
Symbiosis-stage associated alterations in quorum sensing autoinducer molecules biosynthesis in...
12
REGULAR ARTICLE Symbiosis-stage associated alterations in quorum sensing autoinducer molecules biosynthesis in Sinorhizobium meliloti Jerzy Wielbo & Joanna Golus & Monika Marek-Kozaczuk & Anna Skorupska Received: 6 May 2009 / Accepted: 2 September 2009 # Springer Science + Business Media B.V. 2009 Abstract Sinorhizobium meliloti strains use several N-acylhomoserine lactone (AHL) autoinducers to sense the population density during saprophytic growth. Using a sensitive gfp-based AHL biosensor system, we show that synthesis of short-chain AHL molecules is inhibited (or significantly diminished) during the symbiotic phase of growth and is unde- tectable in the nitrogen fixing bacteroids. In the saprophytic zone V of nodules occupied by intensively proliferating vegetative forms of rhizobia, AHL pro- duction recommenced and bright-green fluorescence was observed concomitantly with increasing population density. Keywords Sinorhizobium-alfalfa symbiosis . Quorum sensing . AHL detection Introduction Quorum sensing (QS) is a cell–cell signaling process based on the production and detection of small diffusible molecules that enables bacteria to regulate gene expression as a function of population density. This ability to communicate and regulate metabolism and communal behavior allows the emergence of bacterial traits detectable only at a particular population level, such as bioluminescence, plasmid conjugal transfer, biofilm formation, as well as the coordination of interactions with higher organisms (Cha et al. 1998; Mc Dougald et al. 2007). Several QS systems have been described in Proteobacteria. Rhizobia, like other Gram-negative bacteria, often use N-acylhomoserine lactones (AHLs) as signal molecules, and a single strain may produce a set of AHLs that vary with respect to the length and substituent at the third carbon of acyl side chain (Cha et al. 1998; Lithgow et al. 2001; Teplitski et al. 2003). Well-characterized S. meliloti strains Rm8530 and Rm41 harbor at least two QS systems: the ExpR/SinI/ SinR and TraR/TraI (Gurich and González 2009; Marketon et al. 2002; Marketon and González 2002). The expR/sinR/sinI QS system in Rm8530 is respon- sible for the production long-chain AHLs (C 12 -HSL to C 18 -HSL) (Gurich and González 2009), whereas tra QS system identified in Rm41 mediates plasmid transfer and it is responsible for production of at least three different short-chain AHLs, 3-oxo-C 8 -HSL, Plant Soil DOI 10.1007/s11104-009-0166-z Responsible Editor: Euan K. James. J. Wielbo (*) : M. Marek-Kozaczuk : A. Skorupska Department of Genetics and Microbiology, Institute of Microbiology and Biotechnology, Maria Curie-Skłodowska University, Akademicka 19 st., 20-033 Lublin, Poland e-mail: [email protected] J. Golus Chair and Department of Biochemistry, Medical University of Lublin, Chodźki 1 st., 20-093 Lublin, Poland
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Symbiosis-stage associated alterations in quorum sensing autoinducer molecules biosynthesis in...

REGULAR ARTICLE
Symbiosis-stage associated alterations in quorum sensingautoinducer molecules biosynthesis in Sinorhizobium meliloti
Jerzy Wielbo & Joanna Golus &
Monika Marek-Kozaczuk & Anna Skorupska
Received: 6 May 2009 /Accepted: 2 September 2009# Springer Science + Business Media B.V. 2009
Abstract Sinorhizobium meliloti strains use severalN-acylhomoserine lactone (AHL) autoinducers tosense the population density during saprophyticgrowth. Using a sensitive gfp-based AHL biosensorsystem, we show that synthesis of short-chain AHLmolecules is inhibited (or significantly diminished)during the symbiotic phase of growth and is unde-tectable in the nitrogen fixing bacteroids. In thesaprophytic zone Vof nodules occupied by intensivelyproliferating vegetative forms of rhizobia, AHL pro-duction recommenced and bright-green fluorescencewas observed concomitantly with increasing populationdensity.
Keywords Sinorhizobium-alfalfa symbiosis .
Quorum sensing . AHL detection
Introduction
Quorum sensing (QS) is a cell–cell signaling processbased on the production and detection of smalldiffusible molecules that enables bacteria to regulategene expression as a function of population density.This ability to communicate and regulate metabolismand communal behavior allows the emergence ofbacterial traits detectable only at a particular populationlevel, such as bioluminescence, plasmid conjugaltransfer, biofilm formation, as well as the coordinationof interactions with higher organisms (Cha et al. 1998;Mc Dougald et al. 2007).
Several QS systems have been described inProteobacteria. Rhizobia, like other Gram-negativebacteria, often use N-acylhomoserine lactones (AHLs)as signal molecules, and a single strain may produce aset of AHLs that vary with respect to the length andsubstituent at the third carbon of acyl side chain (Chaet al. 1998; Lithgow et al. 2001; Teplitski et al. 2003).Well-characterized S. meliloti strains Rm8530 andRm41 harbor at least two QS systems: the ExpR/SinI/SinR and TraR/TraI (Gurich and González 2009;Marketon et al. 2002; Marketon and González 2002).The expR/sinR/sinI QS system in Rm8530 is respon-sible for the production long-chain AHLs (C12-HSLto C18-HSL) (Gurich and González 2009), whereastra QS system identified in Rm41 mediates plasmidtransfer and it is responsible for production of at leastthree different short-chain AHLs, 3-oxo-C8-HSL,
Plant SoilDOI 10.1007/s11104-009-0166-z
Responsible Editor: Euan K. James.
J. Wielbo (*) :M. Marek-Kozaczuk :A. SkorupskaDepartment of Genetics and Microbiology,Institute of Microbiology and Biotechnology,Maria Curie-Skłodowska University,Akademicka 19 st.,20-033 Lublin, Polande-mail: [email protected]
J. GolusChair and Department of Biochemistry,Medical University of Lublin,Chodźki 1 st.,20-093 Lublin, Poland

3-OH-C8-HSL, and C8-HSL (Marketon et al. 2003).AHL molecules can be identified by analyticalmethods, and the majority may also be detected usingbiosensors, i.e. bacterial strains as such or inconjunction with dedicated plasmid constructs. Forexample, in the presence of exogenous AHLs,Chromobacterium violaceum CV026 producespurple pigment, violacein (McClean et al. 1997),Agrobacterium tumefaciens NT1 expresses lacZ genelocated under the control of traG promoter (Shaw etal. 1997), and E. coli JB534-MT102 produces GFPprotein from a gene under luxR/luxI transcriptionalcontrol (Andersen et al. 2001). The sensitivity ofbiosensors may vary, but all of them are useful due totheir reliability in AHL screens (Jafra and van derWolf 2004).
