
Dynein Antagonizes Eg5 by Crosslinking and Sliding Antiparallel Microtubules
Upload
independentCategory
view
0download
0
# 2007 The Authors
Journal compilation # 2007 Blackwell Publishing Ltd
doi: 10.1111/j.1600-0854.2007.00636.xTraffic 2007; 8: 1503–1520Blackwell Munksgaard
A Functional Dynein–Microtubule Network Is Requiredfor NGF Signaling Through the Rap1/MAPK Pathway
Chengbiao Wu1,*, Alfredo Ramirez1,
Bianxiao Cui2, Jianqing Ding1,
Jean-Dominique M. Delcroix1,
Janice S. Valletta1, Jia-Jia Liu1, Yanmin Yang1,
Steven Chu2 and William C. Mobley1
1Department of Neurology and Neurological Sciences,Stanford University School of Medicine, Stanford,CA 94305, USA2Lawrence Berkeley National Laboratory, andDepartments of Physics and Molecular andCellular Biology, University of California,Berkeley, CA 94720, USA*Corresponding author: Chengbiao Wu,[email protected]
Rap1 transduces nerve growth factor (NGF)/tyrosine
receptor kinase A (TrkA) signaling in early endosomes,
leading to sustained activation of the p44/p42 mitogen-
activated protein kinases (MAPK1/2). However, themech-
anisms by which NGF, TrkA and Rap1 are trafficked to
early endosomes are poorly defined. We investigated
trafficking and signaling of NGF, TrkA and Rap1 in PC12
cells and in cultured rat dorsal root ganglion (DRG)
neurons. Herein, we show a role for both microtubule-
and dynein-based transport in NGF signaling through
MAPK1/2. NGF treatment resulted in trafficking of NGF,
TrkA and Rap1 to early endosomes in the perinuclear
region of PC12 cells where sustained activation of
MAPK1/2 was observed. Disruption of microtubules with
nocodazole in PC12 cells had no effect on the activation of
TrkA and Ras. However, it disrupted intracellular traffick-
ing of TrkA and Rap1. Moreover, NGF-induced activation
of Rap1 and sustained activation of MAPK1/2 were mark-
edly suppressed. Inhibition of dynein activity through
overexpression of dynamitin (p50) blocked trafficking of
Rap1 and the sustained phase of MAPK1/2 activation in
PC12 cells. Remarkably, even in the continued presence of
NGF, mature DRG neurons that overexpressed p50
became atrophic and most (>80%) developing DRG neu-
rons died. Dynein- and microtubule-based transport is
thus necessary for TrkA signaling to Rap1 and MAPK1/2.
Key words: axon, DRG, dynein, endosome, MAPK, micro-
tubule, NGF, Rap1, retrograde transport, TrkA
Received 23 May 2007, revised and accepted for publica-
tion 7 August 2007, uncorrected manuscript published
online 9 August 2007, published online 6 September 2007
A compelling body of evidence points to the importance
of retrogradely transported neurotrophin (NT) signals for
neuronal survival and function (1–12). The mechanism(s)
underlying retrograde transport of NT signaling has been
explored (1,6,8,10,12). Under the ‘signaling endosome’
hypothesis, nerve growth factor (NGF) binds to and
activates its receptor tyrosine receptor kinase A (TrkA) at
the axon (AX) terminal, and the NGF–TrkA complex is
internalized through clathrin-mediated and -independent
pathways (13–20). The signaling endosome thus formed
is retrogradedly transported to the cell body (CB) (21).
Despite much support for the hypothesis (21–23), import-
ant questions remain as to how retrograde signals are
trafficked (11,19,24). Indeed, some studies suggested that
retrograde axonal transport of neither the NGF–TrkA
complex nor the phosphorylated form of TrkA, pTrkA, is
required to convey a survival signal to the CB (25,26).
Recent studies point to an important role for dynein-based
transport of NT signals. Trk receptor appears to be asso-
ciated with members of the cytoplasmic dynein family
during retrograde transport. Both the 14 and the 74 kD
chains of dynein could be coprecipitated with Trk in brain
lysates, and both subunits accumulated together with Trk
distal to a ligature site in the rat sciatic nerve (27). Proximity
of TrkA to dynein has been shown by both confocal
imaging (28) and immunoelectron microscopy (29). In
addition, dynein-based transport was shown to be required
for retrograde trafficking of activated Trks and thus played
a critical role in promoting survival of NT-dependent DRG
neurons (30). A role for intact microtubules in transporting
NGF signals in sciatic nerve was also recently shown (28).
Taken together with other reports (31,32), a strong case
can be made for the importance of dynein and intact
microtubules for retrograde transport of NT signals.
Beyond a role for moving the signal within AXs, we
envision the possibility that trafficking events also serve
to regulate the nature of the signals generated, an issue
addressed herein.
NGF induces prolonged activation of the mitogen-activated
protein kinases (MAPK1/2) signaling pathway. While Ras
mediates the transient phase of MAPK1/2 activation,
Rap1, a small guanosine triphosphatase, contributes to
the sustained activation of theMAPK1/2 signaling pathway
(33), which is necessary for NGF-induced differentiation
in PC12 cells (34). Following NGF treatment, Rap1 was
shown to be concentrated and activated in the perinuclear
region (35), and activated Rap1 was found in a long-lasting
complex with both activated TrkA and activated MAPK1/2
(pMAPK1/2) in an early-endosome-enriched fraction (36).
We used PC12 cells and DRG neurons to pursue further
the mechanism of NGF trafficking and signaling. We found
www.traffic.dk 1503
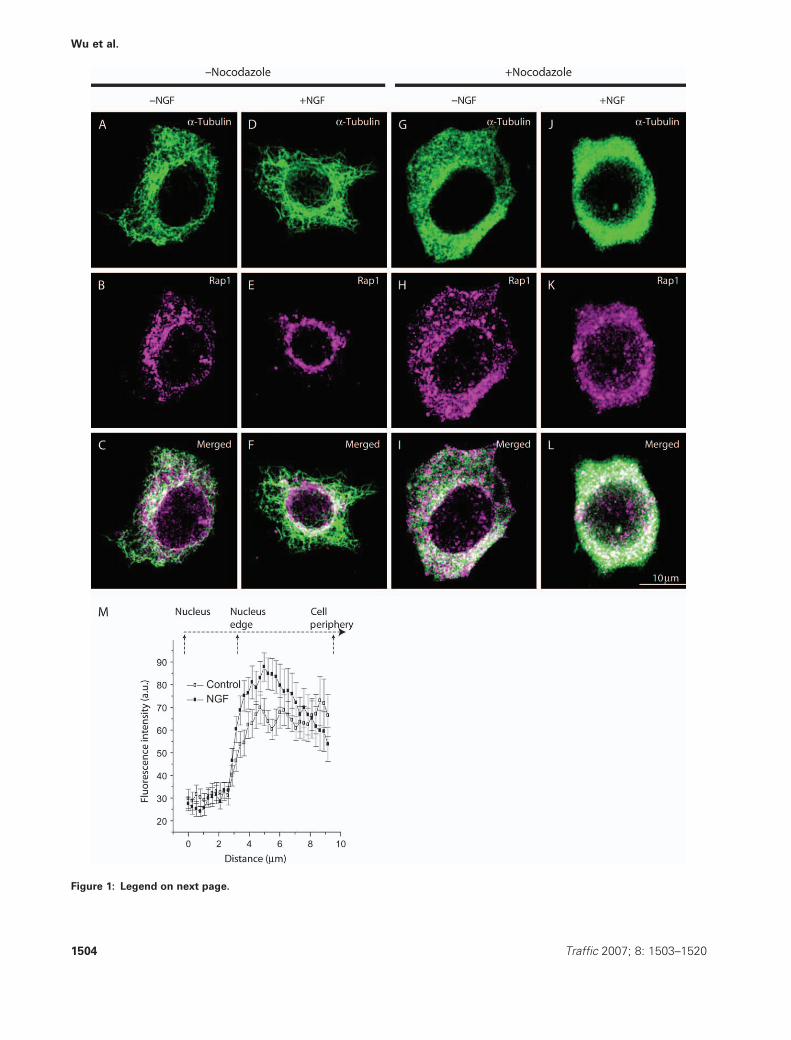
Figure 1: Legend on next page.
1504 Traffic 2007; 8: 1503–1520
Wu et al.

