

MT1-MMP proinvasive activity is regulated by a novel Rab8-dependent exocytic pathway
Upload
independentCategory
view
1download
0
10.1128/MCB.24.12.5447-5458.2004.
2004, 24(12):5447. DOI:Mol. Cell. Biol. Kevin Fogarty, Silvia Corvera and Michael P. CzechAvirup Bose, Stacey Robida, Paul S. Furcinitti, Anil Chawla, PathwayMembrane Fusion in a Regulated Exocytic Unconventional Myosin Myo1c Promotes
http://mcb.asm.org/content/24/12/5447Updated information and services can be found at:
These include:
SUPPLEMENTAL MATERIAL Supplemental material
REFERENCEShttp://mcb.asm.org/content/24/12/5447#ref-list-1at:
This article cites 43 articles, 19 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

MOLECULAR AND CELLULAR BIOLOGY, June 2004, p. 5447–5458 Vol. 24, No. 120270-7306/04/$08.00�0 DOI: 10.1128/MCB.24.12.5447–5458.2004Copyright © 2004, American Society for Microbiology. All Rights Reserved.
Unconventional Myosin Myo1c Promotes Membrane Fusion in aRegulated Exocytic Pathway†
Avirup Bose,1 Stacey Robida,1 Paul S. Furcinitti,1 Anil Chawla,1 Kevin Fogarty,2Silvia Corvera,1 and Michael P. Czech1*
Program in Molecular Medicine1 and Biomedical Imaging Group,2 Department of Physiology,University of Massachusetts Medical School, Worcester, Massachusetts 01605
Received 17 October 2003/Returned for modification 17 November 2003/Accepted 19 March 2004
Glucose homeostasis is controlled in part by regulation of glucose uptake into muscle and adipose tissue.Intracellular membrane vesicles containing the GLUT4 glucose transporter move towards the cell cortex inresponse to insulin and then fuse with the plasma membrane. Here we show that the fusion step is retardedby the inhibition of phosphatidylinositol (PI) 3-kinase. Treatment of insulin-stimulated 3T3-L1 adipocytes withthe PI 3-kinase inhibitor LY294002 causes the accumulation of GLUT4-containing vesicles just beneath the cellsurface. This accumulation of GLUT4-containing vesicles near the plasma membrane prior to fusion requiresan intact cytoskeletal network and the unconventional myosin motor Myo1c. Remarkably, enhanced Myo1cexpression under these conditions causes extensive membrane ruffling and overrides the block in membranefusion caused by LY294002, restoring the display of GLUT4 on the cell exterior. Ultrafast microscopic analysisrevealed that insulin treatment leads to the mobilization of GLUT4-containing vesicles to these regions ofMyo1c-induced membrane ruffles. Thus, localized membrane remodeling driven by the Myo1c motor appearsto facilitate the fusion of exocytic GLUT4-containing vesicles with the adipocyte plasma membrane.
The plasma membranes of living cells undergo constant re-cycling through the dynamic processes of membrane retrievaland membrane insertion. Through these endocytic and exo-cytic events, which include intermediate membrane transportpathways, the relative abundance of specific plasma membranecomponents and the secretion of molecules can be regulated.Complex mechanisms involving protein-protein interactionshave evolved to coordinate such membrane trafficking pro-cesses to ensure that transported membranes are appropriatelytargeted to their specific destinations (36). These mechanismsinclude directed movements on cytoskeletal tracks (39) and theconcerted actions of tethering proteins that anchor transportvesicles to cognate target membranes (27). The membranefusion step in exocytosis is highly dependent on proteins thatjoin membranes in close proximity such that two separate lipidbilayers merge into one (16). One class of proteins that func-tion in this fusion step are the SNAREs (35), of which thesynaptic vesicle proteins synaptobrevin/VAMP (15) and syn-taxin 1 (1), as well as SNAP-25 (13) on the plasma membrane,are best characterized. Although SNAREs are clearly impor-tant for fusion, they may not be involved in directly executingthe fusion reaction. The hypothesis that several other cofactorsare necessary to complete fusion is supported by the observa-tion that at least under some specialized conditions the defi-ciencies of certain SNAREs do not prevent fusion (5, 9, 32, 42).Furthermore, specialized exocytic systems such as those inneuronal synapses contain unique proteins that facilitate tar-
geting or regulation of membrane fusion (28). Thus, the mo-lecular details of membrane fusion processes remain an activearea of investigation.
Among a number of newly discovered proteins that havebeen implicated in the membrane targeting and fusion pro-cesses are the SM family of proteins (38), the septins (12), andRIM (40) and associated proteins. A recent report has alsosuggested the involvement of a class V myosin, myo52, infission yeast in mediating vacuole fusion under osmotic stress,providing the first link between an actin-based motor and ho-motypic membrane fusion (23). This suggestion is particularlyinteresting in light of findings in our laboratory that a myosinI family member (Myo1c) is required for optimal insulin-stim-ulated translocation of intracellular membranes containingGLUT4 glucose transporters to the plasma membrane (3). Inthis membrane trafficking system, GLUT4 recycles betweenintracellular and plasma membrane compartments and insulinacutely stimulates GLUT4 exocytosis through a phosphatidyl-inositol (PI) 3-kinase-dependent pathway (6, 19, 21, 24, 34).The detailed mechanism by which GLUT4-containing mem-branes fuse with the plasma membrane requires interactionbetween syntaxin 4 (t-SNARE) (41) and VAMP-2 (v-SNARE)(7). However, it is not known which components or processesthat function in the GLUT4 recycling pathway are directlydownstream of PI 3-kinase signaling or require the myosinMyo1c.
The aim of the present studies was to characterize the role ofMyo1c in the trafficking pathway of GLUT4-containing mem-branes and its relationship to PI 3-kinase-sensitive steps. Pre-vious work had shown that the expression of high levels ofMyo1c in cultured adipocytes enhances the extent to whichGLUT4 is translocated to the plasma membrane in response toinsulin (3). In other recent studies, PI 3-kinase signaling wasimplicated in the fusion step of exocytosis of GLUT4-contain-
* Corresponding author. Mailing address: Program in MolecularMedicine, Department of Physiology, University of MassachusettsMedical School, 55 Lake Ave. North, Worcester, MA 01605. Phone:(508) 856-2254. Fax: (508) 856-1617. E-mail: [email protected].
† Supplemental material for this article may be found at http://mcb.asm.org/.
5447
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
5448 BOSE ET AL. MOL. CELL. BIOL.
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

ing membranes (25). Consistent with this hypothesis, we reporthere that the blockade of PI 3-kinase inhibits the fusion ofGLUT4-containing membrane vesicles with the plasma mem-brane and causes the accumulation of these vesicles just be-neath the cell surface. Remarkably, high expression of Myo1ccould override the block in membrane fusion caused by PI3-kinase inhibition when insulin is also present. These datasuggest that Myo1c drives a process that promotes the fusion ofGLUT4-containing vesicles with the plasma membrane.