Rhizobium, Sinorhizobium, Mesorhizobium, andBradyrhizobium known by the common name ofrhizobia, are heterotrophic soil bacteria, whichestablish a symbiotic relationship with leguminousplants. Rhizobia synthesize morphogenetic signalmolecules (Nod factors), which elicit new structurescalled nodules on plant roots. The bacteria colonizenodules through tubular structures named infectionthreads, convert themselves into bacteroids, alter theirmetabolism and begin reducing dinitrogen insideplant cells. The reduced nitrogen is transferred toplants in exchange for assimilates (for review seeJones et al. 2007; Perret et al. 2000). In a matureindeterminate nodule formed by some plants amongthem alfalfa, six developmental zones may bedistinguished: the apical meristematic zone I; theinfection zone II in which bacteria are released frominfection threads; the interzone II–III characterized bymajor developmental changes and the accumulationof starch; zone III in which bacteroids fix nitrogen;senescent zone IV, characterized by the degenerationof symbionts; and, finally, saprophytic zone V, inwhich new rounds of bacterial release occur from theremaining infection threads and where saprophyticbacteria proliferate (Timmers et al. 2000; Vasse et al.1990).
It has been suggested that QS in rhizobia may beinvolved in regulation of numerous processes in thenon-symbiotic development stage, such as growth,biofilm formation, toxin production, virulence factorproduction, plasmid transfer, motility and exopoly-saccharide production (for review see Daniels et al.2004; González and Keshavan 2006; González and
Marketon 2003). Studies of the expression of genes,which are regulated by QS signals, suggest that theAHLs might be engaged in some aspects of theRhizobium-plant symbiosis, such as nodulation effi-ciency, nitrogen fixation or symbiosome development(for review see Wisniewski-Dye and Downie 2002;González and Keshavan 2006). The expression ofcinR and cinI genes of QS system and production of atleast three different AHLs autoinducers in bacteroidsof Rhizobium etli CNPAF512 was evidenced byDaniels et al. (2002).
Here, using Sinorhizobium meliloti-alfalfa symbioticsystem as a model, we demonstrate that the synthesis ofshort-chain AHLs, which can be detected by pJBA130sensor plasmid, is inhibited in nitrogen-fixing bacteroidsduring nodule development. Moreover, we show thatthe AHLs are continuously produced by vegetativeforms of bacteria located in the senescent andsaprophytic zones of nodules.
Materials and methods
Bacterial strains
The strains and plasmids used in this study are listedin Table 1. Sinorhizobium meliloti strains have beenisolated from root nodules of alfalfa (Medicago mediaPers. cv. Radius) grown in arable soil in the region ofLublin, Poland (Marek-Kozaczuk et al., submitted). S.meliloti and Agrobacterium tumefaciens were culturedon 79CA (Vincent 1970) or TY medium (Sambrook etal. 1989). Escherichia coli and Chromobacteriumviolaceum CV026 were cultured on TY or LB medium(Sambrook et al. 1989). When needed, tetracycline(Tc) was added to media at the concentration of10 µg ml−1.
Plasmid construct
Standard techniques were used for plasmid isolation,restriction enzyme digestion, cloning and transforma-tion (Sambrook et al. 1989). To construct vectorcontaining constitutively expressed gfp gene, the 1 kbEcoRI-HindIII fragment from plasmid construct pBK-miniTn7-gfp1 (Koch et al. 2001) (gfp1 is identicalwith gfpmut3* (Andersen et al. 1998)) was clonedinto respective restriction sites of vector pME6012(Heeb et al. 2000), resulting in pMEG65. The
Plant Soil

expression of gfp1 gene was directed from PA1/04/03promoter, a Plac derivative (Lanzer and Bujard 1988).Plasmid pJBA130 carrying luxR-PluxI-RBSII-gfpmut3* cassette with AHL-induced promoter ofluxI was isolated from E. coli JB534-MT102. Bothplasmids were introduced into S. meliloti strains byelectroporation as described previously (Wielbo et al.2007). The stability of pJBA130 and pMEG65 in S.meliloti strains under nonselective conditions wasassessed as described previously (Wielbo and Skorupska2001). No segregation of plasmid-free cells wasobserved.
Amplification of QS genes
Four sets of primers were used for amplification oftraI, sinR, sinI, and expR genes (oligonucleotidesequences by J.E. González, personal communication).
For traI amplification primers traI forward 5′-CGCGATCTCGAAGCCCCGGAATG-3′ and traI re-verse 5′-CAGATCGGTAGTTGGAAGGGCG-3′ wereused, and PCR condition were as follows: 95°C for5 min, 30 cycles of 95°C for 30 s, 63°C for 30 s, 72°Cfor 1 min and a final extension 72°C for 5 min.
For sinR amplification primers sinR forward: 5′-ATGGCTAATCAACAGGCTGTCC-3′ and sinR re-verse 5′-GATGGTGGGGATCAGAGCATGTCG-3′were used, and PCR condition were as follows:95°C for 5 min, 30 cycles of 95°C for 30 s, 55°C for30 s, 72°C for 1 min and a final extension 72°C for5 min.
For sinI amplification primers sinI forward 5′-TCGCTACATCGCGTAATCACGC-3′ and sinI re-verse 5′-GATCCTCAGGCGGCGCGTGC CGTTT-3′were used, and PCR condition were as follows: 95°Cfor 5 min, 30 cycles of 95°C for 30 s, 57°C for 30 s,72°C for 1 min and a final extension 72°C for 5 min.
For expR amplification primers expR forward 5′-TCACATGGCCTGGCGGGTTG-3′ and expR reverse5′-CGAATCCGGTCGAGGCGCTG-3′ were used, andPCR condition were as follows: 95°C for 5 min, 30cycles of 95°C for 30 s, 57°C for 30 s, 72°C for 1 minand a final extension 72°C for 5 min.
All PCR reactions were done with DreamTaq™Green PCR Master Mix, Fermentas. As a reference, S.meliloti Rm41 (sinR+, sinI+, expR+, traI+) and S.meliloti Rm8530 (sinR+, sinI+, expR+, traI−) wereused.
