
Identification and characterization of barley RNA-directed RNA polymerases

Translesion Synthesis of 8,5�-Cyclopurine-2�-deoxynucleosidesby DNA Polymerases �, �, and � *□S
Received for publication, April 24, 2013, and in revised form, August 20, 2013 Published, JBC Papers in Press, August 21, 2013, DOI 10.1074/jbc.M113.480459
Changjun You‡, Ashley L. Swanson§1, Xiaoxia Dai‡, Bifeng Yuan‡, Jianshuang Wang‡, and Yinsheng Wang‡§2
From the ‡Department of Chemistry and the §Environmental Toxicology Graduate Program, University of California,Riverside, California 92521-0403
Background: The 8,5�-cyclopurine-2�-deoxynucleosides (cPus) are important types of oxidative DNA damage.Results: cPus exhibit both inhibitory and mutagenic effects on replication; polymerases (Pols) �, �, and � are involved intranslesion synthesis of these lesions.Conclusion: Pols �, �, and � cooperatively promote translesion synthesis across cPu lesions.Significance: This work revealed the effects of cPus on DNA replication in mammalian cells.
Reactive oxygen species can give rise to a battery ofDNAdam-age products including the 8,5�-cyclo-2�-deoxyadenosine (cdA)and 8,5�-cyclo-2�-deoxyguanosine (cdG) tandem lesions. The8,5�-cyclopurine-2�-deoxynucleosides are quite stable lesionsand are valid and reliable markers of oxidative DNA damage.However, it remains unclear how these lesions compromiseDNAreplication inmammalian cells. Previous in vitrobiochem-ical assays have suggested a role for humanpolymerase (Pol)� inthe insertion step of translesion synthesis (TLS) across the (5�S)diastereomers of cdA and cdG. Using in vitro steady-statekinetic assay, herein we showed that human Pol � and a two-subunit yeast Pol � complex (REV3/REV7) could function effi-ciently in the insertion and extension steps, respectively, of TLSacross S-cdA and S-cdG; human Pol � and Pol � could alsoextend past these lesions, albeit much less efficiently. Resultsfrom a quantitative TLS assay showed that, in human cells,S-cdA and S-cdG inhibited strongly DNA replication andinduced substantial frequencies of mutations at the lesion sites.Additionally, Pol�, Pol �, andPol �, but not Pol�, had importantroles in promoting replication through S-cdA and S-cdG inhuman cells. Based on these results, we propose amodel for TLSacross S-cdA and S-cdG in human cells, where Pol � and/or Pol� carries out nucleotide insertion opposite the lesion, whereasPol � executes the extension step.
Living cells are constantly challenged by endogenous andexogenous genotoxins (1). The resultant DNA damage can leadtomutations and altered gene function, cell senescence, or apo-ptosis (2). Tomaintain normal cellular function, multiple DNAdamage surveillance and repair systems have evolved (3).Among them, translesion synthesis (TLS)3 is a damage toler-
ance pathway by which mammalian cells overcome replicationblockages imposed by various types of DNA damage (4, 5). Themost abundant class of TLS polymerases belong to the Y-fam-ily, including DNA polymerase � (Pol �), Pol �, Pol �, and REV1(5). In this regard, Pol�, Pol �, and Pol � usually playmajor rolesin the insertion step of TLS with occasional involvement in thesubsequent extension step (6). In addition, REV1 is widelythought to serve as a scaffold protein to facilitate the recruit-ment of other TLS polymerases, including Pol �, Pol �, Pol �,and, a B-family polymerase, Pol � (7–10). Recently, it was sug-gested that yeast (andpresumablymammalian) Pol � comprisedfour subunits, including REV3, REV7, Pol31, and Pol32 (11, 12).Reactive oxygen species, formed as byproducts of normal
aerobic metabolism as well as from exposure to ionizing radia-tion, can induce a battery of DNA damage products includingthe 5�R and 5�S diastereomers of 8,5�-cyclo-2�-deoxyadenosine(cdA) and 8,5�-cyclo-2�-deoxyguanosine (cdG) (Fig. 1) (13, 14).8,5�-Cyclopurine-2�-deoxynucleosides (cPus) are stable lesionsand are found to be reliablemarkers for oxidative DNAdamage(13, 15, 16). These cPu lesions have been readily detected invitro and in vivo under various conditions (13, 17–19). More-over, it was recently shown that cPu lesions can accumulate inmammalian tissues in an age-dependent and tissue-specificmanner, which may accelerate the natural processes of agingand contribute to the development of cancer, neurodegenera-tion, and other human diseases (13, 16, 18, 20, 21).Unlike most other reactive oxygen species-induced DNA
lesions that are removed by base excision repair, cPu lesions areunique due to the presence of a C8-C5� bond between thepurine base and 2-deoxyribose of the same nucleoside (Fig. 1).This additional covalent bond induces helical distortion toDNA and substantially stabilizes the glycosidic bond againstacid-induced hydrolysis, which may prevent initiation of baseexcision repair by aDNAglycosylase and render the cPu lesionsattractive substrates for nucleotide excision repair (13, 21).Indeed, multiple lines of evidence supports that the cPu lesions
* This work was supported, in whole or in part, by National Institutes of HealthGrant R01 CA101864.
