
MullerMoynaK2021PhD.pdf - OUR Archive (Otago University
363
Comparative anatomy and functional morphology of the forelimb in cetaceans from New Zealand PhD thesis by Moyna K. Müller Geology Department, University of Otago, New Zealand December 2020
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of MullerMoynaK2021PhD.pdf - OUR Archive (Otago University

Comparative anatomy and functional
morphology of the forelimb in
cetaceans from New Zealand
PhD thesis by Moyna K. Müller Geology Department, University of Otago, New Zealand
December 2020

Overleaf: life reconstruction of a pygmy right whale (Caperea marginata) MM2904, a juvenile female, stranded at Karikari Peninsula in Northland, North Island, New Zealand on 05 September 2010.

Dedicated to my family who supported me unwaveringly: Robert Kydd, Katharina Müller, Peter Müller, Lorraine Still
Alan Müller and family, Finlay and Belle - and especially those I can no more thank in person
In memoriam Bert Still (right) Lotti Debrunner
Jeeps, escapades, and practical
jokes (sometimes backfiring) Intrepid adventurer and
a great sport
Teak
Most avid supervisor,
professional sheep wrestler, and expert on bones


V
Table of Contents Abstract ................................................................................................................... VIII Acknowledgements .................................................................................................. IX Introduction ................................................................................................................ 1
Chapter 1: Methods and materials Procedures .................................................................................................................. 8
Protocols and sampling methods ............................................................................ 8 Dissection ................................................................................................................ 9 Photography .......................................................................................................... 14 Preservation and storage ...................................................................................... 16
Terminology ............................................................................................................... 18 Key words and references ..................................................................................... 18 Orientation ............................................................................................................. 19 Anatomy ................................................................................................................ 21
Evaluations ................................................................................................................ 24 Measuring methods ............................................................................................... 24 Flipper shapes ....................................................................................................... 28 Age classification ................................................................................................... 28
Cultural significance and naming of stranded whales ............................................... 30 List of specimens ....................................................................................................... 31 Chapter 2: Pygmy right whale Caperea marginata Introduction ................................................................................................................ 35
History and phylogeny of the genus Caperea ....................................................... 35 Anatomical accounts of the shoulder and forelimb ................................................ 37
Results ....................................................................................................................... 37 Position of the forelimb .......................................................................................... 37 Pectoral limb .......................................................................................................... 38 Osteology .............................................................................................................. 39 Myology ................................................................................................................. 47
Discussion ................................................................................................................. 63 Flipper ................................................................................................................... 63

VI
Scapula and forelimb bones .................................................................................. 69 Musculature ........................................................................................................... 80 Wider implications .................................................................................................. 94 Future Studies ....................................................................................................... 95
Summary .................................................................................................................... 97
Plates ....................................................................................................................... 100 Chapter 3: Arnoux’s beaked whale Berardius arnuxii Introduction .............................................................................................................. 103
Evolution, distribution and habitat of beaked whales ........................................... 103 Historical records and classification of Berardius arnuxii and B. bairdii ............... 105 Anatomy and morphological characteristics of the genus Berardius ................... 108 Anatomical accounts of the shoulder and forelimb in ziphiids ............................. 109
Results ..................................................................................................................... 109 Pectoral limb ........................................................................................................ 109 Osteology ............................................................................................................. 121 Myology ............................................................................................................... 119
Discussion ................................................................................................................ 133 Flipper .................................................................................................................. 133 Scapula and forelimb bones ................................................................................ 136 Musculature ......................................................................................................... 141 Wider implications ................................................................................................ 145 Future Studies ..................................................................................................... 148
Summary .................................................................................................................. 150 Plates ....................................................................................................................... 152 Chapter 4: Functional anatomy Introduction .............................................................................................................. 156
History and phylogeny ......................................................................................... 156 Cetacean evolution .............................................................................................. 157 Anatomical accounts of the shoulder and forelimb .............................................. 163
Results ..................................................................................................................... 164 Position of the forelimb ........................................................................................ 164 Pectoral limb ........................................................................................................ 164

VII
Osteology ............................................................................................................ 169 Myology (G. griseus, P. dioptrica, and C. hectori) .............................................. 188
Discussion ............................................................................................................... 219 Evolution of the forelimb ...................................................................................... 219 Functional aspects of flipper shape ..................................................................... 253
Wider implications ............................................................................................... 289 Future Studies ..................................................................................................... 289
Summary ................................................................................................................. 290 Chapter 5: Literature review Osteology ................................................................................................................ 294
Table 1. Berardius arnuxii and other ‘Berardius’ specimens ............................... 297 Myology ................................................................................................................... 300
Table 2. Cetacean shoulder and limb muscles ................................................... 303 Synthesis and conclusion .................................................................................... 316 Literature cited ....................................................................................................... 319 Appendix Measurements ......................................................................................................... 347
Table 1. Caperea marginata ................................................................................ 347 Table 2. Berardius arnuxii ................................................................................... 349 Table 3. Other species from New Zealand .......................................................... 352

VIII
Abstract The transition of early semi-aquatic archaeocetes to fully aquatic modern whales and dolphins is documented by a wealth of fossil evidence, and one of the most profound and well-studied macroevolutionary events in mammalian history. However, despite a fairly comprehensive understanding of the cetacean evolutionary pathway, only very
few modern accounts have amalgamated arrangements of the soft tissue in the shoulder and forelimb with evolutionary aspects. In this study, a total of 62 specimens comprising 15 species have been investigated and morpho-functional characteristics of the shoulder and forelimb integrated into behavioural, physiological and phylogenetic aspects. A thorough literature research complemented these findings. In Chapter 2 Caperea new osteological characteristics, and for the first time myological arrangements, are described for the pygmy right whale (Caperea marginata), the sole member of the family Neobalaenidae. Some unique traits, only seen in fossil Cetotheriidae, give credence to a possible close affiliation with this extinct family. Other morphological attributes of the forelimb are the unique ability amongst whales to change the contour of the flipper and possibly also correlated functional attributes. Chapter 3 Berardius investigates for the first time soft tissue arrangements of the shoulder and forelimb in Arnoux’s beaked whale (Berardius arnuxii, Ziphiidae). Despite highly derived morphological, physiological and behavioural adaptations to extreme deep-diving, Arnoux’s beaked whales reveal astonishingly unspecialised flipper attributes. Chapter 4 Functional anatomy presents detailed descriptions of soft tissue arrangements in two dolphins (Grampus griseus, Cephalorhynchus hectori) and one
porpoise (Phocoena dioptrica). These and other findings of this study are then analysed and combined with previously published accounts of the cetacean and mammalian forelimb, inclusive fossil specimens. The resulting conclusions are then presented within a phylogenetic context detailing possible locomotory adaptations of early archaeocetes and arrangements of flipper morphology in fully aquatic later archaeocetes and modern cetaceans.

IX
Acknowledgements I would like to thank all those who have supported me with their enthusiasm, advice and actions in my endeavours, without their help the completion of this work would
not have been possible - Thank you
I am greatly indebted to my husband, parents and wider family for the help, support and sacrifices they
have made over the years to make the completion of this thesis possible, especially Robert Kydd, Katharina Müller, Peter Müller, Lotti Debrunner, Bert Still, and Lorraine Still; and for their support and encouragement Alan Müller and family.
I am also enormously thankful to my supervisor R. Ewan Fordyce for his guidance, technical support with material, space, organisation and opportunities, and the odd pep-talk - but most of all for always being there when needed and his great patience.
A big thank you also goes to following people: University of Otago, Geology department staff (former and current) for advice and provision of never-
ending technical demands (besides becoming great friends), especially Andrew Grebneff, Sophie White, Dianne Nyhof, and technical support from John Williams, Damian Walls, Luke Easterbrooke, Hamish Bowman, Stephen Read
University of Otago academic staff for advice, provision of specimens and/or photographic material Steve Dawson, Liz Slooten, Will Rayment, Sarah Wakes
Former colleagues for the many thought-provoking discussions and being good friends Yoshihiro Tanaka, Cheng-Hsiu Tsai, Gabriel Aguirre Fernandez, Felix G. Marx, Bobby Boessenecker, Carolina Loch Santos da Silva, Simone B. Hicks
Otago Museum staff (former and current), for their enthusiasm and great technical support in procuring and processing specimens, particularly Wendy Rowe and Emma Burns, and for their technical support Trudi Webster and Kane Fleury
Southern District Health Board in Dunedin, Radiology staff especially Rebecca Mckenzie and Tracy Holgate for their enthusiasm and going well above and beyond the call of duty. Dunedin Hospital Mammography staff and lab staff of Southland Radiology Rachel Fleck, Josie Parker and Haley Buxton for kindly making themselves and their expertise available on short notice
Tangata whenua, local communities, and many papatipu rūnaka who supported recovery from marine mammal strandings around the South Island: Manawhenua ki Mōhua (Risso’s dolphin from Anaweka), Te Rūnanga o Ōtākou (pygmy sperm whale, spectacled porpoise, from Otago Peninsula, and strap-toothed beaked whale from St Kilda), Kāti Huirapa ki Puketeraki (Arnoux’s beaked whale from Pūrākaunui), Te Rūnanga o Moeraki (pygmy sperm whale and Cuvier’s beaked whale from Katiki Beach), Te Rūnanga o Arrowhead (spectacled porpoise from Timaru), Te Rūnanga o Warawa (spectacled porpoise from Kaitōrete Spit, Birdlings Flat), Waipio (Arnoux’s beaked whale from Sandy Point), Awarua (Arnoux’s beaked whale from Ōmaui), and Ōraka Aparima rūnaka (killer whale from

X
Te Waewae Bay), and for generously hosting numerous hui at Takutai o te Tītī marae with stakeholders around marine mammal stranding protocols, and especially the expertise, guidance and support of Ramari Oliphant Stewart (MNZM), of Te Kauika Tangaroa
Massey University, Palmerston North staff (former and current) for their great support, advice and provision of rare specimens and/or dissection facilities, especially Karen Stockin (now at Albany campus) Wendi Roe and Stewart Hunter, and for encouragement and advice Allan Nutman, Keren Dittmer, Laureline Meynier, Helen McConell
Alastair Watson (retired, Oklahoma State University) for his great dissection skills and insight of anatomy Invermay Agricultural Research Centre staff Geoff Asher and Post Mortem Lab staff for providing hard
to come-by necropsy facilities and provision of specimens Department of Conservation staff for granting me access to or providing specimens and their great
hands-on attitude, particularly Jim Fyffe and Rose Cole and other DOC staff Orca Research Trust staff Ingrid Visser for the provision of specimens, enthusiasm and great insight
into the lives of orcas Te Papa Tongarewa - Museum of New Zealand (current and former) staff Anton van Helden and team
for the provision of necropsy facilities and access to the large osteological collection of the museum University of Maine staff Sentiel Rommel and University of Florida staff Alex Costidis for their insight of
cetacean anatomy Volunteers Rosalie M. Steffen and many other helpers for their enthusiasm and hands-on support

1
Introduction Cetaceans are the only extant mammals that have almost perfectly adapted to nearly every aquatic environment and successfully challenged the supremacy of the fishes in terms of aquatic adaptations and niche diversity: cetaceans have evolved into fast,
agile, powerful and intelligent top predators invading both fresh water (de Muizon, 2009) and marine environments. They inhabit coastal waters as well as the high seas (Fordyce, 2003a; Steeman et al., 2009). They have dispersed worldwide (Fordyce, 2003a; Steeman et al., 2009; Fordyce, 2013) and, despite being air-breathing, successfully conquered the deep sea (Beatty and Rothschild, 2008). Fishes (e.g. Classes Agnatha, Chondrichthyes and Osteichthyes) are the ultimate aquatic vertebrates with a history that reaches back over 500 million years (Cambrian; Shu et al., 2003). The evolution of cetaceans (Order Cetacea) on the other hand spans a mere 1/10th of this time, i.e. about 50 million years (Eocene; Fordyce, 2009a, 2013). Fishes have undergone roughly three major radiation events one of which (Sarcopterygii) gave rise to tetrapods and ultimately mammals during the Late Devonian and Triassic respectively (Chaline, 1990); Daeschler et al., 2006; Long et al., 2006). During the Mesozoic mammals persisted in marginal nocturnal niches (Dong, 1980; Benton, 1983; Prothero, 2015) until the decline of the non-avian dinosaurs and some marine reptiles at the end of this period, about 66 my ago (O’Leary et al., 2013), opened up new opportunities for mammalian diversification and invasion into new ecological niches (Romer, 1966; Alroy, 1999). However, the terrestrial origin and the evolution of mammals as quadrupeds meant that the ancestors of the cetaceans,
cursorial deer-like mammals, were at best only marginally suited to an aquatic environment (Thewissen et al., 2009; Vislobovoka, 2013). It is thus the more remarkable that cetaceans not only accomplished to invade aquatic habitats but excelled in doing so. Morphological, physiological and behavioural adaptations (Gingerich, 2012; Fordyce, 2013; McGowen et al., 2014; Goldbogen et al., 2015) allowed them to successfully coexist and compete with other aquatic predators (Perrin et al., 1973; Heithaus, 2001; Ainley et al., 2006; Kelley and Motani, 2015). Extant cetaceans exploit mammalian physical traits and intelligence to their full advantage. They utilise intelligent hunting strategies such as organised pack hunting

I n t r o d u c t i o n
2
(Pitman and Durban, 2012), or flexible preplanning of dives (Arranz et al., 2018). They also make use of specialised techniques like tail slapping (Simon et al., 2005), intentional stranding (Guinet and Bouvier, 1995; Sargeant et al., 2005) or bubble netting (D'Vincent et al., 1985; Wiley et al., 2011). Indeed, cetaceans grew to be so proficient that they became victims of their own success: while sharks can be serious
threats to smaller cetaceans (Heithaus, 2001), they are no match for the larger species. Ironically, it was members of the physeterid and delphinid families that were ultimately capable of bringing down even the largest whales (Lambert et al., 2010; Gemmell et al., 2015; Whitt et al., 2015; Wellard et al., 2016). In doing so, they also exerted enough evolutionary pressure to bring about multiple other physical and behavioural changes, e.g. anti-predatory behaviours such as migration, distinctive fight / flight responses in the absence / presence of physical adaptations to speed, or loss of ‘whistles’ in mostly smaller cetaceans (Morisaka and Connor, 2007; Ford and Reeves, 2008; Steiger et al., 2008). The evolutionary transformations taking place during the transition from land to sea disconnected modern cetaceans from their closest living relatives so profoundly that whales and dolphins were not recognised as mammals until the turn of the 17th century (Kellogg, 1928; Behrmann, 2002). Despite well-documented fossil records the debate about cetacean ancestry continued (e.g. Flower, 1883; Kellogg, 1936). With the arrival of immunology and DNA analysis cetacean lineage was narrowed down to archaic ungulates (hoofed mammals; e.g. Boyden and Gemeroy, 1950; Stanhope et al., 1996) and eventually nested within artiodactyls (even-toed ungulates): the Cetartiodactyla (e.g. Waddell et al., 1999). Furthermore, some molecular studies regarded cetaceans as a sister group of hippopotamids (Irwin and Árnason, 1994; Gatesy et al., 1997;
Milinkovitch et al., 1998), a view that was refuted by many morphological analyses that allied cetaceans closer to Mesonychia, an extinct taxon of carnivorous ungulates (e.g. Van Valen, 1966; Geisler, 2001 and Geisler and Luo, 1998; Gingerich, 1998; O'Leary and Gatesy, 2008). Eventually, at the beginning of the millennium new fossil evidence of some of the oldest archaic cetaceans emerged linking cetaceans closely to artiodactyls: the ankle structures in the hind limbs (astragalus) of these early whales matched closely those of artiodactyls (Gingerich et al., 2001a; Thewissen et al., 2001). These findings became irrefutable when a few years later corresponding ear and tooth characteristics

I n t r o d u c t i o n
3
of Indohyus, an extinct semiaquatic raoellid artiodactyl, placed cetaceans firmly within the artiodactyl clade (Thewissen et al., 2007; Uhen, 2007b) and thus ultimately eliminated any possible close relationships to Mesonychia. However, to date cetacean phylogeny remains far from resolved and new morphological and molecular data continue to shed new light on the evolutionary history of whales, dolphins and
porpoises (Marx, 2011; Gingerich, 2012). Recent morphological discoveries in fossil and extant cetaceans have added further insight into ecological and behavioural aspects of whales and their allies, both living and extinct (e.g. Geisler et al., 2017; Tsai, 2017; Tsai et al., 2017).
Fig. 1. Phylogeny of Cetacea Phylogenetic relationship of Cetacea and non-cetacean mammals. Symbols: cetacean clades encompassing two or more genera indicated by blue shading. Phylogenetic tree modified from Gatesy et al., 2012. Artwork by C. Buell, except Equus (not referenced) see
<https://equineguelph.ca/learn_objects/evolutiontimeline/equus.html>, Hippopotamus by R. Nicholls.
While there is a variety of aquatic adaptations in early cetaceans, one of the most conspicuous is the transformation of the forelegs into flippers, the loss of hind limbs and the correlated development of flukes (Thewissen et al., 2009; Gingerich, 2012).

I n t r o d u c t i o n
4
With the extreme reduction and eventual loss of the hind limbs, morphological characteristics were also lost and only the discovery of hind leg characteristics in the earliest cetaceans finally presented physical evidence of the close relationship to artiodactyls (Gingerich et al., 2001a; Thewissen et al., 2001). Fossil records of the forelimbs, particularly of fully aquatic cetaceans, are equally rare, albeit for other
reasons: There are no bony connections between the fore leg and torso, or bony features in the skeleton itself that would allow a precise placement and orientation of the shoulder blade and limb once the connective soft tissue has disintegrated (Rommel and Reynolds III, 2009). The forelimb is thus easily separated and lost from the skeleton in a decaying carcass (Liebig et al., 2003). Furthermore, in fully aquatic cetaceans some regions of the shoulder and forelimb do not ossify completely (Calzada and Aguilar, 1996; DiGiancamillo et al., 1998; Stockin et al., 2008), and as a result are usually either lost or not preserved in fossil or osteological specimens (Franc et al., 1995; Horner, 1997; Holliday et al., 2010). The tip of the acromion and coracoid process, as well as the vertebral border of the scapula and sometimes large portions of the olecranon, persist as distinct cartilaginous areas that serves as anchor point for muscles (Howell, 1937). Distal phalanges often remain partly or fully cartilaginous (epiphyseal ankylosis; Galatius and Kinze, 2003; Galatius et al., 2006). Moreover, in fully aquatic cetaceans the digits of the flipper are greatly elongated by repetition of finger elements (hyperphalangy; Cooper et al., 2007a) and can be minute even in large animals, compounding the problem of preservation. Discovery and recognition of the first fossil whales dates back almost 400 years (e.g. Scilla, 1670; Harlan, 1834; Cuvier, 1835; Gibbes, 1847; Duvernoy, 1851b). Anatomical descriptions of extant cetaceans also emerged around this time (e.g. Tyson, 1680; Hunter and Banks, 1787; Stannius, 1849) including some detailed, and often intricately
illustrated, osteological and myological descriptions of the forelimb (e.g. Flower, 1867; Carte and Macalister, 1868; Murie, 1870a; Struthers, 1871). With greater morphological understanding, new concepts of phylogenetic and taxonomic relationships (e.g. Flower, 1883) resulted in comprehensive works either across multiple fossil and living species (e.g. Gray, 1866; van Beneden and Gervais, 1880) or exhaustive anatomical descriptions of one species (e.g. Struthers, 1888). Morphological insight has thus continued to be an important tool in the understanding of evolutionary, physiological, behavioural and ecological aspects.

I n t r o d u c t i o n
5
Despite the poor record of fossil limb elements, some discoveries have yielded exceptionally well-preserved, and sometimes articulated, skeletons of cetaceans and their ancestors, also including limb elements (e.g. Cooper, 20012; Madar, 2007; Gingerich, 2009). However, while many accounts investigating terrestrial / aquatic locomotion in early cetaceans (and correlated physiological, behavioural and
ecological aspects), mostly focus on skull, vertebral and/or hindlimb characteristics (e.g. Buchholtz, 1998, 2001b; Bebej et al., 2012), the role of the forelimb is usually only discussed in broad terms (e.g. Gingerich, 2003b; Gingerich, 2009; Thewissen et al., 2009; Cooper, 2012). There is only one reconstruction of muscular arrangements in the forelimb of archaeocetes (Dorudon; Uhen, 2004), and no in-depth investigation of functional aspects (combined osteological and myological characteristics) investigating comparative and functional morphologies of extant and/or extinct species as models. Moreover, some interpretations of archaeocete locomotion are highly speculative and without comprehensive anatomical support (e.g. terrestrial locomotion in Ambuloceus, Thewissen 1994). One reason for the lack of morphofunctional investigations of the forelimb in extant and extinct cetaceans (despite an abundance of osteological records), is the rarity of myological accounts in extant cetaceans, i.e. in most extant cetaceans, muscular arrangements are not known, and in those species where accounts do exist, they are usually restricted to a single report, often incomplete or incongruous (see Chapter 5 Literature review: Myology). Morphofunctional analyses of the forelimb are further complicated by the lack of data with regards to flipper shape, a crucial component in understanding flipper functionality (see Chapter 1 Methods and materials: Flipper shapes). Flipper shape is determined by both hard- and soft-tissues. However, soft tissue is often not preserved in
osteological collection or has undergone distortion during the preservation process (see Chapter 1 Methods and Materials: Preservation and storage). Likewise, radiographs are rarely published and distal cartilaginous digital elements or the contour of the flipper are usually only little defined or not visible. Given the sparsity of data in the literature (and of specimens in this study), detailed quantitative analyses of flipper phenotypes are as yet not possible. Hence, only a few papers have published analyses including digital arrangements and flipper shapes in view of taxonomic or phylogenetic context; these papers are restricted to a few basic parameters (e.g. flipper shape: elongated, triangular, round or digital arrangement: broad, narrow), thus poorly reflect

I n t r o d u c t i o n
6
the actual variety of flipper shapes and digital arrangements present in extant cetaceans (e.g. Benke, 1993; Sanchez and Berta, 2010). Chapters 2-4 in this study are therefore divided into two principal sections: (1) a descriptive part (Results) and (2) an investigative part (Discussion):
1) Detailed anatomical records of the shoulder and forelimb are reported in selected cetacean species. These records are the direct result of dissections undertaken in this study and are presented in the section Results of each chapter. Preference was given to rare and/or cryptic species that have until now not been described, also including detailed myological examinations (Chapters 2 Caperea marginata, and Chapter 3 Berardius arnuxii). Additionally, three species (Grampus griseus, Phocoena dioptrica, and Cephalorhynchus hectori) were also selected reporting on the musculature of the shoulder and limb, completed by osteological descriptions in various species (Chapter 4).
2) In the section Discussion, literature is critically reviewed taking into account extinct and extant cetacean species (including historical specimens), as well as non-cetacean species. Concepts are then presented (with references to respective authors), and integrated with the results of this study (e.g. Chapter 2 Caperea marginata and Chapter 3 Berardius arnuxii). Evaluation of published data on forelimb characteristics (comparative and functional anatomy, referenced) and findings from dissections undertaken in this study (not referenced) are then utilised to propose new insights (not referenced) into morphofunctional aspects of the forelimb in cetaceans. For example, re-evaluation of osteological characteristics in early archaeocetes and proposition of a possible mode of terrestrial locomotion (e.g. Chapter 4 Functional Anatomy: Ambulocetus); examination of osteological traits to
postulate for the first time muscle arrangements in extinct species (e.g. Chapter 4 Functional anatomy: Indohyus, Maiacetus) or allow a new understanding of already published findings (e.g. Chapter 4 Functional anatomy: Dorudon). Notwithstanding the lack of data (exacerbated by small sample sizes and possible high intra and interspecific variability), known myological forelimb arrangements of extant cetaceans are here presented within an accepted current taxonomic frame, also discussing the potential value of these findings within a phylogenetic context (e.g. Inia geoffrensis). Similarly, digital arrangements and flipper shapes are examined within an accepted taxonomic and phylogenetic framework (see Fig. 1)

I n t r o d u c t i o n
7
and potential pathways leading to the flipper shapes seen in extant cetaceans discussed. Although flipper shapes and forelimb arrangements are predominantly interpreted in view of functional adaptations (based on aerodynamics, see Chapter 1 Methods and Materials: Flipper shapes), other factors, such as phylogenetic, fabricational or environmental constraints (see Seilacher and Gishlick, 2019) are by
no means excluded as possible factors, but not further investigated here. Rather, the focus of this study lies with the description and (mostly functional) interpretation of the immense variability of hard- and soft-tissue arrangements in the cetacean shoulder and forelimb, and correlate these findings to phylogenetic, physiological, behavioural and ecological aspects (Chapter 2-4), completed by a comprehensive literature review (including myology nomenclature; Chapter 5).

8
Chapter 1: Methods and materials
Procedures
Protocols and sampling methods
This study includes a total of 62 animals from stranding events around the coast of the North and the South Island of New Zealand. Sampling methods and numbers were highly dependent on stranding events and subject to access (locality), approval of local communities, and other factors such as time, weather or tides. As a consequence, the total number of specimens investigated in a species are low in this study (usually < 3 individuals, see List of specimens). In some cases, preliminary dissection at the stranding site or suitable facilities was possible, in other cases this was not possible and only a hasty recovery of the forelimb by gross dissection (with or without shoulder blade, skin and/or blubber) was performed. Additional disassociated flippers (with or without scapulae) were also received for the purpose of destructive dissection ( from staff of Massey University, DOC, Orca Research Trust; see also Acknowledgments and Table 3). At the stranding sites, constraints imposed by time, location or other circumstances did not always permit delicate flensing and examination of the whole cadaver, in which case recovery of the flippers was given precedence while the remaining cadaver was roughly flensed, gutted, and buried (Fig. 1a). Interred skeletal remains were later exhumed and investigated if possible, the removed flippers stored frozen until x-rayed and examined in the laboratory of the Palaeontology Group (Geology Department, University of
Otago, Dunedin). In case of advanced decomposition (beach casts) where a detailed dissection was not feasible, priority was given to the recovery and preservation of the limb bones.

M e t h o d s a n d m a t e r i a l s
9
Fig. 1. Fieldwork (a) field dissection on a Mesoplodon layardii on the beach at Ocean View near Dunedin, Otago, South Island; (b) labelling shoulder muscles on Caperea marginata MM1904 at the Museum of New Zealand in Wellington, North Island. Photo (a) R. E. Fordyce
Dissections also included examination of the shoulder and forelimb in terrestrial mammals (Ovis aries), and the lower forelimb in non-cetacean marine mammals (Phocartos hookeri, Mirounga leonia) for comparative purposes. Additional historical osteological specimens from several New Zealand museums were also examined to complement osteological data of the relatively few specimens referred to in Chapter 2 (Caperea) and Chapter 3 (Berardius); they are listed in Table 3 as historical specimens.
Dissections
Recovery of specimens and field dissections
At the site of the stranding event, compilation of standardised measurements (field data sheet) as set out by the Department of Conservation (DOC) was given priority (mostly dimensions of body and body parts). Additionally, gross examination of the stranded whale and documentation of salient features (e.g. injuries, scars, possible post-mortem damage) were carried out including written and photographic documentation by the Palaeontology Group (Department of Geology, University of
Otago, Dunedin) in cooperation with other parties (e.g. DOC, Otago Museum, Dunedin, and volunteers). Subsequent preliminary dissections (often hastily) were usually carried out on larger specimens. If possible, a more thorough dissection was conducted at least on one side of the animal by removing the skin, blubber and the panniculus carnosus, and uncovering superficial shoulder and limb musculature for

M e t h o d s a n d m a t e r i a l s
10
examination and documentation (particularly peripheral musculature). If possible, shoulder and forelimb were subsequently removed for further dissection in the laboratory of the Palaeontology Group or for storage at a suitable facility (e.g. freezers of the Palaenotology Group, commercial freezer storage or at the Otago Museum). In some cases circumstances permitted the recovery of the whole carcass (usually
smaller cetacean species or immature individuals of larger species) by the Palaeontology Group or other parties (e.g. Otago Museum, Dunedin; Museum of New Zealand, Wellington or Department of Marine Sciences, University of Otago). These specimens were gutted at the stranding site and usually stored frozen at a suitable facility. Subsequent dissections of the whole specimen were then performed at suitably equipped facilities (e.g. at University of Otago, Invermay Research Centre, Museum of New Zealand, DOC) by staff of various institutions (see above) and volunteers. Prior to examination, the specimen was gently defrosted by hosing cold water over the cadaver for 1-3 days. In some cases deep layers of muscle tissue did not defrost completely but time limits dictated the completion of the dissection without delay as best as possible (see also Chapter 4 Functional Anatomy: Myology, section Pectoralis abdominalis). Initial dissections of the shoulder and forelimb focused on the documentation of (1) position and orientation of the scapula and forelimb, (2) overall muscular arrangements, and most importantly on (3) extrinsic limb musculature (e.g. panniculus, mastohumeralis, serratus, rhomboideus, latissimus dorsi, pectorales). Time permitting, particular attention was given to the detailed examination and documentation of extrinsic musculature (photographs, sketches, measurements and labelling). Careful and exacting removal of the skin and blubber was paramount when exposing the delicate panniculus. After examination and documentation, the panniculus was
removed, and extrinsic musculature of the shoulder and forelimb exposed and labelled before the limb was separated for later investigation at the laboratory of the Palaeontology Group. Specimens referred to in Chapter 2 (Caperea) and Chapter 3 (Berardius)
In the case of Caperea marginata two separate stranding events involved a 2.96 m long juvenile female stranded on the Karikari Peninsula (north of Kaitaia) in 2010 (MM2904), and a 1.87 m long neonate male stranded near Glink Gully (south of Dargaville) in 2011 (MM2959) on the North Island, New Zealand (Table 1).

M e t h o d s a n d m a t e r i a l s
11
Preliminary dissection at the stranding site was not possible. Instead, dissections were undertaken at the facilities of the Museum of New Zealand (MoNZ), Wellington in 2010 for the juvenile female whale (MM2904, Fig. 1b) and Invermay Research Centre (AgResearch), Dunedin in 2013 for the neonate male (MM2959). To preserve the cadavers both whales were initially frozen, then defrosted for examination and kept
chilled during dissection. They showed little decomposition with MM2959 at a slightly more advanced stage. MM2959 showed severe bruising and extensive haemorrhaging around the shoulder, particularly in the region just anterior to the forelimb extending far into the deep musculature and to some extent also superficial to the scapula. After the initial examination the scapula and flipper were separated, packed airtight and put into freezer storage. Subsequent detailed dissections of the disassociated flippers were later undertaken in the laboratory of the Palaeontology Group at the Geology Department, University of Otago, in Dunedin. In the case of Berardius arnuxii four flippers were recovered from two stranding events (one whale in Pūrākaunui, Dunedin in 2010 and two whales in the New River estuary, Invercargill in 2013) on the South Island, New Zealand (Table 2). Most of the remains of the male Arnoux's beaked whale Purakaunui (from here on referred to by its stranding location Pūrākaunui) were immediately buried, after a rapid necropsy by University of Otago personnel, at the stranding location. The head and left flipper were initially stored frozen until consent for examination by the local runaka of Kati Huirapa was given. After examination the head and left flipper were re-united with the rest of the skeleton at the burial site. Constraints imposed by time, location and circumstances did not permit for the whole
cadaver of the whale stranded at Ōmaui (South bank of New River estuary) to be completely flensed and buried. The specimen was given a rapid necropsy by University of Otago personnel. The head was separated and buried, and the right flipper retained for examination. Some disarticulated bones of the body were later collected from the site and the skull and mandibles recovered in 2016. The right flipper was stored frozen before and after examination. The carcass of the whale stranded at Sandy Point (North bank of New River estuary) was completely flensed by University of Otago personnel and buried immediately after the stranding event; the right and left flipper were retained for examination. The interred

M e t h o d s a n d m a t e r i a l s
12
skeletal remains at the stranding site were later exhumed in 2016, the flippers stored frozen. Both stranding incidents were important for the local communities, and the two whales from Ōmaui and Sandy Point were subsequently named (see below Cultural
significance and naming of whales). They will therefore from here on be referred to by their given names Hine Reti (Ōmaui) and Tangimoana (Sandy Point), and the third whale Purakaunui by the stranding location Pūrākaunui. From the three animals a total of four flippers were retained for examination. In both events, necessarily rapid flensing of the carcasses precluded delicate separation of the forelimb from the trunk and as a result peripheral musculature was severed without prior examination. Subsequent freezing caused further damage, mainly desiccation in the region of the scapula. A general mild decomposition and desiccation was also noted on the flippers, possibly due to exposure during the stranding event. Further isolated flippers of beaked whales were also obtained from the following specimens (Table 3): Mesoplodon grayi, a 5.02 m long female stranded at Kaka Point; Mesoplodon layardii, a 5.90 m long female stranded at Ocean View Beach (Fig. 1b); Ziphius cavirostris, 5.92 m long male stranded at Katiki Point. Several reference specimens were also investigated for comparison (for a more detailed description of some of these specimens see Chapter 4 Functional anatomy). Detailed dissections
In the laboratory, musculoskeletal arrangements of the shoulder and forelimb were examined in detail to establish species specific patterns and find potential intraspecific variations. This included dissection of the shoulder musculature and soft tissues of the flippers. While the dissection of shoulder musculature was comparable to that of terrestrial mammals, dissection of the flippers proofed to be much more difficult: cetacean flippers are encased in a thin layer of blubber and extremely strong, opaque fibrous tissue. This posed two major problems: (1) hazard to the investigator (potential breakage of scalpel blades, particularly prominent in desiccated specimens), (2) damage of tissue to be investigated (not visible until exposed, some tissues are similar in colour, density and with no clear line of separation). Initial trials followed preparation

M e t h o d s a n d m a t e r i a l s
13
techniques described in the literature (e.g. (Burne, 1952). However, boiling of flippers resulted in the breakdown of soft tissue and disintegration of the skeleton. A series of trials eventually established following protocol: frozen specimens were defrosted using running cold to lukewarm water. The soft tissue of the shoulder was then exposed, dissected, examined and photographed before separating the scapula from the flipper
(Fig. 2a). The intact flipper was treated in a hot water bath (70 ºC) for ¼ to one hour (depending on size and condition of the flipper, e.g. state of desiccation or decomposition). Heat and detergents (a 50/50 mix of washing powder and Vircon, a cleaning agent particularly for hazardous spills) were intended to soften the fibrous tissue encasing the flipper and break down oils and fats. During dissection the flipper was briefly re-immersed into the hot water bath to maintain warmth (Fig. 2c).
Fig. 2. Lab work (a) dissecting shoulder muscles of Berardius arnuxii Purakaunui; (b) terrestrial meets ET, more precisely Cephalorhynchus hectori H164; (c) warming flipper of Berardius arnuxii Purakaunui in hot tub prior to dissection. Photo (a) Y. Tanaka, (b) R. M. Steffen
General identification of muscles was based on Strickler, 1978 and Smith, 1976 with additional reference to the myology in baleen whales as described by Carte and Macalister, 1868; Perrin, 1870; Schulte, 1916 and Benke, 1993. Measuring techniques

M e t h o d s a n d m a t e r i a l s
14
followed the methods described by Benke, 1993 and terminology referred to veterinary nomenclature employed by Gasse, 2012. Phalangeal count was based on Padian, 1992. For a more detailed description see section Terminology.
Photography
Radiographs
Prior to x-raying, specimens were thoroughly washed and cleaned (sand is strongly visible in radiographs). Trials revealed that frozen specimens showed slightly more contrast than defrosted specimens, consequently specimens were transported in insulated containers when feasible. Radiographs were then taken at the Dunedin Hospital using a computed radiography Philips Optimus Bucky Diagnostic system with a non-gridded cassette: exposure was 66kV, 12.5 mAs at a source-image distance of 100 to 200 cm. The cassette was a Philips standard, either a long view or a 35 x 43 cassette with standard screen. OsiriX 3.7.1 was used to view the digital radiographs (DICOM files).

M e t h o d s a n d m a t e r i a l s
15
Photographs
Both in the field and in the lab, muscles or muscle groups, were photographed in pairs (with and without scale bar, labels, and/or white string delineating the border of the muscles), and at varying distances (overview and close up) and angles, Recycled, cut up milk containers and waterproof felt pens proofed to be the most resilient and easy to use labelling method, able to cope with bodily fluids while also easily cleanable with alcohol. These labels were also permanently anchored to extrinsic muscles permitting unambiguous identification of these muscles after the detachment of the limb. Photographs were taken either in the field or in the laboratory with a digital Nikon D700 camera. The lenses used were an AF-S DX Nikkor 18-105 mm f/3.5-5.6, AF-S DX Nikkor 35 mm f/1.8 G, AF-S Nikkor 50mm f/1.8 G, and an AF-S micro Nikkor 60 mm f/2.8.
Fig. 3. Camera setup (a) camera stand and elevated specimens on a glass plate; (b) specimen (Cephalorhynchus hectori H197) photographed on the light table; (c) lateral view of the right flipper of Berardius arnuxii Purakaunui with a light source on the other side.
Whenever possible, the camera was mounted on a vertical stand to allow stabilized longer exposure times (around 2 sec) to maximise focal depth (Fig. 3a). The stand also

M e t h o d s a n d m a t e r i a l s
16
provided increased focal distance, minimising peripheral lens distortion. Aside from additional light sources and deflectors, the specimens were either elevated on a glass plate (Fig. 3a), or a light table was used to minimise shadows (Fig, 3b). The light table, or a light source behind the specimen, also allowed x-ray-like imaging of tissue (Fig. 3c). Figures in the following chapters, if not otherwise stated, were taken from
anatomical dissections using the methods described above.
Preservation and storage
When storing specimens frozen, it is imperative to seal them airtight in order to prevent desiccation (mummification). Mummification causes hardening of the tissue and can render specimens hazardous and unsuitable for dissection; in some cases manual preparation was not possible (see above Detailed dissections). Smaller specimens were stored in zip-lock freezer bags, larger items wrapped in glad wrap or in large plastic bags sealed with tape.
Fig. 4. Mounting of specimens (a) temporary storage in clear water (Cephalorhynchus hectori H166); (b) ‘stitching’ of the right flipper of Berardius arnuxii Purakaunui; (c) ‘stiched’ flipper on metal grid (H199 Cephalorhynchus hectori), note foam supports and increased number of loops at the finger tips. Photo (c) Y. Tanaka (author at left helped by volunteer C. C. Fan at right)

M e t h o d s a n d m a t e r i a l s
17
After dissection, bones were manually cleaned (not cooked as is usually recommended) and mounted onto a metal grid. It is also possible to store cleaned specimens temporarily in clean water (up to two days) until further processing (Fig. 4a). Particularly oily specimens were washed in alcohol prior to several final washes in hot soapy water and subsequent rinses in hot clear water.
Fully articulated, undistorted specimens are rare in museums because cartilage shrinks and distorts during the desiccation process. Formaldehyde was not taken into consideration because of the hazardous nature of this substance, There are no descriptions in the literature detailing the desiccation and preparation process. A series of trials resulted in the following protocol for delphinid specimens: the specimen was secured onto a framed wire-grid by looping a few strands of string around the specimen and through the holes of the grid (from here on referred to as ‘stitching’). This was followed by more thorough fastening of each digit separately, with string partly unravelled to broaden the strand (Fig. 4b). Great care was hereby taken to tie digits in their natural position, the string being tight enough to prevent warping but also allowing at the same time shrinkage. Loops of string securing the digits increased in number towards the tip of the digit, as here the cartilage is most prone to warping and shrinkage. Elevated parts of the hand and the tips of the fingers were also supported by foam wedges and firmly secured by string to prevent vertical warping (Fig. 4c). The drying process took place in a well-aerated room over a time-span of 1-3 weeks before the skeleton could be removed. Superficial drying ensued relatively fast and was noticeable within a day (important to prevent mould). However, trials showed that desiccation could not be forced (e.g. fumehood) as cartilage appears to be more prone to cracking when drying occurs too rapidly. Treatment with disinfectant also affected the integrity of the cartilage and is not recommended.
Furthermore, in basal odontocetes and all mysticetes forelimb articulation (distal of the shoulder joint) is not as firmly fused as in delphinids, consequently forelimb elements had a tendency to separate. Preservation of fully or partly articulated specimens is for that reason rarely possible. Instead, conservation of the bony elements was given priority. Cardboard boxes were used for the final storage of specimens. These were laid out with PVC sheet foam to be used as an inert cushion. The same material was also used to ‘stich up’ disarticulated specimens to preserve the original digit position of the hand bones (Fig. 5).

M e t h o d s a n d m a t e r i a l s
18
Fig. 5. Final storage (a) Kogia breviceps OMNZ X2020.75; (b) Globicephala macrorhynchus 10-23Lo. Note string holding finger bones in natural position.
Terminology
Key words and references
Key words within sections are printed in bold letters in order to assist finding sections referred to in other parts of this study, or to emphasise contextual contents. Sections in this study are referenced in italic, inclusive chapter titles, subtitles and/or key words or figures. Authors are referenced either at the end of a sentence or paragraph, or immediately in front of a following list. Where accounts differ greatly, references are inserted immediately behind a statement, either mid-sentence or at the end of a phrase. Lack of references either indicates a summary following well-referenced text, or accounts and personal opinions by the author of this study. Personal opinions and findings are made as obvious as possible by either the use of modal verbs or referring directly to this study. Taxonomic nomenclature, if not otherwise stated, is based on lists published by the Committee of Taxonomy of The Society for Marine Mammology, accessible online at: <https://marinemammalscience.org/species-information/list-marine-mammal-species-subspecies/ >.

M e t h o d s a n d m a t e r i a l s
19
Orientation
Positions and planes in relation to the body
The following definitions are applied to positional terminology relating to the body as a whole, inclusive shoulder and scapula (Fig. 6a). Three perpendicular planes encompass: (1) sagittal plane, division between left and
right side, a parasagittal plane lies parallel to the sagittal plane, (2) dorsal plane, division between up- and underside, (3) transverse plane, division between front and back. A position towards the head is defined as cranial (or anterior), and towards the tail as caudal (or posterior). A position towards the back is referred to as dorsal, and towards the belly as ventral. A position towards the sagittal plane is referred to as medial (or deep), and away from the sagittal plane as lateral (or superficial). The suffix -ad denotes a direction rather than a position, e.g. the scapular spine emerges near the vertebral border and rises ventrad as a small ridge before turning craniad. Alternative terms such as anterior-posterior or deep-superficial are sparingly used either to avoid repetition or as localised descriptions that are intuitively easier to understand. E.g. the subscapularis muscle occupies the medial surface of the scapula; the muscle is on its deep surface (i.e. the lateral surface adhering to the medial surface of the scapula) divided by several strong tendons and superficially (i.e. medial surface facing the ribs) covered by a strong tendinous tissue. Another example are the pectorales major and minor arising from a medial position (sternum) and inserting laterally onto the base of the flipper and coracoid respectively. Although the origin of both muscles is ventral, the origin of the pectoralis major is superficial to that of the pectoralis minor.

M e t h o d s a n d m a t e r i a l s
20
Fig. 6a. Three-dimensional orientation and planes of the body Schematic illustration of orientation of vectors and planes in relation to the body including shoulder and scapula but excluding the forelimb.
Flipper positions and movements
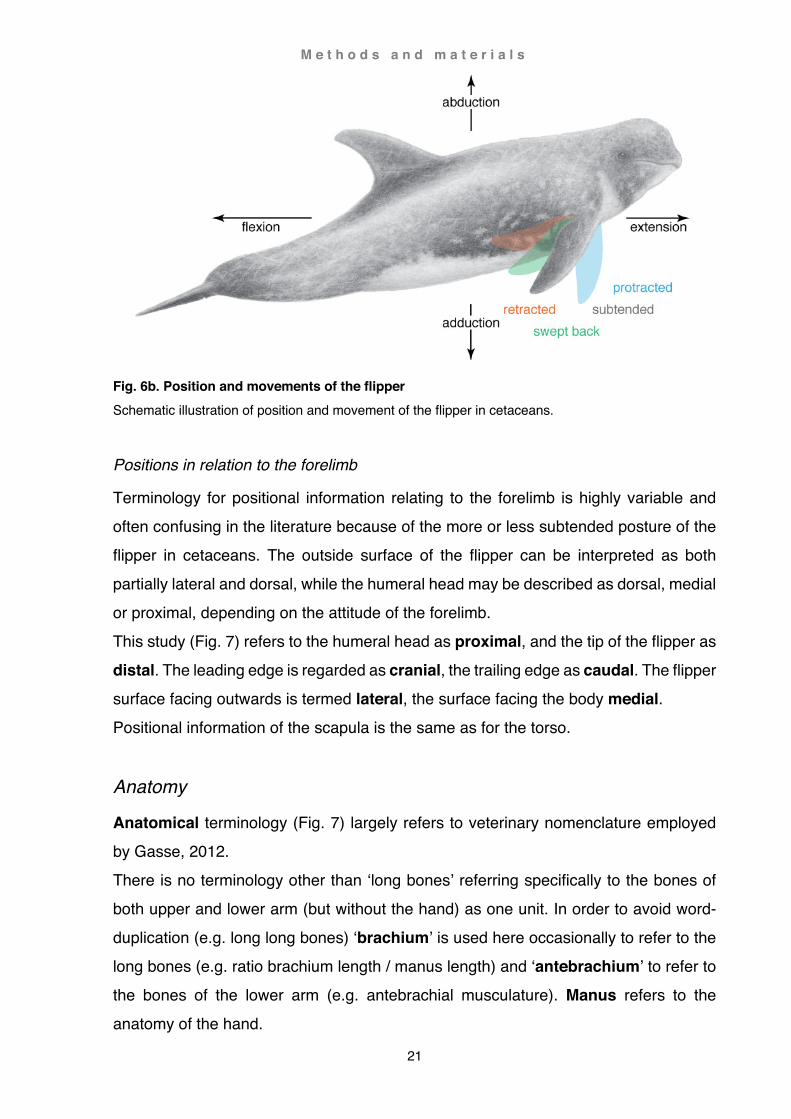
Muscle actions of the limb encompass two axes defined by four vectors (Fig. 6b): (1) flexion, closing of a joint angle in the same plane, (2) extension, opening of a joint angle in the same plane, (3) adduction, closing of a joint angle roughly perpendicular to the plane encompassed by corresponding articulating bones, (4) abduction, opening of a joint angle roughly perpendicular to the plane encompassed by corresponding articulating bones. Protraction of the limb involves in terrestrial mammals opening of all joint angles (shoulder, elbow, wrist, manus), while retraction involves closing of all joint angles. A protracted limb in cetaceans is fully extended at the shoulder joint and can be either ab- or adducted. The natural limit of extension usually does not reach much further craniad than perpendicular to the shoulder joint. A fully retracted limb is positioned close and parallel to the midline of the body. A subtended limb position is a naturally more or less relaxed flipper attitude seen at low speeds. With increased speed, flippers are held more swept back, i.e. further back
and often higher than subtended (Benke, 1993).
M e t h o d s a n d m a t e r i a l s
21
Fig. 6b. Position and movements of the flipper Schematic illustration of position and movement of the flipper in cetaceans.
Positions in relation to the forelimb
Terminology for positional information relating to the forelimb is highly variable and often confusing in the literature because of the more or less subtended posture of the flipper in cetaceans. The outside surface of the flipper can be interpreted as both partially lateral and dorsal, while the humeral head may be described as dorsal, medial or proximal, depending on the attitude of the forelimb. This study (Fig. 7) refers to the humeral head as proximal, and the tip of the flipper as distal. The leading edge is regarded as cranial, the trailing edge as caudal. The flipper surface facing outwards is termed lateral, the surface facing the body medial. Positional information of the scapula is the same as for the torso.
Anatomy
Anatomical terminology (Fig. 7) largely refers to veterinary nomenclature employed by Gasse, 2012. There is no terminology other than ‘long bones’ referring specifically to the bones of both upper and lower arm (but without the hand) as one unit. In order to avoid word-duplication (e.g. long long bones) ‘brachium’ is used here occasionally to refer to the long bones (e.g. ratio brachium length / manus length) and ‘antebrachium’ to refer to the bones of the lower arm (e.g. antebrachial musculature). Manus refers to the anatomy of the hand.

M e t h o d s a n d m a t e r i a l s
22
In the literature, proximal tubercles of the humerus are frequently confused. In this study, the smaller, more cranial tubercle is referred to as greater (aka lateral) tubercle, and the larger, more caudal tubercle as lesser (aka medial) tubercle. The term tubercle is only applied to these two prominent landmarks; all other tuberosities, e.g. the deltoid tuberosity on the shaft of the humerus, are referred to as tuberosities (for
a more detailed description of the tubercles see Chapter 4 Functional anatomy: Functional aspects of flipper shapes, section Inia geoffrensis). Myological terminology is predominantly based on Strickler, 1978 and on Cooper et al., 2007b for antebrachial musculature (for more detailed descriptions of muscles and their synonyms see Chapter 5 Literature review: Myology). The term ‘mastohumeralis’ has been maintained here although the mastoid process (e.g. in Homo) has been homologised in cetaceans by Mead and Fordyce (2009) with the posterior process of the periotic (petrosal bone) which lies isolated within the skull. This process is in cetaceans not visible from outside and therefore not a possible attachment point for this muscle. Furthermore, ‘mastoid’ has also been applied to components of the squamosal and tympanic bulla confusing matters even further (Mead and Fordyce, 2009). Other terms such as ‘cephalohumeralis’ have been synonymised with the trapezius (e.g. Schulte, 1916) and are along with terminology such as ‘brachiocephalicus’, ‘cephalohumeralis’, ‘occipitohumeralis’ either too unspecific or inaccurate. This and the consideration that the term ‘trapezius’ (and other terms such as ‘occipitoscapularis’, ‘cephalic trapezius’, ‘levator anguli scapulae’) has in turn been confused with the rhomboideus capitis (e.g. ‘occipitoscapularis’ in Howell, 1927, but this muscle is the levator scapulae in Pilleri, 1976) would also raise the question of how to re-name the mastoscapularis accordingly.
Overall, ‘mastohumeralis’ is the term used in most of the literature cited in this study. It is also indicative of an area on the skull functionally analogous with the mastoid in other mammals, and consequently excludes other attachment points on or near the skull associated with either the rhomboideus capitis or trapezius.

M e t h o d s a n d m a t e r i a l s
23
Overleaf: Fig. 7. Anatomical terminology Semi-schematic view of the forelimb of Berardius arnuxii, (a) lateral view of the right limb and (b) medial view of the left limb. Bone is represented as lightly shaded areas and cartilage as darker shaded (scapula and long bones) and white (manus) areas; flipper contour is shown in slate grey.

M e t h o d s a n d m a t e r i a l s
24
Similarly, the teres minor is not related to the teres major but a derivate of the deltoideus (Strickler, 1978; Howell, 1937). This muscle appears to have a dual occurrence: (1) as ‘teres minor’ arising near the caudal scapular border (Klima, 1980; Weijuan, 1981; Smith et al., 1976), or as (2) ‘subdeltoideus’ arising near the base of the acromion or coracoid (Schulte, 1916; Schulte and Smith, 1918; Benke, 1993).
Since the two names have equal merits, and are the only labels used in the literature to refer to this muscle, both terms have been retained here (see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscular arrangements, and Chapter 5 Literature review: Myology).
Evaluations
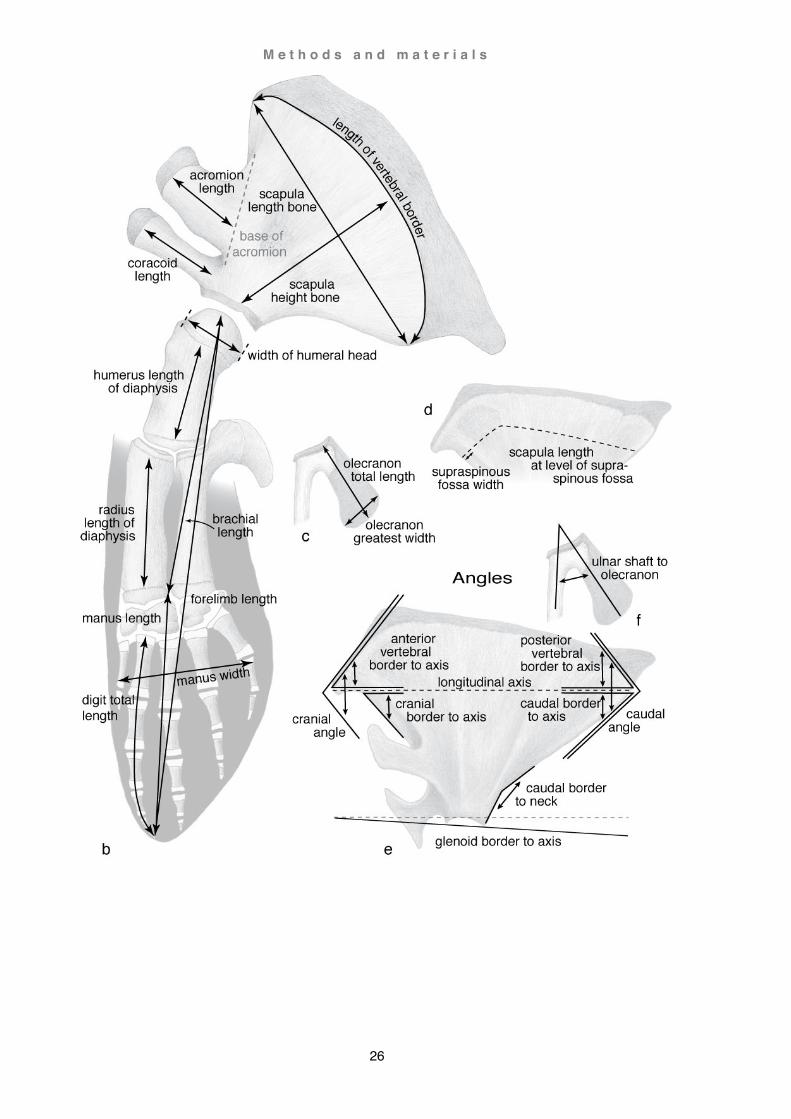
Measuring methods
Measuring techniques followed largely the methods described by Benke, 1993 and Galatius and Kinze, 2003. All measurement parameters used for Caperea marginata and Berardius arnuxii are depicted in Fig. 8. Note, there is a clear distinction between measurements of the body / flipper (soft tissue) and the skeleton / forelimb (hard tissue) which is referred to correspondingly. Measurements of additional specimens were not as detailed. Results are listed in the appendix in the section Measurements. Osteological and myological comparative analyses of the shoulder blade and forelimb are expressed as a ratio in form of fractional or decimal values in a manner comparable to the literature cited in this study (e.g. Beddard, 1901; Benke, 1993; Sanchez and Berta, 2010).

M e t h o d s a n d m a t e r i a l s
25
M e t h o d s a n d m a t e r i a l s
26

M e t h o d s a n d m a t e r i a l s
27
This page and overleaf: Fig. 8. Definition of measuring parameters Semi-schematic view of the left forelimb of Berardius arnuxii, basic measurements in (a) lateral view and (b) medial view; (c) measurements of the olecranon (e.g. Caperea marginata); (d) measurement of the supraspinous fossa in case not reaching the vertebral border of the scapula (e.g. Cephalorhynchus hectori); (e and f) measurements of angles in (e) scapula (e.g. Cephalorhynchus hectori) and (f) olecranon (e.g. Caperea marginata). Bone is represented as lightly shaded areas and cartilage as darker shaded areas; flipper contour is shown in slate grey.

M e t h o d s a n d m a t e r i a l s
28
Flipper shapes
Evaluation of flipper shape includes multiple components (hard- and soft-tissue) composed of multivariate elements (e.g. length, width, depth, or angle of digital elements). Yet, poor preservation of some components (e.g. cartilaginous distal finger elements) also translates in poor representation of reliable data in fossil or osteological collections. Benke (1993) undertook extensive osteological shoulder and forelimb element analyses, and incorporated his findings into a then accepted (now revised) taxonomic framework. However, he does not remark on potential sampling errors caused by the poor preservation of at least some specimens he analysed (e.g. photo fig. 51 posiible distortion and shrinkage of digits, and potentially missing distal digital elements). It is therefore possible that sampling errors could have obscured other characteristics that have significant taxonomic components. Sanchez and Berta (2010) published a comparative analysis of major shoulder and forelimb bones and muscles in odontocetes. However, they did not include mysticetes and omitted forelimb parameters fundamentally influential in the shape of a flipper (e.g. differentiation of
arrangements in preaxial and postaxial digits) resulting in a restricted number of flipper shape categories (circular, triangular, elongated). For these reasons, flipper shape analysis encompasses in this study both quantitative (e.g. ratio of flipper length / body length) and qualitative (see below) parameters. Qualitative parameters have been established in this study from standardised radiographs (see section Photography in this chapter). Parameters from species not listed in the section Materials were sourced wherever possible from either published literature (photos, sketches, illustrations, measurements) or non-peer revied media if no other source was available (mainly photos). However, sampling errors are presumed to be great (small sample size in combination with sampling errors e.g. lack of scale bars, lens distortions, angle of the camera or blurring in photos, and in sketches by possible imprecise contour replication or unknown physical condition of a specimen). The definition of flipper shapes is therefore based on aerodynamic properties (see e.g. Fish and Battle, 1995; Woodward, 2006; McLean, 2012). Performance in a hydrofoil or aerofoil is determined in a planform (flipper silhouette, cross-section excluded) by predominantly 6 attributes that were considered when categorising flipper shapes: aspect ratio, root chord length, shape of leading and trailing edge, and tip of

M e t h o d s a n d m a t e r i a l s
29
the flipper (see also McLean, 2012). Some flipper shapes were more or less congruent with established wing shapes in aeroplanes (e.g. elliptical wing shape in a spitfire and oval flipper in Caperea marginata). In other cases, flipper shapes did not match established wing shapes (e.g. falciform flipper in Globicephala sp.); flipper performances were then evaluated based on the performance properties of the most
similar wing shapes or isolated wing parts described in McLean, 2012. Evaluation of flipper shapes resulted in 6 categories (oar, deltoid, paddle, fusiform, oval and falciform shape) including a total of 13 subcategories (see also Chapter 4 Functional anatomy: Pectoral limb and Functional aspect of flipper shape section Diversity of flipper shapes). However, in some species (specimens), flipper shapes incorporated elements of more than one type (e.g. oval and oar shape in Caperea marginata) and a clear distinction is therefore not always possible. Given the scarcity of records and ostensible intraspecific variability of flipper elements, shape categories specified in this study must be viewed as a tentative proposition to capture the multitude of cetacean flipper shapes as best as possible and are by no means irrefutable.
Age classification
Age determination was based on a combination of body size and epiphyseal and/or carpal development stage (radiographs); age verification using dental GLG (growth layer groups) was not possible. In the absence of accurate data closely related species were compared to gain a rough estimate. Whenever possible, following age categories were used in this study (modified from e.g. Kemp, 1999; Galatius and Kinze, 2003; Galatius et al., 2006): (1) Dependent calves including: neonate (new born), infant (intermediate between neonate and weaner), weaner (calve at the time of weaning) with no epiphyseal fusions in the flipper, no sexual maturity (2) Subadults including juvenile (young subadult, possibly no sexual maturity) and older subadult (possible sexual maturity) with no or very little epiphyseal fusions in the flipper (3) Adults including young adults (possible sexual maturity) and older adults (near certain sexual maturity) with partial and/or fully epiphyseal fusion in the flipper Age estimates in Caperea marginata were based on published data by Kemper (1999) taking into account body length, length of scapula, and epiphyseal fusion of the flipper.

M e t h o d s a n d m a t e r i a l s
30
Total body length in historical osteological specimens (e.g. Otago Museum, Museum of New Zealand, see also Chapter 2 Caperea: Scapula and forelimb bones, Fig. 18, 19b) was assessed on scapular length. Based on these parameters, following age categories were established: neonate (< 1m); juveniles including infant (2.0-3.0 m) and weaner (3.0-3.5 m); subadult (3.5-5.5 m); adult (> 5.5 m); mature (> 6.0). For example,
at a length of 47.4 cm the isolated right scapula of OMNZ VT227 suggested a total length of about 510 cm, confirming the information on the label as a subadult (male). Age estimates in Berardius arnuxii were assessed by a combination of epiphyseal fusion and body length, as well as a comparison to Berardius bairdii. Baird’s beaked whales are at a maximal body length of 12.60 m larger than B. arnuxii at 9.90 m. Sexual differences are not as pronounced in the southern species as they are in the northern species where the female is significantly larger. Neonates of B. arnuxii are estimated to be about 3.0 m in length (McCann, 1975). In Purakaunui and Tangimoana, the epiphyses and carpals were fully developed. The epiphyses were attached to the diaphysis but not completely fused (sensu Galatius and Kinze, 2003). Although in both specimens the developmental stage of the ulnar and radial epiphyses were similar, those of Purakaunui were slightly more advanced. In Hine Reti neither the epiphyses nor the carpals were fully developed or fused. These characteristics, together with total body length, were used to identify Purakaunui as a young adult, Tangimoana as an older subadult and Hine Reti as a juvenile.
Cultural significance and naming of the stranded whales Local communities at the stranding sites were often strongly involved during stranding incidents involving whales (see also Acknowledgements), from failed rescue attempts to subsequent burial and recovery of the remains. This was particularly the case in the stranding event of two Berardius arnuxii beaked whales near Invercargill on the southern coast of the South Island (runaka of Waipio at Sandy Point and Awarua at Ōmaui). During recovery, the remains the two whales were named to acknowledge the close relationship of the whales to the community, the locality and the stranding event. The younger whale who stranded first was named 'Hine Rēti' in reference to her gender (Hine, a female), and stranding location (Oreti near Ōmaui), a site notorious for unpredictable wind and water currents that act like a trap or snare (Rēti). The older

M e t h o d s a n d m a t e r i a l s
31
whale from Sandy Point was named 'Tangimoana', as she was believed to have followed in from the open sea (moana), calling out and crying (tangi) to the distressed younger female. For these reasons, these two whales will be referred to by their given names ‘Hine Rēti’ and ‘Tangimoana’.
The skeletal remains of both whales are currently undergoing preparation (except the flippers, see above Preservation and storage). This process has continued to be a collaborative community initiative, led by Ramari Oliphant Stewart with local volunteers from Waihopai and Awarua runaka, and the support from Southland Museum, University of Otago and Department of Conservation. The skeleton of Tangimoana is intended to be gifted into the collections at the Southland Museum, Invercargill. The final location of the skull, mandibles and right flipper of Hine Reti has yet to be determined.
List of specimens Abbreviations of Institutions UO: Palaeontology Group, Geology Department, University of Otago, Dunedin, NZ OM: Otago Museum, Dunedin, NZ MoNZ: Museum of New Zealand (Te Papa), Wellington, NZ
Table 1. Caperea marginata specimen identification and stranding location Ref. in text MoNZ ID (1)
Age group (2)
Sex Length (3)
Stranding date
Stranding area Coordinates (4)
MM2904
Juvenile Female 296 cm 05 Sep 2010 Karikari Peninsula E coast Northland North Island, NZ
034º 51’ 06” S 173º 21’ 21” E
MM2959
Neonate Male 187 cm 28 Jun 2011 Glink’s Gully
W coast Northland North Island, NZ
036º 08’ 10” S 173º 54’ 05” E
(1) MoNZ: Museum of New Zealand; (2) sensu Baker, 1985; (3) Total length from tip of rostrum to fluke notch; (4) MM2904
peninsula coordinates estimated using NZ Topo Map, Reference System WGS84, MM2959 coordinates provided by MoNZ
The forelimb skeletons of MM2904 and MM2959 have been returned to the Museum of New Zealand

M e t h o d s a n d m a t e r i a l s
32
Table 2. Berardius arnuxii specimen identification and stranding location Provisional
ID Age
group (1) Sex Length
(2) Stranding
date Stranding area (3) Coordinates
(4)
Purakaunui (Pur)
Adult Male 870 cm 22 Dec 2010 Pūrākaunui Inlet Dunedin Otago South Island
045º 44.97’ S 170º 37.52’ N
Tangimoana (Tam)
Subadult Female ~ 700 cm 21 Jan 2013 Sandy Point New River Estuary Invercargill Southland South Island
046º 49’ 47” S 168º 17 ‘ 41” E
Hine Reti
(HiRe) Juvenile Female ~ 650 cm 19 Jan 2013 Beach at Ōmaui
New River Estuary Invercargill Southland South Island
046º 30’ 44” S 168º 16’ 47” E
(1) sensu Kasuya, 2009; (2) Total body length from tip of rostrum to fluke notch; (3) Location names listed in Land Information
New Zealand (LINZ); (4) Coordinates estimated using NZ Topo Map, Reference System WGS84
The forelimb skeleton of Purakanui has been interred with the rest of the skeleton in 2011; Tangimoana and Hine Reti were returned to R. Stewart and runaka of Waipio and Awarua.
Table 3. Additional specimens Sequence of specimen description: stranding incident or repository ID number (if given); investigated body part (flipper(s)(1) or body part); age group and sex; body length; stranding date; stranding location(2); comments (1) Flippers abbreviated to L left flipper and R right flipper
(2) Locations in accordance with Land Information New Zealand (LINZ) place names listed in the New Zealand Gazetteer If not otherwise stated following specimens are currently held by the Palaeontology Group in the Geology Department, University of Otago, Dunedin (UO)
Mysticeti Balaenoptera acutorostrata or B. bonaerensis
ID number not issued; L/R; immature female; 558 cm; 09 Mar 1993; Short Bay, Campbell Point, Otago, South Island; described and depicted by Watson and Fordyce, 1993 (fig. 1d).

M e t h o d s a n d m a t e r i a l s
33
Odontoceti Cephalorhynchus hectori
H164 (stranding incident); L/R; adult female; 144 cm; 07 Sep. 2008; Flowery Creek, West Coast, South Island
H166 (stranding incident); subadult male; 118 cm; 02 Oct 2008; 1 km south of Rapahoe (village), West Coast, South Island
H167 (stranding incident); adult female; 145 cm; 03 Oct 2008; beach near Ross (village), West Coast; South Island
H176 (stranding incident); L/R; adult female; 148 cm; 21 Jan 2009; Katiki Point, Otago, South Island H177 (stranding incident); L/R; neonate male; 76 cm; 04 Feb 2009; Punakaiki (reserve), West Coast,
South Island H180 (stranding incident); L/R; adult female; 158 cm; 17 Mar 2009; Ashworths Ponds, Canterbury,
South Island H182 (stranding incident); L/R; subadult male; 120 cm; 08 May 2009; Conway River, Canterbury,
South Island H197 (stranding incident); L/R; juvenile male; 115 cm; 16 Aug 2010; Kekerengu (village), Marlborough,
South Island H199 (stranding incident); L/R; adult female; 138 cm; 27 Sep 2010; Ruatapu (village), West Coast,
South Island H202 (stranding incident); L/R; subadult male; 125 cm; 20 Nov 2010; Raglan (town), Waikato, North
Island H255 (stranding incident); R; subadult male; 121; 07 Apr 2015; Petit Carenage Bay, Canterbury, South
Island Additional specimens not directly referred to in this study: H169, H171, H181, H183, H186, H193,
H194, H195, H196, H198, H199 (neonate), H200, H204, H205, H207 Grampus griseus
VT3301 (Otago Museum); L/R; adult male; 298 cm; 23 Jun 2013; Raukawa Stream, Tasman, South Island, specimen held by the Otago Museum
Kogia sima VT3311 (Otago Museum); L/R; immature male; 163 cm; 26 Mar 2010; Papanui Inlet, Dunedin, Otago,
South Island, specimen held by the Otago Museum Kogia breviceps
X2020.75 (Otago Museum); L/R; subadult female; 283 cm; 28 Nov 2010; Moeraki Beach, Otago, South Island, specimen held by the Otago Museum
Lagenorhynchus obscurus VT3217 (Otago Museum); L/R; juvenile female; 142 cm; 18 May 2011; Owaka (village), Otago, South
Island, specimen held by the Otago Museum Lissodelphis peronii
VT3010 (Otago Museum); L/R; adult female; 208 cm; 25 Jan 2010; Kaka Point, Otago, South Island, specimen held by the Otago Museum

M e t h o d s a n d m a t e r i a l s
34
Mesoplodon grayi ID number not issued; L/R; adult female; 502 cm; 22 Aug 2010; Kaka Point, Otago, South Island
Mesoplodon layardii ID number not issued; L/R; subadult female; 590 cm; 16 Aug 2008; beach at Ocean View (village),
Brighton, Otago, South Island Phocoena dioptrica
X2020.76 (Otago Museum); L/R; adult male; 215 cm; 17 Sep 2014; Pipikaretu Beach, Dunedin peninsula, Otago, South Island, specimen held by the Otago Museum
X2020.77 (Otago Museum); L/R; adult male; 200 cm; 11 Jul 2014; Caroline Bay, Timaru, Canterbury, South Island, specimen held by the Otago Museum
VT3347 (Otago Museum); L/R; juvenile female; 125 cm; 10 Aug 2015; Bailey’s Beach, Kaitorete Spit, Canterbury, South Island, specimen held by the Otago Museum
Ziphius cavirostris ID number not issued; L/R; adult male; 592 cm; 04 Mar 2013; Katiki Beach, Otago, South Island ID number not issued; L/R; age and sex undetermined; 590 cm; 15 Sep 2010; Long Point (Irihuka),
Otago, South Island
Data deficient specimens received for destructive dissection Balaenoptera edeni: 1 specimen (L/R) Globicephala melas : 5 specimens (09-01Gm L/R; 10-36Gm L/R; 10-40a L/R; 10-51Gm; L/R; 10-55Gm)
and an additional specimen stranded in the Otago Harbour in 2012 (L/R) Globicephala ?macrorhynchus: 2 specimens (10-22Lo L/R; 10-23Lo L/R) Orcinus orca: 2 specimens (Koropepe L/R; Maripi L) Historical specimens Berardius arnuxii
VT220 (Otago Museum); jaw; first registered at Otago Museum around 1840 as beach cast at Taiaroa Head, Otago, South Island
Caperea marginata MM0036 (MoNZ); scapula; subadult female, 465 cm; 1874; skeleton from Stewart Island, South Island,
referred to as the ‘Traill Specimen’ by Hector (1875) and Beddard (1901 MM2049 (MoNZ); scapula; Dec. 1990, Warkwoth, Auckland, North Island MM2232 (MoNZ); scapula; date not known; Ngakengo Stream, Northland, North Island MM2254 (MoNZ); scapula; 18 Nov 1994; Farewell Spit, Tasman, South Island MM2900 (MoNZ); scapula; 20 Dec 1991; Paterson Inlet, Stewart Island, South Island M 1593 (referred to by Hales 1931 and 1961); scapula; animal about 490 cm; Brownlow, Kangaroo
Island, Australia; repository South Australian Museum VT227 (Otago Museum); scapula; subadult male, 10 Aug 1976, Paterson Inlet, Stewart Island, South
Island

35
Chapter 2: Pygmy right whale
Caperea marginata (Gray, 1846)
Introduction Cetaceans represent a morphologically highly disparate lineage derived from land mammals that returned to an aquatic lifestyle about 50 million years ago (Thewissen, 1998). The transition from land to water, i.e. the development from weight-bearing forelimb to flipper, is well documented by fossils (Thewissen et al., 2009), but many morphological characteristics that cannot be detected in fossils have remained little documented or speculative. Freed from the constraints of gravity and the developmental links to the hind legs, there is a persistent anatomical plasticity in the flipper of modern cetaceans, showing substantial skeletal disparities sometimes even
within the same population or pod (Cooper et al., 2007a, see also Chapter 2 Caperea and Chapter 3 Berardius). There is only a small number of morphological studies investigating forelimb musculature in extant cetaceans and the understanding of functional morphology in this group has so far remained largely unresolved. Among mysticetes the lack of data is particularly apparent. The few accounts that include soft tissue are limited to rorquals, Balaenoptera acutorostrata (Carte and Macalister, 1868; Perrin, 1870), Balaenoptera borealis (Schulte, 1916; Benke, 1993), Megaptera novaeangliae and Balaenoptera musculus (Struthers, 1888, 1889; Kükenthal, 1921), and right whales, Balaena mysticetus (Eschricht et al., 1866; Struthers, 1878; Kükenthal, 1922) - with the exception of Benke these papers are all nearly or over a century old. There are no reports on bowhead whales, gray whales or the pygmy right whale. History and phylogeny of the genus Caperea
The pygmy right whale, Caperea marginata, is the least known and most elusive of all baleen whales. Isolated remains, infrequent solitary strandings, and few sightings have so far remained the sole source of information (Kemper et al., 1997; Kemper, 2002), thus sharply contrasting with all other baleen whales that are known from a multitude
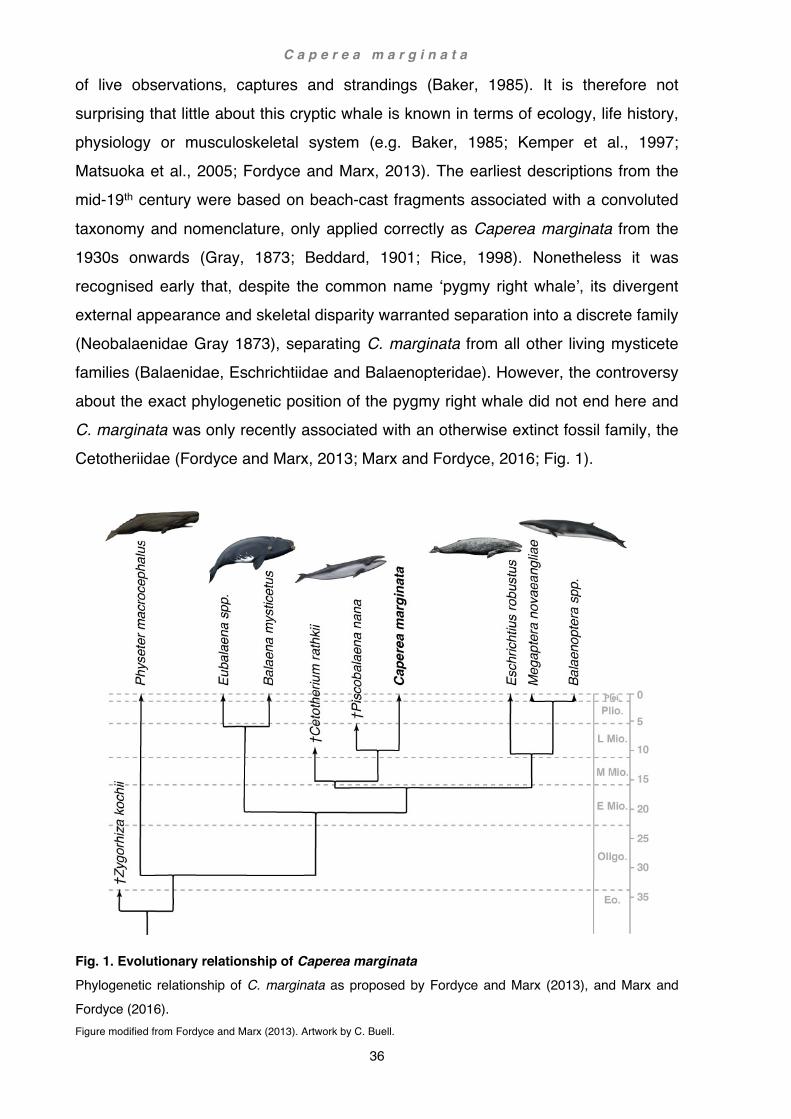
C a p e r e a m a r g i n a t a
36
of live observations, captures and strandings (Baker, 1985). It is therefore not surprising that little about this cryptic whale is known in terms of ecology, life history, physiology or musculoskeletal system (e.g. Baker, 1985; Kemper et al., 1997; Matsuoka et al., 2005; Fordyce and Marx, 2013). The earliest descriptions from the mid-19th century were based on beach-cast fragments associated with a convoluted
taxonomy and nomenclature, only applied correctly as Caperea marginata from the 1930s onwards (Gray, 1873; Beddard, 1901; Rice, 1998). Nonetheless it was recognised early that, despite the common name ‘pygmy right whale’, its divergent external appearance and skeletal disparity warranted separation into a discrete family (Neobalaenidae Gray 1873), separating C. marginata from all other living mysticete families (Balaenidae, Eschrichtiidae and Balaenopteridae). However, the controversy about the exact phylogenetic position of the pygmy right whale did not end here and C. marginata was only recently associated with an otherwise extinct fossil family, the Cetotheriidae (Fordyce and Marx, 2013; Marx and Fordyce, 2016; Fig. 1).
Fig. 1. Evolutionary relationship of Caperea marginata Phylogenetic relationship of C. marginata as proposed by Fordyce and Marx (2013), and Marx and Fordyce (2016). Figure modified from Fordyce and Marx (2013). Artwork by C. Buell.

C a p e r e a m a r g i n a t a
37
Anatomical accounts of the shoulder and forelimb
Apart from a multitude of cranial characters, C. marginata also shares important characteristics of the postcranial skeleton, including some of the forelimb, with the cetotheres. Insight into the morphology of C. marginata could thus elucidate the lifestyle in cetotheres. The earliest and only detailed description of the pygmy right whale skeleton was given by Beddard in 1901. Based on two specimens in the collection of the Natural History Museum in London, Beddard did not give any accounts of the soft tissue in his report. Furthermore, mounted museum specimens or depictions of articulated skeletons are notoriously incongruent where the position of the forelimb is concerned (e.g. Hector, 1875 pl. XVI - fig.1 vs. Beddard 1901 pl. VII). There is no bony connection to the rest of the skeleton or bony features in the skeleton itself that would allow a precise placement and orientation of the shoulder blade once the soft tissue has been removed. Consequently, correct position and orientation of the forelimb can only be ascertained in specimens where the musculoskeletal arrangement is still intact. The stranding of two solitary pygmy right whales on New
Zealand’s shores offered therefore a unique opportunity to investigate the musculoskeletal system of the forelimb in this enigmatic whale.
Results Position of the forelimb
Fig. 2. Position of the forelimb Semi-schematic drawing of Caperea marginata showing the position and orientation of the scapula on the thorax, the forelimb in relation to the body, the axillary fold and the indent of the flipper on the flanks. Note: fused first and second ribs are counted as rib 1. With permission modified from line art by S. Rommel

C a p e r e a m a r g i n a t a
38
The cartilaginous cranial angle of the scapula directly abuts the lateral-most aspect of the exoccipital near the exoccipital-squamosal suture (Fig. 2), a region that is in other mysticetes occupied by the squamosal and in older literature sometimes referred to as ‘mastoid process’ (e.g. Schulte, 1916; see also Mead and Fordyce, 2009). Here the anterior part overlies the splenius and is partly deflected outwards by it. The adjacent
bony cranial angle of the scapula thus lies just forward of the first rib (i.e. fused first and second rib sensu Buchholtz, 2011) at about the height of the costal angle. From here the vertebral border traverses the ribs in caudo-ventrad direction descending at an angle of about 25º from the body axis. Caudally the cartilaginous caudal angle reaches as far back as the 5th rib. Below the scapula there is a shallow, superficial indent of the skin and underlying blubber running caudally as far as the 6th rib. Pectoral limb
In the pygmy right whale, the ratio of body- to forelimb length is about 6 in MM2959 and 8 in MM2904. The contour of the flipper is almost perfectly oblong elliptical
(Fig. 3a) with a length / width ratio of about 5.5 in MM2904 and 6.0 in MM2959. The axillary fold does not quite reach halfway down the trailing edge of the flipper in MM2959 (Fig. 3c) whereas in MM2904 it extends slightly over the midpoint to the height of the carpals, giving the extended flipper a triangular shape more reminiscent of a shark fin (Fig. 3b). There is a distinct indent on the flanks big enough to accommodate the retracted flipper. Both the axillary fold and indent of the flank can also be clearly recognised in live animals as documented in photographs by Ross et al., 1975 in figs. 1 and 3. In MM2959, the lateral surface of the flipper (Fig. 3a) is a medium dark slate grey with faint irregular lighter cloudy patches, and contrasts strongly with the pale grey of the flank. The medial surface of the flipper (Fig. 3c) is similarly pale grey and streaked with hints of pink. At the leading edge the pale grey converges into the darker lateral surface as irregular marbled patches, more prominent towards the tip of the flipper. The proximal trailing edge sharply contrasts with the darker colour of the lateral surface. At the tip, the distal medial surface, and the trailing edge, the pale grey is interspersed with streaks of darker slate grey giving the impression of four digits. In MM2904 the skin shows substantial damage (Fig. 3b), but whether from the stranding event itself or through the successive freezing process could not be determined, and the true colour

C a p e r e a m a r g i n a t a
39
was uncertain. Nevertheless, apart from being overall darker, the contrast in colour between the superficial surface of the flipper and the flank seemed to be less distinct than in MM2959.
Fig. 3. Shape and colour of the flipper (a) Lateral view of MM2959, left flipper, (b) dorsal view of MM2904, left extended flipper showing large axillary skin fold demarcated by dashed line, (c) ventral view of MM2959, left flipper. Note indent of the flank to accommodate flipper, dashed line in a and c. Photos (a-c) R. E. Fordyce
Osteology See also plates I-II and measurements in appendix
Scapula
Two very distinctive profiles of the (bony) shoulder blade were found in this study, one fan-shaped with rounded vertebral border (MM2904, Fig. 5 a,b) and the other trapezoid with tripartite angular vertebral border (MM2959, Fig. 4 a,b). Both profiles are twice as long as high with a corresponding ratio of 2:1 in MM2959 and just under 2:1 in MM2904. The angular vertebral border in MM2959 is divided into three distinct sections: a convex cranial aspect facing dorso-craniad, a level and slightly concave mid-section, and a faintly convex caudal aspect facing dorso-caudad. Occupying about 1/5th of the
vertebral border, the cranial aspect is shorter and, at an angle of about 50º, slightly steeper, while the bulkier caudal aspect occupying about 1/3rd of the vertebral border follows with a less pronounced curvature at a shallower angle of about 40º. Conversely, the fan-shaped blade of MM2904 presents a symmetrical and consistently convex vertebral border.

C a p e r e a m a r g i n a t a
40
Fig. 4. Right scapula of MM2959 (a) lateral view, (b) medial view. Missing cartilage indicated by dashed line.
Fig. 5. Left scapula of MM2904 (a) lateral view, (b) medial view. Note cartilaginous border separated during dissection.
In dorsal view, the scapular blade generally follows a slightly sigmoidal curvature. The cranial part curves most prominently laterad at the vertebral border whereas the caudal part is displaced mediad more ventrally towards the glenoid fossa. Apart from the diminutive scapular spine neither the medial nor the lateral surfaces show prominent ridges or strong variances in thickness. In both animals the cartilaginous (vertebral) border extends relatively far beyond the bony limit of the cranial and caudal aspect yet remains remarkably narrow over the mid-section. In doing so it forms two broad lobes straddling the bony angles of the blade dorso-cranially and dorso-caudally. This is particularly conspicuous in MM2959: here the cranial lobe exceeds the bone by a further 22 mm craniad and the caudal lobe by 28 mm caudad, thus further accentuating the elongation of the bony blade (Fig. 4a,b). Conversely, the narrow cartilaginous central section of the vertebral border
extends only 2 mm above the bony mid-section and strongly emphasises the concave

C a p e r e a m a r g i n a t a
41
curvature of the bone. The resulting ratio of the overall scapular length to height is thus over 2.5:1 in MM2959 and slightly under 2.5:1 in MM2904. In MM2959, the cranial border rises towards the longitudinal axis of the scapula at an angle of about 35º. On the medial side, the outline appears as a gentle sinuous curve, changing from concave at the neck to convex more dorsally. The cranial border in
MM2904 converges with the longitudinal axis at about 30º, slightly less steep but more straight, only interrupted by the short anterior edge of the supraspinous fossa extending beyond the anterior edge as a faintly convex but distinctive curvature. On the lateral cranial surface of the scapula, there is a low but sharp concave ridge, particularly in MM2959, demarcating the scapular spine at the posterior boundary of the supraspinous fossa. The supraspinous fossa is at its greatest width about 1/25th (MM2959) to 1/35th (MM2904) of the (entire bony) vertebral border and gently narrows dorsally into a point. In MM2959 the supraspinous fossa barely reaches the vertebral border, and in MM2904 it is confined to the lower 3/4 of the cranial border. The caudal border of the scapula rises initially steeply from the neck towards the vertebral border but soon curves caudally in alignment, and level with, the ventral edge of the acromion process, both roughly running parallel to the cartilaginous border of the glenoid cavity. In MM2959 this curvature is a gentle arch of about 135º, whereas in MM2904 the neck rises steeper and the curvature towards the caudal border is more pronounced at an angle of about 115º. Cranial and caudal border meet in MM2959 at an angle of about 120º whereas in MM2904 this angle is more open at about 135º. The acromion process is flat and broad, in lateral view rectangular, and only at the tip slightly rounded or delta-shaped. The cartilaginous tip is pentagonal in MM2959 and reduced to a small strip in MM2904. In ventral view, the acromion curves slightly inwards towards the scapular plane but does not cross it.
The coracoid process is rather small, more so in MM2959 where it is completely cartilaginous. Noteworthily, just after the dissection the coracoid in MM2959 curved distinctly upwards, running almost parallel to the anterior border of the scapula, but after drying projected straight forward along the same line as the glenoid border. In ventral view it diverges away from the cranial border, thus creating a U-shaped trough (see also Chapter 4 Functional anatomy: Functional shift of the scapular spine, acromion, coracoid and olecranon). The glenoid fossa is relatively shallow and, in ventral view, oblong-oval. There is no visible glenoid nipple. In lateral view the border of the glenoid cavity rises dorso-caudad

C a p e r e a m a r g i n a t a
42
at an angle of about 25º towards the axis of the scapula and almost parallel to the caudal border, the angle of the bony glenoid border is at about 20º slightly less. Long bones
The humerus (Figs. 6a,e and 7a,e) is short and robust and about 2/3rd to 4/3rd of the radius in length. The hemispherical head is moderately small and situated posterolaterally behind the tubercles; in lateral view, the head does not exceed the width of the distal humeral epiphysis. The greater tubercle, extending as a long ridge antero-medially of the head, is only little separated from the much smaller, concentric, lesser tubercle on the medial aspect of the head. In MM2959 the tuberosity is entirely cartilaginous. The tuberosity of MM2904 is incompletely ossified and only lightly fused to the diaphysis. The distal epiphysis is not fused in either animal and is entirely cartilaginous in MM2959. Dissection revealed a synovial elbow joint. The two facets of the humerus opposing the radius and ulna converge in lateral view in the shape of a fairly symmetrical V at an angle of about 130º. In distal view the two facet elements are
broadly-oblong and along the central line slightly dished. The ulnar facet extends only very slightly onto the posterior edge of the diaphysis. There is no distinct deltopectoral crest or tuberosity. The radius (Figs. 6b,f and 7b,f) is long and straight; the length / width-ratio of the diaphysis is about 5:1 in MM2959 and about 4.5:1 in MM2904. In lateral view, the shaft is slightly wider in the middle. In MM2959, the caudal edge remains fairly straight, but the cranial edge follows a faintly sinuous concave-convex-concave curvature and terminates in a slightly bulbous distension abutting the proximal epiphysis. The shaft in MM2904 is more regular and only slightly distended centrally. Distally the diaphysis is not fully ossified and disintegrates into soft and spongy bone, whereas in MM2959 it is more ossified. In lateral view, the proximal facet is straight and in distal view subrectangular. The proximal epiphysis is not fused in either animal. In MM2904 there is no ossification centre recognisable in the distal epiphysis, whereas in MM2959 radiographs reveal a faint shadow indicating a very early stage of ossification in that area.

C a p e r e a m a r g i n a t a
43
Fig. 6. Right forelimb of MM2959 (a-d) lateral view, (e-h) medial view; (a,e) humerus; (b,f) radius; (c,g) ulna; (d,h) manus. Abbreviations: (2-5) digits 2-5.

C a p e r e a m a r g i n a t a
44
Fig. 7. Left forelimb of MM2904 (a-d) lateral view, (e-h) medial view; (a), (e) humerus; (b), (f) radius; (c), (g) ulna; (d), (h) manus. Abbreviations: (2-5) digits 2-5.

C a p e r e a m a r g i n a t a
45
The ulna (Figs. 6c,g and 7c,g) is slender and comparatively straight; length / width-ratio of the diaphysis decreases from the narrowest proximal point to the widest distal point by about 10:1 to 5:1 in MM2959 and in MM2904 from 8:1 to 4.5:1. Proximally the ulnar shaft widens into a flange attaining a length / width-ratio of about 4:1 in MM2959 and 3:1 in MM2904 to support the large cartilaginous part of the olecranon. The
proximal articular facet is longer than that of the radius and in lateral view straight, except caudally where it follows the curvature of the humeral facet onto the posterior edge of the humerus. In the lateral view the axis of the olecranon diverges ventro-caudad from the axis of the ulna at an angle of about 30º in MM2959 and 40º in MM2904. It is roughly half as long as the ulna, spoon-shaped (MM2959) or spatulate-shaped (MM2904), distally flattened and slightly skewered towards the ulnar shaft. Diaphysis and proximal and distal epiphyses match the developmental state seen in the radius. Manus
Carpals of the proximal row and distal row are present but not ossified (Figs. 6d,h and 7d,h). Each element is delimited by well-defined surface grooves on both the flexor and extensor side with the exception of a nearly indistinguishable border between the ulnar epiphysis and the pisiform; in MM2904 the pinkish carpal elements are also visually clearly discernible by whitish grooves (Fig. 8). The proximal row consists of a radiale, intermedium, ulnare and pisiform; the distal row is possibly a fusion of carpal 2 and 3 (C2+3), and carpal element 4 and 5 (C4+5; see also Chapter 3 Berardius: Osteology, section Manus). The first distal carpal is absent. Fused C4+5 are hexagonal, intermedium and fused C2+3 pentagonal, and radiale and ulnare tetragonal. Apart from the pisiform, there are no accessory carpal elements. In MM2959 the pisiform is long, slender and oblong-spatulate, with a rounded tip; the pisiform is distally curved towards the tip of the flipper. In MM2904 the pisiform is much more robust than in MM2959, and remains broad, straight and short.

C a p e r e a m a r g i n a t a
46
Fig. 8. Carpal arrangement in MM2904 Left manus, lateral view. Note the short and broad outline of the pisiform and the almost invisible border between the ulnar epiphysis and the pisiform.
In the pygmy right whale, the ratio of radius / manus-length is roughly 1:1. The flipper is supported by only four digits: the first digit is absent (see also Cooper et al., 2007a). All four metacarpals are hourglass-shaped, as are the first and second phalanges of digits 2, 3 and 4 (D2-4: Ph1-2), although some are foreshortened and more rectangular-looking (MM2904 D3: Ph2). In MM2904 the first phalanx of digit 5 is rounded (Table 1 and Plate II - Figs. 1-2), and the last two finger elements (phalanges 2 and 3) are not visible in the radiograph but are present as cartilaginous elements. In MM2959 the contour of the first phalanx of digit 5 takes on a trapezoid shape, and the
second a rounded (Table 1 and Plate I - Fig. 2, right forelimb, faint in radiograph) or foreshortened hourglass shape (Table 1 and Plate I - Fig. 1, left forelimb). In digit 2 of MM2959, there is clearly a third, cartilaginous phalanx present which might not be present in MM2904. Phalanx 3 in digits 3 and 4 appears much more variable and can be more or less hourglass-shaped (MM2959), deltoid or hexagonal (MM2904). Beyond phalanx 3, there are no ossified phalanges present in either specimen but two or three cartilaginous elements, the last one (tip) minute and fragile (digits 2 and 3 in MM2904 Ph 4-5, and in MM2959 Ph 4-6). The general phalangeal count is thus for MM2904 0/2/3/3/1 (bony elements only) and 0/2-3?/5/5/3 (unossified elements included) and for MM2959 0/2/3/3/2 and 0/3/6/6/3 respectively (see also Dawson, 2003).

