
2-adrenoceptor blocker synergizes with gemcitabine to inhibit the proliferation of pancreatic cancer...
7
Molecular and Cellular Pharmacology β2-adrenoceptor blocker synergizes with gemcitabine to inhibit the proliferation of pancreatic cancer cells via apoptosis induction Tao Shan a , Qingyong Ma a, ⁎, Dong Zhang a , Kun Guo a , Han Liu a , Fengfei Wang b , Erxi Wu b a Department of Hepatobiliary Surgery, First Affiliated Hospital of Medical College, Xi'an Jiaotong University, 277 West Yanta Road, Xi'an 710061, Shaanxi Province, China b Department of Pharmaceutical Sciences, NDSU, Sudro Hall 203, Fargo, ND 58105, USA abstract article info Article history: Received 6 December 2010 Received in revised form 18 April 2011 Accepted 20 April 2011 Available online 7 May 2011 Keywords: Gemcitabine β2-adrenoceptor blocker Pancreatic cancer Apoptosis NF-κB Bcl-2 The stimulation of β2-adrenoceptor, which is a major mediator for chronic stress-induced cancers, has been implicated in the progression in the number of cancer cells, including pancreatic cancer, which remains one of the most aggressive and lethal diseases worldwide. Whether β-adrenoceptor antagonists potentiate gemcitabine, a standard first-line treatment for advanced pancreatic cancer that offers only modest benefit due to acquired chemoresistance, has not been elucidated. Thus, we studied the antiproliferative and apoptotic effects and the underlying mechanisms of gemcitabine combined with the β2-adrenoceptor blocker ICI 118551 (1-[2,3-(dihydro-7-methyl-1H-iden-4-yl)oxy]-3-[(1-methylethyl)amino]-2-butanol), in human pancreatic cancer BxPC-3 and MIA PaCa-2 cells. Results show that ICI 118551 significantly synergized the antiproliferative and pro-apoptotic effects induced by gemcitabine in both BxPC-3 and MIA PaCa-2 cells (P b 0.05 combination vs. control or gemcitabine alone). When cells were treated with the combination of gemcitabine and ICI 118551, NF-κB activation was blocked; the expression of Bax protein was substantially increased; and Bcl-2 protein was downregulated. Taken together, the data suggest that ICI 118551 potentiates the antiproliferative effects of gemcitabine by inducing apoptosis in pancreatic cancer cells. Our study implies that this combination may be an effective therapeutic strategy for pancreatic cancer. © 2011 Elsevier B.V. All rights reserved. 1. Introduction Pancreatic cancer remains one of the most aggressive and lethal diseases worldwide, with a 5-year survival rate of less than 5%. Even after curative resection, the 5-year survival rate is only 10% (Fryer et al., 2009; Yokoyama et al., 2009). The poor prognosis of pancreatic cancer is attributable to the late manifestation of symptoms, aggressive local invasion, liable metastases, and weak response to chemotherapy (Duffy et al., 2003). Currently, gemcitabine, a standard first-line treatment for advanced pancreatic cancer, is extensively utilized, but offers only a modest benefit due to acquired chemoresistance and multiple adverse effects (Herrmann et al., 2007; Kullmann et al., 2009; Reddy et al., 2008). Thus, it is imperative that we discover novel strategies to identify less toxic agents that can sensitize pancreatic cancer cells to chemotherapy. Considerable evidence shows that NF-κB plays a major role in promoting gemcitabine resistance in pancreatic cancer (Arlt et al., 2003; Li et al., 2004); therefore, the agents that block NF-κB activation could reduce chemoresistance to gemcitabine and perhaps be used in com- bination with gemcitabine as a novel therapeutic regimen for pancreatic cancer. Recent studies have shown that neurotransmitters are involved in the regulation of the number of cancer cells that progress via β-adrenoceptor, which is the major mediator (Antoni et al., 2006; Hu et al., 2010; Thaker et al., 2006; Thaker et al., 2007; Tilan and Kitlinska, 2010). It has also been reported that β-adrenoceptor blockers could inhibit the regulation by blocking β-adrenoceptor, and suppressing NF-κB, PI3K/Akt, and PKA pathways in cancer cells (Audet and Bouvier, 2008). Previously, we have also shown that β-adrenoceptor antagonists are effective inhibitors of cell proliferation and inducers of cellular apoptosis by regulating the expression of NF-κB downstream gene products (Bax, Bcl-2) in pancreatic cancer cells (Zhang et al., 2009). Whether or not β-adrenoceptor antagonists can potentiate the gemcitabine-induced cytotoxicity by inhibiting NF-κB activation has not been elucidated. In this study, we have investigated the mechanisms involved and the effects of the specific β2-adrenoceptor blocker ICI 118551 in combination with gemcitabine. 2. Materials and methods 2.1. Cell cultures and treatments The MIA PaCa-2 and BxPC-3 human pancreatic cancer cell lines (obtained from the American Tissue Type Collection, USA) were main- tained in Dulbecco's modified Eagle's medium (DMEM, GIBCO, USA) supplemented with penicillin (100 U/ml), streptomycin (100 μg/ml), 0.1 mM nonessential amino acids, 0.2 mM glutamine, 1 mM pyruvate, and 10% heat-inactivated fetal bovine serum (FBS) and incubated in 5% European Journal of Pharmacology 665 (2011) 1–7 ⁎ Corresponding author. Tel./fax: + 86 29 8532 3899. E-mail address: [email protected] (Q. Ma). 0014-2999/$ – see front matter © 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.ejphar.2011.04.055 Contents lists available at ScienceDirect European Journal of Pharmacology journal homepage: www.elsevier.com/locate/ejphar
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of 2-adrenoceptor blocker synergizes with gemcitabine to inhibit the proliferation of pancreatic cancer...

