
Zoosporic plant pathogens produce bacterial autoinducer-2 that affects Vibrio harveyi quorum sensing
10
Zoosporic plant pathogens produce bacterial autoinducer-2 that affects Vibrio harveyi quorum sensing Ping Kong 1 , Bobby W.K. Lee 2 , Zhaohui Sunny Zhou 2 , and Chuanxue Hong 1,* 1 Department of Plant Pathology, Physiology and Weed Science, Virginia Polytechnic Institute and State University, Virginia Beach, VA 23455-3363, USA 2 The Barnett Institute of Chemical and Biological Analysis and Department of Chemistry and Chemical Biology, Northeastern University, Boston, MA 02115, USA Abstract The frequent co-isolation of bacteria with Phytophthora and Pythium species suggests possible interspecies communication. Zoospore free fluids (ZFF) from bacteria-free and nutrient-depleted zoospore suspensions were examined to investigate production of autoinducer-2 (AI-2), a bacterial interspecies signal molecule, by zoosporic oomycetes. ZFF from P. nicotianae, P. sojae and Py. aphanidermatum triggered luminescence of Vibrio harveyi AI-2 reporter, indicating the presence of AI-2 in zoospore extracellular products and the potential of cross-kingdom communication between oomycetes and bacteria. Production of AI-2 by zoospores was confirmed by chemical assays. These results provide new insight into the physiology and ecology of oomycetes. Keywords oomycetes; AI-2; quorum sensing; cross-kingdom communication 1. Introduction Phytophthora and Pythium in Oomycota of Stramenopila are phylogenetically related to marine algae but morphologically resemble fungi. Many species in these two genera are destructive pathogens that attack a broad range of economically important agricultural and ornamental crops as well as forest tree species. They produce asexual sporangia that release flagellate zoospores as their primary dispersal and infection agents (Deacon & Donaldson, 1993, Judelson & Blanco, 2005). Zoospores secrete a host of molecules during the homing process, however, with the exception of Ca 2+ and an adhesive protein involved in aggregation, germination, and plant attachment (Deacon & Donaldson, 1993, Reid, et al., 1995, Robold & Hardham, 2005), little is known of the presence of other products and their relevance to zoospore communication. In contrast, the identification of autoinducers or small hormone-like molecules has provided unparalleled insight into cell-to-cell communication and its role in the physiology, ecology, evolution, and pathogenesis of bacteria and a few fungal species (Winans & Bassler, 2008). The vast majority of molecules, such as acyl-homoserine lactones (AHLs) or oligopeptides from bacteria (Waters & Bassler, 2005), and small primary alcohols from fungi (Hogan, 2006), are species specific and used for intraspecific communication. One signal molecule called autoinducer-2 (AI-2) can be produced by half of the known bacterial population (Sun, * Correspondence author: Chuanxue Hong [email protected]. NIH Public Access Author Manuscript FEMS Microbiol Lett. Author manuscript; available in PMC 2011 February 1. Published in final edited form as: FEMS Microbiol Lett. 2010 February ; 303(1): 55–60. doi:10.1111/j.1574-6968.2009.01861.x. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Zoosporic plant pathogens produce bacterial autoinducer-2 that affects Vibrio harveyi quorum sensing

Zoosporic plant pathogens produce bacterial autoinducer-2 thataffects Vibrio harveyi quorum sensing
Ping Kong1, Bobby W.K. Lee2, Zhaohui Sunny Zhou2, and Chuanxue Hong1,*
1Department of Plant Pathology, Physiology and Weed Science, Virginia Polytechnic Institute andState University, Virginia Beach, VA 23455-3363, USA2The Barnett Institute of Chemical and Biological Analysis and Department of Chemistry andChemical Biology, Northeastern University, Boston, MA 02115, USA
AbstractThe frequent co-isolation of bacteria with Phytophthora and Pythium species suggests possibleinterspecies communication. Zoospore free fluids (ZFF) from bacteria-free and nutrient-depletedzoospore suspensions were examined to investigate production of autoinducer-2 (AI-2), a bacterialinterspecies signal molecule, by zoosporic oomycetes. ZFF from P. nicotianae, P. sojae and Py.aphanidermatum triggered luminescence of Vibrio harveyi AI-2 reporter, indicating the presenceof AI-2 in zoospore extracellular products and the potential of cross-kingdom communicationbetween oomycetes and bacteria. Production of AI-2 by zoospores was confirmed by chemicalassays. These results provide new insight into the physiology and ecology of oomycetes.
