
The induction of tuftelin expression in PC12 cell line during hypoxia and NGF induced...
8
The Induction of Tuftelin Expression in PC12 Cell Line During Hypoxia and NGF-Induced Differentiation YOAV LEISER, 1 NECHAMA SILVERSTEIN, 1 ANAT BLUMENFELD, 1 DEKEL SHILO, 1 AMIR HAZE, 1 ELI ROSENFELD, 1 BOAZ SHAY, 1 RINAT TABAKMAN, 2 SHIMON LECHT, 2 PHILIP LAZAROVICI, 2 AND DAN DEUTSCH 1 * 1 Dental Research Laboratory, Faculty of Dental Medicine, Institute of Dental Sciences, The Hebrew University of Jerusalem— Hadassah, Jerusalem, Israel 2 Faculty of Medicine, School of Pharmacy Institute for Drug Research, The Hebrew University of Jerusalem, Jerusalem, Israel The tuftelin protein isoforms undergo post-translation modifications, and are ubiquitously expressed in various tissues in embryos, adults, and tumors. Developmental and pathological studies suggested an apparent correlation between oxygen deprivation and tuftelin expression. The aim of the study was therefore to investigate the effect of a pathological insult (hypoxia) and a physiological growth factor (NGF), which antagonistically regulate HIF1 expression, on tuftelin expression using the neuronal PC12 cell model. In the present study, we first demonstrated the expression of tuftelin in PC12 cells, providing an experimental system to investigate the pathophysiological role of tuftelin. Furthermore, we demonstrated the induction of tuftelin during hypoxia by oxygen deprivation and during chemical hypoxia by cobalt chloride. Down-regulation of HIF1a mRNA blocked hypoxia-induced HIF1a expression, and reduced by 89% hypoxia-induced tuftelin expression. In mice, intraperitoneal injection of cobalt chloride significantly induced tuftelin mRNA and protein expression in the brain. During NGF-mediated PC12 differentiation, tuftelin expression was significantly induced in correlation with neurite outgrowth. This induction was partially blocked by K252a, a selective antagonist of the NGF receptor TrkA, indicating the involvement of the TrkA-signaling pathways in tuftelin induction by NGF. Revealing the physiological role of tuftelin will clarify mechanisms related to the ‘‘hypoxic genome,’’ and NGF-induced neurotrophic and angiogenic effects. J. Cell. Physiol. 226: 165–172, 2010. ß 2010 Wiley-Liss, Inc. Tuftelin was originally identified and characterized in developing and mature extracellular enamel, the unique and highly mineralized epithelial tissue that covers vertebrate teeth (Deutsch et al., 1991). It has now been shown to be expressed also in morula, embryonic stem cells, various normal soft tissues such as brain (specifically in neurons), kidney, adrenal gland, testis, and various cancerous soft tissues (Mao et al., 2001; Deutsch et al., 2002; Leiser et al., 2007). We have found, using quantitative real-time PCR, that tuftelin mRNA expression levels are significantly higher in the kidney and testis compared to brain, liver, lung, and eye (Leiser et al., 2007). Kidney and testis are hovering closely to hypoxia during normal physiological conditions. Low-oxygen tension has also been reported during cartilage/bone mineralization (Hirao et al., 2007). Recently, we found that hypoxia induces tuftelin expression in mouse mesenchymal stem cells (that can differentiate, amongst other cell types, into cartilage and bone; unpublished), as was also found in the transformed cell lines HepG2 and MCF-7 (Saarikoski et al., 2002). Therefore, the rational of this study proposes that tuftelin has an important role during hypoxic conditions and during differentiation. Tuftelin was previously studied mainly at the level of gene and protein expression, with regards to tissue distribution and localization. However, to investigate its physiological role, an in vitro cellular model was needed. Since tuftelin is expressed in the adrenal gland, mainly in the chromaffin medulary cells (neural crest origin) from which the transformed pheochromocytoma PC12 cells were derived, and also in other neuronal tissues such as brain neurons and retina of the eye (Leiser et al., 2007), we decided to explore tuftelin presence and expression in the PC12 neuronal cell model. PC12 cells are characterized by a hypoxia tolerant phenotype that allows them to survive and function in a low- oxygen environment (Seta et al., 2002). In addition, they are a widely used neuronal model to study mechanisms of differentiation by NGF (Greene and Tischler, 1976; Fujita et al., 1989; Vaudry et al., 2002). In the PC12 cell model the oxidative stress insult is achieved by oxygen deprivation and/or oxygen– glucose deprivation using a special ischemic device (Tabakman et al., 2002), or by using cobalt chloride treatment (Kotake-Nara and Saida, 2007). In both cases, induction of HIF1 pathway is regulating oxygen sensitive genes implicated in the adaptive mechanisms, regulating protective processes (Huang and Bunn, 2003). A common model for in vivo induction of hypoxia is intra-peritoneal injection of cobalt chloride (Bergeron et al., 2000; Saarikoski et al., 2002). Therefore, CoCl 2 injection allows in vivo investigation of induction of various genes in relation to this chemical hypoxia insult, in different tissues including the brain. Contract grant sponsor: U.S-Israel Binational Science Foundation (BSF); Contract grant number: 2003-239. *Correspondence to: Dan Deutsch, Faculty of Dental Medicine, Dental Research Laboratory, Institute of Dental Sciences, The Hebrew University, Hadassah, Jerusalem 91120, Israel. E-mail: [email protected] Received 21 July 2009; Accepted 30 June 2010 Published online in Wiley Online Library (wileyonlinelibrary.com.), 23 July 2010. DOI: 10.1002/jcp.22318 ORIGINAL ARTICLE 165 Journal of Journal of Cellular Physiology Cellular Physiology ß 2010 WILEY-LISS, INC.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of The induction of tuftelin expression in PC12 cell line during hypoxia and NGF induced...

ORIGINAL ARTICLE 165J o u r n a l o fJ o u r n a l o f
CellularPhysiologyCellularPhysiology
The Induction of TuftelinExpression in PC12 Cell LineDuring Hypoxia and NGF-InducedDifferentiation
YOAV LEISER,1 NECHAMA SILVERSTEIN,1 ANAT BLUMENFELD,1 DEKEL SHILO,1 AMIR HAZE,1ELI ROSENFELD,1 BOAZ SHAY,1 RINAT TABAKMAN,2 SHIMON LECHT,2 PHILIP LAZAROVICI,2
AND DAN DEUTSCH1*1Dental Research Laboratory, Faculty of Dental Medicine, Institute of Dental Sciences, The Hebrew University of Jerusalem—
Hadassah, Jerusalem, Israel2Faculty of Medicine, School of Pharmacy Institute for Drug Research, The Hebrew University of Jerusalem, Jerusalem, Israel
The tuftelin protein isoforms undergo post-translation modifications, and are ubiquitously expressed in various tissues in embryos, adults,and tumors. Developmental and pathological studies suggested an apparent correlation between oxygen deprivation and tuftelinexpression. The aim of the study was therefore to investigate the effect of a pathological insult (hypoxia) and a physiological growth factor(NGF), which antagonistically regulate HIF1 expression, on tuftelin expression using the neuronal PC12 cell model. In the present study,we first demonstrated the expression of tuftelin in PC12 cells, providing an experimental system to investigate the pathophysiological roleof tuftelin. Furthermore, we demonstrated the induction of tuftelin during hypoxia by oxygen deprivation and during chemical hypoxia bycobalt chloride. Down-regulation of HIF1a mRNA blocked hypoxia-induced HIF1a expression, and reduced by 89% hypoxia-inducedtuftelin expression. In mice, intraperitoneal injection of cobalt chloride significantly induced tuftelin mRNA and protein expression in thebrain. During NGF-mediated PC12 differentiation, tuftelin expression was significantly induced in correlation with neurite outgrowth.This induction was partially blocked by K252a, a selective antagonist of the NGF receptor TrkA, indicating the involvement of theTrkA-signaling pathways in tuftelin induction by NGF. Revealing the physiological role of tuftelin will clarify mechanisms related to the‘‘hypoxic genome,’’ and NGF-induced neurotrophic and angiogenic effects.