European Journal of Pharmacology 665 (2011) 1–7
Contents lists available at ScienceDirect
European Journal of Pharmacology
j ourna l homepage: www.e lsev ie r.com/ locate /e jphar
Molecular and Cellular Pharmacology
β2-adrenoceptor blocker synergizes with gemcitabine to inhibit the proliferation ofpancreatic cancer cells via apoptosis induction
Tao Shan a, Qingyong Ma a,⁎, Dong Zhang a, Kun Guo a, Han Liu a, Fengfei Wang b, Erxi Wu b
a Department of Hepatobiliary Surgery, First Affiliated Hospital of Medical College, Xi'an Jiaotong University, 277 West Yanta Road, Xi'an 710061, Shaanxi Province, Chinab Department of Pharmaceutical Sciences, NDSU, Sudro Hall 203, Fargo, ND 58105, USA
⁎ Corresponding author. Tel./fax: +86 29 8532 3899.E-mail address: [email protected] (Q. Ma).
0014-2999/$ – see front matter © 2011 Elsevier B.V. Aldoi:10.1016/j.ejphar.2011.04.055
a b s t r a c t
a r t i c l e i n f oArticle history:Received 6 December 2010Received in revised form 18 April 2011Accepted 20 April 2011Available online 7 May 2011
Keywords:Gemcitabineβ2-adrenoceptor blockerPancreatic cancerApoptosisNF-κBBcl-2
The stimulation of β2-adrenoceptor, which is a major mediator for chronic stress-induced cancers, has beenimplicated in the progression in the number of cancer cells, including pancreatic cancer, which remains one ofthe most aggressive and lethal diseases worldwide. Whether β-adrenoceptor antagonists potentiategemcitabine, a standard first-line treatment for advanced pancreatic cancer that offers only modest benefitdue to acquired chemoresistance, has not been elucidated. Thus, we studied the antiproliferative andapoptotic effects and the underlying mechanisms of gemcitabine combined with the β2-adrenoceptor blockerICI 118551 (1-[2,3-(dihydro-7-methyl-1H-iden-4-yl)oxy]-3-[(1-methylethyl)amino]-2-butanol), in humanpancreatic cancer BxPC-3 and MIA PaCa-2 cells. Results show that ICI 118551 significantly synergized theantiproliferative and pro-apoptotic effects induced by gemcitabine in both BxPC-3 and MIA PaCa-2 cells(Pb0.05 combination vs. control or gemcitabine alone). When cells were treated with the combination ofgemcitabine and ICI 118551, NF-κB activation was blocked; the expression of Bax protein was substantiallyincreased; and Bcl-2 protein was downregulated. Taken together, the data suggest that ICI 118551 potentiatesthe antiproliferative effects of gemcitabine by inducing apoptosis in pancreatic cancer cells. Our study impliesthat this combination may be an effective therapeutic strategy for pancreatic cancer.
l rights reserved.
© 2011 Elsevier B.V. All rights reserved.
1. Introduction
Pancreatic cancer remains one of the most aggressive and lethaldiseases worldwide, with a 5-year survival rate of less than 5%. Evenafter curative resection, the 5-year survival rate is only 10% (Fryer et al.,2009; Yokoyama et al., 2009). The poor prognosis of pancreatic cancer isattributable to the late manifestation of symptoms, aggressive localinvasion, liablemetastases, andweak response to chemotherapy (Duffyet al., 2003). Currently, gemcitabine, a standard first-line treatment foradvanced pancreatic cancer, is extensively utilized, but offers only amodest benefit due to acquired chemoresistance and multiple adverseeffects (Herrmannet al., 2007;Kullmannet al., 2009; Reddy et al., 2008).Thus, it is imperative that we discover novel strategies to identify lesstoxic agents that can sensitize pancreatic cancer cells to chemotherapy.
Considerable evidence shows that NF-κB plays a major role inpromoting gemcitabine resistance in pancreatic cancer (Arlt et al., 2003;Li et al., 2004); therefore, the agents that block NF-κB activation couldreduce chemoresistance to gemcitabine and perhaps be used in com-binationwith gemcitabine as a novel therapeutic regimen for pancreaticcancer.
Recent studies have shown that neurotransmitters are involvedin the regulation of the number of cancer cells that progress via
β-adrenoceptor, which is the major mediator (Antoni et al., 2006; Huet al., 2010; Thaker et al., 2006; Thaker et al., 2007; Tilan and Kitlinska,2010). It has also been reported that β-adrenoceptor blockers couldinhibit the regulation by blocking β-adrenoceptor, and suppressingNF-κB, PI3K/Akt, and PKA pathways in cancer cells (Audet and Bouvier,2008). Previously, we have also shown that β-adrenoceptor antagonistsare effective inhibitors of cell proliferation and inducers of cellularapoptosis by regulating the expression of NF-κB downstream geneproducts (Bax, Bcl-2) in pancreatic cancer cells (Zhang et al., 2009).Whether or not β-adrenoceptor antagonists can potentiate thegemcitabine-induced cytotoxicity by inhibiting NF-κB activation hasnot been elucidated. In this study, we have investigated themechanismsinvolved and the effects of the specific β2-adrenoceptor blocker ICI118551 in combination with gemcitabine.
2. Materials and methods
2.1. Cell cultures and treatments
The MIA PaCa-2 and BxPC-3 human pancreatic cancer cell lines(obtained from the American Tissue Type Collection, USA) were main-tained in Dulbecco's modified Eagle's medium (DMEM, GIBCO, USA)supplemented with penicillin (100 U/ml), streptomycin (100 μg/ml),0.1 mM nonessential amino acids, 0.2 mM glutamine, 1 mM pyruvate,and 10% heat-inactivated fetal bovine serum (FBS) and incubated in 5%
2 T. Shan et al. / European Journal of Pharmacology 665 (2011) 1–7
CO2 humidified atmosphere at 37 °C. Cells were grown to 80%confluency prior to treatment. The antibodies against NF-kB p65subunit, Bcl-2, Bax, and β-actin were purchased from Santa CruzBiotechnology (Santa Cruz, CA). Annexin V-FITC and propidium iodide(PI), gemcitabine, and ICI 118551 were purchased from Sigma (Beijing,China).
