
Ecology of a Cerrado Lizard Assemblage in the Jalapão Region of Brazil
Upload
independentCategory
view
6download
0
The Ediacaran Assemblage
Abstract
This page describes the geologicaland chronological settings of theEdiacaran forms, the range ofmorphologies of the fossilsthemselves, and concludes withsome remarks about theirrelationships with otherorganisms.
Keywords: Ediacaran fauna, Vendianfossils
Introduction
The name ‘Ediacaran’ has ageochronologic meaning, providingan upper subunit of the Vendian,approximately 620 to 543 Ma (Knollet al. 2004), with a stratotype inSouth Australia. Confusingly, thesame term is also used in abiogenic sense, and in twodifferent ways: Many authors applythe term ‘Ediacaran’ in a broadsense to any Vendian (orEdiacaran) age macrofossil,whereas others restrict the termnarrowly to the unique anddistinctive assemblage ofenigmatic organisms best knownfrom the Ediacara Hills of SouthAustralia, and characterised byproblematic oval, frondose, andspindle-shaped forms of unknownaffinity.
Ediacaran fossils were describedfrom the Fermeuse Formation on theAvalon Peninsula – specifically,Aspidella terranovica was named – by E.Billings in 1872. A secondassemblage was described fromNamibia sixty years later (Gürich1933). Nevertheless, the
Related Topics
Further Reading
Crucible of Creation, The – Simon Conway Morris
Fossils of the Burgess Shale, The – Derek Briggset al .
Snowball Earth - Gabrielle Walker
Garden of Ediacara, The – Mark McMenamin
Related Pages
Vendian Period Vendian-Cambrian
geological column Ediacaran Taxa Ediacaran Localities Ediacaran Survivors Cambrian Explosion
Other Web Sites
Pictures and brief descriptions from Berkeley
Ediacarans of Canda (incl. Mistaken Point) fromQueen’s University
A few more articles about Ediacarans from BellarmineUniversity

assemblage acquired its name froma third and even later discovery,made by Reginald Sprigg in March1946, at an abandonedcopper/lead/zinc mine in theEdiacara Hills, Flinders Range,north of Adelaide in SouthAustralia. Since then, occurrenceshave been located on mostcontinents.
The assemblage comprises marinelife forms first appearing inlatest Precambrian times – about575 Ma, placing them among theoldest multicellular fossils known– and persisting into the basalCambrian. The Ediacaran hey-daypredates by a distinct interval ofperhaps 20 Ma or more, the so-called ‘Cambrian Explosion’ when‘modern’ multicellular life beganto diversify rapidly.
For some years a number of authors(e.g. Seilacher 1984, McMenamin1986) have argued that theEdiacarans were unrelated to anyliving group of organisms; thatthey represented a new kingdom(Vendobionta Seilacher 1992) whichdisappeared around the Vendian-Cambrian boundary, perhaps wipedout by a mass extinction event.However, this view has alwaysencountered opposition and nowappears to have lost much of itssupport.
Geological Setting
Occurrences are scattered at lowpaleolatitudes on every continentexcept (so far) Antarctica.Additionally, the South AmericanMato Grosso occurrence fromsouthwestern Brazil (Hahn et al.1982) is questionable. The best

known are the ‘classic’ localitiesin southern Namibia, the FlindersRange locality in Australia,Mistaken Point in south eastNewfoundland, and on the White Seacoast of northern Russia, butthere are also reported occurencesin Mexico, England, Ireland,Scandinavia, Ukraine, and the UralMountains (read more).
Habitat and Habit
Among the first comprehensivetreatments of the Ediacarans werethose of Martin Glaessner, in the1960s. However, although we maynow agree with many of hisconclusions, his ideas werepredicated on an incorrectinterpretation of thepaleoenvironment: Glaessner (1961;also Glaessner & Wade 1966;Jenkins 1981) believed theRawnsley depositional sequence tohave been semi-emergent ("sandyshoals ... [with] areas oftemporary quiescent conditionsbetween the shifting currenttracks, where fine particles couldsettle until they were coveredagain by sand waves" – Glaessner &Wade 1966, pp. 599-600) and thebody fossil assemblagetransported. Glaessner envisagedthe assemblage as a massstranding, thereby predisposinghimself to accept the radial formsas ‘medusoids.’ More recently,however, Gehling (1991, 2001) hasdemonstrated that the SouthAustralian fossils occur above avalley fill facies, on sandstonepartings within upward-shoaling,delta-front environments betweenstorm- and fair-weather wave base(Gehling 2001, p. 30).
The Australian fossils occur inpreservational windows in theEdiacara Member of the RawnsleyQuartzite, a formation of thePound Subgroup, bounded above bythe Early Cambrian Uratannasequence. The Rawnsleydepositional sequence is developedover an erosional surface havingsome 250 m of relief, wheresoutheasterly directedpaleovalleys are filled withsequences of massive sandstone andlaminated siltstones, passing upinto up into well-beddedsandstone. The fossils occur abovethe valley fill facies, onsandstone partings within upward-shoaling, delta-front environmentsbetween storm- and fair-weatherwave base (Gehling 2001, p. 30).

