
Genomic overview of the phytopathogen Pectobacterium wasabiae strain RNS 08.42.1A suggests...
12
Genomic overview of the phytopathogen Pectobacterium wasabiae strain RNS 08.42.1A suggests horizontal acquisition of quorum-sensing genes Slimane Khayi • Yannick Raoul des Essarts • Ange ´lique Que ˆtu-Laurent • Mohieddine Moumni • Vale ´rie He ´lias • Denis Faure Received: 4 July 2014 / Accepted: 18 September 2014 Ó Springer International Publishing Switzerland 2014 Abstract The blackleg and soft-rot diseases caused by pectinolytic enterobacteria such as Pectobacterium and Dickeya are major causes of losses affecting potato crop in the field and upon storage. In this work, we report the isolation, characterization and genome analysis of the Pectobacterium wasabiae (formely identified as Pecto- bacterium carotovorum subsp. carotovorum) strain RNS 08.42.1A, that has been isolated from a Solanum tuberosum host plant in France. Comparative genomics with 3 other P. wasabiae strains isolated from potato plants in different areas in North America and Europe, highlighted both a strong similarity at the whole genome level (ANI [ 99 %) and a conserved synteny of the virulence genes. In addi- tion, our analyses evidenced a robust separation between these four P. wasabiae strains and the type strain P. wasabiae CFBP 3304 T , isolated from horseradish in Japan. In P. wasabiae RNS 08.42.1A, the expI and expR nucleo- tidic sequences are more related to those of some Pecto- bacterium atrosepticum and P. carotovorum strains (90 % of identity) than to those of the other potato P. wasabiae strains (70 to 74 % of identity). This could suggest a recruitment of these genes in the P. wasabiae strain RNS 08.42.1A by an horizontal transfer between pathogens infecting the same potato host plant. Keywords P. wasabiae Soft-rot Blackleg Quorum- sensing T3SS Virulence Introduction The pectolytic members of the Enterobacteriaceae genera Pectobacterium and Dickeya are causative agents of dam- aging diseases that affect a wide range of plants and crops, including potato (Solanum tuberosum) (Gardan et al. 2003; Samson et al. 2005). On this plant species, they can induce soft rot on tubers, blackleg, and aerial stem rot symptoms that can be associated to wilting and yellowing of the leaves on potato crops (Pe ´rombelon 2002; Czajkowski et al. 2011). Virulence cycle of the soft rot bacteria Pectobacterium or Dickeya alternates between an asymptomatic latent phase and a virulence phase. The virulence phase is characterized by the synthesis and secretion of plant cell wall-degrading enzymes (PCWDEs) such as pectinases, pectate-lyases, cellulases or proteases which provoke the disorganization and maceration of the plant cell wall (Barras et al. 1994). In Pectobacterium species, the pro- duction of PCWDEs is controlled by quorum sensing and Electronic supplementary material The online version of this article (doi:10.1007/s10709-014-9793-2) contains supplementary material, which is available to authorized users. S. Khayi Y. Raoul des Essarts D. Faure (&) Institut des Sciences du Ve ´ge ´tal, UPR2355, Saclay Plant Sciences, Centre National de la Recherche Scientifique, 1, Avenue de la Terrasse, 91 198 Gif-sur-Yvette, France e-mail: [email protected] S. Khayi M. Moumni De ´partement de Biologie, Faculte ´ des Sciences, Universite ´ Moulay Ismaı ¨l, Meknes, Morocco Y. Raoul des Essarts A. Que ˆtu-Laurent V. He ´lias (&) Fe ´de ´ration Nationale des Producteurs de Plants de Pomme de Terre-Recherche De ´veloppement Promotion du Plant de Pomme de Terre (FN3PT-RD3PT), 75008 Paris, France e-mail: [email protected] A. Que ˆtu-Laurent V. He ´lias UMR 1349 IGEPP INRA - Agrocampus Ouest Rennes, 35653 LeRheu, France 123 Genetica DOI 10.1007/s10709-014-9793-2
Transcript of Genomic overview of the phytopathogen Pectobacterium wasabiae strain RNS 08.42.1A suggests...

Genomic overview of the phytopathogen Pectobacteriumwasabiae strain RNS 08.42.1A suggests horizontal acquisitionof quorum-sensing genes
Slimane Khayi • Yannick Raoul des Essarts •
Angelique Quetu-Laurent • Mohieddine Moumni •
Valerie Helias • Denis Faure
Received: 4 July 2014 / Accepted: 18 September 2014
� Springer International Publishing Switzerland 2014
Abstract The blackleg and soft-rot diseases caused by
pectinolytic enterobacteria such as Pectobacterium and
Dickeya are major causes of losses affecting potato crop in
the field and upon storage. In this work, we report the
isolation, characterization and genome analysis of the
Pectobacterium wasabiae (formely identified as Pecto-
bacterium carotovorum subsp. carotovorum) strain RNS
08.42.1A, that has been isolated from a Solanum tuberosum
host plant in France. Comparative genomics with 3 other P.
wasabiae strains isolated from potato plants in different
areas in North America and Europe, highlighted both a
strong similarity at the whole genome level (ANI [ 99 %)
and a conserved synteny of the virulence genes. In addi-
tion, our analyses evidenced a robust separation between
these four P. wasabiae strains and the type strain P.
wasabiae CFBP 3304T, isolated from horseradish in Japan.
In P. wasabiae RNS 08.42.1A, the expI and expR nucleo-
tidic sequences are more related to those of some Pecto-
bacterium atrosepticum and P. carotovorum strains (90 %
of identity) than to those of the other potato P. wasabiae
strains (70 to 74 % of identity). This could suggest a
recruitment of these genes in the P. wasabiae strain RNS
08.42.1A by an horizontal transfer between pathogens
infecting the same potato host plant.
Keywords P. wasabiae � Soft-rot � Blackleg � Quorum-
sensing � T3SS � Virulence
Introduction
The pectolytic members of the Enterobacteriaceae genera
Pectobacterium and Dickeya are causative agents of dam-
aging diseases that affect a wide range of plants and crops,
including potato (Solanum tuberosum) (Gardan et al. 2003;
Samson et al. 2005). On this plant species, they can induce
soft rot on tubers, blackleg, and aerial stem rot symptoms
that can be associated to wilting and yellowing of the
leaves on potato crops (Perombelon 2002; Czajkowski
et al. 2011).
Virulence cycle of the soft rot bacteria Pectobacterium
or Dickeya alternates between an asymptomatic latent
phase and a virulence phase. The virulence phase is
characterized by the synthesis and secretion of plant cell
wall-degrading enzymes (PCWDEs) such as pectinases,
pectate-lyases, cellulases or proteases which provoke the
disorganization and maceration of the plant cell wall
(Barras et al. 1994). In Pectobacterium species, the pro-
duction of PCWDEs is controlled by quorum sensing and
Electronic supplementary material The online version of thisarticle (doi:10.1007/s10709-014-9793-2) contains supplementarymaterial, which is available to authorized users.
