
ActP controls copper homeostasis in Rhizobium leguminosarum bv. viciae and Sinorhizobium meliloti...
11
© 2002 Blackwell Science Ltd ActP controls copper homeostasis in Rhizobium leguminosarum bv. viciae and Sinorhizobium meliloti preventing low pH-induced copper toxicity bium, Rhizobium, Sinorhizobium) fix atmospheric nitrogen in root nodules to a form that can be used for the growth of the legume host. Soil acidity has a detrimental impact on plant productivity by interfering in the infection process or by decreasing the survival of the microsymbiont (Munns, 1986; Howieson et al., 1988; Coventry and Evans, 1989; Recourt et al., 1989; Robson and Bottomley, 1991). However, it is the long-term persistence of the prokaryotic partner in acidic soils between pasture- growing seasons that governs the regeneration of productive pasture legumes. The response to acidity differs among the strains (see review by Barnet, 1991). Mesorhizobium and Bradyrhizobium are quite acid tolerant and grow readily at pH 4.5 or lower, whereas R. leguminosarum can grow at pH levels down to pH 5.0. Sinorhizobium meliloti is one of the most acid-sensitive root nodule bacteria, growing only at pH levels down to pH 5.5. An understanding of these differences is being investigated at the physiological and genetic level (Glenn et al., 1999). Rhizobial mutants with acid-sensitive phenotypes have been identified using Tn5 mutagenesis (Goss et al., 1990; Tiwari et al., 1992). Genes (designated act for ac id-t olerance) identified in S. meliloti in this manner include actR (the regulator module of a two- component sensor–regulator system; Tiwari et al., 1996a), actS (the sensor module; Tiwari et al., 1996a) and actA (a potential lipid-metabolizing protein; Tiwari et al., 1996b). It is interesting that each of these mutants displays a pleiotropic phenotype with sensitivity not only to acid but also to azide and/or certain heavy metals. The actR/S mutants are sensitive to acidity, azide, cadmium or zinc (Glenn et al., 1999), whereas a mutation in actA creates sensitivity to acidity, copper and zinc (Tiwari et al., 1996b). Omission of heavy metals from the medium does not alleviate the acid sensitivity of these mutants. However, acid tolerance may be restored to mutants defective in actR, actS or actA by supplementing the growth medium with high concentrations of calcium (Tiwari et al., 1992; Reeve et al., 1993). By using a different screening strat- egy, we isolated one Tn5-induced mutant of S. meliloti WSM419 (RT3-27) and one of R. leguminosarum bv. viciae WSM710 (WR1-14) that were acid sensitive even in the presence of high concentrations of calcium (10 mM Molecular Microbiology (2002) 43(4), 981–991 Wayne G. Reeve, 1 Ravi P. Tiwari, 1 Neetin B. Kale, 2 Michael J. Dilworth 1 * and Andrew R. Glenn 3 1 Centre for Rhizobium Studies, School of Biological Sciences and Biotechnology, Murdoch University, Murdoch, Perth, Australia 6150. 2 Department of Microbiology, University of Pune, Pune, India. 3 Office of the Pro Vice Chancellor (Research), University of Tasmania, Hobart, Tasmania, Australia 7001. Summary Two ‘calcium-irreparable’ acid-sensitive mutants were identified after mutagenizing Rhizobium legumi- nosarum bv. viciae and Sinorhizobium meliloti with Tn5. Each mutant contains a single copy of the trans- poson which, inserted within the actP gene, prevents expression of a P-type ATPase that belongs to the CPx heavy metal-transporting subfamily. Here, we show that both actP-knockout mutants show sensitivity to copper; omission of this heavy metal from low pH-buffered media restores acid tolerance to these strains. Furthermore, complementation of the mutant phenotype requires only the actP gene. An actP–gusA fusion in R. leguminosarum was transcriptionally regulated by copper in a pH-dependent manner. Downstream to actP in both organisms is the hmrR gene that encodes a h eavy m etal-r esponsive r egula- tor (HmrR) that belongs to the merR class of regula- tory genes. Insertional inactivation of hmrR abolished transcriptional activation of actP by copper ions and increased the basal level of its expression in their absence. These observations suggest that HmrR can regulate actP transcription positively and negatively. We show that copper homeostasis is an essential mechanism for the acid tolerance of these root nodule bacteria since it prevents this heavy metal from becoming overtly toxic in acidic conditions. Introduction The root nodule bacteria (Bradyrhizobium, Mesorhizo- Accepted 5 November, 2001. *For correspondence. E-mail [email protected]; Tel. (+61) 89360 2112; Fax (+61) 89360 6486.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of ActP controls copper homeostasis in Rhizobium leguminosarum bv. viciae and Sinorhizobium meliloti...

© 2002 Blackwell Science Ltd
ActP controls copper homeostasis in Rhizobiumleguminosarum bv. viciae and Sinorhizobium melilotipreventing low pH-induced copper toxicity
bium, Rhizobium, Sinorhizobium) fix atmospheric nitrogenin root nodules to a form that can be used for the growthof the legume host. Soil acidity has a detrimental impacton plant productivity by interfering in the infection processor by decreasing the survival of the microsymbiont(Munns, 1986; Howieson et al., 1988; Coventry andEvans, 1989; Recourt et al., 1989; Robson and Bottomley, 1991). However, it is the long-term persistenceof the prokaryotic partner in acidic soils between pasture-growing seasons that governs the regeneration of productive pasture legumes. The response to aciditydiffers among the strains (see review by Barnet, 1991).Mesorhizobium and Bradyrhizobium are quite acid tolerant and grow readily at pH 4.5 or lower, whereas R.leguminosarum can grow at pH levels down to pH 5.0.Sinorhizobium meliloti is one of the most acid-sensitiveroot nodule bacteria, growing only at pH levels down topH 5.5. An understanding of these differences is beinginvestigated at the physiological and genetic level (Glennet al., 1999).
Rhizobial mutants with acid-sensitive phenotypes have been identified using Tn5 mutagenesis (Goss et al.,1990; Tiwari et al., 1992). Genes (designated actfor acid-tolerance) identified in S. meliloti in this manner include actR (the regulator module of a two-component sensor–regulator system; Tiwari et al.,1996a), actS (the sensor module; Tiwari et al., 1996a) andactA (a potential lipid-metabolizing protein; Tiwari et al.,1996b).
It is interesting that each of these mutants displays apleiotropic phenotype with sensitivity not only to acid butalso to azide and/or certain heavy metals. The actR/Smutants are sensitive to acidity, azide, cadmium or zinc(Glenn et al., 1999), whereas a mutation in actA createssensitivity to acidity, copper and zinc (Tiwari et al., 1996b).Omission of heavy metals from the medium does not alleviate the acid sensitivity of these mutants. However,acid tolerance may be restored to mutants defective in actR, actS or actA by supplementing the growth mediumwith high concentrations of calcium (Tiwari et al., 1992;Reeve et al., 1993). By using a different screening strat-egy, we isolated one Tn5-induced mutant of S. melilotiWSM419 (RT3-27) and one of R. leguminosarum bv.viciae WSM710 (WR1-14) that were acid sensitive evenin the presence of high concentrations of calcium (10 mM
Molecular Microbiology (2002) 43(4), 981–991
Wayne G. Reeve,1 Ravi P. Tiwari,1 Neetin B. Kale,2
Michael J. Dilworth1* and Andrew R. Glenn3
1Centre for Rhizobium Studies, School of BiologicalSciences and Biotechnology, Murdoch University,Murdoch, Perth, Australia 6150.2Department of Microbiology, University of Pune, Pune,India.3Office of the Pro Vice Chancellor (Research), Universityof Tasmania, Hobart, Tasmania, Australia 7001.
