
Effects of Close-to-Nature Transformation on Soil Enzyme ...
Upload
independentCategory
view
7download
0
ORIGINAL ARTICLE
Hojeong Kang AElig Sinkyu Kang AElig Dowon Lee
Variations of soil enzyme activities in a temperate forest soil
Received 4 July 2008 Accepted 27 January 2009 Published online 11 March 2009 The Ecological Society of Japan 2009
Abstract Soil enzyme activities (dehydrogenase ureasephosphatase and arylsulfatase) in a temperate forest soilwere determined in relation to landscape position andseasons Overstory of the area is dominated by Quercusmongolica Kalopanax pictus Carpicus cordata and Acerpseudo-sieboldianum The activities were measured inthree patches namely a north-facing backslope a ridgeand a south-facing backslope in autumn and spring over2 years In addition spatially more detailed analysis forphosphatase was conducted before and after litterfallproduction in six patches Dehydrogenase ureasephosphatase and arylsulfatase activities varied 18ndash185 lg INT-formazan g1 h1 454ndash3470 lg NH4
+
g1 h1 09ndash45 mmol pNP g1 h1 and 07ndash26mmol pNP g1 h1 respectively In general higherenzyme activities were found in the northern aspect thanin the southern aspect This variation appears to be re-lated to differences in chemical properties (eg Fe Aland Mg) of soil as well as distribution of leaf litter Twopatterns were discernible in relation to seasonal varia-tions Dehydrogenase and urease exhibited a positivecorrelation with mean air temperature suggesting thattemperature would be a major controlling variable forthose enzymes In contrast higher activities weredetected in autumn for phosphatase and arylsulfataseactivities which appeared to be closely related to litterproduction and distribution Overall results of this study
indicate that soil enzyme activities in a forest floor areinfluenced by several variables such as temperaturenutrient availability and input of leaf litter which areclosely related to landscape position
Keywords Decomposition AElig Forest floor AElig Extracellularenzyme AElig Seasonal variation AElig Landscape ecology
Introduction
Decomposition and mineralization of organic matter arecritical processes in nutrient cycling in forest ecosystems(Aber and Mellilo 1991) It has been widely acknowl-edged that decomposition and mineralization processesin forest soils exhibit spatial variations For exampleMudrick et al (1994) have reported that higher litterdecomposition rates were found in a northern aspectthan in a southern aspect in a hardwood forest Sucheffects of landscape position could be attributed to dif-ferences in micro-climate (eg temperature and mois-ture) types of vegetation and movement of materials bygravity (eg water and nutrient) Several studies haveaddressed this issue by comparing soil properties ornutrient dynamics in relation to topographic variationssuch as position in a soil catena or aspects of slopes(Schimel et al 1985 Garten Jr et al 1994 Mudrick et al1994) General conclusions of these studies were thatlower areas (ie footslopes or valley floors) exhibitedhigher nutrient contents or microbial activities whichwere accompanied by the long-term changes insoil structures (eg organic matter content soil texturesoil horizon etc) Eviner et al (2006) reported that soilmicrobial P was more strongly affected by slope positionthan by plant species It is also reported that slopeposition is an important determinant for soil nutrientand microbial decomposition (Noorbakhsh et al 2008Sariyildiz et al 2005) However those findings wereobserved at a fairly large scale and hence variousenvironmental conditions may co-vary Further previ-ous studies focused on sediment and water transport
H Kang (amp)School of Civil and Environmental EngineeringYonsei University Seoul KoreaE-mail hj_kangyonseiackrTel +82-2-21235803Fax +82-2-3645300
S KangDepartment of Environmental SciencesKangwon National University Choonchun Korea
D LeeGraduate School of Environmental StudiesSeoul National University Seoul Korea
Ecol Res (2009) 24 1137ndash1143DOI 101007s11284-009-0594-5
along the slope and little information is available aboutlitter redistribution by wind in relation to microbialactivities
In addition to the factors mentioned above wehypothesize that litter redistribution may be anotherimportant factor for microbial activities in temperateforest soils Fresh leaf litter is rich in high-quality carbonsources for microbes compared with woody litter or soilorganic matter Further it was reported that enzymesleached from leaf litter falling on the soil surface con-tributed to the winter peak of annual variation inphosphatase activity (Harrison and Pearce 1979) Assuch leaf litter falling and redistribution by wind andretention by understory vegetation may affect organicmatter decomposition and consequent mineralizationAt our study site northerly wind is dominant over theyear and the understory of south-facing aspect isdominated by Sasa borealis both of which are expectedto affect litter redistribution substantially (Lee et al1999 Yoo et al 2001) Yoo et al (2001) employed colorpapers as a surrogate for leaf litter and estimated themovement of leaf litter on the site Color papers placedon the southern aspect moved 19ndash32 m on averagemostly northerly In contrast those on the northernaspect moved only 4ndash6 m This result suggested that leaflitter would be redistributed from southern aspects tonorthern aspects In particular rugged mountainousareas would exhibit more notable influences by land-scape positions since the factors mentioned above varysubstantially compared with flat landscapes
Microbes in soils play a pivotal role in the decom-position and mineralization by producing variousenzymes (Burns 1982) These enzymes convert largemolecules into low molecular moieties which then canbe assimilated by microorganisms or plants This enzy-matic process is generally regarded as a rate-limiting stepin the decomposition and nutrient cycling (Chrost 1991)Sinsabaugh et al (1991) have demonstrated that enzy-mic analysis could be a valuable tool in a study of litterdecomposition in forest floors It has also been reportedthat enzymic analysis is closely related to biogeochem-istry of wetland soils (Freeman et al 1996) Soil enzymeactivities were also extensively investigated with refer-ence to litter decomposition in agricultural soils (Dillyet al 2007) and a black alder forest (Dilly et al 2001Dilly and Munch 1996) These studies have noted theimportance of soil enzyme activities in relation to litterdecomposition in various ecosystems
Extracellular enzymes can be stabilized by clays andhumus in soil and are involved in decomposition pro-cesses under the influence of edaphic factors (in sensuBurns 1982) Temperature pH nutrient supply andvarious metals have been reported as some of the con-trolling factors of the enzyme activities in upland soils(Sinsabaugh et al 1991)
Considering the substantial differences in environ-mental variables among different landscape positions ina forest floor and between different seasons it isanticipated that soil enzyme activities would vary
according to the variations of those possible controllingvariables In the present study we aimed (1) to deter-mine variations of soil enzyme activities in relation tolandscape position and seasons at a small scale and(2) to elucidate the possible mechanisms of the varia-tions pertinent to litter redistribution in particular Toaddress these issues we conducted both seasonal mon-itoring (spring and autumn) in three patches over2 years and spatially intensive measurement (six pat-ches) beforeafter litter fall production of soil microbialactivities in a temperate forest soil We investigatedurease phosphatase and arylsulfatase activities whichare believed to play important roles in nitrogen phos-phorus and sulphur cycles respectively (Burns 1982)Dehydrogenase activity was also determined as a generalindex for microbial activities To assess chemical prop-erties at the sites we measured metal ions (Fe Al Mg)extractable NH4
+ content in the soils
Materials and methods
Study site
This study was conducted in a temperate mixed hard-wood forest floor in Mt Jumbong (1008 m asl) whichis located in the mid-east of Korea (N 3802cent E 12826cent)The mean monthly temperature varies between 44 and163C and the mean annual precipitation is 1579 mmOverstory of the area is dominated by Quercus mongo-lica Kalopanax pictus Carpicus cordata and Acerpseudo-sieboldianum The area was divided into six pat-ches by landscape positions and understory vegetationnorth-facing footslope (NF) north-facing backslope(NB) ridge (R) south-facing backslope (SB) south-facing backslope with S borealis (SS) and south-facingfootslope (SF)
The total study area is ca 104 m2 of which physicaland geological conditions are similar (ie same bed rockand average rainfall) This allowed us to compare theenzyme activities and chemical properties of the patchesin relation to landscape positions without interferencefrom other factors The soil type was loamy sand(organic carbon content 46 CEC 1225 meq100 g1 pH 45 soil water content 04ndash06 g H2O gsoil1) and no significant differences in soil organiccarbon content CEC and pH were found among soilsamples from the patches Leaf litter production was330ndash360 g m2 year1 and the leaf thickness variedbetween 3 and 15 cm
Samplings
From each patch three soil cores (10 cm diameter and10 cm depth) were collected with a soil corer Thesampling procedures were undertaken in three patches(north-facing backslope ridge and south-facing backs-lope) for eight occasions over 3 years we collected
1138
samples in October and November (Autumn-1) in 1995May and June (Spring-1) September and November in1996 (Autumn-2) In 1997 samples were collected inApril and June (Spring-2) In addition two spatiallyintensive samplings were conducted in all the six patchesin August (before litter fall) and September (after litterfall) 1996 Upon arriving at the laboratory the sampleswere sieved (lt2 mm) and maintained at 4C until theanalyses were conducted
Enzyme assays
Dehydrogenase activity was determined by adding INT(iodonitrozo tetrazolium chloride) as a model substrateand measuring reduction of the chemical spectrophoto-metrically (Trevors et al 1982) Phosphatase and aryl-sulfatase activities were assayed by measuring releasedp-nitrophenol over an hour of incubation from soilsadded with p-nitrophenyl phosphate and p-nitrophenylsulfate as substrates respectively (Eivazi and Tabatabai1990 Tabatabai and Bremner 1970) Urease activity wasdetermined by measuring released NH4
+ from the soilamended with urea described by Kandeler and Gerber(1988) In each assay 2 g of soils was used and at leasttriplicate samples were employed for each location Allthe enzyme activities were presented per dry mass of soilper incubation duration
Chemical analysis
Extractable ammonium concentration in the soil wasdetermined by indophenol method (Emteryd 1989)followed by extraction with 2 M KCl Metal ions weremeasured by an ICP-AES (Perkin-Elmer Plasma 40)after digesting crushed soils (500 lmlt) in mixed acidsolution (HNO3 HF HClO4 = 4 4 1) for 12 h
Statistical analysis
The enzyme activities and NH4+ contents were analyzed
according to a repeated measures ANOVA Differencesin enzyme activities in six patches were determined by aone-way ANOVA followed by Tukeyrsquos test All pro-cesses were conducted on SPSS version 150
Results
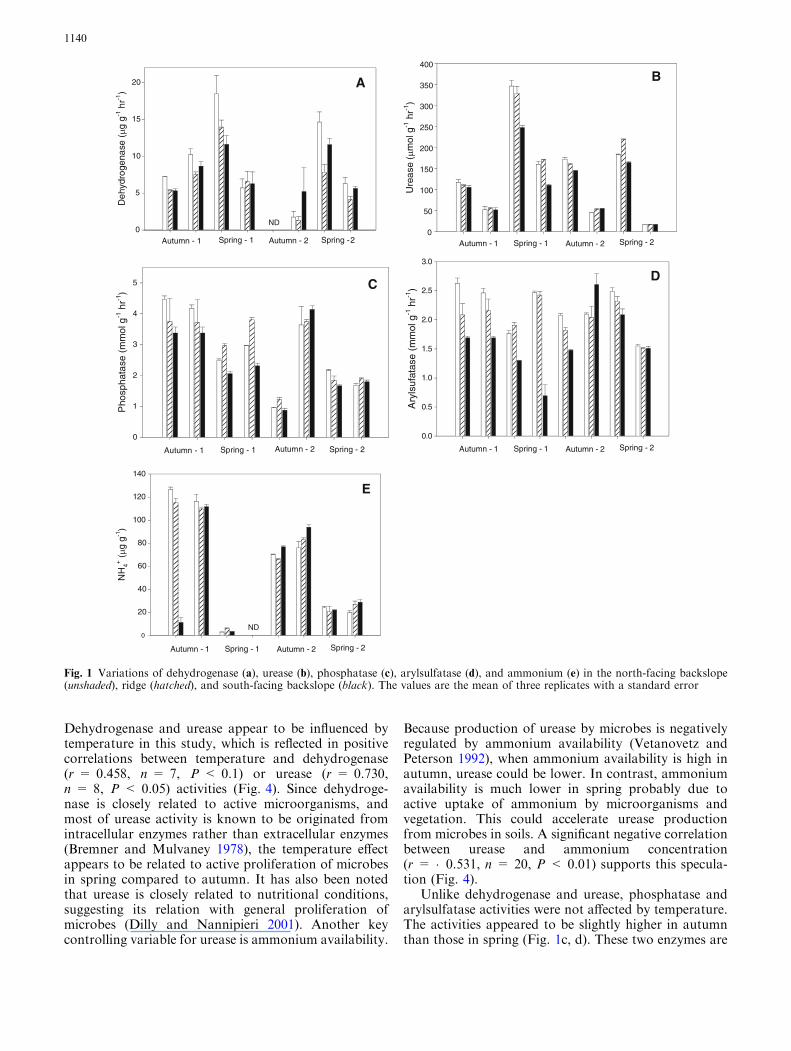
All enzyme activities showed significant variations bysampling time landscape position and the interactionsof the two (Table 1) Overall the enzyme activities werehigher on the north-facing backslope than in the south-facing backslope throughout the studying period withfew exceptions (eg dehydrogenase in autumn-2 andarylsulfatase in autumn-2) (Fig 1)
For the seasonal variations two patterns were dis-cernable Dehydrogenase and urease activities werehigher in spring than those in autumn (Fig 1a b) Incontrast higher activity of phosphatase was generallyobserved in autumn than in spring (Fig 1c) Arylsulfa-tase showed a similar temporal pattern as phosphatasebut it was less discernable (Fig 1d) Extractableammonium concentrations were higher in autumn thanin spring (Fig 1e) However there were no generalpatterns of differences among the three plots