2.2. Proliferation assay
Cell proliferation was determined by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphen-yltetrazolium bromide (MTT, Sigma) uptake method.After the cells were seeded (5×103/well) in 200 μl of DMEM mediuminto 96-well plates and cultured overnight, ICI 118551(1, 10, 50,100 μM), gemcitabine (100 nM) or their combination was added to thecells. When cells were treated with ICI 118551 and gemcitabine, themedium was replaced with DMEM containing ICI 118551 (1, 10, 50,100 μM) and cultured for 3 h, then themediumwas again replacedwithDMEM containing 100 nM gemcitabine and further cultured for 24 h.Then,MTT reagent (5 mg/ml)was added and incubationwas continuedfor an additional 4 h. The reaction was terminated with 150 μldimethylsulfoxide (DMSO, Sigma) per well. Absorbance values weredetermined using an MRX Revelation 96-well multiscanner (DynexTechnologies, Chantilly, VA). The cells cultured in DMEM served as thecontrol group. The cell viability index was calculated according tothe following formula: experimental OD value/control OD value. Theexperiments were repeated three times.
2.3. Detection of cellular apoptosis by flow cytometry
Apoptosis was evaluated with an Annexin V-FITC/PI apoptosisdetection kit according to the instruction of the manufacturer (Pan
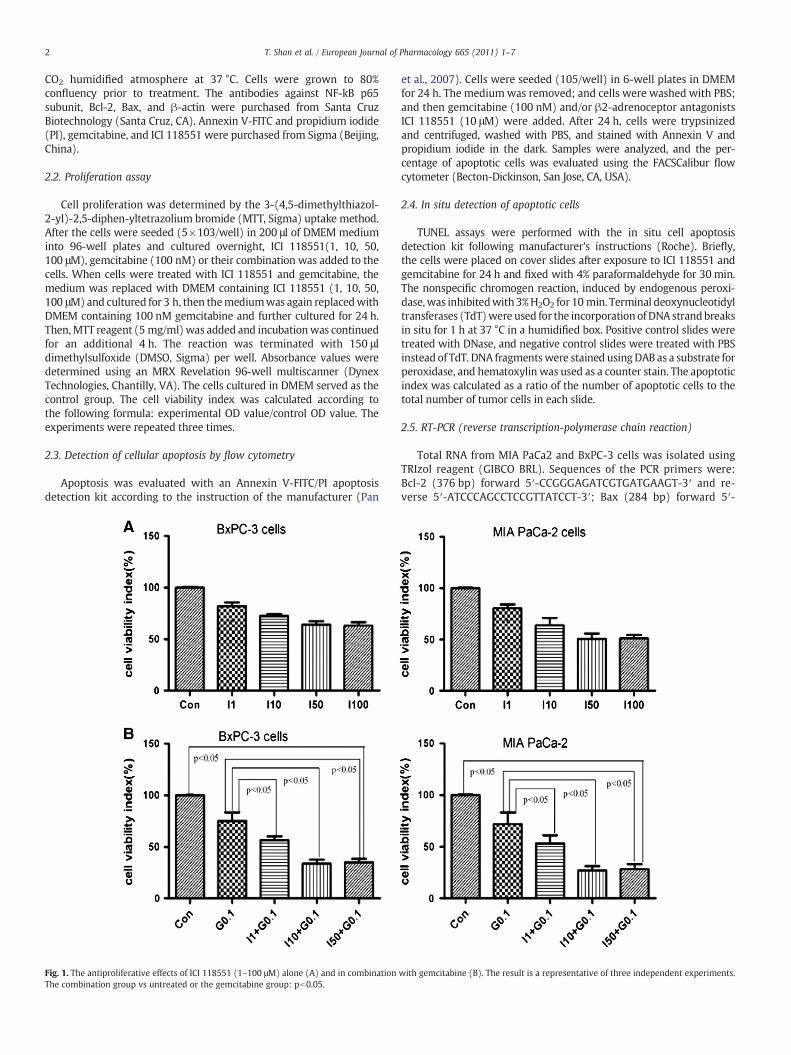
Fig. 1. The antiproliferative effects of ICI 118551 (1–100 μM) alone (A) and in combinationThe combination group vs untreated or the gemcitabine group: pb0.05.
et al., 2007). Cells were seeded (105/well) in 6-well plates in DMEMfor 24 h. The mediumwas removed; and cells were washed with PBS;and then gemcitabine (100 nM) and/or β2-adrenoceptor antagonistsICI 118551 (10 μM) were added. After 24 h, cells were trypsinizedand centrifuged, washed with PBS, and stained with Annexin V andpropidium iodide in the dark. Samples were analyzed, and the per-centage of apoptotic cells was evaluated using the FACSCalibur flowcytometer (Becton-Dickinson, San Jose, CA, USA).
2.4. In situ detection of apoptotic cells
TUNEL assays were performed with the in situ cell apoptosisdetection kit following manufacturer's instructions (Roche). Briefly,the cells were placed on cover slides after exposure to ICI 118551 andgemcitabine for 24 h and fixed with 4% paraformaldehyde for 30 min.The nonspecific chromogen reaction, induced by endogenous peroxi-dase,was inhibitedwith3%H2O2 for 10 min. Terminal deoxynucleotidyltransferases (TdT)were used for the incorporationofDNA strandbreaksin situ for 1 h at 37 °C in a humidified box. Positive control slides weretreated with DNase, and negative control slides were treated with PBSinstead of TdT. DNA fragmentswere stained using DAB as a substrate forperoxidase, and hematoxylin was used as a counter stain. The apoptoticindex was calculated as a ratio of the number of apoptotic cells to thetotal number of tumor cells in each slide.
2.5. RT-PCR (reverse transcription-polymerase chain reaction)
Total RNA from MIA PaCa2 and BxPC-3 cells was isolated usingTRIzol reagent (GIBCO BRL). Sequences of the PCR primers were:Bcl-2 (376 bp) forward 5′-CCGGGAGATCGTGATGAAGT-3′ and re-verse 5′-ATCCCAGCCTCCGTTATCCT-3′; Bax (284 bp) forward 5′-
with gemcitabine (B). The result is a representative of three independent experiments.
