
ALTERATION OF SEAGRASS SPECIES COMPOSITION AND FUNCTION OVER TWO DECADES
Upload
independentCategory
view
1download
0
Variation in fish density, assemblage composition and relative rates of
predation among mangrove, seagrass and coral reef habitats
Paul M. Chittaro, Paolo Usseglio & Peter F. SaleDepartment of Biological Sciences, University of Windsor, 401 Sunset Blvd, N9B 3P4, Windsor, Ont., Canada(e-mail: [email protected])
Received 12 December 2003 Accepted 15 April 2004
Key words: nursery, assemblage, similarity
Synopsis
We tested the hypothesis for several Caribbean reef fish species that there is no difference in nurseryfunction among mangrove, seagrass and shallow reef habitat as measured by: (a) patterns of juvenile andadult density, (b) assemblage composition, and (c) relative predation rates. Results indicated that althoughsome mangrove and seagrass sites showed characteristics of nursery habitats, this pattern was weak. Whilealmost half of our mangrove and seagrass sites appeared to hold higher proportions of juvenile fish (allspecies pooled) than did reef sites, this pattern was significant in only two cases. In addition, only four ofthe six most abundant and commercially important species (Haemulon flavolineatum, Haemulon sciurus,Lutjanus apodus, Lutjanus mahogoni, Scarus iserti, and Sparisoma aurofrenatum) showed patterns of higherproportions of juvenile fish in mangrove and/or seagrass habitat(s) relative to coral reefs, and were limitedto four of nine sites. Faunal similarity between reef and either mangrove or seagrass habitats was low,suggesting little, if any exchange between them. Finally, although relative risk of predation was lower inmangrove/seagrass than in reef habitats, variance in rates was substantial suggesting that not all mangrove/seagrass habitats function equivalently. Specifically, relative risk varied between morning and afternoon,and between sites of similar habitat, yet varied little, in some cases, between habitats (mangrove/seagrass vs.coral reefs). Consequently, our results caution against generalizations that all mangrove and seagrasshabitats have nursery function.
Introduction
Coastal habitats such as coral reefs, mangrovesand seagrass beds are among the most diverse andproductive in the world (Alevizon & Brooks 1975,Heck et al. 2003). Therefore, there are importantscientific and economic reasons to determine whatrole these habitats play in vital ecosystem servicessuch as nutrient cycling, waste treatment and dis-turbance regulation (Costanza et al. 1997). Inaddition, it is important to understand how com-mercially important organisms use these habitatsand how their ecological interactions affect pro-ductivity elsewhere (Munro & Williams 1985).
Due to the dispersive larval and sedentary adultstages inherent to the life cycle of many fishes andcrustaceans there are many factors that can influ-ence the distribution and abundance of organismsamong coastal habitats. For instance, during thepelagic larval phase the distribution and abun-dance of sexually mature individuals (Robertsonet al. 1988), ocean circulation patterns (McIlwain1997, Roberts 1997, Shapiro et al. 1997, Joneset al. 1999, Swearer et al. 1999, Cowen et al. 2000)and larval settling preferences (Victor 1986, Rob-ertson et al. 1988, Leis et al. 2003) all affect theabundance and distribution of larvae. Followingsettlement, the abundance and distribution of
Environmental Biology of Fishes (2005) 72: 175–187 � Springer 2005

organisms is further restricted by competition forlimiting resources, responses to habitat cues (To-limieri 1998) and predation (Caley et al. 1996,Hixon & Carr 1997, Steele 1998). Depending onthe shallow-water habitat where larvae might set-tle, pronounced differences in mortality may oc-cur. This variation in mortality among habitatscan result from differences in quality and/orquantity of resources (Cocheret de la Moriniereet al. 2003) and differences in predator abundancesand/or the frequency of their visits (Shulman 1985,Parrish 1989, Holbrook & Schmitt 2003, Adamset al. 2004). In other words, the magnitude of theimpact that factors such as competition and pre-dation have on an organism’s survival will likelyvary with the habitat to which an individual set-tles. Consequently, certain habitats (referred to asnursery habitats) may increase the probability ofan individual’s survival through the provision offood and shelter as well as supplement nearbypopulations.Mangrove and seagrass habitats are shallow-
water habitats that are often observed to havegreater abundance of juvenile fishes relative tocoral reefs. Frequently, these habitats are referredto as nurseries because of the apparent increase injuvenile survival that these abundance patternssuggest (see Robertson & Duke 1987, Dennis 1992,Halliday & Young 1996, Gillanders 1997, Nagel-kerken et al. 2000a, b). However, Beck et al. (2001)advised that several factors such as growth, sur-vival, density, and movement must be consideredbefore nursery value can be confirmed.The purpose of this study was to assess whether
mangroves and seagrass beds of the Caribbean areimportant nursery habitats for reef fish species bytesting the null hypothesis that there was no dif-ference in nursery value among mangrove, sea-grass and coral reef habitats. We measured (a)patterns of juvenile and adult densities, (b) faunalcomposition, and (c) relative predation rates.
Methods
We conducted this study at three locations: (a) LeeStocking Island, Great Exuma, Bahamas, (b)Mahahual, Mexico, and (c) Turneffe Atoll, Belize.We performed visual censuses (described below) inBahamas during May 2001 and in Mexico and
Belize in June and August 2002, respectively. Wealso conducted tether timer experiments (describedbelow) in Belize during August 2003. We visuallycensused a minimum of one site, comprised of eachhabitat type (i.e. mangrove, seagrass and coral reefhabitats) at each of the three locations for abun-dances of juvenile and adult fishes. We determinedjuvenile and adult status from a combination ofsize range and color patterns based on publishedlife history data and our knowledge of the fauna.We censused seagrass and coral reef habitats atTurneffe Atoll, Belize, at each of six sites, three ofwhich included mangroves. At Lee Stocking Islandwe censused two sites of both mangrove and coralreef habitat, while in Mahahual we monitored onesite that included mangrove, seagrass and coralreef habitat.At each site, were recorded all fishes if observed
within 30 m2 (30 m · 1 m, 5–10 replicates) hap-hazardly located transects. Reef transects wereparallel to the reef crest, while seagrass transectswere within a seagrass bed, 5 m from any otherhabitat. Mangrove transects were parallel to shoreand adjacent to prop roots. Transect width inseagrass and coral reef habitats spanned 0.5 m toeither side of the transect tape, but due to theintricate nature of mangrove roots the transectwidth was 1 m from the centerline of the transecttape into the roots. Censused mangrove habitatswere 1.0–2.0 m deep. Seagrass habitats were 1.5–3.0 m deep. Coral reef habitats were 2.0–4.0 mdeep (7.0 m at Lee Stocking Island, site 2).
