
Epidermal growth factor binding sites in human pituitary macroadenomas
Upload
independentCategory
view
3download
0
�1-Adrenoceptor Expression in Rat Anterior PituitaryGonadotrophs and in Mouse �T3-1 and L�T2Gonadotrophic Cell Lines
Kristel Janssens, Olga Krylyshkina, Nicole Hersmus, Hugo Vankelecom, and Carl Denef
Laboratory of Cell Pharmacology, University of Leuven, Medical School, Gasthuisberg, B-3000 Leuven, Belgium
The rat anterior pituitary expresses �2-adrenoceptors (ARs)on somatotrophs, lactotrophs, and corticotrophs. The presentstudy investigates whether �1-ARs exist in the anterior pitu-itary, in which cell type(s) they are found, and whether theyare regulated by glucocorticoids. As determined by quantita-tive RT-PCR and Western immunoblotting, the rat anteriorpituitary expressed �1-AR mRNA and protein. Unlike the �2-AR, expression decreased to very low levels after 5-d aggre-gate cell culture but was strongly up-regulated in a dose- andtime-dependent manner by dexamethasone (DEX). Glucocor-ticoids attenuated isoproterenol-induced down-regulation of�1-AR mRNA levels. As examined by immunofluorescence con-focal microscopy, �1-AR immunoreactivity was detected in asubpopulation of gonadotrophs, but not in somatotrophs, lac-totrophs, corticotrophs, thyrotrophs, or folliculo-stellatecells. �1-AR-immunoreactivity cells were often surrounded by
cup-shaped lactotrophs. Consistent with these findings, �1-ARmRNA was considerably more abundant in the gonadotrophic�T3-1 and L�T2 cell lines than in the GHFT, GH3, and TtT/GFcell lines. DEX did not affect expression level in the cell lines.DEX also failed to up-regulate �1-AR mRNA levels in aggre-gates from a subpopulation enriched in large gonadotrophsobtained by gradient sedimentation. In contrast, excessiveDEX-dependent up-regulation of �1-AR mRNA was found in asubpopulation enriched in small nonhormonal cells. Thepresent data indicate that �1-AR is expressed in a subpopulationof gonadotrophs with a topographical relationship to lac-totrophs. However, the glucocorticoid-induced up-regulationdoes not seem to occur directly in the gonadotrophs but within(an)other unidentified cell type(s), or is transduced by that celltype on gonadotrophs. (Endocrinology 149: 2313–2324, 2008)
CATECHOLAMINES AND glucocorticoids play an es-sential role in the stress response and contribute to the
pathophysiology of cardiovascular diseases, depression, andanxiety disorders (1), and exert a suppressive influence onreproduction (2). All pituitary hormones participate at di-verse levels during coping with stress (3), and, therefore, itis not surprising that secretion and expression of these hor-mones are modulated by catecholamines and glucocorti-coids. Catecholamines control the secretion of ACTH (4–9),GH (4, 5, 10–14), prolactin (PRL) (4, 15), TSH (4, 16), and LH(4, 17, 18) at the level of the hypothalamus and/or the pi-tuitary. The effects of catecholamines are mediated by �- and�-adrenoceptors (ARs) that are members of a large super-family of G protein-coupled receptors (19).
Glucocorticoids are known to modulate �-AR levels,thereby regulating the cellular responsiveness to cat-echolamines (20). At the pituitary level, glucocorticoids havefast and delayed effects on various hormonal cells. Fast non-genomic effects are mediated by folliculo-stellate (FS) cellsvia translocation of annexin-1 on the cell surface, and resultin inhibition of ACTH, GH, and PRL release by a paracrine
action (21). Genomic effects are established at a slower ratevia the glucocorticoid nuclear receptor. In rat pituitary cellaggregates, Baes and Denef (13) showed a strong potentia-tion by dexamethasone (DEX) of isoproterenol (ISO) andepinephrine-stimulated GH release through �2-ARs. In ad-dition, glucocorticoids enhance GH release in response to�1-AR activation (14). Baes and Denef (13) furthermoreshowed the importance of intercellular communication inthese potentiating effects of DEX. Therefore, the anteriorpituitary represents an interesting model to study the impactof local regulation in catecholamine action.
The �2-AR is the predominant �-AR subtype in the pitu-itary (22), which led research to be mainly focused on thissubtype in previous studies (7, 11, 15, 22–25). The presenceof the �1-AR has been demonstrated, but no precise cell typelocalization was made, although it may participate in theregulation of GH secretion (26). Based on pharmacologicalexperiments, bovine FS cells in culture have expressed both�1- and �2-AR in approximately equal proportions, and re-sponded to �-AR stimulation in terms of cAMP accumula-tion (27). However, no evidence for �1-AR expression couldbe found in rat pituitary cell cultures (4, 5, 10–14).
The increasing evidence that the anterior pituitary is adirect target of peripheral signals related to stress (21), re-production (28), and energy homeostasis (29) revitalized ourinterest in the role of catecholamines at the anterior pituitarylevel. The present study was intended to further investigate�1-AR expression in the anterior pituitary, its responsivenessto glucocorticoids, and its cellular distribution. We used �1-and �2-AR mRNA as a primary read-out system by quan-
First Published Online January 17, 2008Abbreviations: AR, Adrenoceptor; DEX, dexamethasone; DNase, de-
oxyribonuclease; FCS, fetal calf serum; FS, folliculo-stellate; GLM, gen-eralized linear model; �GSU, glycoprotein hormone �-subunit; ir, im-munoreactivity; ISO, isoproterenol; PEX14P, peroxin 14P; POMC,proopiomelanocortin; PRL, prolactin; qRT-PCR, quantitative RT-PCR;SP, side population.Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
0013-7227/08/$15.00/0 Endocrinology 149(5):2313–2324Printed in U.S.A. Copyright © 2008 by The Endocrine Society
doi: 10.1210/en.2007-1396
2313

titative RT-PCR (qRT-PCR) analysis that unequivocally de-tects expression of the receptor, confirmed expression at theprotein level, and determined the cellular distribution byimmunocytochemistry. Furthermore, we investigated �1-ARmRNA and protein expression in various pituitary cell linesrepresentative for each hormonal cell type (GHFT, GH3,AtT-20, �T3-1, and L�T2), and FS cells (TtT/GF). We dem-onstrate that �1-AR is expressed in the anterior pituitary,more precisely in gonadotrophs that are juxtaposed to lac-totrophs. Receptor mRNA level is strongly up-regulated byglucocorticoids, but this glucocorticoid action does not occurdirectly in gonadotrophs but occurs either within anotherunidentified cell type or is transduced by that cell type on thegonadotrophs. In a companion paper (72), we show that the�1-AR constitutively activates adenylate cyclase via a per-tussis toxin-sensitive transduction system, suggesting thehypothesis that its function may be related to the trophicaction of gonadotrophs on lactotrophs.
Materials and MethodsAnimals
Adult (11–12 wk old) male Wistar rats were purchased from ElevageJanvier (Bio-Services, Uden, The Netherlands). The animals were keptin the animal house facilities of the University of Leuven under constanttemperature, humidity, and day-night cycle, and had free access to pelletfood and water. The rats were anesthetized with CO2 and killed bydecapitation. All experiments were conducted in accordance with theGuidelines for Care and Use of Experimental Animals and were ap-proved by the University Ethical Committee.
Anterior pituitary aggregate cell culture
Single cells were obtained by dispersion of the anterior pituitary lobesusing methods described previously (30). Briefly, the pituitaries were cutinto small blocks with a razor blade and enzymatically treated withporcine trypsin [2.5% from ICN Biomedicals (Aurora, OH) or 0.7% fromRoche Diagnostics (Indianapolis, IN)], deoxyribonuclease (DNase) (Sig-ma-Aldrich, Steinheim, Germany), soybean trypsin inhibitor (Sigma-Aldrich), and 2 mm EDTA in Ca2�- and Mg2�-free medium. Aftermechanical dispersion with a polished and narrowed Pasteur pipette,cells were treated again with DNase and centrifuged at 190 g througha 3% BSA layer (fraction V, standard grade; Serva, Heidelberg, Ger-many) to remove cell debris. The cell pellet was then resuspended inserum-free defined culture medium as previously described (30, 31).This medium consists of DMEM/F12 1/1 with selenite and ethanol-amine (prepared as powder by Life Technologies, Inc.-Invitrogen, GrandIsland, NY), supplemented with 0.5% BSA (lyophilized, cell culturegrade or fraction V receptor grade; Serva), 5 mg/liter insulin-Zn (Sigma-Aldrich), 5 mg/liter transferrin (Invitrogen), 50 mg/liter streptomycin(Sigma-Aldrich), 35 mg/liter penicillin (Sigma-Aldrich), 10 mm ethanol,1 mg/liter catalase (Roche Diagnostics), and 1 g/liter NaHCO3. Finalconcentration of iron (Fe2� and Fe3�) was 0.6 �m and of glucose 1.4g/liter (7.8 mm). No phenol red was added to the medium.
To establish three-dimensional aggregate cell cultures, anterior pi-tuitary cells were distributed at 2 � 106/2 ml culture medium in 35-mlnontreated culture dishes (Iwaki, Scitech division, Chiba, Japan), andallowed to re-associate on a gyratory shaker at 63 rpm in a humidified1.5–1.9% CO2 incubator at 37 C. Unless otherwise stated, this mediumwas supplemented with 50 pm T3 (Serva) diluted from a 0.5 �m stocksolution (in 0.9% NaCl) and/or various doses of DEX (Serva) dilutedfrom a 0.8 mm stock solution (in ethanol). Vehicle was added to thecontrols. After 2 d the aggregates of each dish corresponding to a par-ticular experimental treatment were divided equally over three dishes(each corresponding to �666,000 cells) with fresh medium added forfurther analysis with TaqMan real-time qRT-PCR. Analysis was per-formed on the fifth day of culture, and each experiment was repeatedthree times.
Velocity sedimentation at unit gravity
Freshly dispersed cells were allowed to sediment through a 0.3–2.4%BSA (Fraction V, standard grade; Serva) gradient to prepare subpopu-lations enriched for a particular cell type, as previously described (30,32, 33). Eleven fractions of 270 ml were collected from the gradient andcalled fractions A, B, and 1–9. In this study cells from fraction 3 andfraction 7–9 (further indicated as fraction 7 cells) were used. The pro-portion of the different pituitary cell types in the freshly isolated frac-tions was determined previously (10, 13, 32, 34–36). Because analysis of�1-AR mRNA was done at d-5 aggregate cell culture, we reanalyzed theproportions of pituitary cell types in fraction 3 and 7 after 5 d in ag-gregate cell culture by single-cell RT-PCR of mRNA of GH, PRL, pro-opiomelanocortin (POMC), and the glycoprotein hormone �-subunit(�GSU). The method has been described previously in detail (37, 38).
