
SPONGE RESEARCH DEVELOPMENTS Mangrove and coral reef sponge faunas: untold stories about shallow...
14
1 23 ! "# $%&%'(
-
Upload
independent -
Category
Documents
-
view
4 -
download
0
Transcript of SPONGE RESEARCH DEVELOPMENTS Mangrove and coral reef sponge faunas: untold stories about shallow...

1 23

1 23
Your article is protected by copyright andall rights are held exclusively by SpringerScience+Business Media B.V.. This e-offprintis for personal use only and shall not be self-archived in electronic repositories. If youwish to self-archive your work, please use theaccepted author’s version for posting to yourown website or your institution’s repository.You may further deposit the accepted author’sversion on a funder’s repository at a funder’srequest, provided it is not made publiclyavailable until 12 months after publication.

SPONGE RESEARCH DEVELOPMENTS Review paper
Mangrove and coral reef sponge faunas: untold storiesabout shallow water Porifera in the Caribbean
M. Cristina Diaz
Received: 23 February 2011 / Accepted: 14 November 2011! Springer Science+Business Media B.V. 2011
Abstract Sponge faunas from coral reefs and man-
grove ecosystems in the Caribbean have mostly beenstudied from an ecological perspective, with research-
ers considering the effects of physical and biological
factors on their species distribution. To discernevolutionary patterns, this study analyzed the system-
atic composition, taxonomic diversity, and ecologicalproperties (reproductive strategies, size, shape, endo-
symbiosis) of mangrove and reef sponge assemblages
from seven distant Caribbean localities. Speciescomposition was compared by use of cluster analysis
(Sørensen’s), and taxonomic diversity by use of the
biodiversity index average taxonomic distinctness(AvTD). Mangrove and reef-associated sponge faunas
were found to be statistically dissimilar, with the
AvTD values suggesting stronger taxonomic biastoward specific groups in mangroves, irrespective of
geographic distance. Most Demospongiae orders have
30–50%more species in coral reefs than inmangroves.The richest reef genera (Agelas, Aplysina, Callyspon-gia, Petrosia, and Xestospongia) rarely colonize
contiguous mangrove formations. The distribution
and diversity of suprageneric taxa suggest that coral
reef sponge assemblages might represent an olderfauna. This historical interpretation would place man-
grove subtidal habitats as the youngest marine ecosys-
tem, rather than a below-optimum ecosystem. Lifehistory traits support a biological split discussed here
from the perspective of distinct evolutionary historiesand different environmental conditions.
Keywords Mangroves ! Coral reefs ! Sponges !Evolution ! Animal history
Introduction
Coral reef and mangrove ecosystems are among the
most important tropical marine communities. Coral
reefs harbor 4–5% of all known species, and areresponsible for the highest recorded oceanic produc-
tivity (1,500–5,000 gC/m2/year). Mangrove forests
line up to 60–75% of tropical coasts; they constitute‘‘diversity hotspots’’ (Rutzler et al., 2000) and have
been demonstrated to increase reef fish productivity
(Mumby et al., 2007).Sponges may be among the most diverse compo-
nents of benthic fauna in shallow marine Caribbean
systems. Reef sponges may quadruplicate the diversityof hermatypic and soft corals (Dıaz & Rutzler, 2001).
On the other hand, in mangrove islands (leeward side)
at a variety of Caribbean sites, sponges equal orsurpass the richest groups of macroalgae or ascidians,
Guest editors: M. Maldonado, X. Turon, M. A. Becerro &M. J. Uriz / Ancient animals, new challenges: developmentsin sponge research
M. C. Diaz (&)Museo Marino de Margarita, Boca del Rio,Nueva Esparta, Venezuelae-mail: [email protected]
123
Hydrobiologia
DOI 10.1007/s10750-011-0952-5
Author's personal copy

representing from 10 to 70% of the total root epiphyticdiversity (Dıaz & Rutzler, 2009). Furthermore,
sponges share first or second place of dominance in
terms of root and peat area coverage at a variety ofCaribbean mangrove sites (Perez-Vazquez, 2007;
Dıaz & Rutzler, 2009; Guerra-Castro et al., 2011).
Marine sponges are essential to the ecology of theseecosystems, partly because of their unique capacity for
water filtration but also because of the transformation
of nutrients through their metabolic processes andthose of their microbial associates (Dıaz & Rutzler,
2001; Lesser, 2006; de Goeij et al., 2008).
Mangrove and coral reef ecosystems are geograph-ically, and ecologically interconnected throughout the
Caribbean (Rutzler et al., 2000; Dıaz, 2005; Mumby
et al., 2007). Comparative studies between these twofaunas suggest mangrove sponge distributions are
predominantly controlled by abiotic factors (Bingham
& Young, 1995; Farnsworth & Ellison, 1996; Pawliket al., 2007), with intense competition for space
leading to the evolution of high growth capability
(Wulff, 2005, 2009) and chemical defense mecha-nisms (Engel & Pawlik, 2005a). Reef faunas are
believed to be mostly controlled by biotic factors,
mainly fish predation and space competition with thedevelopment of allelopathic defense mechanisms
(Dunlap & Pawlik, 1996, 1998; Pawlik, 1997, 1998;
Wulff, 1997, 2005; Engel & Pawlik, 2005b). Recentstudies suggest that biological features such as growth
rates, recruitment patterns, asexual reproduction, and
root–sponge interactions may vary between spongespecies, and determine the presence, abundance, and
dynamics of sponge populations on mangrove roots
(Farnsworth & Ellison, 1996; Dıaz & Rutzler, 2009;Hunting et al., 2010). However, this analysis of
mangrove and coral reef sponge faunas lacks compre-
hensive systematic comparison. Several questionsremain unanswered, including:
– how similar are these faunas taxonomically?
– are there other ecological strategies besidesgrowth rate, secondary chemistry, antipredatory
defenses, and competitive growth capabilities thatdiverge between these faunas?
