
Vibrio cholerae cytolysin is essential for high enterotoxicity and apoptosis induction produced by a...
11
PATHOGENESIS MICROBIAL Microbial Pathogenesis 44 (2008) 118–128 Vibrio cholerae cytolysin is essential for high enterotoxicity and apoptosis induction produced by a cholera toxin gene-negative V. cholerae non-O1, non-O139 strain Hector Alex Saka, Carla Bidinost, Claudia Sola, Pablo Carranza, Cesar Collino, Susana Ortiz, Jose Ricardo Echenique, Jose´ Luis Bocco Departamento de Bioquı´mica Clı´nica, Facultad de Ciencias Quı´micas, Universidad Nacional de Co´rdoba, Centro de Investigaciones en Bioquı´mica e Inmunologı´a ClBICI-CONICET, Co´rdoba, Argentina Received 24 April 2007; accepted 10 August 2007 Available online 31 August 2007 Abstract Cholera toxin (CT) gene-negative Vibrio cholerae non-O1, non-O139 strains may cause severe diarrhea though their pathogenic mechanism remains unclear. V. cholerae cytolysin (VCC) is a pore-forming exotoxin encoded in the hlyA gene of V. cholerae whose contribution to the pathogenesis is not fully understood. In this work, the virulence properties of a CT gene-negative V. cholerae non-O1, non-O139 strain causing a cholera-like syndrome were analyzed. Inoculation of rabbit ileal loops with the wild type strain induced extensive fluid accumulation, accompanied by severe histopathological damage characterized by villus shortening, lymphangiectasia and focal areas of necrosis. These pathogenic effects were abrogated by mutation of the hlyA gene thus pointing out the main role of VCC in the virulence of the strain. Interestingly, this toxin was capable of triggering apoptosis in human intestinal cell lines due to its anion channel activity. Moreover, the wild type strain also induced increased apoptosis of the intestinal epithelium cells which was not observed upon inoculation of the VCC null mutant strain, indicating that VCC may trigger apoptotic cell death during infection in vivo. Altogether, these results support a main role of VCC in the pathogenesis of the CT gene-negative V. cholerae non-O1, non-O139 strain and identify apoptosis as a previously unrecognized cell death pathway triggered by VCC. r 2007 Elsevier Ltd. All rights reserved. Keywords: Non-O1; Non-O139 Vibrio cholerae; Vibrio cholerae cytolysin; Pore-forming toxin; Apoptosis; Rabbit ileal loop; Enterotoxicity 1. Introduction The Gram-negative bacterium Vibrio cholerae is a natural inhabitant of the aquatic ecosystem capable of causing severe infections in humans [1,2]. V. cholerae is the causative agent of cholera disease, which occurs mainly in developing countries and is transmitted through contami- nated food or drinking-water or by the faecal–oral route through person-to-person contact [3–5]. At least seven cholera pandemics were recorded since 1817 and at present cholera is still a major burden on human health that is becoming increasingly important [5,6]. More than 130,000 cholera cases, which are estimated to represent around 5–10% of actual cases worldwide, were officially reported to the World Health Organization by 52 countries during 2005 [7]. Based on O-antigen structure, V. cholerae strains are grouped into more than 200 serogroups [8,9] from which only O1 and O139 are associated to cholera epidemics and to the production of critical virulence factors as cholera toxin (CT) and toxin co-regulated pilus (TCP); CT is responsible for full-blown cholera whereas TCP plays an essential role in bacterial adherence to the intestinal epithelium [5,10]. The remaining serogroups are referred to as non-O1, non-O139, which mostly lack CT and TCP and are more frequently isolated from aquatic environ- mental sources than from clinical samples. Though it is well documented that V. cholerae non-O1, non-O139 strains are ARTICLE IN PRESS www.elsevier.com/locate/micpath 0882-4010/$ - see front matter r 2007 Elsevier Ltd. All rights reserved. doi:10.1016/j.micpath.2007.08.013 Corresponding author. Tel.: +54 351 433 4164/87; fax: +54 351 433 3048. E-mail address: [email protected] (J.L. Bocco).
Transcript of Vibrio cholerae cytolysin is essential for high enterotoxicity and apoptosis induction produced by a...

ARTICLE IN PRESS
PATHOGENESISMICROBIAL
0882-4010/$ - se
doi:10.1016/j.m
�Correspondfax: +54351 43
E-mail addr
Microbial Pathogenesis 44 (2008) 118–128
www.elsevier.com/locate/micpath
Vibrio cholerae cytolysin is essential for high enterotoxicity andapoptosis induction produced by a cholera toxin gene-negative
V. cholerae non-O1, non-O139 strain
Hector Alex Saka, Carla Bidinost, Claudia Sola, Pablo Carranza, Cesar Collino,Susana Ortiz, Jose Ricardo Echenique, Jose Luis Bocco�
Departamento de Bioquımica Clınica, Facultad de Ciencias Quımicas, Universidad Nacional de Cordoba, Centro de Investigaciones en Bioquımica e
Inmunologıa ClBICI-CONICET, Cordoba, Argentina
Received 24 April 2007; accepted 10 August 2007
Available online 31 August 2007
Abstract
Cholera toxin (CT) gene-negative Vibrio cholerae non-O1, non-O139 strains may cause severe diarrhea though their pathogenic
mechanism remains unclear. V. cholerae cytolysin (VCC) is a pore-forming exotoxin encoded in the hlyA gene of V. cholerae whose
contribution to the pathogenesis is not fully understood. In this work, the virulence properties of a CT gene-negative V. cholerae non-O1,
non-O139 strain causing a cholera-like syndrome were analyzed. Inoculation of rabbit ileal loops with the wild type strain induced
extensive fluid accumulation, accompanied by severe histopathological damage characterized by villus shortening, lymphangiectasia and
focal areas of necrosis. These pathogenic effects were abrogated by mutation of the hlyA gene thus pointing out the main role of VCC in
the virulence of the strain. Interestingly, this toxin was capable of triggering apoptosis in human intestinal cell lines due to its anion
channel activity. Moreover, the wild type strain also induced increased apoptosis of the intestinal epithelium cells which was not observed
upon inoculation of the VCC null mutant strain, indicating that VCC may trigger apoptotic cell death during infection in vivo.
Altogether, these results support a main role of VCC in the pathogenesis of the CT gene-negative V. cholerae non-O1, non-O139 strain
and identify apoptosis as a previously unrecognized cell death pathway triggered by VCC.
r 2007 Elsevier Ltd. All rights reserved.
Keywords: Non-O1; Non-O139 Vibrio cholerae; Vibrio cholerae cytolysin; Pore-forming toxin; Apoptosis; Rabbit ileal loop; Enterotoxicity
1. Introduction
The Gram-negative bacterium Vibrio cholerae is anatural inhabitant of the aquatic ecosystem capable ofcausing severe infections in humans [1,2]. V. cholerae is thecausative agent of cholera disease, which occurs mainly indeveloping countries and is transmitted through contami-nated food or drinking-water or by the faecal–oral routethrough person-to-person contact [3–5].
At least seven cholera pandemics were recorded since1817 and at present cholera is still a major burden onhuman health that is becoming increasingly important
e front matter r 2007 Elsevier Ltd. All rights reserved.
icpath.2007.08.013
ing author. Tel.: +54351 433 4164/87;
3 3048.
ess: [email protected] (J.L. Bocco).