Other occurrences are now widelyunderstood to be in situ marineassemblages, also. In particular,the Mistaken Point and nearbylocalities of south easternNewfoundland are believed topreserve a deep water slopeenvironment.
Lithology
"The sand is of unusual texture;rather like foundry sand, which isa factor in this uncommon mode ofpreservation" (Clarkson 1993, p.59).
"The clay lenses were subsequentlyhighly compacted and altered andare now mostly only thin,lenticular partings between thequartzite flags. Most of thesepartings can be opened only bynatural weathering. They revealfossils mostly on the lowersurfaces of the quartzite flags"(Glaessner & Wade 1966, pp. 599-600).
Chronology
The oldest characteristicEdiacaran fossils are those of theDrook Formation from south-easternNewfoundland, believed to datefrom around 575 Ma, but the oldestof the ‘classical’ localities isthe 565 Ma occurence at MistakenPoint, Newfoundland. Youngest ofthe classical localities is inNamibia, where Ediacarans co-occurwith small shelly faunas and rangeup to the Vendian-Cambrianboundary (543 Ma; see Grotzingeret al. 1995, Martin et al.2000). Thus, paleontologists havehad to come to terms with a"relatively short time frame ofEdiacaran biology. Diverse
Ediacaran assemblages fromAustralia, northern Russia, andNamibia were all deposited withinthe last 15 to 20 million years ofthe Proterozoic Eon" (Knoll &Carroll 1999).
However, there are contenders topush these boundaries in bothdirections. The stratigraphicrange of the Ediacarans sensu amplaessentially equates to the ~100 Marange of Nimbia occlusa. The Twityafossils (pre-Varanger) are theoldest fauna which have beentermed ‘Ediacaran,’ and Booley Bay(Late Cambrian) the youngest: bothinclude forms assigned to Nimbiaocclusa. With the single exceptionof the Twitya Formationoccurrence, all appear to post-date the last Varangianglaciation.
First Appearance
Simple disc-like impressions,interpreted as cnidarian-gradebody fossils, from theintertillite beds of the >635 Ma(Xiao & Laflamme 2008, p. 32)Twitya Formation in the MackenzieMountains, north-western Canada(Hofmann et al. 1990), include the"Ediacaran taxon" Nimbia occlusa andare sometimes described as"Ediacarans." They are simple,circular impressions and, while itis true that many taxa from the‘classic’ Ediacaran localities arelittle more, the Twitya assemblageas a whole does not exhibit themorphological diversity nor thecomplexity of the classicalEdiacaran assemblages. Unless oneregards the Ediacaran assemblageas simply a ‘hold-all’ for anyVendian macrofossil of possiblemetazoan affinity, the Twitya
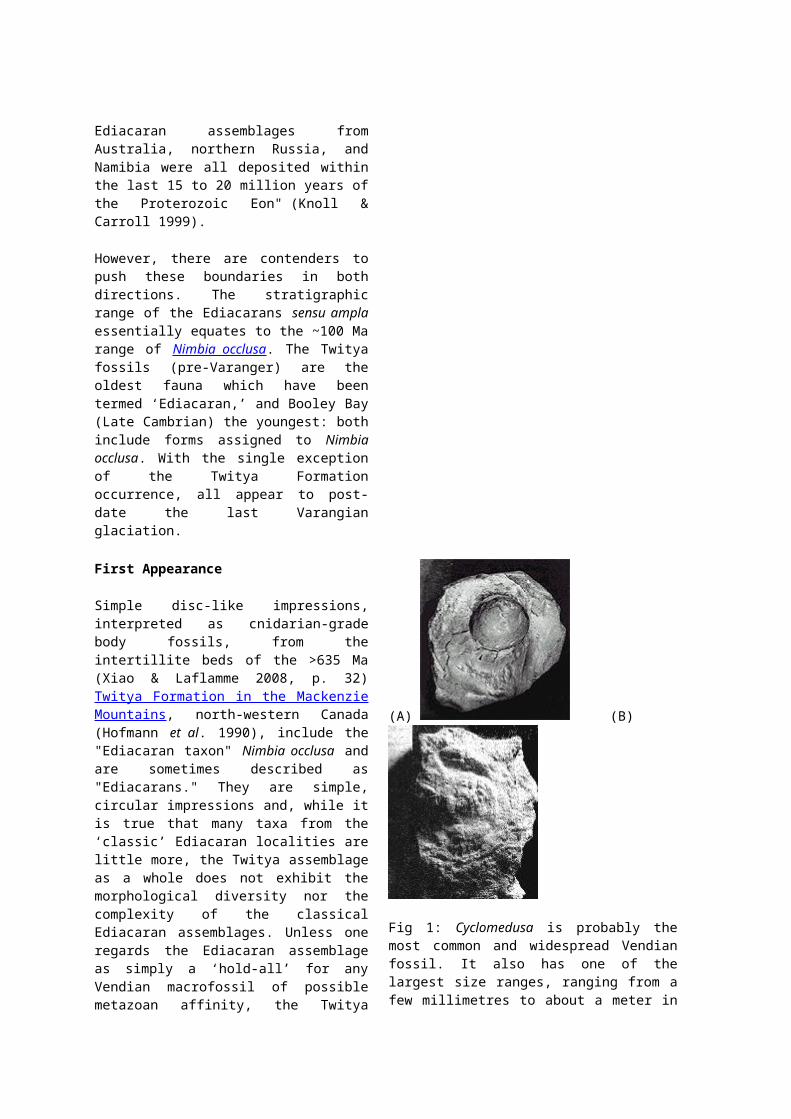
(A) (B)
Fig 1: Cyclomedusa is probably themost common and widespread Vendianfossil. It also has one of thelargest size ranges, ranging from afew millimetres to about a meter in

fossils do not belong here.
In a report of Ediacaran taxa andassociated trace fossils from theClemente Formation of north-western Sonora, Mexico, MarkMcMenamin (1996, p. 4990) rejectsthe authenticity of the Twityaassemblage and explicitly laysclaim to having found "the oldestknown remains of the Ediacaranbiota" himself (see fig. 1A).McMenamin appears committed tothis interpretation (e.g. 1998, pp.204-207) though neither hisassertions for the age of thematerial ("600 million years ormore") nor the biologicalaffinities of his putative fossilshave been embraced with anyenthusiasm by others. The proposedage, in particular, is based uponan extremely tenuous chain ofreasoning, all under-pinned by thesupposed stratigraphic range of asingle, poorly-documented,‘ichnotaxon,’ Vermiforma antiqua(1996, p. 4993), which may, infact, turn out to be a tectonicartefact (Meschede et al. 2000).Thus the Sonora material cannot beseriously considered withoutadditional corroboration and isnot further discussed here.
Claims of 600 Ma plus requirethese fossils to pre-date theVaranger-Marinoan ice age,approximately 600 to 590 Ma, whichmay have been a time of widespreadextinction. "Late Proterozoiccarbon isotopic profiles displaystrong negative as well aspositive excursions. Negativeexcursions are specificallyassociated with the major ice agesthat mark immediately pre-Ediacaran time. Much research iscurrently focused on this unusual
diameter. Formerly thought torepresent a planktonic (floating)jellyfish of some sort, Cyclomedusais now considered by some to havebeen a benthic (bottom-dwelling)polyp, somewhat like a sea anemone,and by others to be the anchoringholdfasts of colonial, softoctocorals. The latter hypothesismay also explain why they are socommon as fossils – because theywere already buried – and positsthat most of the ‘species’ areartefacts of differing preservation.
(A) McMenamin’s form "cf. Cyclomedusaplana Glaessner and Wade" (= Aspidellaterranovica Billings 1872) fromSonora, Mexico: "A discoid fossilpreserved in hyporelief. Noteannular ridge occurrence at themargin (arrowhead) of the centralcone. Greatest dimension of rockspecimen is 6.0 cm. Sample 1 of3/17/95; fossil occurrence isapproximately 75 meters below theClemente Formation oolite, in unit 1of the ClementeFormation" (McMenamin 1996, fig.2A).
(B) Cyclomedusa sp. from the WinterCoast of the White Sea. Thisspecimen is about 5 cm across.[Image courtesy of University ofCalifornia Museum of Paleontology.]

coupling of climate andbiogeochemistry, and bothpaleoceanographic models andclustered phytoplanktonextinctions suggest that these iceages had a severe impact on thebiota – potentially applyingbrakes to early animalevolution" (Knoll & Carroll1999). Although presumed bodyfossils, such as the Twityaassemblage occur earlier, all ofthe diverse Ediacaran fossilassemblages post-date theVaranger-Marinoan ice ages.
Acme
Oldest occurrences, such as thosefrom the Twitya and Drook Formations, are taxonomicallyimpoverished. The assemblagebecomes rich around 565 Ma (e.g. atthe Mistaken Point locality) butdoes not achieve full diversityuntil about 555 Ma. From then itcontinues in full bloom until theVendian-Cambrian boundary afterwhich, although some taxa lingeron, the characteristic assemblageas a whole abruptly disappears. Itis uncertain whether a massextinction event struck at thistime, or if we are simplyobserving the closure of some formof ‘taphonomic window.’ It hasbeen suggested that more widelyspread and deeper bioturbation,evidence for which increasessharply at the base of theCambrian, is incompatible with theunique ‘Ediacaran preservation.’Somehow, I find this viewunconvincing.
A number of ‘Ediacarans’ arereported from the Cambrian. Theyoungest of them all is animpoverished Late Cambrian

assemblage of just two taxa, oneof which is Nimbia occlusa, fromBooley Bay in Co. Wexford,Ireland.
Last Appearance
At the younger end of their range,"Ediacara type" fossils have beenincreasingly reported fromCambrian sediments (see below).Youngest and most exciting ofthese are the Upper CambrianEdiacarans from a turbiditesequence exposed at Booley Bay,near Duncannon in Co. Wexford,Ireland, which includes two taxa:'Ediacaria booleyi' (possibly yetanother variant of Aspidellaterranovica) and the cosmopolitanNimbia occlusa (Crimes, Insole andWilliams 1995). The Booley Bayoccurrence is dated by acritarchsof sufficient "diversity andquantity to constrainbiostratigraphically the relativeage of this succession ... to theupper part of the Upper Cambrian"(Moczydlowska & Crimes 1995, p.125), indicating that at leastsome Ediacarans co-existed with'modern' taxa for perhaps 20 or 30Ma – and certainly throughout theCambrian Explosion.
Morphology

