
Stability of the sponge assemblage of the Mediterranean coralligenous along a millennial span of...
10
ORIGINAL ARTICLE Stability of the sponge assemblage of Mediterranean coralligenous concretions along a millennial time span Marco Bertolino 1 , Barbara Calcinai 2 , Riccardo Cattaneo-Vietti 2 , Carlo Cerrano 2 , Anna Lafratta 2 , Maurizio Pansini 1 , Daniela Pica 2 & Giorgio Bavestrello 1 1 Dipartimento di Scienze della Terra, dell’Ambiente e della Vita, Universit a di Genova, Genova, Italy 2 Dipartimento di Scienze della Vita e dell’Ambiente, Universit a Politecnica delle Marche, Ancona, Italy Keywords Coralligenous; ecosystem stability; Ligurian Sea; Porifera; sponge spicules. Correspondence Marco Bertolino, Dipartimento di Scienze della Terra, dell’Ambiente e della Vita, Universit a di Genova, Corso Europa 26, 16132 Genova, Italy. E-mail: [email protected] Accepted: 4 March 2013 doi: 10.1111/maec.12063 Abstract The Mediterranean coralligenous substratum is a hard bottom of biogenic ori- gin, mainly composed of calcareous algae, growing in dim light conditions. Sponges are among of the most representative taxa of the coralligenous assem- blages, with more than 300 recorded species of different habits: massive, erect, boring and insinuating. When sponges die, their siliceous spicules remain trapped in the biogenic concretion, offering the opportunity to describe the coralligenous spongofauna over a very long span of time, virtually dating back to a large part of the Holocene period. The data reported here were obtained from core samples collected from four coralligenous concretions. Each block was collected in a different locality of the Ligurian Sea: Santo Stefano Shoals, Bogliasco, Punta del Faro (Portofino Promontory) and Punta Manara. Radio- carbon age determinations indicate for these conglomerates a maximal age between 1600 and 3100 years. The spicules trapped in the cores show deep dis- solution marks in the form of circular holes on their surface or present an enlargement of the axial canal. However, their original shape, generally intact, suggests the absence of mechanical injuries and allows a tentative identification at the species level. The analysis of these old spicules reveals an ancient sponge assemblage composed of 30 recognisable species. This indicates that almost one half of the sponge community today settled on coralligenous substrata has been present in the conglomerates for their entire existence. Introduction Coralligenous concretions are calcareous formations of biogenic origin in Mediterranean benthic environments produced by the accumulation of encrusting algae and invertebrates with calcareous structures. Coralligenous communities, together with Posidonia oceanica meadows (Ballesteros 2006), are the most important ‘hot spots’ of biodiversity in the Mediterranean. The few existing radio- carbon datings of the coralligenous assemblages suggest that these concretions had their maximum development between 8000 and 5000 years ago. After that period, appreciable accumulation rates are recorded only for con- cretions situated in relatively shallow waters (10– 35 m depth), whereas the accumulation rates of corallig- enous assemblages deeper than 50 m are negligible (Sar- toretto 1996; Sartoretto et al. 1996; Ballesteros 2006). Sponges, living with different growing strategies in the different micro-habitats offered by the concretions, are one of the most diversified groups of the coralligenous assemblage, with 304 recorded species (Bertolino et al. 2009; Bertolino 2011). Generally, species living on the coralligenous surface (massive, branching or encrusting) are well known, whereas studies of boring and insinuating species, which penetrate deep into the concretion, are rare. In spite of this paucity of knowledge, these sponges play important roles in the stability of the concretions. In fact, the highly destructive boring species play a major Marine Ecology (2013) 1–10 ª 2013 Blackwell Verlag GmbH 1 Marine Ecology. ISSN 0173-9565
Transcript of Stability of the sponge assemblage of the Mediterranean coralligenous along a millennial span of...

ORIGINAL ARTICLE
Stability of the sponge assemblage of Mediterraneancoralligenous concretions along a millennial time spanMarco Bertolino1, Barbara Calcinai2, Riccardo Cattaneo-Vietti2, Carlo Cerrano2, Anna Lafratta2,Maurizio Pansini1, Daniela Pica2 & Giorgio Bavestrello1
1 Dipartimento di Scienze della Terra, dell’Ambiente e della Vita, Universit�a di Genova, Genova, Italy
2 Dipartimento di Scienze della Vita e dell’Ambiente, Universit�a Politecnica delle Marche, Ancona, Italy
Keywords
Coralligenous; ecosystem stability; Ligurian
Sea; Porifera; sponge spicules.
Correspondence
Marco Bertolino, Dipartimento di Scienze
della Terra, dell’Ambiente e della Vita,
Universit�a di Genova, Corso Europa 26,
16132 Genova, Italy.
E-mail: [email protected]
Accepted: 4 March 2013
doi: 10.1111/maec.12063
Abstract
The Mediterranean coralligenous substratum is a hard bottom of biogenic ori-
gin, mainly composed of calcareous algae, growing in dim light conditions.
Sponges are among of the most representative taxa of the coralligenous assem-
blages, with more than 300 recorded species of different habits: massive, erect,
boring and insinuating. When sponges die, their siliceous spicules remain
trapped in the biogenic concretion, offering the opportunity to describe the
coralligenous spongofauna over a very long span of time, virtually dating back
to a large part of the Holocene period. The data reported here were obtained
from core samples collected from four coralligenous concretions. Each block
was collected in a different locality of the Ligurian Sea: Santo Stefano Shoals,
Bogliasco, Punta del Faro (Portofino Promontory) and Punta Manara. Radio-
carbon age determinations indicate for these conglomerates a maximal age
between 1600 and 3100 years. The spicules trapped in the cores show deep dis-
solution marks in the form of circular holes on their surface or present an
enlargement of the axial canal. However, their original shape, generally intact,
suggests the absence of mechanical injuries and allows a tentative identification
at the species level. The analysis of these old spicules reveals an ancient sponge
assemblage composed of 30 recognisable species. This indicates that almost one
half of the sponge community today settled on coralligenous substrata has been
present in the conglomerates for their entire existence.
Introduction
Coralligenous concretions are calcareous formations of
biogenic origin in Mediterranean benthic environments
produced by the accumulation of encrusting algae and
invertebrates with calcareous structures. Coralligenous
communities, together with Posidonia oceanica meadows
(Ballesteros 2006), are the most important ‘hot spots’ of
biodiversity in the Mediterranean. The few existing radio-
carbon datings of the coralligenous assemblages suggest
that these concretions had their maximum development
between 8000 and 5000 years ago. After that period,
appreciable accumulation rates are recorded only for con-
cretions situated in relatively shallow waters (10–
35 m depth), whereas the accumulation rates of corallig-
enous assemblages deeper than 50 m are negligible (Sar-
toretto 1996; Sartoretto et al. 1996; Ballesteros 2006).
Sponges, living with different growing strategies in the
different micro-habitats offered by the concretions, are
one of the most diversified groups of the coralligenous
assemblage, with 304 recorded species (Bertolino et al.