□S This article contains supplemental Tables S1–S8 and Figs. S1–S4.1 Supported by an National Research Service Award T32 Training Grant T32
ES018827.2 To whom correspondence should be addressed: Dept. of Chemistry, Univer-
sity of California, Riverside, CA 92521-0403. Tel.: 951-827-2700; Fax: 951-827-4713; E-mail: [email protected].
3 The abbreviations used are: TLS, translesion synthesis; S-cdA, (5�S)-8,5�-cy-clo-2�-deoxyadenosine; S-cdG, (5�S)-8,5�-cyclo-2�-deoxyguanosine; cPu,
8,5�-cyclopurine-2�-deoxynucleoside; Pol �, DNA polymerase �; Pol �, DNApolymerase �; Pol �, DNA polymerase �; Pol �, DNA polymerase �; SSPCR-CRAB, strand-specific PCR-based competitive replication and adductbypass; RMF, relative mutation frequency; RBE, relative bypass efficiency;ODN, oligodeoxyribonucleotide.
THE JOURNAL OF BIOLOGICAL CHEMISTRY VOL. 288, NO. 40, pp. 28548 –28556, October 4, 2013© 2013 by The American Society for Biochemistry and Molecular Biology, Inc. Published in the U.S.A.
28548 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 40 • OCTOBER 4, 2013

are repaired by nucleotide excision repair, but not by base exci-sion repair or direct enzymatic reversal (22–26).Unrepaired cPu lesions may lead to detrimental biological
consequences. In this regard, S-cdA and S-cdG located in thetemplate DNA inhibit strongly DNA transcription and inducethe generation of mutant transcripts in mammalian cells (22,23, 26). In addition, a single cPu lesion may alter gene expres-sion by preventing the binding of transcription factors and reg-ulatory proteins to their recognition sequences (27, 28). S-cdAand S-cdG can also act as strong inhibitors to DNA replicationand induce nucleobase substitutions at the lesion site in Esche-richia coli cells (25, 29). However, it is still unclear how thesecPu lesions compromise DNA replication in mammalian cells.In addition, although previous biochemical studies have sug-gested a role for Pol � in the TLS across the cPu lesions in vitro(30, 31), it remains unknown whether Pol � is involved in thereplicative bypass of cPu lesions in mammalian cells, andmuchless is known about whether other TLS polymerases, includingPol �, Pol �, and Pol �, participate in the TLS across cPu lesionsin vitro and in mammalian cells.To address these questions, herein we performed in vitro
steady-state kinetic assays to understand the roles of Pol �, Pol�, and Pol � in the replicative bypass of cPu lesions. We alsodeveloped a strand-specific PCR-based competitive replicationand adduct bypass (SSPCR-CRAB) assay and quantitativelyexamined how S-cdA and S-cdG lesions compromise the effi-ciency and accuracy of DNA replication in human cells. More-over, we investigated the relative roles of TLS polymerases,including Pol �, Pol �, Pol �, and Pol �, in the bypass of S-cdAand S-cdG using human cells that are completely or partiallydeficient in one or more of these TLS Pols.