More than 30 different genera havebeen named. Ediacarans are adiverse group and earlier attemptsto pidgeon-hole them into alimited number of phylogenetictypes appear now to have beenmisguided. However, for a quickoverview it may be useful toconsider four broad morphologicalcategories (after Briggs et al.1994, p. 44), bearing in mind thatthese do not indicate evolutionaryrelationship.
1. Most abundant are circularimpressions, some of whichplausibly recall jellyfishand similar medusoidcnidarians, although manyare so simple thatconvergence rather thanaffinity cannot be ruled out(fig. 1). Others of thisform are believed to be theholdfasts of the frond-likeEdiacarans (see below, alsofig. 4A).
2. Next are the trace fossilsof various tracks andburrows made, at least inpart, by bilaterian animals.Although there is evidenceof burrowing, the traces aremostly simple and more orless horizontal (but seeJensen & Runnegar 2005);typically absent is anyevidence for wide-scalechurning up of the sea bedby animals living in thesediment (infauna) (ConwayMorris 1998, p. 30).
3. Third most abundant are anumber of problematicbenthic forms. Whereas someof these seem familiarenough to suggest affinitieswith extant groups such asannelids (e.g. Dickinsonia andSpriggina, see fig. 2;however, also see Dzik and
(A) (B)
Fig. 2: (A) Spriggina floundersiGlaessner – From the Vendian PoundQuartzite of the type locality,Ediacara, South Australia. Overalllength about 10 cm. Specimen fromthe Yale collection (YPM 63257).
(B) Dickinsonia costata Sprigg –Vendian, from the Brachina Gorge,Flinders Ranges, South Australia.Specimen from the Yale collection(YPM 35467). Dickinsonia has beenknown to reach dimensions of up to ametre.[Images courtesy of the PeabodyMuseum of Natural History, YaleUniversity.]

Ivantsov 1999 for a contraryview), echinoderms (e.g.Arkarua), or arthropods (e.g.Diplichnites and Parvancorina,fig. 3A), others such asPraecambridium, Vendia andTribrachidium (fig. 3B) aremore problematic.
4. Least abundant, thoughperhaps most characteristicof the assemblage as awhole, are the attached,frond-like organisms (fig.4A), which some propose haveaffinities with the sea pensand other soft corals.
Leaving aside the trace fossils, atypical Ediacaran of any form hada soft body – there is no evidenceof any skeletal hard parts except,possibly, for the head-shield ofSpriggina (fig. 2A) – yet they mostcommonly occur in silt- andsandstones which typically form inquite turbulent conditions: notthe sort of sediments where onewould ordinarily expect to findgood soft tissue preservation.
Many of the forms display amorphology which has beendescribed as "quilted." Someresearchers consider this to be areal characteristic, whichindicates a phylogeneticrelationship between otherwisedissimilar forms: that all the"Ediacara fossils" are members ofthe same high-level taxon; thatthey form a single clade with asingle bauplan (see below).
Interestingly, it appears thatEdiacaran communities were largelyfree of large predators; nospecies appears to have possesseda jaw apparatus suitable forseizing and tearing prey, and fewfossils show clear evidence of
(A) (B)
Fig. 3: (A) Parvancorina minchami – Acandidate arthropod, possibly atrilobite (see Fortey et al. 1996). Inthis scenario, the central axialridge and the strongly archedanterior ‘lobes’ may be analogous tothe midgut and gastric diverticulae.The scale bar is in centimetres.[Image and interpretation courtesyof Chris Nedin, Department ofIndustry, Science and Resources,Canberra.]
(B) Tribrachidium heraldicum – Fewfossils of Ediacaran animals are socompellingly bizarre as this unusualdisc-shaped form with three-part(triradial) symmetry. Affinitieshave been proposed with either theCnidaria (corals and anemones) orEchinodermata (urchins andstarfish); nor can the possibilitythat it is a holdfast be entirelyeliminated. [Image andinterpretation courtesy ofUniversity of California Museum ofPaleontology.]

predatory damage. A possibleexception, however, are someChinese Cloudina fossils with tinyboreholes, which may simply be adiagenetic effect of may truly beindicative of predation.
Antecedents
Genetic evidence has been used tosuggest significant metazoandiversity far pre-dating theEdiacaran fossils (e.g. Wray,Levinton & Shapiro 1996:"Calibrated rates of molecularsequence divergence were used totest this hypothesis. Sevenindependent data sets suggest thatinvertebrates diverged fromchordates about a billion yearsago, about twice as long ago asthe Cambrian. Protostomesapparently diverged from chordateswell before echinoderms, whichsuggests a prolonged radiation ofanimal phyla.")
Other estimates (e.g. see ConwayMorris 1998, Ayala et al. 1998,Knoll & Carroll 1999) are lower,but still require the existence ofsome animal diversity as early as750 Ma ago, implying that for thefirst 150 Ma or more they left nofossil record. (Inexplicably,Ayala et al. claim that theirresults are "consistent withpaleontological estimates.") Thegeneral rarity of soft-partpreservation may explain this inpart, but one would still expectto find some trace fossils –tracks and burrows – of anyanimals large enough to disturbsea-floor sediments. "Thus, ifthey really were present, we canbe fairly sure that any pre-Cambrian animals would have been

tiny, only a few millimetreslong.... What later triggeredtheir initial emergence as theEdiacaran faunas, and subsequentlythe even more spectacular Cambrianexplosion, remains a significanttopic for debate" (Conway Morris1998, p. 144).
At more than 635 Ma (Xiao &Laflamme 2008), circularimpressions from the TwityaFormation of the MackenzieMountains provide evidence for theearliest macroscopic metazoans,simple cup-shaped organisms,possibly cnidarians, predating theMarinoan Glaciation. Somewhatlater, around 600 Ma, and stillpredating any known Ediacaranassemblage, the Doushantuophosphate deposit in China isslowly yielding a surprisinglydiverse biota, including probablealgae, sponges, cnidarians andbilaterians (read more).
Affinities
Until now we have considered the‘Ediacaran fauna’ in abstractterms, without any attempt todelimit the concept. Realdifficulties stand in our way – itis genuinely difficult to map thecharacters of most Ediacaranfossils onto the body plans ofliving invertebrates; certainlythere are similarities, but theyare "worryingly imprecise" (ConwayMorris 1998, p. 28). Nevertheless,failure to make the attempt is noless reprehensible for having along pedigree.
Initially, Ediacarans wereinterpreted in terms of extant
Phylum Ctenophora?Class unknown
Order unknownFamily Stromatoveridae Shu et al. 2006
StromatoverisPhylum Cnidaria Hatschek 1888
Class Cyclozoa Fedonkin 1983Order unknown
Family Cyclomedusidae Gureev 1987
Aspidella (incl. Cyclomedusa, Ediacaria, Spriggia, Tateana, Tirasiana,etc.)
Class Hydrozoa Owen 1843

