
Inhibition of myeloperoxidase activity by the alkaloids of Peganum harmala L. (Zygophyllaceae)
Upload
independentCategory
view
4download
0
Journal of Photochemistry and Photobiology B: Biology 142 (2015) 250–256
Contents lists available at ScienceDirect
Journal of Photochemistry and Photobiology B: Biology
journal homepage: www.elsevier .com/locate / jphotobiol
tRNA binding with anti-cancer alkaloids–nature of interactionand comparison with DNA–alkaloids adducts
http://dx.doi.org/10.1016/j.jphotobiol.2014.12.0091011-1344/� 2014 Elsevier B.V. All rights reserved.
⇑ Corresponding author at: Quantum Phenomenon and Applications,CSIR-National Physical Laboratory, Dr. K.S. Krishnan Marg, New Delhi 110012,India. Tel.: +91 11 45608366; fax: +91 11 45609310.
E-mail address: [email protected] (R. Mehrotra).
Gunjan Tyagi, Shweta Agarwal, Ranjana Mehrotra ⇑Quantum Optics and Photon Physics, CSIR-National Physical Laboratory, Dr. K.S. Krishnan Marg, New Delhi 110 012, India
a r t i c l e i n f o
Article history:Received 24 October 2014Received in revised form 1 December 2014Accepted 6 December 2014Available online 27 December 2014
a b s t r a c t
Vincristine and vinblastine are potent anti-proliferative compound whose mechanism of action inside acell is not well elucidated and the basis of their differential cellular effect is also unknown. This workfocuses towards understanding the interaction of vincristine and vinblastine with tRNA using spectro-scopic approach. Fourier transform infrared (FTIR) spectroscopy, Fourier transform infrared differencespectroscopy and UV–visible spectroscopy were used to study the binding parameters of tRNA–alkaloidsinteraction. Both the vinca alkaloids interact with tRNA through external binding with some degree ofintercalation into the nitrogenous bases. The alkaloids adduct formation did not alter the A-conformationof the biopolymer and vincristine–tRNA complexes were found to be more stable than that of vinblas-tine–tRNA complexes. The binding constants (K) estimated for VCR–tRNA and VBS–tRNA complexationare 3 � 102 M�1 and 2.5 � 102 M�1 respectively, which suggests low affinity of alkaloids to tRNA. Thestudy recognizes tRNA binding properties of vital vinca alkaloids and contributes to a better understand-ing of their mechanism of action and could also help in identifying the reason behind their diverse actionin a cell.
� 2014 Elsevier B.V. All rights reserved.
1. Introduction
Along with DNA, RNA is widely studied these days as an impor-tant therapeutic target for various molecules [1–8]. It executes asignificant array of functions, which are indispensable for the sur-vival of a cell. RNA is involved in processes like serving as a tem-plate and as an activated intermediate in protein synthesis alongwith being a ribosome component [9–12]. RNA also participatesin phenomenon like gene expression by catalyzing the maturationof mRNAs through ribozymes [13–17] and chromosome-end main-tenance [18]. Disruption of any of these functions can have seriousconsequences on a cell. Several RNA pathways are also linked toapoptosis [19–22] and it is found that the genome of various retro-viruses, which falls into the category of oncovirus, contains onlyRNA [23,24]. The versatility of RNA is derived from its capacity toform profuse three-dimensional structures [25,26]. RNA has vari-ous characterized secondary and tertiary structures [27]. The fold-ing pattern of RNA chain forms the primary element for itschemical reactivity and specific interaction with ligands. All thesediscoveries have led to the expansion of RNA biology and highlightthe potential of targeting RNA for the treatment of myriad of
diseases [28,29]. Further, as RNA lacks repair mechanism andhas structural domains that are more highly conserved, smallmolecules that target RNA are likely to encounter slower drugresistance. These factors make RNA an interesting target, to beexplored, for anti-cancer compounds. A way to screen thesuccessful RNA binding compounds is to study the interaction ofthose compounds with RNA, which has DNA binding properties.There exist some interesting reports, which shows that certainDNA binding molecules exhibit a good amount of RNA affinity[30]. Our interest here is to study the interaction of tRNA with nat-ural anti-cancer compounds, the two vinca alkaloids, vincristine(VCR) and vinblastine (VBS).