EXPERIMENTAL PROCEDURES
Materials and Cell Culture Conditions—Unmodified oli-godeoxyribonucleotides (ODNs), shrimp alkaline phosphatase,chemicals, and [�-32P]ATP were purchased from IntegratedDNA Technologies, U. S. Biochemical Corp., Sigma, andPerkinElmer Life Sciences, respectively. The 12-mer lesion-bearing ODNs 5�-ATGGCGXGCTAT-3� (“X” representsS-cdA or S-cdG) were previously synthesized following pub-lished procedures (29, 32). Human DNA Pol �, Pol �, and atwo-subunit Saccharomyces cerevisiae DNA Pol � complex(Rev3/Rev7) were purchased from Enzymax, and human DNAPol � was provided by Professor Roger Woodgate. All otherenzymes unless otherwise specified were purchased from NewEngland BioLabs. The 293T human embryonic kidney epithe-lial cells were purchased from ATCC. The SV40-transformed
Pol �-deficient XP30RO fibroblasts and the corrected cells(XP30RO� Pol�) were provided by Professor James E. Cleaver(33, 34). Cells were cultured in Dulbecco’s modified Eagle’smedium supplemented with 10% fetal bovine serum (Invitro-gen), 100 units/ml of penicillin, and 100 �g/ml of streptomycin(ATCC), and incubated at 37 °C in 5% CO2 atmosphere.Steady-state Kinetic Measurements—Steady-state kinetic
assays were carried out following previously published proce-dures (35). The primer-template complex consisted of a 20-merS-cdA- or S-cdG-bearing template and a 5�-32P-labeled 13-mer,14-mer (with the correct nucleosides being placed opposite thelesions), or 15-mer (carrying the correct nucleosides oppositethe lesions and their adjacent 5� nucleoside, Figs. 2 and 3)primer (0.05 �M).Template Preparation for in Vivo Replication Studies—We
first constructed a parent vector for S-cdA by modifying thesequence of the original pTGFP-Hha10 plasmid, which con-tains an SV40 origin and is able to replicate in the SV40-trans-formed mammalian cells (36, 37). To this end, a 50-mer ODNwith the sequence 5�-AATTCGCAGCGAGTCATCCATGG-CGAGGTATTGTGGAGTCGATGCATCCG-3� was annealedwith its complementary strand and ligated to an NheI-EcoRIrestriction fragment from the pTGFP-Hha10 plasmid.We nextconstructed the S-cdA-bearing double-stranded shuttle vectorby using a previously described method (Fig. 4A) (36, 37).Briefly, we nicked the parent vector for S-cdA with Nt.BstNBIto produce a gapped vector by removing a 25-mer single-strandedODN, followed by filling the gapwith a 12-mer S-cdA-bearing ODN (5�-ATGGCGXGCTAT-3�, X � S-cdA) and a13-mer unmodified ODN (5�-TGTGGAGTCGATG-3�). Theligationmixture was incubatedwith ethidium bromide, and theresulting supercoiled lesion-bearing plasmidwas purified usingagarose gel electrophoresis. Using the same method, we pre-pared the lesion-free control and competitor vectors for S-cdA,where the 12-mer S-cdA-containing ODN was replaced with a12-merODN(5�-ATGGCGAGCTAT-3�) anda15-merODN(5�-ATGGCGAGCAGCTAT-3�), respectively. The S-cdG-bearingvector as well as its corresponding undamaged control and com-petitor vectors were constructed in a similar fashion.In Vivo Transfection and Plasmid Isolation—The lesion-
bearing or the corresponding non-lesion control plasmids werepremixed with the competitor genome for in vivo transfection,with the molar ratios of competitor vector to control andlesion-bearing genome being 1:1 and 1:19, respectively. The293T human embryonic kidney epithelial cells, XP30RO, andXP30RO � Pol � cells (1 � 105) were seeded in 24-well platesand cultured overnight, after which they were transfected with300 ng of mixed genome by using Lipofectamine 2000 (Invitro-gen) following the manufacturer’s instructions. The cells wereharvested 24 h after transfection, and the progenies of the plas-mid were isolated using the Qiagen Spin Kit (Qiagen), withminor changes (38). The residual unreplicated plasmidwas fur-ther removed by DpnI digestion, followed by digesting theresulting linear DNA with exonuclease III as described else-where (39–41).Highly efficient depletion of specific TLS DNA polymerases
was achieved using siRNAs that were previously validated (42,43). All siRNAs were purchased from Dharmacon: POLK
FIGURE 1. Chemical structures of S-cdA and S-cdG.