phyla, such as cnidarians,annelids, etc. Much of this earlywork was completed by MartinGlaessner. Fellow Australian, JimGehling (1991, p. 182), reportsthat Glaessner left many Ediacaranforms unassigned, and in somecases undescribed, because he"refused to include new taxa inextant phyla without consideredtaxonomic evidence, stating thatfrom a ‘historical perspective,failures are not phyla.’" TheGerman paleontologist, Hans Pflugwas clearly unconvinced, however,and in 1972 erected the new phylumPetalonamae to accommodate some ofthe frondose Ediacaran taxa (Pflug1972).
Vendozoa
Through the mid-1980s to mid-1990s, Adolf Seilacher and someothers further questionedassignments of Ediacaran taxa toliving phyla, and even themetazoan affinities of manyEdiacarans. He contended that, inspite of their apparent diversity,nearly all of the genera share astriking basic uniformity: theyare thin and flattened, round orleaf-like, possess a ridged or‘quilted’ upper surface, and lackclear indications of a mouth orgut. Seilacher believes that theEdiacaran body plan comprisestough organic walls surroundingfluid-filled internal cavities.
Seilacher 1992 recognises threegroups: cryptic bilaterians whichleft only trace fossils, thePsammocorallia – coelenterateswhich utilised sand for aninternal skeleton – and the‘quilted’ Vendobionta, a conceptroughly comparable to Pflug’s
Order unknownFamilies unknown
Eoporpita, Ovatoscutum, Wigwamiella
Class Anthozoa Ehrenberg 1834Order Pennatulacea ??? (Subclass Octocorallia)
?Family Pteridiniidae Richter 1955 [though maybe this family belongs with the Petaliform problematica or with the Trilobozoa?]
Pteridinium (but not Charniodiscus, in my view)
Orders unknownFamilies unknown
Beltanelliformis, Hiemalora, Inaria
Classes unknownOrders unknown
Families unknownNamapoikiaNimbia
Phylum Mollusca Linnaeus 1758Class unknown
Order unknownFamily unknown
Kimberella
Phylum Arthropoda von Siebold & Stannius 1845
Class unknownOrders unknown
Families unknown?Archaeaspinus, Onega, Parvancorina, Praecambridium, Redkinia
Family Sprigginidae Glaessner 1958
Spriggina
Phylum Echinodermata

Petalonamae. The last of thesegroupings has attracted somevigorous criticism: "In proposingthe separation of Ediacaranorganisms from the Metazoa,Seilacher (1989) has attempted tounify them with a commonconstructional model. … Byemphasising the untestedgeneralisation that all [of the‘quilted’] Ediacaran organismswere flat and constructed oftubular elements, anduncomplicated by internal organs,Seilacher (1989, fig. 2) was ableto argue that the observedvariation between taxa was solelybased on modes of growth,involving the addition of tubularelements. … Only with a very broadbrush could all Ediacaranorganisms be represented asfractal growth variations based onthe same units of construction"(Gehling 1991, pp. 192-193, 202).
Gehling’s last point is supportedby observation of considerablevariation between taxa, in thestyle of preservation, indicatingthat there were "different classesof organic construction involvedin the Ediacara fauna" (Gehling &Rigby 1996, p. 185).
The idea that most or allEdiacarans were taxonomicallyclosely related has mostly fallenfrom favour - "It’s clearly nottrue for, say, the BurgessShale. Why should it apply to theEdiacarans?" (Waggoner 1999) - andfew adherants remain.
However, Seilacher’s argument mustbe seen in the context of itstime; it was predicated onknowledge of far fewer Ediacarans(in general, the larger taxa) than
Class ?EdrioasteroideaOrder unknown
Family unknownArkarua
Insertae sedis1. Triradially Symmetric Taxa (= Phylum Trilobozoa Fedonkin 1985)
Classes unknownOrders unknown
Family unknownTriforillonia
Family Albumaresidae Fedonkin 1985
Ablumares, Anafesta, Skinnera
Family Anabaritidae Glaessner 1979
Anabarites
Family Tribrachididae Runnegar 1992
Tribrachidium
2. ‘Metameric’ TaxaClasses unknown
Orders unknownFamily Dickinsonidae Harrington & Moore 1955
Dickinsonia
Family Vendomiidae Keller 1976
?Vendia, Vendomia
Family Yorgiidae Ivantsov 2001
Yorgia3. Petaliform Taxa (= Phylum Petalonamae Pflug 1972; Kingdom Vendobionta Seilacher 1992)
Class Erniettamorpha Pflug 1972Order(s) unknown
Family Erniettidae Pflug1972
Ernietta, ?Swarpuntia

are known today, and underpinnedby the beliefs – prevalent at thetime – that:
1. the Ediacarans died outcompletely well before theVendian-Cambrian boundary(the Kotlin crisis);
2. a considerable age separatedthe disappearance of theEdiacarans and theappearance of the firstcalcareous ‘skeletons’ (theso-called ‘small shellyfauna,’ see below);
3. the assemblages representedmass strandings rather thanin situ associations; andperhaps stemming from this,
4. that coeval trace fossilscould not be attributed tothe Ediacaran taxa.
However, none of these beliefs canbe sustained today except,arguably, the last. Even thatcontention is less tenable than itwas in the 1980s, when theEdiacarans were thought to bemostly large taxa. We now knowthat small taxa make up a largepart of the assemblage; the smallbilaterian forms are potentiallythe missing trace-makers.
Seilacher’s originalconstructional analysis is notdebated a great deal today, andthough it still claims adherents(e.g. see McMenamin 1998), themajority of authors speak of‘Ediacaran metazoans’ and ‘theEdiacaran fauna.’
Recent Views
Bruce Runnegar and MikhailFedonkin (in Schopf & Klein 1992,p. 373) were next to tackle the
Family Pteridiniidae Richter 1955 [though maybe this family belongs with the Anthozoa or even the Trilobozoa?]
?Phyllozoan, Pteridinium
Class Rangeomorpha Pflug 1972Order(s) unknown
Family RangeidaeRangea
Family CharniidaeCharnia, ?Charniodiscus, Paracharnia
4. ‘Vermiform’ Taxa (= Phylum "Vermes" of Runnegar 1992)
?Archaeichnum, ?Cloudina, Cochlichnus, ?Didymaulichnus, Gordia, Harlaniella, Helminthoidichnites, Planolites, Sellaulichnus
5. Serial Growth FormsNeonereites, Palaeopascichnus, Yelovichnus
6. OthersAusia, Bomakiella, Bonata, Lorenzinites, ?Wigwamiella
Table 1: Taxonomic outline of someEdiacaran forms. Blue = form-taxa;red = trace fossils (ichnotaxa).

overall taxonomy of theassemblage. Their approach – byfar the most convincing to date –combines the conservativereference of taxa to modern phyla,where such assignments are notobviously forced, with a pragmaticrecognition of the large number ofundeniably enigmatic forms.Probably the most significantmessage evident in Runnegar &Fedonkin’s classification –obvious today, but enlightening in1992 – is that the Ediacaran faunais not a single, monolithic,taxonomic group. Rather, differentEdiacaran taxa represent a varietyof metazoan (and possibly other)lineages. Even today, some authors(e.g. McMenamin 1998) appearuncommitted to this view. Table 1is an updated version of Runnegar& Fedonkin’s work.
Traces
Although some traces are simple,rather featureless, windingtrails, "others display transverserugae and contain pellets that canbe interpreted as of fecal origin.The bilaterian nature of thesetraces is not in dispute.Furthermore, such traces must havebeen made by worms, some of whichhad lengths measured incentimetres, with through guts,which were capable of displacingsediment during some form ofperistaltic locomotion, implying asystem of body wall musclesantagonized by a hydrostaticskeleton. Such worms are morecomplex than flatworms, whichcannot create such trails and donot leave fecal strings"(Valentine 1995, p. 90).
Sets of paired hypichnial ridges

further hint at an arthropod s.l.presence.
A trace fossil presumed torepresent the radula scratches ofa mollusc is found at Zimnie Goryand in the Ediacara Hills (Martinet al. 2000, p. 844).
'Metameric' (Segmented) Forms
Whereas there may be a generalacceptance that the majority ofEdiacarans are stem metazoans ofsome sort, most are stillnotoriously problematic. One largeand important group of these,arguably the most crucial to ourunderstanding of the overallpattern of metazoan evolution, arethose exhibiting real or apparentmetamerism. Most are small, thoughsome of the dickinsonids can beenormous: up to about a metre.
Some authors, notably M.A.Fedonkin and A. Yu. Ivantsov,argue that many of these organismsare pseudosegmented, with segmentsalternating on either side of themid line, thereby casting doubt ontheir bilaterian affinities.However, their publishedphotographs (e.g. of Archaeaspinusand Yorgia) are often based uponrelatively few specimens and theasymmetry is not always clear (JimGehling, pers. comm.)
In some specimens of Dickinsonia,the segments do not appear tocorrespond across the mid-line onthe dorsal surface. However, asnoted in Gehling 1991 (though theoriginal observation is attributedto Bruce Runnegar), in allspecimens where the ventral sideis preferentially preserved,segments clearly continue across