Table 1 Bacterial strains and plasmids used in this study
Strain Relevant characteristic Source or reference
Sinorhizobium meliloti Wild type alfalfa (Medicago media) nodules isolates Marek-Kozaczuket al. submittedLU02, LU04, LU05, LU06, LU07, LU08,
LU09, LU10, LU11, LU12, LU15, LU17
S. meliloti Rm41 Wild type, producing short-chain AHLs (traI+)and long-chain AHLs (sinI+/sinR+/expR+)
Llamas et al. 2004
S. meliloti Rm8530 Rm1021 expR+ producing long-chain AHLs(sinI+/sinR+/expR+)
Pellock et al. 2002
Chromobacterium violaceum CV026 Kmr, double mini-Tn5 mutant of C. violaceum ATCC31532 McClean et al. 1997
Agrobacterium tumefaciens NT1 tra reporter carrying pDCI41E33 with traG::lacZfusion and traR cloned into pDSK519
Shaw et al. 1997
E. coli JB534-MT102 AHLs reporter harboring pJBA130 Andersen et al. 2001
E. coli DH5α supE44 ΔlacU169 (ϕj80 lacZD M15) hsdR17recA1endA1 Sambrook et al. 1989
E. coli AKN65 gyrA96 thi-1 relA1 Koch et al. 2001Host of pBK-miniTn7-PA1/04/03-RBSII-gfp1-T0–T1
Plasmids
pJBA21Tc Tcr; pJB321 broad-host range vector with parDEand constitutive gusA
Wielbo and Skorupska 2001
pJBA130 Tcr; pME6031-luxR-PluxI-RBSII-gfpmut3*-T0–T1 Andersen et al. 2001
pME6012 Tcr; pVS1-p15A shuttle vector Heeb et al. 2000
pBK-miniTn7-gfp1 Kmr Cmr; pBK-miniTn7-PA1/04/03-RBSII-gfp1-T0–T1 Koch et al. 2001
pMEG65 Tcr; pME6012 with 1 kb EcoRI-HindIII fragmentcontaining gfp originated from pAKN65
This work
Plant Soil

S. meliloti AHLs isolation and detection
AHLs were isolated from 5 ml overnight cultures of S.meliloti by ethyl acetate supernatants extraction (Cha etal. 1998) The samples were separated by thin-layerchromatography on C18 reversed-phase TLC plates(Merck), and overlaid with Chromobacterium viola-ceum CV026 (Lithgow et al. 2001). Synthetic AHLstandards (Sigma-Aldrich) such as N-butyryl-DL-homoserine lactone (C4-AHL), N-hexanoyl-DL-homo-serine lactone (C6-AHL), N-octanoyl-DL-homoserinelactone (C8-AHL) and N-decanoyl-DL-homoserinelactone (C10-AHL) (solutions of 1 mg ml−1 in ethylacetate) were used as markers, and S. meliloti Rm41(short- and long-chain AHLs producer) and S. melilotiRm8530 (long-chain AHLs producer) were used asreference strains.
Plant growth and microscopy
Alfalfa seeds (Medicago media Pers. cv. Radius) weresurface-sterilized, germinated and grown on nitrogen-free Fahråeus medium slants (Vincent 1970). 5-day-old alfalfa seedlings were inoculated with 0.2 ml ofcell suspension at approximate density of 1.0×109
cells/ml. Alfalfa (one plant per tube) was grown in agreenhouse under natural light supplemented withartificial light (14 h day/10 h night, at 24°/19°C). Atspecified times after inoculation, the plants were takenfrom agar slants and microscopic observations wereperformed using a light microscope. Analyses offluorescence and image acquisition were made usingOlympus BX41 microscope equipped with OlympusUSH-103OL mercury lamp and filters U-MWBV2and U-MWIB3. For the observation of fluorescence inroots or nodules younger than 2 weeks, uncut samplesof whole roots were scrutinized under microscope,while 3 to 10 week-old nodules were longitudinally
hand sectioned with a razor blade and only thenanalyzed.
For detection of β-glucuronidase activity, alfalfaroot nodules were stained for β-glucuronidase activityas described earlier (Wielbo et al. 2007).
Results
AHLs production by Sinorhizobium meliloti strains
Twelve Sinorhizobium meliloti strains (Table 1) iso-lated from alfalfa nodules were tested for AHLsproduction using three different bioindicators. All ofthe selected strains produced AHLs detectable by A.tumefaciens NT1. This biosensor detects a broadrange of AHLs, i.e., the 3-oxoacyl-HSLs with sidechains ranging from C4 to C12 and unsubstituted 3-acyl-HSLs with side chains of 6 to 12 carbons (Shawet al. 1997). Two strains, LU09 and LU11, addition-ally produced AHLs which induced production ofpurple pigment in C. violaceum CV026 and triggeredgreen fluorescence of E.coli JB534-MT102 carryingpJBA130 (Table 2). C. violaceum CV026 biosensorcan detect AHLs with relatively short acyl chains, i.e.C6-HSL, C -HSL and C8-HSL but it does not detectC-HSL with longer acyl chains (McClean et al. 1997;Lithgow et al. 2001). The pJBA130 sensor plasmidexhibits the highest sensitivity for 3-oxo-C6-HSL butit is also responsive to various related AHLs albeitwith lower sensitivity (Andersen et al. 1998).