J. Cell. Physiol. 226: 165–172, 2010. � 2010 Wiley-Liss, Inc.
Contract grant sponsor: U.S-Israel Binational Science Foundation(BSF);Contract grant number: 2003-239.
*Correspondence to: Dan Deutsch, Faculty of Dental Medicine,Dental Research Laboratory, Institute of Dental Sciences, TheHebrew University, Hadassah, Jerusalem 91120, Israel.E-mail: [email protected]
Received 21 July 2009; Accepted 30 June 2010
Published online in Wiley Online Library(wileyonlinelibrary.com.), 23 July 2010.DOI: 10.1002/jcp.22318
Tuftelin was originally identified and characterized indeveloping and mature extracellular enamel, the unique andhighly mineralized epithelial tissue that covers vertebrate teeth(Deutsch et al., 1991). It has now been shown to be expressedalso in morula, embryonic stem cells, various normal soft tissuessuch as brain (specifically in neurons), kidney, adrenal gland,testis, and various cancerous soft tissues (Mao et al., 2001;Deutsch et al., 2002; Leiser et al., 2007). We have found, usingquantitative real-time PCR, that tuftelin mRNA expressionlevels are significantly higher in the kidney and testis comparedto brain, liver, lung, and eye (Leiser et al., 2007). Kidney andtestis are hovering closely to hypoxia during normalphysiological conditions. Low-oxygen tension has also beenreported during cartilage/bone mineralization (Hirao et al.,2007). Recently, we found that hypoxia induces tuftelinexpression in mouse mesenchymal stem cells (that candifferentiate, amongst other cell types, into cartilage andbone; unpublished), as was also found in the transformedcell lines HepG2 and MCF-7 (Saarikoski et al., 2002).Therefore, the rational of this study proposes that tuftelin hasan important role during hypoxic conditions and duringdifferentiation.
Tuftelin was previously studied mainly at the level of gene andprotein expression, with regards to tissue distribution andlocalization. However, to investigate its physiological role, an invitro cellular model was needed. Since tuftelin is expressed inthe adrenal gland, mainly in the chromaffin medulary cells(neural crest origin) from which the transformedpheochromocytoma PC12 cells were derived, and also in otherneuronal tissues such as brain neurons and retina of the eye(Leiser et al., 2007), we decided to explore tuftelin presenceand expression in the PC12 neuronal cell model.
� 2 0 1 0 W I L E Y - L I S S , I N C .
PC12 cells are characterized by a hypoxia tolerantphenotype that allows them to survive and function in a low-oxygen environment (Seta et al., 2002). In addition, they are awidely used neuronal model to study mechanisms ofdifferentiation by NGF (Greene and Tischler, 1976; Fujita et al.,1989; Vaudry et al., 2002). In the PC12 cell model the oxidativestress insult is achieved by oxygen deprivation and/or oxygen–glucose deprivation using a special ischemic device(Tabakman et al., 2002), or by using cobalt chloride treatment(Kotake-Nara and Saida, 2007). In both cases, induction of HIF1pathway is regulating oxygen sensitive genes implicated in theadaptive mechanisms, regulating protective processes (Huangand Bunn, 2003). A common model for in vivo induction ofhypoxia is intra-peritoneal injection of cobalt chloride(Bergeron et al., 2000; Saarikoski et al., 2002). Therefore,CoCl2 injection allows in vivo investigation of induction ofvarious genes in relation to this chemical hypoxia insult, indifferent tissues including the brain.

166 L E I S E R E T A L .
During the development of the nervous system,neurotrophic factors such as NGF play a major role in regulatingneuronal survival, proliferation, differentiation, and maturation(Sofroniew et al., 2001). NGF was also found to reduce andeven prevent apoptosis and neuronal death, induced byhypoxia/ischemia events, through activation of the TrkAreceptor-signaling pathway (Tabakman et al., 2004a,b). NGF isimportant not only during CNS development and injury but alsoin the cardiovascular system and craniofacial complexdevelopment and function (Amano et al., 1999; Nosrat et al.,2002; Lazarovici et al., 2006). In view of the wide expression oftuftelin already from very early stages of embryonicdevelopment, and the important role of NGF in controllingtissue innervation during very early embryonic development,we hypothesize that NGF might regulate tuftelin expression.
To explore this possibility we investigated tuftelinexpression in the PC12 neuronal model, and its possibleregulation by NGF and by hypoxia.
In the present study, we have found that tuftelin expression(mRNA and protein) is induced in PC12 cells during hypoxia,and is correlated with HIF1a mRNA expression. NGF alsoinduced tuftelin expression during PC12 cell differentiation, byTrkA-mediated signaling pathways. These findings may suggesta physiological role for tuftelin in neuroprotective pathways insympathetic neurons and in other NGF-mediated processessuch as neuronal differentiation.
Materials And MethodsAnimals and tissue preparation
Three-month old Balb/c mice were subjected to intra-peritoneal injection of 70 mg/kg CoCl2 (experimental) or saline(control). Five animals were included in each group. Mice weresacrificed 12 h after treatment; brain tissue was harvested andimmediately immersed in liquid nitrogen for further mRNA andprotein analysis. All experiments were approved by the AnimalCare Ethical Committee, Faculty of Medicine, The HebrewUniversity of Jerusalem.
PC12 cell culture
PC12 cells were grown in Dulbeco’s modified Eagle’s medium(DMEM), supplemented with 7% fetal calf serum, 7% horseserum, 100mg/ml streptomycin, and 100 U/ml penicillin (Be’itHa’emek, Afula, Israel). The cultures were maintained in ahumidified atmosphere of 6% CO2 at 378C. The medium waschanged twice weekly and the cultures were split at 1:6 ratioonce a week. Unless otherwise specified, about 1� 106 cellswere used. In the experiments involving the use of HIF1asiRNA, about 2� 105 cells were used, according to themanufacturer’s instructions (Ambion, Austin, TX), to maximizethe transfection efficiency. In experiments involving tuftelininduction by NGF, about 4� 105 cells were used, in order toallow neurite elongation. Cells were suspended in DMEM andplated onto 35-mm Petri-dishes coated with rat tail type Icollagen (0.1 mg/ml; Be’it Ha’emek; Tabakman et al., 2002).
