
A New Pleistocene bird assemblage from the Southern Pampas (Buenos Aires, Argentina)
17
A New Pleistocene bird assemblage from the Southern Pampas (Buenos Aires, Argentina) Marcos M. Cenizo a,b, ⁎, Federico L. Agnolin c,b , Lucas H. Pomi d a Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Uruguay 151, 6300 Santa Rosa, La Pampa, Argentina b Fundación de Historia Natural “Félix de Azara”, Departamento de Ciencias Naturales y Antropología, CEBBAD, Universidad Maimónides, Hidalgo 775, C1405BDB Ciudad Autónoma de Buenos Aires, Argentina c Laboratorio de Anatomía Comparada y Evolución de los Vertebrados, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Av. Angel Gallardo, 470, 1405 Buenos Aires, Argentina d Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Calle 122 y 60, 1900 La Plata, Argentina abstract article info Article history: Received 6 February 2014 Received in revised form 5 December 2014 Accepted 5 December 2014 Available online 13 December 2014 Keywords: Marine Isotope Stage 3 Interstadial Avifaunas Luján Formation Pampean Region Paleoenvironments In the present paper we report the most complete fossil avifauna from the southern cone of South America. The specimens here described were collected in the Paso Otero locality, at the middle stream of the Quequén Grande River, Pampean Region (Buenos Aires Province, Argentina). The fossiliferous levels belong to the La Chumbiada Member of Luján Formation (37,800 ± 2300 yr BP, late Pleistocene). The assemblage is represented by 60 spec- imens belonging to at least 22 taxa. The radiocarbon age indicates that Paso Otero birds represent the oldest late Pleistocene avifauna known from the South American continent and the first coming from a clear interstadial event (MIS3). Available evidence indicates similar environmental conditions during the MIS3 interstadial in the Pampean Region of Argentina and the post-Last Glacial Maximum stadials in southeastern Brazil. The absence of significant extinction events in the latest Pleistocene–early Holocene avifaunas of Brazil and Argentina suggests that the main avifaunal composition along southeastern continental lowlands may not be affected by the cyclic retraction and extension of open environments. On the contrary, the fossil record of Peru and Ecuador demonstrates that a large number of taxa, including aquatic and predatory-scavenger birds, have gone extinct along Andean environments at the Pacific coast, and late Pleistocene climatic oscillations may have played a more dramatic impact in these avian communities. © 2014 Elsevier B.V. All rights reserved. 1. Introduction The fossil record of birds in the Pleistocene of South America, includ- ing the Pampean Region of Argentina, is rather patchy and biased, in clear contrast to its well known mammalian faunas (Tambussi, 1995). Most reports belong to isolated and rather fragmentary specimens, and only some scarce authors describe more than a single avian remain from each fossiliferous locality (Tonni and Laza, 1980; Tonni and Fidalgo, 1982; Deschamps, 2005). However, there are some sparse lo- calities along the continent that yielded abundant late Pleistocene birds remains. Regrettably, these continental assemblages are rather distant each other and most of them located at intertropical South America (Winge, 1888; Campbell, 1976, 1979, 1982; Guérin et al., 1993, 1996). In this way, no single representative Pleistocene avifauna was reported up to now for the southern cone. Here we report a rich late Pleistocene avian assemblage from the Pampean Region, at the Paso Otero locality, southern Buenos Aires Province, Argentina (Fig. 1A,B). Previous works in this locality mentioned the presence of some fossil birds, including Pterocnemia cf. P. pennata (D'Orbigny, 1834), Nothura darwinii Gray, 1867, Chloephaga sp., Fulica leucoptera Vieillot, 1817, and Cyanoliseus cf. C. patagonus (Vieillot, 1817) (Tonni and Laza, 1980), together with diverse mamma- lian remains (Prado et al., 1987). Recent work resulted in the recovery of the most abundant and diverse Pleistocene avifauna known from the Pampas and one of the richest vertebrate faunas from this interval in Argentina. Excavations yielded a large amount of avian remains, includ- ing 60 recognizable specimens representing at least 22 taxa. The aim of the present paper is to describe and analyze this assemblage, and to discuss its paleoenvironmental and paleobiogeographical implications. 2. Materials and methods The fossil assemblage from Paso Otero locality (POP1) comprises 1432 skeletal elements corresponding to fishes, anurans, birds, and mammals (Pomi and Tonni, 2011). The excavations of POP1 took place from 2007 to 2009, and covered six grids of 1 m 2 . The systematic arrangement follows the proposals of Dickinson and Remsen (2013, and literature cited therein). Osteological terminology follows Baumel and Witmer (1993) and Livezey and Zusi (2006). Ana- tomical terminology of the quadrate bone is based on Elzanowski et al. Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81 ⁎ Corresponding author at: Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Uruguay 151, 6300 Santa Rosa, La Pampa, Argentina. E-mail address: [email protected] (M.M. Cenizo). http://dx.doi.org/10.1016/j.palaeo.2014.12.009 0031-0182/© 2014 Elsevier B.V. All rights reserved. Contents lists available at ScienceDirect Palaeogeography, Palaeoclimatology, Palaeoecology journal homepage: www.elsevier.com/locate/palaeo
-
Upload
fundacionazara -
Category
Documents
-
view
0 -
download
0
Transcript of A New Pleistocene bird assemblage from the Southern Pampas (Buenos Aires, Argentina)

Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
Contents lists available at ScienceDirect
Palaeogeography, Palaeoclimatology, Palaeoecology
j ourna l homepage: www.e lsev ie r .com/ locate /pa laeo
A New Pleistocene bird assemblage from the Southern Pampas(Buenos Aires, Argentina)
Marcos M. Cenizo a,b,⁎, Federico L. Agnolin c,b, Lucas H. Pomi d
a Facultad de Ciencias Exactas y Naturales, Universidad Nacional de La Pampa, Uruguay 151, 6300 Santa Rosa, La Pampa, Argentinab Fundación de Historia Natural “Félix de Azara”, Departamento de Ciencias Naturales y Antropología, CEBBAD, Universidad Maimónides, Hidalgo 775,C1405BDB Ciudad Autónoma de Buenos Aires, Argentinac Laboratorio de Anatomía Comparada y Evolución de los Vertebrados, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Av. Angel Gallardo, 470, 1405 Buenos Aires, Argentinad Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata, Calle 122 y 60, 1900 La Plata, Argentina
⁎ Corresponding author at: Facultad de Ciencias ExaNacional de La Pampa, Uruguay 151, 6300 Santa Rosa, La
E-mail address: [email protected] (M.M. Ce
http://dx.doi.org/10.1016/j.palaeo.2014.12.0090031-0182/© 2014 Elsevier B.V. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 6 February 2014Received in revised form 5 December 2014Accepted 5 December 2014Available online 13 December 2014
Keywords:Marine Isotope Stage 3InterstadialAvifaunasLuján FormationPampean RegionPaleoenvironments
In the present paper we report the most complete fossil avifauna from the southern cone of South America. Thespecimens here described were collected in the Paso Otero locality, at themiddle stream of the Quequén GrandeRiver, Pampean Region (Buenos Aires Province, Argentina). The fossiliferous levels belong to the La ChumbiadaMember of Luján Formation (37,800 ± 2300 yr BP, late Pleistocene). The assemblage is represented by 60 spec-imens belonging to at least 22 taxa. The radiocarbon age indicates that Paso Otero birds represent the oldest latePleistocene avifauna known from the South American continent and the first coming from a clear interstadialevent (MIS3). Available evidence indicates similar environmental conditions during the MIS3 interstadial inthe PampeanRegion of Argentina and the post-Last GlacialMaximum stadials in southeastern Brazil. The absenceof significant extinction events in the latest Pleistocene–early Holocene avifaunas of Brazil and Argentinasuggests that the main avifaunal composition along southeastern continental lowlands may not be affected bythe cyclic retraction and extension of open environments. On the contrary, the fossil record of Peru andEcuador demonstrates that a large number of taxa, including aquatic and predatory-scavenger birds, have goneextinct alongAndean environments at the Pacific coast, and late Pleistocene climatic oscillationsmay have playeda more dramatic impact in these avian communities.
© 2014 Elsevier B.V. All rights reserved.
1. Introduction
The fossil record of birds in the Pleistocene of South America, includ-ing the Pampean Region of Argentina, is rather patchy and biased, inclear contrast to its well known mammalian faunas (Tambussi, 1995).Most reports belong to isolated and rather fragmentary specimens,and only some scarce authors describe more than a single avian remainfrom each fossiliferous locality (Tonni and Laza, 1980; Tonni andFidalgo, 1982; Deschamps, 2005). However, there are some sparse lo-calities along the continent that yielded abundant late Pleistocenebirds remains. Regrettably, these continental assemblages are ratherdistant each other and most of them located at intertropical SouthAmerica (Winge, 1888; Campbell, 1976, 1979, 1982; Guérin et al.,1993, 1996). In this way, no single representative Pleistocene avifaunawas reported up to now for the southern cone.
Here we report a rich late Pleistocene avian assemblage fromthe Pampean Region, at the Paso Otero locality, southern BuenosAires Province, Argentina (Fig. 1A,B). Previous works in this locality
ctas y Naturales, UniversidadPampa, Argentina.nizo).
mentioned the presence of some fossil birds, including Pterocnemia cf.P. pennata (D'Orbigny, 1834), Nothura darwinii Gray, 1867, Chloephagasp., Fulica leucoptera Vieillot, 1817, and Cyanoliseus cf. C. patagonus(Vieillot, 1817) (Tonni and Laza, 1980), together with diverse mamma-lian remains (Prado et al., 1987). Recentwork resulted in the recovery ofthe most abundant and diverse Pleistocene avifauna known from thePampas and one of the richest vertebrate faunas from this interval inArgentina. Excavations yielded a large amount of avian remains, includ-ing 60 recognizable specimens representing at least 22 taxa. The aim ofthe present paper is to describe and analyze this assemblage, and todiscuss its paleoenvironmental and paleobiogeographical implications.
2. Materials and methods
The fossil assemblage from Paso Otero locality (POP1) comprises1432 skeletal elements corresponding to fishes, anurans, birds, andmammals (Pomi and Tonni, 2011). The excavations of POP1 took placefrom 2007 to 2009, and covered six grids of 1 m2.
The systematic arrangement follows the proposals of Dickinson andRemsen (2013, and literature cited therein). Osteological terminologyfollows Baumel and Witmer (1993) and Livezey and Zusi (2006). Ana-tomical terminology of the quadrate bone is based on Elzanowski et al.

Fig. 1.Geographic location of fossiliferous localities mentioned in the text. A, Late Pleistocene–early Holocene South American avian assemblages: 1. Talara (Piura Department, Peru); 2. LaCarolina (Santa Elena Province, Ecuador); 3. Toca da Janela da Barra do Antonião Cave (Piauí State, Brazil); 4. Minas Gerais caves (Minas Gerais State, Brazil); 5. Paso Otero (Buenos AiresProvince, Argentina). B, Location detail of Paso Otero fossiliferous locality.
66 M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
(2000). Comparisons were based on osteological specimens of extantspecies listed in Appendix 1. The cross in the text (†) indicates extincttaxa.
Bone measurements correspond to personal analyses and datasets published by previous authors: Tinamidae (Cenizo et al., 2012),Anatidae (Campbell, 1979; Louchart et al., 2005), Podicipedidae(Storer, 1976; Bocheński, 1994; Olson, 1995), Phoenicopteridae (Richand Walker, 1983), Threskiornithidae (Miller and Bowman, 1956;Campbell, 1979), Scolopacidae (Emslie, 2004), Cathartidae (Howard,1974; Campbell, 1979; Carr, 1981; Becker, 1986; Stucchi and Emslie,2005; Alvarenga et al., 2008), Accipitridae (Campbell, 1979), Falconidae(Campbell, 1979; Emslie, 1998; Olson, 2008), and Furnariidae (Noriega,1991).
Measurements (in mm) were taken with a digital vernier caliper tothe nearest 0.01mmand are expressed as follows:max–min,mean (n).
2.1 . Measurement abbreviations
D-cm, width of os metacarpale major; D-dist, depth of distal end; D-prox, depth of proximal end; L-ac, length from facies articularis sternalisto processus acrocoracoideus; L-lo, length from condylus lateralis to ca-pitulum oticum; L-pc, length from facies articularis sternalis to
processus procoracoideus; TL, total length; W-II, width of trochleametatarsi II; W-III, width of trochlea metatarsi III; W-IV, width of troch-lea metatarsi IV; W-dist, width of distal end; W-fo, width of faciesarticularis sternalis; W-la: width between the dorsal margin of cotylalateralis and the ventral margin of articular bone; W-qm, width be-tween the cotyla quadratojugalis and the condylus medialis; W-prox,width of proximal end; W-shaft, width of shaft.
2.2 . Institutional abbreviations
MACN, Museo Argentino de Ciencias Naturales, Buenos Aires,Argentina; MLP, Museo de La Plata, La Plata, Argentina.
3. Geographic and geologic setting
POP1 is located at the mid-stream of Quequén Grande River, insouthern Buenos Aires Province, Argentina (38°11′48″S, 59°06′56″ W;Fig. 1A,B). The Quequén Grande River scurry at the Bonaerean Inter-montane Area, from the Tandil Hill System to Atlantic Ocean. This sys-tem, together with the Ventania Hills System, interrupts an extensivegeographic region of plains dominated by grasslands (commonly called“Pampas”). The present-day climate of this area is temperate, with an
67M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
annual mean temperature ranging between 13 and 14.1 °C; annualmean rain is 830mm, andwinds predominate fromnorth/northwestern(Campo de Ferreras and Píccolo, 1997).
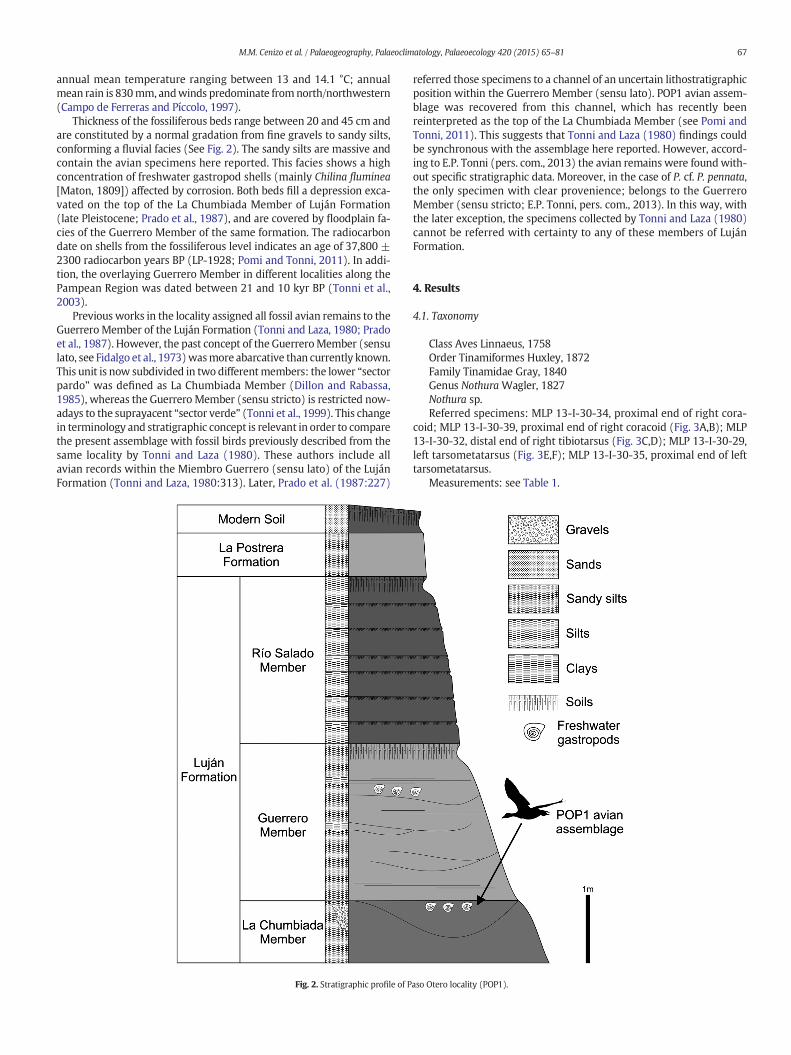
Thickness of the fossiliferous beds range between 20 and 45 cm andare constituted by a normal gradation from fine gravels to sandy silts,conforming a fluvial facies (See Fig. 2). The sandy silts are massive andcontain the avian specimens here reported. This facies shows a highconcentration of freshwater gastropod shells (mainly Chilina fluminea[Maton, 1809]) affected by corrosion. Both beds fill a depression exca-vated on the top of the La Chumbiada Member of Luján Formation(late Pleistocene; Prado et al., 1987), and are covered by floodplain fa-cies of the Guerrero Member of the same formation. The radiocarbondate on shells from the fossiliferous level indicates an age of 37,800 ±2300 radiocarbon years BP (LP-1928; Pomi and Tonni, 2011). In addi-tion, the overlaying Guerrero Member in different localities along thePampean Region was dated between 21 and 10 kyr BP (Tonni et al.,2003).
Previous works in the locality assigned all fossil avian remains to theGuerrero Member of the Luján Formation (Tonni and Laza, 1980; Pradoet al., 1987). However, the past concept of the GuerreroMember (sensulato, see Fidalgo et al., 1973)wasmore abarcative than currently known.This unit is now subdivided in two differentmembers: the lower “sectorpardo” was defined as La Chumbiada Member (Dillon and Rabassa,1985), whereas the Guerrero Member (sensu stricto) is restricted now-adays to the suprayacent “sector verde” (Tonni et al., 1999). This changein terminology and stratigraphic concept is relevant in order to comparethe present assemblage with fossil birds previously described from thesame locality by Tonni and Laza (1980). These authors include allavian records within the Miembro Guerrero (sensu lato) of the LujánFormation (Tonni and Laza, 1980:313). Later, Prado et al. (1987:227)
Fig. 2. Stratigraphic profile of P
referred those specimens to a channel of an uncertain lithostratigraphicposition within the Guerrero Member (sensu lato). POP1 avian assem-blage was recovered from this channel, which has recently beenreinterpreted as the top of the La Chumbiada Member (see Pomi andTonni, 2011). This suggests that Tonni and Laza (1980) findings couldbe synchronous with the assemblage here reported. However, accord-ing to E.P. Tonni (pers. com., 2013) the avian remains were foundwith-out specific stratigraphic data. Moreover, in the case of P. cf. P. pennata,the only specimen with clear provenience; belongs to the GuerreroMember (sensu stricto; E.P. Tonni, pers. com., 2013). In this way, withthe later exception, the specimens collected by Tonni and Laza (1980)cannot be referred with certainty to any of these members of LujánFormation.
4. Results
4.1. Taxonomy
Class Aves Linnaeus, 1758Order Tinamiformes Huxley, 1872Family Tinamidae Gray, 1840Genus Nothura Wagler, 1827Nothura sp.Referred specimens: MLP 13-I-30-34, proximal end of right cora-
coid; MLP 13-I-30-39, proximal end of right coracoid (Fig. 3A,B); MLP13-I-30-32, distal end of right tibiotarsus (Fig. 3C,D); MLP 13-I-30-29,left tarsometatarsus (Fig. 3E,F); MLP 13-I-30-35, proximal end of lefttarsometatarsus.
Measurements: see Table 1.
aso Otero locality (POP1).

