
Using multilevel models to identify drivers of landscape-genetic structure among management areas
14
Using multilevel models to identify drivers of landscape-genetic structure among management areas RACHAEL Y. DUDANIEC,* 1 JONATHAN R. RHODES,* 1 JESSICA WORTHINGTON WILMER, † MITCHELL LYONS,* KRISTEN E. LEE, ‡ CLIVE A. MCALPINE* and FRANK N. CARRICK ‡ *School of Geography, Planning and Environmental Management, The University of Queensland, Brisbane, QLD 4072, Australia, †Natural Environments Program, Queensland Museum, PO Box 3300, South Brisbane, QLD 4101, Australia, ‡Centre for Mined Land Rehabilitation, Sustainable Minerals Institute, The University of Queensland, Brisbane, QLD 4072, Australia Abstract Landscape genetics offers a powerful approach to understanding species’ dispersal patterns. However, a central obstacle is to account for ecological processes operating at multiple spatial scales, while keeping research outcomes applicable to conservation management. We address this challenge by applying a novel multilevel regression approach to model landscape drivers of genetic structure at both the resolution of indi- viduals and at a spatial resolution relevant to management (i.e. local government man- agement areas: LGAs) for the koala (Phascolartos cinereus) in Australia. Our approach allows for the simultaneous incorporation of drivers of landscape-genetic relationships operating at multiple spatial resolutions. Using microsatellite data for 1106 koalas, we show that, at the individual resolution, foliage projective cover (FPC) facilitates high gene flow (i.e. low resistance) until it falls below approximately 30%. Out of six addi- tional land-cover variables, only highways and freeways further explained genetic distance after accounting for the effect of FPC. At the LGA resolution, there was significant variation in isolation-by-resistance (IBR) relationships in terms of their slopes and intercepts. This was predominantly explained by the average resistance distance among LGAs, with a weaker effect of historical forest cover. Rates of recent landscape change did not further explain variation in IBR relationships among LGAs. By using a novel multilevel model, we disentangle the effect of landscape resistance on gene flow at the fine resolution (i.e. among individuals) from effects occurring at coarser resolutions (i.e. among LGAs). This has important implications for our ability to identify appropriate scale-dependent management actions. Keywords: habitat fragmentation, landscape genetics, mammal dispersal, multilevel model, spatial scale, wildlife management Received 22 November 2012; revision received 2 April 2013; accepted 5 April 2013 Introduction Quantifying the mechanisms by which landscape features impede or facilitate dispersal is important for identifying ecological processes that govern species distributions (sensu Slatkin 1993; Macdonald & Johnson 2001). The field of landscape genetics has rapidly enhanced our ability to quantify the effects of landscape features on species’ genetic dispersal (Manel et al. 2003; Storfer et al. 2007). However, the challenge of linking landscape and genetic data at different spatial and temporal resolutions must be met (Landguth et al. 2010; Lowe & Allendorf 2010), because mismatches in the scale of observation and key processes can lead to erroneous conclusions about a species’ sensitivity to landscape features (Anderson et al. 2010). Therefore, it is important to test for the underlying drivers of land- scape-genetic relationships at multiple spatial scales, as Correspondence: Rachael Y. Dudaniec, Fax +61 7 3365 6899; Email: [email protected] 1 Joint first authors. © 2013 John Wiley & Sons Ltd Molecular Ecology (2013) 22, 3752–3765 doi: 10.1111/mec.12359
Transcript of Using multilevel models to identify drivers of landscape-genetic structure among management areas

Using multilevel models to identify drivers oflandscape-genetic structure among management areas
RACHAEL Y. DUDANIEC,* 1 JONATHAN R. RHODES,*1 JESSICA WORTHINGTON WILMER,†
MITCHELL LYONS,* KRISTEN E. LEE,‡ CLIVE A. MCALPINE* and FRANK N. CARRICK‡
*School of Geography, Planning and Environmental Management, The University of Queensland, Brisbane, QLD 4072,
Australia, †Natural Environments Program, Queensland Museum, PO Box 3300, South Brisbane, QLD 4101, Australia,
‡Centre for Mined Land Rehabilitation, Sustainable Minerals Institute, The University of Queensland, Brisbane, QLD 4072,
Australia
Abstract
Landscape genetics offers a powerful approach to understanding species’ dispersal
patterns. However, a central obstacle is to account for ecological processes operating at
multiple spatial scales, while keeping research outcomes applicable to conservation
management. We address this challenge by applying a novel multilevel regression
approach to model landscape drivers of genetic structure at both the resolution of indi-
viduals and at a spatial resolution relevant to management (i.e. local government man-
agement areas: LGAs) for the koala (Phascolartos cinereus) in Australia. Our approach
allows for the simultaneous incorporation of drivers of landscape-genetic relationships
operating at multiple spatial resolutions. Using microsatellite data for 1106 koalas, we
show that, at the individual resolution, foliage projective cover (FPC) facilitates high
gene flow (i.e. low resistance) until it falls below approximately 30%. Out of six addi-
tional land-cover variables, only highways and freeways further explained genetic
distance after accounting for the effect of FPC. At the LGA resolution, there was
significant variation in isolation-by-resistance (IBR) relationships in terms of their
slopes and intercepts. This was predominantly explained by the average resistance
distance among LGAs, with a weaker effect of historical forest cover. Rates of recent
landscape change did not further explain variation in IBR relationships among LGAs.
By using a novel multilevel model, we disentangle the effect of landscape resistance
on gene flow at the fine resolution (i.e. among individuals) from effects occurring at
coarser resolutions (i.e. among LGAs). This has important implications for our ability
to identify appropriate scale-dependent management actions.
Keywords: habitat fragmentation, landscape genetics, mammal dispersal, multilevel model,
spatial scale, wildlife management
Received 22 November 2012; revision received 2 April 2013; accepted 5 April 2013
Introduction
Quantifying the mechanisms by which landscape
features impede or facilitate dispersal is important for
identifying ecological processes that govern species
distributions (sensu Slatkin 1993; Macdonald & Johnson
2001). The field of landscape genetics has rapidly
enhanced our ability to quantify the effects of landscape
features on species’ genetic dispersal (Manel et al. 2003;
Storfer et al. 2007). However, the challenge of linking
landscape and genetic data at different spatial and
temporal resolutions must be met (Landguth et al. 2010;
Lowe & Allendorf 2010), because mismatches in the
scale of observation and key processes can lead to
erroneous conclusions about a species’ sensitivity to
landscape features (Anderson et al. 2010). Therefore, it
is important to test for the underlying drivers of land-
scape-genetic relationships at multiple spatial scales, as
Correspondence: Rachael Y. Dudaniec, Fax +61 7 3365 6899;
Email: [email protected] first authors.
© 2013 John Wiley & Sons Ltd
Molecular Ecology (2013) 22, 3752–3765 doi: 10.1111/mec.12359

well as to incorporate variables that represent dynamic
landscape or demographic processes (e.g. Anderson
et al. 2010; Murphy et al. 2010).
Analysing data at spatial scales that are both biologi-
cally meaningful and of direct use for conservation
management is a further challenge that many molecular
ecological studies fail to meet (Taylor & Dizon 1999). To
develop useful models for managing landscape connec-
tivity for species, we must be able to statistically link
the spatial scales of relevance for understanding ecolog-
ical processes (e.g. species’ range, habitat patches) with
scales of management relevance (e.g. political, land use
or regional boundaries) (Taylor & Dizon 1999; Pelosi
et al. 2010). Complicating this further is that the
relationships between ecological and landscape patterns
can vary spatially, with this variation being mediated
by the scale at which these patterns are measured
(Whittingham et al. 2007; McAlpine et al. 2008; Rhodes
et al. 2008). Spatial heterogeneity in landscape-genetic
processes can also be influenced by a species’ popula-
tion size, dispersal capacity, life history and the
geographic area under study (e.g. Steele et al. 2009;
Landguth et al. 2010; Dudaniec et al. 2012; Rasic &
Keyghobadi 2012). Therefore, there is a need to explain
spatial heterogeneity in landscape-genetic patterns
across scales that are relevant to both ecology and
management.
A further challenge is that genetic population struc-
ture is a product of both past and contemporary pro-
cesses, such as changes in landscape characteristics or
demography (e.g. Chiucci & Gibbs 2010; Cushman &
Landguth 2010; Dudaniec et al. 2012). For species with
long generation times, or those subject to rapid frag-
mentation effects, genetic structure can change more
slowly than the landscape it inhabits, resulting in time
lags to detect landscape effects (Landguth et al. 2010;
Murphy et al. 2010). The majority of studies focus on
historical rather than contemporary landscape-change
effects on genetic structure, but in some cases, rapid
genetic responses to recent landscape change have been
detected (e.g. Balkenhol & Waits 2009; Zellmer &
Knowles 2009). Thus, the strength, rate and pattern of
past ‘legacy effects’ may determine the degree to which
current landscape influences on genetic dispersal can be
detected and explained. Quantifying to what extent
‘legacy effects’ determine landscape-genetic patterns at
scales relevant to management can therefore improve
conservation planning decisions by elucidating the
underlying mechanisms that drive species’ dispersal.
One approach to addressing these issues in landscape
genetics is to link multilevel (hierarchical) regression
with a spatial–temporal landscape-genetics approach.
Multilevel regression models provide a powerful frame-
work for representing relationships among variables
simultaneously at different resolutions based on the
specification of random effects to represent hierarchical
structure (McMahon & Diez 2007; Qian et al. 2010). As
such, they offer an elegant way to incorporate relation-
ships at resolutions relevant to both management and
ecological processes and to account for any mismatches
between them (Cumming et al. 2006; Pelosi et al. 2010).