Translesion Synthesis of 8,5�-Cyclopurine-2�-deoxynucleoside
OCTOBER 4, 2013 • VOLUME 288 • NUMBER 40 JOURNAL OF BIOLOGICAL CHEMISTRY 28549

SMARTpool (L-021038), POLI SMARTpool (L-019650), POLHSMARTpool (M-006454), REV3L SMARTpool (L-006302),and siControl non-targeting pool (D-001210). The 293T cellswere seeded in 24-well plates at 40–60% confluence level andtransfected with�25 pmol of siRNAs for each gene using Lipo-fectamine 2000 (Invitrogen). After a 48-h incubation, 300 ng ofmixed genome was co-transfected into the cells together withanother aliquot of siRNA by using Lipofectamine 2000. Theprogenies of the plasmid were isolated from the cells 24 h aftertransfection as described above.Real-time Quantitative RT-PCR—Total RNA was extracted
from the cells 48 h after transfectionwith siRNAusing theTotalRNA Kit I (Omega), and subjected to DNase I treatment withthe DNA-free kit (Ambion) to eliminate the DNA contamina-tion. cDNA was generated by using Moloney murine leukemiavirus reverse transcriptase (Promega) and an oligo(dT)16primer. Real-time quantitative RT-PCR for evaluating thesiRNA knockdown efficiency was performed by using the iQSYBR Green Supermix kit (Bio-Rad) and gene-specific primersfor POLK, POLI, POLH, REV3L, or the control geneGAPDH asdescribed elsewhere (22), and the primers for real-time RT-PCR shown in supplemental Table S1.Western Blot Analysis—Western analysis was performed
with a total of 40 �g of whole cell lysate. Antibodies that specif-ically recognize human Pol �, Pol �, Pol �, or REV3L were pur-chased from Santa Cruz Biotechnology and used at a 1:10,000dilution. Human �-actin antibody (Abcam) was used at a1:5,000 dilution. Horseradish peroxidase-conjugated second-ary goat anti-mouse antibody (SantaCruz Biotechnology), don-key anti-goat antibody (Santa Cruz Biotechnology), and goatanti-rabbit antibody (Abcam) were used at a 1:10,000 dilution.PCR, PAGE, and LC-MS/MS Analyses—The progeny genomes
arising from in vivo replicationwere amplified in a PCR (GoTaqgreenmastermix, Promega) containing a pair of primers whoseproducts cover the initial lesion site and span 8 DpnI recogni-tion sites. The primers were 5�-GCTAGCGGATGCATCGAC-TCCACAACAG-3� and 5�-GGCTCCCTTTAGGGTTCCGA-TTTAGTG-3�, and the PCR amplification started at 95 °C for 2min; then, 35 cycles at 95 °C for 30 s, 65 °C for 30 s, and 72 °C for1.5min, and a final 5-min extension at 72 °C. The PCRproductswere purified using a QIAquick PCR Purification Kit (Qiagen)and stored at �20 °C until use.For PAGE analysis, a portion of the PCR fragments was
treated with 5 units of NcoI and 1 unit of shrimp alkaline phos-phatase at 37 °C in 10 �l of New England Biolab buffer 3 for 1 h,followed by heating at 80 °C for 20min to deactivate the shrimpalkaline phosphatase. The abovemixture was then treated in 15�l of New England Biolabs buffer 3 with 5 mM DTT, ATP (50pmol of cold, premixed with 1.66 pmol of [�-32P]ATP), and 5units of T4 polynucleotide kinase. The reaction was continuedat 37 °C for 1 h, followed by heating at 65 °C for 20 min todeactivate the T4 polynucleotide kinase. To the reaction mix-ture 5 units of SfaNI was subsequently added, and the solutionwas incubated at 37 °C for 1 h, followed by quenchingwith 15�lof formamide gel loading buffer. Themixturewas loaded onto a30%polyacrylamide gel (acrylamide:bis-acrylamide� 19:1) andproducts were quantified by phosphorimager analysis. Similarto a previously described method for assessing the impact of
DNA lesions on transcription (22, 44), we determined theeffects of DNA lesions on replication efficiency and fidelity bythe “relative bypass efficiency” (RBE) and “relative mutationfrequency” (RMF) values, respectively. Briefly, the RMF valuewas determined from the relative amounts of different productsarising from replication of the lesion-containing genome. TheRBE value was calculated using the following formula, %RBE �(lesion signal/competitor signal)/(non-lesion control signal/competitor signal) (45, 46).LC-MS/MSwas used to further identify the replication prod-
ucts arising from S-cdA- or S-cdG-bearing substrates similar tothose described elsewhere (22, 29, 37, 44). Briefly, PCR prod-ucts were treated with 50 units of NcoI and 20 units of shrimpalkaline phosphatase in 250 �l of New England Biolabs buffer 3at 37 °C for 4 h, followed by heating at 80 °C for 20 min. To theresulting solution was added 50 units of SfaNI, and the reactionmixture was incubated at 37 °C for 4 h followed by extraction
FIGURE 2. Translesion synthesis of cPu lesions in vitro. A, the 20-mer tem-plate and primer sequences used for the in vitro replication studies. B and C,relative efficiencies (with respect to the corresponding damage-free sub-strates) of nucleotide incorporation across and past S-cdA (B) and S-cdG (C) byhuman Pol �, �, and �, and a two-subunit yeast Pol � complex (REV3/REV7).The efficiencies of Pol �-mediated nucleotide insertion opposite the lesionsite or the neighboring 5� nucleotide of S-cdA or S-cdG were published pre-viously (31).