68 M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
Description and comparisons: the size and proportions of the speci-mens arewithin the range of small-sized tinamids of the genusNothura.Additionally, the following features allow its assignment to this genus(Bertelli, 2002; Bertelli and Chiappe, 2005; Cenizo et al., 2012):
Coracoids MLP 13-I-30-34 and MLP 13-I-30-39: 1) foramenpneumaticum reduced on the dorsal surface (well developed in the re-maining Tinamidae, except Taoniscus); 2) facies articularis clavicularisdorsoventrally low and anteroposteriorly elongate, with a straightventral margin and a reduced tuberculum brachiale (wide facies inNothocercus, Eudromia, Rhynchotus, and Tinamotis; facieswith a concaveventral margin and a well-developed tuberculum brachiale in Tinamus,Crypturellus and Eudromia formosa [Lillo, 1905]); 3) impressio ligamentiacrocoracohumeralis donot converges distallywith the facies articularisclavicularis (converges with the facies in Tinamus, Crypturellus,Nothoprocta cinerascens [Burmeister, 1860], Tinamotis, and Eudromia);4) conspicuous distal projection of the processus acrocoracoideus withrespect to the facies articularis humeralis (at the same level or slightlymore distally projected in Crypturellus, Eudromia, and Tinamotis); 5) -absent foramen pneumaticum on the cotyla scapularis (present inCrypturellus, Eudromia, Tinamotis ingoufi [Oustalet, 1890], Rhynchotus,and Nothoprocta); 6) narrow processus acrocoracoideus (wide inEudromia, Tinamotis, and Nothocercus); 7) distinctive scar (impressiomusculi biceps brachii) on the dorsal surface of the processusacrocoracoideus (absent in Tinamus, Nothocercus, and Crypturellus).
Tibiotarsus MLP 13-I-30-32: 1) condylus lateralis transversely widerthan the condylus medialis (subequal in Nothoprocta); 2) in anteriorview, suboval condylus lateralis (proximally acute in Tinamus andNothocercus, subcircular in Tinamotis); 3) condylus medialis less proxi-mally extended than the condylus lateralis (much shorter in Rhynchotus,E. formosa, Tinamus,Nothocercus and some Crypturellus spp.; subequal inTinamotis); 4) groove extending along the caudal margin of thedepressio epicondylaris medialis (reduced or poorly developed inTinamus and Tinamotis, deep depressio on the cranial margin inCrypturellus and Nothocercus); 5) absent transverse groove on the prox-imal margin of area intercondylaris (present in Tinamus, Crypturellusand Nothocercus); 6) distal margin of the pons supratendineus at thesame level than the proximal margin of the condylus lateralis (distallylocated in Tinamus and Nothocercus, more proximal in Tinamotis);7) proximal margin of the condylus medialis slightly convex and adja-cent to the distal opening of the canalis extensorius (strongly convexand separated from the distal opening by a conspicuous depression inNothocercus, Rhynchotus, Eudromia and Tinamotis).
Tarsometatarsi MLP 13-I-30-29 and MLP 13-I-30-35: 1) inplantar view, subtriangular-shaped hypotarsus (square-shaped inNothocercus); 2) cristamedialis hypotarsi present (absent in Eudromia);3) two cristae intermediae hypotarsi (only one in Tinamotis); 4) in prox-imal view, subcircular sulcus hypotarsi medialis, with a verywidemedi-al opening (very wide sulcus in Eudromia, medial opening narrower inTinamus, Nothocercus, Crypturellus and Taoniscus); 5) slightly divergenttrochleae metatarsorum, with trochlea II distally extended at the samelevel of the incisiura intertrochlearis lateralis (highly divergenttrochleae with much more distally extended trochlea II in Tinamotis).
Remarks: NISP = 5, MNI = 2. Tonni and Laza (1980) reported thepresence of a tarsometatarsus (MLP 71-X-17-15) referable to the livingN. darwinii from an indeterminate level of the GuerreroMember (sensulato) of the Luján Formation at the POP1. Regrettably, the poorly re-solved taxonomywithin genusNothura, togetherwith its homogeneouspostcrania, does not allowus to refer the POP1 specimens to any species.Currently, the studied area is inhabited by N. darwinii and Nothuramaculosa (Temminck, 1815), both indistinguishable on the basis of iso-lated postcranial remains. These species are inhabitants of open countryareas inmost central and northern Argentina (Narosky and Di Giacomo,1993).
Genus Eudromia Geoffroy, 1832Eudromia sp.
Referred specimens: MLP 13-I-30-28, left femur (Fig. 3G,H); MLP13-I-30-30, distal end of right femur; MLP 13-I-30-21, proximal endof right tarsometatarsus; MLP 13-I-30-33, proximal end of righttarsometatarsus.
Measurements: see Table 1.Description and comparisons: the size and proportions of the speci-
mens correspond to the range of large-sized tinamids of the genusEudromia. However, the size of the distal end of the femur MLP 13-I-30-30 is larger than the maximum size observed in this genus, andapproaches the larger genera Tinamus and Tinamotis. However, the fol-lowing characters indicate that the new specimens are referable toEudromia (Bertelli, 2002; Bertelli and Chiappe, 2005; Cenizo et al.,2012; and pers. obs.): 1) robust and short femoral shaft (similar toTinamotis, more robust and shorter in Tinamus, elongate and thin in re-maining Tinamidae); 2) proximodistally elongate and laterallyprojected trochlea fibularis (shorter in Tinamus, not projected inTinamotis); 3) bifurcation of the intermuscularis caudalis line locatednear the mid-shaft (distally positioned in Tinamus); 4) strongly proxi-mally extended trochanter femoris (sharedwith Tinamus and Tinamotis;much lower in remaining Tinamidae; see also Tonni, 1977); 5) in caudalview, condylus medialis transversely wide and slightly caudally orient-ed (narrower and strongly caudally projected in Tinamotis); 6) inmedialview, proximal extension of the caudal articular surface of the condylusmedialis approaches the level of its cranial counterpart (as in Tinamus,Tinamotis, Nothoprocta ornata [Gray, 1867], and Nothoprocta perdicaria[Kittlitz, 1830]; extended more proximally than its cranial surface inother tinamids).
The proximal tarsometatarsus MLP 13-I-30-21 is referred toEudromia on the basis of the following characters (Bertelli, 2002;Cenizo et al., 2012): 1) subtriangular-shaped hypotarsus (square-shaped in Nothocercus); 2) crista medialis hypotarsi absent (pres-ent in remaining tinamids); 3) two cristae intermediae hypotarsi(only one in Tinamotis); 4) in proximal view, very wide but notsubcircular sulcus hypotarsi medialis (smaller and subcircular inother Tinamidae).
The distal end of tarsometatarsus MLP 13-I-30-33 is very fragmen-tary and lacks the hypotarsus. However, its size and proportions are in-distinguishable from that of Eudromia elegans d'Orbigny and Geoffroy,1832.
Remarks: NISP = 4, MNI = 2. It is possible that the size differenceobserved between the POP1 femur and extant specimens (Table 1)may reflect an artifact due to the low number of available comparativespecimens. However, another possibility is that these differences mayindicate a taxonomical distinction between Pleistocene and extantrepresentatives of the genus. In this way, it is remarkable the recentcommunication of a nearly complete Eudromia skeleton from the latePleistocene, that represents a possible extinct species much largerthan extant ones (Noriega et al., 2012).
Currently, in the area of study the subspecies E. elegans multiguttataConover, 1950 is a common inhabitant of open areas (Navas and Bo,1981).
Order Anseriformes (Wagler, 1831)Family Anatidae Vigors, 1825Anatidae gen. et sp. indet.Referred specimens: MLP 13-I-30-40, distal end of left tibiotarsus;
MLP 13-I-30-51, distal end of left tibiotarsus (Fig. 3I,J).Measurements: see Table 2.Remarks: NISP = 2, MNI = 2. The material represents a mid-sized
anatid similar in proportions to that of the living Anas sibilatrix Poeppig,1829, Anas bahamensis Linnaeus, 1758, Netta peposaca (Vieillot, 1816)and Dendrocygna bicolor (Vieillot, 1816). However, the poor preserva-tion of the remains precludes a more accurate identification.
Subfamily Dendrocygninae Reichenbach, 1849–50