Natural compounds with their chemical diversity and biologicalsignificance have been accepted as potential candidates for thetreatment of various diseases including cancer. Plants containingalkaloids have been recognized as medicinal and their compoundsare in use for centuries as folk medicine without the clear under-standing of their biological targets. VCR and VBS are known anti-cancer compounds whose complete mechanism of action has notbeen deciphered yet. The vinca alkaloids, VCR and VBS are includedin numerous chemotherapy protocols yet the molecular origins oftheir differential antitumor and toxic side effects remain uncertain.Although, they differ in their molecular structure by only onecarbonyl group, VCR and VBS manifest quite different antitumoractivity (Fig. 1). It is known that they modulate tubulin in the cellas primary target, but to what extent the cell-killing and antitumor

G. Tyagi et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 250–256 251
properties of the Catharanthus alkaloids are due to their interac-tions with tubulin and microtubule is still not clear. In a numberof cases, the order of efficacy of the drugs against tumors doesnot correlate with reported binding strengths to tubulin [31]. Thevinca alkaloids can affect a number of different cellular systemsincluding RNA and DNA synthesis, lipid biosynthesis [32–34], cyc-lic nucleotide metabolism, glutathione metabolism [35] and cal-modulin-dependent Ca2+ transport ATPase [36].Interaction ofalkaloids with RNA is less understood then with DNA. Thus, spe-cific experimental studies of alkaloid–RNA interactions arerequired. We sought to characterize the interaction of these twonaturally occurring vinca alkaloids with tRNA using Fourier trans-form infrared (FTIR) and UV–visible absorption spectroscopy.These techniques are used to investigate the interaction of VCRand VBS with tRNA in aqueous solution at physiological pH. Esti-mation of the binding constant and mode of binding of alkaloidswith tRNA has been conducted along with the evaluation of theconformation state of nucleic acid after the formation of alka-loid–tRNA complexes.
2. Materials and methods
2.1. Materials
RNA (tRNA) from Baker’s yeast was purchased from Sigma–Aldrich chemicals (USA). Vincristine and vinblastine were procuredfrom Sigma–Aldrich chemicals (USA). Purity of RNA was estimatedby recording the UV absorbance at 260 and 280 nm and calculatingratio, A260/A280. The ratio was found to be more than 1.9, indicatingthat the RNA is free from protein. Deionized water from the Milli-pore water purification system was used in the preparation ofdesired aqueous solutions. Other chemicals and reagents used inthe study were of analytical grade and used as supplied.
2.2. Preparation of stock solutions
Solution of RNA sodium salt was prepared in 10 mM Tris–HClbuffer (pH 7.4) and kept at 8 �C for 24 h. The solution was stirredat frequent intervals to ensure its homogeneity. Concentration ofRNA stock solutions were measured spectrophotometrically usingexcitation coefficient of 9250 cm�1 M�1 respectively. The final con-centration of RNA stock solutions was 25 mM. VCR or VBS/RNAratios (r) were kept 1/100, 1/80, 1/40, 1/20 and 1/10 for allmeasurements.
2.3. FTIR spectral measurements
FTIR spectra were recorded with Varian 660-IR spectrophotom-eter, equipped with DTGS (deuterated triglycine sulphate) detectorand KBr beam splitter. All spectra were recorded in 10 mM
Fig. 1. Chemical structure of (a) v
Tris–HCl buffer as a solvent (pH of 7.4). Liquid samples were ana-lyzed in attenuated total reflectance mode with ZnSe crystal.Ambient humidity of 45% RH was maintained during the experi-ment. 256 scans were recorded for each sample in the spectralrange of 4000–650 cm�1 with a resolution of 2 cm�1. Backgroundspectra were collected before each measurement. A spectrum ofbuffer solution was recorded and subtracted from the spectrumof RNA and ligand–RNA complexes. A satisfactory buffer subtrac-tion was considered to be achieved, when the intensity of watercombination band at about 2200 cm�1 became zero in all the spec-tra recorded [37–40]. FTIR difference spectra were produced bysubtracting the spectrum of free RNA from the spectrum ofligand–RNA complex, ((RNA solution + ligand solution) � (RNAsolution)).
2.4. UV–visible spectral measurements
The UV visible spectra were recorded on Perkin Elmer spectro-photometer, Lambda 35. Quartz cuvette of 1 cm path length wereused for measurement. Spectra were recorded for free RNA andfor various ligand–RNA complex solutions. For the calculation ofthe binding constant of the reaction occurring between ligandand RNA the method described by Kanakis et al. is used [41]. It ispresumed that after the interaction between ligand [L], which isvincristine and vinblastine and substrate [S] which is RNA in aque-ous solution, the complex [SL] forms [42].
It is also assumed that the substrate and the ligand follow beer’slaw for the absorbance of light.
The absorbance of substrate solution at its total concentrationwith a path length [l] of 1 cm is
A0 ¼ eSlSt ð1Þ
es is the molar absorptivity of DNA.The absorbance of solution consisting of total concentration of
substrate along with total concentration of ligand is
AL ¼ eSl½S� þ eLl½L� þ eSLl½SL� ð2Þ
[S] is the concentration of uncomplexed substrate.[L] is the concentration of uncomplexed ligand.[SL] is the concentration of substrate–ligand complex.eL is the molar absorptivity of ligand.
After combining with the mass balance of substrate and ligand,the absorbance equation can be written as
AL ¼ eSlSt þ eLlLt þ DeSLl½SL� ð3Þ
DeSL ¼ eSL � eS � eL
The absorbance of solution measured against the total concen-tration of ligand as reference is
incristine and (b) vinblastine.