that following NGF treatment, both NGF–TrkA complexes
and Rap1 moved to the perinuclear region of PC12 cells
where they were present in early endosomes in which
persistent activation of MAPK1/2 was also registered. For
both TrkA and Rap1, trafficking to the perinuclear region
was prevented by the disruption of microtubules with
nocodazole. Notably, nocodazole suppressed the activa-
tion of Rap1 and the ensuing persistent activation of
MAPK1/2 without interfering with TrkA or Ras activation.
Disrupting the dynein–dynactin complex by overexpresss-
ing p50, dynamitin (37), also blocked trafficking of Rap1 to
the perinuclear region and inhibited activation of MAPK1/2.
Using a lentiviral system, we showed that overexpressing
p50 inhibited retrograde transport of NGF and activated
TrkA (pTrkA) and NGF-induced activation of MAPK1/2 in
DRG neurons. Even for neurons whose CBs were bathed
in NGF, the consequence of p50 overexpression was the
death of immature neurons and atrophy of mature neur-
ons. Our findings suggest that dynein and microtubules
play an essential role in mediating not just the retrograde
trafficking of activated TrkA but also activation of Rap1
signaling to MAPK1/2 in support of neuronal differentiation
and survival.
Results
Microtubule depolymerization prevents NGF-induced
translocation of Rap1 to the perinuclear region
Microtubules, comprising a- and b-tubulin, form an elab-
orate network that extends throughout the cytoplasm in
virtually all eukaryotic cells. One of the most important
functions of the microtubule is to regulate the location of
membrane-bound organelles. We reasoned that disruption
of the microtubule network might alter NGF-induced
trafficking events. First, we established that NGF treat-
ment resulted in accumulation of NGF, TrkA, Rap1 and
activated MAPK1/2 in early endosomes in the perinuclear
region in PC12 cells (Figures S1 and S2). We then
examined the microtubule network in PC12 cells using
confocal microscopy. Cells, grown on glass coverslips,
were serum starved overnight prior to treatment. The cells
were pretreated with either the nocodazole vehicle
(�nocodazole) or 5 mg/mL nocodazole (þnocodazole) for
2 h at 378C. Cells were then treated with either the NGF
vehicle or 50 ng/mL NGF (þNGF) for 30 min. The samples
were prepared for indirect immunofluorescence and ana-
lyzed using confocal microscopy. Rap1-positive structures
(magenta) and the microtubule network (green) were
visualized and analyzed. (Please note that red color in all
two-color images is presented as magenta and colocaliza-
tion with green signal will be therefore denoted as white
color hereafter.) In cells pretreated with the vehicle only
(�nocodazole), microtubules were distributed throughout
the cell and the Rap1-positive puncta were seen dispersed
throughout the cytoplasm (Figure 1A–C). Treatment with
NGF for 30 min had little or no effect on the distribution of
a-tubulin (Figure 1A versus 1D). However, NGF treatment
resulted in trafficking of Rap1 to the perinuclear region
(Figure 1E versus 1B and Figure 1F versus 1C). The
increase of Rap1 signal in the perinuclear region in NGF-
treated cells was quantified in arbitrary unit (a.u.) of
fluorescence intensity, and the results are presented in
Figure 1M.
Pretreatment with 5 mM nocodazole at 378C for 2 h
resulted in a very different outcome. There was complete
disintegration of the microtubule network in approxi-
mately 95% of cells (Figure 1;þnocodazole). The staining
pattern for a-tubulin changed from the complex network
seen in normal cells (Figure 1A,D) to a pattern that was
diffused and evenly distributed (Figure 1G,J). The NGF
vehicle caused no obvious change in the pattern for Rap1
immunostaining in cells pretreated with nocodazole, that
is the Rap1-positive structures remained dispersed (Fig-
ure 1H versus 1B). However, Rap1 was no longer present
in the perinuclear region in response to NGF treatment
(Figure 1K versus 1E). Indeed, the pattern of Rap1 immu-
nostaining showed little, if any, difference from that seen
in cells treated with the NGF vehicle alone (Figure 1K
versus 1H and Figure 1L versus 1I). These studies sug-
gested that an intact microtubule network is required for
trafficking of Rap1-containing organelles to the perinuclear
region.
Disruption of the microtubule network blocks
translocation of TrkA to the perinuclear region
To examine whether TrkA trafficking was impaired when
microtubules were disrupted, PC12 cells were pretreated
with either 5 mM nocodazole (þnocodazole) or its vehicle
Figure 1: Nocodazole pretreatment blocked NGF-induced trafficking of Rap1 to the perinuclear region. PC12 cells were prepared
as described in Materials and Methods. Following pretreatment with either the nocodazole vehicle alone (�nocodazole) (A–F) or 5 mM
nocodazole (þnocodazole) (G–L) for 2 h, cells were treatedwith either NGF (50 ng/mL) (þNGF) (D–F, J–L) or the NGF vehicle (�NGF) (A–C,
G–I). Indirect immunofluorescence was carried out using confocal microscopy. Tubulin (green) and Rap1 (magenta) were stained with
a mouse antibody to a-tubulin and a rabbit antibody to Rap1, respectively. An Alexa 488 goat anti-mouse IgG conjugate was used to
visualize the microtubules, and an Alexa 568 goat anti-rabbit IgG conjugate was used to detect Rap1. Colocalization of antigens is denoted
by the white signal. Twenty-five cells were examined in each condition, and a typical representative image was shown. The size of the
collection box for all panels is 30 � 30 mm. In (M), the average line fluorescence intensity of Rap1 staining in 16 control cells and 12 NGF-
treated cells was measured. Because cells were different in size, we chose to measure the line fluorescence intensity of 10 mm distance
(2.5 mm from the nuclear edge toward the center of nucleus and 7.5 mm from the nuclear edge toward the cell periphery). The results
show that NGF treatment resulted inapproximately 30% increase in average fluorescence intensity (a.u.) in the perinuclear region
compared with that in the control groups. Standard errors are also presented.
Traffic 2007; 8: 1503–1520 1505
Dynein-/Microtubule-Mediated Trafficking of TrkA for Rap1 Signaling

(�nocodazole) for 2 h. Cells were then treated for 30 min
with either 50 ng/mL NGF (þNGF) or NGF vehicle (�NGF)
(Figure 2). The samples were prepared for confocal
microscopy to analyze the distribution of TrkA (Figure 2).
A corresponding transmitted light imagewas also captured
and superimposed to show the outline of the cell (Fig-
ure 2). In the absence of nocodazole (�nocodazole),
treatment with NGF resulted in the expected increase in
TrkA immunostaining in the perinuclear region (Figure 2B
versus 2A). However, in cells pretreated with nocodazole,
TrkA-positive puncta remained dispersed following NGF
treatment (Figure 2D versus 2C).
To confirm that TrkA trafficking was altered in cells
pretreated with nocodazole, we examined the distribution
of TrkA using subcellular fractionation on OptiPrep step
gradients of 5:10:15:20:25% (36). Serum-starved PC12
cells were pretreated with either 5 mM nocodazole (Fig-
ure 3B) or its vehicle (Figure 3A) prior to treatment with
either NGF (þNGF) or the NGF vehicle (�NGF). Cells were
then rinsed and prepared for subcellular fractionation.
Membrane fractions were collected from the four inter-
faces, and proteins were precipitated, washed and air-
dried. The samples were analyzed using SDS–PAGE/
immunoblotting. As shown previously, plasma mem-
brane proteins such as the receptor for epidermal growth
factor and Ras were concentrated in the light fractions
(fractions 1 and 2), while the early endosomal markers
Rab5 and EEA1 were enriched in the heavy fraction
(fraction 4) (36).
In cells that were pretreated with the nocodazole vehicle
(Figure 3A), NGF treatment did not alter the fractionation
of EEA1, Rab5B, Rap1 or MAPK1/2; all were enriched in
fraction 4 (Figure 3A). Without NGF treatment, TrkA (*)
was mostly detected in fraction 2, with a small amount in
fraction 1 (Figure 3A; �NGF). Following NGF treatment,
TrkA (*) was predominantly seen in fraction 4 (Figure 3A;
þNGF). The phosphorylated form of TrkA (pY490), pTrkA
(indicated by arrow), was detected in the same fraction, as
was pMAPK1/2 (Figure 3A; þNGF). The pattern for the
74 kD dynein intermediate chain (DIC74) was also exam-
ined. It was present in fractions 1 and 4 in the control
sample (Figure 3A; �NGF). NGF treatment caused the
disappearance of DIC74 from fraction 1; instead, it was
now more concentrated in fraction 4 (Figure 3A; þNGF).
These findings are consistent with the earlier results
showing that a significant amount of TrkA is present in
a light buoyant-density fraction prior to NGF treatment (38)
and that NGF treatment for 30 min resulted in the move-
ment of activated TrkA together with pMAPK1/2 to the
heavier, early-endosome-containing fraction (20,36).
The fractionation patterns for Rab5B, Rap1 and MAPK1/2
were not affected by pretreatment with nocodazole (Fig-
ure 3B versus 3A). This treatment did alter the fractionation
Figure 2: Nocodazole pretreatment dis-
rupted NGF-induced intracellular traffick-
ing of TrkA. Following pretreatment with
either the nocodazole vehicle alone
(�nocodazole) (A, B) or 5 mM nocodazole
(þnocodazole) (C, D) for 2 h, PC12 cells were
treated with either NGF (50 ng/mL) (þNGF) (B,
D) or the NGF vehicle (�NGF) (A, C). Cells were
then rinsed, fixed with 100% methanol and
processed for indirect immunofluorescence
using confocal microscopy as described in
Materials and Methods. TrkA (red) was stained
with a mouse antibody to Trk (B3) and with an
Alexa 568 goat anti-mouse IgG conjugate.
A corresponding image from the transmitted
light detector was also captured and super-
imposed onto the fluorescence image of TrkA
to illustrate the outline of the cell. Twenty-five
cells were examined in each condition, and
a typical representative image was shown. The
size of the collection box for all panels is
30 � 30 mm.
1506 Traffic 2007; 8: 1503–1520
Wu et al.

pattern for DIC74 and TrkA following NGF treatment. In
cells treated with nocodazole but not with NGF, DIC74was
present in fraction 3 and more concentrated in fraction 4
(Figure 3B; �NGF). Most TrkA (*) was detected in fraction
2, with a small amount in fraction 1 (Figure 3B; �NGF).
However, when nocodazole-pretreated cells were treated
with NGF, DIC74 was captured in fraction 1 as well as in
fraction 3 (Figure 3B; þNGF). Most TrkA (*) as well as
most pTrkA (indicated by arrow) were now present in
fraction 3 but not in fraction 4 (Figure 3B; þNGF). It is
noteworthy that activation of TrkA (arrows in Figure 3A,B)
was not affected by nocodazole, suggesting that NGF-
induced TrkA activation at the plasma membrane or initial
internalization of activated TrkA was not hindered. Further-
more, nocodazole pretreatment markedly inhibited NGF-
induced activation of MAPK1/2 (i.e. pMAPK1/2) in fraction
4 (Figure 3B versus 3A). Thus, depolymerization of micro-
tubules alters the trafficking of activated TrkA and impairs
its downstream signaling.
We also treated PC12 cells with biotinylated NGF (BtNGF)
and isolated fractions 1 and 4. Lysates of these fractions
were then incubated with streptavidin–agarose beads to
capture BtNGF and its binding proteins. The protein
complex was analyzed by SDS–PAGE and immunoblotting
with pTrkA antibody (pY490). Lysates of fraction 4 from
BtNGF-treated cells, but not from vehicle-treated cells,
contained pTrkA bound to BtNGF (Figure 3C). As a control,
pTrkA was not recovered from fraction 1 of either BtNGF-
or vehicle-treated cells (Figure 3C). We conclude that
NGF–TrkA complexes are present in early endosomes.
To show if pMAPK1/2 activated by NGF in fraction 4 were
accessible to its downstream signaling molecules, we
carried out an in vitro kinase phosphorylation assay using
recombinant Elk-1, a substrate immediate downstream to
pMAPK1/2 in NGF signaling cascade. Fraction 4 from cells
treated with NGF containing pMAPK1/2 was capable of
Figure 3: Nocodazole pretreatment disrupted the intracellu-
lar trafficking of TrkA as analyzed using a step gradient of
OptiPrep. PC12 cells were starved and pretreated with either
vehicle (A, control) or 5 mM nocodazole (B) for 2 h and were
subsequently treated with NGF (þNGF, 50 ng/mL) or the NGF
vehicle (�NGF) for 30 min. Cells were rinsed and homogenized.
The homogenates were centrifuged to remove nuclei. The result-
ing supernatants were fractionated by centrifugation using the
step-density gradient system as described in Materials and
Methods. Four membrane fractions (1, 5:10%; 2, 10:15%; 3,
15:20% and 4, 20:25%) were collected from the indicated
interface, and proteins from each fraction were precipitated in
7% trichloroacetic acid and washed in acetone. The pellets were
then dried and boiled in SDS–PAGE loading buffer. The proteins
were analyzed using SDS–PAGE and immunoblotting. The blots
were independently probed with the indicated antibodies. In
a separate experiment, following treatment with either the
NGF vehicle (�) or 50 ng/mL BtNGF (þ) for 30 min, cells were
fractionated as in (A). Fractions 1 and 4 from the vehicle- and
BtNGF-treated samples were lysed and precipitated with
streptavidin–agarose beads as described inMaterials andMethods.
The precipitates were washed, separated on SDS–PAGE and
blotted with a rabbit antibody to pTrkA (C). In (D), membrane
fractions (fraction 4) were collected from both NGF-treated (þNGF)
and vehicle-treated (�NGF) samples and were reconstituted into an
in vitro Elk-1 phosphorylation assay system. Phospho-Elk-1 and Elk-
1 were detected using specific antibodies. All blots were visualized
using SuperSignal, and the results shown are representative of
at least three independent experiments.
Traffic 2007; 8: 1503–1520 1507
Dynein-/Microtubule-Mediated Trafficking of TrkA for Rap1 Signaling