To determine the range of AHLs produced by S.meliloti isolates in comparison to S. meliloti referencestrains harboring different QS systems with use of C.violaceum CV026 and E.coli JB534-MT102 asindicators, several tests were performed. Using syn-thetic AHLs such as C4-HSL, C6-HSL, C8-HSL andC10-HSL, comparable sensitivity of C. violaceum
Table 2 Production of AHLs in S. meliloti isolates detected by biosensors
Strains AHL biosensor reaction
E. coli JB534-MT102 Chromobacteriumviolaceum CV026
Agrobacteriumtumefaciens NT1
LU02, LU04, LU05, LU06, LU07, LU08, LU09, LU10,LU11, LU12, LU15, LU17
– − +++
LU09, LU11 +++ +++ +++
Plant Soil
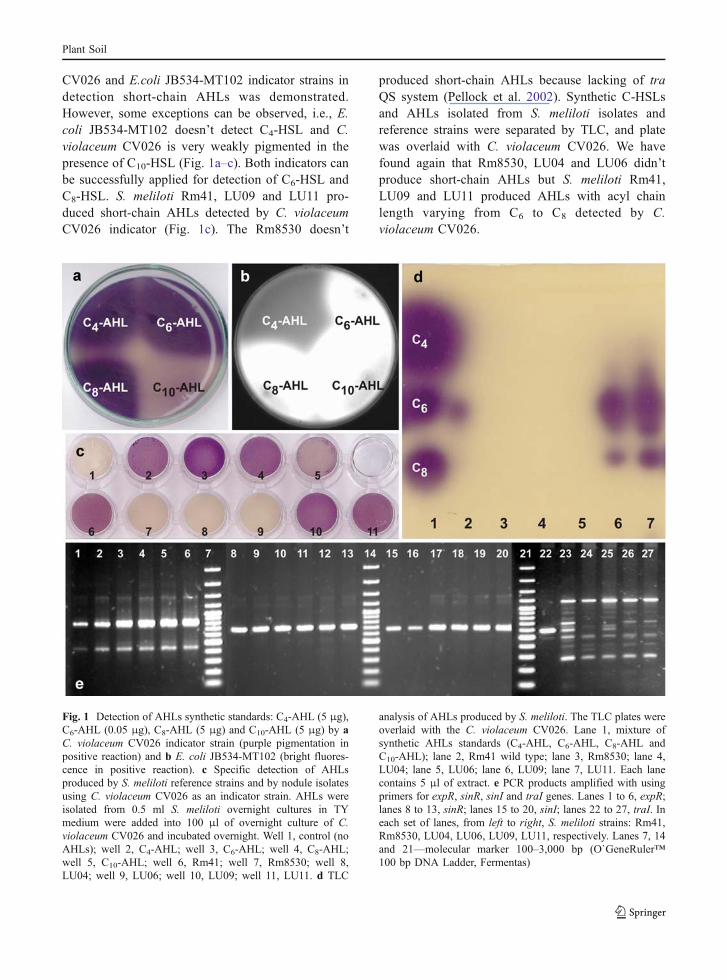
CV026 and E.coli JB534-MT102 indicator strains indetection short-chain AHLs was demonstrated.However, some exceptions can be observed, i.e., E.coli JB534-MT102 doesn’t detect C4-HSL and C.violaceum CV026 is very weakly pigmented in thepresence of C10-HSL (Fig. 1a–c). Both indicators canbe successfully applied for detection of C6-HSL andC8-HSL. S. meliloti Rm41, LU09 and LU11 pro-duced short-chain AHLs detected by C. violaceumCV026 indicator (Fig. 1c). The Rm8530 doesn’t
produced short-chain AHLs because lacking of traQS system (Pellock et al. 2002). Synthetic C-HSLsand AHLs isolated from S. meliloti isolates andreference strains were separated by TLC, and platewas overlaid with C. violaceum CV026. We havefound again that Rm8530, LU04 and LU06 didn’tproduce short-chain AHLs but S. meliloti Rm41,LU09 and LU11 produced AHLs with acyl chainlength varying from C6 to C8 detected by C.violaceum CV026.
Fig. 1 Detection of AHLs synthetic standards: C4-AHL (5 μg),C6-AHL (0.05 μg), C8-AHL (5 μg) and C10-AHL (5 μg) by aC. violaceum CV026 indicator strain (purple pigmentation inpositive reaction) and b E. coli JB534-MT102 (bright fluores-cence in positive reaction). c Specific detection of AHLsproduced by S. meliloti reference strains and by nodule isolatesusing C. violaceum CV026 as an indicator strain. AHLs wereisolated from 0.5 ml S. meliloti overnight cultures in TYmedium were added into 100 μl of overnight culture of C.violaceum CV026 and incubated overnight. Well 1, control (noAHLs); well 2, C4-AHL; well 3, C6-AHL; well 4, C8-AHL;well 5, C10-AHL; well 6, Rm41; well 7, Rm8530; well 8,LU04; well 9, LU06; well 10, LU09; well 11, LU11. d TLC
analysis of AHLs produced by S. meliloti. The TLC plates wereoverlaid with the C. violaceum CV026. Lane 1, mixture ofsynthetic AHLs standards (C4-AHL, C6-AHL, C8-AHL andC10-AHL); lane 2, Rm41 wild type; lane 3, Rm8530; lane 4,LU04; lane 5, LU06; lane 6, LU09; lane 7, LU11. Each lanecontains 5 μl of extract. e PCR products amplified with usingprimers for expR, sinR, sinI and traI genes. Lanes 1 to 6, expR;lanes 8 to 13, sinR; lanes 15 to 20, sinI; lanes 22 to 27, traI. Ineach set of lanes, from left to right, S. meliloti strains: Rm41,Rm8530, LU04, LU06, LU09, LU11, respectively. Lanes 7, 14and 21—molecular marker 100–3,000 bp (O`GeneRuler™100 bp DNA Ladder, Fermentas)
Plant Soil

Finally, the key genes in particular QS systemssuch as sinR, sinI, expR and traI in S. meliloti LU04,LU06, LU09 and LU11 strains were PCR amplifiedwith the specific primers using Rm8530 (sinI+/sinR+/expR+/traI−) and Rm41 (sinI+/sinR+/expR+/traI+) as areference strains (Fig. 1e). We found that S. melilotiLU04, LU06, LU09 and LU11 strains have sinI, sinRand expR genes like Rm8530 and Rm41. Surprisingly,in spite of short-chain AHL production by Rm41 and S.meliloti LU09 and LU11 strains, specific amplicon fortraI gene was obtained only in the case of Rm41 strain.
To study the AHL production in parallel to theprogression of S. meliloti symbiosis with alfalfa, thesensor plasmid pJBA130 carrying transcriptionalfusion of PluxI-gfpmut3* and luxR gene on the broadhost range plasmid pME6031 was used (Andersen etal. 1998; Andersen et al. 2001). It should be notedthat for both, monitoring the rhizobial AHL produc-tion inside the nodules and for constitutive Gfpproduction, an extremely stable variant of Gfp, namedGfpmut3* with significantly enhanced fluorescencewas used (Andersen et al. 1998).