MATERIALS AND METHODS
Materials and chemicals. LY294002 was purchased from BIOMOL. Mouseanti-Myc (clone 9E10) monoclonal antibody was purchased from NeomarkersInc. Rhodamine-labeled goat anti-mouse antibodies and BODIPY 581/591 werepurchased from Molecular Probes.
DNA constructs. The construction of Myc-GLUT4-GFP, Myc-GLUT4-CFP,and YFP-Myo1c has been described previously (3, 17). The YFP-Myo1c(T)plasmid was constructed by subcloning a Myo1c coding sequence encompassingresidues 767 to 1028, amplified by PCR, into the BamHI and XhoI restrictionsites of pEYFPC1 in frame with yellow fluorescent protein (YFP). The constructwas sequenced, and its expression was verified in COS-1 cells prior to experi-ments with 3T3-L1 adipocytes.
Cell culture, cell treatments, and transfection of differentiated 3T3-L1 adipo-cytes. 3T3-L1 fibroblasts were grown to confluence and differentiated as de-scribed previously (3). Differentiated adipocytes were transfected on the fifth daypostdifferentiation by electroporation as described previously. The cells werethen allowed to recover for 24 h before serum starvation in Dulbecco modifiedEagle medium plus 0.5% bovine serum albumin for 5 h and then treated with thereagents described in the figure legends. Cells were pretreated with either 5 �Mlatrunculin and 50 �M colchicine for 1 h or 100 �M LY294002 for 15 min priorto stimulation with 100 nM insulin for 30 min.
Myc-GLUT4-CFP internalization assay. Differentiated 3T3-L1 adipocyteswere electroporated either with Myc-GLUT4-CFP alone or with YFP-Myo1c, asindicated. After 24 h, cells were stimulated with insulin for 1 h to allow theMyc-GLUT4-CFP to translocate to the cell surface. Cells were then washedtwice with ice-cold phosphate-buffered saline, incubated at 4°C with monoclonalanti-Myc antibodies for 1 h, washed twice with ice-cold phosphate-bufferedsaline, and warmed to 37°C for the indicated times in order to allow the Myc-GLUT4-CFP that had translocated to the cell surface in presence of insulin tointernalize. Cells were then fixed and permeabilized, and the Myc-GLUT4-CFP-expressing cells were detected by staining with rhodamine-conjugated anti-mouse secondary antibodies.
Live-cell imaging. In live-cell imaging experiments, adipocytes were seeded in35-mm-diameter plastic tissue culture dishes with glass coverslip bottoms (Mat-Tek). Images of fluorescently labeled live cells were obtained with an IX 70inverted microscope (Olympus) with a �100 NA 1.4 objective lens, a CoolsnapHQ (Roper Scientific) digital camera, and an excitation filter wheel and shutter(Sutter) in the epifluorescence light path and an emission filter wheel (Sutter) inthe imaging light path. Metamorph Image acquisition and analysis software(Universal Imaging) controlled the hardware and acquired the data. Time lapseimages were taken every 5 s for 10 to 15 min by using rhodamine and YFP filters(Chroma). For Fig. 5, the images were deblurred by using the Metamorphno-neighbors algorithm, in which images are reblurred by convolution with themicroscope point spread function and a fraction of the reblurred image is sub-tracted from the original blurred image. The microscope point spread functionwas calculated from the values of the numerical aperture of the objective lens,the wavelength of the fluorescence, and the size of the image pixels.
Ultrafast microscopy. Differentiated 3T3-L1 adipocytes expressing YFP-Myo1c and Myc-GLUT4-CFP and were imaged 24 h after transfection by usinghigh-speed, three-dimensional microscopy (8, 26, 30, 43). Cyan fluorescent pro-tein (CFP) and YFP donor-acceptor pairs were imaged by using a CoherentArgon laser tuned to 458 nm, a wavelength at which both fluorophores weresimultaneously excited. The 128- by 128-pixel camera field was split into twosubfields of 128 by 60 pixels by using a dual-view module (Optical Insights, SantaFe, N.M.). The donor and acceptor fields were filtered by 480/30 and 535/40emission filters, respectively. This optical configuration insured true simultaneityof donor-acceptor excitation and emission. Two-dimensional images of both CFPand YFP were acquired by using exposure times of 5 ms. Each three-dimensionalimage set consisted of 21 optical sections spaced 250 nm apart, with an additional20 ms being allowed for changing focus to the next optical section. A three-
FIG
.1.
PI3-
kina
sein
hibi
tor
LY
2940
02bl
ocks
insu
lin-s
timul
ated
fusi
onof
GL
UT
4-co
ntai
ning
vesi
cles
with
the
plas
ma
mem
bran
e.(A
)D
iffer
entia
ted
3T3-
L1
adip
ocyt
esex
pres
sing
Myc
-GL
UT
4-G
FP
wer
ese
rum
star
ved
and
wer
eth
enei
ther
left
untr
eate
d(C
ontr
ol),
trea
ted
with
colc
hici
nean
dla
trun
culin
B(C
ontr
ol�
Col
�L
atB
),st
imul
ated
with
insu
linan
dtr
eate
dw
ithL
Y29
4002
follo
wed
byin
sulin
(LY
�In
sulin
),tr
eate
dw
ithco
lchi
cine
and
latr
uncu
linB
follo
wed
byin
sulin
(Col
�L
atB
�in
sulin
),or
trea
ted
with
LY
2940
02fo
llow
edby
insu
linfo
r30
min
and
then
with
colc
hici
nean
dla
trun
culin
Bfo
r1
h(L
Y�
Insu
lin�
Col
�L
atB
).T
hece
llsw
ere
then
fixed
and
stai
ned
with
anti-
Myc
follo
wed
byrh
odam
ine-
labe
led
seco
ndar
yan
tibod
y.(B
)F
our
field
sof
diffe
rent
iate
d3T
3-L
1ad
ipoc
ytes
expr
essi
ngM
yc-G
LU
T4-
GF
Pun
der
seru
mst
arva
tion
oraf
ter
trea
tmen
tw
ithL
Y29
4002
(LY
)an
din
sulin
from
the
expe
rim
ent
pres
ente
din
pane
lAar
esh
own.