However, to our knowledge, the teasing apart of spatial
and temporal elements using multilevel regression
has not previously been applied in landscape-genetic
studies.
Here, we apply a novel multilevel regression model-
ling approach (McMahon & Diez 2007) to the analysis
of landscape-genetic relationships while testing for the
effects of dynamic landscape change. We adopt a land-
scape resistance approach based on circuit theory
(McRae & Beier 2007) combined with microsatellite data
for the koala (Phascolarcotos cinereus), an arboreal marsu-
pial endemic to Australia that is under threat from mul-
tiple ecological stressors. Our study region spans eight
local government areas (LGAs) that are the spatial units
relevant to land-use planning and therefore representa-
tive of the scale of management for koalas. First, we test
hypotheses regarding land-cover effects on isolation by
resistance (IBR) at the individual resolution to derive a
landscape resistance model. Second, using multilevel
regression, we test for variation in IBR patterns at the
LGA resolution while continuing to account for IBR
relationships at the individual resolution. Third, we use
past and recent landscape data to test for drivers of
variation in IBR relationships among LGAs. In doing so,
this study provides a novel approach for simultaneously
analysing landscape-genetic patterns at multiple spatial
scales, while also incorporating landscape-change
effects.
Materials and methods
Study species
The koala, P. cinereus (Phascolarctidae), is an arboreal
folivore endemic to Australia and feeds on a small
number of Eucalyptus species (Ellis et al. 2002). Habitat
loss is a major factor causing population declines
(McAlpine et al. 2006a,b; Rhodes et al. 2008; Smith et al.
2013), which is further exacerbated by drought (Adams-
Hosking et al. 2011; Seabrook et al. 2011), disease, car
injuries and dog attacks (Dique et al. 2003b; Rhodes
et al. 2011). The area and configuration of forest habitat
are important determinants of koala distributions
(Rhodes et al. 2005; McAlpine et al. 2006b). Koalas
generally move via the ground and rarely via the canopy.
Dispersal is male-biased and typically 2–3 km from the
natal site; however, dispersal events of approximately
© 2013 John Wiley & Sons Ltd
MULTILEVEL MODELS AND LANDSCAPE GENETICS 3753

10 km have been observed (Dique et al. 2003b). The
koala is federally listed as Vulnerable in the states of
Queensland, New South Wales and the Australian
Capital Territory (Environment Protection and Biodiver-
sity Conservation Act 1999), and population sizes have
reduced by as much as 70% over the past 15 years in
some areas (Department of Environment and Natural
Resources 2009).
Study area and sampling
The study area covered approximately 20 500 km2 of
southeast Queensland (SEQ), incorporating eight LGAs
(Fig. 1). Land-use planning of relevance to koala conser-
vation is largely conducted at the resolution of LGAs,
which is therefore the appropriate resolution at which
to inform conservation decisions. We obtained tissue
samples (approximately 4 mm2) from 594 individuals
from two wildlife hospitals between 2006 and 2009.
Co-ordinates were recorded for each koala sample by
hospital volunteers using a geographic positioning
system, or mapped post hoc from the nearest road inter-
section or landmark (maximum error approximately
200 m). Each individual was sexed and assigned an age
class (adult, juvenile) based on body mass and tooth
wear (Gordon 1991). These samples were combined
with those from a previous study of koalas conducted
within the same study region and time period (Lee et al.
2010; n = 512) to give a total n = 1106 (mean number of
individuals per LGA: 138.3 � 47.1; range: 8–345). The
mean estimated generation time of koalas has been
estimated at 6 years (range = 3–8 years; Phillips 2000);
therefore, our sampling period is likely to span a single
koala generation.
Genotyping, locus characteristics and genetic structure
All koalas were genotyped at six polymorphic microsat-
ellite loci developed by Houlden et al. (1996) using PCR
conditions in Lee et al. (2010) (see Appendix S1, Sup-
porting information). For each locus, we conducted tests
for Hardy–Weinberg (HW) and linkage equilibrium
(Raymond & Rousset 1995b), the presence of null
alleles, scoring errors and large-allele dropout (van
Oosterhout et al. 2004) (see Appendix S1, Supporting
information). We ensured that including juveniles
(n = 40) did not bias our genetic data by confirming
HW equilibrium in each LGA with and without juve-
niles (Table S1, Supporting information). Observed and
expected heterozygosities were calculated for each locus
in GENALEX v6.41 (Peakall & Smouse 2006). Samples
were tested for a significant difference in pairwise FST
between LGAs and sampling years using MSA
(Dieringer & Schlotterer 2003).
Genetic distances and landscape data
We calculated individual-based genetic distances
between all pairs of individuals using the distance mea-
sure, ar, defined by Rousset (2000), using GENEPOP 4.0
(Raymond & Rousset 1995a), which is suitable for
continuously distributed populations (Rousset 2000;
Landguth et al. 2010). We characterized contemporary
land cover across the study area using data for 2009
(Lyons et al. 2012) and data for roads (highways, freeways
and main roads) for 2008 (Table S2, Supporting informa-
tion). Land cover was mapped at a resolution of
100 m 9 100 m for the following land-cover classes
(described in Table S2, Supporting information), which
we ranked in order of increasing resistance as: (i) forest;
(ii) urban vegetation; (iii) cleared; (iv) urban; (v) main
roads; and (vi) highways/freeways. We assumed that
153°0'0"E
153°0'0"E
27°0'0"S 27°0'0"S
28°0'0"S 28°0'0"S
A
B
CD
E
F
G
H
0 10 20 305Kilometers
Koala sampleCoastlineLocal Government Area
Fig. 1 The location of the study area in Australia is indicated
by a black square. Enlarged map shows local government area
(LGA) boundaries, coastline and koala sampling locations.
LGAs are coded as follows: A = Brisbane City Council
(n = 68), B = Gold Coast City Council (n = 96), C = Ipswich
City Council (n = 44), D = Logan City Council (n = 181),
E = Moreton Bay Regional Council (n = 329), F = Redland City
Council (n = 345), G = Scenic Rim Regional Council (n = 8),
H = Somerset Regional Council (n = 35).
© 2013 John Wiley & Sons Ltd
3754 R. Y . DUDANIEC ET AL.

large water bodies (>100 m in width) represent a complete
barrier, although there is some evidence for koalas
occasionally crossing water bodies (F. Carrick, personal
communication). We also mapped woody foliage pro-
jective cover (FPC) (a measure of canopy cover, Table
S2, Supporting information: Specht 1983) across the
study area for 2009 at the same resolution. A resolution
of 100 m 9 100 m was deemed to be fine enough given
that koalas can move several hundred metres in a given
movement event (Dique et al. 2003a; Rhodes et al. 2005).
We set the extent of the land-cover and FPC data so
that it included a buffer (as recommended by Koen
et al. 2010) of 10 km (using the land-cover data), which
is the estimated maximum dispersal distance for koalas
in the region (Dique et al. 2003a).
Modelling drivers of IBR relationships at theindividual resolution
We estimated resistance across the landscape extent
using an approach similar to Shirk et al. (2010), but we
used linear regression and the log-likelihood (sensu
Hilborn & Mangel 1997), rather than Mantel tests and
correlation coefficients to evaluate alternative resistance
models. We first assumed that resistance was a function
of both FPC and land cover as follows:
ri ¼ 1þ a100� Fi
100
� �c
þ bRLi
� 1
5
� �g
ð1Þ
where ri is the resistance of raster cell i; 0 � Fi � 100
is the percentage FPC of cell i; RLiis the rank of land-
cover type 1 � Li � 6 of cell i (with land-cover types
ranked from lowest to highest resistance); a > 0 and
b > 0 are parameters that determine the maximum pos-
sible resistance values; and c > 0 and g > 0 are parame-
ters that determine the shape of the relationship
between Fi and RLi, respectively, and ri. Equation (1)
explicitly assumes that resistance decreases as FPC
increases and that resistance increases as land-cover
rank increases.
The exponent c determines the shape of the relation-
ship between FPC and resistance, being linear when
c = 1 and nonlinear when c 6¼ 1 (Shirk et al. 2010).
Similarly, the exponent g determines the shape of the
relationship between land-cover rank and resistance.
The parameters a and b determine the maximum possible
resistance value, with 1 + a + b being the maximum
resistance value possible (i.e. when FPC is zero and
land cover is highways/freeways). We estimated the
parameters a, b, c, g from the individual-based genetic
distance data in two steps. First, we set b = 0 (i.e. no
effect of land cover) and chose a range of values for the
parameters a and c (i.e. the influence of FPC on
resistance). These values were 0, 2, 5, 10, 100, 1000 for aand 0.001, 0.01, 0.1, 0.2, 0.5, 1, 2, 5, 10, 100, 1000 for c.For each possible combination of these values, we
calculated the resistance value of each raster cell and
obtained pairwise resistance distances between all indi-
viduals using Circuitscape v3.5.7 (McRae & Beier 2007).
For each combination of a and c, we fitted a linear
regression model to the individual-based genetic dis-
tances, with resistance distance as the explanatory variable
(McRae 2006). To avoid problems with nonindependence
among samples, we used a modified bootstrap proce-
dure (without replacement) (Worthington-Wilmer et al.
2008) in which random sampling was constrained to
maintain independence between our pairwise distances
measures. We generated 1000 bootstrap replicates and
fitted a linear regression to each replicate. Our
estimated values for a and c where those that had the
highest mean log-likelihood across bootstrap replicates.
Second, we repeated the procedure for the b and gparameters while holding the best supported a and cparameters constant. We considered the same range of
values for b and g as for a and c. This allowed us to esti-
mate the parameters b and g (that link land cover to
resistance) conditional on the estimates of the parameters
a and c (that link percentage FPC to resistance).