phosphorylating Elk-1 (Figure 3D; þ), while fraction 4 from
cells that were treated with NGF vehicle did not contain
pMAPK1/2 and was incapable of phosphorylating Elk-1
(Figure 3D;�). Therefore, a population of early endosomes
is capable of transmitting the NGF signal.
Disruption of the microtubule network suppresses
Rap1 and MAPK1/2 signaling
To pursue further the idea that aberrant trafficking of TrkA
and Rap1 would compromise activation of the MAPK1/2
signaling pathway, we analyzed the effects of nocodazole
pretreatment on the activation of Ras and Rap1. We first
confirmed that TrkA was activated to a level similar to that
in control samples when microtubules were disrupted.
Serum-starved PC12 cells were pretreated with either
vehicle (�nocodazole) or 5 mM nocodazole (þnocodazole)
prior to treatment with NGF or the NGF vehicle (Figure 4).
Cells were rinsed, lysed and centrifuged. The supernatant
was immunoprecipitated with a specific antibody to
Trk (MCTrk). The immunoprecipitate was analyzed using
SDS–PAGE/immunoblotting. As shown in Figure 4A, no-
codazole pretreatment did not significantly alter the mag-
nitude or the duration of TrkA activation induced by NGF.
Samples were also prepared to assay the active forms (i.e.
GTP-bound form) of Ras and Rap1 using an established
protocol (36). As shown in Figure 4B, nocodazole pre-
treatment had little effect on GTP loading of Ras. In
contrast, nocodazole significantly suppressed the activa-
tion of Rap1 (Figure 4B). Consistent with this result, there
was also suppression of NGF-induced sustained activation
of MAPK1/2. While the early phase of MAPK1/2 activation
was unaffected by nocodazole, the activation of MAPK1/2
seen at 30 min of NGF treatment was markedly reduced
(Figure 4B). The levels for total MAPK1/2 were not
affected by nocodazole. We also examined activation of
the phosphatidylinositide 3-kinase (PI3K)/Akt pathway fol-
lowing NGF treatment. Nocodazole pretreatment caused
partial suppression of this pathway, as evidenced by
a decrease in the level of phosphorylated Akt (pAkt,
Ser473) but not total Akt (Figure 4B). Thus, disrupting the
microtubule network suppressed NGF-induced activation
of Rap1 signaling, the sustained phase of MAPK1/2
activation and activation of PI3K/Akt.
The overexpression of p50 blocks NGF-induced
retrograde transport of Rap1 and abolishes the
sustained phase of MAPK1/2 activation
DIC74 was found to be associated with the Trk receptor in
mouse brain lysates (27). We confirmed the association of
DIC74 with TrkA in PC12 cells by immunoprecipitation
(data not shown). Thus, cytoplasmic dyneins might play
a role in intracellular trafficking of Trk (30). To examine
what role dynein plays in retrograde trafficking of TrkA and
Rap1 signaling, we inhibited the activity of the dynein–
dynactin complex by overexpressing the 50 kD subunit of
dynactin and dynamitin (p50) (37,39). The overexpression
of p50 causes dissociation of the microtubule- and cargo-
binding subunits of the dynein–dynactin complex, thus
serving to disrupt dynein-based transport (39,40). In a trans-
genic mouse model, p50 overexpression led to marked
inhibition of retrograde axonal transport (41).
We reasoned that p50 overexpression in PC12 cells might
block NGF-induced retrograde trafficking of TrkA and Rap1
and, as a result, inhibit the sustained activation ofMAPK1/2.
To test this idea, we transiently transfected PC12 cells
with a pcDNA3 construct that contained the full-length
human p50 complementary DNA (cDNA) sequence. We
used immunostaining to examine the effects of p50 over-
expression. Forty-eight hours posttransfection, PC12 cells
were serum starved for 16 h prior to receiving treatment
Figure 4: Nocodazole pretreatment selectively inhibited
Rap1 activation and the sustained phase of MAPK1/2 acti-
vation induced by NGF. PC12 cells were starved and treated
with either the vehicle (�nocodazole) or 5 mM nocodazole
(þnocodazole) for 2 h prior to treatment with NGF (50 ng/mL) or
NGF vehicle (0 min) for the indicated time intervals. In (A), TrkA
was immunoprecipitated from all samples with a mouse antibody
to Trk (MCTrk) and was analyzed using SDS–PAGE/immunoblot-
ting. TrkA was detected using B3, and pTrkA was revealed using
a rabbit antibody to pTrkA (pY490). In (B), the level of RasGTP and
Rap1GTP was analyzed as described in Materials and Methods. In
addition, cells were washed and then lysed in RadioImmuno-
Precipitation Assay (RIPA) buffer. Ten micrograms of proteins
from the resulting supernatant of each sample was separated on
SDS–PAGE and analyzed by immunoblotting with a mouse anti-
body to the phosphorylated MAPK1/2. The blot was stripped and
reprobed for the total amount of MAPK1/2 to show equal loading
of protein samples. Samples from the lysates were also immuno-
blotted with a rabbit antibody to phosphorylated Akt (Ser473) and
reprobed with a rabbit antibody to total Akt. The results shown are
representative of at least three independent experiments.
1508 Traffic 2007; 8: 1503–1520
Wu et al.

with either the NGF vehicle (�NGF) or the NGF (þNGF,
50 ng/mL for 30 min) (Figure 5). Cells were then pro-
cessed for immunostaining using a mouse monoclonal
antibody to p50 and a rabbit polyclonal antibody to Rap1.
To better illustrate the perinuclear region, we also co-
stained with Hoechst 33258 to visualize the nucleus by
epifluorescence (Figure 5A/b,d,f,h). In untransfected cells
(Figure 5A/a,b,e,f), NGF induced accumulation of Rap1
Figure 5: The overexpression of p50 inhibited NGF-induced translocation of Rap1 and pMAPK1/2 signals to the perinuclear
region. PC12 cells were transiently transfected with pcDNA3 plasmids that contained the full-length human dynamitin cDNA sequence for
48 h as described inMaterials and Methods. Cells were then reseeded on glass coverslips and starved. Cells were treated with either the
NGF vehicle (�NGF) or NGF (þNGF, 50 ng/mL for 30 min). In both (A) and (B), p50 (green) was immunostained with a specific mouse
antibody (1/500 dilution) and detected with a secondary goat anti-mouse IgG–Alexa 488 conjugate. Rap1 (magenta) in (A) was stained with
a specific rabbit antibody and visualizedwith a secondary goat anti-rabbit IgG–Alexa 568 conjugate. The sample in (A) (b, d, f and h) was also
co-stained with Hoechst 33258 (blue) to mark the nucleus. The resulting samples were captured and analyzed using a Nikon E800
epifluorescence microscope. Similarly, the pMAPK1/2 signals (magenta) in (B) were stained with a specific rabbit antibody to pMAPK1/2
and visualized with a secondary goat anti-rabbit IgG–Alexa 568 conjugate. The p50 (green) was stained as in (A). The resulting samples
were analyzed using a confocal microscope (B). Typically, 25 cells were examined in each condition, and a typical representative image is
shown. The size of the collection box is 60 � 60 mm for (A) and 30 � 30 mm for (B).
Traffic 2007; 8: 1503–1520 1509
Dynein-/Microtubule-Mediated Trafficking of TrkA for Rap1 Signaling