Keywordsoomycetes; AI-2; quorum sensing; cross-kingdom communication
1. IntroductionPhytophthora and Pythium in Oomycota of Stramenopila are phylogenetically related tomarine algae but morphologically resemble fungi. Many species in these two genera aredestructive pathogens that attack a broad range of economically important agricultural andornamental crops as well as forest tree species. They produce asexual sporangia that releaseflagellate zoospores as their primary dispersal and infection agents (Deacon & Donaldson,1993, Judelson & Blanco, 2005). Zoospores secrete a host of molecules during the homingprocess, however, with the exception of Ca2+ and an adhesive protein involved inaggregation, germination, and plant attachment (Deacon & Donaldson, 1993, Reid, et al.,1995, Robold & Hardham, 2005), little is known of the presence of other products and theirrelevance to zoospore communication.
In contrast, the identification of autoinducers or small hormone-like molecules has providedunparalleled insight into cell-to-cell communication and its role in the physiology, ecology,evolution, and pathogenesis of bacteria and a few fungal species (Winans & Bassler, 2008).The vast majority of molecules, such as acyl-homoserine lactones (AHLs) or oligopeptidesfrom bacteria (Waters & Bassler, 2005), and small primary alcohols from fungi (Hogan,2006), are species specific and used for intraspecific communication. One signal moleculecalled autoinducer-2 (AI-2) can be produced by half of the known bacterial population (Sun,
*Correspondence author: Chuanxue Hong [email protected].
NIH Public AccessAuthor ManuscriptFEMS Microbiol Lett. Author manuscript; available in PMC 2011 February 1.
Published in final edited form as:FEMS Microbiol Lett. 2010 February ; 303(1): 55–60. doi:10.1111/j.1574-6968.2009.01861.x.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

et al., 2004) and by some eukaryotic plants (Gao, et al., 2003, Hauck, et al., 2003) althoughits production has not been reported in Fungi and Stramenopila. This molecule facilitatesinterspecific communication among bacteria (Xavier & Bassler, 2005). AI-2 is a collectiveterm for a group of signal molecules derived from 4,5-dihydroxy-2,3-pentanedione (DPD)and is used interchangeably with DPD since conversion of DPD to various forms of AI-2 isa spontaneous ring closure process (Miller, et al., 2004).
The notorious presence of bacteria in Phytophthora and Pythium cultures and stimulation ofPhytophthora zoospore and oospore production by bacterial metabolites (Zentmyer, 1965,Malajczuk, 1983) led us to hypothesize that zoosporic pathogens may produce AI-2 tocommunicate with bacteria. To test this, we analyzed zoospore free fluid (ZFF) frombacterium-free and nutrient-depleted zoospore suspensions for AI-2 activity using a AI-2bacterial reporter strain (Bassler, et al., 1997) and chemical methods (Hauck, et al., 2003,Zhao, et al., 2003). We then discuss AI-2 production pathways and implications of AI-2production in oomycte cross-kingdom communication.
2. Materials and methods2.1. Phytophthora species and culture conditions
Two morphological and phylogenetically distinct Phytophthora species, and a species fromthe closely-related genus Pythium, were used in this study. Phytophthora nicotianae (Syn. P.parasitica) isolate 1B11, P. sojae (genotype I) isolate 23G8, and Pythium aphanidermatumisolate 18H1 were maintained in clarified 20 % vegetable juice medium supplemented with1.5 % agar (CV8A) at 23 °C.
2.2. Preparation of zoospore-free fluid (ZFF)ZFF was prepared from nutrient-depleted zoospore suspensions at high densities. A 5-mm2
CV8A mycelial plug was seeded in 10% CV8 in 90-mm Petri dishes. The dishes wereincubated at 23°C in the dark for 3 days for P. sojae, 4 days for Py. aphanidermatum and 1–2 wks for P. nicotianae to induce sporangia. After the seed plugs and medium wereremoved, the mycelial mats were rinsed five times with sterile distilled water (SDW) toeliminate nutrients from the remaining medium. The drained mycelial mats were incubatedfor 16–18 h for P. sojae and Py. aphanidermatum, and one week for P. nicotianae underfluorescent light at 23°C. When numerous sporangia formed, the mats were rinsed anadditional 5 times with SDW to remove residues from the medium. The dilution factor forthe 10 % CV8 was then 1.08 × 109 as measured experimentally. To induce zoospore release,the mats were flooded with 8 ml of chilled SDW and kept under light until the desiredzoospore density was reached. The density for 1B11 was up to 106 zoospores ml−1 in 1 h;for 23G8 and 18H1 was up to 5×104 and 3×104 zoospores ml−1 in 3 h, respectively. Allprocedures were performed under sterile conditions to prevent bacterial contamination.