Summary
Two ‘calcium-irreparable’ acid-sensitive mutants wereidentified after mutagenizing Rhizobium legumi-nosarum bv. viciae and Sinorhizobium meliloti withTn5. Each mutant contains a single copy of the trans-poson which, inserted within the actP gene, preventsexpression of a P-type ATPase that belongs to the CPxheavy metal-transporting subfamily. Here, we showthat both actP-knockout mutants show sensitivity to copper; omission of this heavy metal from low pH-buffered media restores acid tolerance to thesestrains. Furthermore, complementation of the mutantphenotype requires only the actP gene. An actP–gusAfusion in R. leguminosarum was transcriptionally regulated by copper in a pH-dependent manner.Downstream to actP in both organisms is the hmrRgene that encodes a heavy metal-responsive regula-tor (HmrR) that belongs to the merR class of regula-tory genes. Insertional inactivation of hmrR abolishedtranscriptional activation of actP by copper ions andincreased the basal level of its expression in theirabsence. These observations suggest that HmrR canregulate actP transcription positively and negatively.We show that copper homeostasis is an essentialmechanism for the acid tolerance of these root nodulebacteria since it prevents this heavy metal frombecoming overtly toxic in acidic conditions.
Introduction
The root nodule bacteria (Bradyrhizobium, Mesorhizo-
Accepted 5 November, 2001. *For correspondence. E-mail [email protected]; Tel. (+61) 89360 2112; Fax (+61)89360 6486.

or higher) in low-pH-buffered medium (Reeve et al.,1993). These mutants were therefore labelled ‘calcium-irreparable’ acid-sensitive mutants (Reeve et al., 1993).The growth rates of the mutants at pH 7.0 were similar tothose of the wild types, indicating that the acid sensitivitywas not due to any simple growth defect. Here, we show that a failure to produce a CPx P-type ATPaseresults in the calcium-irreparable acid-sensitive pheno-type. Furthermore, we demonstrate the importance of thisprotein for copper homeostasis and highlight how thechanges in copper availability resulting from pH changesmay affect gene expression and thereby influence acidtolerance. Finally, we demonstrate that transcriptional regulation of actP depends on the heavy metal responseregulator HmrR.
Results and discussion
The ‘calcium-irreparable’ acid-sensitive defects arecaused by Tn5
A single copy of Tn5 resides in both R. leguminosarumWR1-14 and S. meliloti RT3-27 that has been identifiedfrom the hybridization of a digoxigenin (DIG)-labelled Tn5 probe (pTn5-H) (data not shown) to a single fragmentof EcoRI restricted genomic DNA from each mutant.These Tn5-containing EcoRI fragments of 11 and 9 kbwere cloned from WR1-14 and RT3-27, respectively, andrestriction mapped (Fig. 1).
Verification that the single Tn5-induced lesion createdthe ‘calcium-irreparable’ acid-sensitive defect in eachmutant was established using two different methods. A
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 981–991
982 W. G. Reeve et al.
Fig. 1. Restriction maps of the Tn5-containingEcoRI fragments cloned from the mutants R.leguminosarum WR1-14 (A) and S. melilotiRT3-27 (B). The NotI site used for insertion ofthe CAS-GNm cassette (Reeve et al., 1999)to construct a R. leguminosarum actP–gusAfusion in the plasmid clone pWR668 has beenmarked. The site of transposon insertion hasbeen indicated by a Tn5-marked flag (103 bpfrom the end of actP in R. leguminosarumand 225 bp in S. meliloti). The direction ofaacC1 transcription in the CAS-Gm cassettein pWR901 and pWR903 is provided by thedirection of the arrow. The restriction map (C)of the actP gene region in S. melilotiWSM419 was derived from the plasmidpRT327-1. The S. meliloti actP gene wasinactivated in pNK57 by blunting andreligating an intragenic SphI site (representedby an asterisk), whereas the hmrR gene wasinactivated in pNK77 by cloning the CAS-GNm cassette into the intragenic BclI site.

Involvement of ActP and HmrR in copper homeostasis 983
RL38 phage lysate prepared from WR1-14 was used totransduce the kanamycin and streptomycin markers ofTn5 into WSM710; all transductants displayed acid sen-sitivity. A phage that can be used to transduce S. melilotiWSM419 has not yet been identified; 187 stocks of virulent and temperate phages failed to lyse this strain (C. Defives, personal communication). To reconstruct themutation in WSM419, marker-assisted exchange wasused to replace the wild-type allele with one containingTn5 using the plasmid pWR327-200. ReconstructedactP::Tn5 mutants displayed an acid-sensitivity profilesimilar to RT3-27.
The Tn5 insertions in RT3-27 or WR1-14 also causecopper sensitivity
Acid-sensitive mutants of S. meliloti display pleiotropicphenotypic defects, including sensitivities to certain heavymetals (Glenn et al., 1999). The sensitivities of WR1-14and RT3-27 to heavy metals were therefore comparedwith their respective wild types. Addition of CuSO4 to TYplates (at pH 7.0) at concentrations above 0.4 mM pre-vented growth of RT3-27 or WR1-14, whereas WSM419and WSM710 could tolerate 1 and 2 mM, respectively.The metal sensitivity is restricted to copper since themutants displayed the same tolerances on tryptone-yeastmedia (TY) plates as their respective wild types to AgNO3
(25 mM), CdCl2 (200 mM), CoCl2 (500 mM), HgCl2 (10 mM),Ni(NO3)2 (50 mM) or ZnSO4 (500 mM).
The relatively low concentration of copper (2 mM)present in the minimal medium was sufficient to terminategrowth of the actP-defective mutants only at low pH;neither R. leguminosarum WR1-14 nor S. meliloti RT3-27could grow at pH 5.5 or pH 5.7, respectively, unlesscopper was omitted from a minimal medium prepared withhigh-purity reagents. The latter finding is a contrast to theobservations that acid tolerance cannot be restored toactA, actS or actR mutants by removing heavy metals(Glenn et al., 1999). These data indicate that the disrup-tion of genes in RT3-27 and WR1-14 prevents the expres-sion of a copper-export system. Here, we present animportant insight into metal toxicity as the availability of a heavy metal may also change in response to pH-dependent binding.
The Tn5 lesion in RT3-27 or WR1-14 does not impairsymbiotic performance
The mutants RT3-27 and WR1-14 produced normalnodules on Medicago murex and Pisum sativum L.(Wirrega pea) respectively. In contrast, no nodules werefound on uninoculated seedlings. Retention of Tn5in nodule occupants was established by reisolatingkanamycin-resistant clones. The nodules of inoculated
plants were pink–red in colour and plants were healthyand green (in contrast to yellow–light green stunteduninoculated controls), indicative that the mutants werefixing nitrogen. The conditions we have used included acopper supplement of 120 mM in the nutrient solution;under these conditions, the gene that is mutated does not appear to be required for the successful establishmentof a normal nitrogen-fixing nodule. It should be noted,however, that the copper concentration in nodulesincreases with increasing copper application (Snowballand Robson, 1980). The induction of the conserved FixI cation transporter in bacteroids (Kahn et al., 1989)appears to be involved in the uptake and metabolism ofcopper for the proper functioning of the symbioticallyessential cbb3 oxidase (Preisig et al., 1996). It couldtherefore be envisaged that if excessive copper importoccurred via FixI or another transporter a counterbalanc-ing export mechanism might be required. If copper readilycrosses the peribacteroid membrane, this situation couldpotentially be worsened because enveloped bacteroidscould be exposed to enhanced copper toxicity in theapparently acidic environment of the symbiosome (Mellor,1989; Parniske, 2000). In view of the normal nodulationbehaviour of the mutants, this appears to be unlikely.