signal in the perinuclear region. In cells that overexpressed
p50 (Figure 5A/c,d,g,h), NGF treatment failed to cause con-
centration of Rap1 in the perinuclear region (Figure 5A/b–f
versus 5A/d–h). The effect was specific for p50 because
overexpression of enhanced green fluorescent protein
(EGFP) did not affect NGF-induced trafficking of Rap1
(data not shown).
We next investigated the sustained phase of MAPK1/2
activation in cells that transiently overexpressed p50.
Without NGF treatment (�NGF; Figure 5B), the baseline
level for pMAPK1/2 signal in transfected (Figure 5B/c) cells
was similar to that in nontransfected (Figure 5B/a) cells
(Figure 5B/d versus 5B/b). Treatment of nontransfected
(Figure 5B/e) cells with NGF for 30 min showed a marked
increase in pMAPK1/2 in the perinuclear region (Figure 5B/f
versus 5B/b). Similarly, a marked increase in pMAPK1/2
was also seen in cells that were transfected with a plasmid
vector expressing EGFP, pEGFP (data not shown). In
contrast, NGF treatment failed to elicit this response when
p50 was overexpressed (Figure 5B/g,h versus 5B/e,f).
Thus, inhibition of cytoplasmic dynein blocked NGF-
induced retrograde transport of Rap1 and abolished the
sustained phase of MAPK1/2 activation.
The overexpression of p50 blocks retrograde
transport of the NGF–pTrkA complex in
mature DRG neurons in culture
We next examined the trafficking of NGF signals in
cultured neurons. Rat embryonic DRGs were cultured on
collagen-coated cover glasses for 21 days (DIV-21) as
described in Materials and Methods. At this point, the
cultures contain large, mature neurons with well-devel-
oped AXs. At least 94% of neurons were judged to be NGF
responsive as evidenced by immunopositivity for TrkA
(data not shown). Of note, these neurons no longer require
NGF for survival (42). Following NGF deprivation for 24 h,
cells were treated with either 100 ng/mL NGF (Figure 6
B,D,F) or NGF vehicle (Figure 6A,C,E) and were prepared
for immunostaining. NGF treatment resulted in redistribu-
tion of both Rap1 and TrkA to the perinuclear region, where
they significantly colocalized (Figure 6A versus 6B). Con-
sistent with our results for PC12 cells, NGF induced
activation of MAPK1/2 and the signal for pMAPK1/2 over-
lapped with that for Rap1 in the perinuclear region (Fig-
ure 6D). Finally, we confirmed colocalization of Rap1 with
EEA1 in the perinuclear region following NGF treatment
(Figure 6F versus 6E). Based on these observations, we
conclude that NGF treatment of DRGs resulted in an
increase of these signaling proteins within the perinuclear
region as expected.
To show whether or not p50 overexpression blocks
retrograde axonal transport of NGF, we used Campenot
chamber cultures (CAMP10; Figure 7A), in which distal
axons (DAXs) were separated from their CBs and proximal
axons (PAXs) (9,25). DRGs (DIV-21) were transduced with
either a control virus or lenti-p50 or were not transduced,
as described in Materials and Methods. The lentiviral
system is highly efficient and results in sustained trans-
duction of nondividing cells, such as DRG neurons (43).
Following NGF withdrawal together with serum starvation
for 24 h, BtNGF (100 ng/mL) was added to the DAX
chamber. We elected to carry out most observations at
60 min because at this time-point there was robust trans-
port of BtNGF to CBs. Samples were rinsed, fixed and
incubated with a Streptavidin–Alexa 488 conjugate to
detect BtNGF in neuron CBs and PAXs. The fluorescence
signal for BtNGF/Streptavidin–Alexa 488 was measured
and quantified as described in Materials and Methods.
The results showed that p50 overexpression markedly
decreased the amount of BtNGF present in the CB in
comparison with the no-virus control (35 � 60 a.u. versus
210 � 40 a.u.). This represents a decrease of approxi-
mately 83% (p < 0.01; t-test) in retrogradely transported
BtNGF when p50 was overexpressed (Figure 7B). Trans-
duction with a control virus had no significant effect on
transport as judged in relation to nontransduced neurons
(210 � 40 a.u. versus 244 � 57 a.u.; p > 0.05).
To show that retrograde transport of BtNGF/pTrkA was
also blocked by p50 overexpression, we cultured DRGs
(DIV-21) in CAMP15 chambers (Figure 7A). Cells were
transduced with either lenti-EGFP or lenti-p50 or were
not transduced, as described previously. Following NGF
deprivation and serum starvation, BtNGF (100 ng/mL) was
added to the DAX chamber for 60 min. Samples from the
CB were rinsed, collected and lysed. BtNGF in lysates was
precipitated with a streptavidin–agarose conjugate. The
precipitates were washed and analyzed by SDS–PAGE/
Western blotting using a rabbit-specific antibody to mouse
NGF. BtNGFwas detected in the CB chamber in the control
virus sample (Figure 7C). In addition, pTrkA (pY490) was
present in the precipitate, as assessed by reprobing with
a rabbit-specific antibody to pTrkA (pY490) (Figure 7C).
Transduction with lenti-EGFP resulted in relatively small
(approximately 20%) decrease in pTrkA under this condi-
tion (Figure 7C). However, in cells transduced with lenti-
p50, the amount of BtNGF in the CBwasmarkedly reduced
and there was little pTrkA (Figure 7C). To ensure that NGF
activated TrkA in the AXs of these cells, we treated the
DAX chamber with NGF (100 ng/mL) for 15 min and
collected the axonal lysate for Western blotting. The level
of pTrkA induced by NGF in lenti-p50 DAXs was approxi-
mately 75% of controls (Figure 7D). These results show
that p50 overexpression blocks retrograde transport of the
NGF–pTrkA complex in DRG neurons.
Overexpression of p50 in dividing cells causes disintegra-
tion of the Golgi complex (39,44). Because DRG neurons
are postmitotic, we expected that the Golgi would remain
intact. To confirm this, we stained DRG neurons with a
mouse antibody (1/200) that specifically recognizes the
38 kD trans Golgi network protein TGN38. The flag-tagged
p50 was visualized using a rabbit antibody to the flag tag
1510 Traffic 2007; 8: 1503–1520
Wu et al.

(1/100). Alexa 568 goat anti-mouse immunoglobulin G
(IgG) and Alexa 488 goat anti-rabbit IgG conjugates were
used to visualize the respective primary antibodies. We
compared the pattern of TGN38 staining in cells trans-
duced with lenti-p50 with that in cells not transduced
(Figure 7E). There was no apparent change in the
appearance of the overall structures as marked by
TGN38 antibody following viral transduction. Moreover,
there was no apparent loss of DRG neurons during these
experiments.
Although these experiments showed that DRG neuron
survival was not affected by p50 overexpression, we
noticed an apparent decrease in the size of neuronal
somas even when neurons were bathed in NGF. Cellular
images were captured and quantified. In the continuing
Figure 6: NGF-induced trafficking
of Rap1, TrkA and pMAPK1/2 to
the perinuclear region in DRGs
(DIV-21). DRGs (DIV-21) were cul-
tured and starved as described in
Materials and Methods. Cells were
treated with either NGF vehicle
(�NGF; A, C and E) or 100 ng/mL
NGF (þNGF; B, D and F) for 60 min.
Cells were rinsed, fixed and prepared
for immunostaining with indicated
antibodies. Alexa 568 goat anti-rabbit
IgG conjugates were used to visualize
Rap1 (magenta). Alexa 488 goat anti-
mouse IgG conjugates were used to
reveal Trk, pMAPK1/2 and EEA1 (all in
green). Representatives of confocal
microscopic images from at least
three independent experiments were
shown. The size of the collection box
is 50 � 50 mm.
Traffic 2007; 8: 1503–1520 1511
Dynein-/Microtubule-Mediated Trafficking of TrkA for Rap1 Signaling

presence of NGF, p50-overproducing cells were smaller,
with a mean profile of 685 � 200 mm2 (n ¼ 50) compared
with 943 � 193 mm2 (n ¼ 50) for cells transducedwith the
control virus (i.e. a decrease to approximately 73% of the
control, p < 0.01). We conclude that it is possible that p50
overexpression led to a change in cell function, resulting in
cellular atrophy. An alternative explanation is that NGF
signaling from the surface of neuron CBs was inadequate
to sustain cellular differentiation in the absence of the
ability to traffic endosomes.
Overexpression of p50 blocks NGF-induced activation
of MAPK1/2 and results in atrophy in mature DRG
neurons in culture
To examine if p50 overexpression suppressed NGF sig-
naling in mature neurons, DRG neurons (DIV-21) were
Figure 7: Ectopic expression of p50-blocked retrograde transport of NGF–pTrkA signaling in rat DRGs (DIV-21) in Campenot
chambers. A) Diagrams of two types of Campenot nerve cell growth chamber (CAMP10 and CAMP15). In (B), 20 000 cells from
dissociated rat E15-16 DRGs were plated in the inner chamber of CAMP10. Cells were maintained for 21 days and then transduced with
the indicated lentivirus. Seven days posttransduction, both the inner and the outer chambers were washed extensively and starved.
Samples were rinsed again, and 50 ng/mL BtNGF was added to the outer chamber, followed by incubation at 378C for 1 h. BtNGF was
detected using a Streptavidin–Alexa 488 conjugate, and the amount of BtNGF in a.u. that was transported to the CBs and part of the PAXs
from the DAX was quantified as described inMaterials andMethods. In (C), DRGs (DIV-21) were cultured in CAMP15 and transduced as in
(B). Following NGF deprivation, 100 ng/mL BtNGF was added to the axonal chamber, and the samples were incubated at 378C for 1 h.
Samples were rinsed, and the CBs were lysed, collected and cleared by centrifugation. The resulting supernatant was incubated with
streptavidin–agarose conjugate. The precipitates were analyzed using SDS–PAGE/immunoblotting to detect BtNGF and pTrkA as
indicated. In (D), NGF (100 ng/mL) (þ) or NGF vehicle (�) was added to the DAX chamber for 15 min, and the AXs were collected for SDS–
PAGE/immunoblotting analysis with the pTrkA antibody. Representative blots from at least three independent experiments were shown.
In (E), DRG neurons (DIV-21) were transduced with either a control virus or lenti-p50 overnight, and the cultures were continued for an
additional 7 days. Cells were extensively washed and prepared for immunostaining with a mouse antibody to TGN38 and a rabbit antibody
to flag for visualizing p50. Alexa 568 goat anti-mouse IgG conjugates were used to visualize TGN38 (magenta). Alexa 488 goat anti-rabbit
IgG conjugates were used to reveal dynamitin (green). Representatives of confocal microscopic images from at least three independent
experiments were shown. The size of the collection box is 50 � 50 mm.
1512 Traffic 2007; 8: 1503–1520
Wu et al.

cultured on coated cover glasses and transduced with
either a control virus or lenti-p50, as described previously.
Cells were then starved and treated with either NGF or the
NGF vehicle and processed for immunostaining for
pMAPK1/2. As expected, NGF treatment of neurons trans-
duced with a control virus induced a marked increase in
pMAPK1/2 in both CBs and AXs (Figure 8B versus 8A). In
cultures transduced with lenti-p50, approximately 90% of
the neurons overexpressed p50 in both the CB and the
AXs. In these cells, NGF failed to elicit activation of
Figure 8: Overexpression of p50 in mature rat DRG neurons inhibited NGF-induced activation of MAPK1/2. DRGs (DIV-21) were
prepared and transduced as described in Figure 7. Prior to processing for immunostaining and immunoblotting, cells were starved, washed
and treatedwith either the NGF vehicle (�NGF) or NGF (þNGF, 100 ng/mL for 60 min). In both (A) and (B), p50 (green) was immunostained
with a specific mouse antibody and detected with a secondary goat anti-mouse IgG–Alexa 488 conjugate. pMAPK1/2 (magenta) was
stained. The samples were captured and analyzed using a Nikon E800 epifluorescence microscope (A and B). Twenty-five cells were
examined in each condition, and a typical representative image is shown. The scale bar in the CB and AX is 20 mm. In (C), lysates from cells
that were treated with either the NGF vehicle (�NGF) or NGF (þNGF, 100 ng/mL for 60 min) were analyzed using SDS–PAGE/
immunoblotting. The blots were probed with a rabbit antibody to either pMAPK1/2 or the phosphorylated form of TrkA (pY490) or to pAkt.
Blots that fell within the linear range of exposures were scanned and measured using Scion Image Beta 4.0.2 (Scion Corporation). The
relative activity in a.u. was obtained by subtracting the background value in the vehicle-treated sample from that of the NGF-treated
sample. The results shown are representative of at least three independent experiments. In (D), images of 100 cells from each sample
were analyzed, and the cell size was measured using Scion Image Beta 4.0.2 as described inMaterials and Methods. The size distribution
was shown.
Traffic 2007; 8: 1503–1520 1513
Dynein-/Microtubule-Mediated Trafficking of TrkA for Rap1 Signaling