S. Khayi � Y. Raoul des Essarts � D. Faure (&)
Institut des Sciences du Vegetal, UPR2355, Saclay Plant
Sciences, Centre National de la Recherche Scientifique, 1,
Avenue de la Terrasse, 91 198 Gif-sur-Yvette, France
e-mail: [email protected]
S. Khayi � M. Moumni
Departement de Biologie, Faculte des Sciences, Universite
Moulay Ismaıl, Meknes, Morocco
Y. Raoul des Essarts � A. Quetu-Laurent � V. Helias (&)
Federation Nationale des Producteurs de Plants de Pomme de
Terre-Recherche Developpement Promotion du Plant de Pomme
de Terre (FN3PT-RD3PT), 75008 Paris, France
e-mail: [email protected]
A. Quetu-Laurent � V. Helias
UMR 1349 IGEPP INRA - Agrocampus Ouest Rennes,
35653 LeRheu, France
123
Genetica
DOI 10.1007/s10709-014-9793-2

signal molecules of the N-acylhomoserine lactone
(NAHL) class (Smadja et al. 2004; Liu et al. 2008; Crepin
et al. 2012a, b, c). Quorum sensing relies upon the pro-
duction and perception of these signal molecules, the
concentration of which mimics the cell-density, to coor-
dinate gene expression (Fuqua et al. 1994; Whitehead
et al. 2002). In Dickeya, a cell–cell signal is also involved
in the control of virulence factors, but its structure
remains unknown (Nasser et al. 2013). Many studies
demonstrated the involvement of numerous virulence
determinants in soft rot bacteria, such as the cell mem-
brane structure (surface lipopolysaccharides and LPS),
flagella and pilus and secretion systems T1SS,T2SS, T4SS
and T6SS (Toth et al. 2003; Bell et al. 2004; Corbett
et al. 2005; Kim et al. 2009).
Among Pectobacterium, Pectobacterium wasabiae
(Gardan et al. 2003) initially isolated from Japanese
horseradish (wasabi) (Goto and Matsumoto 1987) was
more recently identified on potato in USA, Canada, New
Zealand, Iran, South Africa, Zimbabwe, Finland, Poland
(Ma et al. 2007; Pitman et al. 2010; Baghaee-Ravari et al.
2011; De Boer et al. 2012; Ngadze et al. 2012; Nabhan
et al. 2012; Nykyri et al. 2012; Moleleki et al. 2013; Wa-
leron et al. 2013).
Results of characterization studies conducted on Pec-
tobacterium spp. strains originating from various countries
collected up to 40 years evidenced that P. wasabiae was
not new on potato, and is also present in Peru, Ireland,
Scotland, Ireland, The Netherlands and Germany (De Boer
et al. 2012; Nabhan et al. 2012; Nykyri et al. 2012; Wa-
leron et al. 2013).
Few genomes of P. wasabiae are available in the Gene
Bank database: two of them, P. wasabiae WPP163 and P.
wasabiae SCC3193 are completed in a gapless format,
while those of the others P. wasabiae CFIA1002 (42
contigs) and the type strain P. wasabiae CFBP 3304T (73
contigs) are presented under a draft genome format. The P.
wasabiae strains WPP163, CFIA1002 and SCC3193 were
respectively isolated from potato plants in USA, Canada,
and Finland, respectively (Kim et al. 2009; Koskinen et al.
2012; Nykyri et al. 2012; Yuan et al. 2014).
In this study, we report the isolation, characterization
and genome sequence of the P. wasabiae strain RNS
08.42.1A that was recovered from soft rotted potato stem
in France. Comparative genomics that involved P.
wasabiae isolates from North America, Europe, and
Japan, highlighted the existence of distinctive traits
between strains isolated from potato host and the type
strain isolated from Japanese horseradish. Particularly, the
P. wasabiae strain RNS 08.42.1A exhibited an original
quorum-sensing system that appeared to be possibly
acquired by horizontal transfer from another Pectobacte-
rium species.
Materials and methods
Isolation of the bacterial strain and culture conditions
Pectobacterium wasabiae RNS 08.42.1A was recovered in
2008 from a potato plant (CV Bintje) expressing a backleg
symptom in a glasshouse experiment conducted on Rhi-
zoctonia solani at the INRA Institute of Le Rheu.
Diseased tissues were collected from the plant and
rapidly transferred and incubated into 10 mL of phosphate
buffer (Na2HPO4, 12H2O 0.27 %, NaH2PO4, 2H2O
0.04 %) for 2 h at room temperature under a gentle shaking
to allow bacteria to be released from the tissues. Fifty lL of
diluted bacterial suspensions were plated onto CVP med-
ium (Helias et al. 2012). Plates were incubated 48 h at
26 �C. Bacterial colonies associated to pit formations that
evidenced their pectolytic capability were selected and
transferred onto NBA (nutrient broth agar) medium to be
characterized. The P. wasabiae strain RNS 08.42.1A was
usually cultured in TY medium (tryptone 5 g/L, yeast
extract 3 g/L and agar 1.5 %) at 25 �C.
DNA extraction
DNA extractions were performed from overnight cultures
using a phenol–chloroform purification method followed
by an ethanol precipitation as described by Wilson (1987).
Verification of the quality and quantity of the DNA was
completed using a NanoDrop (ND 1000) device and aga-
rose gel electrophoresis at 1.0 %.
Molecular characterization and genome sequencing
Before genome sequencing, several PCR primers (Table 1)
were used to verify that the strain RNS 08.42.1A indeed
belonged to the P. wasabiae species. They were the ADE
primers identifying Dickeya (Nassar et al. 1996), Y1–Y2
primers identifying Pectobacterium (Darrasse et al. 1994),
Y45–46 primers for detecting Pectobacterium atrosepti-
cum (Frechon et al. 1998) and three couples of primers that
amplify P. wasabiae specific gene markers (Kim et al.
2012; De Boer et al. 2012) and the recombinase subunit A
(recA) gene (Waleron et al. 2002). The primers Y1–Y2 and
recAF-recAR were developed to detect Erwinia carotovo-
rum and the subspecies wasabiae (or Pectobacterium
carotovorum subsp. wasabiae) respectively. However,
since the revision of the classification of the Erwinia genus
by Gardan et al. (2003), Y1–Y2 primers are specific for P.
atrosepticum and P. carotovorum subsp. carotovorum and
recAF-recAR for P. wasabiae.
For genomic sequencing, a shotgun long distance mate-
pair library with an insert size of 8,000 bp was constructed
using the TruSeq (TM) SBS v3 sequencing kit. Sequencing
Genetica
123

of the library was performed using 2 9 100 bp paired-end
read modules by Eurofins Genomics (France). Assembly of
the sequences was carried out using the software CLC
Genomics Wokbench V5.1 (CLC bio, Aarhus, Denmark).
A trimming step was performed to remove the sequences
reads with low quality (limit 0.05), ambiguous nucleotides
(n B 2), and length less than 50 nucleotides. The finishing
of the assembly was carried out using GapFiller version
1.11 to close the gaps. The remaining gaps were resolved
by the mapping of mate-pairs using as a reference the 8 kbp
from each of the contig ends (read length = 0.9, iden-
tity = 0.95). Then, using homemade scripts and fastqse-
lect.tcl from MIRA3 package, the mapped reads for both
orientations (R1 and R2) were retrieved and de novo
assembled (read length = 0.5, identity = 0.8).
ORFs annotation, phylogenetic tree and genomic
comparison
The average nucleotide identity (ANI) values were calcu-
lated as previously proposed (Goris et al. 2007) using the
ANI calculator from the Kostas lab with default settings
(http://enve-omics.ce.gatech.edu/ani/). The synteny com-
parisons were performed using the MAUVE software
(Darling 2004).
The phylogenetic analysis was conducted using the
MEGA software (V6) (Tamura et al. 2013). A multi-locus
sequence analysis (MLSA) was performed using 4 house-
keeping complete genes; fusA, gyrB, recA, and dnaX, from
the P. wasabiae strains SCC9193 (CP003415), CFPB
3304T (AKVS00000000), WP166 (CP001790), and
CFIA1002 (JENG00000000). The strains P. atrosepticum
CFBP 6276 (ASAB00000000), P. atrosepticum SCRI1043
(BX950851), P. carotovorum subsp carotovorum PC1
(CP001657), P. carotovorum subsp carotovorum WPP14
(ABVY00000000), P. carotovorum subsp brasiliensis
PBR1692 (ABVX00000000) and P. carotovorum subsp
carotovorum PCC21 (CP003776), Dickeya dadantii 898T
(CM001976), and Dickeya solani IPO2222T (CM001859)
were also used. After being concatenated in the same order,
the consensus sequences were aligned with ClustalW, and
then a Neighbour-joining tree was created by Bootstrap
method with 1,000 bootstrap replications.
