerborer because they were too dense for the D-section corerto penetrate. The Hiller borer core was sampled in the fielddirectly into glass vials, and the peat blocks and D-sectioncore were wrapped in polythene in the field. All sampleswere stored at 5°C until subsampled for analysis.
Pollen extraction and analysis
A calibrated spoon was used to take 1–2 cm3 subsamplesfrom the cores and prepared for pollen analysis using stan-dard procedures (hot 10% KOH, 40% HF and acetolysis) asoutlined in Moore et al. (1991). Samples were spiked in theinitial processing step with tablets of known amounts ofexotic Lycopodium for calculation of palynomorph concen-trations. Pollen counts were continued until a dryland pollensum of at least 250 grains was reached. Percentage calcu-lations are based on dryland pollen types (including ferns).The area of microscopic charcoal fragments (referred to hereas microcharcoal) was quantified in two classes (, 50 mmand . 50 mm) using the point count method of Clark (1982).The results are expressed as a charcoal index (hits per 11points per field of view, calculated as a percentage of thepollen sum) to avoid spurious fluctuations resulting fromchanging accumulation rates. Pollen results were calculatedand drafted using tilia and tilia.graph (Grimm, 1992).
Humification, density and loss on ignition
The semi-quantitative sodium hydroxide extraction technique(Blackford and Chambers, 1993) was used to determine thedegree of peat decomposition in the core at 5-cm intervals,using 0.2 g dry weight of pulverised peat sample. The per-centage transmission measured at 540 nm on a spectrophoto-meter is a measure of the humic acids present in the sample,higher values of light transmission indicating less humifi-cation.
Wet and dry density and loss on ignition were measuredfor fresh peat samples at 5-cm intervals (Bengtsson and Enell,1986). Bulk density was measured as grams per millilitre offresh peat. Loss-on-ignition samples were dried at 90°C for24 h, weighed and then ashed in a muffle furnace for 4 hat 550°C and loss of sample weight calculated as a percent-age of original dry weight.
Copyright 1999 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 14(3) 239–254 (1999)
Macrofossils
Fresh peat samples were prepared for macrofossil analysisat 5-cm intervals using a modified version of the semi-quantitative methods outlined in Barber (1991) and Barberet al. (1994). Peat samples (1 cm vertical thickness; volume5 ml) were washed with a fine jet of distilled water thougha 125 mm mesh sieve. A thin suspension was placed in aglass Petri dish. Any large clumps of organic matter weregently teased apart to release the component parts. Charcoalfragments (referred to here as macrocharcoal) were separatedinto two groups according to longest dimension: , 0.5–1 mm and . 1 mm. Macrofossils were identified under 10–1003 magnification, and percentage cover estimated at 103magnification for 15 fields of view spaced across the dish.Percentage cover of the various peat and macrofossil compo-nents were estimated, i.e. rare (, 5%), occasional (5–10%),frequent (11–25%), common (26–50%) and abundant (50–75%). Using the median value of each class, the 15 measure-ments per sample were averaged to provide a final percent-age estimate of abundance.
Testate amoebae
Subsamples of fresh peat (10 ml in unhumified Sphagnum,5 ml for other sediments) were processed for testates follow-ing the methods recommended by Hendon and Charman(1997), following Tolonen (1986) and Warner (1990).Samples were boiled in distilled water for 10 min in orderto disaggregate the peat, then washed through 300 mm and15 mm mesh sieves. The contents remaining on the 15 mmmesh sieve were centrifuged at 3000 r.p.m. for 5 min, thesupernatant poured off and a drop of the concentratemounted in glycerol. Testate amoebae were counted at 4003magnification. The following works were used to assist withidentifications: Hoogenraad and de Groot (1948), Van Oye(1956), Grospietsch (1958), Corbet (1973), Ogden and Hed-ley (1980), and Ellison and Ogden (1987), and further noteson identifications are given in Appendix I. At least 100testate amoebae were counted in samples from the top 80cm; concentrations were too low in samples below this levelfor a representative count, and these were scanned onlyfor presence/absence.
Radiocarbon dating
Plant fragments, wood and charcoal were used for dating.The weakly organic silts in the basal sediments of this sitewere not dated because of a high risk of contamination.Plant fragments and charcoal samples were pre-treated withacid and alkali washes and vacuum dried before dating. Anextract of cellulose was used to date the wood sample(NZA 6918). Dating was carried out by accelerator massspectrometry at the Radiocarbon Dating Laboratory, Instituteof Geological and Nuclear Sciences, Lower Hutt.
Results
Sediment stratigraphy and dates
Radiocarbon dates are presented in Table 1. The uppermostdate (NZA 8722; 264 6 63 14C yr BP) is almost certainly

242 JOURNAL OF QUATERNARY SCIENCE
contaminated by younger carbon and therefore is rejected.It is centred on 63.5 cm where the first signs of a sustainedrise of the bracken and charcoal curves occur, an eventassociated with first Maori settlement. However, it is approxi-mately 450 years younger than dates for similar fire eventselsewhere in the Otago district (McGlone and Wilmshurst,in press). It seems likely that the Glendhu region also wouldhave been affected by these fires, which removed forest andscrub cover from nearly all the surrounding districts. Insuch a condensed sequence as that of the Glendhu bog,contamination by down-growth of small roots from the sur-face is always possible, although the selection of identifiablesedge leaves and seeds should have minimised this effect.It is also possible that fires may have removed some of theolder surface peat. The question of when Maori deforestationbegan in this area will be clarified only by further datesfrom other nearby bogs. We have here assumed a date of700 yr BP for this level. Measurements of bulk density,humification and loss on ignition are plotted in Fig. 2.
Stratigraphy as noted in the field is as follows.Depth (cm) Sediment type0–15 Unhumified intact Sphagnum cristatum and
Empodisma minus.15–22 Broken down and compacted unhumified
Sphagnum.22–35 Dark yellow-brown partially humified
Sphagnum.35–47 Finely fibrous partially humified Sphagnum and
sedge.47–62 Finely fibrous grey-brown humified sedge peat
with fine charcoal fragments.62–75 Yellow-brown sedge peat with fine charcoal
fragments.75–100 Grey-brown well humified sedge peat with
fine charcoal fragments.100–140 Well humified red-brown pulpy sedge peat
with wood (,5 mm diameter) and finecharcoal fragments.
140–162 Humified yellow-brown sedge peat with wood.162–175 Dark-brown well humified sedge peat with
coarse (,5 mm diameter) charcoal at 165–167 cm.
175–190 Humified yellow-brown sedge peat.190–205 Fine grey-brown silty peat with few sedge
remains.205–207 Sand and gravel.207–220 Finely fibrous grey-brown silty peat with little
sedge.220–250 Olive-grey fine silt.