Sequence and analysis
Full sequences for 5.1 and 3.2 kb DNA regions surround-ing the Tn5 insertions in the mutants WR1-14 and RT3-27, respectively, showed that the disrupted genes codefor cation-transporting ATPases; they have therefore beendesignated actP (acid-tolerance P-type ATPase). The Tn5insertions are located 103 and 225 bp from the 3¢ end ofthe actP open reading frame (ORF) in R. leguminosarumand S. meliloti respectively; the sequences immediatelysurrounding the insertions are not similar. These Tn5insertions, which are in the C-terminal tenth of the ActPprotein in both cases, apparently result in complete inac-tivation of the protein, possibly suggesting that (if thealtered protein is actually produced) the C-terminaldomain is functionally or structurally important. Severalother genes were identified: hmrR occurred downstreamto actP in both organisms while an additional two partialORFs (treY and ecfR) were located in the R. legumi-nosarum strain sequence (Fig. 1A).
ActP is a P-type ATPase that belongs to the CPx family.The sizes of the ActP proteins deduced from R. legumi-nosarum and S. meliloti DNA sequences are 88 and 86 kDa respectively. The pI of S. meliloti ActP is 6.42whereas that for R. leguminosarum is 5.72. Both proteinsare very similar over their entire lengths: the two ActPproteins show 49% identity (and 65% similarity) over 664amino acids.
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 981–991

A BLASTP search of the databases using the R. legu-minosarum and S. meliloti ActP proteins revealed consid-erable similarity to putative cation-transporting P-typeATPases from numerous organisms. The matches to theR. leguminosarum ActP protein include a putative silverion transporter (SilP; AF067954) from Salmonellatyphimurium (53% identity over 808 amino acids), a puta-tive copper ion transporter (PacS; D16747) from Syne-chococcus spp. (49% identity over 656 amino acids) anda heavy metal ion transporter (U42410) from Proteusmirabilis (47% identity over 671 amino acids). Thematches to the S. meliloti ActP protein include the heavymetal ion-transporting P-type ATPase (U42410) from P.mirabilis (66% identity over 806 amino acids), a heavymetal ion-transporting ATPase (Z99121) from Bacillussubtilis (45% identity over 809 amino acids) and thecopper ion-transporting ATPase (PacS; D16747) fromSynechococcus spp. (47% identity over 756 amino acids).These proteins have been placed into the CPx subfamilyof the P-type ATPases that recognize and transport heavymetals and are distinguished from other ATPases by thepresence of the CP(C/H/S) motif, the P-residue generallylocated 43 amino acids upstream to an E1–E2 ATPasephosphorylation signature site (DKTGTLT) (Solioz andVulpe, 1996). Both rhizobial ActP proteins contain theCPC motif, 41 amino acids distal to the phosphorylationsite. This motif is located in the region considered to forman ion channel; members that transport a similar ion sharemore conserved residues in this region. Hydrophilicityprofiles of both rhizobial ActP proteins calculated from the Kyte–Doolittle algorithm show a typical plot for CPxproteins.
A search for domains (using the programs SCANPROSITE
and PROFILESCAN) in the ActP proteins has identified aheavy metal-associated (HMA) domain in the ActP proteinof S. meliloti (IEGMTCASCVRRVEKAIAAVPGVASANVNL). This sequence matches the consensus found inother CPx-type ATPases involved with heavy metal trans-port [L, I, V or N]–X2–[L, I, V, M, F or A]–X–C–X–[S, T, A,G, C, D, N or H]–C–X3–[L, I, V, F or G]–X3–[L, I orV]–X9–11–[I, V or A]–X–[L, V, F, Y or S] (Bull and Cox, 1994;Lin and Culotta, 1995); residues underlined indicate theselections from the consensus domain sequence, whilethose not underlined are spacer residues. Such an HMAdomain is absent from the R. leguminosarum ActP. TheEscherichia coli HRA-1, HRA-2 and Enterococcus hiraeCopB P-type ATPases also lack the HMA domain, butcontain histidine- and methionine-rich leader sequenceswhich Trenor et al. (1994) have suggested bind to heavymetals. The R. leguminosarum ActP sequence also con-tains a histidine-rich leader which may serve a similarfunction.
In addition, a prokaryotic lipoprotein lipid attachmentsite [D, E, R or K]–X6–[L, I, V, M, F, W, S, T, A or G]–X2–
[L, I, V, M, F, Y, S, T, A, G, C or Q]–[A, G or S]–C– (C isthe lipid attachment site; PROSITE database) found inmembrane-associated proteins is present within the ActPamino acid sequences of R. leguminosarum (LAAVAVLIIAC) and S. meliloti (VNAVAVLIIAC), although the initialconsensus residue is absent.
HmrR is a putative transcriptional regulator. Directlydownstream from actP in R. leguminosarum is an ORFthat encodes a protein of 14.7 kDa and a pI of 5.78,whereas in S. meliloti this protein is 16.3 kDa and has apI of 9.53. The two proteins are 55% identical (and contain74% similarity) over 129 amino acids. It is interesting thatthe exopoly-saccharide (EPS) biosynthetic regulatorExoR is also more acidic in R. leguminosarum (pI 5.5)than in its S. meliloti counterpart (pI 7.2) (Reeve et al.,1997). This ORF downstream from actP in both speciesencodes a protein with highest similarity to transcriptionalregulators of the MerR family; these proteins contain theMerR family helix–turn–helix DNA-binding signature in theN-terminal region (Helmann et al., 1989) (GEASKVSGVSSKMIRYYEQIGLI and GEASERSGLPSKTIRYYEDIGLI in S. meliloti and R. leguminosarum respectively;residues underlined are invariant across the proteinsYBBI, YBBI and PMTR from E. coli, Haemophilus influen-zae and P. mirabilis respectively).
The greatest identity (47% over 126 amino acids for R.leguminosarum and 60% over 128 amino acids for S.meliloti) occurs in the merR family with the heavy metal-dependent transcriptional regulator PMTR of P. mirabilis(Noll et al., 1998). The gene directly downstream to actPin the rhizobia has therefore been named hmrR (heavymetal-response regulator).
Function of TreY and EcfR. Upstream of actP in R. legu-minosarum is a partial ORF (treY) that encodes a protein34% identical over 346 amino acids to a malto-oligosyltrehalose synthase from Brevibacterium helvolum (acces-sion number AAB95368), Arthrobacter sp. (accessionnumber BAA09667) and Rhizobium sp. (accessionnumber BAA11186). Malto-oligosyl trehalose synthasecatalyses the conversion of maltodextrins to malto-oligosyl trehaloses through a transglucosylation reaction(Maruta et al., 1996). The match with TreY occurs overthe entire length of the partially sequenced R. legumi-nosarum protein.
Downstream from hmrR in R. leguminosarum is an ORFfor a protein with strong similarity to sigma factors of the extracytoplasmic factor (ECF) subfamily. The startcodon for this ORF (ecfR) in R. leguminosarum could bean ATG codon or an upstream GTG; both are potential ini-tiation codons, but only the GTG codon is preceded by aribosome binding site (RBS). EcfR shows the greatestidentity to the sigma-E factors (sigma-24) of Mycobac-terium tuberculosis (32% over 162 amino acids of SigH;
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 981–991
984 W. G. Reeve et al.

Involvement of ActP and HmrR in copper homeostasis 985
CAB08314), Streptomyces coelicolor (32% over 161amino acids of SigR; CAB09088), Mycobacterium smeg-matis (31% over 163 amino acids of SigH; AAD41810) andTreponema pallidum (33% over 145 amino acids of RpoE;AAC65087). In addition, the BLASTP search revealed 36%and 34% identity to a stretch of 144 amino acids of SigEfrom M. smegmatis (AA45268) and M. avium (AAC45219).
Typical for ECF sigma factors, the EcfR protein from R.leguminosarum is shorter than other classes of sigmafactors and it contains a fis-type (Kostrewa et al., 1992)helix–turn–helix DNA-binding motif EEQREALHLVAIEDLYSQEAAQ (residues underlined are invariant).