[5,6]. More than 130,000 cholera cases, which are estimatedto represent around 5–10% of actual cases worldwide, wereofficially reported to the World Health Organization by 52countries during 2005 [7].Based on O-antigen structure, V. cholerae strains are
grouped into more than 200 serogroups [8,9] from whichonly O1 and O139 are associated to cholera epidemics andto the production of critical virulence factors as choleratoxin (CT) and toxin co-regulated pilus (TCP); CT isresponsible for full-blown cholera whereas TCP plays anessential role in bacterial adherence to the intestinalepithelium [5,10]. The remaining serogroups are referredto as non-O1, non-O139, which mostly lack CT and TCPand are more frequently isolated from aquatic environ-mental sources than from clinical samples. Though it is welldocumented that V. cholerae non-O1, non-O139 strains are

ARTICLE IN PRESSH.A. Saka et al. / Microbial Pathogenesis 44 (2008) 118–128 119
capable of producing human disease, such as sporadicoutbreaks of watery diarrhea, inflammatory enterocolitis,soft tissue infections and isolated cases of fatal sepsis, theirpathogenic mechanisms remain poorly elucidated [11–13].
Most V. cholerae isolates belonging to O1 biotype El Torand to non-O1, non-O139 serogroups produce a hemolyticexotoxin, named El Tor Haemolysin or V. cholerae
cytolysin (VCC) [14–16], which is encoded in the structuralhlyA gene located at the V. cholerae chromosome II [17].VCC is synthesized and secreted as pro-cytolysin mono-mers of about 80 kDa which are proteolytically activatedby different proteases through removal of the 15 kDaprotoxin domain at the N-terminus [18–20]. Mature VCCforms heptameric oligomers in the presence of cholesteroland ceramide-rich membranes, generating membrane poreswhose internal diameter is 1–2 nm [21–27]. It has beenreported that VCC is capable of causing several cytotoxiceffects such as cell lysis and extensive vacuolization in vitro,depending upon the cell line studied and the toxinconcentration [28–33]. Recently, the vacuolization phe-nomenon associated to VCC action has been demonstratedto be an autophagic response of the cells against this toxinand was related to a protective cell response upon VCCintoxication [34]. Though it has been demonstrated thatpurified fractions of VCC produce fluid accumulation inligated rabbit ileal loops [35], several contradictory dataexist regarding the real role of VCC in the pathogenesiscaused by CT-negative TCP-negative V. cholerae strainsduring infection [29,30,35–39].
In this work we studied a CT-negative, TCP-negative(CT� TCP�) non-O1, non-O139 V. cholerae strain causinga cholera-like syndrome [40,41]. Wild-type strain but notits isogenic VCC-null mutant induced extensive fluidaccumulation in rabbit ileal loops, providing evidencesupporting the involvement of VCC in the pathogenesis.Moreover, a new role of VCC as an apoptogenic moleculewas demonstrated. Severe histological damage and in-creased apoptosis of the intestinal epithelium related toVCC action may account for the diarrheagenic potential ofthe CT� TCP� V. cholerae strain studied in this work.
2. Results
2.1. VCC is associated to the cytotoxic and enterotoxic
properties of a CT� TCP� V. cholerae non-O1, non-O139
strain causing a cholera-like syndrome
Two CT� TCP� V. cholerae non-O1, non-O139 isolatespresenting indistinguishable DNA PFGE-type were ob-tained from the feces of two epidemiologically unrelatedpatients suffering a cholera-like syndrome [40,41]. In orderto investigate which could be the virulence factor(s)involved in the pathogenesis of these isolates, severalmutants were obtained and their cytotoxic properties werestudied. As shown in Fig. 1A, sterile culture supernatantsobtained from the wild type strain (Vc-wt) was able toreduce the viability of different cell types, including the
human colon-derived Caco-2 cell line, in a concentrationdependent manner. On the contrary, the isogenic hlyA nullmutant (Vc-hlyA-), defective in VCC production (supple-mentary data Fig. 8), was unable to cause any detrimentaleffect on cell survival (Fig. 1A). The identification of VCCas the specific bacterial product involved in the cytotoxiceffect observed was confirmed using purified VCC andneutralizing the cytotoxicity of the Vc-wt supernatant witha specific anti-VCC antibody (Fig. 1A). This resultprompted us to evaluate if the enterotoxigenicity of thestrain could be attributed to VCC. To this aim, we used therabbit ileal loop assay and compared the enterotoxic effectproduced by both, Vc-wt and Vc-hlyA-. As shown inFig. 1B, Vc-wt was clearly enterotoxic, inducing significantfluid accumulation into the intestinal loops. The accumu-lated fluid was invariably mucous and haemorrhagic (datanot shown). In contrast, no significant fluid accumulationwas detected with Vc-hlyA-. These results are consistentwith the involvement of VCC in the diarrheagenic proper-ties of Vc-wt.
2.2. VCC is an exotoxin capable of inducing apoptosis in
vitro
Considering the evidence presented above, we decided tofurther study the cytotoxic effects caused by VCC. It wasalready reported that VCC had the capability to inducecytotoxicity, causing cell lysis at high doses and vacuoliza-tion at low doses [28,30,32,33]. In agreement, we observedthat higher concentrations of Vc-wt supernatant inducedlysis and that vacuolization was observed at lowerconcentrations in the human intestinal-derived Caco-2and C2BBe1 cell lines, whereas the Vc-hlyA- was unableto induce any cell damage (Supplementary data Fig. 9).Although the ability to induce apoptosis has been
already reported for several bacterial pore-forming toxinsand has been proposed to be involved in the pathogenesis[42–47], the capability of VCC to induce programmed celldeath has not been communicated so far. Thus, we carriedout several independent experimental approaches todetermine if VCC is a toxin capable of triggering apoptoticcell death.
2.2.1. DNA-fragmentation triggered by VCC
Apoptosis typically involves internucleosomal DNAfragmentation into fragments with a ladder-like patterncorresponding to 180- to 200-bp multiples [48,49]. There-fore, DNA cleavage was analyzed in Caco-2 and C2BBe1cells upon intoxication with VCC. As shown in Fig. 2A,DNA-fragmentation was clearly induced at 4 and 6 h oftreatment when Caco-2 cells were exposed to 1.0% and0.1%v/v of Vc-wt supernatant. Conversely, no DNA-cleavage was observed in response to Vc-hlyA- supernatantor when the Vc-wt supernatant was previously neutralizedwith an anti-VCC antibody. DNA fragmentation was notdetected with the highest concentration of Vc-wt super-natant (10.0%v/v), probably due to rapid cell lysis

ARTICLE IN PRESSC
ell
surv
ival (%
)
Caco-2Cos-7MEF
0
10
20
30
40
50
60
70
80
90
100
Vc-wt
1%Vc-w
t
10% Vc-hlyA-VCC
Control
*
*
*
*
**
* **
Vc-wt 1
0%
VCC-ab
0
Vc-wt
Vc-hlyA-DH5α
PBS
EP1/151
AF
(m
l/cm
)
*
*
CFU/ml (x 109)
recovered
1.32
±0.49
1.17
±0.45
ND ND 1.52
±0.52
2.5
2.0
1.5
1.0
0.5
Fig. 1. Cytotoxicity and enterotoxicity triggered by VCC. (A) Cell survival upon VCC intoxication. Sterile supernatants from BHI broth cultures of the
strains (%v/v) or purified VCC (80 pM) were added to cells, as indicated. As control, 10%v/v of non-inoculated BHI broth was used. After 4 h of
treatment, cell viability was determined using the CellTiter 96 AQuoeous kit (Promega). Vc-wt, clinical Vibrio cholerae wild type non-O1, non-O139 strain;
Vc-hlyA-, isogenic hlyA null mutant from Vc-wt; VCC, purified V. cholerae cytolysin; VCC-ab, rabbit polyclonal specific anti-VCC antibody; BHI, brain
heart infusion broth. Data represented are means7standard deviations of at least three individual experiments. Asterisks indicate significant differences
from the results obtained with the control (Po0.001, Tukey–Kramer Multiple Comparisons Test). (B) Enterotoxicity of Vc-wt and Vc-hlyA- in the rabbit
ileal loop assay. Fluid accumulation (FA) is the amount of accumulated fluid (in milliliters) per length (in centimeters) of ligated rabbit small intestine.