The draft genome sequences were annotated using the
RAST (Rapid Annotation using Subsystem Technology)
(Aziz et al. 2008). Genomic comparisons were conducted
based on RAST functional comparative tool (Aziz et al.
2008). The presence of the genomic clusters in P. wasabiae
RNS 08.42.1A was searched via a nucleotide BLAST
analysis using the sequences of the draft genome as query
against a database constituted by the specific clusters sep-
arately (e-value threshold = 10-30).
Nucleotide sequence accession number
This whole genome shotgun project has been deposited at
DDBJ/EMBL/GenBank under the accession
JMDL00000000. The version described in this paper is
version JMDL01000000.
Results and discussion
Isolation and characterization of P. wasabiae RNS
08.42.1A
The disease symptoms expressed at an early stage of the
plant, as soon as 4 leaflefts have developed. The only stem
Table 1 List of the primers used in this study
Primer Sequence (50–[30) References Taxon PCR assays on P. wasabiae
RNS 08.42.1Aa
ADE1 GATCAGAAAGCCCGCAGCCAG AT Nassar et al.
1996
Dickeya sp. –
ADE2 CTGTGGCCGATCAGGATGGTTTTGTCGTGC
Y1 TTACCGGACGCCGAGCTGTGGCGT Darrasse et al.
1994
P. atrosepticum and P.
carotovorum
1
Y2 CAGGAAGATGTCGTTATCGCGAGT
Y45 TCACCGGACGCCAACTGTGGCGT Frechon et al.
1998
P. atrosepticum –
Y46 TCGCCACGTTTCAGCAGAACAAGT
Phf GGTTCAGTGCGTCAGGAGAG De Boer et al.
2012
P. wasabiae 1
Phr GCGGAGAGGAAGCGGTGAAG
PW7011F CTATGACGCTCGCGGGTTGCTGTT Kim et al.
2012
P. wasabiae 1
PW7011R CGGCGGCGTCGTAGTGGAAAGTC
recAF CCTTCACCATACATAATTTGTATCATGCG Waleron et al.
2002
P. wasabiae 1
recAR CCTTCACCATACATAATTTGGA
a Presence (?) or absence (-) of PCR products using P. wasabiae RNS 08.42.1A total DNA as a target
Genetica
123

presented soft rot symptom appearing green at the basal
level to black at margin of the symptom, at the top level of
the plant (Fig. 1a). A black rotting of the basal leaflet could
also be observed, associated to yellowing of the entire
leaflet (Fig. 1b). PCR-based amplification and sequencing
of some gene markers (Table 1) supported this strain as
belonging to P. wasabiae species. This was a first descrip-
tion of P. wasabiae in France, hence motivating a genome
sequencing and analysis of the strain RNS 08.42.1A.
Pectobacterium wasabiae RNS 08.42.1A genome
sequence
After trimming of the sequences reads, 23 866 905 mate
pair reads were obtained with an average length of 87.8 bp.
The de novo assembly followed by scaffolding and gap
closure generated 27 contigs ([2 000 bp) with an average
coverage of 380 fold. The average length of the contigs
was 177 765 bp, with a maximal contig size of 1 340
431 bp. The N50 contig size reached 448,690 bp. The final
draft genome (5,015,350 bp) consisted in 8 scaffolds
ranging from 2 030 to 3 310 633 bp in size with a G?C
content at 50.0 %. A gene RAST annotation generated a
total number of 4,606 protein coding genes which exhibited
either an assigned (3 398 of them, i.e. 74 %) or hypothet-
ical function (1 208 of them, i.e. 26 %). Among the first
category, 95 genes coding for non-translated RNAs
including 22 ribosomal RNA (rrs) genes were identified
(Table 2). The number of the predicted missing genes
during assembly is estimated to 39 genes (0.8 % of the
total number of protein coding genes). The RNS 08.42.1A
strain exhibited significant number of phage and trans-
posable elements with 48,585 bps in cumulative size.
These acquired horizontal genes play an important role in
the evolution of the bacterial and archaeal genomes
(Gogarten et al. 2002; Lang et al. 2012). The strain RNS
08.42.1A exhibited the largest repertoire of phage elements
amongst all P. wasabiae genomes (Table 2). This feature
suggests that this strain is capable to acquire efficiently
large regions of extracellular DNA.
Positioning P. wasabiae RNS 08.42.1A
within Pectobacterium and Dickeya
A MLSA was performed, based on 4 housekeeping genes
(fusA, gyrB, dnaX, recA) from all P. wasabiae strains
available from the NCBI database. P. wasabiae strain RNS
08.42.1A clustered with all other P. wasabiae strains
(Fig. 2). This analysis also showed that the P. wasabiae
cluster can be divided into 2 phyla, hence separating all the
P. wasabiae strains isolated form potato host plants from
the strain type P. wasabiae CFBP 3304T isolated from
wasabi in Japan. A previous study has proposed that this
phylogenetic separation resulted in the emergence of two
subspecies associated with two different hosts, potato and
horseradish plants (Nykyri et al. 2012).
The existence of the P. wasabiae potato host-cluster is
also supported by the calculation of ANI value. The ANI is
proposed as an alternative to DNA–DNA hybridization in
taxonomic studies: ANI values between genomes of the
same species are above 95 % (Goris et al. 2007). Here, the
ANI values between P. wasabiae RNS 08.42.1A and the
other potato-associated P. wasabiae strains were higher
than 99 %, indicating a very close relatedness of these
strains. In contrast, the ANI value between P. wasabiae
RNS 08.42.1A and P. wasabiae CFBP 3304T reached only
94 %. This value could suggest that the P. wasabiae strains
collected from potato plants would belong to a novel P.
wasabiae subspecies or a novel species (Gardan et al. 2003;
Samson et al. 2005; Nykyri et al. 2012). This hypothesis
however requires additional taxonomic investigations.
Synteny relatedness among P. wasabiae strains
To gain a better view at the relationship of P. wasabiae
strains, we used the MAUVE program to perform a
Fig. 1 Blackleg symptoms in greenhouse plant assay. a Blackleg
symptoms extend from the bottom to the top level of the stem. The
arrow indicates the insertion point of the rotted leaflet observed in
association with stem rotting b Leaf rot symptom appeared as the
blackening of the entire petiole associated with the yellowing and
humid necrosis of the leaflet
Genetica
123

multiple genome alignment of P. wasabiae RNS 08.42.1A
genome and those of P. wasabiae WPP163 and P. wasa-
biae SCC3193 (Fig. 3). Because the genome of the P.
wasabiae RNS 08.42.1A is under a draft-genome format,
we also used a move-contig tool that selects the contig
order to maximize the synteny. The P. wasabiae RNS
08.42.1A genome differs by 9.3 and 8.7 % from P. wasa-
biae WPP163 and SCC3193 genomes, respectively. This
pairwise alignment supports the closeness between these
three P. wasabiae isolates collected from the same host
plant.
PCWDEs and virulence determinants
The genes and clusters that encode virulence factors and
host interacting functions were searched in P. wasabiae
RNS 08.42.1A using the two complete genome sequences
of the P. wasabiae strains WPPP163 and SCC3193 as
references.
Plant cell wall-degrading enzymes of pectolytic bacteria
are responsible for the disorganization and maceration of
the plant cell wall resulting in soft rot and blackleg disease
on several hosts (Hugouvieux-Cotte-Pattat et al. 1996).
