
Vleermuizen op de parkbegraafplaats Schoonselhof
59
1 Vleermuizen op de parkbegraafplaats Schoonselhof Johan De Ridder Daniel Sanders 2015
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Vleermuizen op de parkbegraafplaats Schoonselhof

1
Vleermuizen op de
parkbegraafplaats
Schoonselhof
Johan De Ridder
Daniel Sanders
2015

Auteurs:
Johan De Ridder en Daniel Sanders
Contact:
Natuurpunt Zuidrand Antwerpen
Frans De Cortlaan 42
2610 Wilrijk
Afgiftekantoor Wilrijk 1
www.zuidrand.be
e-mail:
Wijze van citeren:
De Ridder, J. & Sanders D. (2015). Vleermuizen op de parkbegraafplaats Schoonselhof.
Rapport Natuurpunt Zuidrand Antwerpen werkgroep Studie 2015/1.
Foto cover:
Johan De Ridder

Vleermuizen op de parkbegraafplaats
Schoonselhof
Zomerinventarisatie, veldwerk en geluidanalyses: Johan De Ridder en Daniel Sanders
Tekst: Johan De Ridder en Daniel Sanders
Eindredactie en vormgeving: Johan De Ridder
Foto’s: Johan De Ridder
Met dank aan Luc De Bruyn (INBO) voor advies bij de statistische verwerking, Michel Barataud
voor controle van een aantal determinaties en Patrick Dictus voor het regelen van de toelating
om het domein te mogen betreden.
opgemaakt te Borgerhout,
1 april 2015
ir. Johan De Ridder

Inhoud 1. Inleiding ................................................................................................................. 1
1.1 Algemeen ......................................................................................................... 1
1.2 Gebiedsbeschrijving ........................................................................................... 2
1.3 Onderzoeksvragen en te verwachten resultaten .................................................... 2
2. Methodiek .............................................................................................................. 3
2.1 Bepalen vlieg- en jachtactiviteit .......................................................................... 3
2.2 Verkenning jachtgebieden, vliegroutes en zwermlocaties ........................................ 4
2.3 Soortdeterminatie aan de hand van ultrasone geluiden .......................................... 5
2.4 Wintertelling ..................................................................................................... 5
2.5 Verwerking en statistische analyse ...................................................................... 6
3. Resultaten .............................................................................................................. 7
3.1 Algemeen ......................................................................................................... 7
3.2 Vliegactiviteit en jachtactiviteit ........................................................................... 8
4. Bespreking en vleermuizenfuncties .......................................................................... 15
4.1 De functie winterverblijfplaats ........................................................................... 16
4.2 De functie zomer-, kraam- en paarverblijfplaats in gebouwen ............................... 16
4.3 De functie zomer-, kraam- en paarverblijfplaats in bomen .................................... 17
4.4 De functie jachtgebied ..................................................................................... 17
4.5 De functie vliegroute/verbindingsfunctie............................................................. 19
4.6 De functie zwermplaats .................................................................................... 20
4.7 Relatie tussen Fort 7 en begraafplaats Schoonselhof ............................................ 21
5. Aanbevelingen m.b.t. beheer .................................................................................. 22
5.1 Algemeen ....................................................................................................... 22
5.2 Dreven en bossen ........................................................................................... 24
5.3 Vijvers en grachten ......................................................................................... 27
5.4 Beheermaatregelen voor behoud en versterken habitat van laatvlieger .................. 28
5.5 Beheermaatregelen voor behoud en versterken habitat grootoorvleermuis ............. 29
5.6 Versterken van de connectiviteit met nabijgelegen gebieden ................................ 32
6. Aanbevelingen voor verder onderzoek ...................................................................... 33
7. Literatuur ............................................................................................................. 34
8. Bijlagen ............................................................................................................... 38
8.1 Overzicht van tel-data, tijdstippen en weersomstandigheden ................................ 38
8.2 Figuren .......................................................................................................... 39
8.3 Analyse geluidsopnamen .................................................................................. 50

Pagina | 1
1. Inleiding
1.1 Algemeen
De Europese vleermuizen zijn insectenetende zoogdieren met een complexe levenswijze.
Ze stellen hoge eisen aan overwinteringsplaatsen, zomerverblijfplaatsen en
jachtgebieden. Hierdoor zijn ze bedreigd in een steeds grootschaliger en verstedelijkt
cultuurlandschap (Wickramasinghe et al., 2003; Dietz et al., 2011). De belangrijkste
bedreigingen in Vlaanderen zijn gebruik van bestrijdingsmiddelen, vernietiging van
leefomgeving en verstoring tijdens belangrijke fasen in de levenscyclus (Verkem en
Verhagen, 1998; Paelinckx et al., 2009). Vleermuispopulaties zijn vaak gelimiteerd door
de aanwezigheid van jachtmogelijkheden. Grote wijzigingen of verstoringen in
jachtgebieden die een veranderde insectendensiteit met zich meebrengen kunnen
populatieaantallen rechtstreeks beïnvloeden (Vaughan et al., 1997).
Het is bekend dat de Antwerpse forten voor de Vlaamse vleermuispopulaties tijdens de
winter een belangrijke rol spelen. Er overwinteren jaarlijks 12 soorten vleermuizen in de
forten. Voor zeven soorten zijn de forten ronduit cruciaal voor hun voortbestaan in
Vlaanderen (Paelinckx et al., 2009; Boers et al., 2010; Boers et al., 2013). Een aantal
forten werd daarom opgenomen binnen habitatrichtlijngebied. Vlaanderen maakte
gewestelijke instandhoudingsdoelstellingen op voor de vleermuissoorten die voorkomen
in Vlaanderen (Adriaens et al., 2008; Paelinckx et al., 2009). Voor het gebied
“BE2100045 Historische Fortengordels van Antwerpen als vleermuizenhabitat” werden
specifieke instandhoudingsdoelstellingen opgesteld. Het SBZ Historische fortengordel
rond Antwerpen is van essentieel belang voor de goede staat van instandhouding van
vleermuizen in Vlaanderen. Aangezien momenteel meer dan 50% van de getelde
populatie vleermuizen in het grotere netwerk van de fortengordels geteld wordt, werd in
het S-IHD rapport algemeen gesteld dat de doelstelling voor het volledige netwerk moet
bestaan uit 50% van een Vlaamse overwinteringspopulatie waarvoor de goede staat van
instandhouding voor het populatiecriterium uit de LSVI-tabellen (Adriaens et al., 2008)
bereikt wordt voor vleermuizen. Specifiek komt dit neer op een overwinteringspopulatie
van 4000 watervleermuizen, 2000 Brandt-/baardvleermuizen, 1600 franjestaarten, 750
ingekorven vleermuizen en 50 meervleermuizen. De voorgestelde maatregelen dienen
gerealiseerd te worden binnen de Speciale Beschermingszone (SBZ) zelf, terwijl de
vleermuizen eveneens behoefte hebben aan een (na)zomerhabitat, verbindingen tussen
zomer- en winterhabitat, en verbindingen tussen de verschillende forten onderling.
De stedelijke begraafplaats Schoonselhof grenst in het zuidoosten aan het
natuurreservaat Fort 7. Het fort maakt deel uit van de speciale beschermingszone
BE2100045 Historische fortengordels van Antwerpen als vleermuizenhabitat. Een
beperkte oppervlakte van het Schoonselhof (oostelijk deel) ligt eveneens binnen het
habitatrichlijngebied. Voor Fort 7 wordt de volgende doelstelling geformuleerd: het
bereiken van kwaliteitsvolle overwinterings- en zwermplaatsen voor watervleermuis,
franjestaart, Brandt- en baardvleermuis. De Ridder et al. (2014) brachten de recentste
gegevens over vleermuizen in Fort 7 in kaart. Het Schoonselhof kan als ecologisch
beheerde begraafplaats mee bijdragen aan het bereiken van een goede staat van
instandhouding voor de vleermuizen in fort 7. Er zijn echter bijna geen gegevens over
vleermuizen in het Schoonselhof beschikbaar. Deze studie wil een deel van deze
kennislacune wegwerken.

Pagina | 2
1.2 Gebiedsbeschrijving
De beschrijving is gebaseerd op Stijnen et al. (2011). De begraafplaats Schoonselhof is
gelegen te Antwerpen (districten Wilrijk & Hoboken) en is in totaal ongeveer 84 ha groot.
Het domein is in eigendom en beheer van de Stad Antwerpen. Het kasteel Schoonselhof
met bijhorende neerhoeve werd bij ministerieel besluit van 18 november 1991
beschermd als monument. De hele site van het Schoonselhof werd bij ministerieel besluit
van 16 juli 2007 integraal als monument beschermd om redenen van artistieke,
historische en sociaal-culturele waarde. Het Schoonselhof is grofweg op te delen in een
parkgedeelte met bijhorende gebouwen, gazons, vijvers en bos dat zich centraal uitstrekt
over een noordzuidas en een open gebied met begraafplaatsen dat gekenmerkt wordt
door verhoogde perken, bomenrijen, hagen en sloten. Belangrijke landschaps-
ecologische elementen zijn de lovergang (berceau), oude dreven met linde, beuk en eik,
het noordelijk parkbos met twee halve manen (ovaalvormige dubbele bomenrij), het
sterbos, het grachtenstelsel, vijf vijvers en de Hollebeek. De Hollebeek is een waterloop
die van zuidwest naar noordoost door het Schoonselhof stroomt en uiteindelijk in de
Schelde uitmondt. In het parkbos ten noorden van het kasteel heeft de Hollebeek nog
haar oorspronkelijke loop. De bedding werd een meter uitgediept bij de aanleg van de
begraafplaats, om een goede afwatering te verzekeren. In het noordoosten van het
parkbos stroomt de Hollebeek in een duiker onder de Krijgsbaan. Het bosdomein
Schoonselhof bestaat uit 2 bospercelen met een oppervlakte van respectievelijk 11,6 ha
ten noorden van het kasteel en 6,4 ha ten zuiden ervan. Het noordelijke parkbos bestaat
uit ongelijkjarige bestanden waarin leeftijdsklassen van de jongste (<20jaar) tot de
oudste (>160 jaar) vertegenwoordigd zijn. Het sterbos bestaat voor een groot deel uit
hooghout meer dan 120 jaar oud. Het totale volume van het staand dood hout bedraagt
11,3 % van het gemiddeld volume hout in het bosdomein.
1.3 Onderzoeksvragen en te verwachten resultaten
Welke soorten vleermuizen komen voor op de begraafplaats Schoonselhof? Zijn er
seizoensgebonden verschillen in het voorkomen van de soorten? Welke functies voor
vleermuizen kunnen toegekend worden aan het gebied? Hoe verloopt de vlieg- en
jachtactiviteit van vleermuizen over het seizoen?
Het antwoord op deze vragen wordt onderzocht aan de hand van het meten van vlieg- en
jachtactiviteit van vleermuizen op vaste meetpunten in 2014. De tellingen en
soortdeterminatie gebeurde met behulp van batdetectoren.
De studie wil een beeld geven van de verspreiding en soortenrijkdom van vleermuizen in
het onderzoeksgebied. Uit de vaststellingen op terrein en de verzamelde gegevens
worden conclusies getrokken over de verschillende vleermuizenfuncties in het gebied. Er
wordt getracht om o.a. vliegroutes en jachtgebieden van vleermuizen in kaart te
brengen. De rapportering wordt afgesloten met het formuleren van bijkomende of nieuwe
onderzoeksvragen en aanbevelingen voor het beheer in functie van vleermuizen.

Pagina | 3
2. Methodiek
2.1 Bepalen vlieg- en jachtactiviteit
De inventarisatiemethode werd uitgewerkt volgens de basisprincipes van het
vleermuisprotocol van het Vleermuisvakberaad Netwerk Groene Bureaus,
Zoogdiervereniging en Gegevensautoriteit Natuur (2013). Het vleermuisprotocol bestaat
uit een tabel met veldcondities en werkwijzen per vleermuissoort en per gebiedsfunctie,
op grond waarvan de aanwezigheid van soorten en gebiedsfuncties met voldoende
zekerheid kunnen worden vastgesteld. De goede praktijken van de Bat Workers’ Manual
(Mitchell-Jones, 2004), Bat Conservation Trust (UK, 2007) en de werkwijze van Barataud
(Barataud, 2012) werden in de mate van het mogelijke gevolgd.
Er werden 19 meetpunten (SH01 – SH19) aangeduid in het gebied (figuur 1.). De
meetpunten lagen minstens 50 meter uit elkaar. De meetpunten werden telkens op 1
avond volgens een vaste route gelopen. Het veldwerk werd uitgevoerd tussen 17 april en
10 oktober 2014, zodat de verzamelde gegevens betrekking hebben op een zo breed
mogelijk aspect van de seizoensactiviteit van de vleermuizen. Tijdens de studie werd
vlieg- en jachtactiviteit bepaald in trek-, balts- en paartijd, zwerm- en kraamperiode.
Figuur 1. Overzicht meetpunten begraafplaats Schoonselhof (luchtfoto bron: AGIV)

Pagina | 4
Er werd van meetpunt naar meetpunt gewandeld. Per meetpunt werden gedurende 3
minuten in tijdsintervallen van 5 seconden genoteerd of er vliegactiviteit was en van
welke soort dit was. Elk ononderbroken signaal werd geteld als 1 passage. Indien een
signaal langer dan 5 seconden aanhield, werd per bijkomende tijdsperiode van 5
seconden 1 extra passage geteld (Barataud, 2012). Met batdetectors kunnen passages
geteld worden, maar geen aantallen of individuen (Flaquer et al., 2007). Het aantal
passages is indicatief voor zowel vleermuisdensiteit als individuele foerageeractiviteit
(Verboom en Huitema, 1997; Verboom en Huitema, 2010, Wickramasinghe et al., 2003).
Aantallen werden enkel geteld bij visuele waarneming van de vleermuizen. Per meetpunt
werd het aantal vangstmomenten geteld op dezelfde manier als vliegactiviteit. De
vangstmomenten werden uitsluitend vastgesteld door een hoorbare vangstbuzz op de
heterodyne batdetector. De aan- of afwezigheid van sociale geluiden op een meetpunt
werd genoteerd, dus tijdens de 3 minuten waren er sociale geluiden of niet. Bij de
verplaatsing tussen meetpunten werden van de dan waargenomen vleermuizen de soort
en eventuele gedragsaanduidingen genoteerd op kaart. De veldnotities werden genoteerd
op een inventarisatiefiche. De tellingen werden uitgevoerd door minstens twee personen,
omwille van veiligheidsredenen en voor controle van de determinaties. Er werd steeds
getracht om minimaal 20 dagen tussen elk opeenvolgend bezoek aan het gebied te laten.
De tellingen werden uitgevoerd in goede weersomstandigheden. De starttemperatuur
was voor alle tellingen hoger dan 10°C (Bat Conservation Trust, 2007;
Vleermuisvakberaad Netwerk Groene Bureaus, Zoogdiervereniging en Gegevensautoriteit
Natuur, 2013). Alle tellingen werden gestart binnen het uur na de officiële
zonsondergang om een zo representatief mogelijk beeld van de activiteit te krijgen.
Vleermuizenactiviteit en de beschikbaarheid van vliegende insecten die dienen als
potentiële prooi volgen immers een bimodale curve met maxima tijdens de avond- en
ochtendschemering (Kusch et al., 2004). Vleermuizen zoals gewone dwergvleermuis, die
jachtvlucht (aerial hawking) als jachtstrategie gebruiken zijn bv. afhankelijk van de
avondpiek van beschikbare insecten. De jachtactiviteit van deze soorten neemt af later
op de avond (Kapfer & Aron, 2007).
2.2 Verkenning jachtgebieden, vliegroutes en zwermlocaties
Aanvullend aan het bepalen van vlieg- en jachtactiviteit werd vleermuisactiviteit
opgevolgd in 4 deelzones: zone kasteelvijver, Neerhoeve en lindendreef (L14), het
noordelijk parkbos, het sterbos (incl. dreef L17b, L19b en L24) en de grachten en dreven
(dreef L29, L30) in de overgangszones met fort 7. De nummering van de dreven is
overgenomen uit het beheerplan en wordt weergegeven op figuur 1 in de bijlagen.
Doelstelling was extra informatie verzamelen om vliegroutes, jachtgebieden of eventuele
zwermlocaties in kaart te brengen. Het noordelijk parkbos was ’s nachts moeilijk te
betreden waardoor mogelijks een vertekend beeld is ontstaan. Vleermuizenactiviteit werd
hier vooral opgevolgd langs de randen en in de beukendreef (L39). Ook voor het sterbos
werd de vleermuizenactiviteit vooral opgevolgd langs de randen en in de dreven. Er werd
op 7 tijdstippen (zie bijlagen tabel 4.) een bezoek gebracht op de verschillende locaties.
Er werden gedurende minstens 30 minuten observaties gedaan in de deelzones. Van de
waargenomen vleermuizen werd de soort en eventuele gedragsaanduidingen genoteerd
op kaart. De veldnotities werden genoteerd op een inventarisatiefiche. Het terreinbezoek
werd uitgevoerd door twee personen, omwille van veiligheidsredenen en voor controle
van de determinaties. De waarnemingen werden uitgevoerd in goede