C a p e r e a m a r g i n a t a
47
Table 1. Phalangeal shape and ossification MM2904 MM2959
D2 D3 D4 D5 D2 D3 D4 D5
MC ⧖ ⧖ ⧖ ⧖ MC ⧖ ⧖ ⧖ ⧖
Ph 1 ⧖ ⧖ ⧖ O Ph 1 ⧖ ⧖ ⧖ ⏢
Ph 2 ⧖ ⧗ ⧖ c Ph 2 ⧖ ⧖ ⧖ O / ⧗
Ph 3 c? D ⎔ c Ph 3 c ⧗ ⧖ c
Ph 4 c c Ph 4 c c Ph 5 c c Ph 5 c c Ph 6 Ph 6 c c
Abbreviations: D2-5 digits 2-5, MC metacarpal row, Ph1-6 phalangeal row 1-6
Symbols, ossified elements: ⧖ hourglass shape, ⧗ foreshortened hourglass shape, O round, ⏢
trapezoid, D deltoid, ⎔ hexagonal; c cartilaginous elements; ? element not found but possibly present.
Myology
All details given for Caperea marginata are based on the results of the dissections described in Chapter 1 Methods and materials: Dissections. The results are illustrated below in figures 9-13. Unless otherwise stated, general muscular arrangements and actions in Cetacea apply to Caperea marginata (see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscle distribution, and Chapter 5 Literature review: Myology, Table 2). Identification and terminology of muscles are predominantly based on Strickler, 1978 and Cooper et al., 2007b, and muscle action on Benke, 1993 or Smith et al., 1976. Panniculus carnosus (Fig. 9)
In the course of the dissection the exact anterior and posterior borders could not be
established but as a whole the muscle followed the general cetacean pattern (e.g. Stannius, 1849; Schulte, 1916; Schulte and Smith, 1918; Howell, 1927, 1930b). As a general rule, the panniculus extends as an uninterrupted, thin muscular sheet that roughly covers the areas corresponding to the body cavities. It is divided into three major regions: (1) throat and neck, (2) shoulder and chest, and (3) trunk region, the latter two further separated into a dorsal and a ventral division. In the region of the neck, the panniculus arises just behind the gape of the mouth, ascending steeply dorsad in the area posterior to the occiput. Here the fasciculi

C a p e r e a m a r g i n a t a
48
completely surround the auditory meatus (meatus acusticus) before inserting into a broad aponeurosis that covers the dorsal midline and the epaxial musculature. Inclination of the fasciculi and anterior border of the ventral division could not be established. In the region of the shoulder and chest, the dorsal and ventral divisions of the
panniculus are separated by the flipper. Dorsally, the fasciculi cover the entire shoulder ventrad to the thick fascia separating the hypaxial from the epaxial musculature, across the scapula and down to the shoulder joint where they insert into a tough raphe covering the proximal portion of the flipper. The fasciculi in this area are inclined dorso-caudad, arising at an angle of about 65º from the lateral midline. Passing ventrad of the flipper, the fasciculi cover the sternum and the pectoralis major, inserting into the aponeurosis at the base of the flipper in common with the pectoralis major. Caudally, the fasciculi insert into the raphe of the axillary skin fold. In the trunk region, the raphe of the axillary fold continues caudally as a thin band separating the two divisions of the panniculus. The inclination of the fasciculi is here about 45º towards the longitudinal axis in dorso-craniad (ventral division) and ventro-craniad (dorsal division) directions and thus about 90º towards each other. Dorsally, the boundary of the panniculus initially follows the fascia between the hypaxial and epaxial musculature before gently arching ventrad towards the vent. The ventral division continues uninterrupted, ventrally delineated by the ventral midline and dorsally by the lateral raphe. Caudally the panniculus could be traced back to about mid-vent.
Fig. 9. Panniculus Semi-schematic drawing of the panniculus showing fasciculi extent and fasciculi inclination, lateral view. Parallel slanted lines show inclination of the fasciculi, broken lines indicate inferred extent of the muscle and fasciculi inclination With permission modified from line art by S. Rommel

C a p e r e a m a r g i n a t a
49
Muscles of the shoulder and forelimb
Fig. 10. Lateral forelimb musculature Superficial musculature of the shoulder and forelimb in Caperea marginata; semi-schematic illustration of MM2904.

C a p e r e a m a r g i n a t a
50
Fig. 11. Origin and insertion of the lateral forelimb musculature Superficial musculature of the shoulder and forelimb in Caperea marginata; schematic illustration of MM2904. Areas in light grey represent bone, medium grey cartilage, and dark grey corresponds to the silhouette of the flipper. Abbreviations: (2-5) digits 2-5.

C a p e r e a m a r g i n a t a
51
Fig. 12. Medial forelimb musculature Deep musculature of the shoulder and forelimb in Caperea marginata; semi-schematic illustration of MM2904.

C a p e r e a m a r g i n a t a
52
Fig. 13. Origin and insertion of the medial forelimb musculature Deep musculature of the shoulder and forelimb in Caperea marginata; schematic illustration of MM2904. Areas in light grey represent bone, medium grey cartilage, and dark grey corresponds to the silhouette of the flipper. Abbreviations: (2-5) digits 2-5. Colour index of muscle groups (sensu Strickler, 1978): brown, costo-spino-scapular; red+orange, latissimus-subscapular; purple, deltoid; yellow, triceps; green, suprascapular; blue, flexor; turquoise, trapezius; pink, manus.

C a p e r e a m a r g i n a t a
53
Levator scapulae
Caperea marginata General cetacean condition
Origin Absent Ventral arch of atlas Insertion Absent Lateral cranial scapular angle, aponeurotic on
deltoid Action Slight cranial ward movement of scapula
No muscle corresponding to the levator scapulae in other cetaceans was found in C. marginata. This muscle is therefore here regarded as absent (see also Musculature in this chapter and Chapter 5 Literature: Myology, Table 2). Serratus anterior - anterior division (Figs. 12, 13)
Caperea marginata General cetacean condition (if present)
Origin Angle of first rib Transverse process of C7 Insertion Medial cranial scapular angle Medial cranial scapular angle Action Rotation of the scapula ventrad Slight cranial ward movement of scapula
Arises from the angle of the first rib (fused first and second rib sensu Buchholtz, 2011) and inserts onto the medial cartilaginous cranial angle of the scapula. Here, the muscle occupies the anterior 1/4 (MM2959) to 1/8th (MM2904) of the cartilaginous vertebral border and, although crossing the subscapularis ventrally, does not insert onto it.
Serratus anterior - posterior division (Figs. 12, 13)
Caperea marginata General cetacean condition
Origin 3-5th ribs 2-4th ribs Insertion Medial cartilaginous vertebral border
of scapula Medial cartilaginous vertebral border of scapula
Action Adducts scapula ventrad, rotation craniad
A strong, broad sheet of this muscle arises from the ventral portion of the 3rd to 5th ribs
and from the aponeurosis of the adjacent intercostal musculature. From there, the muscle converges dorsad towards the scapula where it inserts onto the medial cartilaginous caudal angle and as aponeurosis onto the adjacent surface of the subscapularis.

C a p e r e a m a r g i n a t a
54
Rhomboideus capitis
Caperea marginata General cetacean condition (if present)
Origin absent Supraoccipital Insertion absent Lateral cranial scapular angle Action Slight craniad movement of scapula (Klima et
al., 1980)
Rhomboideus vertebralis (Figs. 10, 11)
Caperea marginata General cetacean condition
Origin 3-5th ribs Epaxial fascia between T1-6 Insertion Edge of cartilaginous vertebral border
of scapula Vertebral border of scapula
Action Adducts scapula dorsad
A single sheet arises from the dorsal portion of the 3rd to 5th ribs and the aponeurosis covering the adjacent intercostal musculature. Ventrally the rhomboideus inserts laterally onto the edge of the anterior 3/4 of the cartilaginous vertebral border. The rhomboideus does not extend further caudad than the bony caudal angle of the scapula. Latissimus dorsi (Figs. 10, 12, 13)
Caperea marginata General cetacean condition
Origin 6-9th ribs 4-6th ribs Insertion Medio-caudal on humerus neck Mid medio-caudal edge of humerus Action Flexes humerus
Arises as a flat, thin, broad tendinous sheet from the dorsal portion of the 6th to 9th ribs, a few fibres also arising from the 10th rib, and the aponeurosis of the adjacent intercostal musculature. From the point of origin, the muscle descends to a mid-ventro-
lateral position inserting onto the flipper. Muscular fibres arise as a thin sheet mid-lateral forward of the 6th rib and converge to a flat belly towards their insertion. In MM2959 the muscle passes about 3 cm below and parallel to the caudal border of the scapula. In MM2904 the arrangement of the muscle and scapula differs slightly: here a section of the dorsal portion of the latissimus dorsi partially covers the cartilaginous caudal angle, including the underlying teres major, before merging with the latter ventrally. The latissimus dorsi inserts in common with the teres major onto the medio-

C a p e r e a m a r g i n a t a
55
caudal surface of the humeral neck, distal to the subscapularis (Fig. 13) and mediad of the triceps (humeral head) insertion (Fig. 11). Teres major (Figs. 10, 11, 12, 13)
Caperea marginata General cetacean condition
Origin Lateral cartilaginous vertebral border of scapula
Caudal border of scapula
Insertion Medio-caudal on humerus neck Mid medio-caudal edge of humerus Action Flexes and/or rotates humerus inwards
Arises just short of the caudal 1/3rd of the lateral cartilaginous vertebral border and caudal scapular angle also including the caudal border and intermuscular fascia in common with the subscapularis. In MM2904 the caudo-ventral part of the teres major lies under cover of the latissimus dorsi before distally merging with the fibres of the
latter. The teres major, in common with the latissimus dorsi, inserts onto the medio-caudal surface of the humeral neck. Subscapularis (Figs.12, 13)
Caperea marginata General cetacean condition
Origin Medial surface of scapula Medial surface of scapula Insertion Lesser tubercle of humerus Lesser tubercle of humerus Action Adduction and inward rotation of humerus
Occupies the entire (medial) subscapular fossa and intermuscular fascia between the caudal angle and posterior glenoid border in common with the teres major. The subscapularis does not encroach onto the cartilaginous surface of the vertebral border. Radiating from the vertebral border, four larger and multiple smaller incomplete tendinous intersections divide the muscle into 5 slips. Converging into a flat tendon, the subscapularis is firmly attached to the capsule of the shoulder joint and inserts onto the lesser tubercle between the insertions of the more distal teres major / latissimus dorsi (Fig. 13) and the more cranial insertion of the coracobrachialis (Fig. 13).

C a p e r e a m a r g i n a t a
56
Deltoideus (Figs. 10, 11)
Caperea marginata General cetacean condition
Origin Cranial lateral surface of scapula Cranial lateral surface of scapula Insertion Lateral surface of humerus
Cranial edge of radius Lateral surface of humerus
Action Abducts and/or extends and reverses inward rotation of humerus
The deltoideus arises from the anterior 2/5th of the lateral cartilaginous vertebral border of the scapula, the cranial angle, the acromion and the thick fascia spanning between the latter two. Like the subscapularis it is firmly attached to the periosteum covering the scapular surface. Caudally the deltoideus overlaps the infraspinatus and it is firmly attached to the aponeurosis between the two. At the cranial border the deltoideus completely overlies the supraspinatus, extending a few millimetres beyond the tip of
the acromion to which it is attached. Ventrad of the acromion, the medial surface of the deltoid is attached to the intramuscular fascia between it and the supraspinatus. Distally the coarse fascicles of the deltoideus converge into a tough flat tendon, which initially appears on the surface. At their insertion the tendinous fibres broadly attach to the anterior border and preaxial lateral surface of the humerus immediately below the insertion of the infraspinatus, narrowing rapidly caudad to a small band across the postaxial area below the humeral neck. Under cover of this tendon, the muscle remains fleshy to the point of insertion. There is a second, rudimentary belly arising as tendinous fibres from the distal anterior border of the humerus adjacent to the insertion of the first. This secondary belly is slightly displaced towards the medial humeral surface and inserts just below the epiphysis onto the proximal anterior border of the radius. Teres minor / Subdeltoideus
Caperea marginata General cetacean condition (if present)
Origin Absent Mid-lateral surface of scapula Insertion Absent Proximal lateral surface of humerus Action Rotates humerus inwards
Among mysticetes, this muscle has only been reported for Balaenoptera borealis (Schulte, 1916; labelled as ‘supraspinatus’ in Benke, 1993) and is not present in C.

C a p e r e a m a r g i n a t a
57
marginata (see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscle distribution). Triceps brachii (Figs. 10, 11, 12, 13)
Caperea marginata General cetacean condition (if present)
Origin Caudal on neck of scapula Caudo-lateral edge of humerus Absent
Caudal on neck of scapula (long head) Lateral on humerus shaft (external head) Medial on humerus shaft (medial head)
Insertion Olecranon Olecranon Action Flexes and rotates humerus inwards (long head)
Not known (humeral heads)
There are two heads present in C. marginata. The scapular (long) head arises from the capsule of the shoulder joint and the postaxial neck of the scapula as a long, broad
and fleshy band. Distally the fasciculi appear on the surface as a tough flat tendon surrounding the fleshy interior of the muscle and, after crossing the teres major superficially, embrace the other head of the triceps laterally at its distal portion and insertion. Here the tough tendinous fibres insert onto the whole lateral surface and postero-proximal edge of the olecranon tip, concealing the insertion of the humeral (triceps) head. The fleshy part of the muscle continues under cover of the tendinous fibres down to the edge but not onto the surface of the olecranon. The humeral (external) head arises between the insertions of the deltoideus and teres major as a narrow band slightly displaced towards the lateral plane along the entire caudal border of the humerus. A strong tendinous sheet covers a broad and well-developed fleshy muscle belly inserting onto the proximal edge and the whole lateral surface of the olecranon, here under cover of the scapular (triceps) head. The fleshy belly extends down towards the proximal edge of the olecranon but does not encroach onto its lateral surface. Supraspinatus (Figs. 11, 12, 13)
Caperea marginata General cetacean condition
Origin Supraspinous fossa of scapula Supraspinous fossa of scapula Insertion Greater tubercle of humerus Greater tubercle of humerus Action Extends and rotates humerus outwards

C a p e r e a m a r g i n a t a
58
Arises from the supraspinous fossa, the fascia between the cranial angle and the acromion, the medial surface of the acromion, and the medial surface of the coracoid. The muscle inserts as a tendon onto the cranial, proximal aspect of the greater tubercle anterior to the insertion of the infraspinatus (lateral) and proximal to the mastohumeralis (medial).
Infraspinatus (Figs. 10, 11)
Caperea marginata General cetacean condition
Origin Mid-lateral surface of scapula Caudo-lateral surface of scapula Insertion Proximal cranio-lateral on humerus Cranio-lateral surface of humerus Action Abducts humerus
Interposed between the deltoideus in front and the teres major at the back, the infraspinatus covers just under 1/3rd of the superficial cartilaginous border but is only
partially visible between the gap of the former two. The muscle extends between the cranial 2/5th and the caudal 1/3rd of the scapular vertebral border. In outline, the anterior part of the infraspinatus is initially only little obscured by the posterior portion of the deltoideus. It descends towards the anterior glenoid border in a straight line to about 1/3rd (MM2904) to 1/2 (MM2959) of the scapular height. At this point the anterior border turns craniad and thus forms a bay outlined by a gentle arch in MM2904 that is much more pronounced in MM2959. This bay is delineated by a multitude of vaguely defined slips reaching as far as the posterior border of the supraspinous fossa in MM2959. The outline of this bay is not well defined in MM2904, as opposed to MM2959 where the anterior demarcation line is much more distinct. Turning once again ventrad, the anterior margin aligns with the anterior glenoid border and continues in a straight line towards its point of insertion. Caudally, the infraspinatus abuts onto the anterior portion of the teres major but is not obscured by it. It remains in contact with the teres major down to the caudal scapular border and, in the region of the neck, is intercepted by the origin of the scapular head of the triceps. Crossing the shoulder joint laterally, the infraspinatus is, at this point, fully concealed by the deltoideus. Remaining under the cover of the deltoideus, the infraspinatus inserts as a broad tendinous band onto the lateral surface of the humerus just distal to the humeral head, embracing the head as a broad collar.

C a p e r e a m a r g i n a t a
59
Pectoralis minor
Caperea marginata General cetacean condition
Origin Absent Sternum Insertion Absent Tip of coracoid process Action Rotates scapula ventrad
The pectoralis minor is regarded as absent in mysticetes (Strickler, 1978). This postulation is supported by this study where no muscle corresponding to the pectoralis minor in odontocetes has been found in C. marginata (see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscle distribution, Table 2). Pectoralis major (not depicted)
Caperea marginata General cetacean condition
Origin Sternum Sternum Insertion Medial base of flipper Medial on flipper Action Adducts and/or flexes humerus
The origin of the pectoralis major can be traced along the linea alba and the superficial surface of the sternum as a broad, thin sheet of musculature, that could be traced back to the ventral aspect of the third rib. The fasciculi maintain a transverse direction throughout. Insertion is onto connective tissue broadly across the medial base of the flipper, along the proximal edge of the olecranon and the axial raphe in common with
the panniculus. Pectoralis abdominalis (not depicted)
Caperea marginata General cetacean condition
Origin Absent Costal cartilages 4-6 Insertion Absent Medial, posterior base of flipper Action Flexes humerus (inferred sensu Strickler, 1978)
Not identified but could have been removed during field recovery. This muscle occurs only sporadically in cetaceans and is possibly absent in Caperea marginata.

C a p e r e a m a r g i n a t a
60
Coracobrachialis (Figs. 11, 12, 13)
Caperea marginata General cetacean condition
Origin Medial surface of coracoid process Medial surface of coracoid process Insertion Greater tubercle of the humerus Just distal to lesser tubercle of the humerus Action Adducts humerus Adducts and slightly flexes humerus
The coracobrachialis arises as a round and short but (compared to other cetaceans) comparatively well-developed muscle from the tip and adjacent surfaces of the coracoid process. On the medial side, it occupies the whole width and the cranial 1/3rd of the coracoid process, whereas on the lateral surface it is restricted to the lower corner of the tip by the insertion of the supraspinatus above. Crossing the subscapularis, the muscle inserts as a tendon onto the posterior aspect of the greater tubercle, flanked by the insertions of the subscapularis caudally and the supraspinatus and mastohumeralis cranially.
Omohyoideus (Figs. 10, 12, 13)
Caperea marginata General cetacean condition
Origin Cranial border of scapula Aponeurotic on cranial angle of scapula Insertion Greater horn of hyoid Hyoid Action Craniad movement of the scapula (or possibly
caudad movement of the hyoid respectively)
The omohyoid is present as a fleshy belly arising from the mid-scapular cranial border and is anchored to the edge of the whole cranial border by fibrous tissue. From here it emerges between the supraspinatus and the deltoid and, crossing the neck obliquely and mediad, inserts into the greater horn of the hyoid. Mastohumeralis (Figs. 10, 12, 13)
Caperea marginata General cetacean condition
Origin Exoccipital Exoccipital Insertion Greater tubercle Mid-cranial edge of humerus Action Extends and rotates humerus inwards
The mastohumeralis arises from the lateral portion of the exoccipital and inserts as a round tendon onto the medial anterior aspect of the greater tubercle distal to the supraspinatus.

C a p e r e a m a r g i n a t a
61
Extensors (Figs. 10, 11) Extensor digitorum communis
Caperea marginata General cetacean condition (if present)
Origin Lateral interosseus membrane Lateral interosseus mambrane of radius and ulna Insertion Digits 2-5 Digits 2-5 Action Slight extension of digits
The extensor digitorum communis arises from the elbow joint and adjacent radial and ulnar surfaces, as well as the interosseus membrane in between. The fleshy belly is slightly displaced towards the radial surface. Distally, the muscle fibres form a flat tendon about midway between the long bones of the forearm, splaying out wide across the carpals and dividing into four tendinous slips, one for each digit. The middle two tendons follow the axial line of their respective digits, the preaxial slip follows the postaxial border of digit 2, and the postaxial slip continues along the preaxial border of
digit 5. All four slips divide quite symmetrically onto their respective digits, the preaxial slip turning towards the tip of the flipper over metacarpal 2 slightly more abruptly than the postaxial slip. The tendons are united by fibrous tissue to the perichondrium of the phalanges; the fibrous tissue is particularly firm at the enlarged interphalangeal joints. Flexors (Figs. 10, 12, 13) Flexor digitorum radialis (Figs. 12, 13)
Caperea marginata General cetacean condition (if present)
Origin Medial postaxial surface of radius Medial postaxial edge of radius Insertion Fused with flexor communis digitorum Absent or fused with flexor communis digitorum Action Slight flexion of preaxial digits
The flexor digitorum radialis arises as a slender muscle from the elbow joint, the medial postaxial surface of the radius, the interosseus membrane, and the intermuscular fascia of the adjacent flexor ulnaris. The muscular fibres reach distally as far as the middle of the long bone before converging to a flattish tendon descending in a gentle curve craniad towards the anterior carpals. Attaining the intermedium, it fuses with the tendon of the flexor communis digitorum. The tendon remains fused over the carpals and splits again over the proximal epiphysis and diaphysis of metacarpal 2, one slip continuing craniad, the other turning more distad following digit 2. Distally, both slips radiate in a fan-like manner to insert along the fibrous tissue of the leading edge of the

C a p e r e a m a r g i n a t a
62
flipper, the shorter and more proximal slip anterior to metacarpal 2, and the longer more distal slip anterior to the first phalanx to the tip of digit 2. Flexor digitorum communis (Figs. 12, 13)
Caperea marginata General cetacean condition (if present)
Origin Medial proximal surface of ulna Medial proximal surface of ulna Insertion Digits 2-5 Digits 2-5 Action Slight flexion of digits
The flexor digitorum communis, or flexor digitorum ulnaris, arises from the medial proximal surface of the ulna and the bony base of the olecranon. Further distally it is also firmly attached to the interosseus membrane and the intermuscular fasciae shared with the flexors digitorum radialis anteriorly and carpi ulnaris posteriorly. The fleshy fasciculi converge about midway on the cranial edge of the ulnar shaft, as a flattish
tendon, from where they continue distad until they fuse anteriorly with the tendon of the radial digital flexor over the distal radial epiphysis and preaxial carpals. The caudal part of the tendon splits into three slips over the carpals. The two axial slips split in the region of the intermedium` and follow digits 3 and 4, continuing roughly in the line of the main tendon. The third, most caudal, slip diverges in the region of the ulnare in a gentle curve towards the trailing edge of the flipper, before following the axis of digit 5. In the region of the carpals, the tendons remain comparatively thick and fleshy, and only gradually transform distally of the metacarpals into flat tendons. The proximal, more bulky part of the tendons is covered by a thick synovial sheet and is still functionally contractile in the region of the metacarpals. The flat tendons further distad are firmly anchored to the digits by fibrous tissue, particularly over the enlarged interphalangeal joints. Flexor carpi ulnaris (Figs. 11, 12, 13)
Caperea marginata General cetacean condition
Origin Distal olecranon Distal olecranon Insertion Proximal pisiform Proximal carpus Action Not known
The flexor carpi ulnaris arises from the anterior medial edge of the cartilaginous base of the olecranon and the intermuscular fascia to the flexor digitorum ulnaris. Distally

C a p e r e a m a r g i n a t a
63
the fleshy fibres converge into a thick, flat tendon before inserting onto the proximal edge of the pisiform.
Discussion Forelimb morphology of C. marginata reveals a remarkable general similarity to that of extant rorquals, rather than that of balaenids with which it was initially associated with (e.g. Hector, 1878; also see phylogeny Fig. 1). Some unique features (among extant cetaceans) seen in the pygmy right whale, particularly of the shoulder blade, can also be seen in Piscobalaena nana (Family Cetotheriidae), an extinct fossil whale from the early Pliocene (5.3-3.6 my; Bouetel and de Muizon 2006). The findings also confirm the only detailed description of the forelimb in C. marginata, described and depicted by F. E. Beddard in 1901 (pl. VII) and by Hector in 1875 (pl. XVI - fig. 1 and pl. XVII - fig. 10). This specimen is the skeleton of a subadult female initially recovered
and described by Mr. Charles Traill, further elucidated by Hector in 1875 and then by Beddard in 1901. It is now held at the Museum of New Zealand (MoNZ, MM0036, photograph available at https://collections.tepapa.govt.nz/object/560376). In his observations Beddard also recognised character traits of the pygmy right whale flipper as closer to those of rorquals rather than balaenids, namely flipper shape and size, tetradactyly and a ratio of humerus / radius length of 4:6. Investigations of the soft tissue, not available to Beddard, show a similar general pattern comparable with that of the minke whale (Balaenoptera acutorostrata, Carte and Macalister, 1868), but some specific arrangements of individual muscles are more congruent with that of the sei whale (Balaenoptera borealis, Schulte, 1916).
Flipper
With a total length / forelimb length ratio of about 6, MM2959, the smaller male neonate, associates closer with balaenids rather than with most rorquals (Fig. 14). Only marginally lower, this ratio is also seen in the fossil cetotheriid P. nana. In the larger and older female, MM2904, the ratio is considerably higher and with a ratio of 8 much more similar to that of rorquals (except humpback whale, Megaptera novaeangliae). Furthermore, when considering potential allometric development, this ratio may be

C a p e r e a m a r g i n a t a
64
even higher in fully grown adults. If correct, pygmy right whales, together with fin whales (Balaenoptera physalus), would have some of the smallest flippers found in extant baleen whales.
Fig. 14. Ratio of total skeleton length to forelimb length (based on Hector, 18751; Benke, 19932; Bouetel and de Muizon, 20063) Left column, colour key: Orange - Caperea marginata; Blue - Balaenopteridae; Dark grey – Eschrichtiidae4; Green - Balaenidae; Ochre - Piscobalaena nana; Right column, light grey - Total length of the skeleton (1) C. marginata MM0036; (2) Balaenopteridae and Balaenidae; (3) P. nana, estimates based on the length of the long bones,
the dentary and/or scapula using estimation values for Caperea marginata (see Kemper, 1999); (4) Estimate of TL/FL based on
photographic record provided by R. E. Fordyce of a specimen displayed in the Osteological Hall of the Smithonian Natural History
Museum, USA.

C a p e r e a m a r g i n a t a
65
This is supported by MM0036, the specimen described by Hector (1875) and Beddard (1901). Judging by the total length of the stranded whale, given as a 14 ft 6 in long female (442.0 cm) and a forelimb length as 16 in (40.6 cm), the animal was subadult (for age classification criteria see Chapter 1 Methods and materials). Accordingly, Hector’s measurements of the skeleton lead to an extremely high ratio of nearly 10.9,
a figure that by far exceeds the ratio of 8.6 seen in fin whales. The length / width ratio of the forelimb in MM2959 lies in between that of minke (Balaenoptera acutorostrata) and fin whale (Balaenoptera physalus) while MM2904 associates closer with blue whales (Balaenoptera physalus) overall (Fig. 15). Hector’s subadult female pygmy right whale (MM0036) on the other hand, exhibits distinctively wider flipper proportions at a ratio in between blue whales and gray whales (Eschrichtius robustus). Although blue whales have the broadest flippers within rorquals (Family Balaenopteridae), the only other tetradactyl family, the grey whales (Family Eschrichtiidae), are clearly separated from the balaenopterids.
Fig. 15. Ratio of forelimb length to forelimb width (based on Hector, 18751; Benke, 19932) Left column, colour key: Orange - Caperea marginata; Blue - Balaenopteridae; Dark grey - Eschrichtiidae3; Green - Balaenidae; Right column, light grey - Total length of the skeleton (1) C. marginata MM0036; (2) Balaenopteridae and Balaenidae; (3) Estimates of FL/FW and TL based on photographic record
provided by R. E. Fordyce of a specimen displayed in the Osteological Hall of the Smithsonian Natural History Museum, USA.

C a p e r e a m a r g i n a t a
66
This disparity forms a gap that is filled by Hector’s specimen. The flippers of C. marginata are thus broad when compared to rorquals, of medium width when compared to all tetradactyl baleen whales (Families Balaenopteridae and Eschrichtiidae), and fairly narrow compared to pentadactyl mysticetes (Family Balaenidae).
The ratio of humerus length / radius length is higher in the older female MM2904 (juvenile) than in the younger male MM2959 (neonate) and ranks just above that of gray whales (Eschrichtius robustus) and the cetotheriid P. nana SAS1617 (Fig. 16). Similar proportions have also been reported by Beddard (1901) for an immature animal of unknown sex and an adult male. MM2959 on the other hand groups closer to Hector’s pygmy right whale specimen (MM0036), minke whales (Balaenoptera acutorostrata) and P. nana SAS892. Nevertheless, the differences between these two groups are small, the ratio ranging from 0.67 to 0.74. The ratio in rorquals (including fin whales) ranges from 0.53 to 0.66 and is thus much closer to that of the pygmy right whales examined in this study, and possibly also gray whales and cetotheriids than to balaenids where it encompasses a range from 0.93 to 1.06 (data of balaenopterids, gray whales and balaenids based on Andrews, 1915 and Benke, 1993). Remarkably, despite having a lower humerus / radius length ratio than MM2904 or Hector’s specimen (MM0036), MM2959 has also the longest humerus proportionally to body size (except humpback whale), proportions that are more akin to those of cetotheriids. Hector’s specimen (MM0036) on the other hand, with a humerus / radius ratio only fractionally above that of MM2959, exhibits the shortest humerus to body size proportions of all whales investigated. A possible explanation of MM2959’s seemingly contradictory proportions could be
found in allometric growth patterns (Kemper, 1999). Very young animals may have comparatively long and narrow flippers with a long humerus as seen in MM2959. Growth patterns in cetaceans show that the humerus attains maturity earlier than the more distal bones (Galatius et al., 2006). This leads to a reduced growing rate of the whole flipper when compared with the growing rate of the whole body. These changed patterns would also explain the decreased ratio of forelimb length to forelimb width seen in MM2904, since flipper width is defined by the carpal elements that are still growing in younger animals. Radius and ulna, and finally carpals and phalanges may reach the end of their growing phase while the animal is still increasing in body size

C a p e r e a m a r g i n a t a
67
before becoming fully mature. This would explain the extremely short (to body size), but relatively broad (to forelimb length), flipper seen in Hector’s specimen. It does not explain the higher ratio of humerus / radius length in MM2904 compared to MM2959 and Hector’s specimen. This discrepancy, however, is much smaller than the other variables and may reflect individual variance rather than ontogenetic or interspecific
variation (which could not be established because of the small sample size in this study).
Fig. 16. Ratio of humerus length to radius length (based on Hector, 18751 and Beddard, 19011; Benke, 19932; Bouetel and de Muizon, 20063; Andrews, 19144) Left column, colour key: Orange - Caperea marginata; Blue - Balaenopteridae; Dark grey - Eschrichtiidae; Green - Balaenidae; Ochre - Piscobalaena nana; Right column, light grey - Total length of the skeleton (1) C. marginata MM0036; (2) Balaenopteridae and Balaenidae, (3) P. nana, (4) Eschrichtiidae.
Overall, rorquals exhibit a narrow but short flipper and a humerus that is about half to two thirds the length of the radius (except humpback whales; see Benke, 1993). Pygmy right whales have a tendency to short but moderately wide flippers with a somewhat shorter humerus than radius. A moderately short humerus / radius ratio is also seen in gray whales and P. nana, but gray whales tend to have longer and wider flippers than pygmy right whales. P. nana may have had much longer flippers than both pygmy right whales and gray whales, but it is not known how wide they were.

C a p e r e a m a r g i n a t a
68
Balaenids on the other hand have long and extremely wide flippers supported by a humerus that is about as long as the radius. Finally, humpback whales have the longest but also narrow flippers. The ratio of humerus / radius length is typical for rorquals, yet compared to body size the humerus
is extremely long, to facilitating the growth of such long flippers. The overall oblong-elliptical shape of the flipper in C. marginata deviates little in its aspect ratio between MM2959 and MM2904, whereas the ratio of body to flipper length changes significantly from 6.1 in the neonate MM2959 (1.75 m total length) to 8.0 in the juvenile MM2904 (2.75 m total length), and 11.5 in Hector’s specimen MM0036 (4.65 m total length) resulting in a very short but moderately narrow flipper. In mysticetes, flippers play an important role during feeding manoeuvres by counteracting drag-induced torque of the body during water and prey intake (Cooper et al., 2008). The narrow and elongated flippers in rorquals are suspected to be linked to high speed in lunge feeding (Goldbogen et al., 2006), whereas the broad flippers in balaenid whales are regarded as aiding manoeuvrability and counteracting pitch during feeding at low speed (Woodward et al., 2006). Although flipper proportions of the pygmy right whales are closer to those of rorquals, they are nonetheless also close to those of grey whales and thus midway between the more narrow flippers of rorquals and the broad flippers of balaenid whales. Intermediate flipper proportions may indicate a more generalised pattern tailored to a wide range of functions (together with other flipper and body characteristics often only sporadically explored in the literature; see also Chapter 4 Functional anatomy: Functional aspects of flipper shape). Grey whales, for example, are unique in their wide
range of prey selection and feeding methods. They are essentially bottom feeders that can also skim feed and are able to negotiate shallow lagoons yet migrate vast distances in open water (Pike, 1962; Rice and Wolman, 1971). Sightings of C. marginata also suggest pelagic habitat with possible seasonal inshore occurrence in shallow waters and lagoons, particularly of juveniles (Ross et al., 1975). Pygmy right whales might not undertake defined migrations like other baleen whales but sightings far offshore (Matsuoka et al., 1996; Kemper, 2002; Matsuoka et al., 2005) indicate that they may still travel long distances. Consequently, it would make sense if young whales, residing closer to shore, have longer flippers to aid manoeuvrability but adult

C a p e r e a m a r g i n a t a
69
animals, roaming offshore, have short flippers to facilitate higher speed (see also Fish, 2002). The rounded tip of the pygmy right whale flipper (Fig. 3a,c; see also Reeves et al., 2002 and Ross et. al, 1975 figs. 1-3) is distinctively different from the pointed tip of rorquals and grey whales. A possible explanation could be that short and moderately
narrow flippers, in combination with the body indent on the flanks to accommodate the flippers, would reduce drag, and a non-tapering flipper with blunt tip would provide more torque when executing tight turns or more lift when feeding at lower speeds. Observations of a pygmy right whale in a shallow lagoon confirmed extensive body flexing (albeit in contradiction to Buchholtz, 2011) and the ability to perform rapid starts and tight turns (Ross et al., 1975). Because C. marginata did not exceed speeds over 5 km/h, drag due to high speed may not be a major factor while executing manoeuvres in a lagoon and consequently tapered flippers may not be as crucial. While flippers were closely adducted during moderate and fast swimming, they were also extended up to 120º (subtended) during slow manoeuvres and surfacing (Ross et al., 1975). Flippers this far extended also reveal the large axillary skin fold (Fig. 3b). This additional surface area might act like the landing flaps on an airplane by providing increased root chord length of the flipper and thus augmenting additional lift when surfacing and feeding, or producing additional torque during manoeuvres (see also Chapter 4 Functional anatomy: Functional aspects of flipper shape). Additional consideration has also to be given to the large olecranon protruding deep into the axillary skin fold. It possibly not only supports the trailing edge of the skin fold like a strut: its associated muscles, namely the triceps and flexor carpi ulnaris, might also act like the wire stays of a biplane by tensioning the soft tissue in this area.
Scapula and forelimb bones
The overall length / height ratio of the scapula in MM2959 and MM2904 surpasses that of any extant baleen whale, the closest being the fin whale, and is matched by possibly only a few members of the cetothere family such as Piscobalaena nana (Fig. 17, 19l). While the scapula of MM2959 (Fig. 19j) approximates the uniquely elongated trapezoid contour of P. nana, in MM2904 (Fig. 19a) the scapula approaches the more generally observed fan-shaped contour of other mysticetes.

C a p e r e a m a r g i n a t a
70
In P. nana, the tripartite vertebral border is distinctly demarcated by a subtly convex dorso-craniad-facing anterior aspect and dorso-caudad-facing posterior aspect, separated by a comparatively straight and level midsection. Depending on the specimen, the cranial and caudal border form a shallow angle of 125º to 155º and merge into the neck of the scapula in a gentle and shallow curve (Table 2).
In MM2959 the anterior and posterior aspects of the vertebral border are not as steep and pronounced as in P. nana; the cranial and caudal border meet at a slightly steeper angle of about 120º in MM2959 and 130º in MM2904.
Fig. 17. Ratio of scapula length to scapula height (based on Benke, 19931; Bouetel and de Muizon, 20062; Andrews, 19143) Left column, colour key: Green - Balaenidae; Dark grey - Eschrichtiidae3; Blue - Balaenopteridae; Orange - Caperea marginata; Ochre - Piscobalaena nana; Right column, light grey - Total length of the skeleton (1) Balaenopteridae and Balaenidae, (2) P. nana, (3) Eschrichtiidae.

C a p e r e a m a r g i n a t a
71
MM2959 and P. nana also agree in following characteristics: - The anterior aspect of the vertebral border is slightly (P. nana) to clearly (MM2959)
shorter than the caudal aspect - The coracoid process is slender and finger-shaped - The acromion process is rectangular (as far as could be ascertained in P. nana),
and its axis parallel to the caudal border of the scapula, albeit in P. nana at a more dorsal level, while in MM2959 the ventral border of the acromion is in line with the scapular caudal border
Further congruence between the two, and some rorquals, is - The ratio of humerus / radius length is roughly 4:6
P. nana differs in some features from MM2959: - The angle between the axis of the scapula and the anterior or posterior aspect of
the vertebral border is greater than in MM2959 - The central aspect of the vertebral border is slightly convex as opposed to MM2959
where it is concave - The neck of the scapula is indistinct whereas in MM2959 it is more pronounced - The angle between the glenoid border and axis of the scapula appears much more
shallow than in MM2959 - The humeral head is larger (wider than the distal epiphysis) than in MM2959
(narrower than the distal epiphysis) - A prominent deltopectoral crest is indistinct in MM2959 - In P. nana the ulnar facet extends onto the postero-distal edge of the humeral
diaphysis opposing the olecranon of the ulna, whereas in MM2959 this facet is straight and extends only very slightly onto the posterior edge of the humerus
- The proximal third of the radial diaphysis is in P. nana markedly curved whereas
in MM2959 the diaphysis is straight - The ulna of P. nana is fairly robust (smallest diameter almost as wide as smallest
diameter of radius) as opposed to a relatively delicate ulna in MM2959 (the smallest diameter of the ulna is only half as wide as the smallest diameter of the radius).
Comparing the angular scapular outline of MM2959, a neonate male, with the fan-shaped scapula of MM2904, a young female (Fig. 18, 19a,j), first impressions might suggest the two specimens represent different stages in ontogenetic development or

C a p e r e a m a r g i n a t a
72
sexual dimorphism. Sexual dimorphism is contradicted by a number of other specimens, demonstratively Hector’s subadult female MM0036 (Fig.18, 19i). In outline fairly angular, the scapula of this specimen also resembles MM2959, albeit with a more gently curved posterior scapular neck (Hector, 1875 pl. XVII - fig. 10). Contrary to MM2959, though, in MM0036 it is the posterior aspect that is shorter and steeper and
the anterior aspect that is longer and ascending at a shallower angle. Similar to MM2959, the central section of the vertebral border is slightly concave. The acromion process is tapered and not square as in MM2959. Despite these small differences, both MM0036 and MM2959 are more similar than either is to MM2904.
Table 2. Comparison of the forelimb in Piscobalaena nana and Caperea marginata MM2959
Character Piscobalaena nana MM2959
Ratio of scapula length to height 1.87-2.12 1.76-2.04 Central aspect of vertebral border slightly rounded slightly depressed Length of anterior aspect to length of posterior aspect of vertebral border
slightly shorter clearly shorter
Angle between anterior or posterior aspect of vertebral border and axis of scapula
~ 60º-70º ~ 40º-50º
Orientation of acromion process to caudal border
parallel, dorsal parallel, in line
Coracoid process slender, finger-shaped slender, finger-shaped Angle between cranial and caudal border 125º - 155º 120º Neck of scapula indistinct more pronounced Angle between glenoid border and axis of scapula
~ 10º ~ 20º-25º
Width of humeral head to distal epiphysis larger smaller Deltopectoral crest prominent indistinct Ratio of length of humerus to radius 4 : 6 4 : 6 Ulnar facet of humeral diaphysis extended, curved restricted, straight Radius shape curved straight Smallest diameter of radius to ulna 1 : 1 2 : 1
The rounded vertebral border of MM2904, on the other hand, closely matches that of a subadult specimen held at the Otago Museum (Fig. 18, 19b - OMNZ VT 227), but
here the acromion and coracoid process are long, slender and tapered. Both
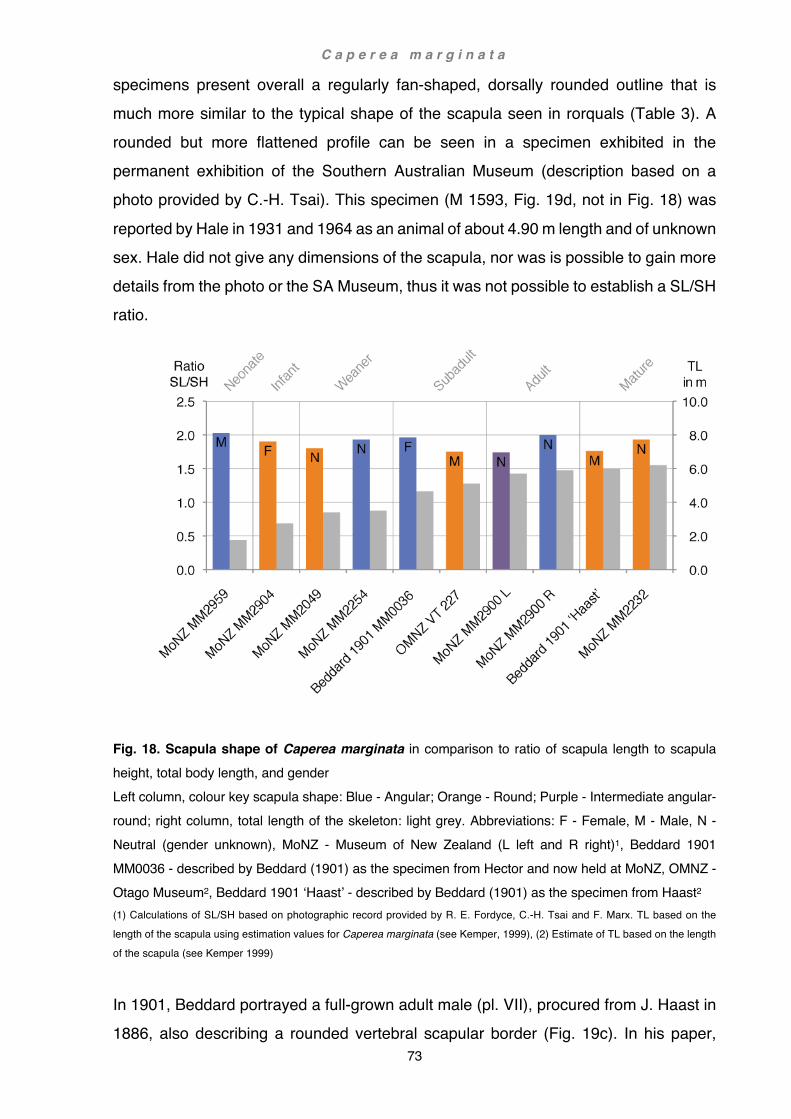
C a p e r e a m a r g i n a t a
73
specimens present overall a regularly fan-shaped, dorsally rounded outline that is much more similar to the typical shape of the scapula seen in rorquals (Table 3). A rounded but more flattened profile can be seen in a specimen exhibited in the permanent exhibition of the Southern Australian Museum (description based on a photo provided by C.-H. Tsai). This specimen (M 1593, Fig. 19d, not in Fig. 18) was
reported by Hale in 1931 and 1964 as an animal of about 4.90 m length and of unknown sex. Hale did not give any dimensions of the scapula, nor was is possible to gain more details from the photo or the SA Museum, thus it was not possible to establish a SL/SH ratio.
Fig. 18. Scapula shape of Caperea marginata in comparison to ratio of scapula length to scapula height, total body length, and gender Left column, colour key scapula shape: Blue - Angular; Orange - Round; Purple - Intermediate angular-round; right column, total length of the skeleton: light grey. Abbreviations: F - Female, M - Male, N - Neutral (gender unknown), MoNZ - Museum of New Zealand (L left and R right)1, Beddard 1901 MM0036 - described by Beddard (1901) as the specimen from Hector and now held at MoNZ, OMNZ - Otago Museum2, Beddard 1901 ‘Haast’ - described by Beddard (1901) as the specimen from Haast2 (1) Calculations of SL/SH based on photographic record provided by R. E. Fordyce, C.-H. Tsai and F. Marx. TL based on the
length of the scapula using estimation values for Caperea marginata (see Kemper, 1999), (2) Estimate of TL based on the length
of the scapula (see Kemper 1999)
In 1901, Beddard portrayed a full-grown adult male (pl. VII), procured from J. Haast in 1886, also describing a rounded vertebral scapular border (Fig. 19c). In his paper,

C a p e r e a m a r g i n a t a
74
Beddard points out that the scapula of the adult male shows a lower SL/SH ratio (1.76, Fig. 18 - Beddard 1901 ‘Haast’) than Hector’s subadult female (2.0, Fig. 18 - MM0036 Hector) and argued that the higher ratio in the younger animal was a reflection of incomplete ossification of the vertebral border. He did not comment on the difference in shape. An angular profile (i.e. a flattened or dipping midsection of the vertebral
border) does not attain the same height as a round profile of the same length and consequently has a higher length / height ratio. The occurrence of similar high proportions over a wide range of ages, from very young (MM2959) to adult specimens (Fig. 18, 19k - MoNZ MM2900 R), also speaks against that argument. A similar age distribution can also be found in specimens that show a lower ratio. This does not exclude a decreasing SL/SH ratio in general during the ontogenetic development of an individual, but rather demonstrates a wide intraspecific variability, separated roughly into two types that seem to occur in both sexes (e.g. MM2959 - angular, male vs. Hector’s specimen MM0036 - angular, female or MM2904 - round, female vs. Haast’s specimen - round, male).
Table 3. Categories of scapular shape in baleen whales Species Type Age Gender Reference
Caperea marginata trapezoid neonate male MM2959 Piscobalaena nana trapezoid adult female Bouetel and de Muizon 2006, SAS 1617 Piscobalaena nana trapezoid adult male Bouetel and de Muizon, SAS 892 Caperea marginata fan-shaped juvenile female MM2904 Balaenopteridae fan-shaped - - Benke 1993, figs. 16-20 Eschrichtiidae fan-shaped to
triangular - - Andrews 1914, fig. 19
Balaenidae triangular - - Benke 1993, figs. 12-14
Although the sample size represented here is too small to ascertain clear correlations, potential aspects might nonetheless be considered. Apart from independent individual (ecophenotypic) variability, the two scapula types could be regarded in view of ontogenetic development or a combination of gender and age. Both scapula types (i.e. angular vs. round vertebral border) occur in younger and older specimens alike. The young female MM2904 (round) is opposed by Hector’s subadult female (MM0036,
angular). The young male MM2959 (angular) is opposed by Haast’s fully grown male (round). This might imply that growth patterns in females and males would progress in

C a p e r e a m a r g i n a t a
75
opposite directions (in females from round to angular and in males from angular to round). Furthermore, the cartilaginous border above the anterior and posterior part of the vertebral border is very broad, yet very narrow above the mid-section, effectively elongating and levelling the contour of the rounded border or enhancing that of the angular border. In consequence, partial or complete ossification of this cartilage would
not result in a rounded vertebral border, a shape that is seen in Haast’s fully grown specimen. Of interest is hereby also the occurrence of intermediate profile types (Table 4). While MM2904 has a distinctive fan-shaped contour, in MM2232 (Fig. 18, 19e) the vertebral border is overall less curved. In MM2049 (fig. 18, 19f) the posterior part of the vertebral border is rounded as in MM2959, but the midsection is almost straight and the anterior part only little rounded. In MM2254 (Fig. 18, 19h) on the other hand, the anterior portion is flattened and the curvature little defined, the midsection is slightly dipping and the posterior part markedly angular. However, the most perplexing profile can be seen in an adult specimen of unknown sex from Stewart Island held at the Museum of New Zealand (MM2900). The right scapula can be readily identified as the angular type by the typically slanted anterior and posterior aspect; the middle part is straight but below the highest point of the cranial and caudal angle (Fig. 18, 19k - MoNZ MM2900 R). In contrast, the left scapula of this specimen appears to be a puzzling combination of both types: the anterior part distinctively rounded and posterior prominently angular; the convex cranial portion of the vertebral border rises high above the longitudinal axis of the scapula and merges smoothly into the anterior part of the midsection; the posterior part of the midsection subsequently dips strongly ventrad amplifying the sharp angle of the caudal part, giving the vertebral border on the whole a rather distorted sinuous
outline (Fig. 18, 19g - MoNZ MM2900 L). These two scapulae, however, have been gnawed on by rats, which could to some extent explain their different profile. Yet, there are other strong discrepancies, such as a different angle of the neck leading into the caudal border of the scapula, a dissimilar acromion (left kinked and tapered, right much straighter and square) and coracoid process (left more slender but longer), the left scapula slightly shorter than the right scapula, and an altogether incongruent silhouette. Such strong dissimilarities between the right- and left-hand side are rare in cetaceans (e.g. Galatius, 2005, 2006; Gómez-Campos et al., 2010) and the question thus arises if the two shoulder blades belong to the same animal, and if so, are the

C a p e r e a m a r g i n a t a
76
dissimilarities potentially either pathological or a response to differential mechanical stresses?
Fig. 19. Scapulae of Caperea marginata and Piscobalaena nana (a) MM2959 (this study), (b) VT227 (OM), (c) modified from Beddard, 1901, plate VII, (d) M1593 (SAM), (e) MM2232 (MoNZ), (f) MM2049 (MoNZ), (g) MM2900 (MoNZ), (h) MM2254 (MoNZ), (i) MM0036, modified from Hector, 1875, plate XVII, fig. 10, (j) MM2959 (this study), (k) MM2900 (MoNZ), (l) SAS892, modified from Bouetel and de Muizon, 2006, fig. 26. Profiles: (a-f) rounded, (g) intermediate, (h-l) angular; (a,d,e,g,i,l) left scapula, (b,c,f,h,j,k) right scapula Abbreviations: (MoNZ) Museum of New Zealand, (OM) Otago Museum, (SAM) South Australian Museum. Scale bars (c,i) estimate
from measurements given by Beddard (1901), (d) measurements not available.
Photos (b,f) F. Marx, (d,f) C. H. Tsai, (e,g,h,k) R. E. Fordyce

C a p e r e a m a r g i n a t a
77
Table 4. Categories of scapular shape in Caperea marginata Specimen Profile Collection Locality Storage Location
MM2904 (infant female) Marked rounded Glink’s Gully, North Is. Museum of New Zealand VT227 Rounded Patterson Inlet, Stewart Is. Otago Museum Beddard 1901 Haast Rounded Not specified Not known M1593 Flat rounded Kangaroo Is., Australia South Australian Museum MM2232 Flat rounded Wharewara Bay, North Is. Museum of New Zealand MM2049 Flat rounded Warkworth, North Is. Museum of New Zealand MM2900 left scapula Round-angular Patterson Inlet, Stewart Is. Museum of New Zealand MM2254 Flat angular Farewell Spit, South Is. Museum of New Zealand MM0036 Hector Angular North end Stewart Island Museum of New Zealand MM2959 (neonate male) Angular Karikari Peninsula, North Is. Museum of New Zealand MM2900 right scapula Marked angular Patterson Inlet, Stewart Is. Museum of New Zealand
In general, the most notable differences between individual animals, e.g. the shape of the acromion (rectangular vs. tapered), are likely to be related to individual variability, whereas other discrepancies, often more subtle in nature, are much harder to pinpoint (mainly due to the small sample sizes in this study); e.g. in some odontocete species the angle of the curvature between the scapular neck and the cranial or caudal border (shallow vs. acute) appears to be age-dependent (pers. observation). Additionally, gender differences may also play a role in some traits (e.g. size or robustness). Despite some evident variations, the scapula in C. marginata also shows relatively constant features: high length / width ratio of at least 1.75; a small, short supraspinous fossa; a broad, rectangular or tapered acromion; a small and slender coracoid and the
slanting border of the glenoid cavity. The elongation of the bony shoulder blade is substantially enhanced by the cartilaginous vertebral border and is probably, among extant mysticetes, the most extreme in the pygmy right whale (see also Hector, 1875; Beddard, 1901, Benke, 1993 and Chapter 4 Functional anatomy: Functional aspects of flipper shape, section Neocetes). Greater length of the cetacean scapula has been associated with increased muscular strength at the expense of agility (Benke, 1993). The extreme elongation of the shoulder blade in C. marginata appears therefore peculiar, since in the pygmy right whale the flippers are small and seemingly not exposed to the high forces that have been observed in the much faster swimming rorquals. Additionally, C. marginata has

C a p e r e a m a r g i n a t a
78
been described as keeping the flippers closely adducted to the body when swimming faster (Ross et al., 1975). However, it is not known how fast C. marginata is actually capable of swimming or executing rapid manoeuvres. One explanation may lie in the low ratio of humerus length to radius length, a ratio that is quite similar to that seen in rorquals (Fig. 16). A short humerus permits fast reposition
of the flipper but also necessitates strong musculature to compensate for the decreased level of leverage momentum. Elongation of the shoulder blade would therefore not just facilitate more room for muscles but also enhanced leverage (see also Smith and Savage, 1956 and Chapter 4 Functional anatomy: Functional aspects of flipper shapes). Cranial elongation of the blade would allow the anterior fibres of the deltoideus a stronger leverage forward while the posterior fibres would maintain upwards pull (Klima et al., 1980; Benke, 1993). Caudal elongation would also allow a better leverage of the teres major (Smith and Savage, 1956) counteracting the deltoideus and thus, in concert with other muscles, improved stabilisation of the abducted and extended flipper. Strengthening of the peripheral muscles, such as the rhomboideus and serratus, might also enhance flipper movements by rotating the scapula more forcefully. More muscle power may also be needed to counteract additional loading incurred by the exposed axial skin fold when the flippers are extended. Another peculiarity seems to be the angle of the glenoid border relative to the longitudinal axis of the scapula, a trait that is strongly associated with the alignment of the scapula on the body: while the longitudinal axis of the scapula descends ventro-caudad towards the longitudinal axis of the body. The border of the glenoid fossa rises by about the same amount towards the caudal angle of the scapula, thus positioning
the glenoid cavity facing the longitudinal axis of the body at a right angle (see also Chapter 4 Anatomy: Positioning of the forelimb, Fig. 2 and Osteology, Fig. 12b,c). This position is a necessity, since an orientation of the longitudinal axis of the scapula parallel to the longitudinal axis of the body would position the glenoid fossa facing backwards and in consequence rotate the forelimb caudad. Such an arrangement would reduce the already limited range of possible extension forward even further. Functional implications arising from the tilt of the scapula are not as apparent in cetaceans as they are in terrestrial mammals. In mammalian quadrupeds the position and orientation of the scapula on the torso are highly influenced by ecological and

C a p e r e a m a r g i n a t a
79
physical constraints, the latter dominated by the need to support the body against gravitation and overcome inertia in locomotion, particularly in large and heavy animals (Gregory, 1912; Smith and Savage, 1956). This is no longer the case in cetaceans. Unchained from the limits imposed by gravitation, cetaceans developed a unique ‘stance’ where the forelimb is no longer carried vertically under the body to support the
weight but is either retracted to the body or subtended at an angle (Benke, 1993). As a result, muscles that operate in quadrupeds predominantly in a parasagital plane counteracting gravity and inertia, facilitate in cetaceans movement in all directions to accommodate three-dimensional movement of the body in water. Position and orientation of the shoulder and forelimb in cetaceans are thus a mixture of phylogenetic inheritance and new adaptations (Thewissen and Fish, 1997; Thewissen and Bajpai, 2001; Houssaye et al., 2015; see also Chapter 4 Functional anatomy: Evolution of the forelimb). Possible functional explanations of the oblique orientation of the scapula may be to facilitate a position of the flippers down and forward or improve muscle efficiency. A mere vertical shift of the scapula ventrad might diminish effectiveness in some muscles associated with the head, throat and ribs. A lower caudal aspect of the scapula on the other hand would effect forward and downward shift of the shoulder joint and effectively shorten some of the more peripheral muscles, e.g. the mastohumeralis arising craniad or the teres major between adducted humerus and posterior portion of the scapula. A shortening of those muscles alone would have little effect unless other factors such as improved leverage angle would also come into play (see also Chapter 4 Functional Anatomy: Evolution of the forelimb). The extreme caudal elongation of the scapula allows for a large and powerful teres
major. Together with the latissimus dorsi this muscle pulls the flipper backwards and inwards towards the body. Once the flipper is retracted, the humerus is placed directly below teres major. Since muscles in situ remain partly stretched and thus act as ligaments (Gregory, 1912), very little energy would have to be expended by the whale to maintain this position. The orientation of the scapula may thus also be linked to energy-efficient muscle arrangements holding the flipper in place and the drag reducing position of the associated indent running along the flank. Additionally, repositioning of the flippers down and forward would also augment body roll and steering performance (Gerstner, 1999; Fish, 2002). Powerful use of the flippers

C a p e r e a m a r g i n a t a
80
and nimbleness are crucial components of steering and undoubtedly an important factor for a whale that has been described by Ross et al (1975) for its agility and ability to execute rapid starts and turns in a shallow lagoon.
Musculature
In general, the shoulder and forelimb musculature of C. marginata is fairly consistent with patterns described in rorquals by Schulte (1916) and Benke (1993) for the sei whale (Balaenoptera borealis), and Carte and Macalister (1868) and Perrin (1870) for the minke whale (Balaenoptera acutorostrata). There are no further detailed accounts of the shoulder musculature in baleen whales, although a somewhat similar arrangement of flipper musculature has been recorded for the humpback and blue whale (Megaptera novaeangliae, Balaenoptera musculus) by Struthers (1888, 1889) and Kükenthal (1921), and for the Bowhead whale (Balaena mysticetus) by Struthers (1878) and Kükenthal (1922). In the following pages, references to B. borealis and B. acutorostrata will refer to the
detailed accounts of Schulte (1916) and Carte and Macalister (1868) unless stated otherwise. Some muscles correspond well in all three species, others differ substantially, and some are unique for the pygmy right whale (Table 5). There are several muscles corresponding to each other equally in all three species: rhomboideus vertebralis, subscapularis, pectoralis major, coracobrachialis, omohyoideus and serratus anterior - posterior division (called ‘serratus anticus’ by Schulte (1916) and ‘serratus magnus’ by Carte and Macalister (1886)). In C. marginata, only one muscle corresponds better with that of B. acutorostrata: Carte and Macalister (1868) described a muscle in this species as ‘arising from the transverse process of the last cervical’ which they called ‘levator anguli scapulae’, yet a similar muscle in the fin whale was described by Schulte (1916) as ‘arising from the costal process of the second cervical vertebra’ which is similarly referred to as ‘levator scapulae’. Strickler (1978) argued that the true levator scapulae arises from the anterior-most cervical vertebrae, while the serratus anterior - anterior division arises either from the cranial-most ribs or caudal-most cervical vertebrae. Strickler concluded

C a p e r e a m a r g i n a t a
81
that Carte and Macalister’s ‘levator anguli scapulae’ corresponds to the serratus anterior - anterior division but Schulte’s ‘levator scapulae’, described as innervated from C2, is consistent with the true levator scapulae. Schulte’s levator scapulae, inserting onto the medial surface of the cranial angle, is thus quite different from that reported for most other whales (odontocetes) where it usually, if present, inserts
laterally (e.g. Howell, 1930b; Smith et al., 1976; Strickler, 1978; Klima et al., 1980) and consequently should not be mistaken for the serratus anterior - anterior division which typically inserts at this site. However, although in C. marginata the insertion of the serratus anterior - anterior division is identical with that of the minke whale, its origin from the fused first and second ribs (sensu Buchholtz, 2011) differs from that of the latter. Some muscles in C. marginata are generally congruent with those of B. borealis, such as the latissimus dorsi, reported by Schulte (1916) as emerging from the aponeurosis covering the dorsal extensor muscles with slips arising from the 7th to 9th ribs, and running parallel below the caudal border of the scapula. Quite a different arrangement was reported by Benke (1993) for the same species and by Carte and Macalister (1868) for B. acutorostrata, where it is described as arising from the spines of the thoracic and anterior lumbar vertebrae. The arrangement in C. marginata is very similar to Schulte’s account of B. borealis, the muscle arising mostly from the region of the 6th to 9th ribs with a few fibres coming from the 10th. In MM2959 the latissimus dorsi passes below the scapula to its point of insertion comparable to Schulte’s depiction in pl. XLIV - fig. 2. In MM2904 part of the scapular caudal angle and associated muscles are concealed by the passing latissimus dorsi (Fig. 10). Origin and insertion are identical in both MM2959 and MM2904.
In most odontocetes, the teres major originates at the caudal border and adjacent deep surface of the scapula (Strickler, 1978). This is also the case in the minke whale as reported by Carte and Macalister (1868). In the sei whale, however, the teres major encroaches onto the caudal lateral surface of the scapula, more or less displacing the infraspinatus cranially, an arrangement that is also found in the pygmy right whale. Although Perrin’s (1870) descriptions agree with those of Carte and Macalister (1868), in their depiction, figure 1 - Tr.Mg., the arrangement of the teres major corresponds better with that found in the sei whale and pygmy right whale.

C a p e r e a m a r g i n a t a
82
Table 5: Comparative forelimb myology in three baleen whales Muscle Caperea marginata Balaenoptera
borealis(1,2) Balaenoptera
acutorostrata(3,4) Levator scapulae absent present absent Serratus anterior - anterior division
origin first rib absent origin last cervical vertebra as ‘levator anguli scapulae’(3)
Serratus anterior - posterior division
present present as ‘serratus anticus’(1)
present as ‘serratus magnus’(3)
Rhomboideus vertebralis
present present present
Latissimus dorsi origin ribs origin ribs(1)
or vertebrae(2) origin vertebrae
Teres major origin caudal lateral surface of scapula
origin caudal lateral surface of scapula
origin caudal border of scapula
Subscapularis present present present Deltoideus secondary belly secondary belly(1)
or single belly(2) single belly
Teres minor absent present as ‘subdeltoid’(1) or
‘supraspinatus’(2)
absent
Triceps two heads two heads three heads Supraspinatus origin supraspinous
fossa origin supraspinous
fossa origin cranial border of
scapula Infraspinatus origin muscle shape
irregular origin muscle shape
trapezoid origin muscle shape
deltoid Pectoralis major present present present Coracobrachialis present present two heads(4) Omohyoideus present present present Trapezius absent present absent Mastohumeralis origin exoccipital origin paramastoid origin squamosal Extensor digitorum communis
present present present
Flexor digitorum radialis
merged with flexor digitorum communis
merged with fdc (1) or discrete(2)
discrete(3) or merged(4)
Flexor digitorum communis
merged flexor digitorum radialis
merged with fdr (1)
or discrete(2) discrete(3)
or merged(4) Flexor carpi ulnaris origin base of
olecranon, insertion pisiform
origin base of olecranon, insertion
pisiform
origin tip of olecranon, insertion metacarpal 5(3)
or origin base of olecranon,
insertion distal ulna(4) Flexor palmaris longus
absent absent present(3)
Flexor sublimis digitorum
absent absent present(4)
(1) Schulte, 1916, (2) Benke, 1993, (3) Carte and Macalister, 1868, (4) Perrin, 1870 Text within table refers to either main differences / similarities between species or alternate nomenclature of muscles, similar muscular arrangements between species are highlighted in bold letters

C a p e r e a m a r g i n a t a
83
While there is little variability of the deltoideus within the three species, in the pygmy right whale there is a tiny belly arising from the distal cranial border of the humerus and inserting onto the proximal cranial border of the radius just below the epiphysis. There is no description of such a muscle in the literature and it is possible that this little belly is not part of the deltoideus but a vestigial remnant of another muscle. There are
potentially two muscles to consider: the biceps brachii and the brachialis. In its primitive condition the brachialis has two heads, one originating from the lateral neck of the humerus inserting medially onto the radius, the other originating medial and distal to the deltoid insertion and inserting medially onto the ulna (Howell, 1937). Based on osteological features of the long bones, Uhen (2004) also inferred the presence of this muscle in Dorudon atrox, a medium-sized archaeocete from the Late Eocene, about 40 my old. He surmised that the brachialis arose from the lateral, proximal side of the humerus and inserted onto the cranial, proximal border of the radius. The second muscle, the biceps, has in its basic form also two heads, one arising from the coracoid and inserting onto the radius, the other arising from the supraglenoid border and inserting onto the ulna (Howell, 1937). Uhen proposed that in D. atrox this muscle originated from the supraglenoid border and inserted onto the radial tuberosity, a ridge just medial and distal to the proximal epiphysis. Accordingly, the closest contender in C. marginata would ostensibly be Uhen’s brachialis as seen in D. atrox, since the muscle in question in C. marginata also arises from the humerus and inserts onto the cranial, proximal border of the radius. However, attachment locations of muscles can be quite variable (Howell, 1937) and homologies without assessment of associated innervation uncertain (Strickler, 1978). Alternatively, it is also conceivable that instead of a vestigial muscle the expansion of the deltoideus in rorquals and C. marginata, by elongation of the scapula, also necessitated extension at the insertion site. Such a
condition has been reported for B. borealis. Schulte (1916) described the tendon of the deltoideus as inserting onto the whole preaxial border of the humerus, also extending onto the elbow capsule and radius. Plate XLVI - fig. 6 also shows the insertion of the deltoideus as well advanced onto the proximal half of the cranial and preaxial lateral surface of the radius. Furthermore, he reported ligamentous bands representative of both heads of the biceps crossing obliquely across the coracobrachialis and latissimus-teres insertion, but he did not associate the fibers distad of the deltoideus with vestiges of the brachialis. Overall, a distinct slip arising from an extended deltoideus insertion

C a p e r e a m a r g i n a t a
84
similar to that of the sei whale might be still more plausible than a vestigial brachialis which is otherwise regarded as absent in extant cetaceans (Cooper et al., 2007b). Carte and Macalister (1868) described in B. acutorostrata a three-headed triceps (long head, internal head and external head) whereas Perrin (1870) reported only two heads
(long head and external head) for the same species, as did Schulte (1916) and Benke (1993) for B. borealis, a configuration that is also found in C. marginata. The supraspinatus in B. acutorostrata, as described by Carte and Macalister (1866), appears split, with one part arising from the anterior border of the cranial scapular angle and the other superficially from the ventral aspect of the acromion. In B. borealis, the organisation of this muscle corresponds much more with that observed by Perrin (1870) in B. acutorostrata and most odontocetes. Here the supraspinatus arises as one consolidated muscle from the supraspinous fossa, the medial surface of the acromion including the intermuscular fascia spanning between it and the cranial angle and the lateral surface of the base and deep tip of the coracoid (Schulte, 1916). This is also the case in C. marginata, and while in B. borealis the supraspinous fossa is comparatively large, it is considerably smaller in the pygmy right whale. In the neonate MM2959 the widest part of supraspinous fossa is about 1/25th of the length of the vertebral border, whereas in the older MM2904 it is only about 1/35th. Furthermore, in MM2904 the supraspinous fossa does not reach the vertebral border but is confined to the lower 3/4 of the cranial border. In general, the arrangements of the digital extensor and all flexors are similar in all three whales; however, particular accounts vary in detail: Schulte (1916) described the
extensor in B. borealis as arising from the lateral surface of the elbow joint capsule, interosseus membrane and adjacent surfaces of radius and ulna, at this point moderately displaced towards the radius, an arrangement that is congruent with that of C. marginata. A similarly large origin, but with slightly different insertion, has also been described by Perrin (1870) for B. acutorostrata. On the other hand, a more reduced extensor muscle, which is largely confined to the interosseus space between the radius and ulna, has been described by Carte and Macalister (1868) for the same species, and by Benke (1993) for B. borealis. The insertion is almost identical in all three species, namely by separation of the tendons onto all four digits in the area of

C a p e r e a m a r g i n a t a
85
the carpals, with adaxial displacement at the height of the metacarpals of digits 2 and 5, particularly in B. acutorostrata, and to some degree also in C. marginata. Only Perrin’s (1870) account differs somewhat in that respect: describing an additional slip of the ‘index’ finger onto the metacarpal. However, in fig. 1 the cranial-most tendon is numbered ‘IV’, followed by number ‘III’ and ‘II’ further caudad, and the two caudal-most
slips of the ‘index’ finger (not numbered). It is thus evident that in Perrin’s paper the ‘index’ finger is digit 5 and finger ‘IV’ is actually digit 2. Hence, it is the tendon of digit 5 that has an additional slip, and the tendon of digit 2 that differs from other accounts by dividing approximately at mid-radial height instead of the carpals.
Extensor digitorum communis Origin Insertion
C. marginata broad split at carpals, slightly adaxial, digits 2-5 B. acutorostrata (Schulte, 1916) broad split at carpals, slightly adaxial, digits 2-5 B. borealis (Benke, 1993) reduced split at carpals, slightly adaxial, digits 2-5 B. acutorostrata (Carte and
Macalister, 1868) reduced split at carpals, digits 2-5, markedly adaxial
B. acutorostrata (Perrin, 1870) very broad split of digit 2 at mid-radius, split of digits 3-5 at carpals, tendon of digit 5 with additional slip
The flexor digitorum radialis arises in all three species from the proximal, postaxial diaphysis of the radius, interosseus membrane and fascia in common with the flexor digitorum communis. In C. marginata and B. borealis, as reported by Schulte (1916), the tendon of this muscle joins the tendon of the flexor digitorum communis in the wrist
area before leading onto the second digit. The same situation has also been reported by Perrin (1870) for B. acutorostrata. A slightly different arrangement has been described by Carte and Macalister(1868) for the same species, and by Benke (1993) for B. borealis: here the tendon of the flexor digitorum radialis remains separated and terminates inserting onto the proximal phalanx in B. acutorostrata, or the entire length of the second digit in B. borealis.

C a p e r e a m a r g i n a t a
86
Flexor digitorum radialis Origin Insertion
C. marginata prox. radius merged with fdc onto digit 2 B. acutorostrata (Schulte, 1916) prox. radius merged with fdc onto digit 2 B. borealis (Benke, 1993) prox. radius separated from fdc onto phalanx 1 of digit 2 B. acutorostrata (Carte and
Macalister, 1868) prox. radius separated from fdc onto digit 2
B. acutorostrata (Perrin, 1870) prox. radius merged with fdc onto digit 2 Abbreviations: prox. proximal, fdc flexor digitorum communis
The flexor digitorum communis (or ulnaris) of C. marginata is, in origin and insertion, somewhat different from that described in Schulte’s (1916) or Benke’s (1993) account of B. borealis. In the sei whale, this muscle arises from the distal part of the humerus, the capsule of the elbow joint, the proximal shaft of the ulna, the interosseus ligament, and the intermuscular fascia adjacent to the flexor digitorum radialis but is clearly separated from the flexor carpi ulnaris. In C. marginata the muscle is broader and arises slightly more distally from the proximal surface of the ulna and bony base of the olecranon but shares the intermuscular fascia of both the flexores digitorum radialis and carpi ulnaris. The same arrangement has also been described in Carte and Macalister’s (1868) account of B. acutorostrata. Perrin’s (1870) report and figure 2 are in this respect somewhat different: there the flexor digitorum communis also borders on the flexor carpi ulnaris but is cranially intercepted by the flexor digitorum sublimis, separating it from the flexor digitorum radialis. No flexor digitorum sublimis has been
reported by the other authors, nor noted in C. marginata. The insertion pattern of this muscle is also slightly different from report to report. The most similar arrangement to C. marginata is that reported by Schulte in B. borealis, where the tendon of digit 2 is joined by the flexor digitorum radialis. However, in B. borealis the undivided tendon of the flexor digitorum communis largely remains on the postaxial side, splitting just proximal to the carpals into four perfectly aligned tendons following the four digits. This causes the tendon of digit 2 to turn sharply craniad while the tendon of digit 4 remains relatively straight. In C. marginata the position of the undivided tendon is more axial, and even though it splits further distad in the wrist area, the split is more symmetrical. Consequently, the tendons of digits 2 and 4 both separate in a gentle curve, initially following a more adaxial path over the proximal digits. Perrin’s description of B. acutorostrata also largely corresponds with that of Schulte, with the distinction of the

C a p e r e a m a r g i n a t a
87
additional flexor digitorum sublimis splitting into two branches, the cranial branch also fusing with the flexor digitorum communis. On the other hand, in Benke’s (1993) report of B. borealis and Carte and Macalister’s (1868) report of B. acutorostrata, the flexor digitorum radialis remains separated from the flexor digitorum communis. In Benke’s B. borealis, the split of the flexor digitorum communis appears symmetrical, and only
three tendons insert perfectly aligned along digits 3 to 5. In contrast, Carte and Macalister reported four tendons. The tendon of digit 2 is strongly adaxially displaced and exclusively inserts onto the terminal phalanx (with a slip to the intermediate phalanx), the proximal phalanx being occupied by the separate tendon of the flexor digitorum radialis. On digit 3 the cranial axial tendon appears displaced to a much lesser degree, namely from the metacarpal by the preaxial tendon of digit 2, and inserts onto all phalanges. The tendon of digit 4 is perfectly aligned along the entire digit. Similar to digit 3, the tendon of digit 5 is also displaced from the metacarpal bone, in this case by the tendon of the flexor carpi ulnaris and palmaris longus, these two tendons inserting onto metacarpal 5 instead. No palmaris longus has been reported by the other authors or noted in C. marginata.
Flexor digitorum communis Origin Insertion
C. marginata adjacent to fcu merged with fdr, symmetric B. acutorostrata (Schulte, 1916) distant from fdu merged with fdr, asymmetric B. borealis (Benke, 1993) distant from fdu separated from fdr, 3 tendons
onto digits 3-5, symmetric B. acutorostrata (Carte and
Macalister, 1868) adjacent to fcu separated from fdr, 4 tendons
onto digits 2-5, symmetric B. acutorostrata (Perrin, 1870) adjacent to fcu but separated
by fds from fdr merged with fdr + fds,
asymmetric Abbreviations: fcu flexor carpi ulnaris, fdr flexor digitorum radialis, fds flexor digitorum sublimis
While the origin of the flexor carpi ulnaris in C. marginata is more or less similar to that reported by Benke (1993) in B. borealis, and Perrin (1870) and Carte and Macalister (1868) in B. acutorostrata, the insertion corresponds better with that of Schulte’s account in B. borealis. Benke, Perrin, and Carte and Macalister describe the origin of this muscle as arising at the base of the olecranon and in B. acutorostrata as closely adjacent to the flexor digitorum communis. In B. borealis, although widely separated from the flexor digitorum communis, the muscle is also reported by Benke as arising at the base of the olecranon, whereas in Schulte’s report it arises from the

C a p e r e a m a r g i n a t a
88
distal border near the tip of the olecranon. Similarly to C. marginata, insertion of the flexor carpi ulnaris is also onto the pisiform in Schulte’s account of B. borealis but is reported by Benke to insert onto the connective tissue of the carpus. In B. acutorostrata Carte and Macalister reported the tendon as inserting onto metacarpal 5, here deflecting the postaxial slip of the flexor digitorum communis away from the metacarpal
further distad onto the phalanges. Perrin on the other hand, placed the insertion of the tendon onto the distal end of the ulna.
Flexor carpi ulnaris Origin Insertion
C. marginata adjacent to fdc, base of ole pisiform B. acutorostrata (Schulte, 1916) distant from fdc, tip of ole pisiform B. borealis (Benke, 1993) distant from fdc, base of ole connective tissue of carpus B. acutorostrata (Carte and
Macalister, 1868) adjacent to fdc, base of ole metacarpal 5
B. acutorostrata (Perrin, 1870) adjacent to fdc, base of ole distal end of ulna Abbreviations: fdc flexor digitorum communis, ole olecranon
C. marginata also presents divergent and in some cases quite unique arrangements of muscles. For instance, while the origin of the infraspinatus is reported to be similar in most cetaceans (Strickler, 1978), there can be considerable differences in profile and extent of this muscle. In all three whales, it is interposed between the deltoideus in front and the teres major behind. In B. acutorostrata, as reported by Carte and Macalister (1868), the infraspinatus is constrained anteriorly by the deltoideus and
posteriorly by the caudal border of the scapula and is thus largely visible. The same condition has also been documented for B. borealis by Benke (1993). In contrast, Perrin’s (1870) figure 1 depicts the infraspinatus in B. acutorostrata as mostly obscured by the deltoideus but visible posteriorly where it encroaches substantially onto the teres major. Schulte’s illustration (1916) of B. borealis on the other hand, shows the anterior portion of the infraspinatus to be concealed by the deltoideus and the posterior portion displaced by the encroaching teres major. Consequently, the infraspinatus is only partially visible through the gap between the former two, a condition that is also seen in C. marginata. Furthermore, the outline and extent of the infraspinatus differ substantially among the three whale species: triangular in B. acutorostrata, trapezoidal in B. borealis, and irregular in C. marginata where the anterior border extends craniad as a large lobe beneath the deltoideus.

C a p e r e a m a r g i n a t a
89
In C. marginata the origin of the mastohumeralis is unique and quite divergent from that known in rorquals. In B. acutorostrata the muscle is reported as arising from the transverse process of the anterior cervical vertebrae and the paramastoid process (paraoccipital process, see Mead and Fordyce, 2009), whereas in B. borealis it arises
from the ‘mastoid process’ of the squamosal. In C. marginata the mastohumeralis arises from the lateral portion of the exoccipital - bearing in mind that in C. marginata the exoccipital extends uniquely (in baleen whales) laterad to a point perhaps analogous to Schulte’s ‘mastoid process’ in B. borealis (see also Mead and Fordyce, 2009). In B. acutorostrata, the mastohumeralis inserts onto the proximo-medial preaxial surface of the humerus, while in B. borealis it splits into two heads, one joining the trapezius inserting onto the greater tubercle, and the other inserting onto the fascia of the anterior medial portion of the humerus at the base of the flipper. In C. marginata the arrangement is slightly different in that there is only one tendinous head, albeit also inserting onto the greater tubercle between the supraspinatus and coracobrachialis, a condition that is also seen in odontocetes (e.g. Schulte and Smith, 1918; Howell, 1927; Smith et al., 1976). Muscles absent in C. marginata are as follows: the levator scapulae and the trapezius, present in B. borealis, but not in B. acutorostrata; the teres minor in B. borealis, frequently described in B. borealis as subdeltoid (e.g. Carte and Macalister, 1868) or reported by Benke (1993) as infraspinatus, which is absent in the minke whale; and in the minke whale the flexor palmaris longus reported by Carte and Macalister (1868) and the flexor digitorum sublimis reported by Perrin (1870).
Overall, muscular arrangement in C. marginata agrees better with that of B. acutorostrata, but some detailed morphology of specific muscles corresponds better with those seen in B. borealis, though occasionally also presenting unique configurations not found in either of them. Congruent patterns in C. marginata and B. acutorostrata are: the absence of the levator scapulae, teres minor and trapezius, and the presence of the serratus anterior - anterior division. Morphologies in C. marginata different from B. acutorostrata and more comparable to B. borealis are: the latissimus dorsi in its origin from the ribs; the teres major

C a p e r e a m a r g i n a t a
90
encroaching onto the lateral scapular surface; a deltoid with vestigial second belly; a triceps with two heads; an undivided supraspinatus with origin from the supraspinous fossa; and the insertion pattern of the digital flexors in combination with the absence of the flexors digitorum sublimis or palmaris longus. Unique arrangements in C. marginata are: the origin of serratus anterior - anterior
division arising from the fused first and second ribs, the irregular outline of the infraspinatus and the mastohumeralis arising from the exoccipital. In C. marginata, elongation of the scapula and re-organisation of the musculature allow a broadening of the most powerful muscles, and in consequence greater power output (Gregory, 1912; Smith, 1956; Benke, 1993). Laterally the infraspinatus extends further underneath the deltoid, and caudally the teres major encroaches from the caudal border of the scapula onto the lateral surface. Medially the elongation of the scapula provides a broader base for the most powerful muscle, the subscapularis. The elongation of the cartilaginous vertebral border effects a further shift of muscles towards the extreme ends of the shoulder blade, with possible improvement of leverage important for muscles rotating the scapula, e.g. the serratus and possibly also the rhomboideus. A larger area for attachment and improved leverage might also play a role in muscles controlling limb movement, namely deltoideus and teres major whilst still allowing the supraspinatus ample space. While in B. acutorostrata the retention of some shoulder muscles that are lost in B. borealis must be regarded as plesiomorphic, and the absence of other muscles as derived, the similarity in shoulder morphology with C. marginata cannot be regarded as proof of close kinship (Marx, 2011; Marx and Fordyce, 2016). Conversely,
phylogenetic affinity cannot be entirely excluded either, as demonstrated by Cooper et al. (2007b) on muscle robustness in the cetacean flipper. Nonetheless, although the arrangement of the musculature in mysticetes is highly variable, it is not unique. A wide range of muscular variability in the forelimb has also been recorded in a great number of other mammals (Howell, 1937), where it has been shown in some to be subjected to strong adaptive pressure (Campbell, 1936; Clifford, 2010). Accordingly, congruent muscle attachments between B. borealis and C. marginata will also have to be considered not only in terms of individual variability, but also of functionality. Individual variability is particularly apparent in the muscular arrangement of the mysticete manus:

C a p e r e a m a r g i n a t a
91
here patterns of a few individuals can be more congruent between different species than within the same species (e.g. separated vs fused flexors digitorum radialis and - communis in C. marginata, B. borealis and B. acutorostrata). Other muscle arrangements in the cetacean forelimb, and possible associated osteological configurations, may be more the result of adaptive pressure, as demonstrated in other
mammalian species (Rose et al., 2014; Fernández-Monescillo et al., 2018). If so, a tentative supposition about lifestyle and ecological niche may be possible. Flipper to body ratio, flipper shape, osteological and myological similarities between the pygmy right whale and the sei- and minke whales contrast strongly with the pygmy right whale’s cranial features, redolent of bowhead- and right whales. Streamlined body shape and small and narrow flippers are regarded as typical for the fast gulp-feeding rorquals, whereas a large skull, together with wide flippers, are associated with the slow skim-feeding right whales, and the medium-wide flippers in combination with low-aspect-ratio flukes of the gray whale are believed to be correlated with slow and exact manoeuvring in a structurally complex environment (Woodward et al., 2006). C. marginata shows a perplexing mixture of these traits: the arched jaw line, unpleated throat and fairly long baleen plates demarcate the pygmy right whale as a skim- rather than gulp- or bottom-feeder; the moderately narrow flippers and rather low aspect ratio of the flukes (about 4.4 in MM2904) imply a rather modest cruising speed, yet the small head and flippers, comparable to the fast rorquals, suggest a whale that may also be able to maintain a rather fast cruising speed. This is supported by other postcranial features, such as the fusion of the cervical vertebrae, broad ribs, overlapping transverse processes, and a low count of lumbar vertebrae (Buchholtz, 2011), suggesting a body rigidity that has in other marine vertebrates been associated with a
high cruising speed (Buchholtz, 2001a; Blake, 2004). Yet, observed extensive body flexing and rapid turns (Ross et al., 1975) ostensibly contradict these osteological findings (Buchholtz, 2011). Furthermore, the ability to perform sharp turns has been associated with long flippers, particularly those of the humpback whale (Woodward et al., 2006), and is in direct contradiction with the short flippers of the pygmy right whale. However, moderately wide flippers and a low aspect ratio of the fluke, typified in the gray whale (Woodward, 2006), support life observations of the pygmy right whale performing rapid starts and turns in a shallow lagoon (Ross et al., 1975). The large axillary fold (particularly well visible on the abducted and extended flipper) and a blunt

C a p e r e a m a r g i n a t a
92
flipper tip may compensate for the lack in flipper length by generating more lift and/or improved flow patterns. This in turn might also enhance the ability to counteract pitch when feeding, a factor that has been linked to the large and wide flippers in right whales (Woodward et al., 2006). Furthermore, seasonal change in the abundance of some zooplankton in coastal upwelling areas was postulated as a potential factor dictating
migratory patterns from off-shore to near-shore areas in C. marginata (Sekiguchi et al., 1992). Small body size and manoeuvrability would undoubtedly favour the pygmy right whale when exploiting food resources in a confined environment not open to the less-manoeuvrable rorquals or the much larger right- and bowhead whales. With a length of up to 6.45 m C. marginata is the smallest of all mysticetes (Baker, 1985) and in size only comparable to minke whales (Balaenoptera bonaerensis and B. acutorostrata) or the calves of larger mysticetes, both of which have been reported to be frequently the target of killer whale attacks. Smaller cetaceans, such as dolphins and porpoises, are also vulnerable to predation from sharks, killer whales and seals (Ford and Reeves, 2008; Turnbull, 2012; Leopold et al., 2015). Their typical response is grouping, high-speed directional flight and retreat into shallow water for refuge. The larger mysticetes have been reported to adopt essentially two different strategies: (1) flight: reported for the faster-swimming rorquals (e.g. minke-, sei-, fin- or blue whales). This strategy consists of outrunning the killer whales by rapid (20-40 km/h) directional swimming away from the danger but, if caught, there is little or no physical self-defence. In the majority of observed cases, seeking refuge in shallow water ended in death by either stranding or predation (Jefferson et al., 1991; Ford and Reeves, 2008). (2) fight: reported for the slower swimming gray whales, humpback whales and right- and bowhead whales. This strategy consists of active physical defence using mostly
flippers and tail, and sometimes the head. It has been observed bothj in individuals, by mothers defending their calf, or in coordination by the whole group, and was, combined with retreat into shallow water, often successful (Ford and Reeves, 2008). This tactic also included escaping detection by hiding in kelp or behind breaking surf, or quiet, low-profile breathing and, if caught, by rolling the vulnerable underside up and out of the water. These species also undertake long migration, mostly along the shore, from protected and relatively safe low-latitude calving grounds to high-latitude feeding grounds (Corkeron and Connor, 1999; Ford and Reeves, 2008). Apart from these

C a p e r e a m a r g i n a t a
93
general strategies, specific behavioural changes may also play a role, e.g. abandonment of an area with a high occurrence of killer whales (Sironi et al., 2008). Based on observations in the wild, pygmy right whales ostensibly combine several strategies: although not undertaking large-scale migrations, they appear to feed in the colder offshore waters of the southern hemisphere and move into more temperate
shallow coastal waters to give birth. After weaning the juveniles probably move southwards to exploit the plankton-rich waters (Kemper, 2002). Observations also suggest that pygmy right whales are pelagic, roaming temperate to subantarctic waters, possibly in small to fairly large groups (Matsuoka et al., 1996; Matsuoka et al., 2005) with seasonal movements inshore (Ross et al., 1975). If so, the pelagic habitat of pygmy whales would preclude a quick retreat into shallow water, and the small size of body and flippers appear to be rather unsuited for defence, thus favouring a flight strategy. This is supported by a number of adaptations correlated with speed (streamlined body shape, body rigidity, and small, strong flippers with body indent). Although the pygmy right whales of this study (a neonate and a juvenile) were found to have a rather low fluke aspect ratio, better suited to rapid starts than prolonged speed (see also Woodward et al., 2006), killer whales are known to tire fairly rapidly and to be outrun by the similar-sized minke whales (Ford and Reeves, 2008). On the other hand, adult pygmy right whales may have higher-ratio flukes than juveniles, better suited for sustained high speed, and it is not known if they are also able to porpoise like fleeing minke whales (Ford and Reeves, 2008). Shoaling into large groups may also be a defence mechanism, but it remains speculative if the whales would remain as a group under attack or rather disperse like fin whales (Ford and Reeves, 2008). Furthermore, pygmy right whales might apply yet a slightly different strategy: instead of fleeing in a straight line at all costs, similar to deer hunted by a
pack of wolves, C. marginata may well be able to draw on its ability to execute fast and tight turns and dodge attacks at the last moment like hare. In favour of this strategy would be the pygmy right whale’s small size (less inertia than larger and heavier whales) and a moderately wide flipper with axillary fold (increased lift). This strategy is not open to the faster rorquals since they are not agile enough, or the slow right- and bowhead whales. Conversely, the apparent seasonal inshore movement of the pygmy right whale would favour techniques associated with the fight strategy (Ross et al., 1975; Corkeron and Connor, 1999). The ability to rapidly start and perform tight turns would enable the

C a p e r e a m a r g i n a t a
94
pygmy right whale to manoeuvre in shallow water without getting stranded. Paradoxically, pygmy right whales seem to be rather solitary when near shore and exhibit cryptic behaviour (inconspicuous breathing and swimming) or shoaling behaviour with other cetacean species (Baker, 1985) rather than group defence.
Overall, it seems that the shoulder morphology of C. marginata has been influenced by at least two opposing major factors: (1) skim feeding, requiring large and wide flippers to counteract sustained pitching forces when feeding, and possibly also the need for precise manoeuvring in shallow water; (2) predator avoidance in open water, requiring small and narrow flippers to reduce drag and maintain high speed efficiently. However, the pygmy right whale has been reported not only to feed on the comparatively slow copepods, but also on the faster euphausiids (Kemper, 2002), and in doing so may also, like the sei whale, use more than one feeding technique and a wider range of food resources (Reeves et al., 2002). It is thus conceivable that adaptations to speed may have also arisen from the need to engulf prey efficiently, particularly if competition with the fast and larger rorquals is a factor (Kemper, 2002). Nonetheless, the result is a strong, short but moderately wide flipper, complemented by a large axillary fold, possibly enabling the pygmy right whale to switch from one strategy to the other in an instant.
Wider implications
C. marginata is a little-known and poorly understood species that possesses a mixture of characteristics found in either bowhead and right whales on one hand, or grey whales and rorquals on the other hand, as well as some features that are unique or only shared with the extinct cetotheriid Piscobalaena nana (see also Chapter 4 Functional anatomy: Diversity of flipper shapes). Up to date the controversy about which characters are the result of convergence and which truly represent phylogenetic relationships has continued: Morphological data have generally associated the pygmy right whale with right and bowhead whales (e.g. Steeman, 2007; Bisconti, 2012, 2014; el Adli et al., 2014) whereas molecular data supported a closer relation with grey whales and rorquals (e.g. Saskai et al., 2005; McGowen et al., 2009; McGowen et al., 2014). Yet total evidence studies (combined morphological and molecular data) support a closer relationship with grey whales and rorquals (e.g.

C a p e r e a m a r g i n a t a
95
Deméré and Berta, 2008; Geisler et al., 2011; Marx, 2011). However, there are a handful of other studies that have revealed new contradictory evidence: a morphological study associating the pygmy right whales with grey whales and rorquals (Fitzgerald, 2012), a molecular study regarding C. marginata as a basal relative of the right and bowhead whales (Agnarsson and May-Collado, 2008) and morphological or
total evidence studies placing C. marginata within the extinct family Cetotheriidae (Fordyce and Marx, 2013; Tsai and Fordyce, 2014a, b) or in close proximity to either Cetotheriidae or grey whales and rorquals (Marx et al., 2013). Morphological studies have so far been dominated by evaluation of cranial characters and little attention has been given to postcranial character traits. Yet some postcranial features bear a striking resemblance to those attributed to grey and rorqual whales (e.g. length/width ratio of flipper, tetradactyly) or cetotheriids (e.g. scapular shape), and by doing so ostensibly contradict right and bowhead whale affiliations. Other inconsistencies may rather arise from variables linked to a particular individual specimen and not from the sampling concept: although it is well known that ontogenetic development recapitulates phylogenetic pathways (e.g. hindlimb development in cetaceans, Thewissen et al., 2006; tooth development in mysticetes, Deméré et al., 2008; McGowen et al., 2014), it has only been recently demonstrated that different ontogenetic stages can confuse placement of phylogenetic relationships in cetaceans (Tsai and Fordyce, 2014b). Another consideration lies with homoplastic character traits that, by definition, reflect modifications triggered by adaptational pressures. Although not relevant in taxonomical terms, recognition of these factors can lead to a better understanding and differentiation of homologous and analogous character traits and their developmental pathways, which in turn could lead to a better understanding of phylogenetic affinities, and lifestyle and habitat preferences in living and extinct
species. All these considerations are important regarding the phylogenetic status of C. marginata but may equally contribute to the understanding of the lifestyle and behaviour in extinct fossil species.
Future studies
Shoulder morphology and anatomy of the flipper portray C. marginata as a whale with a strong shoulder musculature capable of employing moderately slim but blunt flippers forcefully. Although pygmy right whales have been reported to execute fast and tight

C a p e r e a m a r g i n a t a
96
turns, it is neither known whether or if so, how these whales utilise their flippers when doing so, nor the techniques used when feeding. The large axillary fold and the transverse position of the shoulder blade rise additional morphofunctional questions. Computed models integrating different positions of the flipper on the torso as well as varying flipper shapes and orientations may also reveal altered flow patterns affecting
efficiency and manoeuvrability, thus highlighting adaptive patterns as a response to selective pressures as documented by Pilleri et al. (1976) for Delphinus and Platanista. It is unclear how the distinctive angular type of the shoulder blade in some pygmy right whales is related to the extinct cetotheriid Piscobalaena nana and other cetotheriids (see also Marx and Fordyce, 2015) or extinct mysticetes in general (e.g. Tranatocetus, Gol'din and Steeman, 2015). Elongation of the scapula is undoubtedly enhanced by the angular shape of the scapula and goes hand in hand with a muscular arrangement typical for fast-swimming baleen whales; it may also be linked to the large olecranon, which in turn might be essential for the efficiency of the axillary fold. Apparent adaptations for both speed and manoeuvrability ostensibly imply a strong influence on shoulder and forelimb morphology and are also reflected in other postcranial elements (Buchholtz, 2011). Yet, fan-shaped scapulae, characteristic for rorquals, also occur in pygmy right whales and the co-existence of both shape types is suggestive of at least some morphofunctional flexibility permitting both types to persist equally. This can also be said for other elements of the forelimb. Transition to an aquatic life has radically modified both bone and soft tissue in the flipper. In spite of that, proportions of the humerus (Benke, 1993) and neuromuscular anatomy (Cooper et al., 2007b) are best predicted by phylogenetic distribution. Understanding the relationship between adaptive and hereditary factors in C. marginata may also help to resolve the enigma of the unusually broad ribs unique to the pygmy right whale (Buchholtz, 2011) and yet
another extinct cetotheriid, Cetotherium riabinini (Gol'din et al., 2013). Further studies of the forelimb skeleton in other baleen whales, and particularly of the hitherto undescribed shoulder muscles of gray whales and right and bowhead whales, may reveal morphofunctional correlations that can be used to separate homoplastic from synpomorphic character traits which in turn may either be linked to genetic data (e.g. direct link between hox gene and hyperphalangy, Wang et al., 2009) or osteological and fossil material (e.g. potential link between axillary fold, enlarged olecranon and elongated and/or angular shape of scapula).