Cell line culture
GHFT cells, L�T2 cells, �T3-1 cells (all from P. Mellon, University ofCalifornia, San Diego, CA), and AtT20 cells (ATCC; LGC Promochem,Teddington, UK) were cultured in Advanced DMEM/F12 mixture (In-vitrogen) supplemented with GlutaMAX, streptomycin, penicillin (In-vitrogen), and 10% fetal calf serum (FCS) (Cambrex Bio Science, Ver-viers, Belgium). GH3 cells (ATCC; LGC Promochem) and TtT/GF cells(Riken Cell Bank, Tsukuba, Ibaraki, Japan) were cultured in AdvancedDMEM/F12 supplemented with GlutaMAX, streptomycin, penicillin,and 12.5% horse serum (Invitrogen), �2.5% FCS and 10% horse serum�2.5% FCS, respectively. All cells were seeded in 75 cm2 culture flasksand trypsinized twice a week using TrypLE (Invitrogen), except TtT/GFcells, which were released using a mixture of TrypLE and trypsin, andwere dispersed in a way similar to pituitary cell dispersion. Cultureswere maintained in a humidified 5% CO2 incubator at 37 C. Accordingto the experimental design, the medium was supplemented with 80 nmDEX or with vehicle during 3–4 d before experimentation.
Immunofluorescent staining
To avoid interpretation problems of immunostaining signals due toautofluorescent red blood cells, rats were anesthetized with CO2 andperfused subsequently with a PBS solution and a 4% paraformaldehyde(Riedel-deHaen, Seelze, Germany)-PBS solution. Complete pituitarieswere then dissected and further fixed using 4% paraformaldehyde (2 hat 4 C), rinsed with Ca2�- and Mg2�-free PBS, and embedded in 2%agarose (ICN Biomedicals) (in PBS). The samples were mounted on aVibratome (Leica VT 1000 S; Leica Microsystems, Wetzlar, Germany),and sections of 40 �m were made. The slices were transferred to 24-wellplates and stored in PBS at 4 C until additional processing. The completestaining procedure was performed at room temperature. The floatingsections were rinsed with PBS and permeabilized with 0.5% saponin(Sigma-Aldrich) in PBS (2 � 10 min). To block nonspecific binding sites,sections were incubated with 20% normal goat serum (Sigma-Aldrich).Sections were treated overnight with the primary antibodies for the�1-AR and one of the pituitary hormones. Affinity purified rabbit an-tibody A272 against rat �1-AR (1:500 final dilution) was obtained fromSigma-Aldrich. The antibody was raised against a synthetic peptide(Gly-Asp-Arg-Pro-Arg-Ala-Ser-Gly-Cys-Leu-Ala-Arg-Ala-Gly) derivedfrom amino acids 394–408 of mouse and rat �1-AR C-terminal domain.Specificity for �1-AR was shown by the manufacturer for both immu-noblotting and immunofluorescent staining of sections of mouse kidneydistal tubule cells, and further tested in the present study by antibodyomission and preadsorption experiments (see Results). We also foundpositive staining on sections of rat kidney. The guinea pig antiseraagainst rat �GSU (1:1000), LH� (1:4000), TSH� (1:4000), GH (1:1000), andPRL (1:1000) were obtained from Dr. A. F. Parlow through the NationalHormone and Pituitary Program (Harbor-University of California LosAngeles Medical Center, Torrance, CA), and monoclonal mouse anti-body against human POMC (1–50) (1:100) was obtained from Biogenesis(Poole, UK). As negative controls, primary antibodies were omitted.After excessive rinsing, sections were incubated with secondary anti-bodies for 90 min while being shielded from light. Secondary goatantirabbit Alexa 488 (1:1000), goat antiguinea pig Alexa 555 (1:2000), andgoat antimouse Alexa 555 (1:1000) antibodies were purchased fromInvitrogen. TOPRO-3 (1:100) (Invitrogen) was added during 10 min to
2314 Endocrinology, May 2008, 149(5):2313–2324 Janssens et al. • �1-AR in Rat Pituitary Gonadotrophs

stain the nuclei. All steps were performed in 0.5% saponin-PBS. Afterthorough rinsing with PBS, sections were mounted in Vectashield (Vec-tor Laboratories, Burlingame, CA) on glass slides and covered with glasscoverslips. Sections were scanned using a confocal laser-scanning mi-croscope (LSM 510; Zeiss, Zaventem, Belgium) and were analyzed usingthe Zeiss LSM Image Browser as previously described (39).
Western blotting
Membrane proteins were extracted according to the method of Bou-langer et al. (40). Treatment of cell lines and anterior pituitary cellaggregates was performed as described previously, except that the ag-gregates were treated with 80 nm DEX instead of 20 nm. All cells werepooled and spun down at 150 g for 20 sec and rinsed with ice-cold PBS.Subsequently, the pellet was sonicated for 30 sec in 500 �l TAME buffer[50 mm Tris-acetate (pH 7.4), 5 mm MgCl2, 5 mm EDTA, and 250 mmsucrose] to which complete protease inhibitor (Hoffmann-La Roche Ltd.,Basel, Switzerland) was added. This mixture was centrifuged for 10 minat 800 g. The supernatant was spun down at 45,000 g for 30 min. Thepellet was resuspended in TAME buffer, and quantification of the pro-tein content was done with the Pierce Micro BCA Protein Assay ReagentKit (Perbio Science, Erembodegem, Belgium). All steps were performedat 4 C. Proteins were denaturated in loading buffer containing Tris-HCl,glycerol, sodium dodecyl sulfate, bromophenol blue, and 2-mercapto-ethanol, and subsequently boiled for 5 min. Thirty to 75 �G protein ofthe samples was loaded on a sodium dodecyl sulfate/polyacrylamidegel (12%), as well as a broad-range prestained protein marker (NewEngland Biolabs, Ipswich, MA). After electrophoresis, proteins wereblotted on an Immobilon-P membrane (Millipore, Brussels, Belgium).The blots were blocked for nonspecific binding using 5% BSA in Tris-buffered saline to which 0.1% Tween-20 was added. A rabbit antirat�1-AR (1:250) primary antibody was obtained from Sigma-Aldrich (seeImmunofluorescent staining section above). The antibody was added over-night in Tris-buffered saline to which 0.1% Tween 20 was added (4 C),after which an alkaline-phosphatase-coupled antirabbit secondary an-tibody was added for 1 h. After incubating the blots with ECF substrate(GE Healthcare, Diegem, Belgium), chemiluminescence was measuredusing the Storm 840 Phosphor/FluorImager, and analyzed using Im-ageQuaNT 5.0 software (Molecular Dynamics, Sunnyvale, CA). To checkequal loading of samples, the blots were stripped with methanol, and theamount of the peroxisomal membrane protein peroxin 14P (PEX14P)was detected with rabbit anti-PEX14P (41) as the primary antibody.
qRT-PCR
Pituitary cell aggregates were collected on d-5 culture and transferredto 0.5 ml TriPure RNA isolation reagent (Hoffmann-La Roche Ltd.). Celllines were trypsinized and transferred to 1 ml TriPure. RNA isolationand RT were performed as described before (31), with some minormodifications. In brief, the RNA solution was treated with Rnase-freeDNase I (1 U/�l; Invitrogen) to destroy contaminating genomic DNA.The RT reaction was performed on 200 ng RNA in a final volume of 20�l RT mixture. qRT-PCR was performed as described previously (42)using TaqMan Universal PCR Master Mix (Applied Biosystems, Lennik,Belgium). Primers to amplify rat GH and PRL cDNA, and rat as well asmouse �1- and �2-AR cDNA (Table 1) were purchased from Invitrogen,and TaqMan probes were obtained from Eurogentec (Seraing, Belgium).
The primers for GH and PRL were designed over an intron. Given theabsence of introns in �1- and �2-AR genes, precautions had to be takento avoid amplification of genomic DNA. First, as mentioned, a DNasetreatment was performed on the RNA extract. Second, for each sampleto be analyzed, a negative RT control (i.e. RT in the absence of reversetranscriptase) was included. Amplification signal was not different frombackground in all the negative RT controls. Normalization of the mRNAlevel was performed against 18S rRNA, measured using the TaqManRibosomal RNA Control Reagents Kit (Applied Biosystems).
To quantify receptor mRNA levels, the equivalent of 17 ng initialRNA was used for qPCR. To quantify receptor expression, too highamounts of pituitary cDNA were needed to set up a standard curve.Therefore, a standard of receptor cDNA was made as follows. �1- and�2-AR fragments were amplified by PCR from pituitary cDNA. Theamplified fragment was loaded onto a 2% agarose gel, extracted usingGenElute Gel Extraction kit (Sigma-Aldrich), ligated into 2.1 Topo vector(Topo TA Cloning kit; Invitrogen), and the plasmids used to transformTOP10F’ Escherichia coli cells. The cells were plated on LB agar withampicillin overnight. Colonies were picked and transferred to liquid LBampicillin. Plasmid DNA was isolated using the boiling-prep-methodand checked by enzyme restriction for insertion of the PCR fragment,after which plasmids were purified by phenol/chloroform extractionand sequenced by Lark (Essex, UK). Plasmid DNA (containing thereceptor cDNA) was quantified using PicoGreen to calculate copy num-bers. This stock was serially diluted (from 105 to one copy) to use asstandard curve in qRT-PCR. To analyze GH mRNA levels, the RT prod-uct was further diluted in such a way that the equivalent of 170 pg initialRNA was added to the final 25 �l qPCR mixture. For quantificationpituitary cDNA was used in a dilution series from 10 ng to 0.1 pg andrun together with the samples to be analyzed.
The mRNA values measured were normalized for 18S rRNA content.The validity of 18S rRNA as internal control for qRT-PCR was verifiedagainst other housekeeping genes (�-actin and glyceraldehyde-3-phos-phate dehydrogenase). Cycle threshold standard curves for all house-keeping gene products were parallel, and relative expression levelsobtained for pituitary hormone mRNAs were similar, regardless of thehousekeeping gene used for normalization. All samples were analyzedin triplicate.
Statistical analysis
All experiments were independently repeated at least three times.Values were expressed as mean � sem. Most data were log or ln trans-formed because of heterogeneity of variance. Data were compared byone-way, two-way, or three-way generalized linear model (GLM)ANOVA with the Tukey-Kramer comparison test, depending on theexperimental design. The statistical package used for all experimentswas NCSS (Statistical Solutions Ltd., Cork, Ireland).