Overall phosphatase and urease activities were lowerbefore the litterfall production (BF) than those after thelitterfall production (AF) (Fig 2) Phosphatase activitiesranged between 15 and 42 mmol g1 h1 after the lit-terfall production while the same enzyme activity ran-ged between 16 and 22 mmol g1 h1 before thelitterfall production Similar trends were observed forurease activity 28ndash43 NH4
+ lg g1 h1 (AF) and22ndash28 lg g1 min1 (BF) No spatial differences werediscernible before the litterfall production (Fig 2)However significantly different enzyme activities werefound among patches after the litterfall production insuch a way that enzyme activities were the highest innorth-facing footslope and the lowest in south-facingbackslope without S borealis (Fig 2) In contrast theactivities in south-facing backslope with S borealsiswere not different from north-facing backslope ridge orsouth-facing footslope (Fig 2)
Al Fe and Mg contents were lower on the north-facing slope than those on the south-facing slope(Fig 3) In contrast Ca content was higher on thenorth-facing slope than that on the south-facing slope
Discussion
Several environmental variables are known to affect soilenzyme activities Temperature can influence enzymeactivity by direct modification of enzyme kinetics aswell as indirect influence on microbial proliferation
Table 1 A repeated-measure ANOVA table for the enzyme activ-ities and ammonium contents
Dependent Independent df F value P value
Dehydrogenase Position 2 534 0000Months 7 440 0000Position middot months 14 128 0003
Urease Position 2 660 0000Months 7 479 0000Position middot months 14 177 0000
Phosphatase Position 2 634 0000Months 7 6214 0000Position middot months 14 143 0000
Arylsulfatase Position 2 62 0004Months 7 523 0000Position middot months 14 21 0032
Ammonium Position 2 71 0002Months 7 12084 0000Position middot months 14 65 0000
1139
Dehydrogenase and urease appear to be influenced bytemperature in this study which is reflected in positivecorrelations between temperature and dehydrogenase(r = 0458 n = 7 P lt 01) or urease (r = 0730n = 8 P lt 005) activities (Fig 4) Since dehydroge-nase is closely related to active microorganisms andmost of urease activity is known to be originated fromintracellular enzymes rather than extracellular enzymes(Bremner and Mulvaney 1978) the temperature effectappears to be related to active proliferation of microbesin spring compared to autumn It has also been notedthat urease is closely related to nutritional conditionssuggesting its relation with general proliferation ofmicrobes (Dilly and Nannipieri 2001) Another keycontrolling variable for urease is ammonium availability
Because production of urease by microbes is negativelyregulated by ammonium availability (Vetanovetz andPeterson 1992) when ammonium availability is high inautumn urease could be lower In contrast ammoniumavailability is much lower in spring probably due toactive uptake of ammonium by microorganisms andvegetation This could accelerate urease productionfrom microbes in soils A significant negative correlationbetween urease and ammonium concentration(r = 0531 n = 20 P lt 001) supports this specula-tion (Fig 4)
Unlike dehydrogenase and urease phosphatase andarylsulfatase activities were not affected by temperatureThe activities appeared to be slightly higher in autumnthan those in spring (Fig 1c d) These two enzymes are
Deh
ydro
gena
se(
gg-1
hr-1
)
0
5
10
15
20 A
ND
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Ure
ase
(m
olg-1
hr-1
)
0
50
100
150
200
250
300
350
400
B
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Pho
spha
tase
(mm
olg-1
hr-1
)
0
1
2
3
4
5 C
Autumn - 1 Spring - 2Autumn - 2Spring - 1
Ary
lsuf
atas
e(m
mol
g-1hr
-1)
00
05
10
15
20
25
30
D
NH
4+(
gg-1
)
0
20
40
60
80
100
120
140
ND
E
Fig 1 Variations of dehydrogenase (a) urease (b) phosphatase (c) arylsulfatase (d) and ammonium (e) in the north-facing backslope(unshaded) ridge (hatched) and south-facing backslope (black) The values are the mean of three replicates with a standard error
1140
known to be mainly extracellular and stabilized andhence less affiliated with microbial biomass compared todehydrogenase or urease Further it was reported thatenzymes leached from leaf litter falling on the soil sur-face contributed winter peak of the annual variation ofphosphatase activity (Harrison and Pearce 1979) Like-wise higher activities of phosphatase and phenol oxi-dase were found in riparian peatlands in late autumnwhich was attributed to litter quality and quantity(Bonnett et al 2006) Sinsabaugh et al (1991) has alsoreported that maximum enzyme activities were detectedin several weeks after the production of litter As suchlitter production and distribution may offset effects fromother controlling variables such as temperature Previ-
ous study has shown that the site is dominated bynortherly wind and consequent movement of leaf littertoward northern slope (Lee et al 1999 Yoo et al 2001)This contributes the general trends of higher enzymeactivities in northern aspect than in southern aspect inthis study The contribution of leaf litter to enzymeactivity is further supported by the finding that spa-tial differences in enzyme activities emerged only after
Landscape positions
N-Foot N-Back Ridge S-Back S-Sasa S-Foot
Pho
spha
tase
(mm
ole
g-1hr
-1)
0
1
2
3
4
5 a
b
aa a
b b
bb
b
ab
c
Fig 2 Variations of phosphatase activities before the litterfallproduction (unshaded) and after litterfall production (black) Thevalues are mean of three replicates with a standard error Bars withdifferent letters are significantly different among the same enzymesat P lt 005
Ions
Al Fe Ca K Mg
Con
tent
s(m
gg-1
)
0
2
4
6
8
10
a
a
a
a
a
c
b
b
c
bc
ba
a b
Fig 3 Metal ion concentrations in the north-facing backslope(unshaded) ridge (hatched) and south-facing backslope (black) Thevalues are mean of three replicates with a standard error Thevalues are significantly different for each ion at P lt 001 exceptAl which is different at P lt 005
Temperature ( C)
0 5 10 15 20 25
Deh
ydro
gena
se(
gg-1
hr-1
)
0
4
8
12
16
A
r = 0458 n = 7 P lt 01
Temperature ( C)
0 5 10 15 20 25
Ure
ase
(m
olg-1
hr-1
)
0
50
100
150
200
250
300
350
B
r = 0730 n = 8 P lt 005
Urease activity ( mol g-1 hr-1)
0 100 200 300 400
Am
mon
ium
(g
g-1)
0
20
40
60
80
100
120
140
r = -0531 P lt 001 n = 20
C
Fig 4 Relationships between dehydrogenase and temperature (a)urease and temperature (b) and urease and ammonium (c)
1141
litterfall production (Fig 2) It is also interesting to notethat differences in enzyme activities between south-facing backslope without S borealis and south-facingbackslope with S borealis were only found after thelitterfall while such differences were absent beforethe litterfall production (Fig 2) Because areas withS borealis can maintain much larger amount of leaflitter than areas without S borealis understory vegeta-tion may influence biogeochemical processes in forestfloor through leaf litter distribution (Lee et al 1999)
Metal ions notably Fe and Al have reported toinhibit soil enzyme activities by absorbing to the activesite (Ekenler and Tabatabai 2002) In contrast Ca2+
and Mg2+ content often exhibited a positive correlationwith enzyme activities because those are known to re-move inhibition of humic materials by enzymendashhumicinteractions (Freeman et al 1996) Our results exhibitedthat two inhibitory ions (Al Fe) were higher whilst Mgand K contents were lower in soil of the southern-facingslope (Oh et al 1999) As such higher concentration ofinhibitory ions and lower concentration of activationions partially account for the lower enzyme activities inthe southern-facing slope
In this study we assessed the changes in enzymeactivities only However previous studies have revealedthat microbial composition such as bacteria versus fungimay vary over time (Moore-Kucera and Dick 2008Kubartova et al 2007) This would be closely related toenzyme activities because different types of microor-ganisms are responsible for the production of specificenzymes
Landscape position has been reported to affect soilproperties litter decomposition nitrogen dynamics andphosphorus availability (Brubaker et al 1994 Klem-mendson and Wienhold 1991 Mudrick et al 1994Garten Jr et al 1994) These studies focused on thedifferences in organic matter water and nutrient con-tents which were caused by flow of water and soil ero-sions In addition to these factors our results indicatedthat production and redistribution of leaf litter are otherkey factors affecting biogeochemical cycles in a forestfloor Furthermore not only the physical movement ofleaf litter mediated by wind but also understory vege-tation which may impede the movement could play animportant role in this dynamics
Acknowledgments H Kang is grateful to AEBRC EcoRiver21 forfinancial support
References
Aber JD Mellilo JM (1991) Terrestrial ecosystems SaundersCollege Publishing Philadelphia
Bonnett SAF Ostle N Freeman C (2006) Seasonal variations indecomposition processes in a valley-bottom riparian peatlandSci Total Environ 370561ndash573 doi101016jscitotenv200608032
Bremner JM Mulvaney RL (1978) Urease activity in soils InBurns RG (ed) Soil enzymes Academic Press New York
Brubaker SC Jones AJ Frank K Lewis DT (1994) Regressionmodels for estimating soil properties by landscape position SoilSci Soc Am J 581763ndash1767
Burns RG (1982) Enzyme activity in soil location and a possiblerole in microbial ecology Soil Biol Biochem 14423ndash427 doi1010160038-0717(82)90099-2
Chrost RJ (1991) Environmental control of the synthesis andactivity of aquatic microbial ectoenzymes In Chrost RJ (ed)Microbial enzymes in aquatic environments Springer BerlinHeidelberg New York pp 29ndash59
Dilly O Munch JC (1996) Microbial biomass content basal res-piration and enzyme activities during the course of decompo-sition of leaf litter in a black alder (Alnus glutinosa (L) Gaertn)forest Soil Biol Biochem 281073ndash1081 doi1010160038-0717(96)00075-2
Dilly O Nannipieri P (2001) Response of ATP content respirationrate and enzyme activities in an arable and a forest soil tonutrient addition Biol Fertil Soils 3464ndash72 doi101007s003740100375
Dilly O Bartsch S Rosenbrock P Buscot F Munch J-C (2001)Shifts in physiological capabilities of the microbiota during thedecomposition of leaf litter in a black alder (Alnus glutinosa(Gaertn) L) forest Soil Biol Biochem 33921ndash930 doi101016S0038-0717(00)00239-X
Dilly O Munch J-C Pfeiffer E-M (2007) Enzyme activities andlitter decomposition in agricultural soils in northern centraland southern Germany J Plant Nutr Soil Sci 170197ndash204 doi101002jpln200622044
Eivazi F Tabatabai MA (1990) Factors affecting glucosidase andgalactosidase activities in soils Soil Biol Biochem 22891ndash897doi1010160038-0717(90)90126-K
Ekenler M Tabatabai MA (2002) Effects of trace elements on b-glucosaminidase activity in soils Soil Biol Biochem 341829ndash1832 doi101016S0038-0717(02)00167-0
Emteryd O (1989) Chemical and physical analysis of inorganicnutrients in plant soil water and air Department of Forest SiteResearch Swedish University of Agricultural Sciences Umea
Eviner VT Chapin FS Vaughn CE (2006) Seasonal variations inplant species effects on soil N and P dynamics Ecology 87974ndash986 doi1018900012-9658(2006)87[974SVIPSE]20CO2
Freeman C Liska G Ostle NJ Lock MA Reynolds B Hudson J(1996) Microbial activity and enzymic decomposition processesfollowing peatland water table drawdown Plant Soil 180121ndash127 doi101007BF00015418
Garten CT Jr Huston MA Thoms CA (1994) Topographic vari-ation of soil nitrogen dynamics at Walker Branch watershedTennessee For Sci 40497ndash512
Harrison AF Pearce T (1979) Seasonal variation of phosphataseactivity in woodland soils Soil Biol Biochem 11405ndash410 doi1010160038-0717(79)90054-3
Kandeler E Gerber H (1988) Short-term assay of soil ureaseactivity using colorimetric determination of ammonium BiolFertil Soils 658ndash72 doi101007BF00257924
Klemmendson JO Wienhold BJ (1991) Aspect and species influ-ences on nitrogen and phosphorus availability in Arizonachaparral soils Soil Sci Soc Am J 551735ndash1740
Kubartova A Moukoumi J Beguiristain T Ranger J Berthelin J(2007) Microbial diversity during cellulose decomposition indifferent forest stands I Microbial communities and environ-mental conditions Microb Ecol 54393ndash405 doi101007s00248-007-9286-2
Lee D Yoo G Oh S Shim JH Kang S (1999) Significance ofaspect and understory type to leaf litter redistribution in atemperate hardwood forest Korean J Biol Sci 3143ndash147
