
Computation of angular velocity and acceleration tensors by direct measurements

324
Ann. N.Y. Acad. Sci. 956: 324–337 (2002). © 2002 New York Academy of Sciences.
Vergence-Mediated Modulation of the Human Horizontal Angular VOR Provides Evidence of Pathway-Specific Changes in VOR Dynamics
DAVID M. LASKER,a STEFANO RAMAT,b,c JOHN P. CAREY,a ANDLLOYD B. MINORa
aDepartment of Otolaryngology—Head & Neck Surgery, The Johns Hopkins University School of Medicine, Baltimore, Maryland 21287, USAbDepartment of Neurology, The Johns Hopkins University School of Medicine,Baltimore, Maryland 21287, USAcDipartimento di Informatica e Sistemistica, Università di Pavia, Pavia, Italy
ABSTRACT: The horizontal vestibulo-ocular reflex (VOR) evoked by passive,high-acceleration, head-on-body rotations (head thrusts) while viewing a far(124-cm) or near (15-cm) target was recorded (scleral search coil) in four sub-jects with normal vestibular function and in one subject with unilateral vesti-bular hypofunction. For responses in the subjects with normal vestibularfunction, the latency of responses relative to the onset of head movement was7.5 ± 1.5 ms for the VOR and 21.6 ± 1.2 ms for the vergence-mediated increasein VOR gain. The gain of the VOR at the peak of the velocity response whileviewing a far target was 1.01 ± 0.06; while viewing a near target, it was 1.25 ±0.08 (p <0.003). The responses were modeled with two pathways based on thedifferent latencies. The “far-viewing” pathway was represented by a constantgain term. The “near-viewing” pathway was represented by a first-order leadterm, a gain that was dependent on viewing distance, and a delay. Analysis ofthe responses revealed that the lead term was greater for the adducting thanthe abducting eye. In the subject with unilateral vestibular hypofunction, ip-silesional responses showed no change in VOR gain with respect to viewing dis-tance. Contralesional responses retained the vergence-dependent increase ingain. A bilateral model was developed based on the data from the subjects withnormal vestibular function. Simulations of this model when inputs were elimi-nated from one side predict the changes observed in the subject with unilateralvestibular hypofunction. The response asymmetries arise because the near-viewing pathway is more susceptible to inhibitory cutoff than is the far-viewingpathway.
KEYWORDS: vestibulo-ocular reflex; vergence; modulation; vestibular function
Address for correspondence: Dr. Lloyd Minor, Department of Otolaryngology—Head & NeckSurgery, Johns Hopkins University School of Medicine, 601 N. Caroline St., Rm. 6253, Balti-more, MD 21287-0910. Voice: 410-955-3403; fax: 410-955-0035.

325LASKER et al.: VESTIBULO-OCULAR REFLEX DYNAMICS
INTRODUCTION
In studies of the horizontal angular vestibulo-ocular reflex (VOR) evoked byhigh-frequency, high-acceleration rotations in squirrel monkeys, we identified twopathways involved in control of the reflex.1 A linear pathway is responsible for theconstant gain of the reflex across the range of stimulus frequencies of 0.5–15 Hz forpeak stimulus velocities of 20 degrees/s. The linear pathway has a phase lead thatrises with frequency. This phase lead compensates for the phase lag that would beexpected based on the 7-ms delay in the reflex, thereby keeping the VOR relativelyin phase, but opposite in direction, to head velocity.
Our studies in squirrel monkeys also provide evidence of a nonlinear pathwaythat augments the responses provided by the linear pathway. There are two compo-nents of the nonlinearity: (1) a frequency- and velocity-dependent rise in gain and(2) a rectification due to inhibitory cutoff. The phasic dynamics of the nonlinearpathway match those that have been recorded in irregularly discharging afferents.
The gain of the nonlinear pathway is more modifiable than is the gain of the linearpathway. A selectively greater increase in the gain of the nonlinear than the linearpathway on the intact side after unilateral plugging of the three semicircular canals2
or unilateral labyrinthectomy3 is responsible for the recovery of contralesional gainafter these lesions. The VOR following spectacle-induced adaptation provides com-pelling evidence in support of these two pathways.4 In responses to steps of acceler-ation (3,000 degrees/s2 acceleration reaching a peak velocity of 150 degrees/s)following long-term adaptation to 2.2 × magnifying spectacles, the VOR gain duringthe step of acceleration rises to almost equal the power of the lenses. By contrast, theVOR gain at the velocity plateau of the stimulus averaged 1.5, a value commonly ob-served in previous studies of the VOR that employed stimuli that were of lower fre-quency and acceleration.5,6
We sought to determine if there were similar pathways in the human VOR. Wehypothesized that responses for viewing near in comparison to far targets for rapid,unpredictable, head-on-body rotations (head thrusts) might reveal similar pathway-specific effects. Our findings indicate that responses when viewing the near targetmanifest a phase lead. This lead is greater in the adducting than in the abducting eye.These changes in VOR dynamics with viewing distance were absent in the ipsile-sional responses for a patient with unilateral vestibular hypofunction. Future studiesare needed to evaluate the effect of changes in the axis of rotation with respect to theotoliths on response dynamics.