Therefore, our model was constructed such that the
effect of land cover was considered after the effect of
FPC had been accounted for. We examined uncertainty
in our parameter estimates by examining the selection
frequency for each parameter value across bootstrap
replicates. Analyses were conducted in R version 2.14
(http://www.r-project.org/).
Incorporating drivers of IBR relationships at theLGA resolution
To test for and explain variation in the IBR relationship
at resolutions relevant to management (i.e. the LGA
resolution), we used mixed-effects, multilevel linear regres-
sion models (Pinheiro & Bates 2000) (Fig. 2). Based on the
estimated resistance surface at the individual resolution,
we modelled the relationship between genetic distance
and resistance distance using the linear mixed-effects
regression model:
arði;jÞ ¼ sj þ mjRi;j þ ei;j ð2Þ
where ar(i,j) is genetic distance for individual pair i in
LGA pair j; Ri,j is the resistance distance for individual
pair i in LGA pair j; τj is a normally distributed random
effect for LGA pair j with mean τ and variance r2s ; υj is
a normally distributed random effect for LGA pair j
with mean υ and variance r2m ; and ei,j are normally
distributed residuals with mean zero and variance r2.
We represented LGAs as ‘pairs’ to reflect that, for a pair
© 2013 John Wiley & Sons Ltd
MULTILEVEL MODELS AND LANDSCAPE GENETICS 3755

of individuals, each individual will be located in a spe-
cific LGA. Note that we consider all pairs simulta-
neously, although an LGA pair can be with itself or a
different LGA. We allowed for the intercept of the IBR
relationship to vary at the LGA resolution through τjand allowed for the slope to vary at the LGA resolution
through υj.We tested for the statistical significance of variation
in the intercept and slope of IBR relationships among
LGAs (i.e. significance of the random effects τj and υj)against a null hypothesis that there was no variation
in the intercept and/or slope among LGAs. This was
achieved using likelihood ratio tests based on simu-
lated values from the finite sample distribution (Crain-
iceanu & Ruppert 2004). We conducted these tests
using a stepwise procedure in which we first tested
for the significance of the intercept random effect, τj,and then tested for the significance of the slope ran-
dom effect, υj. For each test, we conducted significance
tests for each of 1000 bootstrap replicates of the
genetic distances. We rejected the null hypothesis if
the null hypothesis was rejected (at a significance level
of 0.05) in more than 5% of the individual bootstrap
replicates.
To test for variables measured at the resolution of
LGAs that explain variation in IBR relationships among
LGAs, we considered four predictor variables: (i)
proportion of the landscape that was forest in 1972 (rep-
resenting past habitat amount); (ii) recent rates of
change in the proportion of the landscape that was forest
(between 1988 and 2009); (iii) recent rates of change in
the proportion of the landscape that was urban
(between 1988 and 2009); and (iv) average resistance
distance between LGA pairs (representing a measure of
effective isolation due to the landscape between LGA
pairs). Predictor variable 1 tests whether, at equilib-
rium, LGAs with historically low amounts of habitat
have: (a) steeper IBR slopes due to lower population
densities and higher population divergence, and (b)
lower intercepts due to a smaller panmictic area (Peter-
son & Denno 1998; Novembre & Slatkin 2009). However,
rapid landscape change may create nonequilibrium
patterns (i.e. weaker IBR relationships) if there has
not been enough time for populations to achieve
equilibrium, although this will depend on the species’
generation time and dispersal behaviour relative to the
rate of landscape change (Anderson et al. 2010). To
test for landscape-change effects, we therefore examined
whether the rates of change in both forest (i.e.
representing habitat) and urban cover (i.e. representing
nonhabitat) were important predictors of variation in
IBR relationships among LGAs (predictor variables 2
and 3). Finally, as expected for isolation by distance, we
predict lower IBR slopes at high-resistance distances
than at low-resistance distances to be indicative of non-
equilibrium regional patterns, possibly due to genetic
drift (e.g. Slatkin 1993; Hutchison & Templeton
1999). Therefore, we examined whether IBR relation-
ships varied at the LGA resolution as a function of
the resistance distances separating them (predictor
variable 4).
For each pair of LGAs, we created a minimum con-
vex polygon around the koala sampling locations con-
tained in each LGA pair and buffered this by 3.5 km
(mean dispersal distance, Dique et al. 2003a). We then
calculated the amount of forest and urban cover within
INDIVIDUALRESOLUTION
Predictor variables ( j)
Regression coefficients
Random intercept ( j)
Regression coefficients
Variance of random-effect ( 2 ) Variance of random-effect ( 2 )
Random slope ( j)
Estimated genetic distance ( r (i, j))
Variance of residuals ( 2)
Measured genetic distance ( r (i, j))
Resistance (Ri,j)
LGA RESOLUTION
Residuals ( i,j)
Fig. 2 Directed acyclic graph represent-
ing the multilevel regression model used
to identify drivers of IBR relationships at
the individual and LGA resolutions. Rect-
angular boxes represent predictor variables
and ovals represent model parameters
(dotted ovals represent the variance
components of the model and solid ovals
represent all other components of the
model). The ‘Individual Resolution’
component represents Equation 1, where
‘Resistance’ is resistance distance, ‘esti-
mated genetic distance’ is the expected
genetic distance given the model and
‘measured genetic distance’ is the actual
measured genetic distance. The ‘LGA
Resolution’ component represents Equa-
tions 3 and 4, where ‘Predictor variables’
are the four variables incorporated as
drivers of genetic distance at the LGA
resolution.
© 2013 John Wiley & Sons Ltd
3756 R. Y . DUDANIEC ET AL.

the buffered convex polygons for each LGA pair using
land-cover data from Lyons et al. (2012) for 1972 and
each year from 1988 to 2009. Rates of change in the
amount of forest and urban land cover were estimated
from the slope of a linear regression fitted to the
proportion of the area inside each buffered convex
polygon consisting of urban or forest cover for the
21 years between 1988 and 2009, with time as the
explanatory variable. The proportion of forest cover
within each convex polygon in 1972 was also calculated,
but these data were mapped at a coarser resolution
(60 m 9 60 m) than the data for 1988–2009 (25 m 9 25 m).
Therefore, we were unable to incorporate the 1972 data
into the calculation of rates of change, but instead used
these data to estimate variation among LGA pairs in
terms of past forest amount. Finally, for each LGA pair,
we calculated the average resistance distance (based on
our resistance surface) between all individuals in each
pair of LGAs (for all combinations of the eight LGAs,
n = 36 pairs). All variables were standardized to have a
mean of zero and standard deviation of one prior to
further analysis.
We tested whether the above four predictor variables
were important determinants of variation in IBR inter-
cepts and slopes at the LGA resolution by explicitly
incorporating them into our multilevel model as
predictors for the intercept, τj, and slope, υj, random
effects (sensu McMahon & Diez 2007). We did this by
redefining the intercept and slope random effects in
Equation (2) to be functions of the predictor variables
such that
sj ¼ h0Xj þ nj ð3ÞAnd
mj ¼ w0Xj þ xj ð4Þwhere τj and υj are the intercept and slope random
effects for local government pair j; h and w are vectors
of regression coefficients; Xj is a vector of the predictor
variables for local government pair j; and ξj and xj are
normally distributed random effects for local govern-
ment pair j, with means of zero and variances r2n and
r2x. This formulation allowed us to test explicitly for
drivers of IBR patterns at the LGA resolution, while still
accounting for patterns at the individual resolution. We
constructed 16 different models of all combinations of
the predictor variables assuming that, in every model,
the predictors for variation in the intercept were the
same as for those in the slope.
Each of the 16 regression models was fitted to the
1000 bootstrap replicates of the individual-based genetic
distances. We then conducted model selection within an
information theoretic framework to identify the relative
importance of each of the predictor variables in explaining
variation in IBR relationships among LGAs (Akaike
1998; Burnham & Anderson 2002). For each bootstrap
replicate, we identified the model with the lowest
Akaike’s Information Criterion (AIC) and then, for each
model, calculated the proportion of replicates in which
it was the best model (Burnham & Anderson 2002). This
provided a robust estimate of the probability that each
model is the most parsimonious model (Burnham &
Anderson 2002). We then calculated the 95% confidence
set of models, the relative importance of each variable
based on Akaike weights and model-averaged coeffi-
cients and unconditional standard errors based on the
bootstrap replicates (Burnham & Anderson 2002). For
each model, we also calculated the amount of variation
in the data explained by the explanatory variables and
random effects based on the measure r20 described in
Xu (2003). To assess the potential effect of differences in
sample sizes among LGAs we quantified the influence
of each LGA on model parameter estimates using
Cook’s Distance (Cook 1977). Cook’s Distance provides
a measure of the influence of a particular data point (or
set of data points) by estimating how much removing
that data point (or set of data points) changes the
parameter estimates in the model. We calculated the
mean and 95% confidence intervals of Cook’s Distance
for each LGA pair across bootstrap replicates for a
model containing the highest ranked variables. All
statistical analyses for the multilevel models were con-
ducted in R version 2.14 using the packages ‘lme4’,
‘RLRSim’ and ‘influence.ME’.
Results
Error-checking and locus characteristics
A complete set of genotypes for all six polymorphic loci
was obtained for 1106 individual koalas. There were no
significant differences in pairwise FST between years of
sampling within LGAs (all P > 0.05 following Bonfer-
roni correction), thus, years were pooled for all analy-
ses. None of the loci were consistently out of HW
equilibrium or consistently showed evidence for null
alleles across the eight LGAs (Table S1, Appendix S2,
Supporting information). Results of HW equilibrium
tests were almost identical when juveniles were
excluded (n = 40, Table S1, Supporting information).