Pagina | 5
weeromstandigheden. De starttemperatuur was voor alle waarnemingen hoger dan 10°C
(Bat Conservation Trust, 2007; Vleermuisvakberaad Netwerk Groene Bureaus, Zoogdier-
vereniging en Gegevensautoriteit Natuur, 2013). Alle waarnemingen werden gestart
binnen het kwartier na de officiële zonsondergang om een zo representatief mogelijk
beeld van de activiteit te krijgen. Om de aanwezigheid van vliegroutes te bepalen
plaatsten twee waarnemers zich met batdetectoren op een verschillende plek langs de
dreven, zodat de vliegrichting eenvoudig kon worden bepaald, en om te vermijden dat
terug-vliegers dubbel werden geteld. Een vleermuis op route vliegt rechtlijnig, en er
worden nauwelijks vangstmomenten waargenomen. Er werden geen effectieve tellingen
op vliegroutes uitgevoerd.
2.3 Soortdeterminatie aan de hand van ultrasone geluiden
De activiteitstellingen werden uitgevoerd met 2 heterodyne batdetectoren Pettersson
D100 (Pettersson Elektronics AB, Uppsala, Zweden). De Pettersson D100 werd afgesteld
op 20 kHz en 45 kHz. Vleermuizen werden op het terrein gedetermineerd met 1 of 2
time-expansie batdetectoren Pettersson D240x (Pettersson Elektronics AB, Uppsala,
Zweden). De Pettersson D240x werd ingesteld op het heterodyne kanaal op 34 kHz. Bij
waarneming van vleermuizenactiviteit, werd met de D240x over een brede range van
frequenties gescand om de nulfrequentie te achterhalen. Werken met de Pettersson
D240x maakt het mogelijk geluiden op te nemen en vertraagd terug af te spelen zodat
determinaties op het terrein correcter kunnen gebeuren (o.a. door te luisteren naar
piekfrequentie en bandbreedte). De opnames werden gemaakt in .wav-formaat met een
Roland R05 Edirol. De geluidsopnames werden gedigitaliseerd en geanalyseerd met
Batsound 4 (Pettersson Elektronik AB, Uppsala, Zweden). Bij twijfelgevallen werd enkel
gedetermineerd tot op genus-niveau. Van de signalen werd piekfrequentie (FME), initiële
frequentie (FI), eindfrequentie (FT) en pulslengte (ms) bepaald van een tiental
representatieve signalen. De soorten werden gedetermineerd op basis van
referentiewerken en -geluiden (Van De Sijpe, 1999; Barataud, 2012; Russ, 2012;
Middleton et al., 2014) en determinatiegrafieken (Barataud, 2012). Het is tot op zekere
hoogte mogelijk om vleermuizen te determineren aan de hand van ultrasone geluiden,
zeker indien er een combinatie van heterodyne en time-expansie technieken wordt
gebruikt. Visuele waarnemingen (grootte, kleur, gedrag) kunnen extra informatie bieden
om tot determinatie te komen (Ahlén en Baagøe, 1999). Voor de analyse van de
opnames en de soortdeterminatie werd teruggevallen op de methode beschreven door
Barataud (2012). De vleermuizen van het genus Myotis lijken sterk op elkaar, zowel
uiterlijk als wat ultrasone geluiden betreft. De Myotis–geluiden werden in deze studie
meestal slechts (gedetermineerd en) verwerkt op genus-niveau. Op basis van de
geluidsopnames werd toch getracht te determineren op soortniveau.
2.4 Wintertelling
Het opvolgen van soorten en aantallen vleermuizen in de Neerhoeve gebeurde door
visuele inspectie. Er werd gebruik gemaakt van een goede zaklamp en een spiegeltje om
in moeilijke holten en spleten te zoeken (Dijkstra en Korsten, 2005). De te onderzoeken
locatie werd 1 maal in de winter onderzocht. De beste periode voor het tellen van
vleermuizen is van 15/12 tot 01/03. Waarnemingen buiten deze periode blijken in de
praktijk niet goed vergelijkbaar (Willems et al., 2009, Dijkstra en Korsten, 2005).
Bij de wintertelling worden niet alle aanwezige dieren gevonden. De telfout is afhankelijk
van (het gedrag van) een soort. Verschillende soorten vleermuizen kruipen weg in
scheuren en spleten. Sommige overwinteringsplaatsen zijn slechts gedeeltelijk te

Pagina | 6
controleren op zicht. Het S-IHD rapport van de speciale beschermingszone ‘BE2100045
Historische fortengordels van Antwerpen als vleermuizenhabitat’ gaat uit van een
zoekfout van 30 tot 50%.
2.5 Verwerking en statistische analyse
De telgegevens en puntwaarnemingen werden verwerkt in een GIS-omgeving (QGIS 2.6
Brighton) voor ruimtelijke interpretatie en kunnen bij de auteurs opgevraagd worden. De
nummering van de dreefsegmenten (bijlagen, figuur 1.) en bospercelen (bijlagen, figuur
2.) werd overgenomen uit het beheerplan (Stijnen et al., 2011). De nummering van de
grafperken werd overgenomen van het plan van het Schoonselhof (bijlagen, figuur 3.,
bron: website stad Antwerpen). De gegevens worden weergeven op de middenschalige
kleurenorthofoto’s van de provincie Antwerpen, opname 2010 (bron: AGIV).
In deze studie werd vliegactiviteit gedefinieerd als het aantal tijdsintervallen met
passages per tijdseenheid (i.e. gedurende de 3 minuten waarnemingstijd). Bij de
verwerking werd geen onderscheid gemaakt tussen aantallen passages afkomstig van 1
vleermuis of van meerdere vleermuizen. Vleermuizen zijn zeer opportunistisch in hun
jachtgedrag. De foerageeractiviteit van vleermuizen hangt af van de verschillende
jachtstrategieën van vleermuizensoorten en van de beschikbaarheid van insecten. Als de
activiteit van insecten toeneemt, neemt de vliegactiviteit van vleermuizen toe (Rydell et
al., 1996; Kusch et al., 2004; Cel’uch & Kaňuch, 2004; Lee & McCracken, 2004).
Jachtactiviteit werd eveneens gedefinieerd als het aantal tijdsintervallen met
vangstmomenten per tijdseenheid (i.e. gedurende de 3 minuten waarnemingstijd).
Vleermuisactiviteit wordt beïnvloed door de reproductieve status van de vleermuizen. De
verzamelde gegevens werden daarom voor de bespreking onderverdeeld in 3 perioden:
april tot midden juni, midden juni tot einde juli en augustus tot oktober. Deze perioden
komen overeen met de onderscheidde fasen in de voorplantingscyclus van de meeste
algemene vleermuissoorten in Europa: drachtperiode of kraamtijd, lactatiefase en post-
lactatiefase. In deze studie werden de begrippen vliegroute en jachtgebied gehanteerd.
Een vliegroute is een vaste route van een vleermuis of een groep van vleermuizen vanaf
een verblijfplaats naar een foerageergebied of tussen verblijfplaatsen en vice versa. Het
jachtgebied is een gebied waar een vleermuis of een groep van vleermuizen foerageert
(Vleermuisvakberaad Netwerk Groene Bureaus, Zoogdiervereniging en
Gegevensautoriteit Natuur, 2013). Foerageergebied werd gebruikt als synoniem voor
jachtgebied.
Variatie in vliegactiviteit en jachtactiviteit werden geanalyseerd met “generalised linear
mixed models” (GLMM) (Zuur et al., 2009). Beide activiteiten werden gekwantificeerd als
het aantal tijdsintervallen van 5 seconden over een periode van 3 minuten dat er
activiteit werd waargenomen. Activiteit is dus de proportie van tijdsintervallen waarin
activiteit plaatsvindt en varieert tussen 0 (geen activiteit) en 36 (activiteit tijdens alle
intervallen). De afhankelijke variabele in de analyse is een proportie en dus niet normaal
maar binomiaal verdeeld (= generalised model). Om activiteitsvariatie in de tijd te
bepalen werden de opnamedatums als verklarende factor in het model opgenomen.
Vermits opnames gedaan op een bepaald meetpunt niet onafhankelijk zijn (een bepaald
punt kan immers meer activiteit hebben omdat het een betere foerageerplaats is) moet
daarvoor gecorrigeerd worden. Daarom wordt meetpunt als een random effect in het
statistisch model opgenomen (= mixed model). Wanneer het verschil in activiteit tussen
verschillende meetpunten wordt geanalyseerd zijn opnames gemaakt op eenzelfde dag
niet onafhankelijk. Bij deze analyse wordt dan ook opnamedag als random effect in het

Pagina | 7
statistisch model gestoken. Wanneer variatie in functie van de tijd geanalyseerd wordt,
wordt er van uit gegaan dat er geen lineair verband is. Er wordt bijvoorbeeld minder
activiteit verwacht net na of net voor de winter en meer activiteit in de lente en/of
zomer. Daarom wordt hier gebruik gemaakt van “generalised additive mixed models”
(GAMM) (Zuur et al., 2009). Additive models gebruiken “splines”, een aaneenschakeling
van stukjes polynomen, om de regressielijn te bepalen waardoor die een vloeiende curve
kan beschrijven. Bij het vergelijken van meetpunten is dit niet nodig vermits meetpunt
een categorische variabele is.
Alle analyses werden uitgevoerd met het statistisch programma R (versie 3.0.2,
R Development Core Team 2013). Voor GLMM’s werd gebruik gemaakt van de glmer
procedure van het lme4 pakket (Bates et al., 2013), voor de GAMM van de gamm
procedure uit het mgcv pakket (Wood, 2014).
3. Resultaten
3.1 Algemeen
Er werden tellingen van vlieg- en jachtactiviteit uitgevoerd gedurende 7 avonden (zie
bijlagen tabel 4.). Aanvullende inventarisaties van deelgebieden werden eveneens
uitgevoerd gedurende 7 avonden (zie bijlagen tabel 5.).
Tabel 1. geeft de tijdens de inventarisatieperiode waargenomen soorten weer. Er werden
minstens 8 soorten vleermuizen waargenomen: gewone dwergvleermuis (Pipistrellus
pipistrellus), ruige dwergvleermuis (Pipistrellus nathusii), laatvlieger (Eptesicus
serotinus), rosse vleermuis (Nyctalus noctula), baardvleermuis (Myotis mystacinus),
watervleermuis (Myotis daubentonii), franjestaart (Myotis nattereri) en gewone
grootoorvleermuis (Plecotus auritus).
Tabel 1. Overzicht van de waargenomen soorten (17 april tot 10 oktober 2014). Status rode lijst
(zomer) volgens Maes et al., 2014 Soort Ned. naam Status rode lijst (zomer)
IUCN Rode Lijst Europa
Pipistrellus
pipistrellus
Gewone
dwergvleermuis
Momenteel niet in
gevaar Least Concern
Pipistrellus nathusii Ruige
dwergvleermuis
Momenteel niet in
gevaar Least concern
Eptesicus serotinus Laatvlieger Kwetsbaar Least concern
Myotis mystacinus Baardvleermuis Onvoldoende data Least concern
Myotis nattereri Franjestaart Onvoldoende data Least concern
Myotis daubentonii watervleermuis Bijna in gevaar Least concern
Plecotus auritus Gewone
grootoorvleermuis Bijna in gevaar Least concern
Nyctalus noctula Rosse vleermuis Kwetsbaar Least concern
Niet alle vleermuizen werden op soort- of op genus-niveau gedetermineerd, zo werden
bijvoorbeeld de grootoorvleermuizen door geluidsanalyse meestal niet tot op soortniveau
gedetermineerd. Het betreft gewone grootoorvleermuis of grijze grootoorvleermuis.
Beide soorten komen in Vlaanderen voor, maar gewone grootoorvleermuis is het meest
algemeen van de twee soorten (Verkem et al., 2003, Paelinckx et al., 2009). Een aantal
opnamen konden via de methode Barataud (2012) geïdentificeerd worden als gewone
grootoorvleermuis (identificatieniveau: zeker of waarschijnlijk). In de studie worden de
waarnemingen verder als gewone grootoorvleermuis benoemd.

Pagina | 8
Iets meer dan de helft van de opnames van Myotis-vleermuizen konden tot op soort
worden gedetermineerd met de methode Barataud (2012). De determinatie gebeurde op
basis van auditieve kenmerken of op basis van een combinatie van auditieve kenmerken
en visuele waarneming van bepaalde kenmerken en gedrag. De opnamen van
vleermuizen die gebruik maken van het akoestisch type abs moy (afwezigheid van
energiepiek met gemiddelde eindfrequentie) konden worden gedetermineerd als
watervleermuis (identificatieniveau: zeker of waarschijnlijk) door aanvullende visuele
waarnemingen van jachtgedrag en door de aanwezigheid van overgangen in akoestisch
type (abs moy naar abs ht) en de aanwezigheid van een sinusoïdaal verloop in de
fundamentele puls. In een aantal opnames werd uitsluitend het akoestisch type am moy
waargenomen. De vleermuizen vlogen in een relatief open milieu. De pulsduur was meer
dan 5 ms en het pulsinterval (meer dan 90 ms) was relatief lang. De vleermuizen werden
geïdentificeerd als baardvleermuis (identificatieniveau: zeker). In een aantal opnames
werd een overgang van het akoestisch type am moy naar am ht (explosieve start met
hoge eindfrequentie) waargenomen. De pulsduur was bovendien meer dan 5 ms. De
vleermuizen werden geïdentificeerd als baardvleermuis (identificatieniveau: zeker).
Franjestaart werd gedetermineerd door de aanwezigheid van FM-signalen van het
akoestisch type claq bas, en een grote bandbreedte.
Een groot deel van de waarnemingen met batdetectoren van Myotis-soorten konden
echter (nog) niet op soort gedetermineerd worden. De opnamen waarin uitsluitend het
akoestisch type am moy (explosieve start met gemiddelde eindfrequentie) gebruikt
wordt, werden benoemd als watervleermuis/baardvleermuis. Volgens Barataud (2012)
wordt dit akoestisch type gebruikt door baardvleermuis in een vrij open milieu met een
traag ritme en door watervleermuis in een korte fase bij benadering van het bladerdak,
of tijdens jacht in een dichte begroeiing. In de verdere besprekingen worden deze
waarnemingen behandeld als Myotis spp.
De gedetermineerde Myotis-vleermuizen hebben de volgende verhouding: 79%
watervleermuis, 15% baardvleermuis en 6% franjestaart.
3.2 Vliegactiviteit en jachtactiviteit
Tabel 2. geeft de tijdens de zomerinventarisatie op de meetpunten waargenomen soorten
en het aantal passages per soort op de meetpunten weer. Er werden in 2014 op de
meetpunten 1381 passages van vleermuizen geteld, verdeeld over minstens 5 soorten
vleermuizen: gewone dwergvleermuis, watervleermuis, ruige dwergvleermuis, rosse
vleermuis en laatvlieger. Een aantal soorten werd enkel waargenomen tijdens de
verplaatsingen tussen de meetpunten, maar niet op de meetpunten zelf. Het gaat over
franjestaart en baardvleermuis. De vliegactiviteit van gewone dwergvleermuis was het
hoogst en was verantwoordelijk voor meer dan 90% van de totale activiteit in het
studiegebied. Watervleermuis en ruige dwergvleermuis en watervleermuis vertoonden
respectievelijk 5,5% en 1,5% van de vliegactiviteit. Barataud (2012) geeft aan dat in
Europa 50 – 80% van de waarnemingen met batdetectors uit dwergvleermuizen bestaan.
In andere studies van gebieden in Vlaanderen werden gelijkaardige aantallen soorten
waargenomen en was het aandeel gewone dwergvleermuis vergelijkbaar (Galens en
Dekeukeleire, 2011; Nyssen, 2012; De Ridder en Sanders, 2013; De Ridder et al., 2014).