METHODS
Subjects
Three-dimensional eye movements were recorded from four subjects (aged 20–50 years) with normal vestibular function and one patient with left vestibular hypo-function. This patient, a 48-year-old woman, had intractable vertigo from left Me-niere’s disease and was treated with intratympanic gentamicin at an outsideinstitution. A wick was inserted in her left tympanic membrane, and she placed gen-tamicin drops in the ear twice per day for 3 weeks. She developed signs of left ves-

326 ANNALS NEW YORK ACADEMY OF SCIENCES
tibular hypofunction: right beating spontaneous nystagmus, right beating nystagmusafter horizontal head shaking, and hypometric horizontal VOR evoked by rapid headthrusts to the left. She was tested 3 months after receiving the gentamicin. All sub-jects gave informed consent for eye movement recording through a protocol ap-proved by the Joint Committee on Clinical Investigation of the Johns HopkinsUniversity School of Medicine.
Recording of Eye and Head Movements
The movements of both eyes were recorded around all three axes of rotation (hor-izontal, vertical, and torsional) with the magnetic search coil technique. The fieldcoil system consisted of a cubic frame with a side length of 1.02 m producing threeorthogonal magnetic fields (frequencies: 55.5, 83.3, and 42.6 kHz; intensity:0.088 G). The dual search coils (manufactured by Skalar, Delft, The Netherlands)yielded two sensitivity vectors each characterized by voltages induced in one of thetwo coils by the three orthogonal fields. An in vitro calibration was performed beforeeach experiment in which voltage offsets were nullified by placing the coils into ametallic tube that completely shielded the coil from the magnetic fields. The coil wasthen placed on a gimbal system located in the center of the magnetic coil frame. Coilgains were determined by aligning the sensitivity vectors of each coil with each ofthe three magnetic fields. The output signals of the coils were filtered with a band-width of 0–90 Hz and sampled at 1,000 Hz with 12-bit resolution. The annuli wereplaced on each eye after administration of a topical anesthetic (proparacaine HCl0.5%). Head rotations were sensed with a dual search coil embedded in a bite barthat was molded to the subject’s dental occlusion. Further details of the calibrationand recording procedures can be found in Straumann et al.7
Head Rotation Paradigms
The subject, seated with the head centered in the magnetic field, was instructedto gaze at a light-emitting diode (LED) located either 124 cm or 15 cm directly infront of the eyes. The examiner stood behind the subject and grasped the head overthe temporoparietal area. The axis of rotation was approximately centered with re-spect to the otoliths. The head was kept stationary in a comfortable, “upright” posi-tion prior to each head thrust. This position placed Reid’s stereotactic line (inferiororbital rim to superior external auditory canal) 7 degrees nose-up from the earth-horizontal plane. From this position, the examiner rapidly rotated the head by 10–20degrees in the horizontal plane. Peak acceleration for these stimuli ranged from 800–3,000 degrees/s2 and peak velocity was from 80–150 degrees/s. Head thrusts wererandomized in terms of direction.
Analysis of Eye and Head Movements
Successive head rotations in each direction and for each viewing condition werealigned based on the onset of the head rotation. The onset of the head and eye move-ments was determined to be the points at which the velocity signals deviated fromthe mean value measured before onset of the stimulus for head and eye velocity, re-spectively, by >2 SD. The difference in these two onset points measured for each trialwas defined as response latency. Latency for the vergence-induced increase in VOR

327LASKER et al.: VESTIBULO-OCULAR REFLEX DYNAMICS
gain was measured as the time at which the eye velocity while viewing a near targetdiffered from the eye velocity while viewing a far target by 1 SD.
Gain was determined as the ratio of eye to head velocity at the peak of head ve-locity. Comparisons of gain based on viewing condition and direction of the headthrust were made with t tests.
Modeling of Responses
Mathematical models of the VOR were formulated in Simulink® (The Math-Works). The Dormand-Prince method with a fixed step size of 0.001 s was used forsimulations of the ordinary differential equations.
RESULTS
Responses to rightward and leftward head thrusts under near- and far-viewingconditions in one normal subject are shown in FIGURE 1. The responses were sym-metric for both directions of rotation and for each viewing condition. Averaged re-sponses for each viewing distance and ideal responses for each eye are shown inFIGURE 2A and B. The response in the far-viewing condition (FIG. 2A) more closely
FIGURE 1. Head velocity (dashed traces) and eye velocity (left eye, black; right eye,gray) for head thrusts while viewing far (124 cm; A,C) or near (15 cm; B,D) target in Subject3. (A,B) Responses for head thrusts to the left. (C,D) Responses for head thrusts to the right.On the velocity axis, positive values represent eye or head movement to the right and nega-tive values represent eye or head movement to the left.

328 ANNALS NEW YORK ACADEMY OF SCIENCES
matches the ideal response than does the response in the near-viewing condition(FIG. 2B).
Our analysis of these responses involved calculation of the latency of the VOR aswell as the latency of the vergence-induced change in the VOR. We also calculatedthe gain (defined as peak eye velocity/peak head velocity) for each viewing condi-tion. From these parameters of latency and gain, we developed mathematical modelsto fit and analyze our data.