the mid-line, so offset on thedorsal side must be a product offlattening. It is only in rarespecimens of Dickinsonia elongata thatthe alternate insertion andoverlap of segments along the midline is difficult to explain.
"Fedonkin (1983, 1984, 1985a,1986) has not only attempted toassess the phyletic relationshipsof Ediacaran taxa, but has tackledthe problem of comparativemorphology. However, his approachhas been strongly dependent on atwo dimensional body plan analysisof symmetry. This concept of"promorphology" almost entirelydisregards the original threedimensional architecture,palaeobiology, and ontogeny of theorganisms. Bergström (1990, figure2) illustrated four taxa withapparent alternation of regularelements on each side of the axis;but in each case the sketchesrepresent unrestored images offlattened animals. Order withinsymmetry groups may be useful inclassification of minerals, but inorganisms, the superficialsymmetry of body plans may be asecondary product of adaptation todifferent life styles" (Gehling1991, p. 203.)
Metazoan Origins

Some similar organisms – whichpost-date the major Ediacaranbiotas and are generally betterpreserved – have been identifiedwith conventional zoologicaltaxa. Conway Morris 1998 cites asexamples Thaumaptilon (fig. 4B),which he believes is aconventional pennatulacean (pp.28-29), Mackenzia (pp. 83-84), andEmmonsaspis (p. 134, note7). Advocates of this viewpointexplain the unusual ‘Ediacaranpreservation’ by suggesting apaucity of burrowing andscavenging organisms to disturbthe remains, once buried.
However, it remains true that thedifferences between the Ediacaranand overlying Cambrian faunas arefar more striking than anysimilarities.
There can be little doubt, on thebasis of trace evidence alone,that bilaterian metazoans existedin the Vendian. Unfortunately, itis equally true that therelatively few body fossils knownfrom the late Precambrian do notshed much light on the sequence ofevolutionary advances that led tothe famously diverse Cambriantaxa. There are a few sign-posts,however:
Sponges are widelyrecognised (e.g. Nielsen2001, pp. 30, 506-507) to bethe most primitive of livingmetazoans, occupying a basalposition in metazoanphylogeny, as a sister groupto all other Metazoa. Thustheir first occurrence inthe fossil record is ametric of particularinterest. However, only rareoccurrences of Precambrian
(A) (B)
Fig. 4: (A) Charniodiscus arboreus – Oneof the frond-like Ediacaran fossilsconsidered by some to be a‘conventional’ cnidarian, possibly apennatulacean. Collected from theEdiacaran Member, RawnsleyQuartzite, Bunyeroo Gorge, FlindersRanges, South Australia. Specimenfrom the South Australian MuseumCollection (SAM P19690). Overalllength 40cm. [Image courtesy of theSouth Australian Museum.]
(B) Thaumaptilon walcotti Conway Morris1993 – From the Middle CambrianStephens Formation Burgess Shale.Proposed by Conway Morris as apossible pennatulacean (sea pen).However, this view is by no meansuniversally accepted; for example,

sponges have been reported.The earliest record is ofpresumed sponge remains fromthe Doushantuo phosphates,dated around 570 Ma (Li et al.1998; read more), and theearliest described speciesis Paleophragmodictyareticulata from the ?555 MaEdiacara locality (Gehling &Rigby 1996; read more).However, sponges could haveoccurred earlier and notbeen recognised; spiculesare not necessarilydiagnostic, even in livingsponges (Dr. Allen Collins,pers. comm.)
Fossils of the TwityaFormation are generallypresumed to be cnidarians,or at least as metazoans ofcnidarian grade."Interpretation as colonialaggregates of prokaryotes(e.g. Nostoc-like balls) ispossible but is difficult toreconcile with themorphology and relativelyhigh relief of the remains,their occurrence at thebottom of turbidite beds,and the lack of acarbonaceous film outliningthem, particularly in viewof the of the fact thatcarbonaceous compressionsare present in theformation" (Hofmann et al.1990, p. 1202). Of principalsignificance is thisoccurrence of cnidarian-grade metazoans in pre-Varanger sediments, sincethe Varanger glaciation issometimes cited as anevolutionary 'bottleneck'which arrested metazoanevolution.
Nielsen 2001 notes (p. 59) that thebranches of both Charniodiscus andThaumaptilon "were united with amembrane which makes theinterpretation dubious on functionalgrounds, and the structurestentatively interpreted as polypsare very small and show notentacles." Specimen from the USNational Museum collection (USNM468028). Overall length about 20 cm.If this animal is a descendant ofCharniodiscus and its Ediacaranallies, as proposed by ConwayMorris, the holdfast has beenmodified from the original bulkydisc, possibly to facilitatewithdrawal into aburrow. [Reproduction of fig. 2Efrom Conway Morris 2000.]

In preserving evidence ofbilaterians, the Vendianrecord provides constraintson the protostome-deuterostome split. IfKimberella is indeed amollusc, as suggested byFedonkin & Waggoner 1997, orthe Ediacara/Zimnie Gorytraces are correctlyinterpreted as radulascratches, we have evidencefor derived protostomes at555 Ma. Similarly, if Arkaruaadami (from the PoundSubgroup, South Australia;Gehling 1987) is correctlyinterpreted as anechinoderm, we have evidencefor a derived deuterostomeof similar age. In eithercase, it follows that the P-D split must have occurredwell before 555 Ma, which isin accordance with most'molecular clock' studies.
Nutritional Hypotheses
Many Ediacarans appear to havebeen very thin. Some researchers,particularly those from theSeilacher camp, propose that theirtissues may have housed symbioticalgae. Mark McMenamin (1986, 1998)coined the phrase ‘garden ofEdiacara’ to encapsulate theconcept and he has largelychampioned this theory. However,other workers such as BruceRunnegar (1992, p. 83) point outthat the hypothesis is bothdifficult to test and unlikelyanyway, because some of thefossils appear to have beendeposited below storm wave base –about 100 m – where the intensityof sunlight is very muchdiminished, perhaps below usefullimits for such organisms.

Major Vendian Occurences
575 Ma: The Drook Formation
An impoverished but characteristicEdiacaran assemblage occurs in theupper beds of the Drook Formation,south-eastern Newfoundland, 1,500m stratigraphically below thewell-known Mistaken Point fossils.These are the oldest of the large,architecturally complex fossilsfound so far (Anderson 1978;Hofmann et al. 1979; King 1980;Narbonne & Gehling 2003). Thepublished age constraints on thesefossils are from 595 Ma (Varangianglacial diamictites of theGaskiers Formation) to 565 Ma(well-dated Ediacaran fossils atMistaken Point occurring 1.5 kmstratigraphically higher).Unpublished data noted in Walker2003, p. 220, indicates an age of575 Ma.
Current-aligned frondsattributable to the cosmopolitanEdiacaran, Charnia masoni, and thoseof a large (up to nearly 2 m inlength) new species, Charnia wardi,occur on the shaley tops ofturbidite beds under volcanicashes. Their position above theglacial marine rocks of theGaskiers Formation (595 Ma)provides our earliest window onlife following the Varanger iceage.
~570 to 560 Ma: ‘Old’ Trace Fossils
Crimes in Cowie & Brasier 1989, p.167, lists the earliest occurringtrace fossils as Planolites,Didymaulichnus, Arenicolites, Neonereitesand Gordia.
[Also get the ref. and see what’s inthe Bonahaven Formation, westernScotland (Brasier & McIlroy 1998,Jl. Geol. Soc. London, 155: 5-12).]