Bars: Vc-wt and Vc-hlyA- are as indicated in (A); DH5a, Escherichia coli DH5a (negative control strain); PBS, phosphate buffer saline pH 7.2 (negative
control); EP1/151, cholera toxin-producing Vibrio cholerae O1 Biotype El Tor (positive control strain). Bars represent means7standard deviations of at
least three individual experiments. Asterisks indicate significant differences from the results obtained with the negative control (Po0.001, Tukey–Kramer
Multiple Comparisons Test). The number of viable counts for each V. cholerae strain recovered from the ileal loops is shown at the bottom and expressed
as means7standard deviations. ND: not determined.
H.A. Saka et al. / Microbial Pathogenesis 44 (2008) 118–128120
(Fig. 2A and Supplementary data, Fig. 9). DNA-fragmen-tation was also observed after 6 h of treatment withpurified VCC (80 pM) (Fig. 2A). As shown in Fig. 2B,similar results were obtained with the C2BBe1 cell line.Additional cell lines such as COS-7, J-774, HeLa and CHOcells were also tested obtaining essentially the same resultsas for Caco-2 cells (data not shown).
It is well established that VCC oligomerizes in themembrane of the cells and forms moderately anion-selective transmembrane pores [21,24,50,51]. Thus, weexplored if VCC channel activity was also related to theinduction of DNA fragmentation and apoptotic cell death.It has been demonstrated that DIDS is a compound able toblock VCC channel activity [28]. It was previouslydemonstrated that VCC channel activity was essential forVCC cytotoxicity [28]. Accordingly, blocking VCC channelactivity with DIDS (2mM) impaired VCC to causecytotoxicity in several cell lines (Supplementary dataFig. 10). Interestingly, DNA fragmentation was also preventedin cells treated with DIDS, indicating that VCC channelactivity is required to induce apoptotic cell death (Fig. 2C).
2.2.2. VCC increases the sub-diploid DNA content of Caco-
2 cells
Increments in the sub-diploid DNA content is typicallydetected in apoptotic cells that can be identified by their
reduced fluorescence due to DNA degradation and leakagefrom the cells [52,53]. Sub-diploid cells are detected in abroad hypodiploid or sub-G1 region on a histogram. Thus,we used flow cytometry to analyze the DNA content ofCaco-2 cells treated with Vc-wt or Vc-hlyA- supernatantsor with purified VCC as another approach to detect VCC-induced apoptotic cell death. Following treatment ofconfluent Caco-2 cells with Vc-wt or Vc-hlyA- supernatants(1.0%v/v) or with purified VCC (80 pM) during 4–6 h, themean percentage of hypodiploid cells (M1 region) wasassessed (Fig. 3). No significant differences were detectedbetween BHI-treated (negative control) and Vc-hlyA-supernatant-treated cells. In contrast, cells treated withVc-wt supernatant or with purified VCC showed asignificant increase in the proportion of hypodiploid cells,as it was the case for UV-radiated cells (positive control).VCC-induced increments in sub-diploid DNA content werealso detected in J-774 cells and in COS-7 cells (data notshown).
2.2.3. VCC induces caspase-3 activation
The caspase cascade is a relevant biochemical pathwayinvolved in apoptosis. In this context, caspase-3 is a criticaldownstream protease that, once activated, cleaves severalcellular substrates and activates the endonucleases involvedin DNA fragmentation [54,55]. To ascertain whether the
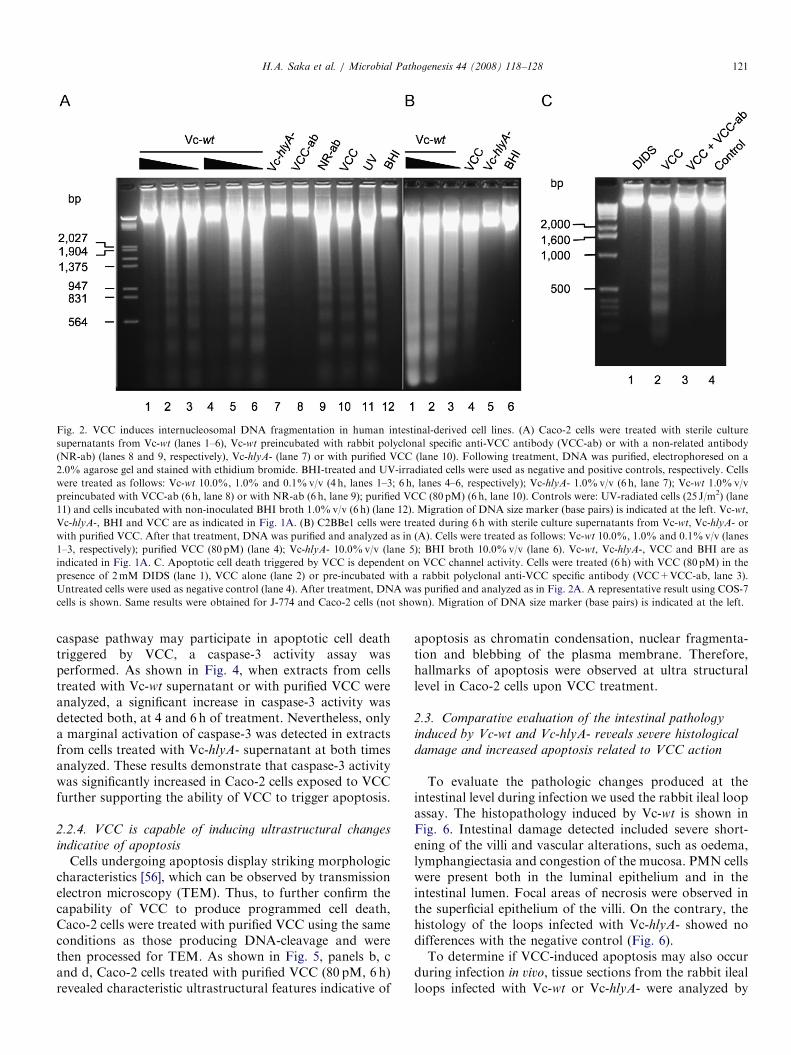
ARTICLE IN PRESS
Fig. 2. VCC induces internucleosomal DNA fragmentation in human intestinal-derived cell lines. (A) Caco-2 cells were treated with sterile culture
supernatants from Vc-wt (lanes 1–6), Vc-wt preincubated with rabbit polyclonal specific anti-VCC antibody (VCC-ab) or with a non-related antibody
(NR-ab) (lanes 8 and 9, respectively), Vc-hlyA- (lane 7) or with purified VCC (lane 10). Following treatment, DNA was purified, electrophoresed on a
2.0% agarose gel and stained with ethidium bromide. BHI-treated and UV-irradiated cells were used as negative and positive controls, respectively. Cells
were treated as follows: Vc-wt 10.0%, 1.0% and 0.1%v/v (4 h, lanes 1–3; 6 h, lanes 4–6, respectively); Vc-hlyA- 1.0%v/v (6 h, lane 7); Vc-wt 1.0%v/v
preincubated with VCC-ab (6 h, lane 8) or with NR-ab (6 h, lane 9); purified VCC (80 pM) (6 h, lane 10). Controls were: UV-radiated cells (25 J/m2) (lane
11) and cells incubated with non-inoculated BHI broth 1.0%v/v (6 h) (lane 12). Migration of DNA size marker (base pairs) is indicated at the left. Vc-wt,
Vc-hlyA-, BHI and VCC are as indicated in Fig. 1A. (B) C2BBe1 cells were treated during 6 h with sterile culture supernatants from Vc-wt, Vc-hlyA- or
with purified VCC. After that treatment, DNA was purified and analyzed as in (A). Cells were treated as follows: Vc-wt 10.0%, 1.0% and 0.1%v/v (lanes
1–3, respectively); purified VCC (80 pM) (lane 4); Vc-hlyA- 10.0%v/v (lane 5); BHI broth 10.0%v/v (lane 6). Vc-wt, Vc-hlyA-, VCC and BHI are as
indicated in Fig. 1A. C. Apoptotic cell death triggered by VCC is dependent on VCC channel activity. Cells were treated (6 h) with VCC (80 pM) in the
presence of 2mM DIDS (lane 1), VCC alone (lane 2) or pre-incubated with a rabbit polyclonal anti-VCC specific antibody (VCC+VCC-ab, lane 3).