To obtain ZFF, zoospore suspensions were filtered through sterile miracloth to removemycelia, sporangia and other structures, then vortexed briefly to facilitate chemical release.The suspensions were then filtered through a 0.2 µm syringe filter to remove the cysts. ZFFwas used fresh or stored at −20°C.
2.3. Detection of AI-2 in ZFF using bioluminescence assayThe bacterial AI-2 reporter Vibrio harveyi BB170 [luxN::TnS] (ATCC BAA-1117) wasused to test the activity of ZFF and detect the presence of AI-2. The assay was conductedusing a combined protocol based on procedures described previously (Bassler, et al., 1997,DeKeersmaecker & Vanderleyden, 2003). Briefly, BB170 was cultured overnight in MBmedium then diluted 10,000× into AB medium. Aliquots of 90 µl from the resulting
Kong et al. Page 2
FEMS Microbiol Lett. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

overnight culture were dispensed into each well of a 96-well plate, followed by the addition(10 µl per well) of test solutions. The plate was then incubated at 30°C with aeration. Lightproduction was monitored using a CCD camera after 3 h incubation for a period of 8 h, andIntegrated Optical Density (IOD) was measured using LabWorks Image Acquisition andAnalysis Software (UVP, California, USA). Each assay included triplicate wells of ZFFsamples with negative and positive controls. The negative controls included SDW and 10,000 × diluted CV8. The positive controls consisted of a 10-fold dilution series from a 550µM stock solution of enzymatically synthesized 4,5-dihydroxy-2,3-pentanedione (DPD),produced and quantified as described previously [17]. The experiment was repeated twice.For quantification, a standard curve was generated based on IOD measured at 6 h ofincubation with the DPD dilution series. The standard curve was then used to plot the IODfrom treatments to obtain AI-2 concentrations.
2.4. Chemical analysis of presence of AI-2 in ZFFTo confirm the presence of AI-2 (DPD) in ZFF and rule out false positives from thebioassay (DeKeersmaecker & Vanderleyden, 2003), ZFF samples were tested for DPD-derived quinoxaline generated via the chemical reaction with 1,2-diaminobenzene (Hauck,et al., 2003, Zhao, et al., 2003). Test solutions were mixed with 10 mM 1,2-diaminobenzeneindividually. After incubation overnight at 37 °C at pH 4.5, the resulting solution wasextracted three times with an equal volume of ethyl ether. The organics were concentratedby rotary evaporation and then dissolved in methanol (500 µl).
The extracts were analyzed using liquid chromatography-mass spectrometry (LC-MS) forDPD-derived quinoxaline on a Surveyor HPLC system coupled to a Finnagan LCQ Deca XPmass spectrometer (Thermo Fisher Scientific, San Jose, CA). Samples were loaded on a self-packed reversed-phase column (75 µm i.d. × 15 cm, Magic C18 resin, 3 µm particle size,200 Å pore size, Michrom Bioresources, Auburn, CA). The column was equilibrated with1% acetonitrile (solvent A) and 0.1% formic acid in water (solvent B) and eluted with thefollowing solvent gradient starting from 1% solvent A for 10 min and increasing to 25%solvent A over 25 min, then to 50% solvent A over 5 min, and finally a constant 50%solvent A for 5 min. The flow rate was kept at a constant 160 µl min−1.
Data from LC-MS were processed using Xcalibar Data System 2.0 (Thermo FisherScientific, San Jose, CA). Quinoxaline was identified by extracted-ion chromatogram andfragmentation pattern analyses (Hauck, et al., 2003). Additional confirmation was made byco-elution with a DPD-derived quinoxaline standard prepared from the synthesized DPD. Toquantify DPD-derived quinoxaline, the peak density at m/z 205 was plotted using acalibration curve generated from the synthetic DPD samples of known concentrations.
