
Response to salinity stress of Rhizobium leguminosarum bv. viciae strains in the presence of...
13
ORIGINAL ARTICLE Response to salinity stress of Rhizobium leguminosarum bv. viciae strains in the presence of different legume host plants ValeriaVentorino & Rosanna Caputo & Stefania De Pascale & Massimo Fagnano & Olimpia Pepe & Giancarlo Moschetti Received: 22 March 2011 /Accepted: 13 July 2011 # Springer-Verlag and the University of Milan 2011 Abstract We investigated the effect of residual salts from the previous summer ’ s irrigation on two non-irrigated cover crops—broad bean and common vetch—and on their rhizobial symbiontics. Before sowing, seeds were inoculat- ed with a salt-tolerant strain and a salt-sensitive strain of Rhizobium leguminosarum biovar viciae. An increase in the electrical conductivity of the saturated-soil extract from 2.0 dS m -1 to 6.0 dS m -1 caused a severe reduction of broad bean biomass, while growth of common vetch was almost unaffected by the salinity level. Our results clearly indicate that common vetch as a cover crop may increase the availability of nitrogen in soil more than broad bean also in saline environments. A high ability of both inoculated strains to nodulate Vicia plants was observed under controlled conditions. In contrast, inoculated strains were not able to nodulate plants in salinized soils, showing lower competitiveness compared to natural rhizobial population. Indeed, the new isolates from nodules were salt-sensitive when tested in vitro. The complex interaction among bacteria, plant and soil determined the survival of endog- enous salt sensitive rhizobial strains and limited the success of inoculation with exogenous salt resistant rhizobia. Keywords Broad bean (Vicia faba L. var. major) . Common vetch (Vicia sativa L.) . Rhizobium leguminosarum . Irrigation water salinity . Residual soil salinity . Salt-resistant strain Introduction Soil salinization is increasing in many areas of the world and causes major problems for the productivity of agricul- tural crops (Parida and Das 2005). Nearly 40% of the world’ s land surface can be categorized as having potential salinity problems (Zahran 1999). In coastal regions of Mediterranean areas, summer crops are often irrigated with saline water. As a consequence, salts accumulate along the soil profile if rainfalls are not sufficient to remove them from the root-zone. The long-term consequences of saline irrigation on soil physical fertility are well documented (De Pascale et al. 2003a; 2003b). The saturation level of sodium is considered the main cause of clay dispersion and degradation of aggregate stability, with the consequent reduction in permeability, porosity and hydraulic conduc- tivity of soils (Amézketa 1999). Cover crops are fundamental, sustainable tools used to manage soil fertility (Lu et al. 2000). Cover crop incorporation into soil may improve physical character- istics, including water infiltration (Cassman and Rains 1986), aggregation and porosity (MacRae and Mehuys 1985) and reduction of soil crust formation (Mitchell et al. 2000). They are commonly leguminous (Fabaceae family), i.e., able to establish a symbiotic association with certain V . Ventorino : O. Pepe Dipartimento di Scienza degli Alimenti, Sezione di Microbiologia Agraria, Università degli Studi di Napoli “Federico II”, via Università 100, 80055 Portici, Italy R. Caputo : S. De Pascale : M. Fagnano Dipartimento di Ingegneria Agraria e Agronomia del Territorio, Università degli Studi di Napoli “Federico II”, via Università 100, 80055 Portici, Italy G. Moschetti (*) Dipartimento DEMETRA, Microbiology Section, Università degli Studi di Palermo, Viale delle Scienze 13, 98128 Palermo, Italy e-mail: [email protected] Ann Microbiol DOI 10.1007/s13213-011-0322-6
Transcript of Response to salinity stress of Rhizobium leguminosarum bv. viciae strains in the presence of...

ORIGINAL ARTICLE
Response to salinity stress of Rhizobium leguminosarum bv.viciae strains in the presence of different legume host plants
Valeria Ventorino & Rosanna Caputo &
Stefania De Pascale & Massimo Fagnano &
Olimpia Pepe & Giancarlo Moschetti
Received: 22 March 2011 /Accepted: 13 July 2011# Springer-Verlag and the University of Milan 2011
Abstract We investigated the effect of residual salts fromthe previous summer’s irrigation on two non-irrigated covercrops—broad bean and common vetch—and on theirrhizobial symbiontics. Before sowing, seeds were inoculat-ed with a salt-tolerant strain and a salt-sensitive strain ofRhizobium leguminosarum biovar viciae. An increase in theelectrical conductivity of the saturated-soil extract from 2.0dS m−1 to 6.0 dS m−1 caused a severe reduction of broadbean biomass, while growth of common vetch was almostunaffected by the salinity level. Our results clearly indicatethat common vetch as a cover crop may increase theavailability of nitrogen in soil more than broad bean also insaline environments. A high ability of both inoculatedstrains to nodulate Vicia plants was observed undercontrolled conditions. In contrast, inoculated strains werenot able to nodulate plants in salinized soils, showing lowercompetitiveness compared to natural rhizobial population.Indeed, the new isolates from nodules were salt-sensitivewhen tested in vitro. The complex interaction among
bacteria, plant and soil determined the survival of endog-enous salt sensitive rhizobial strains and limited the successof inoculation with exogenous salt resistant rhizobia.
Keywords Broad bean (Vicia faba L. var. major) . Commonvetch (Vicia sativa L.) . Rhizobium leguminosarum .
Irrigation water salinity. Residual soil salinity. Salt-resistantstrain
Introduction
Soil salinization is increasing in many areas of the worldand causes major problems for the productivity of agricul-tural crops (Parida and Das 2005). Nearly 40% of theworld’s land surface can be categorized as having potentialsalinity problems (Zahran 1999). In coastal regions ofMediterranean areas, summer crops are often irrigated withsaline water. As a consequence, salts accumulate along thesoil profile if rainfalls are not sufficient to remove themfrom the root-zone. The long-term consequences of salineirrigation on soil physical fertility are well documented (DePascale et al. 2003a; 2003b). The saturation level of sodiumis considered the main cause of clay dispersion anddegradation of aggregate stability, with the consequentreduction in permeability, porosity and hydraulic conduc-tivity of soils (Amézketa 1999).
Cover crops are fundamental, sustainable tools used tomanage soil fertility (Lu et al. 2000). Cover cropincorporation into soil may improve physical character-istics, including water infiltration (Cassman and Rains1986), aggregation and porosity (MacRae and Mehuys1985) and reduction of soil crust formation (Mitchell et al.2000). They are commonly leguminous (Fabaceae family),i.e., able to establish a symbiotic association with certain
V. Ventorino :O. PepeDipartimento di Scienza degli Alimenti, Sezione di MicrobiologiaAgraria, Università degli Studi di Napoli “Federico II”,via Università 100,80055 Portici, Italy
R. Caputo : S. De Pascale :M. FagnanoDipartimento di Ingegneria Agraria e Agronomia del Territorio,Università degli Studi di Napoli “Federico II”,via Università 100,80055 Portici, Italy
G. Moschetti (*)Dipartimento DEMETRA, Microbiology Section,Università degli Studi di Palermo,Viale delle Scienze 13,98128 Palermo, Italye-mail: [email protected]
Ann MicrobiolDOI 10.1007/s13213-011-0322-6

bacteria such as rhizobia and so enrich soil nitrogen, thanksto biological nitrogen contribution, and reduce the require-ment for seasonal N-fertilizers (Perret et al. 2000; Spaink2000). Broad bean (Vicia faba L. var. major Harz., cv.‘Dulce’) and common vetch (Vicia sativa L.) are veryinteresting legume cover crops used to improve the nitrogenbudget of Mediterranean cropping systems (Guiducci et al.2004).
The importance of biological nitrogen fixation inagriculture has been demonstrated in a huge number ofstudies, particularly those that study the impact of soilmanagement on rhizobial populations in arid regions(Howieson and Ballard 2004), as well as by those that dealwith the isolation and characterization of drought- and salt-resistant strains (Chen et al. 2000; Jenkins 2003; Yan et al.2000). It is widely recognized that indigenous rhizobia playan important role in the dominance of Fabaceae in poor andarid soils (Zahran 1999). In fact, the salt tolerance of broadbean was found to be correlated with the response of therespective rhizobia strains to salt stress (Katerji et al. 2003).Unfortunately, information about salt resistance of commonvetch and other vetch species are scarce (UC SAREP2008). However, harsh soil conditions can have a dramaticimpact on soil bacteria (Fierer et al. 2003; Griffiths et al.2003), including rhizobia (Zahran 1999). In fact, thelegume–Rhizobium association is adversely affected by soilsalinity, which can preclude bacterial multiplication andnodulation, reducing crop yield (Ogutcu et al. 2010). Insaline soils, the nitrogen content decreases since thebiological nitrogen fixation (BNF) and the mineralizationof organic nitrogen decrease (Van Horn et al. 2001).