These sigma-24 proteins transcribe genes whose prod-ucts are required for responses to oxidative stress in S.coelicolor (Paget et al., 1998) and both heat shock and oxidative stress in M. tuberculosis (Fernandes et al., 1999). Interestingly, SigE in M. smegmatis has beenshown to promote survival against acidic pH in addition to high temperature, detergents and oxidative stress (Wu et al., 1997).
The phenotypic defects are not caused by a polar effecton hmrR expression
Since the hmrR gene is located downstream to actP inboth organisms and is transcribed in the same direction,it became prudent to investigate whether the acid andcopper sensitivity was caused by a Tn5 polar effect onhmrR expression.
Complementation of RT3-27. Ten individual clones wereisolated from a DNA library of S. meliloti WSM419 thatcould complement both the acid- and copper-sensitivephenotype of RT3-27. Digestion of each complementingplasmid with EcoRI released a 4.1 kb fragment that wascut by HindIII into 2.5 and 1.6 kb fragments. The patternobserved corresponds to that obtained from the rhizobialDNA region in the clone pRT3-27 (Fig. 1C). A 4.8 kbHindIII fragment (in pNK52; Fig. 1) restores copper toler-ance to RT3-27 but a 0.9 kb deletion between the EcoRVsites (in bold in Fig. 1) prevented complementation reveal-ing that the actP ORF does indeed extend into the EcoRVfragment, as predicted by sequence analysis.
Disruption of hmrR does not prevent complementationby the plasmid pNK77 while the generation of a frameshift mutation in actP (actP SphI site inactivated; Fig. 1C)destroyed the ability of pNK57 to complement.
Mobilization of pNK52 (containing S. meliloti actP andhmrR) into WR1-14 restores growth of this mutant onJMM plates buffered at pH 5.5 (containing 2 mM CuSO4)and also on TY plates at pH 7.0 containing 0.75 mMCuSO4. Despite the difference in the proposed metal-binding domains for the ActP proteins, it is evident thatActP possessing the HRA domain may be substituted with
ActP containing a HMA domain to serve the same func-tion in R. leguminosarum.
Complementation of WR1-14. The plasmid pWR880 was obtained by forcing spontaneous precise excision ofTn5 from actP by selecting for copper-tolerant colonies of WR1-14 that originally contained pWR873. Using thisstrategy, precise excision (identified from the number ofcopper-tolerant colonies) could be detected approxi-mately once in every 107 cells. All the copper-tolerantcolonies had lost Tn5 from the plasmid, indicating thatmultiple copies of the wild-type actP gene provided agrowth advantage to the cells on copper-containing platesover those cells that lost Tn5 from the single copy actPlesion already in WR1-14. Since the hmrR gene may beco-transcribed with actP in R. leguminosarum, we exam-ined the complementation pattern of plasmids derivedfrom pWR880. The plasmids pWR901 and pWR903 con-tained only a functional actP gene and were still able torestore the wild-type phenotype to WR1-14.
These observations indicate that the phenotypicdefects in both organisms result directly from the loss ofthe ActP protein and not from a polar effect on the expres-sion of the hmrR gene.
Expression of actP
An actP–gusA fusion present in the plasmid pWR668 wastranscriptionally induced by copper in R. leguminosarumgrown in MJMM-S (see Experimental procedures) con-taining glutamate as the nitrogen source (Fig. 2A). Tran-scription of actP increased further at low pH, parallelingthe observed increase in copper sensitivity of the mutantsat low pH. The expression appears to be specific tocopper since no such response was observed after cellexposure to silver or zinc (data not shown). The lack ofinduction in response to silver indicates that the consid-erable similarity of ActP to SilS does not extend to func-tion, in line with the lack of any increased sensitivity tosilver for both actP mutants.
Transcription is enhanced at low pH presumablybecause greater protonation of glutamate at acid pH than at neutral pH results in a decrease in copper chel-ation and increased free copper concentrations. In a liquidminimal medium containing 3 mM glutamate and 10 mMcopper, the free copper concentration has been calcu-lated to change from 0.00021 to 0.052 mM with a pH shiftfrom 7.0 to 5.5. For the same total concentration of copperin the medium, but with 10 mM NH4Cl replacing the glu-tamate, the range of free copper concentrations has beencalculated to change from 0.47 to 8.7 mM for the same pHchange (P. May, personal communication). In line with the theoretical calculations, cells were far more sensitiveto added copper in a medium containing NH4Cl, where
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 981–991
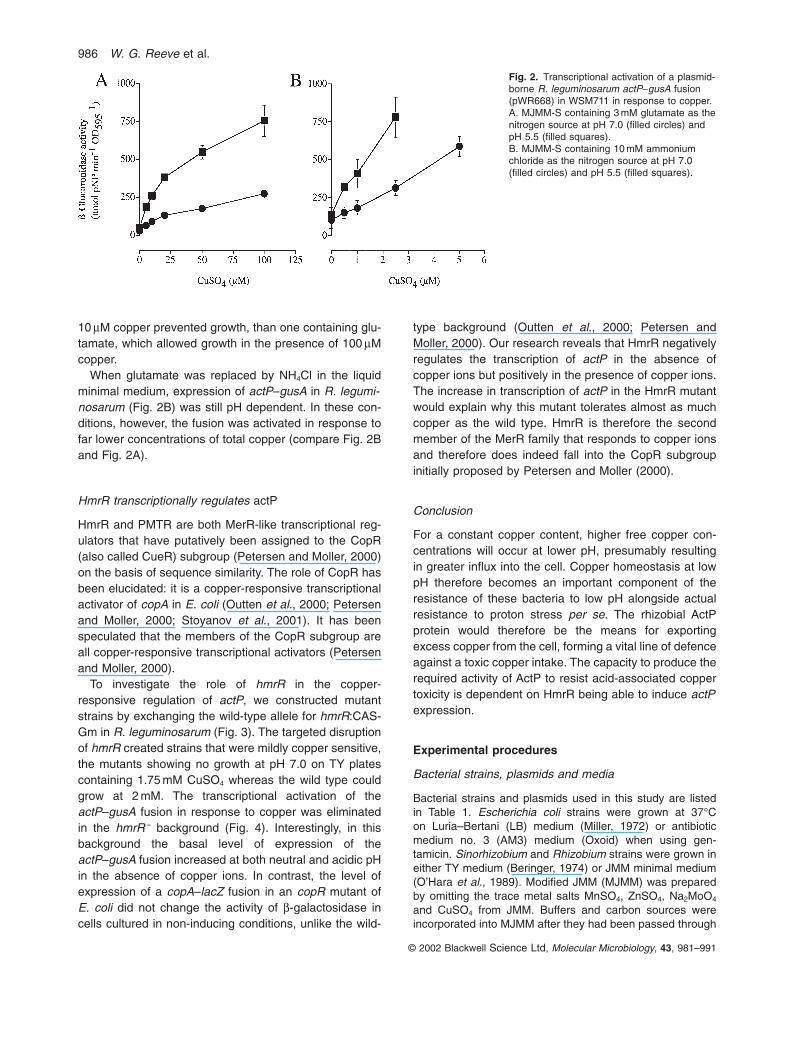
10 mM copper prevented growth, than one containing glu-tamate, which allowed growth in the presence of 100 mMcopper.
When glutamate was replaced by NH4Cl in the liquidminimal medium, expression of actP–gusA in R. legumi-nosarum (Fig. 2B) was still pH dependent. In these con-ditions, however, the fusion was activated in response tofar lower concentrations of total copper (compare Fig. 2Band Fig. 2A).