The lower Elkera Formation in theGeorgina Basin, central Australia,contains the ichnofossil Planolitesballandus, pre-dating the AustralianEdiacaran fauna (Walter et al. 1989,p. 218) though the genus iselsewhere known to range up intothe Phanerozoic (Crimes in Cowie &Brasier 1989, p. 169). Planolitesballandus is a sub-horizontal,simple, straight to gently curved,unbranched, smooth, cylindricalburrow-fill, sometimes with finelongitudinal striations,approximately 1 mm wide (Walter etal. 1989, p. 239).
Didymaulichnus is another horizontalburrow-fill, comprising a double-lobed, slightly sinuous to tightlycurved, convex hyporelief, ~10 mmwide by ~3 mm deep, the lobesseparated centrally by a distinct,shallow furrow, and ranging upinto the Atdabanian. Arenicolites ishighly unusual – possibly uniqueamong Vendian traces – insofar asit is not confined to thehorizontal plane. It comprises aU-shaped tube, approximately 10 mmdeep. It, too, ranges up into thePhanerozoic.
565 Ma: Mistaken Point – Oldest ofthe ‘Classic’ Ediacaran Assemblages
The oldest of the diverseEdiacaran assemblages yetdescribed is that from MistakenPoint, eastern Newfoundland, wherefossils are spectacularlypreserved on large beddingsurfaces along the sea-cliffs ofthe Avalon Peninsula. Zircons frominterbedded ash have been dated at565 ± 3 Ma (Benus 1988).

The Mistaken Point assemblagecontains a few cosmopolitan taxasuch as Charnia and Aspidella, butmost are either endemic or sharedonly with the Charnwood Forestlocality in central England (King1980).
?565 Ma: Charnwood Forest
The volcaniclastic turbidites ofthe Charnian Supergroup contain anEdiacaran fauna comprisingfrondose forms, either as simplefronds or multi-fronded balls,discs which are usually ovoid andcontain variable numbers ofconcentric rings, worm burrows,and other forms which do not fitinto these groups. Worm burrowtraces occur in the lower BrandGroup of Charnwood Forest. Thediscovery of Charnia masoni in 1957has become an essential part ofEdiacaran folk lore; Aspidella (asCyclomedusa) and Pseudovendia havealso been reported from this site(Brasier in Cowie & Brasier 1989,p. 85).
Age constraints on the CharnwoodForest assemblage are poor.Taxonomic similarities withMistaken Point suggest an agearound 565 Ma; a K-Ardetermination from a nearbyporphyroid emplacement yielded adate of 583 ± 25 Ma.
555 Ma: White Sea
The two most abundant and diverseEdiacaran trace and body fossilassemblages are those from theWhite Sea coast of Russia and fromthe Flinders Ranges in SouthAustralia, which together accountfor 60% of the well-described

Ediacaran taxa.
"Many exposures in the White Searegion contain known Ediacaranbiotas; however, the best fossiloccurrences are found along theshoreline cliffs at Zimnie Gory.These unmetamorphosed andnondeformed (except for present-day cliff-face slumping)siliciclastic rocks belong to theuppermost Ust’ Pinega Formationand form the northern flank of theMezen Basin along the southeastflank of the Baltic Shield"(Martin et al. 2000, p. 842).Zircons from a volcanic ash in thelower part of the sequencepreserved between Medvezhiy andYeloviy Creeks (ibid., fig. 2)yielded a date of 555.3 3 Ma,the minimum age for the "oldestdefinitive triploblasticbilaterian, Kimberella, and theoldest well-developed tracefossils; and it documents thatspectacularly diverse andpreserved Ediacaran fossils formedmore than 12 million years beforethe base of the Cambrian" (ibid.,p. 843).
The fossil Kimberella, originallydescribed from southern Australiabut subsequently found elsewhere,including from the White Sea innorthern Russia, has beenpersuasively reconstructed as abenthic bilaterian animal with anon-mineralised, univalved shell,resembling a mollusc (Fedonkin &Waggoner 1997). Thisinterpretation provides evidencefor the existence of largetriploblastic metazoans in thePrecambrian, and requires theorigin of the higher groups ofprotostomes to have occurred deepin the Precambrian, at least prior

to ~560 Ma.
Presumed radula scratchings arefound at Zimnie Gory, as isHiemalora, another problematic formwhich cannot with certainty becategorised as either a bodyfossil or a feeding trace (Martinet al. 2000, p. 844). Irrespective,this evidence establishes theexistence of actively crawlingorganisms, almost certainlybilaterians, and almost certainlyabove the grade of planariansbecause of the implied hydrostaticskeleton.
Most provocative of all theEdiacaran forms are thoseexhibiting real or apparentmetamerism (e.g. Dickinsonia andSpriggina). Most are small, thoughsome of the dickinsonids can beenormous: up to ~1 metre. Severalauthors – notably M.A. Fedonkin(e.g. 1986) and A. Yu. Ivantsov(e.g. 2001) – argue that many ofthese organisms arepseudosegmented, with segmentsalternating on either side of themid line, thereby casting doubt ontheir bilaterian affinities.However, their approach has beenstrongly dependent on a twodimensional body plan analysis ofsymmetry, with little regard forthe original three dimensionalarchitecture of the organisms.Similarly, Bergström (1990, figure2) illustrates four taxa withapparent alternation of regularelements on each side of the axis;but in each case the sketchesrepresent unrestored images offlattened animals. James Gehlingleaves little doubt where hestands on the matter: "Toreconstruct the small Ediacaransegmented taxa as other than

vagile metazoans requires anappeal to the absurd" (Gehling1991, p. 205; also see pp. 199 and203.)
?555 Ma: South Australia
Material from the Ediacara Hills(Flinders Ranges) has still notbeen precisely dated; it isassumed to be approximately coevalwith the White Sea fossils, in theregion of 555 Ma (see below), butit could be as young as the +1 to+2‰ 13C interval, dated at 549 to543 Ma in southern Namibia (Martinet al. 2000, p. 844). It is theassemblage from this site that ismost widely associated with theEdiacaran biota.
The Ediacara Hills locality isalso the provenance of theearliest taxonomically-resolvedporiferan, Paleophragmodictyareticulata, and the possibleechinoderm, Arkarua adami.
Although best known for the‘classical’ body fossils, theregion also provides interestingtraces. One ichnotaxon has beeninterpreted as the rudulascratchings of a mollusc (possiblyKimberella).
[Needs a general statement aboutdiscovery and early work on bodyfossils.]
549 to 543 Ma: The Nama Group
The Nama Group is a thick (> 3 km)shallow marine and fluvialforeland basin succession,partitioned into northern andsouthern sub-basins by anintervening arch, across whichmost stratigraphic units thin,located in southern Namibia. Theage range of the Ediacaranassemblages from the Nama Group is
[Grotzinger et al. 1995 also mentions"complex spiral burrows" – find outmore.]

the interval 548.8 1 to 543.3 1 Ma (Grotzinger et al. 1995).
In addition to typical Ediacarantaxa, such as the cosmopolitanPteridinium, the shelly fossilCloudina first appears slightlybelow the earliest Ediacaranfossils, extends throughout theEdiacaran range, and into theCambrian. A second shelly taxon,Namacalathus (the "goblet-shapedshelly fossils" of Grotzinger et al.1995) coexists with Cloudina fromat least 545 Ma through into theCambrian.
549 to 543 Ma: Southwestern Mongolia
Ediacaran faunas and, notably,unquestionable hexactinellid(glass sponge) spicules, have beenreported from limestones justabove a phosphorite-chert-blackshale marker bed in the upperTsagaan Gol Formation ofsouthwestern Mongolia (Brasier etal. 1997). The dates have beenestablishedchemostratigraphically, based oncarbon and 87Sr/86Sr isotopecorrelations.
Disappearance of the Ediacaran Assemblage
Decline
Although some taxa are now knownto have persisted, and others mayhave evolved into different forms,most of the Ediacarans simplyvanish from the fossil record nearthe beginning of the Cambrian. Thecharacteristic assemblage persistsin full bloom – at least in