Rhizobia strains differ in their ability to cope withabiotic stresses, and use distinct mechanisms for osmoticadaptation when exposed to salt stress, such as intracellularaccumulation of low-molecular-weight organic solutes(Miller and Wood 1996; Zahran 1999) or changes in cellmorphology and size and modifications in the pattern ofextracellular polysaccharides (Lloret et al. 1995, 1998;Soussi et al. 2001).
Some strains of Rhizobium spp. are salt tolerant and,under stress conditions, root-associated beneficial bacteriacan help to improve plant growth and nutrition of their hostplants (Upadhyay et al. 2011). Reduced nodule conductanceto O2 diffusion is considered a major factor responsible forthe inhibition of N2 fixation by soil salinity. Salt tolerancewas associated with higher stability of O2 nodule conduc-tance of the tolerant Rhizobia at increasing soil salinity(L’taief et al. 2007).
In saline environments, inoculation of crops with tolerantsymbiotic strains of Rhizobium may improve their stresstolerance (Bianco and Defez 2009). Legume seeds arecommonly inoculated with peat cultures or liquid inoculantbefore planting (Deaker et al. 2004). The aim of inoculation
is to provide sufficient numbers of viable effective rhizobiato induce rapid colonization of the rhizosphere, so thatnodulation can take place as soon as possible aftergermination (Catroux et al. 2001).
In this study we evaluated the effects of salinity onthe overall performance of two field grown leguminouscover crops (broad bean and common vetch), inoculatedwith salt-tolerant and salt-sensitive strains of Rhizobiumleguminosarum biovar viciae. The two rhizobia strainswere evaluated for their ability to compete in soilcontaining a naturalized population of rhizobia and theirpotential use in sustainable agriculture projects.
Materials and methods
Bacterial strains
Several strains previously isolated in the Campania region(Southern Italy) were tested for their potential use forinoculating legumes in experimental fields (Moschetti et al.2005). The strains Rhizobium leguminosarum biovar viciaeSAAN1 and Rhizobium leguminosarum biovar viciaePOHY2B1 isolated from Lathyrus annus in Montecorice(Salerno, Italy) and from Vicia hybrida in Portici (Naples,Italy), respectively, were used in this study. R. leg. bv. viciaeSAAN1 is a salt-tolerant strain able to grow up to 2% (w/v)(0.34 M) NaCl, while R. leg. bv. viciae POHY2B1 is a salt-sensitive strain able to grow up to 0.1% (w/v) NaCl(Moschetti et al. 2005).
Plant nodulation tests
The strains were evaluated for their symbiotic potentialwith broad bean (Vicia faba L. var. major Harz., cv.‘Dulce’) and common vetch (Vicia sativa L). The strainswere grown on yeast mannitol (YM) medium (Vincent1970) at 28°C for 3 days in a rotary shaker at 150 revmin−1. Strain cultures were harvested at the late exponentialphase of growth. Routine plating on YM agar supplementedwith Congo red (Graham 1969) was performed to confirmpurity. Seeds were surface sterilized by 5 min immersion in3% H2O2 then washed in sterile distilled water. Subse-quently, the broad bean seeds were scarified by 90 minimmersion in 75% H2SO4, washed in sterile distilled waterand incubated in sterile distilled water for 24 h in the dark.The seeds were coated with bacterial suspension (approx-imately 105 cells seed−1) and sowed in standard hydropon-ics (20 seeds per treatment). Plants were grown undercontrolled conditions with a constant temperature of 21°Cand a 12 h day−1 photoperiod, and watered with nitrogen-free nutrient solution (Priefer et al. 2001). At 40 days aftersowing, plants were harvested and effective nodules were
Ann Microbiol

counted. The effectiveness of the nodules for nitrogenfixation was estimated from the pink of the nodules and thedark green of the leaves compared to control plants(without inoculation) (Moschetti et al. 2005). All experi-ments were performed in triplicate.
Study site and experimental treatments
The research was carried out in Southern Italy at theUniversity of Naples experimental farm (40° 31′N, 14°58′E) on a clay loam soil that was irrigated with salinewater during the summer since 1988 (De Pascale et al.2005).
Soil physical-chemical and hydrologic properties at thebeginning of the experiment were: 42% sand; 27% loamand 31% clay content; 1.57% organic matter; 0.09% totalN; 7.1 pH; water content of 0.35 cm3 cm−3 at field capacity(in situ) and 0.17 cm3 cm−3 at −1.5 MPa. Salinitytreatments consisted of five NaCl concentrations (weight/volume) in the irrigation water [NSC=0%, SW1=0.125%,SW2=0.25%, SW3=0.5%, and SW4=1% equivalent toelectrical conductivities (EC) at 25°C of 0.5, 2.3, 4.4, 8.5and 15.7 dS m−1, respectively]. Saline waters were obtainedby adding commercial sea salt (Na+ 12.3, K+ 3.8, Ca2+
0.02, Mg2+ 0.04, Cl− 14.4, SO42− 0.03 mol kg−1) to the
irrigation water (Na+ 0.53, K+ 0.05, Ca2+ 1.55, Mg2+ 0.84,Cl− 0.38, SO4
2− 0.15, HCO3− 4.73 mol m−3). Addition of
commercial sea salt (instead of pure NaCl) allowed us toclosely reproduce the sea water contamination of irrigationwater that often occurs in the coastal areas of southern Italy(De Pascale et al. 2003a, 2003b).
When the long-term salt tolerance assessment projectoriginally started, the experimental layout was a ran-domized block design with three replicates. Each blockincluded five treatments: NSC, SW1, SW2, SW3 andSW4; the average electrical conductivity of thesaturated-soil extract (ECe) in the 0–0.6 m soil layer atthe beginning of the experiment was 1.9 dS m−1. Since theobjective of this study was to investigate the long-termeffects of salinization, the salinity treatments, randomlyassigned within each block when soil salinization wasbegun, were reassigned to the same experimental fieldplots in subsequent years. Therefore, from the begin-ning, each experimental plot of 100 m2 has receivedirrigation water with the same salinity (Maggio et al.2004). After 16 years of recurrent summer irrigations withsaline waters, during the fall/winter period, the effectsof residual soil salinity were evaluated on two rain-fedleguminous crops (broad bean and common vetch)inoculated with salt-tolerant and salt-sensitive strains ofRhizobium leguminosarum biovar viciae. The irrigatedsummer crop that preceded broad bean and common vetchwas tomato.
Seed inoculation in experimental field
The salt-tolerant strain SAAN1 (I1) and the salt sensitivestrain POHY2B1 (I2) of R. leg. bv. viciae, were inoculatedon seeds of broad bean and common vetch. The strainswere grown as described above. Seeds were coated withliquid inoculants applied at 3.0 ml kg−1 seed (approximately105 bacteria seed−1) or with a physiological solution(negative control, I0). In winter (7 December), 24 h afterinoculation, seeds were sowed in rows 0.8 m apart to reach adensity of 9 seeds m−2 of broad bean and 180 seeds m−2 ofcommon vetch. At 2 days after sowing, plots were irrigatedwith cell suspension (in physiological solution) of SAAN1,POHY2B1 (105 cells ml−1) or with physiological solution(negative control).
To ascertain the success of the inoculums, a total of 20pre-treated seeds per treatment were sowed in a growthchamber, and a nodulation test under standard hydroponicswas performed as above described.
Soil measurements
Before sowing and also once a month along the crop cycle,inter-row bulk soil samples (0.4 m from the plant) weretaken from each plot (6 per treatment per 6 months)considering the 0–0.4 m soil layer for electrical conductiv-ity (ECe) and pH measurements of the saturated-soil extract(Kalra and Maynard 1991). On soil samples (after drying at60°C), total nitrogen was determined using the Kjeldhalmethod, and N-NO3 and N-NH4 were measured by HACHDR/2000 Spectrophotometer at 500 nm and 425 nm,respectively. Before sowing and after harvesting, soilaggregate stability was determined by using the wet sievemethod developed by Kemper and Rosenau (1986). Soilaggregate stability was expressed as the mean weightdiameter of the water-stable aggregates (MWD-WSAs). Atthe same time, soil bulk density (Bd) was measured onundisturbed soil core collected using 100 cm3 metalcylinder samplers. Soil cores were oven-dried at 105°Cfor 24 h and weighed.
Data were analyzed by ANOVA using a factorial design(species x residual soil salinity x strain) with threereplications and means were separated using the LSD test.
Plant measurements
In spring, at flowering stage (11 Ma), plants were cut at thesoil surface, counted and weighed. Fresh and dry mass(after oven-drying at 60°C) yield were measured.