Latency of the Responses
The responses to head thrusts under near- and far-viewing conditions from thefour normal subjects are shown in FIGURE 3. For the display and analysis of thesedata, head thrusts were divided into two groups based on the mean acceleration from10–40 ms after the onset of the stimulus. The latency of the VOR during the far-tar-get viewing condition (124 cm, FIG. 1A) measured 7.7 ± 1.2 ms for rotations in theleftward direction and 7.4 ± 2.0 ms in the rightward direction (p >0.7). The latencymeasured during the near-target viewing condition measured 7.2 ± 1.7 ms for rota-tions in the leftward direction and 7.8 ± 1.0 for rotations in the rightward direction(p >0.6). There was no difference in latency of the onset of the VOR at the two dif-ferent target distances (p >0.5).
The average latency, measured from the onset of the head movement, for the ver-gence-induced change in gain measured 21.6 ± 1.2 ms in the four subjects.
FIGURE 2. Averaged responses for head thrusts while viewing far (A) or near (B) tar-get in Subject 2. Rightward eye velocity (resulting from leftward head thrust) is positive,and leftward eye velocity (resulting from rightward head thrust) is negative. Data are dis-played as a shaded window depicting the region of 1 SD surrounding the mean. Responsesshown by gray shading are for the right eye and those with black shading are for the left eye.The solid gray and black traces show ideal responses for the right and left eyes, respectively.Note that the ideal responses for the left and right eye are the same for far target viewing andtherefore only the black trace is noted on the plot. For near-target viewing, the ideal re-sponse of the abducting eyes has a larger gain than does that of the adducting eye.8 This dif-ference arises because of the location of the axis of rotation posterior to the eyes. Asdescribed in TABLE 2, the gain values for the two eyes were similar in these subjects. Thetrajectory of the responses for the adducting eye shows phasic dynamics that are more prom-inent than those recorded from the abducting eye.

329LASKER et al.: VESTIBULO-OCULAR REFLEX DYNAMICS
Dynamics of the Vestibulo-Ocular Reflex
The VOR in response to high acceleration head rotations is shown in FIGURE 1.Peak gains were measured by dividing the peak eye velocity by the peak head veloc-ity. The results for the four subjects during the far-target viewing condition areshown in TABLE 1. The gain for leftward rotations measured 1.02 ± 0.06 for the lefteye and 1.02 ± 0.07 for the right eye (p >0.5). The gain for rightward rotations mea-sured 1.0 ± 0.07 for left eye and 0.99 ± 0.03 for the right eye (p >0.7). After poolingthe data for the two eyes, the gain for head thrusts in the leftward direction was 1.01± 0.06 and the gain for head thrusts in the rightward direction was 1.01 ± 0.05(p >0.9).
The results for the four subjects during the near-target viewing condition areshown in TABLE 2. The gain for leftward rotations when measured from the left eyewas 1.24 ± 0.09 and when measured from the right eye was 1.26 ± 0.09 (p >0.5). Thegain for rightward rotations when measured from the left eye was 1.28 ± 0.06 andwhen measured from the right eye was 1.19 ± 0.08 (p >0.3). After pooling the datafor the left and right eyes, the gain measured 1.25 ± 0.08 for the leftward directionand 1.24 ± 0.08 for the rightward direction (p >0.8). There was a significant differ-ence between the gains for the far-target viewing condition and the near-target view-ing condition (p <0.003).
FIGURE 3. Latency of the vergence-mediated change in gain of the VOR. Data fromthe four subjects with normal vestibular function were pooled. Leftward eye velocity (result-ing from rightward head movements) was inverted so that traces in both directions could becombined. The responses were then divided into two groups according to the peak acceler-ation of the head during the stimuli. The group of responses to lower acceleration stimulihad peak head acceleration of 800–1,200 degrees/s2 (lower traces), whereas those in thehigher acceleration group had peak acceleration of 1,300–2,000 degrees/s2 (upper traces).Average responses for far targets are shown by the solid traces, and average responses fornear targets are shown by the dashed traces. Each trace represents an average of 60–80 stim-ulus repetitions. The X represents the time at which the near-target eye velocity differs by 1SD from the far-target velocity.

330 ANNALS NEW YORK ACADEMY OF SCIENCES
As observed for the responses during far-target viewing (FIG. 2A), there were dif-ferences in the trajectories of the eye velocity for the adducting in comparison withthe abducing eye. The adducting eye had a velocity profile with a larger lead thanthat of the abducting eye (FIG. 2B). This effect was more prominent in the near-targetthan in the far-target condition.
In the subject tested following intratympanic gentamicin (FIG. 4), gains weremeasured while viewing a far and a near target for head thrusts delivered towards (ip-silesional) and away from (contralesional) the treated side. The gain in the contra-lesional direction during the far-viewing condition measured 0.69 ± 0.04 and was0.87 ± 0.08 during the near-viewing condition (p <0.0001). As noted in responsesfrom normal subjects, the adducting eye for the responses to contralesional headthrusts showed more phasic dynamics than the abducting eye. The gain measured inthe ipsilesional direction during the far-viewing condition was 0.38 ± 0.05 and was0.36 ± 0.06 during the near-viewing condition (p >0.15). Responses for the adduct-ing and abducting eye were similar for ipsilesional rotations.