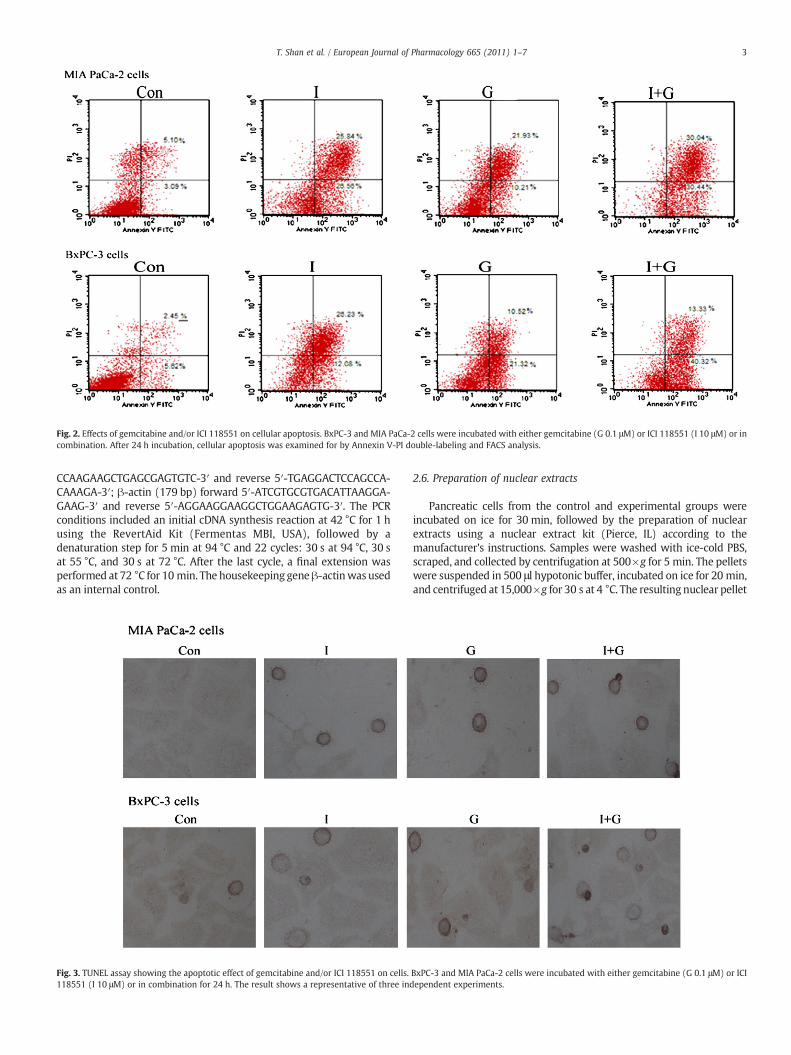
Fig. 2. Effects of gemcitabine and/or ICI 118551 on cellular apoptosis. BxPC-3 and MIA PaCa-2 cells were incubated with either gemcitabine (G 0.1 μM) or ICI 118551 (I 10 μM) or incombination. After 24 h incubation, cellular apoptosis was examined for by Annexin V-PI double-labeling and FACS analysis.
3T. Shan et al. / European Journal of Pharmacology 665 (2011) 1–7
CCAAGAAGCTGAGCGAGTGTC-3′ and reverse 5′-TGAGGACTCCAGCCA-CAAAGA-3′; β-actin (179 bp) forward 5′-ATCGTGCGTGACATTAAGGA-GAAG-3′ and reverse 5′-AGGAAGGAAGGCTGGAAGAGTG-3′. The PCRconditions included an initial cDNA synthesis reaction at 42 °C for 1 husing the RevertAid Kit (Fermentas MBI, USA), followed by adenaturation step for 5 min at 94 °C and 22 cycles: 30 s at 94 °C, 30 sat 55 °C, and 30 s at 72 °C. After the last cycle, a final extension wasperformed at 72 °C for 10 min. The housekeeping geneβ-actinwas usedas an internal control.
Fig. 3. TUNEL assay showing the apoptotic effect of gemcitabine and/or ICI 118551 on cells.118551 (I 10 μM) or in combination for 24 h. The result shows a representative of three ind
2.6. Preparation of nuclear extracts
Pancreatic cells from the control and experimental groups wereincubated on ice for 30 min, followed by the preparation of nuclearextracts using a nuclear extract kit (Pierce, IL) according to themanufacturer's instructions. Samples were washed with ice-cold PBS,scraped, and collected by centrifugation at 500×g for 5 min. The pelletswere suspended in 500 μl hypotonic buffer, incubated on ice for 20 min,and centrifuged at 15,000×g for 30 s at 4 °C. The resulting nuclear pellet
BxPC-3 and MIA PaCa-2 cells were incubated with either gemcitabine (G 0.1 μM) or ICIependent experiments.