Immunohistochemistry
Adrenal gland slices were mounted on microscopic cover slipscoated with mounting media (Zymed, Histostain Bulk kit;Zymed LAB-SA Detection System, San-Francisco, CA). PC12cells were grown on chamber slides (Lab-Tek, Nunc) coatedwith 0.1 mg/ml collagen type 1 (Be’it Ha’emek). Tissue sliceswere de-paraffinized with Xylene, and re-hydrated in a gradedseries of ethanol. The immuno-staining protocol included thefollowing steps: (i) Peroxidase quenching of the tissue and cells,which was performed by submerging the tissue in 1 ml of30% H2O2 diluted in methanol at a ratio of 9:1 for 10 min.(ii) Blocking the non-specific binding of the tissue or cells by 10%
JOURNAL OF CELLULAR PHYSIOLOGY
non-immune goat serum, by submerging the tissue or cells in200ml PBS for 15 min at room temperature. (iii) Overnightincubation at 48C in a humidified chamber with the polyclonalanti-tuftelin antibodies LF74 and LF75 (Deutsch et al., 1991),diluted 1:200 in 200ml PBS. (iv) After rinsing four times in200 ml PBS, the preparations were incubated with biotinylatedanti-rabbit second antibody for 10 min. (v) For visualization,100ml horseradish peroxidase conjugate was added for 10 min.After two rinses with PBS, counter staining of the preparationswas performed by 5 min staining with hematoxylin. Thepreparations were washed with distilled water for 2 min,followed by a wash with PBS, mounting of the coverslips onmicroscopic slides, drying, and microscopic visualization.
Western blotting
Total protein was extracted from frozen whole brain tissue orPC12 cells after homogenization in lysis buffer (0.25 M sucrose,20 mM Tris pH 7.6, 1.5 mM MgCl2, 10% glycerol, 1 mM EDTA,100 mM PMSF and PI cocktail� 50). Equal amounts of totalprotein, determined by the Bradford assay, were loaded onto4–20% Tris–glycine precast gels (Novex). SDS–PAGE wasperformed using a minicell apparatus (Invitrogen, Carlsbad, CA)at 150 V for 50 min using Tris-running buffer (25 mM TrispH 8.8, 1% (v/v) SDS, 0.2 M glycine). Following SDS–PAGE,proteins were transferred onto Protran1 nitrocellulosemembranes (Schleicher & Schuell BioScience, Dassel,Germany) by electrophoretic blotting (BioRad apparatus;BioRad, Munich, Germany) at 19 mA for 60 min. Thenitrocellulose membranes were incubated in ‘‘superblotto’’buffer (10% (v/v) PBS; 1 M glucose; 3% (w/v) BSA; 1% (w/v) milkpowder; 10% (v/v) glycerol; 0.5% (v/v) Tween-20TM) shaking at378C for 2 h. The membranes were than incubated shakingovernight at room temperature with tuftelin polyclonalantibodies LF 74, LF 75 (Deutsch et al., 1991), diluted in 1% (w/v)gelatine–Tris-buffered saline, followed by incubation in alkalinephosphatase-conjugated anti-rabbit IgG (Promega, Madison,WI; diluted 1:7,500) for 2 h at room temperature. Visualizationwas performed with BCIP/NBT reagents (Promega). Bandoptical density intensity measurements at 595 nm werecalculated by the Tina 2.0 software. Results were calculated asband intensity after reduction of background density.
Quantitative real-time PCR
Total RNA was extracted by homogenizing the frozen braintissues or PC12 cells in TRI-REAGENT (MRC, Cincinnati, OH).RNA isolation was performed using the TRI-REAGENTstandard protocol. Total RNA extracts were subjected toDNase treatment (DNA free; Ambion) to eliminate anypossible DNA contamination. Each sample was checked forquality on 1% agarose gel prior to reverse transcription, and thetotal RNA concentration was determined using the NanoDropND-1000 spectrophotometer (NanoDrop Technologies,Wilmington, DE). Total RNA was subjected to reversetranscription according to the manufacturer’s protocol(superscript RNase H- Reverse Transcriptase kit; Invitrogene,Carlsbad, CA). Real-time quantitative PCR was performedusing the LightCycler apparatus (Roche, Basel, Switzerland).The real-time quantitative PCR reactions were performed in avolume of 10ml of a mixture containing the reverse transcribedcDNA product, tuftelin specific oligonucleotide primers(forward 50-TGGACCCTAGCATGAGT-30, reverse50-CGTTCTTGATCCGAAGC-30, amplicon size 312 bp) and1ml of DNA Master SYBR green mix (Roche MolecularBiochemistry, Mannheim, Germany, including; Taq DNApolymerase, reaction buffer and dNTPs). The fluorescent signalwas plotted in real time against temperature to produce amelting curve for each sample. At the end of the run the

T U F T E L I N I N D U C T I O N B Y H Y P O X I A A N D N G F 167
specificity of the reaction was determined by analysis of thePCR products on a 1–3.5% agarose gel (Leiser et al., 2007).
In the following experiments, several modifications to theabove protocol were used: (i) hypoxia effect on HIF1ainduction, (ii) the effect of siRNA on hypoxia-induced HIF1aand tuftelin expression, (iii) The effect of NGF on tuftelinexpression. These modifications include: (i) absence ofDNase treatment, (ii) reverse transcription was performedusing high-capacity cDNA reverse transcription kit (AppliedBiosystems, Austin, TX) according to the manufacturer’sprotocol, (iii) real-time quantitative PCR was performed usingApplied Biosystem’s 7300 Real-Time PCR System with AppliedBiosystem’s TaqMan1 AOD (Assays-On-Demand) and MasterMixes (TaqMan1, Gene Expression Assays, AOD;HIF1a¼RN00577560_M1, beta actin[ACTB]¼RN00667869_M1, Assay Made to Order, AMO,Tuftelin [TUFT1]¼RN01762781_M1). The real-timequantitative PCR reactions were performed in a volume of20ml of a mixture containing 40 ng of the reverse transcribedcDNA product, TaqMan master mix and AOD/AMO. Theresults were normalized to beta actin and shown as foldincrease as compared to their respective controls.
Transfection and siRNA
To knock down HIF1a, the standard Amine Transfection Agentprotocol was followed (Ambion). In brief, 10 nM of Silencer1
Select Pre-designed siRNA, s131712, cat # 4390771 andSilencer1 Select Negative Control (scrambled)#1 siRNA[SCRAM] cat # 4390843 (Ambion), diluted in Gibco1 OPTI-MEM1I (Invitrogen) was combined with siPORTTM Aminetransfection agent (cat # AM4502) diluted in Gibco1 OPTI-MEM1I (Invitrogen). This mixture was added to PC12 cells andincubated for 24 h, at 378C under 6% CO2. The cells were thensubjected to hypoxic insult for 24 h in a hypoxic chamber (seebelow), followed by RNA extraction and LDH assays.
LDH release cell death assay
Cell death was measured at the end of the hypoxia period byquantifying the activity of released LDH into the medium, using acommercial kit (Pointe, Canton, MI). LDH activity wasdetermined spectrophotometrically at 340 nm by following therate of conversion of oxidized nicotinamide adeninedinucleotide (NADþ) to the reduced form (NADH). TotalLDH (extracellularþ intracellular) was obtained by freezingand thawing the cultures. The basal LDH release measured incultures maintained under normoxia was subtracted from allthe experimental values. Hypoxia-induced cell death wasexpressed as percentage of released LDH from total LDHactivity (Tabakman et al., 2002).
In-vitro hypoxia
To induce the oxygen deprivation insult, PC12 cells wereintroduced into a hypoxic device, as was previously described(Tabakman et al., 2002). Briefly, the hypoxic device is composedof two connected chambers, maintained at 378C by circulatinghot water into which Petri-dishes with PC12 cells are placed.The air in the chambers is replaced with a mixture of 99% N2
and 1% O2 bubbled from a tank through a cylinder containingwater, and passed through the system at a reduced flow rate toprevent evaporation of the medium. The oxygen level withinthe device is 1% (as measured online by an oxygen analyzer),representing a hypoxic insult. Control cells were kept in thehumidified chamber in an atmosphere containing 21% O2
(normoxia). Cells were harvested and treated as describedabove.