Mathematical Models of the Data
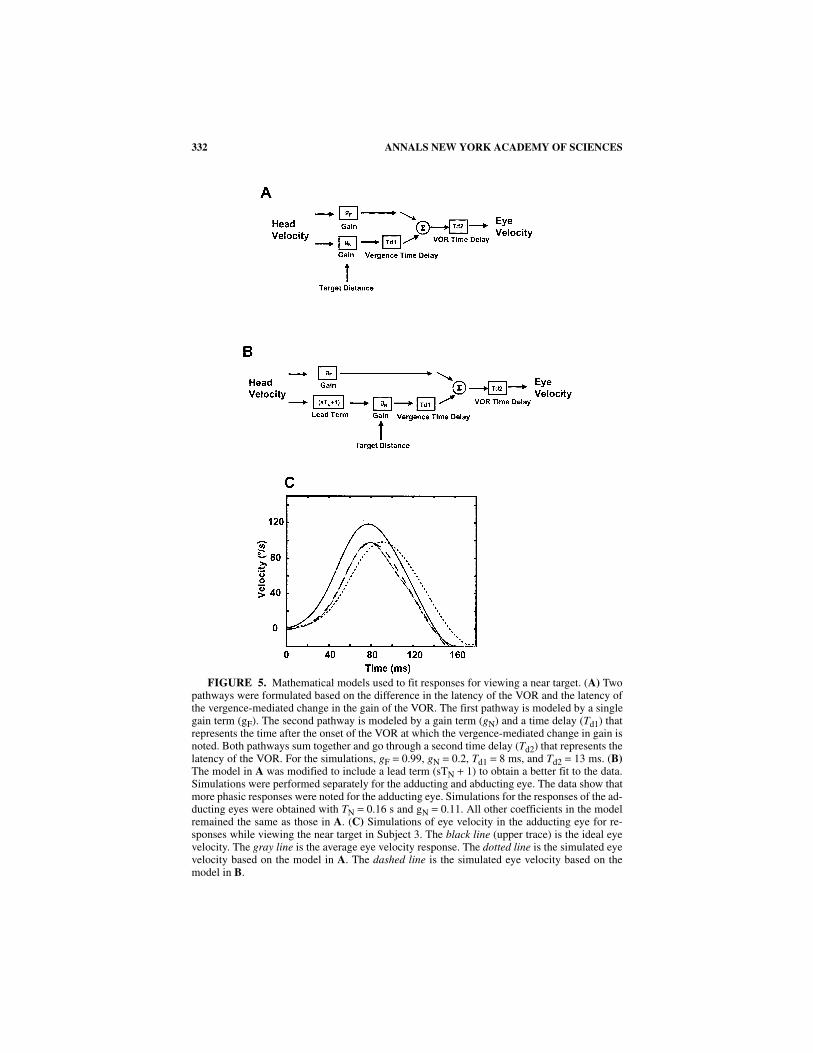
We used mathematical models with two pathways to analyze the responses fornear and far viewing (FIG. 5A). The pathways differ in their dynamic properties andin latency relative to the onset of the head movement. The far-viewing pathway hasa constant gain. The near-viewing pathway has a gain that is inversely proportionalto viewing distance (higher gain for near targets) and a time delay that is determinedby the onset of the viewing distance-dependent increase in gain. The response pre-dicted from this model for the near-viewing condition used in this study is shown inFIGure 5C (dotted trace). The predicted responses from this model lag behind the re-corded data during the head movement.
TABLE 1. Response gain for far-target viewing conditions
Subject No.
Left eye Right eye
Leftward Rightward Leftward Rightward
1 0.95 1.00 0.92 0.952 1.09 1.06 1.07 1.033 1.05 1.10 1.04 1.004 1.00 0.93 0.95 0.99
TABLE 2. Response gain for near-target viewing condition
Subject No.
Left eye Right eye
Leftward Rightward Leftward Rightward
1 1.21 1.36 1.24 1.242 1.35 1.26 1.38 1.253 1.27 1.30 1.22 1.084 1.14 1.21 1.19 1.20
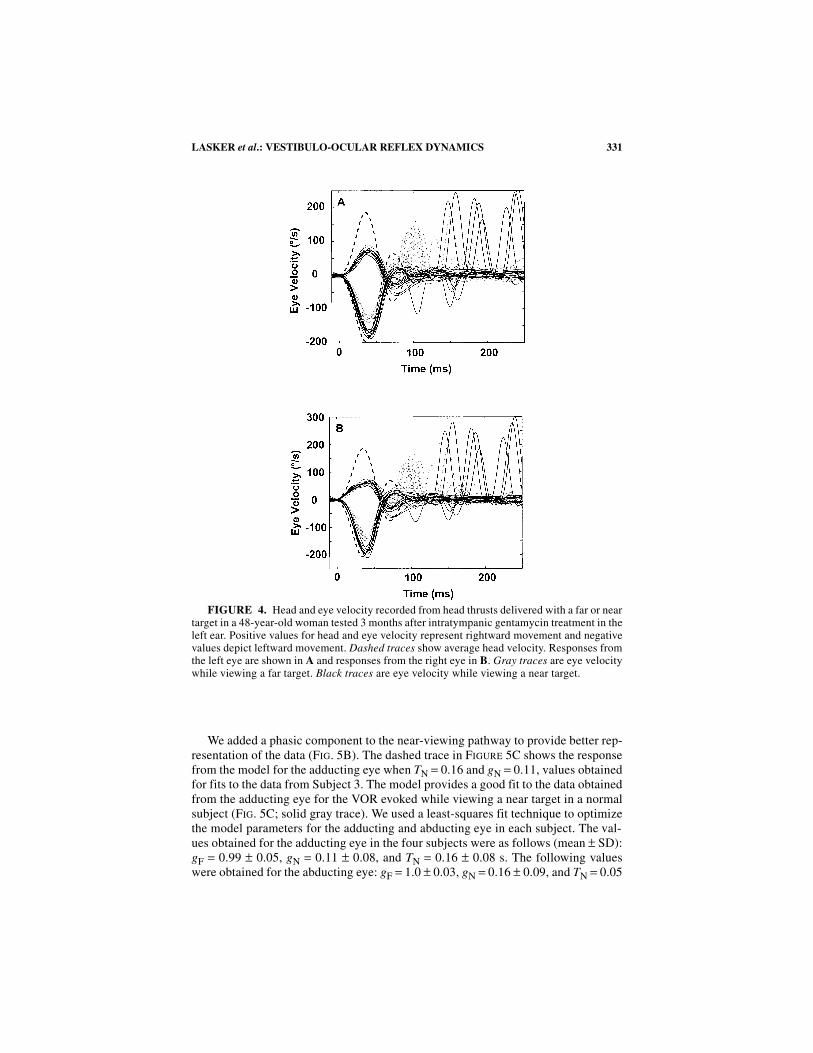
331LASKER et al.: VESTIBULO-OCULAR REFLEX DYNAMICS
We added a phasic component to the near-viewing pathway to provide better rep-resentation of the data (FIG. 5B). The dashed trace in FIGURE 5C shows the responsefrom the model for the adducting eye when TN = 0.16 and gN = 0.11, values obtainedfor fits to the data from Subject 3. The model provides a good fit to the data obtainedfrom the adducting eye for the VOR evoked while viewing a near target in a normalsubject (FIG. 5C; solid gray trace). We used a least-squares fit technique to optimizethe model parameters for the adducting and abducting eye in each subject. The val-ues obtained for the adducting eye in the four subjects were as follows (mean ± SD):gF = 0.99 ± 0.05, gN = 0.11 ± 0.08, and TN = 0.16 ± 0.08 s. The following valueswere obtained for the abducting eye: gF = 1.0 ± 0.03, gN = 0.16 ± 0.09, and TN = 0.05
FIGURE 4. Head and eye velocity recorded from head thrusts delivered with a far or neartarget in a 48-year-old woman tested 3 months after intratympanic gentamycin treatment in theleft ear. Positive values for head and eye velocity represent rightward movement and negativevalues depict leftward movement. Dashed traces show average head velocity. Responses fromthe left eye are shown in A and responses from the right eye in B. Gray traces are eye velocitywhile viewing a far target. Black traces are eye velocity while viewing a near target.