Pagina | 9
Tabel 1. Overzicht van de waargenomen soorten en aantal passages tijdens de totale
inventarisatieperiode (24 april tot 10 oktober 2014)
Soort # passages op
meetpunten
% passages op
meetpunten
% passages
tussen
meetpunten
% passages op
meetpunten Fort
7 (2013)
Pipistrellus
pipistrellus 1243 90 60,2 87,5
Myotis daubentonii 77 5,5 5 5
Pipistrellus nathusiii 21 1,5 7,7 5,3
Nyctalus noctula 15 1 5,3 0
Eptesicus serotinus 11 1 10,1 0,1
Plecotus spp. 9 0,6 9 1
Myotis spp. 5 0,4 2,3 1,1
Als de waarnemingen tussen de meetpunten worden bekeken, dan valt het hoger aandeel
waarnemingen van gewone grootoorvleermuis (9%), laatvlieger (10,1%), ruige
dwergvleermuis (7,7%) en rosse vleermuis (5,3%) op.
Aangezien enkel een ultrasone methode werd gebruikt voor de inventarisatie zal er een
zekere overschatting en onderschatting van de aanwezigheid van bepaalde soorten zijn.
Soorten die QCF-signalen gebruiken zijn luider dan soorten die exclusief FM-signalen
gebruiken. Soorten die tussen vegetatie jagen gebruiken steilere signalen, die over
kortere afstanden worden waargenomen (Brigham et al., 2004). Flaquer et al. (2007)
stelden bij een vergelijking van methoden vast dat bij inventarisatie met batdetectors
holte-bewonende vleermuizen werden onderschat ten opzichte van bijvoorbeeld soorten
die in menselijke structuren leven. Een groot deel van de meetpunten is bovendien
gesitueerd langs de rand van het gebied, langs overgangen van habitats en langs
potentieel foerageergebied van vleermuizen. Dit zou ook een bias van de gegevens
kunnen geven. Lesiński et al. (2011) onderzochten de hypothese dat de vliegactiviteit
van vleermuizen in bossen hoger is in delen dichter bij de randen o.a. door de
aanwezigheid van kraamkolonies in bomen en door foerageergedrag langs de randen van
vleermuizen van binnen en buiten het bos. Bij laatvlieger werd een tendens
waargenomen, maar enkel bij gewone grootoorvleermuis kon door hen een significant
verschil aangetoond worden. Onderzoek in Nederland toonde een verband aan tussen
activiteit van gewone dwergvleermuis en bomenrijen (Verboom, 1998; Verboom &
Huitema, 1997 & 2010). In het onderzoek werd een verminderende activiteit
waargenomen naarmate de afstand tot bomenrijen toenam.
Figuur 2. geeft de relatieve verdeling van de vliegactiviteit over de waargenomen soorten
per meetpunt weer. Het aantal waargenomen soorten per meetpunt is erg variabel. Er
werden 1 tot 5 soorten per meetpunt waargenomen. Op de meetpunt SH04 en SH06
werden 5 soorten waargenomen. Op de meetpunten SH05, SH07 en SH19 werden 4
soorten waargenomen. Gewone dwergvleermuis werd op alle meetpunten waargenomen.
Het voorkomen van ruige dwergvleermuis, Myotis-soorten, grootoorvleermuis, rosse
vleermuis en laatvlieger is sterk afhankelijk van meetpunt.

Pagina | 10
Figuur 2. Relatieve verdeling van de activiteit en de waargenomen soorten per meetpunt (Md:
watervleermuis, Es: laatvlieger, Nn: rosse vleermuis, Msp: Myotis-soorten, Plsp: grootoorvleermuis,
Pn: ruige dwergvleermuis; Pp: gewone dwergvleermuis).
Vliegactiviteit van gewone dwergvleermuis werd waargenomen van begin april tot
midden oktober. Om de verdeling van de vliegactiviteit over de inventarisatieperiode te
testen werd gewerkt via een “generalised additive mixed model” (GAMM) met binomiale
error structuur. De afhankelijke variabele is activiteit en de verklarende variabele is
datum. Meetpunt werd als random effect in het model ingebracht om te corrigeren voor
de afhankelijkheid van de data binnen een meetpunt. Het model fit significant (F1,132 =
2.571, p <0.05). Uit figuur 3. kan afgeleid worden dat de vliegactiviteit het hoogst is in
de periode 15 april tot 15 juni (kraamtijd) en in de periode tussen 15 september en 15
oktober (post-lactatiefase). De gemiddelde vliegactiviteit bedraagt dan iets meer dan 5
passages per 3 minuten (per meetpunt). Midden oktober worden de laagste waarden
waargenomen (2 passsages per 3 minuten). Wijzigingen in de seizoensactiviteit van
gewone dwergvleermuis werd door verschillende onderzoekers vastgesteld. Er worden
verschillende verklaringen gegeven aan de variatie in de vliegactiviteit over het seizoen,
o.a. omwille van wijzigingen in de reproductieve cyclus, verschuivingen van jachthabitat
en nazomerzwermactiviteit. Kapfer et al. (2007) stelden een significant lagere
vliegactiviteit vast van gewone dwergvleermuis tijdens de kraamtijd dan in de lactatie-
en post-lactatiefase. Jonker et al. (2010) namen een duidelijk patroon waar in de
seizoenactiviteit van gewone dwergvleermuis. De vliegactiviteit nam toe gedurende de
zomer, en was het laagst in de nazomer. De toename wordt verklaard door de hogere
energiebehoeften van lacterende vrouwtjes, en doordat juveniele vleermuizen individueel
beginnen te jagen. De Ridder & Sanders (2013) stelden voor het gebied Klaverblad vast
dat de vliegactiviteit het hoogst was in de post-lactatiefase. In het aangrenzende fort 7
werd de hoogste vliegactiviteit vastgesteld in de kraamtijd (De Ridder et al., 2014).

Pagina | 11
Figuur 3. Gemiddelde vliegactiviteit van gewone dwergvleermuis over het vliegseizoen. De zwarte
lijn is de trend (± SE) voor alle meetpunten gecombineerd. Elke oranje lijn is een meetpunt.
De vliegactiviteit was verschillend tussen de meetpunten. Figuur 4. geeft de gemiddelde
vliegactiviteit op elk meetpunt weer. Er is een significant verschil in gemiddelde
vliegactiviteit tussen de meetpunten (χ218 = 758.79, p < 0.001). De meetpunten SH16
en SH19 hebben de hoogste gemiddelde vliegactiviteit (meer dan 20 passages per 3
minuten). Er werd gedurende het hele seizoen hoge vliegactiviteit waargenomen op deze
meetpunten. Het meetpunt SH16 ligt aan een gracht op de grens met Fort 7. Op het
meetpunt werd bij elk bezoek hoge activiteit waargenomen. Het meetpunt SH19 ligt ter
hoogte van de kasteelvijver. Hier werden telkens meerdere jagende dieren
waargenomen. De meetpunten SH02, SH04, SH06 en SH15 hebben een hoge gemiddeld
vliegactiviteit (meer dan 12 passages per 3 minuten). Het meetpunt SH02 bevindt zich
op een vliegroute van gewone dwergvleermuis. De meetpunten SH04, SH06 en SH15
bevinden zich in de omgeving van een open wateroppervlak met voldoende beschutting.
Op andere meetpunten werd soms ook hoge vliegactiviteit geteld, maar deze vertoonden
meer variatie in de loop van het jaar. Dit werd niet statistisch getest in deze studie, maar
de verschillen in punten worden indicatief aangegeven. In de kraamperiode hadden o.a.
meetpunten SH12, SH15, SH16, SH17, SH18 en SH19 een hoge vliegactiviteit. In de
lactatiefase was dit het meetpunt SH02. In de post-lactatiefase waren dit de meetpunten
SH01, SH02, SH06, SH07, SH16 en SH19.

Pagina | 12
Figuur 4. Gemiddelde vliegactiviteit van gewone dwergvleermuis op de meetpunten.
Jachtactiviteit van gewone dwergvleermuis werd waargenomen van begin april tot
midden oktober. Om de verdeling van de vliegactiviteit over de inventarisatieperiode te
testen werd gewerkt via een “generalised lineair mixed model” (GLMM) met binomiale
error structuur. De afhankelijke variabele is activiteit en de verklarende variabele is
datum. Het model zonder random effect gaf het beste resultaat. Het model fit significant
(χ26 = 143.57, p < 0.001). Figuur 5. geeft de gemiddelde jachtactiviteit in het
onderzoeksgebied weer. De gemiddelde jachtactiviteit neemt geleidelijk af over het
verloop van het seizoen. Het patroon is echter niet uitgesproken. De gemiddelde
jachtactiviteit varieert van 2,7 - 1,5 vangstmomenten per 3 minuten.
In figuur 6. wordt de gemiddelde jachtactiviteit per meetpunt weergegeven. Er was een
significant verschil in activiteit tussen de meetpunten (χ218 = 450.39, p < 0.001). Op de
meetpunten SH15, SH16, en SH19 werd de hoogste gemiddelde jachtactiviteit
waargenomen. Op 6 meetpunten werd geen (of zeer beperkt) jachtactiviteit
waargenomen. Op sommige tijdstippen werd op meetpunten de maximale jachtactiviteit
geteld (36 vangstmomenten op 3 minuten). Vermoedelijk is dit een overschatting van de
jachtactiviteit als gevolg van de telmethode. De overgangen van heterodyne signalen van
meerdere te gelijk jagende vleermuizen klinken vaak zoals vangstmomenten
(mondelinge mededeling Ralf Gyselings).

Pagina | 13
Figuur 5. Gemiddelde jachtactiviteit van gewone dwergvleermuis over het vliegseizoen. De zwarte
lijn is de trend (± SE) voor alle meetpunten gecombineerd. Elke oranje lijn is een meetpunt.
Figuur 6. Gemiddelde jachtactiviteit van gewone dwergvleermuis op de meetpunten.

Pagina | 14
De vlieg- en jachtactiviteit van andere soorten was vaak beperkt tot enkele punten. Er
waren bovendien onvoldoende telgegevens (passages) om een relevante statistische
analyse te kunnen uitvoeren. De bespreking is beperkt tot een kwalitatieve bespreking.
Ruige dwergvleermuis werd op 7 meetpunten waargenomen in het voorjaar (april) en
het najaar (augustus tot oktober). Het ging zowel over jagende dieren als dieren op
route. Als de passages tussen de meetpunten mee in rekening worden gebracht, werd
ruige dwergvleermuis gedurende het hele jaar in het gebied waargenomen. Er werden
een beperkt aantal sociale geluiden van ruige dwergvleermuis waargenomen in de
omgeving van het kasteel. De activiteit van ruige dwergvleermuis was steeds in de
omgeving van water: de kasteelvijver en achterliggende grachten, de langgerekte vijver
in het noordelijk parkbos, de spiegelvijver, en een aantal afwateringsgrachten.
Laatvlieger werd op de meetpunten SH01, SH03, SH06 en SH07 waargenomen. Er
waren eveneens een groot aantal passages van laatvlieger tussen de meetpunten. Het
ging zowel over jagende dieren, als over dieren op een vliegroute of verplaatsend tussen
kernjachtgebieden. De vliegactiviteit werd waargenomen van april tot september.
Laatvlieger werd jagend aangetroffen in de omgeving van de grote kasteelvijver en de
lindendreef (L14) voor de Neerhoeve, en in de dreven (L3, L10b, L12b, L13, L16, L18)
en boven de grafperken (perk N, U, L, W2, groot militair ereperk, perk 5). De jagende
dieren konden vaak voor een langere periode geobserveerd worden o.a. ter hoogte van
grafperk W2, grafperk N en het monument 'Hulde Mevr. Leopold De Wael – Van Gend''
op L10b. Er werden maximaal 2 dieren gelijktijdig (visueel) waargenomen. Laatvlieger
werd zich verplaatsend tussen jachtgebieden aangetroffen in de dreven L1, L2, L5, L13,
L14, L17b, L18 en L30 en boven perk 3 en 4 en het groot militair ereperk.
Rosse vleermuis werd op 7 meetpunten waargenomen. Er waren heel wat
puntsgewijze waarnemingen over het gehele domein. De vliegactiviteit werd vastgesteld
van april tot midden juni. Het waren meestal relatief korte waarnemingen, o.a. van
overvliegende dieren of dieren jagend op grote hoogte. Rosse vleermuis werd langdurig
jagend waargenomen boven perk U en Z en boven het groot militair ereperk.
Watervleermuis werd waargenomen op 6 meetpunten SH04, SH06, SH09, SH16, SH17
en SH19. Het betrof telkens jagende dieren boven een afwateringsgracht of de
kasteelvijver of dieren op verplaatsing tussen twee jachtgebieden. De waarnemingen
werden gedaan zowel tijdens als tussen en na de telling. De vlieg- en jachtactiviteit was
relatief beperkt en vaak voor slechts korte duur. Waarnemingen werden gedaan in de
periode midden april tot september. Watervleermuis werd enkel zich verplaatsend tussen
jachtgebieden en/of op route waargenomen in de beukendreef L17b. Op 3 september
werden er minimaal 10 dieren zwermend boven de gracht ter hoogte van meetpunt SH16
waargenomen. 79% van de gedetermineerde Myotis-soorten waren watervleermuizen.
Baardvleermuis en franjestaart werden niet met zekerheid waargenomen op de
meetpunten tijdens de tellingen, maar wel tijdens de inventarisatie van de deelgebieden.
Op 27 juni 2014 waren er twee passages van jagende franjestaart in de beukendreef in
het noordelijke parkbos (segment L39a). Dit waren de enige zekere waarnemingen van
franjestaart voor het Schoonselhof. In dezelfde dreef werd baardvleermuis vliegend op
route waargenomen op 3 september 2014. Er was eveneens beperkte jachtactiviteit.
Baardvleermuis werd eveneens op route waargenomen in beukendreef L17b op 25

Pagina | 15
september. Het ging over 1 passage. Op 3 september werden er minimaal 2 dieren
zwermend boven de gracht ter hoogte van meetpunt SH16 waargenomen.
Andere Myotis-soorten werden slechts beperkt waargenomen tijdens de tellingen op
meetpunten. Het ging telkens over 1 of 2 passages. Deze waarnemingen konden niet op
soort geïdentificeerd worden. De waarnemingen bleven beperkt tot meetpunten SH03,
SH06 en SH07. Tussen de meetpunten werd eveneens slechts beperkte activiteit van
Myotis-soorten waargenomen. De waarnemingen werden gedaan over het hele
waarnemingsseizoen maar waren sterk locatie-gebonden en beperkt tot de dreven: L5,
L15b, L17b, L19a, L24, L25, L29, L30, L39a en L39b. Langs de dreven L5, L17b L24
werden waarnemingen van dieren op route gedaan.
Vliegactiviteit van grootoorvleermuis werd van april tot oktober waargenomen. Het
ging telkens over enkele passages op 7 meetpunten (SH04, SH05, SH07, SH10, SH13,
SH17, SH18). De vliegactiviteit van grootoorvleermuis was groter tussen de telpunten
dan op de meetpunten. 9% van de waarnemingen tussen meetpunten werden
gedetermineerd als grootoorvleermuis. Tijdens de inventarisatie van de deelgebieden
werd eveneens hoge activiteit van grootoorvleermuis vastgesteld. Het ging over 13% van
de waarnemingen. Het ging over zowel foeragerende dieren, als dieren op route. Er
werden 78 passages van grootoorvleermuis geteld over de gehele inventarisatie.
Jachtactiviteit van grootoorvleermuis werd waargenomen in de dreven: L9, L10a, L10b
L14, L17a, L17b, L19b, L22, L24, L29, L33 en L34. Er was ook heel wat jachtactiviteit in
de omgeving van de Neerhoeve en boven de kasteelgracht. Grootoorvleermuis op route
werd waargenomen in de dreven: L14, L17b, L19a, L19b, L24, L29, L30. Sociale geluiden
van grootoorvleermuis werden vastgesteld in de dreven L5, L14, L19b, L29 en L33.
4. Bespreking en vleermuizenfuncties
Tabel 3. geeft een overzicht van de waargenomen vleermuizenfuncties op de
begraafplaats Schoonselhof. Het beeld is momenteel nog versnipperd, aangezien de
vleermuizeninventarisatie enkel uitgevoerd werd over één seizoen en enkel met
batdetectors. Er werd slechts een beperkt onderzoek uitgevoerd naar voorkomen van
winterverblijfplaatsen en vliegroutes en geen specifiek onderzoek naar aanwezigheid van
zwermlocaties.
Tabel 3. Overzicht van waargenomen vleermuizenfuncties op de begraafplaats Schoonselhof (‘X’
aanwezig, ‘O’ beperkt of waarschijnlijk aanwezig, ‘-‘ (waarschijnlijk) niet aanwezig, ‘?’ onbekend)
Zomer-
Verblijf
Nazomer
zwerm-
locatie
Paar-
Verblijf
Winter-
verblijf
Jacht-
gebied
vliegroute/
verbindings-
route
Gewone dwergvleermuis X X O ? X X
Ruige dwergvleermuis ? ? O ? X ?
Laatvlieger ? ? ? ? X ?
Rosse vleermuis ? ? ? ? X n.v.t.
Gewone grootoorvleermuis X ? O X X X
Watervleermuis ? X ? ? X X
Baardvleermuis O O ? ? O O
Franjestaart O ? ? ? O O