C a p e r e a m a r g i n a t a
97
Summary The stranding of two pygmy right whales, Caperea marginata, in 2010 and 2011 on New Zealand’s shores offered a unique opportunity to investigate the musculoskeletal arrangement of the shoulder in this enigmatic whale. Dissections were carried out on a 2.96 m long juvenile female (MM2904) and a 1.87 m long neonate male (MM2959) and radiographs taken. Flipper shape and relative dimensions correlate well with those of rorquals. The pygmy right whale has, in relation to body size, together with the fin whale (Balaenoptera physalus) some of the shortest flippers among baleen whales. The flipper itself is moderately narrow and about midway between the short but narrow flippers of rorquals and the broad and long flippers of bowhead and right whales. However, unlike the oar-shaped flipper with pointed tip seen in rorquals, the flipper of C. marginata is oblong elliptical with rounded tip. Attached to the trailing edge of the flipper and supported by a large olecranon is an atypically-large axillary skin fold, only fully exposed on the
extended limb. Examination of osteological characteristics, also including additional historical specimens, revealed unusual features in the shoulder morphology of C. marginata, namely two extremely-elongated types of shoulder blade. Type 1 has a fan-shaped contour with rounded vertebral border (6 specimens) whereas type 2 is trapezoid with tripartite angular vertebral border (3 specimens) - although one specimen showed an intermediate scapular shape on the right side and a strongly angular shape on the left side. While the fan-shaped type is typical for rorquals and grey whales, albeit not as elongated, the trapezoid outline is only seen in one other species, the extinct cetotheriid Piscobalaena nana. In both types the elongation of the scapula is exaggerated by the cartilaginous vertebral border, making it proportionally the longest shoulder blade in cetaceans. Other characteristics of the skeleton correspond well with both rorquals and P. nana, such as the ratio of humerus / radius length of roughly 4:6 and tetradactyly of the manus. Probably quite unusual for baleen whales is the distinctive slant of the glenoid border rising caudo-dorsad towards the caudal border of the scapula. This slant is strongly correlated to the oblique ventro-caudad orientation of the shoulder blade on the torso. The combined indirect proportional inclination of the

C a p e r e a m a r g i n a t a
98
shoulder blade and glenoid border thus result in the alignment of the glenoid cavity (and limb) at right angles to the long axis of the body. In comparison with the minke whale (Balaenoptera acutorostrata) and sei whale (Balaenoptera borealis), the overall pattern of the shoulder musculature in the pygmy right whale agrees better with that of the minke whale, while detailed morphology of
specific muscles correspond better with those of the sei whale (see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscular arrangements, Table 2). Congruence of the musculature in the minke whale is mainly due to the absence of the levator scapulae, teres minor and trapezius, and the presence of the serratus anterior - anterior division. Specific configurations of muscles divergent from the minke whale, and congruent with the whale, are the latissimus dorsi in its origin from the ribs, the teres major encroaching onto the lateral scapular surface, a deltoid with a rudimentary second belly, a triceps with two heads, an undivided supraspinatus with origin from the supraspinous fossa, and the extensor and flexors of the manus merging into four digital tendons inserting onto the full length of the digits. Unique arrangements in the pygmy right whale include the origin of the serratus anterior - anterior division from the first rib, the irregular outline of the infraspinatus and the mastohumeralis arising from the exoccipital. Morphofunctional implications inferred from the position of the scapula in combination with the musculoskeletal arrangement and flipper shape are not clear. The slanting orientation of the scapula places the limb forward and down. In a whale that is known for its agility and ability to execute tight turns, despite very small flippers, albeit moderately slim and blunt, powerful use of the flippers and nimbleness would possibly be a crucial component of steering, and placement of the flippers to augment body roll and steering performance may be an important factor.
Elongation of the scapula and improved organisation of muscles allow a broadening of the main muscles and possibly improved leverage. Long scapulae have been correlated with increased muscle strength in fast-swimming baleen whales, facilitating a more powerful use of the flippers, particularly at speed. Furthermore, additional loading from the exposed axial skin fold on the extended flippers may also necessitate powerful musculature. However, the true nature of the trigger leading to a more powerful shoulder musculature remains speculative. Overall, morphological characters of the pygmy right whale appear to be contradictory and hard to reconcile with each other. On one hand, a streamlined, rigid body, and

C a p e r e a m a r g i n a t a
99
small flippers with body indent to reduce drag, imply fast locomotion, whereas wide flippers are regarded as necessary to either counteract pitch when skim-feeding or to manoeuvre precisely in shallow water and confined spaces. However, feeding methods and speed of the pygmy right whale are not known, and while C. marginata’s arched jaw line, unpleated throat and fairly long, fine baleen plates demarcate it more
as a slow skim- rather than a fast gulp-feeder, speed and agility may give the pygmy right whale an advantage when competing with the fast rorquals. Small size and solitary lifestyle make C. marginata vulnerable to predation. Cryptic behaviour, coupled with speed and agility, suggest a strategy to retreat fast into shallow water or flee before being detected. In open water, where this might not always be possible, pygmy right whales have been documented to gather in larger groups. However, small size and small flippers make it unlikely for C. marginata to adopt a fight strategy. In this case pygmy right whales might react rather like rorquals by rapidly dispersing individually; with the added twist that pygmy right whales may be able to also dodge attacks at the last moment, an option that is not open for the less manoeuvrable rorquals. The result of these opposing requirements is a strong but short and moderately wide flipper, complemented by a large axillary fold, possibly enabling the pygmy right whale to switch from one requirement to the other in an instant, thus making the musculoskeletal arrangement of the shoulder in C. marginata unique within extant cetaceans.
Caperea marginata MM2904 Life reconstruction of a juvenile female pygmy right whale stranded at Glink’s Gully, west coast of Northland, North Island, New Zealand on 05 September 2010

C a p e r e a m a r g i n a t a
Plate I
Plate I - Figs. 1-2. Radiographic image of the left (Fig. 1) and right (Fig. 2) forelimb of C. marginata MM2959.

C a p e r e a m a r g i n a t a
Plate II
Plate II -Fig. 1. Radiographic image of the left forelimb of C. marginata MM2904.
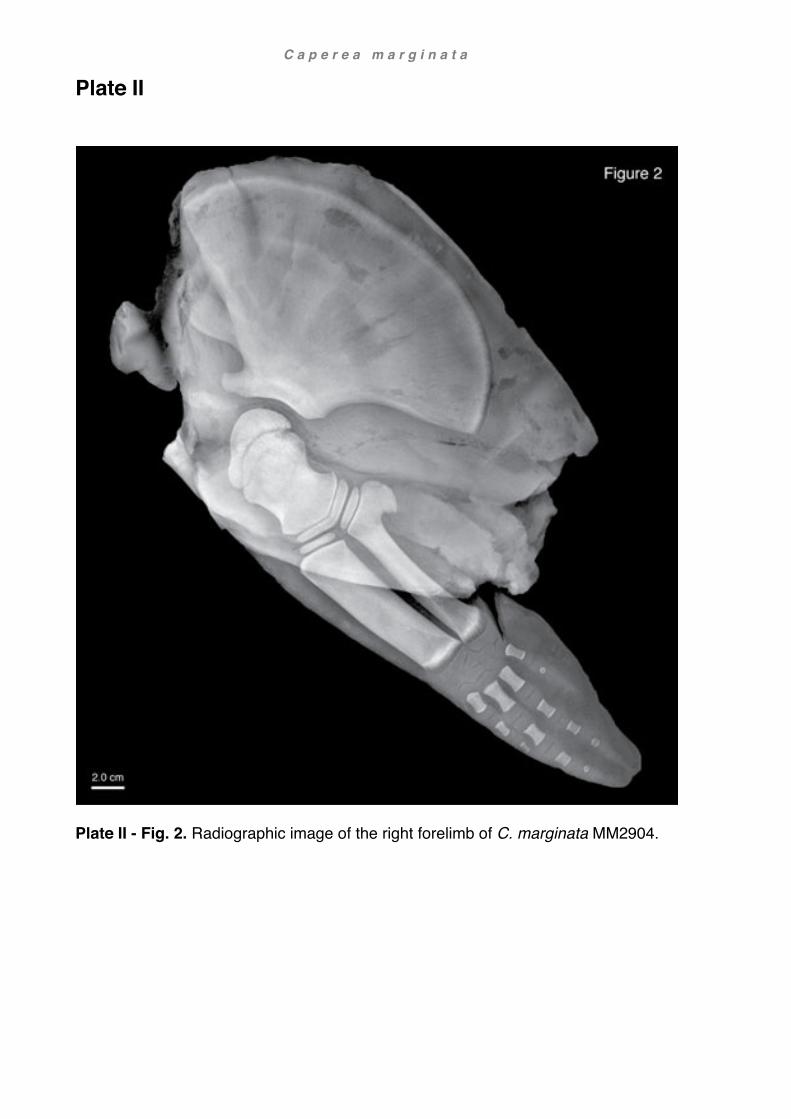
C a p e r e a m a r g i n a t a
Plate II
Plate II - Fig. 2. Radiographic image of the right forelimb of C. marginata MM2904.

103
Chapter 3: Arnoux’s beaked whale
Berardius arnuxii Duvernoy, 1851
Introduction Beaked whales (Family Ziphiidae) are the most speciose but also the least known of all toothed whales (Odontoceti). They usually live in small, widely dispersed groups and lead highly pelagic and cryptic lives (Balcomb, 1985; Mead, 2009; Madsen et al., 2014). Ziphiids are the most extreme deep-diving mammals (Tyack et al., 2006; Schorr et al., 2014) and hunt predominantly squid and fish (MacLeod et al., 2003) using a highly sophisticated sonar system and suction feeding to capture their prey (Werth, 2006; Johnson et al., 2008; Baumann-Pickering et al., 2014). In order to avoid predators, beaked whales also dive silently near the water surface and emerge
inconspicuously, spending as little time at the surface as possible between dives (Aguilar de Soto et al., 2012). As a consequence, ziphiids have never been targeted by large-scale whaling industries (apart from a few individuals targeted by local whalers), and have proven difficult to study in the wild and impossible to keep in captivity (Mead, 2009). Therefore, beaked whales have remained poorly understood despite their generally large body size, broad geographical distribution, and high species number. The stranding of three Arnoux’s beaked whales in Southern New Zealand (one whale in 2010, two whales in 2013; see also Chapter 1 Methods and materials) thus offered an excellent opportunity to examine for the first time the musculoskeletal system of the shoulder and forelimb in this poorly known species, particularly with regard to phalangeal counts, general soft tissue and myological characteristics. Evolution, distribution and habitat of beaked whales
Many beaked whale species are best known from stranded animals or fossil remains. Morphological studies of beaked whales have revealed Berardius and Tasmacetus as generally the most basal ziphiids, usually placing Tasmacetus basal to Berardius (e.g.

B e r a r d i u s a r n u x i i
104
Moore, 1968; Lambert et al., 2013; Bianucci et al., 2016 - but not Bianucci et al., 2007; see also Fig. 1). This has also been supported by recent molecular and total evidence analyses, though some studies place Berardius more basal than Tasmacetus (e.g. McGowen et al., 2009 molecular; Gatesy et al., 2012 total evidence; Geisler et al., 2011 total evidence, not weighted - but not Agnarsson and May-Collado, 2008
molecular; Geisler et al., 2011 total evidence, weighted). General morphological traits observed in fossil and extant ziphiids show a trend towards elevation of the vertex (synvertex), asymmetry of the narial region, enlarged pterygoid sinuses on the ventral region of the skull, general shortening and compaction of the rostrum, reduced dentition, adaptations of the vertebral column and ribs including fusion of neck vertebrae, decreased size of the flipper and increase in body size (Balcomb, 1985; Heyning and Mead, 1996; Lambert et al., 2013; Lambert et al., 2015; Bianucci et al., 2016). Emerging in the Early Miocene, as early as 17.5 my ago, ziphiids rapidly diversified including stem ziphiids referred to by Bianucci et al. (2016) as the Messapicetus clade, and crown ziphiids. The last stem ziphiids disappeared during the Pliocene about 3.5 my ago whereas crown ziphiids are represented by 6 extant genera (Tasmacetus, Berardius, Ziphius, Mesoplodon, Hyperoodon, Indopacetus; Bianucci et al., 2016). Interestingly, both groups evolved similar features independently of each other. The loss of functional teeth, vertex elevation (synvertex) and asymmetry of the premaxillary sac fossae are regarded as homologous but convergent traits (homoplasy). Analogous traits on the other hand are the closure of the mesorostral groove by pachyosteosclerosis of the premaxillae in stem ziphiids and by the vomer in crown ziphiids, as well as distinctive crests of the premaxillae in stem ziphiids and of the maxillae in crown ziphiids (Bianucci et al., 2016).
Bianucci et al. (2016) explain the parallel evolution of these convergent traits by four distinctive radiation events. The earliest radiation of the stem ziphiids took place in the South-East Pacific and North Atlantic, where they probably inhabited shallow, nutrient-rich coastal waters. Later stem ziphiids successively adapted to deep diving and suction-feeding in the North Atlantic and Mediterranean Sea. The majority of the crown ziphiids also occupied a similar habitat and lifestyle but initially inhabited the southern seas from where they ostensibly dispersed to all other oceans, possibly during the cooler Pleistocene.
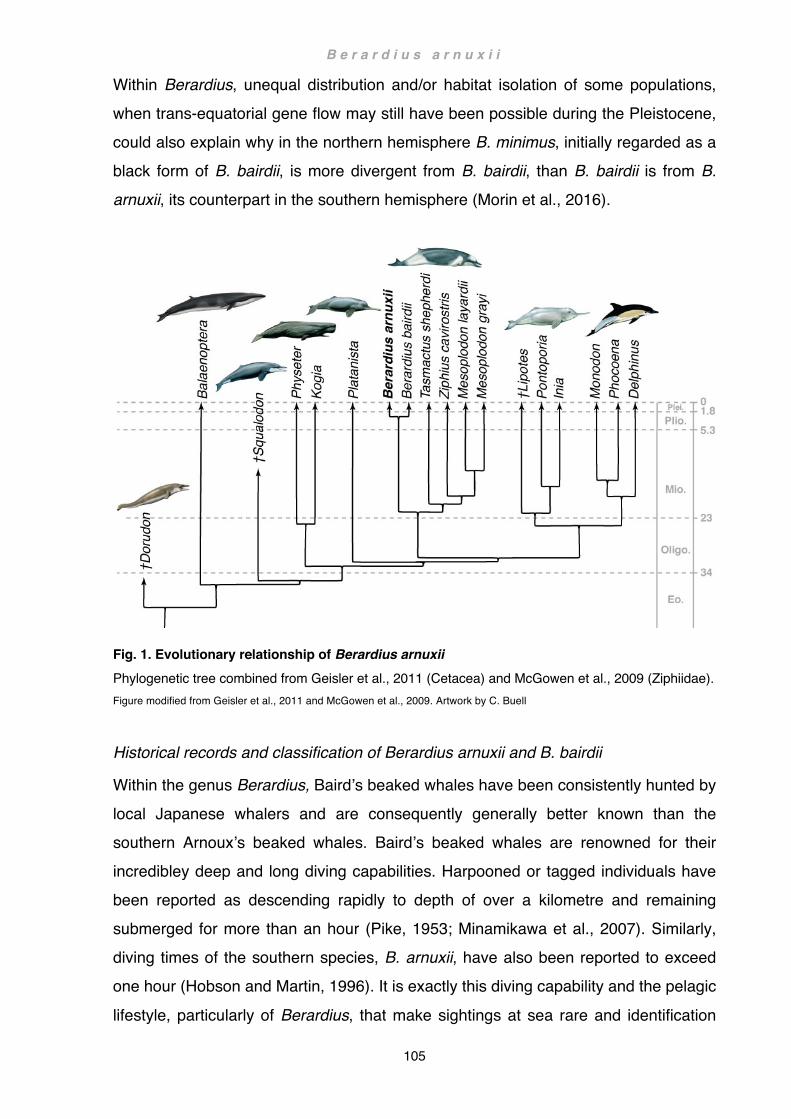
B e r a r d i u s a r n u x i i
105
Within Berardius, unequal distribution and/or habitat isolation of some populations, when trans-equatorial gene flow may still have been possible during the Pleistocene, could also explain why in the northern hemisphere B. minimus, initially regarded as a black form of B. bairdii, is more divergent from B. bairdii, than B. bairdii is from B. arnuxii, its counterpart in the southern hemisphere (Morin et al., 2016).
Fig. 1. Evolutionary relationship of Berardius arnuxii Phylogenetic tree combined from Geisler et al., 2011 (Cetacea) and McGowen et al., 2009 (Ziphiidae). Figure modified from Geisler et al., 2011 and McGowen et al., 2009. Artwork by C. Buell
Historical records and classification of Berardius arnuxii and B. bairdii
Within the genus Berardius, Baird’s beaked whales have been consistently hunted by local Japanese whalers and are consequently generally better known than the southern Arnoux’s beaked whales. Baird’s beaked whales are renowned for their incredibley deep and long diving capabilities. Harpooned or tagged individuals have been reported as descending rapidly to depth of over a kilometre and remaining submerged for more than an hour (Pike, 1953; Minamikawa et al., 2007). Similarly, diving times of the southern species, B. arnuxii, have also been reported to exceed one hour (Hobson and Martin, 1996). It is exactly this diving capability and the pelagic
lifestyle, particularly of Berardius, that make sightings at sea rare and identification

B e r a r d i u s a r n u x i i
106
difficult (Mead, 2009). Arnoux’s beaked whales, never targeted at all by whalers, have therefore remained one of the most obscure beaked whale species despite their large size. The first extant specimen of an Arnoux’s beaked whale (in older literature sometimes referred to as porpoise whale), a lower jaw, was recovered in the 1840s from Taiaroa
Head (Otago Peninsula, South Island, New Zealand) and subsequently donated to the Otago Museum 70 years later (ID number: OMNZ VT220; Oliver, 1922; McCann, 1975). Yet it was not until 1851 when Duvernoy described a skull from Akaroa, Canterbury (South Island, New Zealand) presented to him in 1846 at the Museum d’Histoire Naturelle in Paris, that both genus and species were scientifically recognised (Robineau, 1989). Duvernoy named the new genus after Mr. Berard, the captain of the corvette ‘Rhin’ who presented the skull to the museum, and the species after the surgeon of the ship, Mr. Arnoux, who collected the specimen and briefly described the animal and the circumstances of its capture: hence Berardius arnuxii Duvernoy, 1851. Since then, the spelling of the species name has variably appeared as ‘arnouxii’ (e.g. Owen 1870, Marelli, 1920), ‘arnuxi’ (e.g. Taylor, 1957) or etymologically and grammatically correctly as ‘arnouxi’ (e.g. Flower 1872, Beddard 1900). Still, the original spelling of B. arnuxii Duvernoy, 1851 will take precedence here with reference to the International Code of Zoological Nomenclature (Rice, 1998). Subsequent later records also confused other whale species with this genus, some genuinely belonging to the genus Mesoplodon, some associated with the genus Mesoplodon but actually belonging to Berardius (i.e. B. arnuxii), and vice versa. These specimens were often identified by their stranding or storage locations which were themselves not always consistently cited (see Chapter 5 Literature review: Osteology,
Table 1). For example, the holotype of Mesoplodon hectori (Gray, 1871). The specimen was originally described as a young individual of ‘Berardius arnuxii’ by Hector in a note attached to a paper published by Knox in 1870. Hector referred to the specimen as a beach cast on the West Coast near Porirua Harbour (the modern day Awarua-o-Porirua Harbour: reference to Land Information New Zealand, LINZ) and as having been prepared by Knox (Knox, 1870). In the same year Hector again published the same account as part of an inventory, listing various specimens held at the Colonial Museum in Wellington (now Museum of New Zealand; Hector, 1870). In 1871 Knox

B e r a r d i u s a r n u x i i
107
published a more detailed description with plates referring to the specimen as ‘ziphid [sic] whale, second specimen’ and the stranding location as ‘Titai Bay, Cook’s Strait’. Confusingly, he referred in the text also to plate XIII - fig. 1 which depicts a specimen of Berardius arnuxii he referred to otherwise as ‘ziphid [sic] whale, first specimen’ captured in ‘Porirua Harbour’. According to LINZ, Titahi Bay is an old Maori settlement
and has been a whaling station since the early 1800’s. It is situated just south-east of Awarua-o-Porirua Harbour and north of Wellington Harbour on the North Island. In the same paper, Hector also added some comments but without assigning any of the specimens to a particular genus or species (Knox, 1871). Finally Gray realised in 1871, based on Knox’s and Hectors’s papers of the same year, that differences between the skull of Berardius arnuxii and Hector’s specimen were too great to be explained by age or sex, and proposed a new species ‘Berardius hectori’ (Gray, 1871), which was in 1872 correctly consigned to Mesoplodon hectori by Turner (Turner, 1872). However the confusion did not end here: various publications cited the stranding location also as Tatai, Taitai, Tetai or Titahi Bay and the species as Berardius hectori (Gray, 1873; Hector, 1873a; Gray, 1874a, b), Mesoplodon knoxi (Hector, 1873b) or Paikea hectori (Oliver, 1922). Even as late as 1975 McCann deemed M. hectori to be a juvenile of Berardius arnuxii (McCann, 1962, 1975; for a list of above mentioned specimens see also Chapter 5 Literature review: Osteology, Table 1). Nonetheless, it was not until two decades after Duvernoy’s first account of Berardius arnuxii that Flower published a detailed description of the whole skeleton in 1872. Yet another decade passed until a similar northern whale, B. bairdii, was described by Stejneger in 1883 on the basis of a skull. At last True described and figured a complete specimen of Baird’s beaked whale in 1910. Subsequently the validity of Arnoux’s and
Baird’s beaked whales as two discrete species was disputed by Slipp and Wilke (1953) and Pike (1953). This dispute, however, can now be laid to rest (de Queiroz, 2007; Taylor et al., 2017), and the two species are now classified as Berardius bairdii Stejneger, 1883 and Berardius arnuxii Duvernoy, 1851 (List of Marine Mammals 2014 of the Society for Marine Mammalogy). Moreover, in 2016 Morin et al. argued for a third, sympatric species of Berardius bairdii recently named Sato’s beaked whale Berardius minimus Yamada, Kitamura and Matsuishi, 2019.

B e r a r d i u s a r n u x i i
108
Anatomy and morphological characteristics of the genus Berardius
Apart from cranial characteristics, few accounts of the skeleton of Berardius exist and no detailed study of the forelimb has so far been produced. Evolutionary considerations are mostly restricted to character traits of the skull, or are very unspecific. General plesiomorphic characteristics of Berardius are
- Less prominent elevation of the vertex (synvertex; Moore, 1972; Bianucci et al., 2016) including shape and orientation of nasale and premaxillary crest (Moore, 1972)
- Absence of mesorostral ossification (Moore, 1972) - Intermediate state reflected in the tooth reduction to 4 vestigial teeth, placing
Berardius between Tasmacetus (fully functional dentition) and other beaked whales (two vestigial teeth; Bianucci et al., 2016)
The only apomorphic feature within ziphiids is the extremely large body size, also seen in Ziphius and Hyperoodon (Lambert et al., 2013).
There are only two noteworthy descriptions of the postcranial skeleton and/or soft tissue in Berardius bairdii, largely consisting of body measurements including fins (True, 1910; McCann, 1975), postcranial bones except forelimb (True, 1910; McCann, 1975) and fragmentary accounts of forelimb bones including the scapula (McCann, 1975) and long bones (True, 1910) but missing the bones of the manus. Remarkably, more has been published about the postcranial skeleton of Berardius arnuxii (see Chapter 5 Literature review: Osteology, Table 1). Some publications are short notes (Hale, 1962), many by different authors referring to the same specimens (Hector, 1870; Knox, 1870; Gray, 1871; Flower, 1872; van Beneden and Gervais, 1880). Some are followed up by a sometimes more detailed description in later publications, including measurements of the flippers and/or figures of the forelimb, yet incomplete (Knox, 1871) or highly unlikely (van Beneden and Gervais, 1880) phalangeal counts. There are only three reports giving an altogether not unrealistic phalangeal count; one is without accompanying figures or a detailed description, making it hard to validate (Hector, 1878). Some of the most persuasive osteological accounts have been published by Flower (1872) and Marelli (1920). Flower briefly lists characteristics of the genus before describing in more detail four historic specimens

B e r a r d i u s a r n u x i i
109
(history of the specimens, reference to other authors, osteology), including the holotype. He continues with aspects of osteological characteristics of the species. This also includes a detailed general description and figures of the forelimb (pl. XXVII and XXVIII) and a phalangeal count of 2/5/4/4/3 (p. 231, including carpal 1, metacarpals and phalanges), also mentioning his suspicion that some terminal phalanges were
probably missing (Flower, 1872). Marelli finally published the most comprehensive morphological account of a specimen stranded on the banks of the Rio de la Plata, Argentina, South America. He gives an exhaustive account of body attributes, including measurements of the flippers, and osteological descriptions and measurements including those of the forelimb with two figures, but no phalangeal counts (Marelli, 1920). Anatomical accounts of the shoulder and forelimb in ziphiids and other odontocetes
There are no accounts detailing the soft tissue of the shoulder and/or flipper in Berardius. However, Struthers published in 1873 a short description of flipper
musculature in Hyperoodon bidens (synonym of Hyperoodon ampullatus (Forster 1770), see World Register of Marine Species) and twelve years later, Turner published a comprehensive description of the flipper in Sowerby’s beaked whale (Mesoplodon bidens). Presented with an isolated flipper, Turner did not include muscles pertaining to the shoulder. Nevertheless, he described the intrinsic soft tissue of the flipper in minute detail, including two figures depicting the lateral and medial aspect of the dissected flipper (Turner, 1885). Turner also analysed and compared patterns of the wrist elements (carpals) in two ziphiid and several delphinid species (Turner, 1909). Other noteworthy publications of ziphiid osteological material include those of van Beneden and Gervais (1880), Kükenthal (1893a), and Benke (1993).
Results
Pectoral limb
In Berardius arnuxii, the ratio of body to flipper length ranges from 10.3 (adult male) to about 13.5 (juvenile and subadult female; for specimen ID see Chapter 1 Methods and

B e r a r d i u s a r n u x i i
110
materials). In contour, the flipper is deltoid-shaped, fairly short, broad and angular (Fig. 2a). The leading edge follows a wide, moderately convex arch, distally more so than proximally; the tip is blunt; the trailing edge initially diverges from the leading edge of
the flipper at about 10-15º before, at midpoint, gently turning towards the tip where it meets the leading edge at an angle of about 70º. Consequently, the widest point of the flipper is about 2/3rd distad along the leading edge and about midpoint along the trailing edge. The base of the flipper is very stout and bulky, in cross-section oval, the axillary skin fold very thick and fleshy. The fleshy base merges rapidly into a thin trailing edge distally (Fig. 2d). The leading edge is remarkably wide and blunt at the base of the flipper, narrowing only gradually towards the tip (Fig. 2c).
Fig. 2a-b. Flipper contour and indent in the flank (a) lateral view of retracted right flipper, Tangimoana; (b) caudo-ventral view showing indent in the flank to accommodate the flipper (white dashed line), Purakaunui. Photos (a-b) R. E. Fordyce
Fig. 2 c-d. Flipper profile (c) cranio-ventral view onto leading edge also showing indent in flank (dashed white line), Hine Reti; (d) caudo-dorsal view of the fleshy base and the trailing edge onto the lateral surface of the right extended flipper, Hine Reti.

B e r a r d i u s a r n u x i i
111
Photos (c-d) R. E. Fordyce
Colouration of the flippers is a uniformly dark grey, tinged with a cloudy grey-brown to red-brown laterally, faintly demarcating the manus, but almost black around the edges and medially. The skin may take on a deep black or, when dried out, a lighter greyish hue after death. Shallow indents in the flanks accommodate the retracted flippers
(Fig. 2b, c).
Osteology See also plates I-III and measurements in appendix
Scapula
In lateral view, the flat and bony scapular blade of the Arnoux’s beaked whale takes on the classical fan-shaped outline seen in most cetaceans (Figs. 3a-d, 12a-g).
- The vertebral border rises as a shallow, convex arch only little above the cranial and caudal angle.
- Cranial and caudal borders are straight or slightly concave - The neck of the scapula is sturdy and merges into the caudal border at a
comparatively shallow angle of 130º-150º - The lateral surface of the scapula is divided into four divisions by three blunt ridges
roughly demarcating: (1) supraspinous fossa (2) origin of the deltoid (3) infraspinous fossa (4) origin of the teres major
- The supraspinous fossa is moderately large, about 1/10th of the scapular width.
The three fossae further caudad each occupy about 1/3rd of the remaining scapular surface
- The scapular spine, defining the caudal border of the supraspinous fossa, initially emerges from the cranial angle as a barely distinguishable, smooth ridge. From here it converges towards the acromion and gradually emerges as a pronounced crest. This crest merges distally into the proximal dorsal border of the acromion process.
- The acromion process is long, broad and fairly angular in outline, the tip is wider than the base.

B e r a r d i u s a r n u x i i
112
- The coracoid process is well developed, long and moderately flared at the tip. - There are six or seven barely recognisable smooth ridges on the medial surface
of the scapular blade that correspond to the tendinous divisions of the subscapularis, the most cranial and caudal ridges are the least prominent. In Tangimoana and Hine Reti the medial scapular surface is otherwise smooth.
The scapula of Hine Reti matches this description closely, except in slight differences of the acromion, coracoid and the scapular neck, the latter lacking a clear separation from the caudal border (Fig. 12b). In Purakaunui, the supraspinous fossa is slightly smaller, about 1/15th of the scapular width. The scapular spine, emerging from the blade as a blunt ridge, progresses immediately dorsad of the acromion into a small but sharp convex flange that is strongly embedded in the connective tissue spanning between the cranial angle and the acromion (prescapular ligament; Figs. 3a,b, 12c). The scapular neck of Tangimoana, not unlike that of Hine Reti, is also barely set apart from the caudal border. The supraspinous fossa is much smaller than in the other specimens, about 1/25th of the scapular width. As in the other specimens, the scapular spine arises rather indistinctly, but then transforms fairly high up on the scapular blade into a sharp ridge that continues as a thin bony sheet across the scapular border before merging into the dorsal margin of the acromion (Fig. 12a).
Fig. 3 a-b. Scapula (a) lateral view and (b) medial view of left scapula, Purakaunui Dashed lines: (a) ridges separating (sf) supraspinous fossa, (od) origin of deltoid, (if) infraspinous fossa, (otm) origin of the teres major; (b) Dashed lines: smooth ridges corresponding to tendinous intersections of the subscapularis

B e r a r d i u s a r n u x i i
113
In B. arnuxii the acromion process is in lateral view usually broad, more or less spatulate, i.e. the tip is in outline squarish and slightly wider than the base and does not quite reach as far forward as the cranial most point of the scapula. The base of the acromion is solid and tapers cranially sharply into a long, thin blade. Initially, the base rises laterad above the scapular surface before turning mediad, the anterior part
crossing the scapular plane in a wide arch (Fig. 3c). The acromion process of Hine Reti largely fits the general description, except that the bone maintains the same width throughout from base to tip (Fig. 12b). In Purakaunui, the dorsal border of the acromion follows a sinuous path, concave at the base and convex towards the tip, and in consequence, the tip is slightly rounded dorsad. The ventral border is straight and runs parallel to the longitudinal axis of the coracoid. The dorsal and ventral borders diverge gradually from base to tip, the tip attaining about 4/3rd of the width at the base (Figs. 3a,b, 12c). In Tangimoana, a thin bony sheet arises from the scapular spine and merges with the entire dorsal margin of the acromion, adding considerably to the height of the process. The ventral border of the acromion also extends as a thin bony sheet, albeit not as wide, giving the acromion a distinctive ventral ‘kink’ at about midpoint (Fig. 12a). The coracoid process is comparatively uniform in all three specimens. In lateral view, the coracoid is long and straight but strongly built. It diverges ventrad away from the acromion at an angle of about 25º, its longitudinal axis roughly parallel to the glenoid border. The coracoid is almost as long as the acromion but only about 2/3rd in width, the tip also slightly broader (Fig. 3a-b). It is oval in cross section, and tapers gradually towards the tip. Like the acromion, it is also medially offset, running roughly parallel to the acromion: however it remains straight in its entire length (Fig. 3c).
In Hine Reti, the coracoid is not quite as distinctly spatulate, mainly because the bone remains rather broad throughout. The coracoid process in Purakaunui and Tangimoana correspond well to the general description, particularly when taking the cartilaginous tip into account (Figs. 3d, 12a). In ventral view, the glenoid cavity is fairly shallow and slightly ellipsoid. There is a distinct glenoid nipple present (Fig. 3c). The glenoid border rises towards the longitudinal axis of the scapula at roughly 20º.

B e r a r d i u s a r n u x i i
114
Fig. 3c-d. Acromion process, coracoid process and cartilaginous border (c) ventral view onto glenoid cavity of left scapula, Purakaunui; (d) lateral view of left scapula, Tangimoana; horizontal dashed line: boundary of bony vertebral border; vertical dashed lines: anterior and posterior tendinous border of (Inf) infraspinatus
The cartilaginous vertebral border attaches onto the bony vertebral border as a broad, strong and tough ribbon (Figs. 3d, 12a). During dissection it detached, and it was therefore not possible to undertake accurate measurements. Cranially the cartilaginous border rises at a steep, caudo-dorsally slanting angle to about 4.5 cm
(Purakaunui: females a little less) above the bony border, from where it continues relatively level to the caudal-most point. Here it initially overshoots the bony border before turning sharply ventro-craniad and subsequently turning again ventrad, approaching the bony border at this point at a right angle (relative to the scapular longitudinal axis). The confluence of dense fibrous tissue, bony and cartilaginous elements at the caudal angle gives this area a peculiar, rectangular appearance, and gives the caudal cartilaginous protuberance of the vertebral border the impression of a broad finger pointing backwards. Long bones
The humerus (Fig. 4a,g) and radius are about equally long in the Arnoux’s beaked whale. The hemispherical head of the humerus is moderately large and, in lateral view, the diameter does not exceed that of the distal epiphysis. The greater tubercle is only little separated by a shallow sulcus from the lesser tubercle further caudad. There is a pronounced crista leading towards the deltoid tuberosity.

B e r a r d i u s a r n u x i i
115
In Purakaunui, proximal epiphyses are only moderately firmly fused to the diaphysis (sensu Galatius and Kinze, 2003), while fusion of the distal epiphyses appears more advanced, although still faintly visible. In Tangimoana and Hine Reti, proximal epiphyses separated during preparation. In the slightly older Tangimoana fusion of distal epiphyses appeared more advanced (moderately firm) than in Hine Reti where
they were mostly cartilaginous and only weakly attached. Dissection revealed a synovial elbow joint with joint capsule, synovial fluid and articular cartilage. The two facets of the humerus opposing the radius and ulna converge in lateral view in the shape of a symmetrical V at an angle of about 135º. In proximal view the two facet elements are broadly-oblong and along the central line slightly dished. The ulnar facet extends slightly onto the posterior edge of the diaphysis forming a small facet towards the olecranon. Laterally, along the caudal border of the diaphysis, a markedly rugose triangular area corresponds to the origin of the humeral head of the triceps. A distinct deltoid tuberosity arises about halfway along the anterior humeral shaft. The radius (Fig. 4b,h) is long and straight, the caudal border narrows slightly mid-shaft but expands again towards the proximal epiphysis; length / width-ratio of the diaphysis is about 2.5:1. In Purakaunui the proximal epiphysis is firmly ossified and fused to the shaft, albeit the fusion line still visible. In Hine Reti the epiphyses are not entirely ossified and were easily detachable. Tangimoana shows an intermediate state with the epiphyses fully ossified but not quite as firmly fused to the diaphysis as seen in Purakaunui. In all three animals, the articular surface is in proximal view oblong and slightly dished, perfectly matching that of the humerus. The distal epiphyses are only in Purakaunui and Tangimoana fully ossified, albeit slightly less fused in Tangimoana. In Hine Reti the radiographs revealed oblong ossification centres surrounded by
cartilage (Plate III - Fig. 1). The ulna is fairly robust and straight (Fig. 4c,i); length / width-ratio of the diaphysis from the narrowest proximal point to the widest distal point is between 6:1 (Purakaunui) and 5:1 (Tangimoana). Proximally the ulnar shaft widens into a flange attaining a length / width-ratio of about 1.9 (Hine Reti) to 2.3 (Purakaunui) to support the moderately long cartilaginous part of the olecranon. The proximal articular facet is slightly smaller than that of the radius and in lateral view straight. Caudally the olecranon follows the curvature of the humeral facet onto the posterior edge of the humerus where it also
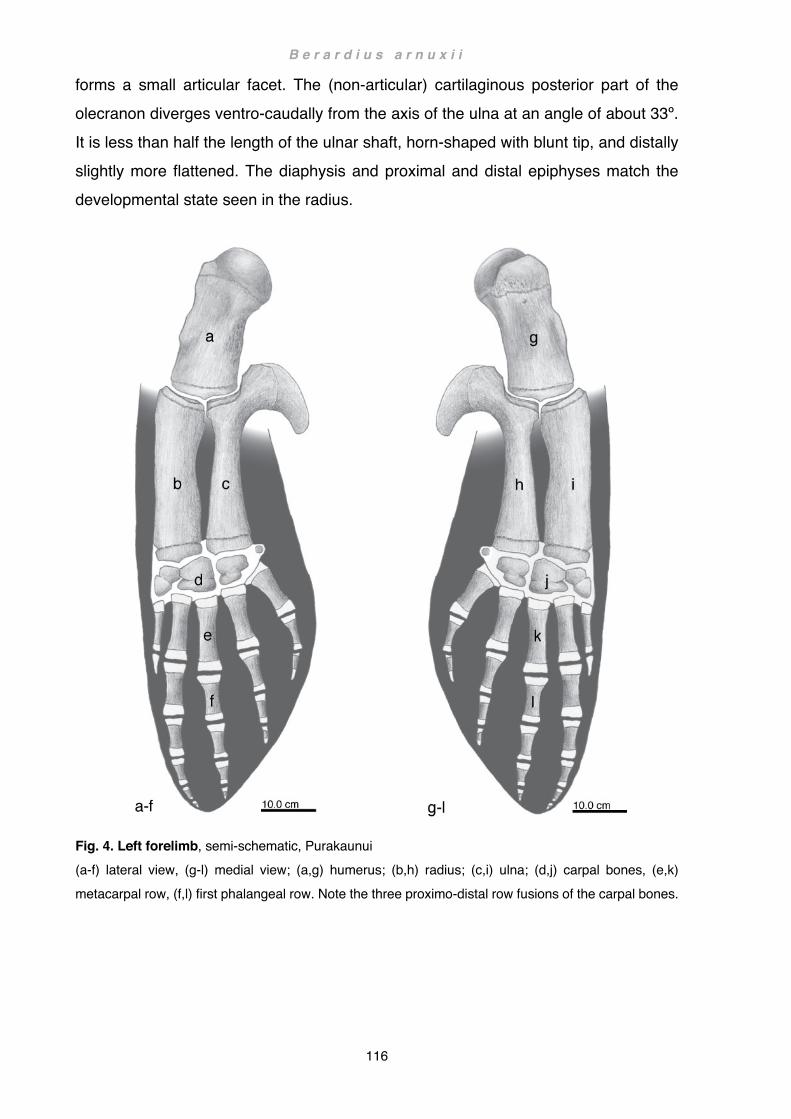
B e r a r d i u s a r n u x i i
116
forms a small articular facet. The (non-articular) cartilaginous posterior part of the olecranon diverges ventro-caudally from the axis of the ulna at an angle of about 33º. It is less than half the length of the ulnar shaft, horn-shaped with blunt tip, and distally slightly more flattened. The diaphysis and proximal and distal epiphyses match the developmental state seen in the radius.
Fig. 4. Left forelimb, semi-schematic, Purakaunui (a-f) lateral view, (g-l) medial view; (a,g) humerus; (b,h) radius; (c,i) ulna; (d,j) carpal bones, (e,k) metacarpal row, (f,l) first phalangeal row. Note the three proximo-distal row fusions of the carpal bones.

B e r a r d i u s a r n u x i i
117
Manus
In all three investigated specimens the manus is slightly shorter than the brachium. There are seven or possibly eight carpals present (Fig. 4d,j). Proximal carpals can be readily identified as radiale, intermedium and ulnare. Distal elements include carpal 1 (C1), ostensibly a fusion of carpals 2 and 3 (C2+3) and what is generally regarded as fused carpals 4 and 5 (C4+5; Flower, 1885; Turner, 1885; Eales, 1954; Rommel, 1990; Cooper et al., 2007a), and a pisiform. However, fused C4+5 is somewhat shorter than C2+3 and articulates predominantly with metacarpal IV (MC IV): thus it is here regarded as a singular C4 - albeit variably fused with other carpal elements (see discussion below). The pisiform is distinguishable in all three animals but only in Tangimoana bilaterally ossified (Fig. 5c,d). Carpal fusions are present in all four flippers and show distinctive fusion patterns. In Purakaunui the three carpal bones of the proximal row are fused with those of the distal row, i.e. radiale with carpal 1, intermedium with carpal 2+3 and ulnare with carpal
4 (Fig. 5a). Alternatively, a slight protrusion at the caudo-distal edge of the ulnare may be interpreted as a fused carpal 5 (Fig. 5a arrow, see also Chapter 4 Functional anatomy: Osteology, Fig. 16d). A U-shaped fusion of the ulnare with carpal 4 via a possibly vestigial carpal 5 is bilaterally present in the Tangimoana (Fig. 5c,d arrow and dashed lines) but only the left flipper appears to have an additional small fusion of the intermedium with the ulnare (Fig. 5c arrow). A broad fusion between intermedium and ulnare can be detected in Hine Reti. Distal carpal 1, however, remains entirely cartilaginous (Fig. 5b). Considering the incomplete ossification of the carpal bones in this animal, it is possible that perhaps carpals 2+3 and 4 would have also fused with those of the proximal row, and a potentially vestigial carpal 5 may have become more apparent at a later stage in life.

B e r a r d i u s a r n u x i i
118
Fig. 5 a-b. Carpal arrangement, radiograph (a) left flipper, Purakaunui; (b) right flipper, Hine Reti. Abbreviations: (1-4) distal carpals 1-4; (I and V) metacarpals I and V; (int) intermedium; (ra) radiale; (ul) ulnare; dashed line: border between fused carpals or border of carpal 1 (not ossified) in Hine Reti; arrow: possible vestigial carpal 5.
Fig. 5 c-d. Carpal arrangement radiograph (c) left flipper and (d) right flipper, Tangimoana. Abbreviations: (1-4) distal carpals 1-4; (I and V) metacarpals I and V; (int) intermedium; (pi) pisiform; (ra) radiale; (ul) ulnare; arrows with dashed lines: fusion between ulnare and carpal 4 via possibly vestigial carpal 5; arrow without lines: fusion between ulnare and intermedium.
All five metacarpals are hourglass shaped (Fig. 4e, k), as are most of the phalanges (or nearly so), except some more distal elements, which are either rounded bones or cartilaginous (Fig. 4f, l). Phalangeal count of bony elements in Purakaunui is 0/5/4/3/2: the most distal bony element of digit 2 (phalanx 5) appears as a small, oblong ossification centre, tipped on its side by the curled edge of the flipper; dissection also
revealed additional cartilaginous phalangeal elements, 8-10 mm long, completing digits 1, 3, 4 and 5. It is possible that there is also a tiny additional finger element in digit 2, but the hardened peripheral tissue of the flipper made confirmation impossible. Phalangeal counts of Tangimoana and Hine Reti are identical in both animals with

B e r a r d i u s a r n u x i i
119
0/4/4/3/2. In Tangimoana the distal-most bony elements of digits 3 and 4 (phalanges 4 and 3) remain rounded whereas in Hine Reti rounded elements occur in digits 2, 3 and 4 (phalanges 4, 4 and 3). Here too, dissection revealed an additional cartilaginous phalangeal element in digits 2 to 5. However, the metacarpal of digit 1 terminates in a round, truncated epiphyseal cartilage. The general phalangeal count of all digital
elements in B. arnuxii is thus: 0-1/5-6?/5/4/3.
Myology
Panniculus carnosus
Not investigated Muscles of the shoulder and forelimb
All details given for Berardius arnuxii are based on the dissections described in Chapter 1 Methods and materials: Dissections. The results are illustrated below in Figs. 6-7.
Unless otherwise stated, general muscular arrangements and actions in Cetacea apply to Berardius arnuxii (see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscle distribution, and Chapter 5 Literature review: Myology, Table 2).Identification and terminology are predominantly based on Strickler, 1978 and Cooper et al., 2007b, and muscle action on Benke, 1993 or Smith et al., 1976.
Overleaf: Fig. 6. Origin and insertion of the lateral forelimb musculature Semi-schematic illustration, composite of Purakaunui and Tangimoana Areas in light grey represent bone, medium grey (long bones) and white (manus) cartilage, and dark grey corresponds to the silhouette of the flipper. Abbreviations: (1-5) digits 1-5. Note fenestration of the extensor digitorum communis tendon over the carpals and short slip over the joint of metacarpal II and first phalanx. Overleaf: Fig. 7. Origin and insertion of the medial forelimb musculature Semi-schematic illustration, composite of Purakaunui and Tangimoana Areas in light gray represent bone, medium grey (long bones) and white (manus) cartilage, and dark grey corresponds to the silhouette of the flipper. Abbreviations: (1-5) digits 1-5. Note slip of flexor digitorum communis tendon over the joint of metacarpal II and first phalanx. Colour index of muscle groups (sensu Strickler, 1978): brown, costo-spino-scapular; red+orange, latissimus-subscapular; purple, deltoid; yellow, triceps; green, suprascapular; blue, flexor; turquoise, trapezius; pink, manus.

B e r a r d i u s a r n u x i i
120
Fig. 6. Origin and insertion of the lateral forelimb musculature
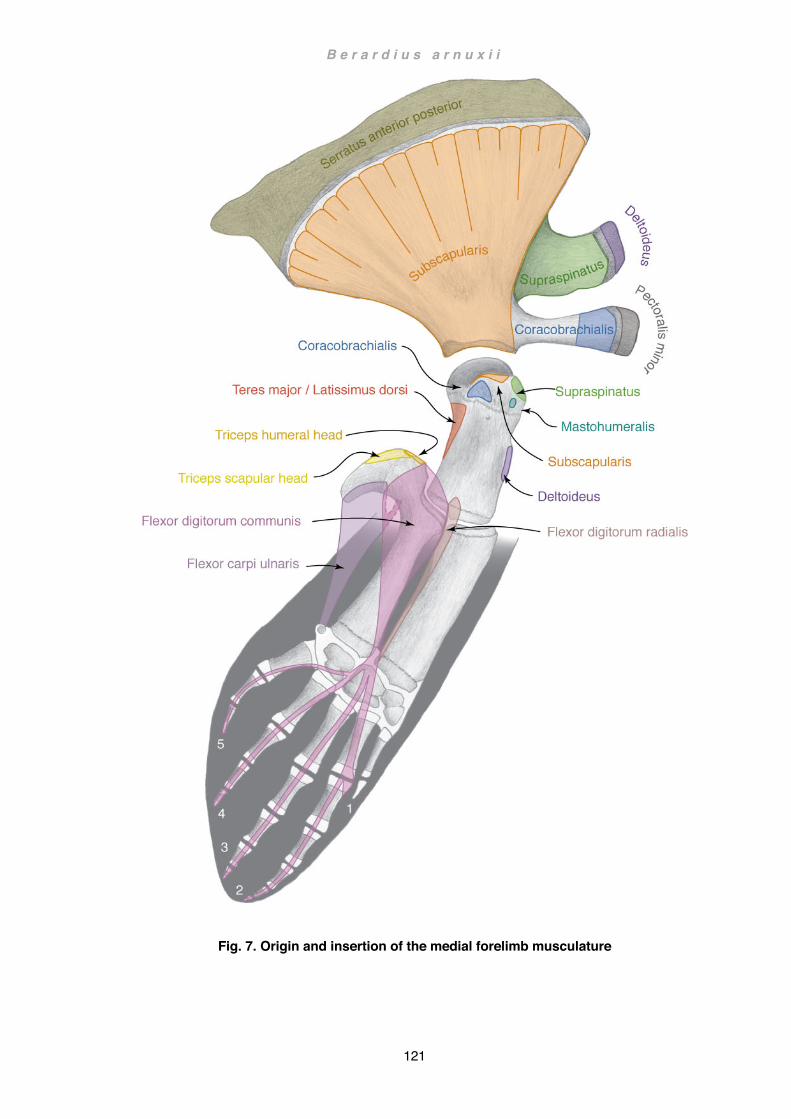
B e r a r d i u s a r n u x i i
121
Fig. 7. Origin and insertion of the medial forelimb musculature

B e r a r d i u s a r n u x i i
122
Levator scapulae
Berardius arnuxii General cetacean condition
Origin Absent? Ventral arch of atlas Insertion Absent? Lateral cranial angle of scapula
Aponeurotic on deltoid Action Slight craniad movement of scapula
Not identified but could have been removed during field recovery. Some fibres attached to the lateral surface of the scapular cranial angle on either shoulder of Tangimoana could not be unambiguously identified. In consequence, misidentification with parts of the panniculus cannot not be excluded. However, no similar fibres were observed in the other two specimens, particularly not in Purakaunui where a hasty personal field observation was possible before the removal of the limb. Furthermore, this muscle also appears to be absent in Mesoplodon layardii and Ziphius cavirostris (personal
observations). For these reasons the levator scapulae is here regarded as absent in ziphiids (see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscle distribution). Serratus anterior - anterior division
Berardius arnuxii General cetacean condition (if present)
Origin Absent? Transverse process of C7 Insertion Absent? Medial cranial angle of scapula Action Slight craniad movement of scapula
Described by Strickler (1978) as absent in almost all odontocetes; could not be identified in B. arnuxii (see serratus anterior - posterior division below) and is here regarded as absent. Serratus anterior - posterior division (Fig. 7)
Berardius arnuxii General cetacean condition
Origin Not identified 2-4th ribs Insertion Medial cartilaginous vertebral border Medial cartilaginous vertebral border of scapula Action Adducts scapula ventrad, rotation craniad
A thin sheet of muscle, inserting into the medial aspect of the cartilaginous vertebral border of the scapula, extends from behind the slanting cranial edge near the cranial angle to the tip of the caudal protuberance at the caudal angle. It stretches out as a

B e r a r d i u s a r n u x i i
123
long, narrow strip between the top edge of the cartilaginous border and the subscapularis. No clear divisions could be identified, so this band of muscle may also include the anterior division of the serratus, but without a definite area of origin identification could not be unambiguously determined.
Rhomboideus capitis
Berardius arnuxii General cetacean condition (if present)
Origin Absent? Supraoccipital Insertion Absent? Lateral cranial angle of scapula Action Slight craniad movement of scapula (Klima et
al., 1980)
As with the levator scapulae, this muscle could have been removed during field recovery: however it too appears to be consistently absent in both Mesoplodon layardii
and Ziphius cavirostris and is here also regarded as absent in Arnoux’s beaked whales (personal observations, see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscle distribution). Rhomboideus vertebralis (Fig. 6)
Berardius arnuxii General cetacean condition
Origin Not identified Epaxial fascia between T1-6 Insertion Lateral cartilaginous vertebral border Lateral vertebral border of scapula Action Adducts scapula dorsad
This muscle inserts laterally as a long, narrow strip onto the whole length and width of the scapular cartilaginous vertebral border. No divisions were identified. Latissimus dorsi (Fig. 7)
Berardius arnuxii General cetacean condition
Origin Not identified 4-6th ribs Insertion Medio-caudal on humerus neck Mid medio-caudal edge of humerus Action Flexes humerus
The latissimus dorsi is in the area of insertion intimately intermingled with the fibres of the teres major. Here the teres major inserts into a more proximal part, whereas the latissimus dorsi inserts into the more distal part of the medio-caudal region of the humeral neck just below the humeral head.

B e r a r d i u s a r n u x i i
124
Teres major (Figs. 6, 7)
Berardius arnuxii General cetacean condition
Origin Caudal 1/4th of lateral scapular blade Caudal border of scapula Insertion Medio-caudal on humerus neck Mid medio-caudal edge of humerus Action Flexes and/or rotates humerus inwards
Arises from the caudal quarter of the lateral scapular surface, also including the caudal border and intermuscular fascia in common with the subscapularis. The teres major inserts just above, but in common with, the latissimus dorsi onto the medio-caudal surface of the humeral neck. Subscapularis (Fig. 7)
Berardius arnuxii General cetacean condition
Origin Medial surface of scapula Medial surface of scapula Insertion Lesser tubercle of humerus Lesser tubercle of humerus Action Adducts and/or rotates humerus inwards
Occupies the entire (medial) subscapular fossa and intermuscular fascia between the caudal angle and posterior glenoid border in common with the teres major. The subscapularis does not encroach onto the cartilaginous surface of the vertebral border. Radiating from the vertebral border, six to seven larger and multiple smaller incomplete tendinous intersections divide the muscle into 7 or 8 slips. Converging into a flat
tendon, the subscapularis is firmly attached to the capsule of the shoulder joint and inserts onto the proximal surface of the lesser tubercle just above the more distal insertion of the coracobrachialis. Deltoideus (Figs. 6, 7)
Berardius arnuxii General cetacean condition
Origin Cranial ½ of lateral scapula surface Cranial lateral surface of scapula Insertion Lateral surface of humerus Lateral surface of humerus Action Abducts and/or extends and reverses inward
rotation of humerus
The deltoideus arises from the anterior half of the lateral scapular surface, the cranial angle, the acromion, and the thick fascia spanning between the latter two (prescapular fascia). Caudally the deltoideus overlaps the infraspinatus onto which it is firmly attached by aponeurosis. At the cranial border the deltoideus completely overlies and

B e r a r d i u s a r n u x i i
125
obscures the supraspinatus, also attaching at the tip of the acromion. Ventrad of the acromion, the deep surface of the deltoid is attached to the intramuscular fascia between it and the supraspinatus. Distally the coarse fascicles of the deltoideus convert into a tough flat tendon, which initially appears on the surface of the muscle. At their insertion, the tendinous fibres broadly attach to the anterior border and preaxial
lateral surface of the humerus immediately below the insertion of the infraspinatus. Here they extend from the deltoid tuberosity caudad, narrowing rapidly into a small band across the humeral neck. Under cover of the superficial tendon, the muscle remains fleshy to the point of insertion, demarcated by a triangular area over the cranio-lateral part of the humerus. Teres minor / Subdeltoideus
Berardius arnuxii General cetacean condition (if present)
Origin Absent Mid-lateral surface of scapula Insertion Absent Proximal lateral surface of humerus Action Rotates humerus inwards
Sometimes reported as Subdeltoideus, this muscle is uncommon in cetaceans (Strickler, 1978). In one individual (Hine Reti), a muscle appeared to closely match in size and position the subdeltoideus as described in other odontocetes (e.g. Kogia breviceps Schulte and Smith, 1918 fig. 7). However, further examination revealed that this muscle was continuous with the deltoideus at a deep level. Furthermore, dissections of the other two specimens (Purakaunui, Tangimoana) showed no partitioning of the deltoideus or the presence of any muscle that could have been interpreted as subdeltoideus. For these reasons the subdeltoideus is here regarded as absent in B. arnuxii (see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscle distribution)

B e r a r d i u s a r n u x i i
126
Triceps brachii (Figs. 6, 7)
Berardius arnuxii General cetacean condition (if present)
Origin Caudo-lateral neck of scapula Caudo-lateral edge of humerus Absent
Caudal neck of scapula (long head) Lateral on humerus shaft (external head) Medial on humerus shaft (medial head)
Insertion Olecranon Olecranon Action Flexes and rotates humerus inwards (long head)
Not known (humeral heads)
There are two triceps heads present in B. arnuxii. The scapular (long) head is massive compared to other cetaceans and arises from the postaxial lateral surface of the scapular neck and the capsule of the shoulder joint as a long, broad and fleshy band. Distally the fasciculi appear on the surface as a tough flat tendon surrounding the fleshy interior of the muscle. Further distad, after crossing the teres major laterally, the muscle
inserts onto the proximal edge of the olecranon, caudad to the insertion of the humeral head of this muscle. The second head (external) head arises between the insertions of the deltoideus (lateral) and teres major (medial) as a narrow band slightly displaced towards the lateral plane along the neck of the humerus. A strong tendinous sheet covers a broad and well-developed fleshy muscle belly inserting onto the proximal edge craniad to the scapular head of this muscle. A medial head was not observed. Supraspinatus (Figs. 6, 7)
Berardius arnuxii General cetacean condition
Origin Supraspinous fossa of scapula Supraspinous fossa of scapula Insertion Greater tubercle of humerus Greater tubercle of humerus Action Extends and rotates humerus outwards
Arises from the supraspinous fossa, the prescapular fascia between the cranial angle and the acromion, the medial surface of the acromion, and the medial surface of the coracoid. The muscle, now covered by a tendinous sheet, has a fleshy insertion onto the cranial aspect of the greater tubercle, craniad to the insertions of the infraspinatus (lateral) and coracobrachialis (medial), as well as proximad to the mastohumeralis (medial). Laterally the tendinous fibres covering the muscle directly adjoin the insertion point of the infraspinatus.

B e r a r d i u s a r n u x i i
127
Infraspinatus (Fig. 6)
Berardius arnuxii General cetacean condition
Origin Lateral postaxial 1/4 of scapula Caudo-lateral surface of scapula Insertion Proximo-lateral on humerus Cranio-lateral surface of humerus Action Abducts humerus
Interposed between the deltoideus in front and the teres major at the back, the infraspinatus covers about a quarter of the lateral scapular surface but is only partially visible in the gap between the former two. In outline, the anterior part of the infraspinatus is initially only little obscured by the posterior portion of the deltoideus. Descending in a straight line towards the anterior glenoid border, it is joined halfway by a slip, which arises further craniad and is entirely concealed by the deltoid. Both, the slip and the main mass of infraspinatus, then descend under the cover of the deltoideus directly towards the point of insertion. Caudally, the infraspinatus abuts onto
the anterior portion of the teres major but is not obscured by it. It remains in contact with the teres major descending towards the glenoid border. In the region of the neck, it is intercepted by the origin of the scapular head of the triceps. Crossing the shoulder joint laterally, the infraspinatus inserts entirely under cover of the deltoideus as a broad tendinous band with few fleshy fibres onto the lateral surface of the humerus, embracing the head of the humerus in the manner of a collar. Further distad, the tendinous fibres extend to the infraspinous fossa of the humerus. Pectoralis minor (Figs. 6, 7)
Berardius arnuxii General cetacean condition
Origin Not identified Sternum Insertion Tip of coracoid Tip of coracoid process Action Rotates scapula ventrad
The origin of this muscle could not be identified but a severed muscle belly inserting onto the tip of the coracoid perfectly corresponds with the description of the pectoralis minor in other cetaceans.

B e r a r d i u s a r n u x i i
128
Pectoralis major
Berardius arnuxii General cetacean condition
Origin Not identified Sternum Insertion Medial base of flipper Medial on flipper Action Adducts and flexes humerus
The origin of this muscle could not be identified. A broad, thin sheet of musculature inserting across the medial base of the flipper is consistent with the description of the pectoralis major in other cetaceans. However, preservation state and the fragmentary condition of the material made it difficult to distinguish between remnants of the panniculus and those of the pectoralis major. Pectoralis abdominalis (not depicted)
Berardius arnuxii General cetacean condition
Origin Absent? Coastal cartilage 4-6 Insertion Absent? Medial, posterior base of flipper Action Flexes humerus (inferred sensu Strickler, 1978)
Not identified but could have been removed during field recovery. This muscle occurs only sporadically in cetaceans (Strickler, 1978) and is possibly absent in B. arnuxii (see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscle distribution).
Coracobrachialis (Figs. 6, 7)
Berardius arnuxii General cetacean condition
Origin Coracoid process Medial surface of coracoid process of scapula Insertion Lesser tubercle Just distal to lesser tubercle of humerus Action Adducts and slightly flexes humerus
The coracobrachialis arises as a round and short but strong muscle immediately behind the tip of the coracoid process and adjacent surface further caudad, occupying
roughly the medial, cranial half of the coracoid process. Crossing the subscapularis mediad, the muscle inserts as a tendon onto the posterior aspect of the lesser tubercle, flanked by the insertions of the subscapularis proximad and the supraspinatus and mastohumeralis craniad.

B e r a r d i u s a r n u x i i
129
Omohyoideus
Berardius arnuxii General cetacean condition
Origin Absent Aponeurotic on cranial angle of scapula Insertion Absent Hyoid Action Craniad movement of scapula (or possibly
caudad movement of the hyoid respectively)
It is possible that the omohyoideus was removed during field recovery, but no traces were found at the expected point of origin. Mastohumeralis (Fig. 7)
Berardius arnuxii General cetacean condition
Origin Not identified Exoccipital Insertion Greater tubercle Mid-cranial edge of humerus Action Extends and rotates humerus inwards
Origin of the mastohumeralis could not be established; however, the muscle inserts as a round tendon onto the medio-anterior aspect of the greater tubercle, just distad to the insertion of the supraspinatus. Extensors Extensor digitorum communis (Fig. 6)
Berardius arnuxii General cetacean condition (if present)
Origin Lateral interosseus space Lateral interosseus space of radius and ulna Insertion Digits 2-4 Digits 2-5 Action Extension of digits
The extensor digitorum communis, or extensor digitorum ulnaris, arises from the elbow joint and adjacent radial and ulnar surfaces, as well as the membrane ligament in between. The fleshy belly is covered by a thick tendinous sheet and is at its origin
predominantly displaced towards the ulnar surface. Distally, the muscle soon splits and the fibres form two flat tendons about midway along the long bones of the forearm. The radial portion is wider and extends over the radius and interosseus space between radius and ulna. Over the wrist the tendon broadens even further, splaying out widely across the carpals. In the region of the axial carpals, the posterior portion of the tendon splits and re-joins itself a couple of times, creating small fenestrations. Immediately

B e r a r d i u s a r n u x i i
130
distal to this area, the tendon divides into three equally sized slips (preaxial, axial and postaxial digital tendons) inserting into digits 2-4. The preaxial digital tendon is slightly displaced axially over metacarpal II but otherwise completely aligned with the digit. The axial and postaxial digital tendons gain alignment over the metacarpals and remain fully aligned with their respective digits. The tendon
of digit 2 also diverts into a small, thin slip over the cranial part of the first joint between metacarpal and first phalanx. The ulnar portion of the flexor remains restricted to the ulnar surface and postaxial carpals where it curves towards the trailing edge of the flipper to align with digit 5. The digital tendon of the ulnar portion is also slightly axially displaced and gains complete alignment with digit 5 from the first phalanx onwards to its insertion at the tip. Flexors Flexor digitorum radialis (Fig. 7)
Berardius arnuxii General cetacean condition (if present)
Origin Medial postaxial surface of radius Medial postaxial edge of radius Insertion Fused with flexor digitorum
communis Absent or fused with flexor digitorum communis
Action Flexion of preaxial digits
The flexor digitorum radialis arises as a slender muscle from the elbow joint, the medial postaxial surface of the radius, and the intermuscular fascia of the adjacent flexor digitorum communis. The muscular fibres reach down as far distad as the middle of the long bone before converging to a flattish, thin tendon descending towards wrist. Attaining the posterior distal radial epiphysis, it fuses with the more massive tendon of the flexor digitorum communis. From here on, the two tendons remain fused to form a broad tendon extending onto the carpals. Flexor digitorum communis (Fig. 7)
Berardius arnuxii General cetacean condition (if present)
Origin Medial proximal surface of ulna Medial proximal surface of ulna Insertion Digits 2-5 Digits 2-5 Action Flexion of digits
The flexor digitorum communis, or flexor digitorum ulnaris, arises from high up on the olecranon, the medial proximal surface of the ulna, and the elbow joint. Further distad it is also firmly attached to the interosseus membrane and the intermuscular fasciae

B e r a r d i u s a r n u x i i
131
shared with the flexores digitorum radialis anteriorly, and carpi ulnaris posteriorly. The fleshy fasciculi converge about midway on the anterior edge of the ulnar shaft as a thick, flattish tendon, whence they continue distad. At the level of the distal radial and ulnar epiphyses the tendon fuses with the tendon of the radial digital flexor. Immediately distad, over the intermedium, the tendon splits into a wider, anterior
portion and a narrower posterior portion. The anterior portion immediately splits again into two slips aligning with digits 2 and 3 (preaxial and axial digital tendons). The caudal portion diverges, initially undivided, from the anterior portion and splits further distad, at the height of the distal carpal elements, into two slips that insert onto digits 4 and 5 (anterior and posterior postaxial digital tendons). The preaxial digital tendon, emerging from the anterior portion fused with the radial tendon, continues in a straight line diagonally across the carpals and metacarpal II towards the leading edge of the flipper. Reaching the distal epiphysis of metacarpal II, it splits once more. One short but strong slip continues without changing direction and expands fan-like over the metacarpal-phalangeal joint. The other slip, now much narrower, diverges in a longitudinal direction and, following digit 2, inserts onto the tip of the finger. The axial digital tendon is more or less aligned with digit 3 and follows this digit to the tip without interruption. The anterior postaxial digital tendon, emerging from the posterior portion of the ulnar flexor, is only slightly axially displaced over metacarpal IV and follows digit 4 perfectly aligned to the tip. In order to align with digit 5, the posterior postaxial digital tendon initially turns sharply caudad before turning distad. Following the anterior edge of metacarpal V, it gains full alignment about the middle of the first phalanx and continues in a wide arch to the tip of digit 5.
Flexor carpi ulnaris (Fig. 7)
Berardius arnuxii General cetacean condition
Origin Distal olecranon Distal olecranon Insertion Proximal pisiform Proximal carpus Action Not known
The flexor carpi ulnaris arises from the medial surface of the olecranon and the intermuscular fascia to the flexor digitorum communis, including the area from the distal
bony base of the olecranon to the distal cartilaginous edge at the tip. Under cover of the radial flexor, a small slip separates from the main belly of the flexor carpi ulnaris to

B e r a r d i u s a r n u x i i
132
allow passage of a large blood vessel, possibly the collateral ulnar artery or part of the rete articulare cubiti. The smaller slip might also represent the relic of the humeral head of this muscle reportedly present in some other cetartiodactyls (Constantinescu et al., 1992). Distally the fleshy fibres converge into a thick, flat tendon before inserting onto the proximal edge of the pisiform.
In general, the fleshy antebrachial flexor / extensor muscles retain tendon sheaths over the manus; in the region of the carpals they remain comparatively thick and fleshy, and only transform gradually into flat tendons distad of the metacarpals. Laterally, the digital tendons are of about equal width and robustness, whereas medially there is a marked reduction of width and robustness from the anterior-most to the caudal most tendons. All tendons are joined to the perichondrium of the phalanges by fibrous tissue, the fibrous tissue is particularly firm at the enlarged interphalangeal joints where it extends beyond the immediate joint. Dissection also revealed several digital flexor tendons inserting in the connective tissue distal to the terminal phalangeal cartilage. A tiny interosseus muscle was also found at the cranial edge of metacarpal V in Tangimoana.

B e r a r d i u s a r n u x i i
133
Discussion Shoulder and forelimb morphology of Berardius arnuxii are in general congruent with that observed in other odontocetes, with the exception of a few distinctive patterns characteristic for the family and/or the species (see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscle distribution). Flipper shape and associated terminology are discussed in Chapter 1 Methods and materials. Taxonomic considerations are based on Bianucci et al., 2007 for Ziphiidae, and on Geisler et al., 2011 for Cetacea in general (Fig. 1). Taxonomic nomenclature is also discussed in Chapter 1 Methods and materials. Flipper
The three specimens of Berardius arnuxii show body / flipper length ratios (soft tissue) of 10.3-10.9. These ratios are close to that of Mesoplodon grayi (10.6); while the other two beaked whale species investigated in this study (Ziphius cavirostris, Mesoplodon layardii, Fig. 8a), encompass a ratio of about 12. The proportions of the soft tissue are also reflected in the skeletal proportion of total length / forelimb length. In the three B. arnuxii specimens this ratio is 8.5-9.1, and in the other beaked whales 9.1-9.9 (Fig. 8a). Both proportions show that the flippers of B. arnuxii are moderately long and comparable with those of Mesoplodon grayi. These data also roughly agree with other osteological data (Benke 1993, fig. 143), where the proportions seen in B. arnuxii also group with those of other beaked whales (Fig. 9a). The data also reveal that in comparison with odontocetes in general, beaked whale flippers are moderately short. Still, among odontocetes short flippers are not
unique: they can also be found amongst Delphinidae (yellow), Monodontidae (orange) or Physeteridae (grey, Fig. 9a). Similarities also emerge with respect to the ratio of forelimb length / forelimb width, including both soft and hard tissue. In B. arnuxii this ratio is 2.5-2.7 (soft tissue) and 3.7-3.9 (hard tissue) and in the other three beaked whale species 2.8-3.1 and 4.1-4.5 respectively (Fig. 8b). The data indicate that B. arnuxii has slightly broader flippers than the other three species of beaked whales, particularly when compared with

B e r a r d i u s a r n u x i i
134
Mesoplodon species. These values are also congruent with data reported by Benke (1993, fig. 141) except here Ziphius cavirostris seems to have slightly wider flippers than B. arnuxii. The data also show beaked whale flippers as fairly narrow when compared with other odontocetes, a feature they share mainly with some porpoises and dolphins (orange and yellow, Fig. 9b).
a b
c d
Fig. 8. Flipper proportions in selected odontocetes stranded in New Zealand (a) ratio of total length / forelimb length; (b) ratio of forelimb length / forelimb width; (c) humerus length / radius length; (d) brachial length / manus length. (a-b) Left columns soft tissue, right columns hard tissue, (c-d) left columns hard tissue, right columns total length. Colour key: Gray, Kogiidae; Purple, Ziphiidae; Orange, Phocoenidae; Yellow, Delphinidae; Red, Berardius arnuxii specimens, Hine Reti (HiRe), Tangimoana (Tan, average of left and right flipper) and Purakaunui (Pur). For taxonomic nomenclature see Chapter 1 Methods and materials.
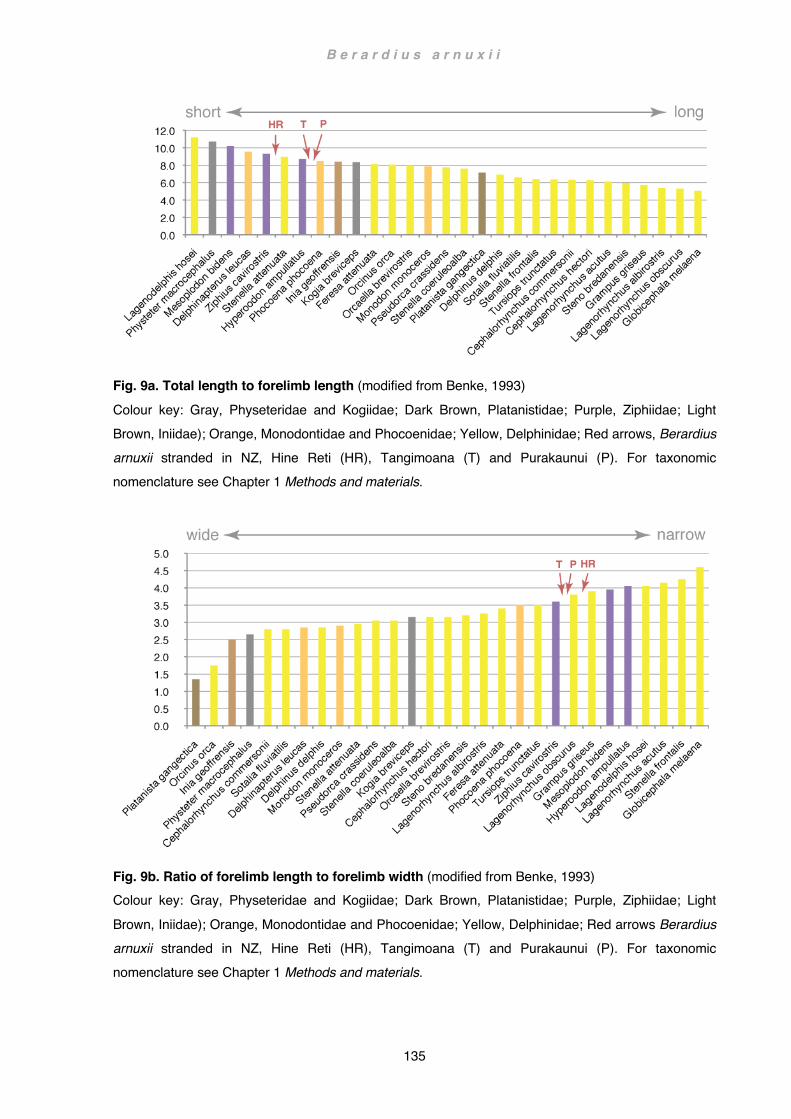
B e r a r d i u s a r n u x i i
135
Fig. 9a. Total length to forelimb length (modified from Benke, 1993) Colour key: Gray, Physeteridae and Kogiidae; Dark Brown, Platanistidae; Purple, Ziphiidae; Light Brown, Iniidae); Orange, Monodontidae and Phocoenidae; Yellow, Delphinidae; Red arrows, Berardius arnuxii stranded in NZ, Hine Reti (HR), Tangimoana (T) and Purakaunui (P). For taxonomic nomenclature see Chapter 1 Methods and materials.
Fig. 9b. Ratio of forelimb length to forelimb width (modified from Benke, 1993) Colour key: Gray, Physeteridae and Kogiidae; Dark Brown, Platanistidae; Purple, Ziphiidae; Light Brown, Iniidae); Orange, Monodontidae and Phocoenidae; Yellow, Delphinidae; Red arrows Berardius arnuxii stranded in NZ, Hine Reti (HR), Tangimoana (T) and Purakaunui (P). For taxonomic nomenclature see Chapter 1 Methods and materials.

B e r a r d i u s a r n u x i i
136
Although beaked whale flippers are similar with regard to both flipper length and width, it is the shape that ultimately separates ziphiid from other odontocete flippers. While a typical dolphin or porpoise flipper (e.g. Phocoena phocoena) may encompass similar values, the shape of the flipper is very different: falcate in most dolphins and fusiform in porpoises; in ziphiids the contour is deltoid (see also Chapter 4 Functional anatomy:
Diversity of flipper shapes). Ziphiid flippers are also quite different from the typical paddle- or fusiform shaped flippers of sperm whales or narwhals (see Chapter 4 Functional anatomy: Diversity of flipper shapes, Fig. 30). Ziphius and Mesoplodon flippers show a tendency towards an oar profile. The almost subrectangular flippers of B. arnuxii on the other hand appear to be broader than in most ziphiids and thus take on a more paddle-shaped outline (Fig. 13a-d), yet ziphiid flippers maintain intermediate length / width proportions. Flipper characteristics are ultimately determined by the proportion and arrangement of limb elements rather than by simple length and width dimensions. A major factor affecting the contour of a flipper is the manus, that is, the proportion of the hand in relation to other limb elements and arrangements of carpal and phalangeal elements (see also Chapter 4 Functional anatomy: Diversity of flipper shapes). Ziphiids, and especially Arnoux’s beaked whales, are distinguished by long brachial elements and a relatively short manus. There are no osteological data for comparison in Benke (1993) but the small sample of specimens from New Zealand nevertheless shows a clear incongruity between ziphiids and non-ziphiids (Fig. 9d). Interestingly, a long humerus does not necessarily translate into overall long arm proportions (e.g. Kogia Fig. 9c,d). Scapula and limb bones
In general, shape and robustness of the scapulae in all beaked whales investigated are quite similar: the scapular blade is in lateral view fan-shaped; the arch of the vertebral border dorso-ventrally compressed; the blade itself is quite sturdy, a relief only vaguely present; the supraspinous fossa is fairly small (in B. arnuxii about 1/14th to 1/25th of the vertebral border length); both the acromion and coracoid are well developed (see also Chapter 4 Functional anatomy: Functional shift of the scapular spine, acromion, coracoid and olecranon).
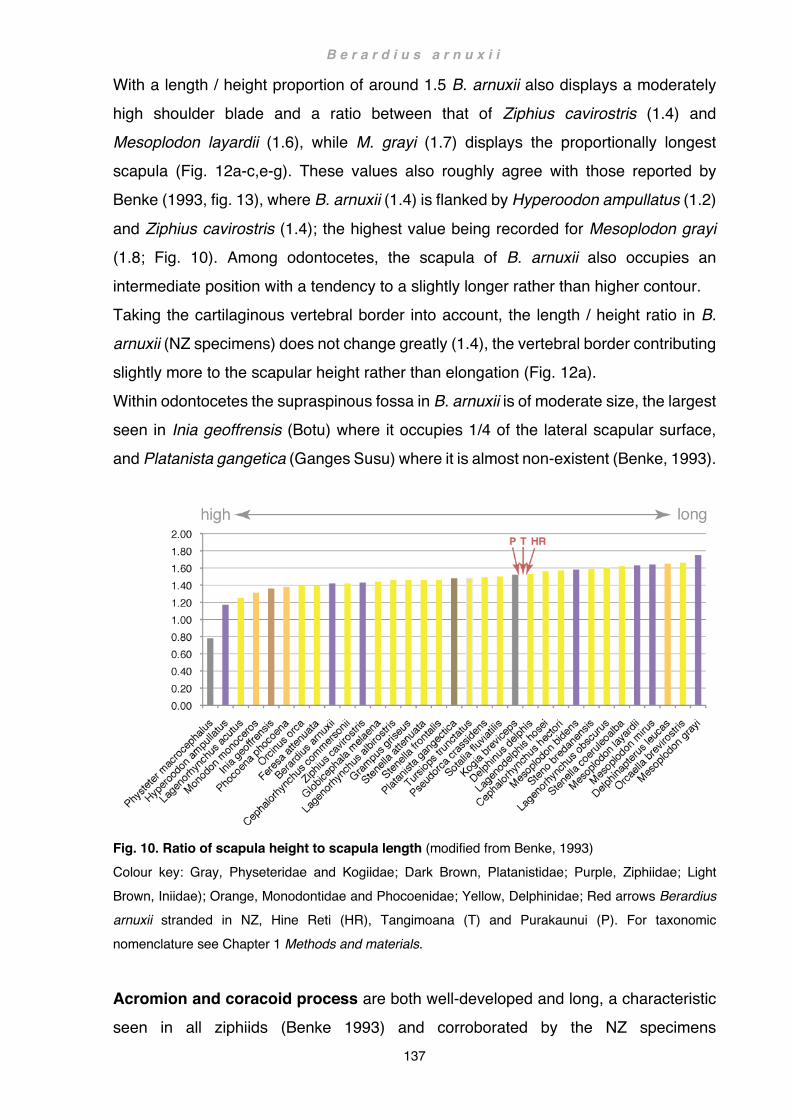
B e r a r d i u s a r n u x i i
137
With a length / height proportion of around 1.5 B. arnuxii also displays a moderately high shoulder blade and a ratio between that of Ziphius cavirostris (1.4) and Mesoplodon layardii (1.6), while M. grayi (1.7) displays the proportionally longest scapula (Fig. 12a-c,e-g). These values also roughly agree with those reported by Benke (1993, fig. 13), where B. arnuxii (1.4) is flanked by Hyperoodon ampullatus (1.2)
and Ziphius cavirostris (1.4); the highest value being recorded for Mesoplodon grayi (1.8; Fig. 10). Among odontocetes, the scapula of B. arnuxii also occupies an intermediate position with a tendency to a slightly longer rather than higher contour. Taking the cartilaginous vertebral border into account, the length / height ratio in B. arnuxii (NZ specimens) does not change greatly (1.4), the vertebral border contributing slightly more to the scapular height rather than elongation (Fig. 12a). Within odontocetes the supraspinous fossa in B. arnuxii is of moderate size, the largest seen in Inia geoffrensis (Botu) where it occupies 1/4 of the lateral scapular surface, and Platanista gangetica (Ganges Susu) where it is almost non-existent (Benke, 1993).
Fig. 10. Ratio of scapula height to scapula length (modified from Benke, 1993) Colour key: Gray, Physeteridae and Kogiidae; Dark Brown, Platanistidae; Purple, Ziphiidae; Light Brown, Iniidae); Orange, Monodontidae and Phocoenidae; Yellow, Delphinidae; Red arrows Berardius arnuxii stranded in NZ, Hine Reti (HR), Tangimoana (T) and Purakaunui (P). For taxonomic nomenclature see Chapter 1 Methods and materials.
Acromion and coracoid process are both well-developed and long, a characteristic seen in all ziphiids (Benke 1993) and corroborated by the NZ specimens

B e r a r d i u s a r n u x i i
138
(Fig. 12a-c,e-g). In B. arnuxii the acromion is also very high throughout its entire length, in Tangimoana and Hine Reti square and in Purakaunui (Fig. 12c) spatula- to spoon-shaped, the latter more resembling that of B. bairdii (Fig. 12d) where this trait is even more accentuated. In M. layardii (Fig. 12f) the tongue-shaped acromion tapers towards the tip whereas in Z. cavirostris (Fig. 12e) and M. grayi (Fig. 12g) the acromion is
subrectangular with a prominent kink, a feature also seen to some degree along the ventral boder in Tangimoana (Fig. 12a). However, the acromion of neither Z. cavirostris nor M. grayi is as high as in Tangimoana, and in M. grayi the tip does not reach as far as the cranial-most point of the scapula (bearing in mind that the scapula of M. grayi is by comparison very long). Overall, the long bones in this study appear fairly homogeneous within B. arnuxii. Benke (1993) regarded the characteristics of the long bones, particularly of the humerus, as species specific and in general more congruent within families. Humeral characteristics of the beaked whales investigated in this study also fit well within his description of this family: the humeral head is comparatively small, the neck not very prominent, there is a crista leading from the well-developed greater tubercle towards the deltoid tuberosity, radius and ulna are long and fairly straight, similar to those of rorquals, there is a large and distinct fan-shaped olecranon, and the distal ends of the humerus, radius and ulna do not flare. A similar-sized fan-shaped olecranon is also present in mysticetes, where it sometimes supports an extensive cartilaginous part. In B. arnuxii the horn-shaped cartilaginous part is comparatively narrow, compact and moderate in size. The overall size of the olecranon in Arnoux’s beaked whales is thus only moderate within cetaceans but large within toothed whales (see also Chapter 4 Functional anatomy: Functional shift of the scapular spine, acromion, coracoid and
olecranon). In the NZ specimens the ratio of humerus length to radius length (0.95 - 1.10) encompasses that given by Benke (1993) for B. arnuxii (1.04; fig. 11). This ratio places the humerus of B. arnuxii, together with other ziphiids, as moderately short among odontocetes, but longer than in most delphinids. A striking plesiomorphy reported here for B. arnuxii is the synovial elbow joint, a characteristic that was also present in the fossil archaeocete Dorudon atrox (Uhen, 2004; see also Chapter 4 Functional anatomy: Fully aquatic archaeocetes). In dorudontids the elbow joint had only restricted mobility but remained functional as a

B e r a r d i u s a r n u x i i
139
hinge joint (Thewissen and Bajpai, 2001). In B. arnuxii this joint has lost almost all flexibility and can no longer be regarded as functional because of the interlocking nature of the long bone joints. Yet, at least slight movement of the joints is regarded as a prerequisite to normal joint development since complete immobilisation would result in joint fusion or malformation (Drachman and Sokoloff, 1966; Murray and
Drachman, 1969). A synovial joint capable of slight jostling movement was also described for Mesoplodon bidens by Turner in 1885 (Turner, 1885).
Fig. 11. Ratio of humerus length to radius length (modified from Benke, 1993) Colour key: Gray, Physeteridae and Kogiidae; Dark Brown, Platanistidae; Purple, Ziphiidae; Light Brown, Iniidae); Orange, Monodontidae and Phocoenidae; Yellow, Delphinidae; Red arrows Berardius arnuxii stranded in NZ, Hine Reti (HR), Tangimoana (T) and Purakaunui (P). For taxonomic nomenclature see Chapter 1 Methods and materials.
The manus is similar in all investigated ziphiids: a row of 3 proximal carpals (radiale, intermedium, ulnare) with a 4th element (pisiform) sometimes not present or only as a cartilaginous element. The distal carpal row is more variable, usually consisting of 3 large carpal elements (C1, fused C2+3 and C4 or C4+5). C1 is cartilaginous in Hine Reti (Fig. 5b). In Purakaunui it is present as a slightly conical, elongated bone fused to the radiale, the two elements demarcated by a notch on either side (Fig. 5a), whereas in Tangimoana this bone is more elongated and rather resembles metacarpal bones (Fig. 5c, d). Similarly elongated first carpal elements are

B e r a r d i u s a r n u x i i
140
not unique and have also been reported for other ziphiids (e.g. Mesoplodon bidens) and delphinids (Turner, 1909). A slight elongation of this carpal element is also present in Ziphius cavirostris and Mesoplodon grayi but not in Mesoplodon layardii from NZ (Fig. 13b-d). Fusion of wrist bone elements, particularly of C2+3, has also been widely observed in
other odontocetes (Turner, 1910) and occurs also in all NZ Berardius specimens (Fig. 13a-d). Other fusion patterns include elements between the proximal and distal row, also seen in Purakaunui (Fig. 5a), or cranio-caudal fusion of carpals within the same row, albeit mostly of distal elements (Turner, 1909). Slightly unusual is thus the fusion of the two proximal elements intermedium + ulnare in Hine Reti (Fig. 5b). Probably more unusual is the distinct U-shaped fusion of the ulnare with C4+5? in Tangimoana (Fig. 5c, d) and in Flower’s New Brighton specimen (Flower, 1872 pl. XVIII fig. 10). Contrary to the usual mammalian three distal carpals, some odontocetes possess five distinct distal carpal units, each articulating with their respective metacarpal (e.g. in Hyperoodon ampullatus C1 with MC I, C2 with MC II etc.; Turner, 1885). It is therefore conceivable that in Berardius arnuxii a reduced C5 persists. In Purakaunui C5 may be present as a knob-like, flattish protrusion at the caudo-distal edge of the ulnare (Fig. 5a). In Tangimoana and in Flower’s New Brighton specimen, C5 may occur as a small, wedge-shaped fusion between ulnare and C4 (Fig. 5c, d). An arrangement very similar to that of Purakaunui has also been reported by Turner for a Sowerby’s beaked whale (Mesoplodon bidens) from Dalgerty Bay, Scotland (Turner, 1909). Here C4 is also cranio-caudally shorter than the fused C2+3 and articulates almost solely with MC IV; MC V articulates directly with the ulnare and pisiform. Turner thus concluded that in this animal, contrary to other M. bidens specimens he investigated, this element has to be regarded as C4, and not as the
more usual fusion of C4+5. This arrangement can also be seen in the NZ specimens of B. arnuxii, although here MC V includes to some degree articulation with the caudal edge of C4. The central part of MC V, however, articulates either with the knob-shaped protrusion of the ulnare or the wedge-shaped fusion between ulnare and C4. A rudimentary C5 is possibly also present in Mesoplodon grayi as a distinct bulge arising from the proximal epiphysis of MC V (Fig. 13c). The general phalangeal count of all digital elements is in B. arnuxii 0-1/5-6?/5/4/3, the distal-most elements often only persisting as cartilage. As in all cetaceans with broad

B e r a r d i u s a r n u x i i
141
flippers, the first digit is the most appressed digit (onto digit 2), digits 3 to 4 are successively more spread apart, and digit 5 not straight but widely arched caudad. Musculature
Most muscles correspond with the general condition and topography in odontocetes (Chapter 4 Functional anatomy: Phylogenetic patterns of muscle distribution). Exceptions include the absence of the levator scapulae, and a triceps brachii retaining a powerful scapular head in addition to the humeral head. In Mesoplodon bidens the triceps is reported to have retained three heads (Turner, 1885). Fleshy antebrachial flexor / extensor muscles with tendon sheaths over the manus have also been observed in Mesoplodon bidens (Struthers, 1873; Turner, 1885). However, the extensor muscles in M. bidens are ostensibly much more fenestrated than in Berardius arnuxii. Furthermore, only a slip of the flexor digitorum radialis fuses distally with the flexor digitorum communis: the anterior portion extends over digit 1. In B. arnuxii the flexor digitorum radialis fuses completely with that of the flexor digitorum
communis and there is no tendon over digit 1. A synovial joint (hinge joint) and extensive antebrachial flexor and extensor muscles have also been proposed for the archaeocete Dorudon atrox (Uhen, 2004). Although Dorudon is not a direct ancestor of modern cetaceans and has a somewhat different arrangement of the forelimb, a synovial joint and functional musculature are both plesiomorphic characteristics that are also seen in ziphiid flippers, albeit reduced. Arrangements of soft tissue in fossils can hereby be inferred from correlated hard tissue structures in extant aquatic or semiaquatic quadrupeds. (Howell, 1937; Uhen, 2010; Geisler et al., 2011; Gatesy et al., 2012; Gingerich, 2012; see also Chapter 4 Functional anatomy: Evolution of the forelimb and Functional aspects of flipper shapes). Notable are also several digital flexor tendons inserting into the connective tissue distal to the terminal phalangeal cartilage. Noteworthy is also the large teres major encroaching onto the caudal border of the scapula, displacing the infraspinatus further craniad. In consequence the large and powerful deltoid muscle arises to a large degree from the prescapular fascia between the cranial angle and the acromion, and from the acromion process itself, both acting as an additional surface for the deltoid muscle to adhere to.