Micro- and macrofossil stratigraphy andinterpretation
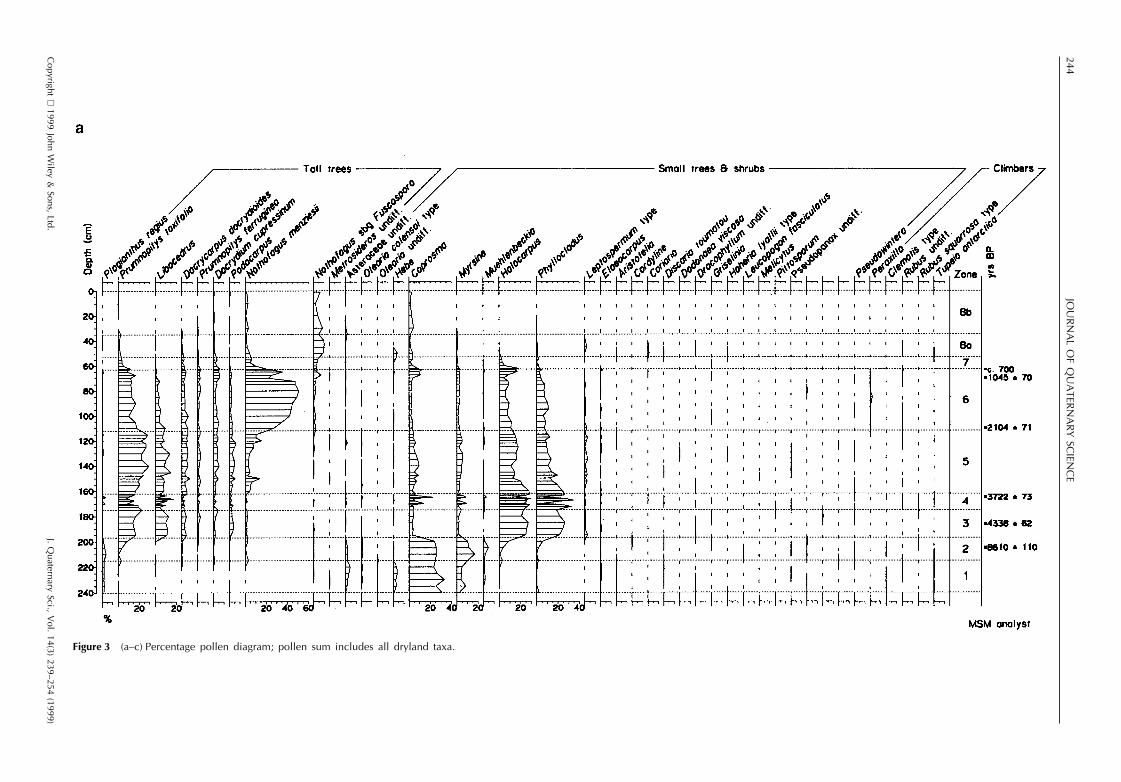
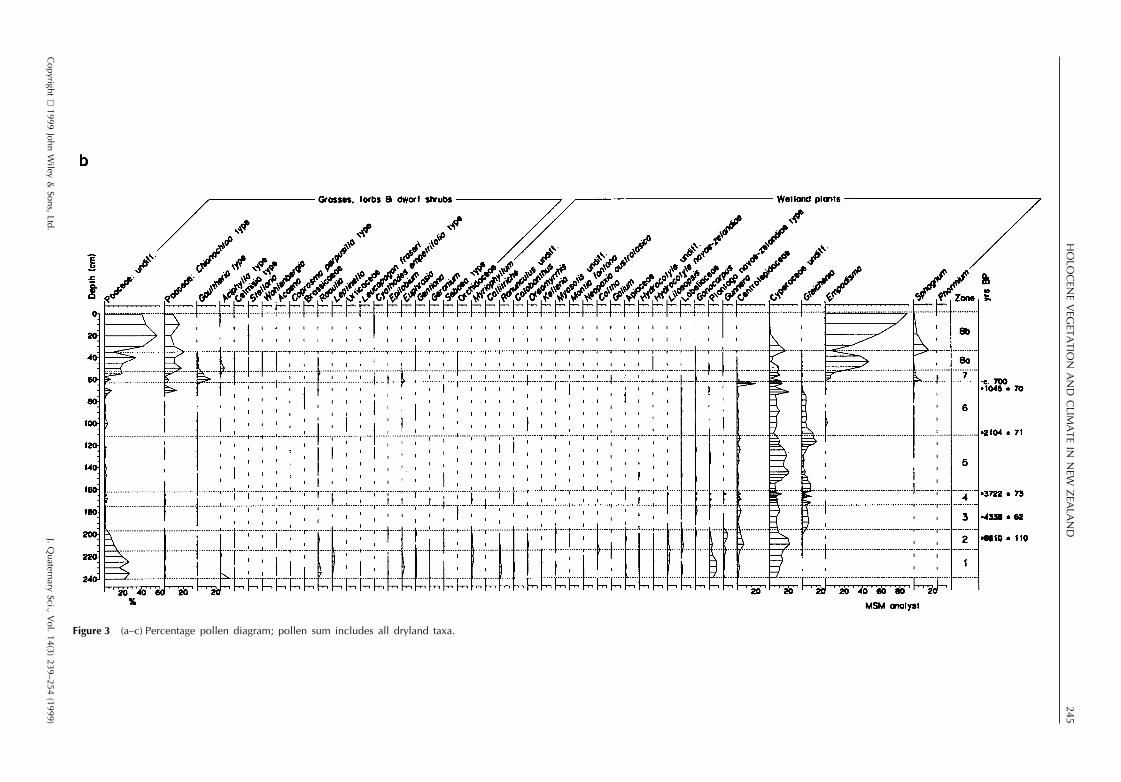
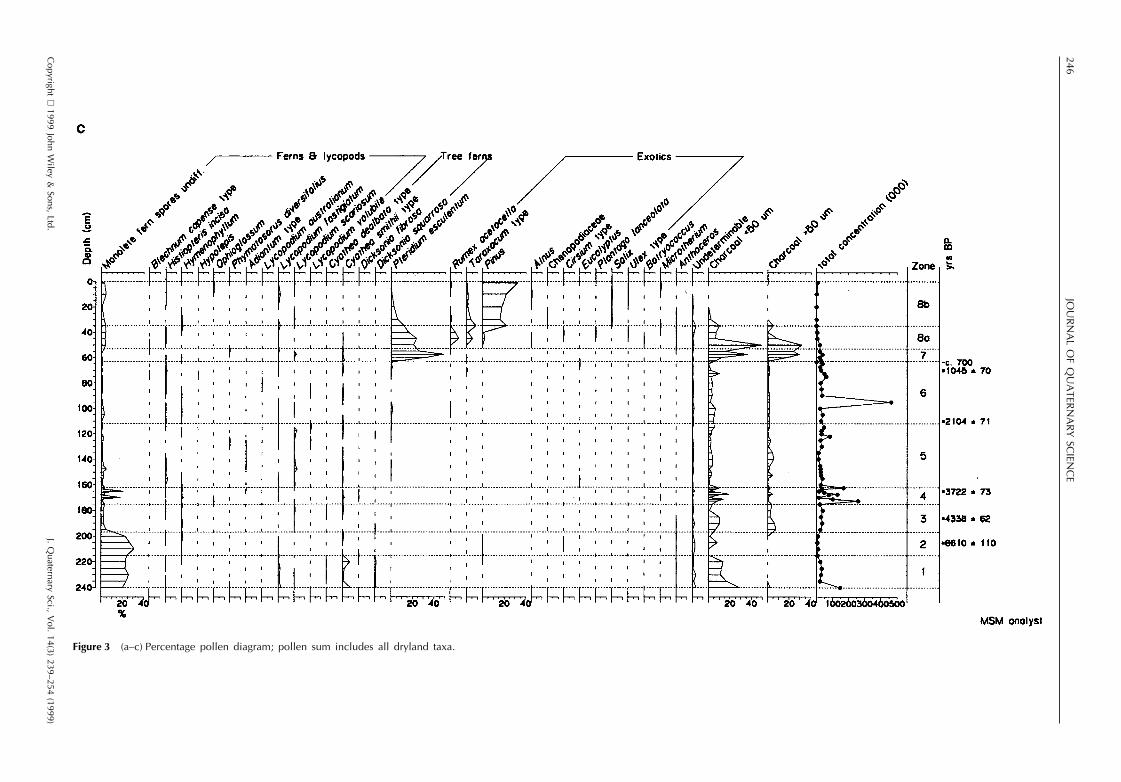
The results of pollen, spore and microcharcoal analyses arepresented in Fig. 3a–c, testate amoebae and desmids in Fig.4 and macrofossils in Fig. 5. The profile has been dividedinto eight zones, based on changes in the local landscapedominants in the pollen record. Estimated dates for the zoneboundaries are calculated by linear extrapolation from thesix radiocarbon dates. The sediment accumulation was esti-mated as beginning at ca. 12 000 yr BP on the basis ofradiocarbon dates for similar profiles in the Southland–Otago region.