There was no evidence for scoring error, stuttering
or large-allele dropout across loci. Genetic diversity of
koalas was high, with 8–19 alleles per locus (mean =13.3 � 1.6 SE) and observed heterozygosity ranged from
0.5 to 1.0 across loci and LGAs (Table S1, Supporting
information). Pairwise FST between LGAs ranged
between 0.002 and 0.107 (mean = 0.047 � 0.006, Table
S3, Supporting information).
© 2013 John Wiley & Sons Ltd
MULTILEVEL MODELS AND LANDSCAPE GENETICS 3757
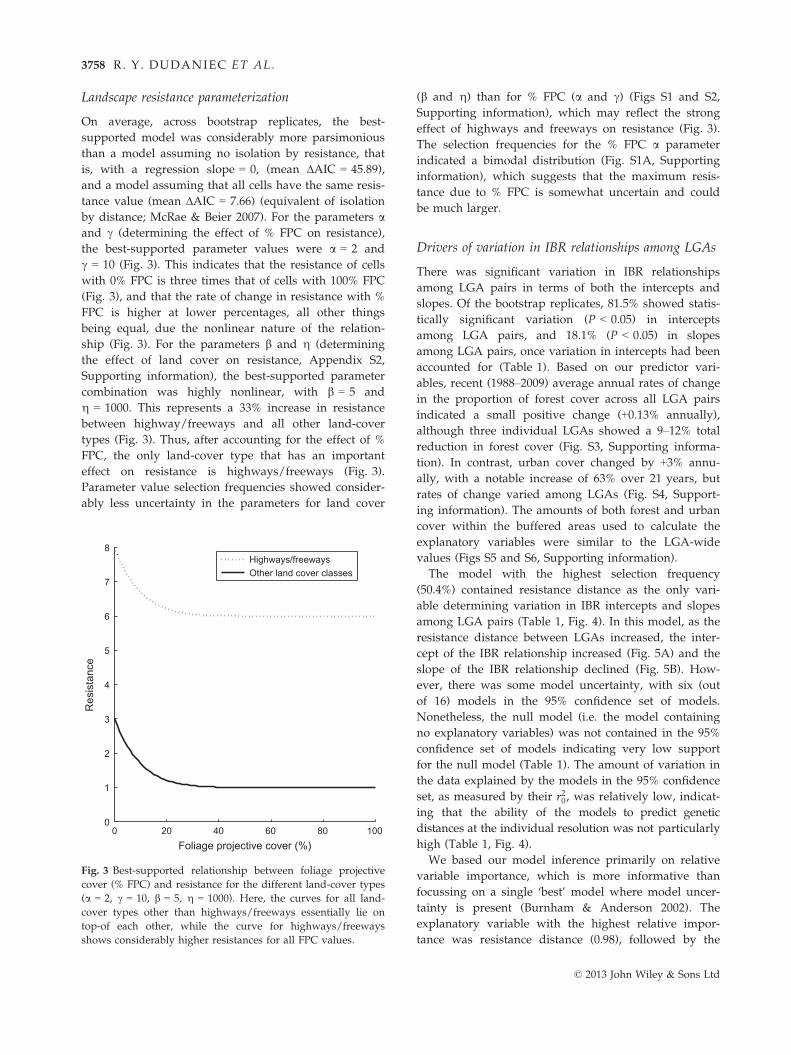
Landscape resistance parameterization
On average, across bootstrap replicates, the best-
supported model was considerably more parsimonious
than a model assuming no isolation by resistance, that
is, with a regression slope = 0, (mean ΔAIC = 45.89),
and a model assuming that all cells have the same resis-
tance value (mean ΔAIC = 7.66) (equivalent of isolation
by distance; McRae & Beier 2007). For the parameters aand c (determining the effect of % FPC on resistance),
the best-supported parameter values were a = 2 and
c = 10 (Fig. 3). This indicates that the resistance of cells
with 0% FPC is three times that of cells with 100% FPC
(Fig. 3), and that the rate of change in resistance with %
FPC is higher at lower percentages, all other things
being equal, due the nonlinear nature of the relation-
ship (Fig. 3). For the parameters b and g (determining
the effect of land cover on resistance, Appendix S2,
Supporting information), the best-supported parameter
combination was highly nonlinear, with b = 5 and
g = 1000. This represents a 33% increase in resistance
between highway/freeways and all other land-cover
types (Fig. 3). Thus, after accounting for the effect of %
FPC, the only land-cover type that has an important
effect on resistance is highways/freeways (Fig. 3).
Parameter value selection frequencies showed consider-
ably less uncertainty in the parameters for land cover
(b and g) than for % FPC (a and c) (Figs S1 and S2,
Supporting information), which may reflect the strong
effect of highways and freeways on resistance (Fig. 3).
The selection frequencies for the % FPC a parameter
indicated a bimodal distribution (Fig. S1A, Supporting
information), which suggests that the maximum resis-
tance due to % FPC is somewhat uncertain and could
be much larger.
Drivers of variation in IBR relationships among LGAs
There was significant variation in IBR relationships
among LGA pairs in terms of both the intercepts and
slopes. Of the bootstrap replicates, 81.5% showed statis-
tically significant variation (P < 0.05) in intercepts
among LGA pairs, and 18.1% (P < 0.05) in slopes
among LGA pairs, once variation in intercepts had been
accounted for (Table 1). Based on our predictor vari-
ables, recent (1988–2009) average annual rates of change
in the proportion of forest cover across all LGA pairs
indicated a small positive change (+0.13% annually),
although three individual LGAs showed a 9–12% total
reduction in forest cover (Fig. S3, Supporting informa-
tion). In contrast, urban cover changed by +3% annu-
ally, with a notable increase of 63% over 21 years, but
rates of change varied among LGAs (Fig. S4, Support-
ing information). The amounts of both forest and urban
cover within the buffered areas used to calculate the
explanatory variables were similar to the LGA-wide
values (Figs S5 and S6, Supporting information).
The model with the highest selection frequency
(50.4%) contained resistance distance as the only vari-
able determining variation in IBR intercepts and slopes
among LGA pairs (Table 1, Fig. 4). In this model, as the
resistance distance between LGAs increased, the inter-
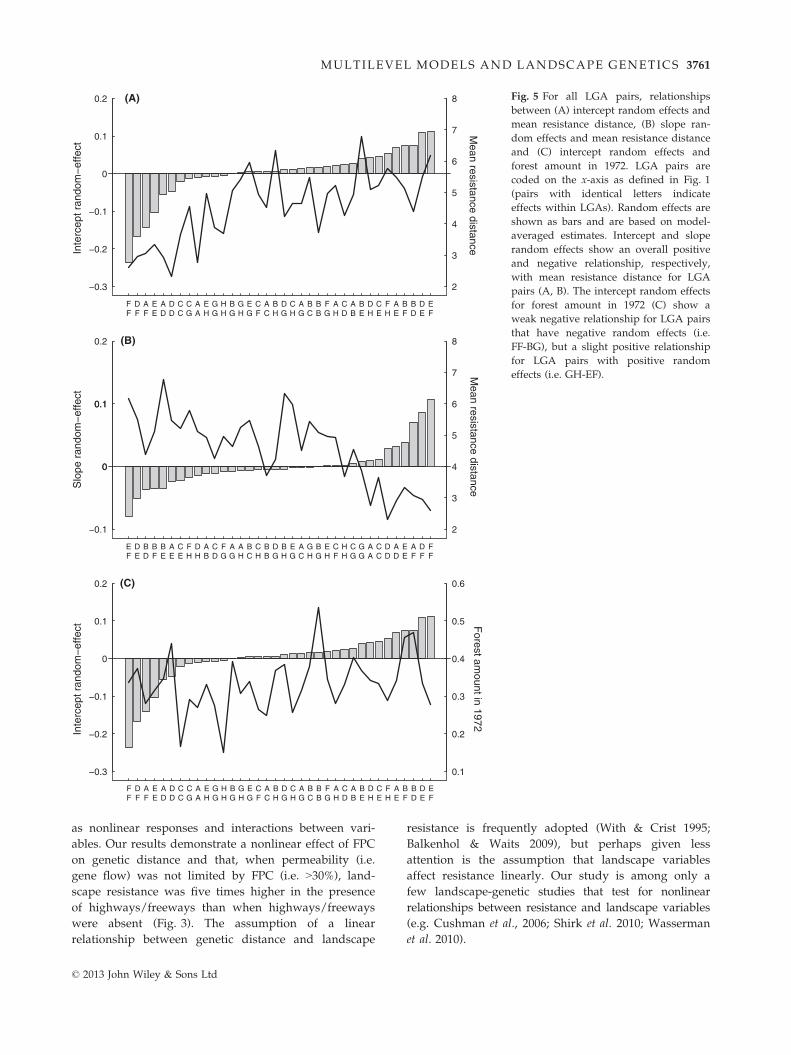
cept of the IBR relationship increased (Fig. 5A) and the
slope of the IBR relationship declined (Fig. 5B). How-
ever, there was some model uncertainty, with six (out
of 16) models in the 95% confidence set of models.
Nonetheless, the null model (i.e. the model containing
no explanatory variables) was not contained in the 95%
confidence set of models indicating very low support
for the null model (Table 1). The amount of variation in
the data explained by the models in the 95% confidence
set, as measured by their r20, was relatively low, indicat-
ing that the ability of the models to predict genetic
distances at the individual resolution was not particularly
high (Table 1, Fig. 4).