Untreated cells were used as negative control (lane 4). After treatment, DNA was purified and analyzed as in Fig. 2A. A representative result using COS-7
cells is shown. Same results were obtained for J-774 and Caco-2 cells (not shown). Migration of DNA size marker (base pairs) is indicated at the left.
H.A. Saka et al. / Microbial Pathogenesis 44 (2008) 118–128 121
caspase pathway may participate in apoptotic cell deathtriggered by VCC, a caspase-3 activity assay wasperformed. As shown in Fig. 4, when extracts from cellstreated with Vc-wt supernatant or with purified VCC wereanalyzed, a significant increase in caspase-3 activity wasdetected both, at 4 and 6 h of treatment. Nevertheless, onlya marginal activation of caspase-3 was detected in extractsfrom cells treated with Vc-hlyA- supernatant at both timesanalyzed. These results demonstrate that caspase-3 activitywas significantly increased in Caco-2 cells exposed to VCCfurther supporting the ability of VCC to trigger apoptosis.
2.2.4. VCC is capable of inducing ultrastructural changes
indicative of apoptosis
Cells undergoing apoptosis display striking morphologiccharacteristics [56], which can be observed by transmissionelectron microscopy (TEM). Thus, to further confirm thecapability of VCC to produce programmed cell death,Caco-2 cells were treated with purified VCC using the sameconditions as those producing DNA-cleavage and werethen processed for TEM. As shown in Fig. 5, panels b, cand d, Caco-2 cells treated with purified VCC (80 pM, 6 h)revealed characteristic ultrastructural features indicative of
apoptosis as chromatin condensation, nuclear fragmenta-tion and blebbing of the plasma membrane. Therefore,hallmarks of apoptosis were observed at ultra structurallevel in Caco-2 cells upon VCC treatment.
2.3. Comparative evaluation of the intestinal pathology
induced by Vc-wt and Vc-hlyA- reveals severe histological
damage and increased apoptosis related to VCC action
To evaluate the pathologic changes produced at theintestinal level during infection we used the rabbit ileal loopassay. The histopathology induced by Vc-wt is shown inFig. 6. Intestinal damage detected included severe short-ening of the villi and vascular alterations, such as oedema,lymphangiectasia and congestion of the mucosa. PMN cellswere present both in the luminal epithelium and in theintestinal lumen. Focal areas of necrosis were observed inthe superficial epithelium of the villi. On the contrary, thehistology of the loops infected with Vc-hlyA- showed nodifferences with the negative control (Fig. 6).To determine if VCC-induced apoptosis may also occur
during infection in vivo, tissue sections from the rabbit ilealloops infected with Vc-wt or Vc-hlyA- were analyzed by

ARTICLE IN PRESS
Fig. 3. Increased sub-diploid DNA content of Caco-2 cells intoxicated
with VCC. Sterile culture supernatants from Vc-wt, Vc-hlyA- (1.0%v/v
each one) or purified VCC (80 pM) were added to cells. Following
treatment, cells were trypsinized, fixed and stained with propidium iodide
and fluorescence distribution was analyzed by flow cytometry. A
statistically significant increase (Po0.01) in hypodiploid nuclei (M1
region) was detected in cells treated with Vc-wt culture supernatant and
with purified VCC. Results are representative of at least three independent
experiments. Approximately 10,000 cells were analyzed per experiment. As
negative control, BHI-treated cells (1.0%v/v, 6 h) and cells exposed only
to cell culture medium (not shown) were used. As positive control, UV-
radiated cells (25 J/m2) were used. Vc-wt, Vc-hlyA-, VCC and BHI are as
indicated in the legend of Fig. 1A.
Caspase-3 activity
**
*
*
0
Fold
activation *
Vc-hlyA- Vc-wt VCC UV
*
4h 6h
7.0
6.0
5.0
4.0
3.0
2.0
1.0
Time
Fig. 4. Caspase-3 activation in VCC-intoxicated Caco-2 cells. Confluent
Caco-2 cells were exposed to sterile culture supernatants (1.0%v/v) from
Vc-wt, Vc-hlyA- or to purified VCC (80 pM). Following treatment,
caspase-3 activity was measured at 4 h and 6 h using a colorimetric kit
according to manufacturer’s instructions. Data are presented as times of
caspase-3 activity with respect to the negative control (fold activation) and
are the mean7standard deviations of at least three individual experi-
ments. Asterisks indicate statistically significant increases in caspase-3
activity (Po0.001, Tukey-Kramer Multiple Comparisons Test). As
negative and positive controls, BHI-treated (1.0%v/v, 6 h) and UV-
radiated (25 J/m2) cells were used, respectively. Vc-wt; Vc-hlyA-; VCC and
BHI are as in the legend of Fig. 1A.
H.A. Saka et al. / Microbial Pathogenesis 44 (2008) 118–128122
TUNEL in situ. As shown in Fig. 7, villi from loopsinfected with Vc-wt presented a greater number of cellslocalized to the epithelium showing TUNEL positive nucleithan loops inoculated with Vc-hlyA- strain (Po0.001).
3. Discussion
The mechanism involved in the pathogenesis producedby CT� TCP� V. cholerae strains is still unclear [11–13].
These strains have been isolated from patients sufferingcholera-like syndrome [41,57]. However, no single viru-lence factor has yet been identified to explain theirenterotoxicity [38]. VCC is a conserved pore-forming toxinof V. cholerae whose role in pathogenesis is not wellelucidated [29,30,35–39]. In this work we provide evidenceindicating that the high enterotoxic potential of a CT�
TCP� V. cholerae strain producing a cholera-like syndromewas dependent on VCC, thus supporting a role for thispore-forming toxin in the pathogenesis of the diarrhea. Wealso found that the wild-type strain culture supernatantdisplayed apoptogenic properties, which can be attributedto VCC. Moreover, results obtained using DIDS indicatedthat the apoptotic cell death triggered by this toxin isrelated to VCC anion channel activity.In an independent study, Zitzer and co-workers did not
detect DNA-fragmentation of the intestine 407 cell lineafter treatment with VCC [33]. Nevertheless, in the presentwork apoptosis triggered by VCC was clearly demon-strated by several independent experimental approaches.It has been reported that VCC shares structural and in
some extent functional characteristics with other pore-forming toxins such as alpha-toxin and leukocidin F (fromStaphylococcus aureus) and with aerolysin (from Aero-
monas hydrophila) [58]. These toxins, including VCC, have

ARTICLE IN PRESS
Fig. 5. Hallmarks of apoptosis of Caco-2 cells upon VCC intoxication. Caco-2 cells were exposed to 80 pM of purified VCC (b–d) and then processed for
transmission electron microscopy. Cells undergoing ultrastructural changes indicative of apoptosis show chromatin condensation (b–d), nuclear
fragmentation and blebbing of the plasma membrane (b). For comparison, normal Caco-2 cells were processed (a). Black bars: 1 mm.