Simultaneously with monitoring ongoing AHLproduction using pJBA130, the progress the alfalfainfection and nodule development was assessed usingthe plasmid pMEG65, which is based on the samereplication system as pJBA30 but contains a consti-tutively expressed gfp gene. Plasmids pJBA130 andpMEG65 were introduced into S. meliloti LU09 andLU11 strains. Both strains harboring these plasmidsexhibited bright-green fluorescence when werecultured on 79CA and TY agar media, suggestingthat the AHLs are produced by the bacterial cells(Fig. 3m, n). The introduction of pJBA130 andpMEG65 into LU09 and LU11 strains did not changethe growth rate and ability of these strains to secreteAHLs (data not shown).
AHL production by S. meliloti during symbiosis
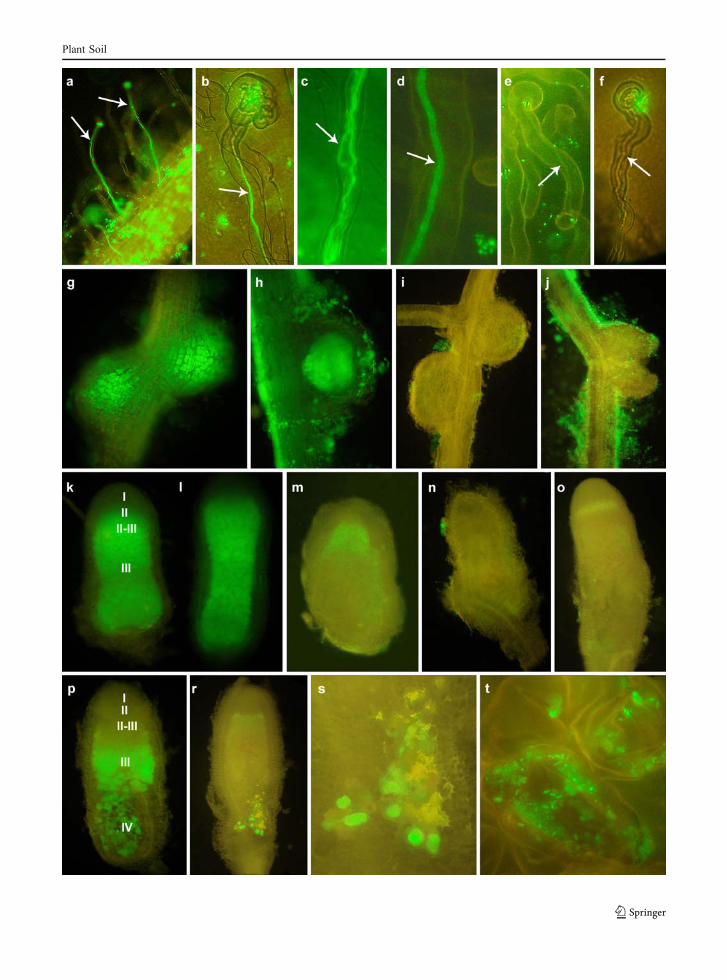
Alfalfa seedlings were inoculated with LU09 andLU11 harboring pJBA130 or pMEG65 in parallel.Samples of plant roots and nodules were taken atvarious time intervals post inoculation and greenfluorescence was monitored. After 3 days, gfpexpression was observed on the surface of root hairsof plants inoculated with bacteria carrying pMEG65(Fig. 2a–d) as well as pJBA130 (Fig. 2e, f). The firstdifference in expression of the plasmids encoded gfp
gene was noticed in bacteria filling infection threads.Specifically, LU09 and LU11 carrying pMEG65 withconstitutively expressed Gfp were clearly visibleinside the infection threads (Fig. 2a–d), whereas thesame strains carrying pJBA130 with gfp under thecontrol of AHL-dependent promoter exhibited signif-icantly less intensive fluorescence (Fig. 2e). More-over, several non-fluorescent infection threads (Fig. 2f)were seen in plants inoculated with rhizobia carryingpJBA130. The differences in gfp expression intensifiedin the course of symbiosis. Strains carrying pMEG65were visible in 10-day-old nodules (Fig. 2g, h),whereas strains carrying pJBA130 were not visible inany of the nodules examined, despite of gfp-expressingbacteria on root surface (Fig. 2i, j). We concluded that,beginning with the early step of nodule development,the synthesis of the rhizobial AHLs detected bypJBA130 has either been stopped or the AHLs weresynthesized at concentrations below the threshold ofbioindicator sensitivity i.e., 5 nM N-acyl homoserinelactone (Andersen et al. 2001).
The developing 3–5 week-old nodules elicited byLU09 and LU11 pMEG65 tagged strains showedbright-green fluorescence (Fig. 2k, l) in all develop-mental zones. On the other hand, no gfp expressionwas recorded in nodules elicited by strains taggedwith pJBA130 (Fig. 2n, o). As an exception, weakfluorescence was observable in the infection zone ofnot numerous 4 week-old nodules (Fig. 2m). Thedegree of nodule development was similar regardlessof the inoculating strain.
Fig. 2 Fluorescent microscopy evaluation of different stages ofalfalfa symbiosis with Sinorhizobium meliloti strains LU09(pMEG65) with constitutively expressed gfp gene and LU09(pJBA130) carrying AHLs-dependent promoter fused with gfpgene. Root hairs with infection threads developed in alfafainoculated with LU09(pMEG65) (a–d) and LU09(pJBA130)(e, f). Bright-green fluorescence of 10 day-old nodules inducedby LU09(pMEG65) (g, h) and lack of fluorescence inside thenodules induced by LU09(pJBA130) (i, j); 5 week-old noduleselicited by LU09(pMEG65) (k, l) in which developmentalzones with fluorescent bacteroids are seen and the nodulesinfected by LU09(pJBA130) that exhibited a weak fluorescencein the infection zone (m) or did not exhibit any fluorescence (n,o). 7 week-old nodules occupied by LU09(pMEG65) (p) andLU09(pJBA130) (r) with green fluorescence in senescencezone IV. Extended fragment of senescence zone IV of noduleoccupied by LU09(pJBA130) with plant cells packed withgreen fluorescent vegetative bacteria (s) and single plant cellswith rhizobia (t). Nodule zones: I—meristem, II—infectionzone, II–III—interzone, III—nitrogen fixation zone, IV—senescent zone. Thick arrow—infection thread
b
Plant Soil
Plant Soil

Bright fluorescence of pMEG65-carrying strains wasstill maintained in 7-week-old nodules in which thesenescence zone IV has already been developed(Fig. 2p). Surprisingly, green fluorescence appearedagain in zone V of 7 week-old-nodules induced byLU09 and LU11 each harboring pJBA130 (Fig. 2r)showing AHL-mediated gfp expression in this zone.Also, the plant cells were filled with rhizobia resem-bling in shape and size vegetative forms and there wereno visible bacteroids in vicinity of these brightlyfluorescent cells (Fig. 2s, t). In some instances,autofluorescence of thickened plant cell walls (probablydue to accumulating phenolic compounds) wasobserved (Fig. 2r, s).