(C)
Cel
lstr
eate
dw
ithth
ere
agen
tsm
entio
ned
abov
efo
rpa
nelA
wer
eco
unte
dfo
rbo
thG
FP
and
Myc
rim
s.A
tlea
st20
0ce
llsfo
rea
chco
nditi
onw
ere
coun
ted
and
scor
edbl
indl
yfo
rG
FP
and
Myc
rim
s.T
heim
ages
wer
eal
ltak
enat
the
sam
eex
posu
re.T
hepe
rcen
tage
sof
cells
with
GF
Pan
dM
ycri
ms
are
show
n.T
here
sults
refle
ctav
erag
esfo
rth
ree
iden
tical
expe
rim
ents
.
VOL. 24, 2004 Myo1c AND GLUT4 5449
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

FIG
.2.
Myo
1cis
requ
ired
tolo
caliz
ein
sulin
-stim
ulat
edG
LU
T4-
cont
aini
ngve
sicl
escl
ose
toth
epl
asm
am
embr
ane.
(A)
Diff
eren
tiate
d3T
3-L
1ad
ipoc
ytes
expr
essi
ngM
yc-G
LU
T4-
CF
Pan
dY
FP-
Myo
1c(T
)w
ere
seru
mst
arve
dan
dei
ther
left
untr
eate
d(C
ontr
ol),
trea
ted
with
insu
lin(I
nsul
in),
ortr
eate
dw
ithL
Y29
4002
follo
wed
byin
sulin
(LY
�In
sulin
).T
hece
llsw
ere
then
fixed
and
stai
ned
with
anti-
Myc
follo
wed
byrh
odam
ine-
labe
led
seco
ndar
yan
tibod
y.C
ells
expr
essi
ngbo
thY
FP-
Myo
1c(T
)an
dM
yc-G
LU
T4-
CF
Par
esh
own.
(B)
Cel
lsex
pres
sing
only
Myc
-GL
UT
4-C
FP.
(C)
Cel
lsfr
ompa
nels
Aan
dB
wer
eco
unte
dfo
rM
ycri
ms
atth
ece
llsu
rfac
es.(
D)
Cel
lsfr
ompa
nels
Aan
dB
wer
eco
unte
dfo
rC
FP
sign
als
atth
ece
llsu
rfac
es.M
ore
than
100
cells
wer
esc
ored
blin
dly
for
Myc
and
CF
Pri
ms.
The
data
repr
esen
tav
erag
esfo
rth
ree
sim
ilar
expe
rim
ents
.
5450 BOSE ET AL. MOL. CELL. BIOL.
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

dimensional image set was acquired in less than 600 ms. In this manner, three-dimensional images were acquired every 10 s for 1,000 continuous seconds (16.7min).
The images were first corrected for camera dark current. Next, since bothfluorophores were simultaneously excited by the 458-nm laser line, cells express-ing CFP only and cells expressing only YFP were imaged to determine both therelative alignment of the CFP and YFP subfields and the percentages of YFPfluorescence in the CFP channel and of CFP fluorescence in the YFP channel(data not shown). YFP bleed-through into the CFP image was negligible. CFPbleed-through into the spatially corresponding YFP pixel (after realignment) wasdetermined as follows: YFP � YFP � (1.09 � CFP). Finally, the haze originatingfrom light sources outside the in-focus plane of the cell was reduced by imagerestoration (7). The microscope point spread function was empirically deter-mined by imaging 190-nm fluorescent beads with the exact same optical config-uration used for the CFP-YFP data.
For analysis of the concentration of plasma membrane-associated GLUT4 inruffling versus nonruffling areas shown in Fig. 6, a three-dimensional time seriesimage set of Myc-GLUT4-CFP and YFP-Myo1c was first subjected to intensitythresholding. Separate thresholds for the Myc-GLUT4-CFP and YFP-Myo1cimages were chosen such that 95% of the cytosolic pixels were excluded in bothcases. Then, the average fluorescence intensity of the above-threshold pixels inthe GLUT4 image series in regions with and without apparent ruffling wascomputed for each time point. The selected intensity threshold value was halvedand doubled to recalculate the GLUT4-CFP fluorescence in order to investigatethe sensitivity of the analysis (data not shown). The results were essentially thesame.
RESULTS AND DISCUSSION
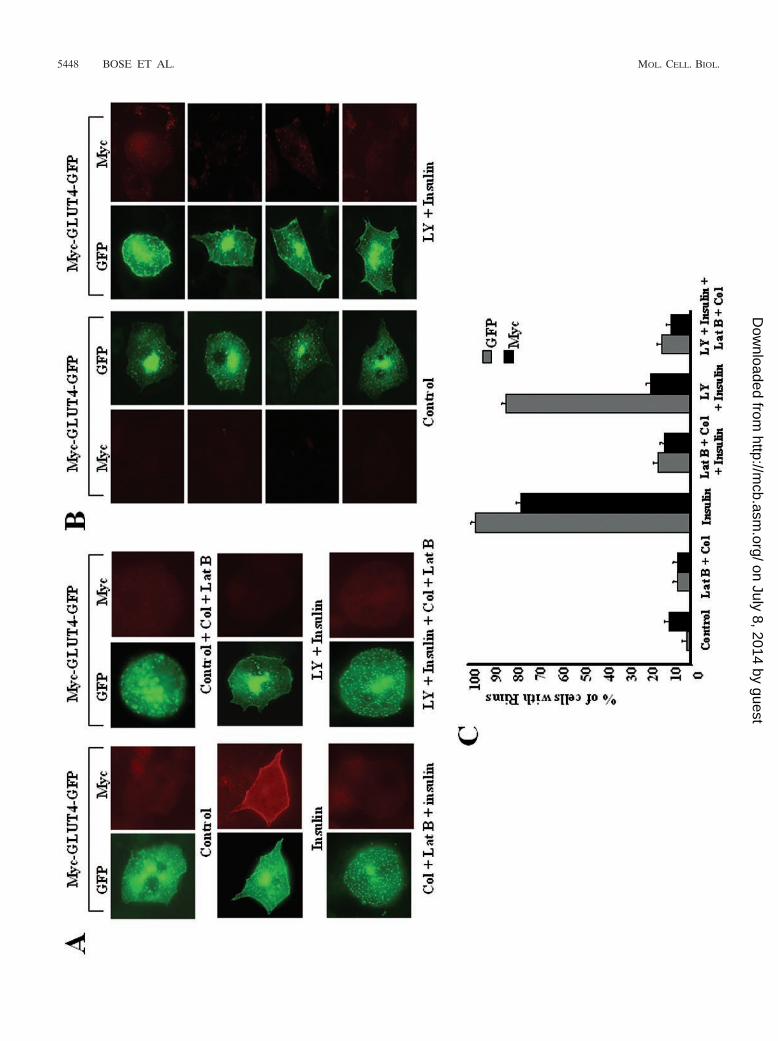
PI 3-kinase is required for fusion of GLUT4-containing ves-icles with the plasma membrane. To differentiate betweenGLUT4 vesicles that translocate to the cell periphery in re-sponse to insulin and those that actually fuse with the plasmamembrane, we employed a GLUT4 construct (Myc-GLUT4-EGFP) containing a Myc epitope in an exofacial loop of thetransporter and a green fluorescent protein (GFP) fusion at itscytoplasmic COOH terminus (17). The fusion of intracellularvesicles containing this construct with the plasma membraneexposes the Myc epitope to reaction with Myc antibody addedto intact adipocytes. As shown in Fig. 1, little or no Mycantibody was detected on the cell surfaces of unstimulated3T3-L1 adipocytes expressing this Myc-GLUT4-EGFP pro-tein. Insulin treatment of these cells acutely translocates intra-cellular Myc-GLUT4-EGFP to the cell surface, as evidencedby the rim of GFP signal around the cell periphery and thestriking anti-Myc staining of the cell surface. Consistent withprevious studies (33), the anti-Myc signal is markedly reducedwhen 3T3-L1 adipocytes are treated with the PI 3-kinase in-hibitor LY294002 prior to stimulation with insulin, indicatingthat PI 3-kinase is required for this insulin effect. However,upon careful examination, we noted that in many of these cellstreated with LY294002 and insulin, a significant rim of GFPsignal could be observed in spite of the absence of anti-Mycstaining (Fig. 1A and B) (33). This rim was not observed incells treated with the inhibitor alone. These results are consis-tent with previous data suggesting that GLUT4-containing ves-icles can translocate to the cell periphery in response to insulinthrough movements on cytoskeletal tracks even in the absenceof PI 3-kinase activity (33). Thus, fusion of the vesicles with theplasma membrane is apparently inhibited by LY294002.