Copyright 1999 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 14(3) 239–254 (1999)
Zone 1 (240–215 cm; 12 000–9580 yr BP)
Poaceae (undifferentiated types with pollen size , 40 mm),Asteraceae and the shrubs Coprosma, Myrsine and Hebedominate the pollen. Tree pollen is , 5%, consisting mainlyof Plagianthus regius. Abundant wetland pollen types includeCyperaceae, Centrolepidaceae, Plantago novae-zelandiae,Gunnera, Lilaeopsis and Myriophyllum. Juncus pusillus typeseeds are present. No testate amoebae were recorded. Thereis abundant charcoal in both the micro- and , 0.5–1 mmmacroclass range. The sediments are silty, with loworganic concentration.
An open mosaic of stunted shrubs and short tussockgrassland occupied the catchment. Fire occurred in thecatchment, as large sized charcoal is unlikely to have beentransported any distance (Whitlock and Millspaugh, 1996).The high influx of silt suggests incomplete vegetation cover.Very little forest was present, and groves of Plagianthus arelikely to have formed the only substantial forest cover. Therichly diverse wetland flora indicates a low turf of spreadingherbs, sedges and rushes on a damp, silty substrate.Myriophyllum indicates the presence of shallow pools andrivulets.
Zone 2 (215–197 cm; 9580–6900 yr BP)
Aboreal pollen percentages (mainly Prumnopitys taxifoliaand Nothofagus menziesii) double in this zone from , 5%to 10–15%. Myrsine and conifer scrub (Halocarpus andPhyllocladus) increase whereas Poaceae decline steadily.The main wetland types of the last zone remain common,with Centrolepidaceae and Cyperaceae increasing. Juncuspusillus type seeds and leaves of Sphagnum cristatum occur.No testates were recorded. Charcoal is scarce. Sedimentsare increasingly organic.
Forest and scrub increased during zone 2. Dense scrubspread in the local catchment while patches of podocarpand Nothofagus menziesii forest established in the region.A similar wetland vegetation to that of zone 1 persisted, butthe presence of Sphagnum suggests an increasingly stable,higher water table.
Zone 3 (197–175 cm; 6900–4000 yr BP)
Pollen percentages of tall podocarp trees, Libocedrus andPhyllocladus, rise sharply at the beginning of this zone fromca. 20% to 60%. Halocarpus also increases abruptly, butother scrub types and grass sink to low levels, only Copro-sma, Myrsine and Muehlenbeckia maintain a consistent pres-ence. The high diversity of wetland pollen types is muchdiminished, and Centrolepidaceae, Cyperaceae and Gleich-enia dominate. Juncus pusillus type seeds and Halocarpusbidwillii leaves occur, but the macrofossil record is domi-nated by sedge remains. A dessication-resistant testate amo-eba, Hyalosphenia subflava, occurs, but no other types. Bothmicro- and macrocharcoal are common in all categories.Sediments are highly organic peats from the first few centi-metres of this zone upwards. Rates of sediment accumulationacross the zone 2–3 boundary were slow (ca. 0.04 mm yr21)increasing with the commencement of peat growth by nearlyan order of magnitude (ca. 0.34 mm yr21), a growth ratethat was maintained with little variation until zone 7.
Prumnopitys taxifolia–Dacrycarpus dacrydioides dominantbroadleaved podocarp forest grew further downslope (below300–400 m altitude on the basis of their present-day range),

243HOLOCENE VEGETATION AND CLIMATE IN NEW ZEALAND
Table 1 Radiocarbon dates from Glendhu Bog. Calibrations made using INSCAL 4.0 (Rodger Sparks, pers. comm., 1998)
Depth (cm) Date 14C yr NZA number Dated material Calibrated age 68%BP confidence interval
years ad/bc
63.5 264 6 63a 8722 Selected sedge leaves and ad 1527–1556 plusCarex seeds ad 1633–1672 plus
ad 1782–1796 plus1947–1953
70 1045 6 70 6896 Selected sedge remains ad 900–903 plus, 120 mm (Empodisma root ad 964–1030hairs removed)
110 2104 6 71 6918 Wood 194–32 bc
165 3722 6 73 6919 Charcoal fragments of 2198–1979 bc
Halocarpus bidwilli
186 4338 6 62 8721 Halocarpus bidwilli leaves and 3028–2973 bc plusCarex seeds 2931–2888 bc
205 8610 6 110 6890 Fine organics 7849–7822 bc plus. 125 mm 7703–7506 bc
aProbably contaminated with younger carbon; most likely age for this depth is c. 700 14C years BP.
Figure 2 Bulk density, loss on ignition and humification curves. No values recorded for humification below 185 cm, because of highvalues of inorganic matter.
while subalpine–montane low forest of Libocedrus bidwillii–Podocarpus hallii and conifer scrub occupied the Glendhucatchment. Nothofagus menziesii is poorly represented inthe modern pollen rain, and the continuing , 5% pollenrepresentation indicates it was well established in the generalarea. A much lower diversity wetland cover established, inwhich stunted shrubs of Halocarpus bidwillii and Leptosper-mum formed an open canopy over a ground cover of thewetland fern Gleichenia, Sphagnum and sedge. Macro-charcoal indicates local fires, corroborated by the find ofPhyllocladus charcoal dated at 4500 6 50 14C yr BP(WkB5534) at 40 cm depth under surface colluvium at thefoot of the southwest slope above the bog (T. Webb, pers.comm.)
Copyright 1999 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 14(3) 239–254 (1999)
Zone 4 (175–162 cm; 4000–3650 yr BP)
This zone is distinguished by three well-defined charcoalpeaks, each of which is followed by a sharp decline in thepollen percentages of Libocedrus and the lower two also bydeclines of Phyllocladus, Halocarpus and Gleichenia, andincreases of Plagianthus regius, Muehlenbeckia, Coprosma,Myrsine, Poaceae and monolete fern spores. Sphagnum cris-tatum and Drepanocladus fluitans leaves, and Halocarpusbidwillii stems now occur regularly, whereas sedge remainsdecrease. The peat has low light transmission.
At least three major fires burnt through the local catchmentand across the bog, reducing the cover of tall trees andconifer scrub, and inducing brief reversions to shrub–
244JO
UR
NA
LO
FQ
UA
TERN
AR
YSC
IENC
E
Figure 3 (a–c) Percentage pollen diagram; pollen sum includes all dryland taxa.
Copyright
1999John
Wiley
&Sons,
Ltd.J.
Quaternary
Sci.,V
ol.14(3)
239–254(1999)
245H
OLO
CEN
EV
EGETA
TION
AN
DC
LIMA
TEIN
NEW
ZEA
LAN
D
Figure 3 (a–c) Percentage pollen diagram; pollen sum includes all dryland taxa.
Copyright
1999John
Wiley
&Sons,
Ltd.J.
Quaternary
Sci.,V
ol.14(3)
239–254(1999)
246JO
UR
NA
LO
FQ
UA
TERN
AR
YSC
IENC
E
Figure 3 (a–c) Percentage pollen diagram; pollen sum includes all dryland taxa.
Copyright
1999John
Wiley
&Sons,
Ltd.J.
Quaternary
Sci.,V
ol.14(3)
239–254(1999)

247H
OLO
CEN
EV
EGETA
TION
AN
DC
LIMA
TEIN
NEW
ZEA
LAN
D
Figure 4 Percentage testate amoebae diagram.