Pagina | 16
4.1 De functie winterverblijfplaats
Er zijn geen gegevens uit literatuur bekend van gebruik van gebouwen of andere
objecten van het Schoonselhof als winterverblijfplaats.
In kader van deze studie werd één verkennende wintertelling uitgevoerd in de
vierkantshoeve De Neerhoeve. De andere gebouwen (kasteel, toegangsgebouw),
bakstenen bruggen en duikers en de monumentale grafmonumenten werden niet
gecontroleerd op aanwezigheid van overwinterende vleermuizen. De wintertelling werd
uitgevoerd op 17 januari 2014. Een groot deel van de zolderruimte kon omwille van
veiligheidsredenen niet betreden worden. De kelderruimte stond onder water, waardoor
deze ook niet betreden kon worden, op een kleine kelder na. Er werden drie exemplaren
gewone grootoorvleermuis gevonden in het noordwestelijk gedeelte van de zolder. De
dieren werden op soort gedetermineerd op basis van visuele kenmerken: lichtgekleurde
tragus en de aanwezigheid van 4 knobbels op een korte snuit. De temperatuur op de
zolder bedroeg 7,3°C, de relatieve luchtvochtigheid bedroeg 67%. De vondst van delen
van een onderarm en middenhandsbeentjes (geen soortdeterminatie) wijst op een al
langduriger gebruik als verblijfplaats. Tijdens de telling werden eveneens op
verschillende locaties (o.a. zolder en gelijkvloerse verdieping) kleine hoeveelheden van
uitwerpselen van (vermoedelijk) dwergvleermuizen gevonden.
4.2 De functie zomer-, kraam- en paarverblijfplaats in gebouwen
Er zijn geen gegevens uit literatuur bekend van gebruik van gebouwen of andere
objecten van het Schoonselhof als zomerverblijfplaats of kraamverblijfplaats.
Er werden op verschillende tijdstippen uitvliegende gewone dwergvleermuis
waargenomen ter hoogte van het noordwestelijk deel van Neerhoeve. De exacte
uitvlieglocatie kon niet bepaald worden. Er werden kleine hoeveelheden uitwerpselen van
dwergvleermuis gevonden in de Neerhoeve, zowel op de gelijkvloerse verdieping als op
de zolderverdieping. Dit wijst op het gebruik van het gebouw als zomerverblijfplaats
door dwergvleermuis. Van grootoorvleermuis is bekend dat ze vaak overwinteren in hun
zomerverblijfplaats. De aanwezigheid van een overwinteringsplaats op de zolder van de
Neerhoeve kan dus wijzen op de aanwezigheid van een zomerverblijfplaats. De houten
luiken en de gevel van de Neerhoeve worden als nachtrustplaats gebruikt door gewone
dwergvleermuis, en vermoedelijk ook door grootoorvleermuis en laatvlieger.
In augustus en september 2014 werden baltsvluchten waargenomen van gewone
dwergvleermuis ter hoogte van de Neerhoeve en de kasteelvijver, en baltsvluchten van
ruige dwergvleermuis ter hoogte van het kasteel. Sociale geluiden van grootoorvleermuis
werden waargenomen in de lindendreef ter hoogte van de Neerhoeve. Dit kan dus wijzen
op de aanwezigheid van een kraamverblijfplaats. Op figuur 7 worden de locaties
weergeven waar sociale geluiden van vleermuizen werden waargenomen.

Pagina | 17
Figuur 7. Locaties met waarnemingen van sociale geluiden: gewone dwergvleermuis (lichtblauw),
ruige dwergvleermuis (donkerblauw), watervleermuis (paars), grootoorvleermuis (geel). (bron
luchtfoto: AGIV)
4.3 De functie zomer-, kraam- en paarverblijfplaats in bomen
Er zijn geen gegevens uit literatuur bekend van gebruik van bomen op het Schoonselhof
als zomerverblijfplaats of kraamverblijfplaats.
Er werden geen zomer- of kraamverblijfplaatsen in bomen vastgesteld. Er werd echter
nog geen exhaustief onderzoek naar gevoerd. Sociale geluiden van grootoorvleermuis
werden vastgesteld in de dreven L5, L14, L19b, L29 en L33 (zie figuur 7). De
aanwezigheid van sociale geluiden kan wijzen op de nabijheid van een kolonieplaats of
kolonieplaatsen. In verschillende dreven werden eveneens sociale geluiden en/of
baltsvluchten van gewone dwergvleermuis waargenomen. Er werden tijdens de
inventarisatie geen roepbomen van rosse vleermuis waargenomen.
4.4 De functie jachtgebied
Een jachtgebied is een gebied waar een vleermuis of een groep vleermuizen jagen.
Vleermuizen maken vaak gebruik van verschillende kernjachtgebieden die ze op een
avond na elkaar bezoeken. Minstens 6 soorten vleermuizen gebruiken delen van het

Pagina | 18
Schoonselhof als jachtgebied: gewone dwergvleermuis, ruige dwergvleermuis,
laatvlieger, rosse vleermuis, watervleermuis en gewone grootoorvleermuis. Franjestaart
en baardvleermuis gebruiken het gebied waarschijnlijk als jachtgebied, maar het beeld is
nog erg onvolledig en versnipperd omwille van een beperkt aantal (gedetermineerde)
waarnemingen. Gewone dwergvleermuis werd jagend aangetroffen in het gehele gebied.
De vleermuizen jagen in de omgeving van het kasteel en de Neerhoeve, in het bos, langs
de bosranden, in de bosdreven, boven de afwateringsgrachten, de kasteelvijver, de
langgerekte vijver met 2 verbredingen aan de uiteinden. Rosse vleermuis werd jagend
aangetroffen in het hele gebied. Vaak ging het over dieren die op grote hoogte aan het
jagen waren. Rosse vleermuis werd langdurig jagend waargenomen boven perk U en Z,
boven het groot militair ereperk. Voor de andere soorten worden indicatief zones met een
hogere concentratie aan jachtactiviteit aangeduid. De waargenomen jachtgebieden zijn
verschillend van soort tot soort.
De kernjachtgebieden van gewone
grootoorvleermuis worden aangeduid op figuur
8. Gewone grootoorvleermuis werd jagend
aangetroffen in:
- De omgeving van de grote kasteelvijver
en de lindendreef voor de Neerhoeve (L14)
- De dreven tussen het sterbos en Fort 7
(L17b, L19b, L24, L29). Het betreft telkens
beuken-, kastanje- en eikendreven van
meer dan 100 jaar oud.
- Dreef L5
De kernjachtgebieden van watervleermuis,
franjestaart, en baardvleermuis worden
aangeduid op figuur 9. Watervleermuis werd
jagend aangetroffen boven:
- De grote kasteelvijver
- Grenssloot ter hoogte van perk P en L
- Grensloot ter hoogte van perk 5
Er is slechts beperkt zicht op het gebruik van
delen van het domein als jachtgebied van
franjestaart en baardvleermuis. Franjestaart
en baardvleermuis werden met zekerheid
jagend waargenomen in de oostelijke
beukendreef (L39a) in het noordelijk parkbos.
Baardvleermuis werd eveneens jagend
waargenomen in de beukendreef (L17b) ten
oosten van het sterbos. Beiden soorten zijn
uitgesproken bosbewonende soorten. Het
aandeel jagende vleermuizen in boshabitat
wordt in de regel onderschat bij
inventarisaties met batdetectors. Vleermuizen
jagen vaak hoog in de kruinen, en worden
Figuur 8. Kernjachtgebieden van grootoor-
vleermuis (bron luchtfoto: AGIV)
Figuur 9. Kernjachtgebieden van water-
vleermuis (blauw) en overige Myotis - soorten
(bruin) (bron luchtfoto: AGIV)

Pagina | 19
onvoldoende gedetecteerd door
batdetectors (Rieger & Nagel, 2007), of
zijn niet op soort te determineren
(omwille van zachte echolocatiegeluiden
of omwille van gebrekkige kwaliteit van
de geluidsopnames).
De kernjachtgebieden van laatvlieger
worden aangegeven op figuur 10.
Laatvlieger werd jagend aangetroffen in:
- De omgeving van het kasteelvijver
en Neerhoeve, incl. lindendreef voor
de Neerhoeve (L14) en het
Columbarium.
- In de dreven (L1, L2, L10a, L10b,
L11a, L11b, L12) en boven de
grafperken (perk N, O, U, W2, groot
militair ereperk, perk 3, 4 en 5)
4.5 De functie vliegroute/verbindingsfunctie
Een vliegroute is een vaste route van een vleermuis of een groep van vleermuizen vanaf
een verblijfplaats naar een foerageergebied of tussen verblijfplaatsen en vice versa. Er is
nog verder onderzoek noodzakelijk om een volledig zicht te krijgen op de aanwezigheid
van vliegroutes op de begraafplaats. Op basis van de verzamelde gegevens, worden
voorlopig volgende conclusies genomen. Er zijn vier vliegroutes vastgesteld in het
gebied. De eerste route loopt langs de dreef van geknotte zomerlinden (L1). De route
wordt voornamelijk gebruikt door gewone dwergvleermuis en laatvlieger.
Dwergvleermuizen die verblijven in huizen buiten het Schoonselhof bereiken
vermoedelijk hun jachtgebieden via deze dreef. Grootoorvleermuis gebruikt de
lindendreef (L14) als vliegroute van de Neerhoeve naar de kasteelvijver. Laatvlieger werd
eveneens op route waargenomen aan de buitenkant van de dreef. Een vliegroute loopt in
noord-zuid richting langs de dreef van oude beuken (L17b) en vervolgens west-oost via
de kastanje-dreef (L24). De vliegroute wordt gebruikt door Myotis-soorten
(watervleermuis, baardvleermuis), laatvlieger, gewone dwergvleermuis en gewone
grootoorvleermuis. De vliegroute werd bij avondschemer in beide richtingen gebruikt.
Dit is een indicatie van de aanwezigheid van verschillende verblijfplaatsen. Het is niet uit
te sluiten dat de verblijfplaatsen in de dreven zelf gelokaliseerd zijn.
De dreef (L39a) wordt als vliegroute gebruikt door Myotis-soorten met o.a.
baardvleermuis en franjestaart. Er is mogelijks een verblijfplaats van deze soorten in de
dreef of om het omliggende bos aanwezig.
Figuur 10. Kernjachtgebieden van laatvlieger (bron
luchtfoto: AGIV)

Pagina | 20
4.6 De functie zwermplaats
Zwermen wordt omschreven als het bezoeken van een groot aantal vleermuizen van
winterverblijven vóór de winterslaap. Aan het zwermgedrag worden twee functies
toegeschreven: voortplanting en verkenning van de mogelijkheden voor overwintering.
Zwermen is een essentieel onderdeel in de levenscyclus van vleermuizen, en
zwermlocaties zijn van groot belang voor de bescherming van de vleermuizen (Dietz et
al., 2011, Vleermuisvakberaad Netwerk Groene Bureaus, Zoogdiervereniging en
Gegevensautoriteit Natuur, 2013). Er wordt eveneens een andere vorm van zwermen
waargenomen: het ochtendzwermen. Vleermuizen zwermen dan enige tijd in de directe
omgeving van de verblijfplaats voordat ze deze binnengaan om te slapen.
Op 3 september werd zwermgedrag van vleermuizen waargenomen boven één van de
afwateringsgrachten op de grens met Fort 7 (zie figuur 12). Er werden minimaal 10
watervleermuizen, 10 dwergvleermuizen en 2 baardvleermuizen zwermend
waargenomen. Er werd geen ander zwermgedrag waargenomen. De zwermlocatie is
gelegen vlakbij een vliegroute van vleermuizen op Fort 7 richting het reduit en richting
de fortgracht. De vliegroute wordt gebruikt door Myotis-soorten (baardvleermuis,
watervleermuis), gewone grootoorvleermuis en gewone dwergvleermuis (De Ridder et
al., 2014). Aangezien er geen ochtendtellingen werden uitgevoerd, werd geen
ochtendzwermgedrag vastgesteld.
Figuur 11. Waargenomen vliegroutes op het Schoonselhof (bron luchtfoto: AGIV)

Pagina | 21
Figuur 12. Vastgestelde zwermlocatie op de grens met Fort 7 (Bron: 51°09’57.33” N 4°22’16.73” O
543 m. GOOGLE EARTH 7 juli 2013. 15 januari 2015.)
4.7 Relatie tussen Fort 7 en begraafplaats Schoonselhof
De meeste vleermuizen maken gebruik van een netwerk van (kern)jachtgebieden. De
grootte van de jachtgebieden en de nabijheid van voldoende zomerverblijven,
kraamverblijven en winterverblijfplaatsen is van belang om een voldoende grote en
stabiele populatie te ontwikkelen. August et al. (2014) schatten dat de home-range voor
een kolonie van watervleermuis of franjestaart tussen 10 en 30 ha bedraagt. De home-
range voor grootoorvleermuis is maximaal 40 ha. Verschillende auteurs geven aan dat de
dieren meestal op 500 – 1000 meter van hun verblijfplaats foerageren (Entwistle et al.,
1998 en 2000; Dietz et al., 2011). Fort 7 en begraafplaats Schoonselhof zijn daarom
vermoedelijke onderdeel van een functioneel geheel.
Beide gebieden zijn belangrijk leefgebied binnen de levenscyclus voor vleermuizen. Er
wordt gedurende het hele voorjaar, zomer en nazomerseizoen activiteit van vleermuizen
waargenomen. Tabel 2. geeft de soortenverdeling van Fort 7 en Schoonselhof weer zoals
waargenomen op de vaste meetpunten (De Ridder et al., 2014). We stelden vast dat het
aandeel watervleermuis en gewone dwergvleermuis gelijkaardig is. Op het Schoonselhof
werd echter in verhouding meer activiteit van laatvlieger en rosse vleermuis
waargenomen. Minstens 4 soorten gebruiken beiden uitgesproken als jachtgebied:
gewone dwergvleermuis, ruige dwergvleermuis, watervleermuis en gewone
grootoorvleermuis. Laatvlieger, rosse vleermuis en baardvleermuis worden in beide
gebieden waargenomen. Hert Schoonselhof is een belangrijk jachtgebied voor laatvlieger.
De activiteit van laatvlieger en rosse vleermuis is vermoedelijk hoger op het
Schoonselhof dan op Fort 7. Voor baardvleermuis is dit omgekeerd.
Zwermgedrag van gewone dwergvleermuis, baardvleermuis en watervleermuis werd
waargenomen op de grens tussen Fort 7 en het Schoonselhof. Zwermgedrag van
gewone dwergvleermuis, baardvleermuis, watervleermuis, laatvlieger en
grootoorvleermuis werd waargenomen in en rond het reduit van Fort 7. De afstand van
de Neerhoeve tot aan het reduit van Fort 7 bedraagt in vogelvlucht ongeveer 700 meter.