HmrR transcriptionally regulates actP
HmrR and PMTR are both MerR-like transcriptional reg-ulators that have putatively been assigned to the CopR(also called CueR) subgroup (Petersen and Moller, 2000)on the basis of sequence similarity. The role of CopR hasbeen elucidated: it is a copper-responsive transcriptionalactivator of copA in E. coli (Outten et al., 2000; Petersenand Moller, 2000; Stoyanov et al., 2001). It has beenspeculated that the members of the CopR subgroup areall copper-responsive transcriptional activators (Petersenand Moller, 2000).
To investigate the role of hmrR in the copper-responsive regulation of actP, we constructed mutantstrains by exchanging the wild-type allele for hmrR:CAS-Gm in R. leguminosarum (Fig. 3). The targeted disruptionof hmrR created strains that were mildly copper sensitive,the mutants showing no growth at pH 7.0 on TY platescontaining 1.75 mM CuSO4 whereas the wild type couldgrow at 2 mM. The transcriptional activation of theactP–gusA fusion in response to copper was eliminatedin the hmrR – background (Fig. 4). Interestingly, in thisbackground the basal level of expression of theactP–gusA fusion increased at both neutral and acidic pHin the absence of copper ions. In contrast, the level ofexpression of a copA–lacZ fusion in an copR mutant ofE. coli did not change the activity of b-galactosidase incells cultured in non-inducing conditions, unlike the wild-
type background (Outten et al., 2000; Petersen andMoller, 2000). Our research reveals that HmrR negativelyregulates the transcription of actP in the absence ofcopper ions but positively in the presence of copper ions.The increase in transcription of actP in the HmrR mutantwould explain why this mutant tolerates almost as muchcopper as the wild type. HmrR is therefore the secondmember of the MerR family that responds to copper ionsand therefore does indeed fall into the CopR subgroup initially proposed by Petersen and Moller (2000).
Conclusion
For a constant copper content, higher free copper con-centrations will occur at lower pH, presumably resulting in greater influx into the cell. Copper homeostasis at lowpH therefore becomes an important component of theresistance of these bacteria to low pH alongside actualresistance to proton stress per se. The rhizobial ActPprotein would therefore be the means for exportingexcess copper from the cell, forming a vital line of defenceagainst a toxic copper intake. The capacity to produce therequired activity of ActP to resist acid-associated coppertoxicity is dependent on HmrR being able to induce actPexpression.
Experimental procedures
Bacterial strains, plasmids and media
Bacterial strains and plasmids used in this study are listed in Table 1. Escherichia coli strains were grown at 37°C on Luria–Bertani (LB) medium (Miller, 1972) or antibioticmedium no. 3 (AM3) medium (Oxoid) when using gen-tamicin. Sinorhizobium and Rhizobium strains were grown ineither TY medium (Beringer, 1974) or JMM minimal medium(O’Hara et al., 1989). Modified JMM (MJMM) was preparedby omitting the trace metal salts MnSO4, ZnSO4, Na2MoO4
and CuSO4 from JMM. Buffers and carbon sources wereincorporated into MJMM after they had been passed through
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 981–991
986 W. G. Reeve et al.
Fig. 2. Transcriptional activation of a plasmid-borne R. leguminosarum actP–gusA fusion(pWR668) in WSM711 in response to copper.A. MJMM-S containing 3 mM glutamate as thenitrogen source at pH 7.0 (filled circles) andpH 5.5 (filled squares).B. MJMM-S containing 10 mM ammoniumchloride as the nitrogen source at pH 7.0(filled circles) and pH 5.5 (filled squares).

Involvement of ActP and HmrR in copper homeostasis 987
a CPG/8-hydroxyquinoline column, as described by Carsonet al. (1992) to prepare MJMM-S. This medium containedeither glutamate (3 mM) or NH4Cl (10 mM) as the nitrogensource. Media were supplemented with the following con-centrations of antibiotics (mg ml–1): ampicillin (100), chloram-phenicol (20), gentamicin (30; 7.5 for E. coli), kanamycin (50),streptomycin (200; 30 for E. coli) and tetracycline (20; 12.5for E. coli). Agar was added at a concentration of 1.5% (w/v)to solidify media.
Nodulation
Seeds of Pisum sativum L. (Wirrega pea) or Medicago murexwere surface sterilized, germinated and sown as describedby Reeve et al. (1999). Immediately after planting, peaseedlings were inoculated with a Rhizobium culture (eitherWSM710 or WR1-14) and medic seedlings with a Sinorhizo-bium culture (either WSM419 or RT3-27). Plants werewatered with sterile nutrient solution, and isolates were
recovered from surface-sterilized crushed nodules asdescribed by Reeve et al. (1999).
DNA manipulation and analysis
Plasmid or genomic DNA isolation, transformation and DNAmanipulation techniques have been described previously(Reeve et al., 1999). DNA sequencing and analysis were asreported by Tiwari et al. (1996b). Potential proteins wereidentified using the BLASTP program and the algorithm gen-erated by Altschul et al. (1997). Protein motifs were identifiedusing the programs SCANPROSITE and PROFILESCAN locatedon the ExPASy Molecular Biology Server provided as aservice by the Swiss Institute of Bioinformatics (SIB).
Plasmid mobilization
Plasmid DNA was mobilized from non-RP4-containing E. coli strains (such as DH5a or DH10B) into Rhizobium or
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 981–991
Fig. 3. Insertional inactivation of hmrR.A. Transposon excision. Tn5 was excisedfrom pWR873 as described in theExperimental procedures section to constructpWR880.B. Cassette insertion and subcloning. A 1.3 kbblunted HindIII fragment containing the CAS-Gm cassette was subcloned from pWRTACT-G2 into the blunted SphI site of hmrR in twoorientations. The NotI fragments containingthe hmrR:CAS-Gm mutation were subclonedinto pJQ2WS (filled arrows represent residualaacC1 gene) to form pWR1033 andpWR1037.C. Marker exchange. The latter plasmids weremobilized into WSM710 and a doublecrossover forced by plating the mating mixtureonto JMM minimal medium containinggentamicin and sucrose (5% w/v). HindIII-restricted genomic DNA from the fivestreptomycin and spectinomycin-sensitivemutants WR1880 (1), WR1881 (2), WR1882(3), WR1884 (4), WR1885 (5) and WSM710(6) was hybridized to DIG-labelledpWR114L–Hp/E to probe for hmrR (D) andDIG-labelled pJQ200SK to probe for thegentamicin resistance gene aacC1 (E).Disruption of hmrR with CAS-Gm shouldresult in a 1.2 kb increase (evident in D andE, lanes 2–5) in size of the 4.0 kb wild-typeHindIII fragment (C; and D, lane 6). The sizeof the 1.2 kb HindIII fragment (C; lowerhybridizing band in D) should not be disruptedduring the crossover process. The size offragments of the molecular weight markerfrom the top are approximately 23, 9.4, 6.6,5.1, 4.9, 4.4, 4.2, 3.5, 2.3, 2.0, 1.9, 1.5, 1.3,0.9, 0.8 and 0.5 kb.

© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 981–991
Table 1. List of strains and plasmids used
Strains/plasmids Relevant characteristics Source/reference
StrainsE. coli
BW20767 RP4-2-tet:Mu-1kan::Tn7 integrant leu-63::IS10 recA1 Metcalf et al. (1996)creC510 hsdR17 endA1 zbf-5 uidA(DMluI):pir+ thi
DH5a F– f80dLacZDM15 recA1 endA1 gyrA96 thi-1 hsdR17 Life Technologies(rK–mK+) supE44 relA1 deoR D(lacZYA–argF)U169
DH10B F– mcrA D(mrr–hsdRMS–mcrBC) f80dLacZDM15 lacX74 endA1 Life TechnologiesrecA1 deoR D(ara, leu)7697 araD139 galU galK nupG rpsL
HB101 F– thi-1hsdS20 (rB–mB
–) supE44 recA13 ara-14 Boyer and Roulland-Dussoix (1969)leuB6 proA2 lacY1 rpsL20 (Smr) xyl-5 mtl-1
MT616 pro-82 thi-1 hsdR17 supE44 endA1 recA56 (pRK600); Cmr Finan et al. (1986)S. meliloti
RT3-27 actP::Tn5 mutant of WSM419; Cus, Cmr, Kmr This studyWSM419 Acidt Sardinian isolate; Cmr, Cut J. Howiesona
R. leguminosarum bv. viciaePW711 Spontaneous Smr mutant of WSM710 Reeve et al. (1999)WR1-14 actP::Tn5 mutant of WSM710; Cus, Kmr, Smr This studyWR1870 hmrR:Gm mutant of WSM710; Cut, Gmr This studyWR1871 hmrR:Gm mutant of WSM710; Cut, Gmr This studyWR1880 WR1870 (pMUE668); Tcr, Gmr, Kmr This studyWR1885 WR1875 (pMUE668); Tcr, Gmr, Kmr This studyWSM710 Acidt Japanese isolate; Cut J. Howieson
PlasmidspBR01 pFUS1 derivative constructed by replacing the PstI/NotI fragment Beau Fennera
containing gusA with the PstI/NotI MCS from pUK21; Tcr
pBR322 Cloning vector; Apr Tcr Bolivar et al. (1977)pCRS433 Broad host range vector; Tcr Reeve et al. (1999)pCRS487 pUT::mTn5-GNm; Apr, Kmr Reeve et al. (1999)pFUS1 pCRS433 derivative containing a promoterless gusA; Tcr Reeve et al. (1999)pJQ2WS pJQ200SK derivative constructed by replacing a blunted EcoRV/SphI Sophie Rome
fragment containing aacC1 with a blunted HindIII fragment containingWSm/Sp; Sm/Spr, Sucs
pJQ200SK Gene replacement vector; Gmr, Sucs Quandt and Hynes (1993)pNK52 pSW213 containing actP and hmrR on a 4.8 kb HindIII fragment; Tcr This studypNK57 pFUS1 containing a mutated actP and wild-type hmrR on a
4.2 kb HindIII/ApaI fragment (Fig. 1); Tcr This studypNK77 pFUS1 containing a 4.8 kb S. meliloti HindIII fragment carrying actP This study
and hmrR:CAS-GNm (inactivation of the hmrR BclI site); Tcr, Kmr
pRK2013 Helper plasmid; Kmr Figurski and Helinski (1979)pRT327 RT3-27 EcoRI fragment containing Tn5 cloned in pBR322; Apr, Kmr This studypRT327-1 Complementing plasmid clone from WSM419 genomic library; Tcr This studypSW213 Broad host range cloning vector; Tcr Chen and Winans (1991)pTn5-H HindIII fragment containing nptII from Tn5 in pUC18; Apr, Kmr This studypWR053 HindIII fragment of pWR114 containing right inverted repeat of Tn5 This study
and associated rhizobial flanking sequence in pBR322; Apr
pWR114 Tn5-containing EcoRI fragment of WR1-14 in pBR322; Apr, Kmr, Tcr This studypWR114L–Hp/E HpaI–EcoRI fragment containing a portion of Tn5 IS50L and This study
associated rhizobial DNA from pWR1-14 in pGEM-7Zf(+); Apr
pWR327-200 Tn5-containing EcoRI fragment from RT3-27 in pJQ200SK; Gmr, Kmr This studypWR657 NotI fragment containing CAS-GNm cloned from pCRS487 into This study
the NotI site of pWR053; Apr, Kmr
pWR668 HindIII–EcoRI blunted fragment containing the actP–gusA fusion This studyfrom pWR657 in a blunted XhoI site of pCRS433; Tcr, Kmr
pWR873 EcoRI fragment containing Tn5 from WR1-14 in pBRO1; Tcr, Kmr This studypWR880 pWR873 devoid of Tn5; Tcr This studypWR901 pWR880 derivative containing actP and hmrR:CAS-Gm This study
(inactivation of the hmrR SphI site); Tcr, Gmr
pWR903 pWR880 derivative containing actP and hmrR:CAS-Gm This study(inactivation of the hmrR SphI site); Tcr, Gmr
pWR1033 NotI fragment containing CAS-Gm cloned from pWR901 into This studyNotI pJQ2WS; Sucs, Gmr, Smr, Spr
pWR1037 NotI fragment containing CAS-Gm cloned from pWR903 into This studyNotI pJQ2WS; Sucs, Gmr, Smr, Spr
PhageRL38 Generalized transducing phage Buchanan-Wollaston (1979)
a. Centre for Rhizobium Studies.Acid tolerant (Acidt); Cu sensitive (Cus) or tolerant (Cut); resistance to ampicillin (Apr), chloramphenicol (Cmr), gentamicin (Gmr), kanamycin (Kmr),streptomycin (Smr), sucrose (Sucs) or tetracycline (Tcr).

Involvement of ActP and HmrR in copper homeostasis 989
Sinorhizobium by performing a triparental mating with eitherHB101 (pRK2013) or MT616. Plasmids were mobilized fromBW20767 using a biparental mating. Both methods used theconjugation protocol described by Reeve et al. (1999).
Phage transduction
The method of Buchanan-Wollaston (1979) was used to prepare phage RL38 stocks and transduce WSM710.EcoRI-digested genomic DNA of the wild-type andkanamycin-resistant transductants was probed with DIG-labelled pWR114 to show that the mutants contained a singlecopy of Tn5 in actP.
Reconstruction of a S. meliloti actP mutant
An EcoRI digest of pRT327 was ligated to a partial EcoRIdigest of pJQ200SK and the reaction mixture transformedinto E. coli BW20767. Transformants were selected for gen-tamicin and kanamycin resistance and replica patched forsucrose sensitivity. In this way, the 9.0 kb EcoRI fragmentcontaining Tn5 (from pRT327) was cloned into the EcoRI site in the multiple cloning site of pJQ200SK to constructpWR327-200. The latter construct was mobilized fromBW20767 into WSM419 to exchange the wild-type actPgene in S. meliloti WSM419 with actP::Tn5 via homologousrecombination. Transconjugants were selected on JMMplates (pH 7.0) containing chloramphenicol, streptomycin,kanamycin and sucrose (10% w/v) and then patched for gentamicin sensitivity. The replacement of actP for actP::Tn5
was confirmed by hybridization of EcoRI-digested DNA of putative actP mutants to DIG-labelled pRT327.
DNA hybridizations
DNA labelling, Southern hybridization and probe detectionwere as described previously (Tiwari et al., 1996b).
Complementation analysis
S. meliloti. A DNA library of WSM419 (Tiwari et al., 1996b)was mobilized from DH10B into S. meliloti RT3-27. Sinorhi-zobium transconjugants were selected on TY plates contain-ing chloramphenicol, kanamycin, tetracycline and 0.75 mMCuSO4. Plasmid DNA was isolated from Sinorhizobiumtransconjugants using alkaline lysis (Sambrook et al., 1989)and then transformed into BW20767. Plasmid DNA wasextracted from these transformants, purified by CsCl isopyc-nic centrifugation and mapped for restriction sites. One complementing plasmid (designated as pRT327-1) that con-tained the S. meliloti actP wild-type gene was mobilized from BW20767 into S. meliloti RT3-27 and R. leguminosarumWR1-14. Transconjugants were selected on JMM plates con-taining kanamycin and tetracycline. The ability of plasmidpRT327-1 to complement the actP defect in the two mutantswas assessed by comparing phenotypes of transconjugantsagainst the wild type on JMM plates in the presence andabsence of copper or on low-pH-buffered JMM plates.