Moore-Kucera J Dick RP (2008) PLFA profiling of microbialcommunity structure and seasonal shifts in soils of a Douglas-fir chronosequence Microb Ecol 55500ndash511 doi101007s00248-007-9295-1
Mudrick DA Hoosein M Hicks RR Jr Townsend EC (1994)Decomposition of leaf litter in an Appalachian forest effects ofleaf species aspect slope position and time For Ecol Manage68231ndash250
1142
Noorbakhsh S Schoenau J Si B Zeleke T Qian P (2008) Soilproperties yield and landscape relationships in south-centralSaskatchewan Canada J Plant Nutr 31539ndash556 doi10108001904160801895076
Oh S Yoo G Shim JH Kang S Lee D (1999) Spatial heterogeneityof soil K Ca Mg Fe and Al generated by leaf litter redistri-bution in a temperate hardwood forest Kor J Ecol 22205ndash210
Sariyildiz T Anderson JM Kucuk M (2005) Effects of tree speciesand topography on soil chemistry litter quality and decom-position in northeast Turkey Soil Biol Biochem 371695ndash1706doi101016jsoilbio200502004
Schimel D Stillwell MA Woodmansee RG (1985) Biogeochemis-try of C N and P in a soil catena of the shortgrass steppeEcology 66276ndash282 doi1023071941328
Sinsabaugh RL Antibus RK Linkins AE (1991) An enzymicapproach to the analysis of microbial activity during plant litterdecomposition Agric Ecosyst Environ 3443ndash54 doi1010160167-8809(91)90092-C
Tabatabai MA Bremner JM (1970) Arylsulfatase activity of soilsSoil Sci Soc Am Proc 34225ndash229
Trevors JT Mayfield CI Inniss WE (1982) Measurement of elec-tron transport system (ETS) activity in soil Microb Ecol 8163ndash168 doi101007BF02010449
Vetanovetz RP Peterson JC (1992) Effect of carbon source andnitrogen on urease activity in a Sphagnum peat mediumCommun Soil Sci Plant Anal 23379ndash388 doi10108000103629209368596
Yoo G Park E-J Kim S-H Lee H-J Kang S Lee D (2001)Transport and decomposition of leaf litter as affected by aspectand understory in a temperate hardwood forest Korean J BiolSci 5319ndash325
1143

along the slope and little information is available aboutlitter redistribution by wind in relation to microbialactivities
In addition to the factors mentioned above wehypothesize that litter redistribution may be anotherimportant factor for microbial activities in temperateforest soils Fresh leaf litter is rich in high-quality carbonsources for microbes compared with woody litter or soilorganic matter Further it was reported that enzymesleached from leaf litter falling on the soil surface con-tributed to the winter peak of annual variation inphosphatase activity (Harrison and Pearce 1979) Assuch leaf litter falling and redistribution by wind andretention by understory vegetation may affect organicmatter decomposition and consequent mineralizationAt our study site northerly wind is dominant over theyear and the understory of south-facing aspect isdominated by Sasa borealis both of which are expectedto affect litter redistribution substantially (Lee et al1999 Yoo et al 2001) Yoo et al (2001) employed colorpapers as a surrogate for leaf litter and estimated themovement of leaf litter on the site Color papers placedon the southern aspect moved 19ndash32 m on averagemostly northerly In contrast those on the northernaspect moved only 4ndash6 m This result suggested that leaflitter would be redistributed from southern aspects tonorthern aspects In particular rugged mountainousareas would exhibit more notable influences by land-scape positions since the factors mentioned above varysubstantially compared with flat landscapes
Microbes in soils play a pivotal role in the decom-position and mineralization by producing variousenzymes (Burns 1982) These enzymes convert largemolecules into low molecular moieties which then canbe assimilated by microorganisms or plants This enzy-matic process is generally regarded as a rate-limiting stepin the decomposition and nutrient cycling (Chrost 1991)Sinsabaugh et al (1991) have demonstrated that enzy-mic analysis could be a valuable tool in a study of litterdecomposition in forest floors It has also been reportedthat enzymic analysis is closely related to biogeochem-istry of wetland soils (Freeman et al 1996) Soil enzymeactivities were also extensively investigated with refer-ence to litter decomposition in agricultural soils (Dillyet al 2007) and a black alder forest (Dilly et al 2001Dilly and Munch 1996) These studies have noted theimportance of soil enzyme activities in relation to litterdecomposition in various ecosystems
Extracellular enzymes can be stabilized by clays andhumus in soil and are involved in decomposition pro-cesses under the influence of edaphic factors (in sensuBurns 1982) Temperature pH nutrient supply andvarious metals have been reported as some of the con-trolling factors of the enzyme activities in upland soils(Sinsabaugh et al 1991)
Considering the substantial differences in environ-mental variables among different landscape positions ina forest floor and between different seasons it isanticipated that soil enzyme activities would vary
according to the variations of those possible controllingvariables In the present study we aimed (1) to deter-mine variations of soil enzyme activities in relation tolandscape position and seasons at a small scale and(2) to elucidate the possible mechanisms of the varia-tions pertinent to litter redistribution in particular Toaddress these issues we conducted both seasonal mon-itoring (spring and autumn) in three patches over2 years and spatially intensive measurement (six pat-ches) beforeafter litter fall production of soil microbialactivities in a temperate forest soil We investigatedurease phosphatase and arylsulfatase activities whichare believed to play important roles in nitrogen phos-phorus and sulphur cycles respectively (Burns 1982)Dehydrogenase activity was also determined as a generalindex for microbial activities To assess chemical prop-erties at the sites we measured metal ions (Fe Al Mg)extractable NH4
+ content in the soils
Materials and methods
Study site
This study was conducted in a temperate mixed hard-wood forest floor in Mt Jumbong (1008 m asl) whichis located in the mid-east of Korea (N 3802cent E 12826cent)The mean monthly temperature varies between 44 and163C and the mean annual precipitation is 1579 mmOverstory of the area is dominated by Quercus mongo-lica Kalopanax pictus Carpicus cordata and Acerpseudo-sieboldianum The area was divided into six pat-ches by landscape positions and understory vegetationnorth-facing footslope (NF) north-facing backslope(NB) ridge (R) south-facing backslope (SB) south-facing backslope with S borealis (SS) and south-facingfootslope (SF)
The total study area is ca 104 m2 of which physicaland geological conditions are similar (ie same bed rockand average rainfall) This allowed us to compare theenzyme activities and chemical properties of the patchesin relation to landscape positions without interferencefrom other factors The soil type was loamy sand(organic carbon content 46 CEC 1225 meq100 g1 pH 45 soil water content 04ndash06 g H2O gsoil1) and no significant differences in soil organiccarbon content CEC and pH were found among soilsamples from the patches Leaf litter production was330ndash360 g m2 year1 and the leaf thickness variedbetween 3 and 15 cm
Samplings
From each patch three soil cores (10 cm diameter and10 cm depth) were collected with a soil corer Thesampling procedures were undertaken in three patches(north-facing backslope ridge and south-facing backs-lope) for eight occasions over 3 years we collected
1138
samples in October and November (Autumn-1) in 1995May and June (Spring-1) September and November in1996 (Autumn-2) In 1997 samples were collected inApril and June (Spring-2) In addition two spatiallyintensive samplings were conducted in all the six patchesin August (before litter fall) and September (after litterfall) 1996 Upon arriving at the laboratory the sampleswere sieved (lt2 mm) and maintained at 4C until theanalyses were conducted
Enzyme assays
Dehydrogenase activity was determined by adding INT(iodonitrozo tetrazolium chloride) as a model substrateand measuring reduction of the chemical spectrophoto-metrically (Trevors et al 1982) Phosphatase and aryl-sulfatase activities were assayed by measuring releasedp-nitrophenol over an hour of incubation from soilsadded with p-nitrophenyl phosphate and p-nitrophenylsulfate as substrates respectively (Eivazi and Tabatabai1990 Tabatabai and Bremner 1970) Urease activity wasdetermined by measuring released NH4
+ from the soilamended with urea described by Kandeler and Gerber(1988) In each assay 2 g of soils was used and at leasttriplicate samples were employed for each location Allthe enzyme activities were presented per dry mass of soilper incubation duration
Chemical analysis
Extractable ammonium concentration in the soil wasdetermined by indophenol method (Emteryd 1989)followed by extraction with 2 M KCl Metal ions weremeasured by an ICP-AES (Perkin-Elmer Plasma 40)after digesting crushed soils (500 lmlt) in mixed acidsolution (HNO3 HF HClO4 = 4 4 1) for 12 h
Statistical analysis
The enzyme activities and NH4+ contents were analyzed
according to a repeated measures ANOVA Differencesin enzyme activities in six patches were determined by aone-way ANOVA followed by Tukeyrsquos test All pro-cesses were conducted on SPSS version 150
Results
All enzyme activities showed significant variations bysampling time landscape position and the interactionsof the two (Table 1) Overall the enzyme activities werehigher on the north-facing backslope than in the south-facing backslope throughout the studying period withfew exceptions (eg dehydrogenase in autumn-2 andarylsulfatase in autumn-2) (Fig 1)
For the seasonal variations two patterns were dis-cernable Dehydrogenase and urease activities werehigher in spring than those in autumn (Fig 1a b) Incontrast higher activity of phosphatase was generallyobserved in autumn than in spring (Fig 1c) Arylsulfa-tase showed a similar temporal pattern as phosphatasebut it was less discernable (Fig 1d) Extractableammonium concentrations were higher in autumn thanin spring (Fig 1e) However there were no generalpatterns of differences among the three plots
Overall phosphatase and urease activities were lowerbefore the litterfall production (BF) than those after thelitterfall production (AF) (Fig 2) Phosphatase activitiesranged between 15 and 42 mmol g1 h1 after the lit-terfall production while the same enzyme activity ran-ged between 16 and 22 mmol g1 h1 before thelitterfall production Similar trends were observed forurease activity 28ndash43 NH4
+ lg g1 h1 (AF) and22ndash28 lg g1 min1 (BF) No spatial differences werediscernible before the litterfall production (Fig 2)However significantly different enzyme activities werefound among patches after the litterfall production insuch a way that enzyme activities were the highest innorth-facing footslope and the lowest in south-facingbackslope without S borealis (Fig 2) In contrast theactivities in south-facing backslope with S borealsiswere not different from north-facing backslope ridge orsouth-facing footslope (Fig 2)
Al Fe and Mg contents were lower on the north-facing slope than those on the south-facing slope(Fig 3) In contrast Ca content was higher on thenorth-facing slope than that on the south-facing slope
Discussion
Several environmental variables are known to affect soilenzyme activities Temperature can influence enzymeactivity by direct modification of enzyme kinetics aswell as indirect influence on microbial proliferation
Table 1 A repeated-measure ANOVA table for the enzyme activ-ities and ammonium contents
Dependent Independent df F value P value
Dehydrogenase Position 2 534 0000Months 7 440 0000Position middot months 14 128 0003
Urease Position 2 660 0000Months 7 479 0000Position middot months 14 177 0000
Phosphatase Position 2 634 0000Months 7 6214 0000Position middot months 14 143 0000
Arylsulfatase Position 2 62 0004Months 7 523 0000Position middot months 14 21 0032
Ammonium Position 2 71 0002Months 7 12084 0000Position middot months 14 65 0000
1139