Results�1- and �2-AR mRNA is differentially expressed in theanterior and neuro-intermediate pituitary ex vivo
Intact anterior pituitary as well as neuro-intermediate lobeof adult male rats expressed both �1- and �2-AR mRNA (Fig.
TABLE 1. Primer and probe oligonucleotide sequences for qRT-PCR and accession numbers in GenBank for rat GH (rGH), rat �1 (r�1)-,rat (r�2)-, mouse �1 (m�1)-, and mouse �2 (m�2)-ARs
GenBank accession no. Primers Probe
rGH U62779 F:5�-CTCGGACCGCGTCTATGAGA-3� 5�-FAM-CCGTCTTCCAGCTCCTGCATCAGAG-TAMRA-3�B:5�-TGAGGATCTGCCCAATACGG-3�
r�1-AR NM_012701 F:5�-GCTGCTACAACGACCCCAAG-3� 5�-FAM-CTCGTCCGTCGTCTCCTTCTACGTGC-TAMRA-3�B:5�-TTCACCTGTTTCTGGGCCTC-3�
r�2-AR NM_012492 F:5�-TGACCAAGAATAAGGCCCGA-3� 5�-FAM-CGACACAATCCACACCATTAGGATGACCA-TAMRA-3�B:5�-CAGTCGATGGCTTGCTTGTG-3�
m�1-AR L10084 F:5�-GAAGGCGCTCAAGACACTGG-3� 5�-FAM-CAGAGCGTGAACACACCCATGATGATG-TAMRA-3�B:5�-CCAGGTCGCGGTGGAA-3�
m�2-AR X15643 F:5�-CCTCAGCCAGGTGGAGCA-3� 5�-FAM-CCACGGACTCCGAAGGTCCTCCA-TAMRA-3�B:5�-TGATGCCTAAAGTCTTGAGGGC-3�
Janssens et al. • �1-AR in Rat Pituitary Gonadotrophs Endocrinology, May 2008, 149(5):2313–2324 2315
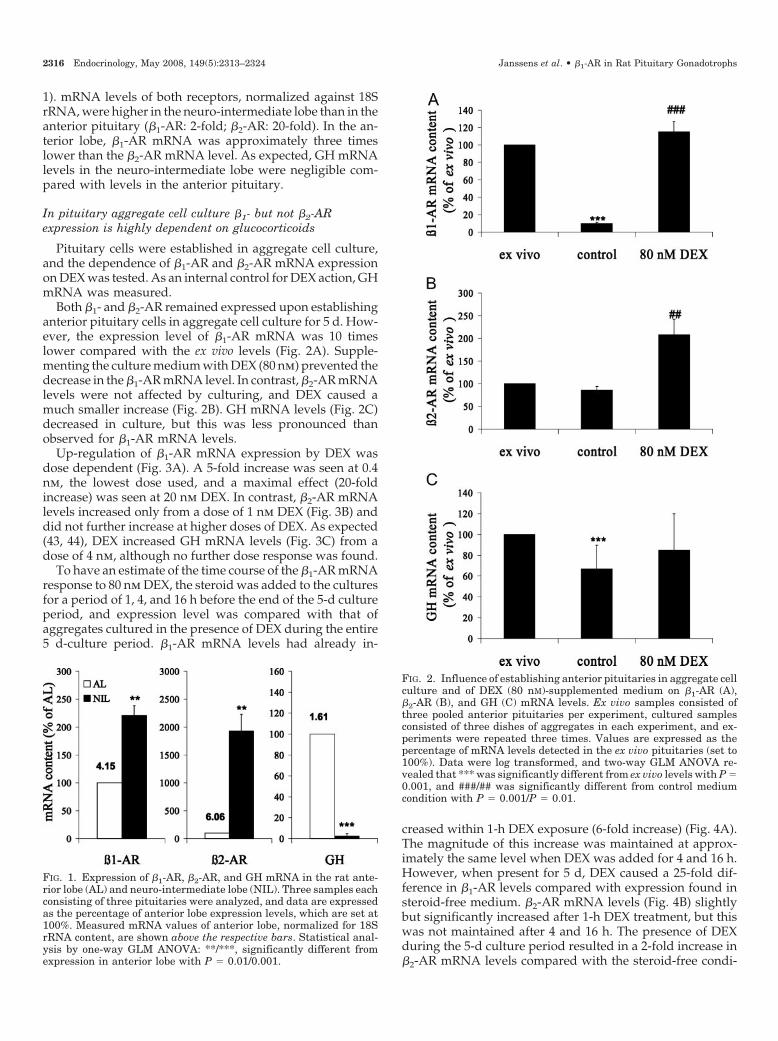
1). mRNA levels of both receptors, normalized against 18SrRNA, were higher in the neuro-intermediate lobe than in theanterior pituitary (�1-AR: 2-fold; �2-AR: 20-fold). In the an-terior lobe, �1-AR mRNA was approximately three timeslower than the �2-AR mRNA level. As expected, GH mRNAlevels in the neuro-intermediate lobe were negligible com-pared with levels in the anterior pituitary.
In pituitary aggregate cell culture �1- but not �2-ARexpression is highly dependent on glucocorticoids
Pituitary cells were established in aggregate cell culture,and the dependence of �1-AR and �2-AR mRNA expressionon DEX was tested. As an internal control for DEX action, GHmRNA was measured.
Both �1- and �2-AR remained expressed upon establishinganterior pituitary cells in aggregate cell culture for 5 d. How-ever, the expression level of �1-AR mRNA was 10 timeslower compared with the ex vivo levels (Fig. 2A). Supple-menting the culture medium with DEX (80 nm) prevented thedecrease in the �1-AR mRNA level. In contrast, �2-AR mRNAlevels were not affected by culturing, and DEX caused amuch smaller increase (Fig. 2B). GH mRNA levels (Fig. 2C)decreased in culture, but this was less pronounced thanobserved for �1-AR mRNA levels.
Up-regulation of �1-AR mRNA expression by DEX wasdose dependent (Fig. 3A). A 5-fold increase was seen at 0.4nm, the lowest dose used, and a maximal effect (20-foldincrease) was seen at 20 nm DEX. In contrast, �2-AR mRNAlevels increased only from a dose of 1 nm DEX (Fig. 3B) anddid not further increase at higher doses of DEX. As expected(43, 44), DEX increased GH mRNA levels (Fig. 3C) from adose of 4 nm, although no further dose response was found.
To have an estimate of the time course of the �1-AR mRNAresponse to 80 nm DEX, the steroid was added to the culturesfor a period of 1, 4, and 16 h before the end of the 5-d cultureperiod, and expression level was compared with that ofaggregates cultured in the presence of DEX during the entire5 d-culture period. �1-AR mRNA levels had already in-
creased within 1-h DEX exposure (6-fold increase) (Fig. 4A).The magnitude of this increase was maintained at approx-imately the same level when DEX was added for 4 and 16 h.However, when present for 5 d, DEX caused a 25-fold dif-ference in �1-AR levels compared with expression found insteroid-free medium. �2-AR mRNA levels (Fig. 4B) slightlybut significantly increased after 1-h DEX treatment, but thiswas not maintained after 4 and 16 h. The presence of DEXduring the 5-d culture period resulted in a 2-fold increase in�2-AR mRNA levels compared with the steroid-free condi-
FIG. 1. Expression of �1-AR, �2-AR, and GH mRNA in the rat ante-rior lobe (AL) and neuro-intermediate lobe (NIL). Three samples eachconsisting of three pituitaries were analyzed, and data are expressedas the percentage of anterior lobe expression levels, which are set at100%. Measured mRNA values of anterior lobe, normalized for 18SrRNA content, are shown above the respective bars. Statistical anal-ysis by one-way GLM ANOVA: **/***, significantly different fromexpression in anterior lobe with P � 0.01/0.001.
FIG. 2. Influence of establishing anterior pituitaries in aggregate cellculture and of DEX (80 nM)-supplemented medium on �1-AR (A),�2-AR (B), and GH (C) mRNA levels. Ex vivo samples consisted ofthree pooled anterior pituitaries per experiment, cultured samplesconsisted of three dishes of aggregates in each experiment, and ex-periments were repeated three times. Values are expressed as thepercentage of mRNA levels detected in the ex vivo pituitaries (set to100%). Data were log transformed, and two-way GLM ANOVA re-vealed that *** was significantly different from ex vivo levels with P �0.001, and ###/## was significantly different from control mediumcondition with P � 0.001/P � 0.01.
2316 Endocrinology, May 2008, 149(5):2313–2324 Janssens et al. • �1-AR in Rat Pituitary Gonadotrophs
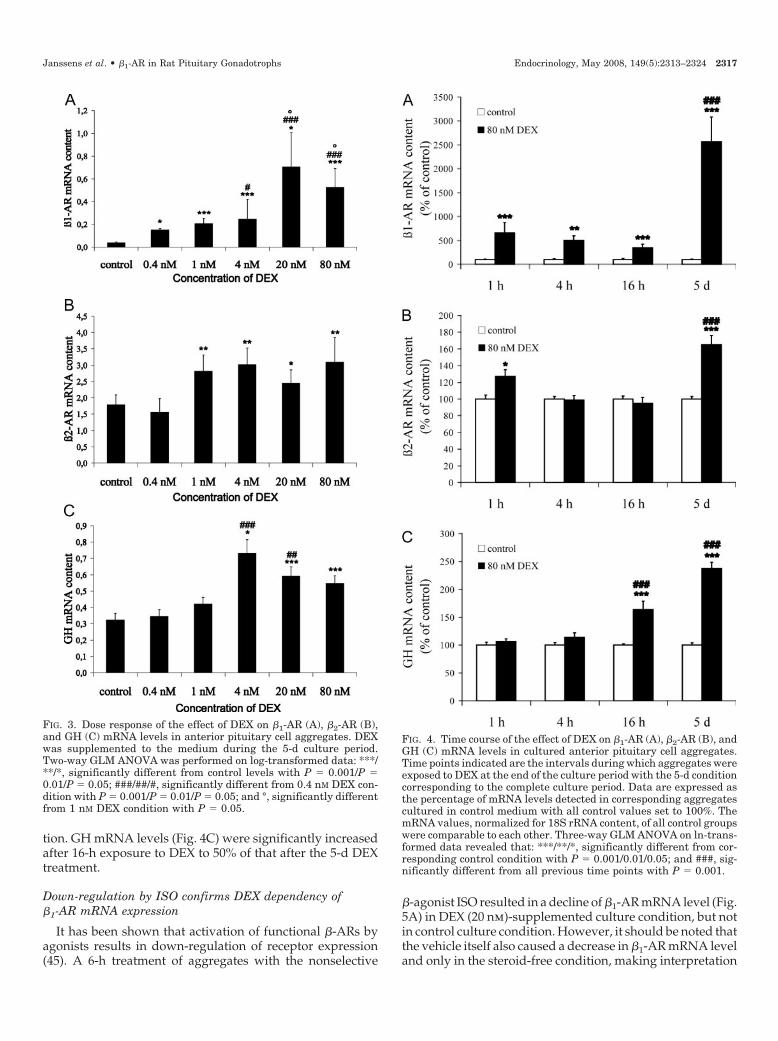
tion. GH mRNA levels (Fig. 4C) were significantly increasedafter 16-h exposure to DEX to 50% of that after the 5-d DEXtreatment.