This paper reviews and analyzes distributional and
ecological data on the sponges of Caribbean man-groves and coral reefs with the objective of under-
standing novel aspects of the ecological and
evolutionary relationships among these faunas. Five
aspects of Caribbean sponge biology and ecology arediscussed:
1. sponge species distributionswithin each ecosystem;
2. taxonomic composition;3. reproductive strategies—viviparity versus oviparity;
4. morphology (size and shape); and
5. distribution of sponge-cyanobacteria symbioses.
Materials and methods
Taxonomic composition of sponge assemblages
of Caribbean mangroves and coral reefs
Faunal surveys, taxonomic revisions, gray literature,
and two unpublished surveys by the author andanother colleague (Dr Pedro Alcolado, Cuba) recently
conducted in either mangrove or coral reef habitatswere reviewed to gather data on the distribution of
species for each habitat. Validity of species names and
their classification were checked against the WorldPorifera Database (Van Soest et al., 2010). The
information was gathered into a species-by-country-
by-habitat (either mangrove and/or coral reef) matrixwith values of presence (1) or absence (0) for each
species. Caribbean sites included seven countries:
Belize (Rutzler et al., 2000; Dıaz & Rutzler, 2009;Rutzler unpublished data), Cuba (Alcolado, 2002,
unpublished data), Curacao and Bonaire (Van Soest,
1978, 1980, 1984, 2009), Jamaica (Hechtel, 1965;Lehnert & van Soest, 1996, 1998), Little Cayman
(Diaz, unpublished data), Panama (Clifton et al., 1997;
Dıaz, 2005), and Venezuela (Alvarez & Dıaz, 1985;Pauls, 2003; Amaro, 2005; Alvizu, 2006; Perez-
Vazquez, 2007; Ramirez, 2010; Diaz et al. unpub-
lished data). Total numbers of species were used tointerpret the distribution of sponge taxa (genera,
families, and orders) among mangrove and open reef
environments. Data from cryptic coral reef studiesfrom Curacao and Bonaire (Kobluk & Van Soest,
1989; Van Soest, 2009) were included as a compar-
ative habitat reference. The species matrix wasanalyzed using Sørensen’s similarity index and then
projected using MDS ordination to identify similar
faunas within and among habitats (Clarke, 1993). Thenull hypothesis of no relationship within and among
habitats was tested by analysis of similarities
(ANOSIM) using all possible permutations (n = 464).
Hydrobiologia
123
Author's personal copy

The biodiversity index average taxonomic distinctness(AvTD) and its associated statistical test (taxonomic
distinctness test or TAXDTEST) were used to evaluate
taxonomic diversity for each habitat by locality(Clarke & Warwick, 1998; Warwick & Clarke,
2001). AvTD is a measure of the average extent to
which individuals in an assemblage are related to eachother, on the basis of a taxonomic hierarchy. This
index was used to test the null hypothesis that the
AvTD for a particular habitat in a locality (mangrove,open or cryptic coral reef) is within the expected range
considering the local species richness and the regional
pool of species. This test was carried out using 1,000simulations of AvTD values calculated from random
subsets of species from the regional pool. Comparison
of the AvTD for each habitat within each countryenables comparison of the evolutionary relatedness
of species within each habitat. All statistical anal-
yses were conducted using Primer v6 (Primer-E,Plymouth, UK) (Clarke & Gorley, 2006).
Species richness was compared at suprageneric
levels to evaluate possible predominance of genera,families, or orders in mangroves or coral reefs at
different Caribbean sites.
Ecological patterns
The incidence of specific biological traits, for examplereproductive strategies (viviparous vs. oviparous lar-
vae), and ecological patterns, for example the inci-
dence of cyanobacteria–sponge associations, andpredominance of shapes or size ranges, were reviewed
using published data and observations by the author.
Results
Taxonomic composition of Caribbean mangrove
and coral reef sponge faunas
A list of 414 species was gathered in a taxonomic
matrix of species presence in (1) or absence from (0)mangrove and open reef habitats in six countries, and
cryptic reef habitats in two countries. The list can be
found at the World Porifera Database web site (http://www.marinespecies.org/porifera/porifera.php?p=
sourcedetails&id=149269) and at the Porifera Tree
of Life (PorToL) website (http://www.portol.org/resources). A complementary taxonomic list that
contains classification and authorship information forall Caribbean species can be found at the World
Porifera Database site (http://www.marinespecies.org/
porifera/porifera.php?p=sourcedetails&id=149270)and at the PorToL site.
Sørensen similarity analysis (Fig. 1) showed that
mangrove and coral reef faunas from distant Carib-bean regions have significant dissimilarity in species
composition (ANOSIM test, R = 0.685, P\ 0.05).
All mangrove sites studied share more speciesbetween them than with nearby reefs (i.e., the
mangroves in Panama share the highest similarity
with faunas from mangroves sites throughout theCaribbean than with Panama’s reef sponge fauna).
Two groups are distinguished: one from the man-
groves, with the highest similarities in species com-position comprising Belize, Cuba, Panama and
Venezuela, and another from the reef composed of
all of the studied coral reef sites and maintaineddespite the disparity of their species richness values
(80–236 species).
All of the reef sites have taxonomic diversity thatfalls within the expected range of values considering
the reported species richness (Fig. 2). However, the
richest mangrove sites (i.e. Panama, Venezuela, andBelize) had AvTD values that were lower than
expected (95% confidence test). These results suggest
that the predominance of specific sponge taxa (i.e.Chalinidae andMycalidae) in mangrove systems is not
a random pattern but that there is a taxonomic bias
toward specific groups.
Suprageneric species distributions
among Caribbean mangrove and coral reef spongefauna
The habitat distributions of the species listed in thedata matrix shows that 320 species have been reported
from open reef habitats (61% not shared with the
mangroves) and while 145 species have been reportedfrom mangrove habitats (24% of species not shared
with the reefs). Therefore the reef sponge fauna istwice as rich as that found in the mangrove habitats.
The number of species within each of 13 orders of
Demospongiae orders and two orders of Calcarea isshown in Fig. 3. Species richness within each order
ranges from 1 to 65, but is consistently 30–50% higher
in open coral reefs than in mangroves for almost allorders considered.