B e r a r d i u s a r n u x i i
142
Spatial requirements to accommodate the deltoideus could therefore be one possible explanation for the extraordinary length of the acromion process in ziphiids (see also Chapter 4 Functional anatomy: Functional shift of the scapular spine, acromion, coracoid and olecranon). A long acromion process would also increase the angle of insertion of the anterior deltoid fibres onto the humerus. An increased angle of insertion
results in a higher rotational component and thus a stronger pulling force (Gregory, 1912). This might be an important factor for the whale to overcome intense drag forces acting on a bulky flipper, particularly when abducting and extending the flipper forward against the flow of the water. A short but well-developed coracobrachialis muscle and the presence of the pectoralis minor could conversely explain a long and compact coracoid process to efficiently stabilise the shoulder joint when the flipper is extended.
Overleaf: Fig. 12. Shape variation of ziphiid scapulae Semi-schematic drawings of (a,b) right and (c-g) left ziphiid scapulae, lateral views (a) Berardius arnuxii, Tangimoana; (b) Berardius arnuxii, Hine Reti; (c), Berardius arnuxii, Purakaunui, (d) Berardius bairdii, Tokyo Museum, (e) Ziphius cavirostris, NZ, (f) Mesoplodon layardii NZ, (g) Mesoplodon grayi NZ. Key: bone: light grey; cartilage: dark grey. Drawings not to scale. Abbreviations: (NZ) specimens from New Zealand, see Chapter 1 Methods and materials: List of specimens. 12(d) based on Photograph by R. E. Fordyce
Overleaf next page: Fig. 13. Shape and skeletal arrangement of ziphiid flippers Semi-schematic drawings of four beaked whale flippers. Left flipper, lateral view of (a) Berardius arnuxii Purakaunui; (b) Ziphius cavirostris, NZ (reconstructed from fractured specimen); (c) Mesoplodon grayi, NZ; (d) Mesoplodon layardii, NZ. Abbreviations: (NZ) specimens from New Zealand, see Chapter 1 Methods and materials: List of specimens.
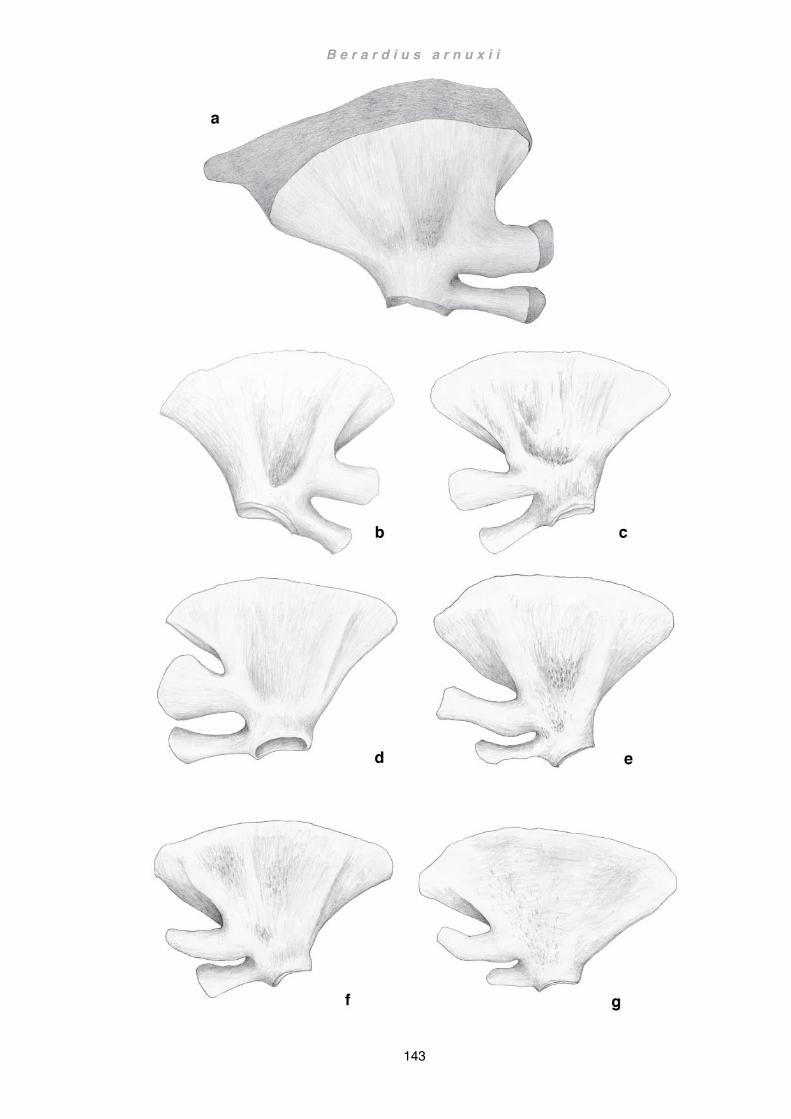
B e r a r d i u s a r n u x i i
143
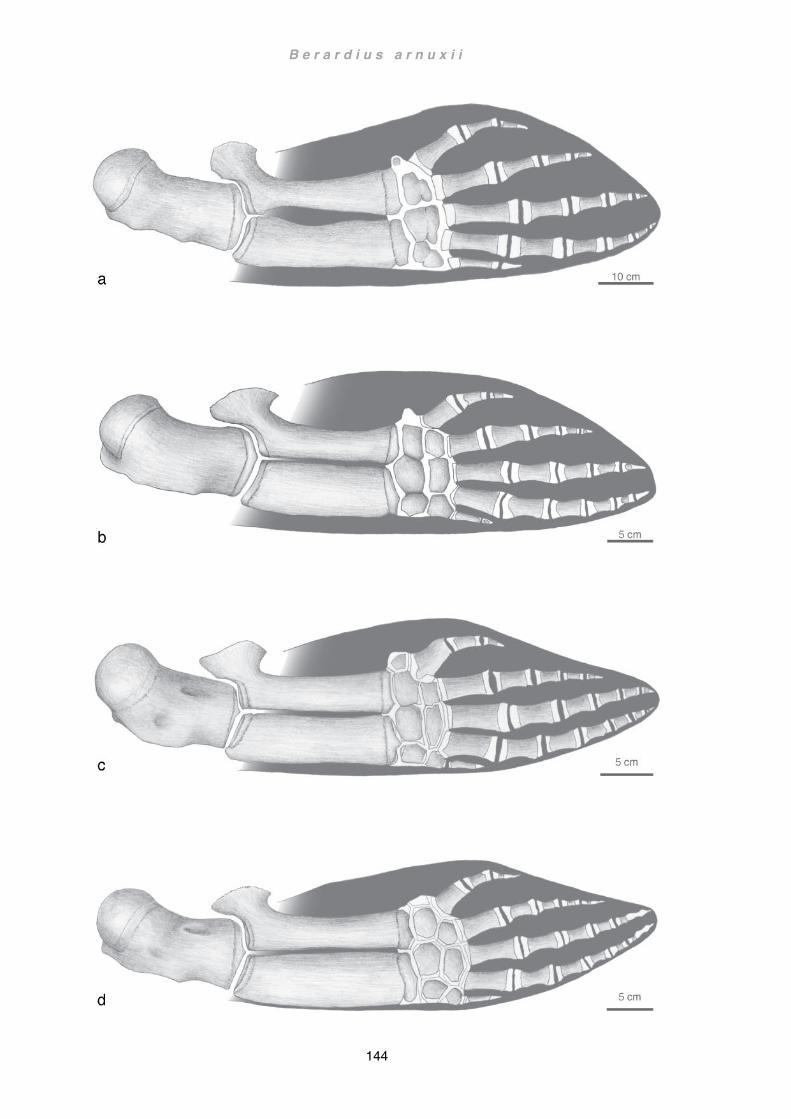
B e r a r d i u s a r n u x i i
144

B e r a r d i u s a r n u x i i
145
Wider implications
Beaked whales are arguably the most extreme diving mammals (Quick et al., 2020). They feed on mesopelagic, bentho-pelagic or batho-benthic fish and squid (MacLeod et al., 2003; Madsen et al., 2013). Some species are capable of diving to a depth of almost 3000 m, some can remain submerged for over 2 hrs, and routinely dive to over 1000 m depth (Schorr et al., 2014). To do so they have evolved a range of highly specialised adaptations such as suction feeding (Heyning and Mead, 1996), specialised echolocation (Johnson et al., 2004; Madsen et al., 2005; Johnson et al., 2008), and anatomical, physiological and behavioural adaptations to deep diving (e.g. Tyack et al., 2006; Minamikawa et al., 2007; López et al., 2015; Williams et al., 2015). All these adaptations stand in contrast to a seemingly archaic flipper morphology (Benke, 1993; Uhen, 2004; Sanchez and Berta, 2010; Gatesy et al., 2012; see also Chapter 4 Functional anatomy: Functional aspects of flipper shapes):
- The flipper is dorso-ventrally little compressed, moderately elongated and with only
a slightly curved leading edge - Synovial elbow joint (albeit interlocked) - Greater and lesser tubercle well developed on humerus
- Ratio humerus / radius ³ 1
- Humerus head projects over the ulnar edge - The long bones are not flared at distal ends - Fleshy antebrachial musculature with large interosseus gap between radius and
ulna - Fan-shaped olecranon - Functional tendons of the manus - Broad arrangement of digits in Berardius and to a lesser degree in other ziphiids
However, Gerstner (1999) demonstrated that morphological differences among perch-like fishes (Ord. Perciformes, Cl. Actinopterygii) are more often correlated with behavioral differences than with locomotor performance alone, and in this respect the diving behavior of beaked whales appears to be even more extreme than that of the deep diving-sperm whales. Sperm whales perform similar diving feats, but their larger body size also acts as a much bigger oxygen reservoir. The much smaller beaked

B e r a r d i u s a r n u x i i
146
whales on the other hand exceed their aerobic dive limit by as much as two times and require a much longer recovery time to metabolise accumulated excessive lactic acid (Tyack et al., 2006; Minamikawa et al., 2007). Additionally, sperm whales optimise their foraging time by near vertical descent and ascent angles (Watwood et al., 2006), whereas beaked whales dive at a much shallower angle, particularly when ascending
(Tyack et al., 2006; Arranz et al., 2011). As a consequence, beaked whales spend day and night either on recovery or on foraging dives (Schorr et al., 2014). Reducing the costs of diving is therefore paramount. One adaptation is the indent on the flank to accommodate the flippers (True, 1910; Heyning and Mead, 2009; Pitman, 2009). Retracted, the flipper is smoothly incorporated into the body contour and offers little resistance to the water flow. Although not documented, it is possible that beaked whales may not use their flippers, or use them only rarely, while diving - in other words, there is no need for the flipper itself to be streamlined. Different diving behaviors between sperm whales and beaked whales may also impact on their social structure (and/or vice versa). Sperm whales live in small groups (Acevedo-Gutiérrez, 2009; Whitehead, 2009) and, when under attack from killer whales, form rosettes with the head at the center and the tail pointing outwards or vice versa. They also leave this formation to assist a distressed group member back into the formation (Pitman et al., 2001; Gemmell et al., 2015; Whitt et al., 2015). Beaked whales also congregate in mostly smaller groups (Acevedo-Gutiérrez, 2009) but are reputedly the group to abandons a stricken group member when harpooned by whalers or when a distressed individual to dives deep (Pike, 1953). Like sperm whales, they are also reported to be a target of killer whale attacks (Jefferson et al., 1991; Gemmell et al., 2015; potential bite marks from killer whales on Hine Reti). Against such an
attack a single individual is almost defenseless, particularly when recovering from a deep dive, when vertical flight into the deep or horizontal flight close to the water surface is physiologically impossible (Tyack et al., 2006; Minamikawa et al., 2007). This is supported by the only hitherto observed and documented orca attack on a beaked whale (Wellard et al., 2016). For beaked whales remaining undetected is consequently the most important factor (Aguilar de Soto et al., 2012) and fight or flight strategies, i.e. the need to be either fast or have a good defense strategy, are of little importance. In sperm whales, a short and broad flipper is ideal to assist positioning within the close proximity of individuals in a rosette. Pilot whales on the other hand are

B e r a r d i u s a r n u x i i
147
fast-swimming hunters (Aguilar de Soto et al., 2008) with compressed, streamlined falcate flippers. Although persecuted by killer whales (Jefferson et al., 1991) they have nevertheless been documented to also mob orcas successfully (de Stephanis et al., 2014). Some beaked whales might be able to hide behind ice floes in higher latitudes, a strategy that would favor broad flippers similar to those of beluga (Kleinenberg et al.,
1969). It is nonetheless questionable if these occurrences would be frequent enough to select for adaptations in the flipper. As a result, flipper morphology may not be strongly influenced by either diving strategies or predation pressure. Thus possible selective pressures on the flippers have to be explained otherwise. Beaked whales most likely don’t use their flippers when travelling vertically through the water column, but rather when catching prey at depth: just before prey capture a Blainville’s beaked whale (Mesoplodon densirostris) has been shown to perform a body roll (Madsen et al., 2005, fig. 5A). It is not clear if this roll is in the manner of ‘capsize and right’ on the same side or a barrel roll, but is nevertheless a manoeuvre the whale could not perform without the use of the flippers since the animal uses the flukes to accelerate (Fish, 1997). The body roll may be necessary to approach the prey quickly and efficiently since ventral flexing is faster than the sole use of flippers (Madsen et al., 2005) while maintaining speed throughout the manoeuvre (about 1.2 m/s or 5 km/h). Some toothed whales use a ventrally-directed sonar beam. This is achieved either by beam steering, i.e. without head-movement, as postulated for the false killer whale (Pseudorca crassidens, Au et al., 1995) and the Risso’s dolphin (Grampus griseus, Philips et al., 2003), or by scanning, i.e. through movement of the head, as seen in Blainville’s beaked whales (Shaffer et al., 2013). A downward-directed sonar beam has been suggested to reduce echoes from the water surface (Koblitz et al., 2016).
However, a ventrally directed beam (without vertical head movement) would also be energetically efficient for detecting prey ventrad, since the whale could maintain a straight head-body attitude along the axis of the swimming direction and, by keeping scanning movements of the head to a minimum, avoid unnecessary drag. The direction of the sonar beam in beaked whales is not known but it would naturally assist whales hunting benthic prey and facilitate in this manner the energetically efficient down-slope hunting reported of these whales (Tyack et al., 2006). It would also explain the whale performing a body roll when pursuing prey attempting to escape upwards (Madsen et al., 2005). Prey capture requiring manoeuvres while maintaining speed necessitate the

B e r a r d i u s a r n u x i i
148
use of flippers (Fish and Rohr, 1999; Fish, 2002). However, in contrast to pilot whales, beaked whales perform these tasks rather slowly (Madsen et al., 2005; Aguilar de Soto et al., 2008). Both species use suction feeding and do not have to apprehend prey with their teeth first. The difference in hunting speed is rather explained by the differences of speed of the fleeing prey (Aguilar de Soto et al., 2008). Drag increases exponentially
with speed (Fish et al., 2008) and is therefore much more of a factor in pilot whales that have adapted a more ‘fast and furious’ hunting strategy (Aguilar de Soto et al., 2008) rather than the highly energy-preserving slow mode of the beaked whales (Tyack et al., 2006). While beaked whales predominantly target slow but low-caloric species in the lower, less oxygenated, deep scattering layer (Arranz et al., 2011), pilot whales pursue more energy-rich but faster prey (Spitz et al., 2011). As a consequence pilot whales are highly dependent on mobile control surfaces, i.e. flippers, to provide induced instabilities for fast turning (Fish, 1997, 2002) and wing-like, highly swept-back flippers to minimise drag (Weber et al., 2009b). The low-aspect-ratio flippers of beaked whales on the other hand, are more efficient at lower speeds and precise manoeuvres which also include extensive body-roll (Fish, 1997; Woodward, 2006; see also Chapter 4 Functional anatomy: Diversity of flipper shapes). Beaked whales probably use their well-developed antebrachial muscles in isometric contraction to stiffen the flipper while pilot whales rather rely on structural stiffness (Cooper et al., 2007b). The question thus arises if the retention of plesiomorphic flipper morphologies is of any advantage or rather merely not detrimental? Beaked whales have not always been deep-divers: early stem ziphiids are regarded as shallow-water inhabitants that may have slowly been displaced in the late Miocene by the faster and more efficient dolphins (Lambert et al., 2015). Later ziphiids responded by specialising on slower, but also much deeper-living prey (Bianucci et al., 2016). Thus, the most
important changes encompassed adaptations to deep diving and not to speed as seen in most modern odontocetes (namely dolphins), permitting ziphiids to retain a plesiomorphic flipper morphology more suited to a general-purpose applications rather than the lifestyle of a specialist.
Future studies
In ziphiids, highly specialised adaptations to deep diving appear to be ostensively unlinked from archaic flipper morphologies. Although increased interest and advances

B e r a r d i u s a r n u x i i
149
in technology have in more recent years significantly increased the knowledge of physiological, ecological and behavioural aspects in these whales, little is known of how (or if) beaked whales utilise their flippers when hunting at depth. The predictions made here in this study are purely speculative but could (if confirmed) contribute to the understanding of economical and behavioural aspects in beaked whales.
Suction cups providing biomechanical data (Dtags recording sound, depth, three-dimensional orientation and heading of a whale) and/or carrying cameras have been successfully attached to whales in a number of studies (e.g. López, 2015; Aguilar de Soto, 2008; Watwood, 2006; Tyack, 2006; Aoki, 2015). These instruments would also be able to offer information on the manner of when and how flippers are employed, and in conjunction with integrated computed 3D models exploring flow patterns of the flippers, give a more detailed insight into the energetics involved in dives and associated imposed physiological restraints. While forelimb characteristics of sperm whales and pygmy sperm whales appear to be more archaic in some aspects, shoulder and forelimb morphology of beaked whales are likely more congruent with that of dorudontids and basilosaurids overall. Ziphiid flippers thus possibly present the best extant model for the reconstruction of soft tissue in early fully-aquatic, fluked cetaceans (pelagicetes), potentially also enabling further insight into physiological and behavioural aspects of extinct species.
Berardius arnuxii Purakaunui Life reconstruction of male adult Arnoux’s beaked whale stranded at Pūrākaunui Inlet, near Dunedin, South Island, New Zealand on 22 December 2010

B e r a r d i u s a r n u x i i
150
Summary Two strandings events of Arnoux’s beaked whales in Southern New Zealand (2010, 2013) offered an excellent opportunity to examine for the first time the musculoskeletal arrangements of the shoulder and forelimb in this poorly known species. Dissections were carried out on three specimens after radiographs were taken: a young adult male (Purakaunui, 8.70 m, left flipper), a subadult female (Tangimoana, about 7.00 m, left and right flipper), and a juvenile female (Hine Reti, about 6.50 m, right flipper). Shoulder and forelimb morphology of Berardius arnuxii correlate well in general well with those observed in other odontocetes, with the exception of a few distinctive patterns characteristic for the family and/or the species. Beaked whale flippers are moderately short and wide compared to other odontocetes but distinctive in their deltoid shape. Among ziphiids, the flipper of B. arnuxii is comparatively broad and appears more paddle-shaped than the flippers of the other beaked whales in this study.
Shape and robustness of the scapulae in all beaked whales investigated are quite similar. The greatest variance was observed in the ratio of length / height of the scapular blade, ranging from somewhat longer than high (Z. cavirostris) to almost twice as long as high (M. grayi), with B. arnuxii in between. In beaked whales, the scapular blade itself is quite sturdy, with a relief only vaguely present, the supraspinous fossa fairly small, and acromion and coracoid processes well-developed. Among ziphiids, the scapula of B. arnuxii is distinguishable by a comparatively broad rectangular to spatulate acromion and a long but sturdy coracoid, features that appear even more exaggerated in B. bairdii. The forelimb in beaked whales encompasses besides a fully functional shoulder joint, also a synovial elbow joint with fully formed articular surfaces. However, the interlocking nature of the elbow joint articulation would almost completely prevent any movements of the forearm. Among beaked whales, characteristics of the long bones have been found to be more homogeneous than those of the scapula. With a ratio of humerus / radius length of about 1.0, the humerus in B. arnuxii is typical for ziphiids, and moderately long within odontocetes yet longer than that of delphinids. Overall length of the long bones, together with a relatively short manus with low phalangeal

B e r a r d i u s a r n u x i i
151
count (for odontocetes) are defining characteristics that give ziphiid flippers their typical deltoid shape. Arrangements of carpal elements in individuals of B. arnuxii are very diverse and show various inter- and intrarow fusion patterns involving both proximal and distal elements. Tangimoana showed a most intriguing U-shaped fusion pattern of the ulnare with the
distal carpal 4 and possibly 5. A similar pattern has also been described in another individual by Flower in 1872. Phalangeal count of digital elements in B. arnuxii is typical for beaked whales and comparatively low for odontocetes with 0/4-5/4/3/2 (bony elements) and 0-1/5-6?/5/4/3 (including cartilaginous elements) respectively. Muscular arrangements correspond well with the general condition and topography in odontocetes. Exceptions are the absence of the levator scapulae and the presence of the triceps brachii retaining a powerful scapular head in addition to the humeral head. A long and sometimes broad acromion may be linked to the teres major encroaching onto the caudal lateral surface of the scapula and spatial requirements of the hypertrophied deltoideus, the acromion process hereby providing additional surface for attachment (together with the prescapular fascia) and increased leverage moment. Similarly, a long and sturdy coracoid supports well-developed coracobrachialis and pectoralis minor muscles. In the manus, functional, albeit reduced, antebrachial flexor and extensor muscles in all ziphiids investigated suggests at least some flexibility of the hand. Despite highly specialised adaptations to deep diving beaked whale flippers retain several plesiomorphic characteristics (e.g. a comparatively long humerus, antebrachial
extensors and flexors; see also Chapter 4 Functional anatomy: Discussion). It is not known if, how or when beaked whales use their flippers at depth. Indents in the flanks suggest that ziphiids retract flippers when diving and in this manner achieve hydrodynamic efficiency. However, some data (Madsen et al., 2005) suggest that beaked whales perform manoeuvres when hunting at depth that necessitate the use of flippers. The low-aspect-ratio flippers of beaked whales may hereby be most efficiently utilised at lower speeds for precise manoeuvres that also include extensive body-roll.

B e r a r d i u s a r n u x i i
Plate I
Plate I - Fig. 1. Radiographic image of the left forelimb of B. arnuxii Purakaunui.
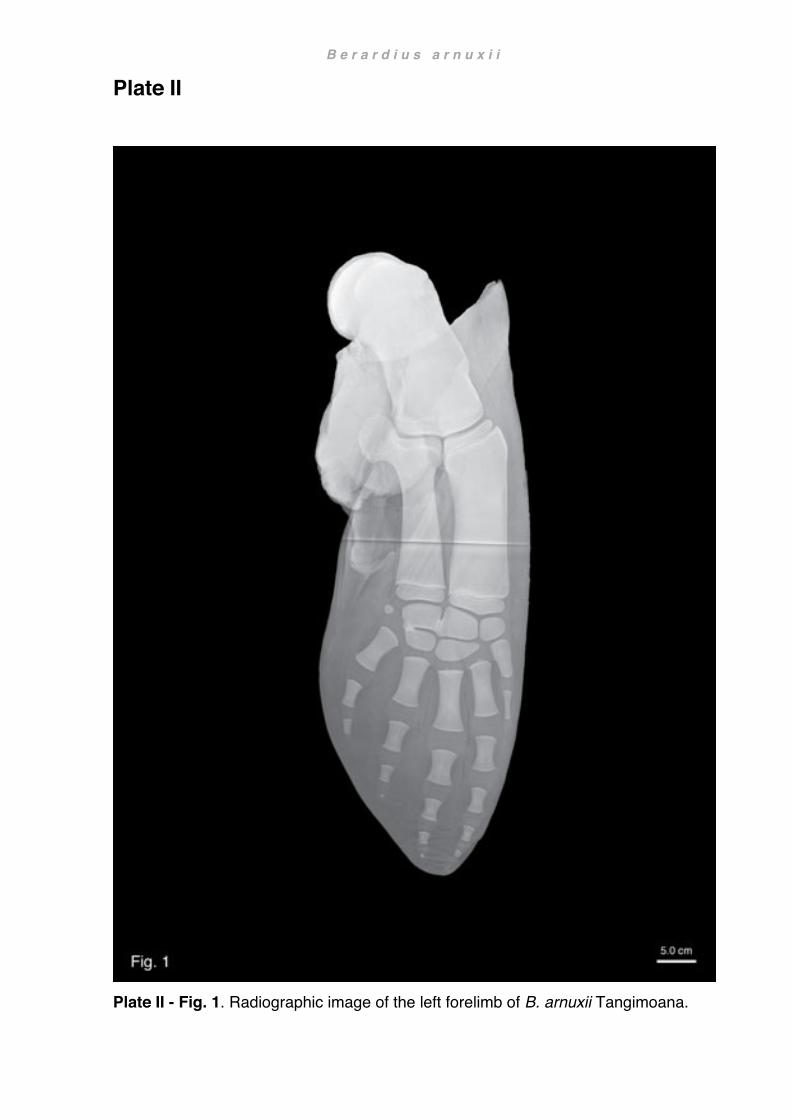
B e r a r d i u s a r n u x i i
Plate II
Plate II - Fig. 1. Radiographic image of the left forelimb of B. arnuxii Tangimoana.

B e r a r d i u s a r n u x i i
Plate II
Plate II - Fig. 2. Radiographic image of the right forelimb of B. arnuxii Tangimoana.

B e r a r d i u s a r n u x i i
Plate III
Plate III - Fig. 1. Radiographic image of the right forelimb of B. arnuxii Hine Reti.

156
Chapter 4: Functional anatomy
Introduction Cetaceans belong to a tetrapod lineage that returned to an aquatic lifestyle during the early Eocene (Kellogg, 1928; Thewissen, 1998; Fordyce and de Muizon, 2001; Thewissen, 2009; Thewissen et al., 2009; Cozzi et al., 2016). They are the only obligate aquatic mammals that have conquered the high seas; the other fully aquatic mammals being the slow-swimming sea cows (Sirenia) that inhabit rivers and shallow coastal areas (Reeves et al., 2002). Extant whales, dolphins and porpoises are the most diverse group amongst aquatic mammals and are able to exploit a wide range of habitats from littoral to pelagic environments such as riverine or coastal shallow waters, the open ocean or the deep sea (Berta et al., 2015). History and phylogeny
The lineage from terrestrial ancestors to modern oceangoing cetaceans is well documented by fossil records, yet essential questions remain unresolved. Modern anatomical data support a close cetacean relationship to ungulates (hoofed animals) and until recently regarded whales as a sister group of artiodactyls (odd-hoofed animals; O'Leary and Geisler, 1999; Geisler, 2001). This was, however, contrary to molecular data supporting cetacean derivation from within the artiodactyl taxon (O'Leary and Geisler, 1999; see also fig. 1). One of the most obvious reasons for this discrepancy is the marked discontinuity in morphology between extant cetaceans and their closest living relatives, and the incongruence between observed morphological characters and independent molecular data sets (Berta et al., 2015). Already over 2000 years ago Aristoteles documented that cetaceans are air-breathing, give birth to live young, suckle their calves and provide parental care (Romero, 2012). However, it was not until the late 17th and 18th century that they were finally recognised as mammals by John Ray (1693) and Carolus Linnaeus (1758). Subsequently in the 19th and early 20th centuries cetaceans were associated with various mammalian

A n a t o m y
157
clades. However, it was John Hunter (1787) who crucially noted a resemblance with ruminants and 100 years later William Flower (1883) who consequently deduced a close relationship with artiodactyls. In the 1960s Everhard Slijper (1966) allied cetaceans with artiodactyls while Leigh Van Valen (1966) favored Mesonychia, extinct carnivorous hoofed mammals. Conversely, some molecular studies in the 1990s
nested cetaceans within Artiodactyla (Cetartiodactyla; Montgelard et al., 1997; Waddell et al., 1999) with Hippopotamidae as their closest relatives (Gatesy, 1998; Nikaido et al., 1999; Waddell et al., 1999). Morphological studies on the other hand still tended to support a closer link to Mesonychia (Gingerich, 1998; Gingerich and Uhen, 1998) and some more recent morphological studies still maintained a closer relation to Perissodactyla (Gingerich, 2015). However, at the beginning of the new millennium new morphological evidence emerged that substantiated a strong link between the earliest archaeocetes and artiodactyls: both groups shared a uniquely configured ankle bone (astragalus) not present in Mesonychia or Perissodactyla (Gingerich et al., 2001a; Thewissen et al., 2001), while archaic whales also possessed ear bones (tympanic bullae) synapomorphic for cetaceans (Gingerich et al., 2001a). The corroboration of morphological and genetic data gave renewed credence to the concept of hippos as the sister group of cetaceans, both nestled within Artiodactyla (Uhen, 2010; Geisler et al., 2011; Gatesy et al., 2012; McGowen et al., 2014).
Cetacean evolution (Fig. 1)
Archaeocetes first emerged in the Indo-Pakistan region during the Early Eocene about 53 my ago. The most archaic whales, the Pakicetidae, were essentially semiaquatic mammals known from the Early to the Middle Eocene (53-47 my; Thewissen and Bajpai, 2001). These early whales were digitigrade with typically artiodactyl cursorial limbs, so were capable of terrestrial locomotion (Madar, 2007). Yet, they also showed already specialised adaptations to their freshwater habitat such as well-developed underwater hearing, increased limb bone density (osteosclerosis, hypermineralisation) serving as ballast to counteract buoyancy (Walley, 1983; McGowen et al., 2014), possibly webbed feet and a strong tail. These adaptations suggest a predominately aquatic life and pakicetids were most likely capable of either foot-powered swimming or walking at the bottom of a river or pool similar to hippos (Gray et al., 2007; Madar, 2007; Thewissen et al., 2007; Cooper et al., 2012; Gingerich, 2012; McGowen et al., 2014). However, such an aquatic lifestyle was not unique to pakicetids and is also

A n a t o m y
158
proposed for some of their close artiodactyl relatives, e.g. the raoellid Indohyus, suggesting that these adaptations occurred before the emergence of the first archaic whales (Thewissen et al., 2007). Like the Pakicetidae, the larger Ambulocetidae, about 49 my ago, were also capable
of terrestrial locomotion (Uhen, 2010) but showed progressive aquatic modifications with shorter, but broader limbs using pelvic paddling and undulation like otters (Thewissen and Hussain, 1996; Thewissen and Fish, 1997; Madar et al., 2002). Ambulocetids lived in both freshwater environments and coastal areas near or in river estuaries. There they probably lay in wait to ambush terrestrial prey, similar to modern crocodiles (Rose, 1985; Thewissen et al., 1994; Thewissen and Hussain, 1996). Remingtonocetidae were coeval with ambulocetids and were also amphibious (Gingerich et al., 1995; Bajpai and Thewissen, 2000; Bajpai et al., 2009), but unlike pakicetids or ambulocetids, remingtonocetids were the first really true marine cetaceans foraging near the coast in bays and swamps (Bajpai and Thewissen, 1998; Clementz et al., 2006; Thewissen and Schneider, 2009). With their long body, elongated skull and rostrum, they resembled gharials (Gavialis gangeticus) but hunted probably somewhat different prey than their modern freshwater counterparts (Bebej et al., 2007; Bajpai et al., 2011; Bebej et al., 2012).
Overleaf: Fig. 1. Phylogeny of Cetacea Phylogenetic relationship of Cetacea and non-cetacean mammals Phylogenetic tree modified from Gatesy et al., 2012, and Uhen, 2014a (Protocetidae + Remingtonocetidae), Uhen, 2018
(Basilosauridae), Lambert et al., 2017 (Mysticeti); classification of Delphinida and Delphinoidea sensu Geisler et al., 2011. Artwork
by C. Buell, except Equus (not referenced), see <https://equineguelph.ca/learn_objects/evolutiontimeline/equus.html>,
Hippopotamus by R. Nicholls, Phocoena by Encyclopaedia Britannica 2010, Lissodelphis by Encyclopaedia of Marine Mammals
2018.

A n a t o m y
159

A n a t o m y
160
Like their more basal predecessors, the Protocetidae also arose in the Indo-Pakistan region about 48 my ago but spread rapidly across North Africa, Europe, North America and western South America (Uhen et al., 2008). Despite only inhabiting coastal marine environments like reefs, their wide geographical range indicates that they were accomplished swimmers. A shift of the nasal openings (nares) posterad and cranial
asymmetry combined with well-developed underwater hearing supports this notion (Uhen, 2010; Fahlke et al., 2011; McGowen et al., 2014). Strong, broad hind limbs and a powerful tail were used for pelvic paddling and undulation (Uhen, 2008). Vertebral morphology also suggests a nascent modern, highly efficient undulatory swimming mode (Bianucci and Gingerich, 2011) but there is no fossil evidence indicative of flukes (Uhen, 2008, 2010). Skeletal remains of early protocetids rather suggest a life-style similar to modern pinnipeds, highly adapted to aquatic life but mating and giving birth on land (Gingerich et al., 2009). The evolution of later protocetids was marked by improved aquatic adaptations and gradual decrease of terrestrial competence until the link to land was finally lost in the latest protocetids (Gingerich et al., 2001b; Gingerich et al., 2005; Uhen, 2014a). Based on skeletal traits of the hip, limbs and particularly of the spinal column, Uhen (2008) proposed a new (but paraphyletic) clade Pelagiceti comprising Basilosauridae and Neoceti, both highly efficient obligate aquatic swimmers with flippers and flukes. Basilosauridae emerged in the Middle Eocene 40 my ago and are found worldwide (Gingerich et al., 1990; Fordyce, 2009b; Uhen, 2009a). Basilosaurids (sensu lato) are regarded as either a sister group to dorudontids or a paraphyletic group (Gatesy et al., 2012). They include the sometimes large and very elongated basilosaurines; the dorudontines, giving rise to modern whales and dolphins (Neoceti); and the little known
kekenodontids from New Zealand (Uhen, 2010). Although they share characteristics with basal Neoceti, such as vestigial hind limbs and flattened, long forelimb bones with limited articulation (Uhen, 1998, 2004), they lacked sophisticated echolocation or baleen plates (Bajpai et al., 2009). Modern cetaceans, the Neoceti, are a monophyletic group divided into Odontoceti (toothed whales, dolphins and porpoises) and Mysticeti (baleen whales). Neoceti arose more than 30 my ago and are either defined by their ability either to echolocate (odontocetes) or to filter-feed (mysticetes; Fordyce, 2009c). Both clades show

A n a t o m y
161
distinctive telescoping of the skull (Miller, Gerrit S., 1923): baleen whales exhibit a marked lengthening in the back of the skull (occipital region, particularly right whales), whereas in toothed whales it is the upper jaw bones (maxillae) that mostly elongate (most prominently in beaked whales), or a combination of the two (sperm whales and Ganges River dolphin). Sophisticated echolocation has also been linked to the highly
asymmetric skull in odontocetes (Fahlke et al., 2011; Geisler et al., 2014). The most archaic baleen whale, Mystacodon, is represented by a single species that appeared about 36 my ago (Lambert et al., 2017) closely followed by the later toothed Llanocetidae from Antarctica and New Zealand, appearing about 34 my ago. They were large animals with a skull in appearance intermediate between basilosaurids and more modern mysticetes, and had a broad, flat rostrum with widely spaced heterodont teeth. Interestingly, baleen, regarded as a key adaptation in older literature, was not present in Llanocetus (Fordyce, 2003b), the animal is rather interpreted as a suction-assisted benthic or raptorial feeder (Lambert et al., 2017; Fordyce and Marx, 2018). Later archaeomysticetes were interpreted as macrophages (Mammalodontidae; Fitzgerald, 2006) and yet another group (Aetiocetidae) had besides small teeth minute lateral palatal foramina and sulci, interpreted as homologous with similar but larger structures found in modern mysticetes (Deméré and Berta, 2008; Deméré et al., 2008). Although the oldest preserved baleen is much younger (Esperante et al., 2008), structures such as these are in modern whales associated with blood vessels supplying tissue from which baleen develops and give strong evidence that these early baleen whales also had some form of baleen. The earliest toothless mysticetes (Eomysticetidae) are about 26 my old and had relatively delicate skulls with long, flat rostra and nares midway between the tip of the
snout and eyes (Sanders, 2002; Uhen, 2007a; King et al., 2009; Boessenecker and Fordyce, 2014, 2015). Later chaeomysticetes (toothless mysticetes) include the right and bowhead whales (Balaenidae), the most basal extant baleen whales, that diverged about 30 my ago (Marx and Fordyce, 2015). They can be readily identified by their stout body shape, and disproportionally large head with distinctively arched, narrow rostrum and long, slender baleen plates (Bannister, 2009). Pygmy right whales (Neobalaenidae) are the most enigmatic living baleen whales and are represented by only one extant species

A n a t o m y
162
(Caperea marginata; Bannister, 2009). Their phylogenetic relationship has so far remained a matter of debate as indeed has that of their close extinct relatives, the cetotheriids (Fordyce, 2009b; Marx et al., 2013). Pygmy right whales are the smallest of all living baleen whales and incorporate physical attributes of both right whales (large head with arched upper jaw) and rorquals (streamlined body shape, four digits), as well
as some unique features (broadened ribs; Baker, 1985; Kemper, 2009; see Chapter 2 Caperea). Gray whales (Eschrichtiidae) are also in appearance intermediate between balaenids and balaenopterids. They have a moderately stout body shape, a narrow, triangular head, and slightly arched rostrum. Grey whales have a broad spectrum of feeding techniques including skimming like right whales, gulping like rorquals and a unique method of suction feeding (Jones and Swartz, 2009). Rorqual and humpback whales (Balaenopteridae) belong to the most recently diverged lineage (Marx and Fordyce, 2015). Rorquals are large mysticetes, built for speed with a streamlined body shape and short, narrow flippers (Ridgway, 1972; Goldbogen, 2006). They have short, coarse baleen plates and throat grooves ideal for engulfing large volumes of water when lunge-feeding (Goldbogen et al., 2007; Jefferson et al., 2015). Humpback whales are only represented by one species (Megaptera novaeangliae). They too, are large animals but stand apart with disproportionately long flippers that are important for social contact, fish herding and great maneuverability (Howell, 1930a; Edel and Winn, 1978; Benke, 1993; Fish and Battle, 1995). The earliest odontocetes appeared about 32 my ago and belonged to several, quite dissimilar families (Xenorophidae also including Archaeodelphis, Simocetidae, Agorophiidae, and others), yet with ancestral features patently similar to basilosaurids (Fordyce, 2009c). On the other hand, apomorphic skull characteristics suggest at least
nascent high-frequency echolocation was already present in most of these early toothed whales (Fleischer, 1976; Geisler et al., 2014; Pyenson, 2017). In many groups polydont-heterodont dentition is present; others evolved homodonty or greatly reduced the number of teeth (Uhen, 2009b). Most modern odontocetes swallow prey whole, a method that has also been postulated for most early odontocetes (Bianucci, 2005). The phylogeny of the odontocete crown groups has remained so far problematic (de Muizon, 1990). Early extinct modern odontocetes comprise Squalodontidae, Waipatiidae and other potential groups like Prosqualodontidae, about 25 my old (Fordyce, 1994; Tanaka and Fordyce, 2014). The box-headed pygmy sperm whale

A n a t o m y
163
and sperm whale (Kogiidae, Physeteridae) are regarded as the most basal extant crown odontocetes, followed by the long-snouted Indian Ganges River Dolphin (Platanistidae; Gatesy et al., 2012; Tanaka and Fordyce, 2015), the latter associated with the extinct Squalodelphinidae and Allodelphinidae. Beaked whales (Ziphiidae) are the most diverse family amongst the larger living toothed whales and are distinguished
by an elongated rostrum (de Muizon, 1990; Fordyce, 1994, see also Chapter 3 Berardius). Delphinida are a well-supported clade including the extinct Kentriodontidae, the recently extinct Chinese Yangtze ‘river dolphin’ (Lipotidae), and the South American Franciscana ‘river dolphin’ (Pontoporiidae) and Amazon ‘river dolphin’ or botu (Iniidae; McGowen et al., 2014). Extant crown delphinoids include beluga and narwhal (Monodontidae), porpoises (Phocoenidae), and the speciose and diverse ocean-going dolphins (Delphinidae) such as killer whales (Orcinus orca, Orcininae), (dorsal-) finless right whale dolphins (Lissodelphininae), pilot whales (Globicephalinae), and bottlenose or common dolphins (Delphininae; McGowen et al., 2009; McGowen et al., 2014). Anatomical accounts of the shoulder and forelimb
There are a multitude of descriptions detailing osteology or myology of the shoulder and forelimb in cetaceans. Those encompassing mysticetes are referred to in Chapter 2 Caperea, Chapter 3 Berardius focuses on beaked whales, and Chapter 5 Literature review enumerates a wide selection of works on osteology and myology too numerous to discuss here. It is therefore rather surprising, that despite a fairly comprehensive understanding of the evolutionary pathway of cetaceans, only very few modern accounts detailing the soft tissue of the shoulder and/or flipper take evolutionary aspects into consideration (e.g. Uhen, 2004; Cooper et al., 2007a; Cooper et al. 2007b; Sanches and Berta, 2010). Other papers investigating biomechanics (Kleinenberg, 1969) or behavioural aspects (e.g. Ford, 2008, Lambert 2015) refer only superficially or in very specific terms to evolutionary aspects. Chapter 4 endeavours to integrate phylogeny, behaviour, ecology, kinematics and other potential facets influencing the anatomical arrangement of the shoulder and forelimb seen in whales and dolphins today.

A n a t o m y
164
Results For more detailed descriptions of the forelimb in mysticetes and beaked whales see Chapter 2 Caperea and Chapter 3 Berardius.
Position of the forelimb
The position of the forelimb is defined by the position and angle of the shoulder blade
on the rib cage. Mounted museum specimens or depictions of articulated skeletons are notoriously incongruent in this respect. There is no bony connection to the rest of the skeleton or bony features in the skeleton itself that would allow a precise placement and orientation of the shoulder blade once the soft tissue has been removed (Rapp, 1837). Consequently, correct position and orientation of the forelimb can only be ascertained in specimens where the musculoskeletal arrangement is still intact. In such specimens the bony cranial angle of the scapula generally lies just forward of the first rib at about the height of the costal angle (Fig. 2). However, the cartilaginous border may project farther in species with pronounced elongated shoulder blades. From here the vertebral border transverses the rib cage in caudo-ventrad direction descending at an angle of around 25º from the body axis (e.g. Caperea marginata). In Grampus griseus this angle is steeper at 30º, whereas in Berardius arnuxii it is shallower at 20º. However, this angle was found to always reflect the angle between the glenoid border and the longitudinal axis of the shoulder blade, positioning the limb at a right angle to the longitudinal axis of the body.
Pectoral limb
The ratio of body length to flipper length varies significantly in the cetaceans investigated, from < 5.5 - 6.5 (C. hectori, L. obscurus, G. griseus), and 7-9 (C. marginata, C. hectori, K. breviceps), to 10-12 < (C. marginata, P. dioptrica, ziphiids). Similarly, the shape of the flipper shows great variability. Nonetheless, six distinctive major shapes could be identified (see also Chapter 1 Methods and materials: flipper shapes and Functional aspects of flipper shapes, section Diversity of flipper shapes, Fig. 30 in this chapter):

A n a t o m y
165
Fig. 2. Position of the forelimb Semi-schematic drawing of Grampus griseus showing position and orientation of the scapula on the thorax and orientation of the forelimb in relation to the body. Skeleton adapted after van Beneden, 1880
1) Oar-shaped flippers occur typically in baleen whales (Fig. 3a). This shape is typified by a narrowing, elongated proximal part of the flipper (supported by the long bones) and a moderately tapered distal part (supported by the manus); the ratio of long bones to manus is about 4/3 or higher. Oar-shaped flippers are long and narrow; forelimb length can range from very long (about 1/3rd of the skeletal length in humpback whales; Benke, 1993) to very short (nearly 1/12th of the skeletal length in pygmy right whale MM0036; this study). The widest part is confined to the basis of the flipper where it is supported by a large olecranon. In this flipper shape, the leading edge is longer than the trailing edge and in most cases the flipper is more than three times longer than wide. The distal trailing edge is usually straight but can also be faintly concave or convex. In rorquals like Balaenoptera acutorostrata and B. edeni, the loss of digit 1 and more or less appressed digits in the manus extrapolate the taper of the more proximal flipper section. In contrast, the tip of the flipper in Caperea marginata is quite blunt and postaxial digits are little appressed resulting in a more oval flipper contour. Unique amongst cetaceans, C. marginata has also a huge olecranon that supports an
extensive axillary fold. 2) Deltoid-shaped flippers are the hallmark of beaked whales (Fig. 3c). In these flippers axial and postaxial digits progressively diverge towards the trailing edge, digit 5

A n a t o m y
166
being the most divergent, angulated and ‘kinked’ caudo-distally. Consequently, the broadest part of the flipper is distal to the mid-section, and from there the flipper tapers much more towards the tip. These flippers are nonetheless fairly long with a slightly lower long bones / manus ratio than baleen whales, about 1/1. The deltoid shape is epitomised in Mesoplodon grayi and M. layardii, whereas Ziphius cavirostris flippers
are slightly wider with a shape in between that of Mesoplodon and Berardius. Berardius arnuxii has the broadest flippers amongst beaked whales. These flippers are almost paddle-shaped with axial and postaxial digits distinctively splayed, particularly digit 5. 3) Paddle-shaped flippers are most conspicuous by their low length/width ratio. They are typified by a narrow proximal portion and a wide mid-section that is either strongly tapered towards the tip (Fig. 3b) or truncated. The widest part of the flipper is hereby determined by the taper of the distal portion: a widening angle at the tip shifts the planform of the flipper from triangular (e.g. Kogia) to spatulate (e.g. Physeter, Platanista), while angles in between result in intermediate shapes (e.g. Pontoporia). Long bones / manus ratios lie in between 4/3 and 3/4, and planforms can approach dimensions nearly as wide as long (Platanista; Benke 1993). Paddles are distinguished by extreme separation of the axial and postaxial digits in the manus. They can be quite large (in right whales 1/6th of the body length; Benke, 1993) or comparatively small (in sperm whales 1/10th of the body length; Benke, 1993), and are represented here by the pygmy sperm whale (Kogia sima and K. breviceps). 4) Fusiform-shaped flippers can appear similar to paddle-shaped flippers but are much more smoothly curved. The leading edge follows a constant curve, while the trailing edge takes on a sinuous contour (Fig. 3d) forming the widest part of the flipper
about 2/3 between base and tip. Fusiform flippers can be broad (e.g. Lipotes, Delphinapterus, Phocoena) or only moderately wide (e.g. Eschrichtius, Monodon), and are here represented by the spectacled porpoise (Phocoena dioptrica). 5) Oval-shaped flippers have similar proportions similar to paddle-shaped flippers, but the entire trailing edge is strongly convex, and the tip is blunt (Fig. 3e). A typical oval-shaped flipper is represented here by the Hector’s dolphin (Cephalorhynchus hectori), but contours can also attain more extreme forms either exaggerating either width (Orcinus orca) or length (Caperea marginata).

A n a t o m y
167
6) Falcate-shaped (falciform) flippers have a short but markedly wide proximal potion, and an extremely pronounced taper distad of the wrist area (Fig. 3f). They are typified by a distinct crescent-shaped (concave) distal trailing edge. This shape is achieved by a comparatively short brachium and a sometimes extremely elongated manus
(extreme hyperphalangy in Globicephala melas; Cooper et al., 2007a) resulting in a long bones / manus ratio of about 2/3 (G. sp., G. griseus) to 3/4 (L. peronii, L. obscurus). Classical examples of this shape are present in Lissodelphis peronii, Globicephala melas, G. macrorhynchus, and Lagenorhynchus obscurus. Grampus griseus flippers also fit this description, if not quite as pronounced.
Overleaf: Fig. 3. Flipper shape From top to bottom row and left to right: (a) oar-shaped flipper of Balaenoptera acutorostrata, B. edeni, Caperea marginata (MM2904); (b) Deltoid-shaped flipper of Ziphius cavirostris (Katiki Beach), Mesoplodon grayi (Kaka Point), M. layardii (Ocean View), Berardius arnuxii (Purakaunui); (c) paddle-shaped flipper of Kogia sima (VT3011); (d) fusiform-shaped flipper of Phocoena dioptrica (X2020.77); (e) oval flipper of Orcinus orca (Maripi), Cephalorhynchus hectori (H199 adult), (f) falcate flipper of Lissodelphis peronii, Globicephala macrorhynchus (10-22Lo), Lagenorhynchus obscurus, Grampus griseus. (a-f) left flipper, lateral view. Semi-schematic illustrations based on radiographs and dissections.

A n a t o m y
168

A n a t o m y
169
Osteology See also measurements in appendix
Scapula
The classic cetacean scapula is more or less shaped like a Japanese ogi (folding fan) and, like the fan, medio-laterally compressed (e.g. Caperea marginata Fig. 4; see also
Benke, 1993). All three borders can show considerable inter- and sometimes intraspecific variability. Acromion and coracoid processes are also flattened and rise only slightly above or below the plane of the blade. Like two handles, they project forward and surpass in some cases the cranial-most tip of the scapula (e.g. Lissodelphis peronii, Fig. 8c or Ziphius cavirostris, Fig. 8d).
Fig. 4. Classical fan-shaped scapula Left scapula of Caperea marginata (MM2904), lateral view
The vertebral border rises as a convex arch above the cranial and caudal angles. The convexity can vary between evenly convex (e.g. Kogia breviceps, Fig. 5a), asymmetrical or irregular (Lissodelphis peronii, Fig. 6b) to comparatively flat or wavy with distinct central dip (Caperea marginata, Fig. 5c). In Globicephala melas and
Cephalorhynchus hectori (Fig. 5b) the vertebral border appears occasionally crenated and in Phocoena dioptrica sinuous (Fig. 6a). Above the vertebral border, the cartilaginous border, sometimes referred to as the suprascapula (e.g. Gegenbaur, 1872; DePalma, 2008), persists life-long as a distinctive cartilaginous section of the bony blade (Flower, 1885). However, it is generally not preserved in osteological collections and consequently rarely included in anatomical descriptions. In most cases the cartilaginous border forms a small, regular

A n a t o m y
170
band reflecting the general outline of the shoulder blade. In other cases, it forms a highly variable band that drastically the changes contour and surface area of the scapula. In Caperea marginata this border extends far beyond the scapular angles and substantially elongates an already long shoulder blade even further (Fig. 5c). Notably, this is true despite the patently divergent contour of the bony blade between the two
individuals of this study (Fig. 4 vs. Fig. 7d).
Fig. 5. Vertebral border (a) Kogia breviceps (X2020.75), with classical convex vertebral border; (b) Crenated vertebral border of Cephalorhynchus hectori (H182) with notch clearly visible; (c) Elongated, flattened vertebral border in Caperea marginata (MM2959), note cartilaginous vertebral border greatly enlarging the length and surface area of the scapula. (a-c) lateral view, left scapula. Abbreviations: (ac) acromion; (cor) coracoid; white broken line in (a) longitudinal axis; black broken line in (c) missing cartilaginous border.
Cranial and caudal borders of the scapula are more or less straight and can vary from faintly concave, to sinuous or slightly convex. They are usually fairly equal in length and roughly encompass a similar angle as the vertebral border (approach angle
to longitudinal axis), usually between 30º and 60º. However, the convex vertebral border and the variable curvature of the cranial or caudal borders, particularly if the scapular angles are blunt, make accurate measurements of angles often difficult (e.g. K. breviceps, Fig. 5a). An unusually kinked cranial border can be seen in Phocoena dioptrica (Fig. 6a). The kink truncates the cranial angle and foreshortens the contour of the shoulder blade. In consequence, the cranial angle appears at 105º wide open and approach angles of

A n a t o m y
171
cranial and vertebral borders are quite asymmetric (75º and 30º). This imbalance is even more exaggerated at the caudal angle where the vertebral border approaches the longitudinal axis almost parallel (about 5º) and the gradient of the caudal border dominates (70º). Yet, cranial and caudal border appear balanced, more so as the cartilaginous border equipoises the asymmetry of the vertebral border, giving the
scapula as a whole a fairly symmetric profile. Exactly the opposite is true for Lissodelphis peronii (Fig. 6b). Here the contour of the scapula appears caudally lop-sided and extremely asymmetric, yet approach angles of the vertebral and caudal borders are symmetrical (both 45º). Anterior to the cranial angle there is a broad protuberance that has fused with the acromion. The location of the fusion corresponds to a more or less prominent tuberosity that serves in other cetaceans as strong attachment point for the supraspinatus muscle (e.g. C. hectori, Fig. 8a; G. griseus, Fig. 7a). The broad apophysis in L. peronii obscures the anterior edge of the cranial border and makes it difficult to delineate its exact course. However, the inferred contour of the cranial border appears similar in outline to that of P. dioptrica. The difference of the blade profile between P. dioptrica and L. peronii is thus mainly defined by the relative length of the cranial to the caudal border and their approach angles. In P. dioptrica both approach angles are fairly equal (75º and 70º) and produce a symmetrical contour. In L. peronii these angles are greatly disparate (80º and 45º) and produce a disproportionately skewed silhouette. Significantly, these characteristics are not reflected by the approach angles of the vertebral border which are in P. dioptrica askew (30º and 5º) and acute, but in L. peronii more symmetrical (60º and 45º) and rounded. Essentially, the angle between the cranio-vertebral and caudo-vertebral border
(P. dioptrica 105º and 75º, L. peronii 120º and 90º) reflects characteristics of the scapular shape poorly, particularly so if the shape of the cartilaginous border is disparate from that of the vertebral border. Remarkably, the difference between those two angles (30º) is in both specimens equal (Fig. 6a-b).

A n a t o m y
172
Fig. 6. Cranial and caudal borders (a) Phocoena dioptrica, note the distinct kink of the cranial border; (b) Lissodelphis peronii, note the much longer caudal border and the protuberance (pt) at the cranial angle. Lateral view (a) left scapula and (b) right scapula, mirrored. Abbreviations (ac) acromion; (cor) coracoid; (pt) protuberance; white broken line demarcates longitudinal axis of the scapula; grey areas represent approach angles.
The neck of the scapula lies just above the glenoid fossa. Anteriorly it is the attachment point for the acromion and coracoid. Posteriorly it merges into the caudal border, at a comparatively shallow angle and barely distinguishable (e.g. in L. peronii with a short neck at 160º, Fig. 6b) to relatively acute and well-defined (e.g. K. breviceps with a long neck at 105º, Fig. 5a). The ratio of length to height is in the cetacean scapula typically about 3/2 to 4/3, i.e. rather a little longer than higher. The scapula of Grampus griseus (Fig. 7a) and Phocoena dioptrica (Fig. 6a) encompass this attribute perfectly with a ratio of 4/3. A somewhat longer scapula profile with a ratio of 3/2 is represented in Lagenorhynchus obscurus (Fig. 7b). This ratio is also seen in Globicephala melas (Fig. 7c) and
Berardius arnuxii (Fig. 9c), although here the almost straight cranial and caudal border give the scapula a deceptively tall look - not forgetting that in reality the proportions of the scapula in odontocetes are usually further shortened by the cartilaginous border. Slightly longer than 3/2 is the shoulder blade of Cephalorhynchus hectori (Fig. 8a). Within ziphiids, Mesoplodon grayi has the longest scapula with a ratio not quite reaching 2/1(Fig. 8e). Slightly longer, the fan-shaped shoulder blade of Caperea marginata (Fig. 8g) and the lop-sided scapula of Lissodelphis peronii (Fig. 8c) attain a ratio of nearly 2/1. However, the proportionally longest scapula has been recorded for another specimen of C. marginata with quite a different, polygonal-shaped scapular
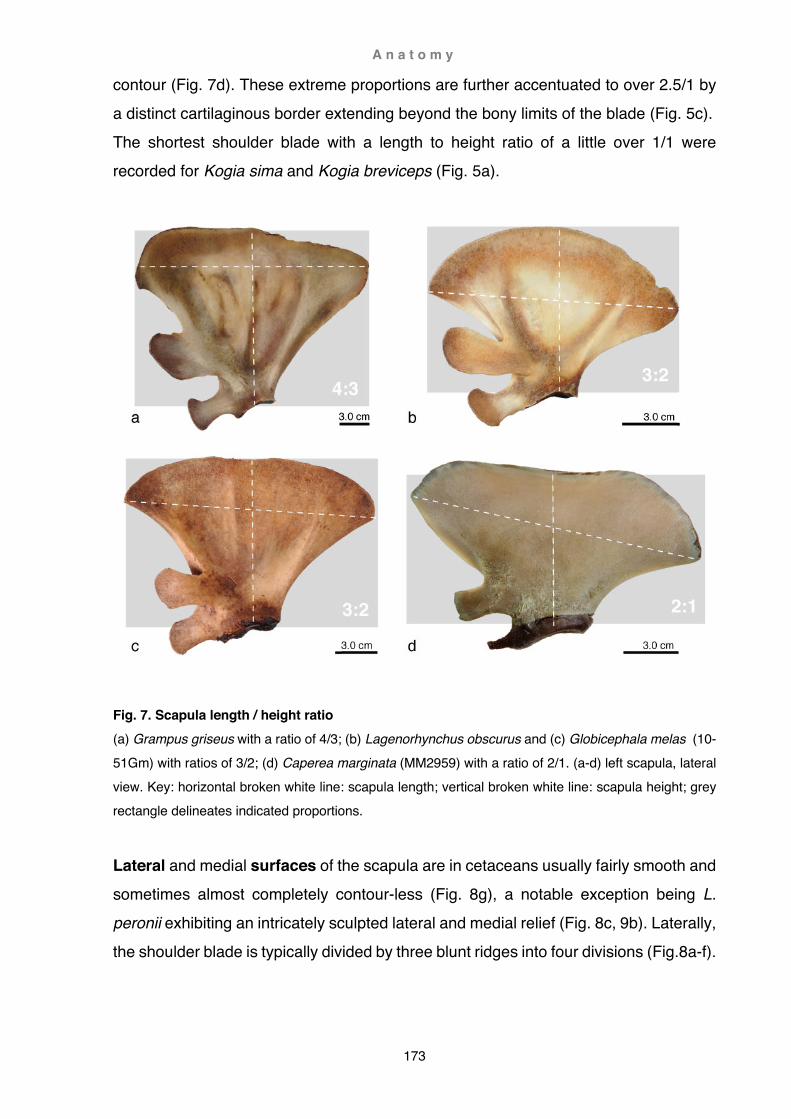
A n a t o m y
173
contour (Fig. 7d). These extreme proportions are further accentuated to over 2.5/1 by a distinct cartilaginous border extending beyond the bony limits of the blade (Fig. 5c). The shortest shoulder blade with a length to height ratio of a little over 1/1 were recorded for Kogia sima and Kogia breviceps (Fig. 5a).
Fig. 7. Scapula length / height ratio (a) Grampus griseus with a ratio of 4/3; (b) Lagenorhynchus obscurus and (c) Globicephala melas (10-51Gm) with ratios of 3/2; (d) Caperea marginata (MM2959) with a ratio of 2/1. (a-d) left scapula, lateral view. Key: horizontal broken white line: scapula length; vertical broken white line: scapula height; grey rectangle delineates indicated proportions.
Lateral and medial surfaces of the scapula are in cetaceans usually fairly smooth and sometimes almost completely contour-less (Fig. 8g), a notable exception being L. peronii exhibiting an intricately sculpted lateral and medial relief (Fig. 8c, 9b). Laterally, the shoulder blade is typically divided by three blunt ridges into four divisions (Fig.8a-f).

A n a t o m y
174
These ridges roughly demarcate: (1) supraspinous fossa (2) origin of the deltoid (3) infraspinous fossa (4) origin of the teres major
The supraspinous fossa can be a small, narrow but distinctly grooved furrow along the cranial border (e.g. G. melas , Fig. 7c) that sometimes terminates in a marked tuberosity (e.g. C. hectori; Fig. 8a) or not be indented at all (e.g. G. griseus, Fig. 7a). In one specimen the fossa merges into a large protuberance bridging the cranial angle and acromion (L. peronii, Fig. 8c, 9b). Occasionally, the fossa veers off craniad and never reaches the tip of the cranial angle (e.g. C. marginata, Fig. 8g). In beaked whales the fossa is generally quite indistinct, and the scapular spine flattens out towards the vertebral border (e.g. Z. cavirostris, Fig. 8d; M. grayi, Fig. 8e; M. layardii, Fig. 8f). The scapular spine, defining the caudal border of the supraspinous fossa, initially emerges from the cranial angle, often as a barely distinguishable, smooth ridge. From here it converges towards the acromion and gradually emerges as a pronounced crest. This crest merges distally into the proximal border of the acromion process. At the point of fusion this crest is sometimes enlarged to a bony flange that can in extreme cases reach halfway towards the tip of the acromion (B. arnuxii, Fig. 10c). The origin of the deltoid lies just behind the scapular spine. Generally, this muscle occupies roughly 1/3rd of the scapular surface. Its fossa however, is usually much smaller since the posterior portion of the deltoid more or less overlaps the anterior portion of infraspinatus muscle (aponeurosis). Where the deltoid and infraspinous fossae are larger, the origin of the teres major remains constricted to a narrow band at
the caudal border of the scapula (e.g. P. dioptrica, Fig. 8b). Separation of the deltoid and infraspinous fossae is typically either by a vague and broad bulge (e.g. C. hectori, Fig. 8a) or a somewhat sharper but little prominent ridge (e.g. P. dioptrica, Fig. 8b; ziphiids, Fig. 8d-f); in some species there is no discernible demarcation (e.g. C. marginata, Fig. 8g).

A n a t o m y
175
Fig. 8. Lateral surface relief (a) Cephalorhynchus hectori (EF005), note cranial and caudal tuberosities; (b) Phocoena dioptrica (X2020.76); (c) Lissodelphis peronii; (d) Ziphius cavirostris (Katiki Beach); (e) Mesoplodon grayi; (f) Mesoplodon layardii; (g) Caperea marginata (MM2904). (a,c) lateral view, right scapula mirrored and (b, d-g) left scapula. Abbreviations: (ac) acromion; (cor) coracoid; (if) infraspinous fossa; (od) origin deltoid; (otm) origin teres major; (pt) protuberance; (sf) supraspinous fossa; broken lines demarcate ridges.
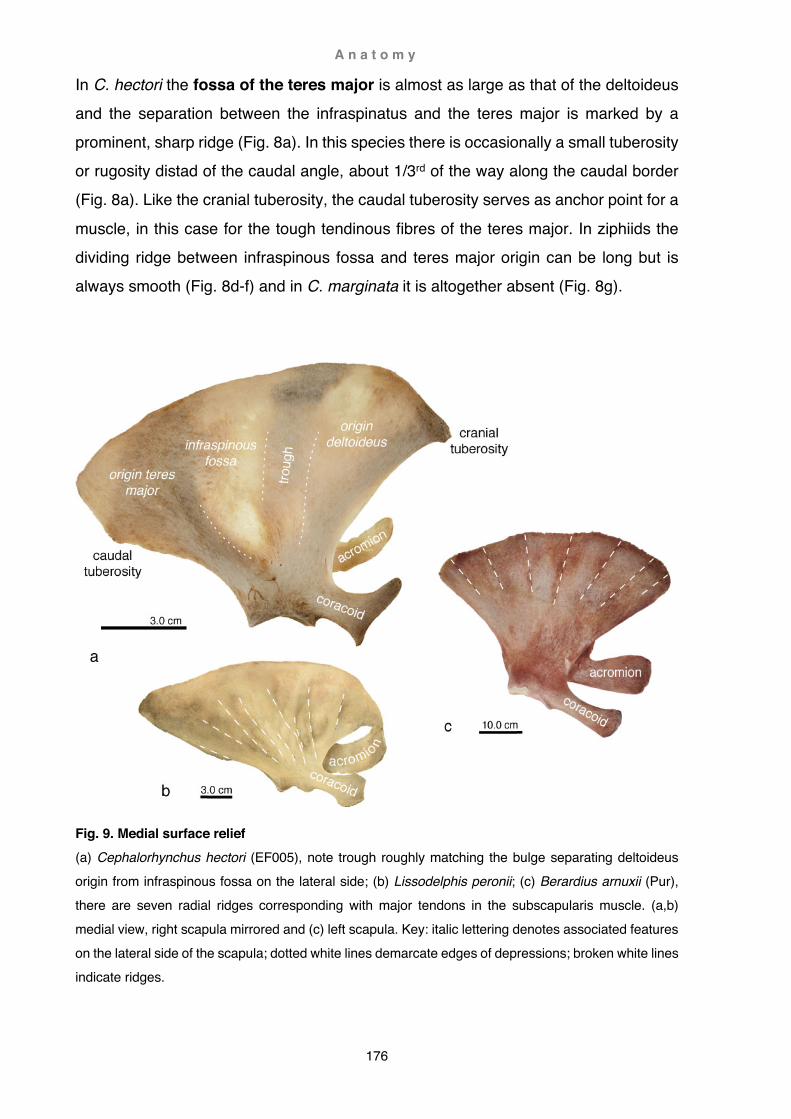
A n a t o m y
176
In C. hectori the fossa of the teres major is almost as large as that of the deltoideus and the separation between the infraspinatus and the teres major is marked by a prominent, sharp ridge (Fig. 8a). In this species there is occasionally a small tuberosity or rugosity distad of the caudal angle, about 1/3rd of the way along the caudal border (Fig. 8a). Like the cranial tuberosity, the caudal tuberosity serves as anchor point for a
muscle, in this case for the tough tendinous fibres of the teres major. In ziphiids the dividing ridge between infraspinous fossa and teres major origin can be long but is always smooth (Fig. 8d-f) and in C. marginata it is altogether absent (Fig. 8g).
Fig. 9. Medial surface relief (a) Cephalorhynchus hectori (EF005), note trough roughly matching the bulge separating deltoideus origin from infraspinous fossa on the lateral side; (b) Lissodelphis peronii; (c) Berardius arnuxii (Pur), there are seven radial ridges corresponding with major tendons in the subscapularis muscle. (a,b) medial view, right scapula mirrored and (c) left scapula. Key: italic lettering denotes associated features on the lateral side of the scapula; dotted white lines demarcate edges of depressions; broken white lines indicate ridges.

A n a t o m y
177
The medial surface of the scapula does not usually have such definite divisions, except in some cetacean species where the relief reflects the negative contour of the lateral side. In C. hectori, the bulge separating the origin of the deltoideus from the infraspinous fossa on the lateral side roughly corresponds with a trough on the medial side of the scapula (Fig. 9a). In ziphiids, low radial ridges ostensibly correlate with
tough tendons running through the subscapularis muscle (e.g. B. arnuxii, Fig. 9c). The scapula of L. peronii shows a series of prominent parallel or diverging and well-defined ridges (Fig. 9b), possibly also correlated to anchor points of septa dividing the subscapularis. The medio-laterally compressed acromion process is flat and broad in cetaceans. The base of the acromion rises barely above the plane of the blade, while the tip may dip mediad below the scapular plane (e.g. B. arnuxii, Fig. 12a). In outline it varies from spoon-shaped (e.g. L. obscurus, Fig. 10a; K. breviceps, Fig. 5a) to spatulate (e.g. C. hectori, Fig. 10b) to extremely broad (particularly B. arnuxii Tam, Fig. 10c) or long and narrow, with distinctive kink and extending as far as the cranial angle in Cuvier’s beaked whales (Z. cavirostris, Fig. 10d).
Fig. 10. Acromion process (a) Lagenorhynchus obscurus with spoon-shaped acromion; (b) Cephalorhynchus hectori (H167), with spatulate acromion; (c) Berardius arnuxii (Tam) with extremely broad acromion; (d) Ziphius cavirostris (Katiki Beach) with long and distinctively kinked acromion. Lateral view, (a-b, d) ventral left scapula and (c) right scapula mirrored. Key: white broken line (c) demarcating fused border of flange.
The coracoid process is usually smaller and particularly at the base more cylindrical
than the acromion. It can be rod-shaped (e.g. M. grayi, Fig. 11a), spatulate, spoon-shaped (e.g. L. obscurus Fig. 11b) or distally strongly flared (e.g. C. hectori, Fig. 11c).
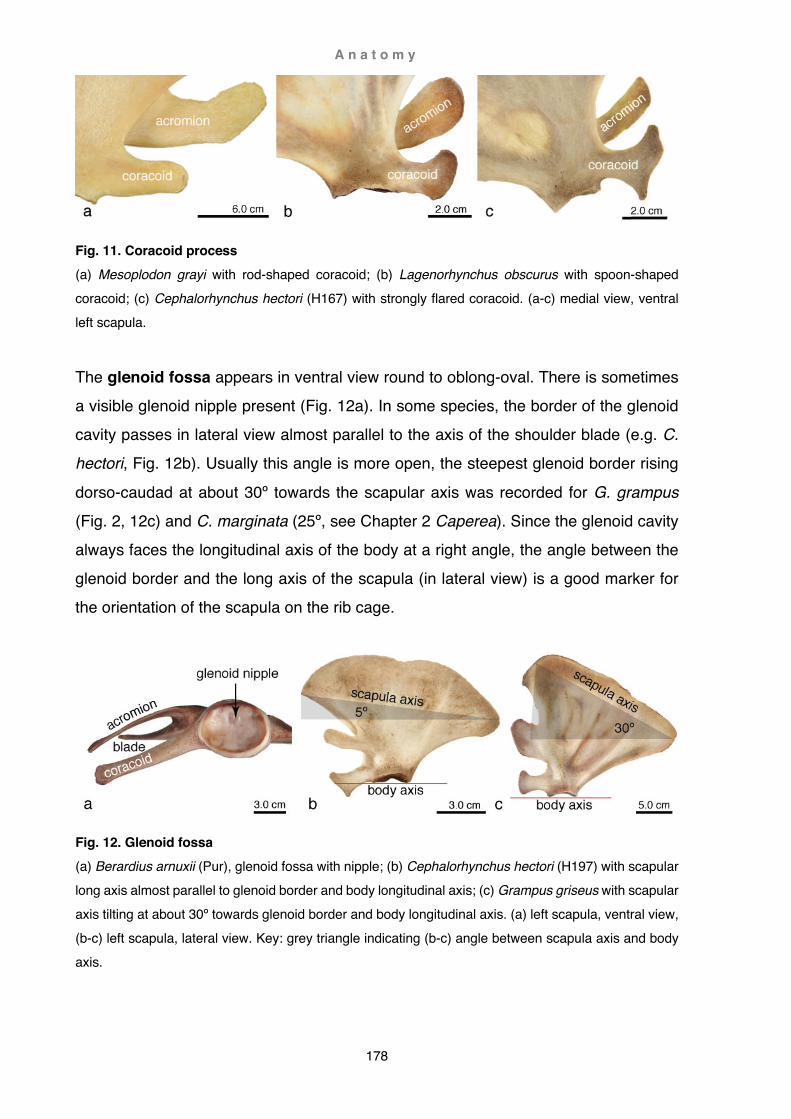
A n a t o m y
178
Fig. 11. Coracoid process (a) Mesoplodon grayi with rod-shaped coracoid; (b) Lagenorhynchus obscurus with spoon-shaped coracoid; (c) Cephalorhynchus hectori (H167) with strongly flared coracoid. (a-c) medial view, ventral left scapula.
The glenoid fossa appears in ventral view round to oblong-oval. There is sometimes a visible glenoid nipple present (Fig. 12a). In some species, the border of the glenoid cavity passes in lateral view almost parallel to the axis of the shoulder blade (e.g. C. hectori, Fig. 12b). Usually this angle is more open, the steepest glenoid border rising
dorso-caudad at about 30º towards the scapular axis was recorded for G. grampus (Fig. 2, 12c) and C. marginata (25º, see Chapter 2 Caperea). Since the glenoid cavity always faces the longitudinal axis of the body at a right angle, the angle between the glenoid border and the long axis of the scapula (in lateral view) is a good marker for the orientation of the scapula on the rib cage.
Fig. 12. Glenoid fossa (a) Berardius arnuxii (Pur), glenoid fossa with nipple; (b) Cephalorhynchus hectori (H197) with scapular long axis almost parallel to glenoid border and body longitudinal axis; (c) Grampus griseus with scapular axis tilting at about 30º towards glenoid border and body longitudinal axis. (a) left scapula, ventral view, (b-c) left scapula, lateral view. Key: grey triangle indicating (b-c) angle between scapula axis and body axis.

A n a t o m y
179
Long bones
The cetacean humerus is robust and relatively short. A hemispherical head, the caput humeri, is situated laterally or postero-laterally behind the tubercles. In some cetacean species the tubercles can be distinct as a greater (tuberculum major) and lesser tubercle (tuberculum minor, e.g. B. arnuxii, Fig. 13b-d), but in most species they are fused into one so-called proximal tubercle (Benke, 1993). In Cephalorhynchus hectori, the proximal tubercle is very little separated from the humeral head (Fig. 13a). In Berardius arnuxii the tubercles are separated by a deep sulcus from the head and from each other by a shallow groove (Fig. 13b-d). In this species there is also a pronounced humeral tuberosity on the antero-lateral surface of the humeral diaphysis (Fig. 13b). This tuberosity is very prominent in Kogia breviceps (Fig. 15d). On the lateral surface of the humeral shaft, just below the head, a more or less distinct groove, the fossa infraspinata humeri, marks the insertion point of the infraspinatus muscle. This fossa is large and deep in beaked whales (Fig. 13d).
Fig. 13. Humeral head (a) Cephalorhynchus hectori (H202) with only one so-called proximal tubercle; (b-d) Berardius arnuxii (Tam) with deep sulcus separating tubercles from humeral head. (a) left humerus, medio-caudal view; (b-d) right humerus, (b) cranial view, (c) proximal view, (d) lateral view. Key: white broken line (a) demarcating sulcus; grey broken lines delineating (b) greater tubercle, (d) border of infraspinous fossa.
Distally the humerus is flattened and cranio-caudally flared; its two distal facets are V-
shaped and interlocked with the two opposing facets of the radius and ulna. In distal view, the two facet elements are broadly-oblong and along the central line slightly dished. The ulnar facet extends in some species onto the posterior edge of the

A n a t o m y
180
diaphysis (e.g. C. hectori, Fig. 14a, B. arnuxii, Fig. 14b). In Caperea marginata and beaked whales, a synovial joint with fully formed capsule, synovial fluid and articular cartilage was found to be present (B. arnuxii, Fig. 14b). In delphinids this joint is a tightly interlocked hyaline synarthrosis covered by dense fibrous tissue (e.g. G. melas, G. macrorhynchus, C. hectori, Fig. 14a).
Fig. 14. Elbow joint (a) Cephalorhynchus hectori (H166) elbow joint is formed by tightly interlocked grey, hyaline articular cartilage, note the ulnar facet encroaching onto the caudal edge of the humerus; (b) Berardius arnuxii (Tam) synovial elbow joint with membrane between radius and ulna still intact (arrow); (c) Caperea marginata (MM2959) articular surface, above humerus and below radius and ulna. (a) left elbow joint, lateral view; (b) right elbow joint mirrored, lateral view; (c) right elbow joint with view onto articular surfaces. Abbreviations: (ol) olecranon; (syn) synovial joint, (ha rad) radial articular surface of humerus; (ha uln) ulnar articular surface of humerus.
Radius and ulna are long and slender in rorquals and C. marginata (Figs. 15a, 16a). In ziphiids (B. arnuxii, Fig. 16b) and delphinoids (L. obscurus, Fig. 15b; P. dioptrica, Fig. 15c, G. melas , Fig. 16c) the two forearm bones are more solid and shorter; the shortest antebrachial bones were recorded in K. simus and K. breviceps (Fig. 15d). The shortest ratio of humerus length / radius length was observed in the neonate neobalaenid C. marginata at about 2/3 (Fig. 15a). In L. peronii and L. obscurus (Fig. 15b) this ratio is less than 3/4, and in G. griseus, C. hectori and P. dioptrica (Fig. 15c) about 3/4 and/or slightly more. Beaked whales attain a ratio of around 1/1, whereas a ratio of 4/3 in K. breviceps (Fig. 15d) denotes a humerus longer than the radius.

A n a t o m y
181
Fig. 15. Ratio of humerus length / radius length (a) Caperea marginata (MM2959); (b) Lagenorhynchus obscurus; (c) Phocoena dioptrica (X2020.77); (d), Kogia breviceps (X2020.75). (a-d) left forelimb, lateral view; Abbreviations: (1-5) digits 1-5; (car) carpals; (dt) deltoid tuberosity; (hum) humerus; (is) interosseous space; (ol) olecranon; (p) pisiform; (rad) radius; (ul) ulna; white dotted line in (a) demarcates border between bone and cartilage; light and dark grey background represent (light grey) humerus length and (dark grey) radius and ulna length; fractional numbers (grey) indicate length ratio of humerus / long bones.
A rudimentary olecranon is present in delphinoids (e.g. C. hectori Fig. 14a, P. dioptrica Fig. 15c, G. melas Fig. 16c) but is larger in beaked whales (e.g. B. arnuxii, Fig. 16b) and well formed in baleen whales (Fig. 3a). In C. marginata the cartilaginous portion is huge (almost half the ulnar length), and either spoon-shaped (Fig. 15a) or spatulate (Fig. 16a). Long bones in baleen whales and basal toothed whales are separated by a large and elongated interosseus space (C. marginata Fig. 15a, K. breviceps Fig. 15d, B. arnuxii Fig. 16b). Benke (1993) correlated the presence of this space with the presence of antebrachial musculature. However, the interosseus space is also present in delphinoids (albeit smaller, Figs. 14a, 15c, 16c) that show no traces of antebrachial

A n a t o m y
182
musculature but instead carry bundles of blood vessels lengthwise along a markedly grooved cranial ulnar edge. With shrinkage of the cartilage during the preservation process this gap may close completely and is consequently not visible anymore in osteological collections (Figs. 15b, 19c,d and e,f).
Fig. 16. Olecranon (a) Caperea marginata (MM2904); (b) Berardius arnuxii (HiRe); (c) Globicephala melas (Otago Harbour). (a-c) left forelimb, lateral view; Abbreviations: (is) interosseous space; (ol) olecranon; (rad) radius; (ul) ulna.
Manus
There are usually five to seven ossified carpals present in the manus. Proximal carpals can be readily identified as radiale, intermedium and ulnare. Distal carpal elements (C1-5) are more difficult to determine. In many dolphin and porpoise species the distal C1 is regarded as lost (Galatius et al., 2006; Cooper et al., 2007a), and C2 and C3 variably present or lost, i.e. either C 2 present and C 3 lost (Cooper et al., 2007a) or C2 lost and C3 present (Galatius et al., 2006). Carpals 4+5 are regarded as fused (Cooper et al., 2007a). This pattern is well represented in K. breviceps (Fig. 15d). Here a large C2 articulates with MC II and MC III. However, size and position of this wrist element could be interpreted rather as a fused C2+3 rather than a single carpal. Fused C4+5 articulates with MC IV and MC V and C1 is either not present or possibly fused to MCI. The carpal arrangement in G. griseus is slightly different (Turner, 1909). Here C1 and C2 are both present and well aligned with their respective metacarpal elements, C1 articulating entirely with MC 1 and C2 almost exclusively with MC II. Fused C3+4

A n a t o m y
183
articulate with both MC III distally and MC IV caudo-distally, and C5 is absent (Fig. 17a). The position of the wrist bones in C. hectori is different again. Here C2 is slightly displaced towards MC III and articulates with both MC II and MC III (Fig. 17b). Radiographs also show frequently semi-fused or fully detached additional carpal bones
in association with C2, possibly a centrale (Fig. 17c).
Fig. 17. Carpals of the wrist (a) Grampus griseus, C1 and C2 well aligned with respective MC I and MC II; (b) Cephalorhynchus hectori (H180), C 2 displaced towards MC III; (c) C. hectori (H164) with additional semi-fused distal carpal element, possibly centrale; (d) Berardius arnuxii (Pur) with three inter-row and one intra-row fusions; (e) B. arnuxii (HiRe) with two intra-row fusions; (f) B. arnuxii (Tam) with typical U-shaped fusion of ulnare and carpal 4, note the ossified pisiform. (a-b, d-e) Radiographs of left manus, mirrored, (c, f) radiographs of right manus. Abbreviations: (1) carpal 1; (2+3) fused carpals 2 and 3; (I – V) metacarpals 1-5; * cartilaginous pisiform; (int) intermedium; (p) partly ossified pisiform; (rad) radiale; (ul) ulnare; white broken line indicates border of carpal elements; arrow in (c, d and f) pointing towards additional distal carpal element.
Carpal bones in ziphiids are highly variable (Turner, 1909), yet some patterns appear to be more or less confined to B. arnuxii. These are fusions either between proximal and distal carpals (e.g. ulnare and C4, Fig. 17d, f), or within the proximal carpal row (fusion of the intermedium and ulnare, Fig. 17e). Although these patterns have not been reported in other ziphiids, they are ostensibly also present in belugas
(Delphinapterus leucas; Kleinenberg et al., 1969). The most striking carpal

A n a t o m y
184
arrangement, and possibly unique to B. arnuxii, is a U-shaped fusion of the ulnare and carpal 4 (Fig. 17f), also reported by Flower (1872) and van Beneden (1880). Notable is also an additional (fused) distal carpal element (Fig. 17d, f), possibly C5. Other patterns are more common, such as C1 that appears variably either as an ossified short element (Fig. 17d), or as a trapezoid-shaped and subtly conical cartilaginous
element (Fig. 17e) or bone (Fig. 17f). The pisiform is usually cartilaginous (Fig. 17d-e) or only partly ossified (Fig. 17f). The smallest ratio (2/5) of manus length / pectoral limb length is represented by B. arnuxii as well as C. marginata (Fig. 18a), the only mysticete in this study where such measurements were possible. On the opposite side of the spectrum is G. melas with a hand spanning in excess of 2/3 of the total forelimb length (Fig. 18b), closely followed by G. macrorhynchus (Fig. 3f) and G. griseus (Fig. 3f) with a little under 2/3. At a value of about 3/5 the manus of K. breviceps (Fig. 18c), K. sima (Fig. 3b), L. peronii (Fig. 3f, 18d), L. obscurus (Fig. 3f) and P. dioptrica (Fig. 3d) are slightly smaller, although in K. sima and P. dioptrica the broad arrangement of the digits gives an overall impression of a rather shorter manus. Measurements in C. hectori (11/20, Fig. 18e), reveal a manus just over half of the forelimb length (Fig. 18e). In Z. cavirostris and O. orca (Fig. 18f) the hand was found to be equally long or slightly shorter than the arm bones (1/2). Overall, ziphiids had the shortest manus among odontocetes, the longest seen in Z. cavirostris (1/2), followed by M. grayi and M. layardii (9/20) and trailed by B. arnuxii (Fig. 18g). The shortest manus occurred in the youngest individual of B. arnuxii (HiRe) with a value of 3/7. Phalangeal counts reveal that cetaceans have unusually supernumerary finger elements in the second and third (odontocetes) or third and fourth (mysticetes) digits (hyperphalangy), and often fewer than the plesiomorphic mammalian phalangeal count
of 2/3/3/3/3 in the first and last digit. In tetradactyl mysticetes the first digit is lost altogether (Cooper et al., 2007a). Cetaceans also show a distinct lack of ossification in the flipper, particularly of the distal phalanges (e.g. Galatius et al., 2006; Mellor et al., 2009). This can lead to confused phalangeal counts, since non-ossified distalmost finger elements are rarely preserved in osteological collections and thus not included in counts. Equally confusing, sometimes metacarpals are also included in phalangeal counts. To confound matters even more, cetacean digits have also, besides the hourglass-shaped elements, differently shaped ‘fingertips’ (Fig. 19a). These tips are not an

A n a t o m y
185
Fig. 18. Ratio of manus length / limb length (a) Caperea marginata, trailing edge of flipper outlined (MM2959); (b). Globicephala melas (KS09-01); (c) Kogia breviceps (X2020.75); (d) Lissodelphis peronii; (4) Cephalorhynchus hectori (H199), note only partly fused distal carpals 3 and 4; (f) Orcinus orca (Maripi); (d) Berardius arnuxii (Pur). (a, d, g) radiograph of left limb, mirrored; (b-c, e-f) radiographs of right limb. Abbreviations: coloured background representing fractional total limb length of (green) brachium, and (blue) manus.

A n a t o m y
186
outgrowth of the terminal element but appear to be jointed, so should be regarded as an element of their own (Fig. 19b). Nevertheless, they are generally not referred to in the literature, hence it is not clear if they have been taken into account or not. In odontocetes, the first digit usually comprises fairly solid, conical elements and the
tip is rounded (Fig. 19c,e). However, in some cases the tip is reduced to a short, domed cap and it is not possible to differentiate a joint between the tip and the previous element. The two postaxial digits are overall much flatter and the tip often a broad, conspicuous triangular element (Fig. 19e). This is also often the case with the third digit, but to a much lesser degree (Fig. 19e). The tip of the second digit, however, is formed by a most delicate, narrow triangular element that is frequently so thin it becomes almost translucent (Fig. 19b,c,e). This pattern is similar in mysticetes, albeit in tetradactyl species it is digits 3 and 4 with pronounced hyperphalangy. As a general rule, the most terminal finger elements are not visible on radiographs and only delicate dissection of well-preserved specimens can reveal the often minute fingertips or distal cartilaginous finger elements. Even though it is not always possible to unambiguously ascertain joint boundaries between the terminal elements, with careful preparation terminal elements can be preserved. Yet, distortion of the cartilage can once again obscure original shape and articulation of individual elements and digits as a whole (Fig. 19d with little distortion, Fig. 19f strongly distorted, also note closed interosseus gap). All following phalangeal counts (exclusive of metacarpals) include cartilaginous finger elements and are thus often higher than stated in the literature. Because of the uncertainty surrounding the fingertips, they are not included here. In odontocetes, the second digit comprises the most numerous phalangeal elements
(Cooper et al., 2007a). With a maximum of 13 elements, G. melas (Fig. 19e-f) had the highest count, followed by G. macrorhynchus (Fig. 3f), L. peronii (Fig. 3f) and L. obscurus (Fig. 3f, 15b) with a count of 10 elements. G. griseus (Fig. 3f), C. hectori (Fig. 3e), and K. breviceps (Fig. 15d) follow with a somewhat lower number of around 8-9 phalanges. 5-7, the lowest number, can be seen in K. sima (Fig. 3c), P. dioptrica (Figs. 3d, 15c), O. orca (Fig. 3e) and beaked whales (B. arnuxii, Fig. 3b). In the only representative of the mysticetes, C. marginata, both the third and fourth digits were the longest with 5-6 elements (15a).
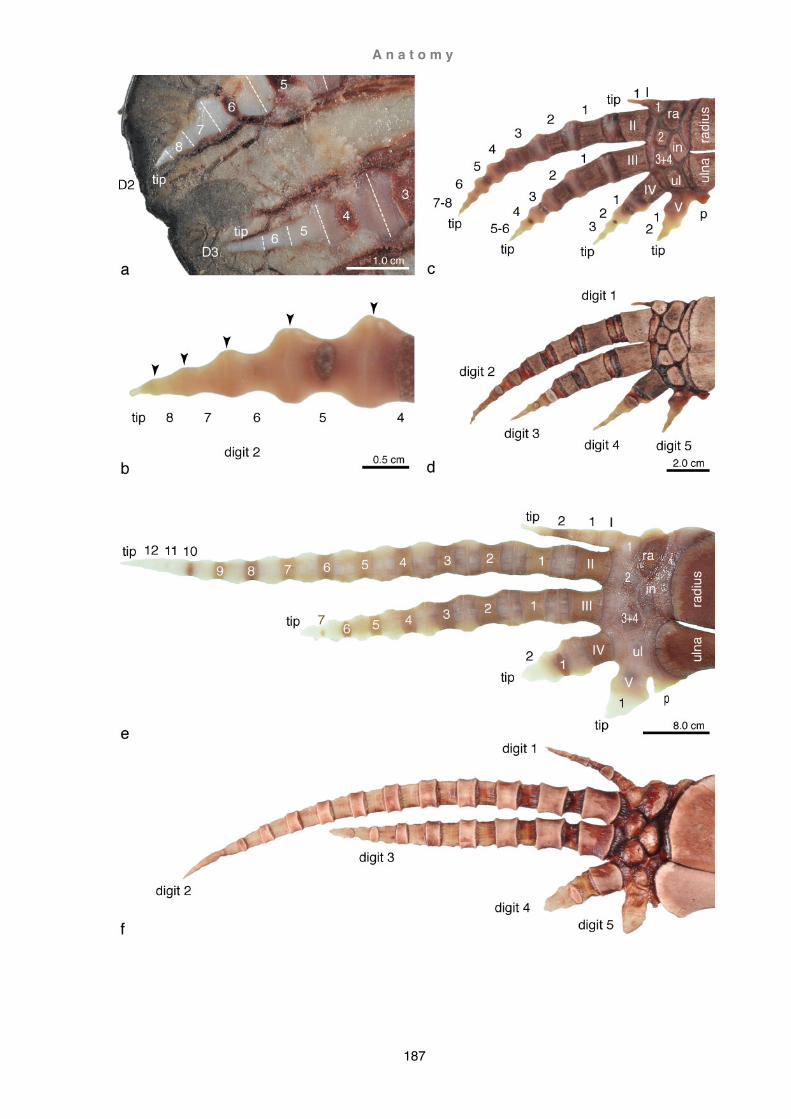
A n a t o m y
187

A n a t o m y
188
Overleaf: Fig. 19. Phalanges (a) Cephalorhynchus hectori (H176), digits 2 and 3 in situ; (b-d) Cephalorhynchus hectori (H197), terminal-most phalanges of digit 2, (c) manus immediately after dissection, (d) preserved manus; (e-f) Globicephala melas (09-01Gm), (e) manus immediately after dissection, (f) preserved manus, note distortion of wrist and phalanges due to cartilage shrinkage and closure of interosseus space. (a) right manus, medial view; (b-f) left manus, lateral view. Abbreviations: phalangeal count numbers in (a,e) white, (b,c) black; distal carpals numbered in (c,e) white; (I-V) metacarpals; (in) intermedium; (ra) radiale; (p) pisiform; (ul) ulnare; white broken lines delineate joints; black arrows indicate joints.
Myology
All details given for Grampus griseus, Phocoena dioptrica, and Cephalorhynchus hectori are based on the results of dissections described in Chapter 1 Methods and materials: Dissections and illustrated below in Figs. 20-23. C. hectori material was in poor condition and consisted of a separated shoulder-flipper unit; origins of peripheral muscles are therefore not described for this species. General cetacean forelimb anatomy, as listed in this chapter in Table 2, is predominantly based on Strickler
(1978), Smith et al. (1976), and Cooper et al. (2007b). Muscle action, if not otherwise stated, is based on the results of this study, and on Benke (1993) and Smith et al. (1976). For a more detailed review of forelimb musculature see also Chapter 5 Literature review: Myology, Table 2. Panniculus carnosus (Fig. 20a-d)
The exact anterior borders of the panniculus in G. griseus could not be established but as a whole corresponded with the muscle seen in other cetaceans (see also Chapter 3 Caperea) and covers three major regions of the body: (1) throat and neck, (2) chest and shoulder, and (3) trunk region. The muscle is roughly divided into four portions (left-right and dorsal-ventral). In the region of the shoulder the muscle is interrupted by
fenestrations for the flipper, and across the throat by a triangular aponeurosis. Left and right side of the panniculus remain separated whereas ventral and dorsal divisions merge across the lateral neck area. Arising approximately from the corner of the mouth, the dorsal division of the muscle describes a long arch just behind the external auditory meatus, rising towards the middorsal line until reaching its climax above the axilla. From here it starts a slow descent towards the genital orifices across the side of the shoulder and trunk.

A n a t o m y
189
Ventrally the panniculus emerges seamlessly from the intermandibular fasciculi and is interrupted at the throat by a triangular aponeurosis. The muscle emerges again in the area between the flippers and continues further caudad terminating in the region of the genital orifices. In the area of the throat and neck the panniculus forms an uninterrupted sheet covering the intermandibular region, parts of the throat, and the lateral neck area. Ventrally the panniculus arises in between the two rami of the lower jaw and continues towards the throat and neck area, the orientation of the fasciculi changing from horizontal to vertical. In the region of the throat, the panniculus splits at the midventral line and inserts into a triangular aponeurosis. The apex of the triangular aponeurosis separates the muscle increasingly towards the insertion of flipper. The base of the fibrous triangle reaches across the chest and delineates the cranial border of the pectoralis major. From this juncture, a small strip also covers the area immediately anterior to the flipper, merging at this point with the aponeurosis of the shoulder. Rising laterally across the neck, the direction of the muscle fasciculi gradually changes from transverse to dorso-craniad. The aponeurosis of the shoulder covers the area immediately anterior and dorsad of the flipper, while the dorsal and caudal aspects of the shoulder are concealed by muscle tissue. A slight concentration of fasciculi is achieved with increased inclination in the area where the fasciculi of the shoulder merge into the fasciculi of the trunk. From here the fasciculi descend ventro-cranially to insert into the axllary raphe. In the area of the trunk, the muscle is divided into four portions, two on each side, separated by mid-dorsal and mid-ventral aponeuroses. Each side in turn is divided by a lateral aponeurosis dividing the muscle into a dorsal and a ventral division.
The dorsal division emerges uninterrupted from the shoulder with fasciculi rising from the lateral raphe slightly more dorso-caudadally inclined. The anterior fibres oppose the posterior fibres of the pectoralis major in the region of the axilla and insert aponeurotically as a raphe into the caudal aspect of the humerus (axillary raphe). Further caudad, this raphe merges into a narrow lateral raphe. The ventral division is a continuation of the pectoralis major; a clear division, however, could not be established as the chest of the specimens was still frozen at the time of dissection. Rising from the ventral raphe, the inclination of the fasciculi increases from transverse in the chest region to dorso-craniad further caudad.

A n a t o m y
190
Fig. 20. Panniculus Semi-schematic illustrations of panniculus showing fasciculi extent and inclination. Grampus griseus (a) lateral view, (b) ventral view; Phocoena dioptrica (c) lateral view, (d) ventral view. Parallel slanted lines show inclination of the fasciculi, broken lines indicate inferred extent of the muscle and fasciculi inclination.

A n a t o m y
191
In Phocoena dioptrica the panniculus is largely congruent with that of the harbour porpoise (Phocoena phocoena) as described by Stannius in 1849, and similar to that seen in Risso’s dolphin. There are four main divisions covering the trunk area, separated by a thin raphe ventrally and a very wide lateral raphe over the flanks. In the shoulder region, just above the humerus, there is a small interruption of fasciculi, but
no gap separating the fasciculi of the neck from those of the shoulder. There is, however, a prominent bulge of the shoulder aponeurosis that covers the anterior part of the flipper base. Contrary to Risso’s dolphin, the aponeurosis of the chest is small and restricted to the sternum.
Muscles of the shoulder and forelimb

A n a t o m y
192
Fig. 21. Origin and insertion of peripheral forelimb musculature Semi-schematic illustration of Grampus griseus. Extent of muscles symbolised by shaded areas outlined in bold. Skeleton adapted from van Beneden, 1880
Colour index of muscle groups Figs. 21-24 (sensu Strickler, 1978): brown, costo-spino-scapular; red+orange, latissimus-subscapular; purple, deltoid; yellow, triceps; green, suprascapular; blue, flexor; turquoise, trapezius; pink, manus.
Overleaf: Figs. 22-24. Origin and insertion of lateral and medial forelimb musculature Semi-schematic illustration of (a) lateral and (b) medial muscle origins and insertions in (21) Grampus griseus (VT3301), (22) Phocoena dioptrica (X2020.77), (23) Cephalorhynchus hectori (H199). Areas in light grey represent bone, medium grey (long bones) and white (manus) cartilage, and dark grey corresponds to the silhouette of the flipper. Abbreviations: (1-5) digits 1-5.

A n a t o m y
193
Fig. 22 a. Lateral forelimb musculature of Grampus griseus

A n a t o m y
194
Fig. 22 b. Medial forelimb musculature of Grampus griseus

A n a t o m y
195
Fig. 23 a. Lateral forelimb musculature of Phocoena dioptrica

A n a t o m y
196
Fig. 23 b. Medial forelimb musculature of Phocoena dioptrica
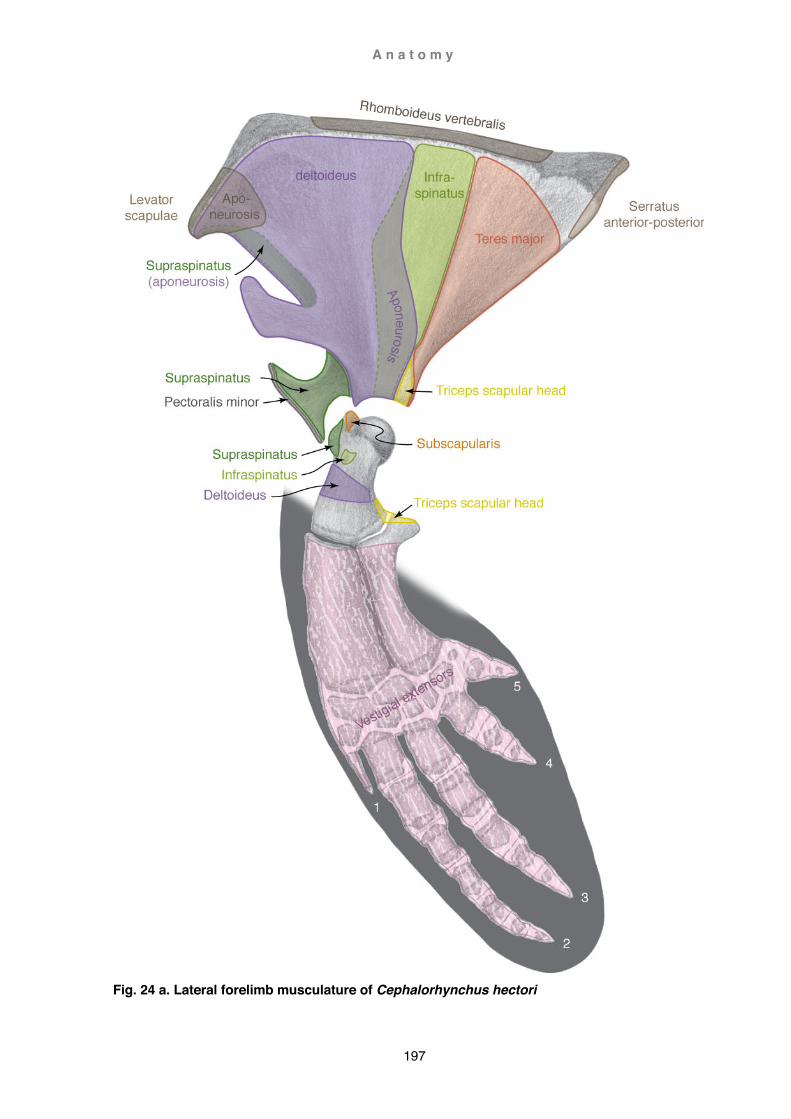
A n a t o m y
197
Fig. 24 a. Lateral forelimb musculature of Cephalorhynchus hectori

A n a t o m y
198
Fig. 24 b. Medial forelimb musculature of Cephalorhynchus hectori

A n a t o m y
199
Levator scapulae (Figs. 23a-b, 24a)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Absent Transverse process of atlas Not known Ventral arch of atlas
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Absent Lateral cranial angle Lateral cranial angle Lateral cranial angle of scapula, aponeurotic on deltoid
Action Slight craniad movement of scapula
In G. griseus a muscle inserts deep to the rhomboideus vertebralis onto the
cartilaginous edge of the cranial angle, a configuration which could be interpreted as similar to that described by Schulte (1918) in Kogia breviceps. However, two aspects stand against this assumption: (1) the muscle seen in G. griseus is sheet-like and not a flat belly; (2) it can be traced back to the first rib, a position more in accordance with the serratus anterior - anterior division (see description of this muscle below). The levator scapulae is for these reasons deemed absent in G. griseus.
The levator scapulae arises in P. dioptrica from the fused atlas-axis transverse process and inserts laterally as a massive flat belly onto the aponeurosis of the deltoid in the region of the cranial angle. From here long tendinous fibres spread caudad over almost the whole width of the infraspinatus, anchoring the levator scapulae firmly onto the lateral side of the shoulder blade musculature. Anteriorly, the muscle overlaps the cranial edge of the scapula and inserts mediad slightly onto the subscapularis as a thin fleshy membrane.
In C. hectori the muscle inserts onto the lateral cranial angle as a small, thin belly and spreads aponeurotically across the area of the anterior portion of the deltoideus. From here the muscle sends long tendinous fibres further caudad across the deltoideus and infraspinatus. However, the origin of this muscle is not known and consequently the rhomboideus capitis, with origin from the back of the skull rather than the anterior cervicals, must be considered as another possible candidate. Against this stands, apart
from a few exceptions, a very constant presence and arrangement of the levator scapulae in cetaceans (see Table 2). For these reasons, the levator scapulae is

A n a t o m y
200
tentatively regraded as present, and the rhomboideus capitis as absent in Hector’s dolphins. Serratus anterior - anterior division (Figs. 21, 22b, 24b)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
First rib or posterior cervicals Absent Not known Transverse process of cervical vertebra C7
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Medial cranial angle Absent Medial cranial angle Medial cranial angle of scapula
Action Slight craniad movement of scapula
In G. griseus the anterior division of the serratus anterior inserts as a thin but strong sheet into the deep cranial angle of the scapula. It is interposed between the subscapularis on the deep side and the anterior part of the rhomboideus vertebralis, which is slightly displaced ventrad from the dorsal edge and confined to the lateral side of the cartilaginous border. The origin of this muscle could not be exactly determined due to the frozen state of the deeper musculature. Even so, it could be traced back to the area of the first rib, not excluding a deeper origin from posterior cervicals and is therefore regarded as the anterior division of the serratus anterior.
A serratus anterior - anterior division is not present in P. dioptrica.
In C. hectori a muscle inserts onto the medial cranial angle and foremost part of the vertebral border as a bulky fleshy sheet, extending caudad about halfway along the cartilaginous vertebral border. Apart from G. griseus, a similar arrangement was also observed in Caperea marginata (this study), Balaenoptera acutorostrata (Carte and Macalister, 1868), †Lipotes vexillifer (Weijuan, 1981), Pontoporia blainvillei (Strickler, 1978), Phocoena phocoena (Smith, 1976), Delphinus delphis and Tursiops truncatus (Sokolov and Rodionov, 1974; Benke, 1993; see also Table 2).