Fig. 6. Intestinal pathology of rabbit ileal loops infected with Vc-wt and Vc-hlyA-. Tissue sections from rabbit ileal loops were stained with H&E and
photomicrographs were taken. Panels a–c: low magnification images of intestinal villi after inoculation with Vc-wt, Vc-hlyA- or PBS are shown. Severe
disruption of villous architecture, shortening of the villi and lymphangiectasia were produced by Vc-wt (panel a) while normal histologic appearance of
intestinal villi were observed both, in the loops infected with Vc-hlyA- (panel b) or in the PBS-inoculated control (panel c). Panel d: a quantitative analysis
of the villi length was performed using the Image-Pro Plus 4.5 software. At least 50 villi were analyzed for each experimental condition and the mean of
villus length7standard deviation was expressed in arbitrary units (a.u.). Asterisk indicates a statistical significant difference compared to the PBS-
inoculated control (Po0,001; Tukey–Kramer Multiple Comparisons Test). Panels e–h: high magnification images of intestinal villi after inoculation with
Vc-wt, Vc-hlyA- or PBS are shown. Focal area of necrosis (arrow, panel e), PMN cells infiltration (inset, panel e), PMN cells in the lumen (asterisk, panel f)
and lymphangiectasia (arrow, panel f) were observed in Vc-wt inoculated loops. Normal intestinal epithelium is observed in Vc-hlyA- (panel g) and PBS-
inoculated loops (panel h), where arrows indicate the presence of goblet cells. Black bars: 20 mm (lower magnification) or 100mm (higher magnification).
H.A. Saka et al. / Microbial Pathogenesis 44 (2008) 118–128 123
in common their so-called cytolytic domain, a region of thetoxin molecule encompassing roughly 250 amino acids.Interestingly, alpha-toxin was shown to evoke the typicalapoptotic alterations such as caspase activation and DNAfragmentation in various cell types [59–63]. More recently,it has been communicated that the staphylococcal leuko-cidin has the capability to induce cell death via necrosis athigh doses and via apoptosis at low doses [64]. In the caseof aerolysin, its ability to produce apoptosis has also beeninformed, at least in T lymphoma cells [47]. Thus, thecapability of VCC to induce apoptosis is in agreement withresults reported for bacterial pore-forming toxins structu-rally related to VCC.
Experiments performed in rabbit ileal loops allowed usto evaluate the effects produced by the CT� TCP�
V. cholerae strain during infection in vivo. Results obtainedusing TUNEL in situ indicated that the wild type strain butnot its isogenic VCC-null mutant induced increasedapoptosis in the intestinal epithelium, thus supporting themain contribution of VCC to produce apoptosis in thecontext of the rabbit intestine lumen. Together withincreased apoptosis, the histologic damage detected in theintestinal epithelium related to VCC action (i.e., shorteningof the villi and focal areas of necrosis) may account forthe production of the mucous and hemorrhagic fluidobserved in the rabbit ileal loops and for the diarrheagenic

ARTICLE IN PRESS
0
5
25
Control
Vc-wt
Vc-hlyA-
Apopto
tic n
ucle
i / vill
i
*
Vc-wt Vc-hlyA- Control
20
15
10
A B
Fig. 7. Vc-wt but not Vc-hlyA- induces apoptosis in the rabbit ileal loop assay. (A) TUNEL staining (red) of tissue sections from rabbit ileal loops
inoculated with Vc-wt, Vc-hlyA- and with PBS, are shown. Sections were counterstained with DAPI DNA binding dye (blue). (B) Apoptotic nuclei were
quantified in at least 50 different villi for each experimental condition and then expressed as the mean of apoptotic nuclei per villus7standard deviation
(black bars). Asterisk indicates a statistical significant difference compared to the PBS-inoculated control (Po0.001; Kruskal–Wallis non-parametric
multiple comparisons test). White bars: 100mm (upper panels) or 20 mm (lower panels).
H.A. Saka et al. / Microbial Pathogenesis 44 (2008) 118–128124
properties of the strain studied in this work. It is importantto mention that previous studies about the effects ofpurified VCC in vivo, reported that this toxin had thecapability to induce fluid accumulation in rabbit ilealloops, increased vascular permeability of rabbit and miceskin and rapid death of mice [14,35,39]. However, in thosereports a detailed description of the intestinal pathologywas not provided and the investigation of apoptosisinduction was not addressed. Furthermore, only purifiedVCC was tested, with the consequent limitation of using anarbitrary concentration of toxin that may not reflect thereal situation produced during infection.
This is, to our knowledge, the first report demonstratingthe participation of apoptosis as a cell death pathwaytriggered by VCC. Experiments using DIDS indicate thatpore formation is required for eliciting VCC-mediatedapoptosis. However, a more detailed analysis is still necessaryto determine how the generation the anion-channels leads tothe induction of apoptotic cell death. Apoptosis induction byVc-wt was detected in vitro but additionally in vivo whereas noeffect was caused by the isogenic VCC-null mutant strain,thus pointing out to the possible relevance of this phenom-enon in the pathogenesis. Further investigation of themechanistic details involved in apoptosis triggered by VCCwill contribute to gain knowledge about the pathogenesisinduced by this pore-forming toxin.
4. Materials and methods
4.1. Bacterial strains, supernatant preparation and VCC
The V. cholerae non-O1, non-O139 isolates (strainsVC175 and VC588) were isolated from two epidemiologi-cally unrelated patients suffering a cholera-like syndrome,
as previously described [40,41]. The isolates were positivefor hlyA and toxR genes and negative for ctxA, zot, ace,tcpA and stn genes as determined by PCR [40]. Forstudying purposes, the strain VC175 (designated as Vc-wt)was used. An isogenic hlyA null mutant from VC175(designated as Vc-hlyA-) was obtained by insertionalmutagenesis using the pLOF/Km suicide delivery vectoras described [34]. For analysis of fluid accumulation inrabbit ileal loops, a V. cholerae O1 biotype El Tor positivefor ctxA, zot, ace and tcpA genes isolated from a cholerapatient, designated as EP1/151 (CIBICI-CONICETV. cholerae collection), was used as positive control strain,while Escherichia coli DH5a was used as negative controlstrain. To obtain the sterile culture supernatants, singlecolonies of Vc-wt and Vc-hlyA- were inoculated into brainheart infusion (BHI) broth and incubated at 37 1C for 16 hunder shaking conditions (120 rpm). These cultures werethen centrifuged at 17,000� g for 10min at 4 1C; thesupernatants were sterilized through 0.22 mm low bindingprotein filters (Millipore Corp.) and kept on ice until use.Purified VCC was a gift of Dr. Chinen and obtained asdescribed [14]. When indicated, sterile culture supernatantfrom Vc-wt or purified VCC was preincubated with equalvolume of rabbit polyclonal specific anti-VCC antibody[65] (30min at room temperature) prior to treatment ofcells. For blockade of VCC anion channel activity, thecompound DIDS (4-acetamido-40-isothiocyanatostilbene-2,20-disulphonic acid), purchased from SIGMA, was usedas previously described [28].
4.2. Cell culture
Cells were cultured in DMEM supplemented with fetalbovine serum (20%v/v for Caco-2 and 10%v/v for

ARTICLE IN PRESSH.A. Saka et al. / Microbial Pathogenesis 44 (2008) 118–128 125
C2BBe1, MEF and COS-7 cells), L-glutamine (2mM),sodium bicarbonate (1.5 g/L), D-glucose (4.5 g/L), penicil-lin G (100U/ml) and streptomycin (100 mg/ml). For Caco-2and C2BBe1 cells, non-essential amino acids (0.1mM) andsodium pyruvate (1.0mM) were added. Cultures wereperformed at 37 1C and 5% CO2 in humidified atmosphere.
4.3. Cytotoxicity assay and quantification of cell viability
A cytotoxicity assay to determine cell viability aftertreatment with Vc-wt or Vc-hlyA- or with purified VCCwas performed using the Cell titer aqueous non-radioactivecell proliferation assay (Promega, Madison, WI, USA).This method allows the colorimetric determination of cellsin cytotoxicity or in proliferation assays, based on thedetection of the bioreduction of a tetrazolium compoundinto a formazan product in the presence of phenazinemethosulfate, which acts as an electron-coupling reagent.As the reaction is mediated by dehydrogenases present onlyin metabolically active cells, the amount of the formazanproduct is directly proportional to the number of livingcells in culture. The method was performed according tothe manufacturer’s instructions (Technical Bulletin 169,Promega). Briefly, 2.5� 104 cells were seeded onto 96-wellplates and incubated for 24 h at 37 1C and 5% CO2 inhumidified atmosphere. After 4 h of treatment, theabsorbance was determined at 490 nm using a Bio-Radmicroplate reader (Hercules, CA, USA).