Copyright
1999John
Wiley
&Sons,
Ltd.J.
Quaternary
Sci.,V
ol.14(3)
239–254(1999)

248JO
UR
NA
LO
FQ
UA
TERN
AR
YSC
IENC
E
Figure 5 Plant macrofossil remains presented as estimated percentage abundance.
Copyright
1999John
Wiley
&Sons,
Ltd.J.
Quaternary
Sci.,V
ol.14(3)
239–254(1999)

249HOLOCENE VEGETATION AND CLIMATE IN NEW ZEALAND
grassland, similar to that of zones 1–2. High humificationbut continuing steady peat growth argues for a relatively drybut productive bog surface. The arrival of Sphagnum crista-tum may have been favoured by the partial destruction ofthe woody cover on the bog.
Zone 5 (162–112 cm; 3650–2180 yr BP)
Tall podocarp forest pollen percentages increase, Prumnopi-tys taxifolia reaching its highest levels in the upper half ofthe zone. Conifer scrub slowly declines from 51 to 24%.Nothofagus menziesii increases throughout the zone, withperiods of rapid increase followed by partial regression.Leptospermum pollen is consistently represented. Leaves ofthe aquatic moss Drepanocladus fluitans, and Sphagnumcristatum, Halocarpus bidwillii stems and Juncus pusillustype seeds are recorded. Testate amoebae Hyalospheniasubflava and Nebela certesi are present. Desmid zygosporesof cf. Micrasterias mahabuleshwarensis var. mahabuleshwar-ensis and M. denticulata types (Croasdale and Flint, 1986)make their first appearance in this zone. Microcharcoal iswell represented but macrocharcoal falls to its lowest levels.Sediment light-transmission increases in the lower 20 cm ofthe zone, but then reverts to values close to those of zone 4.
Nothofagus menziesii played a dynamic and increasinglyimportant role in local forests during this zone, progressivelyreplacing Libocedrus and conifer scrub. Its periods of rapidincrease occur at times of decreasing humification and there-fore surface wetness. Further indications of increasing surfacewetness are the abundance of the aquatic moss Drepano-cladus fluitans and Sphagnum cristatum here and in zone 6.
Sphagnum is a major peat-former for only brief periods:it cannot tolerate a fluctuating water table regime (Johnson,1988), and this may explain why it was abundant onlysporadically. Desmids are freshwater algae that have almostentirely aquatic habitat requirements and are unable to with-stand desiccation (Brook, 1981). Sexual reproduction is rela-tively rare in desmids and the formation of zygospores byconjugation is thought to be a means of survival whendesiccation threatens (Coesel, 1974). Presence of theirzygospores therefore suggests ephemeral, shallow pools.
Low abundance of Sphagnum and fluctuating water tablesexplains the low diversity of testate amoebae (Warner, 1987).The only testate amoeba taxon that is consistently recordedin zones 5 and 6, Hyalosphenia subflava, favours relativelydry, shrubby bogs in New Zealand surface samples(Charman, 1997). Hyalosphenia subflava has been reportedas characteristic of peatlands that have been drained or areovergrown with heather in Finland (Tolonen, 1986), andof virgin xerophilous treed Sphagnum peatlands in Canada(Warner, 1987). Warner (1988) suggests that H. subflava isadapted to well-drained bogs, major seasonal fluctuations ofwater table and dry late summers.
The continuing presence of the shrub conifer Halocarpusbidwillii and the southern heath species Leptospermum sco-parium on the bog confirms the inferences made from thetestate amoebae.
Zone 6 (112–63 cm; 2180–700 yr BP)
Nothofagus menziesii pollen rises steadily to its peak valuesof 40–50% of the pollen sum. Pollen of the mistletoe Perax-illa is recorded only in this zone. Prumnopitys taxifoliadecreases to half its peak values, and long-distance-derivedNothofagus subgenus Fuscospora pollen is recorded consist-
Copyright 1999 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 14(3) 239–254 (1999)
ently for the first time. Leptospermum falls to much lowerpercentages than in zone 5. Macrocharcoal (but notmicrocharcoal) increases, including two marked peaks (at100–95 cm and 80 cm), the lower coinciding with increasedgrass and Pteridium. At the zone top (70–65 cm) a minorpeak of charcoal accompanies a major but temporary declinein Nothofagus menziesii, Libocedrus and Halocarpus. Halo-carpus bidwillii stems, Nothofagus menziesii leaves, Empod-isma root hairs and pollen, and Juncus pusillus seeds arerecorded. Sedge remains are at their lowest levels. Sphagnumcristatum leaves are common, and Drepanocladus fluitansleaves are present in some samples. There is a continuinggood representation of desmid zygospores, but a low abun-dance and diversity of testate amoebae at the top of thezone.
Nothofagus menziesii was now the dominant tree speciesin the catchment and very likely in the montane and subalp-ine forests. Records of its leaves, and the occurrence of thepoorly dispersed pollen of Peraxilla, a mistletoe that mostoften has Nothofagus as a host, suggests it grew right at thesite. Conifer forest and scrub species remained prominent inthe catchment, and Halocarpus bidwillii grew on the bog,although the decline in Leptospermum indicates that thewoody cover was not as complete. The bog was at itsmost complex; pollen, macrofossils, charcoal and desmidzygospores indicating constantly changing vegetation andsurface wetness, and minor local fires. A major fire at aroundca. 1100 yr BP (70–65 cm) partially destroyed the localNothofagus forest and permitted the expansion of tussockgrassland. Libocedrus bidwillii did not recover, but Notho-fagus menziesii forest re-expanded, and Halocarpus andCoprosma scrub became abundant. This fire had a majoreffect on bog vegetation. Centrolepis cushions dominatedfor perhaps as long as 400 yr after this event and one ofthe previous dominants, Gleichenia, did not recover. Peatgrowth was slow after the fire (ca. 0.2 mm yr21).
Zone 7 (63–53 cm; 700 yr BP–ad 1860)
All forest taxa fall sharply, with the exception of long-distance-dispersed Nothofagus subgenus Fuscospora; scrubis greatly reduced with the exception of Hebe and Gaul-theria; and charcoal of all size classes becomes abundant,and charcoalised moss leaves occur. There is a sustainedrise of Pteridium and Poaceae, the small grain undifferen-tiated types expanding first, followed by the Chionochloatype. Empodisma pollen increases, whereas Sphagnum leavesare absent. Micrasterias denticulata type zygospores declineand do not occur above this zone. The previously mostabundant testate amoeba, Hyalosphenia subflava, declines;Heleopera sylvatica dominates, and Arcella discoides andHeleopera sphagni are present.
This zone records the terminal destruction of forest andmontane scrub by fire. High levels of micro- and macrochar-coal in this zone and charcoalised leaves of Sphagnumindicate that fires swept the surface of the bog frequently.The rate of peat growth during zone 7 is uncertain becauseof the problematic date for the beginning of Polynesiandeforestation. If the 264 14C yr BP date is accepted, peataccumulation is a rapid 0.58 mm yr21. However, if the mostlikely date for forest burn-off was ca. 700 yr BP (as elsewherein Otago and Southland, and as accepted here) peat accumu-lation was a slow 0.17 mm yr21 in this zone, similar to therate since the fire at 1100 yr BP. As both bulk density andpercentage light transmission are low compared with theirvalues in zone 8, it seems most probable that the accumu-

250 JOURNAL OF QUATERNARY SCIENCE
lation rate remained low in zone 7. Empodisma begins todominate the bog vegetation in zone 7, whereas Drepano-cladus fluitans and Juncus pusillus type drop out. Gaultheriatype pollen, probably representing Pernettya macrostigma, asprawling subshrub that can tolerate drier bog conditions,was common. The testate fauna represents mixed moisturetolerances, with the wetter types represented by Arcelladiscoides, Difflugia oblonga, Heleopera sphagni and Nebelacertesi, and the relatively dry species by Heleopera sylvatica.Desmid semi-cells and zygospores were also present butrare. Water tables therefore rose, but only slightly.