MAPK1/2 in either the CB or the AX (Figure 8B ver-
sus 8A).
To examine the effect of p50 overexpression on NGF
signaling, we harvested cell lysates from cultures treated
with either the vehicle or the NGF (100 ng/mL) for 60 min.
Samples were analyzed using SDS–PAGE, followed by
immunoblotting with an antibody to pTrkA (pY490). As
shown in Figure 8C, there was a small (approximately
17%) and statistically insignificant reduction (p > 0.5) in
the level of pTrkA induced by NGF in p50-overproducing
cells compared with cells transduced with a control virus.
Thus, consistent with previous results (Figure 7D), p50
overexpression did not appear to significantly affect TrkA
activation. Next, we measured the level of pMAPK1/2.
While a robust response was evident in cells that were
transduced with the control virus, activation of MAPK1/2
by NGF was virtually completely inhibited in neurons that
overexpressed p50 (Figure 8C). NGF-mediated activation
of the PI3K/Akt was also suppressed, albeit to a lesser
extent (Figure 8C). Therefore, as for PC12 cells, over-
expression of p50 in DRG neurons blocked or severely
inhibited the sustained phase of MAPK1/2 activation
induced by NGF while having little effect on the activation
of TrkA.
To address more thoroughly what role dynein-based trans-
port has on NGF signaling, we examined the neuronal
soma size, a well-recognized phenotype regulated by NGF
signaling inmature DRG neurons (45) and in NGF-dependent
basal forebrain cholinergic neurons (46,47). Rat DRG
neurons cultured for DIV-21 on cover glasses were trans-
duced with either a control virus or lenti-p50 in the
continuous presence of NGF. Seven days later, cells were
imaged using confocal microscopy. In control samples, cell
size varied from 200 to 1800 mm2, with a mean value of
1010 � 28 mm2. In cells that overproduced p50, the mean
value was significantly decreased to 651 � 28 mm2
(p < 0.01) (Figure 8D). Overexpression of p50 thus re-
sulted in severe atrophy (to approximately 65% of control
value), despite the fact that NGF was continuously present
in the medium bathing the surface of CBs and their AXs. If
compromised NGF signaling and not some other conse-
quence of p50 overexpression was responsible, the same
effect should be shown with simple removal of NGF from
the bathing medium. This is what was observed. The NGF
withdrawal from mature DRG neurons (DIV-21) for 5 days
resulted in a decrease in somal size to approximately 70%
of the NGF-treated control. We conclude that interrupting
dynein function through p50 overexpression markedly
inhibited the NGF signaling events needed to maintain
the somal size of mature DRG neurons.
Overexpression of p50 in developing DRG neurons
caused cell death even in the presence of NGF
Because the survival of developing DRGs is critically
dependent on the availability of NGF (42), we asked
whether or not overexpression of p50 in developing DRG
neurons would result in cell death in the presence of NGF.
Embryonic rat DRGs (E15-16) were dissected, dissociated
and cultured for 7 days (DIV-7). Cultures were then trans-
duced with either a control virus or lenti-p50 and main-
tained for additional 7 days in the continuous presence of
NGF (100 ng/mL). Under these conditions, we routinely
achieved >95% transduction efficiency. Cultures were
rinsed and apoptotic neurons detected using the In Situ
Cell Death Detection Kit, TMR red (Roche). This assay
specifically labels DNA strand breaks (‘nicks’) in genomic
DNA produced during apoptosis. Apoptotic cells, whose
nuclei were fluorescently labeled with TMR red, were
visualized and counted. Transmitted light images were
simultaneously collected and superimposed onto the cor-
responding confocal images to allow for visualization of
cells. Representative images (Figure 9) show cells trans-
duced with no virus (Figure 9A), a control virus (Figure 9B)
or lenti-p50 (Figure 9C). The DRG neurons transduced with
lenti-p50 showed massive cell death (Figure 9C), whereas
relatively few apoptotic cells were observed in either of the
control cultures (Figure 9A,B). Scoring the percentage of
apoptotic cells showed a marked and highly significant
(p < 0.01) increase when neurons were transduced with
lenti-p50 (Figure 9D). Thus, even though NGF (100 ng/mL)
was continuously present throughout the culture period,
transduction with lenti-p50 resulted in the death of approx-
imately 85% of neurons. Consistent with other recent
findings (30), we conclude that dynein-based transport of
NGF signaling appear to play a critical role in promoting
survival of developing DRG neurons.
Discussion
Herein, we show that intact microtubules and functional
dynein are critical for the ability of NGF signaling to induce
sustained activation of MAPK1/2 and to support the
survival and maintenance of DRG neurons. The underly-
ing trafficking events induced by NGF treatment included
translocation of TrkA and Rap1 to the perinuclear region
where they were present in early endosomes. Import-
antly, the trafficking events supported by microtubules
and dynein were necessary not just for NGF signals
created and transported retrogradely within AXs but also
when signals arose at the level of CBs. The data are
evidence that endosomal trafficking plays an important
role in both the creation of NGF signals and their intracel-
lular transport.
Our working hypothesis posits that several distinct steps
are required for generating the TrkA/Rap1 signaling endo-
some. First, NGF binds to TrkA at the cell surface to
activate the receptor. Second, the NGF–pTrkA complex is
internalized through either clathrin-mediated or non-
clathrin-mediated pathways (18,19,48). Third, the newly
formed NGF–pTrkA signaling endosome is likely docked
onto the microtubule network through the binding of both
the intermediate and the light chains of the dynein
1514 Traffic 2007; 8: 1503–1520
Wu et al.

complex (30), thus linking it to the retrograde transport
system.
The studies reported herein provide evidence for another
step in trafficking of the NGF–pTrkA complex, leading to
‘maturation’ of the NGF–pTrkA signaling complex into one
that is now competent to persistently activate MAPK1/2.
Rap1 plays a key role in transducing and sustaining the
NGF signal from TrkA to MAPK1/2. Disruption of the
microtubule network by nocodazole or the dynein–
dynactin complex through ectopic expression of p50,
dynamitin, prevents the genesis of a fully functional
NGF–TrkA–Rap1 signaling endosome.
There is compelling evidence that the NGF–pTrkA–Rap1
signaling endosome contributes critically to sustained
activation of MAPK1/2, a pathway important for PC12 cell
differentiation (33,36). The current study provides some
key evidence that following NGF treatment, TrkA and Rap1
are trafficked to signaling endosomes in the perinuclear
region. This process likely represents an important step in
delivering the NGF signal to the nucleus (9,48,49). There
are significant potential advantages to the compartmental-
ization of TrkA signaling in endosomes. First, the signal
would be specific for NGF. Second, by concentrating the
signaling machinery used to transmit TrkA signals, the
probability of conducting signals that are robust and of high
fidelity may be increased. Third, providing the signal near
the nucleus a local source of downstream signals, such as
those propagated by activated MAPK1/2, would facilitate
effective delivery of signals through diffusion.
Similar observations were made for the c-Jun NH(2)-
terminal kinase (JNK) signaling pathway in neurons (50).
The JNK-interacting protein-1, JIP1, that serves as a scaf-
fold for the stress-activated JNK pathway was found to be
highly concentrated in the neurite tips of primary hippo-
campal neurons. On anoxic stress (oxygen and glucose
deprivation), JIP1 as well as activated JNK were trans-
located to the perinuclear region (50,51). Ablation of JIP1
prevented JNK activation and rendered neurons less
susceptible to stress-induced apoptosis. Trafficking of
signaling proteins from the cell periphery to the perinuclear
region may contribute critically to the ability to transmit
a variety of signals that modulate neuronal survival and
function.
The mechanistic basis for trafficking events involving TrkA
is the topic of recent studies (18–20). Our findings suggest
that the cytoplasmic motor dynein plays an important role
in trafficking of the NGF–pTrkA complex and in the
activation of the MAPK1/2 and PI3K signaling pathways.
It remains to be determined how the dynein–dynactin
complex interacts with TrkA and, perhaps, with elements
Figure 9: Ectopic expression of p50
in early developing cultured rat DRG
neurons (DIV-7) caused apoptosis.
DRG neurons were isolated and cul-
tured for 7 days as described. Cells
were transduced with no virus (A),
a control virus (B) or lenti-p50 (C) over-
night, and the cultures were continued
for an additional 7 days in the presence
of 100 ng/mL NGF. Seven days post-
transduction, samples were pro-
cessed, and apoptotic cells were
detected using the In Situ Cell Death
Detection Kit, TMR red. The samples
were analyzed and images captured
using a confocal microscope, and the
nuclei of apoptotic cells were specifi-
cally labeled and revealed with
magenta fluorescence. Images from
transmitted light detector were simul-
taneously collected and superimposed
onto their corresponding fluorescence
images. The size of the collection box
is 500 � 500 mm. In (D), we scored
the number of apoptotic DRG neurons
in each condition. The percentile of
apoptotic cells was shown.
Traffic 2007; 8: 1503–1520 1515
Dynein-/Microtubule-Mediated Trafficking of TrkA for Rap1 Signaling