Density
To test the null hypothesis of no differences amonghabitats in terms of nursery potential, we com-pared the proportional abundance of juvenilesamong habitats. A habitat with more juvenilesrelative to adults (as expected in nursery habitats)will yield greater juvenile proportions compared toa habitat with more adults and/or fewer juveniles.We used univariate ANOVA’s to compare pro-portional abundance of juveniles among habitats(mangrove, seagrass and coral reef) at each siteand if necessary followed by Bonferroni post-hoctests. We investigated two levels of biological res-olution using this analysis. First, we compared theproportional abundance of juveniles among habi-tats at the assemblage level (i.e. pooling species
176

within habitats), followed by comparisons of theproportional abundance of juveniles among habi-tats for each of the most abundant and commer-cially important species. Data were arcsinetransformed to improve normality.
Assemblage composition
Assuming that mangrove and seagrass habitatsoperate as nurseries, then a high species overlap isexpected between these habitats and reefs as a re-sult of movements of fishes among the habitats.Here we tested the null hypothesis that species arenot shared among habitats (and hence a lack ofnursery potential) by examining similarity inassemblage structure among habitats using Jac-card’s coefficient of similarity. Specifically, Jac-card’s coefficient of similarity (Sj) compares faunasbetween reefs (a) and mangroves (b) and reefs (a)and seagrass beds (b) by comparing censuses fromeach pair of habitats:
Sj ¼c
aþ bþ c;
where a and b are the number of species presentonly in habitat A and B, respectively; and c is thenumber of species that are common to both hab-itats (Krebs 1989). Because rare species will pro-duce underestimates of faunal similarity werestricted the analyses to the most abundant andcommercially relevant species. Within a site, allpossible comparisons between five replicate cen-suses from mangrove, seagrass and coral reefhabitats would yield ten faunal similarity valuesper pair of habitats. Jaccard’s coefficient of simi-larity values range from 0.0, when the comparedcensuses have no species in common, to 1.0 whenall species are shared. Other diversity indices areavailable that take into account the abundance ofindividuals, but they were not used sincesubstantial variability in abundances likely exists,and our interest was in the composition of species.
Relative predation rates
Assuming that mangrove and seagrass habitats arenurseries then these habitats will likely have lowerrates of predation relative to that on reefs. Here wetested the null hypothesis of no difference in rela-tive predation rates between habitats by examining
the removal times of tethered fish. Specifically, atTurneffe Atoll, Belize, live juvenile Haemulonchrysargyreum (smallmouth grunt) ranging in sizefrom 3 to 6 cm total length, were used as baitfishin timed experiments within mangrove/seagrassbeds (mangrove and seagrass habitats were notmutually exclusive at the chosen sites and thusthey were combined into a ‘mangrove/seagrass’designation) and coral reefs at each of two sites(sites 2 and 3). We used H. chrysargyreum sincethey were observed in all habitats, were numeri-cally abundant and easy to catch. Experimentswere performed in the morning (09:00–11:00 h)and afternoon (14:00–16:30 h) and consisted ofusing fish tethered to activated stopwatch tethertimers to assess relative rates of predation (a) be-tween habitats (within sites and time of day), (b)between morning and afternoon (within sites andhabitat), and (c) between sites (within habitat andtime of day).The tether timer design was modified from that
used by Ha (1996) and Danilowicz & Sale (1999).In short, a stopwatch with chronometer was fixedinside a 30 ml nalgene container and a magneticswitch (i.e. reed switch and actuating magnet) wasattached to the start/stop function of the watch.Fish were tethered with a monofilament line (‘6 lbtest’), which in turn was connected to the start/stopswitch. Consequently, if a fish was taken the switchwould open, causing the watch to stop. Tethertimers were anchored in a given habitat byattaching them to a 30 cm long piece of 2 cmdiameter PVC pipe that was forced into the sub-stratum. Preliminary tests were conducted to en-sure that the movement of the baitfish would notresult in the accidental stopping of the clock. Weplaced eight tethers, each with baitfish that were6 cm in total length (maximum size used in exper-iments), and therefore those most likely capable ofdislodging the plate, in an area devoid of predators(large sand patch) and visually monitored for10 min with an observer 3 m away. Observationsconfirmed that baitfish removals or clock stop-pages were not the result of fish behavior.Each experiment started with the collection of
fish to be used as bait. Juveniles of Haemulonchrysargyreum were collected using a gill net (1 cmmesh), monitored for stress for 20 min and thenattached to a tether timer by placing a tiny hook(0.5 cm long) through the dorsal musculature.
177

Total length was recorded for each fish. The tethertimer was started and the timer with baitfish wasplaced in a given habitat. Tether timers were set upconcurrently in mangrove/seagrass and coral reefhabitats at one site, with no more than 60 minseparating the start time between habitats. Inmangrove/seagrass areas, timers were placedwithin a seagrass bed and near the margin of amangrove. On the reefs, tether timers were placedin sediment a maximum of 30 cm from a coralhead. We placed between 3 and 8 replicate tethertimers within each habitat; each replicate sepa-rated by a minimum of 4 m. Experiments wereconcluded at the end of 90 min, at which time weremoved all tether timers and recorded durationsfor those tethers that had lost their fish. Tetheringexperiments constrain behavior of the baitfish,therefore they can only assess relative risk of pre-dation.We did statistical analysis of tether timer dura-
tion data using survival analysis in the Statisticasoftware package (StatSoft, Inc. 2001). This com-pares the survival/failure times between two sam-ples taking into account censored and completeddata. Censored observations arise when thedependent variable represents the time to a ter-minal event, which does not always occur beforethe end of a fixed duration trial (90 min in thiscase). Fish still attached to a tether at the end of atrial yield censored observations while those re-moved by predators yield completed observations.Pairwise comparisons were conducted using Cox’sF-test between (a) habitats (within site and time ofday), (b) morning and afternoon (within site andhabitat), and (c) sites (within habitat and time ofday).
Results
In total, we censused approximately 7200 indi-viduals of 82 species. Common species, accountingfor 76% of the total sample, wereHalichoeres poeyi(11.9% of total abundances), Lutjanus apodus(11.5%), Haemulon flavolineatum (10.4%), Scarusiserti (9.6%), Family Atherinidae (8.5%), H. bi-vittatus (5.5%), Stegastes leucostictus (4.4%), H.sciurus (3.9%), L. mahogoni (3.4%), Abudefdufsaxatilis (2.8%), Sparisoma aurofrenatum (2.2%),and H. garnoti (2.2%).