Yield response to salinity was evaluated according to theMaas and Hoffman threshold-slope model Y=100−s(ECe−T), where Y is the relative yield expressed as percentage(%); s the slope [yield reduction (%) per unit increase in
Ann Microbiol

salinity above T], ECe the time-weighted average electricalconductivity of the soil saturation extract taken from theroot zone (0–0.4 m soil profile) (dS m−1); T thethreshold, i.e., the maximum soil salinity that does notreduce yield below that obtained under non-saline con-ditions (Maas and Hoffman 1977).
Plant samples were dried and ground, and their Ncontent was determined by the Kjeldhal method and usinga HACH DR/2000 Spectrophotometer.
NaCl tolerance of rhizobial isolates
Forty root nodules, located in the first 5 cm of the primaryroot, were excised randomly from 2-month-old broad beanand common vetch plants (20 plants per species), andrhizobia were isolated from fresh nodules by the “Hotelisolation method” (Vincent 1970). Single colonies werepicked and checked for purity by repeated streaking on YMagar medium supplemented with Congo red. All isolateswere incubated at 28°C and kept at −20°C in YM brothwith 25% (v/v) glycerol until analysis.
The rhizobial isolates were tested for their tolerance tosalinity on YM agar medium and in liquid culturessupplemented with NaCl at concentrations of 1%, 2%,3%, 4% and 5% (w/v). The standard YM medium with0.1% (w/v) NaCl was used as control. Plates and tubes wereinoculated with 10 μl bacterial suspension at a concentra-tion of approximately 107 cells ml−1. The measurementsconsisted of visual observation of the diameter of thecolonies after 7 and 14 days, and the degree of turbidityafter 2 and 5 days of incubation at 28°C. These tests werecarried out in triplicate.
RAPD fingerprinting and statistical analysis
Total genomic DNA from the isolates was extracted usingInstaGene Matrix (Bio-Rad Laboratories, Hercules, CA)according to the manufacturer’s instructions. RAPD-PCRwas performed in a total volume of 25 μl containing 3 μl(approximately 20 ng) bacterial DNA, 1 × buffer (Invi-trogen, Carlsbad, CA), 3.5 mM l−1 MgCl2, 0.4 mM l−1 ofeach of the four dNTPs, 0.6 μM l−1 primer and 2.5 U Taqpolymerase (Invitrogen, Carlsbad, CA). The 10-mer ran-dom primers used for genomic strain typing were CC1 (5′-AGC AGC GTG G -3′) and PRIMM239 (5′-CTG AAGCGG A -3′), with 70% and 80% G-C content, respectively(Moschetti et al. 2005). The amplifications were carried outin a PTC-100 thermocycler (MJ Research, Watertown, MA)and template DNAwas denatured for 1 min at 94°C; thenthe PCR was carried out for 40 cycles (1 min at 94°C,1 min at 31°C, 2 min at 72°C, for each cycle). Finally, a 7-min extension period at 72°C was performed. Amplifiedproducts were resolved on 2% and 1.5% (w/v) agarose-
TBE gel electrophoresis, respectively. The 1 Kb Plus DNALadder (Invitrogen) was used as molecular weight marker.The reference strains were used as controls.
Bands were detected automatically using the softwarePhoretix 1 advanced version 3.01 (Phoretix International,Newcastle upon Tyne, UK).
In order to describe the rhizobial population in theexperimental field (Ventorino et al. 2007), the richness,diversity and dominance indexes were calculated using thefollowing formulae:
(1) Margalef ’s index of genotypes richness (R):
R ¼ S� 1ð Þ= lnNð Þwhere S is the number of genotypes and N is thenumber of isolates collected.
(2) Shannon-Weaver index of diversity (H′):
H0 ¼ �ΣPi lnPi
where Pi is the relative abundance of each speciescalculated according to the following equation:
Pi ¼ ni=N
where ni is the number of each genotype and N thetotal number of genotypes.
(3) Simpson index of dominance (D):
D ¼ ΣPi2
Results
Soil
Total rainfall during the broad bean and common vetchgrowing season reached 647 mm, and the average dailytemperature ranged between 6 and 17°C.
After harvest of the tomato crop (in September), the ECein the 0–0.4 m soil layer ranged between 2.3 (NSC) and 9.6dS m−1 (SW4). Mean ECe values decreased throughout thefall–winter season and, by the end of the legumes growthperiod, rainfall was sufficient to reduce the ECe in the 0–0.4 soil layer to 1.8 and 5.9 dS m−1, in NSC and SW4,respectively (data not shown). Before transplanting, the soilpH in the 0–0.4 m soil layer ranged between 6.8 (NSC) and8.0 (SW4), it was generally higher in the higher salinitytreatments and remained rather stable during the wintergrowth cycle (data not shown).
On average, time-weighted ECe and pH during thegrowing season were not affected by either the crop orthe inoculation treatment, whereas they were bothaffected significantly by the residual soil salinity(Table 1).
Ann Microbiol

The diameter of water-stable aggregates (MWD-WSA)in the 0–0.4 m soil layer decreased strongly at increasingresidual soil salinity, showing a 39% reduction in SW4 ascompared to NSC (Fig. 1). MWD-WSAs increased fromfall (September, before transplanting) to spring (May, theend of the experiment) from 0.79 to 0.83 mm, thanks to thereduction in soil salinity due to the leaching rainfalls. As aconsequence of degradation of structural stability, the bulkdensity increased (from 1.23 to 1.38 t m−3) at increasingresidual soil salinity (Table 1).
Soil total nitrogen content was not affected by crop orinoculation treatments, yet it decreased slightly in SW3 andSW4 (Table 1).
Yield
According to Guiducci et al. (2004), both fresh and dryyield were on average higher in common vetch than in
broad bean. Moreover, the total nitrogen accumulated bycrop was 239 kg ha−1 in vetch compared to 118 kg ha−1 inbroad bean (Table 2).
Yield was affected more by the residual soil salinity innon-irrigated broad-bean than in common vetch (Table 2).The fresh yield of common vetch in SW1, SW2 and SW3treatments (30.4 t ha−1, on average) was not significantlydifferent from the non-salinized control treatment (31.0 tha−1), whereas it was 19% smaller in SW4 (Table 2). Thedry mass yield of common vetch resembled the patternobserved for the fresh yield with a significant 20% decreasein SW4 relative to the control (Table 2). In contrast, theyield of broad bean was reduced markedly in terms of bothfresh and dry plant weight, which were 67 and 74% less,respectively, in SW4 compared to NSC (Table 2). Plant drymatter percentage increased with soil salinity in bothspecies, but the differences were very high and statisticallysignificant in broad bean (+5.1% in NSC compared toSW4), while the increase was slight and non significant incommon vetch (+ 2.1% in NSC compared to SW4). Thesedifferences may be due to the well known osmotic effect ofsalinity that reduces water uptake, thus increasing drymatter concentration, in more sensitive plant species(Munns et al. 1995).
The total amount of nitrogen accumulated by the plantswas lower in broad bean than in common vetch withsignificant differences in salinized treatments and itdecreased in broad bean at increasing soil salinity from215 kg ha−1 (NSC) to 60 kg ha−1 (SW4) (Table 2).