A n a t o m y
201
Serratus anterior - posterior division (Figs. 21, 22b, 23a-b 24a-b)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
2-4th ribs 1-3rd or 2-4th ribs Not known 2-4th ribs
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Medial posterior cartilaginous vertebral border Medial cartilaginous vertebral border Medial posterior cartilaginous vertebral border Medial cartilaginous vertebral border of scapula
Action Adducts scapula ventrad, slight rotation craniad
In G. griseus this muscle arises on the second rib and, under the concealment of the
latissimus dorsi, the third and fourth ribs. At the point of insertion, the muscle occupies a triangular area restricted to the deep posterior cartilaginous vertebral border of the scapula. The base of the triangle abuts the posterior border of the caudal cartilaginous angle, also encroaching onto the posterior edge between the deep and superficial surfaces of the scapula. Anteriorly the insertion narrows to a point along the posterior two-thirds of the cartilaginous border, but does not encroach onto its dorsal edge. Ventrally the muscle inserts onto the fascia of the subscapularis corresponding to about one quarter of the dorsal bony scapular area and about two thirds of the posterior vertebral scapular border.
In P. dioptrica this muscle arises from the first to third or second to fourth ribs and inserts onto the medial surface of the cartilaginous vertebral border. Along the cranial 2/3rd of the border, the muscle runs as a thin strip just below the top edge, widening ventrad towards the caudal 1/3rd of the border. Here it encroaches onto the medial surface and caudal edge of the bony blade, displacing the posterior part of the subscapularis and, on the lateral side, the teres major.
The posterior division of the serratus anterior inserts in C. hectori just behind the anterior division onto the medial surface of the cartilaginous vertebral border and caudal angle. It runs along the top edge of the posterior half of the cartilaginous border and gradually widens towards the caudal angle, there encroaching onto the bony blade
and slightly displacing the subscapularis. Similar to P. dioptrica, the muscle also straddles the posterior border of the caudal angle, displacing the teres major further

A n a t o m y
202
ventrad. The dorsal border of the teres major also appears to be displaced ventrad by the posterior division of the serratus anterior in Balaenoptera borealis and Tursiops truncatus (Sokolov and Rodionov, 1974; Benke, 1993). Rhomboideus capitis (Figs. 21, 22a-b)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Temporal crest Absent Absent Supraoccipital
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Lateral cranial angle Absent Absent Lateral cranial angle of scapula
Action Slight craniad movement of scapula
In G. griseus a thin but fleshy muscle arises from the angle formed by the nuchal and temporal crest (sensu Fordyce, 1994; Mead and Fordyce, 2009) above the temporal fossa, diminishing rapidly to scant tendinous fibres. Approaching the lateral cranial angle of the shoulder blade, the muscle becomes fleshy just before inserting onto the fascia covering the deltoid. A few long tendinous fibres extend further onto the fascia of the deltoid further caudad. Compared to Kogia (Schulte and Smith, 1918) this muscle appears to be vestigial in G. griseus.
This muscle is not present in P. dioptrica.
In C. hectori the rhomboideus capitis is most likely absent (see also levator scapulae).

A n a t o m y
203
Rhomboideus vertebralis (Figs. 21, 22a, 23a, 24a)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Epaxial fascia ribs 2-5 Epaxial fascia ribs 1-6 Not known Epaxial fascia between thoracic vertebrae T1-6
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Lateral cartilaginous vertebral border Lateral cartilaginous vertebral border Lateral cartilaginous vertebral border Lateral vertebral border of scapula
Action Adducts scapula dorsad
In G. griseus only the anterior-most portion of this muscle arises aponeurotically on the
epaxial musculature, whereas the remaining portion ascends from the fascia over the second to fifth ribs approaching the sixth rib. Midway, the muscle is divided into an anterior, superficial portion (serratus vertebralis superficialis) that slightly overlaps the posterior, deeper portion (serratus vertebralis profundus). Although the partition is not very conspicuous and not discernible at all at the point of insertion, the two sections are marked by a directional change of the fasciculi from slightly dorso-caudad anteriorly to a more dorso-ventrad direction posteriorly. At the insertion, the muscle straddles the posterior cartilaginous vertebral edge of the scapular border to about midway from where it deviates by a few millimetres onto the superficial side of the cartilaginous scapular border. At its posterior end the rhomboideus vertebralis does not quite reach the caudal angle and is separated from the encroaching serratus anterior - posterior division, by a thin tendinous interception. Craniad, the rhomboideus vertebralis conceals the caudal portion of the serratus anterior - anterior division by inserting onto the superficial surface of the vertebral border, rather than the edge, but remains restricted to a narrow strip at the upper limit of the cranial angle.
In P. dioptrica the rhomboideus vertebralis is also divided into two portions (superficialis and profundus), chiefly recognisable by a change of direction of the fasciculi. The superficial portion arises over the fascia of ribs 1-3, the profundus portion over rib 2 approaching rib 4. The two portions were not discernible anymore once the
limb was separated from the trunk. The muscle inserts over the whole length of the lateral vertebral border of the scapula as a long and narrow strip, also straddling the

A n a t o m y
204
top edge of this border. However, it does not quite reach the caudal edge of the cartilaginous border from where it is displaced by the serratus anterior - posterior division.
In C. hectori the rhomboideus vertebralis inserts as a long, narrow strip over 2/3rd of the lateral vertebral border. The muscle is displaced from the cranial angle by the
serratus anterior - anterior division and from the caudal angle by the posterior division of the serratus anterior. Latissimus dorsi (Figs. 21, 22b, 23b, 24b)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
4-6th ribs with fibres to 11th rib 5-8th or 9th ribs Not known 4-6th ribs
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Caudal border below humeral neck Caudal border below humeral neck Caudal border below hummeral neck Mid medio-caudal edge of humerus
Action Flexes humerus
In G. griseus the muscle arises fleshy just posterior of the 4th rib crossing the 5th and 6th rib but not quite reaching the 7th rib from where tendinous bands extend caudally to the 11th rib. Insertion is in common with the teres major along the medial mid-posterior edge of the humerus deep to the triceps.
This muscle arises in P. dioptrica from the 3rd to 5th ribs, also extending tendinous fibres to the 8th or 9th rib. It inserts in common with the teres major just below the humeral head on the caudal aspect of the humerus.
In C. hectori the latissimus dorsi also follows the usual cetacean pattern and inserts in common with the teres major along the posterior edge of the humerus just below the humeral head.

A n a t o m y
205
Teres major (Figs. 22a-b, 23a-b, 24a-b)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Lateral posterior 1/4 of scapula Caudal border of scapula Lateral posterior 1/3 of scapula Caudal border of scapula
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Mid-caudal aspect of humerus Mid-caudal aspect of humerus Mid-caudal aspect of humerus Mid-medio-caudal edge of humerus
Action Flexes and/or rotates humerus inwards
The teres major in G. griseus occupies the posterior lateral surface of the scapula,
arising from the posterior quarter of the (bony) vertebral border, the caudal border and superficial surface of the caudal angle. The muscle is clearly demarcated by a deep fossa on the scapular surface but also encroaches anteriorly beyond this boundary aponeurotically onto the infraspinatus, broadening at its widest extent over a third of the posterior lateral scapular surface. Posteriorly the muscle straddles the caudal scapular border, separated from the subscapularis by a tendinous sheet. The insertion onto the posterior deep edge of the humeral shaft (in common with the latissimus dorsi) is typical for odontocetes.
In P. dioptrica the teres major is much more confined than in the two dolphins and only covers about 1/7th of the scapular surface. Moreover, the muscle does not reach the (bony) vertebral border and instead terminates considerably lower than the deltoideus and infraspinatus, extending at the most about 5/6th of the scapular height. The upper 1/6th above the teres major is covered by a smooth, tendinous fascia and the outermost caudal edge is occupied by the posterior division of the serratus anterior. Craniad, the anterior border of the teres major tightly abuts the infraspinatus, while the posterior portion of the muscle adheres to the caudad-facing surface of the scapular neck, also slightly impinging onto the medial posterior-most fringe of the blade. Across the post-scapular area, the teres major is separated from the subscapularis by a tough, fibrous tendinous sheet. Insertion is in common with the latissimus dorsi along the posterior-
facing area of the humerus just below the humeral head.

A n a t o m y
206
In C. hectori this muscle is relatively large, occupying about 1/3rd of the posterior lateral scapular surface, caudally deviating in a sweeping curve towards the lower caudal border. As in P. dioptrica, the area above the teres major is covered by a smooth, tendinous fascia and the outermost caudal edge is occupied by the posterior division of the serratus anterior. Ventrally the teres major is displaced caudad by the scapular
head of the triceps, adhering instead to the caudal-facing surface of the neck and the lateral-most posterior margin of the shoulder blade. Insertion is in common with the latissimus dorsi caudo-medially onto the neck just below the humeral head. Subscapularis (Figs. 22a-b, 23a-b, 24a-b)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Entire medial surface of scapula Entire medial surface of scapula Entire medial surface of scapula Medial surface of scapula
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Proximal + lateral surface of tubercle Proximal + lateral surface of tubercle Proximal + lateral surface of tubercle Caudal surface of humeral tubercle
Action Fixation of shoulder joint Adducts and/or rotates humerus inwards
In G. griseus the subscapularis covers the entire deep surface of the scapular blade and anterior adjacent area between coracoid and cranial angle, also encroaching substantially onto the supraspinatus. The whole muscle is interlaced with tough tendons strongly attached to the surface of the bone, forming seven major dorso-ventrally fan-shaped demarcations, some of which are bifurcated. Superficial tendinous bands also strongly attach on the vertebral border of this muscle. Insertion is tendinous onto the deep (more lateral) and proximal surface of the humeral tubercle.
In P. dioptrica origin and insertion of the subscapularis largely correspond with that of G. griseus. The muscle is divided by multiple tendons, yet they are not as well developed and consistent as in G. griseus. Curiously, in P. dioptrica the subscapularis is divided into a larger anterior bay and a smaller posterior bay, the latter displaced
ventrally by the posterior division of the serratus anterior. Ventrally, insertion of the

A n a t o m y
207
muscle is congruent with that of G. griseus. In one specimen (OM X2020.77), a slip continues posteriorly and further mediad encircles the insertion of the coracobrachialis almost entirely like a thick, fleshy finger (see Fig. 23b).
Abutting the supraspinatus anteriorly, the subscapularis of C. hectori covers most of the medial scapular blade. Cranially, the muscle is slightly displaced by the anterior
division of the serratus anterior, and caudal-most by its posterior division. In this manner, the subscapularis is divided into two equally broad, shallow bays - the anterior bay slightly lower than the posterior bay. Superficial tendinous bands cover the caudal half of the subscapularis, but no deep divisions adhering to the blade have been observed. Insertion is congruent to that of the other two species. Deltoideus (Figs. 22a-b, 23a-b, 24a)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Lateral cranial ½ of scapula Lateral cranial ½ of scapula Lateral cranial ½ of scapula Cranial lateral surface of scapula
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Lateral mid to distal humerus shaft Lateral mid to distal humerus shaft Lateral mid humerus shaft Lateral surface of humerus
Action Whole muscle abducts humerus Cranial part extends and reverses inward rotation of humerus
In G. griseus the deltoideus arises from the anterior half of the scapula and encroaches posteriorly as an aponeurosis onto the infraspinatus, most prominently towards the glenoid fossa. Craniad, the deltoid extends over the tip of the acromion mediad towards the coracoid, also completely concealing the supraspinatus above and below the acromion. Ventrally, deltoid connective tissue intermingles with those of the pectoralis minor and pectoralis major. Over the scapular blade, the deltoid is covered by a strong tendinous sheet and interlaced with five tough tendinous bands attaching firmly in a fan-shaped pattern across the acromial and scapular surface, the fifth band (on the left side, sixth band on the right on the right) attaching dorsally as a V-shaped divergence.
Insertion is onto a bulbous tuberosity at the distal anterior border of the humerus and

A n a t o m y
208
the superficial surface of the shaft, where the muscle forms a cuff traversing antero-posteriorly. Anteriorly this cuff extends across the cranial edge of the humerus and is mirrored on the deep surface by the pectoralis major, the only separation of these two muscles being recognisable by a small tendinous, non-fleshy interruption on the deep surface just behind the anterior border.
The deltoideus of P. dioptrica is large, covering over half of the scapular surface anterior to the infraspinatus. It arises from the scapular blade, acromion and the pre-scapular aponeurosis covering the supraspinatus. Cranially the muscle extends mediad and also inserts onto the tip of the coracoid. Caudally the deltoid encroaches increasingly onto the infraspinatus. From the point of insertion to about the height of the acromion, the deltoid is covered by a strong tendinous sheet that traverses the surface of the muscle cranio-dorsad. Contrary to the Risso’s dolphin, there are no tendinous bands adhering to the aponeurosis of the scapular blade. Under the cover of the tendinous sheet, the deltoid inserts as a fleshy band just below the midpoint on the lateral humeral surface. The remainder of the muscle inserts tendinously and covers the entire lower half of the bone, also extending anteriorly to some extent onto the medial surface.
The deltoideus of the Hector’s dolphin is also large, covering over half of the scapular surface anterior of the infraspinatus. It arises from the scapular blade, acromion and the pre-scapular aponeurosis covering the supraspinatus. Notably in the specimen investigated (UO H255), both the deltoideus and the underlying supraspinatus extend considerably further craniad than the tip of the acromion, but do not pass the tip of the coracoid. Caudally the deltoid encroaches onto the infraspinatus. From the point of insertion to about the height of the acromion, the deltoid is covered by a strong tendinous sheet that traverses the surface of the muscle cranio-dorsad, similar to that
seen in the other dolphin and porpoise. As in the spectacled porpoise, there are no tendinous bands adhering to the aponeurosis of the scapular blade. Under the cover of the tendinous sheet, the deltoid inserts as a fleshy band at about the midpoint on the lateral humeral surface, the remainder of the muscle inserts tendinously and covers the lower half of the bone.

A n a t o m y
209
Teres minor / Subdeltoideus (not depicted)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Absent Absent Absent Mid-lateral surface of scapula
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Absent Absent Absent Proximal lateral surface of humerus
Action Rotates humerus inwards
This muscle is usually absent in odontocetes but has been reported as subdeltoideus
in Kogia breviceps (Schulte and Smith, 1918), and as teres minor in Phocoena phocoena (Smith et al., 1976), Inia geoffrensis (Klima et al., 1980) and †Lipotes vexillifer (Weijuan, 1981; see also Chapter 4 Functional anatomy: Phylogenetic patterns of muscle distribution). It is absent in G. griseus, P. dioptrica and C. hectori. Triceps brachii (Figs. 22a-b, 23a-b, 24a-b)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Scapular head: caudal scapular neck Possibly humeral head: posterior humeral shaft, vestigial Humeral head: posterior humeral shaft, vestigial Scapular head: caudal scapular neck Scapular head: caudal neck of scapula (long head) Humeral head: lateral on humerus shaft (external head) Humeral head: medial on humerus shaft (medial head)

A n a t o m y
210
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Scapular head: posterior distal humeral edge and olecranon Humeral head not discernible Humeral head: olecranon, vestigial Scapular head: proximal edge of olecranon Scapular or humeral heads: olecranon
Action Scapular head flexes and rotates humerus inwards Humeral head not known
In G. griseus the triceps brachii is present as a scapular head in accordance with Murie’s (1870b) description of this species. The scapular head arises from a narrow, triangular area latero-caudal on the scapular neck, between the teres major caudo-mediad and the infraspinatus craniad. The muscle inserts along the posterior edge of the humerus as a broad, thin slip from just below the posterior border of the humerus head to the top of the olecranon. In other cetacean species, fibres extending from the proximal base of the olecranon to the proximal caudal border of the humerus have been regarded as the humeral head of the triceps (Carte and Macalister, 1868; Turner, 1885; Schulte, 1916). In Risso’s dolphin there is no clear division of fasciculi into two heads and the origin of the triceps scapular head might be much more extensive (Murie’s interpretation) than usually seen. Alternatively, the scapular head could be interpreted as strictly confined to the olecranon and scapula, an arrangement that has been described in P. phocoena by Sokolov and Rodionov, 1974. Consequently, the fasciculi extending from the proximal base of the olecranon and along the caudal border of the humerus to just below the humeral head could be regarded as an additional triceps humeral head.
There is no scapular head of the triceps present in P. dioptrica, but a few thin fibres may be regarded as indicating a vestigial humeral head. Alternatively, these fibres
might be part of the teres major.
A scapular head of the triceps arises just above the glenoid fossa wedged between the deltoideus and infraspinatus anteriorly and the teres major posteriorly. From here the triceps passes the conjoined teres major and latissimus dorsi laterad to insert onto the posterior distal-most border of the humerus and proximal part of the olecranon. A humeral head was not observed in Hector’s dolphin.

A n a t o m y
211
Supraspinatus (Figs. 22a-b, 23a-b, 24a-b)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Cranial scapular border Supraspinous fossa of scapula Supraspinous fossa of scapula Supraspinous fossa of scapula
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Cranial on humeral tubercle Cranial on humeral tubercle Cranial on humeral tubercle Greater tubercle of humerus
Action Fixation of shoulder joint Extends and rotates humerus outwards
In G. griseus the supraspinatus arises from an extremely reduced, nondescript supraspinous fossa. Laterally and medially, the muscle’s only bony supports are the scapular processes. The main attachment points for this muscle are therefore the prominent spur of the cranial angle, the cranial scapular border, acromion and coracoid processes, pre-scapular septum and thick lateral and medial fibrous tendinous sheets. Laterally the supraspinatus is completely concealed by two aponeuroses: one overlies the pre-scapular septum and spans from the cranial tuberosity of the scapula to the dorsal border of the acromion; the other extends from the ventral border of the acromion to about the level of the glenoid fossa and does not quite entirely cover the supraspinatus cranially. This is also the case medially, but here the sheets only extend as far forward as the tip of the coracoid process, leaving the anterior dorsal portion of the muscle exposed. The supraspinatus inserts onto the cranial aspect of the humeral tubercle.
The supraspinous fossa is fairly wide in P. dioptrica when compared to the delphinids, but neither the fossa nor the muscle reaches the vertebral border. Laterally, the supraspinatus is entirely covered by the deltoideus, medially by the subscapularis and further ventrally by the coracobrachialis. Cranially, the supraspinatus is concealed by the levator scapulae dorsally and pectoralis minor ventrally. As observed in G. griseus, the muscle is wrapped in a thick fibrous tendinous sheet. Distally the supraspinatus inserts onto the cranial aspect of the proximal tubercle.

A n a t o m y
212
Although the supraspinous fossa is fairly narrow in C. hectori, the muscle is well developed. Laterally, the supraspinatus is completely covered by the deltoideus, but medially not entirely concealed by the subscapularis. Ventrally the supraspinatus is obscured by the coracobrachialis and the pectoralis minor further craniad. A thick, coarse tendinous sheet was noted on the lateral side between the deltoideus and
supraspinatus but not between the other muscles. As in the other dolphin and the porpoise, the supraspinatus inserts distally onto the cranial aspect of the proximal tubercle.
Infraspinatus (Figs. 22a, 23a, 24a)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Infraspinous fossa Infraspinous fossa Infraspinous fossa Caudo-lateral surface of scapula
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Lateral humeral shaft Lateral humeral shaft Lateral humeral shaft Cranio-lateral surface of humerus
Action Fixation of shoulder joint Abducts humerus
The infraspinatus arises in G. griseus from a distinct infraspinous fossa on the lateral surface of the scapula. The infraspinous fossa extends from mid-scapula caudally covering about a quarter of the scapular surface. As in other cetaceans, both the deltoideus and the teres major encroach onto this muscle substantially, the deltoid covering about one quarter of its width anteriorly, and the teres major up to its midline posteriorly. Like the deltoideus, the infraspinatus is also covered by a strong tendinous sheet but divided by only a singular tough tendinous band attaching to the scapular surface. The muscle inserts onto the antero-lateral aspect of the humerus just below the humeral head and under cover of the deltoideus.
Wedged between the deltoid anteriorly and the teres major posteriorly, the infraspinatus occupies the third quarter of the scapular surface in P. dioptrica. Craniad,
the infraspinatus extends forward under cover of the deltoideus; in one specimen some

A n a t o m y
213
fibres almost attain the scapular spine near the dorsal edge of the acromion. Contrary to G. griseus, the infraspinatus is caudally very little obscured by the teres major. Distally the muscle inserts onto the mid-lateral surface of the humerus, the area in some specimens being marked by an infraspinous fossa.
The infraspinatus is relatively narrow in C. hectori, covering about 1/8th of the scapular
surface. Cranially the infraspinatus is covered by the deltoideus, and caudally the muscle is confined (but not obscured) by the teres major and scapular head of the triceps. Distally the muscle inserts below the humeral head onto the middle of the lateral surface of the humerus. Pectoralis minor (Figs. 22b, 23b, 24b)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Lateral anterior edge of sternum Lateral tip of sternum Not known Sternum
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Tip of coracoid process Tip of coracoid process Tip of coracoid process Tip of coracoid process
Action Rotates scapula ventrad
The pectoralis minor arises in G. griseus as a moderately wide sheet from the lateral anterior edge of the sternum. Continuing as a sheet, the muscle inserts onto the coarse aponeurosis of the supraspinatus, the tip of the coracoid process, and further ventrad onto the connective tissue of the supraspinatus, there also intermingling with the connective tissue surrounding the deltoid, mastohumeralis and pectoralis major.
Evisceration of the carcasses prior to dissection made it difficult to trace the origin of this muscle in P. dioptrica, possibly from the anterior lateral tip of the sternum under cover of the pectoralis major. The muscle inserts inconspicuously onto the medial surface of the outermost coracoid tip and onto the deltoideus dorsally as well as the coarse aponeurosis of the supraspinatus ventrally.

A n a t o m y
214
The origin of the pectoralis minor could not be traced in C. hectori, but a muscle inserting onto the tip of the coracoid corresponds with this muscle in other cetaceans. From the outer-most tip of the coracoid, the muscle extends onto the deltoideus dorsally as well as abutting the pectoralis major ventrally, similar to the condition seen in Risso’s dolphin.
Pectoralis major (Figs. 22b, 23b, 24b
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Ventral surface of sternum Not known Not traced Sternum
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Medial collar on humerus Central medial humeral shaft Medial collar on humerus Medial on flipper
Action Whole muscles adducts and caudal part flexes humerus
In G. griseus the separation of the panniculus and pectoralis major was difficult due to the frozen state of the tissue in the ventral region. The muscle arises on the ventral surface of the sternum extending to the midline. At its insertion the pectoralis major forms the medial part of the collar that is laterally continued by the deltoideus. Marked by a thin tendinous interruption the muscle inserts aponeurotically just medial to the anterior border of the humerus, its anterior edge also separated further distally from the fibres of the pectoralis minor by a small tendinous band. Crossing the humeral medial surface transversely, the muscle inserts caudally into the layer of fibrous blubber covering the proximal caudal portion of the ulna.
The pectoralis major arises in P. dioptrica from the midline of the sternum from the first costo-sternal joint to about the level of the third rib. The muscle inserts onto the medial surface of the humerus, anteriorly abutting the pectoralis minor and posteriorly adhering to the subscapularis before converging with the fibres of the panniculus into the axillary raphe of the flipper. In two specimens, the pectoralis major also inserted a
slip onto the proximal tubercle just distal of the coracobrachialis.

A n a t o m y
215
In C. hectori, the pectoralis major inserts transversely on the medial surface of the humerus, anteriorly abutting the pectoralis minor before converging with the fibres of the panniculus into the axillary raphe of the flipper similar to the arrangement seen in Risso’s dolphin.
Pectoralis abdominalis (not depicted)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Absent Absent Absent? Coastal cartilage 4-6
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Absent Absent Absent? Medial, posterior base of flipper
Action Flexes humerus (inferred sensu Strickler, 1978)
In G. griseus the deeper frozen layers of muscle only allowed a rough dissection. There was no muscular layer discernible deep to the pectoralis major and caudal to the forelimb; thus the pectoralis abdominalis is here tentatively regarded as absent.
The origin of this muscle could not be ascertained in P. dioptrica (because of damage by prior evisceration) and no corresponding fibres at the insertion point were noticed, so this muscle is provisionally regarded as absent.
In C. hectori, there was no muscle inserting in the region reported for this muscle in other cetaceans (see Strickler, 1978).

A n a t o m y
216
Coracobrachialis (Figs. 22b, 23b, 24b)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Medial surface of coracoid Medial surface of coracoid Medial surface of coracoid Medial surface of coracoid process of scapula
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Medial central tubercle Medial central tubercle Medial central tubercle Just distal to lesser tubercle of humerus
Action Fixation of shoulder joint Adducts and slightly flexes humerus
The coracobrachialis arises in G. griseus from the medial surface of the coracoid, caudal to the pectoralis minor. The muscle inserts between the subscapularis proximally, mastohumeralis distally, and supraspinatus cranially.
In P. dioptrica a well-developed coracobrachialis arises just behind the pectoralis minor from the central medial surface of the coracoid process. The muscle inserts onto the proximal tubercle behind the supraspinatus and mastohumeralis; in one specimen it is almost entirely surrounded by the insertion of the subscapularis in one specimen.
The coracobrachialis in C. hectori has almost the same arrangement as in the other two species. The muscle arises from the medial surface of the coracoid process and inserts onto the top central aspect of the proximal tubercle.

A n a t o m y
217
Omohyoideus (not depicted)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Absent Absent Likely absent Aponeurotic on cranial angle of scapula
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Absent Absent Likely absent Hyoid
Action Craniad movement of scapula (or possibly caudad movement of the hyoid respectively)
No corresponding muscle was found in G. griseus.
The insertion of this muscle could not be ascertained in P. dioptrica (because of damage by prior evisceration) and the muscle is regarded as most likely absent.
In C. hectori, a muscle near the cranial edge of the scapula was interpreted as severed parts of the panniculus rather than the omohyoideus; the latter is regarded as absent. Mastohumeralis (Figs. 22b, 23b, 24b)
Species described
Origin G. griseus P. dioptrica C. hectori Cetacea in general
Paroccipital process (exoccipital) Paroccipital process (exoccipital) Not known Exoccipital
Insertion G. griseus P. dioptrica C. hectori Cetacea in general
Medio-craniad aspect of tubercle Medio-craniad aspect of tubercle Medio-craniad aspect of tubercle Mid-cranial edge of humerus
Action Extends and rotates humerus inwards
The mastohumeralis arises in G. griseus as a round tendon from the paroccipital process (of the exoccipital) and inserts tendinous between supraspinatus (craniad) and coracobrachialis (proximad).

A n a t o m y
218
In P. dioptrica, the mastohumeralis arises as a round tendon from the paroccipital process and crosses the neck region as a rounded, fleshy belly. The muscle inserts as a round tendon onto the antero-distal portion of the proximal tubercle, between but distal to the insertions of the supraspinatus and coracobrachialis.
The mastohumeralis inserts in C. hectori as a round tendon onto the antero-distal
portion of the proximal tubercle, behind the insertion of the supraspinatus and below the coracobrachialis.
Extensors and flexors (Fig. 25)
No flexors or extensors were found in the lower forelimb of any of the three species. However dense fibrous tissue covered brachium and carpals and tendinous bands along the digits may be regarded as vestigial extensors and flexors in all three species.
Fig. 25. Flexors and extensors of the manus Vestigial flexors and extensors of the manus in Phocoena dioptrica (X2020.77). (a) extensors, lateral view; (b) enlargement of lateral digit 5; (c) flexors, medial view. Note fibres inserting beyond tip of digit 5 (arrow).

A n a t o m y
219
Discussion
Evolution of the forelimb: osteological and myological aspects of fossil and extant species
The transition from land to sea in cetacean evolutionary history is well documented by fossils. In more recent years, the combination of diverse evidence (anatomical,
genomic, physiological, behavioural) has increasingly clarified phylogenetic affinities among cetaceans and their relationships to other extant and fossil mammalian taxa (Gatesy et al., 2012). Morpho-functional comparison of extant taxa has hereby played a crucial role in the reconstruction of lifestyle, physiology and behaviour of extinct whales. In early archaeocetes, aspects of locomotion involving the vertebral spine and hindlimbs were of most interest (e.g. Thewissen and Fish, 1997; Buchholtz, 1998, 2001b; Thewissen et al., 2007; Gingerich, 2012). The focus of this study is integration of these aspects with emphasis on the shoulder and forelimb, and to amalgamate these findings with the diversity found in extant cetaceans. Drag-based swimming as a catalyst
Indohyus (Raoellidae, Fig. 1) is arguably the closest known relative of cetaceans (Thewissen et al., 2007; Spaulding et al., 2009) and was in size, appearance and lifestyle comparable to the extant mousedeer or chevrotains (Tragulus, Hyomoschus; Tragulidae). Both are small (about fox-sized) and stocky, have prominent canine teeth and cursorial ungulate limbs with hooves. Tragulids always remain near freshwater, feeding on fruits and flowers. Indohyus was more aquatic and either fed on water plants or came ashore to feed on terrestrial plants, a habit more like that of the much larger hippopotamus (Thewissen et al., 2007; Cooper et al., 2012). Indohyus is often depicted in a mousedeer-like, unguligrade stance but was actually, like all pentadactyl Eocene artiodactyls, digitigrade (Fig. 26; Clifford, 2010). Forelimbs had five fingers (pentadactyl) and hindlimbs four toes (tetradactyl). Fore- and hind-feet were equipped with foot pads similar to those seen in dogs (Clifford, 2010; Cooper et al., 2012). While some studies propose that only the more robust, central two digits were weight-bearing (paraxonic digits 3 and 4; Cooper et al., 2012), others argue that all four digits were weight-bearing with the main axis passing through digit 3 (mesaxonic; Clifford, 2010).

A n a t o m y
220
Similar to dogs, Indohyus also had fully developed antebrachial musculature and tendons, and was therefore not as ‘light-footed’ as mousedeer (Clifford, 2010). Despite some differences to tragulids, Indohyus had many cursorial characteristics typical for later fossil and modern artiodactyl limbs (Smith and Savage, 1956; Cooper et al., 2012):
- Long and gracile appendicular elements; hindlimbs longer than the forelimbs - Medio-laterally stabilised shoulder and elbow joints (as well as hip and knee joints),
limiting the ability for supination, pronation or medio-lateral translation - A triangular shoulder blade, the scapular spine moderately developed and without
bifurcation - A deep humeral olecranon fossa allowing great extension of the forearm - An ankle with typical artiodactyl double-trochleated astragalus However, morphological characteristics that set it apart from other ungulates (mostly in the skull) were more reminiscent of those in early cetaceans (Thewissen et al., 2007; Cooper et al., 2012): - Incipient osteosclerosis in limb bones and other postcranial elements - Metatarsals and phalanges slightly longer than typical for early artiodactyls These characteristics indicate increased aquatic adaptations. Osteosclerosis in postcranial elements may have been used as ballast to counteract buoyancy in the same manner as extant mammals do that on walk on the river bed rather than swim (e.g. Hippopotamus; Thewissen et al., 2007). Heavy bones likely facilitated invasion of freshwater habitats, maybe in a similar way as chevrotains do. Besides hiding, water chevrotain avoid imminent danger by diving underwater (Dubost, 1975), yet they lack osteosclerosis of the postcranial bones (Thewissen et al., 2009; Cooper et al., 2012). Elongated metatarsals and phalanges are a hallmark of pelvic paddlers such as otters
or desmans which use the large surface of their hindfeet for primary propulsion in water (Thewissen et al., 2001; Gingerich, 2003b). In Indohyus, incipient metapodial elongation and interphalangeal flexibility may have allowed for better weight distribution on soft substrates like mud rather than being a specific aquatic adaptation. Overall limb and vertebral characteristics suggest that Indohyus probably employed quadrupedal paddling similar to terrestrial mammals rather than pelvic paddling like otters (Cooper et al., 2012).

A n a t o m y
221
However, up to date there are no myological publications examining either modern tragulids or reconstructing Indohyus shoulder and limb musculature; predictions of locomotory pattern have thus remained highly speculative. Nonetheless, comparison of gait patterns and forelimb musculature in modern cursorial mammals could provide insight. When moving fast on land, Indohyus most likely adopted a dog-like leaping
gallop, also seen among artiodactyls in pigs and tragulids, and to some extent in smaller deer and antelope (see also Terrestrial gait patterns in this chapter). Leaping is more cost-effective in small mammals than the rapid movement of legs utilised in the horse-like gallop of larger artiodactyls. The main muscles flexing the brachium are hereby the latissimus dorsi and teres major (Smith and Savage, 1956). There are no fossil indicatications of size and extent of the latissimus dorsi, but this muscle is universally present in all tetrapods. In artiodactyls the latissimus dorsi is well-developed and usually inserts in common with the teres major (Howell, 1937). On the other hand, osteological correlates of intrinsic shoulder and limb muscles are strongly indicative of muscular size and performance.
Fig. 26. Indohyus Semischematic musculoskeletal reconstruction of Indohyus. Selected muscles discussed in text; note digitigrade stance with all four main digits supported by large digital pads. Modified from Thewissen et al., 2007 fig. 5

A n a t o m y
222
In highly cursorial modern ungulates, e.g. horse, limb bones are slender and the scapula, humerus and forearm encompass wide, open angles. Foot movements are fast (Gregory, 1912), proximal limb bones short, distal limb elements reduced or lost, and the remaining elements fused. The stance is unguligrade and distal muscles are reduced to tendons that store and release energy efficiently (Smith and Savage, 1956;
Clifford, 2010). The scapula is in contour a high triangle with the long axis nearly vertical; the scapular spine is low and divides the blade into a somewhat smaller supraspinous fossa and larger infraspinous fossa. Sharp scapular angles serve as points of insertion for the two parts of the serratus (Smith and Savage, 1956). The teres major, attached at the caudal border of the caudal angle and inserting onto the posterior border of the humerus, passes closely by the humeral head (fulcrum). In combination with a long limb, such an arrangement results in a low mechanical advantage (low leverage) but fast movement of the distal limb. The triceps brachii (capiti longus, laterale and mediale) inserts on a moderately long olecranon and arises below the teres major on the scapular caudal border, and from the mid- to upper lateral and medial region of the humerus (Sisson, 1910). A moderately long olecranon hereby provides good leverage for the well-developed triceps (Smith and Savage, 1956). During gallop a the greatest moment of resistance occurs when the foot touches the ground (Gregory, 1912). At that moment, the leg is protracted (the humerus extended) and the latissimus dorsi and teres major are both stretched and checked by the resistance of the leg on the ground. As the leg moves backwards resistance diminishes and acceleration of the limb increases. In doing so, contractile forces of the muscles are maximised, a principle that also applies for the back legs (Gregory, 1912). Another important muscle indirectly involved in the movement of the limb is the serratus. In ungulates, this muscle is split in two: the cervical part inserting into the cranial angle,
the thoracic part into the caudal angle of the scapula. Together the two parts assist the fore- and backwards movement of the limbs. At the beginning of the power stroke, the cervical part of the serratus lowers, rotates and advances the shoulder blade, while the thoracic part acts as a stabiliser. This action effectively rises the torso higher off the ground as the leg moves backwards. Up- and forward recovery of the shoulder blade is subsequently accomplished by the cervical part of the trapezius during the forward stroke of the leg (Smith and Savage, 1956). Other shoulder muscles, like the deltoideus and pectorales, serve as accessory muscles as well as stabilising the glenohumeral

A n a t o m y
223
joint (along with the supraspinatus, infraspinatus and subscapularis) to ensure optimal parasagittal movement of the leg (Smith and Savage, 1956). However, modern locomotory models of early pentadactyl artiodactyls are not found among modern ungulates but rather among canids (Clifford, 2010). In the canine leaping gallop, acceleration of the limbs is of much less importance. Instead, priority
lies in rising the center of gravity. In the leaping gallop there are two, instead of one, suspension phases where no limb contacts the ground. The increased duration of suspension is achieved by actively flexing the back rather than by expansion and contraction of passive ligaments and leg-driven propulsive acceleration. In consequence, less effort is needed to move the limbs and most of the energy is used in raising the center of gravity. The extreme adaptations of extant cursorial ungulates are therefore not seen to the same extent in canids or Indohyus, namely: digitigrade instead of unguligrade stance; moderate tapering and (weight) reduction of the distal limb elements, particularly in Indohyus with heavier than usual limb bones (osteosclerosis); less sharp angles of the scapula, and a shorter olecranon process (Fig. 26). Overall Indohyus was ostensibly an inconspicuous Eocene artiodactyl with forelimb musculature possibly quite similar to extant canids, yet it also possessed some unique aquatic adaptations that set it apart from other artiodactyls and in close relationship to archaeocetes. However, the defining characteristics separating early whales from early artiodactyls appear to be less a matter of osteology and more an inferred dietary switch documented by a combination of dental characteristics and stable isotope analyses (Thewissen et al., 2007). Pakicetids (Pakicetidae, Fig. 1) are regarded as the first and most archaic cetaceans.
They include three : Pakicetus, Nalacetus and Ichthyolestes (Thewissen et al., 2009). The oldest known whale, Himalayacetus subathuensis (53.5 my), was originally also classified as a pakicetid but is now regarded as an (Bajpai and Gingerich, 1998; Uhen, 2010). Some pakicetids grew considerably larger than Indohyus, like the mastiff-sized Pakicetus and the greyhound-sized Nalacetus, but the fox-sized Ichthyolestes was about the size of Indohyus (Madar, 2007). Wolf-like in appearance, with a long snout and tail, pakicetids show very little morphological difference from Indohyus. Both were digitigrade, aquatic waders with cursorial adaptations (Thewissen et al., 2001; Uhen, 2007b):

A n a t o m y
224
- Revolute zygapophyses of the lumbar vertebrae - A roughly similar, somewhat pear-shaped scapula with simple spine bearing midway
a small, flat and caudally directed lappet-shaped superior process (called tuber spinae in modern ungulates; Sisson, 1910; Gasse, 2012)
- Relatively long and slender medio-laterally stabilised limb bones
- Radius and ulna not fused, ulna with short olecranon - Slender, unfused metapodials - Double-pulleyed astragalus These features led initially to the conclusion that pakicetids were no more aquatic than a tapir (Thewissen et al., 2001). However, despite the discovery of more than 350 postcranial elements, no articulated specimens are known, and it was only later that differences (some small but significant) became more evident. These also included, besides the vertebral column and hip, characteristics of the forelimb (Madar, 2007; Cooper et al., 2012; Gingerich, 2012): - The glenohumeral surface of the scapula allows greater mobility, although the cubital
joint remains restricted to flexion and extension only - Shorter limbs overall show advanced osteosclerosis and trabecular hyperostosis - Complete loss of ability to supinate or pronate forearm - Metapodials are more elongated and metacarpal flexion is not as limited as in
raoellids, i.e. the degree of flexion is equal to that of extension (up to 180º in total) The most conspicuous difference, though, is the posture of the head. While Indohyus habitually held the head erect above the level of the thorax, position and articular facets of the occipital condyles and atlas indicate a much more level posture in pakicetids (Cooper et al., 2012). Pakicetid forelimb anatomy overall resembles that seen in Indohyus, yet also shows
small but significant differences. The dorsal border of the scapula is rounded (Madar, 2007), more reminiscent of that seen in dogs. However, the scapula of canines is overall long and narrow, whereas in pakicetids it is more pear-shaped. Moreover, in ungulates the cartilaginous dorsal border, which is not preserved in fossils, also forms a rounded dorsal border similar to that of pakicetids (Sisson, 1910). In both raoellids and pakicetids, the scapular spine is well developed with a broad, caudally directed spinal tuber and a supraspinous fossa that is larger than the infraspinous fossa (Madar, 2007), an arrangement typical for cursorial mammals (Smith and Savage, 1956). Both taxa also show a prominent supraglenoid tubercle, i.e. attachment point for the biceps

A n a t o m y
225
brachii (a flexor of the forearm); and a prominent infraglenoid tubercle, indicating a well-developed scapular head of the triceps (flexion of the humerus and retraction of the limb). The coracoid process is hook-shaped, the bulbous tip offering good anchorage for the coracobrachialis (for glenohumeral stabilisation). The acromion process is short and rod-like or spatulate, and projects past the lateral border of the
glenoid fossa (Madar, 2007). In Indohyus, the tip is directed craniad while in pakicetids the tip is oriented ventrad (Cooper et al., 2012), an indication that in Indohyus the acromial part of the deltoideus might have played a slightly more important role in forelimb protraction than in pakicetids (Smith and Savage, 1956). In both Indohyus and pakicetids, the humeral head is large and subspherical, but the greater tubercle in pakicetids does not surpass the head as it does in Indohyus (Cooper et al., 2012). Together with a larger glenoid surface, the lower elevation of the tubercles could be interpreted as reflecting somewhat greater shoulder joint mobility in pakicetids (Larson and Stern Jr., 1989; Cooper et al., 2012). While the profile of the humerus in Indohyus does not deviate from the typical cursorial artiodactyl pattern (proximo-distal tapering), the humeral shaft is nonetheless osteosclerotic, a trait also seen in pakicetids. Conversely, the humeral diaphysis in pakicetids appears overall proximally narrower proximally than distally. The epicondyles are larger in pakicetids, yet there is comparatively little surface for muscles rotating the forearm. As in Indohyus, a posterior tuberosity indicates the origin of forearm flexors (Madar, 2007; Cooper et al., 2012). In pakicetids, the supratrochlear foramen is absent, whereas in Indohyus and Tragulus it appears as a perforation of the triangular olecranon fossa (Cooper et al., 2012). An explanation for the presence of the foramen in various taxa has hitherto remained elusive. It is exclusively present in mammals. In humans, it is more prevalent in ancient peoples, varies greatly between races and generally occurs more frequently in females
and on the left-hand side (Singhal and Rao, 2007). It is also very common in primates other than humans (Erdogmus et al., 2014). In ungulates, the presence of the foramen has been explained by way of impingement of the olecranon onto the olecranon fossa, either caused by impact when running stiff-legged (pronghorn antelope), or pressure when feeding whilst pinning food to the ground and using the straightened limb as a brace to tear off pieces (pigs and peccaries; Lamb, 1890). If correct, the absence of the supratrochlear foramen in pakicetids may also indicate a change of locomotion and/or food-processing.

A n a t o m y
226
The radius and ulna are not fused but reveal a flat radio-ulnar articular surface, and the radius is not bowed (albeit moderately flared); i.e. the bones of the lower arm exhibit characteristics incompatible with pronation or supination. The ulna is shorter than the humerus (Madar, 2007), atypical for cursorial artiodactyls (Polly, 2007) and archaeocetes (Gingerich et al., 2001a; Gingerich et al., 2009; Gingerich and Zalmout,
2019; Lambert et al., 2019). Longitudinal depressions on the radial surface indicate the presence of extensors and flexors of the manus, although not quite as markedly on the extensor surface of pakicetids. The olecranon process of the ulna, insertion of the triceps (for extension of the forearm), is more robust in pakicetids. The medullary cavity of the shaft is small or absent and filled with hyperostotic trabecular bone, the cortical bone, and is heavily osteosclerotic (Madar, 2007; Cooper et al., 2012). In the manus of both Indohyus and pakicetids, metacarpals III and IV are robust and weight-bearing, as are the central digits in pakicetids. Unlike modern canids, articulation of the proximal phalanges shows that the central digits also appeared to have a wider range of movement than the lateral digits. Short phalangeal crests may indicate the presence of webbing between the digits (Madar, 2007; Cooper et al., 2012). Contrary to earlier findings, pakicetids were considerably more aquatic than raoellids (Gingerich, 2003b). Despite a multitude of cursorial adaptations, advanced cortical hypertrophy and increased trabecular density throughout the skeleton indicate poor cursorial capabilities but effective ballast - ideal adaptations for walking on the floor of stream beds and lagoons (Gingerich, 2012). A robust supraglenoid tubercle and olecranon indicate strong flexion and extension of the forearm, useful for limb movement through water. However, there appears to be no major modification to the
acromion (acromial head of deltoideus) or extension of the scapular caudal angle (teres major) to improve limb protraction and retraction. Strong forearm flexion and extension was likely used to assist hindlimb and tail musculature; together they imply quadrupedal paddling and/or undulatory locomotion assisted by the tail and forelimbs for propulsion and steering when swimming (Madar, 2007). Larger than pakicetids, and with long and powerful jaws, eyes positioned high on the skull, a strong neck and sturdy, short limbs with large hands and even larger feet, as well as a long and muscular tail, ambulocetids (Fig. 1) looked like a cross between a

A n a t o m y
227
crocodile and a sea lion (Thewissen and Hussain, 1996; Thewissen et al., 2009). A heavy rib cage and limbs served for neutral buoyancy but negated any remnant cursorial capabilities that might have been still present in pakicetids. Ambulocetus lived in marine environments and was most likely an ambush predator, possibly quite similar to crocodiles (Thewissen and Hussain, 1996). Himalayacetus on the other hand
predates other known ambulocetids by over 4 my and had still had some ties to freshwater habitats (Bajpai and Thewissen, 1998; Bajpai et al., 2009). Terrestrial gait patterns
Although these early cetaceans showed some aquatic adaptations, they essentially retained terrestrial locomotion and thus also terrestrial gait patterns that differ greatly from the mode of propulsion seen in modern cetaceans. To understand the transition from terrestrial to aquatic locomotion, it is important to understand the fundamental terrestrial gait patterns that underly the different modes of aquatic propulsion. Terrestrial gaits are divided into three basic categories, referred to here as walk, trot,
and gallop. Walk is slow but very stable, gallop is fast but also much more unstable and energy-consuming. Trot is an intermediate gait between walk and gallop. Despite a great variety and sometimes complicated sequence patterns, walk and trot include astonishingly simple basic locomotor parameters. Stride length, duration of ground contact and suspension above ground are basically equal for in each foot. Fore- and hind-feet act hereby as a coupled pair where one foot always strikes the ground at half stride of the other (50% of the stride length) and thus mirrors movement of the contra-lateral limb. Walk and trot have therefore been termed ‘symmetrical gaits’ and can be defined by only two parameters: (1) duration of ground contact, (2) phase shift of the ipsilateral forelimb compared to the hind limb (lag). Sequence patterns usually commence with the left hind limb; measurement of the ground contact duration is also taken from hind limbs (forefeet sometimes have slightly shorter ground contact; Hildebrand, 1989). Walking gaits are characterised by various combinations of tetra-, tri- and bipedal phases and long ground contact duration (> 50%). Because these patterns provide great stability, they can assume a wide variability and continuity across quadruped vertebrates (Hildebrand, 1989).

A n a t o m y
228
The trot is much faster and is distinguished by short (if any) tripedal, comparatively longer bi- and unipedal phases with extended suspension of the foot, and with increased speed often also a floating phase (suspension of the body with no foot contact to the ground). As a result, only four major more or less well-delineated
patterns emerge: (1) ‘pace’ (25% ground contact and 0% lag), (2) ‘trot (50% lag), (3) ‘lateral-sequence gait’ (up to 25% lag), and (4) ‘diagonal-sequence gait’ (over 50% but under 80% lag). These gaits are all continuous along decreasing ground contact duration but separated by discontinuous phase shift. Phase shift of the forelimb is expressed as a fraction of stride duration, i.e. the time the forelimb strikes after the initial ground contact of the hindlimb (time lag). Lateral-sequence gaits are hereby the most flexible gaits, both in ground contact duration and phase shift, as well as speed (Hildebrand, 1965). Variability in the Islandic horse for example, is not just within the breed but also seen in individuals using a range of different sequences at different speeds (Nicodemus and Clayton, 2003). In that manner, even the pony-sized Islandic horse can attain up to 50 km/h when ‘tölting’, a speed that is usually associated with gallop (Kimball, 2006). True diagonal-sequence running gaits are primarily known in small deer and antelope, thus may have also been employed by Indohyus (Hildebrand, 1965, 1989). The gallop is a typical running gait where the unifying parameters of the walk and trot are broken and much more variable, hence the gait is referred to as ‘asymmetrical’. Here, contralateral fore- and hind limbs work as functional couplets that are in timing and position more closely linked to each other than to the ipsilateral limb. There are three basic locomotory patterns: (1) horse-gallop, (2) leaping-gallop, (3) bound (Smith
and Savage, 1956; Hildebrand, 1989). Horse-gallop, referred to by Hildebrand (1989) as ‘transverse gallop’, is generally used by large cursors (like horse, large antelopes, bison, rhino) adapted to covering distance economically rather than at top speed. Typical for this gait, the floating phase occurs while all limbs are gathered under the torso. Leaping-gallop, termed by Hildebrand ‘rotary gallop’, can be seen in sprinters like small antelopes as well as cursorial carnivores and is optimised for manoeuvrability and speed. Leaping-gallop was likely used by Indohyus when speed was of the essence (see section Indohyus above). At top speed, cursorial carnivores experience

A n a t o m y
229
two floating phases, one with limbs gathered and one with limbs extended (e.g. fast-running cheetah). A highly compressed form of the leaping gait is the ‘pronk’ of deer where all four feet strike the ground almost simultaneously. The bound is characterised by a series of leaps with an extended floating phase. It is used by small but strong-legged animals (e.g. mice). Typically, the limbs of a couplet
strike the ground almost simultaneously; if there is a leading forefoot, the pattern is called half-bound (e.g. mink or weasel; Williams, 1983; Hildebrand, 1989). Contrary to commonly held views, forelimbs do not just merely support the trunk but provide locomotory power equal to that of the hindlimbs (Gregory, 1912). Powered propulsion of the forefoot is most crucial in gaits where maintenance of the forward momentum by the forelimb is essential, i.e. the strike of a single forelimb is followed by a floating phase as it can be in the tölt. Stride flexibility and the ability to provide powerful strokes with both fore- and hindlimbs are crucial prerequisites for aquatic locomotion, especially during the initial transition from land to water. Aquatic locomotion
Based on Hildebrand’s analysis, Fish (1996) investigated changes in morphology and swimming mode of modern terrestrial, semiaquatic and aquatic mammals. He studied biomechanical and energetic performance in these animals and linked primitive neuromotor patterns associated with terrestrial gaits to aquatic locomotor patterns. Fish concluded that modern semiaquatic analogues best represent original transitional stages of extant fully aquatic mammals. He proposed progressive stages of aquatic adaptation from drag-based paddling, with origin in symmetric terrestrial gaits, to lift-based oscillation, with origin in asymmetric terrestrial gaits. He inferred further that steady increase of swimming speed, power and efficiency were commensurate selective factors in the transition from land to sea (Fish, 2000). Quadruped paddling is a drag-based swimming mode used by terrestrial and semiaquatic quadrupedal mammals and employs symmetrical terrestrial gait patterns in vertical motions. Terrestrial mammals are typified by a stable lumbus and a sacrum of usually four fused vertebrae, with a sacroiliac joint providing support on land and little or no aquatic adaptation of the limbs (Buchholtz, 1998). The ‘dog paddle’ is an archetypical modified lateral sequence run, whereas opossums rather utilise a

A n a t o m y
230
diagonal sequence run (Fish, 1993a). This locomotory mode allows essentially terrestrial quadrupeds, which lack additional buoyancy of non-wettable fur, to hold their nostrils above the water by generating lift with the forelimbs (Fish, 1996). Bipedal paddling is a modified form of quadrupedal paddling employed by semiaquatic mammals. Increased swimming efficiency is achieved by the elimination of ipsilateral
limb interference and additional buoyancy of non-wettable fur. It also frees one pair of limbs for other purposes (stabilisation, manoeuvring, tactile reception, food capture and processing). Polar bears (Ursus maritimus) and water opossums (Chironectes minimus) are a classic examples of alternate pectoral and pelvic paddling respectively (Fish, 1993a; Domning, 2002). In bipedal rowing the motion of the limbs is horizontally directed. This allows simultaneously thrust and generation of downward force to counteract buoyancy. Typical representatives of this swimming mode are platypus (Ornithorhynchus anatinus, alternate pectoral rowing) and muskrat (Ondatra zibethicus, alternate pelvic rowing when submerged; Fish, 1996, 2000). Although predominantly drag-based, simultaneous pelvic paddling is based on an asymmetrical terrestrial gait pattern. Similar to the bound, the hindlimb pair is moved as a transverse couplet in conjunction with spinal and caudal undulation. Contrary to fully terrestrial mammals, lumbar vertebrae of the North American river otter (Lontra canadensis) are quite long and flexible. Proportionally, caudal vertebrae are even longer and most flexible at about mid-tail (Buchholtz, 1998). While undulation of the tail generates lift, the tail of river otters is tapered, and thus contributes little to the overall momentum (Fish, 1994; Fish, 2016; Thewissen and Bajpai, 2001). In pelvic undulation, drag-based paddling motions of the limbs are of much lesser importance or insignificant. Instead, hindfeet have transformed into hydrofoils powered by a travelling wave through the spine (Fish, 1996; Thewissen and Bajpai, 2001). Sea otters
(Enhydra lutris) have a short tail and large, highly asymmetric feet that generate most of the (lift-based) thrust (Thewissen and Fish, 1997). Accordingly, lumbar vertebrae are longer and more flexible than caudal vertebrae. Sea otters also show incipient elongation of posterior thoracic vertebrae gradually approaching lumbar dimensions, a tendency termed ‘lumbarisation’ by Buchholtz (1998). A broad, dorsoventrally flattened tail is the hallmark of caudal undulation. Here, sinusoidal waves of the spine power the tail, which provides the main thrust. Giant South American freshwater otters (Pteronura brasiliensis) or beavers (Castor canadensis) use this form of aquatic locomotion almost exclusively (Fish, 2000; Thewissen and Bajpai, 2001). The only

A n a t o m y
231
extant mammals using a form of lateral undulation are the African otter shrew (Potamogale velox), muskrat (Ondatra zibethicus) and desman (Desmana moschata), but here undulation is confined solely to the tail (Fish, 1996, 2000; Ivlev et al., 2010; Fish, 2016). However, despite ostensibly clearly-defined categories, there is often no sharp distinction between aquatic locomotory modes. Furthermore, semiaquatic
mammals often use more than one type of swimming. The North American river otter may use quadrupedal and alternate pectoral paddling when moving about on the water surface, but utilises alternate pelvic paddling for rectilinear swimming, and switches to simultaneous pelvic paddling and caudal undulation when submerged (Fish, 1994; Thewissen and Fish, 1997). Muskrats and desmans use a combination of alternate pelvic paddling and auxiliary lateral undulation of the tail when swimming on the water surface, but resort to alternate pelvic rowing when submerged (Fish, 1996, 2000; Ivlev et al., 2010; Fish, 2016). Morphological specialisations of the limbs in semiaquatic mammals using paddling or undulatory propulsion include short and robust humerus (and femur), elongation of the digits, increased propulsive surface by way of interdigital webbing or hair fringes, and often increased bone density (Fish, 1996, 2000). Forelimbs are hereby not used in most species for propulsion but rather for steering or other tasks unrelated to aquatic locomotion, such as prey capture, transport and processing or hauling out on ice (Levermann et al., 2003; Cooper, 2009; Hocking et al., 2017). In caudal oscillation spinal movements become centred around one point, producing a standing wave. The propulsive surface is usually a highly asymmetric, often rigid hydrofoil where the angle of attack is controlled by a joint (Fish, 1996; Thewissen and Fish, 1997; Thewissen and Bajpai, 2001). Although lift-based oscillation is typically seen in pelagic marine mammals where steady swimming is essential (Fish, 2000),
incipient forms can be seen in neritic living sirenians (Buchholtz, 1998; Thewissen and Bajpai, 2001). Here the posterior thoracic, lumbar and anterior caudal vertebral centrum length forms a single broad peak indicating a rather vague centre of flexion. Manatees (Trichechus manatus) have a broad, rounded flukes but almost no vertebral correlates indicating its presence. In the somewhat more efficiently swimming dugong (Dugong dugong), with typically V-shaped flukes, a slightly lower width / height ratio than more anterior caudal vertebrae suggests a peduncle, and a marked widening of the posterior-most caudal centra correlates with the presence of flukes. Slow-swimming cetaceans like belugas (Delphinapterus leucas) also show a single, albeit

A n a t o m y
232
narrow peak in length of centra at the lumbar-caudal transition, markedly laterally-compressed caudal centra in the area of the peduncle and a height / width reversal at the point where the flukes join the peduncle (ball vertebra). Whereas the vertebral columns of sirenians and belugas reflect unimodal flexibility, in fast-swimming cetaceans, like the short-beaked common dolphin (Delphinus delphis), it is bimodal.
Here ‘lumbarisation’ resulted in the shortening of lumbar, ‘sacral lumbar’ (sacral vertebrae that are no longer distinguishable from lumbar vertebrae) and anterior caudal vertebrae, in concert with an increased lumbar vertebral count (greater than thoracic). Vertebral length attains a maximum in the area of the thorax and as a second, sharp peak in the area of the peduncle where the centra of the vertebrae are also strongly laterally compressed. Transition to the flukes is clearly marked by height / width reversal and shorter, but much wider posterior-most caudal vertebrae (Buchholtz, 1998). From drag- to lift-based swimming
Over the years, understanding of evolutionary processes moulding early cetacean aquatic adaptions has gained increased insight, yet has also remained controversial. Fish (1996) stipulated that the change from paddling to undulatory swimming is the key to the evolutionary transition from terrestrial to fully aquatic habit. Furthermore, simultaneous paddling of the hind feet, with its root in asymmetrical terrestrial bounding gaits, is integral to undulation. Fish (1994) hypothesised that the incipient undulatory swimming mode of river otters (Lutra canadensis), coupled with simultaneous pelvic paddling, demonstrated a possible route in the evolutionary pathway of early cetaceans from drag-based paddling to lift-based undulation and ultimately oscillatory propulsion of modern cetaceans. Thewissen and Fish (1997) substantiated these earlier findings after close examination of skeletal remains of Ambulocetus and comparing them to modern semiaquatic analogues. Central to their argument are the large, nearly symmetrical feet in form of a high triangle, indicating drag-based rather than lift-based propulsion. A powerful tail possibly also provided some propulsion, but the main thrust was imparted by the hind feet. They concluded that such an arrangement is most similar to modern river otters (Lontra canadensis). Buchholtz (1998) confirmed these findings when examining vertebral characteristics in extinct and extant mammals. She found that Ambulocetus

A n a t o m y
233
did not possess an undulatory torso but rather lumbar flexion with foot propulsion, and that the tail likely played a secondary role in swimming, i.e. vertebral traits that matched most closely those of river otters. Lutrine aquatic locomotion was also the locomotion mode proposed for later archaeocetes such as remingtonocetids and protocetids (see below).
While large, webbed hands would be consistent with river otter swimming, e.g. quadrupedal paddling on the surface and as a steering aid when submerged, locomotion and posture of the forelimb on land has so far remained much more ambiguous in Ambulocetus. The scapula and humerus are largely unknown, and antebrachial and manual characteristics appear somewhat conflicting. Elbow and wrist arrangement show a more or less typical artiodactyl configuration: supination of the forearm was not possible, and a transversely broad distal end of the radius allowed the hand to be placed transversely to the longitudinal axis of the body (Thewissen and Hussain, 1996). A robust and long olecranon allowed for powerful extension of the forearm, but the arm itself remained relatively rigid. Although the hand was not as large as the foot and had mobile digits, parasagittal orientation of the short limbs and plantigrade stance would have precluded efficient or fast locomotion on land. Thewissen et al. (1996) therefore suggested therefore possible lateral rotation of the limbs similar to that of eared seals (Otariidae). However, eared seals have a profoundly different forelimb arrangement, using parasagittal elbow flexion in combination with a laterally deflected manus, besides intrinsic movements of the lower limb, to propel them forward (for a more detailed description see Protocetids below; English, 1976), a movement Ambulocetus would not have been able to perform. Remingtonocetidae (Fig. 1) are regarded as the subsequent stage in cetacean
evolution (Thewissen and Bajpai, 2009; Gatesy et al., 2012). They are predominantly known from cranial and axial remains. Their most salient feature is their long and narrow skull, particularly the rostrum. They were the first whales entirely disassociated from freshwater, living in turbid, near-coastal salty and brackish waters as ambush predators lurking on the ocean floor (Bajpai et al., 2011; Bebej et al., 2015). Accordingly, the long snout with its nasal openings at the tip was interpreted as a way to retain water vapour to limit dehydration, breathing while submerged or the site of mechanoreceptors (Bajpai et al., 2011). One of the best-preserved specimens, Remingtonocetus domandaensis, also includes still-articulated vertebrae of the

A n a t o m y
234
precaudal column. These show that R. domandaensis had the longest neck vertebrae of any (extinct or extant) whale, and uniquely imbricating transverse processes indicative of neck musculature and ligaments comparable to eared seals. Like eared seals, on land Remingtonocetus carried the head above the level of the body, but held it low and extended when submerged for improved streamlining of the body contour
(Bebej et al., 2007; Bebej et al., 2012). Although remingtonocetids had 3 to 4 fused sacrals and were able to support their body weight on land, their terrestrial capabilities were nonetheless limited (Bajpai et al., 2011; Bebej et al., 2012; Bebej et al., 2015). Vertebral remains of Remingtonocetus indicate only moderate lumbar mobility (Bebej et al., 2012), whereas in Rayanistes they are consistent with a more mobile lumbus and other characteristics indicative of improved stroke length of the hindlimbs and powerful pelvic paddling (Bebej et al., 2015). It is not clear if the tail of Remingtonocetus contributed to the propulsion or was rather used as an inertial stabiliser (Bebej et al., 2012). In contrast, the spinal column of Kutchicetus, the smallest remingtonocetid, presents a bimodal vertebral length maximum similar to that of river otters. The first maximum is located in the lumbar area and a second, even higher peak, in about the mid-caudal position. In combination with relatively wide vertebrae, these characteristics indicate a stocky body build and caudal undulation as a propulsory system, possibly using a dorsoventrally flattened tail comparable to that of modern Giant South American river otters (Pteronura brasiliensis). This was also the mode of aquatic locomotion proposed for Andrewsiphius (Bajpai and Thewissen, 2000; Thewissen et al., 2009). Even though vertebral characters portray remingtonocetids as utilising powerful tail undulation and/or hind limb propulsion, many critical features of the limbs, and particularly of the forelimbs and their locomotory role, are unknown (Thewissen et al., 2001).
Proximal limb elements are short and hyperostotic (Bajpai et al., 2011). Distal limb elements as well as the scapula, however, are largely unknown. The humerus of Kutchicetus and Andrewsiphius is curiously medially bowed, the head strongly convex, medio-laterally compressed and somewhat posteriorly oriented (Thewissen et al., 2009). This suggests a somewhat habitually-retracted position of the humerus and a locomotion pattern mostly in the parasagittal plane (Howell, 1930a), possibly akin to terrestrial artiodactyls and the earliest archaeocetes (pakicetids, ambulocetids. However, the bowed contour of the brachium (Thewissen and Bajpai, 2009) would have placed the elbow close and medio-ventrad to the chest, suggestive of a narrow

A n a t o m y
235
stance on land, but in water it might have helped to ‘streamlinie’ the forelimbs against the body. A deep olecranon fossa (Thewissen and Bajpai, 2009 fig. 10) indicates a great range of elbow extension, while the lack of a radial fossa precluded strong elbow flexion. A large medial epicondyle suggests well-developed manual flexors, whereas the lateral epicondyle is relatively indistinct (Thewissen and Bajpai, 2009). Taken
altogether, these characteristics tentatively suggest that remingtonocetids, or at least Kutchicetus and Andrewsiphius, may have swum chiefly with forelimbs retracted and close to the body, only occasionally using the forelimbs for additional thrust in the manner of Giant South American river otters (Pteronura brasiliensis) do. However, only further discovery of forelimb elements could support or refute a particular mode of locomotion with more certainty, and it remains to be seen if characteristics of the limbs reflect known axial characteristics that are ostensibly commensurate with Pteronura.
Protocetidae (Fig. 1) are regarded as a polyphyletic group, originally established by Stromer (1908) and later subdivided into two subfamilies (Indocetinae and Protocetinae) by Gingerich et al. (1993). However, revision of ‘Indocetus’ reassigned particularly postcranial remains to Remingtonocetus (Gingerich et al., 1995; Gingerich et al., 2001a). Subsequently Gingerich et al. (2005) established three subfamilies (Protocetinae, Makaracetinae, Georgiacetinae) based on cranial characteristics and functionality of the sacro-iliac joint, Makaracetus being in its own subfamily mainly because of the specimen’s highly unusual facial features suggestive of a short, muscular proboscis. For the sake of simplicity protocetids will here be divided into only two clades: (1) early semiaquatic protocetids (Protocetinae + Makaracetus sensu Gingerich et al., 2005) retaining a sacro-iliac hip joint and more or less capable of terrestrial locomotion, and (2) fully-aquatic later protocetids (Georgiacetinae sensu Gingerich et al., 2005) that have lost the connection between sacral vertebrae and innominate bones, and were not able to support their body weight on land. Protocetids as a whole were the first whales to disperse beyond the boundaries of the
continuously closing Tethys Seaway and attain a nearly circum-equatorial distribution. Initially thought to have migrated north along the coast of Europe, and westwards via Greenland before heading south along the East Coast of North America (Geisler and Uhen, 2005; Uhen, 2014b), a new fossil from the Pacific Coast of South America (Peregocetus from Peru) raised the possibility that protocetids may have instead migrated first to West Africa and from there directly crossed to South America before

A n a t o m y
236
heading north to North America (Geisler, 2019; Lambert et al., 2019). All accounts, however, agree that protocetid dispersal involved multiple events, including the return to their place of origin, Indo-Pakistan. To do so, protocetids had to be accomplished swimmers and this raised the question of whether protocetids had flukes associated with flippers in the manner of modern cetaceans or had webbed feet more similar to
otters (see also Functional aspect of flipper shape, section Fully aquatic archaeocetes in this chapter). One of the best-preserved protocetids, and the only one including a more or less complete series of caudal vertebrae, is Maiacetus inuus, represented by what have been interpreted as a female skeleton with fetus and a male skeleton (Gingerich et al., 2009). Posterior caudal vertebrae of Maiacetus show a height / width reversal (Ca13 and Ca14) that has been correlated to the presence of a fluke (Bajpai and Thewissen, 2014). However, the reversal is not particularly strong and Maiacetus was a heavy-set animal, in size and proportions comparable to Rodhocetus, with a fully fused sacrum, strong hindlimbs, and elongated metapodials rather suggestive of webbed feet utilising foot-powered propulsion (Buchholtz, 1998; Gingerich et al., 2009). Nonetheless, caudal vertebrae with bifurcated processes in Peregocetus pacificus, one of the earliest whales in the New World and with proportions similar to Maiacetus, suggest the presence of a paddle-like fluke (Lambert et al., 2019). Functional anatomy of the forelimb, more reminiscent of Pakicetus or cursorial artiodactyl in general than basilosaurids or later archaeocetes, remains nonetheless elusive. Some of the most salient features of the shoulder and forelimb are the following (Gingerich et al., 2009; a 3D skeletal reconstruction of Maiacetus can also be accessed at UMORF, University of Michigan Online Repository of Fossils at <umorf.ummp.lsa.umich.edu/wp/>):
- The scapula forms a high, elongated triangle, albeit with convex dorsal border (similar to Pakicetus)
- The scapular blade is unequally divided (supraspinous fossa smaller than infraspinous fossa) by a spine merging into a broad, elevated and caudally directed superior spinal process
- Narrow neck and shallow glenoid fossa - The humerus is long, the humeral head posteriorly oriented with a large, laterall-
broad greater tubercle positioned cranio-laterally, and a robust lesser tubercle cranio-medially; neither tubercle appears to surpass the humeral head in height

A n a t o m y
237
- The humerus is transversely broad distally to accommodate a large medial epicondyle, a shallow trochlea for ulnar articulation, and deep olecranon and radial fossae
- Radius and ulna are shorter than the humerus (similar to Pakicetus; brachium shorter than antebrachium in modern artiodactyls)
- The manus is shorter than the pes (similar to Pakicetus) Despite these general similarities there are also some small, but significant differences: - Cranial and caudal angles of the scapula are distinctly flared and acute, particularly
the caudal angle; the suprascapular notch is more accentuated - A long, cranio-ventrally oriented acromion process is paralleled medially by a short,
knobby coracoid while in Pakicetus the acromion is ventro-laterally oriented and divergent from the coracoid process
- Similar to Pakicetus, neither epicondyles (medial convex, lateral concave) nor the humeral deltoid tuberosity appear to be well-developed. However, the humeral shaft is flattened and medially bowed. A laterally oriented raised wedge, occupying about 2/3rd of the proximal shaft and delineated by the deltoid crest laterally and anterior border cranially, would have offered a large attachment site for the deltoideus.
- The ulnar shaft is flattened; a well-developed, spatulate dorso-caudally-oriented olecranon process terminates in a strong tuberosity, the attachment site for the triceps brachii (olecranon proximally oriented in Pakicetus)
- Carpal elements are interlocked and as a whole transversely arched (concave ventrally) including a long, strongly ventrally projecting pisiform
- Digits 3 and 4 are the most robust; terminal phalanges of digits 2 (and probably 3 and 4) are blunt, suggestive of hoofs, while digits 1 and 5 are pointed and hoof-less
With radius and ulna fixed in pronated position, Maiacetus was not able to supinate.
However, deep olecranon and radial fossae permitted strong extension and flexion of the forearm and a long pisiform provided a great mechanical advantage for the flexor carpi ulnaris.
Shoulder morphology in protocetids
Osteological correlates reveal that Maiacetus had a well-developed forelimb musculature (Fig. 27). Although not massive, the supraspinatus was well-developed, and inserted onto a relatively low greater tubercle. In some marine mammals (sea lion,

A n a t o m y
238
sea cows), this tubercle is enormous and projects well beyond the humeral head. Increased height of this process purportedly also results in improved leverage of the supraspinatus and greater force of extension, commensurate with the proportionally larger area this muscle occupies on the scapular surface (Howell, 1929, 1930a; English, 1977). However, in some species where both strength and mobility of the
shoulder joint appear to be equally important factors, mobility comes at the cost of the mechanical advantage a higher tubercle would offer. Consequently, other muscles are more heavily recruited, although a comparatively larger supraspinatus may still somewhat offset the reduced leverage of a lower tubercle (Larson and Stern Jr., 1989; O'Leary and Rose, 1995). Other palpable protractors would be the mastohumeralis and acromio-deltoid. The former is a large sheet in sea lions or a broad, incompletely divisible band in modern ungulates (Sisson, 1910; Howell, 1930a; Taylor, 1978) and most likely was also present in protocetids. The acromio-deltoid is a subdivision of the deltoideus, which in most mammals comprises either three separate divisions, clavo- (or cleido-) deltoideus, acromio-deltoideus, and spino-deltoideus, or various combinations of complete or partial fusion thereof. The acromial part, however, is absent in most modern artiodactyls and the clavicular is part fused with the cleidomastoid, now called mastohumeralis, which is also present in pinnipeds and sirenians (cephalohumeralis in Howell, 1937). In modern cetaceans, sirenians and pinnipeds the deltoideus has become hypertrophied and appears as one single, massive muscle (Murie, 1872; Howell, 1930a; English, 1977; Domning, 1978). However, in eared seals (Otariidae) this muscle is confined to the posterior half of the shoulder blade (English, 1977), whereas
in cetaceans it occupies predominantly the anterior half. These differences reflect divergent functionality of muscle and limb morphology between those two taxa. In eared seals the glenohumeral joint is habitually flexed and cannot be extended beyond 90º; the acromion is small and little more than the continuation of the spine. Consequently, the deltoideus is situated above and slightly caudad to the massive humeral deltoid crest, its point of insertion, and thus acts mainly as abductor and outward rotator of the forelimb. Additionally, superficial fibers of the acromiotrapezius bypass the spine and insert further caudad onto the aponeurosis of the deltoideus instead, thus forming a synchronized composite muscle able to maintain tension over

A n a t o m y
239
a wider range of movement. As a result, protraction of the forelimb is not facilitated by the deltoideus but rather by the mastohumeralis, aided by the supraspinatus and rotators of the scapula (levator scapulae, rhomboideus capitis and serratus anterior-posterior; Howell, 1937; English, 1977).
Functional implications of scapular morphology
Scapular characteristics of modern eared seals and cetaceans differ greatly, the former with a marked scapular spine and underdeveloped acromion, the latter with indistinct or no spine and generally large, broad acromion. While the presence of the trapezius correlates with a well-defined scapular spine, morphological and functional aspects of the deltoid are greatly influenced by the acromion (Howell, 1930a). Yet, Howell saw ‘no apparent reason for its existence’ when referring to the acromion (or coracoid) in cetaceans. Significantly, the comparatively long, cranio-ventrad projection of the acromion in Maiacetus also stands in stark contrast to that seen in earlier archaeocetes (e.g. Pakicetus), thus possibly indicating a functional change of this bone within among
archaeocetes. There are no modern ungulate analogues congruent with the shoulder morphology of Maiacetus. However, similar osteological characteristics of the shoulder and upper limb, including a well-developed acromion process, can be found in arboreal and fossorial taxa such as †Arctocyon primaevus, binturong (Arctictis binturong), tamandua (Tamandua sp.), and armadillo (Dasypus sp.), and to some degree in the aquatic manatee (Trichechus sp.; Howell, 1930a; Smith and Savage, 1956; Taylor, 1978; Argot, 2013). While shoulder and limb morphology in modern cetaceans and manatee appear unique and widely separated from their closest terrestrial relatives (Howell, 1930a), arboreal and/or fossorial taxa show gradual divergence with increased specialisation across a multitude of species, allowing a more detailed morphofunctional analysis between modern taxa and functional interpretation in fossils (Argot, 2013). Arboreal and fossorial taxa need powerful abductors, adductors, and flexors but strictly fossorial species show generally none of the adaptations of the hindlimb or tail seen in arboreal species (Argot, 2013). For comparative purposes regarding the forelimb in this study, the distinction between arboreal and fossorial taxa is therefore not crucial.

A n a t o m y
240
Consequently, alterations of the scapula and upper arm in either ecotype may serve as a model for analogous adaptations seen in the evolutionary history of cetaceans. Arctocyon is a basal ungulate from the Palaeocene of Europe and North America with a forelimb showing some moderate adaptations towards an arboreal-scansorial habit.
The scapular spine rises high above the scapular blade to accommodate well-developed supraspinatus and infraspinatus muscles, and terminates in a diamond-shaped, slightly cranio-ventrally-projecting acromion that does not extend beyond the glenoid cavity (Argot, 2013). A similar arrangement can also be seen in binturong or †Plesiotypotherium achirense and †Mesotherium cristatum, two Late Miocene mesotheriid notoungulates from Bolivia (Fernández-Monescillo et al., 2018). Here, the acromion is slightly more accentuated and, apart from a small metacromion, also somewhat better comparable to Maiacetus. However, overall scapular and humeral characteristics are best matched by the extant binturong. Contrary to binturong, the spine in Maiacetus appears to be foreshortened and does not reach the vertebral border. Consequently, the superior spinal process sits remarkably close to the shoulder joint and most likely was functionally analogous to the metacromion in binturong. Furthermore, although much more gracile and longer than in Maiacetus, the humerus of the binturong presents neither a prominent deltoid ridge or tuberosity, nor elevated tubercles (Argot, 2013). A similar arrangement is possibly also present in a partially preserved forelimb of Peregocetus, an early protocetid from the early Late Lutetian of Peru (42.6 my; Lambert et al., 2019). In binturong or Plesiotypotherium, protraction of the forelimb is mostly facilitated by the
mastohumeralis and acromio-deltoid, and aided by the supraspinatus. Retraction of the forelimb is accomplished by the teres major, latissimus dorsi and spino-deltoideus (Fernández-Monescillo et al., 2018). In more specialised species like the tamandua (Tamandua sp.), elongation and anterior medial curvature of the acromion place the acromio-deltoideus cranial and dorsal of the humerus. This arrangement provides enhanced leverage for the acromio-deltoid, while the spino-deltoid assists in outward humeral rotation (Taylor, 1978). In some armadillo species (Dasypus sp.) a ventrally-directed acromion extends laterad past the shoulder joint, and in that manner facilitates a swooping action of the forelimb

A n a t o m y
241
away from the body, scooping up soil while avoiding already piled up earth further behind (Smith and Savage, 1956). The cranio-ventral position of the acromion in Maiacetus provides excellent leverage but at the same time also limits extension of the limb (Taylor, 1978). Similar traits can also be seen in other semiaquatic mammals such as †Puijila darwini or
†Potamotherium valletioni, arguably stem pinnipeds or mustelids from the Early Miocene of Canada. In these taxa, the scapula forms a high, asymmetrical triangle similar to armadillos, where the ventrally-expanded caudal angle provides enhanced muscular power and mechanical advantage for the teres major. The acromion, reminiscent of Maiacetus, is on the other hand suggestive of strong but comparatively linear arm protraction and retraction (Smith and Savage, 1956; Northover, 2011; Fernández-Monescillo et al., 2018). The scapular blade of the manatee, triangular in shape, with prominent spine extending into a lappet-shaped superior process and a long, tapering acromion, appears to be overall fairly similar to that of Maiacetus. Nonetheless, the shoulder blade of the manatee is not as gracile and dorso-ventrally elongated as that of Maiacetus, supraspinous and infraspinous fossae are more or less of equal size, cranial and caudal angles are not flared, and the scapular neck is much shorter and at an angle to the scapular spine. Furthermore, in Maiacetus the cranio-ventrally directed acromion runs parallel to the plane of the blade, whereas in manatee it is much more laterally directed. The humerus too, appears to some degree similar, albeit stockier than that of Maiacetus. The humeral shaft is not particularly flared and there is only a moderately developed deltoid tuberosity. The main difference is the high, continuous greater and lesser tubercle in the manatee.
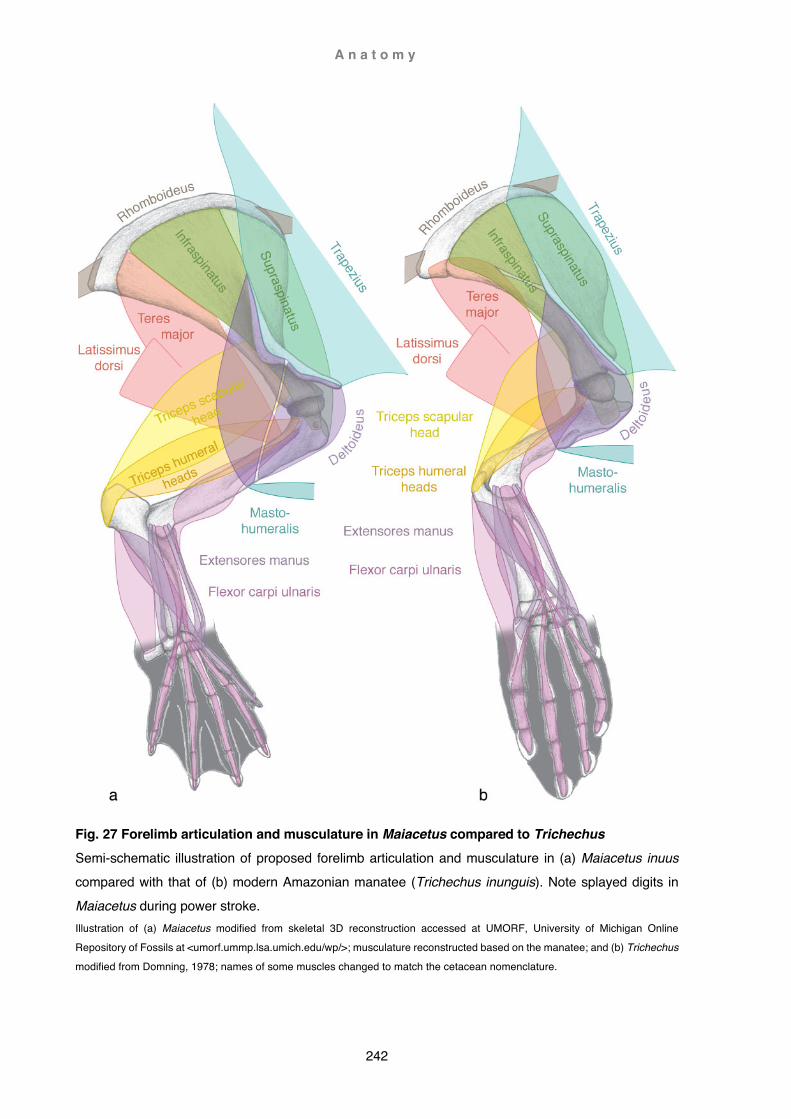
A n a t o m y
242
Fig. 27 Forelimb articulation and musculature in Maiacetus compared to Trichechus Semi-schematic illustration of proposed forelimb articulation and musculature in (a) Maiacetus inuus compared with that of (b) modern Amazonian manatee (Trichechus inunguis). Note splayed digits in Maiacetus during power stroke. Illustration of (a) Maiacetus modified from skeletal 3D reconstruction accessed at UMORF, University of Michigan Online
Repository of Fossils at <umorf.ummp.lsa.umich.edu/wp/>; musculature reconstructed based on the manatee; and (b) Trichechus
modified from Domning, 1978; names of some muscles changed to match the cetacean nomenclature.

A n a t o m y
243
In the antebrachium, the olecranon of the ulna is underdeveloped; radius and ulna are fused at either end, and jointly form the humeral articular surface proximately; supination of the forearm is consequently not possible. Although there is some variability, attachment of the deltoid is fairly congruent in all three species (Trichechus manatus, T. inunguis, T. senegalensis). Obliquely triangular, the deltoideus arises from
the scapular spine and acromion to insert about midshaft onto the lateral rugosity of the humerus. Similar to eared seals seals, the muscle is more or less continuous, and only partially divided by a deep longitudinal tendon (Murie, 1872; Quiring and Harlan, 1953; Domning, 1978). The muscular configuration in manatee thus raises the question, whether the deltoideus in protocetids may also have been consolidated, rather than split into two distinctive parts. The answer may lie not so much in scapular characteristics as in the manner how the forelimb was utilised. Functional implications of forelimb morphology
While there are some similarities of the forelimb in Maiacetus and manatee, differences
become more significant when viewed in ecological and evolutionary context. Manatees are heavily-set, large, fully aquatic mammals that predominantly rely on their broad paddle-like tails for propulsion. They are herbivores and graze by cropping various water plants, although they are also able to manipulate food with the flipper (Howell, 1930a; Domning, 1978; Heithaus and Dill, 2009). Maiacetus was comparable in size and weight but utilised its large hindfeet for propulsion and was essentially a carnivore / piscivore predator of open, coastal waters (Gingerich et al., 2009; Bajpai and Thewissen, 2014). Consequently, speed and agility would have been important. This is supported by changes in the axial and appendicular skeleton towards more efficient aquatic locomotion (Buchholtz, 1998). Early protocetids had four completely fused (synostosis) sacral vertebrae and a fully functional sacroiliac joint (Maiacetus). In later protocetids hindfeet were further enlarged and sacrals began gradually to disassociate, first by synchondrosis (Rodhocetus), then by ‘caudalisation’, i.e. the posterior-most sacral became in appearance and functionally an anterior caudal vertebra (with haemal processes, Qaisracetus). ‘Caudalisation’ of more anterior sacrals continued (Natchitochia) until only the first sacral remained functional, retaining expanded auricular processes. Articulation with the ilium was subsequently lost (Aegicetus) and the first sacral

A n a t o m y
244
became effectively a sacral lumbar (without haemal processes, Georgiacetus). Later protocetids are characterised by an increased ‘lumbarisation’ of the more posterior sacral caudals (basilosaurids). These changes enhanced mobility of the axial skeleton and facilitated more efficient aquatic locomotion: from alternate pelvic paddling (Maiacetus), via simultaneous pelvic paddling and dorsoventral pelvic undulation
(Georgiacetus) to caudal oscillation (Dorudon and modern cetaceans; Uhen, 2014b; Gingerich and Zalmout, 2019). Locomotory models of archaeocetes are predominantly based on lutrine swimming styles (Lontra, Enhydra, Pteronura) yet forelimb morphology remains contradictory. Otters are capable of pronation and supination; they are also able to manipulate their food and also use their forelimbs to great advantage when manoeuvring in water (Fish, 1994; Fabre et al., 2014; Jefferson et al., 2015). The hand of protocetids is transversely fixed into a pronated position and equipped with hooves (Lambert et al., 2019), which would make manipulation of food unlikely. Also of relevance is the huge size difference, potentially affecting overall manoeuvrability. Drag forces scale with linear dimensions, whereas manoeuvrability, i.e. turning radius, is dependent on body mass and swimming velocity (provided all other parameter remain constant). Manoeuvrability is strongly correlated to the size of propulsory surfaces utilised to generate thrust. Undoubtedly, these factors would have accounted for the disproportionally large hands and feet in protocetids (Fish, 1997; Webb, 1997; Gingerich, 2003b). While Maiacetus has body and limb proportions closer to lutrines, Rodhocetus proportions are more akin to those of the desman, including larger feet. This led to the conclusion that Rodhocetus was a slightly more specialised swimmer than Maiacetus and used pelvic paddling with forelimbs folded against the body during rapid swimming (Gingerich et al., 2001a, Gingerich et al., 2009). A medially-bowed, flattened humeral shaft may have
contributed to fit the limb snug against the torso, as well as affecting streamlining when protracted. While configuration of the humeral head and tubercles are more or less congruent with earlier archaeocetes, the anteriorly-directed acromion suggests more powerful protraction of the forelimb. The flaring scapular angles, particularly the extended caudal angle, are reminiscent of the crescent-shaped caudal angle seen in fossorial armadillos (Dasypus sp.), and portend the fan-shaped contour seen in later cetaceans. Extension of the caudal angle is linked to increasingly strong limb retraction, and the rounded dorsal border of the scapula calls to mind increased efficiency of scapular rotators (Taylor, 1978). A large olecranon indicates powerful

A n a t o m y
245
extension of the forearm, and deep humeral olecranon and radial fossae allowed great range of flexion and extension. Freed from the restrictions of a ventrally-overhanging acromion process, Maiacetus would also have a potentially greater range of humeral abduction. Nevertheless, overall characteristics of shoulder and forelimb suggest a predominantly parasagittal locomotor pattern redolent of fossorial-scansorial
mammals, and a possible coincident differentiation of the deltoideus into a pars acromialis and pars spinalis. Aquatic locomotion in protocetids
So far, the exact role the forelimb played in early aquatic locomotion is still unclear. It is unlikely forelimbs would have been used as flippers, yet analysis of the forelimb in another early protocetid from the Upper Lutetian of Senegal suggests that the forelimb was actively used during swimming. Vautrin (2019) compared the long pisiform of the wrist in this whale to that of marine turtles, which use a lift-based flapping motion of the forelimb to generate thrust (Rivera et al., 2013). Yet the ‘kinked’ forelimb of turtles is
not comparable to that of mammals. However, Rivera (2013) describes yet another turtle, the pig-nosed freshwater turtle from Australia (Carettochelys insculpta), utilising hybrid rowing-flapping motions of the flipper to generate a combination of lift- and drag-based thrust. Equally important, sea lions also utilise a hybrid three-phase system using the flipper alternately as a hydrofoil or as a paddle. Although the majority of the thrust is generated by drag-force, oscillation of the flippers is used to reduce drag during the recovery phase and initiate the propulsion phase using lift-based thrust (Feldkamp, 1987). Some sea cows (Trichechus inunguis) may also use a similar locomotory pattern, although it is unclear why other sea cow species do not (Domning, 1978). From the descriptions in various papers, it is not apparent how exactly the wrist of other protocetids is arranged three-dimensionally, yet at some stage the forearm rotated from a pronated state to the semi-supinated position seen in basilosaurids and later cetaceans. Although pronate, the strongly arched wrist seen in Maiacetus may be an incipient form of this rotation. Furthermore, the first digit of protocetids is already strongly reduced, and if appressed would have given the hand a buttressed leading edge similar to that seen in later cetaceans. Decreased finger mobility in the Senegal whale may also indicate an overall more hydrofoil-like shape of the forelimb. The long pisiform might have hereby not only provided the flexor carpi ulnaris with a powerful

A n a t o m y
246
mechanical advantage (Gingerich et al., 2009; Vautrin et al., 2020) but also have allowed it to serve as a tensor, causing the caudal border of the forearm to become rigid, a function that is in sea lions performed by the antebrachial fascial musculature (English, 1976).
Terrestrial locomotion in protocetids
Because of the large webbed hands and feet, terrestrial locomotion of early archaeocetes has been compared to that of eared seals (Otariidae) (Thewissen and Hussain, 1996; Gingerich et al., 2001a). However, on land inability to supinate would have prevented protocetids from placing the forelimb in the manner fur seals and sea lions do. The forearm of sea lions (Otariinae) is held midway between supination and pronation and, although not completely fixed, these forearm movements are limited. On land, the manus is placed plantigrade on the ground with digits laterally directed. This is facilitated by very mobile carpals and a wrist that is bevelled towards the ulnar side, allowing the fur seal to maintain parasagittal elbow flexion. However, propulsion
comes mainly from a series of finely tuned, regionalised carpal and digital movements that translate into a transversely (to the hand) directed lever propelling the animal forward. These movements are only possible because of highly modified antebrachial and manual musculature and ligaments (Howell, 1930a; English, 1976, 1977). In contrast, protocetid foot posture is digitigrade (Gingerich et al., 2001a), lifting the limb further off the ground than fur seals. Besides, to direct the manus laterally, protocetids would have had to rotate the limb at the shoulder joint to an almost impossible 90º angle. It is more likely that they held the forelimb more or less obliquely with elbows turned inwards, and thus elbow flexion would also have been oblique. Some indication might be the medial bow of the humeral shaft, the asymmetrical humeral trochlea, or a large pisiform allowing increased wrist flexion (Vautrin et al., 2020). Whatever the case, terrestrial locomotion of early protocetids was likely not the rolling gait of sea lions but might have more resembled exaggerated waddling where digits are flexed during pronation and extended just before placed on the ground. Functional shift of the scapular spine, acromion, coracoid and olecranon
With the loss of terrestrial competence, late protocetids became fully aquatic and consequently lost constraints imposed on the limbs to function as a body support on

A n a t o m y
247
land. In 1997 Thewissen and Fish proposed a model in which early cetaceans transitioned directly from pelvic paddling to tail undulation (without a pelvic undulation stage), and in 2019 Gingerich et al. presented a specimen supporting this theory. Aegicetus gehennae from the earliest Priabonian of Egypt is about twice the size of Maiacetus with vertebral characteristics similar to those of lutrines. Although the first
sacral bears an auricular process, there is no corresponding articular surface on the ilium and Aegicetus was consequently not able to support its body weight on land. Compared to earlier protocetids, the limbs are relatively small (scaled to comparable body size) and the hindlimb not able to flex during the recovery stroke, but forelimb and manus are proportionally larger relative to the hindlimbs. Gingerich et al. concluded that Aegicetus swam similarly to some later basilosaurids, possibly using a combination of dorsoventral and lateral mid-body to tail undulation, and that the forelimb had gained a greater importance as a stabilising hydrofoil. However, they do not specify how this was achieved and their figures are somewhat confusing. Fig. 14 depicts a lateral, mirrored view of the shoulder blade and forelimb bones, but the humerus is clearly the medial, un-mirrored side and tubercles are not labelled correctly. Both, incorrect articulation and/or mislabelling are a re-occurring problem in the literature and are for these reasons discussed further below (Functional aspect of flipper shapes, section Neocetes including Inia geoffrensis). Fig. 15a,b appear to be unrelated specimens (with the same identification number). However, a detailed 3D rendering of the right humerus (labelled ‘left humerus’), can be viewed at UMORF (see above, Protocetidae) and is used here for a description. The humerus looks roughly similar to that of Maiacetus but is not as gracile. The most significant points are the strong posteriorad orientation of the head and the height of the greater tubercle, both of which would have prevented strong extension of the upper arm. Humeral lateral
torsion orientes the trochlear facet at an angle of 45º and would have deflected the forearm laterally away from the body, particularly when flexed. Deep olecranon, radial and possibly coronoid fossae would have allowed great ulnar extension and flexion. The olecranon (Gingerich and Zalmout, 2019 fig. 14) is large and square as in Maiacetus but oriented at an almost right angle to the ulnar shaft instead of dorso-caudad, which could be interpreted as a possible indication of functional change or altered forelimb posture (see below Functional aspects of flipper shapes, orientation of the olecranon).