2009; Bertolino 2011). Generally, species living on the
coralligenous surface (massive, branching or encrusting)
are well known, whereas studies of boring and insinuating
species, which penetrate deep into the concretion, are
rare. In spite of this paucity of knowledge, these sponges
play important roles in the stability of the concretions. In
fact, the highly destructive boring species play a major
Marine Ecology (2013) 1–10 ª 2013 Blackwell Verlag GmbH 1
Marine Ecology. ISSN 0173-9565

role in the recycling of the carbonates stored in the
coralligenous concretions (Davies 1983; Sorokin 1993;
Becker & Reaka-Kudla 1997). In contrast, insinuating spe-
cies, filling the crevices of the concretions, enhance the
cohesion of the conglomerate (Cerrano et al. 2001).
The skeletons of most demosponges (excluding horny
sponges and a few other species) are composed of dis-
crete siliceous elements called spicules, which show a
large variety of shapes and sizes; these are commonly
used in sponge identification. Sponge spicules enrich the
fine sediment fraction of biogenic silica when dead ani-
mals decay (R€utzler & Macintyre 1978). In Antarctic
waters, sponge spicules can form mats 1.5 m or more
thick (Koltun 1968; Dayton 1979; Barthel 1992; Cattaneo-
Vietti et al. 1999; Gutt 2007).
The numbers and types of sponge spicules in dated
sediment core samples have been used to reconstruct the
dynamics of freshwater sponge assemblages in lakes or
lagoons (Paduano & Fell 1997; Volkmer-Ribeiro et al.
2007). Bavestrello et al. (1996) evaluated the contribu-
tion of this form of low-soluble silica to the bottom
sediments of different areas of the Ligurian Sea. Micro-
scopical analysis of the fine fraction of the sediments,
coming from several stations, demonstrated that sponge
spicules represent nearly the total BSi (biogenic silica),
other siliceous remains, such as diatom and silicoflagel-
late tests, constituting a negligible fraction. Similar
results were recorded from cores taken from a Posidonia
oceanica meadow in the Mediterranean (Bertolino et al.
2011).
When sponges living in coralligenous crevices die, their
siliceous spicules likely become trapped in the biogenic
concretion, offering the unique possibility to describe
the coralligenous spongofauna over a very long span of
time, virtually dating back to a large part of the Holocene
period.
The aim of this paper is to describe the ancient sponge
assemblages on the base of spicules recorded inside blocks
of coralligenous material, assuming that spicules present
inside the conglomerates may have approximately the
same age of the surrounding bio-deposed carbonates. The
comparison of these ancient sponge assemblages with the
living ones recorded on the surface of the concretions
allowed an estimation of the stability level of the corallig-
enous community along a millennial span of time.
Material and Methods
The coralligenous blocks were collected by SCUBA diving
from four localities of the Ligurian Sea: at the Santo Stef-
ano Shoals (Western Ligurian Riviera), Bogliasco, Punta
del Faro (Portofino Promontory) and Punta Manara
(Eastern Ligurian Riviera) (Fig. 1). In the first three
localities, the conglomerate blocks came from the hori-
zontal rim at the base of the vertical cliffs, at a depth
ranging of 30–40 m. At Bogliasco, the sampled block was
from a vertical pinnacle arising from a coralligenous bank
on a flat bottom at 15 m depth. The portions of con-
glomerate were detached using hammer and chisel. For
each block, with a volume of 30–40 l, all the living
sponge species present on the surface (massive, erected or
encrusting) were collected and identified to species level
(Fig. 2A). Sponges without spicules (horny sponges,
Chondrosia reniformis, Oscarella sp.) were identified but
were not considered in the following analysis because of
the impossibility of checking their presence in the ancient
assemblages.
Each block was cut in 2-cm-thick slices running from
the surface toward the plane of junction with the rocky
substratum, perpendicular to the plane of development of
the coralline algae (Fig. 2A,B). Each side of the slices was
observed under stereomicroscope to detect the living,
endolithic (boring or insinuating) sponges present in the
first layer (1–2 cm thick) of the conglomerates (Fig. 2C).
These specimens were identified to species level.
Finally, from the central slice of each block, one verti-
cal sample 9 cm long extending from the surface (consid-
ered the youngest part of the conglomerate) to the face
Fig. 1. The four studied localities along the
Ligurian Coast: St. Stefano Shoal (1),
Bogliasco (2), Punta del Faro (3) and Punta
Manara (4).
2 Marine Ecology (2013) 1–10 ª 2013 Blackwell Verlag GmbH
Stability of Mediterranean coralligenous sponge assemblage Bertolino, Calcinai, Cattaneo-Vietti, Cerrano, Lafratta, Pansini, Pica & Bavestrello

of the block originally attached to the substratum (con-
sidered the oldest part of the conglomerate) was obtained
and divided into nine portions, each 2 9 2 9 1 cm
(4 cm3) (Fig. 2D). Each small portion of the vertical sam-
ple was placed in hydrogen peroxide (240 vol) that was
changed three times at intervals of 24 h. This method
released all of the sediment entrapped in the cavities of
the conglomerate.
The average amounts of sediment obtained from the
different localities were 0.45 � 0.09 g, 0.8 � 0.12 g,
0.33 � 0.06 g and 0.21 � 0.06 g for St. Stefano Shoal,
Bogliasco, Punta del Faro and Punta Manara, respectively.
The conglomerate pieces were removed and dried while
the sediments were treated with boiling nitric acid to
eliminate the carbonatic fraction, rinsed twice in distilled
water and twice in alcohol 95%, and dried. For each sam-
ple of sediment, three replicates of 50 mg were suspended
in 300 ll of Bioclear solvent and mounted on a micro-
scope slide. Microscopic analysis of the spicules contained
in the sediment was done to identify the sponge species
living in ancient times. We have identified only the spe-
cies characterized by peculiar, diagnostic spicules. Finally,
the conglomerate fragments were gold-sputtered (Balzers
Union Evaporator) and observed by scanning electron
microscopy (SEM Philips XL 20).
After the sediment extraction, three subsamples, the most
superficial, the deepest and an intermediate one (Fig. 2D),
were used for dating by 14C using high-resolution
mass spectrometry at the Center of Dating and Diagnostic
(CEDAD) of the University of Salento (http://cedad.unisal-
ento.it).
Statistical analysis
Sponge data were analysed using PAST 2.15, a software
package of computer programs including common sta-
tistical and modelling functions (Hammer et al. 2001).
Data on presence/absence of sponge species were com-
piled prior to analysis. Cluster analysis was used to
check for similarities between the ancient and living
sponge assemblages in the four localities (St. Stefano
Shoal, Bogliasco, Punta del Faro, Punta Manara). This
analysis was run using the average linkage algorithm
(UPGMA method) of Bray–Curtis distances among
samples.
Results
Living sponge assemblages
The living sponge species recorded on and inside the
blocks collected in the four localities are listed in Table 1.
Of a total of 75 determined species (excluding horny
sponge species and other species lacking spicules), 46
(61.3%) were encrusting, 14 (18.6%) insinuating, 10
(13.3%) massive and seven (9.3%) boring.
The assemblages of the different blocks are strongly
diversified: only the insinuating Dercitus (Stoeba) plicatus
was present in the blocks of all the four localities. Com-
paring pairs of localities hosting coralligenous rims (St.