JOURNAL OF CELLULAR PHYSIOLOGY
In-vitro chemical hypoxia
PC12 cells were exposed to 100mM cobalt chloride for 24 h.Cells were harvested and tuftelin mRNA expression levelswere quantified using the real-time PCR. Tuftelin proteinexpression was analyzed by Western blotting. Control sampleswere treated with the same volume of phosphate-bufferedsaline (PBS) for 24 h.
NGF-induced differentiation in PC12 cells
Mouse beta NGF (50 ng/ml; Alomone Labs, Jerusalem, Israel)was applied daily to PC12 cell culture to induce differentiation.Cells were harvested after different exposure intervals to NGF(from 1 to 5 days), and tuftelin mRNA expression levels werequantified using the real-time quantitative PCR. Results werecompared to control cultures of undifferentiated PC12 cells. Toinhibit NGF-induced differentiation, PC12 cultures were pre-treated with K252a (Fermentek, Jerusalem, Israel) at a finalconcentration of 200 nM (Koizumi et al., 1988), 30 min prior tothe daily application of NGF. The levels of tuftelin mRNA werequantified using real-time PCR, 5 days after the beginning of theexperiment.
Neuronal differentiation analysis
To assess NGF-induced neuronal differentiation, neuriteoutgrowths length was measured using the SigmaScanPro 5.0program as was previously described by us (Katzir et al., 2002).Briefly, the length of neurite outgrowth and the diameter ofeach cell were measured. Thereafter, by dividing the totallength of a specific cell by its diameter, an elongation factor,denoted elongation (E) parameter was generated. Theexperiments were performed at least three times in duplicates.In each well three random areas were photographed. In eacharea, 15–25 cells were measured, that is, at least 270 cells weremeasured for each time point. The micrographs were acquiredin 2560� 1920 pixel dimensions and with a resolution of300 pixels/inch.
Statistical methods
In general, in vitro experiments were performed three times(N¼ 3) and experimental groups were measured in triplicates.In the in vivo experiments five animals were included in eachgroup (N¼ 5), and experimental groups were measured intriplicates (n¼ 3). To overcome normal gross fluctuations ingene expression level observed in cultures or betweenindividual animals, mRNA expression levels were normalized tothe expression of the reference gene beta-actin, and presentedas fold increase over control. When only two groups of resultswere compared, the Student’s t-test was used.
For multiple comparisons, the comparisons between thedifferent groups were performed using the ANOVA test, withpost hoc tests and the Dunnett’s correction of the significancelevel for multiple pairwise comparisons. In addition, the non-parametric Kruskal–Wallis ANOVA test was applied, and theMann–Whitney test was applied for pairwise comparisons, withthe Bonferroni correction of the significance level for multiplepairwise comparisons. P-values> 0.01 were considered asstatistically significant.
ResultsTuftelin expression in the adrenal gland and inundifferentiated PC12 cells
The expression of tuftelin mRNA and protein was previouslyshown in many mammalian species including human, bovine,rabbit, mouse, and rat (Deutsch et al., 1995, 2002; Leiser et al.,2007). The expression of tuftelin mRNA in chromaffin cells of

168 L E I S E R E T A L .
mouse adrenal gland was previously detected using in situhybridization (Leiser et al., 2007). Expression of tuftelin mRNAin PC12 cells was first established by RT-PCR, usingoligonucleotides designed for amplification of the full-lengthtuftelin mouse mRNA (MacDougall et al., 1998; GenBank No.AF047704). The tuftelin mRNA isoform in PC12 cells (ratorigin) was found by us to contain 13 exons with exon 2 splicedout. This is a common isoform of tuftelin in mouse and humansoft tissues (liver and kidney) as well as in human tooth bud(Mao et al., 2001).
The expression of tuftelin protein was studied using tuftelinantibodies (LF74, LF75; Deutsch et al., 1991). Tuftelin proteinexpression (red staining) was detected in chromaffin cells in themedulla of the mouse adrenal gland (Fig. 1C, black arrow).Fainter staining was seen in the inner cortex of the adrenal gland(Fig. 1C, white arrow). Rat chromaffin cells are the source of theneuronal model cell line PC12 that was used in the presentstudy. Due to the ubiquitous expression of tuftelin in differenttissues and species and the cross reactivity of tuftelinantibodies, tuftelin protein expression was studied in PC12cells. Tuftelin protein expression was detected mainly in thecytoplasm of undifferentiated PC12 cells (Fig. 1A). No reactionwas detected in the control PC12 cells, using PBS instead of thefirst antibody (Fig. 1B).
Hypoxia-induced expression of tuftelin and HIF1a genesin a time-dependent manner
To induce oxygen deprivation (1% O2), PC12 cells wereintroduced into a hypoxic device from 2 to 24 h. Tuftelin andHIF1a expression were quantified at each time-point using real-time PCR and the results were normalized to the expression ofb-actin. Hypoxia significantly stimulated by 5.2-fold tuftelinmRNA expression in PC12 cells after 24 h (Fig. 2A), and tuftelinprotein expression (data not shown), as compared to control(normoxia). During this time frame cell death, as indicated byLDH assay, was<2% (data not shown), indicating the inductionof tuftelin by hypoxia and not by cell death.
HIF1a was proposed, but not shown, as a potential inducerof tuftelin gene expression during hypoxia in HepG2 and MCF7transformed cell lines (Saarikoski et al., 2002). Therefore, weinvestigated the effect of hypoxia on HIF1a mRNA expression(Fig. 2B), as was described above for tuftelin. The level of HIF1ain normoxic cells was low, as was shown for tuftelin expressionunder normoxia. The expression of HIF1a mRNA wassignificantly induced after 6 and after 24 h of exposure tohypoxia (2- and 3.9-fold, respectively), as compared tonormoxia.
Fig. 1. Tuftelin expression in undifferentiated PC12 cells and in the mourevealedthattuftelin isexpressed inthecytoplasmofratundifferentiatedPreplacing the first antibody. C: Tuftelin expression was also detected in chrFainterstainingwasseen inthe innercortexof theadrenalgland(whitearroavailable at wileyonlinelibrary.com.]
JOURNAL OF CELLULAR PHYSIOLOGY
It is well-established that the Vascular Endothelial GrowthFactor (VEGF) expression is induced during hypoxia via theHIF1a-mediated pathway (Ferrara, 2005; Wang et al., 2007),and was previously studied in PC12 cells (Levy et al., 1996).Hence, VEGF was used as a positive control for hypoxiainduction. VEGF mRNA levels were quantified after incubationof PC12 cells in the hypoxic device for different time spans ascompared to cells grown under normal oxygen level (Fig. 2C).The expression of VEGF was significantly induced by eight-fold,already 2 h after incubation in the hypoxic device. The increasein VEGF mRNA further progressed in a time-dependentmanner, reaching 18.8-fold induction, compared to the controlPC12 cells at normoxia, at the latest time-point studied after24 h of hypoxia, confirming the presence of hypoxic conditionsthroughout the experiment.