Density
Due to the difficulty in differentiating juvenile andadults of the family Atherinidae we did not includethem in this analysis. Results indicated that thenull hypothesis of no differences among habitats interms of the proportional abundance of juvenileswas rejected, but did not suggest that mangrovesor seagrass habitats provide nursery function.Specifically, univariate ANOVAs showed only twoof nine sites with significantly higher proportionalabundance of juveniles in seagrass (Turneffe Atollsite 2; F1,10 ¼ 5.94, p < 0.05) or mangrove (LeeStocking Island site 1; F1,8 ¼ 7.07, p < 0.05)habitats relative to coral reefs (Figure 1). How-ever, we observed significantly higher proportionalabundance of juveniles in mangroves compared toseagrass habitats (but not reef habitats) at Turne-ffe Atoll site 6 (F2,15 ¼ 5.14, p < 0.05) andMahahual site 1 (F2,20 ¼ 3.51, p < 0.05). Alter-natively, significantly lower proportional abun-dance of juveniles were indicated in mangrovecompared to reef and seagrass habitats at TurneffeAtoll site 3(F2,13 ¼ 8.26, p < 0.01).Analyses of the densities of juveniles and adults
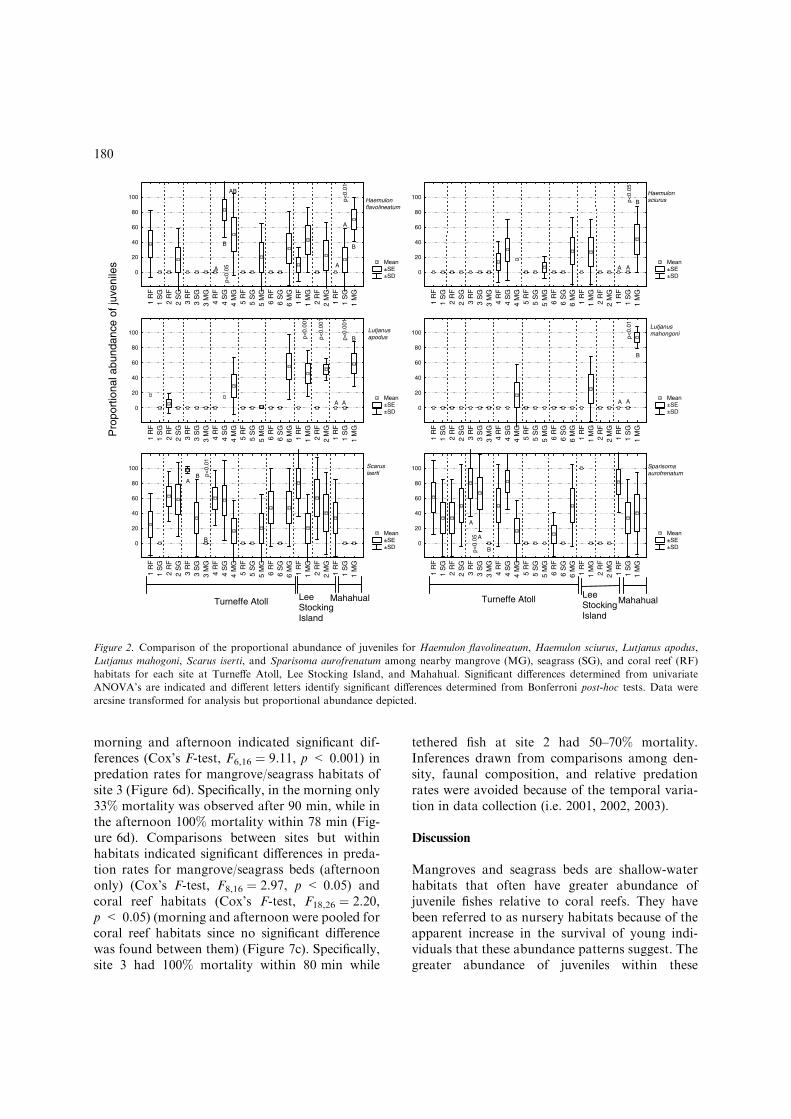
for the six most abundant and commerciallyimportant species (H. flavolineatum, H. sciurus, L.apodus, L. mahogoni, S. iserti, and Sp. aurofrena-tum) revealed only seven of 54 instances (six spe-cies and nine sites) where the proportionalabundance of juveniles were significantly higher inmangroves and/or seagrass habitat(s) than oncoral reefs. Specifically, H. flavolineatum, H. sciu-rus, L. apodus, and L. mahogoni showed signifi-cantly higher proportional abundance of juvenilesin mangroves and/or seagrass habitat(s) relative tocoral reef habitats, at Mahuhual site 1(F2,14 ¼ 8.67, p < 0.01; F2,14 ¼ 5.99, p < 0.05;F2,14 ¼ 31.13, p < 0.001; F2,14 ¼ 173.01,p < 0.01, respectively) (Figure 2). This same pat-tern was also observed for H. flavolineatum atTurneffe Atoll site 4 (F2, 16 ¼ 7.41, p < 0.05) andL. apodus at Lee Stocking Island sites 1 and 2(F1,8 ¼ 21.41, p < 0.001 and F1,8 ¼ 138.67, p <0.001, respectively). However, Scarus iserti and Sp.aurofrenatum showed the opposite pattern, signif-icantly lower juvenile proportions in mangrovescompared to coral reef and seagrass habitats, atTurneffe Atoll site 3 (F2,14 ¼ 12.31, p < 0.01 andF2,14 ¼ 6.89, p < 0.05, respectively).
178

Assemblage composition
An asymptote in cumulative species was observedfor each habitat as additional transects were in-cluded. This asymptote indicates that we ade-quately characterized the fauna of each habitatand thus permitted comparisons of faunas betweenhabitats. Results indicated that the null hypothesisof all (or greater than 50%) species shared amonghabitats was rejected. Jaccard’s coefficient of sim-ilarity indicated that relatively few species were incommon between reef and mangrove(Cij ¼ 0.22 ± 0.18) and reef and seagrass(Cij ¼ 0.23 ± 0.20) comparisons when only the 11most abundant species (A. saxatilis, Ha. bivittatus,Ha. garnoti, Ha. poeyi, H. flavolineatum, H. sciu-rus, L. apodus, L. mahogoni, S. iserti, Sp. auro-frenatum, and St. leucostictus) were considered(Figure 3a). Further refining the analysis to in-clude only those species that were both abundantand commercially important (H. flavolineatum, H.sciurus, L. apodus, L. mahogoni, S. iserti, and Sp.aurofrenatum), indicated an average Jaccard simi-
larity of 0.25 (±0.23) and 0.20 (±0.29) for com-parisons between reef and mangrove and reef andseagrass, respectively (Figure 3b).