Stefano Shoal, Punta Manara, Punta del Faro) the num-
ber of shared species ranged from 5 to 15. Comparing
Bogliasco, hosting coralligenous banks, with the other
A
D
B
C
Fig. 2. Protocol for processing the
coralligenous blocks. From a newly collected
block (A) all the sponges living on the surface
were recorded. The block was then cut into
2- cm thick slices (A–B) and the insinuating or
boring specimens (arrow) observed in each
slice were collected (C). From the central slice
of the block, a vertical core sample 9 cm high
was obtained and divided (D) into nine
portions, each 2 9 2 9 1 cm. Each portion
was used for the ancient spicules extraction.
The most superficial, the deepest and the
intermediate portions (arrows) were used for
radiocarbon dating.
Marine Ecology (2013) 1–10 ª 2013 Blackwell Verlag GmbH 3
Bertolino, Calcinai, Cattaneo-Vietti, Cerrano, Lafratta, Pansini, Pica & Bavestrello Stability of Mediterranean coralligenous sponge assemblage
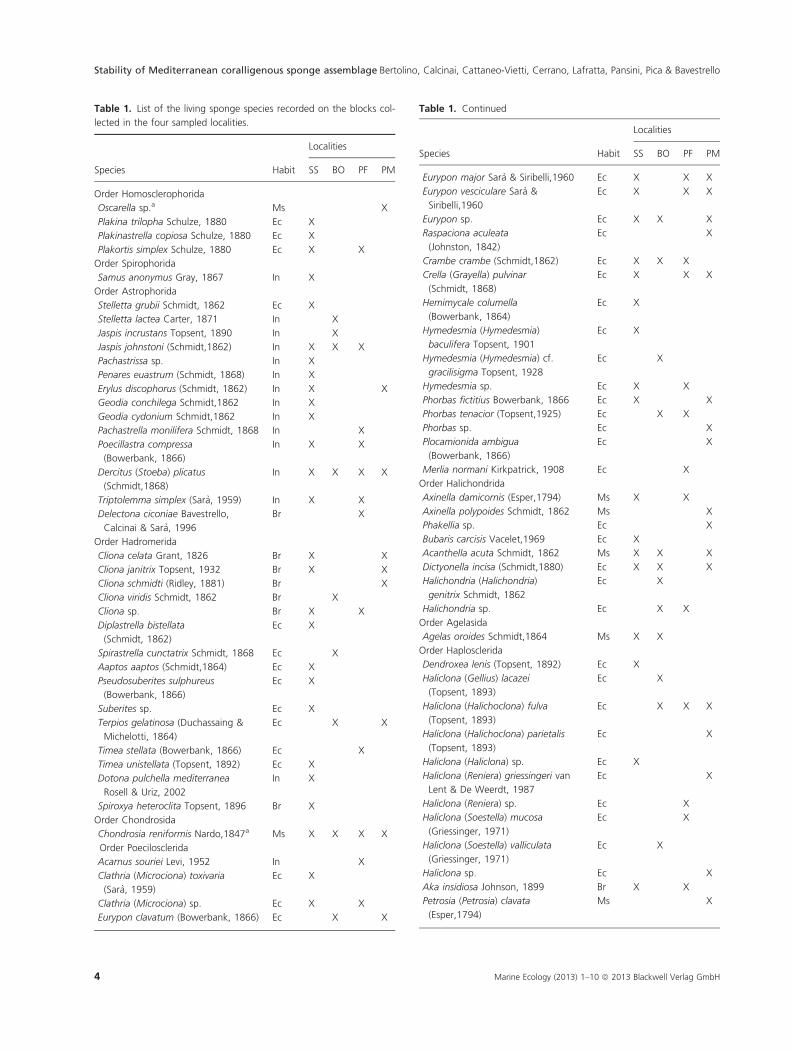
Table 1. List of the living sponge species recorded on the blocks col-
lected in the four sampled localities.
Species Habit
Localities
SS BO PF PM
Order Homosclerophorida
Oscarella sp.a Ms X
Plakina trilopha Schulze, 1880 Ec X
Plakinastrella copiosa Schulze, 1880 Ec X
Plakortis simplex Schulze, 1880 Ec X X
Order Spirophorida
Samus anonymus Gray, 1867 In X
Order Astrophorida
Stelletta grubii Schmidt, 1862 Ec X
Stelletta lactea Carter, 1871 In X
Jaspis incrustans Topsent, 1890 In X
Jaspis johnstoni (Schmidt,1862) In X X X
Pachastrissa sp. In X
Penares euastrum (Schmidt, 1868) In X
Erylus discophorus (Schmidt, 1862) In X X
Geodia conchilega Schmidt,1862 In X
Geodia cydonium Schmidt,1862 In X
Pachastrella monilifera Schmidt, 1868 In X
Poecillastra compressa
(Bowerbank, 1866)
In X X
Dercitus (Stoeba) plicatus
(Schmidt,1868)
In X X X X
Triptolemma simplex (Sar�a, 1959) In X X
Delectona ciconiae Bavestrello,
Calcinai & Sar�a, 1996
Br X
Order Hadromerida
Cliona celata Grant, 1826 Br X X
Cliona janitrix Topsent, 1932 Br X X
Cliona schmidti (Ridley, 1881) Br X
Cliona viridis Schmidt, 1862 Br X
Cliona sp. Br X X
Diplastrella bistellata
(Schmidt, 1862)
Ec X
Spirastrella cunctatrix Schmidt, 1868 Ec X
Aaptos aaptos (Schmidt,1864) Ec X
Pseudosuberites sulphureus
(Bowerbank, 1866)
Ec X
Suberites sp. Ec X
Terpios gelatinosa (Duchassaing &
Michelotti, 1864)
Ec X X
Timea stellata (Bowerbank, 1866) Ec X
Timea unistellata (Topsent, 1892) Ec X
Dotona pulchella mediterranea
Rosell & Uriz, 2002
In X
Spiroxya heteroclita Topsent, 1896 Br X
Order Chondrosida
Chondrosia reniformis Nardo,1847a Ms X X X X
Order Poecilosclerida
Acarnus souriei Levi, 1952 In X
Clathria (Microciona) toxivaria
(Sar�a, 1959)
Ec X
Clathria (Microciona) sp. Ec X X
Eurypon clavatum (Bowerbank, 1866) Ec X X
Table 1. Continued
Species Habit
Localities
SS BO PF PM
Eurypon major Sar�a & Siribelli,1960 Ec X X X
Eurypon vesciculare Sar�a &
Siribelli,1960
Ec X X X
Eurypon sp. Ec X X X
Raspaciona aculeata
(Johnston, 1842)
Ec X
Crambe crambe (Schmidt,1862) Ec X X X
Crella (Grayella) pulvinar
(Schmidt, 1868)
Ec X X X
Hemimycale columella
(Bowerbank, 1864)
Ec X
Hymedesmia (Hymedesmia)
baculifera Topsent, 1901
Ec X
Hymedesmia (Hymedesmia) cf.