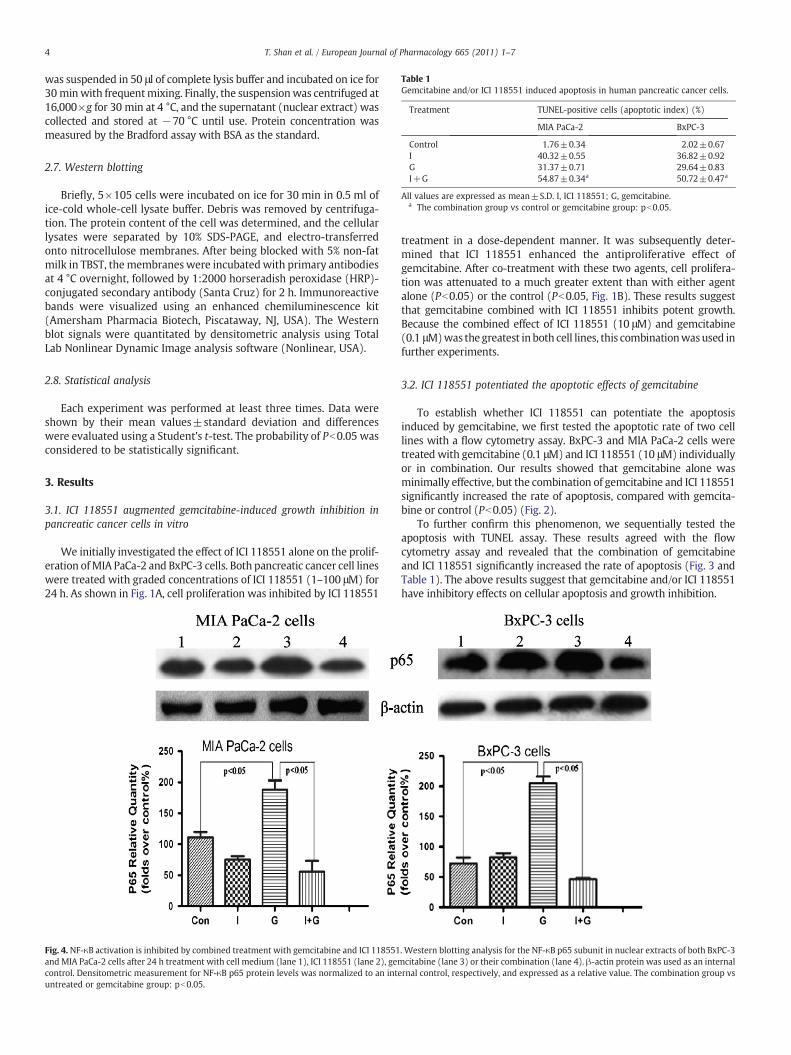
Table 1Gemcitabine and/or ICI 118551 induced apoptosis in human pancreatic cancer cells.
Treatment TUNEL-positive cells (apoptotic index) (%)
MIA PaCa-2 BxPC-3
Control 1.76±0.34 2.02±0.67I 40.32±0.55 36.82±0.92G 31.37±0.71 29.64±0.83I+G 54.87±0.34a 50.72±0.47a
All values are expressed as mean±S.D. I, ICI 118551; G, gemcitabine.a The combination group vs control or gemcitabine group: pb0.05.
4 T. Shan et al. / European Journal of Pharmacology 665 (2011) 1–7
was suspended in 50 μl of complete lysis buffer and incubated on ice for30 minwith frequentmixing. Finally, the suspensionwas centrifuged at16,000×g for 30 min at 4 °C, and the supernatant (nuclear extract) wascollected and stored at −70 °C until use. Protein concentration wasmeasured by the Bradford assay with BSA as the standard.
2.7. Western blotting
Briefly, 5×105 cells were incubated on ice for 30 min in 0.5 ml ofice-cold whole-cell lysate buffer. Debris was removed by centrifuga-tion. The protein content of the cell was determined, and the cellularlysates were separated by 10% SDS-PAGE, and electro-transferredonto nitrocellulose membranes. After being blocked with 5% non-fatmilk in TBST, themembranes were incubatedwith primary antibodiesat 4 °C overnight, followed by 1:2000 horseradish peroxidase (HRP)-conjugated secondary antibody (Santa Cruz) for 2 h. Immunoreactivebands were visualized using an enhanced chemiluminescence kit(Amersham Pharmacia Biotech, Piscataway, NJ, USA). The Westernblot signals were quantitated by densitometric analysis using TotalLab Nonlinear Dynamic Image analysis software (Nonlinear, USA).
2.8. Statistical analysis
Each experiment was performed at least three times. Data wereshown by their mean values±standard deviation and differenceswere evaluated using a Student's t-test. The probability of Pb0.05 wasconsidered to be statistically significant.
3. Results
3.1. ICI 118551 augmented gemcitabine-induced growth inhibition inpancreatic cancer cells in vitro
We initially investigated the effect of ICI 118551 alone on the prolif-eration ofMIA PaCa-2 and BxPC-3 cells. Both pancreatic cancer cell lineswere treated with graded concentrations of ICI 118551 (1–100 μM) for24 h. As shown in Fig. 1A, cell proliferation was inhibited by ICI 118551
Fig. 4. NF-κB activation is inhibited by combined treatment with gemcitabine and ICI 118551and MIA PaCa-2 cells after 24 h treatment with cell medium (lane 1), ICI 118551 (lane 2), gemcontrol. Densitometric measurement for NF-κB p65 protein levels was normalized to an inteuntreated or gemcitabine group: pb0.05.
treatment in a dose-dependent manner. It was subsequently deter-mined that ICI 118551 enhanced the antiproliferative effect ofgemcitabine. After co-treatment with these two agents, cell prolifera-tion was attenuated to a much greater extent than with either agentalone (Pb0.05) or the control (Pb0.05, Fig. 1B). These results suggestthat gemcitabine combined with ICI 118551 inhibits potent growth.Because the combined effect of ICI 118551 (10 μM) and gemcitabine(0.1 μM)was the greatest in both cell lines, this combinationwasused infurther experiments.
3.2. ICI 118551 potentiated the apoptotic effects of gemcitabine
To establish whether ICI 118551 can potentiate the apoptosisinduced by gemcitabine, we first tested the apoptotic rate of two celllines with a flow cytometry assay. BxPC-3 and MIA PaCa-2 cells weretreated with gemcitabine (0.1 μM) and ICI 118551 (10 μM) individuallyor in combination. Our results showed that gemcitabine alone wasminimally effective, but the combination of gemcitabine and ICI 118551significantly increased the rate of apoptosis, compared with gemcita-bine or control (Pb0.05) (Fig. 2).
To further confirm this phenomenon, we sequentially tested theapoptosis with TUNEL assay. These results agreed with the flowcytometry assay and revealed that the combination of gemcitabineand ICI 118551 significantly increased the rate of apoptosis (Fig. 3 andTable 1). The above results suggest that gemcitabine and/or ICI 118551have inhibitory effects on cellular apoptosis and growth inhibition.