Using nucleotide BLAST searches, we have identified all
known or putative pectinases, cellulases and proteases that
were previously described to be shared among the P.
wasabiae species (Nykyri et al. 2012). No less than 99 %
of identity for each gene was calculated, highlighting a
high conservation of these virulence genes amongst
members of the P. wasabiae species. Briefly there were 18
genes coding for pectinases including pectate, pectin lya-
ses, pectine methyl and acetyl esterases and polygalactu-
ronases. Aside, 16 genes coding for proteases and 3 genes
for cellulases were detected (Table 3). The PehK and
Table 2 Some characteristics of the P. wasabiae strains and their genome sequences
Strain Number
of rrs
Number
of tRNA
Phages
elements (bp)
Host plant Year Origin References
P. wasabiae RNS 08.42.1A 22 73 48,585 Potato 2010 France This work
P. wasabiae CFBP 3304T 23 60 58,488 Eutrema wasabi 1985 Japan Goto and Matsumoto (1987)
P. wasabiae CFIA1002 22 73 47,847 Potato 2007 Canada Yuan et al. (2014)
P. wasabiae SCC3193 22 77 19,899 Potato 1980 Finland Koskinen et al. (2012)
P. wasabiae WPP163 22 75 10,536 Potato 2004 USA Kim et al. (2009)
Fig. 2 MLSA-based (fusA, gyrB, recA, dnaX) relation tree and ANI values using P. wasabiae RNS 08.42.1A as a reference. The sequences were
aligned with ClustalW, and then a Neighbour-joining tree was created by Bootstrap method with 1,000 bootstrap replications
Genetica
123

HrpW potential virulence factors and a putative specific
pectate lyase present in other Pectobacterium species
(Nykyri et al. 2012) were absent in P. wasabiae RNS
08.42.1A. It remains unclear whether the lack of PehK,
HrpW and the pectate lyase present in other Pectobacte-
rium has an effect on the virulence of P. wasabiae (Kim
et al. 2009).
Other Pectobacterium virulence determinants were also
shared by all the P. wasabiae strains except the two
putative LPS (liposaccharides) encoding clusters described
in SCC3193 (LPS1:W5S_4520-4538 and
LPS2:W5S_3001-3022) which contained a set of genes not
found in RNS 08.42.1A strain. A total of 19 genes are
composing the LPS1 cluster and 22 genes the LPS2 cluster
of the strain SCC3193. The search of these clusters by
BLASTn in the RNS 08.42.1A genome revealed the con-
servation of only 8 genes for LPS1 and 6 genes for LPS2,
with high similarity (Table 4). In order to check if these
missing LPS genes were absent in P. wasabiae RNS
08.42.1A or just misassembled, we mapped trimmed mate
pair reads of P. wasabiae RNS 08.42.1A strain against each
LPS cluster of SCC3193 strain. As a result, no read mat-
ched the missing genes from LPS clusters.
Interestingly these two LPS clusters were completely
conserved in WPP163 strain and partially present in RNS
08.42.1A and CFIA1002 but they were absent in the type
strain CFBP 3304T. The LPS is a key component of the
outer membrane of the gram-negative bacteria which
comprises three components: the lipid A, an oligosaccha-
ride core and usually a polysaccharide O antigen (Reeves
and Wang 2002). Nykyri et al. (2012) suggested that the
variation of LPS composition within Pectobacterium may
indicate an adaptation of the strains to different
environments.
P. wasabiae RNS 08.42.1A displays an original expI–
expR1 system
The genes coding for the regulation of virulence in soft rot
bacteria have been largely studied and characterized (Toth
et al. 2006). The complex regulatory network of the
P. wasabiae strain described by Nykyri et al. (2012) was
used to search for such genes in RNS 08.42.1A strain. All
these genes were highly conserved, with a percentage of
identity above 99 %, except those of the quorum sensing
(QS) system expI–expR1 that display a lower similarity
(Table 5). The ExpI protein synthesizes diffusible auto
inducers of the NAHL class, while ExpR1 acts as a
repressor (Nasser et al. 1998; Jones et al. 1993). Synteny
and nucletotide identify of the genomic block containing
the determinants of the QS regulatory system (25 kbp) are
highly conserved among the P. wasabiae strains, although
QS locus expI–expR exhibits a low similarity (Fig. 4a).
This feature suggests horizontal acquisition of the expI–
expR only in the P. wasabiae strain RNS 08.42.1A.
Regarding the protein sequence, ExpI and ExpR of P.
wasabiae RNS 08.42.1A are phylogenetically closest to
those of P. atrosepticum and P. carotovorum than those of
others P. wasabiae (Fig. 4b). The discrepancy between the
MLSA-phylogeny (Fig. 2) and ExpI–ExpR1-phylogeny
(Fig. 4a) reinforces the horizontal transfer hypothesis in the
P. wasabiae strain RNS 08.42.1A. The ExpI–ExpR phy-
logeny also designates the other Pectobacterium potato
pathogens as potential donors of the expI–expR genes of P.
Fig. 3 Synteny between P. wasabiae strains RNS 08.42.1A, SCC3193 and WPP163. Pairwise alignments of the genomes were generated using
Mauve
Genetica
123
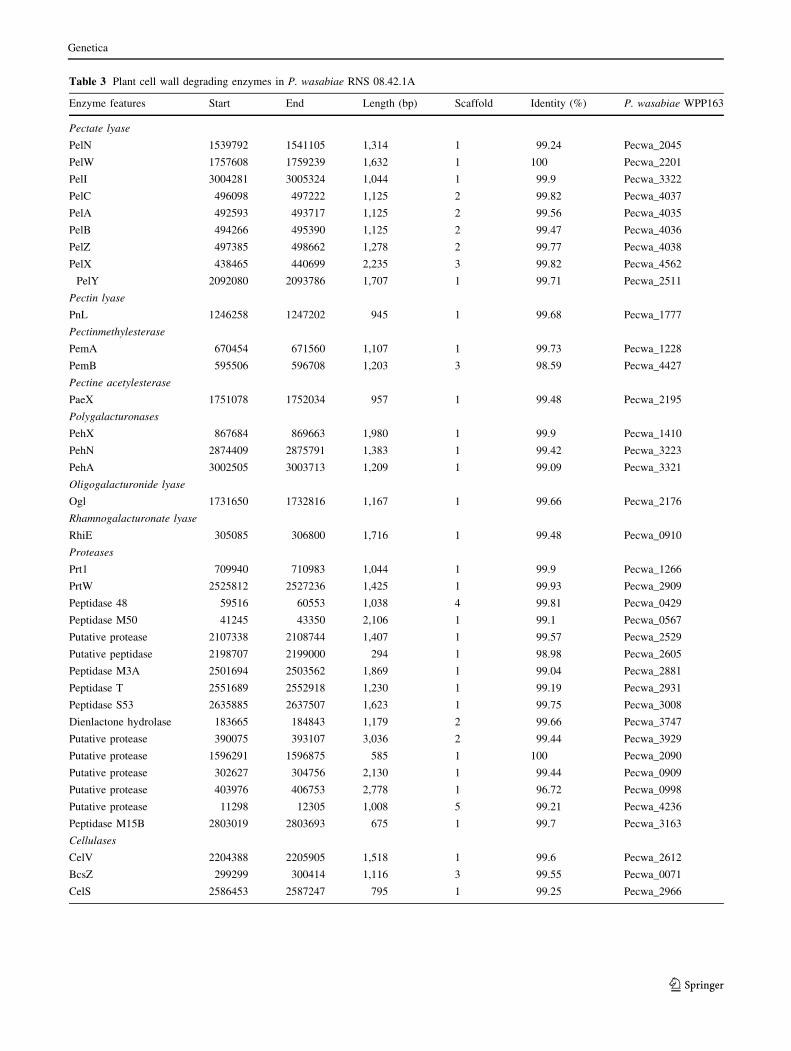
Table 3 Plant cell wall degrading enzymes in P. wasabiae RNS 08.42.1A
Enzyme features Start End Length (bp) Scaffold Identity (%) P. wasabiae WPP163
Pectate lyase
PelN 1539792 1541105 1,314 1 99.24 Pecwa_2045