Fig. 3. Fossil Tinamidae and Anatidae from Paso Otero locality. Nothura sp. (Tinamidae): proximal end of right coracoid (MLP 76-I-30-34) in dorsal (A), and lateral (B) views; distal end ofright tibiotarsus (MLP 76-I-30-32) in cranial (C), and medial (D) views; left tarsometatarsus (MLP 76-I-30-29) in dorsal (E), and plantar (F) views. Eudromia sp. (Tinamidae): left femur(MLP 76-I-30-28) in cranial (G), and caudal (H) views. Anatidae indet.: distal end of left tibiotarsus (MLP 76-I-30-51) in cranial (I), andmedial (J) views.Dendrocygna sp. (Anatidae): prox-imal end of right carpometacarpus (MLP 76-I-30-48) in ventral (K), and dorsal (L) views. Chloephaga sp. (Anatidae): distal end of right humerus (MLP 76-I-30-53) in cranial (M), and cau-dal (N) views. Anatini indet. 1: left tarsometatarsus of a juvenile specimen (MLP 76-I-30-50) in dorsal (O), and plantar (P) views. Anatini indet. 2: distal end of right tibiotarsus (MLP 76-I-30-49) in cranial (Q), and medial (R) views. cf. Callonetta sp. (Anatidae): right carpometacarpus (MLP 76-I-30-42) in ventral (S), and dorsal (T) views; distal end of right tarsometatarsus(MLP 76-I-30-44) in dorsal (U), and plantar (V) views. Anas cf. A. sibilatrix (Anatidae): left ulna (MLP 76-I-30-4) in dorsal (W), and ventral (X) views. Scale bars = 10 mm.
69M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
Genus Dendrocygna Swainson, 1837Dendrocygna sp.Referred specimens: MLP 13-I-30-48, proximal end of right
carpometacarpus (Fig. 3K,L); MLP 13-I-30-47, proximal end of leftfemur.
Measurements: see Table 2.Description and comparisons: on the basis of size and proportions,
the carpometacarpus MLP 13-I-30-48 belongs to a mid-sized anatid. Al-though fragmentary, is referred to Dendrocygna by having a continuousdorsal ring of the trochlea carpalis and lack of a prominent caudal notchpresent in other anatids (Woolfenden, 1961; Livezey, 1986; Livezey andMartin, 1988;Worthy and Lee, 2008). Additionally, Dendrocygna differsfrom other available anatids in the following combination of features:1) in dorsal view, gracile and very proximocranially extended processusextensorius; 2) straight apex of the processus extensorius, with the dis-tal margin more cranially projected than the proximal one; 3) in proxi-mal view, short and stout processus extensorius; 4) in distal view,robust processus pisiformis; 5) poorly caudally extended ventral ringof the trochlea carpalis.
The femur MLP 13-I-30-47 pertains to a mid-sized anatid similar insize and proportions to Dendrocygna spp., Lophonetta specularioides(King, 1828), andN. peposaca. Although thematerial is very incomplete,the referral to Dendrocygna is based on the following combination of
traits: 1) cranially projected trochanter femoris; 2) in medial view,intermuscularis caudalis line delimited proximally by a wide, elongate,and shallow medial depression; 3) trochanter femoris is rounded incontour when viewed laterally.
Remarks: NISP = 2, MNI = 1. The genus Dendrocygna is currentlyrepresented in the area of study by D. viduata and D. autumnalis(Narosky and Di Giacomo, 1993). These taxa are highly similar in sizeand proportions, precluding a specific referral of the fossil specimens.
Subfamily Anatinae Leach, 1820Tribe Tadornini Reichenbach, 1849–50Genus Chloephaga Eyton, 1838Chloephaga sp.Referred specimen: MLP 13-I-30-53, distal end of right humerus
(Fig. 3M,N).Measurements: MLP 13-I-30-53, W-dist: 22.5 (Chloephaga picta
[Gmelin, 1789]: 19.9–24.7 [8]; Coscoroba coscoroba [Molina, 1782]:23.9–26.3 [5]; Cygnus melancoryphus [Molina, 1782]: 24.8–27.0 [6]).
Description and comparisons: the specimen belongs to a largeanatid. Its morphology and proportions are only similar to that ofsheldgeese of the genus Chloephaga, being slightly smaller than theswans C. coscoroba and C. melancoryphus. The humerus MLP 13-I-30-53 is includedwithin Chloephaga on thebasis of the following characters
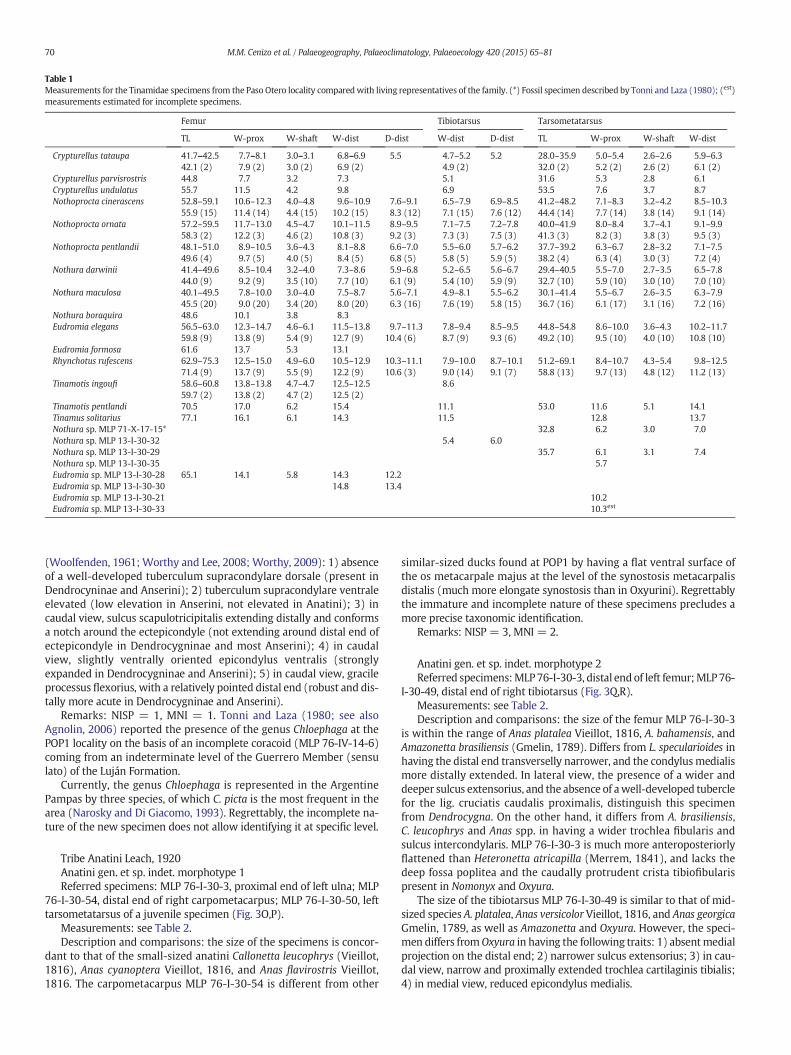
Table 1Measurements for the Tinamidae specimens from the Paso Otero locality comparedwith living representatives of the family. (*) Fossil specimen described by Tonni and Laza (1980); (est)measurements estimated for incomplete specimens.
Femur Tibiotarsus Tarsometatarsus
TL W-prox W-shaft W-dist D-dist W-dist D-dist TL W-prox W-shaft W-dist
Crypturellus tataupa 41.7–42.5 7.7–8.1 3.0–3.1 6.8–6.9 5.5 4.7–5.2 5.2 28.0–35.9 5.0–5.4 2.6–2.6 5.9–6.342.1 (2) 7.9 (2) 3.0 (2) 6.9 (2) 4.9 (2) 32.0 (2) 5.2 (2) 2.6 (2) 6.1 (2)
Crypturellus parvisrostris 44.8 7.7 3.2 7.3 5.1 31.6 5.3 2.8 6.1Crypturellus undulatus 55.7 11.5 4.2 9.8 6.9 53.5 7.6 3.7 8.7Nothoprocta cinerascens 52.8–59.1 10.6–12.3 4.0–4.8 9.6–10.9 7.6–9.1 6.5–7.9 6.9–8.5 41.2–48.2 7.1–8.3 3.2–4.2 8.5–10.3
55.9 (15) 11.4 (14) 4.4 (15) 10.2 (15) 8.3 (12) 7.1 (15) 7.6 (12) 44.4 (14) 7.7 (14) 3.8 (14) 9.1 (14)Nothoprocta ornata 57.2–59.5 11.7–13.0 4.5–4.7 10.1–11.5 8.9–9.5 7.1–7.5 7.2–7.8 40.0–41.9 8.0–8.4 3.7–4.1 9.1–9.9
58.3 (2) 12.2 (3) 4.6 (2) 10.8 (3) 9.2 (3) 7.3 (3) 7.5 (3) 41.3 (3) 8.2 (3) 3.8 (3) 9.5 (3)Nothoprocta pentlandii 48.1–51.0 8.9–10.5 3.6–4.3 8.1–8.8 6.6–7.0 5.5–6.0 5.7–6.2 37.7–39.2 6.3–6.7 2.8–3.2 7.1–7.5
49.6 (4) 9.7 (5) 4.0 (5) 8.4 (5) 6.8 (5) 5.8 (5) 5.9 (5) 38.2 (4) 6.3 (4) 3.0 (3) 7.2 (4)Nothura darwinii 41.4–49.6 8.5–10.4 3.2–4.0 7.3–8.6 5.9–6.8 5.2–6.5 5.6–6.7 29.4–40.5 5.5–7.0 2.7–3.5 6.5–7.8
44.0 (9) 9.2 (9) 3.5 (10) 7.7 (10) 6.1 (9) 5.4 (10) 5.9 (9) 32.7 (10) 5.9 (10) 3.0 (10) 7.0 (10)Nothura maculosa 40.1–49.5 7.8–10.0 3.0–4.0 7.5–8.7 5.6–7.1 4.9–8.1 5.5–6.2 30.1–41.4 5.5–6.7 2.6–3.5 6.3–7.9
45.5 (20) 9.0 (20) 3.4 (20) 8.0 (20) 6.3 (16) 7.6 (19) 5.8 (15) 36.7 (16) 6.1 (17) 3.1 (16) 7.2 (16)Nothura boraquira 48.6 10.1 3.8 8.3Eudromia elegans 56.5–63.0 12.3–14.7 4.6–6.1 11.5–13.8 9.7–11.3 7.8–9.4 8.5–9.5 44.8–54.8 8.6–10.0 3.6–4.3 10.2–11.7
59.8 (9) 13.8 (9) 5.4 (9) 12.7 (9) 10.4 (6) 8.7 (9) 9.3 (6) 49.2 (10) 9.5 (10) 4.0 (10) 10.8 (10)Eudromia formosa 61.6 13.7 5.3 13.1Rhynchotus rufescens 62.9–75.3 12.5–15.0 4.9–6.0 10.5–12.9 10.3–11.1 7.9–10.0 8.7–10.1 51.2–69.1 8.4–10.7 4.3–5.4 9.8–12.5
71.4 (9) 13.7 (9) 5.5 (9) 12.2 (9) 10.6 (3) 9.0 (14) 9.1 (7) 58.8 (13) 9.7 (13) 4.8 (12) 11.2 (13)Tinamotis ingoufi 58.6–60.8 13.8–13.8 4.7–4.7 12.5–12.5 8.6
59.7 (2) 13.8 (2) 4.7 (2) 12.5 (2)Tinamotis pentlandi 70.5 17.0 6.2 15.4 11.1 53.0 11.6 5.1 14.1Tinamus solitarius 77.1 16.1 6.1 14.3 11.5 12.8 13.7Nothura sp. MLP 71-X-17-15* 32.8 6.2 3.0 7.0Nothura sp. MLP 13-I-30-32 5.4 6.0Nothura sp. MLP 13-I-30-29 35.7 6.1 3.1 7.4Nothura sp. MLP 13-I-30-35 5.7Eudromia sp. MLP 13-I-30-28 65.1 14.1 5.8 14.3 12.2Eudromia sp. MLP 13-I-30-30 14.8 13.4Eudromia sp. MLP 13-I-30-21 10.2Eudromia sp. MLP 13-I-30-33 10.3est
70 M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
(Woolfenden, 1961;Worthy and Lee, 2008; Worthy, 2009): 1) absenceof a well-developed tuberculum supracondylare dorsale (present inDendrocyninae and Anserini); 2) tuberculum supracondylare ventraleelevated (low elevation in Anserini, not elevated in Anatini); 3) incaudal view, sulcus scapulotricipitalis extending distally and conformsa notch around the ectepicondyle (not extending around distal end ofectepicondyle in Dendrocygninae and most Anserini); 4) in caudalview, slightly ventrally oriented epicondylus ventralis (stronglyexpanded in Dendrocygninae and Anserini); 5) in caudal view, gracileprocessus flexorius, with a relatively pointed distal end (robust and dis-tally more acute in Dendrocygninae and Anserini).
Remarks: NISP = 1, MNI = 1. Tonni and Laza (1980; see alsoAgnolin, 2006) reported the presence of the genus Chloephaga at thePOP1 locality on the basis of an incomplete coracoid (MLP 76-IV-14-6)coming from an indeterminate level of the Guerrero Member (sensulato) of the Luján Formation.
Currently, the genus Chloephaga is represented in the ArgentinePampas by three species, of which C. picta is the most frequent in thearea (Narosky and Di Giacomo, 1993). Regrettably, the incomplete na-ture of the new specimen does not allow identifying it at specific level.
Tribe Anatini Leach, 1920Anatini gen. et sp. indet. morphotype 1Referred specimens: MLP 76-I-30-3, proximal end of left ulna; MLP
76-I-30-54, distal end of right carpometacarpus; MLP 76-I-30-50, lefttarsometatarsus of a juvenile specimen (Fig. 3O,P).
Measurements: see Table 2.Description and comparisons: the size of the specimens is concor-
dant to that of the small-sized anatini Callonetta leucophrys (Vieillot,1816), Anas cyanoptera Vieillot, 1816, and Anas flavirostris Vieillot,1816. The carpometacarpus MLP 76-I-30-54 is different from other
similar-sized ducks found at POP1 by having a flat ventral surface ofthe os metacarpale majus at the level of the synostosis metacarpalisdistalis (much more elongate synostosis than in Oxyurini). Regrettablythe immature and incomplete nature of these specimens precludes amore precise taxonomic identification.
Remarks: NISP = 3, MNI = 2.
Anatini gen. et sp. indet. morphotype 2Referred specimens:MLP76-I-30-3, distal end of left femur;MLP76-
I-30-49, distal end of right tibiotarsus (Fig. 3Q,R).Measurements: see Table 2.Description and comparisons: the size of the femur MLP 76-I-30-3
is within the range of Anas platalea Vieillot, 1816, A. bahamensis, andAmazonetta brasiliensis (Gmelin, 1789). Differs from L. specularioides inhaving the distal end transverselly narrower, and the condylusmedialismore distally extended. In lateral view, the presence of a wider anddeeper sulcus extensorius, and the absence of awell-developed tuberclefor the lig. cruciatis caudalis proximalis, distinguish this specimenfrom Dendrocygna. On the other hand, it differs from A. brasiliensis,C. leucophrys and Anas spp. in having a wider trochlea fibularis andsulcus intercondylaris. MLP 76-I-30-3 is much more anteroposteriorlyflattened than Heteronetta atricapilla (Merrem, 1841), and lacks thedeep fossa poplitea and the caudally protrudent crista tibiofibularispresent in Nomonyx and Oxyura.
The size of the tibiotarsus MLP 76-I-30-49 is similar to that of mid-sized species A. platalea, Anas versicolor Vieillot, 1816, and Anas georgicaGmelin, 1789, as well as Amazonetta and Oxyura. However, the speci-men differs fromOxyura in having the following traits: 1) absentmedialprojection on the distal end; 2) narrower sulcus extensorius; 3) in cau-dal view, narrow and proximally extended trochlea cartilaginis tibialis;4) in medial view, reduced epicondylus medialis.

Table 2Measurements for the Anatidae specimens from the Paso Otero locality comparedwith living similar-sized representatives of the family in southern South America. (†) Fossil taxa from the late Pleistocene of Talara, Peru (Campbell, 1979); (est) mea-surements estimated for incomplete specimens.
Ulna Carpometacarpus Femur Tibiotarsus Tarsometatarsus
TL W-prox W-dist W-prox D-prox W-dist W-prox D-prox W-dist D-dist W-dist D-dist TL W-prox W-dist
Dendrocygna viduata 103.5–105.0 10.1–10.6 8.1–9.1 11.3–11.6 6.1 7.1–7.3 10.6–11.6 9.6 10.9–12.2 9.2 9.3–9.5 10.0 60.2–60.8 9.9–11.0 10.5–11.9104.2 (2) 10.4 (2) 8.6 (2) 11.4 (2) 7.2 (2) 11.0 (4) 11.5 (4) 9.4 (3) 60.5 (4) 10.5 (4) 11.1 (4)
Dendrocygna bicolor 10.3–11.0 10.7–11.9 8.4–9.5 51.8 9.4–10.3 9.4–10.410.7 (2) 11.3 (2) 9.0 (2) 51.8 (2) 9.9 (2) 9.9 (2)
Oxyura vittata 56.2–58.3 5.9–6.5 5.5–5.8 7.2–7.7 3.6–4.1 4.8–5.1 8.9–9.9 5.9–6.6 8.6–8.9 6.9–7.3 7.6–8.1 7.3–7.9 31.4–33.9 7.9–8.6 7.5–8.256.7 (4) 6.1 (4) 5.7 (4) 7.5 (4) 3.8 (4) 4.9 (4) 9.5 (4) 6.3 (4) 8.8 (4) 7.0 (4) 7.9 (4) 7.5 (4) 32.8 (4) 8.2 (4) 7.9 (4)
Nomonyx dominicus 55.6 6.3 5.8 7.6 4.6 8.7 7.9 6.8 27.3 6.8 6.9Heteronetta atricapilla 55.8–63.0 7.2–8.0 7.0–7.4 8.7–9.3 4.2–4.4 5.2–6.2 7.7–8.3 6.0–6.6 7.3–8.0 6.0–6.2 6.2–6.8 6.9–7.1 30.1–33.1 6.7–6.9 7.1–7.2
58.8 (4) 7.7 (4) 7.1 (4) 9.0 (4) 4.4 5.8 (4) 8.1 (4) 6.3 (3) 7.5 (4) 6.1 (3) 6.5 (4) 7.0 (3) 31.5 (4) 6.8 84) 7.2 (4)Netta peposaca 81.1–82.1 10.4–10.5 9.8 12.3 6.9–7.0 10.2–10.7 11.2–11.3 8.8–9.0 42.3–44.3 9.1–9.6 8.9–9.4
81.6 (2) 10.5 (2) 9.8 (2) 12.3 (2) 7.0 (2) 10.5 (2) 11.3 (2) 9.0 (4) 43.2 (4) 9.4 (4) 9.2 (4)Amazonetta brasiliensis 64.3–66.3 8.3–8.9 8.1–8.6 10.3 4.6–4.9 6.7 7.8–9.3 7.5 8.9–9.1 7.2 6.9–7.5 7.6 37.0–38.4 7.3–7.8 7.7–7.9
65.3 (2) 8.6 (2) 8.4 (2) 4.8 (2) 8.6 (2) 9.0 (2) 7.2 (2) 37.7 (2) 7.5 (2) 7.8 (2)Callonetta leucophrys 54.7–58.3 6.8–7.5 6.9–6.9 8.8 3.9–4.1 5.6 6.8–7.9 6.4–6.6 7.4–8.0 5.6–6.2 5.8–6.6 6.6–6.7 29.6–31.4 6.5–7.1 6.4–7.0
56.5 (2) 7.1 (2) 6.9 (2) 4.0 (2) 7.3 (3) 6.5 (2) 7.6 (3) 5.9 (2) 6.1 (3) 6.6 (2) 30.3 (3) 6.7 (3) 6.6 (3)Lophonetta specularioides 82.2–94.7 10.7–11.8 9.5–10.8 13.4–13.6 5.9–7.1 7.7–8.0 9.9–12.9 10.3 11.7–12.9 9.7 9.4–10.4 10.5 51.0–56.9 10.3–11.3 10.4–11.0
89.0 (5) 10.9 (5) 10.1 (5) 13.5 (2) 6.6 (5) 7.8 (2) 11.3 (4) 12.5 (4) 10.0 (4) 53.5 (4) 10.8 (4) 10.7 (4)Anas versicolor 56.2–57.8 7.6–8.3 7.2–7.6 9.5–9.8 4.9–5.1 5.5–6.5 7.9–8.7 6.2–6.8 7.8–8.1 6.2–6.6 6.4–8.0 7.0–7.4 31.9–35.4 6.9–7.6 6.5–7.6
56.9 (7) 7.9 (7) 7.4 (7) 9.7 (7) 5.0 (7) 6.0 (7) 8.2 (10) 6.5 (4) 8.0 (9) 6.3 (4) 6.8 (10) 7.1 (4) 33.0 (10) 7.1 (10) 7.1 (10)Anas sibilatrix 72.6–81.5 8.9–11.4 9.1–10.8 12.3–14.6 6.4–8.6 6.9–7.2 9.5–11.3 8.0–8.7 9.9–10.7 7.9–8.2 6.8–9.2 8.7–9.0 38.5–43.1 8.0–9.3 8.0–9.4
76.8 (13) 10.4 (15) 10.7 (15) 13.6 (16) 7.4 (16) 7.0 (7) 10.9 (26) 8.4 (10) 10.3 (27) 8.0 (10) 8.4 (21) 8.8 (7) 40.6 (25) 8.5 (25) 8.2 (26)Anas georgica 9.2–10.4 9.5–10.4 7.6–8.2 36.9–42.5 8.2–8.6 8.2–8.9
9.9 (10) 10.0 (10) 7.9 (8) 40.3 (7) 8.4 (7) 8.6 (7)Anas flavirostris 53.9–59.2 7.3–7.8 6.3–7.7 8.9–9.9 5.2–5.3 4.9–6.2 7.9–9.4 6.4–7.1 7.9–8.7 6.2–6.9 6.2–7.5 6.6–7.0 31.9–36.2 6.2–7.4 6.1–7.5
56.1 (9) 7.5 (9) 7.2 (9) 9.4 (10) 5.2 (6) 5.6 (10) 8.6 (15) 6.7 (6) 8.3 (15) 6.4 (6) 6.7 (15) 6.8 (6) 33.6 (15) 6.8 (15) 6.5 (15)Anas platalea 63.9–68.2 8.2–9.1 8.2–8.9 10.6–11.1 5.3–5.7 6.1–7.6 8.4–10.1 7.7–8.2 8.5–9.4 6.9–7.6 6.7–7.9 7.4–8.6 37.2–40.2 7.8–8.6 7.6–8.1
65.9 (9) 8.6 (9) 8.5 (9) 10.8 (9) 5.5 (5) 7.0 (9) 9.4 (9) 7.9 (5) 9.0 (9) 7.2 (5) 7.4 (9) 8.0 (5) 38.9 (9) 8.0 (9) 7.9 (9)Anas cyanoptera 50.5–61.1 6.6–8.5 6.7–8.2 9.4–9.8 3.9–5.0 5.1–6.1 6.1–9.2 6.8–7.4 6.5–8.9 6.5–7.0 5.4–7.1 7.2–7.4 31.3–36.6 6.1–7.2 5.5–7.3
55.0 (9) 7.5 (9) 6.7 (9) 9.5 (5) 4.6 (7) 5.7 (5) 8.3 (12) 7.1 (5) 8.2 (12) 6.7 (5) 6.6 (12) 7.3 (5) 34.6 (12) 6.9 (12) 6.8 (12)Anas bahamensis 8.6–9.3 9.1–9.5 8.6–8.9
8.9 (2) 9.3 (2) 8.8 (2)“Callonetta” talarae† 53.0–53.6 6.8 6.5–6.7 4.0 6.1–6.5 30.8 6.6 6.3
53.3 (2) 6.6 (3) 6.3 (2)Nannonetta invisitata† 54.7–58.0 7.1–7.7 6.3–7.6 4.1–4.6 7.5–8.0 8.0 6.5–6.9 33.6–38.5 6.8–7.4 7.2–7.8
56.3 (4) 7.4 (11) 7.0 (14) 4.4 (12) 7.7 (5) 8.0 (3) 6.6 (5) 35.3 (7) 7.1 (9) 7.5 (7)Anas amotape† 56.0 7.6 7.6 4.3 7.0 6.3–6.5 35.6 7.3 7.2
4.3 (3) 6.5 (6)Anas sanctaehelenae† 6.0 29.5 6.6 6.2Anatidae indet. MLP 13-I-30-40 8.5 8.6Anatidae indet. MLP 13-I-30-51 8.8 9.3Dendrocygna sp. MLP 13-I-30-48 10.1 5.7Dendrocygna sp. MLP 13-I-30-47 11.4 10.2Anatini indet. 1 MLP 76-I-30-54 5.0Anatini indet. 1 MLP 76-I-30-50 32.8est 7.0est 7.1est
Anatini indet. 2 MLP 76-I-30-3 9.1 7.3Anatini indet. 2 MLP 76-I-30-49 7.4 7.8cf. Callonetta sp. MLP 76-I-30-42 8.9 4.4 5.8cf. Callonetta sp. MLP 76-I-30-44 6.4cf. A. sibilatrix MLP 76-I-30-4 75.6 9.5 8.6
71M.M
.Cenizoetal./Palaeogeography,Palaeoclim
atology,Palaeoecology420
(2015)65
–81