A n a t o m y
248
There are many almost completely known fossil remains of basilosaurids and one of the best known is Dorudon atrox. Uhen (2004) described its skeletal characteristics in great detail and deduced a likely muscular arrangement and functional implications. For that reason, only the most salient points will be addressed here. Derived characteristics
- Scapula: triangular, fan-shaped blade with arched dorsal border; two ill-defined spines separating three lateral fossae (supraspinous, infraspinous, teres); a large, blade-like and well-developed anteriorly-directed acromion; a sturdy coracoid process forming together with the acromion a U-shaped through; a shallow glenoid fossa
- Humerus: short with hemispherical head that is not strongly caudally oriented; both tubercle (greater and lesser) situated medially to head; single trochlea accommodating radius and ulna
- Brachium and antebrachium: medio-laterally aligned and habitually more extended - Radius and ulna: flattened and antero-posteriorly arranged; small range of forearm
flexion and extension - Manus: flattened and fixed in semi-supinated position; large, fused carpals; very
limited range of manus flexion or extension Uhen (2004) concluded that forelimb motions and use were very similar to extant cetaceans and that the limb served as a hydrofoil, paddle (stopping or starting) or for social touching (see below Functional aspects of flipper shapes, Dorudontinae). One of the most striking differences to protocetid forelimb morphology is the dramatic reduction of the scapular spine. Smith (1956) associated the absence of the spine in the cetacean scapula with the need to facilitate greater abduction of the flipper. Yet,
there is a well-defined spine in eared seals, animals that propel themselves in water with predominately medio-lateral strokes of the foreflipper (Howell, 1930a). Howell (1937) investigated the shoulder morphology across a multitude of vertebrates and correlated the presence of the scapular spine with the presence of the trapezius. In ungulates the trapezius inserts onto the tuber spinae where as a whole the muscle elevates the shoulder blade, or may impart cranio-dorsal (cervical part) or caudo-dorsal (thoracic part) pull to enhance movements of the foreleg (see also above Drag-based swimming as a catalyst, section Indohyus; Sisson, 1910; Smith and Savage, 1956). In sea lions the trapezius is strongly associated with the deltoideus, forming a composite

A n a t o m y
249
muscle resulting in greater functional length and enhanced performance throughout the upstroke of the flipper (English, 1977). The flipper in cetaceans, however, is not involved in propulsion but used as a rudder. Consequently, some muscles are used more as stabilisers to resist deflection rather than initiate locomotion (Kleinenberg et al., 1969). The cranial orientation of the acromion in protocetids (enhancing active
protraction of the acromio-deltoid during locomotion) may thus be viewed as exaptation for the deltoid to act as a cranial and dorsal stabiliser of a shorter and hydrodynamically more efficient forelimb. This functional change may have negated the need for the trapezius or alternatively, selection pressure may have favoured a larger deltoideus. Pressure drag on the leading edge of a flipper necessitates strong stabilisation to prevent backwards dislodgment of the forelimb but also against dislocation of the shoulder joint (Benke, 1993; Fish, 1993b). In modern cetaceans, stabilisation against backward collapse of the flipper is mainly accomplished by the mastohumeralis, supraspinatus and deltoideus. This would probably not have been very different in Dorudon. On the other hand, Dorudon was also able to flex the forearm (to some degree) and may have in this way reduced torque on the shoulder, a technique some seabirds employ when diving (Fish, 2016). With the reduction of the scapular spine, the deltoideus gained broader insertion on the scapular blade (Uhen, 2004). Likewise, the scapular blades of other dorudontines like Chrysocetus healyorum also appear to have similar arrangements (Uhen and Gingerich, 2001 fig. 9a). Myological reconstruction of Dorudon shows the deltoid as confined on the scapular blade between supraspinatus and infraspinatus, and further craniad limited to the acromion process, somewhat akin to that seen in most terrestrial mammals (Uhen, 2004 fig. 117b). Yet in modern cetaceans the deltoideus completely covers the supraspinatus
and gains strong aponeurotic attachment on the prescapular fascia spanning between the cranial scapular angle and the dorsal border of the acromion. This fascia adheres to a sharp edge on the dorsal border at the base of the acromion, described by Uhen (2004) as the forward slanting part of the spine in Dorudon. It is thus quite possible that such a fascia also existed in Dorudon and the deltoideus consisted of a very large, consolidated muscle covering the entire supraspinatus rather than somewhat disjointed attachment areas (Fig. 28). A flattened and broadly shaped acromion are consistent with this notion. Consequently, in modern cetaceans the cranial limit of the

A n a t o m y
250
deltoideus is ultimately dependent on the extent of the cranial angle and the forward projection of the acromion process (see also Results: Myology in Chapters 2-4). This appears also to be true for the supraspinatus. In most mammals, the supraspinous fossa is a good osteological correlate for the extent of this muscle, but
this is not the case in cetaceans. As with the deltoideus, the supraspinatus adheres to the deep surface of the acromial prescapular fascia and is medially confined by the coracoid and the fascia extending between the coracoid and cranial angle. Rommel (1990) therefore regarded the prescapular fascia of the coracoid as a functional supraspinous fossa. However, in modern cetaceans, the anterior extent of the supraspinatus is not only strongly correlated with the anterior extent of the coracoid but also with that of the acromion (Benke, 1993), the acromion possibly playing an important role in preventing laterad dislodgement of the supraspinatus. In Dorudon, a large acromion and coracoid process would have likely also been correlated with an extended and larger supraspinatus. Reduction of the supraspinous fossa in later cetaceans was subsequently compensated by both the acromion and coracoid and their respective fasciae as the new ‘supraspinous fossa’. An anteriorly-projecting acromion hereby provides improved mechanical advantage for the anterior fibres of the deltoideus used in limb rotation and protraction in modern cetaceans (Benke, 1993). In combination with the prescapular fascia, the acromion also facilitates improved stabilisation of the abducted forelimb. In this manner, the acromion would have provided both strong extension and abduction of the humerus in Dorudon, yet without the drawback of a laterally projecting process limiting the range of lateral abduction and possibly also causing a bulge in the shoulder with disadvantageous hydrodynamic effects.
Elongation of the coracoid process in modern cetaceans also provides improved leverage, albeit for the coracobrachialis and pectoralis minor (see also Results: Myology in Chapters 2-4). In ungulates, the coracobrachialis adducts and flexes the humerus (Sisson, 1910), but in modern cetaceans it provides stabilisation of the shoulder joint and is important in preventing shoulder dislocation. Similarly, the pectoralis minor serves in ungulates as adductor and retractor of the limb (Sisson, 1910), but in modern odontocetes it acts as a strong rotator of the scapula. In ziphiids the coracobrachialis inserts more caudad on the humeral tubercle than in the other

A n a t o m y
251
cetaceans investigated. Moreover, ziphiids also have a longer coracoid and thus greater leverage for the pectoralis minor and coracobrachialis. The latter may in this manner also additionally serve as a rotator of the humerus (see also below Phylogenetic patterns of muscle distribution). Position and height of the tubercles are strongly associated with the habitual posture
of the forelimb (Howell, 1930a), and the medial shift of the lesser and greater tubercles from cranio-medial and cranio-lateral to medial and cranio-medial supports the concept of a hypertrophied deltoideus and large supraspinatus in Dorudon. On the other hand, the fossil remains of one dorudontine basilosaurid, Ancalecetus simonsi from the earliest Priabonian of Egypt, reveals a very different shoulder morphology: the teardrop-shaped scapula is dorso-ventrally high and slender. But most astonishingly, the triangular acromion looks as if it had been bent back into a ventro-caudad direction (Gingerich and Uhen, 1996 fig. 19). This animal also had a smaller than usual (for basilosaurids) forelimb, limited motion at the shoulder joint and a solidly fused elbow. According to the authors, the limb was carried flat against the rib cage pointing caudad. However, they were unable to draw functional-morphological conclusions with regards to the unusual orientation of the acromion. In terrestrial mammals, the caudo-ventral position of the scapular spine is usually occupied by the metacromion (if present; Seckel and Janis, 2008). This appears not to be the case in Ancalecetus, where the scapular spine lacks a craniad facing process. Rather, this process has been identified by Gingerich and Uhen (1996) as a ventro-caudad oriented acromion, and consequently would have been an attachment point for the deltoid (maybe hypotrophied?). Position and orientation of this process and associated muscle(s) may have in this manner played a more important role in stabilizing the limb and shoulder joint when the limb was retracted (rather than subtended as is usually
the case in fully aquatic cetaceans; see also Functional aspects of flipper shapes below).

A n a t o m y
252
Fig. 28 Forelimb articulation and musculature in Dorudon compared to Berardius Semi-schematic illustration of proposed forelimb articulation and musculature in (a) Dorudon atrox compared with that of a modern whale (b) Arnoux’s beaked whale (Berardius arnuxii). Illustration of Dorudon modified from Uhen, 2004
Mystacodon selenensis, the geologically oldest neocete and earliest mysticete from the Priabonian of Peru, already has the typically fan-shaped scapula of modern cetaceans (Lambert, 2017 fig. 2). The supraspinous fossa is about as large as in Dorudon, but unusually, a broad and long acromion faces cranio-ventrally. The humerus is longer than the radius or ulna. The humeral head seems to be similarly oriented as in Dorudon, but the robust greater tubercle appears lower and more craniad, rather than cranio-mediad, to the humeral head. Distal radial and ulnar facets form a distinct V-shape when viewed laterally. The elbow joint is ankylosed and not mobile; the olecranon process appears to be reduced to a tuberosity caudal to the semilunar notch. There is a prominent tuberosity on the anterior edge of the radius, not

A n a t o m y
253
known in other cetaceans and without known functional correlation in other mammals (Lambert et al., 2017).
Functional aspects of flipper shape
Fully aquatic archaeocetes
Members of the family Basilosauridae were ostensibly the first fully aquatic cetaceans, having completely severed all links to land and are consequently the only archaeocetes included in the clade Pelagiceti (Uhen, 2008). While later mid-Eocene protocetids were no longer able to support their weight on land, their mode of locomotion remained strongly reminiscent of semiaquatic mammals that are still able to do so. Freed from the constraints imposed by the exigency of terrestrial life, basilosaurid forelimb posture changed from vertical to subtended, and functionally from drag-based paddle to lift-based hydrofoil - a flipper (Uhen, 2004). Basilosaurids were the first animals to feature osteological correlates of flukes similar to those in modern cetaceans, and like modern cetaceans had short neck vertebrae and an extended functional torso (Buchholtz, 1998). Extremely elongated torso vertebrae and a long body shape in the larger members of this family (subfamily Basilosaurinae) suggest undulatory patterns, rather than oscillation, as their mode of locomotion (Buchholtz, 1998). Both forelimb and flukes are comparatively small and possibly played minor roles in steering and propulsion (Kellogg, 1936; Buchholtz, 1998). Lateral undulation as the primary mode of locomotion (e.g. Gingerich, 2003a) has to be considered as a potential factor for the reduction in size of the forelimb. Subtended or horizontally abducted flippers would incur drag and hard-to-control instability, hence would have to be retracted close to the body. Smaller limbs consequently might have just been the result of improved
streamlining of the body (Klima, 1992; Fish, 1998a). While osteological vertebral features are similar to those of limbless lizards, and thus may not preclude a lateral mode of undulation (Buchholtz, 1998), several points contradict this mode of locomotion: (1) Horizontally-oriented flukes would be an impediment when carried passively by a laterally undulating body and tail since they would introduce uncontrolled instability (Fish, 1998a).

A n a t o m y
254
(2) Although lateral movements of the tail (lateral sculling) have been observed in some whales, e.g. by Scoresby (1820), this author also clearly states that this was not the principal mode of locomotion and it was only employed when moving slowly. The only mammals successfully adopting laterally directed propulsion are phocid seals, but these mammals use their hindlimbs as main propulsors, not the tail, and have
undergone profound musculoskeletal changes in the arrangements of trunk and hindlimbs to do so (Howell, 1930a). (3) Horizontally-oriented flippers would be of little use as stabilisers and recoil dampeners of laterally-directed forces incurred by lateral undulation, but would work well in conjunction with dorsoventral undulation. However, basilosaurid flippers were hydrofoils utilising lift-based forces, which work best at higher speeds (Fish, 1996; Weber et al., 2009a); thus directional change was most likely primarily achieved by twisting the long and agile body. Flippers were possibly more used to boost steering by unilateral braking or acceleration and during start-and-stop maneuvers, maybe in a similar manner as some modern cetaceans do (see Benke,1993). These findings are also applicable to the smaller members of the basilosaurids encompassed in the subfamily Dorudontinae. Based on vertebral characteristics, Buchholtz (1998) postulated that these animals swam by caudal oscillation utilising a well-developed fluke similar to modern cetaceans, making them much faster and more efficient swimmers than their larger relatives, possibly reaching speeds over 40 km/h (Uhen, 2004). As a consequence, lift-based forces would have been more pronounced, and flippers were actively used for steering and balancing requirements (Fish, 2002). Interestingly, Ancalecetus had a much smaller forelimb, in proportions more like those of basilosaurines, and limited mobility in shoulder and limb joints (Gingerich and Uhen,
1996). Limb reduction or loss in terrestrial vertebrates (snakes and limbless lizards) is closely linked to changes associated with a new lifestyle (burrowing) and developmental changes associated with body elongation. In whales, hindlimb loss appears to be a more complex interplay between a multitude of selective pressures and morphological or physiological constraints (Bejder and Hall, 2002) that might have also influenced forelimb reduction in Ancalecetus or basilosaurines in general. Despite similar swimming modes, the forelimb of Dorudon differed in several significant aspects from those of modern whales (e.g. Berardius arnuxii; Table 1):

A n a t o m y
255
Table 1. Comparative osteological characteristics
Dorudon atrox Berardius arnuxii
Scapula about as high as long longer than high Humerus longer than radius about as long as radius Elbow joint mobile (hinge-joint, 30-40º) interlocking Olecranon fully ossified
directed caudo-proximally ossified portion directed caudad cartilaginous part directed caudo-distally
Pisiform large small Carpals limited movement very limited movement Digit movement variable, limited (< 30º) very limited Phalangeal count ‘hypophalangy’ (1/2/2/2/2) moderate hyperphalangy (1/5/5/4/3)
Uhen (2004) described the range of shoulder movements in Dorudon as similar to those of modern cetaceans, with the exception of some mobility in elbow- and digital joints that modern cetaceans do not have. On the other hand, he also describes both inward and outward rotation as limited by the short excursion of the teres major and supraspinatus respectively. In his description he does not consider the pectoralis major inserting medially onto the deltopectoral crest located at the cranial border of the humerus (inward rotation) or the long actions of the mastohumeralis (outward rotation). Furthermore, in modern cetaceans both supraspinatus and deltoideus are much more extensive, an arrangement that was likely also present in Dorudon (see above Functional shift of the scapular spine, acromion, coracoid and olecranon). A more
‘modern’ arrangement of hypertrophied muscles would therefore also imply a more powerful muscular force acting on the humerus to both stabilise the glenohumeral joint and maintain positional attitude of the flipper, crucial during bursts of high speed. In modern cetaceans, shoulder muscles act in two ways: (1) In terrestrial mammals, gravitational forces push the humeral head into the glenoid fossa and contribute in this manner to the stabilization of this joint (Gregory, 1912). Cetaceans are neutrally buoyant and do not need to support their body during locomotion, instead drag- and lift forces act perpendicular to the surface of the flipper (Fish, 1993). To prevent dislocation, these forces are counteracted by strong muscles, tendons and fibrous tissue. The muscles performing this task insert in close proximity

A n a t o m y
256
to the joint such as the subscapularis, coracobrachialis, supraspinatus, and infraspinatus (Kleinenberg et al., 1969; Benke, 1993). (2) Muscles stabilising the flipper against uncontrolled instability induced by drag or lift surround the shoulder joint and insert more distad (but within the body wall) onto the humeral shaft (higher mechanical advantage). Stabilisation of the flipper against
perpendicularly (latero-medial)-acting drag- or lift forces is achieved by opposite muscles acting like cable stays on an airplane wing. The most powerful muscle, in both mass and leverage, is the deltoideus - while the subscapularis is large but with less leverage, and the pectorales are not as massive but with better mechanical advantage (Benke, 1993). Resistance forces against the leading edge of the flipper are not as strong (Fish, 1996) and are mainly counteracted by anterior fibers of the deltoid, particularly those with greater mechanical advantage near the tip of a long, cranially-directed acromion. Similarly, elongation of the scapula not only support s more muscle mass but also imparts increased leverage (Smith and Savage, 1956; Kleinenberg et al., 1969; Benke, 1993). Coincident with musculoskeletal changes in the shoulder, comparable changes of the limb would have also taken place with increased swimming performance in Dorudon. One of the major factors was the transformation of the limb into a hydrofoil. Well-designed hydrofoils (e.g. in boats or submersibles) maximise lift while reducing drag. They are highly streamlined and characterised by a strong, blunt leading edge and a tapering trailing edge. Aspect ratio of a hydrofoil or aerofoil (ratio between span and planform area) is hereby a crucial parameter when predicting performance (Fish, 1996; Fish and Rohr, 1999; McLean, 2012).
In Dorudon, the humerus, radius and ulna are relatively long and thus very much part of the flipper - although, like in modern cetaceans, the most proximal part of the humerus would have been ensconced in a thick blubber layer (Uhen, 2004). Some reconstructions of the manus portray basilosaurid flippers with widely splayed digits similar to early archaeocetes (e.g. Martínez-Cáceres and de Muizon, 2011; Martínez-Cáceres et al., 2017), while others show the digits in a more parallel arrangement (Kellogg, 1936; Uhen, 2004) more reminiscent of rorquals or Caperea marginata (see Fig. 3a).

A n a t o m y
257
Although basilosaurids show a remarkable similarity to the forearm in sirenians, manipulative use of the hand would nonetheless have been nonetheless unlikely. Sirenians have more flexible elbow and finger joints associated with a complete set of fully functional antebrachial muscules (Fig. 27), whereas in Dorudon both appear already somewhat reduced and more similar to modern cetaceans (Fig. 28; Murie,
1872; Howell, 1930a; Domning, 1978; Uhen, 2004). While splayed digits in the slower basilosaurines might not have been impossible, shoulder and limb bone morphology suggest a more hydrodynamic flipper in which at least the preaxial digits were aligned with the long axis of the limb and supported a well-defined leading edge. This was most likely the case in the faster dorudontines. Although Uhen (2004) states that the flattened tips of the second phalanges in digits 2-4 did not appear to articulate with additional phalanges, in B. arnuxii (and generally in modern cetaceans) the distalmost osseus phalanges flatten to articulate with a small, broad cartilaginous tip (or multiple cartilaginous elements in other cetaceans), particularly in digits 3-4. In modern cetaceans, hyperphalangy (most prominent in digits 3-4 in mysticetes and 2-3 in odontocetes) is induced by the maintenance of a secondary apical ectodermal ridge initiating digit elongation and addition of supernumerary digit elements. Furthermore, interdigital webbing (proposed to be present in Dorudon by Cooper, 2004) is intricately linked to the maintenance of interdigital mesoderm tissue (lack of cell-death) which is a prerequisite for the heterochronic, prolonged formation of additional phalanges beyond the normal mammalian count. (Richardson and Oelschlager, 2002; Cooper, 2004; Fedak and Hall, 2004; Cooper et al., 2007a). Ossification and epiphyseal development of the manus progress hereby from proximal to distal elements (Cooper, 2004; Galatius et al., 2006), a process that may have already been emergent in
Dorudon. If so, terminal elements of the digits may have remained unossified or only partly ossified and thus not apparent in the fossil record. Overall, the proportions of the forelimb in Dorudon appear superficially similar to those seen in rorquals or C. marginata. Long bones / manus ratio in Dorudon (about 5/3; Uhen, 2004 appendix VII and estimates from fig. 5) is most closely matched by C. marginata (just over 2/3, Fig. 3a). However, in C. marginata the humerus / radius ratio (2/3) is the exact opposite to that of Dorudon (3/2), the closest match being basal odontocetes such as Platanista, Physeter or Kogia (Fig. 15d and Chapter 3 Berardius:

A n a t o m y
258
Fig. 11). However, in these species the hand is significantly longer than the arm (Fig. 3b,18c). The overall best fit is thus found in the limb proportions of B. arnuxii. Here the ratio of both brachium / manus (Fig. 3c, Chapter 3 Berardius, Fig. 7c,d) and humerus / radius (Chapter 3 Berardius, Fig. 11) are about equal and reflect a moderately long humerus-
and somewhat shorter hand. In comparison to body size, limb length in B. arnuxii is similar to that seen in Dorudon, Platanista and Kogia, i.e. neither particularly short nor particularly long. The biggest difference between Dorudon and Berardius, apart from the mobile elbow joint, is probably an overall slightly shorter and bulkier flipper, with more distant humeral muscle insertions and most likely straighter articulation of the digits in Dorudon (Fig. 28). Proportions of the forelimb directly relate to muscle performance of the shoulder and forelimb (Sanchez and Berta, 2010). The two main differences between Dorudon and modern cetaceans are the shift of peripheral muscle insertions on the humerus to within or near the body wall, and immobility of the elbow joint in modern cetaceans. In extant cetaceans, the orientation of the olecranon is somewhat ambiguous and various. If large, the bony part extends perpendicular to the ulnar shaft or in a slightly proximad direction, not unlike in terrestrial mammals. The cartilaginous part, however, can be variably turned downwards towards the distal end of the limb (e.g. rorquals and Caperea, Figs. 3a,15a,16a) or form an asymmetrically flared cap, its long axis directed away caudo-distally (e.g. Berardius, Fig. 16b). In Dorudon the (fully ossified) olecranon points caudo-proximad in the usual manner of terrestrial mammals in caudo-proximad direction. In terrestrial vertebrates, the slant of the olecranon away from the ulnar shaft is
determined by the relative angle between ulna and humerus: a caudally oriented olecranon in a straight limb alters orientation to a more caudo-proximal or even proximo-cranial direction in an increasingly crouched stance (Fujiwara, 2009). A long olecranon hereby provides great mechanical advantage throughout a narrow arc of action, while a short olecranon operates with lesser leverage but throughout a wider arc (Gregory, 1912; Smith and Savage, 1956). However, in extant cetaceans the elbow joint is rigid and the usual limb position subtended, not vertically below the body. An explanation for the variance in size and orientation of this bone may therefore be quite different from that in terrestrial mammals. In Dorudon, the proximo-caudad orientation

A n a t o m y
259
of the olecranon may be an indication of a slightly flexed elbow either habitually or when most under strain. In Berardius (and other ziphiids) the olecranon is moderately large, proximally the cartilaginous cap is short and rounded, and distally elongated (Fig. 16b). When compared with terrestrial mammals, both the humeral and the scapular head of the
triceps are not particularly well-developed in beaked whales. Retraction of the forelimb is thus predominantly provided by the teres major and latissimus dorsi, while extension of the elbow joint is not possible. However, simultaneous isomeric contraction of the flexor carpi ulnaris and humeral head of the triceps may tense the postaxial area of the flipper and thus potentially reduce vortices along the trailing edge. In Caperea, which has a disproportionally long olecranon and large axillary fold, this might be a particularly important factor (see Chapter 2 Caperea: Discussion, section Flipper). The correlation between the presence of these two muscles and size of the olecranon ostensibly supports this notion. Dorudon, on the other hand, was able to flex the elbow joint to some degree, consequently contraction of the humeral head of the triceps functionally extended this joint. This may have been important in Dorudon to stabilise both forearm (biceps, brachialis, - triceps humeral head) and arm (mastohumeralis, deltoideus - triceps scapular head, teres major) by using opposing isometric muscle tension. In the fast-swimming rorquals, a short humerus and consequently proximal muscle insertions within the body wall, provide fast action but little leverage, compensated for by an elongated scapula bolstering muscular performance (Sanchez and Berta, 2010). In Dorudon, the long humerus provides good leverage, and a comparatively high scapula facilitates precise muscle action. However, distal humeral muscle insertions
and a particularly prominent deltopectoral crest would have made the base of the flipper rather bulky (Fig. 28), a feature that has also been noted to some lesser extent in B. arnuxii (see Chapter 3 Berardius: Results, section Pectoral limb), and stands in direct contradiction to the high swimming speed postulated for Dorudon. Against this stand slender limb elements - a long arm and a small hand with digits splayed little or not at all, features that are a hallmark of rorquals, whales that are able to reach top speeds of over 50 km/h and sustained speeds over 40 km/h (Ridgway, 1972). Berardius (and ziphiids in general) are slow swimmers but deep divers, and conserve energy by retracting the flippers closely to the body during dives, while using them only

A n a t o m y
260
sporadically when manoeuvring during a hunt (see Chapter 3 Berardius: Discussion, section Wider implications). Dorudon, on the other hand, was able to flex the elbow joint, and may have in this way reduced drag when swimming fast, a technique that is also used by seabirds when ‘flying’ underwater (Fish, 2016), while a long olecranon and distal humeral muscle insertions produced great leverage and power to keep the
flippers stable. Neocetes (Fig. 30)
There are no descriptions of shoulder and limb musculature in the most basal neocetes, the Balaenidae. There are, however, general osteological accounts (e.g. van Beneden and Gervais, 1880) and descriptions (osteological and myological) of the forelimb or manus in Balaena mysticetus (Eschricht, 1866, Struthers, 1878; Kükenthal, 1922; Benke, 1993). They reveal that these mysticetes have long, but also very wide, paddle-shaped flippers. The scapula is about as high as it is long and carries a small acromion and a small coracoid or none at all. There is a comparatively low scapular
spine in the bowhead whale, demarcating a large supraspinous fossa. In right whales, the scapular surface is smooth. Flippers are supported by a relatively long humerus (about as long as the radius) topped by a large, strongly curved subspherical head protruding caudad. As is typical for most cetaceans, humerus and ulna are flared distally; the olecranon is reduced to a small knob. Compared to the long bones, the manus is huge, very wide at the wrist, and supported by five widely-spaced digits. Digits have comparatively low phalangeal counts (moderate hyperphalangy; see also Cooper et al., 2007a) and are arranged more or less in parallel or in radiating (splayed) straight rows. In Eubalaena glacialis preaxial digits appear slightly appressed, and the caudal-most digits faintly curved craniad (Benke, 1993). Myological accounts of the manus describe extensor and flexor musculature (Struthers, 1878; Kükenthal, 1922). A high scapula and large humeral head are ideally suited to move flippers through a wide range of movements. When protracted, the flipper converts to a distally broad triangle on a narrow proximal stalk, a configuration postulated to be ideally suited for drag-based propulsion allowing precise manoeuvrability at low speeds (Benke, 1993; Fish, 2004; Woodward et al., 2006; Sanchez and Berta, 2010). When swept back, the broad base of the triangle faces along the body line. This gives the flipper a contour similar to that of a shark fin (delta-wing in aviation) and together with a low-aspect-

A n a t o m y
261
ratio, provides maximal lift to counteract huge pitch forces from the massive open mouth when skim-feeding (Woodward et al., 2006; McLean, 2012). While a large supraspinous fossa, long humerus (proportionally to radius), low phalangeal count (for cetaceans) and straight row of digits can be interpreted as plesiomorphic traits, the large humeral head and hand are undoubtedly derived characteristics and adaptions
to their highly specialised lifestyle (Benke, 1993; Sanchez and Berta, 2010). Shoulder and forelimb characteristics of Caperea marginata are more similar to those of balaenopterids, except for the oblong-oval shape of the flipper. The scapula is extremely elongated and occurs in two morphotypes (fan-shaped and angular). The supraspinous fossa is tiny. The humerus is much shorter than the radius, and the humeral head not as large and voluminous as in balaenids. The hand is much shorter than the arm, the wrist forming the widest section of the flipper, apart from the extremely long olecranon that is part of the axillary fold. Phalangeal count in the four digits is low, the digits are more or less arranged parallel, preaxial digits slightly appressed, and the caudal-most digit faintly splayed and curved (see also Chapter 2 Caperea: Osteology). The pygmy right whale shows a multitude of adaptations for both slow and high swimming speeds such as cranial adaptions for (slow) skim-feeding, apparent high body flexibility, an somewhat broadened flippers (compared to rorquals) on the one hand but also an extremely elongated scapula, comparatively narrow flippers (compared to balaenids) dominated by very elongated antebrachial bones, and the loss of digit 1. Plesiomorphic characters are the retention of antebrachial musculature and a parallel alignment of digits.
The moderately long flipper of Eschrichtius robustus falls in respect to length / width ratio between those of balaenids and balaenopterids but is distinctly fusiform in shape and more reminiscent of the deltoid shape seen in ziphiids. The scapula is only a little higher than long, acromion and coracoid are moderately well-developed, and the supraspinatus fossa very small. Humerus length (compared to radius) and humeral head are both moderately large (Andrews, 1914; Cooper, 2009). Gray whales are unique amongst mysticetes in their wide range of diets and ability to switch opportunistically among feeding techniques (skim-feeding like balaenids, gulp-feeding like rorquals or suction-feeding on the sea floor). They also undertake long

A n a t o m y
262
migratory journeys but usually remain near the coast (Jones and Swartz, 2009). Their ability to be ‘jack of all trades’ is also reflected in perfectly intermediate characteristics of shoulder and flipper characteristics (see Chapter 2 Caperea: Discussion). The result is a flipper shape that minimises vortices (acute tip) during their long migrations but also bolsters lift-generation at low speeds and allows the flipper to be used as a paddle
(broad midsection) when manoeuvring in confined spaces (see also Fish and Rohr, 1999; Woodward et al., 2006). Rorquals (Balaenopteridae) include Megaptera novaeangliae and the genus Balaenoptera. Except Megaptera, rorquals are the race horses amongst mysticetes, with a shoulder and flipper morphology geared for speed. The scapula is markedly longer than high, carrying a long and moderately wide acromion; the coracoid is also longer than that of balaenids. The few existing accounts of the cartilaginous border in these whales (e.g. Howell, 1930a fig. 36b or Schulte, 1916 pl. 45-46) show that this border emphasises the elongation of the scapula even more. Although the insertion of the deltoideus extends distally to the elbow joint (e.g. Schulte, 1916; Benke, 1993), the shortening of the humerus resulted in an overall closer insertion to the shoulder joint. This reduced mechanical advantage on the one hand, but on the other hand gained increased hydrodynamic efficiency by incorporating these muscles near or within the body wall. Accordingly, elongation of both the scapula and acromion allow wider muscle insertions and more powerful extension and abduction of the flipper despite reduced humeral leverage, a necessary requirement to stabilise the flipper at higher speeds (Benke, 1993). The extended cartilaginous cranial and caudal angles of the scapula also improve mechanical advantage of rotator muscles such as rhomboideus and serratus (Howell, 1930a, 1937), possibly particularly important for small flipper
adjustments when maintaining stability during feeding. The flipper itself is narrow and elongated (high aspect ratio), the tip terminating in an acute point (reduction of vortices; Webb, 1997; Fish and Rohr, 1999). The main part of the flipper is supported by the most extremely elongated antebrachial bones in cetaceans. Although the large bony base of the olecranon suggests a large cartilaginous extension, this is not the case in rorquals. The cartilaginous part is rather moderate in size and does not reach as far distally as it does in C. marginata (Fig. 3a and e.g. B. physalus, Benke, 1993 fig. 18 or B. musculus, van Beneden and Gervais, 1880 pl. XII-XIII, fig. 34). The manus of rorquals is often markedly smaller than the arm (Fig. 3a and e.g. van Beneden and

A n a t o m y
263
Gervais, 1880 pl. XII-XIII, B. acutorostrata fig. 7, B. physalus fig. 20), and digits 2-4 are appressed, while digit 5 may be faintly splayed and curved. Megaptera novaeangliae, on the other hand, stands apart with extremely elongated (up to 1/3rd of the body length) and flexible flippers. Extreme elongation of the flipper skeleton occurs late in ontogenetic development (Kükenthal, 1921) and has been
correlated to late phylogenetic occurrence in both most crownward mysticetes (Megaptera) and odontocetes (Globicephala; Cooper, 2004). Humpback whales use their flippers extensively for manoeuvring, utilising highly complex, and sometimes asymmetrical, movements of the flippers. A highly mobile shoulder joint gives the whale enormous freedom of movement, and extremely high-aspect-ratio flippers provide excellent lift-to-drag performance (Benke, 1993; Fish and Rohr, 1999). Humpback whales are thus able to execute sharp turns and stops other rorquals cannot accomplish. They are also known to use their flippers for hunting (fish herding), social touching, and as yet not fully explained manoeuvres such as holding one flipper in the air or slapping the water surface (Benke, 1993). This versatility, however, has profoundly affected the shoulder morphology of Megaptera: the scapula is not as elongated as in other balaenopterids, and uniquely, acromion and coracoid are missing, whereas the humeral head is larger and well-rounded (Benke, 1993). Altogether, this great mobility may come at the cost of strength, but other mechanisms such as tubercles on the leading edge of the flippers (improved flow control), functional antebrachial tendons (active stiffening of the flipper), sweptback attitude of the flippers during fast swimming (reduction of induced drag and distal loading of the flipper, increased stabilisation) and possibly other, as yet not investigated behavioural and musculoskeletal adaptations, ostensibly compensate for these costs (Benke, 1993; Miklosovic et al., 2004; McLean, 2012).
Sperm (Physeteridae) and pygmy sperm whales (Kogiidae) belong to the most basal extant odontocetes. Their forelimb is characterised by a proportionally (to radius) long humerus with a greatly developed deltoid tuberosity and comparatively low phalangeal count (moderate hyperphalangy). The insertion of the deltoid on the cranially-positioned deltoid tubercle is in position similar to that seen in Dorudon. However, the extremely high scapula carrying a massive acromion, together with an overall very short limb are greatly derived characteristics in Physeter. These characteristics are not as prominent in Kogia (Benham, 1902 pl. IV fig. 7), where digit 5 in some specimens

A n a t o m y
264
also displays a marked caudo-distal ‘kink’ (Fig. 18c), whereas digit arrangements in Physeter and some other specimens of Kogia appear more linear. Phalangeal arrangements and numbers have a significant influence on the shape of the flipper. While both flippers can be described as paddle-shaped, the flipper of Physeter is more squared off like a spade, while that of Kogia is triangular (Figs. 3b,18c; Benham, 1902
pl. IV fig. 7; Benke, 1993 figs. 22, 23; see also Cooper et al., 2007a). Both Physeter and Kogia have a relatively large hand (more than half limb length) enlarging the flipper to a broad paddle. However, because of the short limb length in Physeter, the paddle is very small, whereas the bulky, box-shaped head takes up almost a third of the total body length. Steering while swimming is consequently more efficiently done using the head to deflect water (Howell, 1930a). The small flippers are best suited for precise manoeuvres at low speeds, e.g. they may play an important role in maintaining a tight rosette formation when fending off attacking killer whales (see also Chapter 3 Berardius: Discussion, section Wider implications). The only representatives of Platanistidae, the two subspecies of P. gangetica (South Asian- or Ganges- and Indus River dolphin) are renowned for their unusual side-swimming habit (Herald et al., 1969). A detailed account of the functional morphology of this species has been published by Pilleri (1976). He describes these dolphins as not only using their flippers for steering and balance, but also as a sensory organs along the river bed. The scapula in this species is significantly longer than high, and although the surface is featureless, a missing supraspinous fossa does not translate into a missing supraspinatus (‘deltoid’ in Pilleri, 1976); this muscle is rather supported by the medial surface of a well-developed acromion. A strongly curved vertebral border supports a
hypertrophied ‘trapezius’, identified by Strickler (1978) as a rhomboideus capitis, which has fused with the single rhomboideus vertebralis (origin not reported, hence an alternative interpretation could be rhomboideus vertebralis profundus + superficialis but no rhomboideus capitis). These features suggest a powerful ability to rotate the shoulder and thus extend protraction of the limb (Taylor, 1978). The humerus is fairly long when compared to the very wide radius and ulna (Benke, 1993), and acts as insertion point of robust deltoid (‘supraspinatus’ in Pilleri, 1976) and infraspinatus muscles, which hold the flipper in an unusually extended position for cetaceans (horizontally rather than subtended when the dolphin is upright; Pilleri et al., 1976). As

A n a t o m y
265
in many cetaceans, wrist bones vary in number and fusion patterns: in platanistids from 3-8 bones, but generally 5-6 elements. A conspicuous peculiarity is the presence of an additional centrale and the absence of the ulnare (Turner, 1910, 1912; Pilleri and Gihr, 1976a). The most noticeable feature, however, is the large hand dominated by digits 2 and 5. These digits are longer and have a higher phalangeal count (4-5) than the other
digits in the manus (generally 1-4). The digital arrangement in Platanista is thus unique in cetaceans and produces a squared, spade-like flipper contour (Pilleri and Gihr, 1976a). Antebrachial musculature also appears somewhat irregular, in such features as the absence of the flexor digitorum radialis, unusual arrangements of the flexores digitorum communis and carpi ulnaris, and marked reduction of the extensors (Anderson, 1878). Ziphiidae, or beaked whales, are the most speciose family amongst the larger odontocetes. They have very variable height/length scapular dimensions with fossae marked by low ridges or faint elevations. Coracoid and olecranon are well developed, and in some species either very wide or very long. The humeral head is relatively small, long bones are elongated and straight, and the hand smaller than the arm - altogether features that are more reminiscent of balaenopterids rather than odontocetes. Conversely, digit arrangement is more odontocete-like: preaxial digits are more or less appressed, postaxial digits more widedely spaced, and the fifth digit is splayed with a caudo-distal ‘kink’. The phalangeal count is relatively low (moderate hyperphalangy; Figs. 3c, 18g; see also Chapter 3 Berardius: Osteology; Benke, 1993). Kükenthal (1893a), and Turner (1885, 1909, 1912) investigated carpal arrangements and fusion patterns in ziphiids. They regarded hereby the bottlenose whale (Hyperoodon sp.), together with the beluga (Delphinapterus leucas), as key species
for the identification of homologous carpal elements in other cetaceans. Benke (1993) also identified primitive characteristics in the long bones of ziphiids. He considered particularly the features of the humerus as most similar to those seen in basilosaurids and almost identical with those of the extinct family Eurhinodelphinidae, a family that is regarded as a sister taxon (together with other Delphinida) to ziphiids (Fordyce, 2002; Geisler et al., 2011). Although ziphiids are not regarded as the most basal odontocetes (Fig. 1), retention of plesiomorphic characteristics in the forelimb has been corroborated here. One possible explanation may be that, while ziphiids are arguably the most extreme deep divers, selective pressures might have affected other traits

A n a t o m y
266
(behavioural, physiological, morphological) rather than just those of the limb (see Chapter 3 Berardius: Discussion, section Wider implications). Until fairly recently, ‘river dolphins’ have been lumped together in one , but platanistids are now regarded as basal to ziphiids and the other three species of ‘river dolphins’:
†Lipotes (Lipotidae), Pontoporia (Pontoporiidae) and Inia (Iniidae; Fig. 1; de Muizon, 2009; Gatesy et al., 2012). Based on humeral characteristics, Benke (1993) also united ‘river dolphins’ in one cohesive group but conceded that they were not as homogeneous as those of delphinids. Platanista shows hereby the longest humerus (relative to radius) in odontocetes, while Inia and Pontoporia group closer together with Kogia, Monodon, and Delphinapterus. (Benke, 1993 fig. 139, Chapter 3 Berardius, Fig. 11). Similarly, Platanista has the widest radius and ulna (relative to antebrachial length), while Inia and Pontoporia group better together with Physeter and Kogia and some delphinids (Benke, 1993 fig. 146). †Lipotes vexillifer, the baiji has likely become extinct and there is only one myological report, written in Chinese (Weijuan, 1981). The depictions reveal an extensive panniculus and a scapula that is somewhat longer than high. Shoulder and limb musculature are similar in arrangement to that seen in Pontoporia. Slightly divergent muscles are: the deltoideus which appears to be extremely large in the baiji (see also Howell, 1927 Neophocaena phocaenoides), covering the entire lateral side of the scapula, including the rather small infraspinatus (Weijuan, 1981 fig. 5), and the presence of the teres minor (Weijuan, 1981 fig. 8). The shoulder musculature of Pontoporia blainvillei has been extensively described by Strickler (1978). The Franciscana, Toninha or La Plata River dolphin shows a fairly consistent cetacean musculoskeletal arrangement but also some less-common
configurations such as three rhomboideus divisions, two serratus divisions, and the presence of the omohyoideus and pectoralis abdominalis. In this species too, carpal elements (between 5-7, most commonly 6) show various configurations and fusion patterns, including consistent absence of distal carpal 5, sporadic appearance of carpal 3, and in some specimens absence of the radiale (Pilleri and Gihr, 1976c). Inia geoffrensis, the Boto or Amazon River dolphin, appears to be the most peculiar species among ‘river dolphins’. These dolphins are the most agile and nimble cetaceans and use a wide variety of swimming styles including barrel-roll-, yaw-, pitch- and underwater somersault-manoeuvres. The neck and enormously broad, falcate

A n a t o m y
267
flippers are extremely mobile, and both are continuously used for steering. The flippers are also used as oars in rowing motions, and can be lifted up to 45º above the horizontal plane or tilted vertically in either direction (Klima et al., 1980; Benke, 1993). Interestingly, rather than carpal variance, a supernumerary digit 6 has been described in this species (Pilleri and Gihr, 1976b).
Klima et al. (1980) also report highly derived musculoskeletal characteristics, the most pertinent being the unique participation of the sternum in the articulation of the shoulder joint. Greatly differentiated muscular arrangements facilitate hereby a wide range of flipper movements. This is possible because muscular arrangements in the Amazon River dolphin are much less involved in the stabilisation of the shoulder joint, as they are in other cetaceans, but almost solely utilised for locomotory purposes. In their paper, Klima et al. compare those characteristics with Tursiops truncatus, and relate their findings to the very different lifestyles of the two species. Their findings have later been supported by Benke (1993) who also correlated a high scapular shape and differentiated musculature to slow, highly manoeuvrable swimmers that use their mobile flippers profusely, while associating elongate scapula types with fast, pelagic swimmers using only minute movements of the flippers for adjustments. However, Klima et al.’s (1980) findings with regard to the positional characteristics of the greater and lesser tubercles in Inia compared to Homo and Tursiops are erroneous (including fig. 6, described as a right lateral view of the humerus of Tursiops which is actually the left humerus in medial view). In cetacean evolution, the move from land to water has repositioned the limb from initially under the body in semi-aquatic archaeocetes to an oblique lateral (subtended) position in fully aquatic whales (see discussion above). The humeral head has hereby become more aligned with the shaft,
and greater and lesser tubercles have shifted mediad from an initially cranio-lateral and cranio-medial positions (e.g. Indohyus) to a more cranial and/or cranio-medial position (Fig. 29).

A n a t o m y
268
Fig. 29 Position of humeral tubercles Schematic illustration of positional change of tubercles in relation to the humerus head over the course of cetacean evolution, and a comparison with that in Homo (right humeri, proximal view, not to scale). Abbreviations: (bg) bicipital groove, (ct) common tubercle, (gt) greater tubercle, (h) humeral head, (lt) lesser tubercle. (Images modified from following papers: Tursiops, Inia, Homo Klima et al., 1980; Berardius this study; Dorudon Uhen, 2004;
Togocetus Gingerich and Capetta, 2014; Pakicetus Madar, 2007; Indohyus Cooper et al., 2012 - position of the tubercles
approximation only).
During this process, the greater tubercle has become smaller and the lesser tubercle larger (e.g. ziphiids, Inia), whereas in some cetaceans, the two tuberculi have either merged or the greater tubercle regressed, and the remaining tubercle (called ‘common tubercle’ by Klima et al., 1980 or ‘tuberculum proximale’ by Benke, 1993) shifted to a craniad position (e.g. balaenids) or to a mediad position in the most crownward odontocetes (e.g. Tursiops, see Benke, 1993 fig. 108). In primates, the increasingly

A n a t o m y
269
bipedal, upright stance had exactly the opposite effect. In higher primates, the proximal portion of the humerus has undergone an outward twist (humeral torsion), in addition to a re-positioning of the greater and lesser tubercles laterad of the humeral head (Martin, 1933; Larson, 2015), but in this case both tubercles maintained their size. To complicate matters, positional terminology in human anatomy is not congruent with
that of quadrupedal mammals. To avoid confusion, the terms ‘anterior’ and ‘posterior’ are used in Fig. 29 instead of what in quadrupeds is usually referred to as ‘cranial’ and ‘caudal’. Thus, although the angle of radial border to greater tubercle is tantalisingly similar between some cetaceans and primates (Klima et al. 1980 fig. 6), spatial orientation and anatomical position of the radial border and tubercles are not. Monodontidae comprise two species, Delphinapterus leucas, the beluga, and Monodon monoceros, the narwhal. Belugas are slow swimmers and have a short, broad, and very mobile fusiform to paddle-shaped flippers (Kükenthal, 1893b; Struthers, 1895a; Kleinenberg, 1969). Flippers are usually held horizontally, rather than subtended, and are continuously in use. Belugas are also the only other species besides Inia able to lift the flipper up to 45º above the horizontal plane (Benke, 1993). Kleinenberg (1969) wrote a comprehensive monograph on belugas, also including forelimb musculature but particularly focusing on the variance of carpal elements and the high occurrence of polyphalangy and polydactyly (split and multiple digits) in this species. Fedak (2004) proposed bidirectional growth signal reception in the growing tip of a digit as a possible mechanism for the occurrence of polydactyly in species with broad flippers like Inia and Delphinapterus. Broad flippers in basal odontocetes are either spade-like paddles with a proximal trailing edge more or less parallel to digit 4 and/or 5 (Physeter, Platanista) or somewhat more triangular (Kogia), or deltoid-shaped
(beaked whales) with a marked caudo-distal ‘kink’ in digit 5 which supports the caudal-most corner of the trailing edge (. 3b,c and 18c,g; Pouchet and Beauregard, 1889; Pilleri and Gihr, 1976a). Flippers in Inia and Delphinapterus also have a somewhat triangular shape, but phalanges of digit 5 do not show the marked ‘kink’ seen in Kogia (Kleinenberg et al., 1969; Pilleri and Gihr, 1976b). As a result, the caudal-most corner of the flipper may not be as well supported, thus favouring the division of caudal-most elements in the manus that stabilise this area (Kleinenberg et al., 1969). Like belugas, narwhals are also slow swimmers utilising slightly more elongated fusiform and very agile flippers. Digit 5 appears to be slightly more curved than in

A n a t o m y
270
belugas (Eales, 1954; Howell, 1930b). A large, strongly curved and laterally slanted humeral head, and well-developed tuberculi separated from the shaft by a deep sulcus, suggest even greater flipper mobility than that of the beluga. Although the shoulder articulation in the narwhal is not unusual, the hypertrophied lateral (deltoideus, infraspinatus) and medial (subscapularis) shoulder musculature, differentiated into
broad overlapping divisions (Howell, 1930b), vaguely remind of the highly complex musculature in the Amazon River dolphin. Howell also reported vestigial antebrachial musculature in this species (humeral triceps, biceps, extensors, flexors), although only present as a few muscle fibres. Phocoenidae, or porpoises, are the smaller cousins of the dolphins (Delphinidae). Flippers are comparatively small, and the finless porpoise (Neophocaena phocaenoides) is missing the dorsal fin altogether. Porpoises are moderately fast swimmers. Some species are found in coastal and river environments while others prefer pelagic waters (Benke, 1993; Read, 2009). Their most characteristic attribute, a stocky body build and short beak, is attributed to the retention of juvenile traits (paedomorphy; Gol’din, 2004; Mellor et al., 2009; Galatius, 2010). Very little is known of the pelagic spectacled porpoise (Phocoena dioptrica), the species investigated in this chapter (Figs. 20c-d, 23 a-b). The scapula is higher than long (Fig. 8b), which is somewhat unusual for more crownward odontocetes (porpoises and dolphins, see Benke, 1993 fig. 123); acromion and coracoid process are well developed, the coracoid slightly wider. The ratio of long bones/manus is about 3/4 (Fig. 3d) supporting a broadly fusiform flipper. Deltoideus and infraspinatus are comparatively large, while the teres major appears rather underdeveloped. Interestingly, several muscles (levator scapulae, teres major, serratus anterior-
anterior) partially wrap around the edges of the scapular blade onto the opposite surface. This and the division of the subscapularis by several strong septa is in some fossorial mammals seen as a means to maximise available surface area and fascicle numbers (Taylor, 1978). Antebrachial musculature, on the other hand, is reduced to tough tendinous fibrous tissue on both the medial and lateral side of the manus. In the manus, preaxial digits are slightly appressed, and postaxial digits splayed to support the trailing edge. Particularly digit 5 is widely set apart, very broad and flat, and reduced to two cartilaginous phalanges (Fig. 15c).

A n a t o m y
271
In Physeter wrist bones form the widest part of the flipper, whereas in Kogia and more crownward odontocetes it is digit 5, with the exception of Platanista where digits 2 and 5 form a spade-like distal flipper contour (Figs. 3b-f and 18b-g; Pilleri and Gihr, 1976a; Benke, 1993). The widest part of the trailing edge in Kogia and beaked whale flippers is hereby supported by a fairly long 5th digit with a more or less typical caudo-distal
‘kink’, diverting the digit distad along the caudal edge of the flipper. In these cetaceans, phalanges of digit 5 remain fairly robust, albeit also somewhat flattened. While skeletal elements of the La Plata River dolphin (Pontoporia) flipper appear intermediate between the South Asian river dolphin (Platanista) and Amazon River dolphin (Inia), in the South American dolphins, and particularly in belugas and narwhals, the 5th digit is only slightly curved (Kükenthal, 1893b; Struthers, 1895a; Eales, 1954; Howell, 1930b; Kleinenberg et al., 1969; Pilleri and Gihr, 1976c, b; Benke, 1993 figs. 24-25). Consequently digit 5 meets the corner of the trailing edge at a much more open angle, and this area is therefore mainly supported by the terminal phalanx. In porpoises and dolphins, digit 5 has been reduced to few (if any) phalanges, greatly broadened and dorso-ventrally flattened (Figs. 3d-f, 15b-c, 19c-f, 22-24a-b). These are also the last phalanges to ossify and often remain cartilaginous into adulthood (Galatius et al., 2006). The widest area at the trailing edge of the flipper is thus supported by a broad, stiff but flexible, and extremely flat strut akin to the pisiform in mysticetes (Figs. 3a,d-f; see also Chapter 5 Literature: Osteology, section Description of the manus). Delphinidae, dolphins, are the most speciose and diversified odontocete clade. They also encompass the most diversified flipper shapes: from extremely narrow falcate, (e.g. Lissodelphis, Grampus; Fig. 3f) to broader fusiform (e.g. Orcaella) or oval contours (e.g. Orcinus and Cephalorhynchus; Fig. 3e). The two species investigated
in this chapter encompass a typical falcate (Grampus griseus) and oval (Cephalorhynchus hectori) flipper shape. In dolphins and porpoises, preaxial digits are appressed and postaxial digits splayed. Particularly digit 5 is widely set apart (angulated) and supports the caudal-most corner of the trailing edge. Elongation of the flipper is primarily achieved by extreme hyperphalangy (more than 6 phalangeal elements; Cooper et al., 2007a). This has a two-fold effect: (1) additional phalangeal elements also increase cartilaginous components / joints in the flipper, which may improve load distribution and allow further dorso-ventral flattening without compromising strength; (2) a multitude of joints also allows a smooth curvature of the

A n a t o m y
272
leading edge and highly swept falcate profile (Cooper et al., 2007a). Change of flipper shape is achieved by variation of the angle at which postaxial digits diverge and the degree of hyperphalangy in digits 2 and 3. The only exception is Orcinus, where more or less all digits are widely splayed, strongly curved and only moderately hyperphalangic (Fig. 3e-f; see also Chapter 5 Literature review: Osteology, section
Descriptions of the manus). Risso’s dolphins (Grampus griseus) resemble pilot whales (Globicephala sp.) to which they are closely related (Globicephalinae; Gatesy et al., 2012). A bulky head is characterised by a large melon, no beak and few teeth. While pilot whales are black, Risso’s dolphins are grey and often heavily scarred (Ridgway, 1972). The scapula is in proportion longer than high (Fig. 7a). The acromion is short but wide, and larger than the coracoid. There is no supraspinous fossa; instead the well-developed supra-spinatus is dorsally firmly anchored onto a prominent cranial tuberosity. Deltoid and infraspinatus are not as large as in the spectacled porpoise, whereas the teres major is larger and encroaches craniad onto the infraspinatus (Fig. 22a). As in the porpoise, the subscapularis is divided by multiple septa and antebrachial musculature is reduced to tough tendinous fibres (Fig. 22b). Pilot whales are largely teutophagous, fast-swimming, deep-diving dolphins (Aguilar de Soto et al., 2008) with a falcate, and in some species extremely elongated, flipper morphology. The ratio of brachium/manus is even lower than in the porpoise (markedly under 3/4). Risso’s dolphins are also reported to predominantly feed on squid (Ridgway, 1972) but they have a slightly less falcate flipper contour. Nonetheless, overall skeletal similarity of the forelimb between the two globicephalines suggests that Risso’s dolphins may use a hunting technique similar to that of pilot whales. Hector’s dolphins (Cephalorhynchus hectori) on the other hand, appear to be the exact
opposite of the fast-swimming Risso’s dolphin. In appearance almost like porpoises, Hector’s dolphins are moderately fast-swimming, but very agile, coastal inhabitants (Dawson, 1985). Similar to Risso’s dolphin, the scapula is a little longer than high, the supraspinous fossa small (about the same size as in the porpoise) and anchored to a massive tuberosity dominating the cranial scapular angle (Figs. 8a, 9a); deltoid, infraspinatus, and teres major are roughly congruent in proportion to Risso’s dolphin (Fig. 24a), but the humerus is slightly longer and the brachium/manus proportion somewhat closer to 1/1 (Fig. 18e). Hector’s dolphins feed on a variety of small, fast fish they hunt with great skill (Dawson, 1985). In aerodynamics, oval-shaped wings

A n a t o m y
273
produce (at least theoretically) ideal load profiles (elliptical spanload and a constant downwash) and consequently the most optimal lift-distribution (McLean, 2012), factors that may contribute to Hector’s dolphins performing some of the fastest and tightest turning radii observed amongst cetaceans (Fish, 2002).
Diversity of flipper shapes
Overall, cetacean diversity is mirrored by the diversity of flipper morphologies and body designs. Arising from a conceivably rather bulky, broad dorudontine flipper type that was in contour possibly quite similar to that seen in Berardius, cetacean flippers differentiated into a whole range of morphotypes: from extremely broad circular, triangular or spatulate paddles, to high-ratio elongated and/or highly tapered falcate, oval or oar-shaped flipper types (Fig. 30, see also Chapter 1 Methods and materials: Flipper shapes and Pectoral limb in this chapter). Variances of fin and body contours have been correlated with swimming speed and propulsive efficiency, and are the result of a delicate balance between stability and manoeuvrability requirements (Fish,
1998b; Fish and Rohr, 1999; Woodward et al., 2006). However, flipper morphologies also have a phylogenetic component (Sanchez and Berta, 2010), and in order to establish ancestral character states, forelimb arrangements in Dorudon are used here as a model for comparative analysis (Fig. 30). - The scapula is only slightly longer than high (length/height ratio ~1), with a scapular spine demarcating the supraspinous fossa occupying roughly the cranial 1/4 of the scapular blade. - The humeral head is well developed but not overly large, greater and lesser tubercle are discernible and situated cranio-laterad and cranio-mediad to the humeral head. - The humerus is longer than the radius, a long and prominent delto-pectoral crest rises from the cranial edge of the humeral shaft, and distally a mobile hinge joint articulates with the radius and ulna. - Radius and ulna are slightly curved and distally not flared; the olecranon is large, blade-like and proximo-caudally oriented. - The arm is significantly longer than the hand (brachium/manus ratio just under 3/2). - The wrist is composed of seven carpal elements arranged in two rows including a large pisiform caudally.

A n a t o m y
274
- Digits are arranged in a straight line, preaxial digits possibly slightly appressed and postaxial digits somewhat more widely spaced, the phalangeal count is low (typical mammalian or lower). Despite superficially similar shapes, underlying skeletal morphology can reveal
divergent propensities in different clades. Although both mysticetes and odontocetes have broad or elongated flipper shape types, the ways in which these contours are achieved are different. In mysticete paddle-type flippers, enormously cranio-caudally elongated wrist elements support widely spaced digits that are either aligned in more or less straight, parallel lines or slightly splayed, radiating rays. While the pisiform is greatly developed, the olecranon is reduced to a small knob. The flipper is thus widely expanded distally and recedes proximally onto a narrow base, giving the flipper a broad triangular shape. In odontocetes, only Physeter has a somewhat similar arrangement, but here a fairly large olecranon maintains a wide flipper base, giving the flipper a more squarish, spatulate shape. In Kogia and ziphiids, preaxial digits are notably appressed, while postaxial digits are more splayed, particularly digit 5 which appears caudo-distally ‘kinked’. Digit 5 is hereby composed of multiple, little-elongated and only moderately compressed digital elements that support the caudal-most corner of the trailing edge (Fig. 18c,g). Depending on brachium/manus proportions, the flipper takes on a more triangular shape where the hand is large (Kogia), or a deltoid shape strongly influenced by brachial components (ziphiids). Platanista reveals a unique digital arrangement where digits 2 and 5 are longer than the rest, resulting in a spatulate-shaped paddle. As in the more basal taxa, digit 5
supports the thinning caudal area of the flipper by running almost parallel to the trailing edge, but does not show the prominent ‘kink’ seen in ziphiids and Kogia.

A n a t o m y
275
Above: Key to Figure 30 Overleaf: Fig. 30 Characteristics of shoulder and flipper morphology in cetaceans Schematic illustration of cetacean shoulder and flipper morphology projected onto a phylogenetic tree modified from Fig. 1 (not to scale). (Phylogenetic tree based on Fig. 1, data modified from (Benke, 1993) and results of this study in addition to following papers:
Dorudon (Uhen, 2004), Eschrichtius (Andrews, 1914), Physeter (Pouchet and Beauregard, 1889), Platanista (Pilleri and Gihr,
1976a), †Lipotes (Weijuan, 1981), Pontoporia (Pilleri and Gihr, 1976c; Strickler, 1978), Inia (Pilleri and Gihr, 1976b), Monodon
(Howell, 1930b), Delphinapterus (Kleinenberg et al., 1969), hyperphalangy and digit loss (Cooper et al., 2007a).
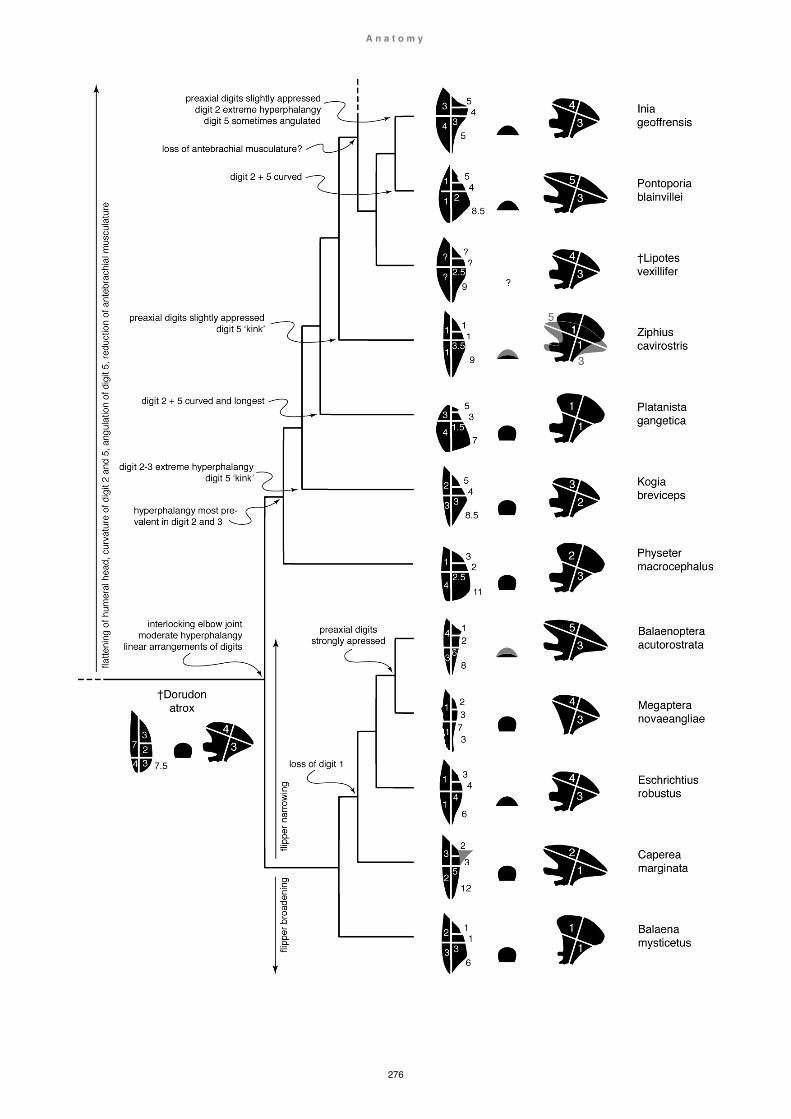
A n a t o m y
276

A n a t o m y
277

A n a t o m y
278
In Pontoporia, Inia, possibly †Lipotes, Monodon and Delphinapterus, flippers are broadly-shaped with a comparatively long, straight or only slightly curved digit 5 supporting the trailing edge. While in the La Plata River dolphin the orientation of digit 5 is similar to that of the South Asian river dolphin, in the former digit 5 approaches the trailing edge of the flipper at a wider angle. This may allow the trailing edge of the
flipper to thin out more, but may also weaken structural integrity in this area and could be a possible causal link to the higher occurrence of polydactyly and polyphalangy seen in the Amazon River dolphin and beluga where this angle is even wider. The most extreme digital arrangement can be seen in the flippers of delphinids and phocoenids (see Osteology, section Manus in this chapter; Benke, 1993; Cooper et al., 2007). Here digit 5 is also strongly caudally displaced but reduced to few (if any) broad phalangeal elements that remain straight and predominantly cartilaginous. In wide paddle-shaped or elliptical flippers, distal elements of digit 4 also remain more or less straight, and flatten into broad terminal elements, while axial and preaxial digits taper into a sharp tip (Figs. 3e, 15c). Such digital arrangements may allow greater thinning out of the postaxial flipper area while maintaining broad and firm, yet flexible, support of the trailing edge. A notable exception is hereby the nearly circular-oval flipper shape in killer whales. Here the central digits are strongly curved and set widely apart, and digits do not reach the trailing edge. Interestingly, the wrist in this species remains predominantly cartilaginous, a trait that has also been reported for mysticetes. It may be that cartilage in this area is better able to mitigate loading strain from the flexible manus onto the rigid bones of the forearm. For the same reason, it might be necessary for mysticetes to maintain active tension control of the outer parts of the flipper, since digits in the manus have fewer phalanges and proportionally more rigid bone than those in
crownward-most odontocetes. A similar divergence in skeletal arrangement of the flipper can also be seen in elongated cetacean flipper types. Balaenopterids show hereby the most extremely elongated, high-aspect-ratio oar-type flippers. A narrow planform of the flipper is achieved by loss of digit 1, narrow carpus, and closely appressed, more or less straight digits 2-5. This flipper type is dominated by highly elongated radial and ulnar components, and a proportionally small manus (Fig. 3a). The pisiform and a fairly large olecranon serve hereby as anchor points for

A n a t o m y
279
the flexor carpi ulnaris, and in conjunction with the humeral triceps, may serve to stabilise the trailing edge of the flipper in a similar manner as cable stays do in wings of vintage airplanes. In a wider oval or fusiform flipper contour, the wrist also widens, and digits are more spaced apart; digit alignment, however, remains fairly straight (e.g. gray whale). In the elongate-oval flipper of the pygmy right whale, the wrist remains
fairly narrow, but digits are set more apart. Here a massive caudo-distally-directed olecranon supports a large axillary fold (Fig. 3a). In contrast, elongated, high-aspect-ratio flippers in odontocetes are dominated by a proportionally large hand. A falcate and highly swept planform is achieved by extreme hyperphalangy of digits 2-3 (see also Fedak and Hall, 2004), and reduction in phalangeal elements of digits 4-5 - apart from digit 1 being reduced or lost across all extant cetacean taxa (Fig. 3f). A high proportion of phalangeal cartilage, particularly distally, may mitigate load stresses onto the carpal elements that can as a result remain fairly rigid and ossified. There are no postulations in the literature why the leading edge in high-aspect-ratio mysticete flippers remains fairly straight but in odontocetes curves caudally to produce a highly swept-back planform. Both clades retract flippers to a more swept-back position with increased speed (Benke, 1993). The most palpable difference between these two groups is their feeding strategy. While dolphins capture their prey without incurring large additional reactional forces, rorquals experience massive and abrupt pitch forces when opening their mouth and engulfing large volumes of water (Cooper et al., 2008). Counteracting these pitch forces may necessitate a sudden increase in lift forces provided by the forward sweeping of the flippers - but at the cost of increased drag forces and energetically less efficient locomotion.
The elliptical planform of the pygmy right whale flipper might prove more efficient in a forward-swept position (McLean, 2012) than the high aspect-ratio flipper of rorquals - important for a skim-feeding whale that has to maintain this position over a much longer timeframe. Proximally, the axillary fold may then also provide additional root chord length and thus cost-effective lift (McLean, 2012). Balaenids on the other hand, may be able to gain enough lift from their low-aspect-ratio flippers held in a swept-back position, assuming an optimal triangular flipper

A n a t o m y
280
shape (large root chord and minimal induced drag) and thus avoid excessive drag forces (Fish and Rohr, 1999; McLean, 2012). Possibly completely different requirements underlay the somewhat similar deltoid shape seen in beaked whales. Berardius may deliberately harness drag forces by
utilising flippers unilaterally as brakes during sharp turns when hunting, as do killer whales or Hector’s dolphins (Fish, 1997; Fish and Rohr, 1999). In this case, drag forces act to pivot the whale around the centre of the turning circle, and in concert with ventral body flexion, improve manoeuvrability greatly (Fish, 1997). While Hector’s dolphins and killer whales are able to perform the tightest turns at higher speeds, by far the smallest absolute turning circles are executed by the Amazon River dolphins and belugas. They are both slow-swimming cetaceans utilising a combination of body flexion, neck mobility and highly mobile broad flippers to great advantage (Fish, 1997; Fish and Rohr, 1999). The directly opposite technique is employed by sperm whales where the head is used to deflect water and flippers are only used when an additional boost is needed or when executing stationary manoeuvres (Howell, 1930a). Webb (1997) postulated that cetaceans are not able to hover or swim at low speeds because of their stiff flipper design. However, many cetaceans do so with great success using a variety of modifications to reduce stability and improve manoeuvrability, such as smaller or absent dorsal fins, buoyancy control, body flexibility and large, broad appendages (Fish, 1997; Fish and Rohr, 1999; Miller, Patrick J. O. et al., 2004). Shoulder blade proportions in these species also have a tendency to emphasise height rather than width. A higher scapula allows longer muscle fibres to
contract over a longer distance, and consequently move the limb through a wider arc in a more controlled manner. Highly differentiated musculature further promotes precision rather than strength (Klima et al., 1980; Benke, 1993). Coincident high mobility at the shoulder joint hereby ostensibly makes up for the lack of fin nimbleness, something fish cannot do (Ahlberg, 1989; Walker, 2004). A much more rigid body design has been postulated for fast swimming cetaceans, producing more rapid turns but also wider turning circles (Fish, 1997). Lift forces increase exponentially with speed and thus muscular strength to maintain control over steering surfaces becomes a major factor (Benke, 1993; Fish and Rohr, 1999). This is

A n a t o m y
281
facilitated by longer-than-high scapular blades and has been related to multipennate musculature and a more sculpted blade surface (Benke, 1993). In odontocetes, a large dorsal fin (instead of pronounced caudal positioning) and higher speeds improve stability but also decrease manoeuvrability, hence flipper movements are augmented by peduncle flexion and fluke depression in unpowered turns or increased banking
angles during powered turns (Fish and Rohr, 1999; Fish, 2002). The most divergent banking angles have been reported for Inia, flexing body and neck rather than banking, and the beluga, a whale without a dorsal fin, banking at a 90º angle (Klima et al., 1980; Fish and Rohr, 1999). However, both delphinids are rather slow-swimming, whereas the southern right whale dolphin (Lissodelphis peronii), also lacking a dorsal fin, is regarded as a fast swimmer (Ridgway, 1972). Although it is not known how much this species banks into a turn, the lack of a dorsal fin and an extremely elongated and sculpted scapula (Figs. 8c, 9b) suggest a need for great strength and flipper control to stabilise rolling and side slip during turning manoeuvres in this species. Phylogenetic patterns of muscle distribution
See also Table 2
Occurrence and complexity of muscles can be highly variable in mammals. Positional congruence of muscular landmarks can therefore not be taken a priori as proof of phylogenetic relationship or muscle identity. Instead, innervation patterns are used to determine muscular associations (Howell, 1930b, 1937; Sokolov and Rodionov, 1974; Strickler, 1978). Yet, that might also prove difficult in some cases: during dissection in this study, a deeply incised portion of the deltoideus in Berardius arnuxii (Hine Reti) corresponded in size and position closely to the subdeltoideus described in Kogia breviceps by Schulte and Smith, 1918 in fig. 7 (see also Chapter 3 Berardius: Myology, section Teres minor / subdeltoideus). This muscle also occurs ostensibly in Balaenoptera borealis (Schulte, 1916; labelled as ‘supraspinatus’ in Benke, 1993 fig. 63a). However, the subdeltoideus, also called teres minor, is not related to the teres major but is a derivative of the deltoideus and as such innervated by the same nerve as the latter (circumflex or axillary nerve; Schulte and Smith, 1918; Howell, 1930b, 1937; Strickler, 1978). Howell (1930b) subsequently suspected Schulte’s subdeltoideus may just be a division of the deltoideus. Moreover, the ‘subdeltoideus’ of this study was not apparent in the other two B. arnuxii individuals and thus would rather support Howell’s suggestion.

A n a t o m y
282
Another notable example is Phocoena phocoena, a species reputedly encompassing an astonishing multitude of varied muscle combinations (including the omocervicalis in addition to the levator scapulae, single and double division of the rhomboideus vertebralis, serratus anterior-anterior, subdeltoideus, costohumeralis in addition to the pectoralis minor, and the humeral and scapular head of the triceps; Rapp, 1837;
Stannius, 1849; Sterling, 1910; Sokolov and Rodionov, 1974; Smith, G. D. J. et al., 1976). However, the harbour porpoise is also one of the most often described species, and the question thus arises whether the great variance of muscular arrangements in this species truly reflects better sampling or is merely the result of inconsistency including a greater number of investigators? Assessments of muscular arrangements in cetaceans are further complicated by muscle misidentification, poor description, language translation or simple omission in the literature (see Chapter 5 Literature review), the latter is particularly significant since absence of muscles is in many cases not recorded either. For example, Strickler (1978) reported the serratus anterior-anterior as absent in odontocetes except in Pontoporia blainvillei. However, Sokolov and Rodionov (1974) described a muscle in Tursiops and Delphinus as arising from the dorsal part of the first rib and inserting deep onto the cranial angle of the scapula - an arrangement congruent with Strickler’s own definition of the serratus anterior-anterior and also very similar to that found in Caperea marginata (this study). Subsequently, a thorough review of older and more recent literature, as well as the results of this study, have revealed this muscle as more prevalent than previously thought (see also Table 2). Nonetheless, there is a core constellation of consistent muscles (in occurrence as well
as differentiation) that appear to be essential in cetaceans (Table 2, highlighted in grey). These muscles are usually large and linked to three basic functions (Howell, 1930a; Smith et al., 1976; Strickler, 1978; Benke, 1993): (1) glenohumeral integrity (subscapularis, supraspinatus, infraspinatus and coracobrachialis) (2) control of flipper attitude, i.e. balance of mobility / stability requirements in dorso-ventral, cranio-caudal and rotational direction: ab- and adduction (deltoideus and pectoralis major), extension and flexion (mastohumeralis, supraspinatus, deltoideus and teres major, latissimus dorsi), plus the same muscles also assist in controlling

A n a t o m y
283
outward and inward rotation of the limb (mastohumeralis, supraspinatus, deltoideus and infraspinatus, teres major, subscapularis) (3) mobility of the scapula (rhomboideus, serratus) supporting protraction and retraction as well as rotation of the limb Muscles in this core group also secondarily support other functions, e.g. the deltoideus
primarily functions as abductor but anterior fibres may act as extensors or rotators, especially in cetaceans with a long acromion (e.g. ziphiids) or differentiated muscle morphology (e.g. Monodon and possibly also Inia). The rhomboideus vertebralis as a whole adducts the scapula dorsad, but fibres at the extreme ends of the vertebral border can act as a rotators, particularly where the muscle is partitioned into two parts (Howell, 1930b; Strickler, 1978; Klima et al., 1980; Benke, 1993). The levator scapulae (craniad movement of scapula) also occurs fairly consistently in cetaceans and thus may be added to the above core group of muscles (Strickler, 1978). Where this muscle is missing, it is ostensibly functionally replaced by the serratus anterior-anterior (C. marginata, B. acutorostrata, G. griseus). A conspicuous exception are the ziphiids where no craniad adduction of the scapula appears to be present. Equally, the pectoralis minor appears to be absent in mysticetes (or at least in C. marginata and rorquals) but present in all odontocetes (Pilleri et al., 1976 fig. 43 and 47 of P. gangetica show a muscle labelled as ‘deltoid’ congruent with the pectoralis minor; Murie, 1873 did not report this muscle in G. melas but it has been found present in this study). Another noteworthy muscle is the reputedly non-functional humeral head of the triceps, persistent in rorquals and pygmy right whale, and quite commonly present in both basal and crownward odontocetes (see above Functional aspects of flipper shapes,
orientation of the olecranon). A host of variably occurring muscles (referred to here as accessory muscle group) act as additional support of the core muscle group: The omocervicalis has been defined by Strickler (1976) as a duplicate of the levator scapulae and has only been described in the most crownward odontocetes (Superfamily Delphinoidea; see also Chapter 5 Literature review: Myology, Table 2). The subdeltoideus (also called teres minor; abduction and inward rotation of the limb; Smith et al., 1976), on the other hand, purportedly occurs only in rorquals (B. borealis,

A n a t o m y
284
Schulte, 1916; Benke, 1993), more basal odontocetes (Turner, 1885; Schulte and Smith, 1918; Klima et al., 1980; Weijuan, 1981; see discussion above), and possibly P. phocoena (Smith et al., 1976). Most puzzling is the occurrence of the rhomboideus capitis (craniad movement of the scapula). This muscle appears to be present in most odontocete families but rather
sporadically. Conversely, descriptions of the mastoscapularis (duplication of the mastohumeralis; Strickler, 1978), costohumeralis and costoscapularis (duplication of the pectoralis minor; Strickler, 1978) are extremely rare and quite likely reflect true absence of this muscle in most cetaceans rather than just omission in the literature. As a result, distribution patterns of accessory muscles within the phylogenetic tree appear to be obscured by high variability, apparent sporadic appearance and poor identification of muscles. For example, Strickler (1978) and Klima (1980) regarded the simultaneous occurrence of both divisions of the serratus anterior (-anterior and -posterior) as plesiomorphic, yet intermittent recurrence of the anterior division throughout modern cetacean families makes determination of ancestral states difficult. There are no osteological correlates for the serratus, and assessment of ancestral states e.g. in Dorudon (if possible at all) is thus encumbered by uncertainty since in basilosaurids variability of at least some muscles would have been potentially high too. Despite the complexity and variability of muscular arrangements, some muscles or muscle groups show clear tendencies throughout the history of modern cetaceans. The trapezius has only been reported for some rorquals (unknown for balaenids) but is considered absent in either odontocetes (Strickler, 1978) or cetaceans altogether
(Howell, 1930b). Within forelimb musculature, distal antebrachial musculature (flexor and extensors of the manus) has experienced gradual reduction (present but reduced in mysticetes and basal odontocetes; e.g. Carte and Macalister, 1868; Turner, 1885; Pouchet and Beauregard, 1889; Schulte, 1916; Schulte and Smith, 1918 and this study) and subsequent loss (more crownward odontocetes; e.g. Howell, 1930a and this study). However, it is not clear at what exact point the loss of functionality occurred. In ziphiids antebrachial musculature is reduced but potentially still functional (see Chapter 3 Berardius: Myology) while in Monodon it has been described as vestigial and non-functional (Howell, 1930b). There are no explicit descriptions of distal

A n a t o m y
285
antebrachial musculature in †Lipotes, Pontoporia or Inia, but given their phylogenetic position proximal to ziphiids the presence of at least vestigial antebrachial musculature is quite possible. On the other hand, omission of detailed descriptions may reflect the perception that the extreme reduction (loss) of distal antebrachial musculature as seen in more crownward odontocetes is ‘normal’ and does not warrant special mentioning
in this group of ‘river dolphins’. In modern cetaceans, more proximal antebrachial musculature has also experienced overall reduction (e.g. triceps) and has partly become vestigial (humeral head) but in some cases is still functional in the most crownward cetaceans (e.g. scapular head in Globicephala, Delphinus and Tursops; Murie, 1873; Sokolov and Rodionov, 1974; Benke, 1993). In other cases, a whole muscle has been reduced (e.g. biceps in rorquals; Carte and Macalister, 1868; Schulte, 1916), become vestigial (possibly Monodon; Howell, 1930b) or was ultimately lost (most odontocetes; Strickler, 1978 and this study). Differentiation patterns of the core muscle group are fairly constant in neocetes (see Chapter 2-4 Caperea, Berardius and Functional anatomy), with the exception of only two species: Pectoral girdle muscle patterns in Inia geoffrensis appear terrestrial-mammal-like in their complexity (e.g. the tri-partition of the sterno-humero-mastoid complex). Klima et al. (1980) therefore regarded the configuration of these muscles as plesiomorphic. However, several aspects stand against this notion: (1) Dorudon and early neocetes were well adapted to their aquatic environment and most likely did not have such differentiated muscular arrangements (see above Functional shift of the scapular spine, acromion, coracoid and olecranon).
(2) the participation of the sternum in the formation of the glenoid cavity of Inia is unique among mammals, and more specifically among cetaceans. Consequently, such highly derived osteological characteristics would most likely also incur de novo, derived muscular differentiations. (3) Ancestral migration patterns suggest initial adaptation to Miocene epicontinental marine environments similar to those postulated for Dorudon and inhabited by many extant dolphins and porpoises (Reeves et al., 2002; Uhen, 2004). Subsequent sea-level regression drained the vast Paranese Sea separating the northern (Amazon) and southern (Paranas) river basins of South America. While the ancestors of Inia adapted

A n a t o m y
286
to the Amazon’s transformation into a freshwater system, the ancestors of Pontoporia followed the receding marine waters from the Paranas into modern-day coastal areas and the estuary of the La Plata River (Hamilton et al., 2001). Consequently, the highly complex muscular arrangements in Inia, although in appearance similar to those in terrestrial mammals, could be at best regarded as
atavistic but are most likely highly derived, convergent adaptations to a new environment (a notion that has also been suggested by Sokolov and Rodionov, 1974 for the sterno-masto-humeral group), while the lack of differentiation in Pontoporia appears to be a retention of traits already well-suited to coastal marine environments. Shoulder musculature in the second species, M. monoceros, was also described as more complex than usual for cetaceans (Howell, 1930b). Here, deltoideus, infraspinatus and subscapularis appear divided and layered into distinctive divisions. Howell also described a greater number of scalenus divisions and pelvic muscles but at the same time cautioned against homologising these muscular arrangements with those in terrestrial mammals.

A n a t o m y
287
Key to Table 2
Abbrev. Genus and species Vernacular name References Mysticeti (baleen whales) Neobalaenidae Cap mar Caperea marginata Pygmy right whale This study Balaenopteridae (rorquals) Bal bor Balaenoptera borealis Sei whale Schulte, 1916
Benke, 1993 Bal acu Balaenoptera acutorostrata Fin whale Carte and Macalister, 1868
Perrin, 1870 Odontoceti (toothed whales) Physeteridae (sperm whales) Phy mac Physeter macrocephalus Sperm whale Pouchet and Beauregard, 1889 Kogiidae (pygmy sperm whales) Kog bre Kogia breviceps Pygmy sperm whale Schulte and Smith, 1918
This study Platanistidae (South Asian river dolphins) Pla gang Platanista gangetica South Asian river dolphin Pilleri et al., 1976Anderson,
1878 Ziphiidae (beaked whales) Ber arn Berardius arnuxii Arnoux’s beaked whale This study Lipotidae (Chinese river dolphins) Lip vex †Lipotes vexillifer Baiji Weijuan, 1981 Pontoporiidae Pon blai Pontoporia blainvillei La Plata River dolphin Strickler, 1978 Iniidae (Amazon River dolphins) Ini geo Inia geoffrensis Amazon River dolphin Klima et al., 1980 Monodontidae (white whales) Mon mon Monodon monoceros Narwhal Howell, 1930b Del leu Delphinapterus leucas Beluga Kleinenberg et al., 1969 Phocoenidae (porpoises) Neo pho Neophocaena phocaenoides Finless porpoise Howell, 1927 Pho pho Phocoena phocoena Harbour porpoise Rapp, 1837
Stannius, 1849 Sokolov and Rodionov, 1974 Smith et al., 1976
Pho dio Phocoena dioptrica Spectacled porpoise This study Delphinidae (dolphins) Lag alb Lagenorhynchus albirostris White-beaked dolphin Murie, 1870a Ceph hec Cephalorhynchus hectori Hector’s dolphin This study Gra gri Grampus griseus Risso’s dolphin Murie, 1870b
This study Glo mel Globicephala melas Long-finned pilot whale Murie, 1873
This study Del del Delphinus delphis Common dolphin Sokolov and Rodionov, 1974
Pilleri et al., 1976 Tur tru Tursiops truncatus Bottlenose dolphin Benke, 1993

A n a t o m y
288

A n a t o m y
289
Wider implications
Over the course of their evolution, cetaceans have dispersed worldwide and adapted to a wide variety of ecological niches. However, despite their well-documented pathway from semiaquatic quadruped to fully aquatic, fluke-powered ocean dweller many osteological aspects of the archaeocete forelimb have remained enigmatic, myological reconstructions absent or incomplete, and functional interpretations often elusive. In this study, a thorough literature review, anatomical comparison of modern mammals and analysis of stranded cetaceans has allowed insight into some of these questions (e.g. terrestrial locomotion and flipper use in protocetids). Dissection of stranded cetaceans has also revealed new, hitherto unknown or undescribed characteristics of hard and soft tissue (e.g. two types of extremely elongated scapulae, axillary fold and extremely large olecranon in C. marginata) underlining the lack of detailed anatomical knowledge of many modern cetaceans. This study has also highlighted some of the inherent problems purely osteological collections pose for anatomical reconstructions (e.g. position of the forelimb) and data analyses (e.g.
shrinkage, distortion or absence of cartilaginous tissue). Behavioural and biological findings have also been integrated and expanded on in this study, emphasising flipper characteristics as an important tool to understand potentially significant aspects of cetacean ecology. Although cetacean forelimb characteristics are overall highly adaptive, some also encompass phylogenetic components and are thus potentially useful in tracing the evolutionary history of cetaceans.
Future studies
To date, the focus of archaeocete studies has firmly remained on character traits of the skull, vertebrae and hindlimbs. While these characteristics are well suited to explain
various behavioural, ecological and aquatic locomotory aspects, forelimbs are also integral to terrestrial locomotion and often show specialisations not seen in hindlimbs (see above Functional implications of scapular morphology). Fundamental questions directly relating to aquatic and terrestrial locomotion, such as the rotation of the wrist from transverse to semi-supinated position, have so far remained unresolved. Similarly, the transition from drag force-based paddle to lift force-based flipper in later archaeocetes, and the role of cartilaginous wrist elements or hyperphalangy in extant cetaceans have also remained elusive.

A n a t o m y
290
In modern cetaceans, theoretical suppositions of flipper shape or muscular arrangements (particularly of the accessory group) have only been tested by a few studies (e.g. Cooper et al., 2008; Sanchez and Berta, 2010). Increased sampling and dissection of almost any cetacean species may provide better understanding of the correct anatomical position of the forelimb, or explain osteological traits (e.g.
elongation of the cartilaginous vertebral border or angle of the glenoid border), muscular arrangements (e.g. antebrachial musculature in the ‘river dolphin’ group) or specialised shoulder morphologies (e.g. skeleto-muscular arrangements that enable killer whales to haul themselves off the beach after intentional stranding; Guinet, 1995), including phylogenetic or behavioural aspects. Furthermore, computed 3D flow analyses over a wide range of flipper shapes may resolve hydrodynamic properties and associated cetacean biology in more detail. This and live Dtag observations of actual flipper use may thus provide important additional data contributing to the overall understanding of the ecological or behavioural profile of a whale species, and ultimately may aid conservation efforts.
Summary During their transition from land to sea cetaceans underwent profound morphological, physiological and behavioural changes. Changes in the forelimbs were hereby strongly influenced by incipient cursorial and aquatic adaptations already present in their closest non-cetacean ancestors (raoellids; Thewissen et al., 2007). Pachyosteosclerosis in the limbs of the earliest cetaceans (pakicetids) aided trim when swimming and negative buoyancy when walking on the river bed (Thewissen et al., 2007; Gingerich, 2012), while strong cursorial forelimb musculature assisted in production of thrust during quadrupedal paddling, as well as counteracting pitch to support the head above water (Fish, 1996). Hooved forelimbs, not capable of supination or pronation, were unable to manipulate food (Cooper et al., 2012; Lambert et al., 2019). As a consequence, early archaeocetes (pakicetids, ambulocetids), postulated to be piscivores or carnivores, likely swallowed at least smaller prey whole (Thewissen and Hussain, 1996; Bianucci, 2005; Thewissen

A n a t o m y
291
et al., 2007) - a characteristic that has persisted into modern odontocetes (with few exceptions like killer whales targeting much larger prey). Aquatic locomotion in later, more aquatic archaeocetes (ambulocetids, remingtonocetids, and particularly protocetids) resembled that of lutrines (Fish, 1996; Thewissen et al., 2009). Forelimbs were equipped with powerful flexor and extensor
musculature and, in combination with large, flexible and fairly symmetrical hands, produced drag force-based thrust, most likely in a parasagittal direction. A mobile shoulder joint allowed sideways thrust to aid manoeuvrability. Large body size, hands and feet, in conjunction with a lack of specialised characteristics of the forelimb and wrist also meant that their terrestrial gait was profoundly different from any modern mammal. Terrestrial locomotion possibly resembled more a waddling stride rather than the rolling gait of otariids or the bounding leap seen in lutrines. Ultimate severance from land and development of a fluke finally enabled the latest archaeocetes (basilosaurids) to develop a lift force-based flipper (Uhen, 2004, and Discussion in this chapter). Concomitantly, several bony processes in the shoulder and forelimb (acromion, coracoid, olecranon) underwent a morpho-functional shift to allow for more powerful and simultaneously also more (dorsoventrally) flexible flipper movements, as well as improved stabilisation of the flipper. Within the flipper, reduction of digit 1, typical for Eocene artiodactyls, manifested itself as a buttress of the leading edge persisting in modern odontocetes but lost in some mysticetes (rorquals; Cooper et al., 2007a). Although musculature and morphology of shoulder and limb were similar to modern cetaceans, there were some small but significant differences. Persistent antebrachial musculature, albeit somewhat reduced, and distal insertion of shoulder musculature on the arm would have made the flipper rather bulky and less hydrodynamic. These features also suggest a rather broad, low-aspect-ratio flipper
compensating for the lack of hydrodynamic properties. Despite powerful musculature stabilising the flipper against backward collapse and shoulder dislocation, the ability to flex the arm may have still played a role in reducing torque forces in arm and shoulder joints. Furthermore, basilosaurids showed no polyphalangy in the hand and might have been less able to cope with shear forces at the leading edge and tip of the flipper. Hyperphalangy is a hallmark of modern cetaceans (Neoceti), albeit only modestly in more basal cetaceans (Cooper et al., 2007a). Yet, despite huge morphological divergence and ecological niche specialisation, the cetacean scapula has remained surprisingly similar in all pelagicetes, mostly with some variation in the contours of the

A n a t o m y
292
blade and shape of the acromion and coracoid. Flipper shapes, on the other hand, are much more divergent, while digit arrangements supporting the flipper are more constant. In this study, six major types of flipper shape have been identified (paddle, fusiform, deltoid, oval, falcate, oar), including 13 sub-types. However, only three digital arrangements support multiple flipper shape types: (1) straight - digits parallel (2)
moderate hyperphalangy - digits 2 and 5 curved, (3) extreme hyperphalangy - preaxial digits highly appressed, digit 5 angulated and truncated, In basilosaurids, mysticetes and basal-most odontocetes (Physeter), digits are more or less linear, and flipper types supported in this manner are paddle, fusiform, (narrow) oval, and oar-shaped. Digits in less basal odontocetes predominantly show moderate hyperphalangy, a slight curvature of the second digit and a moderately splayed digit 5, either with a strongly curved ‘kink’ (Kogia, Ziphiidae) or even curvature (Platanista, †Lipotes, Inia, Monodontidae) supporting the trailing edge of the flipper. The splay (angulation) in digit 5 is particularly prominent in the Amazon River dolphin and beluga and may be linked to the increased occurrence of polyphalangy and polydactyly reported for these two species. Flipper types in this category include paddle, deltoid, and fusiform contours. Extreme hyperphalangy, highly appressed, curved preaxial digits, and flattened, splayed postaxial digits are dominant in crownward-most cetaceans (phocoenids, delphinids). Digit 5 is strongly angulated and truncated, a feature that has also been reported to occasionally occur in the Amazon River dolphin. Broader flipper shapes (e.g. fusiform or oval) are achieved by a greater splay of digits 2-4. The only exception is killer whales where digits 2-5 are all strongly curved, widely spaced, and do not reach the border of the flipper. While the flipper contour in phocoenids is fusiform, in delphinids it is much more variable from oval, fusiform to falcate - the latter the most
prevalent shape in delphinids. Surprisingly, pygmy right whales are also able to alter the contour of the flipper. While the appendage is held in a swept-back position, the axillary fold is retracted, and the flipper assumes a more rorqual-like elongated outline. Protracted, the axillary fold extending to the wrist gives the flipper a more triangular, shark-like fin shape. Pygmy right whales are thus the only cetaceans able to produce an instant shape-shift in the flipper contour, which possibly also correlates with a functional shift of the appendage (possibly from slight attitude adjustments during normal swimming to instant gross production of lift during feeding or extreme manoeuvres).

A n a t o m y
293
Muscular arrangements in cetaceans are roughly divisible into two groups: a core group of muscles that appears to be extremely constant, and a peripheral, highly variable accessory group that is hard to interpret phylogenetically. Core group muscles fulfill essential functions such as (1) maintenance of glenohumeral integrity
(subscapularis, supraspinatus, infraspinatus and coracobrachialis), (2) control of flipper attitude (latissimus dorsi, teres major, subscapularis, deltoideus, supraspinatus, infraspinatus, pectoralis major and maybe also levator scapulae / serratus anterior-anterior group), and (3) mobility of the scapula (rhomboideus and serratus and possibly also in odontocetes pectoralis minor). Proximal muscles in this group serve hereby predominantly glenohumeral integrity, and muscles inserting more distad on the humerus mainly act as stabilisers, while rotators of the scapula assist with protraction / retraction of the limb and indirectly as rotators of the flipper. In this manner, muscles stabilising the flipper are potentially isolated from muscles rotating the flipper, and thus may avoid destabilisation of the flipper when finetuning pitch. In some odontocetes, core group muscles are highly differentiated into split / fused and/or layered subunits (Amazon dolphin, to a lesser degree narwhal and some other odontocetes). Despite the similarity to terrestrial mammals, based on morphological, phylogenetic and historic distribution patterns, differentiation in these muscles (especially in the Amazon River dolphin) are here regarded as most likely highly adaptive, convergent characteristics and not atavistic or plesiomorphic character traits.
Cephalorhynchus hectori Life reconstruction of an adult Hector’s dolphin

294
Chapter 5: Literature review
1. Osteology One of the most comprehensive osteological papers has been written by Benke, (1993). His studies include 79 species (mysticetes and odontocetes) and compile a broad spectrum of scapula- and forelimb measurements, complete with photographic records. He also includes a detailed description of the shoulder and limb musculature in Tursiops truncatus and Balaenoptera borealis. His analyses are supplemented by analyses of video recordings (13 species) and his findings set into taxonomic and functio-morphological context, also including detailed descriptions of cetacean swimming behaviour and flipper motions. Sanchez and Berta (2010) also provide a comparative analysis of the major bones and muscles in odontocete flippers. They relate their findings to the size and shape of flippers, flipper mobility and swimming behaviour. Osteological and soft-tissue characters are then set into a phylogenetic context. The following categories present a selection of pertinent papers and are by no means complete, authors are listed in historical order wherever applicable: One of the earliest and also most exhaustive publications on multiple species of cetaceans is the work of van Beneden (1880) detailing the skeletal characteristics of numerous extant and fossil mysticetes and odontocetes. Other papers comprise less
detailed descriptions of the forelimb or are restricted to only a few species such as Hunter and Banks, 1787 and Rapp, 1837 (general osteology), Eschricht et al., 1866 (B. mysticetus, O. orca, P. crassidens), Malm, 1871 (osteological inventory), Hector, 1875 (C. marginata, M. novaeangliae, B. physalus), Turner, 1912 (H. ampullatus, M. bidens, P. gangetica, M. monoceros, D. leucas). Some also include myological aspects such as Rapp, 1837 (comparative osteology, myology P. phocoena), Anderson, 1878 (O. brevirostris, P. gangetica, B. edeni and myology of the manus P. gangetica) and Sterling, 1910 (comparative functional osteology and myology of P. phocoena).