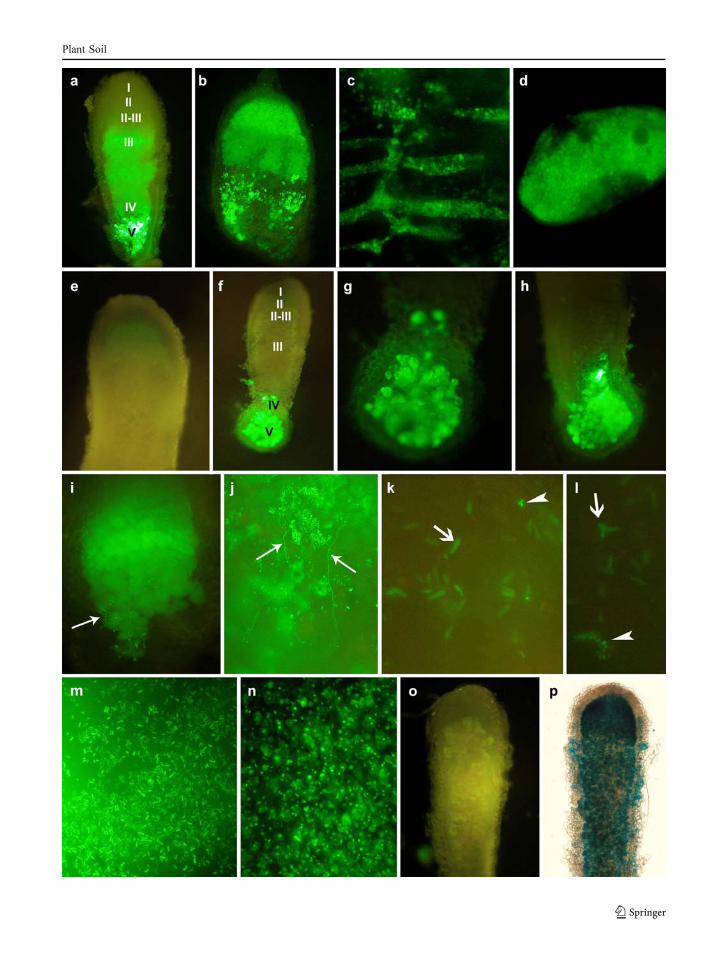
Prolonged growth of nodules up to 10 weeksresulted in expansion of the saprophytic zone V(Fig. 3a, b) where plant cells and intracellular spaceswere filled with fluorescent vegetative forms ofrhizobia tagged with pMEG65, and the bright fluores-cence in infection and symbiotic zones was maintained(Fig. 3c, d). One could also observe fluorescentinfection threads on the surface of 4–5 week-oldnodules (Fig. 3i, j). A considerable difference betweenthe fluorescence of bacteroids and vegetative forms ofrhizobia harboring pMEG65 could be seen in the distalnitrogen fixing zone, i.e., the smaller and brightervegetative cells and the pleomorphic bacteroids withless intense fluorescence (Fig. 3k, l).
Contrary to the above, 10-week-old nodulesoccupied by strain harboring pJBA130 (Fig. 3e–h)did not exhibit green fluorescence outside the sapro-phytic zone V, nevertheless few weakly fluorescentnodules could be observed in the infection zone II(Fig. 3e). In majority of nodules, the saprophytic zoneV was extended, and proliferation of brightly fluores-cent vegetative forms of rhizobia was observed(Fig. 3f–h).
To further investigate whether the lack of gfpexpression in bacteroids inside the 2–5 week-oldnodules resulted from the AHL shortage and not fromthe absence of bacteroids in the nodules, alfalfa plantswere coinoculated with a pair of isogenic strains—LU09 carrying pJBA130, and LU09 with pJBA21Tcstably expressing gusA, in 1:1 ratio. In 5 week-old-nodules, no green fluorescence was detected, signify-ing no gfp expression directed from luxI promoter(Fig. 3o), while ß-glucuronidase activity was present,as was evidenced by histochemical staining of thesecond half of the same nodule (Fig. 3p). Moreover,
mixed population of Gfp+Gus− and Gfp−Gus+ rhizo-bia were recovered from these nodules, indicating thatLU09(pJBA130) bacteria were indeed present in thenodules, but they were not observable due to lack ofthe AHL inducers of gfp expression. Because LU09and LU11 strains behaved identically in the experi-ments performed in planta, the symbiosis of just theLU09 strain with alfalfa has been shown.
Taken together, we conclude that the bacteroids ofS. meliloti LU09 and LU11 did not synthesize short-chain AHL autoinducers at the level detectable byused pJBA30 sensor plasmid, contrary to the vegeta-tive forms of bacteria located in the saprophytic zoneV of nodules, which produced the autoinducers in ahighly efficient manner.
Discussion
QS systems have been described in several species ofrhizobia and it has been shown that QS plays anessential role in nitrogen-fixing rhizobia during theestablishment of interaction with the host plant (for areview see González and Marketon 2003). Plant roothair infection and subsequent nodule formationrequires the clustering of rhizobia around roots andit is possible that QS plays a role in different stages ofsymbiosis. It was also reported that bacterial AHLsmight influence the expression of several plant genes(Mathesius et al. 2003).