Dehydrogenase and urease appear to be influenced bytemperature in this study which is reflected in positivecorrelations between temperature and dehydrogenase(r = 0458 n = 7 P lt 01) or urease (r = 0730n = 8 P lt 005) activities (Fig 4) Since dehydroge-nase is closely related to active microorganisms andmost of urease activity is known to be originated fromintracellular enzymes rather than extracellular enzymes(Bremner and Mulvaney 1978) the temperature effectappears to be related to active proliferation of microbesin spring compared to autumn It has also been notedthat urease is closely related to nutritional conditionssuggesting its relation with general proliferation ofmicrobes (Dilly and Nannipieri 2001) Another keycontrolling variable for urease is ammonium availability
Because production of urease by microbes is negativelyregulated by ammonium availability (Vetanovetz andPeterson 1992) when ammonium availability is high inautumn urease could be lower In contrast ammoniumavailability is much lower in spring probably due toactive uptake of ammonium by microorganisms andvegetation This could accelerate urease productionfrom microbes in soils A significant negative correlationbetween urease and ammonium concentration(r = 0531 n = 20 P lt 001) supports this specula-tion (Fig 4)
Unlike dehydrogenase and urease phosphatase andarylsulfatase activities were not affected by temperatureThe activities appeared to be slightly higher in autumnthan those in spring (Fig 1c d) These two enzymes are
Deh
ydro
gena
se(
gg-1
hr-1
)
0
5
10
15
20 A
ND
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Ure
ase
(m
olg-1
hr-1
)
0
50
100
150
200
250
300
350
400
B
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Pho
spha
tase
(mm
olg-1
hr-1
)
0
1
2
3
4
5 C
Autumn - 1 Spring - 2Autumn - 2Spring - 1
Ary
lsuf
atas
e(m
mol
g-1hr
-1)
00
05
10
15
20
25
30
D
NH
4+(
gg-1
)
0
20
40
60
80
100
120
140
ND
E
Fig 1 Variations of dehydrogenase (a) urease (b) phosphatase (c) arylsulfatase (d) and ammonium (e) in the north-facing backslope(unshaded) ridge (hatched) and south-facing backslope (black) The values are the mean of three replicates with a standard error
1140
known to be mainly extracellular and stabilized andhence less affiliated with microbial biomass compared todehydrogenase or urease Further it was reported thatenzymes leached from leaf litter falling on the soil sur-face contributed winter peak of the annual variation ofphosphatase activity (Harrison and Pearce 1979) Like-wise higher activities of phosphatase and phenol oxi-dase were found in riparian peatlands in late autumnwhich was attributed to litter quality and quantity(Bonnett et al 2006) Sinsabaugh et al (1991) has alsoreported that maximum enzyme activities were detectedin several weeks after the production of litter As suchlitter production and distribution may offset effects fromother controlling variables such as temperature Previ-
ous study has shown that the site is dominated bynortherly wind and consequent movement of leaf littertoward northern slope (Lee et al 1999 Yoo et al 2001)This contributes the general trends of higher enzymeactivities in northern aspect than in southern aspect inthis study The contribution of leaf litter to enzymeactivity is further supported by the finding that spa-tial differences in enzyme activities emerged only after
Landscape positions
N-Foot N-Back Ridge S-Back S-Sasa S-Foot
Pho
spha
tase
(mm
ole
g-1hr
-1)
0
1
2
3
4
5 a
b
aa a
b b
bb
b
ab
c
Fig 2 Variations of phosphatase activities before the litterfallproduction (unshaded) and after litterfall production (black) Thevalues are mean of three replicates with a standard error Bars withdifferent letters are significantly different among the same enzymesat P lt 005
Ions
Al Fe Ca K Mg
Con
tent
s(m
gg-1
)
0
2
4
6
8
10
a
a
a
a
a
c
b
b
c
bc
ba
a b
Fig 3 Metal ion concentrations in the north-facing backslope(unshaded) ridge (hatched) and south-facing backslope (black) Thevalues are mean of three replicates with a standard error Thevalues are significantly different for each ion at P lt 001 exceptAl which is different at P lt 005
Temperature ( C)
0 5 10 15 20 25
Deh
ydro
gena
se(
gg-1
hr-1
)
0
4
8
12
16
A
r = 0458 n = 7 P lt 01
Temperature ( C)
0 5 10 15 20 25
Ure
ase
(m
olg-1
hr-1
)
0
50
100
150
200
250
300
350
B
r = 0730 n = 8 P lt 005
Urease activity ( mol g-1 hr-1)
0 100 200 300 400
Am
mon
ium
(g
g-1)
0
20
40
60
80
100
120
140
r = -0531 P lt 001 n = 20
C
Fig 4 Relationships between dehydrogenase and temperature (a)urease and temperature (b) and urease and ammonium (c)
1141
litterfall production (Fig 2) It is also interesting to notethat differences in enzyme activities between south-facing backslope without S borealis and south-facingbackslope with S borealis were only found after thelitterfall while such differences were absent beforethe litterfall production (Fig 2) Because areas withS borealis can maintain much larger amount of leaflitter than areas without S borealis understory vegeta-tion may influence biogeochemical processes in forestfloor through leaf litter distribution (Lee et al 1999)
Metal ions notably Fe and Al have reported toinhibit soil enzyme activities by absorbing to the activesite (Ekenler and Tabatabai 2002) In contrast Ca2+
and Mg2+ content often exhibited a positive correlationwith enzyme activities because those are known to re-move inhibition of humic materials by enzymendashhumicinteractions (Freeman et al 1996) Our results exhibitedthat two inhibitory ions (Al Fe) were higher whilst Mgand K contents were lower in soil of the southern-facingslope (Oh et al 1999) As such higher concentration ofinhibitory ions and lower concentration of activationions partially account for the lower enzyme activities inthe southern-facing slope
In this study we assessed the changes in enzymeactivities only However previous studies have revealedthat microbial composition such as bacteria versus fungimay vary over time (Moore-Kucera and Dick 2008Kubartova et al 2007) This would be closely related toenzyme activities because different types of microor-ganisms are responsible for the production of specificenzymes
Landscape position has been reported to affect soilproperties litter decomposition nitrogen dynamics andphosphorus availability (Brubaker et al 1994 Klem-mendson and Wienhold 1991 Mudrick et al 1994Garten Jr et al 1994) These studies focused on thedifferences in organic matter water and nutrient con-tents which were caused by flow of water and soil ero-sions In addition to these factors our results indicatedthat production and redistribution of leaf litter are otherkey factors affecting biogeochemical cycles in a forestfloor Furthermore not only the physical movement ofleaf litter mediated by wind but also understory vege-tation which may impede the movement could play animportant role in this dynamics
Acknowledgments H Kang is grateful to AEBRC EcoRiver21 forfinancial support
References
Aber JD Mellilo JM (1991) Terrestrial ecosystems SaundersCollege Publishing Philadelphia
Bonnett SAF Ostle N Freeman C (2006) Seasonal variations indecomposition processes in a valley-bottom riparian peatlandSci Total Environ 370561ndash573 doi101016jscitotenv200608032
Bremner JM Mulvaney RL (1978) Urease activity in soils InBurns RG (ed) Soil enzymes Academic Press New York
Brubaker SC Jones AJ Frank K Lewis DT (1994) Regressionmodels for estimating soil properties by landscape position SoilSci Soc Am J 581763ndash1767
Burns RG (1982) Enzyme activity in soil location and a possiblerole in microbial ecology Soil Biol Biochem 14423ndash427 doi1010160038-0717(82)90099-2
Chrost RJ (1991) Environmental control of the synthesis andactivity of aquatic microbial ectoenzymes In Chrost RJ (ed)Microbial enzymes in aquatic environments Springer BerlinHeidelberg New York pp 29ndash59
Dilly O Munch JC (1996) Microbial biomass content basal res-piration and enzyme activities during the course of decompo-sition of leaf litter in a black alder (Alnus glutinosa (L) Gaertn)forest Soil Biol Biochem 281073ndash1081 doi1010160038-0717(96)00075-2
Dilly O Nannipieri P (2001) Response of ATP content respirationrate and enzyme activities in an arable and a forest soil tonutrient addition Biol Fertil Soils 3464ndash72 doi101007s003740100375
Dilly O Bartsch S Rosenbrock P Buscot F Munch J-C (2001)Shifts in physiological capabilities of the microbiota during thedecomposition of leaf litter in a black alder (Alnus glutinosa(Gaertn) L) forest Soil Biol Biochem 33921ndash930 doi101016S0038-0717(00)00239-X
Dilly O Munch J-C Pfeiffer E-M (2007) Enzyme activities andlitter decomposition in agricultural soils in northern centraland southern Germany J Plant Nutr Soil Sci 170197ndash204 doi101002jpln200622044
Eivazi F Tabatabai MA (1990) Factors affecting glucosidase andgalactosidase activities in soils Soil Biol Biochem 22891ndash897doi1010160038-0717(90)90126-K
Ekenler M Tabatabai MA (2002) Effects of trace elements on b-glucosaminidase activity in soils Soil Biol Biochem 341829ndash1832 doi101016S0038-0717(02)00167-0
Emteryd O (1989) Chemical and physical analysis of inorganicnutrients in plant soil water and air Department of Forest SiteResearch Swedish University of Agricultural Sciences Umea
Eviner VT Chapin FS Vaughn CE (2006) Seasonal variations inplant species effects on soil N and P dynamics Ecology 87974ndash986 doi1018900012-9658(2006)87[974SVIPSE]20CO2
Freeman C Liska G Ostle NJ Lock MA Reynolds B Hudson J(1996) Microbial activity and enzymic decomposition processesfollowing peatland water table drawdown Plant Soil 180121ndash127 doi101007BF00015418
Garten CT Jr Huston MA Thoms CA (1994) Topographic vari-ation of soil nitrogen dynamics at Walker Branch watershedTennessee For Sci 40497ndash512
Harrison AF Pearce T (1979) Seasonal variation of phosphataseactivity in woodland soils Soil Biol Biochem 11405ndash410 doi1010160038-0717(79)90054-3
Kandeler E Gerber H (1988) Short-term assay of soil ureaseactivity using colorimetric determination of ammonium BiolFertil Soils 658ndash72 doi101007BF00257924
Klemmendson JO Wienhold BJ (1991) Aspect and species influ-ences on nitrogen and phosphorus availability in Arizonachaparral soils Soil Sci Soc Am J 551735ndash1740
Kubartova A Moukoumi J Beguiristain T Ranger J Berthelin J(2007) Microbial diversity during cellulose decomposition indifferent forest stands I Microbial communities and environ-mental conditions Microb Ecol 54393ndash405 doi101007s00248-007-9286-2
Lee D Yoo G Oh S Shim JH Kang S (1999) Significance ofaspect and understory type to leaf litter redistribution in atemperate hardwood forest Korean J Biol Sci 3143ndash147
Moore-Kucera J Dick RP (2008) PLFA profiling of microbialcommunity structure and seasonal shifts in soils of a Douglas-fir chronosequence Microb Ecol 55500ndash511 doi101007s00248-007-9295-1
Mudrick DA Hoosein M Hicks RR Jr Townsend EC (1994)Decomposition of leaf litter in an Appalachian forest effects ofleaf species aspect slope position and time For Ecol Manage68231ndash250
1142
Noorbakhsh S Schoenau J Si B Zeleke T Qian P (2008) Soilproperties yield and landscape relationships in south-centralSaskatchewan Canada J Plant Nutr 31539ndash556 doi10108001904160801895076
Oh S Yoo G Shim JH Kang S Lee D (1999) Spatial heterogeneityof soil K Ca Mg Fe and Al generated by leaf litter redistri-bution in a temperate hardwood forest Kor J Ecol 22205ndash210
Sariyildiz T Anderson JM Kucuk M (2005) Effects of tree speciesand topography on soil chemistry litter quality and decom-position in northeast Turkey Soil Biol Biochem 371695ndash1706doi101016jsoilbio200502004
Schimel D Stillwell MA Woodmansee RG (1985) Biogeochemis-try of C N and P in a soil catena of the shortgrass steppeEcology 66276ndash282 doi1023071941328
Sinsabaugh RL Antibus RK Linkins AE (1991) An enzymicapproach to the analysis of microbial activity during plant litterdecomposition Agric Ecosyst Environ 3443ndash54 doi1010160167-8809(91)90092-C
Tabatabai MA Bremner JM (1970) Arylsulfatase activity of soilsSoil Sci Soc Am Proc 34225ndash229
Trevors JT Mayfield CI Inniss WE (1982) Measurement of elec-tron transport system (ETS) activity in soil Microb Ecol 8163ndash168 doi101007BF02010449
Vetanovetz RP Peterson JC (1992) Effect of carbon source andnitrogen on urease activity in a Sphagnum peat mediumCommun Soil Sci Plant Anal 23379ndash388 doi10108000103629209368596
Yoo G Park E-J Kim S-H Lee H-J Kang S Lee D (2001)Transport and decomposition of leaf litter as affected by aspectand understory in a temperate hardwood forest Korean J BiolSci 5319ndash325
1143