Hydrobiologia
123
Author's personal copy

The species richness within the most abundant
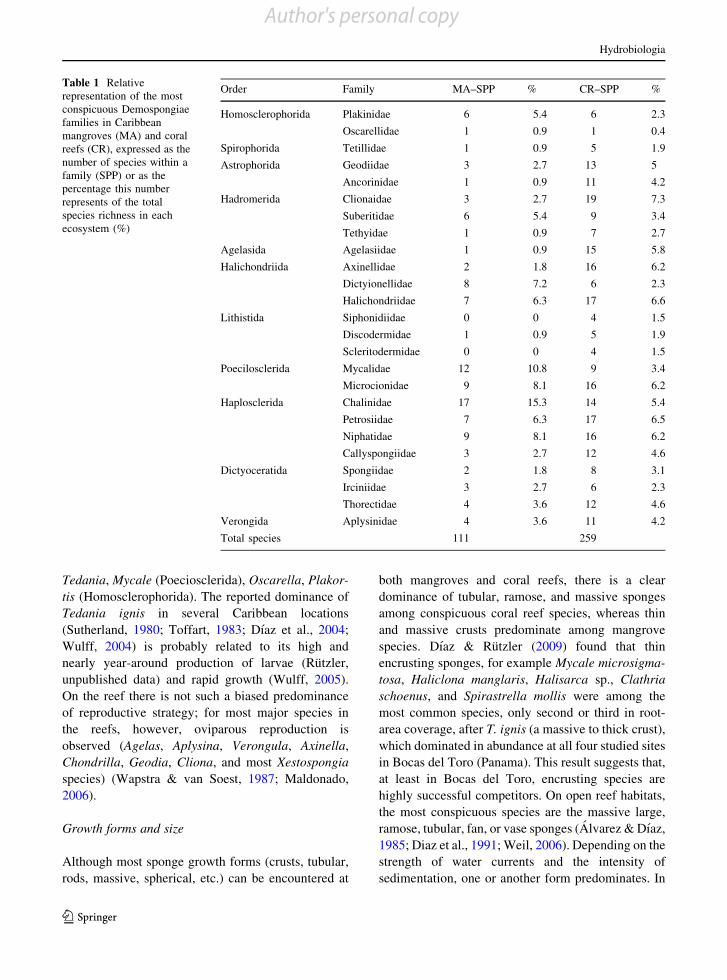
Demospongiae families reported fromCaribbean coralreefs and mangrove habitats is summarized in Table 1.
Most families among Demospongiae orders are richer
in reef habitats than in mangrove habitats; within some
orders, however, there is a clear distinction of which
families diversify within each habitat. For instance, thefamilies Mycalidae (Order Poecilosclerida), Chalinidae
(Order Haplosclerida), and Dictyonellidae (Order
Halichondriidae) are more diverse in the mangroves
Fig. 1 Non-metric multidimensional scaling (nMDS) ordina-tions of Sørensen’s similarities of sponge species compositionamong countries by habitat (species richness is indicated inparentheses). Circles represent data for mangroves (gray), opencoral reefs (black), and cryptic reef habitats in Curacao and
Bonaire (white). Included countries are Panama (P), Jamaica(J), Cuba (C), Venezuela (V), Belize (Twin Cays: B–TC, PelicanCays: B–PC), Little Cayman reefs (LC), and Curacao andBonaire (C–B). The habitats were significantly dissimilar(ANOSIM test, R = 0.685, P\ 0.05)
100
BC CJ
95
J
J
B-LC
P
P
B-PCV
V B-C
Del
ta+ P
B-TC
90
0030020010Number of species
85
Fig. 2 Funnel plot forAvTD at each habitat bycountry. Gray circlescorrespond to reef habitats,black circles to mangrovehabitats, and the open circleto cryptic habitats in Bonaireand Curacao. Funnelsrepresent the simulated 95%confidence intervals for theexpected AvTD values(TAXDTEST)
Hydrobiologia
123
Author's personal copy

than in the reefs, and their relative importance becomes
evident when we analyze their species richness as a
percentage of the total richness reported for mangroves(Table 1). Some families and genera that are emblem-
atic of coral reefs (Aplysinidae, Agelasiidae, Axinelli-
dae, Ancorinidae) are almost absent from mangroveecosystems. The genera of the various lithistid families
have never been observed in mangrove ecosystems.
Within the species-rich order Haplosclerida we findfamilies with many more species on the reefs, for
example the Petrosiidae, Callyspongiidae, and Niphat-
idae, and families radiating in the mangroves, forexample the Chalinidae. This familial bias is repeated
within the order Halichondrida, with the families
Axinellidae and Halichondriidae radiating in the reefs,and the family Dictyonellidae radiating in the man-
groves. If we express the species richness within each
family as a percentage of the total diversity at eachhabitat we find a different picture. For the higher
species numbers found on coral reefs among families
such as Petrosiidae, Niphatidae, and Halichondriidae
the percentage of the total species richness is very similar
to that for both mangroves and reefs. Therefore, those
families, despite having a large number of species onreefs, represent a similar proportion of species richness
in both ecosystems. On the other hand, the taxonomic
bias observed in mangroves for families such asChalinidae, Dictyonellidae, Mycalidae, and Microcion-
idae becomes more evident because their species
richness is a very high portion of the total diversity. Incontrast, the families Agelasiidae, Axinellidae, and
Clionaidae include most of the species on reefs.
Ecological patterns
Reproductive strategies
Most of the dominant demosponge genera (represent-
ing at least six demosponge orders) in Caribbeanmangroves have viviparous larvae. These include
Halichondria, Hymeniacidon, Scopalina (Halichond-
rida), Haliclona, Chalinula (Haplosclerida), Clathria,
Fig. 3 Number of species for 13 Demospongiae and 2 Calcarea orders, for each habitat: Black bars represent coral reef species andgray bars represent mangrove species. The numbers above the bars represent species not shared between habitats
Hydrobiologia
123
Author's personal copy
Tedania, Mycale (Poeciosclerida), Oscarella, Plakor-tis (Homosclerophorida). The reported dominance ofTedania ignis in several Caribbean locations
(Sutherland, 1980; Toffart, 1983; Dıaz et al., 2004;
Wulff, 2004) is probably related to its high andnearly year-around production of larvae (Rutzler,
unpublished data) and rapid growth (Wulff, 2005).