. Western blotting analysis for the NF-κB p65 subunit in nuclear extracts of both BxPC-3citabine (lane 3) or their combination (lane 4). β-actin protein was used as an internalrnal control, respectively, and expressed as a relative value. The combination group vs

5T. Shan et al. / European Journal of Pharmacology 665 (2011) 1–7
3.3. ICI 118551 inhibits NF-κB activation induced by gemcitabine
To explore the molecular mechanism underlying the above effectof ICI 118551 on cells, we investigated whether or not the effect of ICI118551 is associated with the inhibition of NF-κB activation, whichcauses resistance to chemotherapy (Arlt et al., 2003). Western blottingwas performed to evaluate NF-κB p65 subunit protein levels, whichrepresent the active form of NF-κB. As shown in Fig. 4, the combinationtreatment of gemcitabine with ICI 118551 inhibited gemcitabine-induced NF-κB expression.
3.4. ICI 118551 sensitized gemcitabine in down-regulating theexpression of Bcl-2 and increasing the expression of Bax
Expression of Bax and Bcl-2 was detected because both genes arepro-apoptotic and anti-apoptotic molecules downstream of NF-κB(Liptay et al., 2003). RT-PCR analysis (Fig. 5A) indicates that mRNAlevels of Bax and Bcl-2 are significantly increased and suppressed by ICI118551 in combinationwith gemcitabine, respectively.Westernblotting(Fig. 5B) showed that the expression of Bcl-2 was significantly down-regulated in the combination group compared to treatment with an
Fig. 5. ICI 118551 combined with gemcitabine downregulates the expression of Bcl-2, and upthe two reagents for 24 h and harvested for RT-PCR (A) or Western blotting analysis (B). β
individual agent and control, whereas Bax expression was substantiallyincreased (Pb0.05).
4. Discussion
Currently, gemcitabine is the core first-line drug for pancreaticcancer treatment, but its efficacy is often disappointing (Haddock et al.,2007; Heinemann et al., 2008; Ozaki et al., 2007; Zeynalov et al., 2007).To improve its cytotoxic antitumor effect and to identify new reagentsfor pancreatic cancer treatment, we investigated the effects of ICI118551 in combination with gemcitabine on cellular proliferation andapoptosis in human pancreatic cancer cell lines and explored themechanism contributing to these effects. We found that ICI 118551enhanced the apoptotic effects of gemcitabine; these effects of ICI118551 may be associated with suppression of the activity of NF-κBdownregulation of Bcl-2 and upregulation of Bax. These findings extendour understanding of the function and molecular basis of ICI 118551 onthis dismal disease.
ICI 118551 is the most selective β2-adrenoceptor antagonist, and isused to treat cardiovascular diseases. Recently, because literature hasbeen accumulating that reports the involvementof neurotransmitters inthe regulation of cancer cell progression via β-adrenoceptor (of total
regulates the expression of Bax in BxPC-3 and MIA PaCa-2 cells. Cells were treated with-actin protein was used as an internal control.

Fig. 5 (continued).
6 T. Shan et al. / European Journal of Pharmacology 665 (2011) 1–7
β-adrenoceptors, β2-adrenoceptor is the major mediator) (Hasegawaand Saiki, 2002; Yang et al., 2009), clinical trials are beginning toevaluate additional biological activities of the β-adrenoceptor antago-nist in antagonizing cancers (Entschladen et al., 2005; Guo et al., 2009).
Previous studies suggest that β-adrenoceptor blockers can inhibitthe growth of prostate cancer cells and induce apoptosis of coloncancer cells (Masur et al., 2001; Palm et al., 2006). Here, our studyyields similar results and supports previous findings in pancreaticcancer cells. Most importantly, our study is the first report to showthat ICI 118551 enhances the apoptotic effects of gemcitabine incultured pancreatic cancer cells. Gemcitabine alone had a minimaleffect on apoptosis in BxPC-3 and MIA PaCa-2 cell lines. Anotherreport similarly showed that BxPC-3, Capan-1, and PancTu-1 cells areunaffected by gemcitabine (Arlt et al., 2003).
The above bioactivity mechanism of ICI 118551 may involve thesuppression of the activity of NF-κB. NF-κB pathways are key regulatorsof numerous cellular events, such as proliferation, differentiation, andapoptosis (Panet al., 2008;Xiong et al., 2004);moreover, numerous linesof evidence suggest that NF-κB plays a pivotal role in promotinggemcitabine resistance in pancreatic cancer (Bentires-Alj et al., 2003;YamamotoandGaynor, 2001), so it is likely that the inactivationofNF-κBby ICI 118551 can sensitize the cells to gemcitabine. This hypothesis
was verified by the finding that ICI 118551 can significantly sensitizepancreatic cancer cells to gemcitabine-induced apoptosis and stronglysuppress NF-κB p65 expression. These results agree with a previousreport in which MG132, sulfasalazine, and the IκBα suppressorsensitized pancreatic cancer cells to gemcitabine (Arlt et al., 2003).Similarly, genistein has been shown to sensitize prostate cancer cells togemcitabine through the down-regulating of NF-κB (Banerjee et al.,2005); aspirin and gum mastic, respectively, and to augment theantisurvival effect of gemcitabine on human pancreatic cancer throughthe down-regulating of NF-κB (Huang et al., 2010; Ou et al., 2010). Inaddition, more studies are needed to determine the role of NF-κB in theeffect of ICI 118551 on pancreatic cancer cells and to provide evidencethat this mechanism occurs in vivo as well.
The inactivationofNF-κB leads to the expressionof a largenumber oftarget genes, among which are the Bcl-2 family proteins (Pan et al.,2008). Our results show that ICI 118551 downregulates and upregulatesthe expression of Bcl-2 and Bax, respectively. The anti-apoptotic Bcl-2plays an important role in the development of chemotherapy resistancein cancer cells; the pro-apoptotic Bax, which can exist as a homodimer,is also able to form a heterodimer with Bcl-2. Bax/Bcl-2 impacts themitochondrial apoptotic function (Fahy et al., 2005; Korsmeyer et al.,1993). Thereby, our work revealed that enhancement and suppression

7T. Shan et al. / European Journal of Pharmacology 665 (2011) 1–7
of Bax and Bcl-2 expression by ICI 118551 contributes to gemcitabine-induced cytotoxicity and apoptosis.