Down-regulation by ISO confirms DEX dependency of�1-AR mRNA expression
It has been shown that activation of functional �-ARs byagonists results in down-regulation of receptor expression(45). A 6-h treatment of aggregates with the nonselective
�-agonist ISO resulted in a decline of �1-AR mRNA level (Fig.5A) in DEX (20 nm)-supplemented culture condition, but notin control culture condition. However, it should be noted thatthe vehicle itself also caused a decrease in �1-AR mRNA leveland only in the steroid-free condition, making interpretation
FIG. 3. Dose response of the effect of DEX on �1-AR (A), �2-AR (B),and GH (C) mRNA levels in anterior pituitary cell aggregates. DEXwas supplemented to the medium during the 5-d culture period.Two-way GLM ANOVA was performed on log-transformed data: ***/**/*, significantly different from control levels with P � 0.001/P �0.01/P � 0.05; ###/##/#, significantly different from 0.4 nM DEX con-dition with P � 0.001/P � 0.01/P � 0.05; and °, significantly differentfrom 1 nM DEX condition with P � 0.05.
FIG. 4. Time course of the effect of DEX on �1-AR (A), �2-AR (B), andGH (C) mRNA levels in cultured anterior pituitary cell aggregates.Time points indicated are the intervals during which aggregates wereexposed to DEX at the end of the culture period with the 5-d conditioncorresponding to the complete culture period. Data are expressed asthe percentage of mRNA levels detected in corresponding aggregatescultured in control medium with all control values set to 100%. ThemRNA values, normalized for 18S rRNA content, of all control groupswere comparable to each other. Three-way GLM ANOVA on ln-trans-formed data revealed that: ***/**/*, significantly different from cor-responding control condition with P � 0.001/0.01/0.05; and ###, sig-nificantly different from all previous time points with P � 0.001.
Janssens et al. • �1-AR in Rat Pituitary Gonadotrophs Endocrinology, May 2008, 149(5):2313–2324 2317
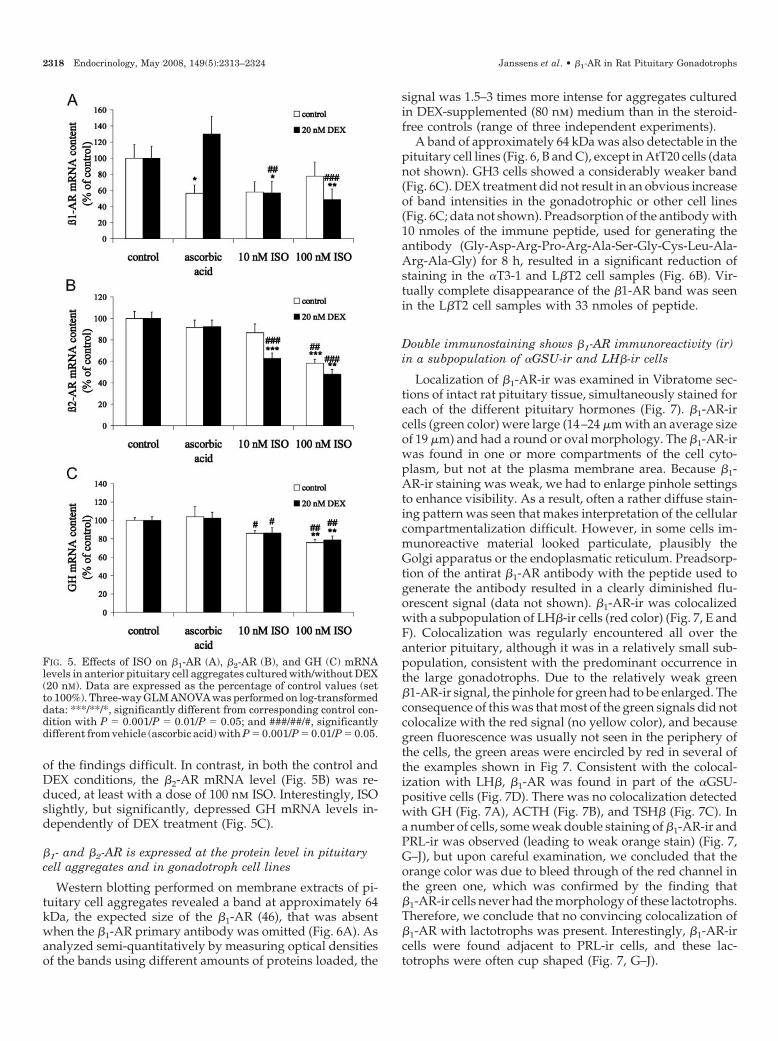
of the findings difficult. In contrast, in both the control andDEX conditions, the �2-AR mRNA level (Fig. 5B) was re-duced, at least with a dose of 100 nm ISO. Interestingly, ISOslightly, but significantly, depressed GH mRNA levels in-dependently of DEX treatment (Fig. 5C).
�1- and �2-AR is expressed at the protein level in pituitarycell aggregates and in gonadotroph cell lines
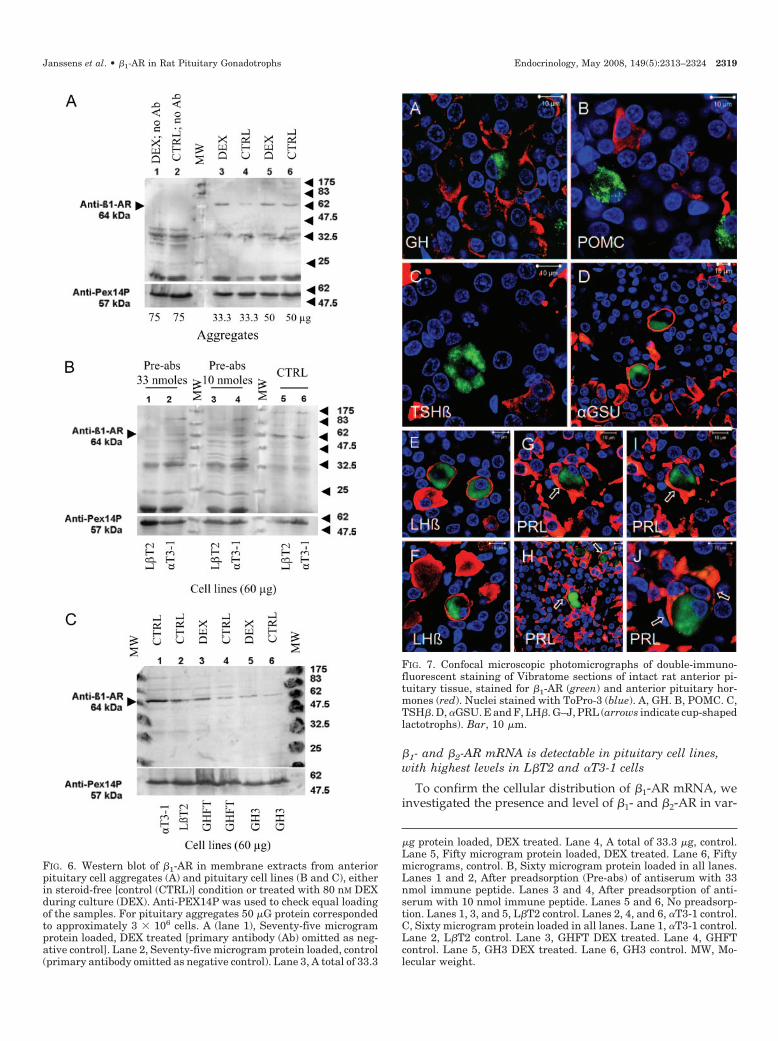
Western blotting performed on membrane extracts of pi-tuitary cell aggregates revealed a band at approximately 64kDa, the expected size of the �1-AR (46), that was absentwhen the �1-AR primary antibody was omitted (Fig. 6A). Asanalyzed semi-quantitatively by measuring optical densitiesof the bands using different amounts of proteins loaded, the
signal was 1.5–3 times more intense for aggregates culturedin DEX-supplemented (80 nm) medium than in the steroid-free controls (range of three independent experiments).
A band of approximately 64 kDa was also detectable in thepituitary cell lines (Fig. 6, B and C), except in AtT20 cells (datanot shown). GH3 cells showed a considerably weaker band(Fig. 6C). DEX treatment did not result in an obvious increaseof band intensities in the gonadotrophic or other cell lines(Fig. 6C; data not shown). Preadsorption of the antibody with10 nmoles of the immune peptide, used for generating theantibody (Gly-Asp-Arg-Pro-Arg-Ala-Ser-Gly-Cys-Leu-Ala-Arg-Ala-Gly) for 8 h, resulted in a significant reduction ofstaining in the �T3-1 and L�T2 cell samples (Fig. 6B). Vir-tually complete disappearance of the �1-AR band was seenin the L�T2 cell samples with 33 nmoles of peptide.