On the reef there is not such a biased predominanceof reproductive strategy; for most major species in
the reefs, however, oviparous reproduction is
observed (Agelas, Aplysina, Verongula, Axinella,Chondrilla, Geodia, Cliona, and most Xestospongiaspecies) (Wapstra & van Soest, 1987; Maldonado,
2006).
Growth forms and size
Although most sponge growth forms (crusts, tubular,
rods, massive, spherical, etc.) can be encountered at
both mangroves and coral reefs, there is a clear
dominance of tubular, ramose, and massive spongesamong conspicuous coral reef species, whereas thin
and massive crusts predominate among mangrove
species. Dıaz & Rutzler (2009) found that thinencrusting sponges, for example Mycale microsigma-tosa, Haliclona manglaris, Halisarca sp., Clathriaschoenus, and Spirastrella mollis were among themost common species, only second or third in root-
area coverage, after T. ignis (a massive to thick crust),
which dominated in abundance at all four studied sitesin Bocas del Toro (Panama). This result suggests that,
at least in Bocas del Toro, encrusting species are
highly successful competitors. On open reef habitats,the most conspicuous species are the massive large,
ramose, tubular, fan, or vase sponges (Alvarez &Dıaz,
1985; Diaz et al., 1991;Weil, 2006). Depending on thestrength of water currents and the intensity of
sedimentation, one or another form predominates. In
Table 1 Relativerepresentation of the mostconspicuous Demospongiaefamilies in Caribbeanmangroves (MA) and coralreefs (CR), expressed as thenumber of species within afamily (SPP) or as thepercentage this numberrepresents of the totalspecies richness in eachecosystem (%)
Order Family MA–SPP % CR–SPP %
Homosclerophorida Plakinidae 6 5.4 6 2.3
Oscarellidae 1 0.9 1 0.4
Spirophorida Tetillidae 1 0.9 5 1.9
Astrophorida Geodiidae 3 2.7 13 5
Ancorinidae 1 0.9 11 4.2
Hadromerida Clionaidae 3 2.7 19 7.3
Suberitidae 6 5.4 9 3.4
Tethyidae 1 0.9 7 2.7
Agelasida Agelasiidae 1 0.9 15 5.8
Halichondriida Axinellidae 2 1.8 16 6.2
Dictyionellidae 8 7.2 6 2.3
Halichondriidae 7 6.3 17 6.6
Lithistida Siphonidiidae 0 0 4 1.5
Discodermidae 1 0.9 5 1.9
Scleritodermidae 0 0 4 1.5
Poecilosclerida Mycalidae 12 10.8 9 3.4
Microcionidae 9 8.1 16 6.2
Haplosclerida Chalinidae 17 15.3 14 5.4
Petrosiidae 7 6.3 17 6.5
Niphatidae 9 8.1 16 6.2
Callyspongiidae 3 2.7 12 4.6
Dictyoceratida Spongiidae 2 1.8 8 3.1
Irciniidae 3 2.7 6 2.3
Thorectidae 4 3.6 12 4.6
Verongida Aplysinidae 4 3.6 11 4.2
Total species 111 259
Hydrobiologia
123
Author's personal copy

the Barrier reef at Belize,where the current is strong andconstant, tube and vase shaped sponges dominate
(Villamizar and Diaz, unpublished data). In reef walls
the stringy or tubular sponges are the most conspicuous.In cryptic coral reef environments, most species are thin
to thick encrusting sponges, or are very small tubular
ones, for example Calcareous species (Kobluk & VanSoest, 1989; Diaz and Rutzler, personal observation).
On mangrove roots, massive growth forms are
common among the halichondrids (Halichondria,Hymeniacidon, and Amorphinopsis), poecilosclerids
(Tedania spp., Clathria spp., Mycale spp.), and dictyo-
ceratids (Spongia, and Hyrtios spp.) (Rutzler et al.,2000; Dıaz & Rutzler, 2009). One might find rods,
branches, and tubes, but usually in smaller sizes than in
reefs. Size rather than shape might be a clear differencebetween these two faunas. Whereas mangrove sponges
might reach sizes up to 10,000s of cm3 (Dıaz &Rutzler,
2009), reef sponges reach sizes of up to 1,000,000s ofcm3, for example the massive Xestospongia muta and
Geodia neptuni, with very high living volumes (Diaz
and Villamizar unpublished data).
Distribution of cyanobacterial sponge associations
Coral reef open habitats, exclusively, contain 80% of
the Caribbean cyanosponges (Dıaz et al., 2007). All
Aplysina, Chondrilla, Erylus, Geodia, Ircinia, Neo-petrosia, Petrosia, Spongia, Svenzea, Verongula,Xestospongia species, and Haliclona walentina are
among the most conspicuous reef species. Only elevenspecies in Caribbean mangroves have been reported
with endobiotic cyanobacteria: Geodia papyracea,G. tumulosa, G. gibberosa, Xestospongia wiedenmay-eri, X. bocatorensis, Spongia pertusa, S. tubulifera,Hyrtios proteus, Chondrilla caribbaensis f. cariben-sis, Ircinia sp., and Haliclona sp.
Discussion
The taxonomic and ecological divergence of spongefauna inhabiting mangrove and open coral reef
habitats is well supported by the current literature.
First, statistically significant dissimilarities of spongespecies composition between mangrove and open
coral reef habitats were observed at a variety of
Caribbean sites. The AvTD analysis further supportsthis distinction and indicates the existence of a strong
taxonomic bias in the sponge fauna of mangrovescompared with coral reefs, which suggests that a
speciation process is associated with the mangrove
habitat itself, operating independently of geographicdistance. Second, throughout the comparison of eco-
logical patterns of these faunas (reproductive strate-
gies, growth forms, cyanobacterial symbioses) cleardivergences are observed. Third, the reef fauna seems
to be more diverse throughout most orders of
Demospongiae, and consistent disjunctive radiationof families and genera is observed, at least in the five
well studied Caribbean regions where these two
ecosystems are contiguous (Belize, Cuba, Jamaica,Panama, and Venezuela). The possible effect of
historical and environmental differences between
mangrove and coral reefs are discussed below aspotential causes of the taxonomic and ecological
distinctiveness of their associated sponge fauna.