252 G. Tyagi et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 250–256
A ¼ eSlSt þ DeSLl½SL� ð4Þ
The stability constant for the formation of complex [SL] can begiven as
KSL ¼ ½SL� ½S�½L�= ð5Þ
Combining Eqs. (4) and (5)
DA ¼ KSL D eSLl½S�½L� ð6Þ
DA ¼ A� A0
From the mass balance equation St = [S] + [SL] we get[S] = St/[1 + KSL[L]], which gives Eq. (7)
DAl¼ StKSLeSL½L�
1þ KSL½L�ð7Þ
This shows the hyperbolic dependence of binding on free ligandconcentration. Linear transformation of Eq. (6) is done by takingthe reciprocal of each side of Eq. (7).
lDA¼ 1
StKSLDeSL½L�þ 1
StDeSLð8Þ
The double reciprocal plot of 1/DA versus 1/[L] is linear and thebinding constant [K] can be estimated by calculating the ratio ofthe intercept to the slope.
3. Results and discussion
3.1. FTIR spectroscopic analysis
3.1.1. Vincristine–tRNA complexInfrared marker bands in the region 1800–700 cm�1 provide
information about various portions of tRNA structure. Carefulobservation of the variations taking place in these bands afterbinding of any ligand gives an important insight into the changesoccurring in the structure of tRNA due to ligand interaction. Theinfrared spectral features for vincristine–tRNA interaction are pre-sented in Fig. 2. The band at 1700 cm�1 in the free tRNA spectrum,related to guanine and uracil bases gained intensity and shifted tohigher frequency at 1708 cm�1 upon VCR binding. The uracil bandat 1649 cm�1 is shifted to 1642 cm�1 while cytosine band posi-tioned at 1490 cm�1 experiences an increased shift and is observedat 1496 cm�1 in VCR–tRNA complex spectrum (r = 1/10). No majorspectral shifting or intensity variation is observed for adenine bandat 1601 cm�1 upon drug interaction. Further, second order deriva-tive spectra clarify the shift and intensity variation of vital infraredbands (Fig. 3). The backbone phosphate (PO2) asymmetric stretch-ing band at 1237 cm�1 presented a small shift of 2 cm�1 with anincrease in its intensity when high concentration of VCR is com-plexed with tRNA. The intensity increase of base and phosphatebands is evident from the difference spectra of various VCR–tRNAcomplexes (Fig. 4). The symmetric PO2 band did not show anychange in its position after VCR binding. The results indicate thatthe formation of VCR–tRNA complex did not alter the conformationof biopolymer as the characteristic marker bands of A-form at 810and 866 cm�1 presented no major alterations in the complex infra-red spectra. The major spectral changes observed for the band at1700 cm�1, 1490 cm�1 and 1649 cm�1 are indicative of VCR bind-ing to guanine, cytosine and uracil reactive sites of tRNA along withsome degree of interaction with the phosphate backbone. Thesespectroscopic findings suggest that the large aromatic structureof VCR interact mainly to guanine and cytosine bases of tRNA.Binding with uracil can be attributed to the local helix meltingafter the reaction with G–C base pair that might have providedadditional binding sites for the VCR–biopolymer interaction. Alltogether, the infrared spectral features suggest external base
binding of fused aromatic ringed structure of ligand to the RNApolymer.
3.1.2. Vinblastine–tRNA complexObservation of prominent infrared spectral features of free
tRNA and VBS–tRNA complexes indicate towards external bindingof ligand via interaction through nitrogenous bases and phosphatebackbone. Evidence for this comes from the shifting of infraredband assigned to guanine (1700–1704 cm�1), cytosine (1490–1493 cm�1) and uracil (1649–1651 cm�1) bases (Figs. 5 and 6).Increase in the intensity of prominent bases is also observed upondrug binding, which can be manifested from the positive featuresin the difference spectra (Fig. 7). Band assigned to asymmetricPO2 underwent a shift of 2 cm�1 from 1237 cm�1 to 1239 cm�1,while symmetric PO2 band did not exhibit any appreciable changeupon drug interaction. Bands at frequency position of 810 cm�1
and 866 cm�1, representative of RNA A-conformation, also didnot experience any shift after VBS complex formation with tRNA.Stable positions of these bands suggest that in the process ofVBS–tRNA binding, tRNA has retained its A-conformation. Minorligand PO2 interaction has also occurred similar to VCR–tRNA com-plexes. These features are typical to the external mode of bindingseen in ligand–biomolecule interactions.