3. Results3.1. Zoospore extracellular products affect bacterial quorum sensing of AI-2 reporter
ZFF triggered luminescence production of Vibrio harveyi AI-2 reporter strain BB170.Intensive light production was observed in ZFF treated wells but not in control wellscontaining SDW and 104 × diluted CV8 at 6 h (Fig. 1A). Based on the light intensityinduced by synthetic AI-2 (Fig. 1B), the concentration of AI-2 in ZFF samples wasestimated between 1.1- 5.5 µM. Within ZFF treatments, ZFFaph displayed the highest lightintensity, followed by ZFFsoj and ZFFnic. Stimulation of light production of V. harveyiindicates zoospore extracellular products regulate quorum sensing of the bacteria andcontain AI-2.
Quantification of AI-2 in ZFF using the DPD standard curve (AI-2=0.376*IOD-28.93)indicated that the amount of AI-2 in ZFF varied with species. ZFFaph contained the highest
Kong et al. Page 3
FEMS Microbiol Lett. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

concentration of AI-2 (1.66 ±0.25 µM) among the three species tested followed by ZFFsoj(1.40 ±0.18 µM), both from zoospores at 104 ml−1 levels, while ZFFnic from zoospores at5×105 ml−1 contained the least AI-2 (0.66 ±0.13 µM). Negative values were obtained for theSDW and CV8 controls, indicating that zoospores produced AI-2, and the AI-2 activity wasnot from residual CV8 broth.
3.2. Confirmation of AI-2 production by chemical methodTo confirm the results from the luminescence assay, ZFFnic and ZFFsoj were analyzedusing chemical methods. The formation of quinoxaline derived from DPD, specifically 2-(1,2-dihydroxyethyl)-3-methylquinoxaline, was detected by liquid chromatography-massspectrometry (LC-MS). First, a peak identical to quinoxaline from synthetic DPD (Fig. 2D)was identified in the extracted ion chromatograms (EIC) at m/z 205 from ZFFsoj (Fig. 2A)and ZFFnic (Fig. 4A middle). Second, quinoxaline products from ZFF coeluted with thequinoxaline standard (Fig. 2C, Fig. 4 top). Lastly, the fragmentation patterns of thequinoxaline derived from ZFF samples were identical to those from synthetic DPD (Fig. 3A,B; Fig. 4. B, C). Furthermore, the quinoxaline peak was not detected in the negative control,a 103 × dilution of the CV8 broth equivalent to 106 times what was left in ZFFs (Fig. 2BFig. 4A bottom). These cumulative results indicate AI-2 originated from zoospores.
Quantification of DPD-derived quinoxaline in ZFF samples also showed variation in AI-2concentrations with species. P. sojae produced a higher amount of AI-2 than P. nicotianae.Based on the standard curve generated from synthetic DPD, AI-2 concentration in ZFFnicfrom a suspension of 106 zoospores ml−1 was 1.1±0.1 µM while in ZFFsoj from asuspension of 5 × 104 zoospores ml−1 was 10.1±2.0 µM (Fig. 4), similar to what wasobserved in the bio-luminescence assay.
DiscussionAI-2 represents an interspecies signaling molecule and is involved in regulation ofluminescence, virulence factor secretion and biofilm formation in bacteria (Vendeville, etal., 2005, Xavier & Bassler, 2005). Here, we demonstrate for the first time the production ofAI-2 by P. nicotianae, P. sojae and Py. aphanidermatum, members of Pythiaceae in theeukaryotic Stramenopila kingdom, which will provide insight into the physiology andecology of zoosporic pathogens.
Detection of AI-2 in zoospore free fluid (Fig. 1–4) raises a question regarding the AI-2production pathway in oomycete species. Currently, there are three known pathways forAI-2 (DPD) production (Winzer, et al., 2002,Hauck, et al., 2003,Nichols, et al., 2009).Within these pathways, the pentose-phosphate pathway used by some plant and bacterialspecies (Hauck, et al., 2003,Tavender, et al., 2008) is most likely to be adopted byoomycetes. The reasons are three fold. First, the LuxS-dependent pathway is only availablein bacterial species (Sun, et al., 2004). Oomycetes, like other eukaryotes, lack LuxShomology in their genome (genome.jgi-psf.org, broad.mit.edu, vmd.vbi.vt.edu). Second, thenoncanonical abiotic/biotic reaction pathway reported in thermal tolerant bacteria requireshydrothermal environments to form DPD (Nichols, et al., 2009). Such conditions are notencountered by these oomycete “water molds”. Lastly, in the pentose-phosphate pathway,DPD is formed spontaneously by converting pentose-phosphates to D-ribulose-5-phosphateusing isomerases (RPI). In searching oomycete genome databases we found that pentosephosphates are common metabolic products, and all four published genome sequences ofPhytophthora species contain conserved sequences for RPI, suggesting zoosporic oomycetesmay form DPD through the central intermediate ribose-5-phosphate. Silencing the RPI geneand testing mutant AI-2 production may provide direct evidence to test this presumption.