Pagina | 22
Fort 7 is een gekende winterverblijfplaats met jaarlijks 20 – 35 overwinterende
vleermuizen. Het aantal overwinterende dieren is jaarlijks stijgende, na een terugval
vanaf de winter 2007 – 2008. Minstens 5 soorten gebruiken Fort 7 regelmatig als
winterverblijfplaats: baard-/Brandt’s vleermuis, watervleermuis, gewone dwergvleermuis,
franjestaart en gewone grootoorvleermuis. Een aantal soorten worden zelden of
onregelmatig overwinterend aangetroffen op Fort 7. Laatvlieger wordt onregelmatig
overwinterend teruggevonden. Het is geen typische fortsoort. De meest algemene
overwinterende soorten vleermuizen in de fortengordel zijn watervleermuis (33%),
baard/Brandts vleermuis (31%) en franjestaart (15%) (Boers et al., 2013). Op Fort 7 is
de meest algemene overwinterende soort de baard/Brandts vleermuis. Watervleermuis
was t.e.m. de winter van 2007 – 2008 de tweede belangrijkste soort, gevolgd door
gewone dwergvleermuis als 3e soort. Vanaf de winter van 2008 – 2009 is gewone
dwergvleermuis de tweede belangrijkste soort.
De Neerhoeve op het Schoonselhof heeft een belang als overwinteringsplaats voor
grootoorvleermuis. Het aantal getelde overwinterende dieren is nog beperkt, maar dit is
in overeenstemming met gekende gegevens van grootoorvleermuis. Bovendien wordt bij
telling van kolonies en winterverblijven het effectieve aantal aanwezige vleermuizen
onderschat. Over de andere gebouwen en constructies is nog te weinig geweten.
Over het belang van beide gebieden als zomerverblijfplaats is momenteel nog
onvoldoende gekend.
5. Aanbevelingen m.b.t. beheer
5.1 Algemeen
De begraafplaats Schoonselhof is, samen met de forten 6, 7 en 8, het domein Klaverblad
en de vallei van de Struisbeek, een belangrijk leefgebied voor vleermuizen in de
stedelijke zuidrand van de stad Antwerpen (district Wilrijk). Een aantal van de
voorkomende vleermuizensoorten kregen de status kwetsbaar (laatvlieger en rosse
vleermuis) en bijna in gevaar (gewone grootoorvleermuis en watervleermuis) op de
Vlaamse Rode lijst. Van een aantal voorkomende soorten zijn onvoldoende data
beschikbaar over de verspreiding in Vlaanderen om de bedreiging voldoende te kunnen
inschatten.
Er werd in 2011 door Arcadis een beheerplan opgemaakt voor het Schoonselhof.
Vleermuizen werden in dit beheerplan niet als specifieke doelsoort of indicatorsoort
opgenomen. In een toekomstig beheerplan, of in een actualisatie krijgt het aanbeveling
om specifieke vleermuissoorten als doelsoort mee op te nemen. Gewone
grootoorvleermuis, laatvlieger, watervleermuis, baardvleermuis en franjestaart komen
hiervoor in aanmerking.
Vleermuizen hebben een complexe seizoensgebonden levenswijze waarbij ze vaak van
jachtgebieden en verblijfplaatsen wisselen. Aangepast beheer voor vleermuizen richt zich
in de eerste plaats dan ook naar het behouden en versterken van de jachtgebieden, de
verblijfplaatsen en de verbindingen er tussen. Alle vleermuizen in Europa zijn
insectenetende zoogdieren. Een leefgebied is interessant als er jachtgebieden met
voldoende insectendensiteit beschikbaar zijn. Het vermijden van het gebruik van
insecticiden als algemene maatregel is daarom aanbevolen.

Pagina | 23
Figuur 4. en tabel 6. in de bijlagen geven een aantal aandachtspunten weer voor het
beheer van de parkbegraafplaats Schoonselhof:
- De dreven L14, L17b, L19b, L24, L25 en L29 worden aangeduid als een belangrijk
element voor vleermuizen. Deze elementen dienen gevrijwaard te worden, en verdere
opvolging in functie van vleermuizen bij kap- en snoeiwerken wordt aanbevolen om
risico op slachtoffers en schade te vermijden.
- De dreven L21, L32 en L39a zijn voorzien om gekapt en hersteld te worden in kader
van het beheerplan. Aanvullend onderzoek op de aanwezigheid van verblijfplaatsen
van vleermuizen voorafgaand de kapwerken is noodzakelijk om risico op slachtoffers
en schade te vermijden.
- De kappingen van het perceel A (noordelijk parkbos, perceel 1g) dienen voorafgegaan
te worden van een aanvullend onderzoek op aanwezigheid van verblijfplaatsen van
vleermuizen.
- De verjonging van het perceel B (sterbos, perceel 2c) dient gefaseerd in de tijd
uitgevoerd te worden, met een behoud van minimum 10 potentiële vleermuisbomen
per ha in het bos of aanpalende dreven. Aanvullend onderzoek voorafgaand aan de
kappingen en opvolging in functie van vleermuizen bij kap- en snoeiwerken wordt
aanbevolen om risico op slachtoffers en schade te vermijden.
- De restauratie en de functie-invulling van de Neerhoeve (perceel C) dient te
gebeuren afgestemd op de vereisten voor een verblijfplaats voor minimaal gewone
grootoorvleermuis.
- De zwermlocatie (perceel D) dient van verstoring (o.a. licht) gevrijwaard te worden.
De oppervlakte open water en randbegroeiing dient behouden te blijven.
In het verdere hoofdstuk 5 van dit rapport wordt ingegaan op deze beheeraanbevelingen
en worden een aantal algemene aanbevelingen m.b.t. het beheer op korte en
middellange termijn meegegeven. Er wordt eveneens ingegaan op specifieke
aandachtspunten m.b.t. habitatvereisten van grootoorvleermuis en laatvlieger.
De levenscyclus van veel soorten is slechts fragmentarisch gekend. De inzichten van
buitenlands onderzoek zijn niet altijd eenvoudig toe te passen in Vlaanderen. De
beschrijving van de habitatvereisten zijn in onderstaande paragrafen gebaseerd op Lange
et al. (1994), Kapteyn (1995), Limpens et al. (1997), Robinson & Stebbings (1997),
Kervyn (1998), Motte et al. (1997), Boonman (2000), Entwistle et al. (2000), Meschede
& Heller (2000), Schober et al. (2001), Simon et al. (2004), Zukal & Ŕehák (2006),
Encarnação (2006, 2007, 2012), Twisk et al. (2010), Dietz et al. (2011) en Buckley et
al. (2012). De aanbevelingen voor beheer en randvoorwaarden werden gebaseerd op
Verkem en Verhagen (1998), Boye en Dietz (2005), Paelinckx, et al. (2009),
Zoogdiervereniging & Probos (2012) en de Nederlandse soortenstandaard voor gewone
grootoorvleermuis, ruige en gewone dwergvleermuis en watervleermuis (Dienst
Regelingen. Ministerie van Economische Zaken, Landbouw & Innovatie, 2011 en
2013). Deze soortenstandaard bevat informatie over de beschermde soort en zijn
leefomgeving, regels en hun toepassing, onderzoek en mogelijke maatregelen.

Pagina | 24
5.2 Dreven en bossen
Watervleermuis, franjestaart, baardvleermuis, rosse vleermuis en gewone
grootoorvleermuis zijn boombewonende vleermuizen. Deze soorten gebruiken bomen als
dagrustplaats, nachtrustplaats, zomerverblijf, kraamverblijf en winterverblijfplaats.
Watervleermuis wordt vooral in grotere dichtheden gevonden waar zowel beschut water
als ouder bos of oudere bomen aanwezig zijn. Voor watervleermuis vormen
verblijfplaatsen in bomen een beperkende factor. De meest gebruikte holten in bomen
zijn verlaten spechtengaten en natuurlijk ontstane inrottingsgaten. Boomkolonies van
baardvleermuis en franjestaart, maar ook ruige dwergvleermuis zitten dan weer vaker
achter losse schors of in boomscheuren. De (kraam)groepen van deze soorten leven in
een metapopulatieverband en gebruiken gelijktijdig meerdere koloniebomen, waartussen
de dieren regelmatig verhuizen. De
koloniebomen zijn niet op elk moment
bezet, en splitsen zich soms op of
voegen zich samen. De wisseling van
verblijfplaatsen gebeurt om de 2 à 5
dagen, maar is sterk afhankelijk van de
soort. Uit onderzoek in het buitenland
bleekt dat een kraamkolonie nood heeft
aan 40 – 70 geschikte holten. De
vleermuizen zijn vaak erg plaatstrouw.
Lučan et al. (2009) stelden vast dat
kraamverblijven van watervleermuizen
gedurende meer dan 1 seizoen gebruikt
werden. 35% van de holten werden zelfs
voor meer dan 5 jaar gebruikt.
Kraamverblijfplaatsen worden gebruikt
vanaf half april tot half september,
waarbij in de periode juni tot en met
augustus ook jongen aanwezig kunnen
zijn. De afstand dat vleermuizen
bewegen tussen twee verblijfplaatsen is
variabel maar ligt tussen de 700 en 1000
meter.
In de dreven en in het noordelijk parkbos en sterbos van het Schoonselhof zijn al
verschillende oude bomen met loshangende schors, scheuren, spleten en holten
aanwezig. Dit zijn potentiële vleermuizenbomen. Bij uitvoering van beheermaatregelen
moet daarom voldoende aandacht uitgaan naar vleermuizen. De uitvoering van
kappingen en snoeiwerken gebeurt dan het best onder begeleiding van een
vleermuisdeskundige, of voorafgegaan door een gerichte controle op aanwezigheid van
vleermuizen. De volgende krachtlijnen voor het beheer kunnen meegegeven worden:
- Oude bomen dienen maximaal behouden te blijven, jongere bomen moeten oud
kunnen worden;
- Gekende verblijfplaatsen van vleermuizen in bomen zo veel mogelijk veilig stellen;
- Herstel of kappen in de dreven of bossen gefaseerd uitvoeren met prioriteit aan niet
vleermuisbomen;
- Bij uitvoering van verjongingen of onderhoudskappen in dreven, specifieke controle
Figuur 13. Potentiële vleermuisboom in dreef L29

Pagina | 25
laten uitvoeren van de geviseerde bomen door een vleermuisdeskundige op
aanwezigheid van vleermuizen om kapslachtoffers te vermijden;
- Kappingen van oude loofbomen uitvoeren in de voor vleermuizen minst gevoelige
periode. Dit is het vroege voorjaar (april) of najaar (half september-eind oktober),
maar alleen als er geen paarverblijven aanwezig zijn.
Het toepassen van criteria en
richtlijnen van duurzaam park- en
bosbeheer heeft in regel een positief
effect op de natuurwaarden van de
begraafplaats, en bijgevolg ook
onrechtstreeks op de aanwezige
vleermuispopulatie. Het middellange en
lange termijnbos- en drevenbeheer
dient zich in de eerste plaats te richten
op het behoud van voldoende oude
bomen (80 tot 120 jaar of ouder) met
voor vleermuizen geschikte holten en
natuurlijke scheuren. Streefbeeld is het
permanent voorzien van 25-30 holten
per hectare oud bosbestand. Dit komt
overeen met behoud van minstens tien
vleermuisbomen (potentiële of
werkelijke) per hectare. Het
beheerplan voorziet voor de verjonging
van het sterbos (bestand 2C, zie
bijlagen, figuur 2 en 4, vlak B) een
eindkap na twee opeenvolgende
groepenkappen en een gedeeltelijk herstel van de radiale dreven met intern
hakhoutbeheer. In het aansluitend bestand (2b) worden reguliere dunnningen in functie
van veiligheid voorzien. De vleermuizenactiviteit werd voornamelijk in de dreven (L17,
L19 en L41) rondom het bos waargenomen en langs het bospad tussen bestand 2b en
2C. De aanwezigheid van verblijfplaatsen in het bos is echter nog niet uit te sluiten. De
groepenkappen wordt daarom best voldoende gespreid in de tijd, zodat gedurende de
gehele verjongingsperiode het streefbeeld van minstens 10 vleermuizenbomen per
hectare behouden blijft. Indien dit niet haalbaar is, bijvoorbeeld omwille van
veiligheidsredenen, dan kan het ophangen van vleermuiskasten een tijdelijke uitkomst
bieden. Vleermuizen jagen vaak langs bosranden, in dreven en open plekken. Het
versterken van de radiale structuur zal dus mogelijks een positief effect hebben, net
zoals het voorzien van een open plek aan de binnenrand van de berceau (bestand 2a).
Voor het noordelijk parkbos (bestand 1) worden voornamelijk groepenkappen aan de
straatzijde en de oevers voorzien met specifiek exotenbeheer, ontwikkelen van
bosranden en verder nulbeheer. Het beheerplan voorziet ook op andere locaties een
aantal maatregelen om het aandeel exoten in bos en dreven te beheersen. Voor jonge
opslag en spontane opslag is dat geen probleem. Bij oudere bomen is hier extra aandacht
nodig. Uit onderzoek blijkt dat het aandeel Amerikaanse eiken met holten 3 à 4 maal
groter is dan het aandeel zomereiken (Strypstein, 2004). Vleermuizenverblijven worden
vaak gevonden in oude Amerikaanse eiken, maar ook in Robinia (bv. ruige
dwergvleermuis). Specifieke aandacht dient dan ook uit te gaan naar vrijwaren van deze
Figuur 14. Vliegroute van vleermuizen in beukendreef
L17b

Pagina | 26
bomen indien er holten, scheuren of loshangende schors aanwezig zijn. De ontwikkeling
van bosranden hebben naar verwachting een positief effect op vleermuizen.
Voor de beukendreef L39a is in het
beheerplan een volledige restauratie
van het dreefstuk tegelijkertijd met de
creatie van een zichtas voorzien. Ter
hoogte van het bosbestand 1g is
hiervoor een eindkap (creëren open
plek) voorzien. De dreef en omgeving
worden gekenmerkt door oude bomen
(>100 jaar oud), waarvan een aantal in
minder goede conditie of aftakelend. In
deze dreef werd met zekerheid
franjestaart en baardvleermuis
aangetroffen. Extra aandacht voor
vleermuizen bij de kappingen en
restauratie is aangewezen.
Boomkolonies van baardvleermuis en
franjestaart zitten vaak achter losse
schors of in boomscheuren.
Baardvleermuis leeft jaarrond in
kolonies. Er wordt een netwerk van
verblijfplaatsen bewoond, die om de 10
à 14 dagen worden gewisseld. Het
jachtgebied ligt vaak binnen een straal
van 1 km rondom de kolonieplaats.
Franjestaart maakt gebruik van een
netwerk van verblijfplaatsen, die om de 4 à 8 dagen worden gewisseld. Het jachtgebied
ligt vaak binnen een straal van 1,5 tot 3 km rond de kolonieplaats. Er kan niet
uitgesloten worden dat verblijfplaatsen aanwezig zijn in de dreef en het bosbestand 1g
(zie bijlagen, figuur 4, vlak A). Kappingen en snoeiwerken kunnen pas gebeuren na een
voorgaande controle door een vleermuizenexpert. Bij het vaststellen van de
aanwezigheid van vleermuizen(verblijfplaatsen) dienen specifieke maatregelen genomen
worden om de bomen te vrijwaren, of om slachtoffers bij het kappen te vermijden (zie
ook algemene krachtlijnen beheer). Bij het aanbrengen van bescherming tegen
zonnebrand, dient er over gewaakt te worden dat de toegangen tot verblijfplaatsen niet
worden afgesloten.
Voor de linden- en eikendreef L21 wordt kruinverzorging op korte termijn voorzien,
gevolgd door een volledige vervanging op lange termijn. Een groot deel van de bomen is
aftakelend. Een controle op aanwezigheid van verblijfplaatsen van ruige dwergvleermuis,
gewone grootoorvleermuis en watervleermuis is aan te bevelen. De zomereikendreef L32
is aftakelend. Het beheerplan voorziet om deze op termijn te vervangen. De dreef sluit
aan op de zomereikendreef L29, waar kruinverzorging wordt voorzien. In beide dreven
werd jachtactiviteit van grootoorvleermuis waargenomen, en in dreef L29 zijn er
indicaties voor de aanwezigheid van verblijfplaatsen van deze soort (sociale roepen in
kraamperiode). In de dreven L14, L17b en L19b werd hoge jachtactiviteit van gewone
grootoorvleermuis vastgesteld. Er zijn eveneens indicaties voor de aanwezigheid van
Figuur 15. Begin van de dreef L39a