3.2. UV–visible absorption spectroscopic analysis
3.2.1. Vincristine–tRNA complexIn UV–visible absorption characterization of RNA–drug binding
the band at 260 nm arises due to p–p⁄ transition of ringed nitrog-enous bases. Changes in absorbance and wavelength shift of260 nm band reflect the structural changes occurring in RNA struc-ture upon ligand complexation. In the present scenario addition ofincreasing concentration of VCR to a constant concentration oftRNA causes increase in its absorption maxima (Fig. 8a). Thishyperchromic effect is attributed to the stacking interaction takingplace between the aromatic chromophores of VCR and pairednitrogenous bases of tRNA resulting in localized distortion. Thislocalized distortion is responsible for the augmented exposure ofnitrogenous bases to incident light resulting in increased absorp-tion [43]. Intercalation of VCR into the structure of RNA can be fur-ther confirmed by careful observation of the absorption spectra ofvincristine in the absence and presence of tRNA (Fig. 8b). Absor-bance maxima of VCR at 220 nm, shifts to 205 nm along with adecrease in its absorption intensity upon VCR–tRNA complex for-mation. The reduced absorbance of vincristine after binding withRNA (hypochromic shift) can be credited to the hindrance causedby the structure of RNA to the chromophores of VCR. The restrainton molecular movements of alkaloid causes a decrease in its capac-ity to absorb light energy. A small bathochromic shift observed inthe UV–visible spectra of VCR–RNA complex can be ascribed to thehydrophobic envelope provided by RNA duplex to VCR after inter-calation that might affect the p–p⁄ transitions of VCR aromaticchromophores. Furthermore, spectral alterations of UV–visiblebands (in Fig. 8b) attribute localized denaturation in tRNA biomol-ecule that results in enhancement of light absorption.
Stability: The stability of VCR–tRNA complexes were determinedby the calculation of binding constant (K). The calculation wasbased on the absorbance values of free tRNA and various VCR–tRNA complexes as a function of the concentration of VCR. Thebinding constant estimated for VCR–tRNA complex is3 � 102 M�1, which suggests low affinity of alkaloids to tRNA [44].
3.2.2. Vinblastine–tRNA complexFig. 9a illustrates the changes occurring in the absorption
profile of free tRNA when it was complexed with VBS. Increasingconcentration of vinblastine exerted an augmented effect on the
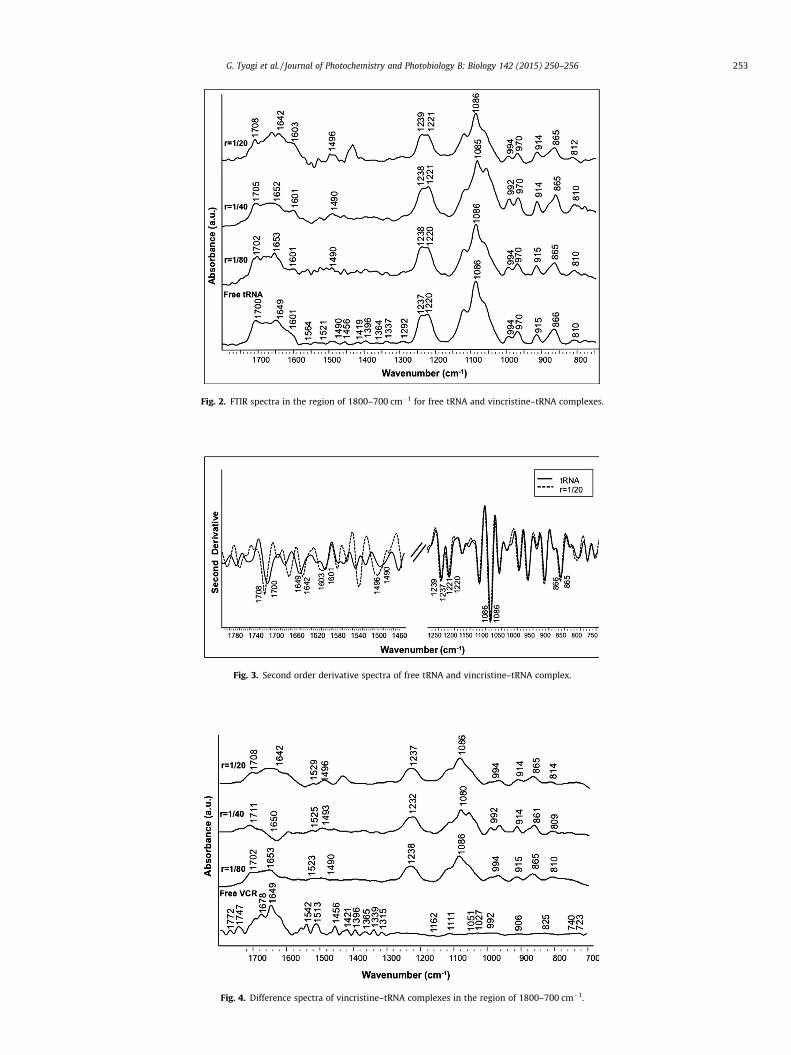
Fig. 2. FTIR spectra in the region of 1800–700 cm�1 for free tRNA and vincristine–tRNA complexes.
Fig. 3. Second order derivative spectra of free tRNA and vincristine–tRNA complex.
Fig. 4. Difference spectra of vincristine–tRNA complexes in the region of 1800–700 cm�1.