Distinct evolutionary histories
The first aspect to consider is that despite thegeographic co-occurrence of these two ecosystems,
it is highly probable that their ‘‘founding taxa’’ and the
ecosystems themselves have very different evolution-ary histories (geological periods in which each eco-
system and its major biological components evolved,
evolutionary age of the lineages of sponge taxa thatflourish at each type of ecosystem, etc.). Modern
hermatypic coral reef ecosystems evolved since the
beginning of the Cenozoic (preceded by the evolutionof photobiosis in scleractinean corals 210 mya and the
later evolution of hervibory and predation 60 mya)
(Wood, 1998). On the other hand, the oldest mangroverecord is of the genus Nypa 80 mya from the Indo-
Pacific (Ellison et al., 1999) and the contemporary
mangrove ecosystems in the Caribbean evolved 5 mya(Lacerda et al., 2002). Therefore, the evolution of
mangroves as marine-terrestrial ecosystems is more
recent than the development and evolution of coralreef ecosystems in the Caribbean basin. Furthermore,
mangrove development around reef ecosystems ispreceded by the growth of a coral reef structure, with
subsequent geo-morphological processes leading to
the environmental conditions (depth and sedimentdeposition) necessary for mangrove colonization.
These geological and geo-morphological timing dif-
ferences support the hypothesis that lineages of coralreef taxa evolved earlier than mangrove taxa, and that
Hydrobiologia
123
Author's personal copy

these taxa are highly derived and adapted to a coralreef environment. This could in turn explain the lack
of successful colonization of neighboring mangrove
systems by major coral reef species (for exampleXestospongia, Agelas, Aplysina, and Axinellid).Species of these common genera in the Caribbean
probably evolved adaptations to exploit, survive in,and colonize coral reef habitats, when mangrove
systems were not the conspicuous marine subtidal
community they are today. The consistently higherdiversity of sponge species, across most sponge
orders, in open reef habitats as opposed to open
mangrove habitats, supports the hypothesis of an oldersponge fauna on the reef. The mangrove sponge fauna
is probably younger and secondarily derived from
previously existing faunas (open and cryptic coral reefhabitats, deep waters, and rocky shores). The work
discussed in this paper revealed significant dissimi-
larities among the prevailing forms observed inmangroves and those from open habitats in coral
reefs. Furthermore, the sponge assemblages associated
with cryptic habitats in Curacao and Bonaire revealedhigher similarity with open reef fauna. However,
several genera which dominate in mangroves, for
example Haliclona, Clathria, and Mycale, are com-mon in reef sciophilous habitats (Kobluk &Van Soest,
1989), and some ecological patterns, for example the
predominance of viviparity, are also common amongcryptic species. Therefore, some of the genera that
successfully colonized mangroves might have been
derived from cryptic or sciophilous habitats.This scenario of the history of these marine
ecosystems would place mangrove subtidal habitats
as the youngest marine ecosystem, rather than a belowoptimum marine ecosystem. Wulff (2005, 2009)
transplanted reef and mangrove sponges on to each
other’s habitats in Belize, and found that some reefspecies thrive when placed on mangrove roots. Pawlik
et al. (2007) repeated the same experiment but in the
Florida Keys and obtained very different results, withall species dying after transplantation. It seems that a
range of effects of abiotic and biotic factors maydifferently affect species survival in the Caribbean.
However, it is probable that constraints in the early
stages of development (recruitment and colonizationmechanisms), and not adult survival, prevent these
emblematic reef species from growing in most man-
grove sites. A higher proportion of reef fauna onmangrove roots has been found in some mangrove
areas adjacent to shallow reef patches (Rutzler et al.,2000). Recruitment facilitation, spongivory pressure,
and the existence of unique environmental conditions
(clear and calm waters) have been suggested asexplanations of this phenomenon (Rutzler et al.,
2000; Wulff, 2005).
Evolutionary relationships of particular taxonomicgroups within mangrove ecosystems have been previ-
ously studied for other animal phyla. Ellison et al.
(1999) depicted evolutionary relationships between avariety of gastropod groups and mangroves. When
they compiled a fossil-occurrence database of gastro-
pods and compared the distribution with that in theirfossil mangrove database they encountered an eco-
logical ‘‘nestedness’’ of gastropods and mangroves
through time. Ellison et al. (1999) concluded, on thebasis of the fossil distributions of both mangroves and
gastropods, and the patterns of their species–area
relationships, that these associations most likely sharea Tethyan origin, with subsequent diversification
patterns related to the tectonic reconfiguration of
shorelines and continents.Currently, we do not have a good sponge fossil
record associated with mangrove ecosystems; however,
one can trace phylogenetic relationships using variousmolecular markers to determine historical relationships
between both faunas across the Caribbean. Duran &
Rutzler (2006) encountered ‘‘nested’’ populations ofChondrilla species (distinct phylotypes), either on reefsor on mangroves, all along the Caribbean. Although the
results of their analysis suggested an ancestral origin ofChondrilla haplotypes from the mangroves, and a
derived origin for the reef populations, their work is the
first proof of the existence of distinct populations andlow gene flow between the two habitats despite
geographic continuity.
Other biological scenarios could explain the taxo-nomic distinctness and higher diversity of open coral
reef sponge taxa compared with mangrove sponge
taxa. A more heterogeneous marine system, forexample a coral reef with more complex biological
interactions, could promote a richer and distinct fauna,which argues against the historical distinctness pro-
posed earlier. Higher species richness can also be
related to the fact that coral reefs extend for severaltens of meters in depth whereas mangroves occur only
between 1 and 3 m deep. Only life-history studies
demonstrating the existence of complex behavior inearly life stages, different success in each habitat, and
Hydrobiologia
123
Author's personal copy

the genetic historical association of specific groupswill clarify these hypotheses.