Zone 8a (53–35 cm; ad 1860–1960)
Exotic taxa occur in this zone for the first time in the profile(apart from occasional contaminants at much lower levels),Poaceae increase and all shrub types and Pteridiumdecrease. Charcoal is abundant in all size classes in the firstfew centimetres, but then declines to low levels at the zoneupper boundary. Sphagnum and Empodisma dominate thepollen. The macrofossil flora is dominated by poorly hum-ified Sphagnum cristatum leaves and Empodisma root hairs,stems and leaves. The diversity of testate amoebae increases;rotifer egg cases (Habrotrocha angusticollis), desmid semi-cells (Cosmarium) and desmid zygospores are abundant. Asharp rise in percentage light transmission and bulk densitypoint to wetter, less humified sediments.
European settlement of the district led to more regularburning, destruction of the previously abundant Pteridiumand scrub, and encouragement of tussock grassland. Testatesbecome more diverse in zone 8: on average 15 taxa wererecorded per sample, compared with only six taxa in thesample at the top of zone 6. The bog surface had becomewetter and the water table higher and more stable, anddominance by Sphagnum cristatum provided a more suitablehabitat for testate amoebae than the previous sedge-dominated associations.
Zone 8b (35–0 cm; ad 1960–1990)
There is a sharp increase in Pinus, but a decline in exoticweeds, and Pteridium vanishes from the record. Charcoalfalls to low levels. Macrofossils are dominated by Sphagnum,Empodisma and fungal hyphae. Difflugia A and E typesincrease in this zone and then decline towards the surface asxerophilous species, including Corythion dubium, Euglypharotunda, Assulina muscorum and Nebela militaris expand.No desmid semi-cells or zygospores were recorded. Bulkdensity and percentage light-transmission rise to the highestvalues for the entire profile in the first 15 cm of this zone,but then fall towards the surface.
The pine pollen curve permits dating of this zone. Plantingof exotic pine forest (mainly Pinus radiata) began at around1900, increased greatly in the 1920s and then began steadycontinuous expansion in the 1940s (New Zealand Ministryof Works, 1967). Exotic forests were established in theTapanui–Blue Mountain area, 25 km west of Glendhu, inthe 1920s. Berwick Forest, 15 km to the east, and moredistant forests in the coastal lowlands were planted in the1950s, and the Glendhu forest, which now surrounds thesite, in the late 1970s and early 1980s. Allowing 15–20years for planted stands to become sexually mature, pinepollen should have first become a significant part of theregional pollen rain in the 1930s, with a major increase inthe 1960s, followed by a very recent increase in the late
Copyright 1999 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 14(3) 239–254 (1999)
1990s. The vegetation changes reflect improved farming andreduction of the indigenous cover. The testate assemblagesand fungal hyphae and peat characteristics indicate relativelydrier bog surfaces towards the present.
Discussion and conclusions
Wetland history and moisture regime
During zones 1 and 2, that is from possibly as early as12 000 yr BP to 6900 yr BP, a silty, damp, sedge turf witha diverse flora occupied the study site of Glendhu basin.Dominance of the sediments by silt and occasional lensesof sand and gravel point to a catchment that was only partlyvegetated, and to a lack of a permanently high water table,although temporary pools formed at times. Such wetlandscan exist under relatively dry climates, and were widespreadduring this period in the southern South Island (McGloneand Bathgate, 1983; McGlone et al., 1997; Vandergoes et al.,1997). A bog with an open scrub canopy formed at thebeginning of zone 3 at around 7000 yr BP. Whereas peatformation occurred earlier between 10 000 and 9000 yr BPalong the Southland and Otago coastlines (McIntyre andMcKellar, 1970; McGlone and Bathgate, 1983) and in highaltitude wetlands with large catchments, it appears to havebeen delayed in bogs with small catchments in inland Otagountil around 7000 yr BP (McGlone et al., 1995, 1997). Amajor increase in annual rainfall is the only likely expla-nation for the formation of the bog.
During zone 4, fires burnt across the bog at least threetimes, temporarily causing reversion to vegetation similar tothat of zones 1 and 2, and initiating first major expansionof Sphagnum cristatum. Zones 5 and 6 (3650–700 yr BP)record a long period of constant fluctuation in surface moist-ure status. Although the bog retained its open scrub cover,and the testate amoebae fauna indicate continuing dry con-ditions, there were brief periods of wetter bog surfaces andshallow temporary ponds formed from time to time. Thedecline of wetland scrub (Halocarpus bidwillii andLeptospermum) and the presence of Empodisma during zone6 (2180–700 yr BP), and decreasing peat humification sug-gest a slight permanent increase in water table caused by along-term increase in annual rainfall.
After the arrival of Maori (most probably around 700 yrBP) the catchment was subjected to frequent fire, perhapsas much as several times a decade. The scrub cover of thebog was gradually eliminated and Empodisma became adominant. However, the dry nature of the bog surface didnot change fundamentally until the 1860s, with the arrival ofEuropean pastoralists. A switch to a Sphagnum–Empodismavegetation cover occurred at that time, and abundance oftestate amoebae indicate a wetter, more stable water regime.This marked change in bog fauna and flora at the time ofEuropean settlement seems to be related primarily toincreased year-round water inflow to the bog. Vegetationcover has a highly significant effect on water yield from acatchment, as demonstrated by the large increases in wateryield when native forests are converted to pasture, reflectingboth the lower evaporation loss from pasture and the lowerpermeability of the soil due to trampling (see discussion inFahey and Rowe, 1992, pp. 272–273). Unmodified tussockcover, however, has evaporative characteristics much thesame as forest (Fahey and Rowe, 1992), which may explainwhy Polynesian deforestation barely affected the bog surface

251HOLOCENE VEGETATION AND CLIMATE IN NEW ZEALAND
wetness. The burning and heavy grazing that accompaniedEuropean pastoralism, however, opened up the previous talltussock and bracken canopy, converting a proportion of thecatchment to pasture. A depression in water yield of 20%per year and a marked decrease in the peak flow rate ofsmall rain storms has accompanied a partial reafforestationwith pine of the adjacent experimental catchment atGlendhu (Fahey and Watson, 1991), showing that significanteffects can be expected with changes in vegetation structure.
There is a trend to somewhat drier conditions in thesurface, indicated by the composition of the testate amoebaefauna, increasing fungal hyphae and peat humification inthe latter part of zone 8b (Fig. 2). This may reflect increasedevaporation resulting from the post-1945 increase in tem-perature of ca. 0.5°C, and prolonged ENSO droughts of thelast two decades, countering the recent overall increase inprecipitation in the southwest of the South Island (Salingeret al., 1996). It would be difficult, however, to separate thiseffect from the possibility that changed catchment manage-ment in the 1970s led to reduced runoff.
Dryland history and climate change
The open stunted grassland–shrublands of zones 1 and 2(ca. 12 000–7000 yr BP) are typical of the late-glacial periodin southern New Zealand (McIntyre and McKellar, 1970;McGlone and Bathgate, 1983; McGlone et al., 1995; Vander-goes et al., 1997). Although the Glendhu site is only 40 kmfrom the coast, the timing of the changes and vegetationcomposition resemble more closely those of the mountainsof the semi-arid interior. As there is no stream input to thesite, the source of the silt and gravel must have been slope-wash off the loess-rich regolith mantle of the catchmentslopes. Present-day sediment yield from the catchment isvery low (1.0 m3 km22 yr21) and derives mostly from bankcollapse and channel scouring during flooding (O’Loughlinet al., 1984). We suggest that the site had patchy vegetationcover and was subject to severe winter frost during zones1 and 2, thus providing a source of fine sediment. A rela-tively dry, frosty climate therefore prevailed at this sitethroughout the late-glacial and early Holocene.