Translesion Synthesis of 8,5�-Cyclopurine-2�-deoxynucleoside
28550 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 40 • OCTOBER 4, 2013

with phenol/chloroform/isoamyl alcohol (25:24:1, v/v). Theaqueous portion was dried with Speed-Vac, desalted withHPLC, and dissolved in 12 �l of water. The ODN mixture wassubjected to LC-MS/MS analysis, as described elsewhere (18, 25,31, 38). An LTQ linear ion trapmass spectrometer (ThermoElec-tron) was set up for monitoring fragmentation of the [M-3H]3�
ions of the 13-mer ODNs (d(CATGGCGMGCTGT), where “M”designates A, T, C, or G).
RESULTS
Roles of TLS Pols in the Bypass of S-cdA and S-cdG in Vitro—We first performed steady-state kinetic assays to determine theability of human Pol � and Pol �, as well as yeast Pol � to insertnucleotides opposite the S-cdA and S-cdG, and their respectiveundamaged substrates (Fig. 2A). In this regard, due to the lackof availability of recombinant mammalian Pol � (6, 47), hereinweused a two-subunit S. cerevisiaePol � complex (REV3/REV7)instead. In this regard, a heterodimeric complex, including theREV3 and REV7 subunits, was previously identified as themin-imal assembly required for catalytic activity of Pol � in vitro (4,11). Our results showed that, like Pol � (31), Pol � was highlyerror-free and efficient in inserting nucleotides opposite thetwo cPu lesions; relative to the corresponding insertion for theunmodified substrates, incorporation of the correct nucleo-tides opposite S-cdA and S-cdG occurred at frequencies of�59and�61%, respectively (Figs. 2,B andC, and 3, andTable 1).Onthe other hand, Pol � and Pol � (REV3/REV7) incorporatednucleotides opposite the cPu lesions with extremely low effi-ciency (Fig. 2, B and C, and supplemental Tables S2 and S3).We next assessed the accuracy and efficiency of nucleotide
incorporation opposite the adjacent 5� nucleoside of the cPulesions, where we employed a 14-mer primer with the correct
dNTP being placed opposite the cPu lesions and their respec-tive controls (supplemental Fig. S1). It turned out that the effi-ciency for nucleotide incorporation opposite the 5� neighbor-ing nucleoside (at�1 position) was very poor with Pol � or Pol �(Fig. 2, B andC, and supplemental Tables S4 and S5). However,Pol � (REV3/REV7) was modestly efficient, albeit error-prone,in extending past the S-cdA:dT and S-cdG:dC pairs (Fig. 2, Band C, and supplemental Tables S6). In this regard, when thecorrect S-cdA:dT pair was placed at the primer-template junc-tion, yeast Pol � (REV3/REV7) preferentially incorporates thecorrect dCMP opposite dG at the �1 position, with misincor-
FIGURE 3. Representative gel images for steady-state kinetic assays measuring nucleotide insertion opposite S-cdA, S-cdG, or corresponding unmod-ified dA or dG by human Pol �. The highest concentrations of dNTPs are indicated below the leftmost lane in each gel image, and no dNTP was added for therightmost lane.