Relative predation rates
Results indicated that the null hypothesis of nodifference in relative predation rate between habi-tats was rejected, although trials in mangrove/seagrass beds had, on average, relatively higherjuvenile survival (42%) than those on coral reefs(15%) after 90 minutes of being tethered (Figure 4).Significant differences (Cox’s F-test, F6,12 ¼ 8.83,p < 0.001) in predation rates were observed be-tween morning trials in mangrove/seagrass andcoral reef habitats at site 3 (Figure 5c). Specifically,in the coral reef at site 3 there was 100% mortalitywithin 80 min, while only 33% mortality after90 min in mangrove/seagrass habitat (Figure 5c).Also important was the number of comparisons(three of four) that showed a lack of significantvariability among habitats, within a site and time(Figure 5a, b and d). Comparisons between
Figure 1. Comparison of the proportional abundance of juveniles among nearby sets of mangrove (MG), seagrass (SG) and coral reef
(RF) habitats for each site at Turneffe Atoll, Lee Stocking Island, and Mahahual. Significant differences determined from univariate
ANOVA’s are indicated and different letters identify significant differences determined from Bonferroni post-hoc tests. Data were
arcsine transformed for analysis but proportional abundance depicted.
179
morning and afternoon indicated significant dif-ferences (Cox’s F-test, F6,16 ¼ 9.11, p < 0.001) inpredation rates for mangrove/seagrass habitats ofsite 3 (Figure 6d). Specifically, in the morning only33% mortality was observed after 90 min, while inthe afternoon 100% mortality within 78 min (Fig-ure 6d). Comparisons between sites but withinhabitats indicated significant differences in preda-tion rates for mangrove/seagrass beds (afternoononly) (Cox’s F-test, F8,16 ¼ 2.97, p < 0.05) andcoral reef habitats (Cox’s F-test, F18,26 ¼ 2.20,p < 0.05) (morning and afternoon were pooled forcoral reef habitats since no significant differencewas found between them) (Figure 7c). Specifically,site 3 had 100% mortality within 80 min while
tethered fish at site 2 had 50–70% mortality.Inferences drawn from comparisons among den-sity, faunal composition, and relative predationrates were avoided because of the temporal varia-tion in data collection (i.e. 2001, 2002, 2003).
Discussion
Mangroves and seagrass beds are shallow-waterhabitats that often have greater abundance ofjuvenile fishes relative to coral reefs. They havebeen referred to as nursery habitats because of theapparent increase in the survival of young indi-viduals that these abundance patterns suggest. Thegreater abundance of juveniles within these
Mean ±SE ±SD
1 R
F1
SG
2 R
F2
SG
3 R
F3
SG
3 M
G4
RF
4 S
G4
MG
5 R
F5
SG
5 M
G6
RF
6 S
G6
MG
1 R
F1
MG
2 R
F2
MG
1 R
F1
SG
1 M
G
0
20
40
60
80
100
p<0.
05A
B B
AB
p<0.
01
A
A
Haemulon flavolineatum
Mean ±SE ±SD
1 R
F1
SG
2 R
F2
SG
3 R
F3
SG
3 M
G4
RF
4 S
G4
MG
5 R
F5
SG
5 M
G6
RF
6 S
G6
MG
1 R
F1
MG
2 R
F2
MG
1 R
F1
SG
1 M
G
0
20
40
60
80
100
p<0.
05
A A
B
Haemulon sciurus
Mean ±SE ±SD
1 R
F1
SG
2 R
F2
SG
3 R
F3
SG
3 M
G4
RF
4 S
G4
MG
5 R
F5
SG
5 M
G6
RF
6 S
G6
MG
1 R
F1
MG
2 R
F2
MG
1 R
F1
SG
1 M
G
0
20
40
60
80
100 Lutjanus apodusp<
0.00
1
p<0.
001
p<0.
001
A A
B
Mean ±SE ±SD
1 R
F1
SG
2 R
F2
SG
3 R
F3
SG
3 M
G4
RF
4 S
G4
MG
5 R
F5
SG
5 M
G6
RF
6 S
G6
MG
1 R
F1
MG
2 R
F2
MG
1 R
F1
SG
1 M
G
0
20
40
60
80
100Lutjanus mahongoni
p<0.
01
AA
B
Mean ±SE ±SD
1 R
F1
SG
2 R
F2
SG
3 R
F3
SG
3 M
G4
RF
4 S
G4
MG
5 R
F5
SG
5 M
G6
RF
6 S
G6
MG
1 R
F1
MG
2 R
F2
MG
1 R
F1
SG
1 M
G
0
20
40
60
80
100 Scarusisertip<
0.01
AB
B Mean ±SE ±SD
1 R
F1
SG
2 R
F2
SG
3 R
F3
SG
3 M
G4
RF
4 S
G4
MG
5 R
F5
SG
5 M
G6
RF
6 S
G6
MG
1 R
F1
MG
2 R
F2
MG
1 R
F1
SG
1 M
G
0
20
40
60
80
100 Sparisomaaurofrenatum
p<0.
05
A
A
B
Pro
port
iona
l abu
ndan
ce o
f juv
enile
s
Turneffe Atoll Turneffe AtollLeeStockingIsland
LeeStockingIsland
Mahahual Mahahual
Figure 2. Comparison of the proportional abundance of juveniles for Haemulon flavolineatum, Haemulon sciurus, Lutjanus apodus,
Lutjanus mahogoni, Scarus iserti, and Sparisoma aurofrenatum among nearby mangrove (MG), seagrass (SG), and coral reef (RF)
habitats for each site at Turneffe Atoll, Lee Stocking Island, and Mahahual. Significant differences determined from univariate
ANOVA’s are indicated and different letters identify significant differences determined from Bonferroni post-hoc tests. Data were
arcsine transformed for analysis but proportional abundance depicted.