In conclusion, our results reveal that ICI 118551 inhibits proliferationand potentiates the apoptosis-inducing effect of gemcitabine inpancreatic cancer cells, probably by inhibiting activation of NF-κB andthe expression of its regulated targets. Thus, the traditional agent ICI118551may prove to be a novel candidate for using in the combinationwith gemcitabine for the chemotherapeutic treatment for patients withpancreatic carcinoma (especially for patients who are subject to chronicstress due to their cancer diagnosis).
Acknowledgements
We wish to extend our thanks to Mary Pull (North Dakota StateUniversity) for her thoughtful reading of themanuscript. This studywasfunded by theMajor Scientific Grand of Shaanxi 13115 (2010ZDKG-49)and partially funded by ND EPSCoR grants (E. Wu).
References
Antoni, M.H., Lutgendorf, S.K., Cole, S.W., Dhabhar, F.S., Sephton, S.E., McDonald, P.G.,Stefanek, M., Sood, A.K., 2006. Opinion—the influence of bio-behavioural factors ontumour biology: pathways and mechanisms. Nat Rev Cancer 6, 240–248.
Arlt, A., Gehrz, A., Muerkoster, S., Vorndamm, J., Kruse, M.L., Folsch, U.R., Schafer, H.,2003. Role of NF-kappa B and Akt/PI3K in the resistance of pancreatic carcinomacell lines against gemcitabine-induced cell death. Oncogene 22, 3243–3251.
Audet, M., Bouvier, M., 2008. Insights into signaling from the beta(2)-adrenergicreceptor structure. Nat. Chem. Biol. 4, 397–403.
Banerjee, S., Zhang, Y., Ali, S., Bhuiyan, M., Wang, Z., Chiao, P.J., Philip, P.A., Abbruzzese,J., Sarkar, F.H., 2005. Molecular evidence for increased antitumor activity ofgemcitabine by genistein in vitro and in vivo using an orthotopic model ofpancreatic cancer. Cancer Res. 65, 9064–9072.
Bentires-Alj, M., Barbu, V., Fillet, M., Chariot, A., Relic, B., Jacobs, N., Gielen, J., Merville,M.P., Bours, V., 2003. NF-kappa B transcription factor induces drug resistancethrough MDR1 expression in cancer cells. Oncogene 22, 90–97.
Duffy, J.P., Eibl, G., Reber, H.A., Hines, O.J., 2003. Influence of hypoxia andneoangiogenesis on the growth of pancreatic cancer. Mol Cancer 2, 12.
Entschladen, F., Drell, T.L., Lang, K., Joseph, J., Zaenker, K.S., 2005. Neurotransmitters andchemokines regulate tumor cell migration: potential for a new pharmacologicalapproach to inhibit invasion and metastasis development. Curr Pharm Design 11,403–411.
Fahy, B.N., Schlieman, M.G., Mortenson, M.M., Virudachalam, S., Bold, R.J., 2005.Targeting BCL-2 overexpression in various human malignancies through NF-kappaB inhibition by the proteasome inhibitor bortezomib. Cancer Chemoth Pharm 56,46–54.
Fryer, R.A., Galustian, C., Dalgleish, A.G., Dalgelish, A.G., 2009. Recent advances anddevelopments in treatment strategies against pancreatic cancer. Curr ClinPharmacol 4, 102–112.
Guo, K., Ma, Q.Y., Wang, L.C., Hu, H.T., Li, J.H., Zhang, D., Zhang, M., 2009. Norepinephrine-induced invasion by pancreatic cancer cells is inhibited by propranolol. Oncol Rep 22,825–830.
Haddock, M.G., Swaminathan, R., Foster, N.R., Hauge, M.D., Martenson, J.A., Camoriano,J.K., Stella, P.J., Tenglin, R.C., Schaefer, P.L., Moore, D.F., Alberts, S.R., 2007.Gemcitabine, cisplatin, and radiotherapy for patients with locally advancedpancreatic adenocarcinoma: results of the north central cancer treatment groupphase II study n9942. J Clin Oncol 25, 2567–2572.
Hasegawa, H., Saiki, I., 2002. Psychosocial stress augments tumor development throughbeta-adrenergic activation in mice. Jap J Cancer Res. 93, 729–735.
Heinemann, V., Boeck, S., Hinke, A., Labianca, R., Louvet, C., 2008. Meta-analysis ofrandomized trials: Evaluation of benefit from gemcitabine-based combinationchemotherapy applied in advanced pancreatic cancer. Bmc Cancer 8.
Herrmann, R., Bodoky, G., Ruhstaller, T., Glimelius, B., Bajetta, E., Schuller, J., Saletti, P.,Bauer, J., Figer, A., Pestalozzi, B., Kohne, C.H., Mingrone, W., Stemmer, S.M., Tamas,K., Kornek, G.V., Koeberle, D., Cina, S., Bernhard, J., Dietrich, D., Scheithauer, W.,2007. Gemcitabine plus capecitabine compared with gemcitabine alone inadvanced pancreatic cancer: a randomized, multicenter, phase III trial of theSwiss Group for Clinical Cancer Research and the Central European CooperativeOncology Group. J Clin Oncol 25, 2212–2217.
Hu, H.T., Ma, Q.Y., Zhang, D., Shen, S.G., Han, L., Ma, Y.D., Li, R.F., Xie, K.P., 2010. HIF-1alpha links beta-adrenoceptor agonists and pancreatic cancer cells under normoxiccondition. Acta Pharmacol Sin 31, 102–110.
Huang, X.Y., Wang, H.C., Yuan, Z., Li, A., He, M.L., Ai, K.X., Zheng, Q., Qin, H.L., 2010.Gemcitabine combined with gum mastic causes potent growth inhibition andapoptosis of pancreatic cancer cells. Acta Pharmacol Sin 31, 741–745.
Korsmeyer, S.J., Shutter, J.R., Veis, D.J., Merry, D.E., Oltvai, Z.N., 1993. Bcl-2/Bax—arheostat that regulates antioxidant pathway and cell-death. Semin Cancer Biol. 4,327–332.