332 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 5. Mathematical models used to fit responses for viewing a near target. (A) Twopathways were formulated based on the difference in the latency of the VOR and the latency ofthe vergence-mediated change in the gain of the VOR. The first pathway is modeled by a singlegain term (gF). The second pathway is modeled by a gain term (gN) and a time delay (Td1) thatrepresents the time after the onset of the VOR at which the vergence-mediated change in gain isnoted. Both pathways sum together and go through a second time delay (Td2) that represents thelatency of the VOR. For the simulations, gF = 0.99, gN = 0.2, Td1 = 8 ms, and Td2 = 13 ms. (B)The model in A was modified to include a lead term (sTN + 1) to obtain a better fit to the data.Simulations were performed separately for the adducting and abducting eye. The data show thatmore phasic responses were noted for the adducting eye. Simulations for the responses of the ad-ducting eyes were obtained with TN = 0.16 s and gN = 0.11. All other coefficients in the modelremained the same as those in A. (C) Simulations of eye velocity in the adducting eye for re-sponses while viewing the near target in Subject 3. The black line (upper trace) is the ideal eyevelocity. The gray line is the average eye velocity response. The dotted line is the simulated eyevelocity based on the model in A. The dashed line is the simulated eye velocity based on themodel in B.

333LASKER et al.: VESTIBULO-OCULAR REFLEX DYNAMICS
± 0.03 s. The principal difference between these parameters is the larger value of TNfor the adducting than for the abducting eye.
We developed a bilateral model to understand the changes in the vergence-depen-dent responses that would be expected in cases of unilateral vestibular hypofunction(FIG. 6). The pathways in this model are similar to those that we used to describechanges in the VOR of the squirrel monkey after unilateral plugging of the threesemicircular canals, unilateral labyrinthectomy, and spectacle-induced adaptation.To model these responses after unilateral vestibular hypofunction, we removed theinput to the left side. One key feature of the model is that the phasic pathway is rap-idly driven into inhibitory cutoff. The dynamics of the far-viewing pathway are sim-ilar to those of regularly discharging afferents, and the dynamics of the near-viewingpathway show an additional component that is similar to the dynamics of irregularlydischarging afferents.