R. leguminosarum. The EcoRI fragment containingactP::Tn5 was subcloned from pWR114 into the broad hostrange vector pBRO1 to construct pWR873. The latter plasmidwas mobilized into WR1-14 and the transconjugants grownto saturation in TY containing tetracycline and streptomycin.Aliquots of the culture were spread onto TY plates contain-ing tetracycline, kanamycin and 0.75 mM CuSO4. PlasmidDNA was extracted from 25 copper-tolerant colonies andtransformed into E. coli DH5a. Transformants were replicapatched onto LB containing tetracycline and then onto LB containing tetracycline and kanamycin. Plasmid DNAwas extracted from tetracycline-resistant and kanamycin-sensitive colonies and the restriction profile compared with pWR873 to ensure that Tn5 had excised from actP. Oneof the complementing plasmids (pWR880) was chosen for further work.
Construction of an actP–gusA fusion
A NotI fragment containing CAS-GNm (Reeve et al., 1999)was cloned from pCRS489 into the NotI site of pWR053 toconstruct pWR657. A HindIII–EcoRI fragment containing theactP–gusA fusion from pWR657 was end-filled and thencloned into a blunted XhoI site of the broad host rangeplasmid pCRS433 to create pWR668. The latter plasmid wasmobilized from E. coli BW20767 into R. leguminosarumbv. viciae PW711, WR1870 or WR1875 by selecting for tetracycline-resistant transconjugants on JMM plates.
Expression studies
Cultures containing pWR668 were inoculated into 5 ml of
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 981–991
Fig. 4. The actP gene is transcriptionally regulated by HmrR in R.leguminosarum in response to copper. b-Glucuronidase expressionfrom the actP–gusA fusion located on the broad host range plasmidpWR668 was measured from the wild-type (PW711) and hmrRmutants (WR1880 and WR1885) in the absence (non-filled bars)and presence (filled bars) of 50 mM CuSO4.

MJMM (glutamate or ammonium chloride as the nitrogensource) at pH 7.0 containing kanamycin. Cultures were grownat 28°C to an optical density at 600 nm (OD600) of approxi-mately 0.4–0.8. Suspensions were centrifuged, concentratedand resuspended in MJMM-S (with kanamycin) in the pres-ence or absence of copper at pH 7.0 or 5.5 to obtain an OD600
of approximately 0.25–0.5 after overnight incubation at 28°C.
Measurement of GUS activity in microplates
A sample of each culture (3 ml) was centrifuged and resus-pended in saline (0.9% NaCl, w/v) to an OD600 of approxi-mately 1. Aliquots (80 ml resuspended culture with 120 mlsaline) were dispensed into flat-bottomed microplate (Sarst-edt Australia) wells and the OD595 measured in a Bio-RadModel 550 Microplate Reader controlled by MICROPLATE
MANAGER software.For the assay, aliquots of 50 and 125 ml were dispensed
in duplicate into four Eppendorf tubes for each culture. Thevolume was adjusted to 125 ml using saline. b-Glucuroni-dase(GUS) buffer (500 ml) and toluene (two drops) were dispensedinto each tube and vigorously vortexed for exactly 10 s (Reeveet al., 1999). The step was performed in Eppendorf tubes toensure complete cell permeabilization by toluene. TheseEppendorf tubes were incubated at 37°C for 30 min with thelids open to evaporate toluene. The tubes were then trans-ferred to 28°C for 5 min. An aliquot of 200 ml from each Eppen-dorf tube was dispensed into a flat-bottomed microplate well.To start the reaction, aliquots of 5 ml of p-nitrophenyl-b-D-glu-curonide (pNPG; 35 mg ml –1in water) were dispensed into thewells using a multichannel pipette. The OD405 was measuredevery 2 min over 1.5 h using a Bio-Rad Model 550 MicroplateReader controlled by MICROPLATE MANAGER software.
Four replicates per strain were assayed from two separateexperiments. b-Glucuronidase specific activity was ex-pressed as nmol p-nitrophenol (pNP) produced per min perOD595 at 28°C.
Accession numbers
The nucleotide sequences derived from R. leguminosarumbv. viciae WSM710 and S. meliloti WSM419 have beenlodged with GenBank and have been assigned the accessionnumbers AF127795 and AF129004 respectively.
Acknowledgements
This work was generously supported by the AustralianResearch Council (ARC). N.B.K. was funded by the JohnCrawford Scholarship Award and AusAid. We would like tothank Peter May for the calculation of the free copper con-centrations in our media, Claude Defives for extensivelytesting WSM419 for phage susceptibility and Sophie Romefor the plasmid pJQ2WS.
References
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang,Z., Miller, W., and Lipman, D.J. (1997) Gapped BLAST and
PSI-BLAST: a new generation of protein database searchprograms. Nucleic Acids Res 25: 3389–3402.
Barnet, Y.P. (1991) Ecology of the root nodule bacteria. InThe Biology and Biochemistry of Nitrogen Fixation. Dilworth, M.J., and Glenn, A.R. (eds). Amsterdam: Elsevier, pp. 199–208.
Beringer, J.E. (1974) R factor transfer in Rhizobium legumi-nosarum. J Gen Microbiol 84: 188–198.
Bolivar, F., Rodriguez, R.L., Greene, P.J., Betlach, M.C., Heynecker, H.L., Boyer, H.W., et al. (1977) Constructionand characterisation of new cloning vehicles. II. A multi-purpose cloning system. Gene 2: 95–113.
Boyer, H.W., and Roulland-Dussoix, D. (1969) A comple-mentation analysis of the restriction and modification ofDNA in Escherichia coli. J Mol Biol 41: 459–472.
Buchanan-Wollaston, V. (1979) Generalized transduction inRhizobium leguminosarum. J Gen Microbiol 112: 135–142.
Bull, P.C., and Cox, D.W. (1994) Wilson disease and Menkesdisease: new handles on heavy-metal transport. TrendsGenet 10: 246–252.
Carson, K.C., Holliday, S., Glenn, A.R., and Dilworth, M.J.(1992) Siderophore and organic acid production in rootnodule bacteria. Arch Microbiol 157: 264–271.
Chen, C.Y., and Winans, S.C. (1991) Controlled expressionof the transcriptional activator gene virG in Agrobacteriumtumefaciens by using the Escherichia coli lac promoter. J Bacteriol 173: 1139–1144.
Coventry, D.R., and Evans, J. (1989) Symbiotic nitrogen fix-ation and soil acidity. In Soil Acidity and Plant Growth.Robson, A.D. (ed.). New York: Academic Press, pp.103–107.
Fernandes, N.D., Wu, Q.L., Kong, D., Puyang, X., Garg, S.,and Husson, R.N. (1999) A mycobacterial extracytoplasmicsigma factor involved in survival following heat shock andoxidative stress. J Bacteriol 181: 4266–4274.
Figurski, D.H., and Helinski, D.R. (1979) Replication of anorigin-containing derivative of plasmid RK2 dependent ona plasmid function provided in trans. Proc Natl Acad SciUSA 76: 1648–1652.
Finan, T.M., Kunkel, B., De Vos, G.F., and Signer, E.R. (1986)Second symbiotic megaplasmid in Rhizobium meliloticarrying exopolysaccharide and thiamine synthesis genes.J Bacteriol 167: 66–72.
Glenn, A.R., Reeve, W.G., Tiwari, R.P., and Dilworth, M.J.(1999) Acid tolerance in root nodule bacteria. In BacterialResponse to pH. Chadwick, D.J., and Cardew, G. (eds).Novartis Foundation Symposium no. 221. London: WileyPublishing, 112–126.
Goss, T.G., O’Hara, G.W., Dilworth, M.J., and Glenn, A.R.(1990) Cloning, characterization, and complementation oflesions causing acid sensitivity in Tn5-induced mutants ofRhizobium meliloti WSM419. J Bacteriol 172: 5173–5179.