Based on these data, we directly tested whether the appear-ance of GLUT4-containing vesicles at the cell periphery wasinfluenced by the disassembly of actin filaments and microtu-bules by using the inhibitors latrunculin B and colchicine, re-
spectively. Figure 1A shows marked dispersion of GLUT4from the perinuclear region of a typical adipocyte in responseto these inhibitors, as well as their ability to decrease theamount of GLUT4 just beneath the plasma membrane in cellstreated with LY294002 and insulin. As previously reported(33), latrunculin B plus colchicine also dramatically inhibitedthe display of the Myc epitope (from Myc-GLUT4-EGFP) onthe cell surface in response to insulin (Fig. 1A). These data arequantified in Fig. 1C and show that over 80% of cells exhibitGFP rims in the presence of the PI 3-kinase inhibitor plusinsulin, whereas less than 20% display such rims when thecytoskeleton is disrupted. Taken together, these data suggestthat insulin treatment of differentiated 3T3-L1 adipocytesleads to the recruitment of GLUT4-containing vesicles fromthe intracellular storage compartment to the cell periphery.This translocation is microtubule and F-actin dependent but isindependent of PI 3-kinase activity. Once these vesicles reachthe cortical region of the cell, they apparently undergo fusionby a process that requires PI 3-kinase activity.
Myo1c is required for accumulation of GLUT4-containingvesicles at the cell cortex. Since pretreatment with LY294002prior to insulin stimulation results in the accumulation ofGLUT4-containing vesicles, presumably anchored to actin fil-aments, near the cortical region of the cell, we sought to testwhether Myo1c is required for this accumulation. A dominant-negative Myo1c(T) construct was expressed in cells that werealso expressing Myc-GLUT4-CFP. This Myo1c(T) truncatedmutant has the ability to bind cargo but is missing the actinbinding region and the motor domain (29). As expected, theexpression of Myo1c(T) significantly inhibited insulin-stimu-lated GLUT4 translocation to the cell surface, as assayed bycounting cells with anti-Myc rims both in the presence and inthe absence of PI 3-kinase activity (Fig. 2A and C). We did notobserve complete inhibition of insulin-stimulated GLUT4translocation in all of the cells expressing Myo1c(T), presum-ably because of the inherent variability in the expression of thedominant-negative Myo1c. As observed before, cells express-ing only Myc-GLUT4-CFP displayed CFP signal around therims of the cells when pretreated with the PI 3-kinase inhibitorLY294002 in the presence of insulin (Fig. 2B and C). However,this cortical accumulation of GLUT4 was significantly inhib-ited in cells expressing the dominant-negative isoform ofMyo1c (Fig. 2A and D). These results suggest that upon insulinstimulation, the movement of GLUT4-containing vesicles tothe cortical region of the cell or their anchoring to the actincytoskeleton prior to fusion requires Myo1c.
Myo1c expression overrides the LY294002-mediated blockin fusion of GLUT4-containing vesicles with the plasma mem-brane. We have recently reported that high expression ofMyo1c augments the insulin-stimulated appearance of GLUT4on the plasma membrane of differentiated 3T3-L1 adipocytes(3). Data presented here (Fig. 1) indicate that PI 3-kinaseregulates the fusion of GLUT4-containing vesicles with theplasma membrane. To test whether Myo1c function can influ-ence the block in fusion caused by the PI 3-kinase inhibitorLY294002, we coexpressed YFP-tagged Myo1c with Myc-GLUT4-CFP in cultured adipocytes. In the absence of insulin,very little GLUT4 was displayed on the cell surface in thepresence or absence of LY294002, as assayed by cell surfaceanti-Myc signal. After insulin stimulation, about 80% of these
VOL. 24, 2004 Myo1c AND GLUT4 5451
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

5452 BOSE ET AL. MOL. CELL. BIOL.
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

intact cells displayed anti-Myc binding at the cell surface, sig-nifying that much of the GLUT4 at the cell surface was in theplasma membrane. Interestingly, the amount of Myc displayedat the cell surface correlated directly with the amount of YFP-Myo1c expression in these cells (Fig. 3B). When these cellswere treated with LY294002 prior to insulin stimulation, mostof the cells expressing YFP-Myo1c displayed cell surface anti-Myc staining, compared to few of the cells expressing Myc-GLUT4-CFP but not YFP-Myo1c (Fig. 3A and C and Fig. SA3in the supplemental material). Quantification of anti-Mycbinding at the cell surface normalized to the expression ofMyc-GLUT4-CFP in these YFP-Myo1c-expressing cells re-vealed about 60% of cells having anti-Myc rims, compared toabout 20% in cells not expressing YFP-Myo1c (Fig. 3D). Thesedata indicate that Myo1c can function to reverse the block inmembrane fusion caused by the blockade of PI 3-kinase activ-ity (Fig. 3C and D).