4.4. DNA cleavage analysis
Confluent cells were obtained in 10 cm Petri dishes andtreated with purified VCC or with sterile culture super-natants from Vc-wt or Vc-hlyA- strains, as indicated inFig. 2. After treatment, cells were washed twice with ice-cold PBS, scraped and collected into eppendorf tubes. ForDNA purification, cells were lysed in 0.7ml of cetyltri-methylammonium bromide (CTAB) buffer (CTAB 2%P/V; NaCl 1.4M; EDTA 20mM, 2-mercapto-ethanol0.2%P/V; Tris �HCl 100mM [pH 8]) for 2 h at 60 1C.DNA was extracted with phenol/chloroform (1:1), pre-cipitated with isopropanol and pelleted at 13,000g for10min. The pellet was washed with 70% ethanol, air dried,resuspended in 20 ml of TE (10mM Tris-Cl, 1mM EDTA[pH 8]) containing 25 mg of RNase A and incubated 1 h at37 1C. Each DNA preparation was electrophoresed in a 2%agarose gel at 60V for 3 h and then stained with ethidiumbromide.
4.5. Flow cytometric analysis of hypodiploid nuclei
The method was based on the use of propidium iodide asdescribed [66]. Briefly, cells treated with purified VCC orwith sterile culture supernatants from Vc-wt or Vc-hlyA-strains (as indicated in Fig. 3) were trypsinized, washedtwice with ice-cold PBS and pelleted by centrifugation(350� g for 3min) prior to fixation and permeabilization
with ice-cold 70%v/v ethanol. The cells were then washedtwice with PBS and stained with propidium iodide(100 mg/ml; Sigma-Aldrich). Fluorescence of individualnuclei was acquired with the Research program in a flowcytometer (Ortho Diagnostics, Raritan, NJ). Events thatfell within the hypodiploid region (indicated as M1 region)were enumerated by the statistics function of the WinMDIV2.8 software (J. Trotter, Scripps Research Institute, LaJolla, CA) and the results were expressed as the percentageof the total events. A minimum of 10,000 events wereanalyzed for each experimental condition and all measure-ments were made at the same instrument setting.
4.6. Caspase-3 activity assay
Caspase-3 activity was determined using the Caspase-3(CPP32) Apoptosis Detection Colorimetric BioGeneTM Kit(United States Biological, Swampscott, MA, USA) follow-ing manufacturer0s instructions. Briefly, Caco-2 cells weretreated with purified VCC or with sterile culture super-natants from Vc-wt or Vc-hlyA- strains (as indicated inFig. 3). After treatment, cells were washed twice with icecold PBS, scraped, pelleted (350� g, 3min) and resus-pended in 50 ml of chilled cell lysis buffer. Caspase-3activity in cytosolic extracts (200 mg) from each experi-mental condition was measured by the addition of thecaspase-3 substrate Ac-DEVD-pNA (N-acetyl-Asp-Glu-Val-Asp-p-nitroanilide) (200 mM). The caspase-3 catalyzedrelease of p-nitroanilide was measured at 405 nm in a Bio-Rad microplate reader (Hercules, CA, USA).
4.7. Transmission electron microscopy
For TEM studies, Caco-2 cells treated or not withpurified VCC (80 pM, 6 h) were resuspended in 2% (v/v)glutaraldehyde, 4% (w/v) formaldehyde in 0.1M cacody-late buffer and then postfixed with 1% osmium tetroxide,dehydrated in increasing concentrations of acetone andembedded in Araldite at 60 1C. Thin sections were obtainedwith a diamond knife in a JEOL JUM-7 ultramicrotome,mounted on nickel grids, counterstained with uranylacetate and lead citrate and finally examined in a ZeissLEO 906E electron microscope.
4.8. Rabbit ileal loop assay
The rabbit ileal loop assay was carried out based on themethod described previously by De and Chatterjee [40,67].Briefly, male New Zealand white rabbits (2–2.5 kg) werestarved for 48 h before surgery. Then, the animals wereanesthetized with subcutaneous injection of ketamine(40mg/kg) and acepromazine (5mg/kg). Midline incisionswere made in the rabbit and the small intestine waswithdrawn and ligated at a distance of approximately10 cm from de ileocaecal region. Ten to twelve loops of6–8 cm were created in each animal by ligation whilepreserving intestinal blood supply. Loops were separated

ARTICLE IN PRESSH.A. Saka et al. / Microbial Pathogenesis 44 (2008) 118–128126
from each other by inter-loop segments of 2–3 cm. Theloops were inoculated with 1ml of bacterial suspension inPBS 1� (Vc-wt or Vc-hlyA-) adjusted to 105UFC/ml [68].Negative control loops were inoculated with 1ml of PBSalone. After 18h, rabbits were euthanized and the intestinesremoved for examination. The volume of fluid accumulatedin each loop was measured and the values were expressed asvolume (ml)/length (cm). Viable counts of V. cholerae strainsrecovered from the fluid accumulated within the loops weredetermined by plating serial dilutions onto TCBS agar. Eachstrain was tested in at least three different animals.
4.9. Histopathology and analysis of villus length
Strips from individual rabbit ileal loops were obtainedand collected in 10% neutral-buffered formalin, embeddedin paraffin and stained with hematoxilin–eosin (H&E)following the standard protocol. To analyze the villuslength, digital images of at least 50 villi per sample fromdifferent sections of tissue were determined using ImagePro Plus 4.5 software. Images were obtained using aNikon’s Eclipse TE2000 fluorescence microscope.
4.10. Determination of apoptosis in vivo by TUNEL in situ
For investigation of in vivo-induced apoptosis, thenumber of apoptotic nuclei per villus in the rabbit ilealloops was determined using Terminal dUTP Nick EndLabeling (TUNEL) in situ. TUNEL reactions wereperformed on thin sections of tissue after deparaffinizationand rehydration following manufacturer’s instructions(In situ Cell Death Detection kit, TMR Red, Roche AppliedScience). The sections were counterstained with the DNAbinding dye 40,6-Diamidino-2-phenylindole 10 mg/ml(DAPI, Sigma). Images were obtained using a Nikon’sEclipse TE2000 fluorescence microscope.
Acknowledgments
We thank Dr. Luis Patrito for his continuous encour-agement and support. We wish to thank Dr. Victor deLorenzo for providing the pLOF/Km plasmid used formutagenesis. We thank Dr. Isabel Chinen for the kindprovision of purified VCC and rabbit polyclonal anti-VCCantibody. This work was supported by the NationalCouncil for Scientific Research and Technology ofArgentina (CONICET), Agencia Nacional de PromocionCientifica y Tecnologica (ANPCyT–PICT 05-13446 toJLB), Secretaria de Ciencia y Tecnica-Universidad Nacio-nal de Cordoba (SECyT-UNC). HAS is a fellow recipientof CONICET. J.L.B. and J.R.E. are career investigatormembers of CONICET.
Appendix A. Supplementary data
Supplementary data associated with this article can be foundin the online version at doi:10.1016/j.micpath.2007.08.013.
References
[1] Colwell RR. Global climate and infectious disease: the cholera
paradigm. Science 1996;274:2025–31.
[2] Islam MS, Drasar BS, Sack RB. The aquatic flora and fauna as
reservoirs of Vibrio cholerae: a review. J Diarrhoeal Dis Res
1994;12:87–96.
[3] Sack DA, Sack RB, Nair GB, Siddique AK. Cholera. Lancet
2004;363:223–33.
[4] Kaper JB, Morris Jr. JG, Levine MM. Cholera. Clin Microbiol Rev
1995;8:48–86.