HIF1a siRNA inhibits tuftelin induction by hypoxia
HIF1 is a major pathway involved in the induction of oxygensensitive genes. To explore the potential relationship betweentuftelin and HIF1a induction under 1% oxygen level, weemployed siRNA-directed against HIF1a. PC12 cells weretransfected with either HIF1a–siRNA, with a scrambled-siRNAto determine the selectivity of siRNA transfection, or were leftuntreated (control). Twenty-four hours later, the transfectedand untreated PC12 cells were introduced into a hypoxic devicefor 24 h. Another group of untreated cells were maintainedunder normoxic conditions for additional 24 h.
The hypoxic conditions induced tuftelin and HIF1aexpression by 3.7- and 1.8-fold, respectively. Transfection withthe scrambled siRNA did not affect tuftelin and HIF1a geneexpression. Transfection with HIF1a-siRNA decreased HIF1aexpression to below the normoxia (control) level. In the sameexperiments, the level of tuftelin expression was decreased by89%, compared with hypoxic cultures or with PC12 cellcultures transfected with the scrambled siRNA (Fig. 3). Culturemedia of all the experimental and control groups were analyzedfor cell death by LDH-leakage assay. The amount of LDHrelease was <2%, indicating the lack of cytotoxicity of thetransfection protocol, of the specific siRNA used, and of thehypoxic conditions.
Chemical hypoxia up-regulated tuftelin expression inPC12 cells
Since hypoxia induced by oxygen starvation can be mimicked byaddition of cobalt chloride, we also treated PC12 cells with100mM cobalt chloride. The chemical hypoxia, caused by
se adrenal gland. A: Immunohistochemistry, using tuftelin antibodies,C12cells (redstaining).B:NoreactionwasobservedwithPBS(control)omaffin cells in the medulla of the mouse adrenal gland (black arrow).w)bar U 10mm.[Colorfigurecanbeviewedintheonline issue,which is

Fig. 2. Induction of tuftelin and HIF1a mRNA expression in undifferentiated PC12 cell cultures during oxygen deprivation. PC12 cells wereexposed to hypoxia for 2–24 h using a specially designed hypoxic chamber. A: Tuftelin, B: HIF1a, C: VEGF mRNA levels were quantified using real-time PCR and presented as mean fold increase over control normalized to the expression level of beta-actin mRNA W S.E. A,C: and W S.D.B: hypoxia (gray); normoxia (black); MP < 0.01, by ANOVA.
T U F T E L I N I N D U C T I O N B Y H Y P O X I A A N D N G F 169
CoCl2 treatment, significantly induced by 2.5-fold tuftelinmRNA, as compared to control (PBS) undifferentiated PC12cells (Fig. 4A). VEGF mRNA levels were quantified as positivecontrol. VEGF expression was significantly induced by 6.9-foldwhen 100mM cobalt chloride was applied to PC12 cells, ascompared to the control cultures (Fig. 4B).
Two major tuftelin protein bands of 64 and 55 kDa weredetected in PC12 cell line by Western blot, using the LF 74anti-tuftelin antibody (Fig. 4C, control). Chemical hypoxia,induced by 100mM cobalt chloride, caused significant increasein the expression of the 64 kDa tuftelin protein, while only amild increase in the expression of the 55 kDa tuftelin band wasdetected (Fig. 4C, CoCl2). Quantification of these two proteinbands revealed that the tuftelin protein was significantlyinduced by 2.9-fold, by 100mM cobalt chloride, as compared tocontrol cultures (Fig. 3B).
Fig. 3. The inhibitory effect of HIF1a siRNA on hypoxia-inducedtuftelin and HIF1a mRNA expression in PC12 cell culture. The cellcultures were transfected with siRNA directed against HIF1a.Control cells were transfected with a scrambled siRNA or were leftuntreated. 24 h later the transfected and untreated cells wereintroduced into a hypoxic device for 24 h, and a group of untreatedcells were left under normoxic conditions for additional 24 h. Tuftelin(diagonal lines bar) and HIF1a (gray bar) mRNA levels werequantified and presented as mean W S.D. MP < 0.01, compared tonormoxia, MMP < 0.01 compared to hypoxia, by Student’s t-test.
JOURNAL OF CELLULAR PHYSIOLOGY
Chemical hypoxia up-regulated tuftelin expression in themouse brain
To confirm that hypoxia-induced tuftelin expression is notlimited to cell lines but also occur in vivo, we employed chemical
Fig. 4. Cobalt chloride-induced chemical hypoxia increased tuftelinmRNA and upregulated tuftelin protein expression in PC12 cultures.A: Tuftelin, B: VEGF mRNA expression were quantified 24 h afterapplication of 100mM cobalt chloride (CoCl2), using real-time PCR.Results (mean W S.E.) were normalized to b-actin mRNA levels andpresented as fold increase over control. C: Western blot analysis oftuftelin in PC12 cells using the anti-tuftelin antibody LF 74 and anti-actin antibody or control PBS. D: Quantification of tuftelin proteinexpression was performed using Tina 2.0 software. Band intensity wasnormalizedtob-actinproteinexpression.MP < 0.01,byStudent’st-test.

Fig. 5. Inductionof brain tuftelinmRNAandup-regulation of tuftelinproteinafter injectionofCoCl2.A:Tuftelin,B:VEGFmRNAlevels12 hafter intraperitoneal injection of 70 mg/kg CoCl2 in Balb/C mice ascompared to injection of PBS (control). Results (mean W S.E.) werenormalizedtob-actinmRNAlevelsandpresentedasfoldincreaseovercontrol. C: Western blot analysis using anti-tuftelin antibodies (LF-74,LF-75 mix) and anti-actin antibodies. D: Quantification of tuftelinprotein expression in the mouse brain using TINA 2.0 software. Bandintensity was normalized to b-actin protein expression. MP < 0.01, byStudent’s t-test.
170 L E I S E R E T A L .
hypoxia, induced by cobalt chloride, in mice. Cobalt chloride(70 mg/kg), injected intra-peritoneally (IP) into Balb/C mice,caused significant induction, by twofold, of tuftelin mRNA in themouse brain 12 h after injection, as compared to control (IPinjection of normal saline; Fig. 5A). VEGF mRNA level was alsosignificantly induced in vivo by CoCl2 by 2.6-fold (Fig. 5B).Western blot analysis revealed an increase in the expression ofthe 64 kDa tuftelin protein in the mouse brain, 12 h after IPinjection of CoCl2, as compared to IP injection of normal saline(Fig. 5C). No substantial change in expression level wasdetected with the minor 36, 45, and 55 kDa tuftelin bands(Fig. 5C). Quantification of the 64 kDa tuftelin band revealedthat the tuftelin protein was significantly induced by sixfold afterIP injection of CoCl2 (Fig. 5D).
NGF-induced tuftelin mRNA expression in relation toPC12 cell differentiation
After demonstrating the induction of tuftelin expression undera hypoxic insult, we proceeded with the investigation of itspotential induction under physiological conditions (normoxia).Candidate growth factors, which were considered for tuftelinstimulation were angiogenic factors such as VEGF and NGF(Lazarovici et al., 2006). PC12 cells release VEGF but do notrespond to VEGF treatment, due to the lack of VEGF receptors(Levy et al., 1996). However, PC12 cells do respond to NGF,
JOURNAL OF CELLULAR PHYSIOLOGY
hence, we studied NGF-induced tuftelin gene expression, inrelation to NGF-induced morphologic differentiation.