The interaction between strain and soil salinity treatmentsrevealed a positive effect of the inoculation on dry mass yieldonly in the non-salinized control (Fig. 2a). A significantdecrease in dry mass yield was observed in SW1 plantsinoculated with SAAN1 (I1) and POHY2B1 (I2), whereasthe dry mass yield of non-inoculated plots (I0), remainedrelatively constant at increasing soil salinity. Therefore, the
Table 1 Time-weighted average electrical conductivity (ECe) and pH,bulk density (Bd) and total N content (Ntot) in the 0–0.4 m soil layeras affected by the two crops common vetch and broad bean. LSDLeast significant difference
pH ECe Bd NtotdS m-1 t m-3 ‰
Cropa (A)
Broad bean 7.36 3.86 1.30 0.99
Common vetch 7.44 3.58 1.31 0.95
ns ns ns ns
Residual soil salinityb (B)
NSC 6.80 1.80 1.23 0.98
SW1 7.20 2.88 1.26 0.98
SW2 7.32 3.55 1.31 0.98
SW3 7.63 4.45 1.34 0.97
SW4 8.05 5.92 1.38 0.96
LSD 0.09 0.35 0.02 0.04
Strainc (C)
I0 7.38 3.57 1.30 0.97
I1 7.41 3.78 1.31 0.96
I2 7.40 3.82 1.30 0.99
ns ns ns ns
Int. AxB ns ns ns ns
Int. BxC ns ns ns ns
Int. AxC ns ns ns ns
Int. AxBxC ns ns ns ns
* P<0.05 , ** P< 0.01, ns non significanta Broad bean (Vicia faba L. var. major Harz., cv. ‘Dulce’) and commonvetch (Vicia sativa L.)b NSC=0%, SW1=0.125%, SW2=0.25%, SW3=0.5%, and SW4=1% NaCl (weight/volume) in the irrigation water during summerc I0, negative control; I1, R. leg. bv. viciae SAAN1; R. leg. bv. viciaePOHY2B1
y = -0.101 x + 1.18
R2 = 0.931
0,0
0,2
0,4
0,6
0,8
1,0
1,2
0 1 2 3 4 5 6 7
Time-weighted average ECe (dS m-1)
MD
W
Fig. 1 Mean weight diameter of water-stable aggregates (MWDWSAs) at increasing soil salinity (ECe). Each point is the mean of 72samples±standard errors
Ann Microbiol
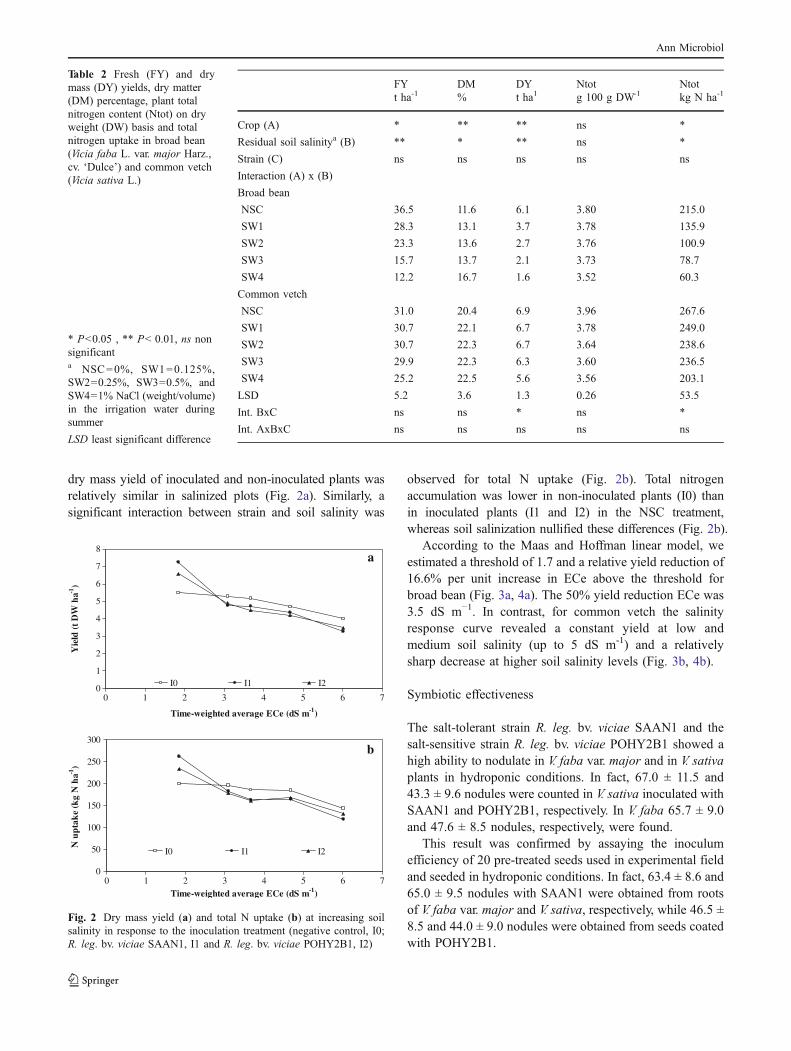
dry mass yield of inoculated and non-inoculated plants wasrelatively similar in salinized plots (Fig. 2a). Similarly, asignificant interaction between strain and soil salinity was
observed for total N uptake (Fig. 2b). Total nitrogenaccumulation was lower in non-inoculated plants (I0) thanin inoculated plants (I1 and I2) in the NSC treatment,whereas soil salinization nullified these differences (Fig. 2b).
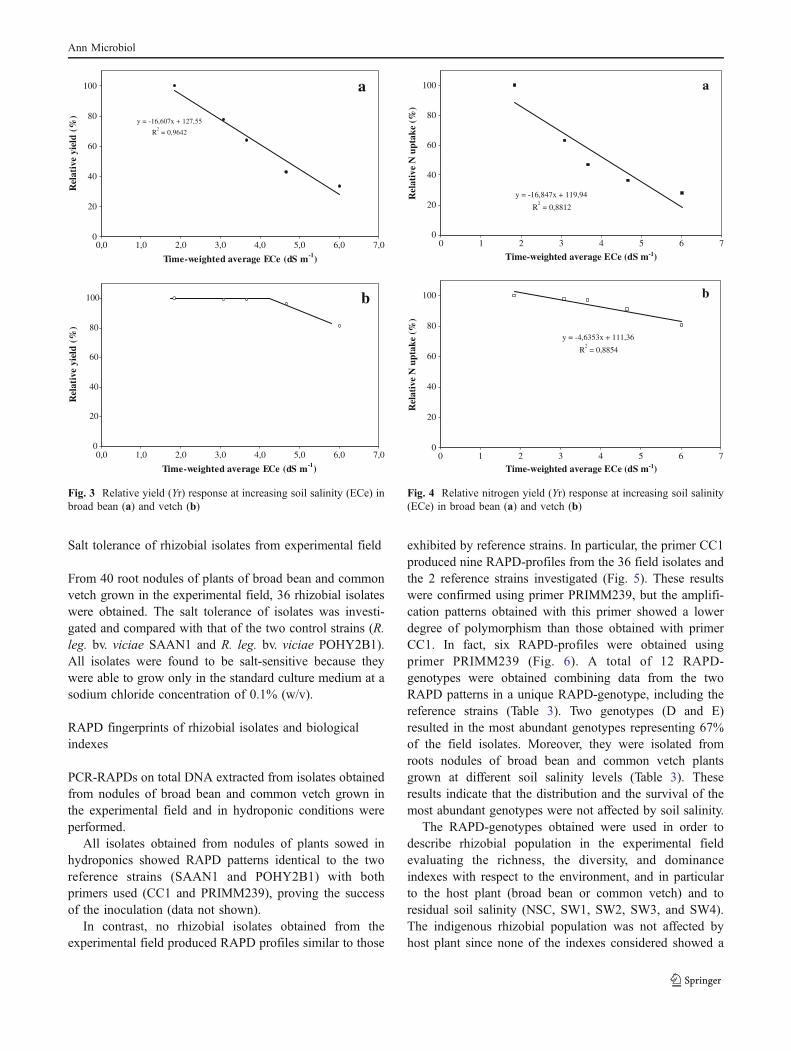
According to the Maas and Hoffman linear model, weestimated a threshold of 1.7 and a relative yield reduction of16.6% per unit increase in ECe above the threshold forbroad bean (Fig. 3a, 4a). The 50% yield reduction ECe was3.5 dS m−1. In contrast, for common vetch the salinityresponse curve revealed a constant yield at low andmedium soil salinity (up to 5 dS m-1) and a relativelysharp decrease at higher soil salinity levels (Fig. 3b, 4b).
Symbiotic effectiveness
The salt-tolerant strain R. leg. bv. viciae SAAN1 and thesalt-sensitive strain R. leg. bv. viciae POHY2B1 showed ahigh ability to nodulate in V. faba var. major and in V. sativaplants in hydroponic conditions. In fact, 67.0 ± 11.5 and43.3 ± 9.6 nodules were counted in V. sativa inoculated withSAAN1 and POHY2B1, respectively. In V. faba 65.7 ± 9.0and 47.6 ± 8.5 nodules, respectively, were found.
This result was confirmed by assaying the inoculumefficiency of 20 pre-treated seeds used in experimental fieldand seeded in hydroponic conditions. In fact, 63.4 ± 8.6 and65.0 ± 9.5 nodules with SAAN1 were obtained from rootsof V. faba var. major and V. sativa, respectively, while 46.5 ±8.5 and 44.0 ± 9.0 nodules were obtained from seeds coatedwith POHY2B1.