Namibia – right up until theVendian-Cambrian boundary afterwhich the assemblage, as a whole,abruptly disappears. It isuncertain whether a massextinction event struck at thistime, or if we are simplyobserving the closure of some formof "taphonomic window" – both havebeen suggested.
One school of thought holdsthat Ediacarans may havebeen largely wiped out –possibly by the supposedKotlin nutrient crisis, seeBrasier 1992 – immediatelyprior to the Vendian-Cambrian boundary.
"In the past few years, evidencehas accumulated for a remarkableperturbation in the carbon cycleclose to the Proterozoic-Cambrianboundary. Globally distributedsedimentary successions document astrong (7 to 9 per mil) but short-lived negative excursion in thecarbon-isotopic composition ofsurface seawater at thestratigraphic breakpoint betweenEdiacaran-rich fossil assemblagesand those that document thebeginning of true Cambriandiversification. The causes ofthis event remain uncertain, butthe only comparable events in themore recent Earth history coincidewith widespread extinction – forexample, the Permo-Triassiccrisis, when some 90% of marinespecies disappeared, is marked byan excursion similar to butsmaller than the Proterozoic-Cambrian boundary event. Anearliest Cambrian increase inbioturbation shuttered thetaphonomic window on Ediacaran

biology. Thus, while Chengjiangand Sirius Passet fossils indicatethat Ediacaran-grade organismswere not ecologically important bythe late Early Cambrian,biostratigraphy admits thepossibility that Ediacarans wereeaten or outcompeted by Cambriananimals. It is biogeochemistrythat lends substance to thehypothesis that Ediacaran andCambrian faunas are separated bymass extinction" (Knoll & Carroll1999, p. 2135).
In Oman, the ‘early’ SSFs, Cloudinaand Namacalathus, are reported togo extinct very shortly after theVendian-Cambrian boundary, at542.0 ± 0.5 Ma (Kerr 2002).
Other researchers observethat a mass extinction eventis not necessary to explainthe disappearance of theEdiacarans from the fossilrecord; conditions maysimply have ceased to befavourable to the unique‘Ediacaran preservation’with the arrival of morenumerous and more diversescavenging and bioturbatingorganisms.
The preservational characteristicsof typical Ediacaran assemblagesare undeniably unusual(‘characteristic’ might be abetter word), and evidence formore widely spread and deeperbioturbation certainly doesincrease sharply at the base ofthe Cambrian. Indeed, as we haveseen, the lower boundary of theCambrian is now defined by theoccurrence of the burrow tracefossil, Trichophycus pedum. However,to offer this as a complete


White Sea coast, and the disc-likeKullingia, occurs in the basalCambrian Uratanna Formation of theFlinders Ranges (Jensen et al.1998).
From Lower Cambrian strata on theDigermul Peninsula, Norway, Crimesand McIlroy 1999 describe thewidely occurring Ediacaranspecies, Nimbia occlusa and Aspidellaterranovica (as Tirasiana sp.), fromapproximately 80 m above the baseof the Vendian–Cambrian boundary(Nemakit-Daldynian), and a furtherspecimen of Aspidella terranovica (thistime as ‘Cyclomedusa’ sp.) fromabout 600 m above the boundary, inrocks of trilobite-bearing age(Atdabanian; as indicated byCruziana).
Ediacarans have been known fromthe Great Basin, California, atleast since 1991. A number oftaxa, including ?Tirasiana disciformis,cf. Swartpuntia, Cloudina-like tubes,and Ernietta plateauensis, have beendescribed from several localities(Horodyski 1991; Hagadorn 1998;Hagadorn et al. 2000). In thisregion, Swartpuntia persists throughseveral hundred metres of section,extending up as far as the Nevadellatrilobite zone (Atdabanian).
Middle Cambrian
Simon Conway Morris (1989, 1993,1998) claims to recogniseEdiacaran forms hiding among‘conventional’ Cambrian faunas. Hecites as examples Thaumaptilon(Conway Morris 1998, pp. 28-29),Mackenzia (ibid., pp. 83-84), andEmmonsaspis (ibid., p. 134, note 7).Thaumaptilon, from the BurgessShale (Middle Cambrian), which

Conway Morris believes to be aconventional pennatulacean, isproposed as a relative of formssuch as Charniodiscus (fig. 4A). Thecomparison is unsatisfying,however. The holdfast ofThaumaptilon in no way resemblesthe disc-shaped structure socharacteristic of many Ediacaranfronds. Moreover, thepennatulacean idea itself requiresfurther testing yet; as Nielsen2001, p. 59, notes the branches ofboth Charniodiscus and Thaumaptilon"were united with a membrane whichmakes the interpretation dubiouson functional grounds, and thestructures tentatively interpretedas polyps are very small and showno tentacles." Also note that the"Burgess Shale fronds lackevidence of the structuralcomplexity found in the primarybranches of Charniodiscus, and maybe structurally closer to otherEdiacaran fronds, such asPteridinium" (Gehling 1991, p. 204).
Upper Cambrian
Youngest and most intriguing arethe Upper Cambrian Ediacarans froma turbidite sequence exposed atBooley Bay, near Duncannon in Co.Wexford, Ireland, which includestwo taxa: ‘Ediacaria’ booleyi and theubiquitous Nimbia occlusa (Crimes etal. 1995). The ‘Ediacaria’ taxon ispreserved three-dimensionallythrough nearly 100 m of sediment.Preservational details suggest theorganism possessed a rigid wall.The Booley Bay occurrence is datedby acritarchs of sufficient"diversity and quantity toconstrain biostratigraphically therelative age of this succession …to the upper part of the UpperCambrian” (Moczydlowska & Crimes

1995, p. 125), indicating that atleast some Ediacarans co-existedwith ‘modern’ taxa for perhaps 20or 30 Ma – and certainly survivedthe Cambrian Explosion.
(Read more.)
Transition to Cambrian Faunas
Whether by mass extinction or someother mechanism, soft-bodiedfossil lagerstätten such as theChengjiang fauna indicate thatEdiacaran-grade organisms were nolonger ecologically significant bythe Botomian (late EarlyCambrian). Although some taxapersisted throughout the Cambrian,as we have seen, most of theEdiacarans simply vanish from thefossil record near the beginningof the Cambrian.
"We cannot tell how abruptly theEdiacaran Faunas became extinct,but only a very small number arerepresented by possiblesurvivors..." (Briggs et al. 1994,p. 46).
"Although most Ediacaran fossilshave no post-Proterozoic record,they were not immediatelysucceeded in lowermost Cambrianrocks by diverse crown groupbilaterians. Earliest Cambrianassemblages contain few taxa, andthe diversity of trace and bodyfossils grew only over aprotracted interval. Hyoliths andhalkierids (extinct forms thoughtto be related to mollusks), trueconchiferan mollusks and, perhaps,chaetognaths enter the recordduring the first 10 to 12 millionyears of the Cambrian, but crown-
Fig 5: The horizontal burrow tracefossil, Trichophycus (formerlyPhycodes) pedum defines the lowerboundary of the Cambrian in thereference section at Fortune Head,southeastern Newfoundland. It hasbeen suggested that newly evolved,burrowing organisms like this mayhave closed the taphonomic door onthe peculiar ‘Ediacaranpreservation’. [Image courtesy ofDr. Gerd Geyer, Institut fürPaläontologie, Bayerische Julius-Maximilians-Universität, Würzburg,Germany.]