72 M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
Remarks: NISP = 2, MNI = 1. The available material is difficultto refer to any previously known anatid from POP1. Moreover, itshows some unique features (e.g., straight caudal margin of the cristatibiofibularis) that precludes a more accurate identification.
Genus Callonetta Delacour, 1936cf. Callonetta sp.Referred specimens: MLP 76-I-30-42, right carpometacarpus
(Fig. 3S,T); MLP 76-I-30-44, distal end of right tarsometatarsus(Fig. 3U,V); MLP 76-I-30-41, right tarsometatarsal shaft.
Measurements: see Table 2.Description and comparisons: the size of the carpometacarpus MLP
76-I-30-42 is within the range of living C. leucophrys, Anas cyanopteraVieillot, 1816, and A. flavirostris. The following combination of charac-ters closely resembles Callonetta: 1) in distal view, transversely narrowand dorsoventrally low facies articularis digitalis major; 2) smalland slightly dorsoventrally elongate facies articularis digitalis minor;3) in caudal view, wide os metacarpale minus, with a deeply concavecaudal surface; 4) in caudal view, very well-developed, deep, andproximodistally elongate fovea carpalis caudalis; 5) in ventral view,shallow pit located distocaudally to the processus pisiformis; 6) veryshort synostosis metacarpalis distalis.
The tarsometatarsi MLP 76-I-30-44 and MLP 76-I-30-41 are distin-guished from Dendrocygna and most South American Tadornini in thepresence of a well-developed sulcus on the trochlea metatarsi II(Cenizo and Agnolin, 2010). The specimens differ from Oxyurini in:1)more elongate, straighter, and transversely narrower shaft; 2) narrowtrochleae metatarsorum; 3) small foramen vasculare distalis; 4) elon-gate plantar surface of trochlea metatarsi III.
Remarks: NISP= 3, MNI= 2. This material cannot be clearly distin-guished from the extinct Callonetta talarae Campbell, 1979, a small duckrestricted to the late Pleistocene of Peru, Ecuador, and the early–middlePleistocene of Argentina (Campbell, 1976, 1979; Agnolin, 2006). Theliving C. leucophrys is a common inhabitant of the Buenos Aires Province(see Narosky and Di Giacomo, 1993).
Genus Anas Linnaeus, 1758Anas cf. A. sibilatrix Poeppig, 1829Referred specimen: MLP 76-I-30-4, left ulna (Fig. 3W,X).Measurements: see Table 2.Description and comparisons: size and proportions ofMLP 76-I-30-4
resemble A. sibilatrix, themore robust long-winged Anas species presentin Argentina. MLP 76-I-30-4 shares with A. sibilatrix the following com-bination of features: 1) very short, stout, and slightly curved shaft (elon-gate, thin and nearly straight shaft in Dendrocygna; very gracile andcurved in Oxyura and Nomonyx, gracile and thin in remaining anatids);2) robust, slightly dorsally projected, and distally extended tuberculumcarpale (more robust in Oxyurini; gracile, more dorsally projected andproximally positioned in remaining Anatidae); 3) slightly developedcondylus ventralis ulnaris (similar to C. leucoprhys, muchmore developedand protrudent in remaining Anatidae); 4) poorly developed incisuratuberculi carpalis (similar to A. versicolor, A. cyanoptera and Oxyurini;very deep in others Anatini); 5) in caudal view, elongate condylusdorsalis ulnaris (shorter and rounded in Dendrocygna and Oxyurini);6) transversely wide facies articularis radiale (reduced in Dendrocygna).
Remarks: NISP = 1, MNI = 1. Living A. sibilatrix occurs along allBuenos Aires Province, including neighboring areas to POP1 (Naroskyand Di Giacomo, 1993).
Order Podicipediformes (Furbringer, 1888)Family Podicipedidae (Bonaparte, 1831)Genus Rollandia Bonaparte, 1856Rollandia rolland Quoy and Gaimard, 1824Referred specimens: MLP 76-I-30-16, left coracoid (Fig. 4A,B); MLP
76-I-30-15, right quadrate.
Measurements: MLP 76-I-30-16, TL: 23.1 (R. rolland: 23.7–26.7 [15];Rollandia microptera [Gould, 1868]: 29.1–34.4 [6]; Podiceps major[Boddaert, 1783]: 39.1–49.3 [12]; Podiceps occipitalis Garnot, 1826:25.4–28.6 [12]; Podiceps gallardoi Rumboll, 1974: 32.7–33.3 [3];Podilymbus podiceps [Linnaeus, 1758]: 27.4–35.9 [42]; Tachybaptusdominicus [Linnaeus, 1766]: 20.3–24.2 [13]). MLP 76-I-30-15, L-lo: 6.3(R. rolland: 6.5–6.9 [6]; P. occipitalis: 6.3–6.5 [3]; P. major: 9.5–10.0 [4]).
Description and comparisons: the coracoid MLP 76-I-30-16 is indis-tinguishable from corresponding elements of living R. rolland, and couldbe clearly distinguished from the only similar-sized sympatric speciesP. occipitalis in the following traits: 1) smaller size; 2) very gracile andelongate shaft; 3) in dorsal view, transversely narrow facies articularissternalis; 4) straighter magro medialis; 5) poorly developed impressiolig. acrocoracohumeralis.
Remarks: NISP = 2, MNI = 1. Current distribution of R. rollandincludes all Argentine territory, being the most abundant greebe alongall its distribution (Olrog, 1979). The species is frequent along theBuenos Aires Province (Narosky and Di Giacomo, 1993).
Order Phoenicopteriformes Fürbringer, 1888Family Phoenicopteridae Bonaparte, 1831Phoenicopteridae gen. et sp. indet.Referred specimen: MLP 76-I-30-19, left trochlea metatarsi III
(Fig. 4C,D).Measurements: MLP 76-I-30-19, W-III: 7.49 (Phoenicopterus
chilensis Molina, 1782: 7.3–7.7 [4]; Phoenicoparrus andinus Philippi,1854: 6.2 [2]; Phoenicoparrus jamesi Sclater, 1886: 7.6 [1]).
Description and comparisons: among wading birds, this specimenmay be referred to the Phoenicopteridae by having very narrow troch-lea metatarsi III, with a deepmedian groove; and an elongate and prox-imally extended plantar articular surface (Rich and Walker, 1983).Moreover, it resembles Phoenicopterus rather than Phoenicoparrus onits robust proportions and, when viewed proximally, the presence of apoorly extended proximally trochlea metatarsi III. However, thesesubtle similarities do not allow us to refer this incomplete specimen toany genus with certainty.
Remarks: NISP = 1, MNI = 1. P. andinus and P. chilensis occur in theBuenos Aires Province; although the former is very scarce and occasion-al, the later is rather abundant andwell represented in a large variety ofaquatic environments (Narosky and Di Giacomo, 1993).
Order Pelecaniformes Sharpe, 1891Family Threskiornithidae Richmond, 1917Genus Plegadis Kaup, 1829Plegadis cf. P. chihi (Vieillot, 1817)Referred specimens: MLP 76-I-30-13, incomplete right
carpometacarpus; MLP 76-I-30-10, distal end of right humerus(Fig. 4E,F).
Measurements: MLP 76-I-30-13, TL: 48.9 (P. chihi: 47.2–58.0 [6];Eudocimus albus [Linnaeus, 1758]: 51.4–57.2 [2]; Theristicus caudatus[Boddaert, 1783]: 68.4–73.4 [4]); D-dist: 7.5 (P. chihi: 6.6–7.3 [6];E. albus: 7.2–7.5 [2]; T. caudatus: 10.4–13.3 [4]). MLP 76-I-30-10, W-dist:13.2 (P. chihi: 13.1–14.8 [4]; Phimosus infuscatus [Lichtenstein, 1823]:14.5 [1]; E. albus: 14.6–17.8 [5]; Eudocimus peruvianus† Campbell, 1979:17.2 [1]; T. caudatus: 21.7–22.4 [2]).
Description and comparisons: the carpometacarpus in Ardeidae andThreskiornithidae is similar in proportions and characters. However, thefollowing features distinguish MLP 76-I-30-16 from Ardeidae and allowsits referral to Threskiornithidae: 1) stout bone shaft (very long and nar-row in most Ardeidae); 2) wide fossa supratrochlearis; 3) in proximalview, trochlea carpalis with awell-defined cranial edge and fossa carpaliscranialis; 4) in proximal view, quadrangular-shaped trochlea carpalis,with a more robust ventral ring; 5) longer synostosis metacarpalisdistalis; 6) in dorsal view, distal extension of the sulcus tendineus crani-ally positioned and distally extended; 7) in dorsal and distal views, con-cave surface on the dorsocaudal portion of the metacarpal II tuberosity

Fig. 4. Fossil waterbirds from Paso Otero locality. Rollandia rolland (Podicipedidae): left coracoid (MLP 76-I-30-16) in ventral (A), and dorsal (B) views. Phoenicopteridae gen. et sp. indet.:left trochlea metatarsi III (MLP 76-I-30-19) in dorsal (C), and plantar (D) views. Plegadis cf. P. chihi (Threskiornithidae): distal end of right humerus (MLP 76-I-30-10) in cranial (E), andcaudal (F) views. cf. Numenius sp. (Scolopacidae): distal end of damaged left tarsometatarsus without trochlea III (MLP 76-I-30-8) in plantar (G), and dorsal (H) views. Fulica rufifrons(Rallidae): complete right coracoid (MLP 76-I-30-25) in dorsal (I), and ventral (J) views. Fulica sp. cf. F. leucoptera-F.rufifrons (Rallidae): distal end of left tibiotarsus (MLP 76-I-30-45)in medial (K), and cranial (L) views. Fulica sp. cf. F. armillata (Rallidae): distal end of left tarsometatarsus (MLP 76-I-30-26) in dorsal (M), and plantar (N) views. Scale bars = 10 mm.
73M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
(sensu Howard, 1929; prominent tubercle in Ardeidae); 8) in distal view,wider facet for digit III (sensu Howard, 1929; narrow in Ardeidae).
Within Threskiornithidae bothMLP 76-I-30-10 andMLP 76-I-30-16,are much smaller than corresponding elements of Platalea ajaja(Linnaeus, 1785) and the ibises Theristicus and Mesembrinibis. The sizeand shape are indistinguishable from that of living P. chihi, being smallerthan Plegadis ridgwayi (Allen, 1876), and E. albus. The humerus MLP 76-I-30-10 shares with P. chihi a tuberculum supracondylare dorsale moredorsally extended than in P. infuscatus.
Remarks: NISP = 2, MNI = 1. P. chihi is very frequent in aquaticenvironments along the grasslands of Buenos Aires Province (Naroskyand Di Giacomo, 1993).
Order Charadriiformes Garrod, 1874Charadriiformes gen. et sp. indet.Referred specimens: MLP 76-I-30-14, distal end of right ulna; MLP
76-I-30-18, proximal end of left femur.Measurements: MLP 76-I-30-14, W-dist: 3.7. MLP 76-I-30-18,
W-prox: 4.2.Description and comparisons: these specimens belong to very small
charadriiform birds that, based on size, may pertain to Calidris orCharadrius. However its incompleteness and uninformative naturedoes not allow us to refer them with more certainty.
Remarks: NISP = 2, MNI = 1.
Family Scolopacidae Rafinesque, 1815Genus Numenius Brisson, 1760cf. Numenius sp.Referred specimens: MLP 76-I-30-8, distal end of left tarsometatar-
sus (Fig. 4G,H); MLP 76-I-30-17, distal end of left tibiotarsus.Measurements: MLP 76-I-30-8, W-dist: 7.8 (Numenius americanus
Bechstein, 1812: 8.4–9.9 [7]; Tringa flavipes [Gmelin, 1789]: 5.0[1]).MLP 76-I-30-17, W-dist (estimated): 5.5.
Description and comparisons: the tarsometatarsus MLP 76-I-30-8may be referred to Scolopacidae on the basis of the following combina-tion of characters (Mayr and Smith, 2001; Ballmann, 2004): 1) reducedforamen vasculare distale, 2) very well-developed and highly raisedfossametatarsi I; 3) very distally extended trochleametatarsi III. Within
scolopacids, the large size is only comparable to the Numenius species.Moreover, the presence of trochlea metatarsi II poorly transverselycompressed and slightly proximally locatedwith respect to the trochleaIV, resembles Numenius, in contrast with other South American taxa (inmid-sized genera such as Tringa and Limosa, the trochlea metatarsi II ishighly transversely compressed and is much proximally positioned).
The tibiotarsus MLP 76-I-30-17 is much larger than the indetermi-nate charadriiform specimen cited above and other genera present inthe Pampean Region. In spite of similarities in size and features withNumenius its referral to this genus is only tentatively due to its poorlypreserved condition.
Remarks: NISP=2,MNI=1.Numenius is represented in the BuenosAires Province by N. phaeopus (Linnaeus, 1758) and N. borealis (Forster,1772). The first one is scarce in the Atlantic coast (Narosky and DiGiacomo, 1993), while N. borealis was a very common inhabitant ofinner aquatic environments along the province by the XIX century(Wetmore, 1939; Gollop et al., 1986) but it is currently near the extinc-tion, or totally extinct (Narosky and Di Giacomo, 1993).
Order Ralliformes Reichenbach, 1852Family Rallidae Vigors, 1825Genus Fulica Linnaeus, 1758Fulica leucoptera Vieillot, 1817Referred specimen: MLP 76-I-30-23, left quadrate bone.Measurements: see Table 3.Description and comparisons: the size of the quadrate MLP 76-I-
30-23 is similar to the corresponding element of F. leucoptera andFulica rufifrons Philippi and Landbeck, 1861, being smaller than Fulicaarmillata (Vieillot, 1817). It could be referred to F. leucoptera and dif-fers from the remaining species Fulica that inhabit the area on thebasis of the following traits: 1) in lateral view, cranioventrallyextended capitulum squamosum (not extended in F. rufifrons);2) short and stout pars otica (gracile and elongate in F. rufifrons, elon-gate in F. armillata); 3) incipient tuberculum subcapitulare (verywell-developed in F. rufifrons and some specimens of F. armillata);4) in ventral view, very robust condylus lateralis (gracile inF. rufifrons).