samples in October and November (Autumn-1) in 1995May and June (Spring-1) September and November in1996 (Autumn-2) In 1997 samples were collected inApril and June (Spring-2) In addition two spatiallyintensive samplings were conducted in all the six patchesin August (before litter fall) and September (after litterfall) 1996 Upon arriving at the laboratory the sampleswere sieved (lt2 mm) and maintained at 4C until theanalyses were conducted
Enzyme assays
Dehydrogenase activity was determined by adding INT(iodonitrozo tetrazolium chloride) as a model substrateand measuring reduction of the chemical spectrophoto-metrically (Trevors et al 1982) Phosphatase and aryl-sulfatase activities were assayed by measuring releasedp-nitrophenol over an hour of incubation from soilsadded with p-nitrophenyl phosphate and p-nitrophenylsulfate as substrates respectively (Eivazi and Tabatabai1990 Tabatabai and Bremner 1970) Urease activity wasdetermined by measuring released NH4
+ from the soilamended with urea described by Kandeler and Gerber(1988) In each assay 2 g of soils was used and at leasttriplicate samples were employed for each location Allthe enzyme activities were presented per dry mass of soilper incubation duration
Chemical analysis
Extractable ammonium concentration in the soil wasdetermined by indophenol method (Emteryd 1989)followed by extraction with 2 M KCl Metal ions weremeasured by an ICP-AES (Perkin-Elmer Plasma 40)after digesting crushed soils (500 lmlt) in mixed acidsolution (HNO3 HF HClO4 = 4 4 1) for 12 h
Statistical analysis
The enzyme activities and NH4+ contents were analyzed
according to a repeated measures ANOVA Differencesin enzyme activities in six patches were determined by aone-way ANOVA followed by Tukeyrsquos test All pro-cesses were conducted on SPSS version 150
Results
All enzyme activities showed significant variations bysampling time landscape position and the interactionsof the two (Table 1) Overall the enzyme activities werehigher on the north-facing backslope than in the south-facing backslope throughout the studying period withfew exceptions (eg dehydrogenase in autumn-2 andarylsulfatase in autumn-2) (Fig 1)
For the seasonal variations two patterns were dis-cernable Dehydrogenase and urease activities werehigher in spring than those in autumn (Fig 1a b) Incontrast higher activity of phosphatase was generallyobserved in autumn than in spring (Fig 1c) Arylsulfa-tase showed a similar temporal pattern as phosphatasebut it was less discernable (Fig 1d) Extractableammonium concentrations were higher in autumn thanin spring (Fig 1e) However there were no generalpatterns of differences among the three plots
Overall phosphatase and urease activities were lowerbefore the litterfall production (BF) than those after thelitterfall production (AF) (Fig 2) Phosphatase activitiesranged between 15 and 42 mmol g1 h1 after the lit-terfall production while the same enzyme activity ran-ged between 16 and 22 mmol g1 h1 before thelitterfall production Similar trends were observed forurease activity 28ndash43 NH4
+ lg g1 h1 (AF) and22ndash28 lg g1 min1 (BF) No spatial differences werediscernible before the litterfall production (Fig 2)However significantly different enzyme activities werefound among patches after the litterfall production insuch a way that enzyme activities were the highest innorth-facing footslope and the lowest in south-facingbackslope without S borealis (Fig 2) In contrast theactivities in south-facing backslope with S borealsiswere not different from north-facing backslope ridge orsouth-facing footslope (Fig 2)
Al Fe and Mg contents were lower on the north-facing slope than those on the south-facing slope(Fig 3) In contrast Ca content was higher on thenorth-facing slope than that on the south-facing slope
Discussion
Several environmental variables are known to affect soilenzyme activities Temperature can influence enzymeactivity by direct modification of enzyme kinetics aswell as indirect influence on microbial proliferation
Table 1 A repeated-measure ANOVA table for the enzyme activ-ities and ammonium contents
Dependent Independent df F value P value
Dehydrogenase Position 2 534 0000Months 7 440 0000Position middot months 14 128 0003
Urease Position 2 660 0000Months 7 479 0000Position middot months 14 177 0000
Phosphatase Position 2 634 0000Months 7 6214 0000Position middot months 14 143 0000
Arylsulfatase Position 2 62 0004Months 7 523 0000Position middot months 14 21 0032
Ammonium Position 2 71 0002Months 7 12084 0000Position middot months 14 65 0000
1139
Dehydrogenase and urease appear to be influenced bytemperature in this study which is reflected in positivecorrelations between temperature and dehydrogenase(r = 0458 n = 7 P lt 01) or urease (r = 0730n = 8 P lt 005) activities (Fig 4) Since dehydroge-nase is closely related to active microorganisms andmost of urease activity is known to be originated fromintracellular enzymes rather than extracellular enzymes(Bremner and Mulvaney 1978) the temperature effectappears to be related to active proliferation of microbesin spring compared to autumn It has also been notedthat urease is closely related to nutritional conditionssuggesting its relation with general proliferation ofmicrobes (Dilly and Nannipieri 2001) Another keycontrolling variable for urease is ammonium availability
Because production of urease by microbes is negativelyregulated by ammonium availability (Vetanovetz andPeterson 1992) when ammonium availability is high inautumn urease could be lower In contrast ammoniumavailability is much lower in spring probably due toactive uptake of ammonium by microorganisms andvegetation This could accelerate urease productionfrom microbes in soils A significant negative correlationbetween urease and ammonium concentration(r = 0531 n = 20 P lt 001) supports this specula-tion (Fig 4)
Unlike dehydrogenase and urease phosphatase andarylsulfatase activities were not affected by temperatureThe activities appeared to be slightly higher in autumnthan those in spring (Fig 1c d) These two enzymes are
Deh
ydro
gena
se(
gg-1
hr-1
)
0
5
10
15
20 A
ND
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Ure
ase
(m
olg-1
hr-1
)
0
50
100
150
200
250
300
350
400
B
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Pho
spha
tase
(mm
olg-1
hr-1
)
0
1
2
3
4
5 C
Autumn - 1 Spring - 2Autumn - 2Spring - 1
Ary
lsuf
atas
e(m
mol
g-1hr
-1)
00
05
10
15
20
25
30
D
NH
4+(
gg-1
)
0
20
40
60
80
100
120
140
ND
E
Fig 1 Variations of dehydrogenase (a) urease (b) phosphatase (c) arylsulfatase (d) and ammonium (e) in the north-facing backslope(unshaded) ridge (hatched) and south-facing backslope (black) The values are the mean of three replicates with a standard error
1140
known to be mainly extracellular and stabilized andhence less affiliated with microbial biomass compared todehydrogenase or urease Further it was reported thatenzymes leached from leaf litter falling on the soil sur-face contributed winter peak of the annual variation ofphosphatase activity (Harrison and Pearce 1979) Like-wise higher activities of phosphatase and phenol oxi-dase were found in riparian peatlands in late autumnwhich was attributed to litter quality and quantity(Bonnett et al 2006) Sinsabaugh et al (1991) has alsoreported that maximum enzyme activities were detectedin several weeks after the production of litter As suchlitter production and distribution may offset effects fromother controlling variables such as temperature Previ-
ous study has shown that the site is dominated bynortherly wind and consequent movement of leaf littertoward northern slope (Lee et al 1999 Yoo et al 2001)This contributes the general trends of higher enzymeactivities in northern aspect than in southern aspect inthis study The contribution of leaf litter to enzymeactivity is further supported by the finding that spa-tial differences in enzyme activities emerged only after
Landscape positions
N-Foot N-Back Ridge S-Back S-Sasa S-Foot
Pho
spha
tase
(mm
ole
g-1hr
-1)
0
1
2
3
4
5 a
b
aa a
b b
bb
b
ab
c
Fig 2 Variations of phosphatase activities before the litterfallproduction (unshaded) and after litterfall production (black) Thevalues are mean of three replicates with a standard error Bars withdifferent letters are significantly different among the same enzymesat P lt 005
Ions
Al Fe Ca K Mg
Con
tent
s(m
gg-1
)
0
2
4
6
8
10
a
a
a
a
a
c
b
b
c
bc
ba
a b
Fig 3 Metal ion concentrations in the north-facing backslope(unshaded) ridge (hatched) and south-facing backslope (black) Thevalues are mean of three replicates with a standard error Thevalues are significantly different for each ion at P lt 001 exceptAl which is different at P lt 005
Temperature ( C)
0 5 10 15 20 25
Deh
ydro
gena
se(
gg-1
hr-1
)
0
4
8
12
16
A
r = 0458 n = 7 P lt 01
Temperature ( C)
0 5 10 15 20 25
Ure
ase
(m
olg-1
hr-1
)
0
50
100
150
200
250
300
350
B
r = 0730 n = 8 P lt 005
Urease activity ( mol g-1 hr-1)
0 100 200 300 400
Am
mon
ium
(g
g-1)
0
20
40
60
80
100
120
140
r = -0531 P lt 001 n = 20
C
Fig 4 Relationships between dehydrogenase and temperature (a)urease and temperature (b) and urease and ammonium (c)
1141
litterfall production (Fig 2) It is also interesting to notethat differences in enzyme activities between south-facing backslope without S borealis and south-facingbackslope with S borealis were only found after thelitterfall while such differences were absent beforethe litterfall production (Fig 2) Because areas withS borealis can maintain much larger amount of leaflitter than areas without S borealis understory vegeta-tion may influence biogeochemical processes in forestfloor through leaf litter distribution (Lee et al 1999)
Metal ions notably Fe and Al have reported toinhibit soil enzyme activities by absorbing to the activesite (Ekenler and Tabatabai 2002) In contrast Ca2+
and Mg2+ content often exhibited a positive correlationwith enzyme activities because those are known to re-move inhibition of humic materials by enzymendashhumicinteractions (Freeman et al 1996) Our results exhibitedthat two inhibitory ions (Al Fe) were higher whilst Mgand K contents were lower in soil of the southern-facingslope (Oh et al 1999) As such higher concentration ofinhibitory ions and lower concentration of activationions partially account for the lower enzyme activities inthe southern-facing slope
In this study we assessed the changes in enzymeactivities only However previous studies have revealedthat microbial composition such as bacteria versus fungimay vary over time (Moore-Kucera and Dick 2008Kubartova et al 2007) This would be closely related toenzyme activities because different types of microor-ganisms are responsible for the production of specificenzymes
Landscape position has been reported to affect soilproperties litter decomposition nitrogen dynamics andphosphorus availability (Brubaker et al 1994 Klem-mendson and Wienhold 1991 Mudrick et al 1994Garten Jr et al 1994) These studies focused on thedifferences in organic matter water and nutrient con-tents which were caused by flow of water and soil ero-sions In addition to these factors our results indicatedthat production and redistribution of leaf litter are otherkey factors affecting biogeochemical cycles in a forestfloor Furthermore not only the physical movement ofleaf litter mediated by wind but also understory vege-tation which may impede the movement could play animportant role in this dynamics
Acknowledgments H Kang is grateful to AEBRC EcoRiver21 forfinancial support
References
Aber JD Mellilo JM (1991) Terrestrial ecosystems SaundersCollege Publishing Philadelphia
Bonnett SAF Ostle N Freeman C (2006) Seasonal variations indecomposition processes in a valley-bottom riparian peatlandSci Total Environ 370561ndash573 doi101016jscitotenv200608032
Bremner JM Mulvaney RL (1978) Urease activity in soils InBurns RG (ed) Soil enzymes Academic Press New York
Brubaker SC Jones AJ Frank K Lewis DT (1994) Regressionmodels for estimating soil properties by landscape position SoilSci Soc Am J 581763ndash1767
Burns RG (1982) Enzyme activity in soil location and a possiblerole in microbial ecology Soil Biol Biochem 14423ndash427 doi1010160038-0717(82)90099-2
Chrost RJ (1991) Environmental control of the synthesis andactivity of aquatic microbial ectoenzymes In Chrost RJ (ed)Microbial enzymes in aquatic environments Springer BerlinHeidelberg New York pp 29ndash59
Dilly O Munch JC (1996) Microbial biomass content basal res-piration and enzyme activities during the course of decompo-sition of leaf litter in a black alder (Alnus glutinosa (L) Gaertn)forest Soil Biol Biochem 281073ndash1081 doi1010160038-0717(96)00075-2