TABLE 1Steady-state kinetic parameters for human Pol �-mediated nucleotideinsertion opposite S-cdA and S-cdG, and unmodified dA and dG
dNTP kcat Km kcat/Km fincmin�1 �M �M�1min�1
S-cdA lesion containingsubstrate
dTTP 32 � 4 0.19 � 0.02 170 � 20 1dGTP 28 � 0.3 2.5 � 0.3 11 � 2 6.5 � 10�2
dCTP 16 � 0.2 63 � 7 0.25 � 0.03 1.5 � 10�3
dATP 21 � 2 4.9 � 0.5 4.3 � 0.5 2.5 � 10�2
dA controldTTP 32 � 4 0.11 � 0.01 290 � 30 1dGTP 11 � 0.5 210 � 20 0.052 � 0.003 1.1 � 10�3
dCTP 40 � 3 400 � 30 0.089 � 0.002 1.8 � 10�3
dATP 10 � 1 310 � 13 0.033 � 0.001 6.9 � 10�4
S-cdG lesion containingsubstrate
dTTP 20 � 2 140 � 20 0.14 � 0.02 7.4 � 10�4
dGTP 28 � 3 95 � 8 3.4 � 0.4 1.8 � 10�2
dCTP 39 � 4 0.21 � 0.02 190 � 20 1dATP 27 � 3 2.8 � 0.3 9.8 � 1 5.2 � 10�2
dG controldTTP 21 � 2 88 � 9 0.24 � 0.03 7.7 � 10�4
dGTP 25 � 3 93 � 10 0.27 � 0.03 8.7 � 10�4
dCTP 28 � 3 0.09 � 0.01 310 � 30 1dATP 23 � 2 58 � 6 0.39 � 0.04 1.3 � 10�3
Translesion Synthesis of 8,5�-Cyclopurine-2�-deoxynucleoside
OCTOBER 4, 2013 • VOLUME 288 • NUMBER 40 JOURNAL OF BIOLOGICAL CHEMISTRY 28551

poration of dAMP, dTMP, and dGMP occurring at frequenciesof 0.65, 1.6, and 35%, respectively (supplemental Table S6).Similarly, Pol � (REV3/REV7)was highly error-prone in extend-ing past the S-cdG:dC pair, with misincorporation of dAMP,dTMP, and dGMP opposite the �1 nucleoside (dG) occurringat frequencies of 10, 21, and 46%, respectively (supplementalTable S6).We further obtained the steady-state kinetic parameters for
yeast Pol � (REV3/REV7)-, as well as human Pol �- and Pol�-mediated nucleotide insertion at the �2 position withrespect to the two cPu lesions. The results showed that yeast Pol� (REV3/REV7) extended past the S-cdA- and S-cdG-bearingsubstrates with high accuracy and efficiencies; relative to thecorresponding insertion for the unmodified substrates, inser-tion of the correct nucleotides opposite the �2 base in theS-cdA- and S-cdG-bearing templates occurred at frequencies of�86 and �68%, respectively (Fig. 2, B and C, and supplementalTable S7). In addition, Pol �-mediated extension past thelesion-containing substrates was moderately efficient, as inser-tion of the correct dGMP occurred at efficiencies of �40% rel-ative to the corresponding extension past the dA- and dG-con-taining substrates (Fig. 2,B andC, and supplemental Fig. S2 andTable S8). On the other hand, human Pol � was also able toextend past S-cdA and S-cdG when the correct nucleosides areplaced opposite the lesions and their adjacent 5� nucleosides,albeit much less efficiently (by �6–10-fold) than the corre-sponding extension for the unmodified substrates (supplemen-tal Table S8).Effects of S-cdA and S-cdG on Replication in Human Cells—To
investigate how S-cdA and S-cdG compromise the efficiencyand fidelity of DNA replication in human cells, we developed anew SSPCR-CRAB assay. This method evolved from the prin-ciple of the traditional competitive replication and adductbypass assay, which has beenwidely used for assessing the cyto-toxic and mutagenic properties of DNA lesions placed in a sin-gle-stranded M13 genome in E. coli cells (45, 46).
We constructed double-stranded shuttle vectors containinga single site-specific lesion (S-cdA or S-cdG), as well as thecorresponding non-lesion control vectors housing an unmodi-fied nucleotide (A or G) at the lesion site. Given that the lesion-bearing strandmay be replicated at a reduced efficiency from itsopposing undamaged strand in mammalian cells (48, 49), weemployed a similar strategy as described previously (37, 48, 49)and incorporated a C/C mismatch two nucleotides away fromthe lesion site to distinguish the replication products formedfrom the two strands (Fig. 4A). The lesion-bearing or undam-aged control vectors were mixed individually with an undam-aged competitor vector at givenmolar ratios and co-transfectedinto human cells (Fig. 4B). In this regard, the competitor vectorcontains three more nucleotides than the control vector situ-ated between the two restriction sites used for SSPCR-CRABassay. The progenies of the plasmids were isolated from humancells 24 h after transfection, and residual unreplicated plasmidswere removed by a combined treatment with DpnI and exonu-clease III as described previously (39–41). The progenygenomes were subsequently amplified using a pair of PCRprimers spanning the lesion site. Notably, one of the primers(P1) carries a G as the terminal 3�-nucleotide corresponding to
the C/Cmismatch site (Fig. 4B), which is used to amplify selec-tively the progeny genomes arising from the replication of thebottom, but not the top strand of the plasmids under appropri-ate conditions (50). In addition, P1 was designed deliberately tocarry a C/A mismatch three bases from its 3�-end (Fig. 4B) toincrease the specificity of strand-specific PCR, as described pre-viously (50). The resulting PCR products were digested withappropriate restriction enzymes, i.e. NcoI and SfaNI, and sub-jected to LC-MS/MS and PAGE analyses.We first asked whether Pol � is involved in TLS across cPu
lesions in human cells. To this end, we performed the SSPCR-CRAB assay using the Pol �-deficient XP30RO fibroblasts andthe corrected cells (XP30RO � Pol �) as hosts for in vivo repli-cation. PAGE analysis of restriction fragments of PCR productsshowed that the RBE values for S-cdA and S-cdG in Pol �-de-ficient XP30RO cells were �3 and �2%, respectively, and the
FIGURE 4. Schematic diagrams showing the procedures for the prepara-tion of the lesion-bearing plasmid (A) and the SSPCR-CRAB assay (B) (see“Experimental Procedures” for details). Only the construction of S-cdA-bearing vector is shown. SV40 ORI and X indicate SV40 replication origin andS-cdA, respectively. The C/C mismatch site is underlined. The cleavage sites ofNt.BstNBI, NcoI, and SfaNI are designated with arrows. P1 represents one ofthe primers for PCR, i.e. d(GCTAGCGGATGCATCGACTCCACAACAG). P1 con-tains a G as the terminal 3�-nucleotide corresponding to the C/C mismatchsite of lesion-bearing genome, and it also contains a C/A mismatch threebases away from its 3�-end for improving the specificity of PCR. M representsthe nucleobase formed at the lesion site after replication, and N designatesthe paired nucleobase of M in the complementary strand.