One interpretation of these findings is that Myo1c potenti-ates fusion by delivering an increased number of exocyticGLUT4-containing vesicles to fusion sites, thus increasing thenumber of possible fusions per unit time. This would tend toincrease the number of fusion events even when fusion ispartially inhibited by LY294002. This explanation is supportedby the fact that there was a small but significant increase in thenumber of fused GLUT4-containing vesicles under basal con-ditions when these cells expressed Myo1c. According to thismodel, in the presence of insulin there are more vesiclespresent at the cell periphery and hence the effect of the ex-pression of Myo1c under these conditions is more pronounced.Another possibility is that Myo1c actually functions in themolecular mechanism of membrane fusion. Opposing theseinterpretations is a potential negative effect Myo1c expressioncould have in reducing the endocytosis of GLUT4 containingvesicles, thereby indirectly increasing the number of these ves-icles at the cell surface.
In order to distinguish between the effects of Myo1c onexocytosis and those on endocytosis, we monitored Myc-GLUT4-CFP internalization from the cell surface in culturedadipocytes expressing high levels of YFP-Myo1c and in thosewith only endogenous Myo1c. In the experiment depicted inFig. 4, adipocytes were initially stimulated with insulin for 30min to allow Myc-GLUT4-CFP to translocate to the cell sur-faces. The cells were then washed to remove insulin, labeledwith anti-Myc antibody at 4°C, warmed at 37°C to allow Myc-GLUT4-CFP internalization, and then fixed at various timepoints. To assess Myc-GLUT4-CFP endocytosis, the cells werethen permeabilized and stained with secondary antibody la-beled with rhodamine. After 5 min of incubation at 37°C, verylittle anti-Myc signal was detected in the cytoplasm of cellsexpressing Myc-GLUT4-CFP with or without YFP-Myo1c,with most of the anti-Myc being displayed at the cell surface.Anti-Myc could be detected within these cells with increasingperinuclear localization after 15- and 40-min incubations, how-ever, and a marked decrease of anti-Myc signal at the cellsurface was observed at these times. However, the amounts ofinternalized anti-Myc in cells expressing both Myc-GLUT4-CFP and YFP-Myo1c and in cells expressing only Myc-GLUT4-CFP were similar (Fig. 4). Although the experimentshown in Fig. 4 is an accepted method for analyzing GLUT4endocytosis, the result obtained 5 min after the removal of
FIG
.3.
Myo
1cex
pres
sion
part
ially
over
com
esth
eL
Y29
4002
-indu
ced
bloc
kin
fusi
onof
GL
UT
4-co
ntai
ning
vesi
cles
.(A
)D
iffer
entia
ted
3T3-
L1
adip
ocyt
esex
pres
sing
Myc
-GL
UT
4-C
FP
and
YF
P-M
yo1c
wer
ese
rum
star
ved
and
then
eith
erle
ftun
trea
ted
(Con
trol
),tr
eate
dw
ithin
sulin
(Ins
ulin
),or
trea
ted
with
LY
2940
02fo
llow
edby
insu
lin(L
Y�
Insu
lin).
The
top
four
row
sof
pane
lssh
owce
llsex
pres
sing
both
Myc
-GL
UT
4-C
FP
and
YF
P-M
yo1c
,an
dth
efif
thro
wof
pane
lssh
owa
cell
expr
essi
ngon
lyM
yc-G
LU
T4-
CF
P.(B
)D
iffer
entia
ted
3T3-
L1
adip
ocyt
esex
pres
sing
Myc
-GL
UT
4-C
FP
and
YF
P-M
yo1c
wer
est
imul
ated
with
insu
lin.T
hece
llsu
rfac
eM
ycsi
gnal
(rho
dam
ine)
inte
nsity
inth
ese
cells
was
quan
titat
edan
dco
mpa
red
with
the
YF
P-M
yo1c
sign
alin
tens
ityin
thes
ece
lls.(
C)
Cel
lssh
own
inpa
nelA
wer
eco
unte
dfo
ran
ti-M
ycri
msa
tthe
cell
surf
aces
.Mor
eth
an10
0ce
llsfo
rea
chco
nditi
onw
ere
scor
edbl
indl
yfo
rM
ycri
ms.
(D)
Qua
ntifi
catio
nof
cell
surf
ace
anti-
Myc
sign
al(r
hoda
min
e)in
tens
ityin
the
cells
show
nin
pane
lA.T
hear
bitr
ary
unit
repr
esen
tsth
era
tioof
the
cell
surf
ace
anti-
Myc
sign
alto
the
tota
lCF
Psi
gnal
inea
chce
ll.T
heda
tare
pres
ent
aver
ages
for
five
sim
ilar
expe
rim
ents
.
VOL. 24, 2004 Myo1c AND GLUT4 5453
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

insulin represents the best estimate of endocytosis, while re-sults obtained after longer times reflect the net results of bothendocytosis and exocytosis. In absence of insulin, the exocyticrate is rather low and is presumed not to influence this assay.These results, combined with the data presented in Fig. 3, areconsistent with the hypothesis that Myo1c potentiates insulin-stimulated Myc-GLUT4-CFP displayed on the cell surface byincreasing the exocytic process rather than inhibiting endocy-tosis. Although these experiments indicate that Myo1c affectsthe exocytosis of GLUT4-containing vesicles, it is not yet clearwhether these effects of Myo1c are specific for GLUT4-con-taining vesicles.
Myo1c expression in differentiated 3T3-L1 adipocytes in-duces active membrane ruffling. An interesting observationmade in the experiments depicted in Fig. 3 and 4 was that theexpression of YFP-Myo1c plus Myc-GLUT4-CFP in differen-tiated 3T3-L1 adipocytes induced dramatic membrane ruffling.Insulin is also known to stimulate actin reorganization and theformation of lamellipodia or membrane ruffles in several celltypes, including 3T3-L1 adipocytes (22), rat1 fibroblasts (10),CHO-T cells (20), and L6 myotubes (37). Actin polymerizationhas been postulated to be a plausible mechanism for drivinglamellipodia and membrane protrusions (2). An alternativemechanism whereby unconventional myosin is responsible fordriving protrusions, with polymerized actin providing the sub-strate on which force is exerted, has been proposed (2, 11). Inorder to better assess Myo1c function in membrane ruffling, weexpressed YFP-Myo1c in differentiated 3T3-L1 adipocytes andobserved these cells by using live-cell fluorescence microscopy.Cultured adipocytes expressing YFP-Myo1c showed dramaticinsulin-independent membrane ruffling (Fig. 5 and accompa-nying movies in Fig. SA1 in the supplemental material), com-pared to little or no ruffling in untransfected control adipo-cytes, as has been observed previously by other groups (18).Cells with high YFP-Myo1c expression did not exhibit in-creased cortical F-actin, as detected by phalloidin staining(data not shown). Insulin also causes membrane ruffling inuntransfected cells but not to the extent seen in YFP-Myo1c-expressing adipocytes (data not shown). Adipocytes expressingYFP-Myo1c were also stained with the intensely fluorescent�-BODIPY 581/591 C5-HPC [4,4-difluoro-5-(4-phenyl-1,3-buta-dienyl)-4-bora-3a,4a-diaza-s-indacene-3-pentanoyl)-1-hexa-decanoyl-sn-glycero-3-phosphocholine], which is commonlyused as a plasma membrane probe. Several areas of these cellsdisplayed continuous ruffles, while the other regions of the cellsurface remained unruffled. Interestingly, the YFP-Myo1c con-centrations in active regions of ruffling membranes were sig-nificantly higher than those in neighboring regions which dis-played little or no ruffling (Fig. 5). In a few cases, we observedareas of the plasma membrane with significant Myo1c concen-trations but which were not ruffling during the period of ob-servation. However, we never observed a membrane ruffle in aregion without significant Myo1c concentrated in that region.These data indicate that in differentiated 3T3-L1 adipocytes,high levels of Myo1c are sufficient to cause extensive mem-brane ruffling without evidence of increased actin polymeriza-tion.