Dilly O Nannipieri P (2001) Response of ATP content respirationrate and enzyme activities in an arable and a forest soil tonutrient addition Biol Fertil Soils 3464ndash72 doi101007s003740100375
Dilly O Bartsch S Rosenbrock P Buscot F Munch J-C (2001)Shifts in physiological capabilities of the microbiota during thedecomposition of leaf litter in a black alder (Alnus glutinosa(Gaertn) L) forest Soil Biol Biochem 33921ndash930 doi101016S0038-0717(00)00239-X
Dilly O Munch J-C Pfeiffer E-M (2007) Enzyme activities andlitter decomposition in agricultural soils in northern centraland southern Germany J Plant Nutr Soil Sci 170197ndash204 doi101002jpln200622044
Eivazi F Tabatabai MA (1990) Factors affecting glucosidase andgalactosidase activities in soils Soil Biol Biochem 22891ndash897doi1010160038-0717(90)90126-K
Ekenler M Tabatabai MA (2002) Effects of trace elements on b-glucosaminidase activity in soils Soil Biol Biochem 341829ndash1832 doi101016S0038-0717(02)00167-0
Emteryd O (1989) Chemical and physical analysis of inorganicnutrients in plant soil water and air Department of Forest SiteResearch Swedish University of Agricultural Sciences Umea
Eviner VT Chapin FS Vaughn CE (2006) Seasonal variations inplant species effects on soil N and P dynamics Ecology 87974ndash986 doi1018900012-9658(2006)87[974SVIPSE]20CO2
Freeman C Liska G Ostle NJ Lock MA Reynolds B Hudson J(1996) Microbial activity and enzymic decomposition processesfollowing peatland water table drawdown Plant Soil 180121ndash127 doi101007BF00015418
Garten CT Jr Huston MA Thoms CA (1994) Topographic vari-ation of soil nitrogen dynamics at Walker Branch watershedTennessee For Sci 40497ndash512
Harrison AF Pearce T (1979) Seasonal variation of phosphataseactivity in woodland soils Soil Biol Biochem 11405ndash410 doi1010160038-0717(79)90054-3
Kandeler E Gerber H (1988) Short-term assay of soil ureaseactivity using colorimetric determination of ammonium BiolFertil Soils 658ndash72 doi101007BF00257924
Klemmendson JO Wienhold BJ (1991) Aspect and species influ-ences on nitrogen and phosphorus availability in Arizonachaparral soils Soil Sci Soc Am J 551735ndash1740
Kubartova A Moukoumi J Beguiristain T Ranger J Berthelin J(2007) Microbial diversity during cellulose decomposition indifferent forest stands I Microbial communities and environ-mental conditions Microb Ecol 54393ndash405 doi101007s00248-007-9286-2
Lee D Yoo G Oh S Shim JH Kang S (1999) Significance ofaspect and understory type to leaf litter redistribution in atemperate hardwood forest Korean J Biol Sci 3143ndash147
Moore-Kucera J Dick RP (2008) PLFA profiling of microbialcommunity structure and seasonal shifts in soils of a Douglas-fir chronosequence Microb Ecol 55500ndash511 doi101007s00248-007-9295-1
Mudrick DA Hoosein M Hicks RR Jr Townsend EC (1994)Decomposition of leaf litter in an Appalachian forest effects ofleaf species aspect slope position and time For Ecol Manage68231ndash250
1142
Noorbakhsh S Schoenau J Si B Zeleke T Qian P (2008) Soilproperties yield and landscape relationships in south-centralSaskatchewan Canada J Plant Nutr 31539ndash556 doi10108001904160801895076
Oh S Yoo G Shim JH Kang S Lee D (1999) Spatial heterogeneityof soil K Ca Mg Fe and Al generated by leaf litter redistri-bution in a temperate hardwood forest Kor J Ecol 22205ndash210
Sariyildiz T Anderson JM Kucuk M (2005) Effects of tree speciesand topography on soil chemistry litter quality and decom-position in northeast Turkey Soil Biol Biochem 371695ndash1706doi101016jsoilbio200502004
Schimel D Stillwell MA Woodmansee RG (1985) Biogeochemis-try of C N and P in a soil catena of the shortgrass steppeEcology 66276ndash282 doi1023071941328
Sinsabaugh RL Antibus RK Linkins AE (1991) An enzymicapproach to the analysis of microbial activity during plant litterdecomposition Agric Ecosyst Environ 3443ndash54 doi1010160167-8809(91)90092-C
Tabatabai MA Bremner JM (1970) Arylsulfatase activity of soilsSoil Sci Soc Am Proc 34225ndash229
Trevors JT Mayfield CI Inniss WE (1982) Measurement of elec-tron transport system (ETS) activity in soil Microb Ecol 8163ndash168 doi101007BF02010449
Vetanovetz RP Peterson JC (1992) Effect of carbon source andnitrogen on urease activity in a Sphagnum peat mediumCommun Soil Sci Plant Anal 23379ndash388 doi10108000103629209368596
Yoo G Park E-J Kim S-H Lee H-J Kang S Lee D (2001)Transport and decomposition of leaf litter as affected by aspectand understory in a temperate hardwood forest Korean J BiolSci 5319ndash325
1143
Dehydrogenase and urease appear to be influenced bytemperature in this study which is reflected in positivecorrelations between temperature and dehydrogenase(r = 0458 n = 7 P lt 01) or urease (r = 0730n = 8 P lt 005) activities (Fig 4) Since dehydroge-nase is closely related to active microorganisms andmost of urease activity is known to be originated fromintracellular enzymes rather than extracellular enzymes(Bremner and Mulvaney 1978) the temperature effectappears to be related to active proliferation of microbesin spring compared to autumn It has also been notedthat urease is closely related to nutritional conditionssuggesting its relation with general proliferation ofmicrobes (Dilly and Nannipieri 2001) Another keycontrolling variable for urease is ammonium availability
Because production of urease by microbes is negativelyregulated by ammonium availability (Vetanovetz andPeterson 1992) when ammonium availability is high inautumn urease could be lower In contrast ammoniumavailability is much lower in spring probably due toactive uptake of ammonium by microorganisms andvegetation This could accelerate urease productionfrom microbes in soils A significant negative correlationbetween urease and ammonium concentration(r = 0531 n = 20 P lt 001) supports this specula-tion (Fig 4)
Unlike dehydrogenase and urease phosphatase andarylsulfatase activities were not affected by temperatureThe activities appeared to be slightly higher in autumnthan those in spring (Fig 1c d) These two enzymes are
Deh
ydro
gena
se(
gg-1
hr-1
)
0
5
10
15
20 A
ND
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Ure
ase
(m
olg-1
hr-1
)
0
50
100
150
200
250
300
350
400
B
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Autumn - 1 Spring - 1 Autumn - 2 Spring - 2
Pho
spha
tase
(mm
olg-1
hr-1
)
0
1
2
3
4
5 C
Autumn - 1 Spring - 2Autumn - 2Spring - 1
Ary
lsuf
atas
e(m
mol
g-1hr
-1)
00
05
10
15
20
25
30
D
NH
4+(
gg-1
)
0
20
40
60
80
100
120
140
ND
E
Fig 1 Variations of dehydrogenase (a) urease (b) phosphatase (c) arylsulfatase (d) and ammonium (e) in the north-facing backslope(unshaded) ridge (hatched) and south-facing backslope (black) The values are the mean of three replicates with a standard error
1140
known to be mainly extracellular and stabilized andhence less affiliated with microbial biomass compared todehydrogenase or urease Further it was reported thatenzymes leached from leaf litter falling on the soil sur-face contributed winter peak of the annual variation ofphosphatase activity (Harrison and Pearce 1979) Like-wise higher activities of phosphatase and phenol oxi-dase were found in riparian peatlands in late autumnwhich was attributed to litter quality and quantity(Bonnett et al 2006) Sinsabaugh et al (1991) has alsoreported that maximum enzyme activities were detectedin several weeks after the production of litter As suchlitter production and distribution may offset effects fromother controlling variables such as temperature Previ-
ous study has shown that the site is dominated bynortherly wind and consequent movement of leaf littertoward northern slope (Lee et al 1999 Yoo et al 2001)This contributes the general trends of higher enzymeactivities in northern aspect than in southern aspect inthis study The contribution of leaf litter to enzymeactivity is further supported by the finding that spa-tial differences in enzyme activities emerged only after
Landscape positions
N-Foot N-Back Ridge S-Back S-Sasa S-Foot
Pho
spha
tase
(mm
ole
g-1hr
-1)
0
1
2
3
4
5 a
b
aa a
b b
bb
b
ab
c
Fig 2 Variations of phosphatase activities before the litterfallproduction (unshaded) and after litterfall production (black) Thevalues are mean of three replicates with a standard error Bars withdifferent letters are significantly different among the same enzymesat P lt 005
Ions
Al Fe Ca K Mg
Con
tent
s(m
gg-1
)
0
2
4
6
8
10
a
a
a
a
a
c
b
b
c
bc
ba
a b
Fig 3 Metal ion concentrations in the north-facing backslope(unshaded) ridge (hatched) and south-facing backslope (black) Thevalues are mean of three replicates with a standard error Thevalues are significantly different for each ion at P lt 001 exceptAl which is different at P lt 005
Temperature ( C)
0 5 10 15 20 25
Deh
ydro
gena
se(
gg-1
hr-1
)
0
4
8
12
16
A
r = 0458 n = 7 P lt 01
Temperature ( C)
0 5 10 15 20 25
Ure
ase
(m
olg-1
hr-1
)
0
50
100
150
200
250
300
350
B
r = 0730 n = 8 P lt 005
Urease activity ( mol g-1 hr-1)
0 100 200 300 400
Am
mon
ium
(g
g-1)
0
20
40
60
80
100
120
140
r = -0531 P lt 001 n = 20
C
Fig 4 Relationships between dehydrogenase and temperature (a)urease and temperature (b) and urease and ammonium (c)
1141
litterfall production (Fig 2) It is also interesting to notethat differences in enzyme activities between south-facing backslope without S borealis and south-facingbackslope with S borealis were only found after thelitterfall while such differences were absent beforethe litterfall production (Fig 2) Because areas withS borealis can maintain much larger amount of leaflitter than areas without S borealis understory vegeta-tion may influence biogeochemical processes in forestfloor through leaf litter distribution (Lee et al 1999)
Metal ions notably Fe and Al have reported toinhibit soil enzyme activities by absorbing to the activesite (Ekenler and Tabatabai 2002) In contrast Ca2+
and Mg2+ content often exhibited a positive correlationwith enzyme activities because those are known to re-move inhibition of humic materials by enzymendashhumicinteractions (Freeman et al 1996) Our results exhibitedthat two inhibitory ions (Al Fe) were higher whilst Mgand K contents were lower in soil of the southern-facingslope (Oh et al 1999) As such higher concentration ofinhibitory ions and lower concentration of activationions partially account for the lower enzyme activities inthe southern-facing slope
In this study we assessed the changes in enzymeactivities only However previous studies have revealedthat microbial composition such as bacteria versus fungimay vary over time (Moore-Kucera and Dick 2008Kubartova et al 2007) This would be closely related toenzyme activities because different types of microor-ganisms are responsible for the production of specificenzymes
Landscape position has been reported to affect soilproperties litter decomposition nitrogen dynamics andphosphorus availability (Brubaker et al 1994 Klem-mendson and Wienhold 1991 Mudrick et al 1994Garten Jr et al 1994) These studies focused on thedifferences in organic matter water and nutrient con-tents which were caused by flow of water and soil ero-sions In addition to these factors our results indicatedthat production and redistribution of leaf litter are otherkey factors affecting biogeochemical cycles in a forestfloor Furthermore not only the physical movement ofleaf litter mediated by wind but also understory vege-tation which may impede the movement could play animportant role in this dynamics
Acknowledgments H Kang is grateful to AEBRC EcoRiver21 forfinancial support
References
Aber JD Mellilo JM (1991) Terrestrial ecosystems SaundersCollege Publishing Philadelphia
Bonnett SAF Ostle N Freeman C (2006) Seasonal variations indecomposition processes in a valley-bottom riparian peatlandSci Total Environ 370561ndash573 doi101016jscitotenv200608032
Bremner JM Mulvaney RL (1978) Urease activity in soils InBurns RG (ed) Soil enzymes Academic Press New York
Brubaker SC Jones AJ Frank K Lewis DT (1994) Regressionmodels for estimating soil properties by landscape position SoilSci Soc Am J 581763ndash1767
Burns RG (1982) Enzyme activity in soil location and a possiblerole in microbial ecology Soil Biol Biochem 14423ndash427 doi1010160038-0717(82)90099-2
Chrost RJ (1991) Environmental control of the synthesis andactivity of aquatic microbial ectoenzymes In Chrost RJ (ed)Microbial enzymes in aquatic environments Springer BerlinHeidelberg New York pp 29ndash59