Double immunostaining shows �1-AR immunoreactivity (ir)in a subpopulation of �GSU-ir and LH�-ir cells
Localization of �1-AR-ir was examined in Vibratome sec-tions of intact rat pituitary tissue, simultaneously stained foreach of the different pituitary hormones (Fig. 7). �1-AR-ircells (green color) were large (14–24 �m with an average sizeof 19 �m) and had a round or oval morphology. The �1-AR-irwas found in one or more compartments of the cell cyto-plasm, but not at the plasma membrane area. Because �1-AR-ir staining was weak, we had to enlarge pinhole settingsto enhance visibility. As a result, often a rather diffuse stain-ing pattern was seen that makes interpretation of the cellularcompartmentalization difficult. However, in some cells im-munoreactive material looked particulate, plausibly theGolgi apparatus or the endoplasmatic reticulum. Preadsorp-tion of the antirat �1-AR antibody with the peptide used togenerate the antibody resulted in a clearly diminished flu-orescent signal (data not shown). �1-AR-ir was colocalizedwith a subpopulation of LH�-ir cells (red color) (Fig. 7, E andF). Colocalization was regularly encountered all over theanterior pituitary, although it was in a relatively small sub-population, consistent with the predominant occurrence inthe large gonadotrophs. Due to the relatively weak green�1-AR-ir signal, the pinhole for green had to be enlarged. Theconsequence of this was that most of the green signals did notcolocalize with the red signal (no yellow color), and becausegreen fluorescence was usually not seen in the periphery ofthe cells, the green areas were encircled by red in several ofthe examples shown in Fig 7. Consistent with the colocal-ization with LH�, �1-AR was found in part of the �GSU-positive cells (Fig. 7D). There was no colocalization detectedwith GH (Fig. 7A), ACTH (Fig. 7B), and TSH� (Fig. 7C). Ina number of cells, some weak double staining of �1-AR-ir andPRL-ir was observed (leading to weak orange stain) (Fig. 7,G–J), but upon careful examination, we concluded that theorange color was due to bleed through of the red channel inthe green one, which was confirmed by the finding that�1-AR-ir cells never had the morphology of these lactotrophs.Therefore, we conclude that no convincing colocalization of�1-AR with lactotrophs was present. Interestingly, �1-AR-ircells were found adjacent to PRL-ir cells, and these lac-totrophs were often cup shaped (Fig. 7, G–J).
FIG. 5. Effects of ISO on �1-AR (A), �2-AR (B), and GH (C) mRNAlevels in anterior pituitary cell aggregates cultured with/without DEX(20 nM). Data are expressed as the percentage of control values (setto 100%). Three-way GLM ANOVA was performed on log-transformeddata: ***/**/*, significantly different from corresponding control con-dition with P � 0.001/P � 0.01/P � 0.05; and ###/##/#, significantlydifferent from vehicle (ascorbic acid) with P � 0.001/P � 0.01/P � 0.05.
2318 Endocrinology, May 2008, 149(5):2313–2324 Janssens et al. • �1-AR in Rat Pituitary Gonadotrophs
�1- and �2-AR mRNA is detectable in pituitary cell lines,with highest levels in L�T2 and �T3-1 cells
To confirm the cellular distribution of �1-AR mRNA, weinvestigated the presence and level of �1- and �2-AR in var-
FIG. 7. Confocal microscopic photomicrographs of double-immuno-fluorescent staining of Vibratome sections of intact rat anterior pi-tuitary tissue, stained for �1-AR (green) and anterior pituitary hor-mones (red). Nuclei stained with ToPro-3 (blue). A, GH. B, POMC. C,TSH�. D, �GSU. E and F, LH�. G–J, PRL (arrows indicate cup-shapedlactotrophs). Bar, 10 �m.
�g protein loaded, DEX treated. Lane 4, A total of 33.3 �g, control.Lane 5, Fifty microgram protein loaded, DEX treated. Lane 6, Fiftymicrograms, control. B, Sixty microgram protein loaded in all lanes.Lanes 1 and 2, After preadsorption (Pre-abs) of antiserum with 33nmol immune peptide. Lanes 3 and 4, After preadsorption of anti-serum with 10 nmol immune peptide. Lanes 5 and 6, No preadsorp-tion. Lanes 1, 3, and 5, L�T2 control. Lanes 2, 4, and 6, �T3-1 control.C, Sixty microgram protein loaded in all lanes. Lane 1, �T3-1 control.Lane 2, L�T2 control. Lane 3, GHFT DEX treated. Lane 4, GHFTcontrol. Lane 5, GH3 DEX treated. Lane 6, GH3 control. MW, Mo-lecular weight.
FIG. 6. Western blot of �1-AR in membrane extracts from anteriorpituitary cell aggregates (A) and pituitary cell lines (B and C), eitherin steroid-free [control (CTRL)] condition or treated with 80 nM DEXduring culture (DEX). Anti-PEX14P was used to check equal loadingof the samples. For pituitary aggregates 50 �G protein correspondedto approximately 3 � 106 cells. A (lane 1), Seventy-five microgramprotein loaded, DEX treated [primary antibody (Ab) omitted as neg-ative control]. Lane 2, Seventy-five microgram protein loaded, control(primary antibody omitted as negative control). Lane 3, A total of 33.3
Janssens et al. • �1-AR in Rat Pituitary Gonadotrophs Endocrinology, May 2008, 149(5):2313–2324 2319

ious pituitary cell lines representative for the different pitu-itary cell types. As shown in Table 2, �1-AR mRNA showedlow levels in TtT/GF (FS cells) and GHFT cells (progenitorsomatotrophs/lactotrophs), midrange levels in L�T2 cells(gonadotrophs) (factor 2 compared with GHFT cells), andhigh levels in �T3-1 cells (gonadotroph precursor) (factor 8compared with L�T2 cells). No expression was found inAtT20 cells (corticotrophs). Levels of �2-AR mRNA differedaccording to cell line as well. Low levels were found in AtT20cells, midrange in GHFT (factor 5 compared with AtT20), andhigher levels in L�T2 and �T3-1 cells (factor 2 compared withGHFT). For both ARs expression was not markedly affectedby DEX (80 nm).
DEX-dependent �1-AR mRNA is confined to asubpopulation of small cells
Because no effect of DEX on �1-AR mRNA level was foundin the cell lines, we hypothesized that the up-regulation of�1-AR mRNA level by DEX in pituitary cell aggregates doesnot occur in gonadotrophs but in another cell type or that itis conferred by another cell type on the gonadotrophs. To testthe latter hypothesis, anterior pituitary cells were fraction-ated into subpopulations, enriched in particular cell types, byvelocity sedimentation at unit gravity through a BSA gra-dient, and subsequently established in aggregate cell cultureand challenged with DEX. We have previously shown thatcells of the top fractions of the gradient are small sized, andconsist mainly of lactotrophs and nonhormonal cells,whereas cells in the bottom fraction of the gradient are largeand enriched in somatotrophs and gonadotrophs (32, 33). Inthe present study, the proportional distribution of the celltypes was verified after 5-d aggregate cell culture by detec-tion of GH, PRL, POMC, and �GSU mRNA in individual cells
after redispersion of the aggregates and fluorescence-acti-vated cell sorting (see Materials and Methods). As shown inTable 3, fraction 7 cells consisted mainly of GH cells (totalnumber of GH mRNA-positive cells: 72%), �GSU cells (12%),whereas fraction 3 cells contained mainly PRL cells (65%) andnonhormonal cells (25%).
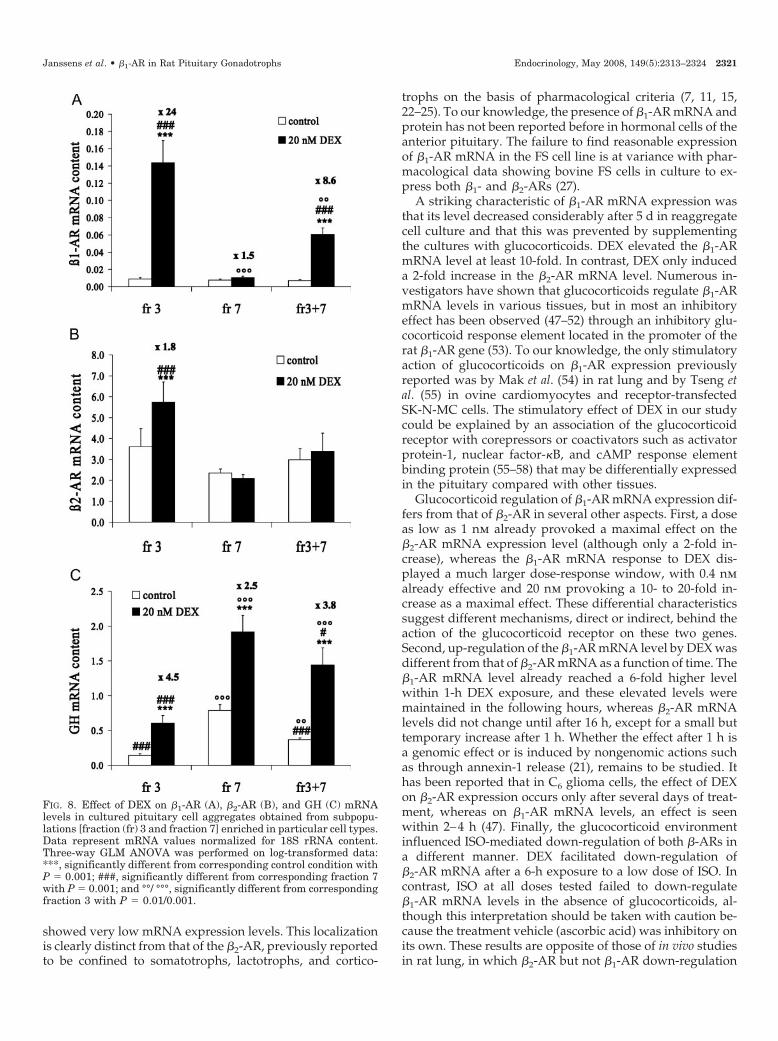
As shown in Fig. 8, in the absence of DEX, absolute �1- and�2-AR mRNA levels within fraction 3 and fraction 7 aggre-gates were comparable. Supplementation with 20 nm DEXcaused a 24-fold increase in the �1-AR mRNA content infraction 3 cell aggregates (Fig. 8A) but was ineffective inaggregates prepared from the bottom fraction 7. Coaggre-gation of fraction 3 and fraction 7 cells in a ratio 3:1 (thatresembles the original ratio in the initial cell population)resulted in a DEX response of �1-AR mRNA that was weak-ened (8.6-fold increase in �1-AR mRNA content) comparedwith that found in fraction 3 cells (Fig. 8A, right panel). Therewas a small effect of DEX on the �2-AR mRNA level infraction 3 (only 1.8-fold increase) (Fig. 8B). Coaggregationabolished that small effect of DEX on �2-AR (Fig. 8B, rightpanel). In contrast, DEX treatment increased GH mRNA lev-els in both fractions 3 and 7, but to a larger extent in fraction3 than in fraction 7 cell aggregates (4.5- and 2.5-fold increase,respectively) (Fig. 8C). In the coaggregates DEX augmentedGH mRNA levels with a magnitude similar to that seen infraction 3 (3.8-fold increase).