0
1
2
3
4
5
6
7
8
Time-weighted average ECe (dS m-1)
Time-weighted average ECe (dS m-1)
Yie
ld (
t D
W h
a-1)
I0 I1 I2
0
50
100
150
200
250
300
0 1 2 3 4 5 6 7
0 1 2 3 4 5 6 7
N u
pta
ke
(kg
N h
a-1)
I0 I1 I2
a
b
Fig. 2 Dry mass yield (a) and total N uptake (b) at increasing soilsalinity in response to the inoculation treatment (negative control, I0;R. leg. bv. viciae SAAN1, I1 and R. leg. bv. viciae POHY2B1, I2)
FY DM DY Ntot Ntott ha-1 % t ha1 g 100 g DW-1 kg N ha-1
Crop (A) * ** ** ns *
Residual soil salinitya (B) ** * ** ns *
Strain (C) ns ns ns ns ns
Interaction (A) x (B)
Broad bean
NSC 36.5 11.6 6.1 3.80 215.0
SW1 28.3 13.1 3.7 3.78 135.9
SW2 23.3 13.6 2.7 3.76 100.9
SW3 15.7 13.7 2.1 3.73 78.7
SW4 12.2 16.7 1.6 3.52 60.3
Common vetch
NSC 31.0 20.4 6.9 3.96 267.6
SW1 30.7 22.1 6.7 3.78 249.0
SW2 30.7 22.3 6.7 3.64 238.6
SW3 29.9 22.3 6.3 3.60 236.5
SW4 25.2 22.5 5.6 3.56 203.1
LSD 5.2 3.6 1.3 0.26 53.5
Int. BxC ns ns * ns *
Int. AxBxC ns ns ns ns ns
Table 2 Fresh (FY) and drymass (DY) yields, dry matter(DM) percentage, plant totalnitrogen content (Ntot) on dryweight (DW) basis and totalnitrogen uptake in broad bean(Vicia faba L. var. major Harz.,cv. ‘Dulce’) and common vetch(Vicia sativa L.)
* P<0.05 , ** P< 0.01, ns nonsignificanta NSC=0%, SW1=0.125%,SW2=0.25%, SW3=0.5%, andSW4=1% NaCl (weight/volume)in the irrigation water duringsummer
LSD least significant difference
Ann Microbiol
Salt tolerance of rhizobial isolates from experimental field
From 40 root nodules of plants of broad bean and commonvetch grown in the experimental field, 36 rhizobial isolateswere obtained. The salt tolerance of isolates was investi-gated and compared with that of the two control strains (R.leg. bv. viciae SAAN1 and R. leg. bv. viciae POHY2B1).All isolates were found to be salt-sensitive because theywere able to grow only in the standard culture medium at asodium chloride concentration of 0.1% (w/v).
RAPD fingerprints of rhizobial isolates and biologicalindexes
PCR-RAPDs on total DNA extracted from isolates obtainedfrom nodules of broad bean and common vetch grown inthe experimental field and in hydroponic conditions wereperformed.
All isolates obtained from nodules of plants sowed inhydroponics showed RAPD patterns identical to the tworeference strains (SAAN1 and POHY2B1) with bothprimers used (CC1 and PRIMM239), proving the successof the inoculation (data not shown).
In contrast, no rhizobial isolates obtained from theexperimental field produced RAPD profiles similar to those
exhibited by reference strains. In particular, the primer CC1produced nine RAPD-profiles from the 36 field isolates andthe 2 reference strains investigated (Fig. 5). These resultswere confirmed using primer PRIMM239, but the amplifi-cation patterns obtained with this primer showed a lowerdegree of polymorphism than those obtained with primerCC1. In fact, six RAPD-profiles were obtained usingprimer PRIMM239 (Fig. 6). A total of 12 RAPD-genotypes were obtained combining data from the twoRAPD patterns in a unique RAPD-genotype, including thereference strains (Table 3). Two genotypes (D and E)resulted in the most abundant genotypes representing 67%of the field isolates. Moreover, they were isolated fromroots nodules of broad bean and common vetch plantsgrown at different soil salinity levels (Table 3). Theseresults indicate that the distribution and the survival of themost abundant genotypes were not affected by soil salinity.
The RAPD-genotypes obtained were used in order todescribe rhizobial population in the experimental fieldevaluating the richness, the diversity, and dominanceindexes with respect to the environment, and in particularto the host plant (broad bean or common vetch) and toresidual soil salinity (NSC, SW1, SW2, SW3, and SW4).The indigenous rhizobial population was not affected byhost plant since none of the indexes considered showed a
y = -16,847x + 119,94
R2 = 0,8812
0
20
40
60
80
100
Time-weighted average ECe (dS m-1)
Rel
ativ
e N
upt
ake
(%)
y = -4,6353x + 111,36
R2 = 0,8854
0
20
40
60
80
100
0 1 2 3 4 5 6 7
0 1 2 3 4 5 6 7
Time-weighted average ECe (dS m-1)
Rel
ativ
e N
upt
ake
(%)
a
b
Fig. 4 Relative nitrogen yield (Yr) response at increasing soil salinity(ECe) in broad bean (a) and vetch (b)
y = -16,607x + 127,55
R2 = 0,9642
0
20
40
60
80
100
Time-weighted average ECe (dS m-1)
Rel
ativ
e yi
eld
(%)
0
20
40
60
80
100
0,0 1,0 2,0 3,0 4,0 5,0 6,0 7,0
0,0 1,0 2,0 3,0 4,0 5,0 6,0 7,0
Time-weighted average ECe (dS m-1)
Rel
ativ
e yi
eld
(%)
a
b
Fig. 3 Relative yield (Yr) response at increasing soil salinity (ECe) inbroad bean (a) and vetch (b)
Ann Microbiol

significant difference in rhizobial population. In fact, therichness index (R), the general index of biodiversity (H’)and the concentration of dominance (D) were identical forbroad bean and common vetch (Table 4). By contrast, thediversity of the natural rhizobia varied with soil salttreatment. In particular, R and H’ increased in broad beanat increasing residual soil salinity ranging between 0 (NSC)and 1.9 or 1.3 (SW3), while it decreased in treatment SW4(Table 4). Moreover, the highest value of microbialbiodiversity was found to be related to the lowestconcentration of dominance in all SW treatments (SW1,SW2, SW3, and SW4). Conversely, in the NSC plots, thelowest value of the diversity index (0) was associated witha higher value of the dominance index (1) (Table 4).
In common vetch, R, H’ and D values did not differ for thefirst three salinity treatments (NSC, SW1 and SW2), whilemicrobial biodiversity increased in SW3 and SW4 (Table 4).
Discussion
After recurrent summer irrigations with saline waters,the SW4 soil presented the characteristics typical of analkaline–saline soil with high soil pH, salt build-upalong the soil profile, increased bulk density and
reduced soil aggregate stability. These results indicatethat increased ECe is only one component of a morecomplex scenario, which includes a high sodiumadsorption ratio, low hydraulic conductivity to water,surface seals, water logging and reduced root zoneaeration (De Pascale et al. 2005; Maggio et al. 2005).These salt-induced modifications of soil physicochemicalproperties may further aggravate the effects of salinizationon crop yield (Emerman and Dawson 1996; Hachicha etal. 2000). In our experiment, growth and yield of a non-irrigated broad bean crop were seriously compromised byresidual soil salinity even during the fall–winter season. Incontrast, soil salinity significantly reduced plant growthand yield of common vetch only in the highest residualsoil salinity treatment. Threshold (1.7 dS m−1) and slope[16.6% (dS m−1)−1] values, estimated for broad beanaccording to Maas and Hoffman (1977), were close tothose found by De Pascale and Barbieri (1997) under
Fig. 6 Randomly amplified polymorphic DNA patterns generated byprimer PRIMM239 of four representative Rhizobium isolates and tworeference strains of R. leg. bv. viciae. Lanes: a reference strainSAAN1, b reference strain POHY2B1, c pattern c, d pattern e, epattern d, f pattern f,M 1 kb Plus DNA Ladder (Invitrogen)
Fig. 5 Randomly amplified polymorphic DNA patterns generated byprimer CC1 of seven representative Rhizobium isolates and tworeference strains of R. leg. bv. viciae. Lanes: a Reference strainSAAN1, b reference strain POHY2B1, c pattern D, d pattern H, epattern E, f pattern I, g pattern C, h pattern F, i pattern G, M 1 kb PlusDNA Ladder (Invitrogen)
Ann Microbiol

similar experimental conditions, but were lower andhigher, respectively, than those reported by other authorsfor the same plant species (Katerji et al. 1992). The ECe50for broad bean was about 60% lower than the correspondingvalue of 8.2 dS m−1 reported by Maas and Hofmann, thusconfirming that, in long-term salinized fields, permanentmodifications of soil physico-chemical properties may reducecrop tolerance to soil salinity and eventually anticipate the onsetof the salinity tolerance threshold (De Pascale et al. 2005; Maasand Grattan 1999). Hyper-osmotic stress, hypoxia, and nutrientimbalance often coexist in soils exposed to long-termsalinization and, if they persist throughout the fall–winterseason, may also modify salt tolerance function of non-irrigatedcrops (Page et al. 1990; Shainberg and Singer 1990).