Table3
Measu
remen
tsfortheFu
licaspecim
ensfrom
thePa
soOtero
localityco
mpa
redwithliv
ingrepresen
tative
sof
thege
nusin
southe
rnSo
uthAmerica.(*)Fo
ssilspecim
ende
scribe
dby
Tonn
iand
Laza
(198
0);P
h-dm
:pha
lanx
prox
imalisdigitimajoris.
Qua
drate
Coraco
idCa
rpom
etacarpu
sPh
-dm
Femur
Tibiotarsu
sTa
rsom
etatarsu
s
L-lo
W-qm
L-ac
L-pc
W-fo
D-prox
D-cm
LTW
-dist
D-dist
W-dist
D-dist
W-prox
D-prox
W-II
W-III
W-IV
Fulicaleuc
optera
7.1–
7.5
3.9–
4.1
30.3–32
.524
.0–27
.03.9–
4.5
4.5–
4.8
2.9–
3.2
13.3–15
.19.5–
10.7
7.8–
8.9
8.0–
8.9
7.7–
8.9
8.2–
9.5
8.1–
10.3
2.7–
3.0
3.3–
3.7
2.9–
3.4
7.2(7
)4.0(7
)31
.6(1
0)25
.1(1
0)4.4(1
0)4.6(7
)3.0(7
)14
.1(7
)10
.2(1
1)8.5(1
1)8.5(1
1)8.5(1
1)8.9(1
1)9.1(1
1)2.8(1
1)3.4(1
1)3.1(1
1)Fu
licarufifron
s6.9–
7.1
3.8–
4.1
27.7–32
.322
.8–27
.33.2–
4.1
3.6–
4.4
2.5–
3.3
12.8–13
.79.9–
11.9
8.3–
10.3
8.1–
9.7
8.2–
9.1
8.4–
9.5
8.5–
10.4
2.6–
2.8
3.3–
3.9
3.0–
3.4
7.0(5
)3.9(5
)29
.6(1
0)24
.7(1
0)3.8(1
0)4.0(9
)2.8(9
)13
.5(9
)10
.7(1
0)9.1(1
0)8.8(9
)8.7(9
)8.8(9
)9.2(9
)2.7(9
)3.6(9
)3.2(9
)Fu
licaarmillata
8.2–
8.3
4.5–
4.9
33.5–41
.126
.7–30
.34.7–
6.7
4.9–
5.7
3.2–
4.0
15.6–16
.711
.3–12
.99.6–
11.7
9.4–
11.5
9.2–
11.5
9.9–
11.8
9.1–
11.7
3.1–
3.9
4.0–
4.6
3.7–
4.0
8.3(3
)4.7(3
)35
.7(1
0)28
.5(1
0)5.5(1
0)5.3(8
)3.5(8
)16
.4(8
)12
.4(9
)10
.7(9
)10
.6(9
)10
.4(9
)10
.9(9
)11
.0(9
)3.5(9
)4.3(9
)3.8(9
)Fu
licagiga
ntea
46.5
38.0
7.0
7.0
4.4
18.9
16.3
16.0
16.2
Fulicaleuc
optera
MLP
71-X
-17-14
*9.0
Fulicaleuc
optera
MLP
76-I-30-23
7.3
4.0
Fulicarufifron
sMLP
76-I-30-25
27.4
22.1
3.9
Fulicarufifron
sMLP
76-I-30-24
3.5
2.5
cf.F.leu
coptera-F.rufifron
sMLP
76-I-30-27
10.8
8.2
cf.F.leu
coptera-F.rufifron
sMLP
76-I-30-45
9.1
8.7
cf.F.leu
coptera-F.rufifron
sMLP
76-I-30-20
9.6
8.4
cf.F.leu
coptera-F.rufifron
sMLP
76-I-30-22
3.8
3.3
cf.F.a
rmillataMLP
76-I-30-56
16.1
cf.F.a
rmillataMLP
76-I-30-26
3.4
4.2
74 M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
Remarks: NISP=1,MNI=1. F. leucoptera is a common inhabitant ofall kinds of aquatic environments along the Pampean Region, includingPOP1 area (Narosky andDi Giacomo, 1993). The presence of this specieswas reported previously from indeterminate levels of the GuerreroMember (sensu lato) of the Luján Formation at POP1, on the basis of aleft tarsometatarsus (MLP 71-X-17-14; Tonni and Laza, 1980).
Fulica rufifrons Philippi and Landbeck, 1861Referred specimens: MLP 76-I-30-25, right coracoid (Fig. 4I,J); MLP
76-I-30-24, incomplete right carpometacarpus.Measurements: see Table 3.Description and comparisons: the specimens are referred to
F. rufifrons on the basis of its small size, and the following combinationof characters:
CoracoidMLP 76-I-30-25: 1) shaft narrower than in F. leucoptera andF. armillata; 2) smaller extremitas omalis than in F. leucoptera andF. armillata; 3) in medial view, processus acrocoracoideus with astraight proximal edge of the facies articularis clavicularis (convex inF. leucoptera and F. armillata); 4) poorly developed tuberculumbrachiale; 5) in lateral view, slightly excavated cotyla scapularis (verydeep in F. leucoptera and F. armillata).
Carpometacarpus MLP 76-I-30-24: 1) in ventral view, poorly devel-oped lamina elliptica articularis (very well-developed in F. leucopteraand F. armillata); 2) in cranial view, very gracile processus pisiformis(stout in F. leucoptera and F. armillata); 3) fossa supratrochleariswith a very well-developed proximal tubercle (poorly developed inF. leucoptera); 4) distally divergent os metacarpale minor (slightlydivergent in F. leucoptera).
Remarks: NISP = 2, MNI = 1. F. rufifrons is a very common inhabi-tant along the aquatic environments of the Buenos Aires Province(Narosky and Di Giacomo, 1993).
Fulica sp. cf. F. leucoptera-F. rufifronsReferred specimens: MLP 76-I-30-27, distal end of left femur; MLP
76-I-30-45, distal end of left tibiotarsus (Fig. 4K,L); MLP 76-I-30-20,proximal end of right tarsometatarsus; MLP 76-I-30-22, distal end ofleft tarsometatarsus.
Measurements: see Table 3.Description and comparisons: the specimens may be referred to
Fulica on the basis of the following features (Wetmore, 1924; Olson,1973; Livezey, 1998; Boles, 2005): 1) femur with a prominenttuberculum m. gastrocnemialis medialis located on the dorsal marginof the crista supracondylaris medialis; 2) presence of a nearly straightto convex slope between epicondylaris medialis and corpus femoris;3) tibiotarsus with robust and laterally oriented condylus medialis;4) very wide sulcus intercondylaris tibialis; 5) tarsometatarsus with alarge and distinct tuberositas m. tibialis cranialis, which shows a flatdorsal surface; 6) robust trochleae metatarsorum; 7) distally extendedtrochlea IV; 8) proximally positioned trochlea II; 9) transverselycompressed shaft proximal to distal trochleae.
Size of new specimens is similar to F. leucoptera and F. rufifrons.Regrettably, both species are very difficult to distinguish on the basisof these isolated elements.
Remarks: NISP = 4, MNI = 1.
Fulica sp. cf. F. armillata Vieillot, 1817Referred specimens:MLP 76-I-30-56, right phalanx proximalis digiti
majoris; MLP 76-I-30-26, distal end of left tarsometatarsus (Fig. 4M,N).Measurements: see Table 3.Description and comparisons: the specimens are highly fragmented;
however, their large size is only approached by F. armillata, being largerthan F. leucoptera and F. rufifrons.
Remarks: NISP = 2, MNI = 1. F. armillata is a common inhabitant ofaquatic environments of the Buenos Aires Province (Narosky and DiGiacomo, 1993).

75M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
Order Cathartiformes Seebohm, 1890Family Cathartidae Lafresnaye, 1839Genus Coragyps Saint-Hilaire, 1853Coragyps atratus (Bechstein, 1793)Referred specimens: MLP 76-I-30-1, proximal end of left tibiotarsus
(Fig. 5A,B); MLP 76-I-30-2, distal end of left tibiotarsus (Fig. 5C,D).Measurements: MLP 76-I-30-2, W-dist: 14.9 (Cathartes aura
[Linnaeus, 1758]: 12.3–15.2 [3]; C. atratus: 12.1–15.9 [83, includes“Coragyps occidentalis”† Miller, 1909]; Sarcoramphus papa [Linnaeus,1758]: 18.0–18.6 [2]; Pleistovultur nevesi† Alvarenga, 2008: 20.5 [1];Geronogyps reliquus† Campbell, 1979: 24.1 [1]; Vultur gryphus Linnaeus,1758: 23.9–27.3 [10]; Gymnogyps howardae† Campbell, 1979: 24.2–25.3[2]; Gymnogyps californianus [Shaw, 1797]: 25.1 [1]; Perugyps diazi†Stucchi and Emslie, 2005: 27.6 [1]); D-dist: 14.3 (C. atratus: 12.7–14.6[20, includes “C. occidentalis”]; C. aura: 13.4–14.2 [2]; S. papa: 17.2–17.3[2]; P. nevesi†: circa 19.2 [1]; G. reliquus†: 22.5 [1]; V. gryphus: 21.8–26.2[8]; G. howardae†: 22.8 [1]; G. californianus: 22.5 [1]; P. diazi†: 26.2 [1]).
Description and comparisons: the size and robustness of MLP 76-I-30-2 are similar to that of small-sized cathartids (C. atratus, Cathartesspp.), being different from larger condor-like vultures (Sarcoramphus,Pleistovultur, Geronogyps, Vultur, Gymnogyps, Perugyps). This specimendiffers from Cathartes spp. (C. aura; Cathartes burrovianus Cassin,1845; and Cathartes melambrotus Wetmore, 1964) and resemblesC. atratus in the following combination of characters: 1) elongate, dorso-ventrally tall, and transversely compressed distal condyles (vs. shorterand transversely wider); 2) in lateral view, rounded condylus lateralis,with a subequal length and depth (vs. much more craniocaudally elon-gated, andwith greater depth than length; Campbell, 1979); 3) roundedlateral edge of condylus medialis (vs. pointed); 4) sulcus extensoriusmedially curved near the proximal end of the pons supratendineus(vs. straight; Campbell, 1979); 5) in distal view, U-shaped and verydeep incisura intercondylaris (vs. V-shaped and shallow incisura;Becker, 1986).
The proximal end of tibiotarsus MLP 76-I-30-1 is poorly preservedand abraded; however, general proportions and size are indistinguish-able from that of living C. atratus.
Remarks: NISP = 2, MNI= 1. Four extinct and five extant genera ofvultures were recorded in late Pleistocene–early Holocene of SouthAmerica (Campbell, 1979; Alvarenga and Olson, 2004; Alvarengaet al., 2008). In the Pampean Region previous records only reportedvery large condors (Tonni and Noriega, 1998; Tambussi and Noriega,1999; Noriega and Tonni, 2007), including the extant V. gryphus andS. papa (Tambussi and Noriega, 1999; Noriega and Areta, 2005). Thepresence of C. atratus in the late Pleistocene of the Pampean Regionadds a new component to this taxonomically diverse scavenger avifau-na. C. atratus is widely distributed in Argentina being common in opencountry areas (Narosky and Di Giacomo, 1993).
Order Accipitriformes Voous, 1973Familia Accipitridae Vieillot, 1816Genus Buteo Lacépède, 1799Buteo sp.Referred specimen: MLP 76-I-30-7, proximal end of left scapula
(Fig. 5E,F).Measurements: MLP 76-I-30-7, D-prox: 14.8 (Rupornis magnirostris
[Gmelin, 1788]: 7.9–10.3 [4]; Parabuteo unicinctus Temminck, 1824:12.6 [1]; Buteo polyosoma [Quoy&Gaimard, 1824]: 15.4 [1];Geranoaetusmelanoleucus [Vieillot, 1819]:16.0–18.5 [6]; Harpyhaliaetus solitarius[Tschudi, 1844]: 19.9 [1]; Amplibuteo hibbardi† Campbell, 1979: 22.4–27.0 [3]).
Description and comparisons: the scapula MLP 76-I-30-7 corre-sponds to a mid-sized Buteoninae hawk of the genus Buteo, beinglarger than the genera Ictinia, Rostrhamus, Rupornis, Leucopternis, andParabuteo, but smaller thanGeranoaetus,Harpyhaliaetus, and the extinctAmplibuteo. Due to its size it is possible that the new specimen may be
related to B. polyosoma. However, its incomplete and uninformativenature precludes a more precise taxonomic identification.
Remarks: NISP = 1, MNI = 1. B. polyosoma is widespread and oftencommon in open habitats in western and southern South America. Inaddition, B. albicaudatus Vieillot, 1816, and B. swainsoni Bonaparte,1838, also are present in the Buenos Aires Province (Narosky and DiGiacomo, 1993).
Order Falconiformes (Sharpe, 1874)Family Falconidae Vigors, 1824Genus CaracaraMerrem, 1826Caracara sp.Referred specimen: MLP 76-I-30-9, distal end of left humerus
(Fig. 5G,H).Measurements: MLP 76-I-30-9, W-dist: 23.2 (Milvago chimango
[Vieillot, 1816]: 11.5–12.5 [7]; Milvago chimachima Vieillot, 1816:11.9–12.0 [2]; Milvago brodkorbi† Campbell, 1979: 13.6–14.6 [6];Phalcoboenus albogularisGould, 1837: 15.3 [1]; Caracara plancus [Miller,1777]: 18.2–22.4 [16]; Caracara cheriway [Jacquin, 1784]: 16.8–18.5[11]).
Description and comparisons: the specimen is referred to Caracarini(being distinguishable fromHerpetotherinae and others Falconinae) onthe basis of the following characters (Noriega et al., 2011; Cenizo et al.,2012): 1) shallow fossa olecrani; 2) poorly developed processusflexorius, not surpassing distal level of condylus ventralis; 3) condylusventralis with elliptical outline; 4) well-defined impressio musculibrachialis distally delimited by a ridge. The material is referred togenus Caracara on the basis of its very large size, being the largest extantcaracarine.
Remarks: NISP= 1, MNI= 1. Recently, a new species was describedfrom late Pleistocene beds of Uruguay, named Caracara major Joneset al., 2013. It represents the largest known falconid, being considerablylarger than C. plancus (Jones et al., 2013). The humerusMLP 76-I-30-9 islarger than extant specimens of the genus, and its referral to C. major isnot improbable. However, until more material become available, thespecimen is here identified only at the generic level. C. plancus is an op-portunistic species widely distributed in Argentina, including the POP1area (Vuilleumier, 1970).
Order Strigiformes Wagler, 1830Family Strigidae Vigors, 1825Genus Asio Brisson, 1760Asio cf. A. flammeus (Pontoppidan, 1763)Referred specimens: MLP 76-I-30-5, articular portion of a right
mandible (Fig. 5I,J); MLP 76-I-30-6, distal end of right carpometacarpus.Measurements:MLP76-I-30-5,W-cla: 5.6 (Tyto alba [Scopoli, 1769]:
6.8–9.7 [11]; Megascops choliba [Vieillot, 1817]: 3.3–3.8 [4]; Athenecunicularia [Molina, 1782]: 3.6–4.1 [8]; Pseudoscops clamator [Vieillot,1807]: 4.1–4.5 [3]; Asio stygius [Wagler, 1832]: 4.8–5.0 [2];A. flammeus: 5.0–5.8 [3]; Bubo virginianus [Gmelin, 1788]: 5.5–6.3 [5];Pulsatrix perspicillata [Latham, 1790]: 7.8 [1]). MLP 76-I-30-6, W-dist:8.1 (T. alba: 6.2–8.5 [18]; Glaucidium nanum [King, 1828]: 5.4–5.5 [2];A. cunicularia: 4.4–5.3 [20]; M. choliba: 5.6–5.7 [3]; A. flammeus: 6.6–8.3 [18]; P. clamator: 8.2–8.6 [3]; B. virginianus: 10.1–12.4 [9]).
Description and comparisons: the mandible MLP 76-I-30-5 is re-ferred to Strigidae and differs from T. alba (Tytonidae), on the basis ofthe following traits (Cenizo and De los Reyes, 2008): 1) in lateralview, dorsoventrally low articular bone, with a strong ventral projec-tion; 2) in dorsal view, very wide and transversely expanded cotylaelateralis et medialis; 3) prominent lateral margin of cotyla medialis;4) medially oriented longitudinal ridge along the ventral margin ofthe mandible (caudally oriented in T. alba). The carpometacarpus MLP76-I-30-6 differs from T. alba and resembles Strigidae in the followingtraits: 1) wide spatium intermetacarpale (reduced in T. alba andA. cunicularia); 2) in distal view, facies articularis digitalis major withan acute cranial margin (slightly developed in A. cunicularia, flattenedin T. alba), 3) in distal view, very well-developed caudal tubercle of