Dilly O Munch JC (1996) Microbial biomass content basal res-piration and enzyme activities during the course of decompo-sition of leaf litter in a black alder (Alnus glutinosa (L) Gaertn)forest Soil Biol Biochem 281073ndash1081 doi1010160038-0717(96)00075-2
Dilly O Nannipieri P (2001) Response of ATP content respirationrate and enzyme activities in an arable and a forest soil tonutrient addition Biol Fertil Soils 3464ndash72 doi101007s003740100375
Dilly O Bartsch S Rosenbrock P Buscot F Munch J-C (2001)Shifts in physiological capabilities of the microbiota during thedecomposition of leaf litter in a black alder (Alnus glutinosa(Gaertn) L) forest Soil Biol Biochem 33921ndash930 doi101016S0038-0717(00)00239-X
Dilly O Munch J-C Pfeiffer E-M (2007) Enzyme activities andlitter decomposition in agricultural soils in northern centraland southern Germany J Plant Nutr Soil Sci 170197ndash204 doi101002jpln200622044
Eivazi F Tabatabai MA (1990) Factors affecting glucosidase andgalactosidase activities in soils Soil Biol Biochem 22891ndash897doi1010160038-0717(90)90126-K
Ekenler M Tabatabai MA (2002) Effects of trace elements on b-glucosaminidase activity in soils Soil Biol Biochem 341829ndash1832 doi101016S0038-0717(02)00167-0
Emteryd O (1989) Chemical and physical analysis of inorganicnutrients in plant soil water and air Department of Forest SiteResearch Swedish University of Agricultural Sciences Umea
Eviner VT Chapin FS Vaughn CE (2006) Seasonal variations inplant species effects on soil N and P dynamics Ecology 87974ndash986 doi1018900012-9658(2006)87[974SVIPSE]20CO2
Freeman C Liska G Ostle NJ Lock MA Reynolds B Hudson J(1996) Microbial activity and enzymic decomposition processesfollowing peatland water table drawdown Plant Soil 180121ndash127 doi101007BF00015418
Garten CT Jr Huston MA Thoms CA (1994) Topographic vari-ation of soil nitrogen dynamics at Walker Branch watershedTennessee For Sci 40497ndash512
Harrison AF Pearce T (1979) Seasonal variation of phosphataseactivity in woodland soils Soil Biol Biochem 11405ndash410 doi1010160038-0717(79)90054-3
Kandeler E Gerber H (1988) Short-term assay of soil ureaseactivity using colorimetric determination of ammonium BiolFertil Soils 658ndash72 doi101007BF00257924
Klemmendson JO Wienhold BJ (1991) Aspect and species influ-ences on nitrogen and phosphorus availability in Arizonachaparral soils Soil Sci Soc Am J 551735ndash1740
Kubartova A Moukoumi J Beguiristain T Ranger J Berthelin J(2007) Microbial diversity during cellulose decomposition indifferent forest stands I Microbial communities and environ-mental conditions Microb Ecol 54393ndash405 doi101007s00248-007-9286-2
Lee D Yoo G Oh S Shim JH Kang S (1999) Significance ofaspect and understory type to leaf litter redistribution in atemperate hardwood forest Korean J Biol Sci 3143ndash147
Moore-Kucera J Dick RP (2008) PLFA profiling of microbialcommunity structure and seasonal shifts in soils of a Douglas-fir chronosequence Microb Ecol 55500ndash511 doi101007s00248-007-9295-1
Mudrick DA Hoosein M Hicks RR Jr Townsend EC (1994)Decomposition of leaf litter in an Appalachian forest effects ofleaf species aspect slope position and time For Ecol Manage68231ndash250
1142
Noorbakhsh S Schoenau J Si B Zeleke T Qian P (2008) Soilproperties yield and landscape relationships in south-centralSaskatchewan Canada J Plant Nutr 31539ndash556 doi10108001904160801895076
Oh S Yoo G Shim JH Kang S Lee D (1999) Spatial heterogeneityof soil K Ca Mg Fe and Al generated by leaf litter redistri-bution in a temperate hardwood forest Kor J Ecol 22205ndash210
Sariyildiz T Anderson JM Kucuk M (2005) Effects of tree speciesand topography on soil chemistry litter quality and decom-position in northeast Turkey Soil Biol Biochem 371695ndash1706doi101016jsoilbio200502004
Schimel D Stillwell MA Woodmansee RG (1985) Biogeochemis-try of C N and P in a soil catena of the shortgrass steppeEcology 66276ndash282 doi1023071941328
Sinsabaugh RL Antibus RK Linkins AE (1991) An enzymicapproach to the analysis of microbial activity during plant litterdecomposition Agric Ecosyst Environ 3443ndash54 doi1010160167-8809(91)90092-C
Tabatabai MA Bremner JM (1970) Arylsulfatase activity of soilsSoil Sci Soc Am Proc 34225ndash229
Trevors JT Mayfield CI Inniss WE (1982) Measurement of elec-tron transport system (ETS) activity in soil Microb Ecol 8163ndash168 doi101007BF02010449
Vetanovetz RP Peterson JC (1992) Effect of carbon source andnitrogen on urease activity in a Sphagnum peat mediumCommun Soil Sci Plant Anal 23379ndash388 doi10108000103629209368596
Yoo G Park E-J Kim S-H Lee H-J Kang S Lee D (2001)Transport and decomposition of leaf litter as affected by aspectand understory in a temperate hardwood forest Korean J BiolSci 5319ndash325
1143
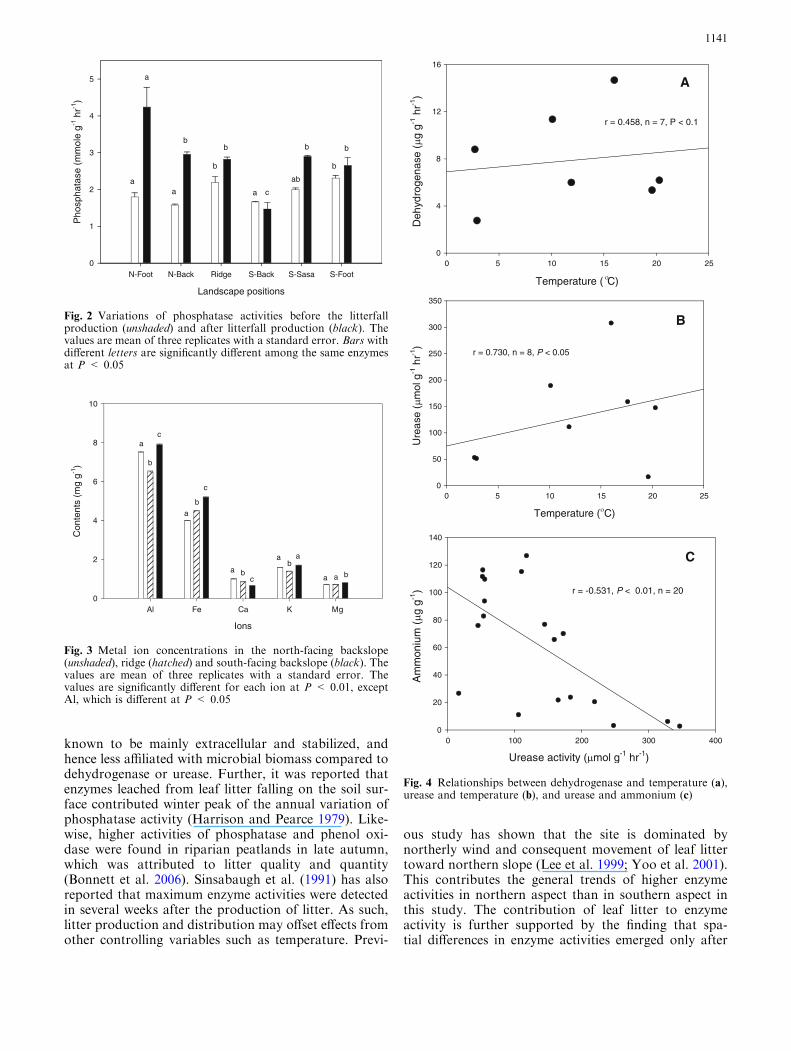
known to be mainly extracellular and stabilized andhence less affiliated with microbial biomass compared todehydrogenase or urease Further it was reported thatenzymes leached from leaf litter falling on the soil sur-face contributed winter peak of the annual variation ofphosphatase activity (Harrison and Pearce 1979) Like-wise higher activities of phosphatase and phenol oxi-dase were found in riparian peatlands in late autumnwhich was attributed to litter quality and quantity(Bonnett et al 2006) Sinsabaugh et al (1991) has alsoreported that maximum enzyme activities were detectedin several weeks after the production of litter As suchlitter production and distribution may offset effects fromother controlling variables such as temperature Previ-
ous study has shown that the site is dominated bynortherly wind and consequent movement of leaf littertoward northern slope (Lee et al 1999 Yoo et al 2001)This contributes the general trends of higher enzymeactivities in northern aspect than in southern aspect inthis study The contribution of leaf litter to enzymeactivity is further supported by the finding that spa-tial differences in enzyme activities emerged only after
Landscape positions
N-Foot N-Back Ridge S-Back S-Sasa S-Foot
Pho
spha
tase
(mm
ole
g-1hr
-1)
0
1
2
3
4
5 a
b
aa a
b b
bb
b
ab
c
Fig 2 Variations of phosphatase activities before the litterfallproduction (unshaded) and after litterfall production (black) Thevalues are mean of three replicates with a standard error Bars withdifferent letters are significantly different among the same enzymesat P lt 005
Ions
Al Fe Ca K Mg
Con
tent
s(m
gg-1
)
0
2
4
6
8
10
a
a
a
a
a
c
b
b
c
bc
ba
a b
Fig 3 Metal ion concentrations in the north-facing backslope(unshaded) ridge (hatched) and south-facing backslope (black) Thevalues are mean of three replicates with a standard error Thevalues are significantly different for each ion at P lt 001 exceptAl which is different at P lt 005
Temperature ( C)
0 5 10 15 20 25
Deh
ydro
gena
se(
gg-1
hr-1
)
0
4
8
12
16
A
r = 0458 n = 7 P lt 01
Temperature ( C)
0 5 10 15 20 25
Ure
ase
(m
olg-1
hr-1
)
0
50
100
150
200
250
300
350
B
r = 0730 n = 8 P lt 005
Urease activity ( mol g-1 hr-1)
0 100 200 300 400
Am
mon
ium
(g
g-1)
0
20
40
60
80
100
120
140
r = -0531 P lt 001 n = 20
C
Fig 4 Relationships between dehydrogenase and temperature (a)urease and temperature (b) and urease and ammonium (c)
1141
litterfall production (Fig 2) It is also interesting to notethat differences in enzyme activities between south-facing backslope without S borealis and south-facingbackslope with S borealis were only found after thelitterfall while such differences were absent beforethe litterfall production (Fig 2) Because areas withS borealis can maintain much larger amount of leaflitter than areas without S borealis understory vegeta-tion may influence biogeochemical processes in forestfloor through leaf litter distribution (Lee et al 1999)
Metal ions notably Fe and Al have reported toinhibit soil enzyme activities by absorbing to the activesite (Ekenler and Tabatabai 2002) In contrast Ca2+
and Mg2+ content often exhibited a positive correlationwith enzyme activities because those are known to re-move inhibition of humic materials by enzymendashhumicinteractions (Freeman et al 1996) Our results exhibitedthat two inhibitory ions (Al Fe) were higher whilst Mgand K contents were lower in soil of the southern-facingslope (Oh et al 1999) As such higher concentration ofinhibitory ions and lower concentration of activationions partially account for the lower enzyme activities inthe southern-facing slope
In this study we assessed the changes in enzymeactivities only However previous studies have revealedthat microbial composition such as bacteria versus fungimay vary over time (Moore-Kucera and Dick 2008Kubartova et al 2007) This would be closely related toenzyme activities because different types of microor-ganisms are responsible for the production of specificenzymes
Landscape position has been reported to affect soilproperties litter decomposition nitrogen dynamics andphosphorus availability (Brubaker et al 1994 Klem-mendson and Wienhold 1991 Mudrick et al 1994Garten Jr et al 1994) These studies focused on thedifferences in organic matter water and nutrient con-tents which were caused by flow of water and soil ero-sions In addition to these factors our results indicatedthat production and redistribution of leaf litter are otherkey factors affecting biogeochemical cycles in a forestfloor Furthermore not only the physical movement ofleaf litter mediated by wind but also understory vege-tation which may impede the movement could play animportant role in this dynamics
Acknowledgments H Kang is grateful to AEBRC EcoRiver21 forfinancial support
References
Aber JD Mellilo JM (1991) Terrestrial ecosystems SaundersCollege Publishing Philadelphia
Bonnett SAF Ostle N Freeman C (2006) Seasonal variations indecomposition processes in a valley-bottom riparian peatlandSci Total Environ 370561ndash573 doi101016jscitotenv200608032
Bremner JM Mulvaney RL (1978) Urease activity in soils InBurns RG (ed) Soil enzymes Academic Press New York
Brubaker SC Jones AJ Frank K Lewis DT (1994) Regressionmodels for estimating soil properties by landscape position SoilSci Soc Am J 581763ndash1767
Burns RG (1982) Enzyme activity in soil location and a possiblerole in microbial ecology Soil Biol Biochem 14423ndash427 doi1010160038-0717(82)90099-2
Chrost RJ (1991) Environmental control of the synthesis andactivity of aquatic microbial ectoenzymes In Chrost RJ (ed)Microbial enzymes in aquatic environments Springer BerlinHeidelberg New York pp 29ndash59
Dilly O Munch JC (1996) Microbial biomass content basal res-piration and enzyme activities during the course of decompo-sition of leaf litter in a black alder (Alnus glutinosa (L) Gaertn)forest Soil Biol Biochem 281073ndash1081 doi1010160038-0717(96)00075-2
Dilly O Nannipieri P (2001) Response of ATP content respirationrate and enzyme activities in an arable and a forest soil tonutrient addition Biol Fertil Soils 3464ndash72 doi101007s003740100375
Dilly O Bartsch S Rosenbrock P Buscot F Munch J-C (2001)Shifts in physiological capabilities of the microbiota during thedecomposition of leaf litter in a black alder (Alnus glutinosa(Gaertn) L) forest Soil Biol Biochem 33921ndash930 doi101016S0038-0717(00)00239-X
Dilly O Munch J-C Pfeiffer E-M (2007) Enzyme activities andlitter decomposition in agricultural soils in northern centraland southern Germany J Plant Nutr Soil Sci 170197ndash204 doi101002jpln200622044
Eivazi F Tabatabai MA (1990) Factors affecting glucosidase andgalactosidase activities in soils Soil Biol Biochem 22891ndash897doi1010160038-0717(90)90126-K
Ekenler M Tabatabai MA (2002) Effects of trace elements on b-glucosaminidase activity in soils Soil Biol Biochem 341829ndash1832 doi101016S0038-0717(02)00167-0
Emteryd O (1989) Chemical and physical analysis of inorganicnutrients in plant soil water and air Department of Forest SiteResearch Swedish University of Agricultural Sciences Umea
Eviner VT Chapin FS Vaughn CE (2006) Seasonal variations inplant species effects on soil N and P dynamics Ecology 87974ndash986 doi1018900012-9658(2006)87[974SVIPSE]20CO2
Freeman C Liska G Ostle NJ Lock MA Reynolds B Hudson J(1996) Microbial activity and enzymic decomposition processesfollowing peatland water table drawdown Plant Soil 180121ndash127 doi101007BF00015418
Garten CT Jr Huston MA Thoms CA (1994) Topographic vari-ation of soil nitrogen dynamics at Walker Branch watershedTennessee For Sci 40497ndash512