L i t e r a t u r e r e v i e w
295
Many papers give a detailed description of the odontocete manus, particularly with respect to variability in carpal elements, such as Weber, 1888 (variability in multiple cetacean species) or Kükenthal, 1893b (D. leucas), Eales, 1954 (M. monoceros, G. macrorhynchus, G. melas, P. phocoena), Kleinenberg et al., 1969 (D. leucas
including myology of the shoulder and forelimb), Watson and Fordyce, 1993 (B. acutorostrata), and del Castillo et al., 2014 (P. blainvillei). Multiple papers have been published by Turner, 1885 (M. bidens incluing myology of the manus), 1893 (B. acutorostrata), 1909 (M. bidens), 1910 (P. gangetica) and three papers by Pilleri and Gihr in 1976 (P. gangetica, P. blainvillei, I. geoffrensis). Other publications focus more on ontogenetic and/or evolutionary patterns in the mammalian or cetacean forelimb and manus, e.g. Howes, 1888 (supernumerary phalanges in mammals), Kükenthal, 1888-1893 (variability of carpal elements, hyperphalangy, polydactyly), Stuart and Morejohn, 1980 (morphometrics P. phocoena), Dawson, 1994 and 2003 (allometry and ossification in cetaceans and Stenella), Calzada and Aguilar, 1996 and DiGiancamillo et al., 1998 (allometry and ossification in S. coeruleoalba), Tanaka, 1999 (asymmetry in humans), Galatius and Kinze, 2003, Galatius 2005-2006, Galatius et al., 2006 (ossification and asymmetry in P. phocoena and L. albirostris), Cooper et al., 2007a (hyperphalangy, polydactyly), Fettuccia et al., 2012 (variability within the genus Sotalia), de Carvalho et al., 2014 and Torres Azevedo et al., 2015 (ossification in S. guianensis). Investigating underpinning mechanisms of limb patterning have been published by authors such as Holder, 1983 (two papers on digit and limb patterning), Klima, 1978, 1990 (clavicles in cetaceans), Bejder and Hall, 2002 (limbs and limblessness),
Richardson and Oelschläger, 2002 and Richardson and Chipman, 2003 (hyperphalangy), and Fedak and Hall, 2004 (hyperphalangy, polydactyly). Particularly Cooper et al., 2007a and Fedak and Hall, 2004 focus on comparative carpal and digital arrangements and ontogenetic patterning (including hyperphalangy, polydactyly, polyphalangy and digit loss). Hyperphalangy in cetaceans and digit loss in tetradactyl mysticetes is set in phylogenetic context, and the resulting flipper shape correlated to function. A handful of papers also describe anomalies of the cetacean forelimb such as Watson et al., 1994-2008 (polydactyly and other anomalies in T. truncatus and I. pacificus),

L i t e r a t u r e r e v i e w
296
Cooper and Dawson, 2009 (digital anomalies), Marigo et al., 2013 (flipper malformation in S. guianensis) and Thomas and Barnes, 2015 (pathologies in extant and fossil marine mammals). Detailed descriptive osteological works of the mysticete shoulder and forelimb have
been published by Murie, 1865 (B. physalus), Dwight, 1872 (B. musculus), Turner, 1893 (B. acutorostrata), Struthers, 1895b (B. mysticetus, B. physalus with emphasis on the carpus), Beddard, 1901 (C. marginata), Andrews, 1914 and 1916 (E. robustus and B. borealis) and Omura, 1975 (B. acutorostrata), while descriptions in other papers are less detailed like Hale, 1931 and 1964 (C. marginata), Marples, 1956 (cetotheres from New Zealand), Gol'din et al., 2013 (Cetotherium riabinini). Some papers also include detailed myological descriptions like Carte and Macalister, 1868 (B. acutorostrata), Struthers, 1871-1889 (B. physalus, M. novaeangliae, and B. musculus), and Benke, 1993 (B. borealis), Shoulder and forelimb in odontocetes have been described more or less in detail by Krefft, 1865 and Benham, 1902 (K. breviceps), Flower in many publications, 1867-1876 (I. geoffrensis, beaked whales, P. macrocephalus), two papers from Haast, 1876 (Z. cavirostris, M. layardii), Struthers, 1895a (D. leucas), Benham, 1902 (K. breviceps), Turner, 1907 (L. acutus), Rommel, 1990 (T. truncatus), de Muizon, 1994 (scapulae of fossil squalodontids) and multiple authors referring to different species of beaked whales (Berardius or Mesoplodon, see Table 1). Papers also including myology have been published by Pouchet and Beauregard, 1889 (P. macrocephalus) and Klima et al., 1980 (T. truncatus with focus on I. geoffrensis).

A p p e n d i x
297
Table 1. Berardius arnuxii Duvernoy 1851 specimens and other southern hemisphere ‘Berardius’ specimens Year Species Reported event
location Additional geographical information
Sex Length of
animal
Specimen Historical repository
and ID Current repository (if
known)
Most important publications Other detailed reports
Short references
New Zealand Waters ~1840 Berardius arnuxii
Duvernoy 1851 Taiaroa Head Otago Peninsula,
Dunedin Otago
Mandible Otago Museum, A14.37 OMNZ VT220 (current)
Oliver, 1922a; McCann, 1975 (ref. as 14.37 and erroneously as 14.31)
1846 Berardius arnuxii Duvernoy 1851
Akaroa Banks Peninsula Canterbury
Male 32 ft (9.75 m)
Skull (Holotype) Paris Museum British Museum, 96.11.20.1
Duvernoy, 1851a; Flower, 1872 (specimen 1, p. 212-213); van Beneden and Gervais, 1880 (specimen without number, p. 387-388, plate 21bis, 23bis)
Gray, 1866; Haast, 1870a,b; Owen, 1870; Beddard, 1900; True, 1910; McLachlan et al., 1966
Hector, 1870, 1873, 1878; Knox, 1870; Marelli, 1920; Oliver 1922a, McCann, 1975
1862 Berardius arnuxii Duvernoy 1851
Porirua Harbour (Bay)
W Coast / Cook Strait
Male 27 ft (8.23 m)
Not preserved Knox, 1871 (specimen 1, p. 126); Flower, 1872 (specimen 2, p. 213); van Beneden and Gervais, 1880 (specimen 1, p. 388)
Ross, 1984 Marelli, 1920; McCann, 1975
1866 Mesoplodon hectori (Gray 1871)
Titai (Tatai, Taitai, Titahi, Tetai) Bay
Porirua Harbour W Coast / Cook
Strait
9 ft 3 in (3.65 m)
Skull (Holotype) Part Skeleton Colonial Museum British Museum, 1677b 76.1.16.3
Hector, 1870; Hector in Knox, 1870; Gray, 1871; Knox, 1871 (specimen 2, p. 126, plate 14’, 15’); Gray, 1873 (p. 111); van Beneden and Gervais, 1880 (specimen 4, 5, p. 388)
Hector, 1873a, b; 1878; Gray in Hector, 1873a Turner, 1872; Gray, 1874a, b; Oliver 1822a,b; McCann, 1962;
McCann, 1975 1868 Berardius arnuxii
Duvernoy 1851 New Brighton River Avon Christchurch Canterbury
Female (uncertain) 30 ft 6 in (9.30 m)
Skeleton Canterbury Museum Royal College of Surgeons
Haast, 1870a,b; Flower, 1872 (specimen 3, p. 213-215, plate 28); van Beneden and Gervais, 1880 (specimen 2, p. 388)
Knox, 1870, 1878; True, 1910; Ross, 1984 Hector, 1870, 1873b; Marelli, 1920; Oliver, 1922a; McCann,
1975 1870 Berardius arnuxii
Duvernoy 1851
Worser’s Bay Port Nicholson Wellington
Harbour
27 ft (8.23 m) Skull and part Skeleton Dominion Museum, 7 MoNZ MM0007 (current)
Knox, 1870; Knox, 1871 (specimen 3, p. 126, plate 13, 16-17); Flower, 1872 (specimen 4, p. 215-216); van Beneden and Gervais, 1880 (specimen 3, p. 388)
Gray, 1871; Hector, 1878; Ross, 1984 Hector, 1873b; Marelli, 1920; Oliver, 1922a,b; McCann, 1975
1873 Berardius arnuxii Duvernoy 1851
Saltwater Creek North of
Christchurch
Skeleton Dominion Museum, 1402
Oliver, 1922a; McCann, 1975
<1874 Mesoplodon hectori (Gray 1871)
Kaikoura
Mandible Dominion Museum
Hector, 1874; Hector, 1878
~1874 Berardius arnuxii Duvernoy 1851
Port Molyneux Clutha district,
Otago
Skull Otago Museum (registered 1874), A339, A.64.03 OMNZ VT 222 (current)
Oliver, 1922a; McCann, 1975 (both ref. erroneoulsy as from Chatham Islands)
1875 Mesoplodon hectori (Gray 1871)
Lyall Bay Wellington
Harbour
Male (adult) 15 ft 6 in (4.72 m)
Skeleton Hector, 1875 Hector, 1878 Buller, 1876; McCann, 1975
1875 Mesoplodon grayi Haast 1876
Waitangi Beach Chatham Islands
Skull, mandibles (two of three syntypes) Otago Museum A50.10, A50.9 OMNZ VT196 (current) OMNZ VT197 (current)
Haast, 1876a (p. 7-13) Haast, 1876a (p. 457-458),b (450-457); Hector, 1878 (p. 339-341, plate 12), Forbes, 1893 (p. 216-236: skull A, F, K,), Oliver, 1922 (p. 572)
1876 Mesoplodon hectori (Gray 1871)
2 Teeth Dominion Museum, 239
McCann, 1975
1877 Berardius arnuxii Duvernoy 1851
Wellington Harbour
Male 27 ft 6 in (8.38 m)
Skeleton Dominion Museum, 183 MoNZ MM0183 (current)
Hector, 1878 True, 1910; Ross, 1984 Oliver, 1922a; McCann, 1975
<1880 Berardius arnuxii Duvernoy 1851
New Zealand Female (uncertain)
Skeleton Paris Museum
van Beneden and Gervais, 1880 (specimen without number, p. 387-392, plate 23, 23bis)
Oliver, 1922a <1920 Berardius arnuxii
Duvernoy 1851 Coast near
Wanganui Skeleton
Wanganui Museum, A339
Oliver, 1922a; McCann, 1975

A p p e n d i x
298
~1924 Berardius arnuxii Duvernoy 1851
Mason Bay (?) Stewart Island
Skull, mandible, scapula Otago Museum (registered 1924), A24.69, VT219 (scapula), VT221 (skull, mandible) OMNZ VT219 (all, current)
McCann, 1975
1929 Berardius arnuxii Duvernoy 1851
Mason Bay Stewart Island
2 Teeth Dominion Museum, 523
McCann, 1975
1929 Berardius arnuxii Duvernoy 1851
Mason Bay Stewart Island
Skeleton Dominion Museum, 416
McCann, 1975
1931 Berardius arnuxii Duvernoy 1851
Te Horo Beach Wellington
Male 28 ft (8.40 m)
McCann, 1975
1934 Berardius arnuxii Duvernoy 1851
Otaki Beach Wellington
Female 28 ft (8.40 m)
4 Teeth Dominion Museum, 528
McCann, 1975
1937 Mesoplodon hectori (Gray 1871)
Plimmerton Porirua Harbour Wellington
10 ft 8 in (3.20 m)
Skeleton Dominion Museum, 614
McCann, 1962 McCann, 1975
1944 Berardius arnuxii Duvernoy 1851
South Tapu Thames Coast
Male 28 ft (8.40 m)
Auckland Museum McCann, 1975
1980 Berardius arnuxii Duvernoy 1851
Pukerua Bay Wellington
Male 29 ft (8.85 m)
2 Teeth Dominion Museum, 1433
McCann, 1975
2010 Berardius arnuxii Duvernoy 1851
Purakaunui Inlet Dunedin, Otago
Adult male 8.70 m
Not preserved This thesis
2013 Berardius arnuxii Duvernoy 1851
Tangimoana, Sandy Point Invercargill
Subadult female ~7.00 m
Skull and pectorals Private
This thesis
2013 Berardius arnuxii Duvernoy 1851
Hine Reti, Beach at Ōmaui Invercargill
Juvenile female ~ 6.50 m
Skull and right pectoral Private
This thesis
New Zealand Waters, data deficient x Berardius arnuxii
Duvernoy 1851 Centre Island Foveaux Strait
Worn tooth Dominion Museum, 528
McCann, 1975
x ?Mesoplodon hectori (Gray 1871)
Chatham Islands 1 Tooth Dominion Museum
Hector, 1873b
x Berardius arnuxii Duvernoy 1851
Pencarrow Wellington Harbour
Male, maybe female and calf below 30 ft (9.00 m)
2 Teeth Dominion Museum 526
McCann, 1975
x Berardius arnuxii Duvernoy 1851
Pencarrow Wellington Harbour
Female, maybe calf of above 20 ft (6.00 m)
Dominion Museum McCann, 1975
x ?Mesoplodon hectori (Gray 1871)
Kaiapoi Beach Wanganui
Skull Canterbury Museum
Hector, 1873b, 1878; McCann, 1962
x ?Mesoplodon hectori (Gray 1871)
Sandy deposits Coast Wanganui
Skull Hector, 1873b
x Berardius arnuxii Duvernoy 1851
East Cape 1 Tooth Dominion Museum, 527
McCann, 1975
x ?Mesoplodon hectori (Gray 1871)
New Zealand 1 Tooth Dominion Museum
Hector, 1873b
x Berardius arnuxii Duvernoy 1851
New Zealand Skull British Museum US Natural Museum, USNM 21511
True, 1910; Oliver, 1922a; McCann, 1975
x ?Mesoplodon hectori (Gray 1871)
New Zealand Skeleton US Nat. Museum?
Oliver, 1922b
x Berardius arnuxii Duvernoy 1851
Skull Dominion Museum, 143
McCann, 1975
x Berardius arnuxii Duvernoy 1851
Skull British Museum, 1949-8.A.1
Oliver, 1922a; McCann, 1975

A p p e n d i x
299
Australian Waters 1935 Berardius arnuxii
Duvernoy 1851 Port Lorne St. Vincent Gulf S Australia
Pregnant female 29 ft (8.84 m)
Skeleton South Australian Museum
Hale, 1939, 1962; MacLachlan et al. 1966; McCann, 1975; Ross, 1984
1977 Berardius arnuxii Duvernoy 1851
Port Davey Tasmania
Beach cast Part Skull Guiler, 1978
1989 Berardius arnuxii Duvernoy 1851
Cape Riche W Australia
Skull West Australian Museum
Groom et al., 2014
Antarctic Waters 1955 Berardius arnuxii
Duvernoy 1851 Carlson Island Crown Prince Gustav Channel
Trapped in ice
Photographs
Taylor, 1957; McCann, 1975
South American Waters 1918 Berardius arnuxii
Duvernoy 1851 Arroyo del
Pescado Los Talas, Rio de
la Plata Argentina
Female 7.80 m
Skeleton Nat. Museum of Buenos Aires
Marelli, 1920; Oliver, 1922a,b; McCann, 1975
1934? Berardius arnuxii Duvernoy 1851
South Shetland Islands
Female British Museum, 1934.6.15.1
McCann, 1975
1949? Mesoplodon hectori (Gray 1871)
Falkland Islands Photograph of skull British Museum, 1949.8.19.1
Fraser, 1950; McCann, 1962
1978 Berardius arnuxii Duvernoy 1851
Tierra de Fuego Argentina
Beach cast 3 specimens Part Skull (693) Vertebrae (472 and 566)
Goodall, 1978
South African Waters 1962 Berardius arnuxii
Duvernoy 1851 Kromme River Jeffery’s Bay East Cape
Male 30 ft 6 in (9.30 m)
Skull Port Elizabeth Museum
McLachlan et al., 1966; Ross, 1984
Abbreviations: MoNZ Museum of New Zealand; OMNZ Otago Museum, New Zealand

L i t e r a t u r e r e v i e w
300
2. Myology Arguably, the overall most-informative paper on cetacean limb musculature has been published by Strickler (1978). In his paper, Strickler gives a general overview of muscular arrangements in cetaceans, as well as a detailed description in P. blainvillei. He also discusses evolutionary development of mammalian musculature arrangements and proposes a standardised nomenclature referring to multiple papers and synonyms. However, all this information is compressed into 13 pages and sometimes hard to follow. For this reason, a summary is presented in Table 2, also amalgamating Strickler’s paper and findings of this study (inclusive literature review). Interestingly, Strickler did not include another informative paper written by Smith et al., 1976. In their paper, they describe and illustrate in detail muscular actions during surfacing manoeuvres of P. phocoena, inclusive those of the flippers. They also give an itemised account of muscular arrangements, as well as associated muscular actions. The paper is complemented by a list of synonyms, albeit without explanations.
Probably the most influential paper on mammalian musculature patterns, ‘Morphogenesis of the shoulder architecture’, has been written by Howell in 1937. In his paper, Howell gives a general account of muscle innervation patterns and associated muscle arrangements encompassing a wide range of extant mammals, including cetaceans. However, only few papers have so far specifically investigated the cetacean forelimb in an evolutionary context, namely Sanchez and Berta (2010) who analysed morpho-functional aspects (inclusive flipper shape), and Cooper et al. (2007b) who published a more detailed paper on neuromuscular arrangements but restricted to antebrachial musculature. Detailed descriptions of mysticete shoulder and forelimb musculature have been published by Carte and Macalister, 1868 and Perrin, 1870 (B. acutorostrata), and Schulte, 1916 and Benke, 1993 (B. borealis). A minor caveat in Benke’s publication is the inclusion of the subdeltoideus into the supraspinatus. One of the earliest reports on cetacean musculature of the arm and manus has been published by Flower, 1865 (B. physalus). Struthers and Kükenthal published many papers predominantly investigating antebrachial characteristics (see section Osteology) amongst them four papers by Struthers from 1871-1889 (B. physalus, B.

L i t e r a t u r e r e v i e w
301
mysticetus, M. novaeangliae, B. musculus) and one paper by Kükenthal in 1922 (B. mysticetus) referring to antebrachial musculature. Publications of odontocete shoulder and forelimb musculature are much more numerous. One of the earliest records was published by Rapp in 1837 and another a
few years later by Stannius in 1849 (P. phocoena). Murie published several papers, two in 1870 (L. albirostris, G. griseus) and one in 1873 (G. melas) that also include detailed illustrations. However, descriptions of the shoulder and forelimb anatomy in these papers are either not detailed enough or contradictory (e.g. text not congruent with figures), and thus unambiguous identification of some muscles is often difficult or impossible (see also comments by Strickler, 1978). Publications in the earlier part of the 20th century include Sterling, 1910 (P. phocoena), comprehensive papers by Schulte and Smith, 1918 (K. breviceps) and Howell, 1927, 1930b (N. phocaenoides, M. monoceros), and a paper by Smirnowsky, 1928 (T. truncatus). Inexplicably, some of the later papers appear to be less coherent, e.g. in Kleinenberg et al., 1969 (D. leucas). Descriptions in this paper are unspecific, and some captions do not match figure labels, i.e. fig. 36 (diagram of wrist and digit arrangements) is clearly back to front, and some parts in fig. 34 (myology of the shoulder) most likely have similar issues (captions do not match either figure labels or text in the article). Similar issues are also apparent in a paper published by Pilleri et al. 1976 (P. gangetica, D. delphis), apart from an obvious misidentification of the supraspinatus with the deltoideus. The same misidentification is also notable in Purves and Pilleri, 1978 (P. crassidens). Similarly, Klima et al., 1980 (I. geoffrensis, T. truncatus) complicate correlation of positional information by the use of an unconventional
position of the forelimb (retracted rather than protracted) and misidentification of osteological landmarks (see also Chapter 4 Functional anatomy: Functional aspects of flipper shape, section Inia geoffrensis). Originally published in Russian, the paper by Sokolov and Rodionov, 1974 (P. phocoena, D. delphis, T. truncatus) is also available in English translation (cited here). Although the translation is a little coarse, the descriptions and figures in this paper are fairly clear and consistent (for congruence in nomenclature see also comments by Strickler, 1978). Regrettably, there seems to be no translation of the Chinese paper by

L i t e r a t u r e r e v i e w
302
Weijuan, 1981 (L. vexillifer), however, muscule names are written in Latin letters and figures are relatively clear (fig. 8b I. geoffrensis?). The predominantly osteological paper by Benke, 1993 also including descriptions of shoulder musculature (T. truncatus, B. borealis) has already been mentioned above.
There are also a few accounts detailing antebrachial musculature in odontocetes: Struthers, 1873 (H. ampullatus), Anderson, 1878 (P. gangetica), Turner, 1885 and Hanke, 1915 (M. bidens), Pouchet and Beauregard, 1889 also including some shoulder musculature (P. macrocephalus). Cooper et al., 2007b related antebrachial musculature of odontocetes and mysticetes to taxonomic distribution.
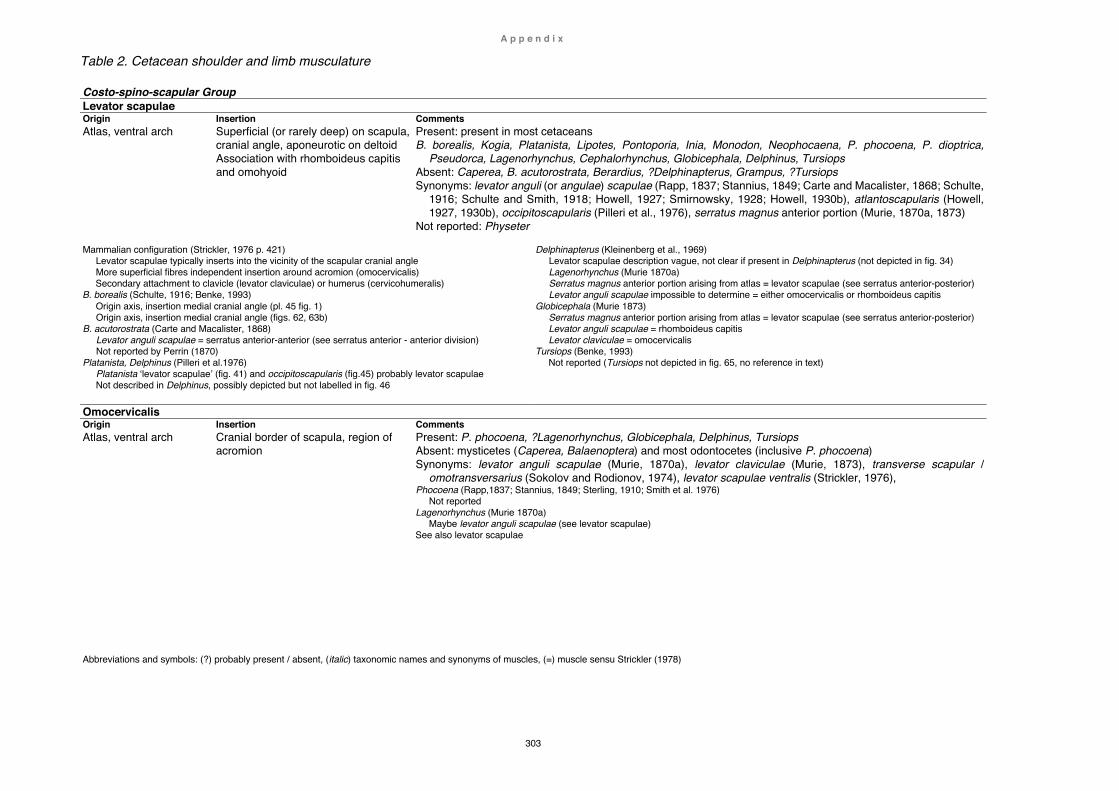
A p p e n d i x
303
Table 2. Cetacean shoulder and limb musculature
Costo-spino-scapular Group Levator scapulae Origin Insertion Comments Atlas, ventral arch Superficial (or rarely deep) on scapula,
cranial angle, aponeurotic on deltoid Association with rhomboideus capitis and omohyoid
Present: present in most cetaceans B. borealis, Kogia, Platanista, Lipotes, Pontoporia, Inia, Monodon, Neophocaena, P. phocoena, P. dioptrica,
Pseudorca, Lagenorhynchus, Cephalorhynchus, Globicephala, Delphinus, Tursiops Absent: Caperea, B. acutorostrata, Berardius, ?Delphinapterus, Grampus, ?Tursiops Synonyms: levator anguli (or angulae) scapulae (Rapp, 1837; Stannius, 1849; Carte and Macalister, 1868; Schulte,
1916; Schulte and Smith, 1918; Howell, 1927; Smirnowsky, 1928; Howell, 1930b), atlantoscapularis (Howell, 1927, 1930b), occipitoscapularis (Pilleri et al., 1976), serratus magnus anterior portion (Murie, 1870a, 1873)
Not reported: Physeter
Mammalian configuration (Strickler, 1976 p. 421) Levator scapulae typically inserts into the vicinity of the scapular cranial angle More superficial fibres independent insertion around acromion (omocervicalis) Secondary attachment to clavicle (levator claviculae) or humerus (cervicohumeralis)
B. borealis (Schulte, 1916; Benke, 1993) Origin axis, insertion medial cranial angle (pl. 45 fig. 1) Origin axis, insertion medial cranial angle (figs. 62, 63b)
B. acutorostrata (Carte and Macalister, 1868) Levator anguli scapulae = serratus anterior-anterior (see serratus anterior - anterior division) Not reported by Perrin (1870)
Platanista, Delphinus (Pilleri et al.1976) Platanista ‘levator scapulae’ (fig. 41) and occipitoscapularis (fig.45) probably levator scapulae Not described in Delphinus, possibly depicted but not labelled in fig. 46
Delphinapterus (Kleinenberg et al., 1969) Levator scapulae description vague, not clear if present in Delphinapterus (not depicted in fig. 34) Lagenorhynchus (Murie 1870a) Serratus magnus anterior portion arising from atlas = levator scapulae (see serratus anterior-posterior) Levator anguli scapulae impossible to determine = either omocervicalis or rhomboideus capitis
Globicephala (Murie 1873) Serratus magnus anterior portion arising from atlas = levator scapulae (see serratus anterior-posterior) Levator anguli scapulae = rhomboideus capitis Levator claviculae = omocervicalis
Tursiops (Benke, 1993) Not reported (Tursiops not depicted in fig. 65, no reference in text)
Omocervicalis Origin Insertion Comments Atlas, ventral arch Cranial border of scapula, region of
acromion
Present: P. phocoena, ?Lagenorhynchus, Globicephala, Delphinus, Tursiops Absent: mysticetes (Caperea, Balaenoptera) and most odontocetes (inclusive P. phocoena) Synonyms: levator anguli scapulae (Murie, 1870a), levator claviculae (Murie, 1873), transverse scapular /
omotransversarius (Sokolov and Rodionov, 1974), levator scapulae ventralis (Strickler, 1976), Phocoena (Rapp,1837; Stannius, 1849; Sterling, 1910; Smith et al. 1976)
Not reported Lagenorhynchus (Murie 1870a)
Maybe levator anguli scapulae (see levator scapulae) See also levator scapulae
Abbreviations and symbols: (?) probably present / absent, (italic) taxonomic names and synonyms of muscles, (=) muscle sensu Strickler (1978)

A p p e n d i x
304
Serratus anterior - anterior division Origin Insertion Comments Vertebra or ribs, C7 or rib 1
Deep on scapula, cranial angle Present: Caperea, B. acutorostrata, Lipotes, Pontoporia, ?Delphinapterus, P. phocoena, Cephalorhynchus, Grampus, Delphinus, Tursiops
Absent: B. borealis, Berardius, Monodon, Neophocaena, P. phocoena, P. dioptrica, ?Lagenorhynchus Synonyms: levator anguli scapulae (Carte and Macalister, 1868), serratus magnus posterior slip first digitation
(Murie, 1870a), serratus / serratus major 1st digitation (Sokolov and Rodionov, 1974), serratus ventralis cervicis (Smith et al., 1976 fig. 3; Benke, 1993 fig. 65), serratus anterior cervicis (Benke, 1993 fig. 65), serratus anterior anterior? (Weijuan, 1981 fig. 6)
B. borealis, Tursiops (Benke, 1993)
Serratus ventralis cervicis (Tursiops fig. 65, no reference in text) Not reported in B. borealis by Schulte (1916) or Benke (1993)
B. acutorostrata (Carte and Macalister, 1868) Levator anguli scapulae = serratus anterior-anterior, origin cervical vertebra C7 Not reported by Perrin (1870)
Lipotes (Weijuan, 1981) Serratus anterior anterior? (fig. 6)
Delphinapterus (Kleinenberg et al., 1969) Possibly ‘rsc’, not annotated (fig. 34)
P. phocoena, Delphinus, Tursiops (Sokolov and Rodionov 1974) Serratus major 1st digitation arising from dorsal part of rib 1 (Delphinus; Tursiops fig. 4 serratus) Serratus major 1st digitation absent (P. phocoena)
P. phocoena (Smith et al., 1976) Serratus ventralis cervicis, origin cervical vertebra C7 (fig. 3)
Lagenorhynchus (Murie, 1870a) Maybe serratus magnus posterior slip, first digitation arising from ribs
Serratus anterior - posterior division Origin Insertion Comments Ribs 2,3,4 Deep on scapula, cartilage vertebral
border Present: present in all cetaceans Synonyms: serratus magnus (Howell, 1927, 1930b; Rapp, 1837; Carte and Macalister, 1868; Murie, 1870a, 1873;
Perrin, 1870), serratus anticus major (Stannius, 1849; Murie, 1873), depressor anguli scapulae (Carte and Macalister, 1868), serratus anterior (Sterling, 1910; Smith et al., 1976; Klima et al., 1980; Weijuan, 1981), serratus anticus (Schulte, 1916; Schulte and Smith, 1918; Howell, 1927), serratus ventralis (Smirnowsky, 1928), serratus / serratus major 2nd-4th digitation (Sokolov and Rodionov, 1974), serratus anterior posterior? (Weijuan, 1981 fig. 6) , serratus and serratus anterior (Benke, 1993 figs. 62+63a,b+65)
Not reported: Physeter, Platanista
B. borealis, Tursiops (Benke, 1993) Serratus origin ribs 4-11, insertion two heads on vertebral scapular border (B. borealis figs. 62 + 63a,b
and text) Serratus anterior (Tursiops fig. 65, no reference in text)
Lipotes (Weijuan, 1981) Serratus anterior posterior? (fig. 6)
Delphinapterus (Kleinenberg et al., 1969) Possibly ‘cr’ annotated as ‘Cerratus’ (fig. 34)
P. phocoena, Delphinus, Tursiops (Sokolov and Rodionov, 1974) Serratus major 2nd-4th digitation arising from central part of ribs 1-4 or 5 (P. phocoena, Delphinus;
Tursiops fig. 4) Lagenorhynchus (Murie, 1870a)
Serratus magnus posterior slip, all or digitation 2-3 arising from ribs Globicephala (Murie 1873)
Serratus magnus = levator scapulae (atlas) + serratus anterior-posterior (ribs 2-3) + epaxial iliocostalis (caudad of rib 4)

A p p e n d i x
305
Rhomboideus capitis Origin Insertion Comments Skull, temporal crest Superficial on scapula, cranial angle Present: Kogia, Platanista, Lipotes, Pontoporia, Inia, ?Delphinapterus, Pseudorca, Grampus, Globicephala,
Delphinus, Tursiops Absent: Caperea B. borealis, B. acutorostrata, Berardius, Monodon, Neophocaena, P. phocoena, P. dioptrica,
?Lagenorhynchus, Cephalorhynchus Synonyms: levator anguli scapulae (Murie, 1870a, 1873), trapezius (Murie, 1870b; 1873; Purves and Pilleri, 1978,
Pilleri et al., 1976 figs. 43-44), occipitoscapularis (Howell, 1927), trapezius capitis (Smirnowsky, 1928), cranial / cephalic trapezius (Sokolov and Rodionov, 1974 fig. 3), rhomboidei (Klima et al., 1980 figs. 12-13), trapezius anterior? (Weijuan, 1981 figs. 5+7)
Platanista and Delphinus (Pilleri et al. 1976)
Platanista: trapezius (text and fig. 43) = rhomboideus capitis, or alternatively absent (see rhomboideus vertebralis)
Delphinus: trapezius (text and fig. 44) = rhomboideus capitis Lipotes (Weijuan, 1981)
Trapezius anterior? (figs. 5+7) Inia (Klima et al., 1980)
Rhomboidei (figs. 12-13 not labelled) = rhomboideus capitis, or alternatively absent (see rhomboideus vertebralis)
Delphinapterus (Kleinenberg et al., 1969) Rhomboideus capitis description vague, ‘rmc’ not annotated (fig. 34)
Neophocaena (Howell 1927) Occipitoscapular (absent in Neophocaena), rhomboideus capitis used in later publications
Lagenorhynchus (Murie 1870a) Maybe levator anguli scapulae (see levator scapulae)
Grampus (Murie 1870b) Trapezius or rhomboideus capitis, confirmed by this study
Globicephala (Murie 1873) Levator anguli scapulae (see levator scapulae)
Tursiops (Smirnovsky, 1928; Sokolov and Rodionov, 1976) Reported as trapezius capitis by Smirnowsky (abb. 1), and cephalic or cranial trapezius by Sokolov and
Rodionov (fig. 3) Not described or depicted by Benke (1993 fig. 65)

A p p e n d i x
306
Rhomboideus vertebralis Origin Insertion Comments Vertebrae, aponeurotic on epaxial muscles at level of 1st to 6th thoracic vertebrae
Not specified Present: present in all cetaceans Single division: Caperea, B. borealis, B. acutorostrata, Berardius, Monodon, Neophocaena, P. phocoena,
Pseudorca, ?Lagenorhynchus, ?Cephalorhynchus, Grampus, Globicephala, Delphinus Two divisions (profundus and superficialis): Kogia, ?Platanista, Lipotes, Pontoporia, ?Inia, Delphinapterus, P.
phocoena, P. dioptrica, Grampus, Delphinus, Tursiops Not reported: Physeter Synonyms (single division): rhomboideus (Carte and Macalister, 1868; Murie, 1873; Sterling, 1910; Schulte, 1916;
Howell, 1927, 1930b; Benke, 1993), rhomboideus scapulae (Murie, 1870a), rhomboideus proprius (Murie, 1870b), rhomboideus / rhomboideus posterior (Pilleri et al., 1976 fig. 43, mislabelled fig. 46), rhomboideus thoracis (Smith et al., 1976; Benke, 1993), rhomboideus anterior (Purves and Pilleri, 1978), trapezius (Klima et al., 1980 figs. 12-13)
Synonyms: (profundus and superficialis): rhomboideus and trapezius dorsi (Smirnowsky, 1928), rhomboideus inferior and superior (Rapp, 1837; Stannius, 1849), rhomboideus vertebralis posterior and rhomboideus vertebralis (Kleinenberg et al., 1969 fig. 34), rhomboid and dorsal trapezius (Sokolov and Rodionov, 1974 fig. 4), rhomboideus and trapezius posterior? (Weijuan, 1981 figs. 5+7)
Not reported: Physeter
Platanista and Delphinus (Pilleri et al. 1976) Platanista: rhomboideus (text and fig. 43) = rhomboideus vertebralis (single division) Alternatively, rhomboideus + trapezius = rhomboideus vertebralis profundus + superficialis
(rhomboideus capitis absent) Delphinus: rhomboideus posterior (text) = rhomboideus vertebralis (single division) Mislabelled as ‘longissimus capitis’ (fig. 46)
Inia (Klima et al., 1980) Trapezius (figs. 12-13) = rhomboideus vertebralis (single division) Alternatively, trapezius + rhomboidei = rhomboideus vertebralis profundus + superficialis (rhomboideus
capitis absent)
Phocoena (Rapp, 1837; Stannius, 1849; Murie, 1873; Sterling, 1910; Sokolov and Rodionov 1974; Smith et al., 1976) Reported as single division by Murie, Sterling, and Smith et al. Reported as two divisions by Rapp, Stannius and Sokolov and Rodionov
Lagenorhynchus (Murie 1870a) Rhomboideus scapulae slip diverting craniad is a reference to the afore mentioned levator anguli scapulae
(see levator scapulae) Murie (1873) confirms ‘there is but a single rhomboideus’ in Lagenorhynchus
Grampus (Murie, 1870b) Rhomboideus proprius not clearly stated as single division, it was found divided in this study
Rhomboideus vertebralis superficialis Vertebrae, aponeurotic on epaxial muscles at level of 1st to 2nd thoracic vertebrae
Superficial on scapula, anterior 1/3 vertebral border
Present: Kogia, Delphinus, Tursiops, P. phocoena, Pontoporia, Kogia Lipotes (Weijuan, 1981)
Trapezius posterior? (figs. 5+7) P. phocoena, Delphinus, Tursiops (Sokolov and Rodionov 1974)
Dorsal trapezius (Tursiops fig. 4), referred to by Strickler (1978) as ‘spinotrapezius’
Rhomboideus vertebralis profundus Vertebrae, aponeurotic on epaxial muscles at level of 2nd to 6th thoracic vertebrae
Superficial on scapula, posterior 2/3 vertebral border
Present: Delphinus, Tursiops, P. phocoena, Pontoporia, Kogia Lipotes (Weijuan, 1981)
Rhomboideus (figs. 5+7) P. phocoena, Delphinus, Tursiops (Sokolov and Rodionov 1974)
Rhomboideus (Tursiops fig. 4)

A p p e n d i x
307
Latissimus-subscapular Group Latissimus dorsi Origin Insertion Comments Ribs 5 to 9 Aponeurotic on inter-costal and epaxial musculature Association with teres major
Humerus, midpoint posterior surface Association with teres major
Present: present in all cetaceans Synonym: grand dorsal (Pouchet and Beauregard, 1889) Cetacean configuration:
Insertion in common with teres major B. borealis (Schulte, 1916)
Origin ribs 7 to 9 B. acutorostrata (Carte, 1868)
Origin from dorsal and anterior lumbar vertebrae Monodon (Howell 1930b)
Origin from vesicular fascia
Teres major Origin Insertion Comments Scapula, caudal border or posterior lateral surface Association with latissimus dorsi
Humerus, midpoint posterior surface Association latissimus dorsi
Present: present in all cetaceans Synonyms: teres (Murie, 1873; Schulte, 1916; Klima et al., 1980), muscle rond (Pouchet and Beauregard, 1889) Not reported: Lagenorhynchus Cetacean configuration:
Origin in odontocetes variable, either caudal border and adjacent deep surface or posterior lateral scapular surface Insertion in common with latissimus dorsi
P. phocoena (Rapp, 1837, Stannius, 1849, Smith et al., 1976) Origin confined to axillary border of scapula
P. phocoena (Sterling, 1910, Sokolov and Rodionov, 1974 + Tursiops, Delphinus) Origin caudal angle and posterior lateral scapular surface
Globicephala (Murie, 1873) Murie regarded single teres major incorrectly as a fusion of teres major with teres minor (see Strickler, 1978)
Subscapularis Origin Insertion Comments Scapula, entire medial surface and tendinous sheet between cranial angle and coracoid
Humerus, lesser or proximal tubercle Present: present in all cetaceans Synonym: sous-scapulaire (Pouchet and Beauregard, 1889) Not reported: Platanista, Lagenorhynchus
Deltoid Group Deltoideus Origin Insertion Comments Scapula, anterior lateral surface of acromion, and aponeurotic on fascia of infraspinatus
Humerus, deltoid tuberosity and/or lateral and anterior aspect of humeral diaphysis
Present: present in all cetaceans Synonyms: deltoïde (Pouchet and Beauregard, 1889) Not reported: Lagenorhynchus Caperea (this srtudy)
Unusual secondary humeral head across cranial cubital joint (see Chapter 2 Caperea) Platanista and Delphinus (Pilleri et al. 1976)
Deltoideus consistently mislabelled as ‘supraspinatus’ and vice versa (figs. 41-46) Delphinapterus (Kleinenberg et al., 1969)
Unusual insertion onto posterior lateral and medial border of humerus, maybe inaccurate (fig. 34) Pseudorca (Purves and Pilleri, 1978)
Deltoideus mislabelled as ‘supraspinatus’ and vice versa (fig. 9)

A p p e n d i x
308
Teres minor / subdeltoideus Origin Insertion Comments Scapula, either posterior border or lateral anterior aspect
Humerus, either posterior aspect of proximal tubercle / posterior humeral border or lateral aspect of greater tubercle
Present: usually absent in cetaceans, reported as teres minor or subdeltoideus (see also Strickler, 1978) Teres minor (as described by Smith et al., 1976, not figured): P. phocoena, Lipotes, Inia Subdeltoideus (as described by Schulte and Smith 1918): B. borealis, Kogia Absent: Caperea, B. acutorostrata, Berardius, Pontoporia, Monodon, ?Delphinapterus, Neophocaena,
P. phocoena, P. dioptrica, Lagenorhynchus, Cephalorhynchus, Grampus, Globicephala, Delphinus, Tursiops Not reported: Physeter, Platanista, Pseudorca
Subdeltoideus B. borealis (Schulte, 1916)
Circumflex innervation, origin anterior scapular neck, insertion lateral aspect of greater humeral tubercle (Schulte, 1916 pl. 46 - fig. 1)
B. borealis (Benke, 1993) ‘Supraspinatus’ origin between deltoideus and infraspinatus, insertion lateral aspect of greater humeral
tubercle (fig. 63a-b, not labelled in fig. 61) B. acutorostrata (Carte and Macalister, 1868)
Not mentioned or depicted in detailed figures (e.g. pl. 5-figs. 2,3) most likely not present Not reported in somewhat less detailed account by Perrin (1870)
Kogia (Schulte and Smith, 1918) Circumflex innervation, origin between deltoideus and infraspinatus, insertion lateral aspect of greater
humeral tubercle (fig. 7) Findings questioned by Howell (1930b), maybe more developed division of the deltoideus or slip of
infraspinatus similar to Monodon Berardius (this study)
Deltoideus in one specimen deeply incised with superficial arrangement similar to Kogia (Schulte and Smith, 1918)
Monodon (Howell, 1930b) Deltoideus and infraspinatus unusually complex, innervation not conclusive for distinction between
deltoideus and subdeltoideus Neophocaena (Howell, 1927)
Reported as absent
Teres minor Lipotes (Weijuan, 1981)
Origin caudal scapular border, insertion proximal aspect of humerus (fig. 8a) Inia (Klima et al., 1980) Origin caudal scapular border, insertion posterior medial humeral border (fig. 7) Delphinapterus (Kleinenberg et al., 1969)
‘Teres minor’ not mentioned in text ‘tmi’ small muscle with origin from medial and ventral anterior scapular area, insertion onto proximal
anterior humerus, more congruent with coracobrachialis (fig. 34) P. phocoena (Rapp, 1837)
Teres minor not defined, reported as absent P. phocoena, Delphinus, Tursiops (Sokolov and Rodionov 1974)
Teres minor not defined, reported as absent in all three species P. phocoena, (Smith et al., 1976)
Origin caudal scapular border, insertion lateral and posterior aspect of humeral tubercle, not depicted (fig. 3)
Globicephala (Murie, 1873) Single teres in Globicephala, Grampus, Lagenorhynchus = teres major Murie’s theory of single teres as composite of teres major and teres minor not supported by Strickler (1978)
Tursiops (Benke, 1993) ‘Triceps’ (fig. 65) rather large muscle in arrangement similar to teres minor in Inia (Klima et al., 1980) Alternatively, humeral and scapular head of triceps (origin not depicted in fig. 65)

A p p e n d i x
309
Triceps Group Triceps Origin Insertion Comments Scapular head, posterior scapular neck above glenoid border Humeral head, posterior border of shaft lateral of teres major/ latissimus dorsi insertion
Olecranon Present: variations of one or two humeral heads and/or one scapular head present in almost all cetaceans 3-headed (two humeral, one scapular): B. acutorostrata 2-headed (humeral, scapular): Caperea, B. borealis, Kogia, Berardius, Grampus, Globicephala, Delphinus,
Tursiops 1-headed (humeral): Pontoporia, Monodon, Neophocaena, P. phocoena, P. dioptrica 1-headed (scapular): P. phocoena, Pseudorca, Lagenorhynchus, Cephalorhynchus, Grampus Absent: Lipotes Not reported: Physeter, Platanista, Inia, Delphinapterus
Cetacean configuration: B. acutorostrata long head described by Carte and Macalister (1868) as irregular mass (fleshy?) and
humeral heads as fibrous Caperea, B. borealis both scapular and humeral head reported as fleshy More basal odontocetes humeral and scapular head fleshy (Kogia, Berardius) Less basal and more crownward odontocetes with more or less fleshy scapular head and/or absent or
vestigial, fibrous humeral head (Pontoporia, Monodon, Neophocaena, Cephalorhynchus, Grampus, Globicephala, Delphinus)
Mysticetes Caperea (this study)
Humeral (external) and scapular head B. borealis (Schulte, 1916)
External head (humeral) and long head (scapular) reported as fleshy, third head absent B. borealis (Benke, 1993)
Second caput (humeral) and caput longum (scapular), not stated if fleshy or fibrous B. acutorostrata (Carte and Macalister, 1868)
Short or inner- and external head (humeral), long head (scapular) Triceps not described, only external- and long head depicted by Perrin (1870 figs. 1-2)
Odontocetes Lipotes (Weijuan, 1981)
Triceps absent (abstract) Pontoporia (Strickler, 1978)
Humeral head origin posterior humeral shaft and somewhat unusual insertion onto distal posterior border of ulna instead of olecranon
Neophocaena (Howell, 1927) Humeral head fibrous, vestigial
P. phocoena, Delphinus, Tursiops (Sokolov and Rodionov 1974) Scapular head vestigial with profusion of connective tissue, humeral head absent in all three species Somewhat unusual insertion onto distal posterior border of humerus in Delphinus
P. phocoena (Stannius, 1849) Humeral head, few fleshy fibres often absent, scapular head not reported Single vestigial humeral head with few muscle fibres also reported by Rapp (1837) and Sterling (1910) Vestigial scapular head but absent humeral head reported by Sokolov and Rodionov (1976)
Pseudorca (Purves and Pilleri, 1978) Triceps not described, scapular head only depicted (fig. 8, not labelled)
Globicephala, P. phocoena, Lagenorhynchus, Grampus (Murie, 1873) Scapular and humeral head diminutive, humeral head more tendinous Scapular head small but not vestigial, humeral head vestigial (this study) Sterling (1910) pointed out insertion onto digit 5 (Murie, 1873 pl. 37, fig. 70) incongruent with triceps Single scapular head cited by Murie for P. phocoena, Lagenorhynchus, Grampus Additional vestigial humeral head in Grampus observed (this study)
Tursiops (Benke, 1993) Triceps (fig. 65) possibly humeral and scapular head (origin not depicted, no reference in text) Alternatively, ‘triceps’ is a rather large muscle in arrangement similar to teres minor in Inia (Klima et al.,
1980)
Suprascapular Group Supraspinatus Origin Insertion Comments Scapula, cranial angle, supraspinous fossa and tendinous sheet between acromion and cranial border
Humerus, cranial aspect of greater or proximal tuberosity
Present: present in all cetaceans Synonym: sus-épieux (Pouchet and Beauregard, 1889) Not reported: reported by all cited authors Cetacean configuration:
Extent of supraspinous fossa variable, particularly in species with very small fossa rugosity at cranial angle serves as strong functional point of insertion
Size of supraspinatus correlated to length of acromion and space between acromion and coracoid process Platanista and Delphinus (Pilleri et al. 1976)
Deltoideus consistently mislabelled as ‘supraspinatus’ and vice versa (figs. 41-46) Pseudorca (Purves and Pilleri, 1978)
Deltoideus mislabelled as ‘supraspinatus’ and vice versa (fig. 9)

A p p e n d i x
310
Infraspinatus Origin Insertion Comments Scapula, lateral surface between deltoideus and infraspinatus
Humerus, lateral mid- shaft and/or infraspinous fossa
Present: present in all cetaceans Synonym: Sous-épineux (Pouchet and Beauregard, 1889) Not reported: Lagenorhynchus Cetacean configuration:
Largely covered anteriorly by deltoideus and sometimes posteriorly by teres major Monodon (Howell 1930b)
Unusually divided tendon near insertion point on humerus
Pectoral Group Pectoralis major Origin Insertion Comments Sternum, extending to ventral midline
Deep on humerus or connective tissue of flipper and axilla
Present in all cetaceans Synonyms: pectoralis (Schulte, 1916; Howell, 1927; Weijuan, 1981 fig. 5; Benke, 1993), ectopectoral clavicular,
sternal and abdominal part (Schulte and Smith, 1918), pectoralis superficialis (Howell, 1930b), pectorales (Kleinenberg et al., 1969; Klima et al., 1980), pectoralis including part of panniculus (Pilleri et al., 1976 figs. 47-48; Purves and Pilleri, 1978 fig. 9)
Not reported: Physeter
Cetacean configuration: There is widespread confusion in the literature surrounding definition and identification of the pectoral
muscles and the panniculus carnosus. As a result, Strickler (1978) proposed following definitions to standardise nomenclature and classification of these muscles:
Pectoralis major: superficial to pectoralis minor and abdominalis, origin broad over the sternum and adjacent costal cartilages extending to the midline, insertion broad onto the medial distal surface of the humerus and axilla of the flipper
Pectoralis minor: deep to pectoralis major, origin narrow from cranio-lateral sternum (manubrium) in the vicinity (and often including) first and second costal cartilages, insertion onto the tip of the scapular coracoid process.
Pectoralis abdominalis: deep to pectoralis major, origin narrow and caudal to the forelimb with attachment from ribs, costal cartilages and fascia never extending to the midline, insertion narrow and deep onto pectoralis major at the posterior border of the distal humerus.
Panniculus carnosus: superficial to, and often intermingling with pectoral muscles, sometimes difficult to separate from underlying muscles and also inserting broad onto medial surface of humerus and/or connective tissue of axilla and flipper.
In most odontocetes the pectoralis major takes origin over the sternum extending cranio-caudally somewhere between manubrium and sterno-costal articulation of ribs 3 to 5 (e.g. Pontoporia, Inia, Monodon, Neophocaena, P. phocoena, P. dioptrica, Lagenorhynchus, Globicephala, Delphinus, Tursiops). In some species the posterior border of this muscle appears caudally extended over the abdomen to a position more or less craniad of the umbilicus (Kogia). The sternum in mysticetes is very small, and although the caudal border of the pectoralis major may be comparable to that of odontocetes, e.g. B. acutorostrata level of rib 5 (Benke, 1993) or Caperea level of rib 3 (this study), this level by far exceeds the caudal border of the sternum and is therefore quite different to odontocetes.
Kogia (Schulte and Smith, 1918) Ectopectoral (clavicular, sternal and abdominal part) = pectoralis major Additional slip of ectopectoral on the right hand = pectoralis abdominalis
Platanista, Pseudorca, Delphinus (Pilleri et al., 1976; Purves and Pilleri, 1978) The caudal extent of the pectoralis major in these species has been described as exceeding by far that of
any other cetaceans. However, these authors also reported difficulties in distinguishing the pectoralis major from the panniculus, only noting a change in thickness. Pilleri et al. (1976) subsequently defined the pectoralis major as the ‘part that lies between the flippers’. Although Strickler (1978) was not able to resolve the issue, this definition is supported by the findings of Sokolov and Rodionov (1974) who distinguished the pectoralis major from the panniculus. They reported the origin of the pectoralis major in Delphinus and Tursiops as confined to the posterior sternum (judging in Tursiops by their fig. 2b extending between costo-sternal ribs 3-5), an area that is much more in accordance with that reported of most odontocetes.
Apart from the pectoralis major, there also seem to be issues with misidentifications of muscles, including the pectoralis minor. It is thus following proposed here: pectoralis major Delphinus (fig. 40), pectoralis major + part panniculus and/or pectoralis abdominalis Delphinus (fig. 48) and Platanista (fig. 47); pectoralis minor maybe an unlabelled muscle just anterior or mediad of the flipper base, Delphinus (figs. 38, 40, 48), Platanista (figs. 37, 49, 47) and maybe a curios amalgamation of pectoralis minor origin + pectoralis major insertion in Platanista (fig. 49 ‘pectoralis minor’).
Lipotes (Weijuan, 1981) Pectoralis major (fig. 5) appears to be congruent with the general odontocete morphology, but another
‘pectoralis major’ (figs. 6, 7) clearly corresponds to Strickler’s (1978) definition of the pectoralis minor P. phocoena (Rapp, 1837) Rapp vaguely described a divided pectoralis major taking origin over the sternum, maybe anterior larger
division = pectoralis major, posterior smaller division = pectoralis abdominalis? No division recorded by Stannius (1849), Sterling (1910), Sokolov and Rodionov (1974), Smith et al. (1976)

A p p e n d i x
311
Pectoralis minor Origin Insertion Comments Sternum, cranio-lateral on or near 1st and 2nd sterno-costal joint Association with costoscapularis Association with costohumeralis
Scapula, tip of coracoid Present: present in almost all odontocetes (inclusive Globicephala, Tursiops) Absent: absent in mysticetes (Caperea, B. borealis, B. acutorostrata) and Neophocaena, ?Globicephala, ?Tursiops Synonyms: petit pectoral (Pouchet and Beauregard, 1889), costo-coracoid (Murie, 1870a), sterno-scapular (Murie,
1870b), entopectoral (Schulte and Smith, 1918), pectoralis profundus (Howell, 1930b) Not reported: ?Platanista, Pseudorca Platanista (Pilleri et al., 1976)
Pectoralis minor maybe regarded as part of pectoralis major (p.113), description too vague to be conclusive (see pectoralis major) Lipotes (Weijuan, 1981)
‘Pectoralis major’ (figs. 6,7, see pectoralis major above) Delphinapterus (Kleinenberg et al., 1969)
Pectoralis minor not mentioned in text, maybe falsely regarded as part of pectoralis major ‘tmi’ (fig. 34) small muscle with origin and insertion in area of medial anterior shoulder joint more congruent with coracobrachialis ‘cbr’ (fig. 34) with potential origin from tip of coracoid and no insertion onto humerus more congruent with perctoralis minor
Globicephala (Murie, 1873) Pectoralis minor appears to be absent (pl. 37 figs. 70, 71) but was found present in this study
Tursiops (Benke, 1993) Not explicitly mentioned as absent but not depicted in detailed figures (fig. 65) Reported as present by Sokolov and Rodionov (1974)
Costoscapularis Origin Insertion Comments Sternum, cranio-lateral on or near 1st and 2nd sterno-costal joint Association with pectoralis minor
Scapula, tip of coracoid Present: Delphinus, Tursiops Absent: absent in almost all cetaceans (inclusive ?Tursiops) Synonym: serratus anterior (Sokolov and Rodionov, 1974 figs. 2,4) Cetacean configuration: Duplicates pectoralis minor, only present if pectoralis minor is also present Delphinus, Tursiops (Sokolov and Rodionov, 1974) Costoscapular or serratus anterior in Delphinus and Tursiops (figs. 2,4) Pectoralis minor not reported by Benke (1993) nor any other muscle with similar attachments
Costohumeralis Origin Insertion Comments Sternum, cranio-lateral on or near 1st and 2nd sterno-costal joint Association with pectoralis minor
Humerus, medial aspect of proximal tubercle
Present: Phocoena, ?Lagenorhynchus Absent: absent in almost all cetaceans (inclusive ?Phocoena) Synonym: costobrachial (Sokolov and Rodionov, 1974) Cetacean configuration:
Regarded by Sokolov and Rodionov (1976) as homologue to costoscapularis in Phocoena (Rapp 1837), and to subsclavius by Stannius (1849)
Phocoena (Rapp, 1837; Stannius, 1849; Sterling, 1910; Sokolov and Rodionov, 1974) Costohumeralis or costobrachialis reported as attaching to humerus instead of scapula This muscle was not reported or figured by Smith et al. (1976) although they did report a pectoralis minor
Lagenorhynchus in Globicephala-paper (Murie, 1873) Reported as present in Lagenorhynchus but absent in Globicephala Strickler (1978) questioned validity of this statement pointing out possible confusion with the costocoracoid (pectoralis minor) reported
for this species in 1870

A p p e n d i x
312
Pectoralis abdominalis Origin Insertion Comments Ribs, costal cartilages and covering fascia caudad of forelimb, not extending to midline
Humerus, caudal edge Present: Kogia, Pontoporia, ?Globicephala, ?Lagenorhynchus Absent: absent in almost all cetaceans Kogia (Schulte and Smith, 1918) ‘Ectopectoral’ additional slip on right side arising from sheath of rectus at level of ribs 4-5 Globicephala, Lagenorhynchus (Murie, 1873) Possibly ‘pectoralis minor’ over sterno-costal rib 4, deep of pectoralis major in Globicephala (pl. 36 fig. 58) Possibly ‘pectoralis minor’ extending from sterno-costal ribs 4-6 in Lagenorhynchus
Trapezius Group Mastohumeralis Origin Insertion Comments Cranium, exoccipital or adjacent squamosal
Humerus, distad to insertions of supraspinatus and coracobrachialis
Present: in all cetaceans Synonym: mastoideohumeralis (Sterling, 1910), cephalohumeralis (Sterling, 1910), brachiocephalicus
(Smirnowsky, 1928; Sokolov and Rodionov, 1974), cephalohumeralis (Rapp, 1837; Murie, 1873), occipitohumeralis (Sterling, 1910; Stannius, 1849)
Not reported: Physeter, Platanista, Lipotes, Lagenorhynchus Cetacean configuration:
Attachments constant dorsal and superficial of sternomastoideus in cetaceans B. borealis (Schulte, 1916)
Single origin from squamosal but double insertion: superficial head (cleidomastoid) insertion into fascia along pectoralis and deltoideus, and a deep head (mastohumeralis) insertion onto distal aspect of greater humeral tubercle after fusion with the trapezius
B. acutorostrata (Carte, 1868) Origin two heads, one arising from atlas and one from paramastoid process
Platanista, Delphinus (Pilleri et al., 1976) Mastohumeralis not mentioned, figured but not labelled (possibly figs. 38,44-46)
Mastoscapularis Origin Insertion Comments Cranium, paroccipital process of exoccipital
Scapula, fascia covering deltoid in vicinity of acromion
Present: Monodon Absent: absent in all cetaceans except Monodon Pontoporia (Strickler, 1978)
Strickler (1978) erroneously cited Neophocaena (Howell, 1927) instead of Monodon (Howell, 1930b)
Sternomastoideus Origin Insertion Comments Sternum, at level of first sterno-costal joint
Cranium, exoccipital or adjacent squamosal
Present: in all cetaceans Cetacean configuration:
Passes deep of mastohumeralis

A p p e n d i x
313
Trapezius Origin Insertion Comments Neck, dorsal aponeurosis of the cervical region
Humerus, between insertion of deltoid and coracobrachialis
Present: B. borealis Absent: B. acutorostrata and odontocetes Synonym: cephalohumeralis (Schulte, 1916) Cetacean configuration:
Strickler (1978) regarded the trapezius as potentially regularly present in mysticetes but not in odontocetes; his statement was based on the lack of proven accessory nerve innervation in the latter
Howell (1930b) considered the trapezius altogether absent in cetaceans B. borealis (Schulte, 1916)
Cephalohumeralis or trapezius proper origin arising from dorsal aponeurosis of cervical region and fusing with the mastohumeralis near the point of insertion (see mastohumeralis above)
Schulte regarded the trapezius as absent in B. acutorostrata of Carte and Macalister (1868)
Flexor Group Coracobrachialis Origin Insertion Comments Scapula, medial ventral edge of coracoid and adjacent surfaces
Humerus, midportion of median tuberosity base
Present: in all cetaceans Absent: ?Inia, Neophocaena Not reported: Platanista, Lagenorhynchus Inia (Klima et al., 1980)
Although Klima et al. (1980) do not mention explicitly the absence of the coracobrachialis, their detailed depictions (figs. 12,14c) nonetheless likely attest to the absence of this muscle in boto
Delphinapterus (Kleinenberg et al., 1969) ‘tmi’ (fig. 34) small muscle with origin and insertion in area of medial anterior shoulder joint more congruent with coracobrachialis ‘cbr’ (fig. 34) with potential origin from tip of coracoid and no insertion onto humerus more congruent with pectoralis minor
Neophocaena (Howell, 1927) Howell stated ‘there is no muscle attachment to either acromion or coracoid’ which excludes the presence of both pectoralis minor and
coracobrachialis, his statement is supported by detailed depictions (figs. 6,13)
Biceps Origin Insertion Comments Antebrachium, distad of greater tubercle
Elbow, medial in the area of the cubital joint
Present: B. borealis, B. acutorostrata, Monodon Absent: absent in almost all cetaceans Cetacean configuration: Vestigial or absent in cetaceans
Medial ventral cervical Group Omohyoideus Origin Insertion Comments Hyoid bone, greater horn Superficial on scapula, cranial angle
aponeurotic on levator scapulae and underlying muscles
Present: present in mysticetes (Caperea, B. borealis, B. acutorostrata) and Platanista, Lipotes, Pontoporia, Inia, Delphinus
Absent: in most odontocetes Cetacean configuration:
Overlaps and partly surrounds levator scapulae
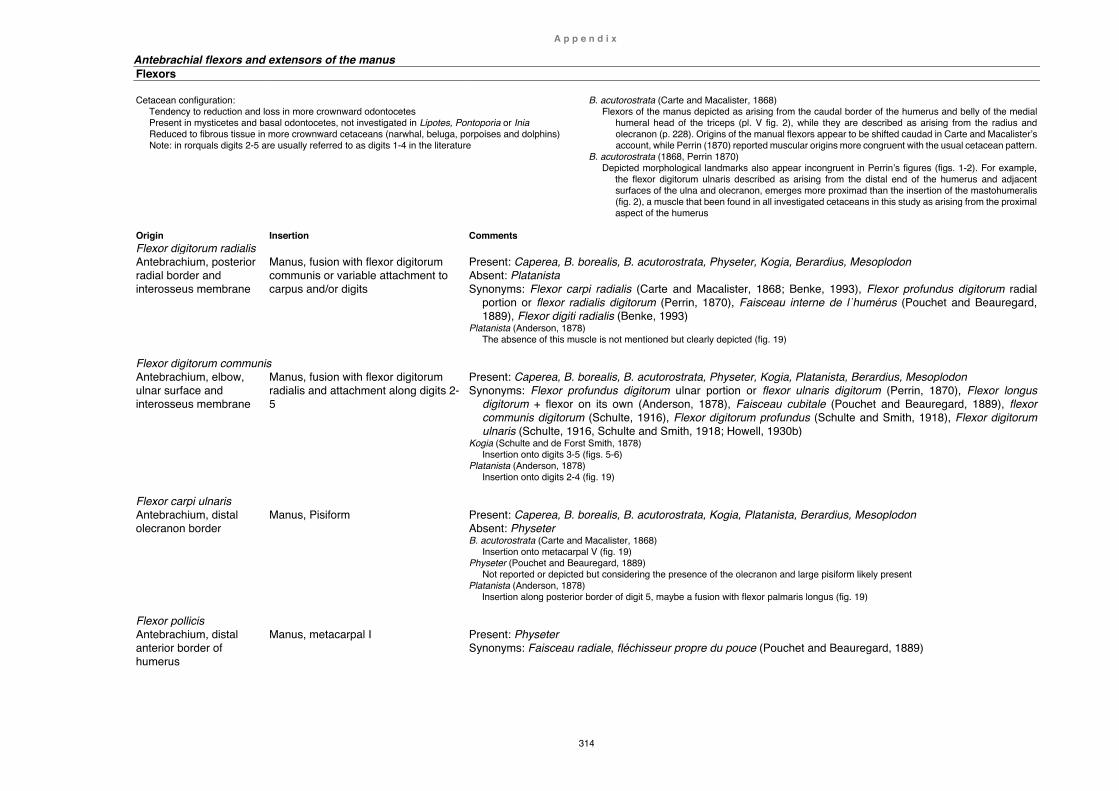
A p p e n d i x
314
Antebrachial flexors and extensors of the manus Flexors Cetacean configuration:
Tendency to reduction and loss in more crownward odontocetes Present in mysticetes and basal odontocetes, not investigated in Lipotes, Pontoporia or Inia Reduced to fibrous tissue in more crownward cetaceans (narwhal, beluga, porpoises and dolphins) Note: in rorquals digits 2-5 are usually referred to as digits 1-4 in the literature
B. acutorostrata (Carte and Macalister, 1868) Flexors of the manus depicted as arising from the caudal border of the humerus and belly of the medial
humeral head of the triceps (pl. V fig. 2), while they are described as arising from the radius and olecranon (p. 228). Origins of the manual flexors appear to be shifted caudad in Carte and Macalister’s account, while Perrin (1870) reported muscular origins more congruent with the usual cetacean pattern.
B. acutorostrata (1868, Perrin 1870) Depicted morphological landmarks also appear incongruent in Perrin’s figures (figs. 1-2). For example,
the flexor digitorum ulnaris described as arising from the distal end of the humerus and adjacent surfaces of the ulna and olecranon, emerges more proximad than the insertion of the mastohumeralis (fig. 2), a muscle that been found in all investigated cetaceans in this study as arising from the proximal aspect of the humerus
Origin Insertion Comments Flexor digitorum radialis Antebrachium, posterior radial border and interosseus membrane
Manus, fusion with flexor digitorum communis or variable attachment to carpus and/or digits
Present: Caperea, B. borealis, B. acutorostrata, Physeter, Kogia, Berardius, Mesoplodon Absent: Platanista Synonyms: Flexor carpi radialis (Carte and Macalister, 1868; Benke, 1993), Flexor profundus digitorum radial
portion or flexor radialis digitorum (Perrin, 1870), Faisceau interne de l˙humérus (Pouchet and Beauregard, 1889), Flexor digiti radialis (Benke, 1993)
Platanista (Anderson, 1878) The absence of this muscle is not mentioned but clearly depicted (fig. 19)
Flexor digitorum communis Antebrachium, elbow, ulnar surface and interosseus membrane
Manus, fusion with flexor digitorum radialis and attachment along digits 2-5
Present: Caperea, B. borealis, B. acutorostrata, Physeter, Kogia, Platanista, Berardius, Mesoplodon Synonyms: Flexor profundus digitorum ulnar portion or flexor ulnaris digitorum (Perrin, 1870), Flexor longus
digitorum + flexor on its own (Anderson, 1878), Faisceau cubitale (Pouchet and Beauregard, 1889), flexor communis digitorum (Schulte, 1916), Flexor digitorum profundus (Schulte and Smith, 1918), Flexor digitorum ulnaris (Schulte, 1916, Schulte and Smith, 1918; Howell, 1930b)
Kogia (Schulte and de Forst Smith, 1878) Insertion onto digits 3-5 (figs. 5-6)
Platanista (Anderson, 1878) Insertion onto digits 2-4 (fig. 19)
Flexor carpi ulnaris Antebrachium, distal olecranon border
Manus, Pisiform Present: Caperea, B. borealis, B. acutorostrata, Kogia, Platanista, Berardius, Mesoplodon Absent: Physeter B. acutorostrata (Carte and Macalister, 1868)
Insertion onto metacarpal V (fig. 19) Physeter (Pouchet and Beauregard, 1889)
Not reported or depicted but considering the presence of the olecranon and large pisiform likely present Platanista (Anderson, 1878)
Insertion along posterior border of digit 5, maybe a fusion with flexor palmaris longus (fig. 19)
Flexor pollicis Antebrachium, distal anterior border of humerus
Manus, metacarpal I Present: Physeter Synonyms: Faisceau radiale, fléchisseur propre du pouce (Pouchet and Beauregard, 1889)

A p p e n d i x
315
Flexor sublimis digitorum Antebrachium, distal humerus
Manus, fusion with tendons of flexors digitorum radialis and communis, and attachment to carpus
Present: B. acutorostrata Absent: B. acutorostrata B. acutorostrata (Carte and Macalister; 1868; Perrin, 1993)
Described and figured by Perrin (fig. 2) but not reported by Carte and Macalister (pl. V fig. 2)
Flexor palmaris longus Antebrachium, elbow, olecranon
Manus, carpals and metacarpal V Present: B. acutorostrata Absent: B. acutorostrata B. acutorostrata (Carte and Macalister; 1868; Perrin 1870)
Described by Carte and Macalister (not depicted in pl. V fig. 2) but reported as absent by Perrin
Extensors Origin Insertion Comments Extensor digitorum communis Antebrachium, elbow and axial borders of radius and ulna, interosseus membrane
Manus, along digits 2-5 Present: Caperea, B. borealis, B. acutorostrata, Physeter, Kogia, Platanista, Berardius, Mesoplodon Synonyms: Extenseur commun des doigts (Pouchet and Beauregard, 1889) Platanista (Anderson, 1878)
Reduced and interwoven with fibrous tissue, insertion along digits 3-5
Extensor carpi ulnaris Antebrachium, elbow, posterior humerus and ulna
Manus, along posterior border of carpus and digit 5
Present: Platanista Platanista (Anderson, 1878)
Insertion along posterior border of digit 5, maybe continuation of triceps or dorsi-epitrochlearis (fig. 19)
Extensor pollicis Antebrachium, distal of deltoid insertion
Proximal digit 1 Present: Physeter Synonyms: Extenseur propre du pouce (Pouchet and Beauregard, 1889)
Interossei Antebrachium, metacarpals
Proximal digits Present: Physeter, Kogia, Berardius, Neophocaena Synonyms: interosseux
Papers referred to by Strickler, 1978 Additional papers (this study) Mysticetes Mysticetes Balaenoptera acutorostrata Carte and Macalister (1868) Balaenoptera acutorostrata Perrin (1870) Balaenoptera borealis Schulte (1916) Balaenoptera borealis Benke (1993) Caperea marginata This study Odontocetes Odontocetes Delphinus delphis Sokolov and Rodionov (1974), Pilleri et al. (1976) Berardius arnuxii This study Globicephala melas Murie (1873) Cephalorhynchus hectori This study Grampus griseus Murie (1870b) Delphinapterus leucas Kleinenberg (1969) Inia geoffrensis Fitzgerald (1970) Thesis Grampus griseus This study Kogia breviceps Schulte and Smith (1918) Inia geoffrensis Klima (1980) Lagenorhynchus albirostris Murie (1870a) †Lipotes vexillifer Weijuan (1981) Monodon monoceros Howell (1930b) Mesoplodon bidens Turner (1885), Hanke (1915) Neophocaena phocaenoides Howell (1927) Phocoena dioptrica This study Platanista gangetica minor Pilleri et al. (1976) Phocoena phocoena Smith et al. (1976), Sterling (1910) Phocoena phocoena Rapp (1837), Stannius (1849), Sokolov and Rodionov (1974) Physeter macrocephalus Pouchet and Beauregard (1889) Pontoporia blainvillei Strickler (1978) Platanista gangetica Anderson (1878) Tursiops truncatus Sokolov and Rodionov (1974), Pilleri et al. (1976) Pseudorca crassidens Purves and Pilleri (1978) Tursiops truncatus Smirnowsky (1928), Benke (1993)

316
Synthesis and conclusion Cetaceans represent a unique lineage derived from semiaquatic, quadruped mammals that returned to an aquatic lifestyle about 50 million years ago. Although the transition from land to sea is well documented by fossils, many morphological characteristics of the forelimb have remained little documented or speculative. Furthermore, investigations of extant cetacean species continue to reveal new insight into the biology and phylogeny of modern and extinct whales. Multiple stranding events on the shores of New Zealand offered therefore a unique opportunity to investigate forelimb morphologies so far undescribed in cetaceans or to complement already published data. Five (out of 15 investigated) cetacean species have been selected in this study and described in detail, also incorporating relevant descriptions of other species in Chapter 4.
The first species, Caperea marginata (Chapter 2) reveals multiple, and previously unknown, traits of the shoulder and forelimb such as two extremely elongated, distinctly shaped scapula types. While fan-shaped shoulder blades are typical in other mysticetes (rorquals), the angular contour found in some pygmy right whales appears to be unique among modern cetaceans. However, the latter shows remarkable similarities with fossil scapulae reported for members of the Cetotheriidae (Piscobalaena nana), a family otherwise believed to have gone extinct about 2.5 my ago. Similarly unique, an enormous cartilaginous olecranon, together with a large axillary skin fold, also offers additional new clues to ecological and behavioural traits not known from other living or extinct baleen whales. Like all beaked whales, Berardius arnuxii (Chapter 3) undertakes some of the most extreme deep diving feats seen in cetaceans and is physically and behaviourally highly adapted to do so, yet this species has retained plesiomorphic flipper characteristics. Incorporating recent publications, a possible explanation of this apparent discrepancy is offered here by way of behavioural traits rather than physical adaptations of the forelimb.

S y n t h e s i s
317
Chapter 4 (Functional Anatomy) presents morphological descriptions of multiple cetacean species, also including detailed myological accounts of one porpoise (Phocoena dioptrica) and two dolphin species (Grampus griseus, Cephalorhynchus hectori). The first part of the discussion focuses on cetacean evolution, particularly on aspects of locomotion and the role of the forelimb during the transition from land to
sea. While the earliest archaeocetes (Indohyus) most likely retained more or less typical Eocene artiodactyl attributes, later protocetids (Maiacetus) show a combination of archaic and new traits. They maybe also utilised a unique swimming style not seen in either older or younger archaeocetes. On land, a somewhat peculiar waddling walk is proposed here rather than the rolling gait of sea lions. Basilosaurids (Dorudon) finally present a lift-based flipper morphology more redolent of modern cetaceans, possibly quite similar to ziphiids (B. arnuxii). Flipper morphology in neocetes experienced further transformation and culminated in the distinctive flipper shapes seen in extant cetaceans today. Comparative morphologies in this study include specimens of both toothed (odontocetes) and baleen whales (mysticetes) encompassing a total of six flipper shapes (paddle, deltoid, fusiform, oval, oar, and falcate). In basilosaurids, mysticetes and basal-most odontocetes (Physeter), digits are more or less linear, and flipper types supported in this manner are paddle, fusiform, oval, and oar-shaped. Digits in less basal odontocetes (Kogia, Ziphiidae, Platanista, †Lipotes, Inia, Monodontidae) show predominantly moderate hyperphalangy, a slight curvature of the second digit, and a moderately splayed digit 5 supporting the trailing edge of the flipper. Flipper types in this category include paddle, deltoid, and fusiform contours. Extreme hyperphalangy, highly appressed, curved preaxial digits and
flattened, splayed postaxial digits are dominant in crownward-most cetaceans (phocoenids, delphinids). Here, digit 5 is strongly angulated and truncated. Broader flipper shapes (e.g. fusiform or oval) are achieved by a greater splay of digits 2-4 (except killer whales). In phocoenids, the flipper contour is generally fusiform (e.g. P. phocoena, P. dioptrica, N. phocaenoides), but can be much more variable in other non-phocoenid delphinids, and ranges from oval (e.g. O. orca, C. hectori), fusiform (M. monoceras, D. leucas) to falcate - the latter the most prevalent shape in the most crownward delphinids (e.g. L. peronii, G. melas, D. delphis).

S y n t h e s i s
318
In the last part of Chapter 4, the analysis of taxonomic distribution of cetacean forelimb muscles show two major groups: a consistent core group (rhomboideus vertebralis, serratus anterior-posterior, latissimus dorsi, teres major, subscapularis, deltoideus, supraspinatus, infraspinatus, pectoralis major, coracobrachialis, mastohumeralis) that is linked to three basic but essential functions (glenohumeral integrity, flipper attitude,
scapula mobility). While this group is more or less strongly developed and (almost) always present, the second muscle group (accessory group) appears in development and occurrence much more varied and inconsistent. Despite ostensible similarities with arrangements seen in terrestrial mammals, the presence and arrangements of muscles in either group are here not regarded as proof of phylogenetic relationship and in many cases eludes explanation altogether. Overall, forelimb morphology of whales and dolphins has undergone profound physical and functional changes in the course of their evolution, and culminated in a high variability of flipper elements and shapes. This process has separated cetacean limb morphology greatly from that of their nearest modern relatives rendering some elements hard to interpret. Despite this, some flipper characteristics appear to be strongly influenced by their evolutionary history and may in the future prove useful in the resolution of phylogenetic affinities. Investigation of modern cetaceans has also revealed formerly unknown characteristics suggestive of particular lifestyles, and thus may contribute to the understanding of cetacean biology and ultimately aid conservation efforts of threatened members in this fascinating clade.

319
Literature Cited Acevedo-Gutiérrez, A. (2009) Group behavior. Encyclopedia of Marine Mammals. 3rd
ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 511-520. Agnarsson, I. and May-Collado, L. J. (2008) The phylogeny of Cetartiodactyla: the
importance of dense taxon sampling, missing data, and the remakable promise of cytochrome b to provide reliable species-level phylogenies. Molecular Phylogenetics and Evolution 48 (3): 964-985.
Aguilar de Soto, N., Johnson, M. P., Madsen, P. T., et al. (2008) Cheetahs of the deep sea: deep foraging sprints in short-finned pilot whales off Tenerife (Canary Islands). Journal of Animal Ecology 77 (5): 936-947.
Aguilar de Soto, N., Madsen, P. T., Tyack, P. L., et al. (2012) No shallow talk: Cryptic strategy in the vocal communication of Blainville's beaked whales. Marine Mammal Science 28 (2): 1-18.
Ahlberg, P. E. (1989) Paired fin skeletons and relationships of the fossil group Porolepiformes (Osteichthyes: Sarcopterygii). Zoological Journal of the Linnean Society 96 (2): 119-166.
Ainley, D. G., Ballard, G. and Dugger, K. M. (2006) Competition among penguins and cetaceans reveals trophic cascades in the western Ross Sea, Antarctica. Ecology and Evolution 87 (8): 2080-2093.
Alroy, J. (1999) The fossil record of North American mammals: evidence for a Paleocene evolutionary radiation. Systematic Biology 48 (1): 107-118.
Anderson, J. (1878) Zoological results of the two expeditions to Western Yunnan in 1868 and 1875 and a monograph of the two cetacean genera, Platanista and Orcella. Anatomical and Zoological Researches, Ed. J. M. D. Anderson (Bernard Quartich; London) 1 (2): 365-951.
Andrews, R. C. (1914) 1. The California gray whale Rhachianectes glaucus Cope. Memoirs of the American Museum of Natural History. Monographs of the Pacific Cetacea 1 (5): 227-287.
Andrews, R. C. (1916) 2. The sei whale. Memoirs of the American Museum of Natural History. Monographs of the Pacific Cetacea 1 (6): 289-388.
Aoki, K., Amano, M., Kubodera, T., et al. (2015) Visual and behavioral evidence indicates active hunting by sperm whales. Marine Ecology Progress Series 523: 233-241.
Argot, C. (2013) Postcranial analysis of a carnivoran-like archaic ungulate: the case of Arctocyon primaevus (Arctocyonidae, Mammalia) from the late Paleocene of France. Journal of Mammalian Evolution 20 (2): 83-114.
Arranz, P., Aguilar de Soto, N., Madsen, P. T., et al. (2011) Following a foraging fish-finder: diel habitat use of Blainville’s beaked whales revealed by echolocation. PLoS ONE 6 (12): 1-10.
Arranz, P., Benoit-Bird, K. J., Southall, B. L., et al. (2018) Risso's dolphins plan foraging dives. Journal of Experimental Biology 221 (4): 1-9.
Au, W. W. L., Pawloski, J. L., Nachtigall, P. E., et al. (1995) Echolocation signals and transmission beam pattern of a false killer whale (Pseudorca crassidens). The Journal of the Acoustical Society of America 98 (1): 51-59.

R e f e r e n c e s
320
Bajpai, S. and Gingerich, P. D. (1998) A new Eocene archaeocete (Mammalia, Cetacea) from India and the time of origin of whales. Proceedings of the National Academy of Sciences 95 (26): 15464-15468.
Bajpai, S. and Thewissen, J. G. M. (1998) Chapter 7: Middle Eocene cetaceans from the Harudi and Subathu formations of India. The emergence of whales: evolutionary patterns in the origin of Cetacea. Advances in Vertebrate Paleobiology, Ed. J. G. M. Thewissen (Plenum Press; New York): 213-233.
Bajpai, S. and Thewissen, J. G. M. (2000) A new, diminutive Eocene whale from Kachchh (Gujarat, India) and its implications for locomotor evolution of cetaceans. Current Science 79 (10): 1478-1482.
Bajpai, S. and Thewissen, J. G. M. (2014) Protocetid cetaceans (Mammalia) from the Eocene of India. Palaeontologia Electronica 17 (3): 1-19.
Bajpai, S., Thewissen, J. G. M. and Conley, R. W. (2011) Cranial anatomy of middle Eocene Remingtonocetus (Cetacea, Mammalia) from Kutch, India. Journal of Paleontology 85 (4): 703-718.
Bajpai, S., Thewissen, J. G. M. and Sahni, A. (2009) The origin and evolution of whales: macroevolution documented on the Indian subcontinent. Journal of Bioscience 34 (5): 673-686.
Baker, A. N. (1985) Chapter 12: Pygmy right whale Caperea marginata (Gray, 1846). Handbook of Marine Mammals. Ed. S. H. Ridgway (Academic Press; London, Orlando, San Diego, New York, Toronto, Montreal, Sydney, Tokyo): 344-354.
Balcomb, K. C. (1985) Chapter 10: Baird's beaked whale Berardius bairdii Stejneger, 1883: Arnoux's beaked whale Berardius arnuxii Duvenroy, 1851. Handbook of Marine Mammals. Ed. S. H. Ridgway: 261-288.
Bannister, J. L. (2009) Baleen whales (Mysticeti). Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 80-89.
Baumann-Pickering, S., Roch, M. A., Brownell, R. L. J., et al. (2014) Spatio-temporal patterns of beaked whale echolocation signals in the North Pacific. PLoS ONE 9 (1): 1-17.
Beatty, B. L. and Rothschild, B. M. (2008) Decompression syndrome and the evolution of deep diving physiology in the Cetacea. Naturwissenschaften 95 (9): 793-801.
Bebej, R. M., Ul-Haq, M., Zalmout, I. S., et al. (2007) Functional interpretation of the neck in Eocene Remingtonocetus from Pakistan (Mammalia, Cetacea, Archaeoceti). Journal of Vertebrate Paleontology 27 (Suppl. 3) Abstract of Papers 67rd. Annual Meeting: 45A.
Bebej, R. M., Zalmout, I. S., El-Aziz, A. A. A., et al. (2015) First remingtonocetid archaeocete (Mammalia, Cetacea) from the middle Eocene of Egypt with implications for biogeography and locomotion in early cetacean evolution. Journal of Paleontology 89 (5): 882-893.
Bebej, R. M., Zalmout, I. S. and Gingerich, P. D. (2012) Morphology and function of the vertebral column in Remingtonocetus domandaensis (Mammalia, Cetacea) from the middle Eocene Domanda Formation of Pakistan. Journal of Mammalian Evolution 19 (2): 77-104.
Beddard, F. E. (1901) Contribution towards the knowledge of the osteology of the pygmy right whale (Neobalaena marginata). Proceedings of the Zoological Society of London 16 (2): 87-108.
Behrmann, G. (2002) Evolution der Wale. Lebensraum Meer, 5th ed. (Centre of Marin Research and Investigations on Cetacea; Bremerhaven) 19: 1-105.

R e f e r e n c e s
321
Bejder, L. and Hall, B. K. (2002) Limbs in whales and limblessness in other vertebrates: mechanisms of evolutionary and developmental transformation and loss. Evolution and Development 4 (6): 445-458.
Benham, W. B. (1902) Notes on the osteology of the short nosed sperm whale. Proceedings of the Zoological Society of London 72 (1): 51-62.
Benke, H. (1993) Chapter 1: Functional and comparative anatomy - Investigations on the osteology and functional morphology of the flipper of whales and dolphins. Investigations on Cetacea, Ed. G. Pilleri (Museum of Natural History Paciano; Paciano, Umbria) 14: 9-252.
Benton, M. J. (1983) Dinosaur success in the Triassic: a noncompetitive ecological model. The Quarterly Review of Biology 58 (1): 29-55.
Benton, M. J. (2015) Vertebrate palaeontology. 4th ed. (John Wiley & Sons; West Sussex, England).
Burne, R. H. (1952) Handbook of R. H. Burne's cetacean dissection. Ed. F. C. Fraser (British Museum, Natural History; London): 1-70.
Chaline, J. (1990) Paleontology of Vertebrates. (Springer; Berlin, Heidelberg): 1-186. Berta, A., Sumich, J. L. and Kovacs, K. M. (2015) Marine mammals: evolutionary
biology. 3rd ed., Eds. K. A. S. Gomez and P. Gonzalez (Academic Press, Elsevier; London, San Diego, Waltham, Oxford): 1-726.
Bianucci, G. (2005) Arimidelphis sorbinii, a new small killer whale-like dolphin from the Pliocene of Marecchia River (central eastern Italy) and a phylogenetic analysis of the Orcininae (Cetacea: Odontoceti). Rivista Italiana di Paleontologia e Stratigrafia 111 (2): 329-344.
Bianucci, G., Di Celma, C., Urbina, M., et al. (2016) New beaked whales from the late Miocene of Peru and evidence for convergent evolution in stem and crown Ziphiidae (Cetacea, Odontoceti). PeerJ 4: 1-55.
Bianucci, G. and Gingerich, P. D. (2011) Aegyptocetus tarfa, n. gen. et sp.(Mammalia, Cetacea), from the middle Eocene of Egypt: clinorhynchy, olfaction, and hearing in a protocetid whale. Journal of Vertebrate Paleontology 31 (6): 1173-1188.
Bianucci, G., Lambert, O. and Post, K. (2007) A high diversity in fossil beaked whales (Mammalia, Odonotoceti, Ziphiidae) recovered by trawling from the sea floor off South Africa. Geodiversitas 29 (4): 561-618.
Bisconti, M. (2012) Comparative osteology and phylogenetic relationships of Miocaperea pulchra, the first fossil pygmy right whale genus and species (Cetacea, Mysticeti, Neobalaenidae). Zoological Journal of the Linnean Society 166: 876-911.
Bisconti, M. (2014) Anatomy of a new cetotheriid genus and species from the Miocene of Herentals, Belgium, and the phylogenetic and palaeobiogeographical relationships of Cetotheriidae s.s. (Mammalia, Cetacea, Mysticeti). Journal of Systematic Palaeontology 13 (5): 377-395.
Blake, R. W. (2004) Fish functional design and swimming performance. Journal of Fish Biology 65 (5): 1193-1222.
Boessenecker, R. W. and Fordyce, R. E. (2014) A new eomysticetid (Mammalia: Cetacea) from the Late Oligocene of New Zealand and a re-evaluation of 'Mauicetus' waitakiensis. Palaeontology 1 (2): 107-140.
Boessenecker, R. W. and Fordyce, R. E. (2015) Anatomy, feeding ecology, and ontogeny of a transitional baleen whale: a new genus and species of Eomysticetidae (Mammalia: Cetacea) from the Oligocene of New Zealand. PeerJ 3: 1-69.

R e f e r e n c e s
322
Botella, H., Blom , H., Dorka, M., et al. (2007) Jaws and teeth of the earliest bony fishes. Nature 448 (7153): 583-586.
Boyden, A. and Gemeroy, D. (1950) The relative position of the Cetacea among the orders of Mammalia as indicated by precipitin tests Zoological Journal of the Linnean Society 35: 145-151.
Buchholtz, E. A. (1998) Chapter 11: Implications of vertebral morphology for locomotor evolution in early cetacea. The emergence of whales: evolutionary patterns in the origin of Cetacea. Advances in Vertebrate Paleobiology, Ed. J. G. M. Thewissen (Plenum Press; New York): 325-350.
Buchholtz, E. A. (2001a) Swimming styles in Jurassic ichthyosaurs. Journal of Vertebrate Paleontology 21 (1): 61-73.
Buchholtz, E. A. (2001b) Vertebral osteology and swimming style in living and fossil whales (Order: Cetacea). Journal of Zoology 253 (2): 175-190.
Buchholtz, E. A. (2011) Vertebral and rib anatomy in Caperea marginata: implications for evolutionary patterning of the mammalian vertebral column. Marine Mammal Science 27 (2): 382-397.
Burne, R. H. (1952) Handbook of R. H. Burne's cetacean dissection. Ed. F. C. Fraser (British Museum, Natural History; London): 1-70.
Butler, P. J. (2001) Diving beyond the limits. Physiology 16 (5): 222-227. Calzada, N. and Aguilar, A. (1996) Flipper development in the Mediterranean striped
dolphin (Stenella coeruleoalba). Anatomical Record 245 (4): 708-714. Campbell, B. (1936) The comparative myology of the forelimb of the hippopotamus,
pig and tapir. American Journal of Anatomy 59 (2): 201-247. Carte, A. and Macalister, A. (1868) On the anatomy of Balaenoptera acutorostrata.
Philosophical Transactions of the Royal Society of London 158: 201-261. Chaline, J. (1990) Paleontology of Vertebrates. (Springer; Berlin, Heidelberg): 1-186. Clementz, M. T., Goswami, A., Gingerich, P. D., et al. (2006) Isotopic records from
early whales and sea cows: contrasting patterns of ecological transition. Journal of Vertebrate Paleontology 26, (2): 355-370.
Clifford, A. B. (2010) The evolution of the unguligrade manus in artiodactyls. Journal of Vertebrate Paleontology 30 (6): 1827-1839.
Constantinescu, G. M., Habel, R. E., Sack, W. O., et al. (1992) Illustrated veterinary anatomical nomenclature. Ed. O. Schaller (Ferdinand Enkel Verlag; Stuttgart): 1-614.
Cooper, L. N. (2004) From Dorudon to finbacks: comparative anatomy and evolution of the baleen whale forelimb. (Department of Biology; Kent State University) PhD thesis: 1-186.
Cooper, L. N. (2009) Forelimb anatomy. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 449-452.
Cooper, L. N., Berta, A., Dawson, S. D., et al. (2007a) Evolution of hyperphalangy and digit reduction in the cetacean manus. Anatomical Record 290 (6): 654-672.
Cooper, L. N. and Dawson, S. D. (2009) The trouble with flippers: a report on the prevalence of digital anomalies in Cetacea. Zoological Journal of the Linnean Society 155: 722-735.
Cooper, L. N., Dawson, S. D., Reidenberger, J. S., et al. (2007b) Neuromuscular anatomy and evolution of the cetacean forelimb. Anatomical Record 290 (9): 1121-1137.
Cooper, L. N., Sedano, N., Johansson, S., et al. (2008) Hydrodynamic performance of the minke whale (Balaenoptera acutorostrata) flipper. Journal of Experimental Biology 211 (12): 1859-1867.

R e f e r e n c e s
323
Cooper, L. N., Thewissen, J. G. M., Bajpai, S., et al. (2012) Postcranial morphology and locomotion of the Eocene raoellid Indohyus (Artiodactyla: Mammalia). Historical Biology 24 (3): 279-310.
Corkeron, P. J. and Connor, R. C. (1999) Why do baleen whales migrate? Marine Mammal Science 15 (4): 1228-1245.
Cozzi, B., Huggenberger, S. and Oelschläger, H. (2016) Chapter 3: Locomotion (including osteology and myology). Anatomy of dolphins. (Elsevier; Amsterdam): 33-89.
Cuvier, G. (1835) Chapter 4: Des ossemens fossiles de narvals et de cetaces voisins des hyperoodons et des cachalots. Recherche sur les ossemens fossiles, où l'on rétablit les caractères de plusieurs animaux dont les révolutions du globe ont détruit les espèces. 4th ed., Ed. E. d'Ocagne (E. d'Ocagne; Paris) 4 (7): 171-248.
D'Vincent, C. G., Nilson, R. M. and Hanna, R. E. (1985) Vocalization and coordinated feeding behavior of the humpback whale in southeastern Alaska. The Scientific Reports of the Whales Research Institute (The Whales Research Institute; Tokio) 36: 41-47.
Daeschler, E. B., Shubin, N. H. and Jenkins, F. A. J. (2006) A Devonian tetrapod-like fish and the evolution of the tetrapod body plan. Nature 440 (7085): 757-763.
Dawson, S. D. (1994) Allometry of cetacean forelimb bones. Journal of Morphology 222 (2): 215-221.
Dawson, S. D. (2003) Patterns of ossification in the manus of the harbor porpoise (Phocoena phocoena): hyperphalangy and delta-shaped bones. Journal of Morphology 258 (2): 200-206.
Dawson, S. M. (1985) Hector's dolphin. The New Zealand Whale & Dolphin Digest: 72-75.
de Carvalho, A. P. M., Lima, J. Y., Azevedo, C. T., et al. (2014) Ossification pattern of estuarine dolphin (Sotalia guianensis) forelimbs, from the coast of the State of Espirito Santo, Brazil. PLoS ONE 10 (5): 1-10.
de Muizon, C. (1990) A new Ziphiidae (Cetacea) from the Early Miocene of Washington State (USA) and phylogenetic analysis of the major groups of odontocetes. Bulletin du Muséum national d'histoire naturelle. Sciences de la terre, paléontologie, géologie, minéralogie 4 (12): 279-326.
de Muizon, C. (1994) Are the squalodonts related to the platanistoids? Proceedings of the San Diego Society of Natural History 29: 135-146.
de Muizon, C. (2009) River dolphins, evolutionary history and affinities. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 979-986.
de Queiroz, K. (2007) Species concepts and species delimitation. Systematic Biology 56 (6): 879–886.
de Stephanis, R., Giménez, J., Esteban, R., et al. (2014) Mobbing-like behavior by pilot whales towards killer whales: a response to resource competition or perceived predation risk? Acta Ethologica 18 (1): 69-78.
del Castillo, D. L., Panebianco, M. V., Negri, M. F., et al. (2014) Morphological analysis of the flippers in the Franciscana dolphin, Pontoporia blainvillei, applying X-ray technique. Anatomical Record 297 (7): 1181-1188.
Deméré, T. A. and Berta, A. (2008) Skull anatomy of the Oligocene toothed mysticete Aetioceus weltoni (Mammalia; Cetacea): implications for mysticete evolution and functional anatomy. Zoological Journal of the Linnean Society 154: 308-352.