gracilisigma Topsent, 1928
Ec X
Hymedesmia sp. Ec X X
Phorbas fictitius Bowerbank, 1866 Ec X X
Phorbas tenacior (Topsent,1925) Ec X X
Phorbas sp. Ec X
Plocamionida ambigua
(Bowerbank, 1866)
Ec X
Merlia normani Kirkpatrick, 1908 Ec X
Order Halichondrida
Axinella damicornis (Esper,1794) Ms X X
Axinella polypoides Schmidt, 1862 Ms X
Phakellia sp. Ec X
Bubaris carcisis Vacelet,1969 Ec X
Acanthella acuta Schmidt, 1862 Ms X X X
Dictyonella incisa (Schmidt,1880) Ec X X X
Halichondria (Halichondria)
genitrix Schmidt, 1862
Ec X
Halichondria sp. Ec X X
Order Agelasida
Agelas oroides Schmidt,1864 Ms X X
Order Haplosclerida
Dendroxea lenis (Topsent, 1892) Ec X
Haliclona (Gellius) lacazei
(Topsent, 1893)
Ec X
Haliclona (Halichoclona) fulva
(Topsent, 1893)
Ec X X X
Haliclona (Halichoclona) parietalis
(Topsent, 1893)
Ec X
Haliclona (Haliclona) sp. Ec X
Haliclona (Reniera) griessingeri van
Lent & De Weerdt, 1987
Ec X
Haliclona (Reniera) sp. Ec X
Haliclona (Soestella) mucosa
(Griessinger, 1971)
Ec X
Haliclona (Soestella) valliculata
(Griessinger, 1971)
Ec X
Haliclona sp. Ec X
Aka insidiosa Johnson, 1899 Br X X
Petrosia (Petrosia) clavata
(Esper,1794)
Ms X
4 Marine Ecology (2013) 1–10 ª 2013 Blackwell Verlag GmbH
Stability of Mediterranean coralligenous sponge assemblage Bertolino, Calcinai, Cattaneo-Vietti, Cerrano, Lafratta, Pansini, Pica & Bavestrello
three localities, the number of shared species ranged from
six to seven.
The sponge fauna recorded on the block from the St.
Stefano Shoal is the most diverse (44 species). In the
other three localities the number of species is very similar
(25, 24 and 21 species for Punta del Faro, Punta Manara
and Bogliasco, respectively). Massive and erect species are
the less common forms (two at Punta del Faro to four at
St. Stefano Shoal), whereas the encrusting species are very
abundant (from 13 at Bogliasco to 24 at St. Stefano
Shoal). Insinuating sponges are numerous at the St. Stef-
ano Shoal (11 species) but show lower values at Punta
del Faro (seven), Bogliasco (four) and Punta Manara
(two). Few boring species are recorded in the studied
blocks; in this case as well, the highest number is
observed at the St. Stefano Shoal (five).
Although the values of species richness are different in
the blocks studied, the encrusting species represent the
most abundant category, ranging from 53% to 66% of
the total. The insinuating species vary from 8% to 27%,
the boring species from 4% to 12.5% and the massive
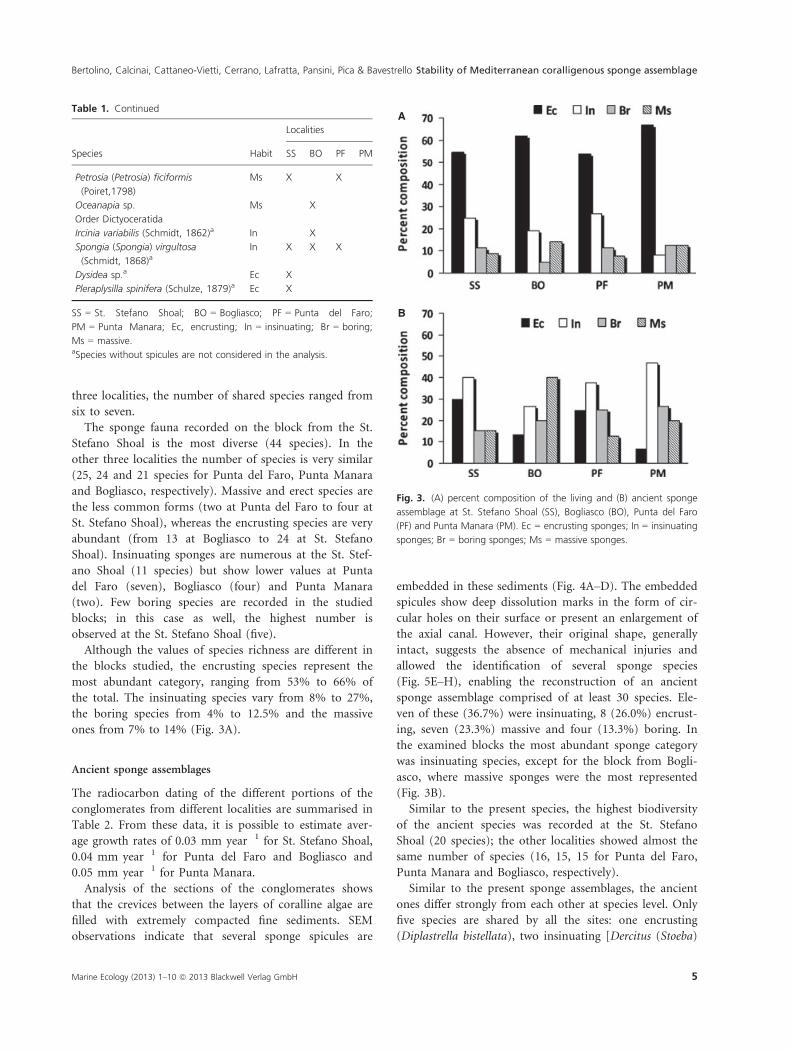
ones from 7% to 14% (Fig. 3A).
Ancient sponge assemblages
The radiocarbon dating of the different portions of the
conglomerates from different localities are summarised in
Table 2. From these data, it is possible to estimate aver-
age growth rates of 0.03 mm year�1 for St. Stefano Shoal,
0.04 mm year�1 for Punta del Faro and Bogliasco and
0.05 mm year�1 for Punta Manara.