Information about salt resistant vetch species is scarce(UC SAREP 2008). In a 30-day experiment, seedlings ofcommon vetch showed an increase in fresh weight withincreasing water salinity up to 1.35 dS m−1 and a decreaseat water salinity levels higher than 2.70 dS m−1, whereasdry weight was not affected by salinity (Orak and Ates2005). Under our experimental conditions, field growncommon vetch showed higher salinity tolerance up to ECeof 5 dS m−1 and a sharp decrease in fresh yield at highersoil salinity levels. Although it is very difficult to comparefield results obtained throughout a complete crop cycle witha short-term experiment performed on seedlings undercontrolled conditions, the pattern of yield decrease inresponse to salinity that we observed in common vetch
Table 3 RAPD genotypes of rhizobial field isolates and reference strains obtained by using primers CC1 and PRIMM239
Profile CC1a Profile 239b RAPD genotype Isolate numberbroad beanc
Isolate numbercommon vetchd
Residual soilsalinitye
Strainsf
C c C 0 2 SW4 SAAN1
D d D 0 1 SW1 SAAN1
0 1 SW2 SAAN1
1 1 SW3 SAAN1
2 2 SW4 SAAN1
1 0 SW3 POHY2B1
0 1 SW4 POHY2B1
1 0 SW3 NTg
1 0 SW4 NT
D e De 0 1 NSC SAAN1
1 0 SW1 SAAN1
1 0 SW4 NT
D f Df 0 1 SW2 SAAN1
1 0 SW3 SAAN1
E e E 1 1 NSC SAAN1
0 1 SW1 SAAN1
1 2 SW4 SAAN1
1 0 SW2 POHY2B1
1 0 SW3 POHY2B1
1 2 SW4 POHY2B1
1 0 SW4 NT
E d Ed 1 0 SW3 POHY2B1
F e Fe 0 1 SW3 SAAN1
G e Ge 1 0 SW2 SAAN1
H e He 0 1 SW3 SAAN1
I d Id 1 0 SW1 SAAN1
A a A Reference strain R. leg. bv. viciae SAAN1
B b B Reference strain R. leg. bv. viciae POHY2B1
a,b RAPD profile obtained by using CC1 and PRIMM239 primers, respectivelyc,d Number of isolates obtained from root nodules of broad bean and common vetch, respectivelye NSC=0%, SW1=0.125%, SW2=0.25%, SW3=0.5%, and SW4=1% NaCl (weight/volume) in the irrigation water during summerf R. leg. bv. viciae strain inoculated in each plots of experimental siteg No inoculated plot
Ann Microbiol

was consistent with earlier results by Orak and Ates (2005).The complexity of the plant response to salinity may haveled to substantial variations of the salt tolerance threshold.
The observed differences confirm that species within thesame gender may have different tolerance to salt stress (vanHoorn et al. 2001), and that evaluation of crop performanceunder “real” field conditions is therefore important toidentify more tolerant species that could be used to extendthe cultivation of leguminous cover crops in sub-optimalareas. Our results clearly indicate that, as a cover crop,common vetch may increase the availability of nitrogen insoil more that broad bean also in saline environments.
Since rhizobial strains are recognized to play animportant role in the impact of soil management in stressedsoils (i.e., arid, poor, and salinized), the effectiveness andcompetitiveness in an artificial salinized field of twoselected rhizobial strains (SAAN1 and POHY2B1) isolatedfrom Campania region were evaluated. Our results showedthat these strains had a high potential ability to nodulateVicia faba and Vicia sativa plants as demonstrated by thesuccess of inoculum estimated by sowing the pre-treatedseeds in hydroponic conditions. RAPD-PCR of rhizobialisolates confirmed that the nodules contained our referencesstrains. Afterwards, the inoculated seeds were sowed in theexperimental field but the reference strains showed loweffectiveness and competitiveness compared to the naturalrhizobial population. In fact, to obtain a result representing
intra-species relationships among all our field isolates andreference strains, data obtained from two RAPD patternswere combined in a unique RAPD-genotype. Nonetheless,no rhizobial field isolates belonged to the reference strains.In our experimental farm, where legume crops wereintroduced in crop rotation in the 1980s, the inoculantsshowed a lower level of infectivity than the indigenousrhizobial population. As demonstrated by Bogino and co-workers (2006), inoculation of peanut (Arachis ipogea L.)plants with Bradyrhizobium spp. gave better results in fieldswhere peanut was sowed for the first time. In fact, selectedstrains inoculated on legume seeds often fail to occupy asignificant proportion of nodules when an indigenouscompetitor rhizobial population is established in the soil(Bogino et al. 2008; Streeter 1994; Toro 1996; VanDillewijn et al. 2001). The competitive advantage of theendogenous population could result from genetic/physio-logical adaptation over the introduced inoculant or from apositional advantage. In fact, the natural bacterial popula-tion already occupies the soil along the root zone, whereasthe artificially inoculated strains usually remain concentrat-ed around the seeds (López-García et al. 2002).
The Shannon-Weaver index of diversity (H′) combinesthe relative abundance of genotypes and the total specificrichness, thus reflecting the distribution of dominanceamongst the major genotypes present in the field. Thenatural rhizobial population was not affected by the kind ofFabaceae, i.e., V. faba or V. sativa, in the experimental field.Indeed, as showed by biological indexes, no significantdifference in the richness, biodiversity, and dominance wasrecorded with respect to the host plant. The low diversity inthe field was also due to the use of legumes in crop rotationfor several years. These results were in agreement withprevious observations (Ventorino et al. 2007) on thediversity of rhizobial natural populations in the volcanicsoil of the Vesuvian area, which decreased after only 3 yearsas a result of bean monocropping. By contrast, the diversityof the natural rhizobial population varied with respect tosoil salt treatments. In fact, salinity alters the diversity andfunctions of microbial communities in soil. Several studiesalong salinity gradients showed decreasing diversity ofmicrobial communities when the salinity increased (Abed etal. 2007; Rothrock and Garcia-Pichel 2005). Interestingly,this study recorded a lower value of biodiversity index H′ inthe non-salinized control (NSC) than that reported byTrabelsi et al. (2009). In fact, their results showed thatsaline soil had higher diversity (in terms of Shannon index)than non-salinized soil for heterotrophic bacteria. We foundthat, in salinized plots, biodiversity increases compared tothe control, but the increase is not proportional to theamount of residual soil salinity. Interestingly, salt stressshowed a similar effect on rhizobial biodiversity and yieldin common vetch. In fact, the diversity index did not vary
Table 4 Richness index (R), diversity index (H’), and dominanceindex (D) estimated for the two host plants
R H’ D
Host planta
Broad bean 2.1 1.6 0.24
Common vetch 2.1 1.5 0.24
Residual soil salinityb
Broad bean
NSC 0 0 1
SW1 1.5 0.7 0.5
SW2 1.8 1.1 0.3
SW3 1.9 1.3 0.3
SW4 1.1 1 0.4
Common vetch
NSC 1.5 0 0.5
SW1 1.5 0 0.5
SW2 1.5 0 0.5
SW3 1.8 1.1 0.3
SW4 0.9 1 0.4
a Broad bean (Vicia faba L. var. major Harz., cv. ‘Dulce’) andcommon vetch (Vicia sativa L.)b NSC=0%, SW1=0.125%, SW2=0.25%, SW3=0.5%, and SW4=1% NaCl (weight/volume) in the irrigation water during summer
Ann Microbiol

in NSC, SW1, and SW2 treatments as much as the relativeyield obtained in the Maas and Hoffman linear model. Inaddition, the most abundant genotypes identified wererecovered in all salinity treatments and in the control plots,showing that the selection and survival of strains belongingto D and E genotypes in field was not affected by salinity.
After 32 years of salt treatments in this experiment, wewould expect that the rhizobia living in those soils to haveevolved to tolerate salt; instead, the saline-tolerance ofrhizobia in liquid or solid culture showed a differentphenotype then the saline-tolerance in the soil (or insymbiosis). In fact, our rhizobial field isolates were foundto be salt sensitive in culture medium, growing only atchloride concentrations of 0.1% (w/v). These results are inagreement with those of Shamseldin et al. (2009), whoreported that the majority of rhizobial strains isolated fromdifferent soils of Egypt were sensitive to salt stress inculture medium, growing only at 0.2% NaCl. Thus, acomplex interaction between bacteria, plant and soil candetermine the survival of salt-sensitive indigenous rhizobialstrains and limit the success of inoculation of exogenoussalt resistant rhizobia. In fact, levels of salinity that inhibitthe symbiosis between legumes and rhizobia are differentfrom those that inhibit the growth of the individualsymbionts (Shamseldin et al. 2009). Moreover, it has beenshown that salt-resistant mutants of Rhizobium havedeficiencies in their symbiotic capacity (Nogales et al.2002). The response and adaptation to environmentalstresses are complex phenomena involving many physio-logical and biochemical processes that may reflect changesin gene expression and in the activity of enzymes andtransport proteins (Djordjevic et al. 2003; Wei et al. 2004).Unsuccessful symbiosis under salt-stress may be due to afailure in the infection process because of the effect ofsalinity on the establishment of rhizobia (Cordovilla et al.1999) since legumes and process nodule initiation are bothmore sensitive to osmotic stress than rhizobia (Velagaleti etal. 1990). However, further work is needed to investigatethe different phenotypes as saline-tolerance of the indige-nous rhizobial isolates exhibited in vitro and in soil in orderto understand the complex interactions that determine thesurvival of salt-sensitive strains in salinized soils.