Discussion
The main finding of the present investigation is that theanterior pituitary expresses �1-AR mRNA and protein in aglucocorticoid-regulated manner, and that expression isfound in a subpopulation of gonadotrophs. No colocalizationwas seen with somatotrophs, corticotrophs, thyrotrophs, andlactotrophs. In confirmation, expression of �1-AR, at themRNA and protein level, was also found in two gonadotrophcell lines, i.e. the gonadotroph precursor �T3-1 and the moremature L�T2 cells, whereas the GHFT (somatotroph precur-sor), GH3 (lactosomatotrophs) cells, and TtT/GF cells (FS)
TABLE 2. ß1- and ß2-AR mRNA levels, normalized to 18S rRNA,in mouse pituitary cell lines cultured either in steroid-free mediumor in medium with 80 nM DEX
�1-AR �2-AR
A�T3-1 control 7.80 � 1.39 59.8 � 10.13�T3-1 DEX 6.21 � 0.94 48.12 � 2.52AtT20 control 4.76 � 2.14AtT20 DEX 2.88 � 1.13L�T2 control 0.92 � 0.25 41.69 � 2.06L�T2 DEX 1.52 � 0.28 35.01 � 3.59GHFT control 0.58 � 0.04 26.66 � 0.97GHFT DEX 0.49 � 0.20 19.42 � 7.43
B�T3-1 control 7.60 � 0.64 29.98 � 2.67�T3-1 DEX 6.27 � 1.80 21.86 � 10.17L�T2 control 1.03 � 0.27 17.99 � 0.25L�T2 DEX 1.47 � 0.46 18.75 � 4.01GHFT control 0.49 � 0.03 7.53 � 0.92GHFT DEX 0.41 � 0.04 7.52 � 2.76TtT/GF control �0.06 6.87 � 1.52TtT/GF DEX �0.03 8.08 � 1.54
Data are means � SEM of three independent experiments, exceptGHFT cells that were analyzed only twice. All experiments in A wererun and analyzed simultaneously. The experiments with TtT/GF cellsshown in B represent three experiments executed separately fromthose in A, but the samples from A were reanalyzed to be able tocompare expression in TtT/GF with expression in the other cell lines.No expression was found in AtT20 cells.
TABLE 3. Proportions of hormonal cells identified on the basis ofhormone mRNA content detected by single-cell RT-PCR in anteriorpituitary aggregates prepared from separated subpopulations andcultured in the absence or presence of 20 nM DEX
Hormone mRNASeparated population
Fraction3
Fraction3 DEX
Fraction7
Fraction7 DEX
GH 3.5 5.3 68.1 72.7PRL 57.6 63.3 5.0 1.4�GSU 0 0.5 10.6 9.1POMC 6.4 2.1 4.3 3.5GH � PRL 4.6 9.0 0.7 0.7GH � PRL � �GSU 0 0.5 0 0GH � �GSU 0 0 1.4 2.8PRL � �GSU 1.7 0.5 0 1.4�GSU � POMC 0 0 0 0.7PRL � POMC 1.2 1.1 1.4 0GH � POMC 0 0 1.4 1.4No hormone 25.0 17.5 7.1 6.3
Nonhormonal cells expressed none of the pituitary hormonemRNAs but expressed L19 mRNA. L19 mRNA is a housekeeping genemRNA, detected as a control for the presence of a cell. Data are from140–190 cells in each population.
2320 Endocrinology, May 2008, 149(5):2313–2324 Janssens et al. • �1-AR in Rat Pituitary Gonadotrophs
showed very low mRNA expression levels. This localizationis clearly distinct from that of the �2-AR, previously reportedto be confined to somatotrophs, lactotrophs, and cortico-
trophs on the basis of pharmacological criteria (7, 11, 15,22–25). To our knowledge, the presence of �1-AR mRNA andprotein has not been reported before in hormonal cells of theanterior pituitary. The failure to find reasonable expressionof �1-AR mRNA in the FS cell line is at variance with phar-macological data showing bovine FS cells in culture to ex-press both �1- and �2-ARs (27).
A striking characteristic of �1-AR mRNA expression wasthat its level decreased considerably after 5 d in reaggregatecell culture and that this was prevented by supplementingthe cultures with glucocorticoids. DEX elevated the �1-ARmRNA level at least 10-fold. In contrast, DEX only induceda 2-fold increase in the �2-AR mRNA level. Numerous in-vestigators have shown that glucocorticoids regulate �1-ARmRNA levels in various tissues, but in most an inhibitoryeffect has been observed (47–52) through an inhibitory glu-cocorticoid response element located in the promoter of therat �1-AR gene (53). To our knowledge, the only stimulatoryaction of glucocorticoids on �1-AR expression previouslyreported was by Mak et al. (54) in rat lung and by Tseng etal. (55) in ovine cardiomyocytes and receptor-transfectedSK-N-MC cells. The stimulatory effect of DEX in our studycould be explained by an association of the glucocorticoidreceptor with corepressors or coactivators such as activatorprotein-1, nuclear factor-�B, and cAMP response elementbinding protein (55–58) that may be differentially expressedin the pituitary compared with other tissues.
Glucocorticoid regulation of �1-AR mRNA expression dif-fers from that of �2-AR in several other aspects. First, a doseas low as 1 nm already provoked a maximal effect on the�2-AR mRNA expression level (although only a 2-fold in-crease), whereas the �1-AR mRNA response to DEX dis-played a much larger dose-response window, with 0.4 nmalready effective and 20 nm provoking a 10- to 20-fold in-crease as a maximal effect. These differential characteristicssuggest different mechanisms, direct or indirect, behind theaction of the glucocorticoid receptor on these two genes.Second, up-regulation of the �1-AR mRNA level by DEX wasdifferent from that of �2-AR mRNA as a function of time. The�1-AR mRNA level already reached a 6-fold higher levelwithin 1-h DEX exposure, and these elevated levels weremaintained in the following hours, whereas �2-AR mRNAlevels did not change until after 16 h, except for a small buttemporary increase after 1 h. Whether the effect after 1 h isa genomic effect or is induced by nongenomic actions suchas through annexin-1 release (21), remains to be studied. Ithas been reported that in C6 glioma cells, the effect of DEXon �2-AR expression occurs only after several days of treat-ment, whereas on �1-AR mRNA levels, an effect is seenwithin 2–4 h (47). Finally, the glucocorticoid environmentinfluenced ISO-mediated down-regulation of both �-ARs ina different manner. DEX facilitated down-regulation of�2-AR mRNA after a 6-h exposure to a low dose of ISO. Incontrast, ISO at all doses tested failed to down-regulate�1-AR mRNA levels in the absence of glucocorticoids, al-though this interpretation should be taken with caution be-cause the treatment vehicle (ascorbic acid) was inhibitory onits own. These results are opposite of those of in vivo studiesin rat lung, in which �2-AR but not �1-AR down-regulation
FIG. 8. Effect of DEX on �1-AR (A), �2-AR (B), and GH (C) mRNAlevels in cultured pituitary cell aggregates obtained from subpopu-lations [fraction (fr) 3 and fraction 7] enriched in particular cell types.Data represent mRNA values normalized for 18S rRNA content.Three-way GLM ANOVA was performed on log-transformed data:***, significantly different from corresponding control condition withP � 0.001; ###, significantly different from corresponding fraction 7with P � 0.001; and °°/ °°°, significantly different from correspondingfraction 3 with P � 0.01/0.001.
Janssens et al. • �1-AR in Rat Pituitary Gonadotrophs Endocrinology, May 2008, 149(5):2313–2324 2321

could be prevented by DEX (54). This suggests tissue-specificregulation by glucocorticoids.
Immunoblotting revealed the presence of �1-AR immu-noreactive protein of the expected size (64 kDa) in membraneextracts of anterior pituitary cell aggregate cultures. Estima-tion of the protein amount in the bands indicated only 1.5–3times more protein in DEX-cultured compared with controlaggregates, which is a magnitude considerably smaller thanthat induced by DEX at the mRNA level. Because expressionof �1-AR-ir itself was modest on Western blot, the smallchanges induced by DEX should be taken with caution. Thesmall magnitude of the DEX effect at the protein level maybe trivial because the level of �1-AR mRNA in steroid-freemedium could decrease faster than that of �1-AR protein.This would result in a seemingly smaller proportional effectof DEX on the protein level than on the mRNA level. Alter-natively, it is possible that DEX increases the �1-AR mRNAlevel without (or with limited) translation into protein. It isreasonable to suspect that other regulatory factors may berequired to further increase �1-AR protein expression at thetranslational level and that these are perhaps not present inthe pituitary aggregate cell cultures.
Virtually all �1-AR-ir material was detected in the cyto-plasmic compartments of relatively large cells with a roundto oval morphology, and not at the cell membrane or in theperipheral areas of the cells. The lack of immunostaining atthe plasma membrane may be due to diminished ir as aconsequence of the folding of the protein into the membraneor of binding of regulatory proteins, particularly because theantibody was raised against a peptide from the C-terminalpart of the protein. Cytoplasmic staining of the �1-AR hasbeen described also in other tissues (59).
The finding of �1-AR expression in gonadotrophs is instriking contrast to the evidence that the up-regulation of�1-AR mRNA levels by DEX was apparently not located inthe gonadotrophs. In a subpopulation of cells, separated bysedimentation at unit gravity and relatively enriched in largegonadotrophs (fraction 7), DEX had no effect on �1-ARmRNA levels, but in a subpopulation poor in gonadotrophs(fraction 3), DEX caused a vast up-regulation (24-fold in-crease) of the �1-AR mRNA level. Fraction 3 contains, besideslactotrophs, FS cells (60) and other nonhormonal cells (39, 61,62) as major populations (Table 3), whereas corticotrophs,thyrotrophs, and somatotrophs are poorly represented. Noevidence for an involvement of lactotrophs and FS cells wasfound because no �1-AR expression was found in lactotrophsor the lactotroph-derived GH3 cells (63) (and this study).Furthermore, only a very low to negligible �1-AR mRNAlevel was found in the FS cell line TtT/GF, of which expres-sion was not inducible by glucocorticoids. There was also noeffect of DEX found on the �1-AR mRNA level in the gona-dotrophic cell lines �T3-1 and L�T2. Thus, it seems that theDEX-induced �1-AR mRNA up-regulation found in the smallcell population (fraction 3) is located in an unknown cell type.A possibility is that these small cells are nonhormonal cellsthat separate in the “side population” (SP) (64). In two in-dependent experiments, we have been able to detect �1-ARmRNA by RT-PCR in a collected population of mouse pi-tuitary SP cells (Vankelecom, H., unpublished observations).Moreover, three independent microarray analysis experi-
ments performed on mouse pituitary SP cells and cells fromthe main population showed statistically significant expres-sion of �1-AR mRNA in both populations (Vankelecom, H.,unpublished observations). The SP enriches in the upperfractions of the unit gravity sedimentation gradient (rat)(Vankelecom, H., unpublished observations). These cells ex-press stem/progenitor cell and early embryonic markers(64). Thus, also nonhormonal cells seem to express �1-AR, atleast at the mRNA level. Because in our immunofluorescentstudies only gonadotrophs were immunopositive for �1-AR,it looks plausible that DEX up-regulates �1-AR mRNA in anonhormonal cell type in fraction 3 without �1-AR mRNAsufficiently translated into �1-AR protein. Perhaps these cellsare committed gonadotroph progenitors. Alternatively, theaction of DEX may occur in a nonhormonal cell type offraction 3 that in turn up-regulates �1-AR in gonadotrophsthrough a paracrine mechanism. In an attempt to test thelatter hypothesis, fraction 3 cells were coaggregated withfraction 7 cells and the coaggregates examined for their re-sponsiveness to DEX. It was found that fraction 3 cells did notconfer the DEX responsiveness of the �1-AR to the fraction7 cells, whereas the magnitude of the GH mRNA responseto DEX seen in fraction 3 cells was indeed transduced to thefraction 7 cells. Unfortunately, we cannot exclude that frac-tion 7 cells may have exerted a negative feedback on thefraction 3 cells in their putative action on �1-AR expressionin gonadotrophs.