We based our model inference primarily on relative
variable importance, which is more informative than
focussing on a single ‘best’ model where model uncer-
tainty is present (Burnham & Anderson 2002). The
explanatory variable with the highest relative impor-
tance was resistance distance (0.98), followed by the
0 20 40 60 80 1000
1
2
3
4
5
6
7
8
Foliage projective cover (%)
Res
ista
nce
Highways/freewaysOther land cover classes
Fig. 3 Best-supported relationship between foliage projective
cover (% FPC) and resistance for the different land-cover types
(a = 2, c = 10, b = 5, g = 1000). Here, the curves for all land-
cover types other than highways/freeways essentially lie on
top-of each other, while the curve for highways/freeways
shows considerably higher resistances for all FPC values.
© 2013 John Wiley & Sons Ltd
3758 R. Y . DUDANIEC ET AL.
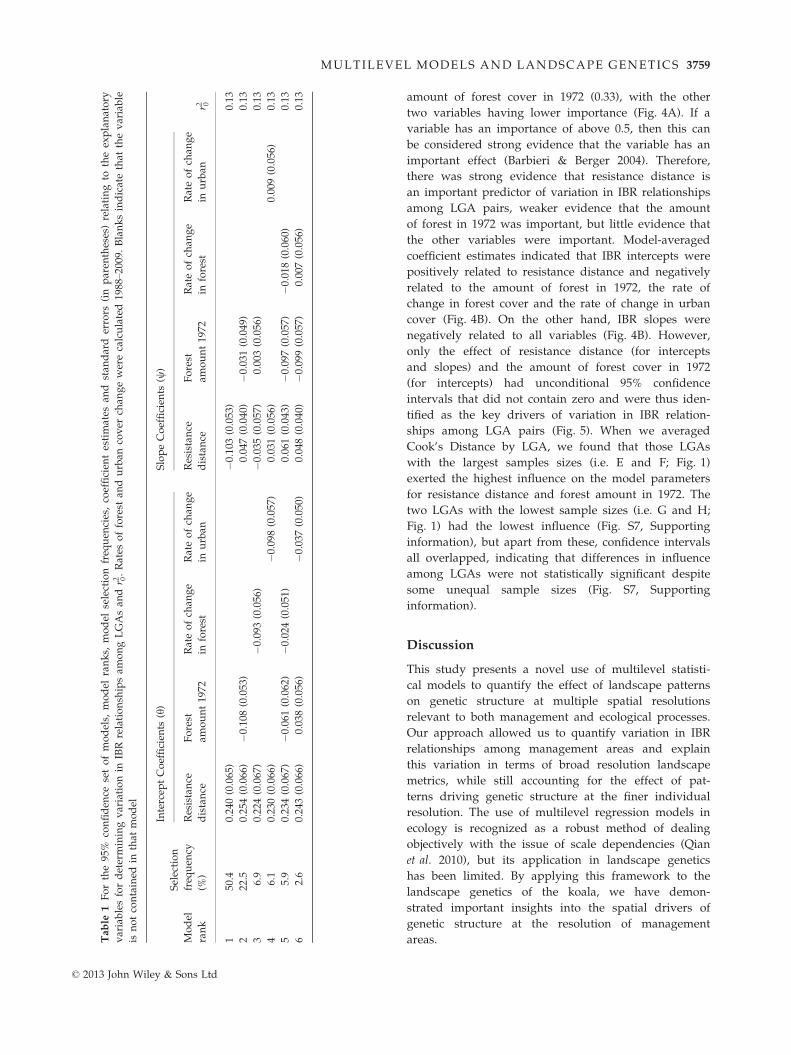
amount of forest cover in 1972 (0.33), with the other
two variables having lower importance (Fig. 4A). If a
variable has an importance of above 0.5, then this can
be considered strong evidence that the variable has an
important effect (Barbieri & Berger 2004). Therefore,
there was strong evidence that resistance distance is
an important predictor of variation in IBR relationships
among LGA pairs, weaker evidence that the amount
of forest in 1972 was important, but little evidence that
the other variables were important. Model-averaged
coefficient estimates indicated that IBR intercepts were
positively related to resistance distance and negatively
related to the amount of forest in 1972, the rate of
change in forest cover and the rate of change in urban
cover (Fig. 4B). On the other hand, IBR slopes were
negatively related to all variables (Fig. 4B). However,
only the effect of resistance distance (for intercepts
and slopes) and the amount of forest cover in 1972
(for intercepts) had unconditional 95% confidence
intervals that did not contain zero and were thus iden-
tified as the key drivers of variation in IBR relation-
ships among LGA pairs (Fig. 5). When we averaged
Cook’s Distance by LGA, we found that those LGAs
with the largest samples sizes (i.e. E and F; Fig. 1)
exerted the highest influence on the model parameters
for resistance distance and forest amount in 1972. The
two LGAs with the lowest sample sizes (i.e. G and H;
Fig. 1) had the lowest influence (Fig. S7, Supporting
information), but apart from these, confidence intervals
all overlapped, indicating that differences in influence
among LGAs were not statistically significant despite
some unequal sample sizes (Fig. S7, Supporting
information).
Discussion
This study presents a novel use of multilevel statisti-
cal models to quantify the effect of landscape patterns
on genetic structure at multiple spatial resolutions
relevant to both management and ecological processes.
Our approach allowed us to quantify variation in IBR
relationships among management areas and explain
this variation in terms of broad resolution landscape
metrics, while still accounting for the effect of pat-
terns driving genetic structure at the finer individual
resolution. The use of multilevel regression models in
ecology is recognized as a robust method of dealing
objectively with the issue of scale dependencies (Qian
et al. 2010), but its application in landscape genetics
has been limited. By applying this framework to the
landscape genetics of the koala, we have demon-
strated important insights into the spatial drivers of
genetic structure at the resolution of management
areas.Table
1Forthe95%
confiden
cesetofmodels,
model
ranks,
model
selectionfreq
uen
cies,coefficien
testimates
and
stan
dard
errors
(inparen
theses)relatingto
theexplanatory
variablesfordeterminingvariationin
IBR
relationsh
ipsam
ongLGAsan
dr2 0.Rates
offorest
andurban
cover
chan
gewerecalculated1988–2009.
Blanksindicatethat
thevariable
isnotcontained
inthat
model
Model
rank
Selection
freq
uen
cy
(%)
Intercep
tCoefficien
ts(h)
SlopeCoefficien
ts(w)
r2 0
Resistance
distance
Forest
amount1972
Rateofch
ange
inforest
Rateofch
ange
inurban
Resistance
distance
Forest
amount1972
Rateofch
ange
inforest
Rateofch
ange
inurban
150.4
0.240(0.065)
�0.103
(0.053)
0.13
222.5
0.254(0.066)
�0.108
(0.053)
0.047(0.040)
�0.031
(0.049)
0.13
36.9
0.224(0.067)
�0.093
(0.056)
�0.035
(0.057)
0.003(0.056)
0.13
46.1
0.230(0.066)
�0.098
(0.057)
0.031(0.056)
0.009(0.056)
0.13
55.9
0.234(0.067)
�0.061
(0.062)
�0.024
(0.051)
0.061(0.043)
�0.097
(0.057)
�0.018
(0.060)
0.13
62.6
0.243(0.066)
0.038(0.056)
�0.037
(0.050)
0.048(0.040)
�0.099
(0.057)
0.007(0.056)
0.13
© 2013 John Wiley & Sons Ltd
MULTILEVEL MODELS AND LANDSCAPE GENETICS 3759
Variation in IBR relationships among managementareas
The significant variation in the IBR relationship among
LGAs was predominantly explained by the average
resistance distance among LGAs (Table 1, Fig. 4). The
IBR relationship became flatter with increasing resis-
tance distance, as indicated by the increase in intercept
(Fig. 5A) and reduction in slope (Fig. 5B). Hence, regio-
nal nonequilibrium processes (e.g. Hutchison & Tem-
pleton 1999; Kittlein & Gaggiotti 2008) were apparent at
the LGA resolution for larger resistance distances
among LGAs. This dampening of the IBR relationship
may indicate a dominating effect of genetic drift (rather
than gene flow) on genetic structure at larger geographic
distances.
Isolation-by-distance patterns can vary according to
spatial extent (Slatkin 1993; Peterson & Denno 1998),
but our findings are in contrast to predictions that
ecological systems at smaller spatial extents are more
likely to exhibit nonequilibrium (i.e. non-IBD) dynamics
than larger extents due to increased detection of sto-
chastic processes (e.g. more frequent colonization or
extinction events) (DeAngelis & Waterhouse 1987). The
nonequilibrium patterns, we observe at larger distances,
may therefore be better explained by the ecological or
behavioural attributes of the study species, habitat
heterogeneity or environmental disturbance (Slatkin 1993).
Although a weaker effect than for resistance distance
(Table 1), there was some evidence for a negative
relationship between ‘forest amount in 1972’ and the
intercept of the IBR relationship (Table 1, Fig. 4). This
suggests higher genetic differentiation between LGAs
with lower historical forest amount, although the rela-
tionship may be nonlinear (Fig. 5C). This relationship
may also be partially shaped by genetic drift and other
factors driving nonequilibrium processes across LGAs.
Importantly, due to the absence of an effect of the rate
of landscape change, it may be inferred that current IBR
patterns are driven predominantly by a ‘legacy effect’,
rather than recent landscape modification. If the current
and historic landscape structure were both important
drivers of variation in IBR relationships among LGAs,
we would expect the rate of change in forest cover to
also be an important predictor. The change in urban
cover was not found to be an important predictor of
variation in IBR relationships despite a rapid increase
in urban cover (1988–2009, Fig. S4, Supporting informa-
tion), which may be explained by the low impact of
urban land cover on resistance, once FPC had been
accounted for.