Harrison AF Pearce T (1979) Seasonal variation of phosphataseactivity in woodland soils Soil Biol Biochem 11405ndash410 doi1010160038-0717(79)90054-3
Kandeler E Gerber H (1988) Short-term assay of soil ureaseactivity using colorimetric determination of ammonium BiolFertil Soils 658ndash72 doi101007BF00257924
Klemmendson JO Wienhold BJ (1991) Aspect and species influ-ences on nitrogen and phosphorus availability in Arizonachaparral soils Soil Sci Soc Am J 551735ndash1740
Kubartova A Moukoumi J Beguiristain T Ranger J Berthelin J(2007) Microbial diversity during cellulose decomposition indifferent forest stands I Microbial communities and environ-mental conditions Microb Ecol 54393ndash405 doi101007s00248-007-9286-2
Lee D Yoo G Oh S Shim JH Kang S (1999) Significance ofaspect and understory type to leaf litter redistribution in atemperate hardwood forest Korean J Biol Sci 3143ndash147
Moore-Kucera J Dick RP (2008) PLFA profiling of microbialcommunity structure and seasonal shifts in soils of a Douglas-fir chronosequence Microb Ecol 55500ndash511 doi101007s00248-007-9295-1
Mudrick DA Hoosein M Hicks RR Jr Townsend EC (1994)Decomposition of leaf litter in an Appalachian forest effects ofleaf species aspect slope position and time For Ecol Manage68231ndash250
1142
Noorbakhsh S Schoenau J Si B Zeleke T Qian P (2008) Soilproperties yield and landscape relationships in south-centralSaskatchewan Canada J Plant Nutr 31539ndash556 doi10108001904160801895076
Oh S Yoo G Shim JH Kang S Lee D (1999) Spatial heterogeneityof soil K Ca Mg Fe and Al generated by leaf litter redistri-bution in a temperate hardwood forest Kor J Ecol 22205ndash210
Sariyildiz T Anderson JM Kucuk M (2005) Effects of tree speciesand topography on soil chemistry litter quality and decom-position in northeast Turkey Soil Biol Biochem 371695ndash1706doi101016jsoilbio200502004
Schimel D Stillwell MA Woodmansee RG (1985) Biogeochemis-try of C N and P in a soil catena of the shortgrass steppeEcology 66276ndash282 doi1023071941328
Sinsabaugh RL Antibus RK Linkins AE (1991) An enzymicapproach to the analysis of microbial activity during plant litterdecomposition Agric Ecosyst Environ 3443ndash54 doi1010160167-8809(91)90092-C
Tabatabai MA Bremner JM (1970) Arylsulfatase activity of soilsSoil Sci Soc Am Proc 34225ndash229
Trevors JT Mayfield CI Inniss WE (1982) Measurement of elec-tron transport system (ETS) activity in soil Microb Ecol 8163ndash168 doi101007BF02010449
Vetanovetz RP Peterson JC (1992) Effect of carbon source andnitrogen on urease activity in a Sphagnum peat mediumCommun Soil Sci Plant Anal 23379ndash388 doi10108000103629209368596
Yoo G Park E-J Kim S-H Lee H-J Kang S Lee D (2001)Transport and decomposition of leaf litter as affected by aspectand understory in a temperate hardwood forest Korean J BiolSci 5319ndash325
1143

litterfall production (Fig 2) It is also interesting to notethat differences in enzyme activities between south-facing backslope without S borealis and south-facingbackslope with S borealis were only found after thelitterfall while such differences were absent beforethe litterfall production (Fig 2) Because areas withS borealis can maintain much larger amount of leaflitter than areas without S borealis understory vegeta-tion may influence biogeochemical processes in forestfloor through leaf litter distribution (Lee et al 1999)
Metal ions notably Fe and Al have reported toinhibit soil enzyme activities by absorbing to the activesite (Ekenler and Tabatabai 2002) In contrast Ca2+
and Mg2+ content often exhibited a positive correlationwith enzyme activities because those are known to re-move inhibition of humic materials by enzymendashhumicinteractions (Freeman et al 1996) Our results exhibitedthat two inhibitory ions (Al Fe) were higher whilst Mgand K contents were lower in soil of the southern-facingslope (Oh et al 1999) As such higher concentration ofinhibitory ions and lower concentration of activationions partially account for the lower enzyme activities inthe southern-facing slope
In this study we assessed the changes in enzymeactivities only However previous studies have revealedthat microbial composition such as bacteria versus fungimay vary over time (Moore-Kucera and Dick 2008Kubartova et al 2007) This would be closely related toenzyme activities because different types of microor-ganisms are responsible for the production of specificenzymes
Landscape position has been reported to affect soilproperties litter decomposition nitrogen dynamics andphosphorus availability (Brubaker et al 1994 Klem-mendson and Wienhold 1991 Mudrick et al 1994Garten Jr et al 1994) These studies focused on thedifferences in organic matter water and nutrient con-tents which were caused by flow of water and soil ero-sions In addition to these factors our results indicatedthat production and redistribution of leaf litter are otherkey factors affecting biogeochemical cycles in a forestfloor Furthermore not only the physical movement ofleaf litter mediated by wind but also understory vege-tation which may impede the movement could play animportant role in this dynamics
Acknowledgments H Kang is grateful to AEBRC EcoRiver21 forfinancial support
References
Aber JD Mellilo JM (1991) Terrestrial ecosystems SaundersCollege Publishing Philadelphia
Bonnett SAF Ostle N Freeman C (2006) Seasonal variations indecomposition processes in a valley-bottom riparian peatlandSci Total Environ 370561ndash573 doi101016jscitotenv200608032
Bremner JM Mulvaney RL (1978) Urease activity in soils InBurns RG (ed) Soil enzymes Academic Press New York
Brubaker SC Jones AJ Frank K Lewis DT (1994) Regressionmodels for estimating soil properties by landscape position SoilSci Soc Am J 581763ndash1767
Burns RG (1982) Enzyme activity in soil location and a possiblerole in microbial ecology Soil Biol Biochem 14423ndash427 doi1010160038-0717(82)90099-2
Chrost RJ (1991) Environmental control of the synthesis andactivity of aquatic microbial ectoenzymes In Chrost RJ (ed)Microbial enzymes in aquatic environments Springer BerlinHeidelberg New York pp 29ndash59
Dilly O Munch JC (1996) Microbial biomass content basal res-piration and enzyme activities during the course of decompo-sition of leaf litter in a black alder (Alnus glutinosa (L) Gaertn)forest Soil Biol Biochem 281073ndash1081 doi1010160038-0717(96)00075-2
Dilly O Nannipieri P (2001) Response of ATP content respirationrate and enzyme activities in an arable and a forest soil tonutrient addition Biol Fertil Soils 3464ndash72 doi101007s003740100375
Dilly O Bartsch S Rosenbrock P Buscot F Munch J-C (2001)Shifts in physiological capabilities of the microbiota during thedecomposition of leaf litter in a black alder (Alnus glutinosa(Gaertn) L) forest Soil Biol Biochem 33921ndash930 doi101016S0038-0717(00)00239-X
Dilly O Munch J-C Pfeiffer E-M (2007) Enzyme activities andlitter decomposition in agricultural soils in northern centraland southern Germany J Plant Nutr Soil Sci 170197ndash204 doi101002jpln200622044
Eivazi F Tabatabai MA (1990) Factors affecting glucosidase andgalactosidase activities in soils Soil Biol Biochem 22891ndash897doi1010160038-0717(90)90126-K
Ekenler M Tabatabai MA (2002) Effects of trace elements on b-glucosaminidase activity in soils Soil Biol Biochem 341829ndash1832 doi101016S0038-0717(02)00167-0
Emteryd O (1989) Chemical and physical analysis of inorganicnutrients in plant soil water and air Department of Forest SiteResearch Swedish University of Agricultural Sciences Umea
Eviner VT Chapin FS Vaughn CE (2006) Seasonal variations inplant species effects on soil N and P dynamics Ecology 87974ndash986 doi1018900012-9658(2006)87[974SVIPSE]20CO2
Freeman C Liska G Ostle NJ Lock MA Reynolds B Hudson J(1996) Microbial activity and enzymic decomposition processesfollowing peatland water table drawdown Plant Soil 180121ndash127 doi101007BF00015418
Garten CT Jr Huston MA Thoms CA (1994) Topographic vari-ation of soil nitrogen dynamics at Walker Branch watershedTennessee For Sci 40497ndash512
Harrison AF Pearce T (1979) Seasonal variation of phosphataseactivity in woodland soils Soil Biol Biochem 11405ndash410 doi1010160038-0717(79)90054-3
Kandeler E Gerber H (1988) Short-term assay of soil ureaseactivity using colorimetric determination of ammonium BiolFertil Soils 658ndash72 doi101007BF00257924
Klemmendson JO Wienhold BJ (1991) Aspect and species influ-ences on nitrogen and phosphorus availability in Arizonachaparral soils Soil Sci Soc Am J 551735ndash1740
Kubartova A Moukoumi J Beguiristain T Ranger J Berthelin J(2007) Microbial diversity during cellulose decomposition indifferent forest stands I Microbial communities and environ-mental conditions Microb Ecol 54393ndash405 doi101007s00248-007-9286-2
Lee D Yoo G Oh S Shim JH Kang S (1999) Significance ofaspect and understory type to leaf litter redistribution in atemperate hardwood forest Korean J Biol Sci 3143ndash147
Moore-Kucera J Dick RP (2008) PLFA profiling of microbialcommunity structure and seasonal shifts in soils of a Douglas-fir chronosequence Microb Ecol 55500ndash511 doi101007s00248-007-9295-1
Mudrick DA Hoosein M Hicks RR Jr Townsend EC (1994)Decomposition of leaf litter in an Appalachian forest effects ofleaf species aspect slope position and time For Ecol Manage68231ndash250
1142
Noorbakhsh S Schoenau J Si B Zeleke T Qian P (2008) Soilproperties yield and landscape relationships in south-centralSaskatchewan Canada J Plant Nutr 31539ndash556 doi10108001904160801895076
Oh S Yoo G Shim JH Kang S Lee D (1999) Spatial heterogeneityof soil K Ca Mg Fe and Al generated by leaf litter redistri-bution in a temperate hardwood forest Kor J Ecol 22205ndash210
Sariyildiz T Anderson JM Kucuk M (2005) Effects of tree speciesand topography on soil chemistry litter quality and decom-position in northeast Turkey Soil Biol Biochem 371695ndash1706doi101016jsoilbio200502004
Schimel D Stillwell MA Woodmansee RG (1985) Biogeochemis-try of C N and P in a soil catena of the shortgrass steppeEcology 66276ndash282 doi1023071941328
Sinsabaugh RL Antibus RK Linkins AE (1991) An enzymicapproach to the analysis of microbial activity during plant litterdecomposition Agric Ecosyst Environ 3443ndash54 doi1010160167-8809(91)90092-C
Tabatabai MA Bremner JM (1970) Arylsulfatase activity of soilsSoil Sci Soc Am Proc 34225ndash229
Trevors JT Mayfield CI Inniss WE (1982) Measurement of elec-tron transport system (ETS) activity in soil Microb Ecol 8163ndash168 doi101007BF02010449
Vetanovetz RP Peterson JC (1992) Effect of carbon source andnitrogen on urease activity in a Sphagnum peat mediumCommun Soil Sci Plant Anal 23379ndash388 doi10108000103629209368596
Yoo G Park E-J Kim S-H Lee H-J Kang S Lee D (2001)Transport and decomposition of leaf litter as affected by aspectand understory in a temperate hardwood forest Korean J BiolSci 5319ndash325
1143

Noorbakhsh S Schoenau J Si B Zeleke T Qian P (2008) Soilproperties yield and landscape relationships in south-centralSaskatchewan Canada J Plant Nutr 31539ndash556 doi10108001904160801895076
Oh S Yoo G Shim JH Kang S Lee D (1999) Spatial heterogeneityof soil K Ca Mg Fe and Al generated by leaf litter redistri-bution in a temperate hardwood forest Kor J Ecol 22205ndash210
Sariyildiz T Anderson JM Kucuk M (2005) Effects of tree speciesand topography on soil chemistry litter quality and decom-position in northeast Turkey Soil Biol Biochem 371695ndash1706doi101016jsoilbio200502004
Schimel D Stillwell MA Woodmansee RG (1985) Biogeochemis-try of C N and P in a soil catena of the shortgrass steppeEcology 66276ndash282 doi1023071941328
Sinsabaugh RL Antibus RK Linkins AE (1991) An enzymicapproach to the analysis of microbial activity during plant litterdecomposition Agric Ecosyst Environ 3443ndash54 doi1010160167-8809(91)90092-C
Tabatabai MA Bremner JM (1970) Arylsulfatase activity of soilsSoil Sci Soc Am Proc 34225ndash229
Trevors JT Mayfield CI Inniss WE (1982) Measurement of elec-tron transport system (ETS) activity in soil Microb Ecol 8163ndash168 doi101007BF02010449
Vetanovetz RP Peterson JC (1992) Effect of carbon source andnitrogen on urease activity in a Sphagnum peat mediumCommun Soil Sci Plant Anal 23379ndash388 doi10108000103629209368596
Yoo G Park E-J Kim S-H Lee H-J Kang S Lee D (2001)Transport and decomposition of leaf litter as affected by aspectand understory in a temperate hardwood forest Korean J BiolSci 5319ndash325
1143
Copyright © 2022 FDOKUMEN




