G. Tyagi et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 250–256 253
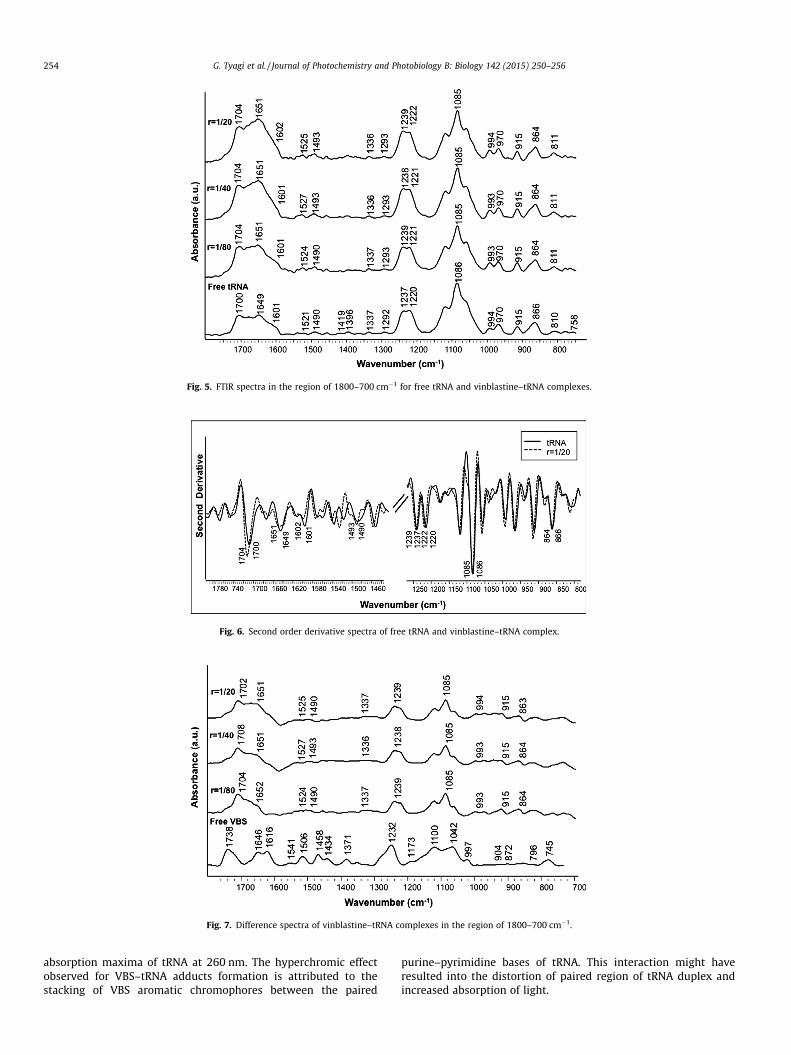
Fig. 5. FTIR spectra in the region of 1800–700 cm�1 for free tRNA and vinblastine–tRNA complexes.
Fig. 6. Second order derivative spectra of free tRNA and vinblastine–tRNA complex.
Fig. 7. Difference spectra of vinblastine–tRNA complexes in the region of 1800–700 cm�1.
254 G. Tyagi et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 250–256
absorption maxima of tRNA at 260 nm. The hyperchromic effectobserved for VBS–tRNA adducts formation is attributed to thestacking of VBS aromatic chromophores between the paired
purine–pyrimidine bases of tRNA. This interaction might haveresulted into the distortion of paired region of tRNA duplex andincreased absorption of light.
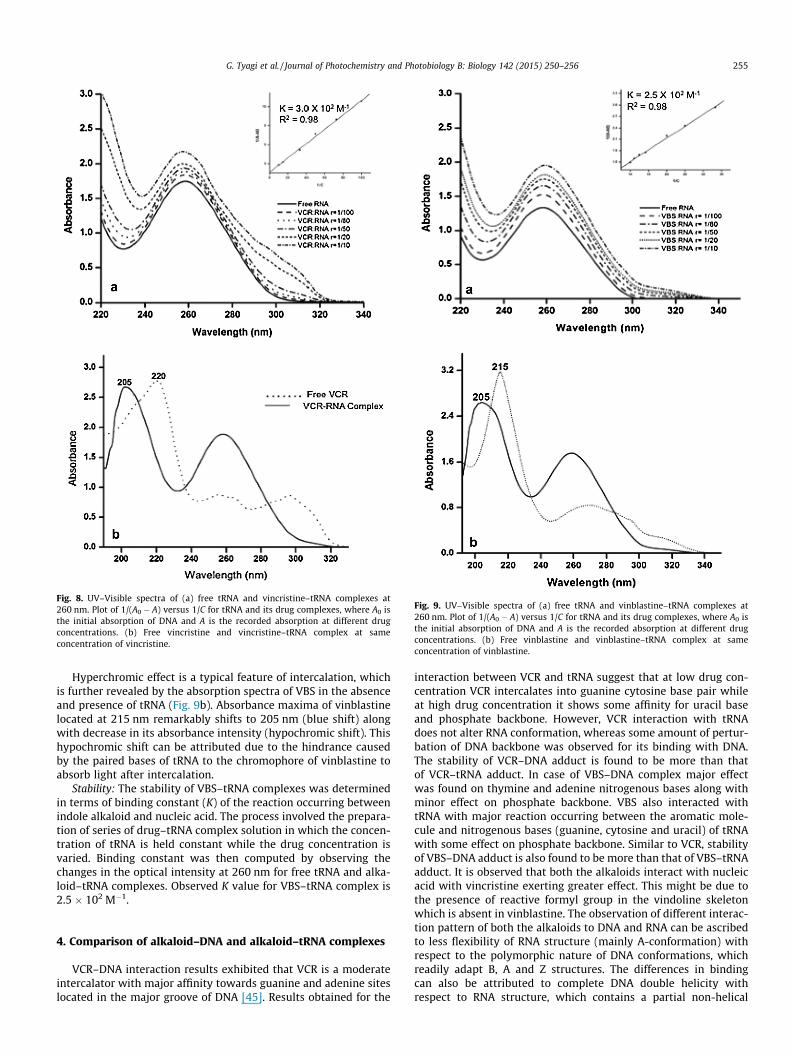
Fig. 8. UV–Visible spectra of (a) free tRNA and vincristine–tRNA complexes at260 nm. Plot of 1/(A0 � A) versus 1/C for tRNA and its drug complexes, where A0 isthe initial absorption of DNA and A is the recorded absorption at different drugconcentrations. (b) Free vincristine and vincristine–tRNA complex at sameconcentration of vincristine.