Depending on the rate at which genetic changes accu-
mulate and respond to impacts of habitat fragmenta-
tion, genetic signatures may take several generations to
become evident within populations (e.g. Holzhauer
et al. 2006; Orsini et al. 2008; Metzger et al. 2009).
According to simulations by Landguth et al. (2010), 1–3
generations are generally sufficient to detect novel land-
scape barriers for species capable of dispersing 10 km
or more, with the number of generations expected to
increase sharply with decreasing dispersal capability.
We found a signature for past habitat availability (i.e.
approximately six generations ago) influencing IBR
patterns at the LGA resolution, but did not find an effect
of landscape change over a period of approximately 3.5
generations (based on Phillips 2000) in koalas, which
disperse 3.5 km on average. Therefore, a possible expla-
nation for the lack of support for an effect of the rate of
landscape change is the limited dispersal capacity of
the koala, relative to the size of the study area, com-
bined with a time frame of analysis that may be too
narrow to detect a strong genetic response.
Interpretation of the best resistance model
The parameter estimation method we use offers a more
defensible approach than the use of ‘expert opinion’
(Koen et al. 2010; Spear et al. 2010) as it allows the
genetic data to inform the best resistance model, as well
Resistance distance Forest amount Forest change Urban change0
0.2
0.4
0.6
0.8
1V
aria
ble
impo
rtanc
e
Resistance distance Forest amount Forest change Urban change−0.1
0
0.1
0.2
0.3
0.4
Coe
ffici
ent e
stim
ate
InterceptSlope
(A)
(B)
Fig. 4 Variable importance (A) and model-averaged coefficient
estimates (with unconditional 95% confidence intervals, CI) (B)
for the effect of resistance distance (CI intercept: 0.111–0.358,
CI slope: �0.095 to 0.27), amount of forest cover in 1972 (CI
intercept: �0.054 to 0.003, CI slope: �0.043 to 0.009), rate of
change in forest cover (1988–2009) (CI intercept: �0.021 to
0.004, CI slope: �0.019 to 0.021), and rate of change in urban
cover (1988–2009) (CI intercept: 0.019–0.000, CI slope: �0.011t o
0.008) on IBR relationships among LGA pairs. Confidence
intervals were significantly different from zero for all variables.
© 2013 John Wiley & Sons Ltd
3760 R. Y . DUDANIEC ET AL.
as nonlinear responses and interactions between vari-
ables. Our results demonstrate a nonlinear effect of FPC
on genetic distance and that, when permeability (i.e.
gene flow) was not limited by FPC (i.e. >30%), land-
scape resistance was five times higher in the presence
of highways/freeways than when highways/freeways
were absent (Fig. 3). The assumption of a linear
relationship between genetic distance and landscape
resistance is frequently adopted (With & Crist 1995;
Balkenhol & Waits 2009), but perhaps given less
attention is the assumption that landscape variables
affect resistance linearly. Our study is among only a
few landscape-genetic studies that test for nonlinear
relationships between resistance and landscape variables
(e.g. Cushman et al., 2006; Shirk et al. 2010; Wasserman
et al. 2010).
−0.3
−0.2
−0.1
0
0.1
0.2
2
3
4
5
6
7
8
FF
DF
AF
EE
AD
DD
CC
CG
AA
EH
GG
HH
BG
GH
EG
CF
AC
BH
DG
CH
AG
BC
BB
FG
AH
CD
AB
BE
DH
CE
FH
AE
BF
BD
DE
EF
(A)
Mean resistance distanceIn
terc
ept r
ando
m−
effe
ct
−0.1
00
0.10.1
0.2
2
3
4
5
6
7
8
EF
DE
BD
BF
BE
AE
CE
FH
DH
AB
CD
FG
AG
AH
BC
CH
BB
DG
BH
EG
AC
GH
BG
EH
CF
HH
CG
GG
AA
CC
DD
AD
EE
AF
DF
FF
(B)
Mean resistance distanceS
lope
ran
dom
−ef
fect
−0.3
−0.2
−0.1
0
0.1
0.2
0.1
0.2
0.3
0.4
0.5
0.6
FF
DF
AF
EE
AD
DD
CC
CG
AA
EH
GG
HH
BG
GH
EG
CF
AC
BH
DG
CH
AG
BC
BB
FG
AH
CD
AB
BE
DH
CE
FH
AE
BF
BD
DE
EF
(C)
Forest am
ount in 1972
Inte
rcep
t ran
dom
−ef
fect
Fig. 5 For all LGA pairs, relationships
between (A) intercept random effects and
mean resistance distance, (B) slope ran-
dom effects and mean resistance distance
and (C) intercept random effects and
forest amount in 1972. LGA pairs are
coded on the x-axis as defined in Fig. 1
(pairs with identical letters indicate
effects within LGAs). Random effects are
shown as bars and are based on model-
averaged estimates. Intercept and slope
random effects show an overall positive
and negative relationship, respectively,
with mean resistance distance for LGA
pairs (A, B). The intercept random effects
for forest amount in 1972 (C) show a
weak negative relationship for LGA pairs
that have negative random effects (i.e.
FF-BG), but a slight positive relationship
for LGA pairs with positive random
effects (i.e. GH-EF).
© 2013 John Wiley & Sons Ltd
MULTILEVEL MODELS AND LANDSCAPE GENETICS 3761

The small number of spatial genetic studies on
nonflying, arboreal mammals show variable movement
sensitivities to forest fragmentation (e.g. in orang-utans:
Goossens et al. 2005; gliders: Taylor et al. 2007; lemurs:
Qu�em�er�e et al. 2010; and possums: Lancaster et al.
2011). Studies on the effect of canopy cover on gene
flow in terrestrial vertebrates show mostly positive rela-
tionships (e.g. Munshi-South 2012; Niedziałkowska et al.
2012), but the effect may be dominated by elevation
(e.g. Wasserman et al. 2010; Dudaniec et al. 2012) or
road cover (e.g. Crosby et al. 2009). Roads represent a
major movement barrier for many taxa (e.g. Balkenhol
& Waits 2009; Jackson & Fahrig 2011), and the impact
of major roads on koalas (via fatalities and injuries) is
well documented (Dique et al. 2003a,b; Rhodes et al.
2011). Notably, our results suggest that highways and
freeways confer much greater resistance to gene flow
than main roads, which were not distinguishable from
other land-cover variables. This may be related to dif-
ferences in traffic volume (e.g. as in Shirk et al. 2010), or
the presence of physical barriers bordering major roads.
Notably, urban areas appeared to have little influence
on resistance once FPC had been accounted for.
Limitations and future research
This study represents the largest sampling effort for
any regional genetic study on the koala (Houlden et al.
1996; Sherwin et al. 2000). Although greater resolution
in genetic patterns may have been obtained with more
loci, the high polymorphism of the six loci used, the
lack of missing genotypes and the large sample size of
individuals greatly increase the resolution of our data
(Landguth et al. 2012). Regarding the sampling of
LGAs, two points should be noted, (i) our inferences
are only well supported for the area where samples
were collected; and (ii) LGAs with small sample sizes
exerted little influence on our overall model because
their contributions to the model likelihood are small. In
addition, variation in sample sizes among LGAs does
not appear to be a major issue for inference, with little
significant variation in influence on parameter esti-
mates among LGAs (Fig. S7, Supporting information).
Further, our approach was designed to understand var-
iation in landscape-genetic relationships among LGAs,
rather than to make inferences about specific LGAs,
which would tend to be unreliable due to potentially
high levels of uncertainty in estimates of individual
LGA random effects (Link & Sauer 1996; Sauer & Link
2002). Although our model detected major landscape
effects, it is possible that we failed to detect other
land-cover effects due to a thematic resolution that was
too coarse (Cushman & Landguth 2010). Future
research may benefit from a finer habitat resolution, for
example, incorporating the distribution of koala-specific
Eucalyptus trees.
Management implications and conclusions
Evaluating different planning strategies to minimize the
impact of landscape change on species connectivity is
critical for conserving biodiversity (Epps et al. 2007).
The variations in IBR relationships identified in this
study provide several scale-dependent insights to guide
conservation decision-making for the koala. At a finer
resolution, targeting the preservation and restoration of
forest cover to a minimum 30% FPC (i.e. of food trees)
and facilitating movement across highways and
freeways are important. At the broader LGA resolution,
improving landscape connectivity within LGAs or
between geographically proximate LGAs may be more
important than among geographically distant LGAs.
This is because the effect of landscape resistance on
gene flow appears to give way to the effect of genetic
drift for LGAs that are separated by larger geographic
distances. Thus, improving connectivity may be best
directed towards resolutions at which gene flow
responds to landscape resistance. Generally increasing
the amount of forest within LGAs may also be an
important strategy for maintaining gene flow. The
choice of analysis scale can dramatically affect the infer-
ences we make about landscape-genetic processes
(Anderson et al. 2010). Multilevel models offer a robust
way to explicitly model landscape-genetic patterns at
resolutions appropriate to both genetic processes and
conservation management.
Acknowledgements
This project was conducted with approval from the Animal
Ethics Committee of the University of Queensland (approval
number GPA/359/09/ARC). The project was funded by the
Australian Research Council (ARC), the Queensland Museum,
Moreton Bay Regional Council, Redland City Council, Logan
City Council, Gold Coast City Council and the Queensland
Department of Environment and Heritage Protection. We
thank Matthew Warren for help with landscape data prepara-
tion, Camryn Allen for providing samples from Burbank and
Greg Simmons for providing Gold Coast samples collected
by Steve Phillips. We are grateful to the staff of the Austra-
lian Wildlife Hospital, Moggill Koala Hospital and the
Queensland Department of National Parks, Recreation, Sport
and Racing, for providing koala tissue samples and associ-
ated data.