To verify a temporal possible correlation between themorphological differentiation of PC12 cells and tuftelinexpression, we treated PC12 cell cultures with 50 ng/ml NGFfor 1–5 days. Treatment with 50 ng/ml NGF, induced PC12 celldifferentiation, expressed by progressive elongation of neuriteoutgrowths (Fig. 6, upper part). The elongation factor (the ratiobetween neurite outgrowth length and cell diameter)represents an increase from a value of 1.0 in untreated cells to avalue of 10 in cells treated with NGF for 5 days. The expressionof tuftelin mRNA was quantified at the same time-points whenneurite elongation was measured. Tuftelin expression wassignificantly decreased after 1 day of NGF treatment.Thereafter, a gradual significant increase in tuftelin expressionwas observed from day 2 up to 5, in a positive correlation withneurite outgrowth elongation (Fig. 6, lower part).
To confirm that tuftelin expression induced by NGF isreceptor-mediated, PC12 cells were treated with K252a, aknown antagonist of NGF-trkA receptor. K252a (200 nm) wereapplied to PC12 cells 30 min prior to application of 50 ng/mlNGF, and supplemented daily for 5 days. Fifty-five percentinhibition of NGF-induced tuftelin expression was detected5 days after the beginning of NGF application (Fig. 7). At thistime point 60% inhibition of NGF-induced neurite outgrowthelongation occurred (data not shown).
Discussion
In the present study, we first demonstrated the expression oftuftelin protein in PC12 cells, providing an experimental systemto investigate the physiological role of tuftelin. Furthermore, wedemonstrated the induction of tuftelin mRNA and proteinduring hypoxia by oxygen deprivation, and during chemicalhypoxia by cobalt chloride. These results are in-line with tuftelininduction during hypoxia in cancerous cell lines (Saarikoskiet al., 2002). The induction of tuftelin mRNA was significantafter 24 h of hypoxic insult (1% O2), which correlated with asignificant induction of HIF1a mRNA after 24 h of oxygendeprivation, indicating a temporal correlation of tuftelin andHIF1a induction. Knockdown of HIF1a by a selective siRNAprobe, down-regulated the expression of HIF1a mRNA tobelow normoxic level, and significantly down-regulated tuftelinmRNA by 89%, demonstrating a correlation between theinduction and down-regulation of these two genes. This findingstrongly supports the concept of the involvement of HIF1a inthe mechanism of induction of tuftelin during hypoxia. Thepresence of two putative HIF consensus DNA binding sites,compatible with hypoxia responsive elements (HRE, 5’-RCGTG-3’) in the tuftelin promoter region, at positions�1296, �27 of the human tuftelin gene (upstream to exon 1;GenBank: AH009496.1), and at positions �130, �8 of themouse tuftelin promoter (GenBank: NC_000069.5), furthersupport this concept.
Although tight regulation of HIF1a stability is believed to becontrolled mainly by its post-translational modifications(Brahimi-Horn et al., 2005), it is becoming clear that in differentorgans and cells, the balance between transcriptional(Wiesener et al., 1998; Ke and Costa, 2006), post-transcriptional, translational and post-translational processing,as well as nuclear trafficking and trans-activation, regulateHIF1a (Semenza, 2000, 2009). More recently Tai et al., (2009)showed that in sympathetic neuronal cell model such as PC12,the hypoxic stress controls adrenergic functions viatranscriptional and translational induction of HIF1a. Therefore,it is tempting to propose that HIF1a induces tuftelin promoteractivity as reported for PNMT promoter (Tai et al., 2009). Themechanism of regulation of tuftelin by HIF1a protein will beinvestigated in future studies.
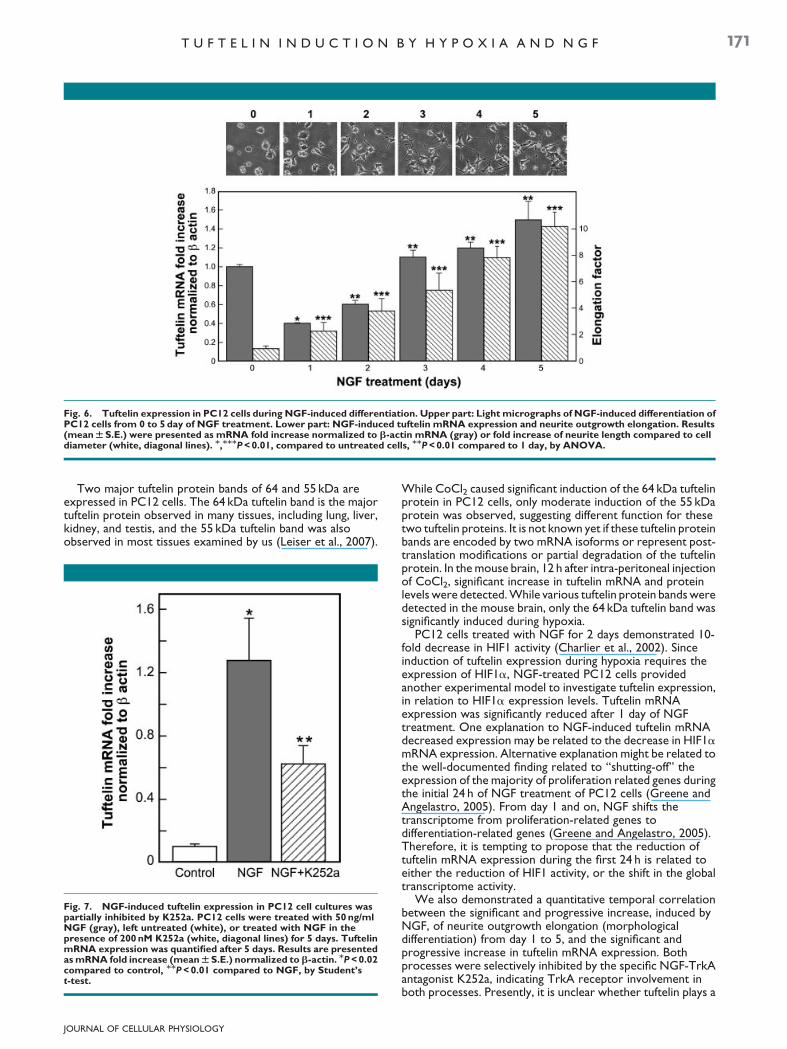
Fig. 6. Tuftelin expression in PC12 cells during NGF-induced differentiation. Upper part: Light micrographs of NGF-induced differentiation ofPC12 cells from 0 to 5 day of NGF treatment. Lower part: NGF-induced tuftelin mRNA expression and neurite outgrowth elongation. Results(mean W S.E.) were presented as mRNA fold increase normalized to b-actin mRNA (gray) or fold increase of neurite length compared to celldiameter (white, diagonal lines). M,MMMP < 0.01, compared to untreated cells, MMP < 0.01 compared to 1 day, by ANOVA.
T U F T E L I N I N D U C T I O N B Y H Y P O X I A A N D N G F 171
Two major tuftelin protein bands of 64 and 55 kDa areexpressed in PC12 cells. The 64 kDa tuftelin band is the majortuftelin protein observed in many tissues, including lung, liver,kidney, and testis, and the 55 kDa tuftelin band was alsoobserved in most tissues examined by us (Leiser et al., 2007).
Fig. 7. NGF-induced tuftelin expression in PC12 cell cultures waspartially inhibited by K252a. PC12 cells were treated with 50 ng/mlNGF (gray), left untreated (white), or treated with NGF in thepresence of 200 nM K252a (white, diagonal lines) for 5 days. TuftelinmRNA expression was quantified after 5 days. Results are presentedas mRNA fold increase (mean W S.E.) normalized to b-actin. MP < 0.02compared to control, MMP < 0.01 compared to NGF, by Student’st-test.