Fig. 9. UV–Visible spectra of (a) free tRNA and vinblastine–tRNA complexes at260 nm. Plot of 1/(A0 � A) versus 1/C for tRNA and its drug complexes, where A0 isthe initial absorption of DNA and A is the recorded absorption at different drugconcentrations. (b) Free vinblastine and vinblastine–tRNA complex at sameconcentration of vinblastine.
G. Tyagi et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 250–256 255
Hyperchromic effect is a typical feature of intercalation, whichis further revealed by the absorption spectra of VBS in the absenceand presence of tRNA (Fig. 9b). Absorbance maxima of vinblastinelocated at 215 nm remarkably shifts to 205 nm (blue shift) alongwith decrease in its absorbance intensity (hypochromic shift). Thishypochromic shift can be attributed due to the hindrance causedby the paired bases of tRNA to the chromophore of vinblastine toabsorb light after intercalation.
Stability: The stability of VBS–tRNA complexes was determinedin terms of binding constant (K) of the reaction occurring betweenindole alkaloid and nucleic acid. The process involved the prepara-tion of series of drug–tRNA complex solution in which the concen-tration of tRNA is held constant while the drug concentration isvaried. Binding constant was then computed by observing thechanges in the optical intensity at 260 nm for free tRNA and alka-loid–tRNA complexes. Observed K value for VBS–tRNA complex is2.5 � 102 M�1.
4. Comparison of alkaloid–DNA and alkaloid–tRNA complexes
VCR–DNA interaction results exhibited that VCR is a moderateintercalator with major affinity towards guanine and adenine siteslocated in the major groove of DNA [45]. Results obtained for the
interaction between VCR and tRNA suggest that at low drug con-centration VCR intercalates into guanine cytosine base pair whileat high drug concentration it shows some affinity for uracil baseand phosphate backbone. However, VCR interaction with tRNAdoes not alter RNA conformation, whereas some amount of pertur-bation of DNA backbone was observed for its binding with DNA.The stability of VCR–DNA adduct is found to be more than thatof VCR–tRNA adduct. In case of VBS–DNA complex major effectwas found on thymine and adenine nitrogenous bases along withminor effect on phosphate backbone. VBS also interacted withtRNA with major reaction occurring between the aromatic mole-cule and nitrogenous bases (guanine, cytosine and uracil) of tRNAwith some effect on phosphate backbone. Similar to VCR, stabilityof VBS–DNA adduct is also found to be more than that of VBS–tRNAadduct. It is observed that both the alkaloids interact with nucleicacid with vincristine exerting greater effect. This might be due tothe presence of reactive formyl group in the vindoline skeletonwhich is absent in vinblastine. The observation of different interac-tion pattern of both the alkaloids to DNA and RNA can be ascribedto less flexibility of RNA structure (mainly A-conformation) withrespect to the polymorphic nature of DNA conformations, whichreadily adapt B, A and Z structures. The differences in bindingcan also be attributed to complete DNA double helicity withrespect to RNA structure, which contains a partial non-helical

256 G. Tyagi et al. / Journal of Photochemistry and Photobiology B: Biology 142 (2015) 250–256
structure, and to minor differences in some of the H-bondingarrangements.
5. Conclusions
The study identifies RNA binding properties of vincristine andvinblastine and contributes to a better understanding of the mech-anism of action of indole alkaloids. Results provided here can notonly help in finding the basis behind diverse actions of indole alka-loids inside a cell, like their role in inhibition of DNA repair, butalso possibly aid in developing analogues for structure activityrelationship studies and to find potentially more active and lesstoxic derivatives. The study can be extended to chemically modifyand/or substitute methyl group of vinblastine in the vindoline moi-ety to some more reactive group. This may enhance the binding ofthe drug with DNA and hence might result in better effectiveness.