GLUT4-containing vesicles concentrate in areas of extensivemembrane ruffling prior to fusion. Taken together, the datadescribed above indicate that Myo1c functions to promote
FIG
.4.
Myo
1cex
pres
sion
does
not
disr
upt
Myc
-GL
UT
4-C
FP
inte
rnal
izat
ion
toth
epe
rinu
clea
rre
gion
sof
3T3-
L1
adip
ocyt
es.
Diff
eren
tiate
d3T
3-L
1ad
ipoc
ytes
eith
ertr
ansf
ecte
dw
ithM
yc-G
LU
T4-
CF
Pal
one
orco
tran
sfec
ted
with
Myc
-GL
UT
4-C
FP
and
YF
P-M
yo1c
wer
est
imul
ated
with
insu
linfo
r30
min
.The
cells
wer
eth
enla
bele
dw
ithan
ti-M
ycan
tibod
iesa
ndth
enw
arm
edto
37°C
for
the
indi
cate
dtim
esto
allo
wth
eM
yc-G
LU
T4-
CF
Pto
unde
rgo
endo
cyto
sis.
The
cells
wer
eth
enpe
rmea
biliz
edan
dth
enst
aine
dw
ithrh
odam
ine-
conj
ugat
edse
cond
ary
antib
ody
tode
tect
tota
lMyc
-GL
UT
4-C
FP.
The
data
are
repr
esen
tativ
eof
thre
esi
mila
rex
peri
men
ts.
VOL. 24, 2004 Myo1c AND GLUT4 5455
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

membrane ruffling as well as the fusion of GLUT4-containingvesicles with the plasma membrane in the presence of insulin(Fig. 1 to 5). One interpretation of these findings is that mem-brane ruffles and the F-actin that is beneath these ruffles mayplay a role in concentrating GLUT4-containing vesicles priorto their fusion with the plasma membrane. This hypothesis haspreviously been suggested based on experiments with culturedL6 muscle cells (31, 37). In order to test whether GLUT4-containing vesicles are recruited to the regions of membraneruffling upon insulin stimulation, we monitored the movementof Myc-GLUT4-CFP in cells also expressing YFP-Myo1c byusing ultrafast microscopy. Three-dimensional images of thesecells were acquired every 10 s for 1,000 continuous seconds,and each set of three-dimensional images consisted of 21 op-tical sections spaced 250 nm apart, thereby accounting for theentire volume of the cell. Figure 6 and the accompanyingmovie in Fig. SA2 in the supplemental material show twodifferentiated 3T3-L1 adipocytes expressing both Myc-GLUT4-CFP and YFP-Myo1c 10 min after insulin stimulation.
Figure 6 (top panel) shows the three-dimensional projectionsencompassing the 21 optical sections of these cells followingimage restoration to remove out-of-focus light at one giventime point in the movie. The average Myc-CFP-GLUT4 fluo-rescence intensity within regions of the cell adjacent to mem-brane ruffles was then computed. This computation was doneby first visually identifying regions in these cells which wereruffling and neighboring nonruffling regions in the same cells.The average Myc-GLUT4-CFP intensities in the three-dimen-sional volume near the ruffling and nonruffling areas of thesame cell at each time point were then measured over theentire length of the movie (1,000 s). The mean and standarddeviation of this Myc-GLUT4-CFP fluorescence intensity ineach of these three-dimensional regions of the cell were thenplotted.
Based on the data presented in Fig. 6 and in Fig. SA2 in thesupplemental material, we can draw two conclusions. First,Myo1c was consistently enriched in regions of active mem-brane ruffles compared to Myo1c in neighboring nonruffling
FIG. 5. Myo1c expression in 3T3-L1 adipocytes causes insulin-independent membrane ruffles. Differentiated 3T3-L1 adipocytes expressingYFP-Myo1c were serum starved. BODIPY 581/591 was added to the media at a final concentration of 1 �M, and the cells were incubated for 20min at 37°C. Membrane ruffling was observed by monitoring the cells live for 10 min, imaging them at 5-s intervals. Shown are six frames, each25 s apart, for both BODIPY 581/591 stain and YFP-Myo1c. The images are all single optical sections from three-dimensional images followingimage restoration.
5456 BOSE ET AL. MOL. CELL. BIOL.
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

domains. Second, Myc-GLUT4-CFP vesicles were also pre-dominantly recruited into these cytoplasmic regions of activemembrane reorganizations. As is evident from Fig. 6, the re-gions of the cell near membrane ruffles were highly enriched inboth Myc-GLUT4-CFP and YFP-Myo1c (also see the accom-panying movies Fig. SA2 in the supplemental material). Thisaggregation of Myc-GLUT4-CFP vesicles near membrane ruf-fles was not observed in the absence of insulin, consistent withthe hypothesis that these ruffles are enriched in exocytic Myc-GLUT4-CFP vesicles. As shown in Fig. 5, YFP-Myo1c was alsopredominantly concentrated in these ruffles. These data areconsistent with the hypothesis that insulin stimulation of cul-tured adipocytes results in the recruitment of exocytic GLUT4-containing vesicles to regions of Myo1c-driven membrane ruf-fling, where they undergo fusion.