PelW 1757608 1759239 1,632 1 100 Pecwa_2201
PelI 3004281 3005324 1,044 1 99.9 Pecwa_3322
PelC 496098 497222 1,125 2 99.82 Pecwa_4037
PelA 492593 493717 1,125 2 99.56 Pecwa_4035
PelB 494266 495390 1,125 2 99.47 Pecwa_4036
PelZ 497385 498662 1,278 2 99.77 Pecwa_4038
PelX 438465 440699 2,235 3 99.82 Pecwa_4562
PelY 2092080 2093786 1,707 1 99.71 Pecwa_2511
Pectin lyase
PnL 1246258 1247202 945 1 99.68 Pecwa_1777
Pectinmethylesterase
PemA 670454 671560 1,107 1 99.73 Pecwa_1228
PemB 595506 596708 1,203 3 98.59 Pecwa_4427
Pectine acetylesterase
PaeX 1751078 1752034 957 1 99.48 Pecwa_2195
Polygalacturonases
PehX 867684 869663 1,980 1 99.9 Pecwa_1410
PehN 2874409 2875791 1,383 1 99.42 Pecwa_3223
PehA 3002505 3003713 1,209 1 99.09 Pecwa_3321
Oligogalacturonide lyase
Ogl 1731650 1732816 1,167 1 99.66 Pecwa_2176
Rhamnogalacturonate lyase
RhiE 305085 306800 1,716 1 99.48 Pecwa_0910
Proteases
Prt1 709940 710983 1,044 1 99.9 Pecwa_1266
PrtW 2525812 2527236 1,425 1 99.93 Pecwa_2909
Peptidase 48 59516 60553 1,038 4 99.81 Pecwa_0429
Peptidase M50 41245 43350 2,106 1 99.1 Pecwa_0567
Putative protease 2107338 2108744 1,407 1 99.57 Pecwa_2529
Putative peptidase 2198707 2199000 294 1 98.98 Pecwa_2605
Peptidase M3A 2501694 2503562 1,869 1 99.04 Pecwa_2881
Peptidase T 2551689 2552918 1,230 1 99.19 Pecwa_2931
Peptidase S53 2635885 2637507 1,623 1 99.75 Pecwa_3008
Dienlactone hydrolase 183665 184843 1,179 2 99.66 Pecwa_3747
Putative protease 390075 393107 3,036 2 99.44 Pecwa_3929
Putative protease 1596291 1596875 585 1 100 Pecwa_2090
Putative protease 302627 304756 2,130 1 99.44 Pecwa_0909
Putative protease 403976 406753 2,778 1 96.72 Pecwa_0998
Putative protease 11298 12305 1,008 5 99.21 Pecwa_4236
Peptidase M15B 2803019 2803693 675 1 99.7 Pecwa_3163
Cellulases
CelV 2204388 2205905 1,518 1 99.6 Pecwa_2612
BcsZ 299299 300414 1,116 3 99.55 Pecwa_0071
CelS 2586453 2587247 795 1 99.25 Pecwa_2966
Genetica
123

wasabiae strain RNS 08.42.1A. Horizontal transfer of the
QS genes was reported in different species such as Pseu-
domonas aeruginosa (Gray and Garey 2001; Lerat and
Moran 2004).
The expI–expR1 genes exhibit another characteristics
which renders the QS system original in P. wasabiae strain
RNS 08.42.1A. The expI–expR1 system in P. wasabiae
RNS 08.42.1A displayed high homology (90 % of identity)
to those of P. atrosepticum SCRI1043 and P. carotovorum
subsp carotovorum PCC21, but a lower identity (70 %) to
that of P. atrosepticum CFBP 6276. Moreover, P. atro-
septicum SCRI1043 ExpI produces N-3-oxo-
hexanoylhomoserine lactone (3O,C6-HSL) as a main QS
signal (Bell et al. 2004), while P. atrosepticum CFBP 6276
ExpI produces N-3-oxo-octanoylhomoserine lactone
(3O,C8-HSL). This sequence similarity suggested that P.
wasabiae RNS 08.42.1A would produce 3O,C6-HSL as a
main QS-signal, but not 3O,C8-HSL. This hypothesis is
confirmed by a previous study reporting that 3O,C6-HSL is
indeed the main signal emitted by P. wasabiae RNS
08.42.1A (Crepin et al. 2012c). The introduction of a
plasmid that encodes a NAHL-lactonase in P. wasabiae
RNS 08.42.1A induced the quenching of the QS signal in
RNS 08.42.1A and a decrease of the virulence symptoms
Table 4 Virulence determinants in P. wasabiae RNS 08.42.1A
Virulence determinantsa Scaffold Start End Identity (%) P. wasabiae SCC3193
Flagella encoding cluster (48/51) Scaffold1 1324372 1369770 99.63 WS5_1760–WS5_1810
Flagella encoding cluster (3/51) Scaffold1 1370307 1370541 95.32
Enterobacterial common antigen (21/21) Scaffold2 666428 686800 99.36 WS5_4355–WS5_4365
Citrate transporter(1/1) Scaffold2 415571 417025 99.45 WS5_4105
Nip(1/1) Scaffold1 891610 892215 99.83 WS5_1316
Svx(1/1) Scaffold1 458640 460508 99.57 WS5_0937
budrab (3/3) Scaffold1 212836 216351 99.57 WS5_0740–WS5_0742
budc(1/1) Scaffold3 45781 46554 99.74 WS5_0317
LPS encoding cluster1(4/19) Scaffold3 648426 652897 98.17 WS5_4520–WS5_4538
LPS encoding cluster 1(2/19) Scaffold3 633689 635512 97.15
LPS encoding cluster 1(2/19) scaffold3 646215 648330 71.42
LPS encoding cluster2(3/22) Scaffold1 1438 3500 89.15 WS5_3001–WS5_3022
LPS encoding cluster2(3/22) Scaffold1 4906 5915 88.47
a Number of genes found/number of genes composing the cluster
Table 5 Virulence regulators in P. wasabiae RNS 08.42.1A
Virulence network or determinant Start End Length Scaffold Identity (%) P. wasabiae SCC3193
PmrAB 470461 472220 1,760 2 99.43 W5S_4173/4174
ExpI 593792 594371 651 3 74.01 W5S_4607
ExpR1 594442 595154 729 3 70.80 W5S_4606
ExpR2 1310935 1311672 738 1 99.86 W5S_1749
KdgR 1733032 1733823 792 1 100 W5S_2118
ExpAS 1154611 1155267 657 1 100 W5S_1457
Hor 2268354 2268791 438 1 100 W5S_2637
RsmA 534243 534428 186 1 100 W5S_1009
ExpAS-2 3266669 3269455 2,787 1 99.86 W5S_3687
LuxS 538608 539123 516 1 99.81 W5S_1019
ExpM 1831462 1832478 1,017 1 99.8 W5S_2224
PehRS-2 1655658 1657118 1,461 1 99.66 W5S_2095
Rcs 2852864 2859034 6,171 1 99.58 W5S_3208/3206
PehRS 1657115 1657804 690 1 99.42 W5S_2096
Genetica
123

on potato tubers (Crepin et al. 2012a). Hence, the 3O,C6-
HSL-mediated QS system is involved in the control of the
virulence functions in P. wasabiae RNS 08.42.1A.