[5] Faruque SM, Albert MJ, Mekalanos JJ. Epidemiology, genetics, and
ecology of toxigenic Vibrio cholerae. Microbiol Mol Biol Rev
1998;62:1301–14.
[6] Pollitzer R, Swaroop S, Burrows W. Cholera. Monogr Ser World
Health Organ 1959;58:1001–19.
[7] World CJ, Yamawaki H, Berk BC. Thioredoxin in the cardiovascular
system. J Mol Med 2006;84:997–1003.
[8] Dziejman M, Serruto D, Tam VC, Sturtevant D, Diraphat P,
Faruque SM, et al. Genomic characterization of non-O1, non-O139
Vibrio cholerae reveals genes for a type III secretion system. Proc Natl
Acad Sci USA 2005;102:3465–70.
[9] Faruque SM, Sack DA, Sack RB, Colwell RR, Takeda Y, Nair GB.
Emergence and evolution of Vibrio cholerae O139. Proc Natl Acad
Sci USA 2003;100:1304–9.
[10] Karaolis DK, Johnson JA, Bailey CC, Boedeker EC, Kaper JB,
Reeves PR. A Vibrio cholerae pathogenicity island associated with
epidemic and pandemic strains. Proc Natl Acad Sci USA
1998;95:3134–9.
[11] Blake PA, Weaver RE, Hollis DG. Diseases of humans (other than
cholera) caused by vibrios. Annu Rev Microbiol 1980;34:341–67.
[12] Finch MJ, Valdespino JL, Wells JG, Perez-Perez G, Arjona F,
Sepulveda A, et al. Non-O1 Vibrio cholerae infections in Cancun,
Mexico. Am J Trop Med Hyg 1987;36:393–7.
[13] Ninin E, Caroff N, El Kouri D, Espaze E, Richet H, Quilici ML,
et al. Nontoxigenic Vibrio cholerae O1 bacteremia: case report and
review. Eur J Clin Microbiol Infect Dis 2000;19:489–91.
[14] Yamamoto K, Al-Omani M, Honda T, Takeda Y, Miwatani T. Non-
O1 Vibrio cholerae hemolysin: purification, partial characterization,
and immunological relatedness to El Tor hemolysin. Infect Immun
1984;45:192–6.
[15] Goldberg SL, Murphy JR. Molecular cloning of the hemolysin
determinant from Vibrio cholerae El Tor. J Bacteriol 1984;160:
239–44.
[16] Honda T, Finkelstein RA. Purification and characterization of a
hemolysin produced by Vibrio cholerae biotype El Tor: another toxic
substance produced by cholera vibrios. Infect Immun 1979;26:
1020–7.
[17] Heidelberg JF, Eisen JA, Nelson WC, Clayton RA, Gwinn ML,
Dodson RJ, et al. DNA sequence of both chromosomes of the
cholera pathogen Vibrio cholerae. Nature 2000;406:477–83.
[18] Nagamune K, Yamamoto K, Naka A, Matsuyama J, Miwatani T,
Honda T. In vitro proteolytic processing and activation of the
recombinant precursor of El Tor cytolysin/hemolysin (pro-HlyA) of
Vibrio cholerae by soluble hemagglutinin/protease of V. cholerae,
trypsin, and other proteases. Infect Immun 1996;64:4655–8.
[19] Nagamune K, Yamamoto K, Honda T. Intramolecular chaperone
activity of the pro-region of Vibrio cholerae El Tor cytolysin. J Biol
Chem 1997;272:1338–43.
[20] Alm RA, Stroeher UH, Manning PA. Extracellular proteins of Vibrio
cholerae: nucleotide sequence of the structural gene (hlyA) for the
haemolysin of the haemolytic El Tor strain 017 and characterization
of the hlyA mutation in the non-haemolytic classical strain 569B. Mol
Microbiol 1988;2:481–8.
[21] Menzl K, Maier E, Chakraborty T, Benz R. HlyA hemolysin of
Vibrio cholerae O1 biotype E1 Tor. Identification of the hemolytic
complex and evidence for the formation of anion-selective ion-
permeable channels. Eur J Biochem 1996;240:646–54.

ARTICLE IN PRESSH.A. Saka et al. / Microbial Pathogenesis 44 (2008) 118–128 127
[22] Zitzer A, Palmer M, Weller U, Wassenaar T, Biermann C, Tranum-
Jensen J, et al. Mode of primary binding to target membranes and
pore formation induced by Vibrio cholerae cytolysin (hemolysin). Eur
J Biochem 1997;247:209–16.
[23] Zitzer A, Walev I, Palmer M, Bhakdi S. Characterization of Vibrio
cholerae El Tor cytolysin as an oligomerizing pore-forming toxin.
Med Microbiol Immunol 1995;184:37–44.
[24] Zitzer A, Zitzer O, Bhakdi S, Palmer M. Oligomerization of Vibrio
cholerae cytolysin yields a pentameric pore and has a dual specificity
for cholesterol and sphingolipids in the target membrane. J Biol
Chem 1999;274:1375–80.
[25] Harris JR, Bhakdi S, Meissner U, Scheffler D, Bittman R, Li G, et al.
Interaction of the Vibrio cholerae cytolysin (VCC) with cholesterol,
some cholesterol esters, and cholesterol derivatives: a TEM study.
J Struct Biol 2002;139:122–35.
[26] Krasilnikov OV, Muratkhodjaev JN, Zitzer AO. The mode of action
of Vibrio cholerae cytolysin. The influences on both erythrocytes and
planar lipid bilayers. Biochim Biophys Acta 1992;1111:7–16.
[27] Krasilnikov OV, Merzlyak PG, Lima VL, Zitzer AO, Valeva A,
Yuldasheva LN. Pore formation by Vibrio cholerae cytolysin requires
cholesterol in both monolayers of the target membrane. Biochimie
2007;89:271–7.
[28] Moschioni M, Tombola F, de Bernard M, Coelho A, Zitzer A,
Zoratti M, et al. The Vibrio cholerae haemolysin anion channel is
required for cell vacuolation and death. Cell Microbiol 2002;4:
397–409.
[29] Alm RA, Mayrhofer G, Kotlarski I, Manning PA. Amino-terminal
domain of the El Tor haemolysin of Vibrio cholerae O1 is expressed in
classical strains and is cytotoxic. Vaccine 1991;9:588–94.
[30] Figueroa-Arredondo P, Heuser JE, Akopyants NS, Morisaki JH,
Giono-Cerezo S, Enriquez-Rincon F, et al. Cell vacuolation caused
by Vibrio cholerae hemolysin. Infect Immun 2001;69:1613–24.
[31] Coelho A, Andrade JR, Vicente AC, Dirita VJ. Cytotoxic cell
vacuolating activity from Vibrio cholerae hemolysin. Infect Immun
2000;68:1700–5.
[32] Mitra R, Figueroa P, Mukhopadhyay AK, Shimada T, Takeda Y,
Berg DE, et al. Cell vacuolation, a manifestation of the El tor
hemolysin of Vibrio cholerae. Infect Immun 2000;68:1928–33.
[33] Zitzer A, Wassenaar TM, Walev I, Bhakdi S. Potent membrane-
permeabilizing and cytocidal action of Vibrio cholerae cytolysin on
human intestinal cells. Infect Immun 1997;65:1293–8.
[34] Gutierrez MG, Saka HA, Chinen I, Zoppino FC, Yoshimori T,
Bocco JL, et al. Protective role of autophagy against Vibrio cholerae
cytolysin, a pore-forming toxin from V.cholerae. Proc Natl Acad Sci
USA 2007;104:1829–34.
[35] Ichinose Y, Yamamoto K, Nakasone N, Tanabe MJ, Takeda T,
Miwatani T, et al. Enterotoxicity of El Tor-like hemolysin of non-O1
Vibrio cholerae. Infect Immun 1987;55:1090–3.