Kullmann, F., Hollerbach, S., Dollinger, M.M., Harder, J., Fuchs, M., Messmann, H., Trojan,J., Gabele, E., Hinke, A., Hollerbach, C., Endlicher, E., 2009. Cetuximab plusgemcitabine/oxaliplatin (gemoxcet) in first-line metastatic pancreatic cancer: amulticentre phase II study. Brit J Cancer 100, 1032–1036.
Li, L., Aggarwal, B.B., Shishodia, S., Abbruzzese, J., Kurzrock, R., 2004. Nuclear factor-kappa B and I kappa B kinase are constitutively active in human pancreatic cells,and their down-regulation by curcumin (diferuloylmethane) is associated with thesuppression of proliferation and the induction of apoptosis. Cancer 101,2351–2362.
Liptay, S., Weber, C.K., Ludwig, L., Wagner, M., Adler, G., Schmid, R.M., 2003. Mitogenicand antiapoptotic role of constitutive NF-kappa B/Rel activity in pancreatic cancer.Int J Cancer 105, 735–746.
Masur, K., Niggemann, B., Zanker, K.S., Entschladen, F., 2001. Norepinephrine-inducedmigration of SW 480 colon carcinoma cells is inhibited by beta-blockers. Cancer Res61, 2866–2869.
Ou, Y.Q., Zhu, W.B., Li, Y., Qiu, P.X., Huang, Y.J., Xie, J., He, S.M., Zheng, X.K., Leng, T.D., Xu,D., Yan, G.M., 2010. Aspirin inhibits proliferation of gemcitabine-resistant humanpancreatic cancer cells and augments gemcitabine-induced cytotoxicity. ActaPharmacol Sin 31, 73–80.
Ozaki, Y., Inoue, K., Muraki, T., Hamano, H., Arakura, N., Ochi, Y., Kawa, S., Kiyosawa, K.,Miyagawa, S., Sano, K., 2007. Pathological evaluation of advanced pancreatic cancerafter genicitabine treatment. Pancreas 35, 85.
Palm, D., Lang, K., Niggemann, B., Drell, T.L., Masur, K., Zaenker, K.S., Entschladen, F.,2006. The norepinephrine-driven metastasis development of PC-3 human prostatecancer cells in BALB/c nude mice is inhibited by beta-blockers. Int J Cancer 118,2744–2749.
Pan, J., Quintas-Cardama, A., Kantarjian, H.M., Akin, C., Manshouri, T., Lamb, P., Cortes, J.E.,Tefferi, A., Giles, F.J., Verstovsek, S., 2007. EXEL-0862, a novel tyrosine kinase inhibitor,induces apoptosis in vitro and ex vivo in human mast cells expressing the KIT D816Vmutation. Blood 109, 315–322.
Pan, X., Arumugam, T., Yamamoto, T., Levin, P.A., Ramachandran, V., Ji, B., Lopez-Berestein, G., Vivas-Mejia, P.E., Sood, A.K., McConkey, D.J., Logsdon, C.D., 2008.Nuclear factor-kappa B p65/relA silencing induces apoptosis and increasesgemcitabine effectiveness in a subset of pancreatic cancer cells. Clin Cancer Res14, 8143–8151.
Reddy, L.H., Marque, P.E., Dubernet, C., Mouelhi, S.L., Desmaele, D., Couvreur, P., 2008.Preclinical toxicology (subacute and acute) and efficacy of a new squalenoylgemcitabine anticancer nanomedicine. J Pharmacol Exp Ther 325, 484–490.
Thaker, P.H., Han, L.Y., Kamat, A.A., Arevalo, J.M., Takahashi, R., Lu, C.H., Jennings, N.B.,Armaiz-Pena, G., Bankson, J.A., Ravoori, M., Merritt, W.M., Lin, Y.G., Mangala, L.S.,Kim, T.J., Coleman, R.L., Landen, C.N., Li, Y., Felix, E., Sanguino, A.M., Newman, R.A.,Lloyd, M., Gershenson, D.M., Kundra, V., Lopez-Berestein, G., Lutgendorf, S.K., Cole,S.W., Sood, A.K., 2006. Chronic stress promotes tumor growth and angiogenesis in amouse model of ovarian carcinoma. Nat Med 12, 939–944.
Thaker, P.H., Lutgendorf, S.K., Sood, A.K., 2007. The neuroendocrine impact of chronicstress on cancer. Cell Cycle 6, 430–433.
Tilan, J., Kitlinska, J., 2010. Sympathetic neurotransmitters and tumor angiogenesis—link between stress and cancer progression. J Oncol 2010, 539706.
Xiong, H.Q., Abbruzzese, J.L., Lin, E., Wang, L.W., Zheng, L.Z., Xie, K.P., 2004. Nf-kappa bactivity blockade impairs the angiogenic potential of human pancreatic cancercells. Int J Cancer 108, 181–188.
Yamamoto, Y., Gaynor, R.B., 2001. Therapeutic potential of inhibition of the NF-kappa Bpathway in the treatment of inflammation and cancer. J Clin Invest 107, 135–142.
Yang, E.V., Kim, S.J., Donovan, E.L., Chen, M., Gross, A.C., Marketon, J., Barsky, S.H., Glaser,R., 2009. Norepinephrine upregulates VEGF, IL-8, and IL-6 expression in humanmelanoma tumor cell lines: Implications for stress-related enhancement of tumorprogression. Brain Behav Immun 23, 267–275.
Yokoyama, Y., Nimura, Y., Nagino, M., 2009. Advances in the treatment of pancreaticcancer: limitations of surgery and evaluation of new therapeutic strategies. SurgToday 39, 466–475.
Zeynalov, R., Musayev, I., Giyasbeyli, S., Gasanzadeh, J., Dadashova, N., Ahadova, N.,2007. Efficiency of gemcitabine and capecitabine in the treatment of advancedpancreatic cancer. Ann Oncol 18, I117.
Zhang, D., Ma, Q.Y., Shen, S.G., Hu, H.T., 2009. Inhibition of pancreatic cancer cellproliferation by propranolol occurs through apoptosis induction the study of beta-adrenoceptor antagonist's anticancer effect in pancreatic cancercell. Pancreas 38,94–100.




