Secretions systems
The protein secretion systems play an important role in the
pathogenicity of the soft rot bacteria. They facilitate
delivery of the virulence determinants across the bacterial
cell wall, outside the cell, to attack host plant and com-
peting bacteria (Charkowski 2006). P. wasabiae species
possesses four of the known gram-negative protein secre-
tion systems and the corresponding genes were highly
conserved in P. wasabiae RNS 08.42.1A. The extracellular
proteases are secreted through a T1SS, while the T2SS
exports the cellulases and pectinases. The role of the T4SS
remains unknown, but in P. atrosepticum it weakly con-
tributes to virulence (Bell et al. 2004). P. wasabiae strains
harbor two T6SS that were highly conserved in P. wasa-
biae RNS 08.42.1A (Nykyri et al. 2012). T6SS would
conduct the delivery of the predicted proteins Hcp
(heamolysin co-regulated proteins) and VgrG (valine-
glycine repeat protein G) which are major virulence
determinants in P. atrosepticum (Liu et al. 2008). We
observed that T3SS was absent in P. wasabiae RNS
08.42.1A, this was also reported for the other P. wasabiae
strains (Kim et al. 2009; Nykyri et al. 2012).
Toxin antitoxin HigA/HigB complex shared
only by RNS 08.42.1A
The Toxin–Antitoxin (TA) system modules are commonly
found on plasmids, as they contribute to their maintenance.
They are also encoded on chromosomes (Gerdes et al.
2005; Magnuson 2007). They play a crucial role in bac-
terial survival during environmental stresses, as they are
implicated in diverse functions such as programmed cell
death, cell division, biofilm formation (Schureck et al.
2014). Using a function based comparison tool from the
RAST server, some TA loci were identified only in P.
wasabiae RNS 08.42.1A and other TA loci are shared just
by the potato isolates (Supplementary data S1). One of
these, the vapBC (virulence associated protein) operon is
shared by P. wasabiae RNS 08.42.1A and the type strains
Fig. 4 The QS regulatory system ExpI–ExpR1 of P. wasabiae RNS 08.23.1A. a Nucleotidic similarity indicating a low identity of expI–expR of
RNS 08.42.1A strain compared to other P. wasabiae strains b ExpI–ExpR1 protein sequences based phylogenetic tree
Genetica
123

CFBP 3304T. It was previously identified on several plas-
mids of pathogenic bacteria where it contributes to their
stability (Gerdes et al. 2005). The cellular targets of the
VapC toxin remain unidentified (Gerdes et al. 2005).
The loci higBA (host inhibition of growth) was present
only in RNS 08.42.1A strain: It is a type II TA system
comprising small adjacent genes that code for a toxin and
an antitoxin proteins. These toxin and antitoxin proteins are
considered unusual compared to other TA systems, because
the toxin encoding gene is located upstream the antitoxin
encoding gene (Gerdes et al. 2005). It is known that the
antitoxins are more labile than the toxins and are readily
degraded under stress conditions, a feature that allows the
toxins to exert their toxic effect. Previous studies reported
that the endoribonuclease HigB protein recognizes AAA
sequences on mRNA. The activity of HigB toxin is
dependent on the association with ribosomes, facilitating
the recognition then the cleavage of specific AAA
sequence (Hurley and Woychik 2009; Bukowski et al.
2011). Exogenous expression of the toxin HigB in Vibrio
cholerae and Escherichia coli led to a drastic growth
limitation and a reduced number of CFU (Budde et al.
2007). Several studies stated the involvement of TA sys-
tems not only in normal bacterial physiology but also in
pathogenicity (Yamaguchi et al. 2011; De la Cruz et al.
2013; Wen et al. 2014). At present, however, there is no
evidence that would suggest the implication of TA systems
in the virulence of soft rot bacteria.
Finally, many genes were found to be specific to P.
wasabiae potato isolates, including genes that encode
maltose, maltodextrin and histidine degradation, or genes
that encode the utilization of galactosamine and that of its
N-acetyl derivative. Also aeorobactin and assembly kit
siderophores were identified only in potato isolate of P.
wasabiae including RNS 08.42.1A strain. They contribute
to iron uptake from the environment, in order to overcome
the starvation phases and enhance bacterial growth. In
relation, it was also shown that siderophores contribute to
the virulence in Dickeya (Boughammoura et al. 2007).
However the role of these features in the ecology and the
virulence of the soft rot bacteria remains unknown. Com-
bined with the whole set of specificity exposed above, these
data support the view that P. wasabiae potato isolates
might delineate a novel species or a novel subspecies.
Conclusions
Several works indicate that P. wasabiae may have been
present on potato fields and misidentified as P. carotovo-
rum in the past, when the biochemical and phenotypic
characteristics where not discriminating enough to allow
the classification of P. wasabiae as a defined species of the
Pectobacterium genus (Nabhan et al. 2012; Nykyri et al.
2012; Waleron et al. 2013). With the data generated by
DNA-based methods, P. wasabiae is now clearly separated
from other clade of soft rot bacteria. The main feature that
separates these bacteria from other Pectobacterium mem-
bers is the lack of a T3SS which is a secondary factor
contributing to pathogenesis in pectolytic bacteria (Toth
et al. 2006). Our study aimed at describing the french P.
wasabiae isolate through whole genome investigation. Our
functional comparative analyses revealed specific traits that
are shared only by potato-associated isolates, a feature that
supports the adaptation to the environment within the host.
Amongst these genes are those encoding maltose, malto-
dextrin, histidine and galactosamine utilization.
Pectobacterium wasabiae RNS 08.42.1A has an original
expI–expR1 complex which showed a low similarity with
those identified in other P. wasabiae strains. It is tempting
to speculate that RNS 08.42.1A has acquired the expI–
expR1 complex from another Pectobacterium by an hori-
zontal gene transfer event.
Acknowledgments This work was supported by a cooperative
project between France and Morocco (PRAD 14-02, Campus France
No. 30229 ZK), the excellence Grant (No. H011/007) awarded by the
Ministry of Higher education of Morocco, and a collaborative project
between Centre National de la Recherche Scientifique (CNRS, Gif sur
Yvette) and Federation Nationale des Producteurs de Plants de
Pomme de Terre-Recherche Developpement Promotion du Plants de
Pomme de Terre (FN3PT-RD3PT, Paris).
References
Aziz RK, Bartels D, Best AA, DeJongh M, Disz T, Edwards RA,
Formsma K, Gerdes S, Glass EM, Kubal M et al (2008) The
RAST server: rapid annotations using subsystems technology.
BMC Genomics 9:75. doi:10.1186/1471-2164-9-75
Baghaee-Ravari S, Rahimian H, Shams-Bakhsh M, Lopez-Solanilla
E, Antunez-Lamas M, Rodrıguez-Palenzuela P (2011) Charac-
terization of Pectobacterium species from Iran using biochem-
ical and molecular methods. Eur J Plant Pathol 129:413–425.
doi:10.1007/s10658-010-9704-z
Barras F, van Gijsegem F, Chatterjee AK (1994) Extracellular
enzymes and pathogenesis of soft-rot Erwinia. Annu Rev
Phytopathol 32:201–234. doi:10.1146/annurev.py.32.090194.
001221
Bell KS, Sebaihia M, Pritchard L, Holden MTG, Hyman LJ, Holeva
MC, Thomson NR, Bentley SD, Churcher LJC, Mungall K et al
(2004) Genome sequence of the enterobacterial phytopathogen
Erwinia carotovora subsp. atroseptica and characterization of
virulence factors. Proc Natl Acad Sci 101:11105–11110. doi:10.