180

habitats has been attributed to several features thatcould enhance survivorship. For instance, greaterabundance of food and food types for developingorganisms (Odum & Heald 1972, Stunz et al. 2002,Cocheret de la Moriniere et al. 2003); complexity,shade and/or turbidity-reduced visibility inherentto the mangrove and seagrass habitats may reducethe efficiency of predation (complexity – Quinn &Kojis 1985, Parrish 1989, Cocheret de la Moriniereet al. 2002, shade – Cocheret de la Moriniere et al.2002, visibility/turbidity – Blaber & Blaber 1980);or predators may frequent mangrove and seagrasshabitats less than coral reefs (Shulman 1985, Par-rish 1989). To confirm whether a habitat is anursery Beck et al. (2001) proposed the examina-tion of several factors, such as density, survival,growth and movement among several habitats that
juveniles occupy. For this study we assessednursery value among three habitats (coral reefs,mangroves, and seagrass beds) at several sitesamong three locations (Belize, Bahamas, andMexico) by using density, faunal composition, andrelative mortality (limited to one location).Assuming that density can be used as an
approximation of the significance of a habitat to agiven life history stage of a species, we can thenassess the relative importance of various habitatsby comparing density among habitats (Dennis1992). Consequently, if mangroves or seagrassbeds have fewer juveniles than other habitats theywould not be considered critical nursery area for agiven species. For instance, Dennis (1992) workingin Puerto Rico, noted that mangroves may not beimportant nurseries since equal or greater
Mean ±SE ±SD
1 2 3 4 5 6 1 2 1
0.0
0.2
0.4
0.6
0.8
1.0
A) REEF vs. MANGROVE: most abundant species
Mean ±SE ±SD
1 2 3 4 5 6 1 2 1
0.0
0.2
0.4
0.6
0.8
1.0
A) REEF vs. SEAGRASS: most abundant species
Mean ±SE ±SD
1 2 3 4 5 6 1 2 1
0.0
0.2
0.4
0.6
0.8
1.0
B) REEF vs MANGROVE: most abundant commercially important species
Mean ±SE ±SD
1 2 3 4 5 6 1 2 1
0.0
0.2
0.4
0.6
0.8
1.0
B) REEF vs SEAGRASS: most abundant commercially important species
Jacc
ard'
s co
effic
ient
of s
imila
rity
Turneffe Atoll Turneffe AtollLee StockingIsland
Lee StockingIsland
Mahahual Mahahual
Figure 3. Jaccard’s coefficient of similarity for reef and mangrove and reef and seagrass comparisons for each site at Turneffe Atoll,
Lee Stocking Island, and Mahahual: (a) represents similarity values produced from habitat comparisons using only the 11 most
abundant species; (b) represents similarity values produced from habitat comparisons using six most abundant and commercially
important species.
181

Per
cent
aliv
e or
dea
d
Dead Alive
AM PM AM PM
Mangrove/seagrass Coral reef
0
20
40
60
80
100A) Site 2
Per
cent
aliv
e or
dea
d
Dead Alive
AM PM AM PM
Mangrove/seagrass Coral reef
0
20
40
60
80
100
A) Site 3p < 0.05betweenam & pm
Figure 4. Percent of baitfish alive or dead at the end of 90-min experiments for each habitat, time of day, and site. (a) Calabash Caye
and (b) Blackbird Caye.
0:00 0:14 0:28 0:43 0:57 1:12 1:26 1:40
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Complete CensoredA) Site 2 - A.M.
Mangrove/Seagrass bed Coral Reef
0:00 0:14 0:28 0:43 0:57 1:12 1:26 1:40-0.1
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Complete Censored
Mangrove/Seagrass Beds Coral Reef
B) Site 2 - P.M.
0:00 0:14 0:28 0:43 0:57 1:12 1:26 1:40
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0C) Site 3 - A.M.: Significant at p < 0.01
Complete Censored Mangrove/Seagrass Bed Coral Reef
0:00 0:14 0:28 0:43 0:57 1:12 1:26 1:40
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
D) Site 3 - P.M.Complete Censored
Mangrove/Seagrass Bed Coral Reef
Cum
ulat
ive
prop
ortio
n su
rviv
ing
Cum
ulat
ive
prop
ortio
n su
rviv
ing
Time (minutes)Time (minutes)
Figure 5. Results of tether timer experiments examining cumulative proportion of individuals surviving between mangrove/seagrass
and coral reef habitats for each of Blackbird Caye (a) am and (b) pm, and Calabash Caye (c) am and (d) pm. Complete and censored
data defined in text.
182