of the NGF–pTrkA–Rap1 signaling complex. Whether or
not the activity of dynein itself is regulated by NGF
signaling is another important question. In addition, the
receptor for transforming growth factor-b (52,53) and the
activated receptor for epidermal growth factor (54) were
recently shown to undergo translocation to the perinuclear
region through early endosomes and that this process was
driven by dynein. It will be important to define whether or
not dynein contributes routinely to the trafficking of the
signals of other NTs and cytokines.
The signaling endosome hypothesis focuses on the retro-
grade transport of neurotrophic signals within AXs. We
expected to show that overexpressing p50 would severely
inhibit retrograde transport of NGF signaling from AX
terminals of DRG neurons. Unexpectedly, we discovered
that ectopic expression of p50 also inhibited NGF signaling
from the somal surface of DRG neurons, leading to the
death of immature neurons and the atrophy of mature ones.
These findings make the case that NGF signaling events,
including at least those that involve MAPK1/2 and PI3K,
need to engage the endosomal system. Indeed, our findings
are evidence that endosomal traffic not only delivers signals
but is also used to create them. It is important to point out
that the present study does not rule out the existence of
other paradigms by which NGF signal can be conveyed to
the CB. We speculate that it is advantageous to have
multiple pathways coexit to support neuronal function.
Disruption of trafficking of NGF signal can readily be
envisioned as adversely affecting neuronal function.
Indeed, an emerging body of data suggests that such
events are impaired or altered in the central nervous
system of patients suffering from neurodegenerative dis-
orders such as Alzheimer’s disease (6,55–62). It is impera-
tive to pursue further the mechanisms of retrograde
trafficking of NGF and other NT signaling and their possible
involvement in neurodegenerative disorders.
Materials and Methods
Antibodies and reagentsMouse antibodies to EEA1, dynamitin (p50), Ras, Rap1 and TGN38 were
purchased from BD Transduction Laboratories. Rabbit antibodies to Rap1
(Krev-1) and EGFP and mouse antibodies to Trk (B3 and MCTrks), DIC74
and pMAPK1/2 were purchased from Santa Cruz Biotechnology, Inc. Rabbit
antibodies to Elk-1, phospho-Elk-1, pTrkA (pY490), Akt, phosphorylated Akt
(Ser473), MAPK1/2 (Erk1/2) and the phosphorylated (i.e. activated) MAPK1/2
were purchased from Cell Signaling, Inc. Rabbit antibody to flag was
purchased from Upstate Biotechnology, Inc. A rabbit antibody to mouse
NGF was purchased from Alomone labs. Control mouse IgGs were
obtained from Chemicon. Nocodazole and n-Octyl-b-D-glucopyranoside
were purchased from Calbiochem-Novabiochem Corp. Nocodazole was
dissolved at a concentration of 5 mg/mL (1000 � stock) in dimethyl
sulfoxide (DMSO). Mouse NGF was purified as previously described (63).
The NGF was stored in 0.2% acetic acid (the NGF vehicle). The BtNGF was
produced according to a published method (64). The efficiency of biotin
labeling (>95%) was verified by retarded mobility of BtNGF versus NGF on
SDS–PAGE. The BtNGF was fully potent in activating TrkA and in inducing
neurite outgrowth as confirmed using PC12 cells (data not shown).
Horseradish peroxidase (HRP) conjugated to goat anti-rabbit or anti-mouse
IgGs was obtained from Jackson Immunoresearch Laboratories, Inc. Alexa
568 or Alexa 488 goat IgG conjugates (anti-mouse or anti-rabbit) and
Hoechst 33258 (10 mg/mL), Streptavidin–Alexa 488 were purchased from
Molecular Probes. Protein G–agarose conjugates and SuperSignal reagents
were obtained from Pierce. OptiPrep was obtained from Life Technologies.
Mouse IgGs against a-tubulin (Clone DM1A) and agarose–glutathione
conjugates and all other chemicals were purchased from Sigma. In Situ
Cell Death Detection Kit, TMR red (Cat# 2 156 792) was purchased from
Roche Applied Science. Campenot nerve cell growth chambers (CAMP10
and CAMP15) were purchased from Tyler Research Corporation.
Cell culture and transient transfectionPC12 cells were maintained in DMEM (4.5 mg/L glucose) supplemented
with 10% horse serum and 5% FBS. Cells were incubated at 378C with 5%
CO2. Cells that were 50–60% confluent and serum starved for 24 h prior to
treatments were used in all experiments. Unless indicated otherwise, all
pretreatments and treatments were carried out by adding growth factors
and other reagents to the media at 378C. The final concentrations for the
nocodazole vehicle (DMSO) and the NGF vehicle (0.2% acetic acid) were
0.1 and 0.0001%, respectively. For transient transfection, the pcDNA3
plasmids containing the full-length, flag-tagged human dynamitin cDNA
sequence (a generous gift from Dr E. Holzbaur, University of Pennsylvania)
were introduced into 70–80% confluent PC12 cells using Lipofectamine
2000 (Invitrogen). For mock transfection, pEGFP-N1 (BD Clontech) was
used as a control. In general, a transfection efficiency of 5–10% was
obtained.
Rat DRG culture and lentiviral transductionThe DRG neurons were isolated aseptically from E15-16 Sprague Dawley
rat following previously established methods (42,65). Dissociated neurons
were plated on either collagen-coated 12-well plates (25 000) or 18-mm
glass coverslips in 12-well plates (10 000/plate) in NGF (100 ng/mL) in
‘maintenance medium’ [(MM), containing sodium phosphate-free MEM
with Earle’s salts, L-glutamine, 10% heat-inactivated fetal bovine serum and
4.5 g/L D-glucose]. Selection medium (MM with antimitotic factors) was
added to the culture next day. The selection was continued for 2 days.
Cultures were then maintained in MM containing NGF (100 ng/mL) for 18
additional days (a total of 21 days), feeding every 3 days. The 21-day-old
cells in vitro are referred as DIV-21. At this time, they contain large, mature
neurons, with an average diameter of 48 mm, at least 94% of which are
immunopositive for TrkA and extend well-developed neurites that also stain
positively for TrkA (data not shown).
For lentiviral transduction, a cDNA encoding either EGFP or dynamitin was
cloned into the pLenti6/V5 expression plasmid using the ViraPower
Lentivirus Expression System (Invitrogen) following the manufacture’s
instruction. The plenti6-EGFP, pLenti6-dynamitin plasmid or the empty
vector was cotransfected together with pLP1, pLP2 and pLP/VSVG into
293FT cells. The lentivirus (lenti-EGFP or lenti-dynamitin or control virus)
was harvested from the supernatant (5000� g for 15 min). The DRG
neurons of DIV-7 or DIV-21 were transduced with the resulting lentivirus
at a multiplicity of infection of approximately 10. A control virus was
produced using the empty pLenti6/V5 vector and was also introduced into
DRGs in parallel experiments. Virus was removed from DRG culture
following an overnight incubation. The DRG neurons were switched back
to MM, and the culture was continued for an additional 7 days. Prior to
experiments, DRG neurons were extensively washed in serum-free, NGF-
free MM and were deprived of NGF for 24 h in serum-free, NGF-free MM
containing a goat antibody against mouse NGF (1:500). After washing in
MM, NGF (100 ng/mL) or equal volume of the vehicle control (0.05% acetic
acid in PBS, pH 7.0) was added for 60 min. Cells were washed and
processed for either immunostaining for confocal analysis or lysed in
1516 Traffic 2007; 8: 1503–1520
Wu et al.

RadioImmunoPrecipitation Assay (RIPA) buffer for SDS–PAGE/immuno-
blotting as described subsequently.
Ras and Rap1 activation assayEstablished methods were used to detect endogenous GTP-bound Ras and
Rap1 proteins as described previously (36). The fusion constructs between
glutathione S-transferase (GST) and either the Rap-binding domain of
RalGDS (RalGDSRBD) or the Ras-binding domain of C-Raf (C-RafRBD)
(gifts from Dr J. L. Bos, Utrecht University, The Netherlands) were over-
expressed in Escherichia coli DH5a cells. The fusion proteins were purified
and used to assay Rap1GTP and RasGTP, respectively. Briefly, an equal
number of treated or untreated PC12 cells were lysed in ice for 30 min in
fishing buffer (FB) (10% glycerol; 1% Nonidet P-40; 50 mM Tris–HCl, pH
7.5; 200 mM NaCl; 2.0 mM MgCl2; 250 mM phenylmethylsulfonyl fluoride;
2 mg/mL aprotinin; 1 mg/mL leupeptin; 10 mg/mL soybean trypsin inhibitor;
10 mM NaF and 1 mM Na3VO4). The samples were centrifuged at 16 000� g,
for 30 min at 48C. Either 5 mg RalGDSRBD/GST or 5 mg C-RafRBD/GST
proteins that were prebound to agarose–glutathione conjugates was added
to the resulting supernatants and incubated at 48C for 60–120 min with
gentle rotation. The beads were washed four times in cold FB and boiled in
SDS–PAGE sample buffer. The amounts of RasGTP and Rap1GTP were
analyzed using SDS–PAGE and immunoblotting.
Cell fractionationFractionation of PC12 cells was carried out using a published protocol (36).
Briefly, an equal number of treated or untreated PC12 cells were rinsed and
harvested by centrifugation (800� g for 5 min). Cells were resuspended
and homogenized by douncing 20 times in a Teflon-coated homogenizer in
1 mL cold homogenization buffer (HB) (250 mM sucrose; 20 mM Tricine–
NaOH, pH7.8; 1 mM ethylenediaminetetraacetic acid and 2 mM MgCl2). The
samples were centrifuged (800� g for 10 min), and the supernatant was
adjusted to 25% OptiPrep with 50% OptiPrep in HB. The resulting mixture
(2 mL in 25% OptiPrep) was placed at the bottom of an Ultra-Clear� Tube
(14 � 89 mm; Beckman Instruments, Inc.) and was overlaid successively
with 2 mL each of 20, 15, 10 and 5% OptiPrep in cold HB. The samples
were centrifuged for 16–18 h at 27 000 r.p.m. at 48C in a SW41 rotor
(Beckman Instruments, Inc.). Membrane fractions were collected from
each of the four interphases of the OptiPrep gradients. For all studies,
proteins were precipitated from the membrane fractions using 7% tri-
chloroacetic acid and washed with acetone. The pellets were air-dried,
boiled in SDS–PAGE loading buffer and analyzed using SDS–PAGE and
immunoblotting.
Elk-1 in vitro phosphorylation assay,
immunoprecipitation, SDS–PAGE and
immunoblottingBriefly, protein samples were separated on 7.5–12.5% gels and proteins
electrotransferred onto polyvinylidene fluoride (PVDF) membranes (NEN
Life Science Products, Inc.). The PVDF membranes were preblotted with
5% nonfat milk (carnation) and probed with primary antibodies, as indi-
cated, at concentrations suggested by the suppliers. The blots were
washed in Tris Buffered Saline Tween 20 (TBST) (20 mM Tris–HCl, pH
7.4; 150 mM NaCl and 0.1% Tween-20), followed by incubation with either
goat anti-mouse or anti-rabbit IgG–HRP conjugates at a dilution of 1/10 000
to 1/40 000. The blots were washed and developed with SuperSignal
(Pierce). For quantification, exposures within the linear ranges were
scanned and determined using the Scion Image program (Scion Corp.).
For in vitro Elk-1 phosphorylation assay, a previously published method was
followed (28).
Indirect immunofluorescence and
confocal microscopyThe PC12 cells were grown for 24–48 h on glass coverslips coated with
matrigel (Becton Dickinson). Cells were starved, washed twice briefly
with cold PBS and fixed with ice-cold 100% methanol for 5 min at �208C.The samples were rinsed thrice with PBS at room temperature. The fixed
cells were preblocked with 1% BSA and 0.2% Triton-X-100 in PBS for
20 min at room temperature. Primary antibodies were diluted at 1/300 to
1/500 in PBS containing 1% BSA and were incubated with the fixed cells
for 1 h at room temperature. The samples were washed with PBS
containing 0.8% BSA thrice and then incubated with goat anti-rabbit– or
anti-mouse–IgG Alexa conjugates (1/600) for 1 h at room temperature.
The coverslips were washed with PBS containing 0.2% BSA thrice,
followed by a rinse with PBS and dH2O. In experiments where nuclear
staining was required (Figure S2C,D and 5A), samples were further
incubated with 2 mg/mL Hoechst 33258 (final concentration) in PBS for
1 min at room temperature, followed by a rinse with PBS and dH2O. The
coverslips were air-dried and mounted in antifade medium for observa-
tion. Images shown in Figure S2C,D and 5A were analyzed using a Nikon
E800 epifluorescence microscope and were captured using a SPOT
charge-coupled device (CCD) camera (Diagnostic Instruments). The
images were processed using SPOT 3.4.3 and Adobe Photoshop 5.0
(Adobe Systems). For confocal microscopy, the images were captured
using a Nikon Eclipse E800 microscope and a Bio-Rad Laser Scanning
System Radiance2000. Using a 60� Plan-Apo immersion objective
(NA1.4), images at the half height of the cell were collected in a
512 � 512 collection box. The images were processed using Confocal
Assistant 4.02 (Bio-Rad) and Adobe Photoshop 7.0 (Adobe Systems).
In experiments in which the periphery of cells was visualized, a bright field
image was captured and superimposed onto the fluorescence image.
Transport of BtNGF in rat DRGs using
Campenot chamberE15-16 rat DRGs were dissected and dissociated, and 20 000 cells were
placed in the middle chamber of a CAMP10 Campenot chamber (Figure 7).
After seven DIV, AXs crossed the barrier and extended into the outer
chamber. Cultures were maintained for 3 weeks in NGF-containing
medium as described previously. The DRGs were then transduced with
lentivirus with different constructs and maintained for an additional 7 days.
Cells were extensively rinsed and starved as described previously. We
routinely checked and discarded those chambers with leakage problem. For
transport assay, BtNGF (100 ng/mL) was added to the DAXs in the outer
chamber and incubated for 60 min at 378C. Cells were rinsed, fixed,
permeabilized and incubated with Streptavidin–Alexa 488 (1/1000 dilution)
for 1 h at room temperature. Samples were rinsed and mounted. The
BtNGF transported to the CB was measured as follows: images of CBs in
the middle chamber were acquired using an inverted Nikon fluorescence
microscope (TE2000U) equipped with Nikon 10� Plan objective. The
fluorescence signal was detected and captured using a CCD camera
(CoolSNAP HQ; Roper Scientific, Inc.), with a detection area of
1392 � 1040 pixels (pixel size: 6.45 � 6.45 mm2). This cooled (�308C)camera has a 12-bit dynamic range, which allows imaging very dim and very
bright objects at the same time without saturation. Binary data were
collected and saved using a home-built software package with 2 � 2
hardware binning and 0.5 seconds exposure time (Stanford University).
Subsequent data analysis was performed using Data Interactive Language
(IDL) software with scripts written in the Chu laboratory. For each fluores-
cence image, the average signal intensity within a selected region of
interest (ROI) encompassing 100 cells was measured. At least four ROIs
were selected to obtain the mean and the standard deviation. A background
value was obtained by incubating a sample with Streptavidin–Alexa 488
conjugate only. This value was subtracted from those obtained from the
samples that were treated with BtNGF. All statistical analyses were carried
out using Student’s t-test.
For detection of BtNGF that was transported retrogradely to the CB/PAXs
by Western blot, 200 000 DRGs were placed in the small chamber of
a CAMP15 apparatus (Figure 7) and maintained for 3 weeks. Following
lentiviral transduction and NGF deprivation, BtNGF (50 ng/mL) was added
into the larger chamber that harbored only the AXs and was incubated for
12 h. The CBs/PAXs were rinsed, lysed and cleared by centrifuge. Forty
microliters of streptavidin–agarose conjugate slurries was added to the
supernatant to precipitate BtNGF. The precipitates were analyzed by
SDS–PAGE/Western blotting.
Traffic 2007; 8: 1503–1520 1517
Dynein-/Microtubule-Mediated Trafficking of TrkA for Rap1 Signaling