Acknowledgements
The authors thank Director, National Physical Laboratory forgranting the permission for publication of the work. G.T. is thankfulto Indian Council of Medical Research and S.A. is thankful toCouncil of Scientific and Industrial Research for providing financialsupport.
References
[1] T. Wang, L. Fan, Y. Watanabe, P. McNeill, G. Moulton, C. Bangur, G. Fanger, M.Okada, Y. Inoue, D. Persing, L523S, an RNA-binding protein as a potentialtherapeutic target for lung cancer, Br. J. Cancer 88 (2003) 887–894.
[2] T. Hermann, Rational ligand design for RNA: the role of static structure andconformational flexibility in target recognition, Biochimie 84 (2002) 869–875.
[3] T. Hermann, E. Westhof, Rational drug design and high-throughput techniquesfor RNA targets, Comb. Chem. High Throughput Screening 3 (2000) 219–234.
[4] E.E. Swayze, E.A. Jefferson, K.A. Sannes-Lowery, L.B. Blyn, L.M. Risen, S.Arakawa, S.A. Osgood, S.A. Hofstadler, R.H. Griffey, SAR by MS: a ligandbased technique for drug lead discovery against structured RNA targets, J. Med.Chem. 45 (2002) 3816–3819.
[5] J. Gallego, G. Varani, Targeting RNA with small-molecule drugs: therapeuticpromise and chemical challenges, Acc. Chem. Res. 34 (2001) 836–843.
[6] K.A. Xavier, P.S. Eder, T. Giordano, RNA as a drug target: methods forbiophysical characterization and screening, Trends Biotechnol. 18 (2000)349–356.
[7] W.D. Wilson, K. Li, Targeting RNA with small molecules, Curr. Med. Chem. 7(2000) 73–98.
[8] Y. Tor, Targeting RNA with small molecules, ChemBioChem 4 (2003) 998–1007.
[9] J.D. Watson, Involvement of RNA in the synthesis of proteins, Science 140(1963) 27.
[10] U.E. Loening, Molecular weights of ribosomal RNA in relation to evolution, J.Mol. Biol. 38 (1968) 355–365.
[11] R.P. Perry, The cellular sites of synthesis of ribosomal and 4S RNA, Proc. Natl.Acad. Sci. U.S.A. 48 (1962) 2179.
[12] R.P. Perry, D.E. Kelley, Messenger RNA–protein complexes and newlysynthesized ribosomal subunits: analysis of free particles and components ofpolyribosomes, J. Mol. Biol. 35 (1968) 37–59.
[13] J.R. Nevins, M.C. Wilson, Regulation of adenovirus-2 gene expression at thelevel of transcriptional termination and RNA processing, Nature 290 (1981)113–118.
[14] M. Grunberg-Manago, Messenger RNA stability and its role in control of geneexpression in bacteria and phages, Annu. Rev. Genet. 33 (1999) 193–227.
[15] L. He, G.J. Hannon, MicroRNAs: small RNAs with a big role in gene regulation,Nat. Rev. Genet. 5 (2004) 522–531.
[16] A. Serganov, D.J. Patel, Ribozymes, riboswitches and beyond: regulation ofgene expression without proteins, Nat. Rev. Genet. 8 (2007) 776–790.
[17] M.U. Kaikkonen, M.T. Lam, C.K. Glass, Non-coding RNAs as regulators of geneexpression and epigenetics, Cardiovasc. Res. 90 (2011) 430–440.
[18] J.L. Osterhage, K.L. Friedman, Chromosome end maintenance by telomerase, J.Biol. Chem. 284 (2009) 16061–16065.
[19] D. Ovcharenko, K. Kelnar, C. Johnson, N. Leng, D. Brown, Genome-scalemicroRNA and small interfering RNA screens identify small RNA modulators ofTRAIL-induced apoptosis pathway, Cancer Res. 67 (2007) 10782–10788.
[20] D. Khaitan, M.E. Dinger, J. Mazar, J. Crawford, M.A. Smith, J.S. Mattick, R.J.Perera, The melanoma-upregulated long noncoding RNA SPRY4-IT1 modulatesapoptosis and invasion, Cancer Res. 71 (2011) 3852–3862.
[21] R.T. Lima, S. Busacca, G.M. Almeida, G. Gaudino, D.A. Fennell, M.H. Vasconcelos,MicroRNA regulation of core apoptosis pathways in cancer, Eur. J. Cancer 47(2011) 163–174.
[22] S.D. Der, Y.-L. Yang, C. Weissmann, B.R. Williams, A double-stranded RNA-activated protein kinase-dependent pathway mediating stress-inducedapoptosis, Proc. Natl. Acad. Sci. 94 (1997) 3279–3283.
[23] P.H. Duesberg, K. Bister, P.K. Vogt, The RNA of avian acute leukemia virusMC29, Proc. Natl. Acad. Sci. 74 (1977) 4320–4324.
[24] W.S. Hayward, Size and genetic content of viral RNAs in avian oncovirus-infected cells, J. Virol. 24 (1977) 47–63.