R e f e r e n c e s
324
Deméré, T. A., McGowen, M. R., Berta, A., et al. (2008) Morphological and molecular evidence for a stepwise evolutionary transition from teeth to baleen in mysticete whales. Systematic Biology 57 (1): 15-37.
DePalma, A. F. (2008) Origin and comparative anatomy of the pectoral limb. Clinical Orthopaedic Related Research (466): 531-542.
DiGiancamillo, M., Rattegni, G., Podestà, M., et al. (1998) Postnatal ossification of the thoracic limb in striped dolphins (Stenella coeruleoalba) (Meyen, 1833) from the Mediterranean Sea. Canadian Journal of Zoology 76 (7): 1286-1293.
Diogo, R., Abdala, V., Aziz, M. A., et al. (2009) From fish to modern humans – comparative anatomy, homologies and evolution of the pectoral and forelimb musculature. Journal of Anatomy 214: 694-716.
Domning, D. P. (1978) The myology of the Amazonian manatee, Trichechus inunguis (Natterer) (Mammalia: Sirenia): muscles of the trunk, thorax, shoulder and forelimb. Acta Amazonica 8 (2) Suppl. 1: 1-81.
Domning, D. P. (2002) The terrestrial posture of desmostylians. Smithonian Contribution to Paleobiology. Cenozoic Mammals of Land and Sea: Tributes to the Career of Clayton E. Ray 93: 313-356.
Dong, Z. M. (1980) Chinese dinosaur faunas and their stratigraphic position. Journal of Stratigraphy 4 (4): 256-263.
Drachman, D. B. and Sokoloff, L. (1966) The role of movement in embryonic joint development. Developmental Biology 14: 401-420.
Dubost, G. (1975) Le comportement du Chevrotain africain, Hyemoschus aquaticus Ogilby (Artiodactyla, Ruminantia). Sa signification écologique et phylogénétique. Zeitschrift für Tierpsychologie 37 (4): 403-448.
Duvernoy, G. (1851a) Espèce Berardius Arnuxii, Nob. (le Bérardien d'Arnoux). Anales de Sciences naturelles, Zoologie. Mémoire sur les charactères ostéologiques des genres nouveaux ou des espèces nouvelles de cétacés vivants ou fossiles 3 (15): 52-68.
Duvernoy, G. (1851b) Mémoire sur les caractères ostéologiques des genres nouveaux ou des espèces nouvelles de cétacés vivants ou fossiles. Anales de Sciences naturelles, Zoologie 3 (15): 1-384.
Dwight, T. J. (1872) Description of the whale (Balaenoptera musculus Auct.) in the possession of the Society: with remarks on the classification of fin whales. Boston Society of Natural History 2 (2): 203-230.
Eales, N. B. (1954) The manus of the narwhal, Monodon monoceros L. Proceedings of the Zoological Society of London 124 (2): 201-212.
Edel, R. K. and Winn, H. E. (1978) Observations on underwater locomotion and flipper movement of the humpback whale Megaptera novaeangliae. Marine Biology 48: 279-287.
el Adli, J. J., Deméré, T. A. and Boessenecker, R. W. (2014) Herpetocetus morrowi (Cetacea: Mysticeti), a new species of diminutive baleen whale from the Upper Pliocene (Piacenzian) of California, USA, with observations on the evolution and relationships of the Cetotheriidae. Zoological Journal of the Linnean Society 170: 400-466.
Ellis, R. and Mead, J. G. (2017) Beaked whales: a complete guide to their biology and conservation. (John S. Hopkins University Press; Baltimore): 1-194.
English, A. W. M. (1976) Functional anatomy of the hands of fur seals. American Journal of Anatomy 147 (1): 1-18.
English, A. W. M. (1977) Structural correlates of forelimb function in fur seals and sea lions. Journal of Morphology 151 (3): 325-352.

R e f e r e n c e s
325
Erdogmus, S., Guler, M., Eroglu, S., et al. (2014) The importance of the supratrochlear foramen of the humerus in humans: an anatomical study. Medical Science Monitor International Medical Journal of Experimental and Clinical Research 20: 2643-2650.
Eschricht, D. F., Reinhardt, J. and Lilljeborg, W. (1866) Recent memoirs on the Cetacea. Ed. W. H. Flower (Ray Society; London) 26: 1-312.
Esperante, R., Brand, L., Nick, K. E., et al. (2008) Exceptional occurrence of fossil baleen in shallow marine sediments of the Neogene Pisco Formation, Southern Peru. Palaeogeography, Palaeoclimatology, Palaeoecology 257 (3): 344-360.
Fabre, A. C., Goswami, A., Peigné, S., et al. (2014) Morphological integration in the forelimb of musteloid carnivorans. Journal of Anatomy 225 (1): 19-30.
Fahlke, J. M., Gingerich, P. D., Welsh, R. C., et al. (2011) Cranial asymmetry in Eocene archaeocete whales and the evolution of directional hearing in water. Proceedings of the National Academy of Sciences 108 (35): 14545-14548.
Fedak, T. J. and Hall, B. K. (2004) Perspective on hyperphalangy: patterns and processes. Anatomical Society of Great Britain and Ireland 204: 151-163.
Feldkamp, S. D. (1987) Foreflipper propulsion in the California sea lion, Zalophus californianus. Journal of Zoology 212 (1): 43-57.
Fernández-Monescillo, M., Quispe, B. M., Pujos, F., et al. (2018) Functional anatomy of the forelimb of Plesiotypotherium achirense (Mammalia, Notoungulata, Mesotheriidae) and evolutionary insights at the family level. Journal of Mammalian Evolution 25 (2): 197-211.
Fettuccia, D. C., da Silva, V. M. F., Rocha, M. S., et al. (2012) Sternum and appendicular skeleton: morphometric differences between the species of genus Sotalia (Cetacea: Delphinidae) Journal of the Marine Biological Association of the United Kingdom 92 (8): 1657-1662.
Fish, F. E. (1993a) Comparison of swimming kinematics between terrestrial and semiaquatic opossums. Journal of Mammalogy 74 (2): 275-284.
Fish, F. E. (1993b) Influence of hydrodynamic design and propulsive mode on mammalian swimming energetics. Australian Journal of Zoology 42: 79-101.
Fish, F. E. (1994) Association of propulsive swimming mode with behavior in river otters (Lutra canadensis). Journal of Mammalogy 75 (4): 989-997.
Fish, F. E. (1996) Transitions from drag-based to lift-based propulsion in mammalian swimming. American Zoologist 36 (6): 628-641.
Fish, F. E. (1997) Biological designs for enhanced maneuverability: analysis of marine mammal performance. Proceedings of the 10th International Symposium for Unmanned Untethered Submersible Technology (Autonomous Undersea Systems Institute; Lee, New Hampshire): 109-117.
Fish, F. E. (1998a) Chapter 10: Biomechanical perspective on the origin of cetacean flukes. The emergence of whales: evolutionary patterns in the origin of Cetacea. Advances in Vertebrate Paleobiology, Ed. J. G. M. Thewissen (Plenum Press; New York): 303-324.
Fish, F. E. (1998b) Comparative kinematics and hydrodynamics of odontocete cetaceans: morphological and ecological correlates with swimming performance. Journal of Experimental Biology 201 (10): 2867–2877.
Fish, F. E. (2000) Biomechanics and energetics in aquatic and semiaquatic mammals: platypus to whale. Physiological and Biochemical Zoology 73 (6): 683-698.
Fish, F. E. (2002) Balancing requirements for stability and maneuverability in cetaceans. Integrative and Comparative Biology 42 (1): 85–93.

R e f e r e n c e s
326
Fish, F. E. (2004) Structure and mechanics of nonpiscine control surfaces. IEEE Journal of Oceanic Engineering 29 (3): 605-621.
Fish, F. E. (2016) Secondary evolution of aquatic propulsion in higher vertebrates: validation and prospect. Integrative and Comparative Biology 56 (6): 1285-1297.
Fish, F. E. and Battle, J. M. (1995) Hydrodynamic design of the Humpback whale flipper. Journal of Morphology 225 (1): 51-60.
Fish, F. E., Howle, L. E. and Murray, M. M. (2008) Hydrodynamic flow control in marine mammals. Integrative and Comparative Biology 48 (6): 788-800.
Fish, F. E. and Rohr, J. J. (1999) Review of dolphin hydrodynamics and swimming performance. SPAWAR Technical Report 1801 (Space and Naval Warfare System Centre; San Diego) App. A1-E8: 1-137.
Fitzgerald, E. M. G. (2006) A bizarre new toothed mysticete (Cetacea) from Australia and the early evolution of baleen whales. Proceedings of the Royal Society 273 (1604): 2955-2963.
Fitzgerald, E. M. G. (2012) Possible neobalaenid from the Miocene of Australia implies a long evolutionary history for the pygmy right whale Caperea marginata (Cetacea, Mysticeti). Journal of Vertebrate Paleontology 32 (4): 976-980.
Fleischer, G. (1976) Hearing in extinct cetaceans as determined by cochlear structure. Journal of Paleontology 50 (1): 133-152.
Flower, W. H. (1865) 11. Observations upon a fin whale (Physalus antiquorum, Gray) recently stranded in Pevensey Bay. Proceedings of the Zoological Society of London 33 (1): 699-705.
Flower, W. H. (1867) IV. Description of the skeleton of Inia geoffrensis and of the skull of Pontoporia blainvillii, with remarks on the systematic position of these animals in the Order Cetacea. Proceedings of the Zoological Society of London 6 (3): 87-116.
Flower, W. H. (1868) XII. On the osteology of the cachalot or sperm-whale (Physeter macrocephalus). Transactions of the Zoological Society of London 6 (6): 309-372.
Flower, W. H. (1872) On the recent ziphoid whales, with a description of the skeleton of Berardius arnouxi. Transactions of the Zoological Society of London 8 (3): 203-234.
Flower, W. H. (1876a) 2. Remarks upon Dr. von Haast's communication on Ziphius novae-zelandiae. Proceedings of the Zoological Society of London 44 (1): 477-478.
Flower, W. H. (1876b) 4. Remarks upon Dr. von Haast's communication on Mesoplodon floweri. Proceedings of the Zoological Society of London 44 (1): 485-486.
Flower, W. H. (1883) On whales, past and present and their probable origin. (Royal Institution of Great Britain; London): 1-17.
Flower, W. H. (1885) An introduction to the osteology of the mammalia. 3rd ed., Ed. H. Gadow (Macmillan and Co.; London): 1-383.
Ford, J. K. B. and Reeves, R. R. (2008) Fight or flight: antipredator strategies of baleen whales. Mammal Review 38 (1): 50-86.
Fordyce, R. E. (1994) Waipatia maerewhenua, new genus and new species (Waipatiidae, new family), an archaic Late Oligocene dolphin (Cetacea: Odontoceti: Platanistoidea) from New Zealand. Contributions in marine mammal paleontology honoring Frank C. Whitmore, Eds. A. Berta and T. A. Deméré (San Diego Society of Natural History; San Diego) 29: 147-176.

R e f e r e n c e s
327
Fordyce, R. E. (2002) Simocetus rayi (Odontoceti: Simocetidae, New Family): a bizarre new archaic Oligocene dolphin from the Eastern North Pacific. Smithonian Contribution to Paleobiology. Cenozoic Mammals of Land and Sea: Tributes to the Career of Clayton E. Ray 93: 185-222.
Fordyce, R. E. (2003a) Chapter 9: Cetacean evolution and Eocene-Oligocene oceans revisited. From greenhouse to icehouse: the marine Eocene-Oligocene transition. Eds. D. R. Prothero, L. C. Ivany, et al. (Columbia University; New York): 154-170.
Fordyce, R. E. (2003b) Early crown-group Cetacea in the southern ocean: the toothed archaic mysticete Llanocetus. Journal of Vertebrate Paleontology 23 (Suppl. 3) Abstract of Papers 63rd. Annual Meeting: 50A-51A.
Fordyce, R. E. (2009a) Cetacean evolution. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 201-207.
Fordyce, R. E. (2009b) Cetacean fossil record. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 207-215.
Fordyce, R. E. (2009c) Neoceti. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 758-763.
Fordyce, R. E. (2013) Cetacea (whales, porpoises and dolphins). eLS (Ecyclopedia of Life Sciences). Major Reference Works (John Wiley & Sons; Chichester): 1-9.
Fordyce, R. E. and de Muizon, C. (2001) Evolutionary history of cetaceans: a review. Secondary Adaptation of Tetrapods to Life in Water. Eds. J.-M. Mazin and V. de Buffrenil (Friedrich Pfeil Verlag; München): 169-233.
Fordyce, R. E. and Marx, F. G. (2013) The pygmy right whale Caperea marginata: the last of the cetotheres. Proceedings of the Royal Society B (Biology) 280 (1753): 2645-2650.
Fordyce, R. E. and Marx, F. G. (2018) Gigantism precedes filter feeding in baleen whale evolution. Current Biology 28 (10): 1670-1676.
Franc, S., Marzin, E., Boutillon, M. M., et al. (1995) Immunohistochemical and biochemical analyses of 20 000 - 25 000-year-old fossil cartilage. European journal of biochemistry 234 (1): 125-131.
Fujiwara, S.-i. (2009) Olecranon orientation as an indicator of elbow joint angle in the stance phase, and estimation of forelimb posture in extinct quadruped animals. Journal of Morphology 270 (9): 1107-1121.
Galatius, A. (2005) Bilateral directional asymmetry of the appendicular skeleton of the harbour porpoise (Phocoena phocoena). Marine Mammal Science 21 (2): 401-410.
Galatius, A. (2006) Bilateral directional asymmetry of the appendicular skeleton of the white-beaked dolphin (Lagenorhynchus albirostris). Aquatic Mammals 32 (2): 232-235.
Galatius, A. (2010) Paedomorphosis in two small species of toothed whales (Odontoceti): how and why? Biological Journal of the Linnean Society 99 (2): 278-295.
Galatius, A., Andersen, M.-B. E. R., Haugan, B., et al. (2006) Timing of epiphyseal development in the flipper skeleton of the harbour porpoise (Phocoena phocoena) as an indicator of paedomorphosis. Acta Zoologica 87: 77-82.
Galatius, A. and Kinze, C. C. (2003) Ankylosis patterns in the postcranial skeleton and hyoid bones of the harbour porpoise (Phocoena phocoena) in the Baltic and North Sea. Canadian Journal of Zoology 81 (11): 1851-1861.
Gasse, H. (2012) Nomina anatomica veterinaria. 5th ed. (Editorial Committee; Hannover, Columbia, Ghent, Sapporo): 1-160.

R e f e r e n c e s
328
Gatesy, J. (1998) Chapter 3: Molecular evidence for the phylogenetic affinities of Cetacea. The emergence of whales: evolutionary patterns in the origin of Cetacea. Advances in Vertebrate Paleobiology, Ed. J. G. M. Thewissen (Plenum Press; New York): 63-111.
Gatesy, J., Amato, G., Vrba, E., et al. (1997) A cladistic analysis of mitochondrial ribosomal DNA from the Bovidae. Molecular Phylogenetics and Evolution 7 (3): 303-319.
Gatesy, J., Geisler, J. H., Chang, J., et al. (2012) A phylogenetic blueprint for a modern whale. Molecular Phylogenetics and Evolution 66 (2): 479-506.
Gegenbaur, C. (1872) Schultergürtel der Wirbelthiere / Brustflosse der Fische. Untersuchungen zur vergleichenden Anatomie der Wirbelthiere (Wilhelm Engelmann; Leipzig) 2: 1-169.
Geisler, J. H. (2001) New morphological evidence for the phylogeny of Artiodactyla, Cetacea, and Mesonychidae. American Museum Novitates 3344: 1-53.
Geisler, J. H. (2019) Whale evolution: dispersal by paddle or fluke. Current Biology 29 (8): 294-R296.
Geisler, J. H., Boessenecker, R. W., Brown, M., et al. (2017) The origin of filter feeding in whales. Current Biology 27 (13): 2036-2042.
Geisler, J. H., Colbert, M. W. and Carew, J. L. (2014) A new fossil species supports an early origin for toothed whale echolocation. Nature 508 (7496): 383-394.
Geisler, J. H. and Luo, Z.-X. (1998) Chapter 6: Relationships of cetacea to terrestrial ungulates and the evolution of cranial vasculature in Cete. The emergence of whales: evolutionary patterns in the origin of Cetacea. Advances in Vertebrate Paleobiology, Ed. J. G. M. Thewissen (Plenum Press; New York): 163-212.
Geisler, J. H., McGowen, M. R., Yang, G., et al. (2011) A supermatrix analysis of genomic, morphological, and paleontological data from crown Cetacea. BMC Evolutionary Biology 11 (112): 1-33.
Geisler, J. H. and Uhen, M. D. (2005) Phylogenetic relationships of extinct cetartiodactyls: results of simultaneous analyses of molecular, morphological, and stratigraphic data. Journal of Mammalian Evolution 12 (1-2): 145-160.
Gemmell, G. L., McInnes, J. D., Heinrichs, S. J., et al. (2015) Killer whale (Orcinus orca) predation on whales in Sri Lankan Waters. Aquatic Mammals 41 (3): 265-271.
Gerstner, C. L. (1999) Maneuverability of four species of coral-reef fish that differ in body and pectoral-fin morphology. Canadian Journal of Zoology 77 (7): 1102-1110.
Gibbes, R. W. (1847) On the fossil genus Basilosaurus, Harlan, (Zeuglodon, Owen) with a notice of specimens from the Eocene green sand of South Carolina. Journal of the Academy of Natural Sciences of Philadelphia 1 (Pt 1): 5-17.
Gingerich, P. D. (1998) Chapter 15: Paleobiological perspectives on Mesonychia, Archaeoceti, and the origin of whales. The emergence of whales: evolutionary patterns in the origin of Cetacea. Advances in Vertebrate Paleobiology, Ed. J. G. M. Thewissen (Plenum Press; New York): 423-449.
Gingerich, P. D. (2003a) Evolution of Eocene Archaeoceti (Cetacea) in relation to skeletal proportions and locomotion of living semiaquatic mammals. Journal of Vertebrate Paleontology 23 (Suppl. 3) Abstract of Papers 63rd. Annual Meeting: 55.
Gingerich, P. D. (2003b) Land-to-sea transition in early whales: evolution of Eocene Archaeoceti (Cetacea) in relation to skeletal proportions and locomotion of living semiaquatic mammals. Paleobiology 29 (3): 429-454.

R e f e r e n c e s
329
Gingerich, P. D. (2012) Evolution of whales from land to sea. Proceedings of the American Philosophical Society 156 (3): 309-323.
Gingerich, P. D. (2015) New partial skeleton and relative brain size in the Late Eocene archaeocete Zygorhiza kochii (Mammalia, Cetacea) from the Pachuta Marl of Alabama, with a note on contemporaneous Pontogeneus brachyspondylus. Contributions from the Museum of Paleontology, University of Michigan 32 (10): 161-188.
Gingerich, P. D., Arif, M. and Clyde, W. C. (1995) New Archaeocetes (Mammalia, Cetacea) from the Middle Eocene Domanda Formation of Sulaiman Range, Punjab (Pakistan). Contributions from the Museum of Paleontology, University of Michigan 29 (11): 291-330.
Gingerich, P. D., Smith, B. H. and Simons, E. L. (1990) Hind limbs of Eocene Basilosaurus: evidence of feet in whales. Science 249 (4965): 154-157.
Gingerich, P. D. and Uhen, M. D. (1996) Ancalecetus simonsi, a new dorudontine archaeocete (Mammalia, Cetacea) from the early late Eocene of Wadi Hitan, Egypt. Contributions from the Museum of Paleontology, University of Michigan 29 (13): 359-401.
Gingerich, P. D. and Uhen, M. D. (1998) Likelihood estimation of the time of origin of Cetacea and the time of divergence of Cetacea and Artiodactyla. Palaeontologia Electronica 1 (2): 1-47.
Gingerich, P. D., Ul Haq, M., Zalmout, I. S., et al. (2001a) Origin of whales from early artiodactyls: hands and feet of Eocene Protocetidae from Pakistan. Science 293 (5538): 2239–2242.
Gingerich, P. D., Ul-Haq, M., Khan, I. H., et al. (2001b) Eocene stratigraphy and archeocete whales (Mammalia, Cetacea) of Drug Lahar in the eastern Sulaiman Range, Balochistan (Pakistan). Contributions from the Museum of Paleontology, University of Michigan 30 (11): 269-319.
Gingerich, P. D., von Koenigswald, W., Sanders, W. J., et al. (2009) New protocetid whale from the middle Eocene of Pakistan: birth on land, precocial development, and sexual dimorphism. PLoS one 4 (2): 1-20.
Gingerich, P. D. and Zalmout, I. S. (2019) Aegicetus gehennae, a new late Eocene protocetid (Cetacea, Archaeoceti) from Wadi Al Hitan, Egypt, and the transition to tail-powered swimming in whales. PLoS ONE 14 (12): 1-56.
Gingerich, P. D., Zalmout, I. S., Ul-Haq, M., et al. (2005) Makaracetus bidens, a new protocetid archaeocete (Mammalia, Cetacea) from the early middle Eocene of Balochistan (Pakistan). Contributions from the Museum of Paleontology, University of Michigan 31 (9): 197-210.
Gol'din, P. E., Startsev, D. and Krakhamalnaya (2013) The anatomy of Cetotherium riabinini Hofstein, 1948, a baleen whale from the late Mioceneof Ukraine. Acta Palaeontologica Polonica 59 (4): 1-56.
Gol'din, P. E. and Steeman, M. E. (2015) From problem taxa to problem solver: a new Miocene family, Tranatocetidae, brings perspective on baleen whale evolution. PLoS ONE 10 (9): 1-19.
Gol’din, P. E. (2004) Growth and body size of the harbour porpoise, Phocoena phocoena (Cetacea, Phocoenidae), in the Sea of Azov and the Black Sea Vestnik zoologii 38 (4): 59-73.
Goldbogen, J. A., Calambokidis, J., Shadwick, R. E., et al. (2006) Kinematics of foraging dives and lunge-feeding in fin whales. Journal of Experimental Biology 209 (7): 1231-1244.

R e f e r e n c e s
330
Goldbogen, J. A., Pyenson, N. D. and Shadwick, R. E. (2007) Big gulps require high drag for fin whale lunge feeding. Marine Ecology Progress Series 349: 289-301.
Goldbogen, J. A., Shadwick, R. E., Lillie, M. A., et al. (2015) Using morphology to infer physiology: case studies on rorqual whales (Balaenopteridae). Canadian Journal of Zoology 93 (9): 687-700.
Gómez-Campos, E., Aguilar, A. and Goodall, R. N. P. (2010) Serrated flippers and directional asymmetry in the appendicular skeleton of the Commerson’s dolphin (Cephalorhynchus commersonii). Tha Anatomical Record 293: 1816-1824.
Gray, J. E. (1866) Catalogue of seals and whales in the British Museum. 2nd ed. (Order of Trustees; London): 1-402.
Gray, J. E. (1871) Notes on the Berardius of New Zealand. Annals and Magazine of Natural History 4 (8): 115-117.
Gray, J. E. (1873) Remarks on some of the species in the forgoing paper. Annals and Magazine of Natural History 4 (11): 107-112.
Gray, J. E. (1874a) List of seals, whales, and dolphins of New Zealand. Transactions and Proceedings of the New Zealand Institute 1873 (6): 87-89.
Gray, J. E. (1874b) Notes on Dr. Hector's paper on the whales and dolphins of the New Zealand seas. Transactions and Proceedings of the New Zealand Institute 1873 (6): 93-97.
Gray, N.-M., Kainec, K., Madar, S. I., et al. (2007) Sink or swim? Bone density as a mechanism for buoyancy control in early cetaceans. Anatomical Record 290 (6): 638-653.
Gregory, W. G. (1912) Notes on the principles of quadrupedal locmotion and the mechanism of the limbs in hoofed animals. Annals of the New York Academy of Sciences 22 (1): 267-294.
Grogan, E. D. and Lund, R. (2004) Chapter 1: The origin and relationships of early Chondrichthyes. Biology of sharks and their relatives. Marine Biology Series, Eds. J. C. Carrier, J. A. Musick, et al. (CRC Press; London, Boca Raton, New York, Washington D.C.): 3-31.
Guinet, C. and Bouvier, J. (1995) Development of intentional stranding hunting techniques in killer whale (Orcinus orca) calves at Crozet Archipelago. Canadian Journal of Zoology 73 (1): 27-33.
Haast, J. (1870a) Preliminary notice of a ziphid whale, probably Berardius arnouxii, stranded on the 16th of December 1868, on the sea beach, near Brighton, Canterbury. Transactions and Proceedings of the New Zealand Institute 1869 (2): 190-192.
Haast, J. (1870b) Preliminary notice of a ziphioid whale, probably Berardius arnuxii, stranded on the 16th of December, 1868, on the sea-beach, near New Brighton, Canterbury, New Zealand. Annals and Magazine of Natural History 4 (6): 348-351.
Haast, J. (1876a) 1. Notes on the skeleton of Ziphius novae-zealandiae. Proceedings of the Zoological Society of London 44 (1): 466-477.
Haast, J. (1876b) 3. Notes on Mesoplodon floweri. Proceedings of the Zoological Society of London 44 (1): 478-485.
Hale, H. M. (1931) The pygmy right whale (Caperea marginata) in South Australian Waters, Part I. Records of the South Australian Museum 4 (3): 314-31.
Hale, H. M. (1962) Occurence of the whale Berardius arnuxi in southern Australia. Records of the South Australian Museum 14 (2): 231-243.
Hale, H. M. (1964) The pygmy right whale (Caperea marginata) in South Australian Waters, Part II. Records of the South Australian Museum 14 (4): 679-194.

R e f e r e n c e s
331
Hamilton, H., Caballero, S., Collins, A. G., et al. (2001) Evolution of river dolphins. Proceedings of the Royal Society 268 (1466): 549-556.
Hanke, H. (1915) Über die Brustflosse von Mesoplodon bidens (Sow.). Anatomischer Anzeiger 48 (2): 59-62.
Harlan, R. (1834) Notice of fossil bones found in the Tertiary formation of the state of Louisiana. Transactions of the American Philosophical Society 4: 397-403.
Hector, J. (1870) List of the bones of seals and whales in the Colonial Museum, Wellington, New Zealand. With notes by Dr. J. E. Gray. Annals and Magazine of Natural History 4 (5): 220-224.
Hector, J. (1873a) Notes on the whales and dolphins of the New-Zealand seas. With Remarks by Dr. J. E. Gray. Annals and Magazine of Natural History 4 (11): 104-107.
Hector, J. (1873b) On the whales and dolphins of the New Zealand seas. Transactions and Proceedings of the New Zealand Institute 1872 (5): 154-170, Pl 1-6.
Hector, J. (1875) Notes on New Zealand whales. Transactions and Proceedings of the New Zealand Institute 1874 (3): 251-265, Pl 16-18.
Hector, J. (1878) Notes on the whales of the New Zealand seas. Transactions and Proceedings of the New Zealand Institute 1877 (10): 331-342.
Heithaus, M. R. (2001) Predator-prey and competitive interactions between sharks (Order Selachii) and dolphins (Suborder Odontoceti): a review. Journal of Zoology 253 (1): 53-68.
Heithaus, M. R. and Dill, L. M. (2009) Feeding Strategies and Tactics. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 414-423.
Herald, E. S., Brownell, R. L. J., Frye, F. L., et al. (1969) Blind river dolphin: first side-swimming cetacean. Science 166 (3911): 1408-1410.
Heyning, J. E. and Mead, J. G. (1996) Suction feeding in beaked whales: morphological and observational evidence. Contributions in Science, Natural History Museum Los Angeles County 464: 1-12.
Heyning, J. E. and Mead, J. G. (2009) Cuvier’s beaked whale, Ziphius cavirostris. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 294-295.
Hildebrand, M. (1965) Symmetrical gaits of horses. Science 150 (3697): 701-708. Hildebrand, M. (1989) The quadrupedal gaits of vertebrates. Bioscience 39 (11): 766. Hobson, R. P. and Martin, A. R. (1996) Behaviour and dive times of Arnoux's beaked
whales, Berardius arnuxii, at narrow leads in fast ice. Canadian Journal of Zoology 74 (2): 388-393.
Hocking, D. P., Marx, F. G., Park, T., et al. (2017) A behavioural framework for the evolution of feeding in predatory aquatic mammals. Proceedings of the Royal Society B (Biology) 284 (1850): 2750-2759.
Holder, N. (1983a) Developmental constraints and the evolution of vertebrate digit patterns. Journal of Theoretical Biology 104 (3): 451-471.
Holder, N. (1983b) The vertebrate limb: patterns and constraints in development and evolution. Development and Evolution. Ed. B. C. Goodwin, Holder, N., Wylie, C. C. (Cambridge University Press; New York): 399-425.
Holliday, C. M., Ridgely, R. C., Sedlmayr, J. C., et al. (2010) Cartilaginous epiphyses in extant archosaurs and their implications for reconstructing limb function in dinosaurs. PLoS One 5 (9): 1-17.
Horner, J. R. (1997) Rare preservation of an incompletely ossified fossil embryo. Journal of Vertebrate Paleontology 17 (2): 431-434.

R e f e r e n c e s
332
Houssaye, A., Tafforeau, P., de Muizon, C., et al. (2015) Transition of Eocene whales from land to sea: evidence from bone microstructure. PloS One 10 (2): 1-28.
Howell, A. B. (1927) Contribution to the anatomy of the Chinese finless porpoise, Neomeris phocaenoides. Proceedings of the American Philosophical Society 70 (13): 1-40.
Howell, A. B. (1929) Contribution to the comparative anatomy of the eared and earless seals (genera Zalophus and Phoca). Proceedings of the United States National Museum 73 (15): 1-142.
Howell, A. B. (1930a) Aquatic Mammals. (Charles C. Thomas; Springfield and Baltimore): 1-338.
Howell, A. B. (1930b) Myology of the narwhal (Monodon monoceros). American Journal of Anatomy 46 (2): 187-215.
Howell, A. B. (1937) Morphogenesis of the shoulder architecture. Part VI. Therian Mammalia. The Quarterly Review of Biology 12 (4): 440-462.
Howes, G. B. (1888) Observations upon the morphology and genesis of supernumerary phalanges, with especial reference to those of the Amphibia. Proceeding of the Zoological Society of London 56 (1): 495-511.
Hunter, J. and Banks, J. (1787) Observations on the structure and oeconomy of whales. By John Hunter, Esq. FRS; Communicated by Sir Joseph Banks, Bart. PRS. Philosophical Transactions of the Royal Society of London 77: 371-450.
Irwin, D. M. and Árnason, Ú. (1994) Cytochrome b gene of marine mammals: phylogeny and evolution. Journal of Mammalian Evolution 2 (1): 37-55.
Ivlev, Y. F., Kuznetsov, A. N. and Rutovskaya, M. V. (2010) Preliminary data on the swimming kinematics of the Russian desman (Desmana moschata L.). Doklady Biological Sciences 431 (1): 144.
Jefferson, T. A., Stacey, P. J. and Baird, R. W. (1991) A review of killer whale interactions with other marine mammals: predation to co-existence. Mammal Review 21 (4): 151-180.
Jefferson, T. A., Webber, M. A. and Pitman, R. L. (2015) Marine mammals of the world: a comprehensive guide to their identification. 2nd ed., Eds. K. A. S. Gomez and P. Gonzalez (Academic Press; Amsterdam): 1-528.
Johnson, M. P., Hickmott, L. S., Aguilar de Soto, N., et al. (2008) Echolocation behaviour adapted to prey in foraging Blainville’s beaked whale (Mesoplodon densirostris). Proceedings of the Royal Society B (Biology) 275 (1631): 133-139.
Johnson, M. P., Madsen, P. T., Zimmer, W. M. X., et al. (2004) Beaked whales echolocate on prey. Proceedings of the Royal Society B (Biology) 271 (Suppl. 6): S383-S386.
Jones, M. L. and Swartz, S. L. (2009) Gray whale Eschrichtius robustus. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 503-511.
Kelley, N. P. and Motani, R. (2015) Trophic convergence drives morphological convergence in marine tetrapods. Biological Letters 11: 1-5.
Kellogg, R. (1928) The history of whales - their adaptation to life in the water. The Quarterly Review of Biology 3 (1): 29-76.
Kellogg, R. (1936) A review of the Archaeoceti. Carnegie Institution of Washington Publication; Washington) 482: 1-366.
Kemper, C. M. (1999) Estimating body length of pygmy right wales (Caperea marginata) from measurements of the skeleton and baleen. Marine Mammal Science 15 (3): 683-700.

R e f e r e n c e s
333
Kemper, C. M. (2002) Distribution of the pygmy right whale, Caperea marginata, in the Australasian region. Marine Mammal Science 18 (1): 99-111.
Kemper, C. M. (2009) Pygmy right whale. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 939-941.
Kemper, C. M., Dulton, J., Forster, B., et al. (1997) Sightings and strandings of the pygmy right whale Caperea marginata near Port Lincoln, South Australia and a review of the Australasian sightings. Transactions of the Royal Society of South Australia 121 (2): 79-82.
Kimball, C. (2006) The complete horse: an entertaining history of horses. Ed. K. Cornell (Voyageur Press; St. Paul, Minnesota): 1-144.
King, C. M., Roberts, C. D., Bell, B. D., et al. (2009) Chapter 24: Phylum Chordata: lancelets, fishes, amphibians, reptiles, birds, and mammals - Cetaceans: whales, dolphins, and porpoises. New Zealand Inventory of Biodiversity, Ed. D. P. Gordon (Canterbury University Press; Christchurch) 1: 503-511.
Kleinenberg, S. E., Yablokov, A. V., Bel’kovich, B. M., et al. (1969) Beluga (Delphinapterus leucas) investigation of the species (Translated from Russian). Israel Program for Scientific Translations (Izdatel'stvo Nauka; Moskva and Jerusalem): 1-376.
Klima, M. (1978) Comparison of early development of sternum and clavicle in striped dolphin and in humpback whale. The scientific reports of the whales research institute (The Whales Research Institute; Tokio) 30: 253-269.
Klima, M. (1990) Rudiments of the clavicle in the embryos of whales (Cetacea). Zeitschrift für Säugetierkunde 55: 202-212.
Klima, M. (1992) Schwimmbewegungen und Auftauchmodus bei Walen und bei Ichthyosauriern I. Anatomische Grundlagen der Schwimmbewegungen. Natur und Museum 122 (1): 1-17.
Klima, M., Oelschläger, H. H. A. and Wünsch, D. (1980) Morphology of the pectoral girdle in the Amazon dolphin Inia geoffrensis with special reference to the shoulder joint and the movement of the flipper. Sonderdruck aus Zeitschrift für Säugetierkunde 45 (5): 288-309.
Klinowska, M. (1991) Dolphins, porpoises, and whales of the world : the IUCN red data book. (IUCN; Gland and Cambridge): 1-429.
Knox, F. J. (1870) On the Balaenidae or whales with baleen. With notes on the Cetacea, in the Colonial Museum, Wellington by Dr. Hector. Transactions and Proceedings of the New Zealand Institute 1869 (2): 21-28, Pl 2a-b.
Knox, F. J. (1871) Observation on the Ziphidae [sic], a family of the Cetacea. With notes by Dr. Hector. Transactions and Proceedings of the New Zealand Institute 1870 (3): 125-129, Pl 13-17.
Koblitz, J. C., Stilz, P., Rasmussen, M. H., et al. (2016) Highly directional sonar beam of narwhals (Monodon monoceros) measured with a vertical 16 hydrophone array. PLoS ONE 11 (11): 1-16.
Krefft, G. (1865) Notice on a new species of sperm whale belonging to the genus Euphystetes of MacLeay. Proceedings of the Scientific Meetings of the Zoological Society of London: 708-713.
Kükenthal, W. (1890a) Cetologische Notiz. Anatomischer Anzeiger 5 (24): 709-710. Kükenthal, W. (1890b) Über die Hand der Cetaceen (dritte Mitteilung). Anatomischer
Anzeiger 5 (2): 44-52.

R e f e r e n c e s
334
Kükenthal, W. (1893a) Chapter II: Die Hand der Cetaceen. Vergleichend-anatomische und entwicklungsgeschichtliche Untersuchungen an Walthieren. Denkschrift der medizinisch-naturwissenschaftlicen Gesellschaft zu Jena (Gustaf Fischer; Jena) 3 (1): 21-70.
Kükenthal, W. (1893b) Mittheilungen über den Carpus des Weißwals. Morphologisches Jahrbuch 19: 56-64.
Kükenthal, W. (1921) Die Brustflosse des Buckelwales (Megaptera nodosa) und ihre Entwicklung. Sitzungsberichte der preussischen Akademie der Wissenschaften 306: 568-588.
Kükenthal, W. (1922) Die Brustflosse des Grönlandswales (Balaena mysticetus L.). Bijdragen tot de Dierkunde 22 (1): 59-64.
Lamb, D. S. (1890) The olecranon perforation. American Anthropologist 3 (2): 159-174.
Lambert, O., Bianucci, G., Post, K., et al. (2010) The giant bite of a new raptorial sperm whale from the Miocene epoch of Peru. Nature 466 (7302): 105-109.
Lambert, O., Bianucci, G., Salas-Gismondi, R., et al. (2019) An amphibious whale from the middle Eocene of Peru reveals early South Pacific dispersal of quadrupedal cetaceans. Current Biology 29 (8): 1352-1359.
Lambert, O., Collareta, A., Landini, W., et al. (2015) No deep diving: evidence of predation on epipelagic fish for a stem beaked whale from the Late Miocene of Peru. Proceedings of the Royal Society B (Biology) 282 (1815): 1530-1537.
Lambert, O., de Muizon, C. and Bianucci, G. (2013) The most basal beaked whale Ninoziphius platyrostris Muizon, 1983: clues on the evolutionary history of the family Ziphiidae (Cetacea: Odontoceti). Zoological Journal of the Linnean Society 167: 569-598.
Lambert, O., Martínez-Cáceres, M., Bianucci, G., et al. (2017) Earliest mysticete from the Late Eocene of Peru sheds new light on the origin of baleen whales. Current Biology 27 (10): 1535-1541.
Larson, S. G. (2015) Rotator cuff muscle size and the interpretation of scapular shape in primates. Journal of Human Evolution 80: 96-106.
Larson, S. G. and Stern Jr., J. T. (1989) Role of supraspinatus in the quadrupedal locomotion of vervets (Cercopithecus aethiops): implications for interpretation of humeral morphology. American Journal of Physical Anthropology 79 (3): 369-377.
Leopold, M. F., Begeman, L., Heße, E., et al. (2015) Porpoises: from predators to prey. Journal of Sea Research 97: 14-23.
Levermann, N., Galatius, A., Ehlme, G., et al. (2003) Feeding behaviour of free-ranging walruses with notes on apparent dextrality of flipper use. BMC Ecology 3 (9): 1-13.
Liebig, P. M., Taylor, T.-S. A. and Flessa, K. W. (2003) Bones on the beach: marine mammal taphonomy of the Colorado Delta, Mexico. Palaios 18 (2): 168-175.
Long, J. A., Young, G. C., Holland, T., et al. (2006) An exceptional Devonian fish from Australia sheds light on tetrapod origins. Nature 444 (7116): 199-202.
López, L. M. M., Miller, P. J. O., Aguilar de Soto, N., et al. (2015) Gait switches in deep-diving beaked whales: biomechanical strategies for long-duration dives. Journal of Experimental Biology 218 (9): 1325-1338.
MacLeod, C. D., Santos, M. B. and Pierce, G. J. (2003) Review of data on diets of beaked whales: evidence of niche separation and geographic segregation. Journal of the Marine Biological Association of the United Kingdom 83 (3): 651-665.

R e f e r e n c e s
335
Madar, S. I. (2007) The postcranial skeleton of early Eocene pakicetid cetaceans. Journal of Paleontology 81 (1): 176-200.
Madar, S. I., Thewissen, J. G. M. and Hussain, S. T. (2002) Additional holotype remains of Ambulocetus natans (Cetacea, Ambulocetidae), and their implications for locomotion in early whales. Journal of Vertebrate Paleontology 22 (2): 4-5-422.
Madsen, P. T., Aguilar de Soto, N., Tyack, P. L., et al. (2014) Beaked whales. Current Biology 24 (16): 728-730.
Madsen, P. T., de Soto, N. A., Arranz, P., et al. (2013) Echolocation in Blainville’s beaked whales (Mesoplodon densirostris). Journal of Comparative Physiology A 199 (6): 451-469.
Madsen, P. T., Johnson, M. P., Aguilar de Soto, N., et al. (2005) Biosonar performance of foraging beaked whales (Mesoplodon densirostris). Journal of Experimental Biology 208 (2): 181-194.
Malm, A. V. (1871) Hvaldjur i Sveriges useer 1869. (P.A. Norstedt & Söner; Stockholm): 1-104.
Marelli, C. (1920) Revisión osteológica de Berardius arnouxii Duv. Anales del Museo Nacionale de Historia Natural de Buenos Aires 30: 411-444.
Marigo, J., Pinto, N. S., Simoes-Lopes, P. C., et al. (2013) Case report of flipper anatomic anomaly of Sotalia guianensis from Sepetiba Bay, Rio de Janeiro. Anatomical Record 296 (7): 1016-1018.
Marples, B. J. (1956) Cetotheres from the Oligocene of New Zealand. Proceedings of the Zoological Society of London 126 (4): 565-580.
Martin, C. P. (1933) The cause of torsion of the humerus and of the notch on the anterior edge of the glenoid cavity of the scapula. Journal of Anatomy 67 (4): 573-582.
Martínez-Cáceres, M. and de Muizon, C. (2011) A new basilosaurid (Cetacea, Pelagiceti) from the late Eocene to early Oligocene Otuma Formation of Peru. Comptes Rendus 10 (7): 517-526.
Martínez-Cáceres, M., Lambert, O. and de Muizon, C. (2017) The anatomy and phylogenetic affinities of Cynthiacetus peruvianus, a large Dorudon-like basilosaurid (Cetacea, Mammalia) from the late Eocene of Peru. Geodiversitas 39 (1): 7-164.
Marx, F. G. (2011) The more the merrier? A large cladistic analysis of mysticetes, and comments on the transition from teeth to baleen. Journal of Mammalian Evolution 18 (2): 77-100.
Marx, F. G., Buono, M. R., Fordyce, R. E., et al. (2013) Juvenile morphology: a clue to the origins of the most mysterious of mysticetes? Naturwissenschaften 100 (3): 257-261.
Marx, F. G. and Fordyce, R. E. (2015) Baleen boom and bust: a synthesis of mysticete phylogeny, diversity and disparity. Royal Society Open Science 2 (4): 1-14.
Marx, F. G. and Fordyce, R. E. (2016) A link no longer missing: new evidence for the cetotheriid affinities of Caperea. PLoS ONE 11 (10): 1-26.
Matsuoka, K., Fujise, Y. and Pastene, L. A. (1996) A sighting of a large school of the pygmy right wale, Caperea marginata, in the southeast Indian Ocean. Marine Mammal Science 12 (4): 594-597.
Matsuoka, K., Pitman, R. L. and Marquez, F. F. C. (2005) A note on a pygmy right whale (Caperea marginata) sighting in the southwestern Pacific Ocean. Journal of Cetacean Research Management 7 (1): 71-73.

R e f e r e n c e s
336
McCann, C. (1962) The taxonomic status of the beaked whale, Mesoplodon hectori (Gray) - Cetacea. Records of the Dominion Museum 4 (9): 83-94.
McCann, C. (1975) A study of the genus Berardius Duvernoy. The Scientific Reports of the Whales Research Institute (The Whales Research Institute; Tokio) 27: 111-137.
McGowen, M. R., Gatesy, J. and Wildman, D. E. (2014) Molecular evolution tracks macroevolutionary transitions in Cetacea. Trends in Ecology and Evolution 29 (6): 1-11.
McGowen, M. R., Spaulding, M. and Gatesy, J. (2009) Divergence date estimation and a comprehensive molecular tree of extant cetaceans. Molecular Phylogenetics and Evolution 53 (3): 891-906.
McLean, D. (2012) Understanding aerodynamics: arguing from the real physics. (John Wiley & Sons; Chichester): 1-550.
Mead, J. G. (2009) Beaked whales. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 94-97.
Mead, J. G. and Fordyce, R. E. (2009) The therian skull. Smithsonian Contributions to Zoology (Smithsonian Institution Scholarly Press; Washington D. C.) 627: 1-248.
Mellor, L., Cooper, L. N., Torre, J., et al. (2009) Paedomorphic ossification in porpoises with an emphasis on the vaquita (Phocoena sinus). Aquatic Mammals 35 (2): 193-202.
Miklosovic, D. S., Murray, M. M., Howle, L. E., et al. (2004) Leading-edge tubercles delay stall on humpback whale Megaptera novaeangliae flippers. Physics and Fluids 16 (5): L39-L42.
Milinkovitch, M. C., Berube, M. and Palsbøll, P. J. (1998) Chapter 4: Cetaceans are highly derived artiodactyls. The emergence of whales: evolutionary patterns in the origin of Cetacea. Advances in Vertebrate Paleobiology, Ed. J. G. M. Thewissen (Plenum Press; New York): 113-131.
Miller, G. S. (1923) The telescoping of the cetacean skull. Smithsonian Miscellaneous Collections (Smithonian Institution; Washington) 76 (5): 1-71.
Miller, P. J. O., Johnson, M. P., Tyack, P. L., et al. (2004) Swimming gaits, passive drag and buoyancy of diving sperm whales Physeter macrocephalus. Journal of Experimental Biology 207 (11): 1953-1967.
Minamikawa, S., Iwasaki, T. and Kishiro, T. (2007) Diving behaviour of a Baird’s beaked whale, Berardius bairdii, in the slope water region of the western North Pacific: first dive records using a data logger. Fisheries Oceanography 16 (6): 573-577.
Montgelard, C., Catzeflis, F. M. and Douzery, E. (1997) Phylogenetic relationships of artiodactyls and cetaceans as deduced from the comparison of cytochrome b and 12S rRNA mitochondrial sequences. Molecular Biology and Evolution 14 (5): 550-559.
Moore, J. C. (1968) Relationships among the living genera of beaked whales with classifications, diagnoses and keys.. Fieldiana, Zoology 53 (4): 209-298.
Moore, J. C. (1972) More skull characters of the beaked whale Indopacetus pacificus and comparative measuements of Austral relatives. Fieldiana, Zoology 62: 1-19.
Morin, P. A., Scott Baker, C., Brewer, R. S., et al. (2016) Genetic structure of the beaked whale genus Berardius in the North Pacific, with genetic evidence for a new species. Marine Mammal Science 33 (1): 96-111.

R e f e r e n c e s
337
Morisaka, T. and Connor, R. C. (2007) Predation by killer whales (Orcinus orca) and the evolution of whistle loss and narrow-band high frequency clicks in odontocetes. European Society for Evolutionary Biology, Journal Compilation 20: 1439-1458.
Murie, J. (1865) 5. On the anatomy of a fin-whale (Physalus antiquorum, Gray) captured near Gravesend. Proceedings of the Zoological Society of London 33 (1): 206-227.
Murie, J. (1870a) Notes on the white-beaked bottlenose, Lagenorhynchus albirostris, Gray. Journal of the Linnean Society, Zoology 11 (50): 141-152.
Murie, J. (1870b) On Risso's grampus, G. rissoanus (Desm.). Journal of Anatomy and Physiology 5 (Pt 1): 118-138.
Murie, J. (1872) V. On the form and structure of the manatee (Manatus americanus). Transactions of the Zoological Society of London 8 (3): 127-220.
Murie, J. (1873) On the organization of the Caaing Whale, Globiocephalus melas. Transactions of the Zoological Society of London 4 (8): 235-302.
Murray, P. D. F. and Drachman, D. B. (1969) The role of movement in the development of joints and related structures: the head and neck in the chick embryo. Journal of Embryology and Eperimental Morphology 22 (3): 349-371.
Nicodemus, M. C. and Clayton, H. M. (2003) Temporal variables of four-beat, stepping gaits of gaited horses. Applied Animal Behaviour Science 80 (2): 133-142.
Nikaido, M., Rooney, A. P. and Okada, N. (1999) Phylogenetic relationships among cetartiodactyls based on insertions of short and long interpersed elements: hippopotamuses are the closest extant relatives of whales. Proceedings of the National Academy of Sciences 96 (18): 10261-10266.
Northover, J. M. (2011) Skeletal morphology and evidence for swimming in a fossil stem pinniped, Puijila darwini, from the Canadian high Arctic. (Department of Earth Science; Carleton University) MSc thesis: 1-151.
O'Leary, M. A., Bloch, J. I., Flynn, J. J., et al. (2013) The placental mammal ancestor and the post-K-Pg radiation of placentals. Science 339 (6120): 662-667.
O'Leary, M. A. and Gatesy, J. (2008) Impact of increased character sampling on the phylogeny of Cetartiodactyla (Mammalia): combined analysis including fossils. Cladistics 24 (4): 397-442.
O'Leary, M. A. and Geisler, J. H. (1999) The position of Cetacea within Mammalia: phylogenetic analysis of morphological data from extinct and extant taxa. Systematic Biology 48 (3): 455-490.
O'Leary, M. A. and Rose, K. D. (1995) Postcranial skeleton of the early Eocene mesonychid Pachyaena (Mammalia: Mesonychia). Journal of Vertebrate Paleontology 15 (2): 401-430.
Oliver, W. R. B. (1922) A review of the Cetacea of the New Zeland seas. Proceedings of the Zoological Society of London 3 (92): 557-585.
Omura, H. (1975) Osteological study of the minke whale from the Antarctic. The scientific reports of the Whales Research Institute (The Whales Research Institute; Tokyo) 27: 1-36.
Padian, K. (1992) A proposal to standardize tetrapod phalangeal formula designations. Journal of Vertebrate Paleontology 12 (2): 260-262.
Perrin, J. B. (1870) Notes on the anatomy of Balaenoptera rostrata. Proceedings of the Scientific Meetings of the Zoological Society London: 805-817.
Perrin, W. F., Warner, R. R., Fiscus, C. H., et al. (1973) Stomach contents of porpoise, Stenella spp., and yellowfin tuna, Thunnus albacares, in mixed-species aggregations. Fishery Bulletin 71 (4): 1077-1092.

R e f e r e n c e s
338
Philips, J. D., Nachtigall, P. E., Au, W. W. L., et al. (2003) Echolocation in the Risso’s dolphin, Grampus griseus. The Journal of the Acoustical Society of America 113 (1): 605-616.
Pike, G. C. (1953) Two records of Berardius bairdi from the coast of British Columbia. Journal of Mammalogy 34 (1): 98-104.
Pike, G. C. (1962) Migration and feeding of the gray whale (Eschrichtius gibbosus). Journal of the Fisheries Board of Canada 19 (5): 815-838.
Pilleri, G. and Gihr, M. (1976a) Chapter 1: Platanistoidea - The function and osteology of the manus of Platanista gangetica and Platanista indi. Investigations on Cetacea, Ed. G. Pilleri (Brain Anatomy Institute; Bern) 7: 109-118.
Pilleri, G. and Gihr, M. (1976b) Chapter 1: Platanistoidea - The manus of the Amazon dolphin, Inia geoffrensis (de Blainville 1817) and remarks concerning socalled ‘polydactyly’. Investigations on Cetacea, Ed. G. Pilleri (Brain Anatomy Institute; Bern) 7: 129-137.
Pilleri, G. and Gihr, M. (1976c) Chapter 1: Platanistoidea - On the manus of the La Plata dolphin, Pontoporia blainvillei. Investigations on Cetacea, Ed. G. Pilleri (Brain Anatomy Institute; Bern) 7: 119-128.
Pilleri, G., Gihr, M., Kraus, C., et al. (1976) Chapter 3: Comparative study of the skin and general myology of Platanista indi and Delphinus delphis in relation to hydrodynamics and behaviour. Investigations on Cetacea, Ed. G. Pilleri (Brain Anatomy Institute; Bern) 6: 89-127.
Pitman, R. L. (2009) Mesoplodont whales (Mesoplodon spp.). Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 721-726.
Pitman, R. L., Ballance, L. T., Mednick, S. I., et al. (2001) Killer whale predation on sperm whales: observations and implications. Marine mammal science 17 (3): 494-507.
Pitman, R. L. and Durban, J. W. (2012) Cooperative hunting behavior, prey selectivity and prey handling by pack ice killer whales (Orcinus orca), type B, in Antarctic Peninsula waters. Marine Mammal Science 28 (1): 16-36.
Polly, P. D. (2007) Chapter 15: Limbs in mammalian evolution. Fins into limbs: evolution, development and transformation. Ed. B. K. Hall (University of Chicago; Chicago): 245-268.
Ponganis, P. J., Kooyman, G. L. and Ridgway, S. H. (2003) Comparative diving physiology. Bennett and Elliot's physiology and medicine of diving. Eds. A. O. Brubakk and T. S. Neuman (Saunders; Edinburgh): 211-226.
Pouchet, G. and Beauregard, H. (1889) Recherches sur le cachalot. Nouvelles archives du Museum d'histoire naturelle 3 (1): 1-90.
Prothero, D. R. (2015) The story of life in 25 fossils. Tales of intrepid fossil hunters and the wonders of evolution. (Columbia University Press; New York): 1-389.
Purves, P. E. and Pilleri, G. (1978) Chapter 4: Anatomy and physiology - The functional anatomy and general biology of Pseudorca crassidens (Owen) with a review of the hydrodynamics and acoustics in Cetacea. Investigations on Cetacea, Ed. G. Pilleri (Brain Anatomy Institute; Bern) 9: 67-227.
Pyenson, N. D. (2017) The ecological rise of whales chronicled by the fossil record. Current Biology 27 (11): 558-564.
Quick, N. J., Cioffi, W. R., Shearer, J. M., et al. (2020) Extreme diving in mammals: first estimates of behavioural aerobic dive limits in Cuvier's beaked whales. Journal of Experimental Biology 223 (18): 1-6.

R e f e r e n c e s
339
Quiring, D. P. and Harlan, C. F. (1953) On the anatomy of the manatee. Journal of Mammalogy 34 (2): 192-203.
Rapp, W. (1837) Die Cetaceen zoologisch-anatomisch dargestellt. (J. G. Cotta’schen Buchhandlung; Stuttgart und Tübingen): 1-183.
Read, A. J. (2009) Porpoises, Overview. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 920-923.
Reeves, R. R., Stewart, B. S., Clapham, P. J., et al. (2002) Guide to marine mammals of the world. (Alfred A. Knopf; New York): 1-528.
Rice, D. W. (1998) Marine mammals of the world. Society for Marine Mammalogy (The Society for Marine Mammals; Lawrence, Kansas) Special Publication Number 4: 1-231.
Rice, D. W. and Wolman, A. A. (1971) The life history and ecology of the gray whale (Eschrichtius robustus). American Society of Mammalogists (The American Society of Mammalogists; Stillwater) Special publication Number 3: 1-142.
Richardson, M. K. and Chipman, A. D. (2003) Developmental constraints in a comparative framework: a test case using variations in phalanx number during amniote evolution. Journal of Experimental Biology 296B (1): 8–22.
Richardson, M. K. and Oelschlager, H. H. A. (2002) Time, pattern, and heterochrony: a study of hyperphalangy in the dolphin embryo flipper. Evolution and Development 4 (6): 435-444.
Ridgway, S. H. ed. (1972) Mammals of the sea: biology and medicine. (Charles C. Thomas; Springfield) R. L. J. Brownell, D. K. Caldwell, et al.: 1-297.
Ridgway, S. H. and Harrison, R. eds. (1985) The sirenians and baleen whales. Handbook of Marine Mammals 3: 1-378.
Rivera, A. R. V., Rivera, G. and Blob, R. W. (2013) Forelimb kinematics during swimming in the pig-nosed turtle, Carettochelys insculpta, compared with other turtle taxa: rowing versus flapping, convergence versus intermediacy. Journal of Experimental Biology 216 (4): 668-680.
Robineau, D. (1989) Les types de cétacés actuels du Muséum National d'Histoire Naturelle: I. Balaenidae, Balaenopteridae, Kogiidae, Ziphiidae, Iniidae, Pontoporiidae. Bulletin Muséum national d'Histoire naturelle, Paris 4 (11): 271-289.
Romer, A. S. (1966) Vertebrate paleontology. 3rd ed. (University of Chicago Press; Chicago): 1-478.
Romero, A. (2012) Chapter 1: When whales became mammals: the scientific journey of cetaceans from fish to mammals in the history of science. New approaches to the study of marine mammals, Ed. A. Romero (InTech; Open access): 3-30.
Rommel, S. (1990) Chapter 2: Osteology of the bottlenose dolphin. The Bottlenose Dolphin. 29-49.
Rommel, S. A. and Reynolds III, J. E. (2009) Skeleton, Postcranial. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 1021-1033.
Rose, J., Moore, A., Russell, A., et al. (2014) Functional osteology of the forelimb digging apparatus of badgers. Journal of Mammalogy 95 (3): 543-558.
Rose, K. D. (1985) Comparative osteology of North American dichobunid artiodactyls. Journal of Paleontology 59 (5): 1203-1226.
Ross, G. J. B., Best, P. B. and Donnelly, B. G. (1975) New records of the pygmy right whale (Caperea marginata) from South Africa, with comments on distribution, migration, appearance, and behavior. Journal of the Fisheries Research Board Canada 32 (7): 1005-1017.

R e f e r e n c e s
340
Sanchez, J. A. and Berta, A. (2010) Comparative anatomy and evolution of the odontocete forelimb. Marine Mammal Science 26 (1): 140-160.
Sanders, A. E. (2002) Paleontology of the Late Oligocene Ashley and Chandler Bridge Formations of South Carolina, 3: Eomysticetidae, a new family of mysticetes. Smithonian Contribution to Paleobiology. Cenozoic Mammals of Land and Sea: Tributes to the Career of Clayton E. Ray 93: 313-356.
Sargeant, B. L., Mann, J., Berggren, P., et al. (2005) Specialization and development of beach hunting, a rare foraging behavior, by wild bottlenose dolphins (Tursiops sp.). Canadian Journal of Zoology 83 (11): 1400-1410.
Saskai, T., Nikaido, M., Hamilton, H., et al. (2005) Mitochondrial phylogenesis and evolution of mysticete whales. Systematic Biology 54 (1): 77-90.
Schorr, G. S., Falcone, E. A., Moretti, D. J., et al. (2014) First long-term behavioral records from Cuvier’s beaked whales (Ziphius cavirostris) reveal record breaking dives. PLoS ONE 9 (3): 1-10.
Schulte, H. v. W. (1916) Anatomy of a foetus of Balaenoptera borealis. Monographs of the pacific Cetacea: II The sei whale (Balaenoptera borealis Lesson) 1 (6): 394-500.
Schulte, H. v. W. and Smith, M. d. F. (1918) The external characters, skeletal muscles, and peripheral nerves of Kogia breviceps (Blainville). Bulletin American Museum of Natural History 38 (2): 7-72.
Scilla, A. (1670) La vana speculazione disingannata dal senso: lettera risponsiva circa i corpi marini, che petrificati si trovano in varii luoghi terrestri. (Andrea Colicchia; Napoli): 1-168.
Seckel, L. and Janis, C. M. (2008) Convergences in scapula morphology among small cursorial mammals: an osteological correlate for locomotory specialization. Journal of Mammalian Evolution 15 (4): 261-279.
Seilacher, A. and Gishlick, A. D. (2019) Morphodynamics. (CRC press; Boca Raton, London, New York): 1-513.
Shaffer, J. W., Moretti, D. J., Jarvis, S., et al. (2013) Effective beam pattern of the Blainville's beaked whale (Mesoplodon densirostris) and implications for passive acoustic monitoring. The Journal of the Acoustical Society of America 133 (3): 1770-1784.
Shu, D.-G., Morris, S. C., Han, J., et al. (2003) Head and backbone of the Early Cambrian vertebrate Haikouichthys. Nature 421 (6922): 526-529.
Simon, M., Wahlberg, M., Ugarte, F., et al. (2005) Acoustic characteristics of underwater tail slaps used by Norwegian and Icelandic killer whales (Orcinus orca) to debilitate herring (Clupea harengus). Journal of Experimental Biology 208 (12): 2459-2466.
Singhal, S. and Rao, V. (2007) Supratrochlear foramen of the humerus. Anatomical Science International 82 (2): 105-107.
Sironi, M., López, J. C., Bubas, R., et al. (2008) Predation by killer whales (Orcinus orca) on southern right whales (Eubalaena australis) off Patagonia, Argentina: effects on behavior and habitat choice. Encuentro del Comité Científico de la Comisión Ballenera Internacional (Documento SC/60/BRG29): 1-18.
Sisson, S. (1910) The anatomy of domestic animals. 2nd ed. (W. B. Saunders; Philadelphia and London): 1-930.
Smirnowsky, B. N. (1928) Einige Daten zur Myologie der Delphine. Anatomischer Anzeiger 66 (11/13): 193-199.

R e f e r e n c e s
341
Smith, G. D. J., Browne, K. W. and Gaskin, D. E. (1976) Functional myology of the harbour porpoise, Phocoena phocoena (L.). Canadian Journal of Zoology 54: 716-729.
Smith, J. M. and Savage, R. J. G. (1956) Some locomotory adaptations in mammals. Journal of the Linnean Society, Zoology 42 (288): 603-622.
Sokolov, V. E. and Rodionov, V. A. (1974) Morphofunctional patterns of shoulder girdle muscles in the Black Sea dolphins. Zoologial Journal 53 (5): 756-768.
Spaulding, M., O’Leary, M. A. and Gatesy, J. (2009) Relationships of Cetacea (Artiodactyla) among mammals: increased taxon sampling alters interpretations of key fossils and character evolution. PLoS ONE 4 (9): 1-14.
Spitz, J., Cherel, Y., Bertin, S., et al. (2011) Prey preferences among the community of deep-diving odontocetes from the Bay of Biscay, Northeast Atlantic. Deep-Sea Research I 58 (3): 273-282.
Stanhope, M. J., Smith, M. R., Waddell, V. G., et al. (1996) Mammalian evolution and the interphotoreceptor retinoid binding protein (IRBP) gene: convincing evidence for several superordinal clades. Journal of Molecular Evolution 43 (2): 83-92.
Stannius, H. F. (1849) Beschreibung der Muskeln des Tümmlers (Delphinus phocaena). Archiv für Anatomie und Physiologie, und wissenschaftliche Medizin 1: 1-41.
Steeman, M. E. (2007) Cladistic analysis and a revised classification of fossil and recent mysticetes. Zoological Journal of the Linnean Society 150: 875-894.
Steeman, M. E., Hebsgaard, M. B., Fordyce, R. E., et al. (2009) Radiation of extant cetaceans driven by restructuring of the oceans. Systematic Biology 58 (6): 573-585.
Steiger, G. H., Calambokidis, J., Straley, J. M., et al. (2008) Geographic variation in killer whale attacks on humpback whales in the North Pacific: implications for predation pressure. Endangered Species Research 4: 247-256.
Sterling, S. (1910) Beiträge zur Kenntnis der Muskulatur des Schultergürtels und der Vorderextremität bei Zahn- und Bartenwalen. Jenaische Zeitschrift für Naturwissenschaft 36 (Neue Folge 39): 667 - 680.
Stockin, K. A., Wiseman, N., Hartman, A., et al. (2008) Use of radiography to determine age class and assist with the post-mortem diagnostics of a Bryde's whale (Balaenoptera brydei). New Zealand Journal of Marine and Freshwater Research 42 (3): 307-313.
Strickler, T. L. (1978) Myology of the shoulder of Pontoporia blainvillei, including a review of the literature on shoulder morphology in the Cetacea. American Journal of Anatomy 152 (3): 419-432.
Struthers, J. (1871) Some points in the anatomy of a great fin-whale. Journal of Anatomy and Physiology 6 (Pt 1): 107–512.
Struthers, J. (1873) Account of rudimentary finger muscles found in a toothed whale (Hyperoodon bidens). Journal of Anatomy and Physiology 8 (Pt 1): 114-119.
Struthers, J. (1878) Account of rudimentary finger muscles found in the Greenland right-whale (Balaena mysticetus). Journal of Anatomy and Physiology 12 (Pt 2): 217-224.
Struthers, J. (1888) On some points in the anatomy of a Megaptera longimana. Journal of Anatomy and Physiology. Part II - The Limbs 22 (Pt 2): 240-282.
Struthers, J. (1889) Memoir on the anatomy of the humpback whale, Megaptera longimana. Reprint from ‘Journal of Anatomy and Physiology’ 1887-1889: 18-60.

R e f e r e n c e s
342
Struthers, J. (1895a) External characters and some parts of the anatomy of a beluga (Delphinapterus leucas). Journal of anatomy and physiology 30 (Pt 1): 124-156.
Struthers, J. (1895b) On the carpus of the Greenland right whale (Balaena mysticetus) and of fin-whales. Journal of Anatomy and Physiology 29 (Pt 2): 145-190.
Stuart, L. J. and Morejohn, G. V. (1980) Developmental patterns in osteology and external morphology in Phocoena phocoena. Age determination of toothed whales and sirenians. Reports of the International Whaling Commission Special Issue 3: 133-142.
Tanaka, H. (1999) Numerical analysis of the proximal humeral outline: bilateral shape differences. American Journal of Human Biology 11: 343-357.
Tanaka, Y. and Fordyce, R. E. (2014) Fossil dolphin Otekaikea marplesi (Latest Oligocene, New Zealand) expands the morphological and taxonomic diversity of Oligocene cetaceans. PLoS ONE 9 (9): 1-30.
Tanaka, Y. and Fordyce, R. E. (2015) A new Oligo-Miocene dolphin from New Zealand: Otekaikea huata expands diversity of the early Platanistoidea. Palaeontologia Electronica 18 (2): 1-71.
Taylor, B. K. (1978) The anatomy of the forelimb in the anteater (Tamandua) and its functional implications. Journal of Morphology 157 (3): 347-367.
Taylor, B. L., Larcher, F., Martien, K. K., et al. (2017) Guidelines and quantitative standards to improve consistency in cetacean subspecies and species delimitation relying on molecular genetic data. Marine Mammal Science 33 Special Issue 1: 132–155.
Thewissen, J. G. M. ed. (1998) The emergence of whales: evolutionary patterns in the origin of Cetacea. Advances in Vertebrate Paleobiology (Plenum Press; New York): 1-477.
Thewissen, J. G. M. (2009) Archaeocetes, Archaic. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 46-48.
Thewissen, J. G. M. and Bajpai, S. (2001) Whale origins as a poster child for macroevolution. BioScience 51 (12): 1037-1049.
Thewissen, J. G. M. and Bajpai, S. (2009) New skeletal material of Andrewsiphius and Kutchicetus, two Eocene cetaceans from India. Journal of Paleontology 83 (5): 635-663.
Thewissen, J. G. M., Cohn, M. J., Stevens, L. S., et al. (2006) Developmental basis for hind-limb loss in dolphins and origin of the cetacean bodyplan. PNAS 103 (22): 8414-8418.
Thewissen, J. G. M., Cooper, L. N., Clementz, M. T., et al. (2007) Whales originated from aquatic artiodactyls in the Eocene epoch of India. Nature 450 (7173): 1190-1195.
Thewissen, J. G. M., Cooper, L. N., George, J. C., et al. (2009) From land to water: the origin of whales, dolphins and porpoises. Evo Edu Outreach 2: 272-288.
Thewissen, J. G. M. and Fish, F. E. (1997) Locomotor evolution in the earliest cetaceans: functional model, modern analogues, and paleontological evidence. Paleobiology 23 (4): 482-490.
Thewissen, J. G. M. and Hussain, S. T. (1996) Ambulocetus natans, an Eocene cetacean (Mammalia) from Pakistan. Courier Forschungsinstitut Senckenberg (Senkenbergische Naturforschende Gesellschaft; Frankfurt a. M.) 191: 1-86.
Thewissen, J. G. M., Hussain, S. T. and Arif, M. (1994) Fossil evidence for the origin of aquatic locomotion in archaeocete whales. Science 263 (5144): 210-212.

R e f e r e n c e s
343
Thewissen, J. G. M. and Schneider, B. J. (2009) Marine mammal evolution. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 692.
Thewissen, J. G. M., Williams, E. M., Roe, L. J., et al. (2001) Skeletons of terrestrial cetaceans and the relationship of whales to artiodactyls. Nature 413 (6853): 277–281.
Thomas, H. W. and Barnes, L. G. (2015) The bone joint pathology osteochondrosis in extant and fossil marine mammals. Contributions in Science, Natural History Museum Los Angeles County 523: 1–35.
Torres Azevedo, C., Lima, J. Y., de Azevedo, R. M., et al. (2015) Thoracic limb bone development in Sotalia guianensis (Van Beneden 1864) along the coastline of Espírito Santo, Brazil. Journal of Mammalogy 96 (3): 541-551.
True, F. W. (1910) An account of the beaked whales of the family Ziphiidae in the collection of the United States National Museum, with remarks on some specimens in other American museums. (United States National Museum, Smithonian Institution; Washington) 73: 1-81.
Tsai, C.-H. (2017) A Miocene breeding ground of an extinct baleen whale (Cetacea: Mysticeti). PeerJ 5: 1-10.
Tsai, C.-H., Collareta, A., Fitzgerald, E. M. G., et al. (2017) Northern pygmy right whales highlight Quaternary marine mammal interchange. Current Biology 27 (19): 1037–1059.
Tsai, C.-H. and Fordyce, R. E. (2014a) Disparate heterochronic processes in baleen whale evolution Evolutionary Biology 41 (2): 299-307.
Tsai, C.-H. and Fordyce, R. E. (2014b) Juvenile morphology in baleen whale phylogeny. Naturwissenschaften 101 (9): 765-769.
Tsai, C.-H. and Fordyce, R. E. (2015) Ancestor-descendant relationships in evolution: origin of the extant pygmy right whale, Caperea marginata. Biology letters 11 (1): 1-4.
Turnbull, S. D. (2012) White shark (Carcharodon carcharias) attack on a harbor porpoise (Phocaena phocaena) in the Bay of Fundy, Canada. Notes of the Northeastern Naturalist 19 (4): 705-707.
Turner, W. (1872) On the occurrence of Ziphius cavirostris in the Shetland Seas, and a comparison of Its skull with that of Sowerby's whale (Mesoplodon Sowerbyi). Transactions of the Royal Society of Edinburgh 26: 759-780.
Turner, W. (1885) The anatomy of a second specimen of Sowerby's whale (Mesoplodon bidens) from Shetland. Journal of Anatomy and Physiology 20 (Pt 1): 144-188.
Turner, W. (1893) The lesser rorqual (Balaenoptera rostrata) in the Scottish seas, with observations on its anatomy. Proceedings of the Royal Society of Edinburgh 1891-1892 (19): 36-75.
Turner, W. (1907) Note on a rare dolphin (Delphinus acutus), recently stranded on the coast of Sutherland. Proceedings of the Royal Society of Edinburgh 1905-1906 (26): 310-319.
Turner, W. (1910) XXXV. - The morphology of the manus in Platanista gangetica, the dolphin of the Ganges. Proceedings of the Royal Society of Edinburgh 1909-1910 (30): 508-514.
Turner, W. (1912) The marine mammals in the Anatomical Museum of the University of Edinburgh. (Macmillan and Co.; London): 1-205.

R e f e r e n c e s
344
Turner, W. M. (1909) XLI. - The skeleton of a Sowerby's whale (Mesoplodon bidens) stranded at St Andrews, and the morphology of the manus in Mesoplodon, Hyperoodon and the Delphinidæ. Proceedings of the Royal Society of Edinburgh 1908-1909 (29): 687-720.
Tyack, P. L., Johnson, M. P., Aguilar de Soto, N., et al. (2006) Extreme diving of beaked whales. Journal of Experimental Biology 209 (21): 4238-4253.
Tyson, E. (1680) Phocaena, on the anatomy of a porpess. (Benjamin Tooke; London): 1-48.
Uhen, M. D. (1998) Chapter 2: Middle to Late Eocene basilosaurines and dorudontines. The emergence of whales: evolutionary patterns in the origin of Cetacea. Advances in Vertebrate Paleobiology, Ed. J. G. M. Thewissen (Plenum Press; New York): 29-61.
Uhen, M. D. (2004) Form, function, and anatomy of Dorudon atrox (Mammalia, Cetacea): an archaeocete from the Middle to Late Eocene of Egypt Papers on Paleontology (University of Michigan; Michigan) 34: 1-222.
Uhen, M. D. (2007a) The earliest toothless mysticete: a chaeomysticetan from the early Oligocene Alsea Formation, Toledo, Oregon. Journal of Vertebrate Paleontology 27 (Suppl. 3) Abstracts of the 67. SVP meeting: 161A.
Uhen, M. D. (2007b) Evolution of marine mammals: back to the sea after 300 million years. Anatomical Record 290 (6): 514-522.
Uhen, M. D. (2008) New protocetid whales from Alabama and Mississippi, and a new cetacean clade, Pelagiceti. Journal of Vertebrate Paleontology 28 (3): 589-593.
Uhen, M. D. (2009a) Basilosaurids. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 91-94.
Uhen, M. D. (2009b) Dental morphology, Evolution of. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 302-307.
Uhen, M. D. (2010) The origin(s) of whales. Annual Review of Earth and Planetary Sciences 38: 189-219.
Uhen, M. D. (2014a) New material of Natchitochia jonesi and a comparison of the innominata and locomotor capabilities of Protocetidae. Marine Mammal Science 30 (3): 1029-1066.
Uhen, M. D. (2014b) New specimens of Protocetidae (Mammalia, Cetacea) from New Jersey and South Carolina. Journal of Vertebrate Paleontology 34 (1): 211-219.
Uhen, M. D. (2018) Basilosaurids and Kekenodontids. Encyclopedia of Marine Mammals. 3rd ed. (Academic Press; London): 78-80.
Uhen, M. D. and Gingerich, P. D. (2001) New genus of dorudontine archaeocete (Cetacea) from the middle�to�late Eocene of South Carolina. Marine Mammal Science 17 (1): 1-34.
Uhen, M. D., Pyenson, N. D., Thomas, D. V., et al. (2008) The oldest cetaceans from the southern hemisphere: new archaeocetes from the Pisco Basin of Southern Peru. Journal of Vertebrate Paleontology 28 (Suppl. 3) Abstracts of the 68. SVP meeting: 154A.
van Beneden, P.-J. and Gervais, P. (1880) Ostéographie des Cétacés vivant et fossiles. Ed. A. Bertrand (Libraire de la Société de Geographie; Paris): 1-658.
Van Valen, L. (1966) Deltatheridia, a new order of mammals. Bulletin American Museum of Natural History (American Museum of Natural History; New York) 132 (1): 1–126.

R e f e r e n c e s
345
Vautrin, Q., Lihoreau, F., Sambou, B., et al. (2020) From limb to fin: an Eocene protocetid forelimb from Senegal sheds new light on the early locomotor evolution of cetaceans. Palaeontology 63 (1): 51-66.
Vislobovoka, I. A. (2013) On the origin of Cetartiodactyla: comparison of data on evolutionary morphology and molecular biology. Paleontological Journal 47 (3): 321-334.
Waddell, P. J., Okada, N. and Hasegawa, M. (1999) Towards resolving the interordinal relationships of placental mammals. Systematic Biology 48 (1): 1–5.
Walker, J. A. (2004) Kinematics and performance of maneuvering control surfaces in teleost fishes. IEEE Journal of Oceanic Engineering 29 (3): 572-584.
Wall, W. P. (1983) The correlation between high limb-bone density and aquatic habits in Recent mammals. Journal of Paleontology 57 (2): 197-207.
Watson, A. G., Bahr, R. J. and Matheson, T. (2008a) Metacarpo-phalangeal anomalies in bottlenose dolphins (Tursiops truncatus). Aquatic Mammals 34 (1): 65-70.
Watson, A. G. and Fordyce, R. E. (1993) Skeleton of two minke whales, Balaenoptera acutorostrata, stranded on the southeast coast of New Zealand. New Zealand Natural Sciences 20: 1-14.
Watson, A. G., Kuo, T.-K., Yang, W.-C., et al. (2008b) Distinctive osteology of distal flipper bones of tropical bottlenose whales, Indopacetus pacificus, from Taiwan: mother and calf, calf with polydactyly. Marine Mammal Science 24 (2): 398-410.
Watson, A. G., Stein, L. E., Marshall, C., et al. (1994) Polydactyly in a bottlenose dolphin, Tursiops truncatus. Marine Mammal Science 10 (1): 93-100.
Watwood, S. L., Miller, P. J. O., Johnson, M. P., et al. (2006) Deep-diving foraging behaviour of sperm whales (Physeter macrocephalus). Journal of Animal Ecology 75 (3): 814-825.
Webb, P. W. (1997) Designs for stability and maneuverability in aquatic vertebrates: what can we learn? Proceedings of the 10th International Symposium for Unmanned Untethered Submersible Technology (Autonomous Undersea Systems Institute; Lee, New Hampshire): 86-108.
Weber, M. (1888) Anatomisches über Cetaceen. Morphologisches Jahrbuch 23: 616-652.
Weber, P. W., Howle, L. E., Murray, M. M., et al. (2009a) Lift and drag performance of odontocete cetacean flippers. Journal of Experimental Biology 212 (14): 2149-2158.
Weber, P. W., Murray, M. M., Howle, L. E., et al. (2009b) Comparison of real and idealized cetacean flippers. Bioinspiration and Biomimetics 4 (4): 1-12.
Weijuan, Z. K. Q. (1981) The myology of the baiji, Lipotes vexillifer (Odontoceti, Lipotidae). Acta Theriologica Sinica 1 (2) Part 1 - Skin mucles, forelimb muscles, tail and trunk muscles: 1-10.
Wellard, R., Lightbody, K., Fouda, L., et al. (2016) Killer whale (Orcinus orca) predation on beaked whales (Mesoplodon spp.) in the Bremer Sub-Basin, Western Australia. PLoS ONE 11 (12): 1-15.
Werth, A. J. (2006) Mandibular and dental variation and the evolution of suction feeding in Odontoceti. Journal of Mammalogy 87 (3): 579-588.
Whitehead, H. (2009) Sperm Whale, Physeter macrocephalus. Encyclopedia of Marine Mammals. 3rd ed., Eds. W. F. Perrin, B. G. Würsig, et al. (Academic Press; London): 1091-1097.
Whitt, A. D., Baran, M. A., Bryson, M., et al. (2015) First report of killer whales harassing sperm whales in the Gulf of Mexico. Aquatic Mammals 41 (3): 252-255.

R e f e r e n c e s
346
Wiley, D., Ware, C., Bocconcelli, A., et al. (2011) Underwater components of humpback whale bubble-net feeding behaviour. Behaviour 148 (5): 575-602.
Williams, T. M. (1983) Locomotion in the North American mink, a semi-aquatic mammal. II. The effect of an elongate body on running energetics and gait patterns. Journal of Experimental Biology 105 (1): 283-295.
Williams, T. M., Fuiman, L. A., Kendall, T., et al. (2015) Exercise at depth alters bradycardia and incidence of cardiac anomalies in deep-diving marine mammals. Nature Communications 6 (1): 1-9.
Woodward, B. (2006) Locomotory strategies, dive dynamics, and functional morphology of the mysticetes: using morphometrics, osteology, and DTAG data to compare swim performances in four species of baleen whales. (Colorado; Colorado State University) PhD thesis: 1-180.
Woodward, B. L., Winn, J. P. and Fish, F. E. (2006) Morphological specializations of baleen whales associated with hydrodynamic performance and ecological niche. Journal of Morphology 267 (11): 1284-1294.
Yu, X., Zhu, M. and Zhao, W. J. (2010) The origin and diversification of osteichthyans and sarcopterygians: rare Chinese fossil findings advance research on key issues of evolution. Bulletin of the Chinese Academy of Science 24: 71-75.
Zhu, M., Yu, X., Wang, W., et al. (2006) A primitive fish provides key characters bearing on deep osteichthyan phylogeny. Nature 441 (7089): 77-80.

347
Appendix 1. Caperea marginata Dimensions in mm if not otherwise stated Specimen MM2959 MM2904 Total length (contour) 1872 2961 Total length (direct) 1745 2745 Dimensions of the flipper soft tissue Flipper length of leading edge 266 316 Flipper length of trailing edge 215 226 Flipper greatest width(1) 62 77 Flipper greatest width(2) 132 186 (1) without axillary fold, (2) including axillary fold Dimensions of the articulated forelimb Forelimb length(1) (cartilage included) 285 342 Forelimb length(1) (bony elements only) 268 316 Brachial length (cartilage included) 164 207 Manus length (cartilage included) 123 136 Manus width(2) (cartilage included) 47 63 Manus width(2) (bony elements only) 40 53 Ratio of total length / forelimb 6.1 8.0 Ratio of forelimb length / manus width 6.1 5.4 Orientation: (1) proximo-distal, (2) cranio-caudal Dimensions of the scapula Scapula length(1) (bone) 148 237 Scapula length (+ cartilage) 198 310 Vertebral border length (bone, contour) 178 317 Scapula height(2) (bone) 73 125 Scapula height (+ cart. vert. border) 75 129 Scapula height (+ cart. vert. + glen. border) 82 136 Scapula length / height ratio (bone) 2.0 1.9 Scapula length / height ratio (+ cartilage) 2.6 2.4 Supraspinous fossa width 7 9 Acromion length 23 36 Acromion height 15 24 Coracoid length 13 27 Coracoid height 5 13 Angle of cranial vert. border to axis 47º - Angle of caudal vert. border to axis 41º - Angle of cranial border to axis 36º 30º Angle of caudal border to axis 27º 19º Angle of caudal border to neck 135º 113º Angle of cranial border to caudal border 119º 135º Angle of glenoid border (bone) to axis 21º 16º Angle of glenoid border (cart.) to axis 27º 22º

A p p e n d i x
348
Ratio of scapula length / height (bone) 2.0 1.9 Ratio of scapula length / height (cart.) 2.6 2.4 Ratio of supraspinous fossa to vert. border 1/25 1/35 Orientation: (1) cranio-caudal, (2) dorso-ventral Dimensions of the brachium Humerus total length 70 98 Humerus length (bone) 60 91 Humerus head height(1) 30 43 Humerus head width(2) 31 41 Humerus max. distal width 33 51 Humerus min. width 27 39 Radius total length 108 127 Radius length (bone) 95 110 Radius max. width 21 29 Radius min. width 19 26 Ulna total length ~ 106 122 Ulna length (bone) ~ 89 109 Ulna max. proximal width 22 38 Ulna max. distal width 18 23 Ulna min. width 9 14 Olecranon total length 54 82 Olecranon greatest width 32 42 Angle of cart. olecranon to ulnar shaft 28º 40º Ratio of radius length to max. width 5/1 4.5/1 Ratio of humerus to radius 2/3 3/4 Ratio ulnar length to max. prox. width 4/1 3/1 Ratio ulnar length to max. distal width 5/1 4.5/1 Ratio ulnar length to min. width 10/1 8/1 Orientation: (1) proximo-distal, (2) cranio-caudal, ~ approximate Dimensions of the manus Carpus length(1) 16 22 Carpus width(2) 48 63 Pisiform length(2) 16 15(4) Digit 2 total length(3) 59 66(4) Metacarpal 2 total length 23 28 Metacarpal 2 length (bone) 11 11 Digit 2 first total phalanx length 18 19 Digit 2 first phalanx length (bone) 8 7 Digit 3 total length 107 114 Metacarpal 3 total length 32 37 Metacarpal 3 length (bone) 17 18 Digit 3 first phalanx total length 25 32 Digit 3 first phalanx length (bone) 11 12 Digit 4 total length 99 108(4) Metacarpal 4 total length 25 30 Metacarpal 4 length (bone) 13 14 Digit 4 first total phalanx length 23 25 Digit 4 first phalanx length (bone) 9 10

A p p e n d i x
349
Digit 5 total length 57 59 Metacarpal 5 total length 23 26 Metacarpal 5 length (bone) 11 11 Digit 5 first total phalanx length 16 17 Digit 5 first phalanx length (bone) 4 2 Orientation: (1) proximo-distal, (2) cranio-caudal; (3) inclusive metacarpals, (4) estimate
2. Berardius arnuxii Dimensions in mm if not otherwise stated Specimen Purakaunui Tangimoana Hine Reti Left Left Right Right Total length (soft tissue) 8700 7000(1) 6500(1) (1) estimate Dimensions of the flipper (soft tissue) Flipper length of leading edge 980 780 783(1) 682(1) Flipper length of trailing edge 722 513 508(1) 503(1) Flipper length body wall to tip 843 651 655(1) 597(1) Flipper greatest width 317 259 256 222 Ratio body / flipper length 10.3 10.7(1) 10.7(1) 10.9(1) Ratio flipper length / flipper width 2.7 2.5 2.6 2.7 (1) estimate (incomplete specimen) Dimensions of the articulated pectoral limb (hard tissue) Forelimb length(1) 1024 796 815 714 Brachial length 556 453 459 410 Manus length(1) 471 347 361 307 Manus width(2) 271 215 215 185 Ratio body / forelimb length 8.5 8.8(3) 8.6(3) 9.1(3) Ratio forelimb length / width 3.8 3.7 3.8 3.9 Ratio brachium / manus 1.2 1.3 1.3 1.3 Orientation: (1) proximo-distal, (2) cranio-caudal; (3) body length estimate Dimensions of the scapula Scapula length(1) (bone) 553 - 421 356 Scapula length (+ cartilage) 603(3) - 472 404 Vert. border length (bone, contour) 634 - 517 427 Length at level of ssp fossa 566 - 463 343 Scapula height(2) (bone) 361 - 276 238 Scapula height (+ cart. border) 401(3) - 316 271(3) Scapula height (+ border + glenoid) 411(3) - 324 281(3) Supraspinous fossa width 42 - 20 34(5) Acromion length 175 - 113 92 Acromion greatest height 94 - 148(4) 58 Acromion least height 62 - 73 56 Coracoid length 155 - 125 89 Coracoid greatest height 71 - 58 50 Coracoid least height 45 - 35 39 Angle of cranial border to axis 35º - 35º 37º

A p p e n d i x
350
Angle of caudal border to axis 41º - 40º 35º Angle of caudal border to neck 150º - 127º 143º Angle of caudal border to neck: 136º (Flower, 1872) and 147º (Benke,1993) Angle of cranial / caudal border 105 - 105 108 Angle of glen. border / axis (bone) 21 - 30 26 Angle of glen. border / axis (cart.) 18 - 25 25 Ratio of sca. length / height (bone) 1.5 - 1.5 1.5 Ratio of sca. length / height (cart.) 1.5 - 1.5 1.5 Ratio of ssp fossa / vert. border (6) 13.5 - 23 10 Orientation: (1) cranio-caudal, (2) dorso-ventral, (3) estimate, (4) approximately, (5) greatest width, (6) length at level of ssp fossa Dimensions of the brachium Humerus total length 284 251 255 224 Humerus length (diaphysis) 210 185 191 161 Humerus head height(1) 62 63 64 56 Humerus head width(2) 119 118 119 103 Humerus max. distal width 133 117 118 109 Humerus min. width 101 96 98 96 Radius total length 298 232 232 206 Radius length (diaphysis) 255 198 195 172 Radius max. width 91 76 78 74 Radius min. width 78 67 70 66 Ulna total length 279 203 206 194 Ulna length (diaphysis) 245 177 176 162 Ulna max. proximal width 137 108 107 85 Ulna max. distal width 84 64 68 60 Ulna min. width 46 41 42 36 Olecranon total length ~ 95 - - - Olecranon greatest width ~ 65 - - - Angle of olecranon to ulnar shaft ~ 33 - - - Ratio of humerus to radius 0.9 1.1 1.1 1.1 Ratio of rad. length / max. width 2.8 2.6 2.5 2.3 Ratio ul. length / max. prox. width 2.0 1.9 1.9 2.3 Ratio ul. length / max. dist. width 3.3 3.2 3.0 3.2 Ratio ulnar length to min. width 6.1 5.0 4.9 5.4 Orientation: (1) proximo-distal, (2) cranio-caudal, ~ approximate Dimensions of the manus Carpus length(1) 79 80 81 52 Carpus width(2) 178 159 168 139 Pisiform length(2) - 12 14 - Digit 1 total length(3) 114 70 82 56 Metacarpal 1 total length 91 52 56 56 Metacarpal 1 length (diaphysis) 57 36 38 35 Digit 1 first phalanx total length 22 18 23 - Digit 1 first ph. length (diaphysis) - - - - Digit 2 total length 402 287 297 248 Metacarpal 2 total length 122 91 95 78 Metacarpal 2 length (diaphysis) 85 70 69 59 Digit 2 first phalanx total length 97 67 69 57 Digit 2 first ph. length (diaphysis) 59 45 47 37 Digit 3 total length 365 260 263 245

A p p e n d i x
351
Metacarpal 3 total length 135 95 98 88 Metacarpal 3 length (diaphysis) 98 76 76 70 Digit 3 first phalanx total length 91 66 67 54 Digit 3 first ph. length (diaphysis) 56 44 44 37 Digit 4 total length 290 206 210 185 Metacarpal 4 total length 113 79 82 71 Metacarpal 4 length (diaphysis) 82 63 75 57 Digit 4 first total phalanx length 80 59 60 50 Digit 4 first ph. length (diaphysis) 51 38 38 35 Digit 5 total length 223 169 166 157 Metacarpal 5 total length 103 79 76 71 Metacarpal 5 length (diaphysis) 72 56 55 50 Digit 5 first total phalanx length 64 51 51 42 Digit 5 first ph. length (diaphysis) 42 33 31 28 Orientation: (1) proximo-distal, (2) cranio-caudal; (3) inclusive metacarpals, (4) estimate
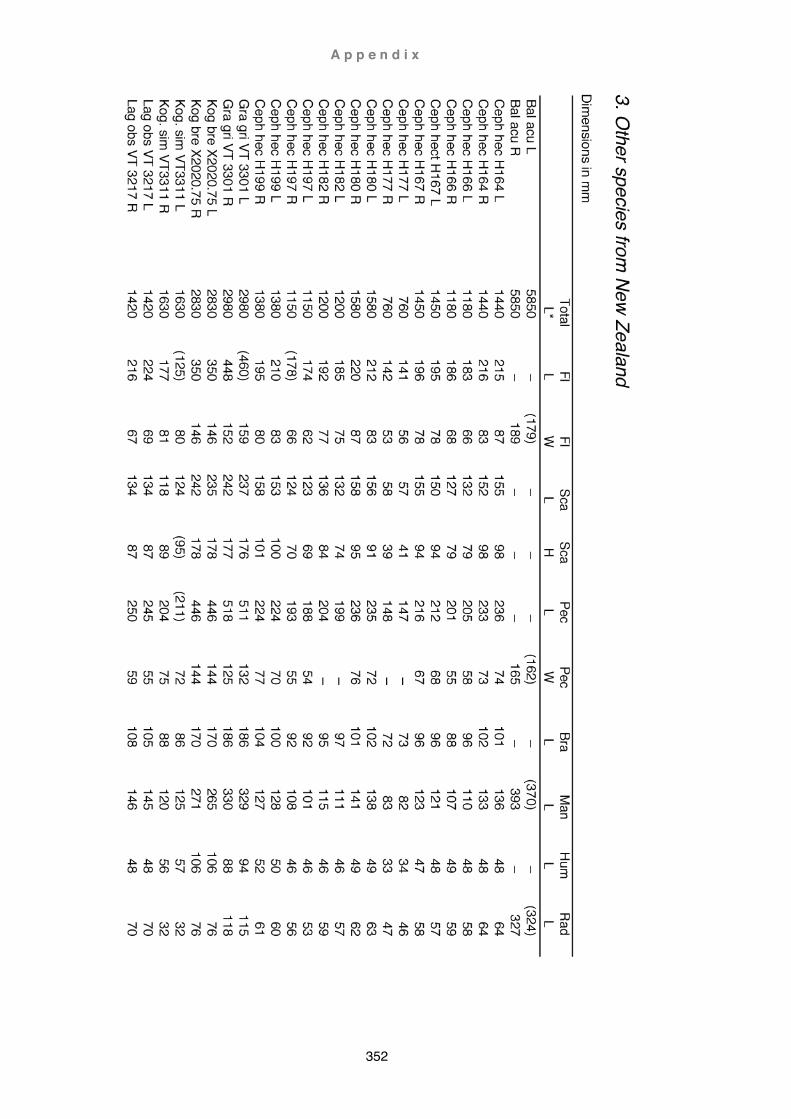
A p p e n d i x
352

A p p e n d i x
353




