Acknowledgments
We thank the members of the Mobley laboratory for their assistance. We
also thank Drs Francisca Bronfman and Mike Fainzilber for their invaluable
advice on the production of BtNGF. These studies are supported through
the grants fromNational Institutes of Health (NS24054, NS38869, AG16999
and NS055371), the John Douglas French Alzheimer’s Foundation, the
McGowan Charitable Trust, the Larry L Hillblom Foundation, the Adler
Foundation and the Deane Johnson Alzheimer’s Disease Fund.
Supplementary Materials
Figure S1: The BtNGFwas internalized into EEA1-positive endosomes
in PC12 cells. The PC12 cells were cultured for 24–48 h on cover glasses
coated with matrigel and were serum starved overnight. Cells were chilled
and incubated with BtNGF (50 ng/mL) for 45 min on ice. Cells were rinsed
and immediately processed for immunostaining (A, C, E and G). A parallel
sample was shifted to 378C for 30 min to allow endocytosis of BtNGF prior
to immunostaining (B, D, F and H). The BtNGF was detected with
a Streptavidin–Alexa 488 conjugate (green, A–D). The following specific
primary antibodies were used: a mouse antibody to Trk, B3 (1/200) for TrkA
(A, B, E and F); a mouse antibody to EEA1 (1/50) for EEA1 (C and D); a rabbit
antibody to Rab5B (1/200) for Rab5B (E, F, G and H) and a mouse antibody
to pMAPK1/2 (1/100) for pMAPK1/2 (G and H). Secondary antibody
conjugated to either Alexa 568 or Alexa 488 was used to visualize the
primary antibody. Colocalization of proteins is denoted by the white signal.
The size of the collection box is 30 � 30 mm.
Figure S2: The NGF-induced trafficking of Rap1, TrkA and pMAPK1/2
to the perinuclear region. The PC12 cells were cultured as described in
Figure 1. Following pretreatment with either the NGF vehicle alone (�NGF)
or the NGF (50 ng/mL) (þNGF), indirect immunofluorescence was carried
out using confocal microscopy as described in Materials and Methods.
Rap1 (A–D and G–L) was immunostained using a specific rabbit antibody
(1/250) and detected with an Alexa 568 goat anti-rabbit IgG conjugate. The
TrkA (E and F and G and H) was detected using B3 (1/200) and visualized
with either an Alexa 568 goat anti-mouse IgG conjugate (E and F) or an Alexa
488 goat anti-mouse IgG conjugate (G and H). The signals for pMAPK1/2
and EEA1 were each detected using a mouse monoclonal antibody specific
for these proteins as shown in Figure 1. The dilutions of primary antibodies
were as follows: 1/100 for pMAPK1/2 and 1/50 for EEA1. An Alexa 488 goat
anti-mouse IgG conjugate was used to visualize these antibodies. Colocal-
ization of antigens is denoted by the white signal. In A, B, E and F,
a corresponding image from the transmitted light detector was simul-
taneously captured and superimposed onto the fluorescence image of Rap1
or TrkA to illustrate the outline of the cell. In (C) and (D), Rap1 (magenta) was
co-stained with Hoechst 33258 to reveal the nucleus (blue). The resulting
samples were captured and analyzed using a Nikon E800 epifluorescence
microscope. Twenty-five cells were examined in each condition (A–L), and
a typical representative image was shown. The size of the collection box is
30 � 30 mm for (A) and (B) and E–L and 60 � 60 mm for (C) and (D).
Supplemental materials are available as part of the online article at http://
www.blackwell-synergy.com
References
1. Riccio A, Pierchala BA, Ciarallo CL, Ginty DD. An NGF-TrkA-mediated
retrograde signal to transcription factor CREB in sympathetic neurons.
Science 1997;277:1097–1100.
2. Riccio A, Ahn S, Davenport CM, Blendy JA, Ginty DD. Mediation by
a CREB family transcription factor of NGF-dependent survival of
sympathetic neurons. Science 1999;286:2358–2361.
3. Watson FL, Heerssen HM, Moheban DB, Lin MZ, Sauvageot CM,
Bhattacharyya A, Pomeroy SL, Segal RA. Rapid nuclear responses to
target-derived neurotrophins require retrograde transport of ligand-
receptor complex. J Neurosci 1999;19:7889–7900.
4. Kuruvilla R, Ye H, Ginty DD. Spatially and functionally distinct roles
of the PI3-K effector pathway during NGF signaling in sympathetic
neurons. Neuron 2000;27:499–512.
5. Miller FD, Kaplan DR. On Trk for retrograde signaling. Neuron 2001;
32:767–770.
6. Sofroniew MV, Howe CL, Mobley WC. Nerve growth factor signaling,
neuroprotection, and neural repair. Annu Rev Neurosci 2001;24:
1217–1281.
7. Watson FL, Heerssen HM, Bhattacharyya A, Klesse L, LinMZ, Segal RA.
Neurotrophins use the Erk5 pathway to mediate a retrograde survival
response. Nat Neurosci 2001;4:981–988.
8. Ginty DD, Segal RA. Retrograde neurotrophin signaling: Trk-ing along
the axon. Curr Opin Neurobiol 2002;12:268–274.
9. MacInnis BL, Campenot RB. Retrograde support of neuronal survival
without retrograde transport of nerve growth factor. Science 2002;
295:1536–1539.
10. Ye H, Kuruvilla R, Zweifel LS, Ginty DD. Evidence in support of signaling
endosome-based retrograde survival of sympathetic neurons. Neuron
2003;39:57–68.
11. Campenot RB, MacInnis BL. Retrograde transport of neurotrophins:
fact and function. J Neurobiol 2004;58:217–229.
12. Kuruvilla R, Zweifel LS, Glebova NO, Lonze BE, Valdez G, YeH, Ginty DD.
A neurotrophin signaling cascade coordinates sympathetic neuron
development through differential control of TrkA trafficking and retro-
grade signaling. Cell 2004;118:243–255.
13. Ehlers MD, Kaplan DR, Price DL, Koliatsos VE. NGF-stimulated
retrograde transport of trkA in the mammalian nervous system. J Cell
Biol 1995;130:149–156.
14. Beattie EC, Zhou J, Grimes ML, Bunnett NW, Howe CL, Mobley WC.
A signaling endosome hypothesis to explain NGF actions: potential
implications for neurodegeneration. Cold Spring Harb Symp Quant Biol
1996;61:389–406.
15. Beattie EC, Howe CL, Wilde A, Brodsky FM, Mobley WC. NGF signals
through TrkA to increase clathrin at the plasmamembrane and enhance
clathrin-mediatedmembrane trafficking. J Neurosci 2000;20:7325–7333.
16. Cooper JD, Salehi A, Delcroix JD, Howe CL, Belichenko PV, Chua-
Couzens J, Kilbridge JF, Carlson EJ, Epstein CJ, Mobley WC. Failed
retrograde transport of NGF in a mouse model of Down’s syndrome:
reversal of cholinergic neurodegenerative phenotypes following NGF
infusion. Proc Natl Acad Sci U S A 2001;98:10439–10444.
17. Neet KE, Campenot RB. Receptor binding, internalization, and retrograde
transport of neurotrophic factors. Cell Mol Life Sci 2001;58:1021–1035.
18. Shao Y, AkmentinW, Toledo-Aral JJ, Rosenbaum J, Valdez G, Cabot JB,
Hilbush BS, Halegoua S. Pincher, a pinocytic chaperone for nerve
growth factor/TrkA signaling endosomes. J Cell Biol 2002;157:679–691.
19. Valdez G, Akmentin W, Philippidou P, Kuruvilla R, Ginty DD, Halegoua S.
Pincher-mediated macroendocytosis underlies retrograde signaling by
neurotrophin receptors. J Neurosci 2005;25:5236–5247.
20. Jullien J, Guili V, Derrington EA, Darlix JL, Reichardt LF, Rudkin BB.
Trafficking of TrkA-green fluorescent protein chimerae during nerve
growth factor-induced differentiation. J Biol Chem 2003;278:8706–8716.
21. Howe CL, Mobley WC. Long-distance retrograde neurotrophic signal-
ing. Curr Opin Neurobiol 2005;15:40–48.
22. Glebova NO, Ginty DD. Growth and survival signals controlling sym-
pathetic nervous system development. Annu Rev Neurosci 2005;28:
191–222.
1518 Traffic 2007; 8: 1503–1520
Wu et al.