Pagina | 27
verblijfplaatsen (sociale roepen en ligging op een vliegroute van grootoorvleermuis,
watervleermuis en andere Myotis-soorten). Kappingen en kroonverzorging kunnen pas
gebeuren na een voorafgaande controle door een vleermuizenexpert. Bij het vaststellen
van de aanwezigheid van vleermuizen(verblijfplaatsen) dienen specifieke maatregelen
genomen worden om de bomen te vrijwaren, of om slachtoffers bij het kappen te
vermijden (zie ook algemene krachtlijnen beheer).
5.3 Vijvers en grachten
Watervleermuizen jagen vlak boven het wateroppervlak van beschutte en vlakke
waterpartijen (Rydell et al., 1999; Warren et al., 2000, Dietz et al., 2011). Gewone
dwergvleermuis en ruige dwergvleermuis worden ook vaak jagend in de buurt van of
boven wateroppervlakten waargenomen. Kwalitatieve jachtgebieden hebben een
voldoende hoge insectendensiteit. In praktijk blijken dit vooral beschutte zones te zijn.
Bij windstil weer wordt de beschutting minder belangrijk. Watervleermuis profiteert
bovendien van eerder eutrofe waters (Vaughan et al. 1997, Warren et al. 2000), waar
een hoger aandeel Chironomidae voorkomen.
Voor watervleermuis wordt aangegeven dat het jachtgebied meestal binnen 2 à 4
kilometer van de verblijfplaats ligt. De kernjachtgebieden hebben oppervlakten van
100m² tot 7500m². De oppervlakte geschikt jachthabitat voor watervleermuis in het
Schoonselhof is relatief beperkt. De kernjachtgebieden voor watervleermuis zijn de
kasteelvijver en enkele voldoende brede en waterhoudende afwateringsgrachten. De
overige grachten zijn onvoldoende waterhoudend, of zijn met kruid dichtgegroeid. Een
aantal grachten lijken geschikt, maar er werden geen jagende dieren boven
waargenomen. Het behoud van de huidige wateroppervlakte is een minimumvereiste
voor het in stand houden van de vleermuizenpopulatie. Bovendien is het noodzakelijk dat
de kwaliteit van de jachtgebieden optimaal is. De kernjachtgebieden van
watervleermuizen moeten zo veel mogelijk gevrijwaard blijven van verlichting. Voor de
kasteelvijver wordt gestreefd naar het behouden van een grote oppervlakte water met
voldoende opgaande oeverbegroeiing. Watervleermuizen jagen niet boven water in
perioden van algenbloei, als het wateroppervlak is dichtgegroeid met kroos of waterlelies
(persoonlijke observaties). Een
waterbeheer gericht op het
natuurlijker maken van het
vissoortenbestand en het verhogen
van het aandeel onder-waterplanten
zal bijdragen aan een gunstiger
jachtbiotoop voor watervleermuis en
andere soorten. Voor de grachten is
het noodzakelijk dat er een voldoende
evenwicht gezocht wordt tussen
grachten met open water en grachten
met begroeiing (riet, gele lis). De
grachten worden best regelmatig
geruimd zodat hier ook een voldoende
kwalitatief jachthabitat kan
ontwikkelen, met weinig hindernissen
voor vleermuizen die jagen boven het
wateroppervlak. Het uitbaggeren van
de waterpartijen zal op lange termijn
hoofdzakelijk een positief effect op
Figuur 16. Jachtgebied van gewone en ruige
dwergvleermuis en watervleermuis

Pagina | 28
vleermuizen hebben, door een groter en gevarieerder aanbod van insecten met een
aquatisch larvaal stadium, via de verbetering van de zuurstofvoorziening in de
bodemlaag. De werkzaamheden worden best uitgevoerd buiten het actieve seizoen van
vleermuizen.
Eén van de grenssloten met Fort 7 (figuur 17) is een zwermplaats voor watervleermuis,
baardvleermuis en gewone dwergvleermuis. De locatie dient gevrijwaard te blijven, o.a.
door regelmatige slibruiming, open houden van het wateroppervlak, verwijderen van
omgevallen bomen uit de sloot. Verlichting van deze zone moet absoluut vermeden te
worden.
Figuur 17. Zwermlocatie op de grens met Fort 7
5.4 Beheermaatregelen voor behoud en versterken habitat van laatvlieger
De begraafplaats Schoonselhof is een belangrijk jachtgebied voor laatvlieger. Een
functionele relatie met Fort 7 en het domein Klaverblad wordt verondersteld. Zowel op
fort 7 als op het domein Klaverblad werden regelmatig jagende laatvliegers
waargenomen. Bij de wintertellingen in Fort 7 worden elk jaar 1 of 2 overwinterende
laatvliegers gevonden. Laatvlieger jaagt in relatief grote jachtgebieden. De literatuur
geeft een home-range aan van 4,6 km² tot 77 km² voor kolonies met dichtheden van 0,5
tot 3 dieren per 100 ha. Het jachtgebied ligt in de regel binnen 1 tot 2 kilometer van de
verblijfplaatsen, maar afstanden tot 7 km werden waargenomen. De dichtstbijzijnde
gekende kolonie laatvliegers is gesitueerd in de kerk van Niel op minder dan 7 km van
het Schoonselhof. Dit is binnen de home-range van laatvlieger. De dieren jagen vaak
direct na het uitvliegen gezamenlijk op zogenaamde sociale foerageerplaatsen, om zich
nadien over het landschap te verspreiden. De verblijfplaatsen zijn meestal gebouwen

Pagina | 29
(zowel in zomer als in winter). De kolonies zijn bovendien erg plaatstrouw. Laatvliegers
jagen meestal maar enkel uren per nacht, de rest van de tijd wordt voornamelijk in de
verblijfplaats of op route doorgebracht. De dieren vliegen uit een half uur tot 3 kwartier
na zonsondergang.
Laatvliegers foerageren in open en halfopen gebieden, boven weilanden, perceelsranden
en op enkele meters van opgaande begroeiing, vaak in aansluiting tot een bosrijk gebied.
Simon et al. (2004) stelden bij telemetrisch onderzoek in Duitsland vast dat weilanden
tot het voorkeursjachthabitat van laatvlieger behoren. Ze stelden bovendien een
positieve correlatie vast tussen de aanwezigheid van kraamkolonies van laatvlieger en
(door runderen) begraasde weilanden. Verboom en Huitema (1997) argumenteren dat
landschappen met een hoge densiteit aan lineaire elementen een meerwaarde hebben
voor laatvlieger. De keuze van jachtbiotopen van laatvlieger is sterk afhankelijk van
prooibeschikbaarheid, en is seizoensgebonden. Studies geven aan dat laatvliegers in de
periode mei – juni jagen langs randen van loofbossen en kleine landschapselementen. Ze
jagen dan o.a. op meikevers (Melolontha-soorten) en junikevers (Amphimallon
solstitiale). In de periode van augustus tot oktober jagen ze meer boven graasweiden op
zoek naar o.a. mestkevers (Aphodius spp.). Ook voor langpootmuggen (Tipulidae) zijn ze
aangewezen op (vochtige) weilanden in juni en in de periode september tot oktober. De
dieren jagen jaarrond op grote nachtvlinders (Heterocera).
De afwisseling van bos, dreven, hagen, droge en vochtige graslanden, bermen en taluds
op het Schoonselhof biedt een ideaal geheel van jachtbiotopen voor laatvliegers. Het
behoud van dit evenwicht is dan ook van belang, net zoals een natuurlijk beheer van
bossen en de ontwikkeling van soortenrijke bosranden. Een haagbeheer gericht op het
bevoordelen van inheemse soorten zal bijdragen aan het creëren van geschikt
jachthabitat. Laatvliegers zouden in hogere dichtheden voorkomen in gebieden met
voldoende oppervlakte grasland en graasweide. Het behoud en versterken van
oppervlakten grasland, bermen en taluds met een extensief beheer op het Schoonselhof
is dan ook van belang om de laatvliegers in de zoogperiode van voldoende stapelvoedsel
te voorzien. Het vermijden van het gebruik van pesticiden om bv. engerlingen te
bestrijden krijgt aanbeveling.
5.5 Beheermaatregelen voor behoud en versterken habitat grootoorvleermuis
Figuur 5. In de bijlagen geeft het habitatgebruik en de vastgestelde vleermuizenfuncties
voor grootoorvleermuis weer op het Schoonselhof. Gewone grootoorvleermuis is een
typische soort van bosgebieden, en in Vlaanderen ook van parken. De zomerverblijven
zijn gesitueerd in holle bomen, maar ook vaak in gebouwen. Het gaat dan meestal over
(kerk)zolders, waar de dieren tussen de pannen, latten en balken worden gevonden. Er
wordt een netwerk van verblijfplaatsen bewoond. De winterverblijven en paarverblijven
zijn gesitueerd in (ijs)kelders, bunkers, forten, etc. Het is een standsoort die slechts
korte afstanden (vaak beperkt tot 1 km) aflegt tussen zomer- en winterverblijfplaats.
Fort 7 is een gekende winterverblijfplaats van grootoorvleermuis. Net zoals bij laatvlieger
is een functionele relatie tussen fort 7 en het Schoonselhof niet uit te sluiten.
Grootoorvleermuizen volgen vliegroutes langs lijnvormige landschapselementen. In de
zomer installeren ze zich in kraamkolonies in boomholten en in warme plekken van
gebouwen. Kraamkolonies van vrouwtjes bezetten vaak een klein grondgebied van
ongeveer 1 km². Het jachtgebied is gelegen binnen een straal van enkele honderden
meter tot 2 km rondom de kolonieplaats. De (kraam)groep leeft in een netwerk van een

Pagina | 30
groot aantal bij elkaar gelegen
verblijfplaatsen, waartussen vaak
verhuisd wordt. Verblijven in bomen
worden soms binnen een gebied van
een paar honderd meter om de 1 tot 5
dagen gewisseld. De dieren zijn meer
plaatstrouw bij verblijf in gebouwen,
maar wisselen dan wel regelmatig van
locatie in het gebouw. In de zomer
leven vrouwtjes in kleine
kraamkolonies (gem. 5-30 dieren).
Mannetjes verblijven in kleine groepjes
in de omgeving van de kraamkolonie.
De kraamtijd is van half juni tot half
augustus. Paargroepen worden
gevormd vanaf augustus. In de periode
van augustus tot oktober vindt een
sterk zwermgedrag plaats. Dit
zwermgedrag werd in 2013 op Fort 7
waargenomen.
Grootoorvleermuizen vliegen laat uit, meestal als het al voldoende donker is. Ze zijn dan
ook gevoelig voor lichtverstoring. De jachtgebieden zijn bos- en parkachtige gebieden en
plantsoenen. Ze jaagt voornamelijk op beschutte plekken in het bos, boven bospaden en
open plekken, langs bosranden, dreven en laag boven (bloeiende) kruidenvegetaties. De
belangrijkste prooien van grootoorvleermuizen zijn nachtvlinders (Noctuidae) en
oorwormen (Dermaptera). Omwille van de prooikeuze zijn ze weinig afhankelijk van
activiteitspieken van insecten en jagen in de regel aan één stuk door. De
kernjachtgebieden van een grootoorvleermuis zijn gewoonlijk kleiner dan 1 ha. Dietz et
al. (2011) geeft een maximum van 4 hectare aan. Gewone grootoorvleermuis werd
jagend aangetroffen op 3 locaties. Het is niet uit te sluiten dat dit 3 groepen
grootoorvleermuizen betreft. Op deze 3 locaties is extra aandacht nodig voor het
vermijden van lichtverstoring, en voor het behoud van de dreven met oude bomen, en
voor de ontwikkeling van vegetatierijke bosranden of bermen. Het gebruik van
bestrijdingsmiddelen dient absoluut vermeden te worden.
In de dreven waar (hoge) jachtactiviteit of sociale geluiden van gewone
grootoorvleermuis werden vastgesteld kunnen kappingen en kroonverzorging pas
gebeuren na een voorafgaande controle door een vleermuizenexpert. Het gaat minimaal
over de dreven: L14, L17b, L19b, L24 en L29.
De Neerhoeve is een vastgestelde zomerverblijfplaats van gewone dwergvleermuis en
een winterverblijfplaats van gewone grootoorvleermuis. Op basis van de huidige studie
kunnen andere vleermuizenfuncties niet uitgesloten worden. Grootoorvleermuizen
worden meestal slechts in beperkte aantallen bij elkaar aangetroffen in de
winterverblijven: alleen of in groepjes van 2 tot 15 dieren. Dit komt overeen met onze
vaststellingen in de Neerhoeve. De winterverblijfplaatsen zijn in gebruik van half oktober
tot en met begin april. De dieren slapen echter niet constant. Vooral dieren die op
locaties met wisselende temperaturen overwinteren worden, ook bij koud weer (<0
graden Celsius), regelmatig wakker en foerageren dan bijvoorbeeld op zolders op
Figuur 18. Jachtgebied van grootoorvleermuis in
eikendreef L29

Pagina | 31
overwinterende vlinders. De gewenste overwinteringstemperatuur bedraagt 2 – 8°C,
maar de dieren kunnen voor enkele dagen koudere temperaturen verdragen.
Zomerverblijfplaatsen zijn bewoond van eind maart tot en met half oktober. Hierbij is
een voldoende grote ruimte waar de dieren vrij kunnen rondvliegen van belang. De
invlieg- en uitvliegopeningen moeten onbereikbaar zijn voor predators. Dezelfde
verblijfplaatsen kunnen ook in de winter gebruikt worden als winterverblijfplaats.
Bij sloop en renovatiewerkzaamheden aan de Neerhoeve kan de functionaliteit van de
voortplantingsplaatsen en van de zomer-, winter- en paarverblijfplaatsen van
vleermuizen in het gedrang komen. Hierdoor zullen voortplantingsplaatsen en vaste rust-
en verblijfplaatsen verdwijnen of niet meer als zodanig kunnen functioneren.
Holten, nissen e.d. die toegang bieden voor vleermuizen worden vaak tijdelijk of
permanent ontoegankelijk gemaakt of verdwijnen zelfs geheel. Ook kunnen de
klimatologische omstandigheden zich in die spleten e.d. als gevolg van de renovatie
wijzigen door ander materiaalgebruik, betere isolatie, minder luchtdoorstroming e.d..
Figuur 19. De Neerhoeve
Vooral de aanwezigheid van het origineel zadeldak met oorspronkelijke gordingen,
spanten en kepers en leien dakbedekking creëren gunstige omstandigheden voor een
verblijfplaats van grootoorvleermuizen. De dieren overwinteren o.a. in spleten en
uitsparingen in en tussen het hout. Er moet zoveel als mogelijk gezocht worden naar
inrichtingsalternatieven die ook voor de gewone grootoorvleermuis een gunstig effect
hebben, bv. het inrichten van duurzame plekken waar de gewone grootoorvleermuis zijn
vaste rust- en verblijfplaats, en een vaste in- en uitvliegopening kan hebben.