numbers of young fish were found in nearby sea-grass and coral reef habitats compared to man-grove and lagoon habitats. In addition, a reviewby Heck et al. (2003), that evaluated the nurseryrole of seagrass beds, noted that approximatelyhalf of the species studied had abundances inseagrass beds that were not significantly greaterthan those of other habitats in surrounding areas.Similarly, work in Curacao by Nagelkerken & vander Velde (2002) indicated that non-estuarinemangroves had equivalent juvenile densities com-pared to that on coral reefs. Our examination ofjuvenile and adult densities in mangrove, seagrassand coral reef habitats at sites in several Carib-bean locations, indicated that densities of juvenileswere not always greater in mangrove and seagrassbeds and were rarely significantly greater. Conse-quently, generalizations about seagrass and man-grove habitats having nursery related functionsshould be avoided if estimates of density are theonly method to confirm nursery potential. How-ever, additional methods should be used inconjunction with density to more accurately assess
whether a habitat operates as a nursery (Becket al. 2003).If a habitat functions as a nursery, then move-
ment of fish must occur between it and nearbyadult populations, thus resulting in assemblagestructure being relatively similar between thesehabitats. For instance, studies conducted in Cura-cao and Florida by Nagelkerken et al. (2000a) andMullin (1995), respectively, observed a relativelylarger number of fish species shared between hab-itats (mangrove and reef in Curacao and mangroveand adjacent open water for Florida) suggestingthe movement between habitats. However, work byThayer et al. (1987) in Florida and Morton (1990)in Australia found relatively few species in com-mon between mangrove and adjacent habitats (alsosee Laroche et al. 1997). Similarly, Blaber andMilton (1990) noted that the role of estuaries asnurseries for coral reef fish in the Solomon Islandswas insignificant since only 8–9% of fish numberswere juveniles of reef species. The results of ourstudy indicated that relatively few species wereconsistently shared between coral reefs and
0:00 0:14 0:28 0:43 0:57 1:12 1:26 1:40
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
A) Site 2 - Coral Reef
Complete Censored
AM PM
0:00 0:14 0:28 0:43 0:57 1:12 1:26 1:40
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Complete Censored
AM PM
B) Site 2 - Mangrove/Seagrass Beds
0:00 0:14 0:28 0:43 0:57 1:12 1:26 1:40
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Complete Censored AM PM
C) Site 3 - Coral Reef
0:00 0:14 0:28 0:43 0:57 1:12 1:26 1:40
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Complete Censored
D) Site 3 - Mangrove/Seagrass BedsSignificant at p < 0.001
AM PM
Cum
ulat
ive
prop
ortio
n su
rviv
ing
Cum
ulat
ive
prop
ortio
n su
rviv
ing
Time (minutes)Time (minutes)
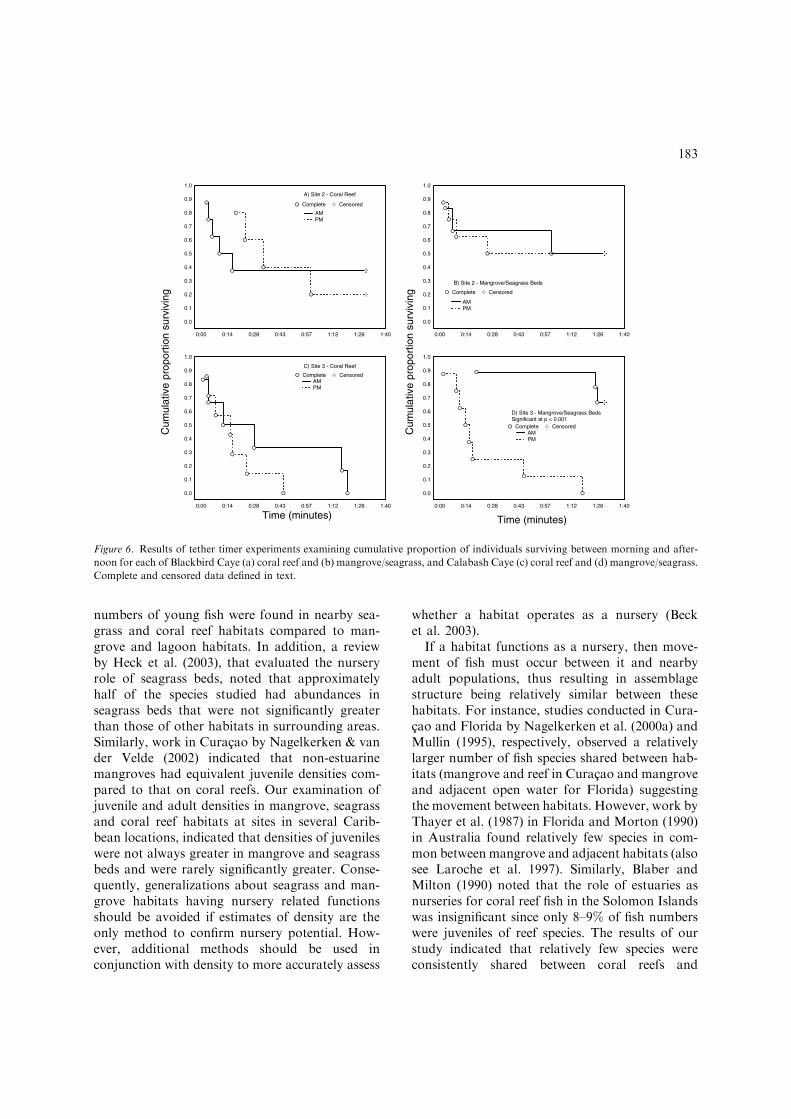
Figure 6. Results of tether timer experiments examining cumulative proportion of individuals surviving between morning and after-
noon for each of Blackbird Caye (a) coral reef and (b) mangrove/seagrass, and Calabash Caye (c) coral reef and (d) mangrove/seagrass.
Complete and censored data defined in text.
183

mangroves or seagrass beds, regardless of theirrelative abundances or commercial importance(Figure 3) and therefore questions of whethermangrove and seagrass habitats function as nurs-eries may be limited to a few species. Consequently,we agree with conclusions made by Thollot (1992)that connectivity between coral reefs and man-
groves or seagrass beds can provide benefits to thereef fish fauna (i.e. food and shelter for juveniles,additional space and feeding for adults) but they donot seem critical to the maintenance of the diverseand complex fish assemblages on coral reefs.Population dynamics of many species are often
influenced by behavioral responses to ecological
0:00 0:14 0:28 0:43 0:57 1:12 1:26 1:40
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Complete Censored BLACKBIRD CALABASH
A) Mangrove/Seagrass Beds (am only)
0:00 0:14 0:28 0:43 0:57 1:12 1:26 1:40
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Complete Censored BLACKBIRD CALABASH
B) Mangrove/Seagrass Beds (pm only)Significant at p < 0.05
0:00 0:14 0:28 0:43 0:57 1:12 1:26 1:40
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1.0
Complete Censored Blackbird Caye Calabash Caye
C) Coral Reef (pooled am and pm)Significant at p < 0.05
Cum
ulat
ive
prop
ortio
n su
rviv
ing
Time (minutes)
Figure 7. Results of tether timer experiments examining cumulative proportion of individuals surviving between sites (Blackbird Caye
and Calabash Caye) for (a) mangrove/seagrass morning, (b) mangrove/seagrass afternoon, and (c) coral reef pooled morning and
afternoon. Complete and censored data defined in text.