In conclusion, the use of common vetch as a cover crop insaline environments may increase the availability of nitrogenin soil more than broad bean. Moreover, the selected rhizobialstrains exhibited lower competitiveness compared to naturalrhizobial population, and the “sustained” production undersalt stress in common vetch was due to combined effect of thepresence of halo-tolerant rhizobia in the field and of theintrinsic salt tolerance of plants.
Acknowledgment This research was supported by a grant from theMinistero delle Politiche Agricole e Forestali (MIPAF, Rome) within
the Programme “Genetic resources of organisms useful for theimprovement of agricultural species and to a sustainable agriculture.”
References
Abed RMM, Kohls K, de Beer D (2007) Effect of salinity changes onthe bacterial diversity, photosynthesis and oxygen consumptionof cyanobacterial mats from an intertidal flat of the Arabian Gulf.Environ Microbiol 9:1384–1392. doi:10.1111/j.1462-2920.2007.01254.x
Amézketa E (1999) Soil aggregate stability: a review. J Sustain Agric14:83–151. doi:10.1300/J064v14n02_08
Bianco C, Defez R (2009) Medicago truncatula improves salttolerance when nodulated by an indole-3-acetic acid-overproducing Sinorhizobium meliloti strain. J Exp Bot60:3097–3107. doi:10.1093/jxb/erp140
Bogino P, Banchio E, Rinaudi L, Cerioni G, Bonfiglio C, Giordano W(2006) Peanut (Arachis hypogaea) response to inoculation withBradyrhizobium sp. in soils of Argentina. Ann Appl Biol148:207–212. doi:10.1111/j.1744-7348.2006.00055.x
Bogino P, Banchio E, Bonfiglio C, Giordano W (2008) Competitive-ness of a Bradyrhizobium sp. strain in soils containing indigenousrhizobia. Curr Microbiol 56:66–72. doi:10.1007/s00284-007-9041-4
Cassman KG, Rains DW (1986) A cropping systems approach tosalinity management in California. Am J Altern Agric 1:115–121
Catroux G, Hartmann A, Revellin C (2001) Trends in rhizobialinoculant production and use. Plant Soil 230:21–30. doi:10.1023/A:1004777115628
Chen WM, Lee TM, Lan CC, Cheng CP (2000) Characterization ofhalotolerant rhizobia isolated from root nodules of Canavaliarosea from seaside areas. FEMS Microbiol Ecol 34:9–16.doi:10.1111/j.1574-6941.2000.tb00749.x
Cordovilla MP, Ligero F, Lluch C (1999) Effects of NaCl on growthand nitrogen fixation and assimilation of inoculated and KNO3
fertilized Vicia faba L. and Pisum sativum L. plants. Plant Sci140:127–136. doi:10.1016/S0168-9452(98)00201-5
De Pascale S, Barbieri G (1997) Effect of soil salinity and top removalon growth and yield of broadbean as green vegetable. Sci Hortic71:147–165. doi:10.1016/S0304-4238(97)00104-0
De Pascale S, Ruggiero C, Barbieri G, Maggio A (2003a) Physiolog-ical response of pepper to salinity and drought. J Am Soc HorticSci 128:48–54
De Pascale S, Maggio A, Ruggiero C, Barbieri G (2003b) Growth,water relation, and ion content of field-grown celery (Apiumgraveolens L. Var. Dulce (Mill.) Pers.) under saline irrigation. JAm Soc Hortic Sci 128:136–143
De Pascale S, Maggio A, Barbieri G (2005) Soil salinization affectsgrowth, yield and mineral composition of cauliflower andbroccoli. Eur J Agron 23:254–264. doi:10.1016/j.eja.2004.11.007
Deaker R, Roughley RJ, Kennedy IR (2004) Legume seed inoculationtechnology—a review. Soil Biol Biochem 36:1275–1288.doi:10.1016/j.soilbio.2004.04.009
Djordjevic MA, Chen HC, Natera S, Van Noorden G, Menzel C,Taylor S, Renard C, Geiger O, Weiller GF (2003) A globalanalysis of protein expression profiles in Sinorhizobium meliloti:discovery of new genes for nodule occupancy and stressadaptation. Mol Plant Microbe Interact 16:508–524.doi:10.1094/MPMI.2003.16.6.508
Emerman SH, Dawson TE (1996) Hydraulic lift and its influence onthe water content of the rhizosphere: an example from sugarmaple, Acer saccharum. Oecologia 108:273–278. doi:10.1007/BF00334651
Ann Microbiol

Fierer N, Schimel JP, Holden PA (2003) Influence of drying-rewettingfrequency on soil bacterial community structure. Microb Ecol45:63–71. doi:10.1007/s00248-002-1007-2
Graham PH (1969) Selective medium for growth of Rhizobium. ApplEnviron Microbiol 17:769
Griffiths RI, Whiteley AS, O’Donnell AG, Bailey MJ (2003)Physiological and community responses of established grasslandbacterial populations to water stress. Appl Environ Microbiol69:6961–6968. doi:10.1128/AEM.69.12.6961-6968.2003
Guiducci M, Bonciarelli U, Stagnari F, Benincasa P (2004) Total Nsupply and profit from several green manure crops. In: Proc ofthe VIII ESA Congress: European agriculture in a global context,Copenhagen, Denmark, 11–15 July, pp 981–982
Hachicha M, Cheverry C, Mhiri A (2000) The impact of long-termirrigation on changes of ground water level and soil salinity innorthern Tunisia. Arid Soil Res Rehab 14:175–182. doi:10.1080/089030600263094
Howieson J, Ballard R (2004) Optimising the legume symbiosis instressful and competitive environments within southern Australia- some contemporary thoughts. Soil Biol Biochem 36:1261–1273. doi:10.1016/j.soilbio.2004.04.008
Jenkins MB (2003) Rhizobial and bradyrhizobial symbionts ofmesquite from the Sonoran Desert: salt tolerance, facultativehalophily and nitrate respiration. Soil Biol Biochem 35:1675–1682. doi:10.1016/j.soilbio.2003.08.014
Kalra YP, Maynard DG (1991) Methods manual for Forest Soil andPlant Analysis. Forestry Canada, Northwest Region, Edmonton,Alberta, Canada. Information Report NOR-X-319
Katerji N, van Hoorn JW, Hamdy A, Bouzid N, El-Sayed Mahrous S,Mastrorilli M (1992) Effect of salinity on water stress, growthand yield of broadbeans. Agric Water Manag 21:107–117.doi:10.1016/0378-3774(92)90086-C
Katerji N, van Hoorn JW, Hamdy A, Mastrorilli M (2003) Salinityeffect on crop development and yield, analysis of salt toleranceaccording to several classification methods. Agric Water Manag62:37–66. doi:10.1016/S0378-3774(03)00005-2
Kemper WD, Rosenau RC (1986) Aggregate stability and sizedistribution. In: Klute A (ed) Methods of soil analysis, part I.Physical and mineralogical methods. Agronomy, No. 9, ASA-SSSA, Madison, pp 425–442
L’taief B, Sifi B, Zaman-Allah M, Drevon JJ, Lachaal M (2007) Effectof salinity on root-nodule conductance to the oxygen diffusion inthe Cicer arietinum–Mesorhizobium ciceri symbiosis. J PlantPhysiol 164:1028–1036. doi:10.1016/j.jptph.2006.05.016
Lloret J, Bolaños L, Lucas MM, Peart JM, Brewin NJ, Bonilla I,Rivilla R (1995) Ionic stress and osmotic pressure inducedifferent alterations in the lipopolysaccharide of a Rhizobiummeliloti strain. Appl Environ Microbiol 61:3701–3704
Lloret J, Wulff BBH, Rubio JM, Downie JA, Bonilla I, Rivilla R(1998) Exopolysaccharide II production is regulated by salt in thehalotolerant strain Rhizobium meliloti EFB1. Appl EnvironMicrobiol 64:1024–1028
López-García SL, Vázquez TE, Favelukes G, Lodeiro AR (2002)Rhizobial position as a main determinant in the problem ofcompetition for nodulation in soybean. Environ Microbiol 4:216–224. doi:10.1046/j.1462-2920.2002.00287.x