Like other regulated exocytic pathways, insulin-stimulatedGLUT4 vesicle trafficking involves budding from an internalstorage compartment and translocation to the cell periphery,followed by fusion. Insulin-stimulated GLUT4 translocation tothe cell surface probably involves microtubules aided by kine-sin motors (14, 33). Results presented here confirm that thecytoskeleton is necessary for the translocation and that PI3-kinase activity is not needed for this process. Our presentand previous results indicate that, once GLUT4 reaches thecell periphery, the GLUT4-containing vesicles associate withactin filaments. A novel finding presented here is the require-ment of Myo1c in this step, which apparently involves GLUT4mobilization and anchoring to the cell cortex (Fig. 2). Al-though direct evidence is lacking, it is possible that the Myo1ccargo domain binds to a putative receptor on the GLUT4-containing vesicles, thereby tethering the vesicle to actin fila-ments. This process is apparently followed by the fusion ofthese vesicles with the plasma membrane, which, as we haveshown, requires PI 3-kinase activity (Fig. 1). Importantly, theexpression of Myo1c can partially overcome the block in fusionin the absence of PI 3-kinase activity (Fig. 3). Coupled with thepreviously reported observation that the expression of Myo1cpotentiates GLUT4 translocation and fusion, these data sug-gest that Myo1c may be involved in the fusion process.
The hypothesis that Myo1c may directly participate in themechanism of membrane fusion is further highlighted by thedramatic membrane ruffling induced by Myo1c in culturedadipocytes (Fig. 5). Insulin treatment of these cells results inthe recruitment of exocytic GLUT4-containing vesicles tothese sites of membrane ruffles (Fig. 6). A recent report sug-gests that exocytic vesicles are directed to membrane rufflesprior to fusion with the plasma membrane (4). Our data thussuggest that, in cultured adipocytes, Myo1c is responsible forlocalized membrane remodeling, which facilitates the fusion ofGLUT4-containing vesicles with the plasma membrane in thepresence of insulin. Future experiments are needed to directlytest this hypothesis.
ACKNOWLEDGMENTS
This work was supported by National Institutes of Health grantsDK063023 to M.P.C. and DK58133 to S.C.
REFERENCES
1. Bennett, M. K., J. E. Garcia-Arraras, L. A. Elferink, K. Peterson, A. M.Fleming, C. D. Hazuka, and R. H. Scheller. 1993. The syntaxin family ofvesicular transport receptors. Cell 74:863–873.
2. Borisy, G. G., and T. M. Svitkina. 2000. Actin machinery: pushing theenvelope. Curr. Opin. Cell Biol. 12:104–112.
3. Bose, A., A. Guilherme, S. I. Robida, S. M. C. Nicoloro, Q. L. Zhou, Z. Y.Jiang, D. P. Pomerleau, and M. P. Czech. 2002. Glucose transporter recy-cling in response to insulin is facilitated by myosin Myo1c. Nature 420:821–824.
4. Bretscher, M. S., and C. Aguado-Velasco. 1998. EGF induces recyclingmembrane to form ruffles. Curr. Biol. 8:721–724.
5. Broadie, K., A. Prokop, H. J. Bellen, C. J. O’Kane, K. L. Schulze, and S. T.Sweeney. 1995. Syntaxin and synaptobrevin function downstream of vesicledocking in Drosophila. Neuron 15:663–673.
6. Bryant, N. J., R. Govers, and D. E. James. 2002. Regulated transport of theglucose transporter GLUT4. Nat. Rev. Mol. Cell Biol. 3:267–277.
7. Cain, C. C., W. S. Trimble, and G. E. Lienhard. 1992. Members of theVAMP family of synaptic vesicle proteins are components of glucose trans-porter-containing vesicles from rat adipocytes. J. Biol. Chem. 267:11681–11684.
8. Carrington, W. A., R. M. Lynch, E. D. Moore, G. Isenberg, K. E. Fogarty,and F. S. Fay. 1995. Superresolution three-dimensional images of fluores-cence in cells with minimal light exposure. Science 268:1483–1487.
FIG. 6. Ultrafast microscopy reveals enrichment of GLUT4 andMyo1c in ruffling areas compared to nonruffling areas. (Top) Three-dimensional projection images of two different cells, showing Myc-GLUT4-CFP distribution. The images shown were selected from asequence of 100 such three-dimensional images of each cell spanning16.7 min (see Materials and Methods). The three-dimensional imageswere first subjected to image restoration to remove out-of-focus light,and then maximum-intensity projections were made by retaining themaximum (brightest) intensity at each pixel position of the 21 opticalsections. Regions of active membrane ruffling (red boxes) were iden-tified visually and compared with regions not showing significant ruf-fling (white boxes). See the supplemental material for complete moviesequences. (Bottom) In order to separate cytosolic from membrane-associated Myc-GLUT4-CFP fluorescence, an intensity threshold waschosen for each cell such that 95% of the Myc-GLUT4-CFP pixels inthe cytosol were below the threshold. Then, for each region of eachcell, the average Myc-GLUT4-CFP fluorescence of just the pixelswhose intensities were above the threshold and within the indicatedregion was computed for each time point. The mean and standarddeviation of the average Myc-GLUT4-CFP concentration for all 100time points were calculated and plotted as shown.
VOL. 24, 2004 Myo1c AND GLUT4 5457
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

9. Deitcher, D. L., A. Ueda, B. A. Stewart, R. W. Burgess, Y. Kidokoro, and T. L.Schwarz. 1998. Distinct requirements for evoked and spontaneous release ofneurotransmitter are revealed by mutations in the Drosophila gene neuro-nal-synaptobrevin. J. Neurosci. 18:2028–2039.
10. Dong, L. Q., L. R. Landa, M. J. Wick, L. Zhu, H. Mukai, Y. Ono, and F. Liu.2000. Phosphorylation of protein kinase N by phosphoinositide-dependentprotein kinase-1 mediates insulin signals to the actin cytoskeleton. Proc.Natl. Acad. Sci. USA 97:5089–5094.
11. Evangelista, M., B. M. Klebl, A. H. Tong, B. A. Webb, T. Leeuw, E. Leberer,M. Whiteway, D. Y. Thomas, and C. Boone. 2000. A role for myosin-I in actinassembly through interactions with Vrp1p, Bee1p, and the Arp2/3 complex.J. Cell Biol. 148:353–362.
12. Faty, M., M. Fink, and Y. Barral. 2002. Septins: a ring to part mother anddaughter. Curr. Genet. 41:123–131.
13. Gerst, J. E. 1999. SNAREs and SNARE regulators in membrane fusion andexocytosis. Cell. Mol. Life Sci. 55:707–734.
14. Imamura, T., J. Huang, I. Usui, H. Satoh, J. Bever, and J. M. Olefsky. 2003.Insulin-induced GLUT4 translocation involves protein kinase C-lambda-mediated functional coupling between Rab4 and the motor protein kinesin.Mol. Cell. Biol. 23:4892–4900.