[36] Fullner KJ, Boucher JC, Hanes MA, Haines 3rd GK, Meehan BM,
Walchle C, et al. The contribution of accessory toxins of Vibrio
cholerae O1 El Tor to the proinflammatory response in a murine
pulmonary cholera model. J Exp Med 2002;195:1455–62.
[37] Levine MM, Kaper JB, Herrington D, Losonsky G, Morris JG,
Clements ML, et al. Volunteer studies of deletion mutants of Vibrio
cholerae O1 prepared by recombinant techniques. Infect Immun
1988;56:161–7.
[38] Faruque SM, Chowdhury N, Kamruzzaman M, Dziejman M,
Rahman MH, Sack DA, et al. Genetic diversity and virulence
potential of environmental Vibrio cholerae population in a cholera-
endemic area. Proc Natl Acad Sci USA 2004;101:2123–8.
[39] Zitzer AO, Nakisbekov NO, Li AV, Semiotrochev VL, Kiseliov Yu
L, Muratkhodjaev JN, et al. Entero-cytolysin (EC) from Vibrio
cholerae non-O1 (some properties and pore-forming activity).
Zentralbl Bakteriol 1993;279:494–504.
[40] Bidinost C, Saka HA, Aliendro O, Sola C, Panzetta-Duttari G,
Carranza P, et al. Virulence factors of non-O1 non-O139 Vibrio
cholerae isolated in Cordoba, Argentina. Rev Argent Microbiol
2004;36:158–63.
[41] Patrito E, Aliendro O, Lesa M. Isolation of Vibrio cholerae non-O1
from patients with acute gastroenteritis. Rev Fac Cien Med Univ Nac
Cordoba 1999;56:85–9.
[42] Aroian R, van der Goot FG. Pore-forming toxins and cellular non-
immune defenses (CNIDs). Curr Opin Microbiol 2007;10:57–61.
[43] Braun JS, Sublett JE, Freyer D, Mitchell TJ, Cleveland JL,
Tuomanen EI, et al. Pneumococcal pneumolysin and H(2)O(2)
mediate brain cell apoptosis during meningitis. J Clin Invest
2002;109:19–27.
[44] Carrero JA, Calderon B, Unanue ER. Listeriolysin O from Listeria
monocytogenes is a lymphocyte apoptogenic molecule. J Immunol
2004;172:4866–74.
[45] Chakrabarti G, Zhou X, McClane BA. Death pathways activated in
CaCo-2 cells by Clostridium perfringens enterotoxin. Infect Immun
2003;71:4260–70.
[46] Menzies BE, Kourteva I. Staphylococcus aureus alpha-toxin induces
apoptosis in endothelial cells. FEMS Immunol Med Microbiol 2000;
29:39–45.
[47] Nelson KL, Brodsky RA, Buckley JT. Channels formed by
subnanomolar concentrations of the toxin aerolysin trigger apoptosis
of T lymphomas. Cell Microbiol 1999;1:69–74.
[48] Steller H. Mechanisms and genes of cellular suicide. Science 1995;
267:1445–9.
[49] Hockenbery D. Defining apoptosis. Am J Pathol 1995;146:16–9.
[50] Olson R, Gouaux E. Crystal structure of the Vibrio cholerae cytolysin
(VCC) pro-toxin and its assembly into a heptameric transmembrane
pore. J Mol Biol 2005;350:997–1016.
[51] Pantano S, Montecucco C. A molecular model of the Vibrio cholerae
cytolysin transmembrane pore. Toxicon 2006;47:35–40.
[52] Darzynkiewicz Z, Bruno S, Del Bino G, Gorczyca W, Hotz MA,
Lassota P, et al. Features of apoptotic cells measured by flow
cytometry. Cytometry 1992;13:795–808.
[53] Nicoletti I, Migliorati G, Pagliacci MC, Grignani F, Riccardi C. A
rapid and simple method for measuring thymocyte apoptosis by
propidium iodide staining and flow cytometry. J Immunol Methods
1991;139:271–9.
[54] Enari M, Sakahira H, Yokoyama H, Okawa K, Iwamatsu A, Nagata
S. A caspase-activated DNase that degrades DNA during apoptosis,
and its inhibitor ICAD. Nature 1998;391:43–50.
[55] Nicholson DW, Ali A, Thornberry NA, Vaillancourt JP, Ding CK,
Gallant M, et al. Identification and inhibition of the ICE/CED-3
protease necessary for mammalian apoptosis. Nature 1995;376:37–43.
[56] Fink SL, Cookson BT. Apoptosis, pyroptosis, and necrosis:
mechanistic description of dead and dying eukaryotic cells. Infect
Immun 2005;73:1907–16.
[57] Mukhopadhyay AK, Saha PK, Garg S, Bhattacharya SK, Shimada
T, Takeda T, et al. Distribution and virulence of Vibrio cholerae
belonging to serogroups other than O1 and O139: a nationwide
survey. Epidemiol Infect 1995;114:65–70.
[58] Olson R, Gouaux E. Vibrio cholerae cytolysin is composed of an
alpha-hemolysin-like core. Protein Sci 2003;12:379–83.
[59] Haslinger-Loffler B, Kahl BC, Grundmeier M, Strangfeld K, Wagner
B, Fischer U, et al. Multiple virulence factors are required for
Staphylococcus aureus-induced apoptosis in endothelial cells. Cell
Microbiol 2005;7:1087–97.
[60] Haslinger B, Strangfeld K, Peters G, Schulze-Osthoff K, Sinha B.
Staphylococcus aureus alpha-toxin induces apoptosis in peripheral
blood mononuclear cells: role of endogenous tumour necrosis factor-
alpha and the mitochondrial death pathway. Cell Microbiol
2003;5:729–41.
[61] Jonas D, Walev I, Berger T, Liebetrau M, Palmer M, Bhakdi S.
Novel path to apoptosis: small transmembrane pores created by
staphylococcal alpha-toxin in T lymphocytes evoke internucleosomal
DNA degradation. Infect Immun 1994;62:1304–12.
[62] Bayles KW, Wesson CA, Liou LE, Fox LK, Bohach GA, Trumble
WR. Intracellular Staphylococcus aureus escapes the endosome
and induces apoptosis in epithelial cells. Infect Immun 1998;66:
336–42.

ARTICLE IN PRESSH.A. Saka et al. / Microbial Pathogenesis 44 (2008) 118–128128
[63] Bantel H, Sinha B, Domschke W, Peters G, Schulze-Osthoff K,
Janicke RU. Alpha-toxin is a mediator of Staphylococcus aureus-
induced cell death and activates caspases via the intrinsic death
pathway independently of death receptor signaling. J Cell Biol
2001;155:637–48.
[64] Genestier AL, Michallet MC, Prevost G, Bellot G, Chalabreysse L,
Peyrol S, et al. Staphylococcus aureus Panton-Valentine leukocidin
directly targets mitochondria and induces Bax-independent apoptosis
of human neutrophils. J Clin Invest 2005;115:3117–27.
[65] Chinen I, Toma C, Honma Y, Higa N, Iwanaga M. Hemolysin
production by Vibrio cholerae as examined by hemolytic and
antigenic activities. Jpn J Trop Med Hyg 1996;24:151–5.
[66] Telford WG, King LE, Fraker PJ. Rapid quantitation of apoptosis in
pure and heterogeneous cell populations using flow cytometry.
J Immunol Methods 1994;172:1–16.
[67] De SN, Chatterje DN. An experimental study of the mechanism of
action of Vibrio cholerae on the intestinal mucous membrane.
J Pathol Bacteriol 1953;66:559–62.
[68] Koley H, Mitra R, Basu A, Mukhopadhyay AK, Saha PK,
Ramakrishna BS, et al. Response of wild-type mutants of Vibrio
cholerae O1 possessing different combinations of virulence genes
in the ligated rabbit ileal loop and in using chambers: evidence
for the presence of additional secretogen. J Med Microbiol 1999;48:
51–7.




