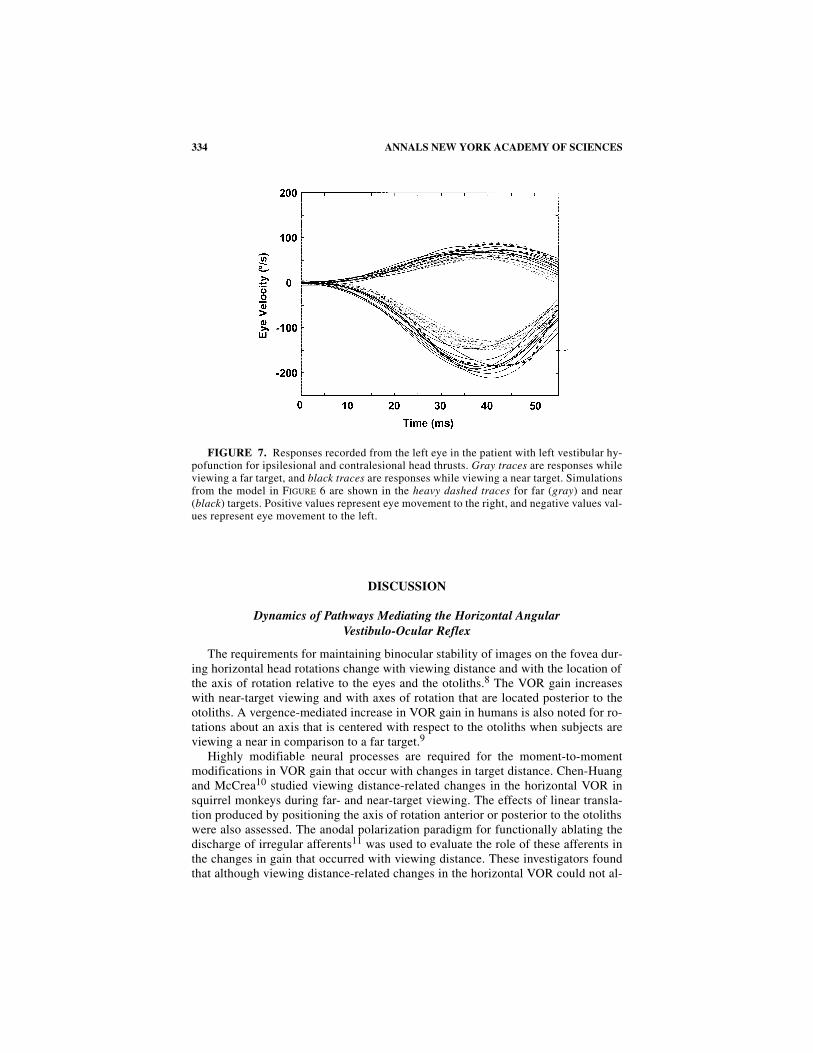
FIGURE 7 shows a comparison of responses for ipsilesional (leftward) and con-tralesional (rightward) head thrusts for near and far viewing in the patient with leftvestibular hypofunction. The eye movements displayed in this figure were recordedfrom the patient’s left eye. Ipsilesional responses did not change with viewing dis-tance. The model predicts this observation (dashed traces, light and dark gray). Thisloss of the increase in gain with near viewing arises because the near-viewing path-way is driven into inhibitory cutoff during the ipsilesional rotations. Contralesionalresponses continue to show vergence-dependent changes in gain.
FIGURE 6. Bilateral model of vergence-mediated changes in VOR. The phasic pathwayhas a lead term that is comparable to the lead noted in the responses of irregularly discharg-ing vestibular-nerve afferents. A cutoff block in this phasic pathway indicates that it makeslittle contribution to inhibitory responses. The following parameters were used in the simu-lations: gF = 0.43, TN = 50 ms, Td1 = 13 ms, and Td2 = 8 ms. For the far-viewing condition,gN = 0.15. For the near-viewing condition, gN = 0.25.
334 ANNALS NEW YORK ACADEMY OF SCIENCES
DISCUSSION
Dynamics of Pathways Mediating the Horizontal AngularVestibulo-Ocular Reflex
The requirements for maintaining binocular stability of images on the fovea dur-ing horizontal head rotations change with viewing distance and with the location ofthe axis of rotation relative to the eyes and the otoliths.8 The VOR gain increaseswith near-target viewing and with axes of rotation that are located posterior to theotoliths. A vergence-mediated increase in VOR gain in humans is also noted for ro-tations about an axis that is centered with respect to the otoliths when subjects areviewing a near in comparison to a far target.9
Highly modifiable neural processes are required for the moment-to-momentmodifications in VOR gain that occur with changes in target distance. Chen-Huangand McCrea10 studied viewing distance-related changes in the horizontal VOR insquirrel monkeys during far- and near-target viewing. The effects of linear transla-tion produced by positioning the axis of rotation anterior or posterior to the otolithswere also assessed. The anodal polarization paradigm for functionally ablating thedischarge of irregular afferents11 was used to evaluate the role of these afferents inthe changes in gain that occurred with viewing distance. These investigators foundthat although viewing distance-related changes in the horizontal VOR could not al-
FIGURE 7. Responses recorded from the left eye in the patient with left vestibular hy-pofunction for ipsilesional and contralesional head thrusts. Gray traces are responses whileviewing a far target, and black traces are responses while viewing a near target. Simulationsfrom the model in FIGURE 6 are shown in the heavy dashed traces for far (gray) and near(black) targets. Positive values represent eye movement to the right, and negative values val-ues represent eye movement to the left.