Fig. 3 Fluorescent microscopy evaluation of the late stages ofalfalfa symbiosis with Sinorhizobium meliloti strains LU09(pMEG65) and LU09(pJBA130) (a–j). Bright-green fluores-cence of 10 week-old nodules induced by LU09(pMEG65) (a,b), bacterial cells in the intracellular spaces of nodule (c) and asingle plant cell fulfilled with rhizobia (d). Very weakfluorescence in the infection zone of 10 week-old nodule (e)and the expanded saprophytic zone V occupied by fluorescentLU09(pJBA130) bacteria (f–h). Brightly fluorescing vegetativebacteria and weakly fluorescing bacteroids observed in sapro-phitic zone V of 10 week-old nodules occupied by LU09(pMEG65) (i, j). Vegetative cells and pleomorphic bacteroids innodules occupied by LU09(pMEG65) (k, l). LU09(pMEG65)and LU09(pJBA130) strains grown on liquid TY medium (m,n). 7 week-old nodules coinfected with LU09(pJBA130) andLU09(pJBA21Tc) (o, p). Lack of fluorescence of LU09(pJBA130) (o) and β-glucuronidase activity of LU09(pJBA21Tc) harboring gusA gene (p). Nodule zones: I—meristem, II—infection zone, II–III—interzone, III—nitrogenfixation zone, IV—senescent zone, V—saprophytic zone. Thickarrow—infection thread, thin arrow—bacteroid, arrowhead—vegetative cell
b
Plant Soil
Plant Soil

Knowledge of AHL production in bacteroids andthe role of QS in nodule development remainfragmentary. Daniels et al. (2002) have observed cinIand cinR expression and the production of at leastthree different AHLs under symbiotic conditions in R.etli bacteroids. However, the AHLs produced undersymbiotic conditions were less diverse than underfree-living growth. Recently, Gurich and González(2009) in the whole-genome expression analysisdemonstrated that the ExpR/Sin quorum-sensingsystem responsible for long-chain AHLs productionis repressed after Medicago sativa invasion by S.meliloti.
Among S. meliloti isolates studied here, weidentified two strains that produced AHLs detectedby several indicator strains what suggested that thesestrains synthesized diverse classes of autoinducers. Infact, the LU09 and LU11 produced three differentshort-chain AHLs that was evidenced by TLCanalysis. However, besides the short-chain AHLsproduction, traI gene was not identified in thesestrains using primers and PCR procedure described inMaterials and Methods. Therefore we can speculate,that the LU09 and LU11 have other not yet identifiedQS system responsible for these AHLs synthesis ortheir traI genes have sequence diversity.
In this work, we made an attempt to study theAHLs production in situ, in developing nodulesduring symbiosis of S. meliloti with alfalfa usingpJBA130 sensor plasmid, in which a promoterless gfpgene was cloned under luxI promoter activated by abroad range of AHLs (Andersen et al. 2001). As acontrol, we used plasmid pMEG65 with constitutivelyexpressed gfp gene. The pJBA130 reporter was usedin our study because of its extreme stability in cells inthe absence of antibiotic selection and the possibilityto observe in situ green protein fluorescence insamples of roots and nodules. S. meliloti LU09 andLU11 strains were selected from among strains of ourcollection of alfalfa nodule isolates. These strainswere identified as S. meliloti by 16S rDNA sequenc-ing (Marek-Kozaczuk et al. submitted). Both strainsproduced diverse AHLs with short and long acylchains as determined using three different biosensors.
The most interesting outcome of our experimentswas that only vegetative forms efficiently producedAHLs during infection and nodule development,and bright-green fluorescence could be observed onthe surface of roots and in the saprophytic zone V
of nodules. In the bacteroids, AHL synthesis wasnot detected with the chosen reporter system. Weakgreen fluorescence was observed in the infectionthreads that suggested progressive quenching theproduction of AHLs. We did not observe fluores-cence in the nitrogen-fixing zone of any nodulesstudied in this work. Nonetheless, because thepJBA130 biosensor does not detect every kind ofAHLs (Andersen et al. 2001), it is possible that somelong-acyl chain AHL synthesis occurs in bacteroidsduring nodule development.
These observations are in accordance with resultsof Daniels et al. (2002) who observed differences inprofiles of AHLs isolated from free-living cellscompared to those isolated from R. etli bacteroids.Moreover, they observed a 10-fold lower expressionof luxI-gfp fusion inside the nodules compared tofree-living conditions, which suggested the changes inthe QS system regulation.
Vegetative forms of bacteria producing AHLslocated in the infection threads are in significantminority in relation to bacteroids that are packedinside nodule cells. This could be one possibleexplanation for the lack of green fluorescence in thedeveloping nodules. We observed vegetative cells inthe vicinity of bacteroids only in the case of plantsinoculated with LU09(pMGE65) (constitutive gfp). Inthe saprophytic zone Vof nodules, vegetative rhizobiaintensively grow, produce AHLs at a high level and asa result of high cell density, bright-green fluorescenceis observed. At this stage of symbiosis, bacteria are“preserved” in the safety compartment of the sapro-phytic zone V where these can proliferate intensivelyin an environment abundant in nutrients delivered bydegenerating plant cells and devoid of other bacteriathat compete with rhizobia. Finally, these rhizobia arereleased into the soil. According to Timmers et al.(2000) saprophytic zone V represents an ecologicalniche where rhizobia take advantage of the interactionand escape from plant control. The significance andmolecular mechanism of inhibition of AHLs produc-tion in bacteroids during nodule development remainsto be elucidated in further experiments, but one mayhypothesize that production of AHLs by bacteroidsthat intensively fix nitrogen is not metabolicallyjustified and, as such, is not likely to be beneficialat this stage of symbiosis. It is reasonable to supposethat nitrogen fixation and quorum sensing pathwayscompete for energy sources inside the nodules.
Plant Soil

Acknowledgement We greatly acknowledge the gift ofplasmids and strains from D. Haas (Université de Lausanne,Lausanne, Switzerland), J.B. Andersen and L. Lambertsen (TheTechnical University of Denmark, Lyngby, Denmark), and S.Jafra (University of Gdańsk, Gdańsk, Poland). Special acknowl-edgements are given for Prof. J.E. González (University of Texas atDallas, Richardson, US) for the bacterial strains, oligonucleotidesand a very helpful discussion. This work was partially supported bya grant from the Ministry of Education and Science no. N N304026734.