References
Adams-Hosking C, Grantham HS, Rhodes JR, McAlpine CA,
Moss PT (2011) Modelling climate-change-induced shifts in
the distribution of the koala. Wildlife Research, 38, 122–130.
© 2013 John Wiley & Sons Ltd
3762 R. Y . DUDANIEC ET AL.

Akaike H (1998) Information theory as an extension of the
maximum likelihood principle. In: Selected Papers of Hirotugu
Akaike (eds Parzen E, Tanabe K & Kitagawa G), pp. 199–213.
Springer, New York.
Anderson CD, Epperson BK, Fortin M-J et al. (2010) Consider-
ing spatial and temporal scale in landscape-genetic studies
of gene flow. Molecular Ecology, 19, 3565–3575.
Balkenhol N, Waits LP (2009) Molecular road ecology: explor-
ing the potential of genetics for investigating transportation
impacts on wildlife. Molecular Ecology, 18, 4151–4164.Barbieri MM, Berger JO (2004) Optimal predictive model selec-
tion. Annals of Statistics, 32, 870–897.Burnham KP, Anderson DR (2002) Model Selection and Multi-
Model Inference: A Practical Information Theoretic Approach, 2nd
edn. Springer-Verlag, New York.
Chiucci JE, Gibbs HL (2010) Similarity of contemporary and
historical geneflow among highly fragmented populations of
an endangered rattlesnake. Molecular Ecology, 19, 5435–5358.Cook RD (1977) Detection of influential observation in linear
regression. Technometrics, 19, 15–18.Crainiceanu CM, Ruppert D (2004) Likelihood ratio tests in
linear mixed models with one variance component. Journal of
the Royal Statistical Society Series B - Statistical Methodology,
66, 165–185.Crosby MKA, Licht LE, Fu J (2009) The effect of habitat frag-
mentation on finescale population structure of wood frogs
(Rana sylvatica). Conservation Genetics, 10, 1707–1718.
Cumming GS, Cumming DHM, Redman CL (2006) Scale mis-
matches in social-ecological systems: causes, consequences,
and solutions. Ecology and Society, 11, 14.
Cushman SA, McKelvey KS, Hayden J, Schwartz MK (2006)
Gene flow in complex landscapes: testing multiple hypotheses
with causal modeling. The American Naturalist, 168, 486–499.Cushman SA, Landguth EL (2010) Scale dependent inference
in landscape genetics. Landscape Ecology, 25, 967–979.Department of Environment and Natural Resources (2009)
Decline of the Koala Coast population: Population Status in 2008.
Queensland Government, Brisbane, Australia.
DeAngelis DL, Waterhouse JC (1987) Equilibrium and nonequi-
librium concepts in ecological models. Ecological Monographs,
57, 1–21.Dieringer D, Schlotterer C (2003) Microsatellite analyser (MSA):
a platform independent analysis tool for large microsatellite
data sets. Molecular Ecology Notes, 3, 167–169.
Dique DS, Thompson J, Preece HJ, de Villiers DL, Carrick FN
(2003a) Dispersal patterns in a regional koala population in
south-east Queensland. Wildlife Research, 30, 281–290.Dique DS, Thompson J, Preece HJ et al. (2003b) Koala mortality
on roads in south-east Queensland: the koala speed-zone
trial. Wildlife Research, 30, 419–426.
Dudaniec RY, Spear SF, Storfer A (2012) Current and historical
drivers of landscape genetic structure differ in core and
peripheral salamander populations. PLoS ONE, 7, e36769.
Ellis W, Melzer A, Carrick F, Hasegawa M (2002) Tree use, diet
and home range of the koala (Phascolarctos cinereus) at Blair
Athol, central Queensland. Wildlife Research, 29, 303–311.
Environment Protection and Biodiversity Conservation Act
(1999) Australian Government, Canberra, Australia.
Epps CW, Wehausen JD, Bleich VC, Torres SG, Brashares JS
(2007) Optimizing dispersal and corridor models using land-
scape genetics. Journal of Applied Ecology, 44, 714–724.
Goossens B, Chikhi L, Jalil F et al. (2005) Patterns of genetic
diversity and migration in increasingly fragmented and
declining orang-utan (Pongo pygmaeus) populations from
Sabeh, Malaysia. Molecular Ecology, 14, 441–456.Gordon G (1991) Estimation of the age of the koala, Phascolarc-
tos cinereus (Marsupialia: Phascolarctidae) from tooth wear
and growth. Australian Mammalogy, 14, 5–12.
Hilborn R, Mangel M (1997) The Ecological Detective. Confronting
Models with Data. Princeton University Press, Princeton, New
Jersey.
Holzhauer SIJ, Ekschmitt K, Sander AC, Dauber J, Wolters V
(2006) Effect of historic landscape change on the genetic
structure of the bush-cricket Metrioptera roeseli. Landscape
Ecology, 21, 891–899.Houlden BA, England P, Eldridge MDB (1996) Paternity exclu-
sion in koalas (Phascolarctos cinereus) using hypervariable
microsatellite loci. Journal of Heredity, 87, 149–152.
Hutchison DW, Templeton AR (1999) Correlation of pairwise
genetic and geographic distance measures: inferring the rela-
tive influences of gene flow and drift on the distribution of
genetic variability. Evolution, 53, 1898–1914.
Jackson ND, Fahrig L (2011) Relative effects of road mortality
and decreased connectivity on population genetic diversity.
Biological Conservation, 144, 3143–3148.Kittlein MJ, Gaggiotti OE (2008) Interactions between envi-
ronmental factors can hide isolation by distance patterns: a
case study of Ctenomys rionegrensis and Uraguay. Proceed-
ings of the Royal Society B: Biological Sciences, 275, 2633–
2638.
Koen EL, Garroway CJ, Wilson PJ, Bowman J (2010) The effect
of map boundary on estimates of landscape resistance to animal
movement. PLoS ONE, 5, e11785.
Lancaster ML, Taylor AC, Cooper SJB, Carthew SM (2011) Lim-
ited ecological connectivity of an arboreal marsupial across a
forest/plantation landscape despite apparent resilience to
fragmentation. Molecular Ecology, 20, 2258–2271.
Landguth EL, Cushman SA, Schwartz MK et al. (2010) Quanti-
fying the lag time to detect barriers in landscape genetics.
Molecular Ecology, 19, 4179–4191.Landguth EL, Fedy BC, Oyler-McCance O et al. (2012) Effects
of sample size, number of markers, and allelic richness on
the detection of spatial genetic pattern. Molecular Ecology
Resources, 12, 276–284.Lee KE, Seddon JM, Corely SW et al. (2010) Genetic variation
and structuring in the threatened koala populations of
Southeast Queensland. Conservation Genetics, 11, 2091–2103.
Link W, Sauer J (1996) Extremes in ecology: avoiding the
misleading effects of sampling variation in summary analyses.
Ecology, 77, 1633–1640.Lowe WH, Allendorf FW (2010) What can genetics tell us
about population connectivity? Molecular Ecology, 19, 3038–3051.
Lyons M, Phinn SR, Roelfsema CM (2012) Long term land
cover and seagrass mapping using Landsat and object-based
image analysis from 1972 to 2010 in the coastal environment
of South East Queensland, Australia. ISPRS Journal of Photo-
grammetry and Remote Sensing, 71, 34–46.Macdonald DW, Johnson DDP (2001) Dispersal in theory and
practice: consequences for conservation biology. In: Dispersal
(eds Clobert J, Danchin E, Dhondt AA & Nichols JD), pp.
358–372. Oxford University Press, Oxford.
© 2013 John Wiley & Sons Ltd
MULTILEVEL MODELS AND LANDSCAPE GENETICS 3763

Manel S, Schwartz MK, Luikart G, Taberlet P (2003) Landscape
genetics: combining landscape ecology and population genet-
ics. Trends in Ecology & Evolution, 18, 189–197.
McAlpine CA, Bowen ME, Callaghan JG et al. (2006a) Testing
alternative models for the conservation of koalas in frag-
mented rural-urban landscapes. Austral Ecology, 31, 529–544.McAlpine CA, Rhodes JR, Bowen ME et al. (2006b) The impor-
tance of forest area and configuration relative to local habitat
factors for conserving forest mammals: a case study of koalas
in Queensland, Australia. Biological Conservation, 132,
153–165.
McAlpine CA, Rhodes JR, Bowen ME et al. (2008) Can multi-
scale models of species’ distribution be generalized from
region to region? A case study of the koala. Journal of Applied
Ecology, 45, 558–567.
McMahon SM, Diez JM (2007) Scales of association: hierarchi-
cal linear models and the measurement of ecological
systems. Ecology Letters, 10, 437–452.McRae BH (2006) Isolation by resistance. Evolution, 60, 1551–
1561.
McRae BH, Beier P (2007) Circuit theory predicts gene flow in
plant and animal populations. Proceedings of the National
Academy of Sciences, USA, 104, 19885–19890.
Metzger JP, Martensen AC, Dixo M et al. (2009) Time-lag in
biological response to landscape changes in a highly dynamic
Atlantic forest region. Biological Conservation, 142, 1166–1177.Munshi-South J (2012) Urban landscape genetics: canopy cover
predicts gene flow between white-footed mouse (Peromyscus
leucopus) populations from New York City. Molecular Ecology,
21, 1360–1378.
Murphy MA, Dezzani R, Pilliod DS, Storfer A (2010) Land-
scape genetics of high mountain frog metapopulations.