Environmental conditions
Various environmental differences must be discussed
as possible contributors to the taxonomic and ecolog-ical patterns presented here. A variety of environmen-
tal conditions in open and cryptic coral reef and
mangrove habitats are characterized qualitatively inTable 2. Marked differences become evident when
one compares general qualitative aspects among them.
Abiotic factors are less likely to differently structurethe reef sponge community because coral reef habitats
have relatively constant, oligotrophic conditions
(Pawlik et al., 2007). It is well known that few spongespecies can overcome air exposure and high temper-
atures (Rutzler, 1995). Daily temperature, tidal, and
salinity variation found in mangrove systems mustrestrict the long term growth and survival of coral reef
sponges that inhabit much more stable environments.
The possible effect of changes in substrate availabilityon the dynamics of its associated benthic fauna has not
yet been evaluated. Major coral reef ‘‘structure
builders’’, hermatypic corals, grow very slowly,1–2 cm per year, whereas mangrove roots grow at
between 10 and 40 cm per year, adding continuously
new substrate to the sub-tidal environment (Dıaz &Rutzler, 2009). These large differences in substrate
dynamics must have important ecological conse-
quences for the fauna inhabiting them. Strategies suchas high growth rates, asexual reproduction, and the
availability of viviparous recruitment stages would befavored in systems such as mangroves, where imme-
diate colonization could be advantageous given the
stochastic appearance of available substrate.Oviparity and broadcasting strategies in sponges
might be favored under conditions of strong unidirec-
tional currents, with wider geographic distributionscommon for organisms inhabiting coral reefs. Short-
ened viviparous larval stages might be favored in
slower current environments and in habitats withmuch restricted geographic distributions.
Cryptic reef species and several mangrove forms
minimize volume to area coverage ratios. Limitedspace between roots and under reef crevices would
favor smaller sizes in both environments. Open reef
species tend to maximize volume-to-area ratios withbodies expanding in the water column, rather than on
the substrate. Associations with symbiotic organisms
are strikingly excluded from cryptic and mangrovespecies, where light conditions are less favorable for
photosynthetic organisms.
The high rate of stochastic and predictable events inmangrove ecosystems (tides temperature change,
drastic substrate change, upwelling, etc.), has probably
caused a myriad of variable conditions that haveprovided an opportunity for the genera that reached
this environment to evolve distinct micro niches at
these sites. The availability of these niches mightexplain the sympatric occurrence of so many unique
species within several genera (for example Haliclona,Mycale), where one can find two or three species of thesame genus co-occurring on a single mangrove root.
Table 2 Generalcharacterization ofenvironmental factors atCaribbean mangrove, andopen and cryptic reefhabitats
Condition Mangrove Open reef Cryptic reef
Temperature daily/seasonalchanges
High No No
Salinity daily/seasonal changes High No No
Sedimentation Low-high Low-high Low
Water circulation Poor Mid to high Poor
Tides Diurnal No No
Wave action Very low High Low
Solar radiation Low High Low
Nutrient levels High Low Low
Substrate type Living wood, peat,mud-sand
Sand, coralrock
Sand, coralrock
Substrate availability Very low Low Very low
Rate of substrate growth 10–40 cm/year 1–2 cm year NA
Predation Low High Low
Hydrobiologia
123
Author's personal copy

Conclusions
Sponge faunas from mangrove and open coral reefhabitats have high distinctness at different taxonomic
levels, in a variety of Caribbean sites, suggesting a
different founding fauna in both marine ecosystems.Distribution and diversity of suprageneric taxa
suggests that the coral reef sponge fauna might be
older, and that the mangrove sponge fauna might beyounger and derived from coral reef cryptic, deep
water, and rocky shores habitats.
The historical aspects of the evolution of coral reefand mangrove fauna must be further explored (e.g.
genetically) to evaluate the hypothetical explanations
proposed here for the strong taxonomic disjunctionbetween these to co-occurring faunas.
Mangroves might be the latest evolutionary frontier,
because they are among the youngest marine ecosystems.The effect of various environmental conditions
(temperature and salinity changes, tidal currents,
substrate affinities) at various life stages of thesesponge species must be studied to evaluate the
contribution of environmental differences to the
ecological distinctness of these two faunas.
Acknowledgments I would like to thank Lic. Paula Young,and Lic. Adriana Perez for their help gathering speciesdistribution data, Dr Pedro Alcolado and Dr Klaus Rutzler forproviding unpublished species lists, Lic. Edlin Guerra forprocessing distribution data and performing the similarity andAvTD analyses, Dr Andrzej Pisera, Dr Klaus Rutzler, Dr RobertThacker, and Dr Rob van Soest for their comments, suggestions,and editorial correction of this paper, anonymous reviewers fortheir valuable comments and corrections, the CCRE-MarineScience Network program, Smithsonian Tropical ResearchInstitute, Bocas del Toro Panama, and NOAA OceanExploration Project ‘‘Cayman Islands Twilight Zone’’ forallowing my studies in mangrove and coral reef ecosystemsduring the last 10 years, the PorTol team and grants forsupporting my attendance at the 8th sponge conference, Gerona,Spain, 2010, and the organizers of the conference. Portions ofthis work were supported by the US National ScienceFoundation under grant 0829986 to Dr Robert Thacker.
References
Alcolado, P. M., 2002. Catalogo de las esponjas de Cuba.Avicennia 15: 53–72.
Alvarez, B. & M. C. Dıaz, 1985. Las esponjas de un arrecifecoralino en el Parque Nacional Archipielago de LosRoques: Taxonomıa y Ecologıa. Universidad Central deVenezuela, Caracas, Venezuela. Trabajo Especial deGrado para optar al Tıtulo de Licenciado en Biologıa.
Alvizu, A., 2006. Estructura comunitaria de esponjas asociadasa dos parches coralinos en la Isla de Cubagua, Venezuela.Universidad de Oriente, Margarita, Venezuela. Trabajopresentado para optar al tıtulo de Licenciado en BiologıaMarina.