15. Jahn, R., and T. C. Sudhof. 1999. Membrane fusion and exocytosis. Annu.Rev. Biochem. 68:863–911.
16. Jahn, R., T. Lang, and T. C. Sudhof. 2003. Membrane fusion. Cell 112:519–533.
17. Jiang, Z. Y., A. Chawla, A. Bose, M. Way, and M. P. Czech. 2002. A phos-phatidylinositol 3-kinase-independent insulin signaling pathway to N-WASP/Arp2/3/F-actin required for GLUT4 glucose transporter recycling. J. Biol.Chem. 277:509–515.
18. Kanzaki, M., and J. E. Pessin. 2001. Insulin-stimulated GLUT4 transloca-tion in adipocytes is dependent upon cortical actin remodeling. J. Biol.Chem. 276:42436–42444.
19. Karylowski, O., A. Zeigerer, A. Cohen, and T. E. McGraw. 2004. GLUT4 isretained by an intracellular cycle of vesicle formation and fusion with endo-somes. Mol. Biol. Cell 15:870–882.
20. Kotani, K., K. Hara, K. Kotani, K. Yonezawa, and M. Kasuga. 1995. Phos-phoinositide 3-kinase as an upstream regulator of the small GTP-bindingprotein Rac in the insulin signaling of membrane ruffling. Biochem. Biophys.Res. Commun. 208:985–990.
21. Lampson, M. A., A. Racz, S. W. Cushman, and T. E. McGraw. 2000. Dem-onstration of insulin-responsive trafficking of GLUT4 and vpTR in fibro-blasts. J. Cell Sci. 113:4065–4076.
22. Martin, S. S., T. Haruta, A. J. Morris, A. Klippel, L. T. Williams, and J. M.Olefsky. 1996. Activated phosphatidylinositol 3-kinase is sufficient to medi-ate actin rearrangement and GLUT4 translocation in 3T3-L1 adipocytes.J. Biol. Chem. 271:17605–17608.
23. Mulvihill, D. P., P. J. Pollard, T. Z. Win, and J. S. Hyams. 2001. MyosinV-mediated vacuole distribution and fusion in fission yeast. Curr. Biol. 11:1124–1127.
24. Okada, T., Y. Kawano, T. Sakakibara, O. Hazeki, and M. Ui. 1994. Essentialrole of phosphatidylinositol 3-kinase in insulin-induced glucose transportand antilipolysis in rat adipocytes. Studies with a selective inhibitor wort-mannin. J. Biol. Chem. 269:3568–3573.
25. Patel, N., A. Rudich, Z. A. Khayat, R. Garg, and A. Klip. 2003. Intracellularsegregation of phosphatidylinositol-3,4,5-trisphosphate by insulin-dependentactin remodeling in L6 skeletal muscle cells. Mol. Cell. Biol. 23:4611–4626.
26. Patki, V., J. Buxton, A. Chawla, L. Lifshitz, K. Fogarty, W. Carrington, R.Tuft, and S. Corvera. 2001. Insulin action on GLUT4 traffic visualized insingle 3T3-l1 adipocytes by using ultra-fast microscopy. Mol. Biol. Cell 12:129–141.
27. Pfeffer, S. 2001. Vesicle tethering factors united. Mol. Cell 8:729–730.28. Pfeffer, S. R. 1999. Transport-vesicle targeting: tethers before SNAREs. Nat.
Cell Biol. 1:E17–E22.29. Reizes, O., B. Barylko, C. Li, T. C. Sudhof, and J. P. Albanesi. 1994. Domain
structure of a mammalian myosin I beta. Proc. Natl. Acad. Sci. USA 91:6349–6353.
30. Rizzuto, R., W. Carrington, and R. A. Tuft. 1998. Digital imaging microscopyof living cells. Trends Cell Biol. 8:288–292.
31. Rudich, A., and A. Klip. 2003. Push/pull mechanisms of GLUT4 traffic inmuscle cells. Acta Physiol. Scand. 178:297–308.
32. Saitoe, M., T. L. Schwarz, J. A. Umbach, C. B. Gundersen, and Y. Kidokoro.2001. Absence of junctional glutamate receptor clusters in Drosophila mu-tants lacking spontaneous transmitter release. Science 293:514–517.
33. Semiz, S., J. G. Park, S. M. C. Nicoloro, P. Furcinitti, C. Zhang, A. Chawla,J. Leszyk, and M. P. Czech. 2003. Conventional kinesin KIF5B mediatesinsulin-stimulated GLUT4 movements on microtubules. EMBO J. 22:2387–2399.
34. Simpson, F., J. P. Whitehead, and D. E. James. 2001. GLUT4—at the crossroads between membrane trafficking and signal transduction. Traffic 2:2–11.
35. Sollner, T. H. 2003. Regulated exocytosis and SNARE function. Mol.Membr. Biol. 20:209–220.
36. Sollner, T. H., and J. E. Rothman. 1996. Molecular machinery mediatingvesicle budding, docking and fusion. Experientia 52:1021–1025.
37. Tong, P., Z. A. Khayat, C. Huang, N. Patel, A. Ueyama, and A. Klip. 2001.Insulin-induced cortical actin remodeling promotes GLUT4 insertion atmuscle cell membrane ruffles. J. Clin. Investig. 108:371–381.
38. Toonen, R. F., and M. Verhage. 2003. Vesicle trafficking: pleasure and painfrom SM genes. Trends Cell Biol. 13:177–186.
39. Vale, R. D. 2003. The molecular motor toolbox for intracellular transport.Cell 112:467–480.
40. Wang, Y., S. Sugita, and T. C. Sudhof. 2000. The RIM/NIM family ofneuronal C2 domain proteins. Interactions with Rab3 and a new class of Srchomology 3 domain proteins. J. Biol. Chem. 275:20033–20044.
41. Yang, C., K. J. Coker, J. K. Kim, S. Mora, D. C. Thurmond, A. C. Davis, B.Yang, R. A. Williamson, G. I. Shulman, and J. E. Pessin. 2001. Syntaxin 4heterozygous knockout mice develop muscle insulin resistance. J. Clin. In-vestig. 107:1311–1318.
42. Yoshihara, M., A. Ueda, D. Zhang, D. L. Deitcher, T. L. Schwarz, and Y.Kidokoro. 1999. Selective effects of neuronal-synaptobrevin mutations ontransmitter release evoked by sustained versus transient Ca2� increases andby cAMP. J. Neurosci. 19:2432–2441.
43. Young, A., R. Tuft, W. Carrington, and S. J. Doxsey. 1999. Centrosomedynamics in living cells. Methods Cell Biol. 58:223–238.
5458 BOSE ET AL. MOL. CELL. BIOL.
on July 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from
Copyright © 2022 FDOKUMEN




