184

processes and therefore habitat use, for example,can often reflect behavioral decisions associatedwith the need to find resources, avoid predators,and/or reproduce (Dalhgren & Eggleston 2000).Assuming that shallow water habitats such asmangrove and seagrass beds are in fact nurseryhabitats, what explanation(s) is/are available tounderstand why juveniles use these habitats.Hypotheses offered to explain the use of nurseryhabitats include those that suggest that reducedpredator efficiency and/or reduced predator abun-dances influence juvenile survivorship and hencetheir numbers (see Shulman 1985, Parrish 1989,Robertson & Duke 1990, Cocheret de la Moriniereet al. 2002). Based on tether experiments at LeeStocking Island, Bahamas, Dahlgren & Eggleston(2000) indicated that algal beds had lower levels ofmortality for Nassau grouper, Epinephelus striatus,and thus suggested why juveniles occupy thesehabitats. Kramer (1991), working on Californiahalibut, Paralichthys californicus, in shallowcoastal and bay habitats of California, indicatedthat juveniles had higher survival during the firstyear of life if they settled along the coast and en-tered and used the bays as nursery areas. However,several studies have argued that seagrass or man-grove habitats do not necessarily function asnursery habitat. For example, Blaber & Milton(1990), working in Solomon Islands, and Morton(1990), in Eastern Australia, suggested that therewas likely little reduction in predation pressurebetween an estuary and coral reefs and betweenmangroves and adjacent habitats, respectively,since top-level piscivores were present in bothhabitats. In addition, work in Senegal by Vidy(2000), showed a lack of support of mangroves asnurseries since there was relatively high abundanceand predation pressure by a cichlid, Hemichromisfasciatus, on young fish, shrimps and crabs.In the present study, we assessed relative pre-
dation rates between mangrove/seagrass and coralreef habitats and found that although juvenilesurvivorship was, on average, 2.7 times greater inmangrove/seagrass habitats (regardless of time ofday) (Figure 4) statements of their nursery poten-tial were avoided. Specifically, the nursery functionof mangroves/seagrass habitats was unclear sincemortality of young individuals varied significantlybetween sites and time of day. Similarly, Bullard &Hay (2002) observed highly variable mortality in
tethered zooplankton within and between coralreef and seagrass habitats, and as a result boththeir study and ours urge that generalizationsregarding the nursery potential of certain habitatsbe reconsidered. This conclusion also identifies theneed of similar studies to be conducted that sub-stantially replicate habitats and thus better assessnursery function.From this study, information regarding the
nursery potential of mangrove and seagrass habi-tats was assessed relative to that of coral reefsacross three locations in the Caribbean. Resultsindicated that based on density, assemblage com-position and relative rates of predation, not allmangrove and seagrass beds appear to offernursery function. Consequently, generalizationsabout the nursery role should be avoided.Clearly, mangroves and seagrass beds may act
as important nurseries for some species and insome locations, but these habitats also have anumber of other intrinsic values, such as sup-porting their own fisheries, controlling sedimentmovement, wave action, and coastal erosion. Theoverall value of a particular mangrove or seagrassbed is likely to depend on a number of factorsincluding its location relative to reefs, other coastalhabitats, and open water, and that value may notalways include functioning as a nursery habitat. Amore cautious and explicit analysis of ecologicalroles of such habitats is required.
Acknowledgements
Special thanks to D. Hogan and C. Mora forinvaluable field assistance, to D. Campbell-Smith(Blue Ha Diving, Mahahual, Mexico) and E.Garcia (Institute of Marine Studies, CalabashCaye, Belize) for logistic support and to S. Bu-dinsky and M. Enns for assistance with the designand production of the tether timers. Financialsupport was provided to P.F. Sale by DonnerFoundation (grant # C29-99 and C17-99), NSERC(grant # 227965) and to P.M. Chittaro by OGSST(2001-2003) and OGS (2003-2004).
References
Adams, A.J., J.V. Locascio & B.D. Robbins. 2004. Microhab-
itat use by a post-settlement stage estuarine fish: Evidence
185

from relative abundance and predation among habitats. J.
Exp. Marine Biol. Ecol. 299: 17–33.
Alevizon, W.S. & M.G. Brooks. 1975. The comparative struc-
ture of two western Atlantic reef fish assemblages. Bull.
Marine Sci. 25: 482–490.
Beck, M.W., K.L. Heck, K.W. Able, D.L. Childers, D.B.
Efleston, B.M. Gillanders, B. Halpern, C.G. Hays, K.
Hoshino, T.J. Minello, R.J. Orth, P.F. Sheridan & M.P.
Weinstein. 2001. The identification, conservation, and man-
agement of estuarine and marine nurseries for fish and
invertebrates. BioScience 51: 633–641.
Blaber, S.J.M. & T.G. Blaber. 1980. Factors affecting the dis-
tribution of juvenile estuarine and inshore fish. J. Fish Biol.
17: 143–162.
Blaber, S.J.M. & A. Milton. 1990. Species composition, com-
munity structure and zoogeography of fishes of mangrove
estuaries in the Solomon Islands. Marine Biol. 105: 259–267.
Bullard, S.G. & M.E. Hay 2002. Plankton tethering to assess
spatial patterns of predation risk over a coral reef and sea-
grass bed. Marine Ecol. Progr. Series 225: 17–28.
Caley, M.J., M.H. Carr, M.A. Hixon, T.P. Hughes, G.P. Jones &
B.A.Menge 1996. Recruitment and the local dynamics of open
marine populations. Ann. Rev. Ecol. Systemat. 27: 477–500.
Cocheret de la Moriniere, E., I. Nagelkerken, H. van der Meij
& G. van der Velde. 2002. What attracts nursery fish to
mangroves: Habitat complexity or shade? pp. 117–130. In: E.
Cocheret de la Moriniere (ed.) Post-settlement Life Cycle
Migrations of Reef Fish in the Mangrove-Seagrass Coral
Reef Continuum. Febodruk BV, Enschede.
Cocheret de la Moriniere, E., B.J.A. Pollux, I. Nagelkerken,
M.A. Hemminga, A.H.L. Huiskes & G. van der Velde. 2003.
Ontogenetic dietary changes or coral reef fishes in the man-
grove-seagrass-reef continuum: Stable isotopes and gut-con-
tent analysis. Marine Ecol. Progr. Series 246: 279–289.
Costanza, R., R. d’Arge, R. de Groot, S. Farber, M. Grasso, B.
Hannon, K. Limburg, S. Naeem, R. O’Neill, J. Paruelo, R.G.
Raskin, P. Sutton & M. van den Belt. 1997. The value of the
world’s ecosystem services and natural capital. Nature 387:
253–260.
Cowen, R.K., K.M.M. Lwiza, S. Sponaugle, C.B. Paris & D.B.
Olson. 2000. Connectivity of marine populations: Open or
closed. Science 287: 857–859.
Dahlgren, C.P. & D.B. Eggleston. 2000. Ecological processes
underlying ontogenetic habitat shifts in a coral reef fish.
Ecology 81: 2227–2240.
Danilowicz, B.S. & P.F. Sale. 1999. Relative intensity of pre-
dation on the French grunt, Haemulon flavolineatum, during
diurnal, dusk, and nocturnal periods on a coral reef. Marine
Biol. 133: 337–343.
Dennis, G.D. 1992. Island mangrove habitats as spawning and
nursery areas for commercially important fishes in the
Caribbean. Proceedings of the 41st Gulf and Caribbean
Fisheries Institute. pp. 205–225.
Gillanders, B.M. 1997. Comparison of growth rates between
estuarine and coastal reef populations of Achoerodus viridis
(Pisces: Labridae). Marine Ecol. Progr. Series 146: 283–287.
Ha, D.S. 1996. An investigation of timed-tethering: A method
for study of coral reef piscivory. M.Sc. Thesis, University of
Windsor. 80 pp.
Halliday, I.A. & W.R.Young. 1996. Density, biomass and
species composition of fish in a subtropical Rhizophora styl-
osa mangrove forest. Marine Freshwater Res. 47: 609–615.