JOURNAL OF CELLULAR PHYSIOLOGY
While CoCl2 caused significant induction of the 64 kDa tuftelinprotein in PC12 cells, only moderate induction of the 55 kDaprotein was observed, suggesting different function for thesetwo tuftelin proteins. It is not known yet if these tuftelin proteinbands are encoded by two mRNA isoforms or represent post-translation modifications or partial degradation of the tuftelinprotein. In the mouse brain, 12 h after intra-peritoneal injectionof CoCl2, significant increase in tuftelin mRNA and proteinlevels were detected. While various tuftelin protein bands weredetected in the mouse brain, only the 64 kDa tuftelin band wassignificantly induced during hypoxia.
PC12 cells treated with NGF for 2 days demonstrated 10-fold decrease in HIF1 activity (Charlier et al., 2002). Sinceinduction of tuftelin expression during hypoxia requires theexpression of HIF1a, NGF-treated PC12 cells providedanother experimental model to investigate tuftelin expression,in relation to HIF1a expression levels. Tuftelin mRNAexpression was significantly reduced after 1 day of NGFtreatment. One explanation to NGF-induced tuftelin mRNAdecreased expression may be related to the decrease in HIF1amRNA expression. Alternative explanation might be related tothe well-documented finding related to ‘‘shutting-off’’ theexpression of the majority of proliferation related genes duringthe initial 24 h of NGF treatment of PC12 cells (Greene andAngelastro, 2005). From day 1 and on, NGF shifts thetranscriptome from proliferation-related genes todifferentiation-related genes (Greene and Angelastro, 2005).Therefore, it is tempting to propose that the reduction oftuftelin mRNA expression during the first 24 h is related toeither the reduction of HIF1 activity, or the shift in the globaltranscriptome activity.
We also demonstrated a quantitative temporal correlationbetween the significant and progressive increase, induced byNGF, of neurite outgrowth elongation (morphologicaldifferentiation) from day 1 to 5, and the significant andprogressive increase in tuftelin mRNA expression. Bothprocesses were selectively inhibited by the specific NGF-TrkAantagonist K252a, indicating TrkA receptor involvement inboth processes. Presently, it is unclear whether tuftelin plays a

172 L E I S E R E T A L .
regulatory role in NGF-induced morphological differentiation,or its increased expression is the consequence of theNGF-induced differentiation process.
Tuftelin was shown to be involved in the developmentalprocesses of various tissues including the mouse developingtooth germ and brain, where it is expressed by the neurons(Leiser et al., 2007). Tuftelin mRNA expression in the mousebrain is significantly increased with age from pre-natal to7 months old mice (data not shown). We have previously alsoshown that tuftelin recruits embryonic mesenchymal cells (Shayet al., 2009), and induces embryonic mesenchymal cellproliferation (unpublished). Hence, tuftelin might be involved inproliferation and differentiation processes. NGF, which isessential for neuronal growth, survival, and differentiation, hasalso been implicated in morphogenetic events during toothdevelopment and injury (Amano et al., 1999; Woodnutt et al.,2000; Magloire et al., 2001; Nosrat et al., 2002; Mizuno et al.,2007). It is therefore tempting to propose that tuftelin inductionby NGF in PC12 cells is reminiscent of physiological processessuch as sympathetic innervation of various organs duringdevelopment (Amano et al., 1999; Ieda et al., 2004; Mizuno et al.,2007; Andres et al., 2008).
In PC12, culture tuftelin was induced both by hypoxia (aninsult) and by NGF (a growth factor) suggesting an importantrole for tuftelin in various cellular pathways. At the molecularlevel there is an overlap between the signaling pathways inducedby hypoxia and NGF, including Ca2þ-Calmodulinpathway, PI3K-Akt pathway, MAPK pathway, and HIF pathway(Seta et al., 2002). An important implication of these results isthat most probably tuftelin induction is associated with one ormore of the above molecular pathways. These findings proposePC12 cells as a cellular model to study tuftelin’s role duringanaerobic metabolic processes and during neuronaldifferentiation.
In conclusion, the present study describes the presence andinduction of tuftelin (mRNA and protein) by hypoxia and NGFin the neuronal PC12 cell model. Revealing the physiologicalrole of tuftelin will clarify mechanisms related to the ‘‘hypoxicgenome’’ (Seta et al., 2002), and NGF-induced differentiation inthe nervous system (Vaudry et al., 2002).
Acknowledgments
We would like to thank Ms. Tali Bdolah-Abram from the Facultyof Medicine, the Hebrew University of Jerusalem, for thestatistical analysis, and Ms. Leah Dafni from the Faculty of DentalMedicine, the Hebrew University of Jerusalem, for her technicalassistance. This study was supported by U.S-Israel BinationalScience Foundation (BSF; 2003-239) to D.D.
Literature Cited
Amano O, Bringas P, Takahashi I, Takahashi K, Yamane A, Chai Y, Nuckolls GH, Shum L,Slavkin HC. 1999. Nerve growth factor (NGF) supports tooth morphogenesis in mousefirst branchial arch explants. Dev Dyn 216:299–310.
Andres R, Herraez-Baranda LA, Thompson J, Wyatt S, Davies AM. 2008. Regulation ofsympathetic neuron differentiation by endogenous nerve growth factor and neurotrophin-3. Neurosci Lett 431:241–246.
Bergeron M, Gidday JM, Yu AY, Semenza GL, Ferriero DM, Sharp FR. 2000. Role of hypoxia-inducible factor-1 in hypoxia-induced ischemic tolerance in neonatal rat brain. Ann Neurol48:285–296.
Brahimi-Horn C, Mazure N, Pouyssegur J. 2005. Signalling via the hypoxia-inducible factor-1alpha requires multiple posttranslational modifications. Cell Signal 17:1–9.
Charlier N, Leclere N, Felderhoff U, Heldt J, Kietzmann T, Obladen M, Gross J. 2002.Hypoxia-induced cell death and changes in hypoxia-inducible factor-1 activity in PC12 cellsupon exposure to nerve growth factor. Brain Res Mol Brain Res 104:21–30.
Deutsch D, Palmon A, Fisher LW, Kolodny N, Termine JD, Young MF. 1991. Sequencing ofbovine enamelin (‘‘tuftelin’’) a novel acidic enamel protein. J Biol Chem 266:16021–16028.
JOURNAL OF CELLULAR PHYSIOLOGY
Deutsch D, Palmon A, Dafni L, Catalano-Sherman J, Young MF, Fisher LW. 1995. Theenamelin (tuftelin) gene. Int J Dev Biol 39:135–143.
Deutsch D, Leiser Y, Shay B, Fermon E, Taylor A, Rosenfeld E, Dafni L, Charuvi K, Cohen Y,Haze A, Fuks A, Mao Z. 2002. The human tuftelin gene and the expression of tuftelin inmineralizing and nonmineralizing tissues. Connect Tissue Res 43:425–434.
Ferrara N. 2005. VEGF as a therapeutic target in cancer. Oncology 69:11–16.Fujita K, Lazarovici P, Guroff G. 1989. Regulation of the differentiation of PC12
pheochromocytoma cells. Environ Health Perspect 80:127–142.Greene LA, Tischler AS. 1976. Establishment of a noradrenergic clonal line of rat adrenal
pheochromocytoma cells which respond to nerve growth factor. Proc Natl Acad Sci USA73:2424–2428.