Increasing rainfall and annual temperatures promoted thespread of forest in the region, between 9600 and 7000 yrBP (zone 2), but low scrub and grassland remained abundantat the site. Mean annual temperatures throughout this periodwere similar to, if not higher than at present (McGlone et al.,1993), and thus lower precipitation is likely to have been theprimary cause of slow forest spread to the area. Prumnopitystaxifolia, the lowland forest dominant, is typical of drierregions of New Zealand (Wardle, 1991). The dominant treesand shrubs in the upland conifer vegetation (Libocedrus,Podocarpus, Phyllocladus and Halocarpus) will tolerate awide range of climate and soil conditions (Wardle 1969,1991). Podocarpus hallii, Phyllocladus alpinus and Halo-carpus bidwillii all tolerate low soil moisture, and thesespecies were once abundant in the dry interior of Otago(McGlone and Moar, 1998). Libocedrus bidwillii is delayedrelative to the other species, and establishes at the site onlyat the time that peat formation begins in the bog. Libocedrusis characteristic of humid to superhumid climates, with highrainfall and frequent fogs, and is often found on wet organicsoils (Ogden and Stewart, 1995).
We conclude that the early Holocene in this area,although warm, had a relatively dry climate that confinedtall forest to the moist coastal zone, allowing scrub–grassland
Copyright 1999 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 14(3) 239–254 (1999)
to persist inland. McGlone et al. (1997) have argued fromthe absence of alpine mires in some central Otago sitesduring the early Holocene that annual rainfall may havebeen as much as a third lower than at present.
Over southeastern Australia, a greatly expanded subtropi-cal high-pressure zone kept full glacial and late-glacial cli-mates dry until 12 000 yr BP. From then on climates becamewetter as the high-pressure zone shifted gradually towardsthe Equator, to lie northwards of its present position by 6000yr BP (Harrison and Dodson, 1993). This scenario fits thesouthern New Zealand vegetation pattern: a southwards dis-placement of the subtropical high during the late-glacial andearly Holocene would have produced dry climates overall,with relatively frosty winters, and stable summers moderatedby mist and fog from enhanced northeasterly airflow.
The continuing increase of Prumnopitys taxifolia through-out the lower half of zone 5 (3700–3000 yr BP) while localforest and scrub types remain static or decline, hints at arising altitudinal limit for lowland forest. This may reflectwarmer summers under a regime of increasingly seasonalclimates (McGlone et al., 1993). Nothofagus menziesii hadbeen present in the general area since at least 9500 yr BP,and must have survived along the Otago coast during theglaciation (McGlone et al., 1996). However, only after thefires of zone 4 (3700–4000 yr BP) did this species makesignificant headway against the local conifer scrub and for-est. Two distinct upsurges followed by regression in the faceof conifer competition occurred, the final local expansionoccurring between 2200 yr BP and 1300 yr BP. At the sametime Nothofagus menziesii expanded into the lower altitudesites, replacing Prumnopitys taxifolia conifer/hardwood for-est. The rapid rise of Nothofagus menziesii at Glendhu ataround 2200 yr BP after an initial increase at around 4000yr BP is matched by similar timing elsewhere in Otagoand Southland (McIntyre and McKellar, 1970; McGlone andBathgate, 1983; McGlone et al., 1995, 1997; Vandergoeset al., 1997). This mid- to late-Holocene increase of well-separated populations of Nothofagus menziesii arguesstrongly for a regional climatic explanation for the change.
Nothofagus menziesii, like other Nothofagus species, toler-ates low winter insolation, low summer humidity, and isfavoured by disturbance (Ogden et al., 1996; Leathwick, inpress; Leathwick et al., 1998). We argue that decreasingwinter solar radiation, mainly through increased rain andcloudiness during the winter months, gave it a competitiveedge over Libocedrus bidwillii and Podocarpus hallii in themontane zone, and over Prumnopitys taxifolia in thelowlands. Increased winter and early spring precipitation—suggested by the ponding of water on the bog surface anddecreasing peat humification—leading to better earlysummer growing conditions may have improved the survivor-ship of fast-growing Nothofagus menziesii seedlings inrelation to slower maturing conifer scrub seedlings.
The northwards retreat of the southern edge of the sub-tropical high from its expanded late-glacial to early Holoceneposition is likely to have increased rainfall through stimulat-ing more frontal activity. East Otago has a well-markedwinter rainfall maximum, owing to a winter increase infrontal activity in the southwesterly flow (de Lisle andBrowne, 1968). We surmise that after 7000 yr BP andintensifying towards 2000 yr BP, southwesterly air flowincreased, bringing wetter, cooler winters.
El Nino–Southern Oscillation events may have begunaffecting New Zealand after 7000 yr BP, increasing thevariability of rainfall and producing prolonged severedroughts (McGlone et al., 1992). The fire outbreaks of zone4 (4000–3700 yr BP) — when several fires burnt across the

252 JOURNAL OF QUATERNARY SCIENCE
local catchment — and continuing fires, culminating in themajor event of 1100 yr BP that temporarily destroyed thelocal forests, are likely to be drought-related. The 1100 yrBP fire occurred at a time of slow peat growth, and theseveral fires during zone 4 coincided with or immediatelyfollowed periods of high concentrations of palynomorphs inthe sediments, hence periods of slow peat growth.
The onset of fire outbreaks and spread of Nothofagusmenziesii follow the expansion of glaciers in the SouthernAlps, which began at around 5000 yr BP after a long period(ca. 9000 to 5000 yr BP) of inactivity (Gellatly et al., 1988).At least 11 episodes of glacial expansion have been recordedsince 5000 yr BP. Behaviour of glacier termini is stronglylinked with circulation changes: southerly and southwesterlyairflows tend to bring cooler temperatures and cloud to theSouthern Alps, and in summer strongly reduce ablation; ifthe subtropical high lies over the southern South Island insummer, rather than further north, ablation is increased(Fitzharris et al., 1992). El Nino–Southern Oscillation press-ure fluctuations lead to stronger westerlies and southwest-erlies and cooler weather in El Nino phases, whereas duringLa Nina phases the air flow anomaly is more northerly andnortheasterly (Gordon, 1985). Therefore, El Nino episodestend to favour glacial advances, especially if they occur insummer, and bring rain to the west and far south andsoutheast of the South Island. La Nina tends to produce adrier far southeastern South Island. Thus, east Otago tendsto be wetter than normal during El Nino events, and drierduring La Ninas (Gordon, 1985). The Glendhu record is ofinsufficient resolution to demonstrate the presence of theshort (ca. 4 yr) ENSO cycles directly. We suggest, however,that ENSO fluctuations are likely to have contributed to thevariability observed in the bog surface wetness between4000 and 1000 yr BP. In particular, runs of unusually wetwinters associated with El Ninos would have led to theformation of pools, whereas unusually dry summers associa-ted with La Ninas lowered water tables and dried out pools.
The major disruption of vegetation caused by early Maorisettlement and then European pastoralism makes it difficultto assess the possibility of climatic change over the last700 yr.