Lu YC, Watkins KB, Teasdale JR, Abdul-Baki AA (2000) Cover cropsin sustainable food production. Food Rev Int 16:121–157.doi:10.1081/FRI-100100285
Maas EV, Grattan SR (1999) Crop yield as affected by salinity. In:Skaggs RW, van Schilfgaarde J (eds) Agricultural drainage.Agronomy monograph 38, ASA-SSSA, Madison, pp 55–110
Maas EV, Hoffman GJ (1977) Crop salt tolerance. J Irrig Drainage Div103:115–134
MacRae RJ, Mehuys GR (1985) The effect of green manuring on thephysical properties of temperate area soils. Adv Soil Sci 3:71–94
Maggio A, De Pascale S, Angelino G, Ruggiero C, Barbieri G (2004)Physiological response of tomato to saline irrigation in long-termsalinized soils. Eur J Agron 21:149–159. doi:10.1016/S1161-0301(03)00092-3
Maggio A, De Pascale S, Ruggiero C, Barbieri G (2005) Physiologicalresponse of field-grown cabbage to salinity and drought stress.Eur J Agron 23:57–67. doi:10.1016/j.eja.2004.09.004
Miller KJ, Wood JM (1996) Osmoadaptation by rhizospherebacteria. Annu Rev Microbiol 50:101–136. doi:10.1146/annurev.micro.50.1.101
Mitchell JP, Shennan C, Singer MJ, Peters DW, Miller RO, Prichard T,Grattan SR, Rhoades JD, May DM, Munk DS (2000) Impacts ofgypsum and winter cover crops on soil physical properties andcrop productivity when irrigated with saline water. Agric WaterManag 45:55–71. doi:10.1016/S0378-3774(99)00070-0
Moschetti G, Peluso AL, Protopapa A, Anastasio M, Pepe O, Defez R(2005) Use of nodulation pattern, stress tolerance, nodC geneamplification, RAPD-PCR and RFLP-16S rDNA analysis todiscriminate genotypes of Rhizobium leguminosarum biovarviciae. Syst Appl Microbiol 28:619–631. doi:10.1016/j.syapm.2005.03.009
Munns R, Schachtman DP, Condon AG (1995) The Significance of aTwo-Phase Growth Response to Salinity in Wheat and Barley.Aust J Plant Physiol 22:561–569. doi:10.1071/PP9950561
Nogales J, Campos R, BenAbdelkhalek H, Olivares J, Llunch C,Sanjuan J (2002) Rhizobium tropici genes involved in free-livingsalt tolerance are required for the establishment of efficientnitrogen-fixing symbiosis with Phaseolus vulgaris. Mol PlantMicrobe Interact 15:225–232. doi:10.1094/MPMI.2002.15.3.225
Ogutcu H, Kasimoglu C, Elkoca E (2010) Effects of rhizobium strainsisolated from wild chickpeas on the growth and symbioticperformance of chickpeas (Cicer arietinum L.) under salt stress.Turk J Agric For 34:361–371. doi:10.3906/tar-0904-34
Orak A, Ates E (2005) Resistance to salinity stress and available waterlevels at the seedling stage of the common vetch (Vicia sativa L.).Plant Soil Environ 51:51–56
Page AL, Chang AC, Adriano DC (1990) Deficiencies and toxicitiesof trace elements. In: Tanji KK (ed) Agricultural salinityassessment and management. Manual and Report EngineeringPractice 71, American Society of Civil Engineers, New York,pp 138–160
Parida SK, Das AB (2005) Salt tolerance and salinity effects on plants.Ecotoxicol Environ Safety 60:324–349. doi:10.1016/j.ecoenv.2004.06.010
Perret X, Staehelin C, Broughton WJ (2000) Molecular basis ofsymbiotic promiscuity. Microbiol Mol Biol Rev 64:180–201.doi:1092-2172/00/$04.0010
Priefer UB, Auag J, Boesten B, Bouhmouch I, Defez R, Filali-MaltoufA, Miklis M, Moawad H, Mouhsne B, Prell J, Schluter A,Senatore B (2001) Characterization of Phaseolus symbiontsisolated from Mediterranean soils and analysis of genetic factorsrelated to pH tolerance. J Biotechnol 91:223–236. doi:10.1016/S0168-1656(01)00329-7
Rothrock MJ, Garcia-Pichel F (2005) Microbial diversity of benthicmats along a tidal desiccation gradient. Environ Microbiol7:593–601. doi:10.1111/j.1462-2920.2004.00728.x
Shainberg I, Singer MJ (1990) Soil response to saline and sodicconditions. In: Tanji KK (ed) Agricultural Salinity Assessmentand Management. Manual and Report Engineering Practice.American Society of Civil Engineers, New York 71:91–112
Shamseldin A, El-Sadani M, Sadowsky MJ, Sun An C (2009) Rapididentification and discrimination among Egyptian genotypes ofRhizobium leguminosarum bv. viciae and Sinorhizobium melilotinodulating faba bean (Vicia faba L.) by analysis of nodC,ARDRA, and rDNA sequence analysis. Soil Biol Biochem41:45–53. doi:10.1016/j.soilbio.2008.09.014
Ann Microbiol

Soussi M, Santamarìa M, Ocaña A, Lluch C (2001) Effects of salinityon protein and lipopolysaccharide pattern in a salt-tolerant strainof Mesorhizobium ciceri. J Appl Microbiol 90:476–481.doi:10.1046/j.1365-2672.2001.01269.x
Spaink HP (2000) Root nodulation and infection factors produced byrhizobial bacteria. Annu Rev Microbiol 54:257–288.doi:10.1146/annurev.micro.54.1.257
Streeter JG (1994) Failure of inoculant rhizobia to overcome thedominance of indigenous strains for nodule formation. Can JMicrobiol 40:513–522. doi:10.1139/m94-084
Toro N (1996) Nodulation competitiveness in the Rhizobium-legumesymbiosis. World J Microbiol Biotechnol 12:157–162.doi:10.1007/BF00364680
Trabelsi D, Mengoni A, Elarbi Aouani M, Mhamdi R, Bazzicalupo M(2009) Genetic diversity and salt tolerance of bacterial commu-nities from two Tunisian soils. Ann Microbiol 59:25–32
UC SAREP (2008) On line cover crop database. <http://www.sarep.ucdavis.edu/cgi-bin/ccrop.EXE/show_crop_14> (30 April 2008)
Upadhyay SK, Singh JS, Singh DP (2011) Exopolysaccharide-producingplant growth-promoting rhizobacteria under salinity condition.Pedosphere 21:214–222. doi:10.1016/S1002-0160(11)60120-3
Van Dillewijn P, Soto MJ, Villadas PJ, Toro N (2001) Construction andenvironmental release of a Sinorhizobium meliloti strain geneti-cally modified to be more competitive for alfalfa nodulation.Appl Environ Microbiol 67:3860–3865. doi:10.1128/AEM.67.9.3860-3865.2001
van Hoorn JW, Katerji N, Hamdy A, Mastrorilli M (2001) Effect ofsalinity on yield and nitrogen uptake of four grain legumes andon biological nitrogen contribution from the soil. Agric WaterManag 51:87–98. doi:10.1016/S0378-3774(01)00114-7
Velagaleti RR, Marsh S, Kramer D (1990) Genotypic differencesin growth and nitrogen fixation among soybean (Glycine max(L.) Merr.) cultivars grown under salt stress. Trop Agric67:169–177
Ventorino V, Chiurazzi M, Aponte M, Pepe O, Moschetti G (2007)Genetic diversity of a natural population of Rhizobium legumi-nosarum bv. viciae nodulating plants of Vicia faba in theVesuvian area. Curr Microbiol 55:512–517. doi:10.1007/s00284-007-9024-5
Vincent JM (1970) A manual for the practical study of root nodulebacteria. Blackwell Scientific, Oxford
Wei W, Jiang J, Li W, Wang L, Yang SS (2004) Isolation of salt-sensitive mutants from Sinorhizobium meliloti and characteriza-tion of genes involved in salt tolerance. Lett Appl Microbiol39:278–283. doi:10.1111/j.1472-765X.2004.01577.x
Yan AM, Wang ET, Kan FL, Tan ZY, Sui XH, Reinhold-Hurek B, ChenWX (2000) Sinorhizobium meliloti associated with Medicagosativa and Melilotus spp. in arid saline soils in Xinjiang, China.Int J Syst Evol Microbiol 50:1887–1891
Zahran HH (1999) Rhizobium-legume symbiosis and nitrogen fixationunder severe conditions in an arid climate. Microbiol Mol BiolRev 63:968–989
Ann Microbiol




