335LASKER et al.: VESTIBULO-OCULAR REFLEX DYNAMICS
ways be evoked and were often small in amplitude, the galvanic anodal polarizationsreduced the increase in gain associated with near targets by an average of 64%.
The findings of Chen-Huang and McCrea have particular significance in terms ofthe putative pathways for processing of signals arising from regularly and irregularlydischarging afferents. In studies using intrasomatic recordings from central vestibularneurons in squirrel monkeys, it was found that 30% of the monosynaptic excitatorypostsynaptic potential to neurons projecting to the extraocular motor nuclei arise fromirregularly discharging afferents.12,13 In contrast, studies of the VOR using a galvanicpolarization paradigm to selectively and reversibly ablate the discharge of irregularafferents did not result in any change in the horizontal VOR for stimuli at 0.5 Hz, ±60 degrees/s; 4 Hz, ± 10 degrees/s; and steps of acceleration (600 degrees/s2).11 Toexplain the apparent discrepancy between the findings in the intrasomatic recordingexperiments and the functional ablation paradigm used in alert monkeys, it was hy-pothesized that there was a convergence of monosynaptic excitatory and polysynapticinhibitory inputs from irregularly discharging afferents onto the central neurons thatproject to the extraocular motor nuclei.11
Evidence in favor of the notion that there are convergent excitatory and inhibitoryinputs from irregularly discharging afferents onto central vestibular neurons wasprovided by extracellular recordings from central vestibular neurons in alert squirrelmonkeys.14 Galvanic anodal polarization led to an increase in response gain in somehorizontal position-vestibular-pause neurons (PVPs) (the interneurons mainly re-sponsible for signals mediating the horizontal VOR) and to a decrease in responsegain in others. When the entire sample of PVPs was considered together, the excita-tory and inhibitory effects almost completely cancelled one another so that there wasno change in the overall signal from PVPs as a consequence of silencing irregularafferents.
Recent studies of the horizontal angular VOR evoked by high-frequency, high-acceleration head rotations in the squirrel monkey have provided evidence of linearand nonlinear (phasic) pathways mediating the reflex.1 The rotations used in thesestudies were delivered in darkness, and the vergence angle was less than 1.5 meter-angle (reciprocal of the fixation distance), the position of the eyes that is observedwhen the animal is not attending to a specific target. The linear pathway is respon-sible for the relatively constant gain and phase of the VOR with respect to frequencyover the range of 0.5–15 Hz for peak head velocity up to 20 degrees/s. The dynamicsof regularly discharging afferents have been shown to closely match those of the lin-ear pathway.15 The nonlinear pathway is phasic (gain and phase lead that rise withfrequency), is highly modifiable, and has two nonlinear properties: inhibitory cutoffand gain that increases with the cube of velocity for frequencies greater than 2 Hz.Irregularly discharging afferents have the phasic dynamics and inhibitory cutoff thatare noted in the nonlinear pathway. The nonlinear pathway became most importantfor large changes in VOR gain (brought about through processes of vestibular com-pensation or adaptation) during rapid head rotations. The frequencies and velocitiesof the rotational stimuli used in the study of Minor and Goldberg11 would have elic-ited little contributionm from the nonlinear pathway.
The findings in the present study show that there is an increase in the phasic con-tribution with near-target viewing. This increase could be related to the changes thatare seen in the squirrel monkey VOR after compensation to unilateral vestibular in-jury. The increase in VOR gain with near viewing was absent for ipsilesional rota-

336 ANNALS NEW YORK ACADEMY OF SCIENCES
tions in the subject with unilateral vestibular hypofunction. Similar results werenoted for the near-viewing responses to passive, whole-body rotations in subjectswith unilateral vestibular hypofunction.7 An increase in gain for responses to thenear target than to the far target was noted for contralesional rotations. These find-ings suggest that the vergence-mediated increase in gain arises from a rectified oripsilateral pathway that makes a substantial excitatory contribution to the responsebut that is rapidly driven into inhibitory cutoff. The model in FIGure 6 accounts forthese effects.
Vergence-Mediated Changes in the Dynamics of the HumanVestibulo-Ocular Reflex
The four subjects with normal vestibular function in this study showed a robustincrease in VOR gain for responses to head thrusts when viewing near in comparisonwith far targets. The rotational stimuli in these studies were head-on-body rotationsdelivered passively (head thrusts) with the axis of rotation approximately centeredwith respect to the otoliths. The axis of rotation for these stimuli is located behindthe eyes, and compensatory eye movements in this condition are expected to be larg-er for the abducting than for the adducting eye. Instead, we observed comparablegains for both eyes (when measured as peak eye velocity/peak head velocity); how-ever, the response for the adducting eye showed a larger phase lead than did theabducting eye.