Deforestation and settlement
Recovery from the 1100 yr BP fire event was partial andshort-lived. Nothofagus menziesii had not reached its pre-fire levels when repeated fire destroyed nearly all the forestand scrub in the Glendhu region, leading to the massiveexpansion of bracken, and then grassland. These fires fol-lowed Maori settlement of the area around 700 yr BP,as indicated by archaeological sites (Anderson, 1991). Theincrease of tall snow tussocks, as represented by Chionoch-loa type pollen, was delayed relative to the other grasses,most probably because of their slow growth rate. Repeatedfire initially encouraged bracken through destroying compet-ing woody species, but then bracken in turn was eventuallysuppressed by long-lived, tall, fire-resistant snow tussocks.Similar sequences of bracken followed by grassland domi-nance after repeated burning are seen throughout the SouthIsland (for instance, McGlone and Basher, 1995). It seemspossible that the extensive fires and rapid destruction offorest during the Maori settlement may have been assistedby continuation of the dry variable conditions that character-ised the late Holocene.
Copyright 1999 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 14(3) 239–254 (1999)
Acknowledgements We thank Barry Fahey, Trevor Webb, AlexWatson and Rob Allen (Landcare Research) for provision of unpub-lished data and helpful discussions about the vegetation, soils andhydrology of the Glendhu catchment. Alison Watkins (LandcareResearch) processed the pollen and testate samples. Elizabeth Flint(Landcare Research) helped with desmid identifications andinterpretation of their ecology, and Dan Charman (PlymouthUniversity) with testate amoebae identifications. We thank BarryFahey and Rob Allen for comments on a draft manuscript, andPeter Kershaw (Monash University) for his helpful referee comments.This work was funded by the Foundation for Research, Science andTechnology, C09525.
References
ANDERSON, A. J. 1991. The chronology of the settlement of NewZealand. Antiquity, 65, 767–795.
BARBER, K. E. 1991. Peat Stratigraphy and Climatic Change: aPalaeoecological Test of the Theory of Cyclic Bog Regeneration.Balkema, Rotterdam.
BARBER, K. E., CHAMBERS, F. M., MADDY, D., STONEMAN, R.and BREW, J. S. 1994. A sensitive high-resolution record of lateHolocene climatic change from a raised bog in northern England.The Holocene, 4, 198–205.
BENGTSSON, L. and ENELL, M. 1986. Chemical Analysis. IN:Berglund, B. E. (ed.), Handbook of Holocene Palaeoecology andPalaeohydrology, 423–451. John Wiley & Sons, Chichester.
BLACKFORD, J. J. and CHAMBERS, F. B. 1993. Determining thedegree of peat decomposition for peat-based palaeoclimatic stud-ies. International Peat Journal, 5, 7–24.
BLISS, L. C. and MARK, A. F. 1974. High-alpine environments andprimary production on the Rock and Pillar Range, Central Otago,New Zealand. New Zealand Journal of Botany, 12, 445–483.
BROOK, A. J. 1981. The Biology of Desmids—Botanical Mono-graphs, 16. Blackwell Scientific Publications, Oxford.
BRUCE, J. G., IVES, D. W. and LEAMY, M. L. 1973. Maps andSections showing the Distribution and Stratigraphy of South IslandLoess Deposits, New Zealand. New Zealand Soil Survey Report 7.
CARTER, J. 1994. Waipori Ecological District: a Survey Report forthe Protected Natural Areas Programme. Department of Conser-vation, Dunedin.
CHARMAN, D. J. 1997. Modelling hydrological relationships oftestate amoebae (Protozoa: Rhizopoda) on New Zealand peat-lands. Journal of the Royal Society of New Zealand, 27, 465–483.
CLARK, R. L. 1982. Point count estimation of charcoal in pollenpreparations and thin sections. Pollen et Spores, 24, 523–535.
COESEL, P. F. M. 1974. Desmids. I. Zygospore formation in nature(with special reference to some unusual records of zygotes). ActaBotanica Neerlandica, 23, 363–368.
CORBET, S. A. 1973. An illustrated introduction to the testateRhizopoda in Sphagnum, with special reference to the area aroundMalham Tarn,Yorkshire. Field Studies, 3, 801–838.
CROASDALE, H. and FLINT, E. A. 1986. Flora of New Zealand:Freshwater algae, Chlorophyta, Desmids with Ecological Com-ments on their Habitats, Vol. 1. Government Printer, Wellington,New Zealand.
DE LISLE, J. F. and BROWNE, M. L. 1968. The Climate and Weatherof the Otago Region, New Zealand. Miscellaneous Publications115, New Zealand Meteorological Service, New Zealand Ministryof Works, Wellington.
ELLISON, R. L. and OGDEN, C. G. 1987. A guide to the studyand identification of fossil testate amoebae in Quaternary lakesediments. Internationale Revue der Gesamten Hydrobiologie, 72,639–652.
FAHEY, B. D. and JACKSON, R. J. 1997. Hydrological impacts ofconverting native forests and grasslands to pine plantations, SouthIsland, New Zealand. Agricultural and Forest Meteorology, 84,69–82.
FAHEY, B. D. and ROWE, L. K. 1992. Land-use impacts. IN: Mosley,

253HOLOCENE VEGETATION AND CLIMATE IN NEW ZEALAND
M. P. (ed.), Waters of New Zealand, 265–284. New ZealandHydrological Society, Wellington.
FAHEY, B. D. and WATSON, A. J. 1991. Hydrological impact ofconverting tussock grassland to pine plantation, Otago, New Zea-land. Journal of Hydrology (New Zealand), 48, 1–15.
FITZHARRIS, B. B., HAY, J. E. and JONES, P. D. 1992. Behaviourof New Zealand glaciers and atmospheric circulation changesover the past 130 years. The Holocene, 2, 97–106.
GELLATLY, A. F., CHINN, T. J. H. and ROTHLISBERGER, F. 1988.Holocene glacier variations in New Zealand: a review. QuaternaryScience Reviews, 7, 227–242.
GORDON, N. D. 1985. The Southern Oscillation: a New Zealandperspective. Journal of the Royal Society of New Zealand, 15,137–155.
GRIMM, E. C. 1992. Tilia and Tilia.graph, Version 2. Springfield, Illi-nois.
GROSPIETSCH, T. 1958. Wechseltierchen (Rhizopoden). Kosmos,Stuttgart.
HARRISON, S. P. and DODSON, J. 1993. Climates of Australia andNew Guinea since 18,000 yr B.P. IN: Wright, H. E., Kutzbach,J. E., Webb III, T., Ruddiman, W. F., Street-Perrrott, F. A. and P.J. Bartlein (eds), Global Climates since the Last Glacial Maximum,265–293. University of Minnesota Press, Minneapolis.
HENDON, D. and CHARMAN, D. J. 1997. The preparation oftestate amoebae (Protozoa: Rhizopoda) samples from peat. TheHolocene, 7, 199–205.
HOOGENRAAD, H. R. and DE GROOT, A. A. 1948. Thecamoebousmoss-rhizopods from New Zealand. Hydrobiologia, 1, 28–44.
JOHNSON, P. N. 1988. Sphagnum Moss in Otago: a DiscussionReport. Botany Division, DSIR, Lincoln, New Zealand.
LEATHWICK, J. R. in press. Are New Zealand’s Nothofagus speciesin equilibrium with their environment? Journal of VegetationScience, 9, 719–732.
LEATHWICK, J. R., BURNS, B. R. and CLARKSON, B. D. 1998.Environmental correlates of tree alpha-diversity in New Zealandprimary forests. Ecography, 21, 235–246.
MARKGRAF, V., DODSON, J. R., KERSHAW, P. A., McGLONE, M.S. and NICHOLLS, N. 1992. Evolution of late Pleistocene andHolocene climates in the circum-South Pacific land areas. ClimateDynamics, 6, 193–211.
McGLONE, M. S. 1983. Polynesian deforestation of New Zealand:a preliminary synthesis. Archaeology in Oceania, 18, 11–25.