Kong et al. Page 4
FEMS Microbiol Lett. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

However, it is possible that other unknown pathways are responsible for the production ofAI-2.
Although it is not clear whether oomycetes use AI-2 to encode information forcommunication within the population to coordinate behaviors such as aggregation and plantinfection, AI-2 production by Pythiaceae species raises the possibility that zoosporicpathogens may use AI-2 as a common signal to communicate with bacteria. Communicationwith bacteria may be beneficial to these pathogens as shown by their ability to survive insoil with a wide range of bacteria and their tolerance to frequent culture contamination bybacteria. It will be interesting to know if this cross-kingdom relationship is bridged by AI-2.In fact, triggering luminescence of Vibrio harveyi by ZFF (Fig. 1) has verified thatoomycetes can communicate with bacteria and affect their quorum sensing through thismolecule. This process may give oomycetes an advantage in fitness and possibly virulence.Bacteria and bacterial metabolites have been shown to stimulate Phytophthora reproduction(Zentmyer, 1965) and contribute to Phytophthora colonization on plants (Yang, et al.,2001).
AcknowledgmentsWe gratefully acknowledge supplies of isolates of Phytophthora and Pythium from Drs. Brett Tyler, MichaelBenson and Gary Moorman; expression strains for AI-2 production from Drs. Kenneth Cornell, Michael Riscoe,Mark Hilgers and Martha Ludwig. We thank Dr. Brett Tyler for assistance with oomycete bioinformatics; andPatricia Richardson for reading this manuscript. This work is supported in part by grants to C. Hong from USDA-CSREES (2005-51101-02337) and to Z. S. Zhou from NIAID/NIH (1R01AI058146). This is publication number939 from the Barnett Institute.
ReferencesBassler BL, Greenberg EP, Stevens AM. Cross-species induction of luminescence in the quorum-
sensing bacterium Vibrio harveyi. J. of Bacteriol. 1997; 179:4043–4045. [PubMed: 9190823]Deacon JW, Donaldson SP. Molecular recognition in the homing responses of zoosporic fungi, with
special reference to Pythium and Phytophthora. Mycol. Res. 1993; 97:1153–1171.DeKeersmaecker SCJ, Vanderleyden J. Constraints on detection of autoinducer-2 (Al-2) signalling
molecules using Vibrio harveyi as a reporter. Microbiology-Sgm. 2003; 149:1953–1956.Gao M, Teplitski M, Robinson JB, Bauer WD. Production of substances by Medicago truncatula that
affect bacterial quorum sensing. Molecular Plant-Microbe Interactions. 2003; 16:827–834.[PubMed: 12971606]
Hauck T, Hubner Y, Bruhlmann F, Schwab W. Alternative pathway for the formation of 4,5-dihydroxy-2,3-pentanedione, the proposed precursor of 4-hydroxy-5-methyl-3(2H)-furanone as wellas autoinducer-2, and its detection as natural constituent of tomato fruit. Biochimica Et BiophysicaActa-General Subjects. 2003; 1623:109–119.
Hogan DA. Talking to themselves: autoregulation and quorum sensing in Fungi. Eukaryotic Cell.2006; 5:613–619. [PubMed: 16607008]
Judelson HS, Blanco FA. The spores of Phytophthora: Weapons of the plant destroyer. NatureReviews Microbiology. 2005; 3:47–58.
Malajczuk, N. Microbial antagonism to Phytophthora. In: Erwin, DC.; Bartnicki-Garcia, S.; Tsao, PH.,editors. Phytophthora: Its Biology, Taxonomy, Ecology, and Pathology. St. Paul, Minnesota: APSPress; 1983. p. 197-218.