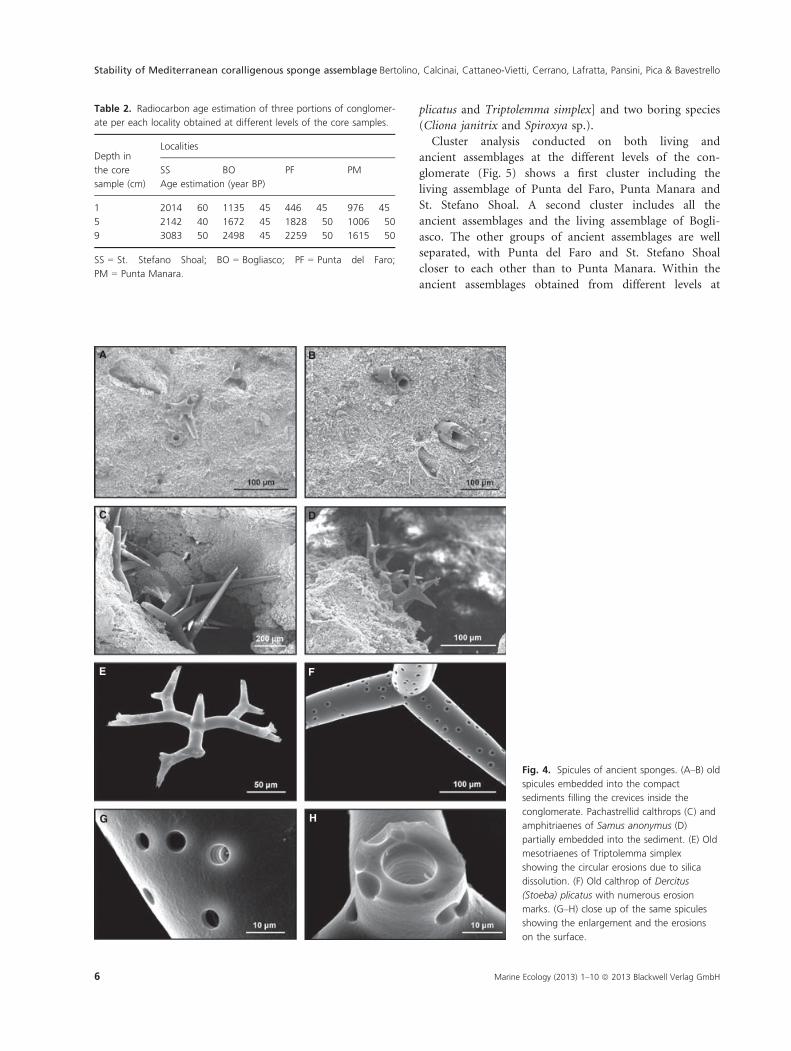
Analysis of the sections of the conglomerates shows
that the crevices between the layers of coralline algae are
filled with extremely compacted fine sediments. SEM
observations indicate that several sponge spicules are
embedded in these sediments (Fig. 4A–D). The embedded
spicules show deep dissolution marks in the form of cir-
cular holes on their surface or present an enlargement of
the axial canal. However, their original shape, generally
intact, suggests the absence of mechanical injuries and
allowed the identification of several sponge species
(Fig. 5E–H), enabling the reconstruction of an ancient
sponge assemblage comprised of at least 30 species. Ele-
ven of these (36.7%) were insinuating, 8 (26.0%) encrust-
ing, seven (23.3%) massive and four (13.3%) boring. In
the examined blocks the most abundant sponge category
was insinuating species, except for the block from Bogli-
asco, where massive sponges were the most represented
(Fig. 3B).
Similar to the present species, the highest biodiversity
of the ancient species was recorded at the St. Stefano
Shoal (20 species); the other localities showed almost the
same number of species (16, 15, 15 for Punta del Faro,
Punta Manara and Bogliasco, respectively).
Similar to the present sponge assemblages, the ancient
ones differ strongly from each other at species level. Only
five species are shared by all the sites: one encrusting
(Diplastrella bistellata), two insinuating [Dercitus (Stoeba)
A
B
Fig. 3. (A) percent composition of the living and (B) ancient sponge
assemblage at St. Stefano Shoal (SS), Bogliasco (BO), Punta del Faro
(PF) and Punta Manara (PM). Ec = encrusting sponges; In = insinuating
sponges; Br = boring sponges; Ms = massive sponges.
Table 1. Continued
Species Habit
Localities
SS BO PF PM
Petrosia (Petrosia) ficiformis
(Poiret,1798)
Ms X X
Oceanapia sp. Ms X
Order Dictyoceratida
Ircinia variabilis (Schmidt, 1862)a In X
Spongia (Spongia) virgultosa
(Schmidt, 1868)aIn X X X
Dysidea sp.a Ec X
Pleraplysilla spinifera (Schulze, 1879)a Ec X
SS = St. Stefano Shoal; BO = Bogliasco; PF = Punta del Faro;
PM = Punta Manara; Ec, encrusting; In = insinuating; Br = boring;
Ms = massive.aSpecies without spicules are not considered in the analysis.
Marine Ecology (2013) 1–10 ª 2013 Blackwell Verlag GmbH 5
Bertolino, Calcinai, Cattaneo-Vietti, Cerrano, Lafratta, Pansini, Pica & Bavestrello Stability of Mediterranean coralligenous sponge assemblage
plicatus and Triptolemma simplex] and two boring species
(Cliona janitrix and Spiroxya sp.).
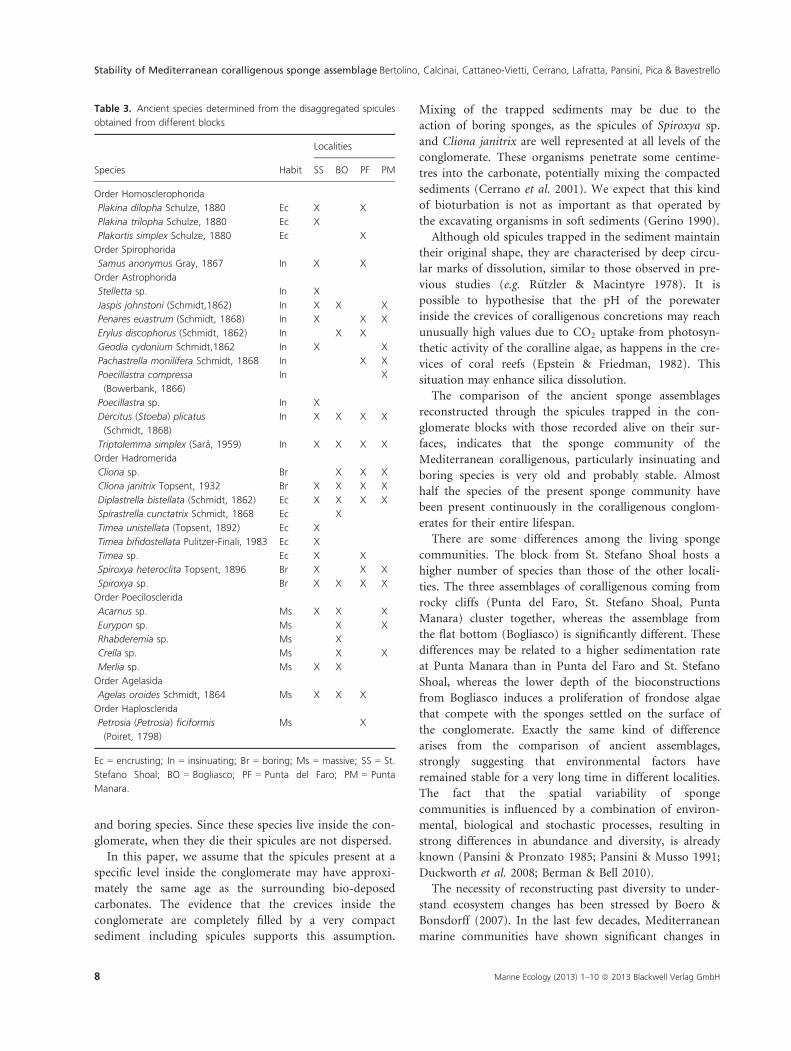
Cluster analysis conducted on both living and
ancient assemblages at the different levels of the con-
glomerate (Fig. 5) shows a first cluster including the
living assemblage of Punta del Faro, Punta Manara and
St. Stefano Shoal. A second cluster includes all the
ancient assemblages and the living assemblage of Bogli-
asco. The other groups of ancient assemblages are well
separated, with Punta del Faro and St. Stefano Shoal
closer to each other than to Punta Manara. Within the
ancient assemblages obtained from different levels at
Table 2. Radiocarbon age estimation of three portions of conglomer-
ate per each locality obtained at different levels of the core samples.