Pagina | 32
Voor verblijfplaatsen zijn onderstaande parameters van belang:
- Microklimaat: temperatuur, temperatuurgradiënt, luchtvochtigheid, afwezigheid van
tocht, en snelheid van opwarmen of afkoelen (bufferwaarde), vochtigheid, geen tocht.
- de in- en uitvliegopeningen zijn bij voorkeur gericht op zuid / zuidwest of noord en de
aanvliegroute moet vrij van obstakels als takken of bomen zijn. Er mag geen lichtbron
vlakbij zijn.
- De verblijfplaats is voldoende ruim zodat er vrije vliegruimte is.
- Ook mag de verblijfplaats niet via bijvoorbeeld een afdakje bereikbaar zijn voor
bijvoorbeeld katten.
- De binnenkant moet ruw zijn (geen glad hout) en niet geverfd.
- Ruimtegebruik en veiligheid (tegen predatoren).
Het gebruik van niet-giftige of aangepaste houtverduurzamingsmiddelen bij
renovatiewerken is een belangrijk aandachtspunt voor het behouden van een
vleermuizenpopulatie.
5.6 Versterken van de connectiviteit met nabijgelegen gebieden
De meeste vleermuizen maken gebruik van een netwerk van (kern)jachtgebieden. De
grootte van de jachtgebieden en de nabijheid van voldoende zomerverblijven,
kraamverblijven en winterverblijfplaatsen is van belang om een voldoende grote en
stabiele populatie te ontwikkelen. Om de parkbegraafplaats Schoonselhof als kwalitatief
vleermuizenhabitat te ontwikkelen, moet er ook voldoende aandacht zijn voor het
versterken van de verbinding met nabijgelegen gebieden. De relatie tussen Fort 7 en de
parkbegraafplaats Schoonselhof werd al in 4.7 besproken. Het Schoonselhof is echter
onderdeel van een groter netwerk van geschikt vleermuizenhabitat. Het park Groen
Neerland ligt op minder dan 2 kilometer afstand van de begraafplaats is een stapsteen
naar Fort 6. Fort 6 en de vallei van de Kleine Struisbeek inclusief de campus van de UA
en het bosgebied Mariënborgh vormen een functioneel geheel voor vleermuizen. De
gebieden liggen binnen een straal van 2,5 kilometer ten oosten van het Schoonselhof.
Kasteeldomein Klaverblad, Fort 8 en park Zorgvliet liggen binnen een straal van 1,5
kilometer ten westen van het Schoonselhof.
De Moerelei vormt de enige verbinding voor vleermuizen tussen de omgeving van het
kasteeldomein Klaverblad en het complex Fort 7 – Schoonselhof (figuur 20). Het is van
essentieel belang om deze laatste open ruimteverbinding te vrijwaren. De zone wordt
momenteel gebruikt als paardenweide door particulieren. De aanplant van geleidende
beplantingen langs deze percelen zal de connectiviteit tussen de gebieden versterken.
Het oostelijk gedeelte van de Moerelei heeft een meer urbaan karakter. Een meer
vleermuisvriendelijke inrichting van de Moerelei is aangewezen. Dit kan o.a. door het
plaatsen van aangepaste, vleermuisvriendelijke straatverlichting en door de aanplant van
geleidende beplantingen.
De verbinding tussen het Schoonselhof en Fort 8 kan versterkt worden door het groener
inrichten van het kruispunt van de Sint-Bernardsesteenweg ter hoogte van de
hoofdingang van het Schoonselhof. Dit kan door bomen langs de bermen aan te
planten en waar mogelijk een middenberm met bomen te creëren (‘hop over’ voor
vleermuizen).

Pagina | 33
Figuur 20. Open ruimteverbinding tussen Klaverblad en Schoonselhof langs de Moerelei (bron:
AGIV)
6. Aanbevelingen voor verder onderzoek
Het onderzoek met batdetectors en een wintertelling gaf alvast een beeld van de
soortenrijkdom van het gebied. Alle vleermuisfuncties van het gebied zijn echter nog niet
volledig in kaart gebracht. Verder onderzoek naar aanwezigheid van
winterverblijfplaatsen, kraamkolonies en zomerverblijfplaatsen kan een beter inzicht
geven in het gebruik van het domein door vleermuizen. Sommige van deze functies
kunnen enkel grondig in kaart gebracht worden door het inzetten van andere
onderzoeksmethoden zoals o.a. vangsttechnieken en telemetrisch onderzoek. Dit is
vooral relevant voor verdere wetenschappelijke onderbouwing, minder in functie van de
beheerplanning. De volgende aanbevelingen voor verder onderzoek (in volgorde van
belangrijkheid) worden gedaan:
- Inspectie van de Neerhoeve op aanwezigheid van zomerverblijfplaats en
kraamkolonies van grootoorvleermuis en laatvlieger, en vaststellen van de in- en
uitvliegopeningen. Dit kan gebeuren door een extra bezoek in het voorjaar aan de
Neerhoeve, eventueel aangevuld met enkele terreinbezoeken om ochtendzwermen
vast te stellen. Deze inspectie is belangrijk om (mogelijke) gevolgen van de
restauratie te kunnen inschatten en/of te vermijden.
- Uitgebreide inventarisatie van oudere bomen met boomholten in functie van
aanwezigheid kraamkolonies en zomerverblijfplaatsen. Prioriteit dient gegeven te
worden aan de dreven waar vervanging wordt vooropgesteld: L21, L24, L32 en L39a,
en de dreven waar hoge vleermuisactiviteit werd vastgesteld: L14, L17b, L19b, L24
en L29. Dit kan gebeuren voorafgaand vooropgestelde beheerwerken.
- Gedetailleerde inspectie van de Neerhoeve als winterverblijfplaats. Dit kan jaarlijks,
2-jaarlijks of 5-jaarlijks gebeuren door vrijwilligers.
- Gedetailleerde inspectie van het kasteel als winterverblijfplaats.
- Inspectie van het kasteel op aanwezigheid van nachtrustplaatsen, en vaststellen van
de in- en uitvliegopeningen.
- Gericht onderzoek naar zwermgedrag op de grens met Fort 7.

Pagina | 34
7. Literatuur
Adriaens D., Adriaens T., & Ameeuw G. (red.) (2008). Ontwikkeling van criteria voor de
beoordeling van de lokale staat van instandhouding van de habitatrichtlijnsoorten.
Rapporten van het Instituut voor Natuur- en Bosonderzoek 2008 (35). Instituut voor Natuur- en Bosonderzoek, Brussel.
Ahlén, I. & Baagøe, H.J., (1999). Use of ultrasound detectors for bat studies in Europe:
experiences from field identification, surveys, and monitoring. Acta Chiropterologica 1 (2): 137-150.
August T.A., Nunn M.A., Fensome A.G., Linton D.M. & Mathews F. (2014). Sympatric
woodland Myotis bats form tight-knit social groups with exclusive roost home ranges. PLoS ONE 9(10): e112225. doi:10.1371/journal.pone.0112225.
Barataud, M. (2012). Ecologie acoustique des chiroptères d’Europe, identification des
espèces, étude de leurs habitats et comportements de chasse. Biotope, Mèze; Muséum national d’Histoire naturelle, Paris (collection Inventaires et biodiversité). 344p.
Bat Conservation Trust (2007). Bat Surveys – Good Practice Guidelines. Bat Conservation Trust, London. 82 p.
Bates, D., Maechler, M., Bolker, B. & Walker, S. (2013) lme4: linear mixed-effects models using S4 classes. R Package version 1.0.5.
Boers, K., Verkem S., & Willems, W. (2010). Verslag fortentelweekend 5-6-7 februari 2010. Natuurpunt Werkgroep Vleermuizen. 12p.
Boers, K., Verkem S., & Willems, W. (2013). Verslag fortentelweekend 1-2-3 februari
2013. Natuurpunt Werkgroep Vleermuizen. 9p.
Boonman, M. (2000). Roost selection by noctules (Nyctalus noctula) and Daubenton's bats (Myotis daubentonii). Journal of Zoology 251: 385-389.
Boye, P. & Dietz, M. (2005). Development of good practice guidelines for woodland
management for bats English Nature Research Reports nr. 661. English Nature, Peterborough. 89p.
Brigham, R. M., et al., (2004). Bat Echolocation Research: tools, techniques and analysis.
Bat Conservation International. Austin, Texas. 167p.
Buckley, D.J., et al. (2012) The spatial ecology of the whiskered bat (Myotis mystacinus)
at the western extreme of its range provides evidence of regional adaptation. Mammal. Biol. http://dx.doi.org/10.1016/j.mambio.2012.06.007
Cel’uch, M. & Kaňuch, P. (2004). Foraging and flight activity of bats in beech-oak forests (Western Carpathians). Folia Oecologica 31(1):8-16.
De Ridder, J. & Sanders, D. (2013). Vleermuizenonderzoek in de omgeving van het
kasteeldomein Klaverblad en de vallei van de Kleine Struisbeek te Wilrijk. Rapport Natuurpunt Zuidrand Antwerpen werkgroep Studie 2013/1.
De Ridder, J., Sanders D., Van Deynze, W. & Waeterschoot, H. (2014). Vleermuizen op
Fort 7: synthese van de beschikbare gegevens en aanzet tot een visie en zoneringsplan. Rapport Natuurpunt Zuidrand Antwerpen werkgroep Studie 2014/1.
Dienst Regelingen. Ministerie van Economische Zaken, Landbouw & Innovatie (2011).
Soortenstandaard Gewone dwergvleermuis Pipistrellus pipistrellus. 77p.
Dienst Regelingen. Ministerie van Economische Zaken, Landbouw & Innovatie (2011). Soortenstandaard Ruige dwergvleermuis Pipistrellus nathusii. 69p.
Dienst Regelingen. Ministerie van Economische Zaken, Landbouw & Innovatie (2013). Soortenstandaard watervleermuis Myotis daubentonii. 72p.
Dienst Regelingen. Ministerie van Economische Zaken, Landbouw & Innovatie (2011). Soortenstandaard gewone grootoorvleermuis Plecotus auritus. 65p.

Pagina | 35
Dietz, C., Helversen O. von, & D. Nill. (2011). Vleermuizen. Alle soorten van Europa en
Noord-West Afrika. Tirion Natuur. 400 p.
Dijkstra, V. & Korsten, E. (2005). Handleiding wintertellingen van vleermuizen. Voor het monitoren van vleermuizen in de winter. Zoogdierenvereniging VZZ. Arnhem. 20p.
Entwistle, A.C., Racey, P.A. & Speakman J.R. (1998). Habitat Exploitation by a gleaning bat, Plecotus auritus. Philosophical Transactions: Biological Sciences. 1342: 921 - 931
Entwistle, A.C., Racey, P.A. & Speakman J.R. (2000). Social and population of a gleaning bat, Plecotus auritus. J. Zool. Lond. 252:11 - 17
Encarnação, J. A., Dietz, M. & Kalko E.K.V. (2006). Small scale distribution patterns of
female and male Daubenton's bats (Myotis daubentonii). Acta Chiropterologica 8(2):403-415.
Encarnação, J.A., Kierdorf, U. & Wolters, V. (2007). Do mating roosts of Daubenton’s bats (Myotis daubentonii) exist at summer sites? Myotis Vol. 43: 31 – 39.
Encarnaçaõ, J.A. (2012). Spatiotemporal pattern of local sexual segregation in a tree-dwelling temperate bat Myotis daubentonii. J Ethol (2012) 30:271–278.
Flaquer, C., Torre, I. & Arrizabalaga, A. (2007). Comparison of sampling methods for
inventory of bat communities. Journal of Mammalogy. 88(2):526-533.
Galens, D. & Dekeukeleire, D. (2011). Onderzoek naar de Franjestaart in de Makegemse
Bossen. Rapport Natuurpunt Studie 2011/15, Mechelen.30p.
Jonker, M. N., De Boer, W.F., Kurvers, R. & Dekker, J. (2010) Foraging and public
information use in common pipistrelle bats (Pipistrellus pipistrellus): a field experiment. Acta Chiropterologica, 12(1): 197–203.
Kapteyn, K., (1995). Vleermuizen in het landschap. Over hun ecologie, gedrag en
verspreiding. Schuyt & Co, Haarlem.
Kervyn, T. (1998). Plan d'action pour conservation de la sérotine commune, du grand
murin et du grand rhinolophe en Région wallonne. Rapport non publié, Université de Liège, 75 pp.
Kusch, J., Weber, C., Idelberger, S. & Koob T., (2004). Foraging habitat preferences of
bats in relation to food supply and spatial vegetation structures in a western European
low mountain range forest. Folia Zoologica 53 (2): 113-128
Lange, R., Twisk, P., van Winden, A., & van Diepenbeek, A. (1994). Zoogdieren van West-Europa (deel vleermuizen). Uitgave KNNV-uitgeverij.
Lee, Y. & McCracken, G. F. (2004). Flight Activity and Food Habits of Three Species of
Myotis Bats (Chiroptera: Vespertilionidae) in Sympatry. Zoological Studies 43(3): 589-597.
Lesiński, G., Olszewski, A. & Popczyk, B. (2011). Forest roads used by commuting and
foraging bats in edge and interior zones. Polish Journal of Ecology 59(3):611-616.
Limpens, H., Mostert, K. & Bongers, W. (red.) (1997). Atlas van de Nederlandse
vleermuizen. Onderzoek naar verspreiding en ecologie. Stichting Uitgeverij KNNV, Utrecht.
Lučan, R.K., Hanák, V. & Horáček, I. (2009). Long-term re-use of tree roosts by European forest bats. Forest Ecology and Management 258: 1301–1306.
Lučan, R.K. & Radil, J. (2010). Variability of foraging and roosting activities in adult
females of Daubenton’s bat (Myotis daubentonii) in different seasons. Biologia 65/6: 1072—1080.
Maes, D., Baert, K., Boers, K., Casaer, J., Crevecoeur, L., Criel, D., Dekeukeleire, D.,
Gouwy, J., Gyselings, R., Haelters, J., Herman, D., Herremans, M., Lefebvre, J., Lefevre,
A., Onkelinx, T., Scheppers, T., Stuyck, J., Thomaes, A., Van Den Berge, K.,
Vandendriessche, B., Verbeylen, G. & Vercayie, D. (2014). De IUCN Rode Lijst van de

Pagina | 36
zoogdieren in Vlaanderen. Rapport Instituut voor Natuur- en Bosonderzoek
INBO.R.2014.1828211 Instituut voor Natuur – en Bosonderzoek, Brussel
Meschede, A. & Heller, K. (2000). Ökologie und schutz von Fledermäusen in Wäldern.
Bundesamt für Naturschutz: Schriftenreihe für landschaftsplege und Naturschuz Heft 77. 374 p.
Middleton, N., Froud, A., & French, K. (2014). Social calls of the bats of Britain and Ireland. Pelagic Publishing, UK. 176p.
Mitchell-Jones, A.J. et al. (2004). Bat Workers’ Manual. Joint Nature Conservation
Committee. Pelagic Publishing, UK. 178p.
Motte, G. & Kervyn, Th. Kervyn (1997). Caractérisation des terrains de chasse d’une
colonie de sérotines communes, Eptesicus serotinus (Schreber, 1774) (Mammalia :
Chiroptera) en Lorraine belge. Implication pour une meilleure conservation des
populations. VIIéme Université d’été de la Nature. (R.N.O.B. - F.N.R.S - Région
Wallonne).
Nyssen, P. (2012). Natura 2000 inventarisaties in Wallonië: overzicht van 5 jaar vleermuizenstudie. Presentatie Vleermuizenstudiedag KBIN – 24 maart 2012
Paelinckx, D., et al. (red.) (2009). Gewestelijke doelstellingen voor de habitats en
soorten van de Europese Habitat- en Vogelrichtlijn voor Vlaanderen. Mededelingen van het Instituut voor Natuur- en Bosonderzoek INBO.M.2009.6, Brussel, 669 p.
Rieger, I. & Nagel, P. (2007). ‘Vertical stratification of bat activity in a deciduous forest’
in ‘The Canopy of a Temperate Floodplain Forest’. p141-149.
Robinson, M. F. & Stebbings, R. E. (1997) Home range and habitat use by the serotine bat, Eptesicus serotinus, in England. Journal of Zoology 243: 117 – 136.
Russ, J. (2012). British bat calls: a guide to species identification. Pelagic Publishing, UK. 192p.
Rydell, J., Entwistle, A. & Racey, P.A. (1996). Timing of Foraging Flights of Three Species of Bats in Relation to Insect Activity and Predation Risk. Oikos Vol. 76, No. 2 pp. 243-252
Rydell, J., Miller, L.A., & Jensen, M.E. (1999). Echolocation constraints of Daubenton’s
Bat foraging over water. Functional Ecology 13, 247–255
Schober, W., Grimmberger, E. & Lina, P. (vert. en bewerk.). (2001). Gids van de
vleermuizen van Europa, Azoren en Canarische Eilanden: met specifieke informatie over de vleermuizen in Nederland en België. Tirion, 263 p.
Simon, M., Hüttenbügel, S. & Smit-Viergutz, Janna (2004). Ecology and conservation of
Bats in villages and towns. Bundesamt für Naturschutz: Schriftenreihe für
Landschaftsplege und Naturschutz Heft 77. Bonn – Bad Godesberg. 263p.
Stijnen, T., Lambrechts, J., Andriessen, W., Hendrickx, P., & Verheijen, W. (2011).
Beheerplan Parkbegraafplaats Schoonselhof. ARCADIS Belgium nv. Antwerpen. 166p.
Strypstein C. (2004). Chiropterologisch onderzoek naar de jachtbiotopen en het aanbod
van dagverblijfplaatsen m.b.t. de bossamenstelling in het domeinbos Grotenhout
(Antwerpen, België). Thesis Vrije Universiteit Brussel, Faculteit Wetenschappen vakgroep
Biologie, 112p.
Twisk, P., van Diepenbeek, A. & Bekker, J. P. (2010). Veldgids Europese Zoogdieren
(deel vleermuizen en redactie). Uitgave KNNV-uitgeverij.
Van De Sijpe, M. (1999). Batdetector opnamen van de voornaamste vleermuizensoorten in Vlaanderen. Natuurreservaten vzw. 39p.
Vaughan N., Jones G. & Harris S. (1997). Habitat use by bats (Chiroptera) assessed by means of a broad-band acoustic method. Journal of Applied Ecology 34, 716-730