group fossils of most otherbilaterian phyla appear later: theearliest body fossils ofbrachiopods, arthropods,chordates, and echinoderms allpost-date the beginning of theperiod by 10 to 25 million years.Trace fossils suggest earlierappearances for some groups,notably arthropods, but theobservation remains that the EarlyCambrian contains considerabletime for the assembly anddiversification of crown groupmorphologies" (Knoll & Carroll1999).
References
Ayala, Francisco José; Rzhetsky, Andrey; Ayala, Francisco J. 1998: Originsof the Metazoan Phyla: Molecular Clocks Confirm Paleontological Estimates. Proceedings ofthe National Academy of Sciences of the USA 95: 606-611.
Blaker, M.R.; Peel, J.S. 1997: Lower Cambrian trilobites from North Greenland. MoGGeoscience v. 35.
Brasier, M.D. 1992: Introduction. Background to the Cambrian Explosion. Journal ofthe Geological Society, London 149: 585-587.
Briggs, Derek E.G.; Erwin, Douglas H.; Collier, Frederick J.; Clark, Chip1994: The Fossils of the Burgess Shale. Smithsonian.
Clarkson, E.N.K. 1993: Invertebrate Paleontology and Evolution (3rd ed.) Chapmanand Hall.
Conway Morris, Simon 1998: The Crucible of Creation. Oxford.
Conway Morris, Simon 2000: The Cambrian "explosion": Slow-fuse or megatonnage?Proceedings of the National Academy of Sciences of the USA 97: 4426-4429.
Crimes, T.P.; Insole, A.; Williams, B.J.P. 1995: A Rigid Bodied Ediacaran Biotafrom Upper Cambrian Strata in Co. Wexford, Eire. Geological Journal 30: 89-109.
Crimes, T.P.; McIlroy, D. 1999: A Biota of Ediacaran Aspect from Lower CambrianStrata on the Digermul Peninsula, Arctic Norway. Geological Magazine, v. 136: 633-642.
Dzik, J.; Ivantsov, A.Y. 1999: An Asymmetric Segmented Organism from the Vendian

of Russia and the Status of the Dipleurozoa. Hist. Biol. 13: 255-268.
Fortey, R.A.; Briggs, D.E.G.; Wills, M.A. 1996: The Cambrian Evolutionary‘Explosion’: Decoupling Cladogenesis from Morphological Disparity. Biological Journal ofthe Linnaean Society 57: 13-33.
Gehling, J.G. 1987: Earliest known echinoderm - a new Ediacaran fossil from the PoundSubgroup of South Australia. Alcheringa 11: 337-345.
Gehling, J.G. 2001: Proterozoic Ediacara Member Within the Rawnsley Quartzite, SouthAustralia. Petroleum Abstracts, 30.
Gehling, James G.; Narbonne, Guy M.; Anderson, Michael M. 2000: The FirstNamed Ediacaran Body Fossil, Aspidella terranovica. Palaeontology 43 (3): 427-456.
Gehling, J.G.; Rigby, J.K. 1996: Long Expected Sponges from the NeoproterozoicEdiacara Fauna of South Australia. Journal of Paleontology, 2, pp. 185-195.
Glaessner, Martin F.; Wade, Mary 1966: The Late Precambrian Fossils from Ediacara,South Australia. Palaeontology 9 (4), pp. 599-628.
Grotzinger, J.P.; Bowring, Samuel A.; Saylor, Beverly Z.; Kaufman, AlanJ. 1995: Biostratigraphic and Geochronologic Constraints on Early Animal Evolution.Science, 270: 598-604.
Gürich, G. 1933: Die Kuibis Fossilen der Nama-Formation von Sudwestafrika.Paläontologische Zeitschrift 15: 137-154.
Hagadorn, J.W. 1998: Restriction of a Late Neoproterozoic Biotype. Unpublished PhDdissertation, University of Southern California, Los Angeles.
Hagadorn, James W.; Fedo, Christopher M.; Waggoner, Ben M. 2000: EarlyCambrian Ediacaran-Type Fossils from California. Journal of Paleontology 74: 731-740.
Hofmann, H. J., Narbonne, G. M., and Aitken, J. D. 1990: Ediacaran Remainsfrom Intertillite Beds in Northwestern Canada. Geology 18: 1199-1202.
Horodyski, R.J. 1991: Late Proterozoic Megafossils from Southern Nevada. GeologicalSociety of America Abstracts with Programs 23: 163.
Jensen, S.; Runnegar, B.N. 2005: A complex trace fossil from the Spitskop Member(terminal Ediacaran-?Lower Cambrian) of southern Namibia. Geological Magazine 142:561-569.
Knoll, Andrew H.; Carroll, Sean B. 1999: Early Animal Evolution: Emerging Views

from Comparative Biology and Geology. Science 284 (5423): 2129-2137.
Knoll, Andrew H.; Walter, Malcolm R.; Narbonne, Guy M.; Christie-Blick,Nicholas 2006: A New Period for the Geologic Time Scale. Science 305:621-622.
Langille, G.B. 1974: Problematic Calcareous Fossils from the Stirling Quartzite, FuneralMountains, Inyo County, California. Gological Society of America Abstracts withPrograms 6: 204-205.
Li, Chia-Wei; Chen, Jun-Yuan; Hua, Tzu-En 1998: Precambrian Sponges withCellular Structures. Science v. 279, issue of 6 February 1998, pp. 879 - 882.
Martin, M.W.; Grazhdankin, D.V.; Bowring, S.A.; Evans, D.A.D.; Fedonkin,M.A.; Kirschvink, J.L. 2000: Age of Neoproterozoic Bilaterian Body and Trace Fossils,White Sea, Russia: Implications for Metazoan Evolution. Science v.288: 841-845.
McMenamin, Mark A.S. 1986: The Garden of Ediacara. Palaios v. 1, pp. 178-182.
McMenamin, M. A. S. 1996: Ediacaran biota from Sonora, Mexico. Proceedings of theNational Academy of Sciences of the USA 93: 4990-4993.
McMenamin, Mark A.S. 1998: The Garden of Ediacara. Columbia.
Moczydlowska, M.; Crimes, T.P. 1995: Late Cambrian Acritarchs and their AgeConstraints on an Ediacaran-Type Fauna from the Booley Bay Formation, Co. Wexford, Eire.Geological Journal 30: 111-128.
Nielsen, Claus 2001: Animal Evolution. Second ed. Oxford University Press.563 pp.
Runnegar, Bruce 1992: Chapter 3 – Evolution of the Earliest Animals. In, Schopf, J.William (ed) Major Events in the History of Life. Jones and Bartlett.
Seilacher, A. 1984: Late Precambrian and Early Cambrian Metazoa: Preservational or RealExtinctions? In Holland, H.D. and Trendall, A.F. (eds.) Patterns of Change inEarth Evolution, pp. 159-168. Springer Verlag.
Seilacher, A. 1992: Vendobionta and Psammocorallia. Journal of the GeologicalSociety, London 149:607-613.
Shu, D.-G.; Conway Morris, S.; Han, J. 2006 in Shu, D-G.; Conway Morris,S.; Han, J.; Li, Y.; Zhang, X.-L.; Hua, H.; Zhang, Z.-F.; Liu, J.-N.;Guo, J.-F.; Yao, Y.; Yasui, K. 2006: Lower Cambrian vendobionts from China andearly diploblast evolution. Science 312: 731-734.
Waggoner, Ben 1999: The Garden of Ediacara, by Mark A. S. McMenamin (book

review). Palaeontologia Electronica, 15 March 1999.
Wray, Gregory A.; Levinton, Jeffrey S.; Shapiro, Leo H. 1996: MolecularEvidence for Deep Precambrian Divergences Among Metazoan Phyla. Science v. 274(5287), issue of 25 Oct 1996, pp. 568 - 573.
Xiao, Shuhai; Laflamme, Marc 2008: On the eve of animal radiation:phylogeny, ecology and evolution of the Ediacara biota. Trends in Ecologyand Evolution 24 (1): 31-40.
Copyright © 2022 FDOKUMEN




