Heck, J.L., G. Hays & R.J. Orth. 2003. Critical evaluation of
the nursery role hypothesis for seagrass meadows. Marine
Ecol. Progr. Series 253: 123–136.
Hixon, M.A. & M.H. Carr. 1997. Synergistic predation, density
dependence, and population regulation in marine fish. Sci-
ence 277: 946–949.
Holbrook, S.J. & R.J. Schmitt. 2003. Spatial and temporal var-
iation in morality of newly settled damselfish: Patterns, causes
and co-variation with settlement. Oecologia 135: 532–541.
Jones, G.P., M.J. Millcich, M.J. Emsile & C. Lunow. 1999. Self
recruitment in a coral reef population. Nature 402: 802–804.
Kramer, S.H. 1991. Growth, mortality, and movements of
juvenile California halibut Paralichthys californicus in shal-
low coastal and bay habitats of San Diego County, Califor-
nia. Fishery Bull. 89: 195–207.
Krebs, C.J. 1989. Ecological Methodology. Harper and Row,
New York, New York. 654 pp.
Laroche, J., E. Baran & N.B. Rasoanandreasana. 1997. Tem-
poral patterns in a fish assemblage of a semiarid mangrove
zone in Madagascar. J. Fish Biol. 51: 3–20.
Leis, J.M., B.M. Carson-Ewart, A.C. Hay & D.H. Cato. 2003.
Coral-reef sounds enable nocturnal navigation by some reef-
fish larvae in some places and at some times. J. Fish Biol. 63:
724–737.
McIlwain, J.L. 1997. Hydrodynamic flows and the flux of larval
fishes across the crest of Ningaloo reef, western Australia.
Proceedings of the 8th International Coral Reef Symposium
2: 1133–1138.
Morton, R.M. 1990. Community structure, density and stand-
ing crop of fishes in a subtropical Australian mangrove area.
Marine Biol. 105: 385–394.
Mullin, S.J. 1995. Estuarine fish populations among red man-
grove prop roots of small overwash islands. Wetlands 15:
324–329.
Munro, J.L. & D. Mc B. Williams. 1985. Assessment and
management of coral reef fisheries: Biological, environmental
and socio-economic aspects. Proceedings of the 5th Interna-
tional Coral Reef Congress 4: 545–581.
Nagelkerken, I., M. Dorenbosch, W.C.E.P. Verberk, E. Coc-
heret de la Moriniere & G. van der Velde. 2000a. Importance
of shallow-water biotopes of a Caribbean bay for juvenile
coral reef fishes: Patterns in biotope association, community
structure and spatial distribution. Marine Ecol. Progr. Series
202: 175–192.
Nagelkerken, I., G. van der Velde, M.W. Gorissen, G.J. Meijer,
T. van’t Hof & C. den Hartog. 2000b. Importance of man-
groves, seagrass beds and the shallow coral reef as a nursery
for important coral reef fishes, using a visual census tech-
nique. Estuarine Coastal Shelf Sci. 51: 31–44.
Nagelkerken, I. & G. van der Velde. 2002. Do non-estuarine
mangroves harbor higher densities of juvenile fish than
adjacent shallow-water and coral reef habitats in Curacao
(Netherlands Antilles)? Marine Ecol. Progr. Series 245: 191–
204.
Odum, W.E. & E.J. Heald. 1972. Tropical analyses of an estu-
arine mangrove community. Bull. Marine Sci. 22: 671–768.
186

Parrish, J.D. 1989. Fish communities of interacting shallow-
water habitats in tropical oceanic regions. Marine Ecol.
Progr. Series 58: 143–160.
Quinn, N.J. & B.J. Kojis. 1985. Does the presence of coral reefs
in proximity to a tropical estuary affect the estuarine fish
assemblage? Proceedings of the 5th International Coral Reef
Congress 5: 445–450.
Roberts, C.M. 1997. Connectivity and management of Carib-
bean coral reefs. Science 278: 1454–1457.
Robertson, A.I. & N.C. Duke. 1987. Mangroves as nursery
sites: Comparisons of the abundance and species composition
of fish and crustaceans in mangroves and other nearshore
habitats in tropical Australia. Marine Biol. 96: 193–205.
Robertson, A.I. and N.C. Duke. 1990. Recruitment, growth
and residence time of fishes in a tropical Australian mangrove
system. Estuarine Coastal Shelf Sci. 31: 723–743.
Robertson, D.R., D.G. Green & B.C. Victor. 1988. Temporal
coupling of production and recruitment of larvae of a
Caribbean reef fish. Ecology 69: 370–381.
Shapiro, D.Y., R.S. Appeldoorn, D.A. Hensley & M. Ray.
1997. Water flow and spawning time in a coral reef fish.
Proceedings of the 8th International Coral Reef Symposium
2: 1121–1126.
Shulman, M.J. 1985. Recruitment of coral reef fishes: Effects of
distribution of predators and shelter. Ecology 66(3): 1056–
1066.
StatSoft, Inc. 2001. STATISTICA (data analysis software sys-
tem), version 6.
Steele, M.A. 1998. The relative importance of predation and
competition in two reef fishes. Oecologia 115: 222–232.
Stunz, G.W., T.J. Minello & P.S. Levin. 2002. Growth of newly
settled red drum Sciaenops ocellatus in different estuarine
habitat types. Marine Ecol. Progr. Series 238: 227–236.
Swearer, S.E., J.E. Caselle, D.W. Lea & R.R. Warner. 1999.
Larval retention and recruitment in an island population of a
coral-reef fish. Nature 402: 799–802.
Thayer, G.W., D.R. Colby & W.F. Hettler Jr. 1987. Utilization
of the red mangrove prop root habitat by fishes in south
Florida. Marine Ecol. Progr. Series 35: 25–38.
Thollot, P. 1992. Importance of mangroves for Pacific reef fish
species, myth or reality? Proceedings of the 7th International
Coral Reef Symposium 2: 934–941.
Tolimieri, N. 1998. The relationship among microhabitat
characteristics, recruitment and adult abundance in the
stoplight parrotfish, Sparisoma viride, at three spatial scales.
Bull. Marine Sci. 62: 253–268.
Victor, B. 1986. Larval settlement and juvenile mortality in a
recruitment-limited coral reef fish population. Ecol. Monogr.
53: 145–160.
Vidy, G. 2000. Estuarine and mangrove systems and the nurs-
ery concept: Which is which? The case of the Sine Saloum
system (Senegal). Wetlands Ecol. Manage. 8: 37–51.
187
Copyright © 2022 FDOKUMEN




