Depth in
the core
sample (cm)
Localities
SS BO PF PM
Age estimation (year BP)
1 2014 � 60 1135 � 45 446 � 45 976 � 45
5 2142 � 40 1672 � 45 1828 � 50 1006 � 50
9 3083 � 50 2498 � 45 2259 � 50 1615 � 50
SS = St. Stefano Shoal; BO = Bogliasco; PF = Punta del Faro;
PM = Punta Manara.
A B
C D
E F
G H
Fig. 4. Spicules of ancient sponges. (A–B) old
spicules embedded into the compact
sediments filling the crevices inside the
conglomerate. Pachastrellid calthrops (C) and
amphitriaenes of Samus anonymus (D)
partially embedded into the sediment. (E) Old
mesotriaenes of Triptolemma simplex
showing the circular erosions due to silica
dissolution. (F) Old calthrop of Dercitus
(Stoeba) plicatus with numerous erosion
marks. (G–H) close up of the same spicules
showing the enlargement and the erosions
on the surface.
6 Marine Ecology (2013) 1–10 ª 2013 Blackwell Verlag GmbH
Stability of Mediterranean coralligenous sponge assemblage Bertolino, Calcinai, Cattaneo-Vietti, Cerrano, Lafratta, Pansini, Pica & Bavestrello

each locality it is not possible to observe a pattern of
similarity related to the hypothesised age of the con-
glomerate.
Discussion
This study shows that spicules trapped inside coralligenous
conglomerates can be used to describe the diversity of
ancient sponge communities. Data on the dating of the
Mediterranean coralligenous and its growth rates are very
scarce (Sartoretto 1996; Sartoretto et al. 1996). The known
accumulation rate of the constructions is considered very
low (0.006–0.83 mm year�1) and oscillates greatly depend-
ing on depth and time period. The higher accumulation
rates (0.20–0.83 mm year�1) were recorded for deep coral-
ligenous concretions and correspond to a period between
8000 and 5000 years BP. More recently, the only apprecia-
ble accumulation rates (0.11–0.42 mm year�1) have been
recorded for coralligenous concretions situated in relatively
shallow waters (10–35 m depth�1) (Sartoretto 1996; Sarto-
retto et al. 1996). The findings of our study strongly agree
with evaluations obtained from constructions recorded in
the same depth range.
The recorded differences in the number of sponge spe-
cies between living and ancient assemblages (Tables 1–3)are due to the obvious difficulty of identifying species
relying only on disaggregated spicules. Moreover, it is evi-
dent, in all the studied localities, that spicules from
encrusting species are rarely preserved inside the con-
glomerate. This is due to the low biomass of these kinds
of sponges and to their position on the conglomerate sur-
face inducing spicule dispersion when sponges decay.
Owing to this, the diversity of encrusting species was
probably higher in ancient times than the present study
suggests. The relatively high number of spicules belonging
to massive sponges is due to the high quantity of spicules
present in these species. When a massive specimen
decays, a large amount of spicules is probably dispersed
and there is a higher probability that a fraction of them
will penetrate the crevices of the coralligenous. About half
of the ancient species recognised from the spicules
trapped inside the conglomerate belong to insinuating
Fig. 5. Dendogram of the four living sponge
assemblages (SS, BO, PF, PM) and the ancient
sponge assemblages at the different levels of
the core samples from St. Stefano Shoal (SS
1–9), Bogliasco (BO 1–9), Punta del Faro (PF
1–9) and Punta Manara (PM 1–9).
Marine Ecology (2013) 1–10 ª 2013 Blackwell Verlag GmbH 7
Bertolino, Calcinai, Cattaneo-Vietti, Cerrano, Lafratta, Pansini, Pica & Bavestrello Stability of Mediterranean coralligenous sponge assemblage
and boring species. Since these species live inside the con-
glomerate, when they die their spicules are not dispersed.
In this paper, we assume that the spicules present at a
specific level inside the conglomerate may have approxi-
mately the same age as the surrounding bio-deposed
carbonates. The evidence that the crevices inside the
conglomerate are completely filled by a very compact
sediment including spicules supports this assumption.
Mixing of the trapped sediments may be due to the
action of boring sponges, as the spicules of Spiroxya sp.
and Cliona janitrix are well represented at all levels of the
conglomerate. These organisms penetrate some centime-
tres into the carbonate, potentially mixing the compacted
sediments (Cerrano et al. 2001). We expect that this kind
of bioturbation is not as important as that operated by
the excavating organisms in soft sediments (Gerino 1990).
Although old spicules trapped in the sediment maintain
their original shape, they are characterised by deep circu-
lar marks of dissolution, similar to those observed in pre-
vious studies (e.g. R€utzler & Macintyre 1978). It is
possible to hypothesise that the pH of the porewater
inside the crevices of coralligenous concretions may reach
unusually high values due to CO2 uptake from photosyn-
thetic activity of the coralline algae, as happens in the cre-
vices of coral reefs (Epstein & Friedman, 1982). This
situation may enhance silica dissolution.
The comparison of the ancient sponge assemblages
reconstructed through the spicules trapped in the con-
glomerate blocks with those recorded alive on their sur-
faces, indicates that the sponge community of the
Mediterranean coralligenous, particularly insinuating and
boring species is very old and probably stable. Almost
half the species of the present sponge community have
been present continuously in the coralligenous conglom-
erates for their entire lifespan.
There are some differences among the living sponge
communities. The block from St. Stefano Shoal hosts a
higher number of species than those of the other locali-
ties. The three assemblages of coralligenous coming from
rocky cliffs (Punta del Faro, St. Stefano Shoal, Punta
Manara) cluster together, whereas the assemblage from
the flat bottom (Bogliasco) is significantly different. These
differences may be related to a higher sedimentation rate
at Punta Manara than in Punta del Faro and St. Stefano
Shoal, whereas the lower depth of the bioconstructions
from Bogliasco induces a proliferation of frondose algae
that compete with the sponges settled on the surface of
the conglomerate. Exactly the same kind of difference
arises from the comparison of ancient assemblages,
strongly suggesting that environmental factors have
remained stable for a very long time in different localities.
The fact that the spatial variability of sponge
communities is influenced by a combination of environ-
mental, biological and stochastic processes, resulting in
strong differences in abundance and diversity, is already
known (Pansini & Pronzato 1985; Pansini & Musso 1991;
Duckworth et al. 2008; Berman & Bell 2010).