A remarkable feature of the �1-AR positive cells was thatthey are located adjacent to lactotrophs. These lactotrophswere often found in clusters surrounding the �1-AR positivecells, and many had a cup-shaped morphology. The sub-population of cup-shaped lactotrophs that is in close asso-ciation with gonadotrophs has been described in the rat (65,66) as well as in other species (67–69), and the selectiveassociation is reconstituted in aggregate cell culture (35).Most interestingly, also calcitonin, a paracrine factor thatdepresses PRL release and gene expression, has been locatedin gonadotrophs that are surrounded by cup-shaped lac-totrophs (70). Therefore, the present findings can be relatedto earlier findings in our laboratory (33) and to that of others(for review, see Ref. 71) that gonadotrophs exert trophicactions on lactotrophs. Further support to the latter hypoth-esis is given in a companion paper (72), in which we showthat in pituitary cell aggregates, the �1-AR is constitutivelycoupled to adenylate cyclase in a pertussis toxin-sensitivemanner. The unliganded receptor constitutively activatescAMP formation, which in turn may activate communicationbetween gonadotrophs and lactotrophs.
In conclusion, the present study shows expression of�1-AR in a subpopulation of gonadotrophs on the basis ofmRNA detection, Western blot, double immunostaining, andspecific cell line analysis. �1-AR mRNA expression is up-regulated by glucocorticoids, but this does not seem to occurin the main gonadotroph cell population itself. Whether thisoccurs within a subpopulation of small nonhormonal cells,or is transduced by the latter cells upon gonadotrophs, re-mains to be studied further. Because �1-AR-positive cells areoften surrounded by cup-shaped lactotrophs, �1-AR mayplay a role in intracrine regulation in gonadotrophs or inparacrine regulation between gonadotrophs and lactotrophs.
2322 Endocrinology, May 2008, 149(5):2313–2324 Janssens et al. • �1-AR in Rat Pituitary Gonadotrophs

The present findings suggest a potential interference of theadrenergic system with the paracrine network in the anteriorpituitary (71), which may be of relevance in the context of thenegative influences of stress on reproduction.
Acknowledgments
We thank Dr. A. F. Parlow (Harbor-University of California LosAngeles Medical Center, Torrance, CA) for providing guinea pig anti-sera against rat glycoprotein hormone �-subunit, LH�, TSH�, GH, andprolactin. We also thank M. Boussemaere, K. Rillaerts, and Y. VanGoethem for their skillful technical assistance.
Received October 11, 2007. Accepted January 10, 2008.Address all correspondence and requests for reprints to: Professor
Carl Denef, Laboratory of Cell Pharmacology, University of Leuven,Medical School, Campus Gasthuisberg (O & N), B-3000 Leuven, Bel-gium. E-mail: [email protected].
This work was supported by grants from the Flemish Ministry of SciencePolicy (Concerted Research Actions) and the Fund for Scientific ResearchFlanders (Belgium) (Fonds voor Wetenschappelijk Onderzoek).
Disclosure Statement: The authors have nothing to disclose.
References
1. Goodman LS, Hardman JG, Limbird LE, Gilman AG 2001 Goodman &Gilman’s the pharmacological basis of therapeutics. New York: McGraw-HillMedical Publishing Division
2. Kalantaridou SN, Makrigiannakis A, Zoumakis E, Chrousos GP 2004 Stressand the female reproductive system. J Reprod Immunol 62:61–68
3. Tsigos C, Chrousos GP 2002 Hypothalamic-pituitary-adrenal axis, neuroen-docrine factors and stress. J Psychosom Res 53:865–871
4. al-Damluji S 1993 Adrenergic control of the secretion of anterior pituitaryhormones. Baillieres Clin Endocrinol Metab 7:355–392
5. Alarcon P, Nunez L, Garcia-Sancho J 2001 Direct actions of adrenergic agentson rat anterior pituitary cells. Pflugers Arch 442:834–841
6. Giguere V, Cote J, Labrie F 1981 Characteristics of the �-adrenergic stimu-lation of adrenocorticotropin secretion in rat anterior pituitary cells. Endocri-nology 109:757–762
7. Mezey E, Reisine TD, Palkovits M, Brownstein MJ, Axelrod J 1983 Directstimulation of �2-adrenergic receptors in rat anterior pituitary induces therelease of adrenocorticotropin in vivo. Proc Natl Acad Sci USA 80:6728–6731
8. Giguere V, Labrie F 1983 Additive effects of epinephrine and corticotropin-releasing factor (CRF) on adrenocorticotropin release in rat anterior pituitarycells. Biochem Biophys Res Commun 110:456–462
9. Boyle M, Yamamoto G, Chen M, Rivier J, Vale W 1988 Interleukin 1 preventsloss of corticotropic responsiveness to �-adrenergic stimulation in vitro. ProcNatl Acad Sci USA 85:5556–5560
10. Swennen L, Baes M, Schramme C, Denef C 1985 Beta-adrenergic stimulationof adenosine-3�,5�-monophosphate (c-AMP) in primary cultures of rat anteriorpituitary cell populations separated by unit gravity sedimentation. Relation-ship to growth hormone and prolactin release and to nonsecreting cells. Neu-roendocrinology 40:78–83
11. Swennen L, Baes M, Denef C 1985 Beta-adrenergic stimulation of adenosine-3�,5�-monophosphate (c-AMP) accumulation and of prolactin and growth hor-mone secretion in rat anterior pituitary cell cultures. Neuroendocrinology40:72–77
12. Perkins SN, Evans WS, Thorner MO, Cronin MJ 1983 �-Adrenergic stimu-lation of growth hormone release from perifused rat anterior pituitary cells.Neuroendocrinology 37:473–475
13. Baes M, Denef C 1987 Evidence that stimulation of growth hormone releaseby epinephrine and vasoactive intestinal peptide is based on cell-to-cell com-munication in the pituitary. Endocrinology 120:280–290
14. Maertens P, Denef C 1987 �-Adrenergic stimulation of growth hormonerelease in perifused rat anterior pituitary reaggregate cell cultures. Mol CellEndocrinol 54:203–211
15. Denef C, Baes M 1982 �-Adrenergic stimulation of prolactin release fromsuperfused pituitary cell aggregates. Endocrinology 111:356–358
16. Peters JR, Foord SM, Dieguez C, Scanlon MF, Hall R 1983 �1-Adrenorecep-tors on intact rat anterior pituitary cells: correlation with adrenergic stimula-tion of thyrotropin secretion. Endocrinology 113:133–140
17. Petrovic SL, Bedran de Castro JC, McCann SM 1987 �-Adrenergic blockersdecrease levels of luteinizing hormone in plasma of orchidectomized rats. BiolReprod 37:539–545
18. Petrovic SL, Bedran de Castro JC, McCann SM 1986 �-Adrenergic agonistsincrease amplitude of LH release in orchidectomized rats. Am J Physiol 251(3Pt 1):E316–E321
19. Lefkowitz RJ 2007 Seven transmembrane receptors: something old, somethingnew. Acta Physiol (Oxf) 190:9–19
20. Collins S, Caron MG, Lefkowitz RJ 1988 �-Adrenergic receptors in hamstersmooth muscle cells are transcriptionally regulated by glucocorticoids. J BiolChem 263:9067–9070
21. John CD, Christian HC, Morris JF, Flower RJ, Solito E, Buckingham JC 2004Annexin 1 and the regulation of endocrine function. Trends Endocrinol Metab15:103–109
22. Petrovic SL, McDonald JK, Snyder GD, McCann SM 1983 Characterizationof �-adrenergic receptors in rat brain and pituitary using a new high-affinityligand, [125I]iodocyanopindolol. Brain Res 261:249–259
23. Baes M, Denef C 1984 �2-Receptors in the rat anterior pituitary mediateadrenergic stimulation of prolactin release. Life Sci 34:1447–1454
24. De Souza EB 1985 �-2-Adrenergic receptors in pituitary. Identification, char-acterization, and autoradiographic localization. Neuroendocrinology 41:289–296
25. Perkins SN, Evans WS, Thorner MO, Gibbs DM, Cronin MJ 1985 �-Adren-ergic binding and secretory responses of the anterior pituitary. Endocrinology117:1818–1825
26. Gabriel SM, Milbury CM, Alexander SM, Nathanson JA, Martin JB 1989 Isostimulation of GH and cAMP: comparison of �-adrenergic- to GRF-stimulatedGH release and cAMP accumulation in monolayer cultures of anterior pitu-itary cells in vitro. Neuroendocrinology 50:170–176
27. Findell PR, Weiner RI 1988 Bovine pituitary folliculo-stellate cells have �-ad-renergic receptors positively coupled to adenosine 3�,5�-cyclic monophosphateproduction. Endocrinology 123:2454–2461
28. Bilezikjian LM, Blount AL, Leal AM, Donaldson CJ, Fischer WH, Vale WW2004 Autocrine/paracrine regulation of pituitary function by activin, inhibinand follistatin. Mol Cell Endocrinol 225:29–36
29. McDuffie IA, Akhter N, Childs GV 2004 Regulation of leptin mRNA andprotein expression in pituitary somatotropes. J Histochem Cytochem 52:263–273
30. Denef C, Maertens P, Allaerts W, Mignon A, Robberecht W, Swennen L,Carmeliet P 1989 Cell-to-cell communication in peptide target cells of anteriorpituitary. Methods Enzymol 168:47–71
31. Langouche L, Hersmus N, Papageorgiou A, Vankelecom H, Denef C 2004Melanocortin peptides stimulate prolactin gene expression and prolactin ac-cumulation in rat pituitary aggregate cell cultures. J Neuroendocrinol 16:695–703