1073/pnas.0402424101
Boughammoura A, Franza T, Dellagi A, Roux C, Matzanke-
Markstein B, Expert D (2007) Ferritins, bacterial virulence and
plant defence. Biometals 20:347–353. doi:10.1007/s10534-006-
9069-0
Budde PP, Davis BM, Yuan J, Waldor MK (2007) Characterization of
a higBA toxin–antitoxin locus in Vibrio cholerae. J Bacteriol
189:491–500. doi:10.1128/JB.00909-06
Genetica
123

Bukowski M, Rojowska A, Wladyka B (2011) Prokaryotic toxin–
antitoxin systems—the role in bacterial physiology and applica-
tion in molecular biology. Acta Biochim Pol 58:1–9
Charkowski AO (2006) The soft rot Erwinia. In: Gnanamanickam S
(ed) Plant-associated bacteria. Springer, Netherlands,
pp 423–505
Corbett M, Virtue S, Bell K, Birch P, Burr T, Hyman L, Lilley K,
Poock S, Toth I, Salmond G (2005) Identification of a new
quorum-sensing-controlled virulence factor in Erwinia caroto-
vora subsp. atroseptica secreted via the type II targeting
pathway. Mol Plant Microbe Interact 18:334–342
Crepin A, Barbey C, Beury-Cirou A, Helias V, Taupin L, Reverchon
S, Nasser W, Faure D, Dufour A, Orange N et al (2012a)
Quorum sensing signaling molecules produced by reference and
emerging soft-rot bacteria (Dickeya and Pectobacterium spp.).
PLoS ONE 7:e35176. doi:10.1371/journal.pone.0035176
Crepin A, Barbey C, Cirou A, Tannieres M, Orange N, Feuilloley M,
Dessaux Y, Burini J-F, Faure D, Latour X (2012b) Biological
control of pathogen communication in the rhizosphere: a novel
approach applied to potato soft rot due to Pectobacterium
atrosepticum. Plant Soil 358:27–37. doi:10.1007/s11104-011-
1030-5
Crepin A, Beury-Cirou A, Barbey C, Farmer C, Helias V, Burini J-F,
Faure D, Latour X (2012c) N-Acyl homoserine lactones in
diverse Pectobacterium and Dickeya plant pathogens: diversity,
abundance, and involvement in virulence. Sensors
12:3484–3497. doi:10.3390/s120303484
Czajkowski R, Perombelon MCM, van Veen JA, van der Wolf JM
(2011) Control of blackleg and tuber soft rot of potato caused by
Pectobacterium and Dickeya species: a review: control of
Dickeya and Pectobacterium species in potato. Plant Pathol
60:999–1013. doi:10.1111/j.1365-3059.2011.02470.x
Darling ACE (2004) Mauve: multiple alignment of conserved
genomic sequence with rearrangements. Genome Res
14:1394–1403. doi:10.1101/gr.2289704
Darrasse A, Priou S, Kotoujansky A, Bertheau Y (1994) PCR and
restriction fragment length polymorphism of a pel gene as a tool
to identify Erwinia carotovora in relation to potato diseases.
Appl Environ Microbiol 60:1437–1443
De Boer SH, Li X, Ward LJ (2012) Pectobacterium spp. associated
with bacterial stem rot syndrome of potato in Canada. Phyto-
pathology 102:937–947
De la Cruz MA, Zhao W, Farenc C, Gimenez G, Raoult D, Cambillau
C, Gorvel J-P, Meresse S (2013) A toxin–antitoxin module of
Salmonella promotes virulence in mice. PLoS Pathog
9:e1003827. doi:10.1371/journal.ppat.1003827
Frechon D, Exbrayat P, Helias V, Hyman LJ, Jouan B, Llop P, Lopez
MM, Payet N, Perombelon MCM, Toth IK (1998) Evaluation of
a PCR kit for the detection of Erwinia carotovora subsp.
atroseptica on potato tubers. Potato Res 41:163–173
Fuqua WC, Winans SC, Greenberg EP (1994) Quorum sensing in
bacteria: the LuxR–LuxI family of cell density-responsive
transcriptional regulators. J Bacteriol 176:269–275
Gardan L, Gouy C, Richard C, Samson R (2003) Elevation of three
subspecies of Pectobacterium carotovorum to species level:
Pectobacterium atrosepticum sp. nov., Pectobacterium betavas-
culorum sp. nov. and Pectobacterium wasabiae sp. nov. Int J
Syst Evol Microbiol 53:381–391. doi:10.1099/ijs.0.02423-0
Gerdes K, Christensen SK, Løbner-Olesen A (2005) Prokaryotic
toxin–antitoxin stress response loci. Nat Rev Microbiol
3:371–382. doi:10.1038/nrmicro1147
Gogarten JP, Doolittle WF, Lawrence JG (2002) Prokaryotic evolu-
tion in light of gene transfer. Mol Biol Evol 19:2226–2238
Goris J, Konstantinidis KT, Klappenbach JA, Coenye T, Vandamme
P, Tiedje JM (2007) DNA–DNA hybridization values and their
relationship to whole-genome sequence similarities. Int J Syst
Evol Microbiol 57:81–91. doi:10.1099/ijs.0.64483-0
Goto M, Matsumoto K (1987) Erwinia carotovora subsp. wasabiae
subsp. nov. Isolated from Diseased Rhizomes and Fibrous Roots
of Japanese Horseradish (Eutrema wasabi Maxim.). Int J Syst
Evol Microbiol 37(2):130–135. doi:10.1099/00207713-37-2-130
Gray KM, Garey JR (2001) The evolution of bacterial LuxI and LuxR
quorum sensing regulators. Microbiology 147:2379–2387
Helias V, Hamon P, Huchet E, Wolf JVD, Andrivon D (2012) Two
new effective semiselective crystal violet pectate media for
isolation of Pectobacterium and Dickeya: isolating pectolytic
bacteria on CVP. Plant Pathol 61:339–345. doi:10.1111/j.1365-
3059.2011.02508.x
Hugouvieux-Cotte-Pattat N, Condemine G, Nasser W, Reverchon S
(1996) Regulation of pectinolysis in Erwinia chrysanthemi.
Annu Rev Microbiol 50:213–257
Hurley JM, Woychik NA (2009) Bacterial toxin HigB associates with
ribosomes and mediates translation-dependent mRNA cleavage
at A-rich sites. J Biol Chem 284:18605–18613. doi:10.1074/jbc.
M109.008763
Jones S, Yu B, Bainton NA, Birdsall M, Bycroft BW, Chhabra SR,
Cox AJ, Golby P, Reeves PJ, Stephens S et al (1993) The Lux
autoinducer regulates the production of exoenzyme virulence
determinants in Erwinia carotovora and Pseudomonas aerugin-
osa. EMBO J 12:2477
Kim H-S, Ma B, Perna NT, Charkowski AO (2009) Phylogeny and
virulence of naturally occurring type III secretion system-
deficient Pectobacterium strains. Appl Environ Microbiol
75:4539–4549. doi:10.1128/AEM.01336-08
Kim MH, Cho MS, Kim BK, Choi HJ, Hahn JH, Kim C, Kang MJ,
Kim SH, Park DS (2012) Quantitative real-time polymerase
chain reaction assay for detection of Pectobacterium wasabiae
using YD repeat protein gene-based primers. Plant Dis
96:253–257
Koskinen JP, Laine P, Niemi O, Nykyri J, Harjunpaa H, Auvinen P,
Paulin L, Pirhonen M, Palva T, Holm L (2012) Genome
sequence of Pectobacterium sp. strain SCC3193. J Bacteriol
194:6004. doi:10.1128/JB.00681-12
Lang AS, Zhaxybayeva O, Beatty JT (2012) Gene transfer agents:
phage-like elements of genetic exchange. Nat Rev Microbiol.
doi:10.1038/nrmicro2802
Lerat E, Moran NA (2004) The evolutionary history of quorum-
sensing systems in bacteria. Mol Biol Evol 21:903–913. doi:10.