McGLONE, M. S. and BASHER, L R. 1995. The deforestation of theupper Awatere catchment, Inland Kaikoura Range, Marlborough,South Island, New Zealand. New Zealand Journal of Ecology, 19,53–66.
McGLONE, M. S. and BATHGATE, J. L. 1983. Vegetation andclimate history of the Longwood Range, South Island, New Zea-land. 12 000 B.P. to the present. New Zealand Journal of Botany21, 292–315.
McGLONE, M. S. and MOAR, N. T. 1998. Dryland Holocenevegetation history, Central Otago and the Mackenzie Basin, SouthIsland, New Zealand. New Zealand Journal of Botany, 36, 91–111.
McGLONE, M. S. and WILMSHURST, J. M. in press. Establishingthe date of Maori environmental impact in New Zealand throughpollen analysis and radiocarbon dating. Quaternary International.
McGLONE, M. S., KERSHAW, A. P. and MARKGRAF, V. 1992. ElNino/Southern Oscillation and climatic variability in Australasianand South American palaeoenvironmental records. IN: Markgraf,V. and Diaz, H. (eds), El Nino: Historical and PaleoclimaticAspects of the Southern Oscillation, 435–462. Cambridge Univer-sity Press, Cambridge.
McGLONE, M. S., SALINGER, M. J. and MOAR, N. T. 1993.Palaeovegetation studies of New Zealand’s climate since the LastGlacial Maximum. IN: Wright, H. E., Kutzbach, J. E., Webb III,T., Ruddiman, W. F., Street-Perrrott, F. A. and Bartlein, P. J.(eds), Global Climates since the Last Glacial Maximum, 294–317.University of Minnesota Press, Minneapolis.
McGLONE, M. S., MARK, A. F. and BELL, D. 1995. Late Pleistocenend Holocene vegetation history, Central Otago, South Island, NewZealand. Journal of the Royal Society of New Zealand, 25, 1–22.
Copyright 1999 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 14(3) 239–254 (1999)
McGLONE, M. S., MILDENHALL, D. C. and POLE, M. S. 1996.History and paleoecology of New Zealand Nothofagus forests.IN: Veblen, T. T., Hill, R. S. and Read, J. (eds), The Ecology andBiogeography of Nothofagus Forest, 83–130. Yale UniversityPress, New Haven.
McGLONE, M. S., MOAR, N. T. and MEURK, C. D. 1997. Growthand vegetation history of Alpine Mires on the Old Man Range,Central Otago, New Zealand. Arctic and Alpine Research, 29,32–44.
McINTYRE, D. J. and McKELLAR, I. C. 1970. A radiocarbon datedpost glacial pollen profile from Swampy Hill, Dunedin, NewZealand. New Zealand Journal of Geology and Geophysics, 13,346–349.
MOORE, P. D., WEBB, J. A. and COLLINSON, M. E. 1991. PollenAnalysis, 2nd edn. Blackwell Scientific Publications, Oxford.
NEW ZEALAND MINISTRY OF WORKS, 1967. Otago Region:National Resources Survey, Part V. Wellington, Town and CountryPlanning Branch, Ministry of Works.
OGDEN, C. G. and HEDLEY, R. H. 1980. An Atlas of FreshwaterTestate Amoebae. British Museum, London.
OGDEN, J. and STEWART, G. H. 1995. Community dynamics ofthe New Zealand conifers. IN: Enright, N. J. and Hill, R. S. (eds),Ecology of the Southern Conifers, 81–119. Melbourne UniversityPress, Melbourne.
OGDEN, J., STEWART, G. H. and ALLEN, R. B. 1996 Ecology ofNew Zealand Nothofagus forests. IN: Veblen, T. T., Hill, R. S.and Read, J. (eds), The Ecology and Biogeography of NothofagusForest, 25–82. Yale University Press, New Haven.
O’LOUGHLIN, C. L., ROWE, L. K. and PEARCE, A. J. 1984.Hydrology of mid-altitude tussock grasslands, upper Waiporicatchment, Otago I — erosion, sediment yields, and water quality.Journal of Hydrology (New Zealand), 23, 45–59.
SALINGER, M. J. 1980. New Zealand climate I. Precipitation pat-terns. Monthly Weather Review, 108, 1892–1904.
SALINGER, M. J., ALLAN, R., BINDOFF, N., HANNAH, J., LAVERY,B., LIN, Z., LINDESAY, J., NICHOLLS, N., PLUMMER, N. andTOROK, S. 1996. Observed variability and change in climate andsea level in Australia, New Zealand and the South Pacific. IN:Bouma, W. J., Pearman, G. I. and Manning, M. R. ( eds), Green-house: Coping with Climate Change, 100–126. CSIRO Publishing,Collingwood, Australia.
TOLONEN, K. 1986. Rhizopod analysis. IN: Berglund, B. E. (ed.),Handbook of Holocene Palaeoecology and Palaeohydrology, 645–666. John Wiley & Sons, Chichester.
VAN OYE, P. 1956. On the Thecamoeban fauna of New Zealandwith description of four new species and biogeographical dis-cussion. Hydrobiologia, 8, 16–37.
VANDERGOES, M. J., FITZSIMONS, S. J. and NEWNHAM, R. M.1997. Late glacial to Holocene vegetation and climate change inthe eastern Takitimu Mountains, western Southland, New Zealand.Journal of the Royal Society of New Zealand, 27, 53–66.
WARDLE, P. 1969. Biological Flora of New Zealand. 4. Phyllocladusalpinus Hook f. (Podocarpaceae) mountain toatoa, celery pine.New Zealand Journal of Botany, 7, 76–95.
WARDLE, P. 1991. Vegetation of New Zealand. Cambridge Univer-sity Press, Cambridge.
WARNER, B. G. 1987. Abundance and diversity of testate amoebae(Rhizopoda: Testaceae) in Sphagnum peatlands in southwesternOntario, Canada. Archiv fur Protistenkunde, 133, 173–189.
WARNER, B. G. 1988. Methods in Quaternary ecology #5. TestateAmoebae (Protozoa). Geoscience Canada, 15, 251–260.
WARNER, B. G. 1990. Testate amoebae (Protozoa). IN: Warner, B.G. (ed), Methods in Quaternary Ecology, 65–74. GeoscienceCanada, Reprint series 5. Geological Association of Canada, StJohns.
WHITLOCK, C. and MILLSPAUGH, S. H. 1996. Testing the assump-tions of fire-history studies: an examination of modern charcoalaccumulation in Yellowstone National Park, USA. The Holocene,6, 7–15.
WOOD, B. L. 1969. Periglacial tor topography in southern NewZealand. New Zealand Journal of Geology and Geophysics, 12,361–375.

254 JOURNAL OF QUATERNARY SCIENCE
Appendix I
Notes on testate amoebae identifications
Amphitrema flavum A type
Tests similar in all aspects to A. flavum, except for deeplyrugulate sculpturing on exterior of test.
Difflugia A type
Test more or less ovoid, lightly coated with mineral particles.Mouth more than 1/3 of test width, without a neck. Length15–25 mm.
Difflugia C type
Test straight sided, mouth almost as wide as test, usually withfungal hyphae and organic xenosomes. Length 25–40 mm.
Difflugia E type
Test pear shaped, narrowing to a neck with a small mouththat is ca. 1/3 the width of the test. Length 20–35 mm.
Difflugia P type
cf. Difflugia oblonga, but mouth and test shape almostcompletely obscured with xenosomes of pollen grains andmineral particles.
Copyright 1999 John Wiley & Sons, Ltd. J. Quaternary Sci., Vol. 14(3) 239–254 (1999)




