Fig. 5. Fossil landbirds from Paso Otero locality. Coragyps atratus (Cathartidae): proximal end of left tibiotarsus (MLP 76-I-30-1) in cranial (A), and proximal (B) views; distal end of lefttibiotarsus (MLP 76-I-30-2) in cranial (C), and distal (D) views. Buteo sp. (Accipitridae): proximal end of left scapula (MLP 76-I-30-7) in medial (E), and lateral views (F). Caracara sp.(Falconidae): distal end of left humerus (MLP76-I-30-9) in cranial (G), and caudal (H) views.Asio cf.A.flammeus (Strigidae): articular portion of a rightmandible (MLP 76-I-30-5) in dorsal(I), and lateral (J) views. Passeriformes indet.: proximal end of left carpometacarpus (MLP 76-I-30-38) in dorsal (K), and ventral (L) views. Furnariidae indet.: distal end of right humerus(MLP 76-I-30-12) in caudal (M), and cranial (N) views. Thraupidae/Emberizidae? indet.: proximal end of left carpometacarpus (MLP 76-I-30-36) in dorsal (O), and ventral (P) views. Scalebars = 10 mm.
76 M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
the facies articularis digitalis major; 4) in distal view, wide spacebetween facies articularis digitalis major et minor (narrow in T. albaand A. cunicularia); 5) in dorsal view, reduced disto-caudal notch ofthe sulcus tendineus (very wide in T. alba).
Both MLP 76-I-30-5 and MLP 76-I-30-6 are larger than Glaucidium,Megascops, and Athene; and smaller than Bubo, Strix, and Pulsatrix. Thesize of the new specimens agrees with mid-sized Asioninae owls(P. clamator and Asio spp.) and shares with A. flammeus the followingcharacters: 1) vertically oriented and not caudally extended caudalmargin of the mandible; 2) short synostosis metacarpale distalis.Their fragmentary nature precludes a more accurate taxonomicdetermination.
Remarks: NISP = 2, MNI = 1. A. flammeus is a very common inhab-itant of open grasslands along the Buenos Aires Province, includingPOP1 area (Narosky and Di Giacomo, 1993).
Order Passeriformes (Linnaeus, 1758)Passeriformes gen. et sp. indet.Referred specimens: MLP 76-I-30-38, proximal end of left
carpometacarpus (Fig. 5K,L); MLP 76-I-30-3, distal end of right ulna.Measurements: MLP 76-I-30-38, D-prox: 4.5.Description and comparisons: the specimens are referred to
Passeriformes due to small size, delicateness, and the carpometa-carpus MLP 76-I-30-38 by the presence of a small cranially protrud-ing process at the proximal end of the sulcus tendinosus on the osmetacarpale majus (Mourer-Chauviré et al., 1989; Mayr, 1998;Mayr and Manegold, 2006a, 2006b; Manegold, 2008). MLP 76-I-30-
38 is reminiscent to Furnariidae in lacking the processus dentiformison the cranial margin of the os metacarpale majus. However, thefragmentary nature of specimens does not allow referring it beyondthe ordinal level.
Remarks: NISP = 2, MNI = 1.
Suborder Tyranni, Wetmore and Miller, 1926Family Furnariidae (Gray, 1840)Furnariidae gen. et sp. indet.Referred specimens: MLP 76-I-30-12, distal end of right humerus
(Fig. 5M,N); MLP 76-I-30-11, right ulna; MLP 76-I-30-37, distal end ofright carpometacarpus.
Measurements: MLP 76-I-30-12, W-dist: 8.3 (Pseudoseisuropsisnehuen† Noriega, 1991: 7.5–8.6 [2]; Pseudoseisura cursor† Tonni andNoriega, 2001: 8.7 [1]; Pseudoseisura gutturalis [d'Orbigny and Lafresnaye,1838]: 6.1 [1]; Pseudoseisura lophotes [Reichenbach, 1853]: 6.9 [1];Drymornis bridgesii [Eyton, 1849]: 7.9–8.0 [2]). MLP 76-I-30-11, LT: 36.5(P. nehuen†: 42.0–44.4 [2]; P. cursor†: 42.0 [1]; P. gutturalis: 30.5 [1];P. lophotes: 35.0 [1]; D. bridgesii: 38.2–40.2 [2]); W-prox: 6.2(P. nehuen†: 6.5–7.0 [2]; P. cursor†: 6.9 [1]; P. gutturalis: 4.8 [1];P. lophotes: 5.63 [1]).MLP 76-I-30-37, D-dist: 4.4 (D. bridgesii: 4.9–5.0 [2]).
Description and comparisons: the specimens are referred to Tyranniand differ from Passeri on the basis of the following characters(Ballmann, 1969; Mourer-Chauviré et al., 1989; Mayr and Manegold,2006a, 2006b; Manegold, 2008; Cenizo et al., 2012): 1) humerus withless developed processus supracondylaris dorsalis; 2) humerus withoutdepression within the sulcus humerotricipitalis; 3) ulna with well-

77M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
developed tuberculum ligamenti collateralis ventralis; 4) ulna withshallow pit for the insertion of the tendon of the m. biceps brachii;5) ulna with rounded and proximodistally short cotyla dorsalis;6) ulna with rounded proximal rim of the cotyla ventralis (proximalnotch absent); 7) carpometacarpus with a distally protruding andfinger-like process on the craniodistal edge of the metacarpal III;8) protrudent caudodistal portion of metacarpal II that overhangs thesulcus interosseus on the carpometacarpus.
The robustness and size of available material is similar to the large-sized extant furnariids of the genus Pseudoseisura and largestDendrocolaptinae woodcreepers. Proportions are similar to that of ex-tinct furnariid P. nehuen and its possible relative P. cursor (Noriega,1991; Tonni and Noriega, 2001). However, some features suggest thatnew specimens may belong to another furnariid group. In this sense,in the carpometacarpus MLP 76-I-30-37 the finger-like process on thecraniodistal edge of the metacarpal III is less developed than in anyother available Tyranni. On the other hand, the presence of a deepdepression on the ventral surface of the synostosis metacarpalisdistalis (Manegold, 2008) shared by MLP 76-I-30-37, Passeri(Mourer-Chauviré et al., 1989; Manegold, 2008), and some Tyranni,such as the dendrocolaptines (e.g., D. bridgesii), suggests that itmay belong to the later group.
Remarks: NISP = 3, MNI = 1. The record of large-sized furnariids inthe Pleistocene of South America is represented by P. nehuen andP. cursor, coming from the lower–middle Pleistocene of Buenos AiresProvince (Noriega, 1991; Tonni and Noriega, 2001), and Pseudoseisuropsiscuelloi Claramunt and Rinderknecht, 2005 from the late Pleistocene ofUruguay. It is worthy to mention that the phylogenetic affinities ofthese species are under debate. P. nehuen was formerly relatedby Noriega (1991) to the genus Pseudoseisura. Posteriorly, Claramuntand Rinderknecht (2005) related Pseudoseisuropsis spp. with theDendrocolaptinae or a basal linaje of Furnariidae. Recently, Stefanini andGómez (2012) proposed that this extinct genus was probably close tothe Philydorini, particularly Automolus. Beyond their phylogeneticposition, extinct large-sized furnariids are very common in the Pleisto-cene outcrops of the Pampean Region (obs. pers.). Presence of largePseudoseisuropsis-like specimens in POP1 and Uruguay, suggests thatthe diversification of these peculiar furnariids may have lasted until latePleistocene.
Currently, two Pseudoseisura species (P. lophotes and P. gutturalis) aredistributed along flat or hilly open semiarid steppes and shrubs inArgentina (Vaurie, 1980). Both species are present, but are uncommon in-habitants in southern Buenos Aires Province (Narosky and Di Giacomo,1993).
Suborder Passeri Linnaeus, 1758Thraupidae/Emberizidae? gen. et sp. indet.Referred specimen: MLP 76-I-30-36, proximal end of left
carpometacarpus (Fig. 5O,P).Measurements: MLP 76-I-30-36, D-prox: 3.4.Description and comparisons: MLP 76-I-30-36 exhibits typical fea-
tures to that of Passeriformes, including (Mourer-Chauviré et al., 1989;Mayr, 1998; Mayr and Manegold, 2006a, 2006b; Manegold, 2008):1) small and cranially protruding process at the proximal end of thesulcus tendinosus on the os metacarpale majus; 2) well-developedprocessus dentiformis on the cranial margin; 3) large processusintermetacarpalis fused to the os metacarpale majus et minus.
MLP 76-I-30-36 may be referred to Passeri by the presence of awell-developed fossa on the ventrocaudal margin of the trochleacarpalis ventralis (see Manegold, 2008), whereas in Tyranni thisfossa is much shallower, and it is distally located. In MLP 76-I-30-36 the proximal margin of this fossa is located at the level of the dis-tal margin of the processus pisiformis, a feature occurring inThraupidae and Emberizidae (in other available Passeri, the proxi-mal margin of this fossa do not approaches the level of the distalmargin of processus pisiformis).
Remarks: NISP=1,MNI=1. The size ofMLP 76-I-30-36 is similar tothe corresponding element of Paroaria coronata (Miller, 1776).
4.2. Taphonomy
Bird remains in POP1 represent 4.18% (NISP birds = 60) of the totalof anatomically and taxonomically determined materials (NISP total =1432; Pomi and Tonni, 2011; Table 4) coming from this fossiliferouslocality. However, compared with the scarcer avian remains comingfrom Pampean Pleistocene localities, the finding of this relativelylarge number of specimens is remarkable. The autopodial elementsare the most commonly preserved bones (carpometacarpi = 15%,tarsometatarsi = 20%), suggesting some kind of differential preserva-tion of robust elements. The high percentage of breakage (84.4%) isconsistent with the fragility of bird remains. Oblique fractures are themost abundant, followed by the irregular. Complete elements aremainly represented by coracoids, ulnae and tarsometatarsi. Consideringonly the long bones, the 37% of the sample presented evidence ofweathering, corresponding to stage 1 of Behrensmeyer et al. (2003).These cracks were developed before the final burial, and show strongdifferences with those produced during the diagenesis. The later arestraight and of glassy aspect, with sharp and straight fissure surfaces.It can be pointed out that the weathering process affected similar waydifferent anatomical units.
Pomi and Tonni (2011) described several scratches on some mam-mals and birds remains from POP1. Based on recent field samples andlaboratory experiments they conclude that the POP1 scratches probablyreflect the behavior of termites (Isoptera). The location of some of thesemarks above weathering cracks indicate that it were produced afterskeletonization of the carcass, with absence of soft tissue and perhapswith already disarticulated bones, previous to the final burial.
The MNI (=30) is relatively low with respect to the number of re-covered taxa (at least 22 taxa representing 13 families). The averagenumber of remains collected for each taxon is very low (=2.4) with re-spect of the total taxonomic diversity. This suggests that more detailedexcavations at POP1 may increase the taxonomic avian diversity ofthis fossiliferous locality.
5. Discussion and conclusions
As pointed out above, avifaunas from the Pleistocene of SouthAmerica are rather scarce, and no single representative assemblagewas known in Argentina until now. Previously reported assemblagesin the continent were located within intertropical northern areas andbelong to relatively young deposits (latest Pleistocene–early Holocene).In this regard, the Talara avifauna (Peru; Campbell, 1976, 1979) hasbeen dated to 13,9 kyr BP (Churcher, 1966), and a similar age was esti-mated for La Carolina avifauna (Ecuador, see Campbell, 1976, 1979,1982). The chronology of the birds listed byWinge (1888) from severalcaves in Minas Gerais State (southeastern Brazil), is difficult to assess,and available data suggest ages between 12 and 8 kyr BP (Dias, 2004;Bueno et al., 2013). In the case of the birds recovered from the cave ofToca da Janela da Barra do Antonião in Piauí State (northeasternBrazil; Guérin et al., 1993, 1996), the dating yielded an age of 9670 ±140 yr BP (Martin, 1996). With an age of circa 38 kyr BP, the POP1assemblage represents the oldest Pleistocene avifauna known fromthe South American continent, and the first one coming from a clearinterstadial event (see below).
POP1 bird taphocenosis is composed of 60 remains representing atleast 22 taxa (Table 4). Based on its biological requirements, 64% of thetaxa are nearly exclusive inhabitants of aquatic environments (14 taxa,53% of collected specimens, MNI = 19), 9% are exclusive terrestrialbirds (2 taxa, 15% of collected specimens, MNI = 4), and 18% are birdsof prey (4 taxa, 10% of collected specimens,MNI=4). The predominanceof aquatic birds is in agreement with sedimentological characteristics.The presence of freshwater snails and the marsh-rat Lundomys molitor

Table 4Synthesis of bird taxa recovered fromPasoOtero locality and their preferred habitats. Hab-itat types: A, aquatic environments; I, indeterminate; M, multiple environments; O, openenvironments (savannas, grasslands and shrub steppes).
Taxa AbundanceNISP (MNI)
NISP% Habitattype
AvesAves indet. 7 (–) 11.7 I
TinamidaeNothura sp. 5 (2) 8.3 OEudromia sp. 4 (2) 6.7 O
AnatidaeAnatidae indet. 2 (2) 3.3 ADendrocygna sp. 2 (1) 3.3 AChloephaga sp. 1 (1) 1.7 AAnatini indet. morphotype 1 3 (2) 5.0 AAnatini indet. morphotype 2 2 (1) 3.3 Acf. Callonetta sp. 3 (2) 5.0 AAnas cf. A. sibilatrix 1 (1) 1.7 A
PodicipedidaeRollandia rolland 2 (1) 3.3 A
PhoenicopteridaePhoenicopteridae indet. 1 (1) 1.7 A
ThreskiornithidaePlegadis cf. P. chihi 2 (1) 3.3 A
Scolopacidaecf. Numenius sp. 2 (1) 3.3 ACharadriiformes indet. morphotype 1 2 (1) 3.3 A
RallidaeFulica leucoptera 1 (1) 1.7 AFulica rufifrons 2 (1) 3.3 AFulica cf. F. leucoptera-F.rufifrons 4 (1) 6.7 AFulica cf. F. armillata 2 (1) 3.3 A
CathartidaeCoragyps atratus 2 (1) 3.3 M
AccipitridaeButeo sp. 1 (1) 1.7 M
FalconidaeCaracara sp. 1 (1) 1.7 M
StrigidaeAsio cf. A. flammeus 2 (1) 3.3 O
FurnariidaeFurnariidae indet. 3 (1) 5.0 I
Thraupidae/EmberizidaeThraupidae/Emberizidae? indet. 1 (1) 1.7 IPasseriformes indet. 2 (1) 3.3 I
Total 60 (30) 100 –
78 M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
(Winge, 1887) (see Prado et al., 1987; Teta and Pardiñas, 2006; Pardiñasand Teta, 2011) reinforces this interpretation.
The ecological requirements of the reported avian taxa indicate theexistence of open grasslands with the presence of freshwater andpermanent ponds, similar to that found today in the Bonaerean Inter-montane Area. The high similarities in ecological requirements andspecies diversity shared between the POP1 assemblage and livingavifaunas in the area, suggest very similar climatic conditions.
The radiocarbon date for POP1 indicates that the fossiliferous levelcorresponding to the La Chumbiada Member was deposited duringthe Marine Isotope Stage 3 (MIS 3, about 60–35 kyr BP; Clapperton,1993; see also Tonni et al., 1999). Abruptwarmglobal episodeswere re-corded during the last part of MIS 3 (e.g., Huber et al., 2006; Hodgsonet al., 2009). In concordance with this, the presence in POP1 of the ro-dent L. molitor (Prado et al., 1987) and termite marks on bones (Pomiand Tonni, 2011), has been employed to sustain that the La ChumbiadaMember was deposited within warm climatic conditions (Teta andPardiñas, 2006; Pardiñas and Teta, 2011; Pomi and Tonni, 2011). How-ever, presence in the same level of some desert-inhabiting mammals(i.e., Lama gracilis [Gervais and Ameghino, 1880] and Lestodelphys sp.;Prado et al., 1987; Pomi and Tonni, 2011) suggests a possible non-analogous assemblage, or alternatively, an average time representedin the deposits during which rapid climatic changes occurred (Pomiand Tonni, 2011). In contrast to the La Chumbiada Member, the
overlaying Guerrero Member is known to have been deposited in a rel-atively cold and arid period, probably the Last Glacial Maximum (LGM,circa 20–18 kyr BP; Tonni et al., 1999), but also other cold-climateevents (21–10 kyr BP, most of the MIS 2 and the beginning of the MIS1, Tonni et al., 2003). The change between the La Chumbiada and Guer-rero members is also represented by a faunal turnover, including a de-crease in frequency and diversity of browsers, and an increase of hugegrazers that weighed more than a ton (Prado et al., 1987; Tonni et al.,1999, 2003). This turnoverwas interpreted as evidence for a vegetation-al shift in response to a more arid and colder climate (Tonni et al., 1999,2003). The previous record of P. cf. P. pennata from the Guerrero Mem-ber at POP1 (MLP 80-VIII-13-1; see Tonni and Laza, 1980) is congruentwith such inferences.
The fragmentary and dissociated nature of most remains found atPOP1 does not allow making a taxonomic identification beyond thegeneric level, and we cannot discard that some specimens may belongto extinct taxa with biological requirements different from livingspecies. Only the finding of new and more complete material mayallow a more accurate determination. However, based on availableevidence, the POP1 assemblage includes taxa that are currently widelydistributed in the southern cone of South America. Moreover, we didnot find any distributional change, non-analogous association, or clearevidence of extinct taxa. In contrast, the late Pleistocene assemblagesof La Carolina and Talara, from the northwest Andes, include a largenumber of birds represent extinct taxa. In the first locality 53 non-passerine species were reported, of which 9 are currently extinct(17%; Campbell, 1976, 1982), and in the second 89 non-passerine taxawere listed, including 21 extinct species (23%; Campbell, 1976, 1982).These localities show homogeneous species richness and composition(39 shared species, seven of which are extinct), indicating a great dealof interchange between them (Campbell, 1976, 1979, 1982).
Like the POP1 assemblage, the Brazilian late Pleistocene–early Holo-cene avifaunas show very few or no extinct species. In theMinas Geraisassemblages, 126 species were recognized, but only a single extincttaxon was described by Winge (1888). New findings from those caves,together with partial overviews ofWinge's original list, allowed the rec-ognition of three additional large and extinct scavenger taxa (Alvarengaand Olson, 2004; Alvarenga et al., 2008; Agnolin, 2009, see below).However, as pointed out by Olson (1985), although Winge was uncer-tain in the identification of at least 50% of the taxa, later authors accept-ed his identifications without a detailed overview. In the Piauí avianassemblage 34 non-passerine species were mentioned, all representingextant taxa (Guérin et al., 1996).
Although the South American late Pleistocene avian record is stillvery patchy and biased, the differences in taxonomic compositionbetween the northwestern Andean region and eastern lowlands re-flect different evolutionary processes in dissimilar geographic areaswith contrasting climatic and geological histories. The avian assem-blages from Peru and Ecuador correspond to a post-LGM event(circa 14 kyr BP), during which a well-defined stadial period oc-curred in the Andes (Clapperton, 1993). Currently, these fossiliferouslocalities are located within subtropical coastal desert to tropicalscrub desert, but during the glacial expansive events, there existedabundant freshwater and marine environments surrounded bysavanna-like vegetation and dry forest inland habitats (Campbell,1982). In this regard it is possible that the cyclic climatic changesalong the Plio-Pleistocene were the main triggering factors of thehigh speciation rates observed in several “hot spot” endemic areasfrom the northwestern Andes (e.g., Fjeldså, 1992; Heindl andSchuchmann, 1998; Myers et al., 2000; Fjeldså and Rahbek, 2006;Ribas et al., 2007). These climatic pulses may have favored orograph-ic isolation, and represent an interesting model which would explainthe diverse extinct taxa found in the late Pleistocene of the northernAndes (Campbell, 1982). The low sea level and taxonomic similari-ties between the Carolina and Talara avifaunas allowed Campbell(1982) to propose that a dry savanna corridor existed along the