Amaro, M. E., 2005. Demospongiae (Porifera) asociadas a su-strato coralino, en la Bahıa de Mochima, Estado Sucre,Venezuela, durante el ano 2004–2005. Universidad deOriente, Cumana, Venezuela. Trabajo de grado para optaral tıtulo de MSc Ciencias Marinas.
Bingham, B. L. & C. M. Young, 1995. Stochastic events anddynamics of mangrove root epifaunal community. MarineEcology 16: 145–163.
Clarke, K. R., 1993. Non-parametric multivariate analyses ofchanges in community structure. Australian Journal ofEcology 18: 117–143.
Clarke, K. R. & R. N. Gorley, 2006. Plymouth Routines inMultivariate Ecological Research, Primer V6: User Man-ual/Tutorial. PRIMER-E Ltd, Plymouth, UK.
Clarke, K. R. & R.M.Warwick, 1998. A taxonomic distinctnessindex and its statistical properties. Journal of AppliedEcology 35: 523–531.
Clifton, K. E., K. Kim & J. Wulff, 1997. A field guide to thereefs of Caribbean Panama with emphasis of Western SanBlas. In Lessios, H. A. & I. G. Macintyre (eds), Proceed-ings of the 8th International Coral Reef Symposium, Vol.1: 167–184.
de Goeij, J. M., H. van den Berg, M. Martine, E. van Oostveen,H. G. Epping & F. C. van Duyl, 2008. Major bulk dissolvedorganic carbon (DOC) removal by encrusting coral reefcavity sponges. Marine Ecology Progress Series 357:139–151.
Dıaz, M. C., 2005. Common sponges from shallow marinehabitats from Bocas del Toro region, Panama. CaribbeanJournal of Science 41: 465–475.
Dıaz, M. C. &K. Rutzler, 2001. Sponge: an essential componentof Caribbean coral reefs. Bulletin of Marine Science 69:535–546.
Dıaz, M. C. & K. Rutzler, 2009. Biodiversity and abundance ofsponges in Caribbean mangrove: indicators of environ-mental quality. Smithsonian Contributions to the MarineSciences 38: 151–172.
Diaz, M. C., B. Alvarez & R. A. Laughlin, 1991. The ecology ofsponges in a fringing coral reef at the National park‘‘Archipelago de los Roques’’, Venezuela II: Communitystructure. In Ruetzler, K. (ed.), New Perspectives in SpongeBiology. Smithsonian Institution Press, Washington, DC:367–375.
Dıaz, M. C., K. P. Smith & K. Rutzler, 2004. Sponge speciesrichness and abundance as indicators of mangrove epiben-thic community health. Atoll Research Bulletin 518: 1–17.
Dıaz, M. C., R. W. Thacker & K. Rutzler, 2007. Haliclona(Soestella) walentinae (Chalinidae, Haplosclerida) andXestospongia bocatorensis (Petrosiidae, Haplosclerida), twonew sponge species from Caribbean Panama with filamen-tous cyanobacterial symbionts. In Custodio, M. R., G. Lobo-Hajdu, E. Hajdu & G. Muricy (eds), Porifera Research:Biodiversity, Innovation, and Sustainability, Vol. 1: 31–36.
Dunlap, M. & J. R. Pawlik, 1996. Video-monitored predation byCaribbean reef fishes on an array of mangrove and reefsponges. Marine Biology 126: 117–123.
Hydrobiologia
123
Author's personal copy

Dunlap, M. & J. R. Pawlik, 1998. Spongivory by parrotfish inFlorida mangrove and reef habitats. Marine Ecology 19:325–337.
Duran, S. & K. Rutzler, 2006. Ecological speciation in aCaribbean marine sponge. Molecular Phylogenetics andEvolution 40: 292–297.
Ellison, A. M., E. J. Farnsworth & R. E. Merkt, 1999. Origins ofmangrove ecosystems and the mangrove biodiversityanomaly. Global Ecology and Biogeography 8: 95–115.
Engel, S. & J. R. Pawlik, 2005a. Interactions among Floridasponges. I. Reef habitats. Marine Ecology Progress Series303: 133–144.
Engel, S. & J. R. Pawlik, 2005b. Interactions among Floridasponges. II. Mangrove habitats. Marine Ecology ProgressSeries 303: 145–152.
Farnsworth, E. J. &A.M. Ellison, 1996. Scale-dependent spatialand temporal variability in biogeography of mangrove rootepibiont communities. Ecological Monographs 66: 45–66.
Guerra-Castro, E., P. Young, A. Perez-Vazquez, S. Carteron &A. Alvizu, 2011. Spatial variability and human disturbanceof sponge assemblages associated with mangrove roots inthe southern Caribbean. Marine & Freshwater Research 62:491–501.
Hechtel, G. J., 1965. A Systematic Study of the Demospongiaeof Port Royal, Jamaica. Peabody Museum of Natural His-tory. Yale University, New Haven, CT.
Hunting, E. R., H. G. van der Geest, A. M. Krieg, M. B. L. vanMierlo & R. W. M. van Soest, 2010. Mangrove–spongeassociations: a possible role for tannins. Aquatic Ecology44: 679–684.
Kobluk, D. R. & R. W. M. van Soest, 1989. Cavity-dwellingsponges in a southern Caribbean coral reef and their pale-ontological implications. Bulletin of Marine Science 44:1207–1235.
Lacerda, L. D., J. E. Conde, B. Kjerfve, R. Alvarez-Leon, C.Alarcon & J. Polanıa, 2002. American mangroves. In deLacerda, L. D. (ed.), Mangrove Ecosystems: Function andManagement. Springer-Verlag, Berlin: 1–62.
Lehnert, H. & R. W. M. van Soest, 1996. North Jamaican deepfore-reef sponges. Beaufortia 46: 53–81.
Lehnert, H. & R.W.M. van Soest, 1998. Shallow water spongesof Jamaica. Beaufortia 48: 71–103.