The necessity of reconstructing past diversity to under-
stand ecosystem changes has been stressed by Boero &
Bonsdorff (2007). In the last few decades, Mediterranean
marine communities have shown significant changes in
Table 3. Ancient species determined from the disaggregated spicules
obtained from different blocks
Species Habit
Localities
SS BO PF PM
Order Homosclerophorida
Plakina dilopha Schulze, 1880 Ec X X
Plakina trilopha Schulze, 1880 Ec X
Plakortis simplex Schulze, 1880 Ec X
Order Spirophorida
Samus anonymus Gray, 1867 In X X
Order Astrophorida
Stelletta sp. In X
Jaspis johnstoni (Schmidt,1862) In X X X
Penares euastrum (Schmidt, 1868) In X X X
Erylus discophorus (Schmidt, 1862) In X X
Geodia cydonium Schmidt,1862 In X X
Pachastrella monilifera Schmidt, 1868 In X X
Poecillastra compressa
(Bowerbank, 1866)
In X
Poecillastra sp. In X
Dercitus (Stoeba) plicatus
(Schmidt, 1868)
In X X X X
Triptolemma simplex (Sar�a, 1959) In X X X X
Order Hadromerida
Cliona sp. Br X X X
Cliona janitrix Topsent, 1932 Br X X X X
Diplastrella bistellata (Schmidt, 1862) Ec X X X X
Spirastrella cunctatrix Schmidt, 1868 Ec X
Timea unistellata (Topsent, 1892) Ec X
Timea bifidostellata Pulitzer-Finali, 1983 Ec X
Timea sp. Ec X X
Spiroxya heteroclita Topsent, 1896 Br X X X
Spiroxya sp. Br X X X X
Order Poecilosclerida
Acarnus sp. Ms X X X
Eurypon sp. Ms X X
Rhabderemia sp. Ms X
Crella sp. Ms X X
Merlia sp. Ms X X
Order Agelasida
Agelas oroides Schmidt, 1864 Ms X X X
Order Haplosclerida
Petrosia (Petrosia) ficiformis
(Poiret, 1798)
Ms X
Ec = encrusting; In = insinuating; Br = boring; Ms = massive; SS = St.
Stefano Shoal; BO = Bogliasco; PF = Punta del Faro; PM = Punta
Manara.
8 Marine Ecology (2013) 1–10 ª 2013 Blackwell Verlag GmbH
Stability of Mediterranean coralligenous sponge assemblage Bertolino, Calcinai, Cattaneo-Vietti, Cerrano, Lafratta, Pansini, Pica & Bavestrello

composition and distribution. These modifications are
considered, directly or indirectly, to be related to the cur-
rent climatic changes, which may influence the spreading
capacity of some species, modifying their range (Astraldi
et al. 1995; Morri & Bianchi 2001; Bianchi & Morri 2004;
Cerrano & Bavestrello 2009). Moreover, a simplification of
the superficial benthic communities has occurred, mainly
in consequence of a sharp reduction of structuring organ-
isms (above all sponges, bryozoans and gorgonians) after a
number of mass mortality events that took place from 1999
to 2006 (Coma et al. 2009; Garrabou et al. 2009; Maldonado
et al. 2010; Cebrian et al. 2011; Di Camillo et al. 2012).
Sponge spicules represent a powerful tool for the tenta-
tive reconstruction of ancient assemblages both from a
quantitative (Bertolino et al. 2011) and qualitative (this
paper) point of view. Data from this study, obtained
from the most diversified assemblage of the Mediterra-
nean Sea, indicate that the diversity of coralligenous
sponges has remained stable over a millennial span of
time. This extended stability may be related to the pecu-
liarly protected inner habitat of the coralligenous crevices.
Conclusion
During the few last decades, Mediterranean marine com-
munities have shown significant changes in taxa composi-
tion and distribution. These modifications can be related,
directly or indirectly, to current climatic changes that, in
turn, have influenced the diffusive capacity of certain spe-
cies modifying their range. It is difficult to interpret the
changes, especially in the coastal environment, where dif-
ferent anthropic disturbances may merge with warming
effects, and where there is an almost complete absence of
historical quantitative data on the composition of benthic
communities. This problem was addressed through quan-
titative studies on the remains (e.g. mineral skeletal struc-
tures) that benthic organisms leave in the sediment after
death. Sponge spicules represent a good potential proxy
for the evaluation of long-term sponge community
dynamics. In this study the sponge spicules trapped inside
the crevices of coralligenous concretions collected in four
localities of the Ligurian Sea were extracted and studied.
In this way, the ancient sponge assemblage of the
coralligenous habitat was described over a millennial span
of time. This assemblage was composed of 30 recognisable
species representing almost half of the sponge community
present today on the same coralligenous concretions.
Acknowledgements
We would like to thank Dr Mario Mori for the helpful
suggestions. This study was conducted within the Ritmare
Project and Prin Cofin 2010–2011 Project.
References
Astraldi M., Bianchi C.N., Gasparini G.P., Morri C. (1995)
Climatic fluctuations, current variability and marine species
distribution: a case study in the Ligurian Sea (north-west
Mediterranean). Oceanologica Acta, 18, 139–149.
Ballesteros E. (2006) Mediterranean coralligenous assemblages:
a synthesis of present knowledge. Oceanography and Marine
Biology, 44, 123–195.
Barthel D. (1992) Do hexactinellids structure Antarctic sponge
associations? Ophelia, 36, 111–118.
Bavestrello G., Cattaneo-Vietti R., Cerrano C., Cerutti S., Sar�a
M. (1996) Contribution of sponge spicules to the
composition of biogenic silica in the Ligurian Sea. Marine
Ecology, 17, 41–50.
Becker L.C., Reaka-Kudla M.L. (1997) The use of tomography
in assessing bioerosion in corals. In: Lessios H.A. &
Macintyre I.G. (Eds), Proceedings of the 8th International
Coral Reef Symposium. Vol. 2. Smithsonian Tropical
Research Institute, Panama: 1819–1824.
Berman J., Bell J.J. (2010) Spatial variability of sponge
assemblages on the Wellington South Coast, New Zealand.
Open Marine Biology Journal, 4, 12–25.
Bertolino M. (2011) Sponges of the Coralligenous Community
in the Mediterranean Sea. PhD Thesis, Universit�a
Politecnica delle Marche. 1–162.
Bertolino M., Bavestrello G., Di Carlo M., Calcinai B. (2009)
Analisi della spongofauna del coralligeno ligure. Biologia
Marina Mediterranea, 16, 248–249.
Bertolino M., Calcinai B., Capellacci S., Cerrano C., Lafratta
A., Pansini M., Penna A., Bavestrello G. (2011) Posidonia
oceanica meadows as sponge spicule traps. Italian Journal of
Zoology, 79, 231–238.
Bianchi C.N., Morri C. (2004) Climate change and biological
response in Mediterranean Sea ecosystems – a need for broad-
scale and long-term research. Ocean Challenge, 13, 32–36.
Boero F., Bonsdorff E. (2007) A conceptual framework for
marine biodiversity and ecosystem functioning. Marine
Ecology, 28(Suppl. 1), 134–145.
Cattaneo-Vietti R., Bavestrello G., Cerrano C., Gaino E.,
Mazzella L., Pansini M., Sara M. (1999) The role of sponges
in the Terra Nova Bay ecosystem. In: Faranda F., Guglielmo
L., Ianora A. (Eds), Ross Sea Ecology Italiantartide
Expeditions (1987–1995). Springer, Berlin Heidelberg New
York: 539–549.