The accentuated phasic response for the adducting eye suggests a role for the as-cending tract of Deiter’s (ATD) in the signals that mediate these effects. The ATDprojects ipsilaterally from secondary vestibular neurons receiving inputs from hori-zontal canal afferents to the medial rectus subdivision of the oculomotor nucleus.The sensitivity of ATD neurons to angular and linear head velocity has been shownto increase for fixation of a near target than a far target.16
These findings are provocative, but additional studies are needed to define the or-igin of the effects we have observed. Experiments in which the axis of rotation is var-ied will help to characterize the asymmetries between adducting and abducting eyemovements. Larger changes in VOR gain with respect to viewing distance would beexpected from responses to rotations about axes that are displaced posterior to theotoliths. The eye movements in the VOR for viewing a near target involve differentprofiles of activation for the medial and lateral rectus in each eye. Both medial rectusmuscles are contracted to achieve vergence prior to the head thrust. During the VOR,the response in the abducting eye involves contraction of the lateral rectus with con-comitant relaxation of the medial rectus. By contrast, the response in the adductingeye involves further contraction of the medial rectus and further relaxation of the lat-eral rectus. Experiments in which the vergence angle and eccentricity of the targetare varied are needed to evaluate the role of the dynamics of the oculomotor plant inthese effects.
ACKNOWLEDGMENTS
This work was supported by National Institutes of Health grants R01 DC02390and K23 DC00196.

337LASKER et al.: VESTIBULO-OCULAR REFLEX DYNAMICS
REFERENCES
1. MINOR, L.B., D.M. LASKER, D.D. BACKOUS & T.E. HULLAR. 1999. Horizontal vestibu-loocular reflex evoked by high-acceleration rotations in the squirrel monkey. I. Nor-mal responses. J. Neurophysiol. 82: 1254–1270.
2. LASKER, D.M., D.D. BACKOUS, A. LYSAKOWSKI, et al. 1999. Horizontal vestibuloocularreflex evoked by high-acceleration rotations in the squirrel monkey. II. Responsesafter canal plugging. J. Neurophysiol. 82: 1271–1285.
3. LASKER, D.M., T.E. HULLAR & L.B. MINOR. 2000. Horizontal vestibuloocular reflexevoked by high-acceleration rotations in the squirrel monkey. III. Responses afterlabyrinthectomy. J. Neurophysiol. 83: 2482–2496.
4. CLENDANIEL, R.A., D.M. LASKER & L.B. MINOR. 2001. Horizontal vestibuloocularreflex evoked by high-acceleration rotations in the squirrel monkey. IV. Responsesafter spectacle-induced adaptation. J. Neurophysiol. 86: 1594–1611.
5. MILES, F.A. & B.B. EIGHMY. 1980. Long-term adaptive change in primate vestibulooc-ular reflex. I. Behavioral observations. J. Neurophysiol. 43: 1406–1425.
6. LISBERGER, S.G. & T.A. PAVELKO. 1986. Vestibular signals carried by pathways sub-serving plasticity of the vestibulo-ocular reflex in monkeys. J. Neurosci. 6: 346–354.
7. STRAUMANN, D., D.S. ZEE, D. SOLOMON, et al. 1995. Transient torsion during and aftersaccades. Vision Res. 35(23/24): 3321–3334.
8. VIRRE, E., D. TWEED, K. MILNER & T. VILIS. 1986. A reexamination of the gain of thevestibuloocular reflex. J. Neurophysiol. 56: 439–450.
9. CRANE, B.T. & J.L. DEMER. 1998. Human horizontal vestibulo-ocular reflex initiation:Effects of acceleration, target distance, and unilateral deafferentation. J. Neurophys-iol. 80: 1151–1166.
10. CHEN-HUANG, C. & R.A. MCCREA. 1998. Contribution of vestibular nerve irregularafferents to viewing distance-related changes in the vestibulo-ocular reflex. Exp.Brain Res. 119: 116–130.
11. MINOR, L.B. & J.M. GOLDBERG. 1991. Vestibular-nerve inputs to the vestibulo-ocularreflex: a functional-ablation study in the squirrel monkey. J. Neurosci. 11: 1636–1648.
12. HIGHSTEIN, S.M., J.M. GOLDBERG, A.K. MOSCHOVAKIS & C. FERNANDEZ. 1987. Inputsfrom regularly and irregularly discharging vestibular nerve afferents to secondaryneurons in the vestibular nuclei of the squirrel monkey. II. Correlation with outputpathways of secondary neurons. J. Neurophysiol. 58: 719–738.
13. BOYLE, R., J.M. GOLDBERG & S.M. HIGHSTEIN. 1992. Inputs from regularly and irregu-larly discharging vestibular nerve afferents to secondary neurons in the squirrel mon-key vestibular nuclei. III. Correlation with vestibulospinal and vestibuloocular outputpathways. J. Neurophysiol. 68: 471–484.
14. CHEN-HUANG, C., R.A. MCCREA & J.M. GOLDBERG. 1997. Contributions of regularlyand irregularly discharging vestibular-nerve inputs to the discharge of central vestib-ular neurons in the alert squirrel monkey. Exp. Brain Res. 114: 405–422.
15. HULLAR, T.E. & L.B. MINOR. 1999. High-frequency dynamics of regularly dischargingcanal afferents provide a linear signal for angular vestibuloocular reflexes. J. Neuro-physiol. 82: 2000–2005.
16. CHEN-HUANG, C. & R.A. MCCREA. 1998. Viewing distance related sensory processingin the ascending tract of Deiters vestibulo-ocular reflex pathway. J. Vestib. Res. 8:175–184.
Copyright © 2022 FDOKUMEN




