23. MacCormickM,Moderscheim T, van der Salm LW,Moore A, Pryor SC,
McCaffrey G, Grimes ML. Distinct signalling particles containing
ERK/MEK and B-Raf in PC12 cells. Biochem J 2005;387:155–164.
24. Saxena S, Bucci C, Weis J, Kruttgen A. The small GTPase Rab7
controls the endosomal trafficking and neuritogenic signaling of the
nerve growth factor receptor TrkA. J Neurosci 2005;25:10930–10940.
25. MacInnis BL, Senger DL, Campenot RB. Spatial requirements for TrkA
kinase activity in the support of neuronal survival and axon growth in rat
sympathetic neurons. Neuropharmacology 2003;45:995–1010.
26. Mok SA, Campenot RB. A nerve growth factor-induced retrograde
survival signal mediated by mechanisms downstream of TrkA. Neuro-
pharmacology 2007;52:270–278.
27. YanoH, Lee FS, KongH, Chuang J, Arevalo J, Perez P, Sung C, ChaoMV.
Association of Trk neurotrophin receptors with components of the
cytoplasmic dynein motor. J Neurosci 2001;21:RC125.
28. Delcroix JD, Valletta JS, Wu C, Hunt SJ, Kowal AS, Mobley WC. NGF
signaling in sensory neurons: evidence that early endosomes carry
NGF retrograde signals. Neuron 2003;39:69–84.
29. Bhattacharyya A, Watson FL, Pomeroy SL, Zhang YZ, Stiles CD,
Segal RA. High-resolution imaging demonstrates dynein-based ves-
icular transport of activated Trk receptors. J Neurobiol 2002;51:
302–312.
30. Heerssen HM, Pazyra MF, Segal RA. Dynein motors transport acti-
vated Trks to promote survival of target-dependent neurons. Nat
Neurosci 2004;7:596–604.
31. Reynolds AJ, Kaasinen SK, Hendry IA. Retrograde axonal transport of
dopamine beta hydroxylase antibodies by neurons in the trigeminal
ganglion. Neurochem Res 2005;30:703–712.
32. Bartlett SE, Reynolds AJ, Tan T, Heydon K, Hendry IA. Differential
mRNA expression and subcellular locations of PI3-kinase isoforms in
sympathetic and sensory neurons. J Neurosci Res 1999;56:44–53.
33. York RD, Yao H, Dillon T, Ellig CL, Eckert SP, McCleskey EW, Stork PJ.
Rap1 mediates sustained MAP kinase activation induced by nerve
growth factor. Nature 1998;392:622–626.
34. Traverse S, Gomez N, Paterson H, Marshall C, Cohen P. Sustained
activation of the mitogen-activated protein (MAP) kinase cascade may
be required for differentiation of PC12 cells. Comparison of the effects
of nerve growth factor and epidermal growth factor. Biochem J 1992;
288:351–355.
35. Mochizuki N, Yamashita S, Kurokawa K, Ohba Y, Nagai T, Miyawaki A,
Matsuda M. Spatio-temporal images of growth-factor-induced activa-
tion of Ras and Rap1. Nature 2001;411:1065–1068.
36. Wu C, Lai CF, Mobley WC. Nerve growth factor activates persistent
Rap1 signaling in endosomes. J Neurosci 2001;21:5406–5416.
37. Paschal BM, Holzbaur EL, Pfister KK, Clark S, Meyer DI, Vallee RB.
Characterization of a 50-kDa polypeptide in cytoplasmic dynein prepa-
rations reveals a complex with p150GLUED and a novel actin. J Biol
Chem 1993;268:15318–15323.
38. Huang CS, Zhou J, Feng AK, Lynch CC, Klumperman J, DeArmond SJ,
Mobley WC. Nerve growth factor signaling in caveolae-like domains
at the plasma membrane. J Biol Chem 1999;274:36707–36714.
39. Burkhardt JK, Echeverri CJ, Nilsson T, Vallee RB. Overexpression of
the dynamitin (p50) subunit of the dynactin complex disrupts dynein-
dependent maintenance of membrane organelle distribution. J Cell Biol
1997;139:469–484.
40. Echeverri CJ, Paschal BM, Vaughan KT, Vallee RB. Molecular charac-
terization of the 50-kD subunit of dynactin reveals function for the
complex in chromosome alignment and spindle organization during
mitosis. J Cell Biol 1996;132:617–633.
41. LaMonte BH, Wallace KE, Holloway BA, Shelly SS, Ascano J, Tokito M,
Van Winkle T, Howland DS, Holzbaur EL. Disruption of dynein/dynactin
inhibits axonal transport in motor neurons causing late-onset pro-
gressive degeneration. Neuron 2002;34:715–727.
42. VogelbaumMA, Tong JX, Rich KM. Developmental regulation of apopto-
sis in dorsal root ganglion neurons. J Neurosci 1998;18:8928–8935.
43. Fleming J, Ginn SL, Weinberger RP, Trahair TN, Smythe JA, Alexander
IE. Adeno-associated virus and lentivirus vectors mediate efficient and
sustained transduction of cultured mouse and human dorsal root
ganglia sensory neurons. Hum Gene Ther 2001;12:77–86.
44. Blocker A, Severin FF, Burkhardt JK, Bingham JB, Yu H, Olivo JC,
Schroer TA, Hyman AA, Griffiths G. Molecular requirements for
bi-directional movement of phagosomes along microtubules. J Cell
Biol 1997;137:113–129.
45. Rich KM, Yip HK, Osborne PA, Schmidt RE, Johnson EM Jr. Role of
nerve growth factor in the adult dorsal root ganglia neuron and its
response to injury. J Comp Neurol 1984;230:110–118.
46. SofroniewMV,CooperJD,SvendsenCN,CrossmanP, IpNY,LindsayRM,
Zafra F, Lindholm D. Atrophy but not death of adult septal cholinergic
neurons after ablation of target capacity to produce mRNAs for NGF,
BDNF, and NT3. J Neurosci 1993;13:5263–5276.
47. Chen KS, Nishimura MC, Armanini MP, Crowley C, Spencer SD,
Phillips HS. Disruption of a single allele of the nerve growth factor
gene results in atrophy of basal forebrain cholinergic neurons and
memory deficits. J Neurosci 1997;17:7288–7296.
48. Howe CL, Valletta JS, Rusnak AS, Mobley WC. NGF signaling from
clathrin-coated vesicles: evidence that signaling endosomes serve as
a platform for the Ras-MAPK pathway. Neuron 2001;32:801–814.
49. Zhang Y, Moheban DB, Conway BR, Bhattacharyya A, Segal RA. Cell
surface Trk receptors mediate NGF-induced survival while internalized
receptors regulate NGF-induced differentiation. J Neurosci 2000;20:
5671–5678.
50. Whitmarsh AJ, Kuan CY, Kennedy NJ, Kelkar N, Haydar TF, Mordes JP,
Appel M, Rossini AA, Jones SN, Flavell RA, Rakic P, Davis RJ.
Requirement of the JIP1 scaffold protein for stress-induced JNK
activation. Genes Dev 2001;15:2421–2432.
51. Lindwall C, Kanje M. Retrograde axonal transport of JNK signaling
molecules influence injury induced nuclear changes in p-c-Jun and
ATF3 in adult rat sensory neurons. Mol Cell Neurosci 2005;29:
269–282.
52. Hayes S, Chawla A, Corvera S. TGF beta receptor internalization into
EEA1-enriched early endosomes: role in signaling to Smad2. J Cell Biol
2002;158:1239–1249.
53. Tang Q, Staub CM, Gao G, Jin Q, Wang Z, Ding W, Aurigemma RE,
Mulder KM. A novel transforming growth factor-beta receptor-
interacting protein that is also a light chain of the motor protein dynein.
Mol Biol Cell 2002;13:4484–4496.
54. Driskell OJ, Mironov A, Allan VJ, Woodman PG. Dynein is required
for receptor sorting and the morphogenesis of early endosomes. Nat
Cell Biol 2007;9:113–120.
55. Cooper JD, Mobley WC. Neurotrophic factors as potential thera-
peutic agents in neuronal ceroid lipofuscinosis. Adv Genet 2001;45:
169–182.
56. Salehi A, Delcroix JD, Mobley WC. Traffic at the intersection of
neurotrophic factor signaling and neurodegeneration. Trends Neurosci
2003;26:73–80.
57. Stokin GB, Lillo C, Falzone TL, Brusch RG, Rockenstein E, Mount SL,
Raman R, Davies P, Masliah E, Williams DS, Goldstein LS. Axonopathy
and transport deficits early in the pathogenesis of Alzheimer’s disease.
Science 2005;307:1282–1288.
58. Goldstein L, Ashford JW, Busciglio J, Crutcher K, Gouras G, Kinoshita J,
Mandelkow E, Sudol M. Live discussion. From here to there: AbetaPP
as an axonal transport receptor–how could this explain neurode-
generation in AD. 15 July 2002. J Alzheimers Dis 2003;5:
483–489.
59. Zhu X, Moreira PI, Smith MA, Perry G. Alzheimer’s disease: an
intracellular movement disorder? Trends Mol Med 2005;11:391–393.
Traffic 2007; 8: 1503–1520 1519
Dynein-/Microtubule-Mediated Trafficking of TrkA for Rap1 Signaling

60. Charrin BC, Saudou F, Humbert S. Axonal transport failure in neurode-
generative disorders: the case of Huntington’s disease. Pathol Biol
(Paris) 2005;53:189–192.
61. Roy S, Zhang B, Lee VM, Trojanowski JQ. Axonal transport defects:
a common theme in neurodegenerative diseases. Acta Neuropathol
(Berl) 2005;109:5–13.
62. Gunawardena S, Goldstein LS. Polyglutamine diseases and transport
problems: deadly traffic jams on neuronal highways. Arch Neurol 2005;
62:46–51.
63. Mobley WC, Rutkowski JL, Tennekoon GI, Gemski J, Buchanan K,
Johnston MV. Nerve growth factor increases choline acetyltransferase
activity in developing basal forebrain neurons. Brain Res 1986;387:53–62.
64. Bronfman FC, Tcherpakov M, Jovin TM, Fainzilber M. Ligand-induced
internalization of the p75 neurotrophin receptor: a slow route to the
signaling endosome. J Neurosci 2003;23:3209–3220.
65. Chan JR, Rodriguez-Waitkus PM, Ng BK, Liang P, Glaser M. Pro-
gesterone synthesized by Schwann cells during myelin formation
regulates neuronal gene expression. Mol Biol Cell 2000;11:2283–2295.
1520 Traffic 2007; 8: 1503–1520
Wu et al.
Copyright © 2022 FDOKUMEN




