Miller ST, Xavier KB, Campagna SR, Taga ME, Semmelhack MF, Bassler BL, Hughson FM.Salmonella typhimurium recognizes a chemically distinct form of the bacterial quorumsensingsignal Al-2. Molecular Cell. 2004; 15:677–687. [PubMed: 15350213]
Nichols JD, Johnson MR, Chou C-J, Kelly RM. Temperature, not LuxS, mediates AI-2 formation inhydrothermal habitats. Fems Microbiology Ecology. 2009; 68:173–181. [PubMed: 19309315]
Reid B, Morris BM, Gow NAR. Calcium-dependent, genus-specific, autoaggregation of zoospores ofphytopathogenic fungi. Exp. Mycol. 1995; 19:202–213.
Kong et al. Page 5
FEMS Microbiol Lett. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Robold AV, Hardham AR. During attachment Phytophthora spores secrete proteins containingthrombospondin type 1 repeats. Current Genetics. 2005; 47:307–315. [PubMed: 15815927]
Sun JB, Daniel R, Wagner-Dobler I, Zeng AP. Is autoinducer-2 a universal signal for interspeciescommunication: a comparative genomic and phylogenetic analysis of the synthesis and signaltransduction pathways? BMC Evol. Biol. 2004; 4
Tavender T, Halliday N, Hardie K, Winzer K. LuxS-independent formation of AI-2 from ribulose-5-phosphate. BMC Microbiology. 2008; 8:98. [PubMed: 18564424]
Vendeville A, Winzer K, Heurlier K, Tang CM, Hardie KR. Making 'sense' of metabolism:Autoinducer-2, LuxS and pathogenic bacteria. Nat. Rev. Microbiol. 2005; 3:383–396. [PubMed:15864263]
Waters CM, Bassler BL. Quorum sensing: cell-to-cell communication in bacteria. Annu. Rev. Cell.Dev. Biol. 2005; 21:319–346. [PubMed: 16212498]
Winans, SC.; Bassler, BL. Chemical Communication Among Bacteria. Washington, DC: ASM Press;2008.
Winzer K, Hardie KR, Burgess N, et al. LuxS: its role in central metabolism and the in vitro synthesisof 4-hydroxy-5-methyl-3(2H)-furanone. Microbiology-Sgm. 2002; 148:909–922.
Xavier KB, Bassler BL. Interference with Al-2-mediated bacterial cell-cell communication. Nature.2005; 437:750–753. [PubMed: 16193054]
Yang CH, Crowley DE, Menge JA. 16S rDNA fingerprinting of rhizosphere bacterial communitiesassociated with healthy and Phytophthora infected avocado roots. Fems Microbiology Ecology.2001; 35:129–136. [PubMed: 11295451]
Zentmyer GA. Bacterial stimulation of sporangium production in Phytophthora cinnamomi. Science.1965; 150:1178–1179. [PubMed: 17742598]
Zhao G, Wan W, Mansouri S, Alfaro JF, Bassler BL, Cornell KA, Zhou ZHS. Chemical synthesis ofS-ribosyl-L-homocysteine and activity assay as a LuxS substrate. Bioorganic & MedicinalChemistry Letters. 2003; 13:3897–3900. [PubMed: 14592470]
Kong et al. Page 6
FEMS Microbiol Lett. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. Induced luminescence production of AI-2 reporter strain Vibrio harveyi BB170Light production was photographed using a CCD camera at 6 h at 30°C after addition of (A)ZFFnic from a zoospore suspension of Phytophthora nicotianae at 5×105 ml−1, ZFFsoj froma suspension of P. sojae at 104 ml−1, ZFFaph from a suspension of Py. aphanidermatum at3×104 ml−1, SDW and 104 × diluted CV8 controls; (B) a dilution series of synthetic AI-2.Each assay contained 3 wells and was repeated twice.