Cebrian E., Uriz M.J., Garrabou J., Ballesteros E. (2011)
Sponge mass mortalities in a warming Mediterranean Sea:
are cyanobacteria-harboring species worse off? PLoS ONE, 6,
e20211.
Cerrano C., Bavestrello G. (2009) Mass mortalities and
extinctions. In: Wahl M. (Ed.). Marine Hard Bottom
Communities. Ecological Studies. Springer, Berlin, 206: 295–
307.
Cerrano C., Bavestrello G., Bianchi C.N., Calcinai B.,
Cattaneo-Vietti R., Morri C., Sar�a M. (2001) The role of
Marine Ecology (2013) 1–10 ª 2013 Blackwell Verlag GmbH 9
Bertolino, Calcinai, Cattaneo-Vietti, Cerrano, Lafratta, Pansini, Pica & Bavestrello Stability of Mediterranean coralligenous sponge assemblage

sponge bioerosion in the Mediterranean coralligenous
accretion. In: Faranda F.M., Guglielmo L. & Spezie G.
(Eds). Mediterranean Ecosystems: Structures and Processes.
Springer-Verlag, Milan: 235–240.
Coma R., Ribes M., Serrano E., Jimeneza E., Salat J., Pascual J.
(2009) Global warming-enhanced stratification and mass
mortality events in the Mediterranean. Proceedings of the
National Academy of Sciences of the United States of America,
106, 6176–6181.
Davies P.J. (1983) Reef growth. In: Barnes D.J. (Ed.).
Perspectives on Coral Reefs. Australian Institute of Marine
Science, Townsville: 69–106.
Dayton P.K. (1979) Observations of growth, dispersal and
population dynamics of some sponges in McMurdo Sound,
Antarctica. In: L�evi C. & Boury-Esnault N. (Eds). Colloques
internationaux du C.N.R.S. 291. Biologie des spongiaires.
Editions du Centre National de la Recherche Scientifique,
Paris: 271–282.
Di Camillo C.G., Bartolucci I., Cerrano C., Bavestrello G.
(2012) Sponge disease in the Adriatic sea. Marine Ecology,
34, 62–71.
Duckworth A.R., Wolff C., Evans-Illidge E., Whalan S., Lui S.
(2008) Spatial variability in community structure of
Dictyoceratida sponges across Torres Strait, Australia.
Continental Shelf Research, 28, 2168–2173.
Epstein S.A., Friedman G.M. (1982) Processes controlling
precipitation of carbonate cement and dissolution of silica
in reef and near-reef settings. Sedimentary Geology, 33, 157–
171.
Garrabou J., Coma R., Bensoussan N., Bally M., Chevaldonn�e
P., Cigliano M., Diaz D., Harmelin J.G., Gambi M.C.,
Kersting D.K., Ledoux J.B., Lejeusne C., Linares C.,
Marschal C., Perez T., Ribes M., Romano J.C., Serrano E.,
Teixido N., Torrents O., Zabala M., Zuberer F., Cerrano C.
(2009) Mass mortality in Northwestern Mediterranean rocky
benthic communities: effects of the 2003 heat wave. Global
change biology, 15, 1090–1103.
Gerino M. (1990) The effects of bioturbation on particle
redistribution in Mediterranean coastal sediment.
Preliminary results. Hydrobiologia, 207, 251–258.
Gutt J. (2007) Antarctic macrozoobenthic communities: a
review and an ecological classification. Antarctic Science, 19,
165–182.
Hammer Ø., Harper D.A.T., Ryan P.D. (2001) PAST:
paleontological statistics software package for education and
data analysis. Palaeontologia Electronica, 4, 1–9. http://
palaeo-electronica.org/2001_2001/past/issue2001_2001.htm.
Koltun V.M. (1968) Spicules of sponges as an element of the
bottom sediments of the Antarctic. In: Scar, Scor, Iapo &
Iubs (Ed.). Scar Symposium on Antarctic Ocean. Scott Polar
Research Institute, Cambridge, 268: 121–123.
Maldonado M., Sanchez-Tocino L., Navarro C. (2010)
Recurrent disease outbreaks in corneous demosponges of
the genus Ircinia: epidemic incidence and defense
mechanisms. Marine Biology, 157, 1577–1590.
Morri C., Bianchi C.N. (2001) Recent changes in biodiversity
in the Ligurian Sea (NW Mediterranean): is there a climatic
forcing? In: Faranda F.M., Guglielmo L. & Spezie G. (Eds).
Structure and Processes in the Mediterranean Ecosystems.
Springer Verlag, Milan: 375–384.
Paduano G.M., Fell P.E. (1997) Spatial and temporal
distribution of freshwater sponges in Connecticut lakes
based upon analysis of siliceous spicules in dated sediment
cores. Hydrobiologia, 350, 105–121.
Pansini M., Musso B. (1991) Sponges from Trawl-Exploitable
Bottoms of Ligurian and Tyrrhenian seas: distribution and
ecology. Marine Ecology, 12, 317–329.
Pansini M., Pronzato R. (1985) Distribution and ecology of
epiphytic Porifera in two Posidonia oceanica (L.) Delile
meadows of the Ligurian and Tyrrhenian Sea. Marine
Ecology, 6, 1–11.
R€utzler K., Macintyre I.G. (1978) Siliceous sponge spicules in
coral reef sediments. Marine Biology, 49, 147–159.
Sartoretto S. (1996) Vitesse de croissance et bio�erosion des
concr�ectionnements ‘corallig�ene’ de M�editerran�ee nord-
occidentale. Rapport avec les variations Holoc�enes du
niveau marin. Th�ese Doctorat d’Ecologie, Universit�e d’
Aix-Marseille, II: 194 pp.
Sartoretto S., Verlaque M., Laborel J. (1996) Age of
settlement and accumulation rate of submarine ‘corallig�ene’
(–10 to –60 m) of the northwestern Mediterranean Sea;
relation to Holocene rise in sea level. Marine Geology, 130,
317–331.
Sorokin Y.I. (1993) Coral Reef Ecology. Springer, Berlin.
Volkmer-Ribeiro C., De Drago E., Parolin M. (2007) Spicules
of the freshwater sponge Ephydatia facunda indicate
lagoonal paleoenvironment at the Pampas of Buenos Aires
Province, Argentina. (Proceedings of the 9th International
Coastal Symposium). Journal of Coastal Research, SI 50,
449–452.
10 Marine Ecology (2013) 1–10 ª 2013 Blackwell Verlag GmbH
Stability of Mediterranean coralligenous sponge assemblage Bertolino, Calcinai, Cattaneo-Vietti, Cerrano, Lafratta, Pansini, Pica & Bavestrello












![Descriptors for Sponge Gourd [Luffa cylindrica (L.) Roem.]](https://static.fdokumen.com/doc/165x107/63187e763394f2252e02b92e/descriptors-for-sponge-gourd-luffa-cylindrica-l-roem.jpg)