32. Denef C, Hautekeete E, De Wolf A, Vanderschueren B 1978 Pituitary ba-sophils from immature male and female rats: distribution of gonadotrophs andthyrotrophs as studied by unit gravity sedimentation. Endocrinology 103:724–735
33. Denef C, Andries M 1983 Evidence for paracrine interaction between gona-dotrophs and lactotrophs in pituitary cell aggregates. Endocrinology 112:813–822
34. Allaerts W, Denef C 1989 Regulatory activity and topological distribution offolliculo-stellate cells in rat anterior pituitary cell aggregates. Neuroendocri-nology 49:409–418
35. Allaerts W, Mignon A, Denef C 1991 Selectivity of juxtaposition betweencup-shaped lactotrophs and gonadotrophs from rat anterior pituitary in cul-ture. Cell Tissue Res 263:217–225
36. Denef C, Hautekeete E, Rubin L 1976 A specific population of gonadotrophspurified from immature female rat pituitary. Science 194:848–851
37. Pals K, Vankelecom H, Denef C 2006 Triiodothyronine expands the lactotrophand maintains the lactosomatotroph population, whereas thyrotrophin-releas-ing hormone augments thyrotroph abundance in aggregate cell cultures ofpostnatal rat pituitary gland. J Neuroendocrinol 18:203–216
38. Pals K, Boussemaere M, Swinnen E, Vankelecom H, Denef C 2006 A pituitarycell type coexpressing messenger ribonucleic acid of proopiomelanocortin andthe glycoprotein hormone �-subunit in neonatal rat and chicken: rapid declinewith age and reappearance in vitro under regulatory pressure of corticotropin-releasing hormone in the rat. Endocrinology 147:4738–4752
39. Krylyshkina O, Chen J, Mebis L, Denef C, Vankelecom H 2005 Nestin-immunoreactive cells in rat pituitary are neither hormonal nor typical folliculo-stellate cells. Endocrinology 146:2376–2387
40. Boulanger L, Andersen PH, Gaudreau P 1999 Development of a site-directedpolyclonal antibody against the pituitary growth hormone-releasing hormonereceptor and its use to estimate GHRH receptor concentration in normal andhypothyroid rats. Neuroendocrinology 70:117–127
41. Amery L, Fransen M, De Nys K, Mannaerts GP, Van Veldhoven PP 2000Mitochondrial and peroxisomal targeting of 2-methylacyl-CoA racemase inhumans. J Lipid Res 41:1752–1759
42. Lu J, Swinnen E, Proost P, De Vreese B, Vankelecom H, Denef C 2002Isolation and structure-bioactivity characterization of glycosylated N-pro-opi-omelanocortin isoforms. J Neuroendocrinol 14:869–879
43. Martinoli MG, Veilleux R, Pelletier G 1991 Effects of triiodothyronine, dexa-methasone and estradiol-17� on GH mRNA in rat pituitary cells in culture asrevealed by in situ hybridization. Acta Endocrinol (Copenh) 124:83–90
44. Treacy MN, Ryan F, Martin F 1991 Functional glucocorticoid inducible en-hancer activity in the 5�-flanking sequences of the rat growth hormone gene.J Steroid Biochem Mol Biol 38:1–15
Janssens et al. • �1-AR in Rat Pituitary Gonadotrophs Endocrinology, May 2008, 149(5):2313–2324 2323

45. Johnson M 1998 The �-adrenoceptor. Am J Respir Crit Care Med 158(5 Pt3):S146–S153
46. Bahouth SW 1992 Effects of chemical and surgical sympathectomy on ex-pression of �-adrenergic receptors and guanine nucleotide-binding proteins inrat submandibular glands. Mol Pharmacol 42:971–981
47. Kiely J, Hadcock JR, Bahouth SW, Malbon CC 1994 Glucocorticoids down-regulate �1-adrenergic-receptor expression by suppressing transcription of thereceptor gene. Biochem J 302(Pt 2):397–403
48. Zhong H, Minneman KP 1993 Close reciprocal regulation of �1- and �2-adrenergic receptors by dexamethasone in C6 glioma cells: effects on cate-cholamine responsiveness. Mol Pharmacol 44:1085–1093
49. Zhong H, Minneman KP 1995 Coexisting �-1 and �-2-adrenergic receptorswith differential receptor reserves in rat C6 glioma cells. J Pharmacol Exp Ther272:1088–1094
50. Dangel V, Giray J, Ratge D, Wisser H 1996 Regulation of beta-adrenoceptordensity and mRNA levels in the rat heart cell-line H9c2. Biochem J 317(Pt3):925–931
51. Feve B, Emorine LJ, Briend-Sutren MM, Lasnier F, Strosberg AD, PairaultJ 1990 Differential regulation of �1- and �2-adrenergic receptor protein andmRNA levels by glucocorticoids during 3T3-F442A adipose differentiation.J Biol Chem 265:16343–16349
52. Guest SJ, Hadcock JR, Watkins DC, Malbon CC 1990 �1- and �2-Adrenergicreceptor expression in differentiating 3T3-L1 cells. Independent regulation atthe level of mRNA. J Biol Chem 265:5370–5375
53. Bahouth SW, Park EA, Beauchamp M, Cui X, Malbon CC 1996 Identificationof a glucocorticoid repressor domain in the rat �1-adrenergic receptor gene.Recept Signal Transduct 6:141–149
54. Mak JC, Nishikawa M, Shirasaki H, Miyayasu K, Barnes PJ 1995 Protectiveeffects of a glucocorticoid on downregulation of pulmonary �2-adrenergicreceptors in vivo. J Clin Invest 96:99–106
55. Tseng YT, Stabila JP, Nguyen TT, McGonnigal BG, Waschek JA, PadburyJF 2001 A novel glucocorticoid regulatory unit mediates the hormone respon-siveness of the �1-adrenergic receptor gene. Mol Cell Endocrinol 181:165–178
56. Barnes PJ 1998 Anti-inflammatory actions of glucocorticoids: molecular mech-anisms. Clin Sci (Lond) 94:557–572
57. Kassel O, Herrlich P 2007 Crosstalk between the glucocorticoid receptor andother transcription factors: molecular aspects. Mol Cell Endocrinol 275:13–29
58. Schoneveld OJ, Gaemers IC, Lamers WH 2004 Mechanisms of glucocorticoidsignalling. Biochim Biophys Acta 1680:114–128
59. Thorlin T, Anders P, Persson I, Eriksson PS, Ronnback L, Hansson E 2000Astrocyte �1-adrenergic receptor immunoreactivity and agonist induced in-creases in [Ca2�]i: differential results indicative of a modified membranereceptor. Life Sci 67:1285–1296
60. Baes M, Allaerts W, Denef C 1987 Evidence for functional communicationbetween folliculo-stellate cells and hormone-secreting cells in perifused an-terior pituitary cell aggregates. Endocrinology 120:685–691
61. Tilemans D, Vijver VV, Verhoeven G, Denef C 1995 Production of trans-ferrin-like immunoreactivity by rat anterior pituitary and intermediate lobe.J Histochem Cytochem 43:657–664
62. Vankelecom H, Matthys P, Denef C 1997 Inducible nitric oxide synthase inthe anterior pituitary gland: induction by interferon-� in a subpopulation offolliculostellate cells and in an unidentifiable population of non-hormone-secreting cells. J Histochem Cytochem 45:847–857
63. Guerrero SW, Minneman KP 1999 Coupling efficiencies of �1- and �2-ad-renergic receptors expressed alone or together in transfected GH3 pituitarycells. J Pharmacol Exp Ther 290:980–988
64. Chen J, Hersmus N, Van Duppen V, Caesens P, Denef C, Vankelecom H 2005The adult pituitary contains a cell population displaying stem/progenitor celland early embryonic characteristics. Endocrinology 146:3985–3998
65. Nakane PK 1970 Classifications of anterior pituitary cell types with immu-noenzyme histochemistry. J Histochem Cytochem 18:9–20
66. Sato S 1980 Postnatal development, sexual difference and sexual cyclic vari-ation of prolactin cells in rats: special reference to the topographic affinity togonadotroph. Endocrinol Jpn 27:573–583
67. Gregory SJ, Brooks J, McNeilly AS, Ingleton PM, Tortonese DJ 2000 Gona-dotroph-lactotroph associations and expression of prolactin receptors in theequine pituitary gland throughout the seasonal reproductive cycle. J ReprodFertil 119:223–231
68. Oguchi A, Tanaka S, Aida T, Yamamoto K, Kikuyama S 1997 Enhancementby prolactin of the GnRH-induced release of LH from dispersed anteriorpituitary cells of the bullfrog (Rana catesbeiana). Gen Comp Endocrinol 107:128–135
69. Tortonese DJ, Brooks J, Ingleton PM, McNeilly AS 1998 Detection of pro-lactin receptor gene expression in the sheep pituitary gland and visualizationof the specific translation of the signal in gonadotrophs. Endocrinology 139:5215–5223
70. Shah GV, Wang W, Grosvenor CE, Crowley WR 1990 Calcitonin inhibits basaland thyrotropin-releasing hormone-induced release of prolactin from anteriorpituitary cells: evidence for a selective action exerted proximal to secretagogue-induced increases in cytosolic Ca2�. Endocrinology 127:621–628
71. Denef C 2008 Paracrinicity: the story of 30 years of cellular pituitary crosstalk.J Neuroendocrinol 20:1–70
72. Janssens K, Boussemaere M, Wagner S, Kopka K, Denef C 2008 �1-Adre-noceptors receptors in rat anterior pituitary may be constitutively active.Inverse agonism of CGP 20712A on basal 3',5'-cyclic adenosine 5'-monophos-phate levels. Endocrinology 149:2391–2402
Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremost professional society serving theendocrine community.
2324 Endocrinology, May 2008, 149(5):2313–2324 Janssens et al. • �1-AR in Rat Pituitary Gonadotrophs
Copyright © 2022 FDOKUMEN




