Molecular Ecology, 19, 3634–3649.Niedziałkowska M, Fontaine MC, Jezdrzejewska B (2012)
Factors shaping geneflow in red deer (Cervus elaphus) in
seminatural landscapes of central Europe. Canadian Journal of
Zoology, 90, 150–162.Novembre J, Slatkin M (2009) Likelihood-based inference in
isolation-by-distance models using the spatial distribution of
low-frequency alleles. Evolution, 63, 2914–2925.
van Oosterhout C, Hutchinson WF, Wills DPM, Shipley P
(2004) MICRO-CHECKER: software for identifying and
correcting genotyping errors in microsatellite data. Molecular
Ecology, 4, 535–538.
Orsini L, Corander J, Alasentie A, Hanski I (2008) Genetic
spatial structure in a butterfly metapopulation correlates better
with past than present demographic structure. Molecular
Ecology, 17, 2629–2642.
Peakall R, Smouse PE (2006) Genalex 6: genetic analysis in
Excel. Population genetic software for teaching and research.
Molecular Ecology Notes, 6, 288–295.Pelosi C, Goulard M, Balent G (2010) The spatial scale
mismatch between ecological processes and agricultural
management: do difficulties come from underlying theoretical
frameworks? Agriculture, Ecosystems and Environment, 139,
455–462.
Peterson MA, Denno RF (1998) The influence of dispersal and
diet breadth on patterns of genetic isolation by distance in
phytophagous insects. American Naturalist, 152, 428–446.Phillips SS (2000) Population trends and the koala conservation
debate. Conservation Biology, 14, 650–659.
Pinheiro J, Bates D (2000) Mixed Effects Models in S and S-Plus.
Springer-Verlag, New York.
Qian SS, Cuffney TF, Alameddine I, McMahon G, Reckhow
KH (2010) On the application of multilevel modeling in envi-
ronmental and ecological studies. Ecology, 91, 355–361.
Qu�em�er�e E, Crouau-Roy B, Rabarivola C, Louis EE, Chikhi L
(2010) Landscape genetics of an endangered lemur (Propithe-
cus tattersalli) within its entire fragmented range. Molecular
Ecology, 19, 1606–1621.
Rasic G, Keyghobadi N (2012) From broadscale patterns to
fine-scale processes: habitat structure influences genetic
differentiation in the pitcher plant midge across multiple
spatial scales. Molecular Ecology, 21, 223–236.
Raymond M, Rousset F (1995a) GENEPOP (Version 1.2): popu-
lation genetics software for exact tests and ecumenicism.
Journal of Heredity, 86, 248–249.Raymond M, Rousset F (1995b) GENEPOP (version 1.2): popu-
lation genetics software for exact tests and ecumenicism.
Journal of Heredity, 86, 248–249.
Rhodes JR, McAlpine CA, Lunney D, Possingham HP (2005) A
spatially explicit habitat selection model incorporating home
range behaviour. Ecology, 86, 1199–1205.Rhodes JR, Callaghan JG, McAlpine CA et al. (2008) Regional
variation in habitat-occupancy thresholds: a warning for con-
servation planning. Journal of Applied Ecology, 45, 549–557.
Rhodes JR, Ng CF, de Villiers DL et al. (2011) Using integrated
population modelling to quantify the implications of multi-
ple threatening processes for a rapidly declining population.
Biological Conservation, 144, 1081–1088.Rousset F (2000) Genetic differentiation between individuals.
Journal of Evolutionary Biology, 13, 58–62.Sauer J, Link W (2002) Hierarchical modeling of population
stability and species group attributes from survey data.
Ecology, 83, 1743–1751.
Seabrook L, McAlpine CA, Baxter G et al. (2011) Drought-driven
change in wildlife distribution and numbers: a case study
of koalas in south-west Queensland. Wildlife Research, 38,
509–524.
Sherwin WB, Timms P, Wilcken J, Houlden B (2000) Analysis
and conservation implications of koala genetics. Conservation
Biology, 14, 639–649.Shirk AJ, Wallin DO, Cushman SA, Rice CG, Warheit KI (2010)
Inferring landscape effects on gene flow: a new model selec-
tion framework. Molecular Ecology, 19, 3603–3619.
Slatkin M (1993) Isolation by distance in equilibrium and non-
equilibrium populations. Evolution, 47, 264–279.
Smith AG, McAlpine CA, Rhodes JR et al. (2013) At what
spatial scales does resource selection vary? A case study of
koalas in a semi-arid region. Austral Ecology, 38, 230–240.Spear SF, Balkenhol N, Fortin M-J, McRae BH, Scribner KIM
(2010) Use of resistance surfaces for landscape genetic
studies: considerations for parameterization and analysis.
Molecular Ecology, 19, 3576–3591.Specht R (1983) Foliage projective covers of overstorey and un-
derstorey strata of mature vegetation in Australia. Australian
Journal of Ecology, 8, 433–439.
Steele CA, Baumsteiger J, Storfer A (2009) Influence of life-history
variation on the genetic structure of two sympatric salamander
taxa.Molecular Ecology, 18, 1629–1639.Storfer A, Murphy MA, Evans JS et al. (2007) Putting the ‘land-
scape’ in landscape genetics. Heredity, 98, 128–142.
© 2013 John Wiley & Sons Ltd
3764 R. Y . DUDANIEC ET AL.

Taylor BL, Dizon AE (1999) First policy then science: why a
management unit based soley on genetic criteria cannot
work. Molecular Ecology, 8, S11–S16.
Taylor AC, Tyndale-Biscoe H, Lindenmayer DB (2007) Unex-
pected persistence on habitat islands: genetic signatures
reveal dispersal of a eucalypt-dependent marsupial through
a hostile pine matrix. Molecular Ecology, 16, 2655–2666.
Wasserman TN, Cushman SA, Schwartz MK, Wallin DO (2010)
Spatial scaling and multi-model inference in landscape
genetics: Martes americana in northern Idaho. Landscape Ecology,
25, 1601–1612.
Whittingham MJ, Krebs JR, Swetnam RD et al. (2007) Should
conservation strategies consider spatial generality? Farmland
birds show regional not national patterns of habitat associa-
tion. Ecology Letters, 10, 25–35.
With K, Crist T (1995) Critical thresholds in species’ responses
to landscape structure. Ecology, 76, 2446–2459.
Worthington-Wilmer J, Elkin C, Wilcox C et al. (2008) The
influence of multiple dispersal mechanisms and landscape
structure on population clustering and connectivity in frag-
mented artesian spring snail populations. Molecular Ecology,
17, 3733–3751.Xu RH (2003) Measuring explained variation in linear mixed
effects models. Statistics in Medicine, 22, 3527–3541.Zellmer AJ, Knowles L (2009) Disentangling the effects of historic
vs. contemporary landscape structure on population genetic
divergence.Molecular Ecology, 18, 3593–3602.
Data accessibility
Final microsatellite data set, koala sample coordinates,
genetic distance matrix, land-cover data, best resistance
layer and R scripts are uploaded as online supporting
information.
JRR, RYD, JWW and CAM designed the research, FNC
contributed to sample collection, JWW contributed
molecular analytical tools, KEL conducted laboratory
work and initial analysis, RYD performed the genetic
analyses, ML provided and processed the landscape
data, ML, RYD and JRR performed the geographical
analyses, JRR performed the statistical modelling, RYD
and JRR wrote the paper, JWW, ML, KEL, CAM and
FNC provided edits to the paper.
Supporting information
Additional supporting information may be found in the online ver-
sion of this article.
Table S1 Locus characteristics for the koala (Phascolarctos cinereus)
within each local government area (LGA, with map ID, Fig. 1)
with sample size (n), coordinates, number of alleles per locus (Na),
observed heterozygosity (Ho), and expected heterozygosity (He).
Table S2 Description of land cover (adapted from Lyons et al.
2012) and foliage projective cover (FPC) variables used in the
landscape genetic analysis for koalas.
Table S3 Pairwise Fst between eight Local Government Areas
(LGAs).
Appendix S1 DNA extraction, genotyping and locus characteris-
tics.
Appendix S2Microsatellite locus tests.
Fig. S1 Parameter value selection frequencies across bootstrap rep-
licates for the (A) a and (B) c parameters that control the relation-
ship between Foliage Projective Cover (FPC) and resistance.
Fig. S2 Parameter value selection frequencies across bootstrap rep-
licates for the (A) b and (B) g parameters that control the relation-
ship between land cover and resistance.
Fig. S3 Percentage of forest cover for the eight Local Government
Areas (LGAs) within the 3.5 km buffers around the sample points
between 1988 and 2009.
Fig. S4 Percentage of urban cover for the eight Local Government
Areas (LGAs) within the 3.5 km buffers around the sample points
between 1988 and 2009.
Fig. S5 Percentage of forest cover within each Local Government
Area (LGA) for 1972 calculated within 3.5 km buffers around sam-
pling points used to calculate predictor variables (grey) and for the
total extent of each LGA (black).
Fig. S6 Percentage urban cover within each Local Government
Area (LGA) for 2009 calculated within 3.5 km buffers around
sampling points used to calculate predictor variables (grey) and
for the total extent of each LGA (black).
Fig. S7 For each LGA, the mean and 95% confidence interval of
Cook’s Distances (Cook 1977; i.e. a measure of the influence
of data point(s) on parameter estimates of a model) across pairs of
LGAs where at least one of the pair is the LGA of interest.
© 2013 John Wiley & Sons Ltd
MULTILEVEL MODELS AND LANDSCAPE GENETICS 3765




