Translesion Synthesis of 8,5�-Cyclopurine-2�-deoxynucleoside
28552 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 40 • OCTOBER 4, 2013
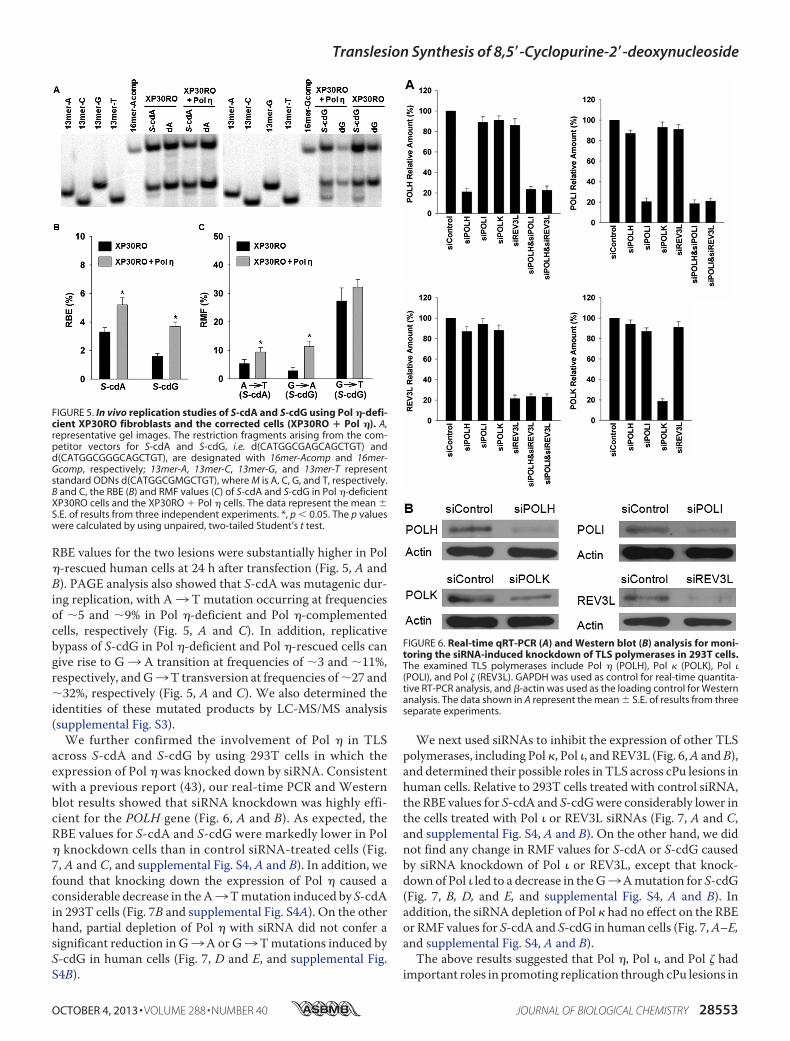
RBE values for the two lesions were substantially higher in Pol�-rescued human cells at 24 h after transfection (Fig. 5, A andB). PAGE analysis also showed that S-cdA was mutagenic dur-ing replication, with A3 T mutation occurring at frequenciesof �5 and �9% in Pol �-deficient and Pol �-complementedcells, respectively (Fig. 5, A and C). In addition, replicativebypass of S-cdG in Pol �-deficient and Pol �-rescued cells cangive rise to G3 A transition at frequencies of �3 and �11%,respectively, andG3T transversion at frequencies of�27 and�32%, respectively (Fig. 5, A and C). We also determined theidentities of these mutated products by LC-MS/MS analysis(supplemental Fig. S3).