Greene LA, Angelastro JM. 2005. You can’t go home again: Transcriptionally driven alterationof cell signaling by NGF. Neurochem Res 30:1347–1352.
Hirao M, Hashimoto J, Yamasaki N, Ando W, Tsuboi H, Myoui A, Yoshikawa H. 2007. Oxygentension is an important mediator of the transformation of osteoblasts to osteocytes. J BoneMiner Metab 25:266–276.
Huang LE, Bunn HF. 2003. Hypoxia-inducible factor and its biomedical relevance. J Biol Chem278:19575–19578.
Ieda M, Fukuda K, Hisaka Y, Kimura K, Kawaguchi H, Fujita J, Shimoda K, Takeshita E, OkanoH, Kurihara Y, Kurihara H, Ishida J, Fukamizu A, Federoff HJ, Ogawa S. 2004. Endothelin-1regulates cardiac sympathetic innervation in the rodent heart by controlling nerve growthfactor expression. J Clin Invest 113:876–884.
Katzir I, Shani J, Regev K, Shabashov D, Lazarovici P. 2002. A quantitative bioassay for nervegrowth factor, using PC12 clones expressing different levels of trkA receptors. J MolNeurosci 18:251–264.
Ke Q, Costa M. 2006. Hypoxia-inducible factor-1 (HIF-1). Mol Pharmacol 70:1469–1480.Koizumi S, Contreras ML, Matsuda Y, Hama T, Lazarovici P, Guroff G. 1988. K-252a: A
specific inhibitor of the action of nerve growth factor on PC12 cells. J Neurosci 8:715–721.Kotake-Nara E, Saida K. 2007. Characterization of CoCl2-induced reactive oxygen species
(ROS): Inductions of neurite outgrowth and endothelin-2/vasoactive intestinal contractorin PC12 cells by CoCl2 are ROS dependent, but those by MnCl2 are not. Neurosci Lett422:223–227.
Lazarovici P, Marcinkiewicz C, Lelkes PI. 2006. Cross talk between the cardiovascular andnervous systems: Neurotrophic effects of vascular endothelial growth factor (VEGF) andangiogenic effects of nerve growth factor (NGF)-implications in drug development. CurrPharm Des 12:2609–2622.
Leiser Y, Blumenfeld A, Haze A, Dafni L, Taylor AL, Rosenfeld E, Fermon E, Gruenbaum-Cohen Y, Shay B, Deutsch D. 2007. Localization, quantification, and characterization oftuftelin in soft tissues. Anat Rec (Hoboken) 290:449–454.
Levy AP, Levy NS, Goldberg MA. 1996. Post-transcriptional regulation of vascular endothelialgrowth factor by hypoxia. J Biol Chem 271:2746–2753.
MacDougall M, Simmons D, Dodds A, Knight C, Luan X, Zeichner-David M, Zhang C, RyuOH, Qian Q, Simmer JP, Hu CC. 1998. Cloning, characterization, and tissue expressionpattern of mouse tuftelin cDNA. J Dent Res 77:1970–1978.
Magloire H, Romeas A, Melin M, Couble ML, Bleicher F, Farges JC. 2001. Molecular regulationof odontoblast activity under dentin injury. Adv Dent Res 15:46–50.
Mao Z, Shay B, Hekmati M, Fermon E, Taylor A, Dafni L, Heikinheimo K, Lustmann J, FisherLW, Young MF, Deutsch D. 2001. The human tuftelin gene: Cloning and characterization.Gene 279:181–196.
Mizuno N, Shiba H, Xu WP, Inui T, Fujita T, Kajiya M, Takeda K, Hasegawa N, Kawaguchi H,Kurihara H. 2007. Effect of neurotrophins on differentiation, calcification and proliferationin cultures of human pulp cells. Cell Biol Int 31:1462–1469.
Nosrat I, Seiger A, Olson L, Nosrat CA. 2002. Expression patterns of neurotrophic factormRNAs in developing human teeth. Cell Tissue Res 310:177–187.
Saarikoski ST, Rivera SP, Hankinson O. 2002. Mitogen-inducible gene 6 (MIG-6), adipophilinand tuftelin are inducible by hypoxia. FEBS Lett 530:186–190.
Shay B, Gruenbaum-Cohen Y, Tucker AS, Taylor AL, Rosenfeld E, Haze A, Dafni L, Leiser Y,Fermon E, Danieli T, Blumenfeld A, Deutsch D. 2009. High yield expression of biologicallyactive recombinant full-length human tuftelin protein in baculovirus-infected insect cells.Protein Expr Purif 68:90–98.
Semenza GL. 2000. HIF-1: Mediator of physiological and pathophysiological responses tohypoxia. Appl Physiol 88:1474–1480.
Semenza GL. 2009. Regulation of vascularization by hypoxia-inducible factor 1. Ann NY AcadSci 1177:2–8.
Seta KA, Spicer Z, Yuan Y, Lu G, Millhorn DE. 2002. Responding to hypoxia: Lessons from amodel cell line. Sci STKE 146:re 11.
Sofroniew MV, Howe CL, Mobley WC. 2001. Nerve growth factor signaling,neuroprotection, and neural repair. Annu Rev Neurosci 24:1217–1281.
Tabakman R, Lazarovici P, Kohen R. 2002. Neuroprotective effects of carnosine andhomocarnosine on pheochromocytoma PC12 cells exposed to ischemia. J Neurosci Res68:463–469.
Tabakman R, Lecht S, Sephanova S, Arien-Zakay H, Lazarovici P. 2004a. Interactions betweenthe cells of the immune and nervous system: Neurotrophins as neuroprotection mediatorsin CNS injury. Prog Brain Res 146:387–401.
Tabakman R, Jiang H, Schaefer E, Levine RA, Lazarovici P. 2004b. Nerve growth factorpretreatment attenuates oxygen and glucose deprivation-induced c-Jun amino-terminalkinase 1 and stress-activated kinases p38alpha and p38beta activation and confersneuroprotection in the pheochromocytoma PC12 model. J Mol Neurosci 22:237–250.
Tai TC, Wong-Faull DC, Claycomb R, Wong DL. 2009. Hypoxic stress-induced changes inadrenergic function: Role of HIF1 alpha. J Neurochem 109:513–524.
Vaudry D, Stork PJ, Lazarovici P, Eiden LE. 2002. Signaling pathways for PC12 celldifferentiation: Making the right connections. Science 296:1648–1649.
Wang Y, Wan C, Gilbert SR, Clemens TL. 2007. Oxygen sensing and osteogenesis. Ann NYAcad Sci 1117:1–11.
Wiesener MS, Turley H, Allen WE, Willam C, Eckardt K-U, Talks KL, Wood SM, Gatter KC,Harris AL, Pugh CW, Ratcliffe PJ, Maxwell PH. 1998. Induction of endothelial PAS domainprotein-1 by hypoxia: Characterization and comparison with hypoxia-inducible factor-1a.Blood 92:2260–2268.
Woodnutt DA, Wager-Miller J, O’Neill PC, Bothwell M, Byers MR. 2000. Neurotrophinreceptors and nerve growth factor are differentially expressed in adjacent nonneuronalcells of normal and injured tooth pulp. Cell Tissue Res 299:225–236.




