Pagina | 37
Verboom, B. (1998). The use of edge habitats by commuting and foraging bats. IBN
Scientific Contributions 10. DLO Institute for Forestry and Nature Research (IBN-DLO), Wageningen.
Verboom, B., & Huitema H. (1997). The importance of linear landscape elements for the
pipistrelle Pipistrellus pipistrellus and the serotine Eptesicus serotinus. Landscape Ecology 12 (2): 117-125.
Verboom, B., & Huitema H. (2010). The Influence of treeline structure and wind
protection on commuting and foraging pipistrelles (Pipistrellus pipistrellus). Lutra 53 (2): 63 - 80
Verkem, S. & Verhagen, R. (1998). Bescherming vleermuizen. Rapport AMINAL/afdeling Natuur/1995/nr 8.
Verkem, S., De Maeseneer, J. Vandendriessche, B., Verbeylen, G. & Yskout, S. (2003).
Zoogdieren in Vlaanderen. Ecologie en verspreiding van 1987 tot 2002. Natuurpunt
Studie & JNM-zoogdierenwerkgroep, Mechelen & Gent, België.
Vleermuisvakberaad Netwerk Groene Bureaus, Zoogdiervereniging en Gegevensautoriteit
Natuur. (2013). Vleermuisprotocol 2013, 27 maart 2013. www.gegevensautoriteitnatuur.nl en www.netwerkgroenebureaus.nl
Warren, R.D., Waters, D.A., Altringham, J.D. & Bullock, D.J. (2000). The distribution of
Daubenton’s bats (Myotis daubentonii) and pipistrelle bats (Pipistrellus pipistrellus)
(Vespertilionidae) in relation to small-scale variation in riverine habitat. Biological Conservation 92: 85-91.
Wickramasinghe, L. P., Harris, S., Jones G. & Vaughan, N. (2003). Bat activity and
species richness on organic and conventional farms: impact of agricultural intensification. Journal of Applied Ecology 40: 984–993
Willems, W., Boers, K., Vandendriessche, B., Vanreusel, W. & Lefevre, A. (2009).
Monitoring handleiding Natuurpunt Module F10 – Wintertelling vleermuizen. 6 p.
Wood, S. (2014) mgcv: Mixed GAM Computation Vehicle with GCV/AIC/REML smoothness estimation. R Package version 1.7.28.
Zoogdiervereniging & Probos. (2012). Laanbeheer en vleermuizen; met oog voor
veiligheid en cultuurhistorie; met bijdragen van: E.A. Jansen, M.H.A. van Benthem, C. de
Groot, P. Twisk & H.J.G.A. Limpens, Rapport 2012.10. Zoogdiervereniging & Probos,
Nijmegen/Wageningen. 69p.
Zukal J., & Řehák, Z. (2006) Flight activity and habitat preference of bats in a karstic area, as revealed by bat detectors. Folia Zoologica 55(3): 273–281
Zuur, A.F., I. E. N., Walker, N.J., Saveliev, A.A. & Smith, G.M. (2009) Mixed effect models and extensions in ecology with R. Springer, New York, U.S.A.

Pagina | 38
8. Bijlagen
8.1 Overzicht van tel-data, tijdstippen en weersomstandigheden
Tabel 4. Overzicht van de waarnemingsdata, locatie, start- en einduur, start-temperatuur en
weersomstandigheden.
Periode Datum Start Einde Start
temp. Weer
Kraam
24/04/2014 21u05 0u15 17°C Bewolkt; lichte wind bij
aanvang
15/05/2014 21u37 0u22 14,5°C Licht bewolkt
12/06/2014 22u00 01u00 21,5°C Open hemel
Lactatie 16/07/2014 22u10 00u40 24,5°C Open hemel
Post-lactatie 27/08/2014 20u43 00u00 20,5°C Bewolkt
17/09/2014 20u05 23u22 22°C Licht bewolkt, warm
10/10/2014 19u53 22u13 16°C Licht bewolkt, frisse wind
Tabel 5. Overzicht van de waarnemingsdata, locatie, start- en einduur, start-temperatuur en weersomstandigheden.
Periode Datum Start Einde Start
temp. Weer
Kraam
17/04/2014 20:58 23:13 17°C Bewolkt, regenachtig,
winderig
25/04/2014 20:58 00:35 18°C Bewolkt, koude wind
22/05/2014 21:26 00:56 20°C Licht bewolkt
Lactatie 27/06/2014 21:51 01:51 20°C
Afwisselend bewolkt,
lichte wind
07/07/2014 21:50 01:0 20,5°C Dicht bewolkt, frisse wind
Post-lactatie 03/09/2014 20:38 23:30 21,5°C Licht bewolkt
25/09/2014 19:51 22:30 15°C Open hemel, winderig

Pagina | 39
8.2 Figuren
De gegevens worden weergeven op de middenschalige kleurenorthofoto's, van de
provincie Antwerpen, opname 2010 (bron: AGIV).
Figuur 21. Dreven of bomenrijen op de parkbegraafplaats Schoonselhof met nummering
(geïnventariseerde dreven: zwart; niet geïnventariseerde: oranje (bron luchtfoto AGIV)

Pagina | 40
Figuur 2. Bospercelen parkbegraafplaats Schoonselhof (bron beheerplan ARCADIS)

Pagina | 41
Figuur 3. Grafperken en groenzones op begraafplaats Schoonselhof (bron: website stad Antwerpen)

Pagina | 42
Figuur 4. Overzicht van locaties met specifieke aandachtspunten voor vleermuizen (bron luchtfoto:
AGIV)

Pagina | 43
Figuur 5. Overzichtskaart habitatgebruik grootoorvleermuis (bron luchtfoto: AGIV). Vliegroute:
zwarte streeplijn, jachtgebied: gele vlakken, overwinteringsplaats: bruine vlakken, waarnemingen
buiten kerngebieden: gele vierkantjes, locaties met sociale geluiden: witte bol met stip

Pagina | 44
Tabel 6. Overzicht van de aanbevelingen m.b.t. beheer met prioritering (waarbij 1 hoogste prioriteit) (P = preventieve maatregel, V = versterkende maatregel, A = aanbevolen maatregel, R = (strikte) randvoorwaarde)
nr. actie locatie type prioriteit
Algemeen
1 Kappingen van oude loofbomen uitvoeren in de voor vleermuizen minst gevoelige periode. Dit is het vroege voorjaar (april) of najaar (half september-eind oktober), maar alleen als er geen paarverblijven aanwezig zijn.
Schoonselhof P 2
Dreven en bossen
2 Aanvullend onderzoek op aanwezigheid van verblijfplaatsen van vleermuizen voorafgaand aan kapwerken
L32 P 1
3 Aanvullend onderzoek op aanwezigheid van verblijfplaatsen van vleermuizen voorafgaand aan snoei - en kapwerken
L14, L17B, L19B, L29
P 2
4 Aanvullend onderzoek op aanwezigheid van verblijfplaatsen van vleermuizen voorafgaand aan snoei- en kapwerken
L21 P 3
5 Aanvullend onderzoek op aanwezigheid van verblijfplaatsen van baardvleermuis en franjestaart voorafgaand aan kapwerken
L39a R 1
6 Aanvullend onderzoek op aanwezigheid van verblijfplaatsen van vleermuizen voorafgaand aan kapwerken bosperceel
1g P 1
7 Aanvullend onderzoek op aanwezigheid van verblijfplaatsen van vleermuizen voorafgaand aan kapwerken
2c P 1
8 De verjonging van het bosperceel 2c dient gefaseerd in de tijd uitgevoerd te worden, met een behoud van minimum 10 potentiële vleermuisbomen per ha in het bos of aanpalende dreven.
2c R 1
9 Ophangen vleermuiskasten in oude bospercelen en dreven waar het minimum van 10 potentiële vleermuisbomen niet gehaald wordt
2c, L17, L19 en L29
A 2
Vijvers en grachten
10 Zwermlocatie vrijwaren van lichtverstoring en hindernissen (o.a. omgevallen bomen)
perceel D; figuur 12
P 1
11 Behoud oppervlakte open water als jachtgebied voor watervleermuis Jachtgebied
figuur 9. A 2
12 Periodiek kruid- en slibruimen van brede grachten als jachtgebied voor watervleermuis
Schoonselhof V 2
13 Kruid- en slibruimingen uitvoeren buiten actief jachtseizoen van vleermuizen Schoonselhof P 2
Beheermaatregelen voor behoud en versterken habitat van laatvlieger
14 Laatvlieger als doelsoort opnemen in actualisatie beheerplan Schoonselhof A 3
15 Vermijden van gebruik van pesticiden bij beheer graslanden Schoonselhof V 1
16 Ontwikkelen van bosranden en randhabitats Schoonselhof V 3
17 Hagen met exotische soorten vervangen door inheems soorten Schoonselhof V 2
Beheermaatregelen voor behoud en versterken habitat van grootoorvleermuis
18 Grootoorvleermuis als doelsoort opnemen in actualisatie beheerplan Schoonselhof A 3
19 Vrijwaren van de dreven (= behoud van de lijnelementen) als vliegroute van grootoorvleermuis (en andere soorten)
L14, L17b, L24 en L29
R 1
20 De restauratie en de functie-invulling van de Neerhoeve afstemmen met de vereisten voor een verblijfplaats voor gewone grootoorvleermuis (m.b.t. temperatuur, in- en uitvliegopeningen, toegankelijke ruimte).
Neerhoeve R 1
21 Het gebruik van niet-giftige of aangepaste houtverduurzamingsmiddelen bij renovatiewerken is een belangrijk aandachtspunt voor het behouden van een vleermuizenpopulatie.
Neerhoeve P 1
22 In de dreven waar (hoge) jachtactiviteit of sociale geluiden van gewone grootoorvleermuis werden vastgesteld kunnen kappingen en kroonverzorging pas gebeuren na een voorafgaande controle door een vleermuizenexpert
L14, L17B, L19B en L29
P 1
23 L24, L25 P 2

Pagina | 45
Figuur 6. Overzicht van locaties met waarnemingen van gewone dwergvleermuis op het Schoonselhof in de periode april tot oktober 2014 (bron
luchtfoto: AGIV).

Pagina | 46
Figuur 7. Overzicht van locaties met waarnemingen van ruige dwergvleermuis op het Schoonselhof in de periode april tot oktober 2014 (bron
luchtfoto: AGIV).
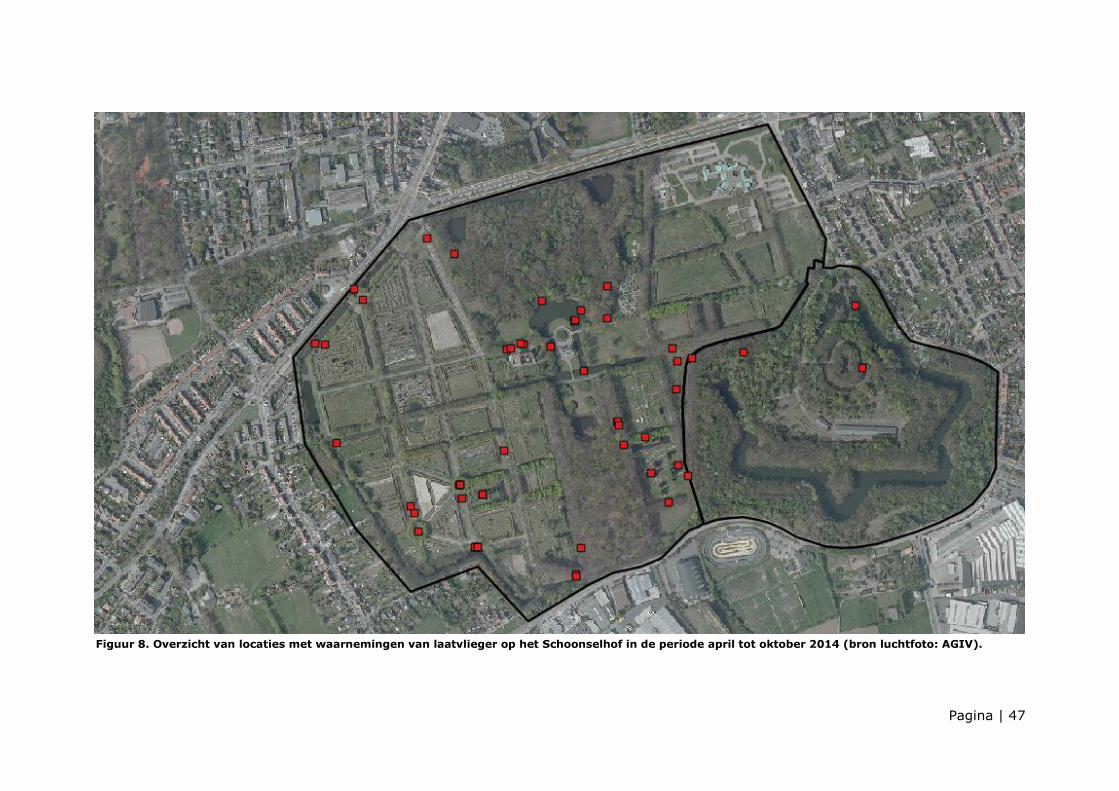
Pagina | 47
Figuur 8. Overzicht van locaties met waarnemingen van laatvlieger op het Schoonselhof in de periode april tot oktober 2014 (bron luchtfoto: AGIV).

Pagina | 48
Figuur 9. Overzicht van locaties met waarnemingen van rosse vleermuis op het Schoonselhof in de periode april tot oktober 2014 (bron luchtfoto:
AGIV).

Pagina | 49
Figuur 10. Overzicht van locaties met waarnemingen van grootoorvleermuis op het Schoonselhof in de periode april tot oktober 2014 (bron luchtfoto:
AGIV).

Pagina | 50
Figuur 11. Overzicht van locaties met waarnemingen van Myotis-soorten op het Schoonselhof in de periode april tot oktober 2014 (bron luchtfoto:
AGIV). Paars: watervleermuis, oranje: franjestaart, bruin: baardvleermuis, holle vierkant: Myotis spp.

Pagina | 51
8.3 Analyse geluidsopnamen
Figuur 22. Sonogram Plecotus spp. (opname 2014_DS_1158, 15 mei 2014)

Pagina | 52
Figuur 23. Sonogram franjestaart (opname 2014_JDR_0146, 27 juni 2014)

Pagina | 53
Figuur 24. Sonogram watervleermuis en gewone dwergvleermuis (opname 2014_JDR_0075, 22 mei 2014)

Pagina | 54
Figuur 25. Sonogram baardvleermuis (opname 2014_DS_1417, 3 september 2014)

Pagina | 55
Figuur 26. Sonogram ruige dwergvleermuis (opname 2014_JDR_0291, 3 september 2014)




