79M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
Pacific coast of northern South America, which also probably favoredsouthward range expansions of North American birds through Cen-tral America to South America. It is possible that this interchangemay also constitute an important factor that influenced bird compo-sition at Peru and Ecuador assemblages. In addition, although thestatus of some genera should be reviewed (Olson, 1985), six generic“paleoendemic” taxa were recognized in both assemblages(Campbell, 1979). The confirmation of its validity could indicatethat the divergence of such taxa may not be the result of recent phe-nomena (i.e., late Pleistocene), but may correspond to older events,possibly related to Andean uplift and gradual formation of new hab-itats in the northern Andes since the Miocene (Hooghiemstra andCleef, 1995; Wijninga, 1995; Hooghiemstra and Van der Hammen,1998).
The avifaunas from caves of Minas Gerais are also post-Late Gla-cial Maximum, and based on the available dating (circa 12–10 kyrBP), may correspond to the stadial event of Younger Dryas (seeLedru, 1993). In fact, faunistic (Voss and Carleton, 1993) and palyno-logical (Ledru, 1993; Behling et al., 2002) evidence suggest that pastclimates in Minas Gerais were more temperate than today, allowingthe development of extensive grasslands. Concordantly, the type lo-cality of themarsh-rat L. molitor is Lagoa Santa (Minas Gerais;Winge,1887), located more than 1450 km north of its current distributionalrange (Pardiñas and Teta, 2011). In contrast, Piauí caves are current-ly within the xerophytic formations of the Caatinga (the dryest areaof Brazil), but birds and mammals from the late Pleistocene–earlyHolocene transition indicate a savannah landscape, with scatteredforested areas, much more humid than at present (Guérin et al.,1996).
In this way, the evidence suggests similar environmental conditionsduring the post-Late Glacial Maximum stadials in southeastern Braziland the MIS3 interstadial in the Pampean Region of Argentina. The ab-sence of significant extinction events (limitedmainly to large scavengerbirds; see below) in the late Pleistocene–early Holocene avifaunas ofBrazil and Argentina reinforces such hypothesis. In contrast, the recordsof Talara and La Carolina demonstrate the extinction of a large numberof aquatic and predatory birds on the coasts of Ecuador and Peru coastsin the last 14 kyr BP. Based on available data, the cyclic retraction andexpansion of open environments during the late Pleistocene climatic os-cillations may have a more dramatic impact on the avian communitiesalong the Pacific coast than on those of southeastern lowlands ofSouth America.
In contrast with most taxa, the late Pleistocene avian extinctions inSouth America drastically affected large scavenger birds (i.e., largecaracaras, vultures and ciconids). In Peru, Ecuador, Brasil, Uruguay,and Argentina the records for this ecological guild indicate total andpartial extinctions (Campbell, 1976, 1979; Alvarenga, 1998; Tonni andNoriega, 1998; Tambussi and Noriega, 1999; Alvarenga and Olson,2004; Noriega and Areta, 2005; Agnolin, 2006; Noriega and Tonni,2007; Alvarenga et al., 2008; Jones et al., 2013), a pattern shared byboth Andean and lowland regions. As a rule, living taxa of this ecologicalguild are not limited to specific environments, and show very widegeographical ranges. Thus, as proposed by Campbell (1982; see alsoEmslie, 1987; Tonni and Noriega, 1998; Noriega and Tonni, 2007), it ispossible that scavenger extinction may be related to the disappearanceof megafauna (which constituted their main food source) rather thanlate Pleistocene climatic oscillations.
Acknowledgments
We acknowledge J. Navas (†), P. Tubaro, and C.P. Tambussi foraccess to the osteological collections under their care. W. Jones, A.Rinderknecht, C.P. Tambussi and H.M.F. Alvarenga made useful com-ments on the identification of some fossil specimens. K. Cockle fortheir assistancewith English version and E.P. Tonni for providing impor-tant stratigraphic data. Finally, we thank Editor Paul Hesse, Rodrigo
Tomassini and an anonymous reviewer for their helpful comments onthe manuscript.
Appendix 1
Comparisons were undertaken with specimens of the followingextant species, which are deposited in MLP and MACN, and authors'collection (MMC): Crypturellus parvirostris, C. tataupa, C. undulatus,Eudromia elegans, E. formosa, Nothoprocta cinerascens, N. ornata,N. pentlandii, Nothura boraquira, N. darwinii, N. maculosa, Rhynchotusrufescens, Tinamotis ingoufi, T. pentlandii, Tinamus solitarius,Dendrocygnabicolor, D. viduata, Coscoroba coscoroba, Cygnus melancoryphus,Chloephaga picta, Tachyeres patachonicus, Lophonetta specularioides,Netta peposaca, Anas sibilatrix, A. georgica, A. cyanoptera, A. flavirostris,A. platalea, A. versicolor, A. bahamensis, Callonetta leucophrys, Amazonettabrasiliensis,Heteronetta atricapilla,Oxyura ferruginea,O. vittata,Nomonyxdominicus, Rollandia rolland, Podiceps major, P. occipitalis, P. gallardoi,Podilymbus podiceps, Tachybaptus dominicus, Syrigma sibilatrix,Egretta alba, E. cocoi, E. thula, Bubulcus ibis, Nycticorax nycticorax, Ciconiamaguari, Theristicus caudatus, T. melanopis, Mesembrinibis cayennensis,Plegadis chihi, P. ridgwayi, Phimosus infuscatus, Platalea ajaja, Jacanajacana,Himantopusmelanurus, Belonopterus chilensis, Charadrius collaris,Tringa flavipes, Calidris fuscicollis, Limosa haemastica, Gallinago gallinago,Thinocorus rumicivorus, Phoenicopterus chilensis, Phoenicoparrus andinus,Aramides ypecaha, A. cajanea, Fulica armillata, F. leucoptera, F. rufifrons,F. gigantea, Rallus sanguinolentus, Pardirallus maculatus, Gallinulachloropus, Porphyrula martinica, Vultur gryphus, Sarcoramphus papa,Cathartes aura, C. burrovianus, Coragyps atratus, Rupornis magnirostris,Buteo polyosoma, Parabuteo unicinctus, Rostrhamus sociabilis,Geranoaetus melanoleucus, Buteogallus urubitinga, Harpyhaliaetuscoronatus, Ictinia plumbea, Circus cinereus, Elanus leucurus, Accipiterstriatus, Milvago chimango, M. chimachima, Caracara plancus,Phalcoboenus australis, Falco femoralis, F. peregrinus, F. sparverius,F. columbarius, Herpetotheres cachinnans, Spiziapteryx circumcincta, Tytoalba, Glaucidium nanum, Megascops choliba, Athene cunicularia,Pseudoscops clamator, Asio stygius, A. flammeus, Bubo virginianus,Pulsatrix perspicillata, Colaptes campestris, Drymornis bridgesii,Lepidocolaptes angustirostris, Geositta cunicularia, Upucerthia dumetaria,Cinclodes fuscus, Furnarius rufus, Phleocryptes melanops, Asthenes baeri,Pseudoseisura lophotes, P. gutturalis, Thammophilus caerulescens,Phytotoma rutila, Xolmis irupero, Hymenops perspicilliatus, Pitangussulphuratus, Tyrannus savanna, Pyrocephalus rubinus, Progne modesta,Troglodytes aedon, Anthus correndera, Mimus saturninus, Turdusrufiventris, Saltator aurantiirostris, Paroaria coronata,Gubernatrix cristata,Sporophila collaris, Catamenia analis, Sicalis luteola, Zonotrichia capensis,Embernagra platensis, Carduelis magellanica, Molothrus badius, Sturnellaloyca.
References
Agnolin, F.L., 2006. Dos nuevos Anatidae (Aves, Anseriformes) del Pleistoceno Inferior-Medio) de Argentina. Stud. Geol. Salmanticensia 42, 81–95.
Agnolin, F.L., 2009. El registro fósil de Ciconia lydekkeri Ameghino, 1891 en el Pleistocenode Sudamérica. Stud. Geol. Salmanticensia 45, 53–58.
Alvarenga, H., 1998. Sobre a ocorrência do condor (Vultur gryphus) no Holoceno da Regiãode Lagoa Santa, Minas Gerais, Brasil. Ararajuba 6, 60–63.
Alvarenga, H.M.F., Olson, S.L., 2004. A new genus of tiny condor from the Pleistocene ofBrazil (Aves: Vulturidae). Proc. Biol. Soc. Wash. 117, 1–9.
Alvarenga, H., Brito, G., Migotto, R., Hubbe, A., Hofling, E., 2008. Pleistovultur nevesi gen. etsp. nov. (Aves: Vulturidae) and the diversity of condors and vultures in the SouthAmerican Pleistocene. Ameghiniana 45, 613–618.
Ballmann, P., 1969. Die vögel aus der altburdigalen spaltenfüllung vonWintershof (West)bei Eichstätt in Bayern. Zitteliana 1, 5–60.
Ballmann, P., 2004. Fossil Calidridinae (Aves: Charadriiformes) from the Middle Mioceneof the Nördlinger Ries. Bonner Zool. Beitr. 52, 101–114.
Baumel, J., Witmer, L.M., 1993. Osteologia. In: Baumel, J., King, A., Breazile, J., Evans, H.,Vanden Berge, J. (Eds.), Handbook of Avian Anatomy: Nomina Anatomica Avium.Publications of the Nuttall Ornithological Club, Massachusetts, pp. 45–132.
Becker, J.J., 1986. A new vulture (Vulturidae: Pliogyps) from the late Miocene of Florida.Proc. Biol. Soc. Wash. 99, 502–508.