Lesser, M., 2006. Benthic–pelagic coupling on coral reefs:feeding and growth of Caribbean sponges. Journal ofExperimental Marine Biology and Ecology 328: 277–288.
Maldonado, M., 2006. The ecology of the sponge larva. Cana-dian Journal of Zoology 84: 175–194.
Mumby, P. J., A. R. Harborne, J. Williams, C. V. Kappel, D.R. Brumbaugh, F. Micheli, K. E. Holmes, C. P. Dahlgren,C. B. Paris & P. G. Blackwell, 2007. Trophic cascadefacilitates coral recruitment in a marine reserve. Proceed-ings of the National Academy of Sciences USA 104:8362–8367.
Pauls, S. M., 2003. Esponjas. In Aguillera, M., A. Azocar & E.Gonzalez (eds), Biodiversidad en Venezuela. Tomo I.Fundacion Polar & Ministerio de Ciencia y Tegnologıa,Caracas: 210–219.
Pawlik, J. R., 1997. Fish predation on Caribbean reef sponges:an emerging perspective of chemical defenses. In Pro-ceedings of the 8th International Coral Reef Symposium,Vol. 2: 1255–1258.
Pawlik, J. R., 1998. Coral reef sponge: do predatory fishes affecttheir distribution? Limnology and Oceanography 43:1396–1399.
Pawlik, J. R., S. McMurray & T. P. Henkel, 2007. Abioticfactors control sponge ecology in Florida mangroves.Marine Ecology Progress Series 339: 93–98.
Perez-Vazquez, A., 2007. Estudio de las comunidades deesponjas sociadas a las raıces del mangle rojo Rhizophoramangle en la Laguna de la Restinga entre julio 2004 yagosto 2005. Universidad de Oriente, Margarita, Venezu-ela. Trabajo presentado para optar al tıtulo de Licenciadoen Biologıa Marina.
Ramirez, I. A., 2010. Taxonomıa de esponjas marinas (Porifera)de la Isla de la Tortuga, dependencia federal, Venezuela.Trabajo de grado de Magister en Ciencias Marinas. Uni-versidad de Oriente, Cumana: 114 pp.
Rutzler, K., 1995. Low-tide exposure of sponges in a Caribbeanmangrove community. Marine Ecology 16: 165–179.
Rutzler, K., M. C. Dıaz, R.W.M. van Soest, S. Zea, K. P. Smith,B. Alvarez & J. Wulff, 2000. Diversity of sponge fauna inmangrove ponds, Pelican Cays, Belize. Atoll ResearchBulletin 476: 230–248.
Sutherland, J. P., 1980. Dynamics of the epibenthic communityon roots of the mangrove Rhizophora mangle, at Bahia deBuche, Venezuela. Marine Biology 58: 75–84.
Toffart, J. L., 1983. Les Peuplements des racines de paletuviersen Guadeloupe (Antilles Francaises), I. Analyse floristiqueet faunistique; methodologie et premier resultats. Bulletind’Ecologie 14: 227–239.
van Soest, R. W. M., 1978. Marine sponges from Curacao andother Caribbean localities. Part I. Keratosa. In Humme-linck, P. W. & L. J. van der Stee (eds), Studies on the Faunaof Curacao and Other Caribbean Islands, Vol. 56. M. Nij-hoff, The Hague: 1–94.
van Soest, R. W. M., 1980. Marine sponges from Curacao andother Caribbean licalities, Part II. Haplosclerida. In Hum-melinck, P. W. & L. J. van der Steen (eds), Studies on theFauna of Curacao and Other Caribbean Islands, Vol. 62.M. Nijhoff, The Hague: 1–173.
van Soest, R. W. M., 1984. Marine sponges from Curacao andother Caribbean localities, Part III. Poecilosclerida. InHummelinck, P. W. & L. J. van der Steen (eds), Studies onthe Fauna of Curacao andOther Caribbean Islands, Vol. 66.M. Nijhoff, The Hague: 1–167.
van Soest, R. W. M., 2009. New sciophilous sponges from theCaribbean (Porifera: Demospongiae). Zootaxa 2107: 1–40.
van Soest, R. W. M., N. Boury-Esnault, J. N. A. Hooper, K.Rutzler, N. J. de Voogd, B. Alvarez de Glasby, E. Hajdu, A.B. Pisera, R. Manconi, C. Schoenberg, D. Janussen, K.R. Tabachnick, M. Klautau, B. Picton & M. Kelly, 2010.World Porifera database. http://www.marinespecies.org/porifera.
Wapstra, M. & R. W. M. van Soest, 1987. Sexual reproduction,larval morphology and behaviour in Demosponges fromthe southwest of the Netherlands. In Vacelet, J. & N. Bo-ury-Esnault (eds), Taxonomy of Porifera. Springer-Verlag,Berlin: 281–307.
Warwick, R. M. & K. R. Clarke, 2001. Practical measures ofmarine biodiversity based on relatedness of species.Oceanography and Marine Biology: An Annual Review39: 207–231.
Hydrobiologia
123
Author's personal copy

Weil, E., 2006. Diversidad y abundancia relativa de corales,octocorales y esponjas en el Parque Nacional Jaragua,Republica Dominicana. Revista de Biologıa Tropical 54:423–443.
Wood, R., 1998. The ecological evolution of reefs. AnnualReviews of Ecology, Evolution, and Systematics 29:179–206.
Wulff, J. L., 1997. Parrotfish predation on cryptic sponges ofCaribbean coral reefs. Marine Biology 129: 41–52.
Wulff, J., 2004. Sponges on mangrove roots, Twin Cays, Belize:early stages of community assembly. Atoll Research Bul-letin 519: 1–10.
Wulff, J. L., 2005. Trade-offs in resistance to competitors andpredators, and their effects on the diversity of tropicalmarine sponges. Journal of Animal Ecology 74: 313–321.
Wulff, J. L., 2009. Sponge community dynamics on Caribbeanmangrove roots: significance of species idiosyncrasies.Smithsonian Contributions to Marine Science 38: 501–514.
Hydrobiologia
123
Author's personal copy




