1093/molbev/msh097
Liu H, Coulthurst SJ, Pritchard L, Hedley PE, Ravensdale M,
Humphris S, Burr T, Takle G, Brurberg M-B, Birch PRJ et al
(2008) Quorum sensing coordinates rrute force and stealth
modes of infection in the plant pathogen Pectobacterium
atrosepticum. PLoS Pathog 4:e1000093. doi:10.1371/journal.
ppat.1000093
Ma B, Hibbing ME, Kim H-S, Reedy RM, Yedidia I, Breuer J, Breuer
J, Glasner JD, Perna NT, Kelman A et al (2007) Host range and
molecular phylogenies of the soft rot enterobacterial genera
Pectobacterium and Dickeya. Phytopathology 97:1150–1163.
doi:10.1094/PHYTO-97-9-1150
Magnuson RD (2007) Hypothetical functions of toxin-antitoxin
systems. J Bacteriol 189:6089–6092. doi:10.1128/JB.00958-07
Moleleki LN, Onkendi EM, Mongae A, Kubheka GC (2013)
Characterisation of pectobacterium wasabiae causing blackleg
and soft rot diseases in South Africa. Eur J Plant Pathol
135:279–288. doi:10.1007/s10658-012-0084-4
Nabhan S, Wydra K, Linde M, Debener T (2012) The use of two
complementary DNA assays, AFLP and MLSA, for epidemic
and phylogenetic studies of pectolytic enterobacterial strains
with focus on the heterogeneous species Pectobacterium
Genetica
123

carotovorum: phylogenetic focus on soft-rot plant pathogens.
Plant Pathol 61:498–508. doi:10.1111/j.1365-3059.2011.02546.x
Nassar A, Darrasse A, Lemattre M, Kotoujansky A, Dervin C, Vedel
R, Bertheau Y (1996) Characterization of Erwinia chrysanthemi
by pectinolytic isozyme polymorphism and restriction fragment
length polymorphism analysis of PCR-amplified fragments of
pel genes. Appl Environ Microbiol 62:2228–2235
Nasser W, Bouillant ML, Salmond G, Reverchon S (1998) Charac-
terization of the Erwinia chrysanthemi expI–expR locus directing
the synthesis of two N-acyl-homoserine lactone signal mole-
cules. Mol Microbiol 29:1391–1405. doi:10.1046/j.1365-2958.
1998.01022.x
Nasser W, Dorel C, Wawrzyniak J, Van Gijsegem F, Groleau M-C,
Deziel E, Reverchon S (2013) Vfm a new quorum sensing
system controls the virulence of Dickeya dadantii: new quorum
sensing signal in Dickeya. Environ Microbiol 15:865–880.
doi:10.1111/1462-2920.12049
Ngadze E, Brady CL, Coutinho TA, van der Waals JE (2012)
Pectinolytic bacteria associated with potato soft rot and blackleg
in South Africa and Zimbabwe. Eur J Plant Pathol 134:533–549.
doi:10.1007/s10658-012-0036-z
Nykyri J, Niemi O, Koskinen P, Nokso-Koivisto J, Pasanen M,
Broberg M, Plyusnin I, Toronen P, Holm L, Pirhonen M et al
(2012) Revised phylogeny and novel horizontally acquired
virulence determinants of the model soft rot phytopathogen
Pectobacterium wasabiae SCC3193. PLoS Pathog 8:e1003013.
doi:10.1371/journal.ppat.1003013
Perombelon MCM (2002) Potato diseases caused by soft rot erwinias:
an overview of pathogenesis. Plant Pathol 51:1–12
Pitman AR, Harrow SA, Visnovsky SB (2010) Genetic characterisa-
tion of Pectobacterium wasabiae causing soft rot disease of
potato in New Zealand. Eur J Plant Pathol 126:423–435. doi:10.
1007/s10658-009-9551-y
Reeves PP, Wang L (2002) Genomic organization of LPS-specific
loci. In: Hacker J, Kaper JB (eds) Pathogenicity islands and the
evolution of pathogenic microbes. Springer, Berlin, Heidelberg,
pp 109–135
Samson R, Legendre JB, Richard C, Fischer-Le Saux M, Achouak W,
Gardan L et al (2005) Transfer of Pectobacterium chrysanthemi
(Burkholder, 1953) Brenner et al. 1973 and Brenneria paradisi-
aca to the genus Dickeya gen. nov. as Dickeya chrysanthemi
comb. nov. and Dickeya paradisiaca comb. nov. and delineation
of four novel species, Dickeya dadantii sp. nov., Dickeya
dianthicola sp. nov., Dickeya dieffenbachiae sp. nov. and
Dickeya zeae sp. nov. Int J Syst Evol Microbiol
55:1415–1427. doi:10.1099/ijs.0.02791-0
Schureck MA, Maehigashi T, Miles SJ, Marquez J, Cho SE, Erdman
R, Dunham CM (2014) Structure of the Proteus vulgaris HigB-
(HigA)2-HigB toxin-antitoxin complex. J Biol Chem
289:1060–1070. doi:10.1074/jbc.M113.512095
Smadja B, Latour X, Trigui S, Burini JF, Chevalier S, Orange N
(2004) Thermodependence of growth and enzymatic activities
implicated in pathogenicity of two Erwinia carotovora subspe-
cies (Pectobacterium spp.). Can J Microbiol 50:19–27. doi:10.
1139/w03-099
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013)
MEGA6: molecular evolutionary genetics analysis version 6.0.
Mol Biol Evol 30:2725–2729. doi:10.1093/molbev/mst197
Toth IK, Bell KS, Holeva MC, Birch PR (2003) Soft rot erwiniae:
from genes to genomes. Mol Plant Pathol 4:17–30
Toth IK, Pritchard L, Birch PR (2006) Comparative genomics reveals
what makes an enterobacterial plant pathogen. Annu Rev
Phytopathol 44:305–336
Waleron M, Waleron K, Podhajska AJ, Lojkowska E (2002) Genotyping
of bacteria belonging to the former Erwinia genus by PCR-RFLP
analysis of a recA gene fragment. Microbiology 148:583–595
Waleron M, Waleron K, Lojkowska E (2013) Occurrence ofPectobacterium wasabiae in potato field samples. Eur J Plant
Pathol 137:149–158. doi:10.1007/s10658-013-0227-2
Wen Y, Behiels E, Devreese B (2014) Toxin-Antitoxin systems: their
role in persistence, biofilm formation, and pathogenicity. Pathog
Dis 70:240–249. doi:10.1111/2049-632X.12145
Whitehead NA, Byers JT, Commander P, Corbett MJ, Coulthurst SJ,
Everson L, Harris AKP, Pemberton CL, Simpson NJL, Slater H
et al (2002) The regulation of virulence in phytopathogenic
Erwinia species: quorum sensing, antibiotics and ecological
considerations. Antonie Van Leeuwenhoek 81:223–231
Wilson K (1987) Preparation of genomic DNA from bacteria. In:
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG,
Smith JA, Struhl K (eds) Protocols in molecular biology. Greene
Publishing and Wiley- Interscience, New York, pp 2.4.1–2.4.5
Yamaguchi Y, Park J-H, Inouye M (2011) Toxin–antitoxin systems in
bacteria and archaea. Annu Rev Genet 45:61–79. doi:10.1146/
annurev-genet-110410-132412
Yuan K, Adam Z, Tambong J, Levesque CA, Chen W, Lewis CT, De
Boer SH, Li X (2014) Draft Genome sequence of Pectobacte-
rium wasabiae strain CFIA1002. Genome Announc
2:e00214–14. doi:10.1128/genomeA.00214-14
Genetica
123




