[25] G. Varani, C. Cheong, I. Tinoco Jr, Structure of an unusually stable RNA hairpin,Biochemistry 30 (1991) 3280–3289.
[26] B.T. Wimberly, D.E. Brodersen, W.M. Clemons, R.J. Morgan-Warren, A.P. Carter,C. Vonrhein, T. Hartsch, V. Ramakrishnan, Structure of the 30S ribosomalsubunit, Nature 407 (2000) 327–339.
[27] H.M. Berman, W.K. Olson, D.L. Beveridge, J. Westbrook, A. Gelbin, T. Demeny,S.-H. Hsieh, A. Srinivasan, B. Schneider, The nucleic acid database. Acomprehensive relational database of three-dimensional structures ofnucleic acids, Biophys. J. 63 (1992) 751.
[28] B. Davis, M. Afshar, G. Varani, A.I. Murchie, J. Karn, G. Lentzen, M. Drysdale, J.Bower, A.J. Potter, I.D. Starkey, Rational design of inhibitors of HIV-1 TAR RNAthrough the stabilisation of electrostatic ‘‘hot spots’’, J. Mol. Biol. 336 (2004)343–356.
[29] J. Zhou, G. Wang, L.H. Zhang, X.S. Ye, Modifications of aminoglycosideantibiotics targeting RNA, Med. Res. Rev. 27 (2007) 279–316.
[30] F.A. Tanious, J.M. Veal, H. Buczak, L.S. Ratmeyer, W.D. Wilson, DAPI (40 ,6-diamidino-2-phenylindole) binds differently to DNA and RNA: minor-groovebinding at AT sites and intercalation at AU sites, Biochemistry 31 (1992) 3103–3112.
[31] K. Gerzon, Dimeric Catharanthus Alkaloids, Anticancer Agents Based onNatural Product Models, Academic Press, New York, 1980, pp. 271–317.
[32] V.A. Bernstam, R.H. Gray, I.A. Bernstein, Effect of microtubule-disrupting drugson protein and RNA synthesis in Physarum polycephalum amoebae, Arch.Microbiol. 128 (1980) 34–40.
[33] W.A. Creasey, Modifications in biochemical pathways produced by the Vincaalkaloids, Cancer Chemother. Rep. Part 1 52 (1968) 501.
[34] W. Creasey, Biochemical effects of vindesine, Br. J. Cancer 44 (1981) 921.[35] W.T. Beck, Increase by vinblastine of oxidized glutathione in cultured
mammalian cells, Biochem. Pharmacol. 29 (1980) 2333–2337.[36] K. Gietzen, A. Wüthrich, H. Bader, Effects of microtubular inhibitors on plasma
membrane calmodulin-dependent Ca2+-transport ATPase, Mol. Pharmacol. 22(1982) 413–420.
[37] S. Agarwal, D. Chadha, R. Mehrotra, Molecular modeling and spectroscopicstudies of semustine binding with DNA and its comparison with lomustine–DNA adduct formation, J. Biomol. Struct. Dyn. (2014) 1–16.
[38] S. Agarwal, D.K. Jangir, R. Mehrotra, Spectroscopic studies of the effects ofanticancer drug mitoxantrone interaction with calf-thymus DNA, J.Photochem. Photobiol., B 120 (2013) 177–182.
[39] S. Agarwal, D.K. Jangir, P. Singh, R. Mehrotra, Spectroscopic analysis of theinteraction of lomustine with calf thymus DNA, J. Photochem. Photobiol., B 130(2014) 281–286.
[40] S. Agarwal, D.K. Jangir, Ranjana Mehrotra, Neelam Lohani, M.R. Rajeswari, AStructural Insight into major groove directed binding of nitrosourea derivativenimustine with DNA: a spectroscopic study, PLoS ONE 9 (2014) e104115.
[41] C. Kanakis, P. Tarantilis, C. Pappas, J. Bariyanga, H. Tajmir-Riahi, M. Polissiou,An overview of structural features of DNA and RNA complexes with saffroncompounds: models and antioxidant activity, J. Photochem. Photobiol., B 95(2009) 204–212.
[42] K.A. Connors, Binding Constants: The Measurement of Molecular ComplexStability, Wiley, New York, 1987.
[43] S. Nafisi, A.A. Saboury, N. Keramat, J.-F. Neault, H.-A. Tajmir-Riahi, Stability andstructural features of DNA intercalation with ethidium bromide, acridineorange and methylene blue, J. Mol. Struct. 827 (2007) 35–43.
[44] A. Shah, R. Qureshi, N.K. Janjua, S. Haque, S. Ahmad, Electrochemical andspectroscopic investigations of protonated ferrocene–DNA intercalation, Anal.Sci. 24 (2008) 1437–1441.
[45] G. Tyagi, D.K. Jangir, P. Singh, R. Mehrotra, DNA interaction studies of ananticancer plant alkaloid, vincristine, using Fourier transform infraredspectroscopy, DNA Cell Biol. 29 (2010) 693–699.
Copyright © 2022 FDOKUMEN




