Helmann, J.D., Wang, Y., Mahler, I., and Walsh, C.T. (1989)Homologous metalloregulatory proteins from both gram-positive and gram-negative bacteria control transcription ofmercury resistance operons. J Bacteriol 171: 222–229.
Howieson, J.G., Ewing, M.A., and D’Antuono, M.F. (1988)Selection for acid-tolerance in Rhizobium meliloti. PlantSoil 105: 179–188.
Kahn, D., David, M., Domergue, O., Daveran, M.L., Ghai, J.,Hirsch, P.R., and Batut, J. (1989) Rhizobium meliloti fixGHI
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 981–991
990 W. G. Reeve et al.

Involvement of ActP and HmrR in copper homeostasis 991
sequence predicts the involvement of a specific pump insymbiotic nitrogen fixation. J Bacteriol 171: 929–939.
Kostrewa, D., Granzin, J., Stock, D., Choe, H.W., Labahn, J.,and Saenger, W. (1992) Crystal structure of the factor forinversion stimulation FIS at 2.0 A resolution. J Mol Biol226: 209–226.
Lin, S.J., and Culotta, V.C. (1995) The ATX1 gene of Sac-charomyces cerevisiae encodes a small metal homeosta-sis factor that protects cells against reactive oxygentoxicity. Proc Natl Acad Sci USA 92: 3784–3788.
Maruta, K., Hattori, K., Nakada, T., Kubota, M., Sugimoto, T.,and Kurimoto, M. (1996) Cloning and sequencing of tre-halose biosynthesis genes from Rhizobium sp. M-11.Biosci Biotechnol Biochem 60: 717–720.
Mellor, R.B. (1989) Bacteroids in the Rhizobium–legumesymbiosis inhabit a plant internal lytic compartment: impli-cations for other microbial endosymbioses. J Exp Bot 40:831–839.
Metcalf, W.W., Jiang, W., Daniels, L.L., Kim, S., Haldimann,A., and Wanner, B.L. (1996) Conditionally replicative andconjugative plasmids carrying lacZa for cloning, muta-genesis and allele replacement in bacteria. Plasmid 35:1–13.
Miller, J.H. (1972) Experiments in Molecular Genetics. ColdSpring Harbor, New York: Cold Spring Harbor LaboratoryPress.
Munns, D.N. (1986) Acid soil tolerance in legumes and rhizobia. In Advances in Plant Nutrition. Tinker, B., andLauchlie, A. (eds). New York: Praeger Scientific, pp. 63–90.
Noll, M., Petrukhin, K., and Lutsenko, S. (1998) Identificationof a novel transcription regulator from Proteus mirabilis,PMTR, revealed a possible role of YJA1 protein in balanc-ing zinc in Escherichia coli. J Biol Chem 273: 21393–21401.
O’Hara, G.W., Goss, T.J., Dilworth, M.J., and Glenn, A.R.(1989) Maintenance of intracellular pH and acid-tolerancein Rhizobium meliloti. Appl Environ Microbiol 55: 1870–1876.
Outten, F.W., Outten, C.E., Hale, J., and O’Halloran, T.V.(2000) Transcriptional activation of an Escherichia colicopper efflux regulon by the chromosomal MerR homo-logue, CueR. J Biol Chem 275: 31024–31029.
Paget, M.S., Kang, J.G., Roe, J.H., and Buttner, M.J. (1998)SigmaR, an RNA polymerase sigma factor that modulatesexpression of the thioredoxin system in response to oxida-tive stress in Streptomyces coelicolor A3(2). EMBO J 17:5776–5782.
Parniske, M. (2000) Intracellular accommodation of microbesby plants: a common developmental program for symbio-sis and disease? Curr Opin Plant Biol 3: 320–328.
Petersen, C., and Moller, L.B. (2000) Control of copper home-ostasis in Escherichia coli by a P-type ATPase, CopA, anda MerR-like transcriptional activator, CopR. Gene 261:289–298.
Preisig, O., Zufferey, R., and Hennecke, H. (1996) TheBradyrhizobium japonicum fixGHIS genes are required forthe formation of the high-affinity cbb3-type cytochromeoxidase. Arch Microbiol 165: 297–305.
Quandt, J., and Hynes, M.F. (1993) Versatile suicide vectors which allow direct selection for gene replacementin Gram-negative bacteria. Gene 127: 15–21.
Recourt, K., Van Brussel, A.A.N., Driessen, A.J.M., andLugtenberg, B.J.J. (1989) Accumulation of nod geneinducer, the flavonoid naringenin, in the cytoplasmic mem-brane of Rhizobium leguminosarum bv. viciae is caused by the pH-dependent hydrophobicity of naringenin. J Bacteriol 171: 4370–4377.
Reeve, W.G., Tiwari, R.P., Dilworth, M.J., and Glenn, A.R.(1993) Calcium affects the growth and survival of Rhizo-bium meliloti. Soil Biol Biochem 25: 581–586.
Reeve, W.G., Dilworth, M.J., Tiwari, R.P., and Glenn, A.R.(1997) Regulation of exopolysaccharide production in Rhizobium leguminosarum WSM710 involves exoR.Microbiology 143: 1951–1958.
Reeve, W.G., Tiwari, R.P., Worsley, P.S., Dilworth, M.J.,Glenn, A.R., and Howieson, J.G. (1999) Constructs forinsertional mutagenesis, transcriptional signal localisationand gene regulation studies in root nodule and other bacteria. Microbiology 145: 1307–1316.
Robson, A.D., and Bottomley, P.J. (1991) Limitations in theuse of legumes in agriculture and forestry. In Biology andBiochemistry of Nitrogen Fixation. Dilworth, M.J., andGlenn, A.R. (eds). Amsterdam: Elsevier, pp. 320–349..
Sambrook, J., Fritsch, E.F., and Maniatis, T. (1989) Molecu-lar Cloning: A Laboratory Manual, 2nd edn. Cold SpringHarbor, New York: Cold Spring Harbor Laboratory Press.
Snowball, K., and Robson, A.D. (1980) The effect of copperon nitrogen fixation in subterranean clover (Trifoliumsubterraneum) New Phytol 85: 63–72.
Solioz, M., and Vulpe, C. (1996) CPx-type ATPases: a classof P-type ATPases that pump heavy metals. TrendsBiochem Sci 21: 237–241.
Stoyanov, J.V., Hobman, J.L., and Brown, N.L. (2001) CueR(YbbI) of Escherichia coli is a MerR family regulator controlling expression of the copper exporter CopA. MolMicrobiol 39: 502–511.
Tiwari, R.P., Reeve, W.G., and Glenn, A.R. (1992) Mutationsconferring acid-sensitivity in the acid-tolerant strains ofRhizobium meliloti WSM419 and Rhizobium legumi-nosarum biovar viciae WSM710. FEMS Microbiol Lett 100:107–112.
Tiwari, R.P., Reeve, W.G., Dilworth, M.J., and Glenn, A.R.(1996a) Acid-tolerance in Rhizobium meliloti strainWSM419 involves a two-component sensor–regulatorsystem. Microbiology 142: 1693–1704.
Tiwari, R.P., Reeve, W.G., Dilworth, M.J., and Glenn, A.R.(1996b) An essential role for actA in acid-tolerance of Rhizobium meliloti. Microbiology 142: 601–610.
Trenor, C., Lin, W., and Andrews, N.C. (1994) Novel bacter-ial P-type ATPases with histidine-rich heavy-metal associ-ated sequences. Biochem Biophys Res Commun 205:1644–1650.
Wu, Q.L., Kong, D., Lam, K., and Husson, R.N. (1997) Amycobacterial extracytoplasmic function sigma factorinvolved in survival following stress. J Bacteriol 179:2922–2929.
© 2002 Blackwell Science Ltd, Molecular Microbiology, 43, 981–991




