80 M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
Behling, H., Arz, H.W., Pätzold, J., Wefer, G., 2002. Late Quaternary vegetational andclimate dynamics in southeastern Brazil, inferences from marine core GeoB 3229-2and GeoB 3202-1. Palaeogeogr. Palaeoclimatol. Palaeoecol. 179, 227–243.
Behrensmeyer, A.K., Stayton, C.T., Chapman, R.E., 2003. Taphonomy and ecology ofmodern avifaunal remains from Amboseli Park, Kenya. Paleobiology 29, 52–70.
Bertelli, S., 2002. Filogenia del orden Tinamiformes. Unpublished PhD Thesis (unpub-lished), Facultad de Ciencias Naturales e Instituto Miguel Lillo, Universidad Nacionalde Tucumán, 357 pp.
Bertelli, S., Chiappe, L.M., 2005. Earliest tinamous (Aves: Palaeognathae) from theMiocene of Argentina and their phylogenetic position. Nat. Hist. Mus. Los Angel.County Contrib. Sci. 502, 1–20.
Bocheński, Z.M., 1994. The comparative osteology of grebes (Aves: Podicipediformes) andits systematic implications. Acta Zool. Cracoviesia 37, 191–346.
Boles, W.E., 2005. A new flightless gallinule (Aves: Rallidae: Gallinula) from the Oligo–Miocene of Riversleigh, Northwestern Queensland, Australia. Rec. Aust. Mus. 57,179–190.
Bueno, L., Schmidt Dias, A., Steele, J., 2013. The Late Pleistocene/Early Holocene archaeo-logical record in Brazil: a geo-referenced database. Quat. Int. 301, 74–93.
Campbell, K.E., 1976. The Late Pleistocene Avifauna of La Carolina, southwestern Ecuador.Smithson. Contrib. Paleobiol. 27, 155–168.
Campbell, K.E., 1979. The non-passerine Pleistocene avifauna of the Talara Tar Seeps,northwestern Peru. R. Ont. Mus. Life Sci. Contrib. 118, 1–203.
Campbell, K.E., 1982. Late Pleistocene events along the coastal plain of northern SouthAmerica. In: Prance, G.T. (Ed.), Biological Diversification in the Tropics. ColumbiaUniversity Press, New York, pp. 423–440.
Campo De Ferreras, A.M., Píccolo, M.C., 1997. El balance hídrico en la cuenca del RíoQuequén Grande, Argentina. Rev. Geofís. 46, 51–66.
Carr, G.A., 1981. An early Pleistocene avifauna from Inglis, Florida. Unpublished PhDThesis, University of Florida, Gainesville, Florida, 162 pp.
Cenizo, M.M., Agnolin, F., 2010. The southernmost records of Anhingidae and a new basalspecies of Anatidae (Aves) from the lower–middle Miocene of Patagonia, Argentina.Alcheringa 34, 493–514.
Cenizo, M.M., de Los Reyes, L.M., 2008. Primeros registros de Tyto alba (Scopoli, 1769)(Strigiformes, Aves) en el Pleistoceno Medio-Tardío de la provincia de BuenosAires (Argentina) y sus implicancias tafonómicas. Rev. Mus. Argent. Cienc. Nat. 10,199–209.
Cenizo, M.M., Tambussi, C.P., Montalvo, C.I., 2012. LateMiocene continental birds from theCerro Azul Formation in the Pampean region (central-southern Argentina).Alcheringa 36, 47–68.
Churcher, C.S., 1966. The insect fauna from the Talara tar seeps, Peru. Can. J. Zool. 44,985–993.
Clapperton, C., 1993. Quaternary Geology and Geomorphology of South America. Elsevier,Amsterdam (779 pp.).
Claramunt, S., Rinderknecht, A., 2005. A new fossil Furnariid from the Pleistocene ofUruguay, with remarks on nasal type, cranial kinetics, and relationships of the extinctgenus Pseudoseisuropsis. Condor 107, 114–127.
Deschamps, C.M., 2005. Late Cenozoic mammal biochronostratigraphy in southwesternBuenos Aires province, Argentina. Ameghiniana 42, 733–750.
Dias, A.S., 2004. Diversificar para poblar: el contexto arqueológico bralsieño en latransición Pleistoceno–Holoceno. Complutum 15, 249–263.
The Howard and Moore Complete Checklist of the Birds of the World. In: Dickinson, E.C.,Remsen, J.V. Jr (Eds.), Non-passerines, 4th ed. vol.1. Aves Press, Eastbourne, U.K(461 pp.).
Dillon, A., Rabassa, J., 1985. Miembro La Chumbiada, Formación Luján (Pleistoceno,provincia de Buenos Aires): Una nueva unidad estratigráfica del valle del río Salado.Prim. Jornadas Geol. Bonaer. Abstr. 27.
Elzanowski, A., Paul, G.S., Stidham, T.A., 2000. An avian quadrate from the Late CretaceousLance Formation of Wyoming. J. Vertebr. Paleontol. 20, 712–719.
Emslie, S., 1987. Age and diet of fossil California condors in Grand Canyon, Arizona.Science 237, 768–770.
Emslie, S.D., 1998. Avian community, climate, and sea-level changes in the Plio-Pleistocene of the Florida Peninsula. Am. Ornithol. Union Ornithol. Monogr. 50,1–113.
Emslie, S.D., 2004. The early and middle Pleistocene avifauna from Porcupine Cave,Colorado. In: Barnosky, A.D. (Ed.), Early and Middle Pleistocene Biodiversity andEnvironmental Change: the Porcupine Cave Fauna from Colorado. University ofCalifornia Press, Berkeley, pp. 127–140.
Fidalgo, F., De Francesco, F., Colado, U., 1973. Geología superficial en las hojasCastelli, M. J. Cobo y Monasterio, provincia de Buenos Aires. V Congr. Geol. Ar-gent. 4, 27–39.
Fjeldså, J., 1992. Biogeographic patterns and evolution of the avifauna of relict high-altitude woodlands of the Andes. Steenstrupia 18, 9–62.
Fjeldså, J., Rahbek, C., 2006. Diversification of tanagers, a species rich bird group,from lowlands to montane regions of South America. Integr. Comp. Biol. 46,72–81.
Gollop, J.B., Barry, T.W., Iversen, E.H., 1986. Eskimo curlew a vanishing species? Saskatch-ewan Natural History SocietySpecial Publication 17 (160 pp.)
Guérin, C., Hugueney, M., Mourer-Chauviré, C., Faure, M., 1993. PaléoenvironnementPléistocene dans l'aire archéologique de Sao Raimundo Nonato (Piaui, Brésil): apportdes mammiféres et des oiseaux. Doc. Lab. Géol. 125, 187–202.
Guérin, C., Curvelo, M.A., Faure, M., Hugueney, M., Mourer-Chauvire, C., 1996. A faunapleistocênicadoPiaui (Nordeste do Brasil). Relaçõespaleoecologicas e biocronologicas.Fumdhamentos 1, 55–103.
Heindl, M., Schuchmann, K.L., 1998. Biogeography, geographical variation and taxon-omy of the Andean hummingbird genus Metallura Gould, 1871. J. Ornithol. 139,425–473.
Hodgson, D., Verleyen, E., Vyverman, W., Sabbe, K., Leng, M.J., Pickering, M.D., Keely, B.J.,2009. A geological constraint on relative sea level in marine isotope stage 3 in theLarsemann Hills, Lambert Glacier Region, East Antarctica (31.366–33.228 cal yr BP).Quat. Sci. Rev. 28, 2689–2696.
Hooghiemstra, H., Cleef, A.M., 1995. Pleistocene climatic change and environmental andgeneric dynamics in the north Andean montane forest and páramo. In: Churchill,S.P., Balslev, H., Forero, E., Luteyn, J.L. (Eds.), Biodiversity and Conservation ofNeotropical montane forests. The New York Botanical Garden, New York, pp. 35–49.
Hooghiemstra, H., Van Der Hammen, T., 1998. Neogene and Quaternary development ofthe Neotropical rain forest: the forest refugia hypothesis, and a literature overview.Earth Sci. Rev. 44, 147–183.
Howard, H., 1929. The avifauna of Emerville shellmound. Univ. Calif. Publ. Zool. 32, 301–394.Howard, H., 1974. Postcranial elements of the extinct condor Breagyps clarki (Miller).
Contrib. Sci. Nat. Hist. Mus. Los Angel. County 256, 1–24.Huber, C., Leuenberger, M., Spahni, R., Flückiger, J., Schwander, J., Stocker, T.F.,
Johnsen, S., Landais, A., Jouzel, J., 2006. Isotope calibrated greenland temperaturerecord over marine isotope stage 3 and its relation to CH4. Earth Planet. Sci. Lett.243, 504–519.
Jones, W., Rinderknecht, A., Migotto, R., Blanco, R.E., 2013. Body mass estimations andpaleobiological inferences on a new species of large caracara (Aves, Falconidae)from the Late Pleistocene of Uruguay. J. Paleontol. 87, 151–158.
Ledru, M.P., 1993. Late Quaternary environmental and climatic changes in Central Brazil.Quat. Res. 39, 90–98.
Livezey, B.C., 1986. A phylogenetic analysis of recent anseriform genera usingmorphologicalcharacters. Auk 105, 681–698.
Livezey, B.C., 1998. A phylogenetic analysis of the Gruiformes (Aves) based on morpho-logical characters, with an emphasis on the rails. Phil. Trans. R. Soc. London (Ser. B)353, 2077–2151.
Livezey, B.C., Martin, L.D., 1988. The systematic position of the Miocene anatid Anas [?]blanchardi Milne-Edwards. J. Vertebr. Paleontol. 8, 196–211.
Livezey, B.C., Zusi, R.L., 2006. Higher-order phylogeny of modern birds (Theropoda, Aves:Neornithes) based on comparative anatomy: I—methods and characters. Bull. Carne-gie Mus. Nat. Hist. 37, 1–544.
Louchart, A., Vignaud, P., Likius, A., Mackaye, H.T., Brunet, M., 2005. A new swan (Aves:Anatidae) in Africa, from the latest Miocene of Chad and Libya. J. Vertebr. Paleontol.25, 384–392.
Manegold, A., 2008. Passerine diversity in the Late Oligocene of Germany: earliest evi-dence for the sympatric coexistence of suboscines and Oscines. Ibis 150, 377–387.
Martin, G., 1996. Pré-história do Nordeste. Universitária-UFPE, Recife.Mayr, G., 1998. Coraciiforme” und “piciforme” Kleinvögel aus dem Mittel–Eozän der
Grube Messel (Hessen, Deutschland). Cour. Forsch.Inst. Senckenb. 205, 1–101.Mayr, G., Manegold, A., 2006a. A small suboscine-like passeriform bird from the early
Oligocene of France. Condor 108, 717–720.Mayr, G., Manegold, A., 2006b. New specimens of the earliest European passeriform bird.
Acta Palaeontol. Pol. 51, 315–323.Mayr, G., Smith, R., 2001. Ducks, rails, and limicoline waders (Aves: Anseriformes,
Gruiformes, Charadriiformes) from the lowermost Oligocene of Belgium. Geobios34, 547–561.
Miller, A.H., Bowman, R.I., 1956. Fossil birds of the late Pliocene of Cita Canyon, Texas.Wilson Bull. 68, 38–46.
Mourer-Chauviré, C., Hugueney, M., Jonet, P., 1989. Découverte de Passeriformes dansl'Oligocène supérieur de France. C. R. Acad. Sci. 309, 843–849.
Myers, N., Mittermeier, R.A., da Fonseca, C.G., Gustavo, A.B., Kent, J., 2000. Biodiversityhotspots for conservation priorities. Nature 403, 853–858.
Narosky, T., Di Giacomo, A.G., 1993. Las aves de la provincia de Buenos Aires. Distribucióny Estatus. Asociación Ornitológica del Plata, Vázquez Mazzini Editores and LOLA,Buenos Aires (127 pp.).
Navas, J.R., Bo, N., 1981. Nuevas aportaciones a la taxonomia de las razas geográficas deEudromia elegans y Eudromia formosa. Rev. Mus. Argent. Cienc. Nat. 11, 33–59.
Noriega, J.I., 1991. Un nuevo género de Furnariidae (Aves: Passeriformes) del PleistocenoInferior-Medio de la Provincia de Buenos Aires. Ameghiniana 28, 317–323.
Noriega, J.I., Areta, J.I., 2005. First record of Sarcoramphus Dumeril 1806 (Ciconiiformes:Vulturidae) from the Pleistocene of Buenos Aires province, Argentina. J. S. Am.Earth Sci. 20, 73–79.
Noriega, J.I., Tonni, E.P., 2007. Geronogyps reliquus Campbell (Ciconiiformes: Vulturidae)en el Pleistoceno tardío de la provincia de Entre Ríos y su significado ambiental.Ameghiniana 44, 245–250.
Noriega, J.I., Areta, J.I., Vizcaíno, S.F., Bargo, M.S., 2011. Phylogeny and taxonomy of thepatagonianMiocene falcon Thegornis musculosus Ameghino, 1895 (Aves: Falconidae).J. Paleontol. 85, 1089–1104.
Noriega, J.I., Soibelzon, L., Beilinson, E., Soibelzon, E., Diederle, J.M., 2012. A large crestedtinamous (Aves, Tinamidae, Eudromia) from the Late Pleistocene of Buenos AiresProvince, Argentina. Ameghiniana 49, R68.
Olrog, C.C., 1979. Nueva lista de la Avifauna Argentina. Oper. Lilloana 27, 1–324.Olson, S.L., 1973. A classification of the Rallidae. Wilson Bull. 85, 381–416.Olson, S.L., 1985. Faunal turnover in South American fossil avifaunas: the insufficiencies of
the fossil record. Evolution 39, 1174–1177.Olson, S.L., 1995. Thiornis sociata Navás, a nearly complete miocene grebe (Aves:
Podicipedidae). Cour. Forschungsinstitut Senckenb. 181, 131–140.Olson, S.L., 2008. A new species of very large, terrestrial caracara from Holocene deposits
in southern Jamaica (Aves: Falconidae). J. Raptor Res. 42, 265–272.Pardiñas, U.F.J., Teta, P., 2011. Fossil history of the marsh rats of the genus Holochilus and
Lundomys (Cricetidae, Sigmodontinae) in southern South America. Estud. Geol. 67,111–129.
Pomi, L.H., Tonni, E.P., 2011. Termite traces on bones from the Late Pleistocene ofArgentina. Ichnos 18, 166–171.

81M.M. Cenizo et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 420 (2015) 65–81
Prado, J.L., Menégaz, A.N., Tonni, E.P., Salemme, M.C., 1987. Los mamíferos de la FaunaLocal Paso Otero (Pleistoceno tardío), provincia de Buenos Aires. Aspectospaleoambientales y bioestratigráficos. Ameghiniana 24, 217–233.
Ribas, C.C., Moyle, R.G., Miyaki, C.Y., Cracraft, J., 2007. The assembly of montane biotas:linking Andean tectonics and climatic oscillations to independent regimes of diversi-fication in Pionus parrots. Proc. R. Soc. B 274, 2399–2408.
Rich, P.V., Walker, C.A., 1983. A new genus of Miocene flamingo from East Africa. Ostrich54, 95–104.
Stefanini, M.I., Gómez, R.O., 2012. Revisión sistemática de Pseudoseisuropsis nehuenNoriega, 1991 (Aves, Furnariidae) del Pleistoceno de provincia de Buenos Aires,Argentina. Ameghiniana 49, R68.
Storer, R.W., 1976. The Pleistocene pied-billed grebes (Aves: Podicipedidae). Smithson.Contrib. Paleobiol. 27, 147–153.
Stucchi, M., Emslie, S.D., 2005. A new condor (Ciconiiformes, Vulturidae) from the LateMiocene / Early Pliocene Piso Formation, Peru. Condor 107, 107–113.
Tambussi, C.P., 1995. Aves. In: Alberdi, M., Leone, G., Tonni, E.P. (Eds.), Evolución biológicay climática de la Región Pampeana durante los últimos cinco millones de años.Monografía del Museo Nacional de Ciencias Naturales, Madrid 7, pp. 145–161.
Tambussi, C.P., Noriega, J.I., 1999. The fossil record of condors (Ciconiiformes: Vulturidae)in Argentina. Smithson. Contrib. Paleobiol. 89, 177–184.
Teta, P., Pardiñas, U.F.J., 2006. Pleistocene record of the marsh rat of the genus Lundomys inSouthern South America: Paleoclimatic significance. Curr. Res. Pleistocene 23, 202–204.
Tonni, E.P., 1977. Los tinámidos fósiles argentinos. I, El género Tinamisornis Rovereto,1914. Ameghiniana 14, 224–232.
Tonni, E.P., Fidalgo, F., 1982. Geología y Paleontología de los sedimentos del Pleistocenoen el área de Punta Hermengo (Miramar, provincia de Buenos Aires, RepúblicaArgentina): aspectos paleoclimáticos. Ameghiniana 19, 79–108.
Tonni, E.P., Laza, J.H., 1980. Las aves de la Fauna Local Paso Otero (Pleistoceno Tardío) dela provincia de Buenos Aires. Su significación ecológica, climática y zoogeográfica.Ameghiniana 17, 313–322.
Tonni, E.P., Noriega, J.I., 1998. Los Cóndores (Ciconiiformes, Vulturidae) de la RegiónPampeana de la Argentina durante el Cenozoico tardío: Distribución, interaccionesy extinciones. Ameghiniana 35, 141–150.
Tonni, E.P., Noriega, J.I., 2001. Una especie extinta de Pseudoseisura Reichenbach, 1854(Passeriformes: Furnariidae) del Pleistoceno de la Argentina: comentarios filogenéticos.Ornitol. Neotropical 12, 29–44.
Tonni, E.P., Cione, A.L., Figini, A.J., 1999. Predominance of arid climates indicated bymammals in the pampas of Argentina during the late Pleistocene and Holocene.Palaeogeogr. Palaeoclimatol. Palaeoecol. 147, 257–281.
Tonni, E.P., Huarte, R.A., Carbonari, J.E., Figini, A.J., 2003. New radiocarbon chronologyfor the Guerrero Member of the Luján Formation (Buenos Aires, Argentina):palaeoclimatic significance. Quat. Int. 109–110, 45–48.
Vaurie, C., 1980. Taxonomy and geographical distribution of the Furnariidae (Aves,Passeriformes). Bull. Am. Mus. Nat. Hist. 166, 1–357.
Voss, R.S., Carleton, M.D., 1993. A new genus forHesperomysmolitorWinge and Holochilusmagnus Hershkovitz, with comments on phylogenetic relationships and oryzomyinemonophyly. Am. Mus. Novit. 3085, 1–39.
Vuilleumier, F., 1970. Generic relations and speciation patterns in the Caracaras (Aves;Falconidae). Breviora 355, 1–29.
Wetmore, A., 1924. Fossil birds from south eastern Arizona. Proc. U. S. Natl. Mus. 64, 1–18.Wetmore, A., 1939. Recent observations on the Eskimo Curlew in Argentina. Auk 56,
475–476.Wijninga, V.M., 1995. A first approximation of montane forest development during the
late Tertiary in Colombia. In: Churchill, S.P., Balslev, H., Forero, E., Luteyn, J.L. (Eds.),Biodiversity and Conservation of Neotropical Montane Forests. The New York BotanicalGarden, pp. 23–34.
Winge, H., 1887. Jordfundne og nulevende Gnavere (Rodentia) fra Lagoa Santa, MinasGeraes, Brasilien: med udsigt over gnavernes indbyrdes slægtskab. F. Dreyer,Kjøbenhavn (178 pp).
Winge, O., 1888. Fugle fra Knuglehuler i Brasilien. E. Museo Lundii 1, 1–54.Woolfenden, G.E., 1961. Postcraneal osteology of the waterfowl. Bull. Fla. State Mus. 6,
1–129.Worthy, T.H., 2009. Descriptions and phylogenetic relationships of two new genera and
four new species of Oligo-Miocene waterfowl (Aves: Anatidae) from Australia. Zool.J. Linnean Soc. 156, 411–454.
Worthy, T.H., Lee, M.S.Y., 2008. Affinities of Miocene waterfowl (Anatidae: Manuherikia,Dunstanetta andMiotadorna) from the St. Bathansfauna, New Zealand. Palaeontology51, 677–708.




