Kong et al. Page 7
FEMS Microbiol Lett. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
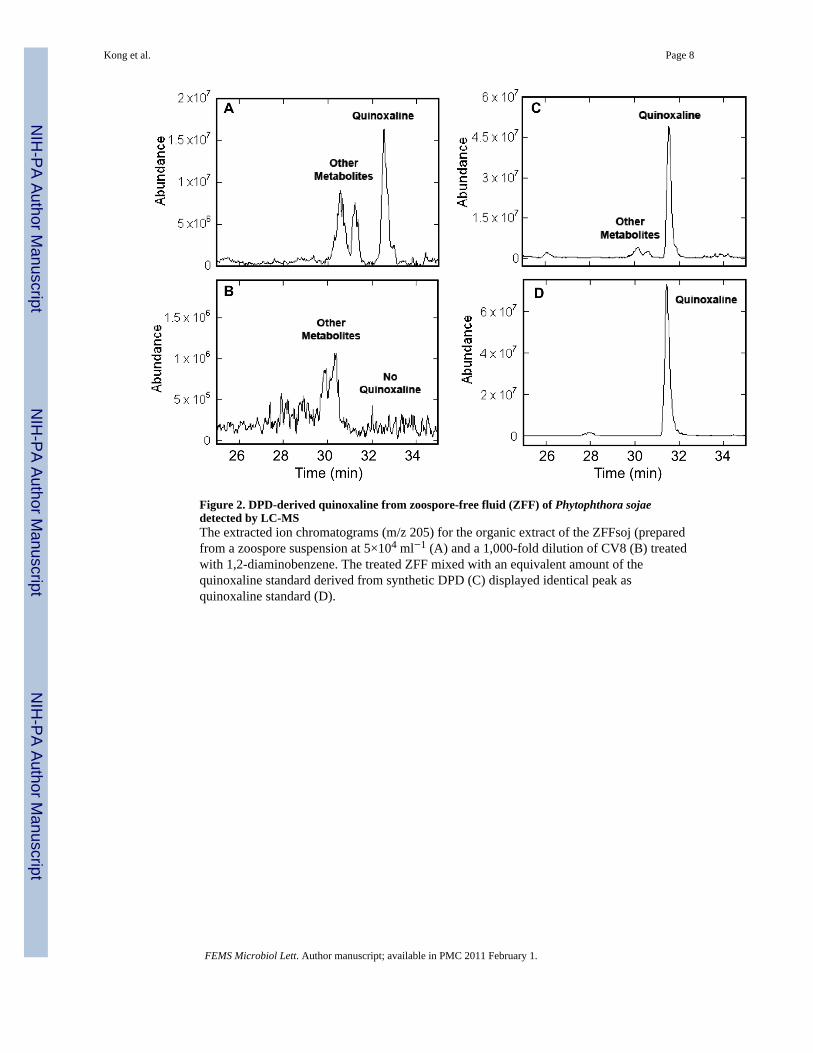
Figure 2. DPD-derived quinoxaline from zoospore-free fluid (ZFF) of Phytophthora sojaedetected by LC-MSThe extracted ion chromatograms (m/z 205) for the organic extract of the ZFFsoj (preparedfrom a zoospore suspension at 5×104 ml−1 (A) and a 1,000-fold dilution of CV8 (B) treatedwith 1,2-diaminobenzene. The treated ZFF mixed with an equivalent amount of thequinoxaline standard derived from synthetic DPD (C) displayed identical peak asquinoxaline standard (D).
Kong et al. Page 8
FEMS Microbiol Lett. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Detection of AI-2 in zoospore-free fluid (ZFF) of Phytophthora sojae using electrosprayionisation-mass spectrometry (ESI-MS)Mass spectra of the DPD-derived quinoxaline in the extract of synthetic DPD (A), ZFFsojfrom a zoospore suspension at 5×104 ml−1 (B), and a 1,000 × dilution of CV8 (C). DPD-derived quinoxaline was detected in the extract of ZFFsoj, but not in that of CV8 negativecontrol, as indicated by presence of the characteristic parent ion of m/z 205 and fragmentions at m/z 187 and 145 as those in authentic DPD samples above the noise level between 30to 35 min (the quinoxaline standard eluted from the column at ~32 min).
Kong et al. Page 9
FEMS Microbiol Lett. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 4. Detection of AI-2 in zoospore-free fluid (ZFF) of Phytophthora nicotianae using thechemical method(A). The extracted ion chromatograms (m/z 205) for the organic extract of the ZFFnic froma zoospore suspension at 106 ml−1 treated with 1,2-diaminobenzene (middle trace), the samesample mixed with an equivalent amount of the quinoxaline standard (top trace), and theorganic extract of a 1,000 × dilution of CV8 treated with 1,2-diaminobenzene (bottomtrace). Mass spectra of the DPD-derived quinoxaline in the extract of ZFFnic from azoospore suspension at 106 ml−1 presenting the characteristic parent ion of m/z 205 andfragment ions at m/z 187 and 145 (C) to those in authentic DPD samples (B).
Kong et al. Page 10
FEMS Microbiol Lett. Author manuscript; available in PMC 2011 February 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript




