References
Andersen JB, Sternberg C, Poulsen LK, Bjorn SP, Givskov M,Molin S (1998) New unstable variants of green fluorescentprotein for studies of transient gene expression in bacteria.Appl Environ Microbiol 64:2240–2246
Andersen JB, Heydorn A, Hentzer M, Eberl L, Geisenberger O,Christensen BB, Molin S, Givskov M (2001) gfp-based N-acyl homoserine-lactone sensor system for detection ofbacterial communication. Appl Environ Microbiol 67:575–585
Cha C, Gao P, Chen YC, Shaw PD, Farrand SK (1998)Production of acyl-homoserine lactone quorum-sensingsignals by Gram-negative plant-associated bacteria. MolPlant-Microbe Interact 11:1119–1129
Daniels R, De Vos DE, Desair J, Raedschelders G, Luyten E,Rosemeyer V, Verreth C, Schoeters E, Vanderleyden J,Michiels J (2002) The cin quorum sensing locus ofRhizobium etli CNPAF512 affects growth and symbioticnitrogen fixation. J Biol Chem 277:462–468
Daniels R, Vanderleyden Y, Michiels J (2004) Quorum sensingand swarming migration of bacteria. FEMS Microbiol Rev28:261–289
González JE, Keshavan ND (2006) Messing with bacterialquorum sensing. Microbiol Mol Biol Rev 70:859–875
González JE, Marketon MM (2003) Quorum sensing innitrogen-fixing rhizobia. Microbiol Mol Biol Rev 67:574–592
Gurich N, González JE (2009) Role of quorum sensing inSinorhizobium meliloti-alfalfa symbiosis. J Bacteriol191:4372–4382
Heeb S, Itoh Y, Nishijyo T, Schnider U, Keel Ch, Wade J, WalshU, O’Gara F, Haas D (2000) Small, stable shuttle vectorsbased on minimal pVS1 replicon for use in Gram-negativeplant-associated bacteria. Mol Plant-Microbe Interact13:232–237
Jafra S, van der Wolf JM (2004) Fast screening method fordetection of acyl-HSL-degrading soil isolates. J MicrobiolMeth 57:415–420
Jones KM, Kobayashi H, Davies BW, Taga ME, Walker GC(2007) How symbionts invade plants: the Sinorhizobium-Medicago model. Nat Rev Microbiol 5:619–633
Koch B, Jensen LE, Nybroe O (2001) A panel of Tn7-based vectors for insertion of the gfp marker gene orfor delivery of cloned DNA into Gram-negative bacteriaat a neutral chromosomal site. J Microbiol Meth 45:187–195
Lanzer M, Bujard H (1988) Promoters largely determine theefficiency of repressor action. Proc Natl Acad Sci USA85:8973–8977
Lithgow JK, Danino VE, Jones J, Downie JA (2001) Analysisof N-acyl homoserine-lactone quorum-sensing moleculesmade by different strains and biovars of Rhizobiumleguminosarum containing different symbiotic plasmids.Plant Soil 232:3–12
Llamas I, Keshavan N, González JE (2004) Use of Sinorhizobiummeliloti as indicator for specific detection of long-chain N-acyl homoserine lactones. Appl Environ Microbiol70:3715–3723
Marketon MM, González JE (2002) Identification of twoquorum-sensing systems in Sinorhizobium meliloti. JBacteriol 184:3466–3475
Marketon MM, Gronquist MR, Eberhard A, González JE(2002) Characterization of the Sinorhizobium melilotisinR/sinI locus and the production of novel N-acylhomoserine-lactones. J Bacteriol 184:5686–5695
Marketon MM, Glenn SA, Eberhard A, González JE (2003)Quorum sensing controls exopolysaccharide production inSinorhizobium meliloti. J Bacteriol 185:325–331
Mathesius U, Mulders S, Gao M, Teplitski M, Caetano-AnollesG, Rolfe BG, Bauer W (2003) Extensive and specificresponses of a eukaryote to bacterial quorum-sensingsignals. Proc Natl Acad Sci USA 100:1444–1449
McClean KH, Winson MK, Fish L, Taylor A, Chhabra SR,Camara M, Daykin M, Lamb JH, Swift S, Bycroft BW,Stewart GSAB, Williams P (1997) Quorum sensing andChromobacterium violaceum: exploitation of violaceinproduction and inhibition for the detection of N-acylhomoserine lactones. Microbiology 143:3703–3711
Mc Dougald D, Rice SA, Kjelleberg S (2007) Bacterial quorumsensing and interference by naturally occurring biomimics.Anal Bioanal Chem 387:445–453
Pellock BJ, Teplitski M, Boinay RP, Bauer WD, Walker GC(2002) A LuxR homolog controls production of symbioti-cally active extracellular polysaccharide II by Sinorhizobiummeliloti. J Bacteriol 184:5067–5076
Perret X, Staehelin C, Spaink HP (2000) Molecular basis ofsymbiotic promiscuity. Microbiol Mol Biol Rev 64:180–201
Sambrook J, Fritsch EF, Maniatis T (1989) Molecularcloning: a laboratory manual. Cold Spring Harbor,New York
Shaw PD, Ping G, Daly SL, Cha C, Cronan JE, RinehardKL, Farrand SK (1997) Detecting and characterizing N-acyl-homoserine lactone signal molecules by thin-layerchromatography. Proc Natl Acad Sci USA 94:6036–6041
Teplitski M, Eberhard A, Gronquist MR, Gao M, Robinson JB,Bauer WD (2003) Chemical identification of N-acylhomoserine lactone quorum-sensing signals produced bySinorhizobium meliloti strains in defined medium. ArchMicrobiol 180:494–497
Timmers AC, Soupène E, Auriac MC, de Billy F, Vasse J,Boistard P, Truchet G (2000) Saprophytic intracellularrhizobia in alfalfa nodules. Mol Plant-Microbe Interact13:1204–1213
Vasse J, de Billy F, Camut S, Truchet G (1990) Correlationbetween ultrastructural differentiation of bacteroids and
Plant Soil

nitrogen fixation in alfalfa nodules. J Bacteriol 172:4295–4306
Vincent JM (1970) A manual for the practical study of theroot-nodule bacteria. Blackwell Scientific Publishers,Oxford
Wielbo J, Skorupska A (2001) Construction of improvedvectors and cassettes containing gusA and antibioticresistance genes for studies of transcriptional activity
and bacterial localization. J Microbiol Methods 45:197–205
Wielbo J, Marek-Kozaczuk M, Kubik-Komar A, Skorupska A(2007) Increased metabolic potential of Rhizobium spp. isassociated with bacterial competitiveness. Can J Microbiol53:957–967
Wisniewski-Dye F, Downie JA (2002) Quorum-sensing inRhizobium. Antonie van Leeuwenhoek 81:397–407
Plant Soil
![A Functional myoInositol Dehydrogenase Gene Is Required for Efficient Nitrogen Fixation and Competitiveness of Sinorhizobium fredii USDA191 To Nodulate Soybean (Glycine max [L.] Merr](https://static.fdokumen.com/doc/165x107/6312da2ab22baff5c40edaa4/a-functional-myoinositol-dehydrogenase-gene-is-required-for-efficient-nitrogen-fixation.jpg)

