We further confirmed the involvement of Pol � in TLSacross S-cdA and S-cdG by using 293T cells in which theexpression of Pol � was knocked down by siRNA. Consistentwith a previous report (43), our real-time PCR and Westernblot results showed that siRNA knockdown was highly effi-cient for the POLH gene (Fig. 6, A and B). As expected, theRBE values for S-cdA and S-cdG were markedly lower in Pol� knockdown cells than in control siRNA-treated cells (Fig.7, A and C, and supplemental Fig. S4, A and B). In addition, wefound that knocking down the expression of Pol � caused aconsiderable decrease in theA3Tmutation induced by S-cdAin 293T cells (Fig. 7B and supplemental Fig. S4A). On the otherhand, partial depletion of Pol � with siRNA did not confer asignificant reduction in G3A or G3Tmutations induced byS-cdG in human cells (Fig. 7, D and E, and supplemental Fig.S4B).
We next used siRNAs to inhibit the expression of other TLSpolymerases, including Pol �, Pol �, and REV3L (Fig. 6,A andB),and determined their possible roles in TLS across cPu lesions inhuman cells. Relative to 293T cells treated with control siRNA,the RBE values for S-cdA and S-cdGwere considerably lower inthe cells treated with Pol � or REV3L siRNAs (Fig. 7, A and C,and supplemental Fig. S4, A and B). On the other hand, we didnot find any change in RMF values for S-cdA or S-cdG causedby siRNA knockdown of Pol � or REV3L, except that knock-down of Pol � led to a decrease in theG3Amutation for S-cdG(Fig. 7, B, D, and E, and supplemental Fig. S4, A and B). Inaddition, the siRNA depletion of Pol � had no effect on the RBEor RMF values for S-cdA and S-cdG in human cells (Fig. 7,A–E,and supplemental Fig. S4, A and B).
The above results suggested that Pol �, Pol �, and Pol � hadimportant roles in promoting replication through cPu lesions in
FIGURE 5. In vivo replication studies of S-cdA and S-cdG using Pol �-defi-cient XP30RO fibroblasts and the corrected cells (XP30RO � Pol �). A,representative gel images. The restriction fragments arising from the com-petitor vectors for S-cdA and S-cdG, i.e. d(CATGGCGAGCAGCTGT) andd(CATGGCGGGCAGCTGT), are designated with 16mer-Acomp and 16mer-Gcomp, respectively; 13mer-A, 13mer-C, 13mer-G, and 13mer-T representstandard ODNs d(CATGGCGMGCTGT), where M is A, C, G, and T, respectively.B and C, the RBE (B) and RMF values (C) of S-cdA and S-cdG in Pol �-deficientXP30RO cells and the XP30RO � Pol � cells. The data represent the mean �S.E. of results from three independent experiments. *, p � 0.05. The p valueswere calculated by using unpaired, two-tailed Student’s t test.
FIGURE 6. Real-time qRT-PCR (A) and Western blot (B) analysis for moni-toring the siRNA-induced knockdown of TLS polymerases in 293T cells.The examined TLS polymerases include Pol � (POLH), Pol � (POLK), Pol �(POLI), and Pol � (REV3L). GAPDH was used as control for real-time quantita-tive RT-PCR analysis, and �-actin was used as the loading control for Westernanalysis. The data shown in A represent the mean � S.E. of results from threeseparate experiments.
Translesion Synthesis of 8,5�-Cyclopurine-2�-deoxynucleoside
OCTOBER 4, 2013 • VOLUME 288 • NUMBER 40 JOURNAL OF BIOLOGICAL CHEMISTRY 28553

human cells. To further ascertain whether these three poly-merases function in the same or different pathways for replica-tive bypass of cPu lesions, we assessed the effects of simultane-ous knockdowns of two of the three TLS Pols (Fig. 6, A and B).Our results showed that knockdown of both Pol � and REV3Lhad an effect similar to that of Pol � alone (Fig. 7, A–E, andsupplemental Fig. S4, C and D). Similarly, we found that thedual knockdown of Pol � and Pol �, or Pol � and REV3L, did notcause further change in TLS frequencies relative to thatobserved upon the knockdown of one of these polymerasesalone (Fig. 7, A–E, and supplemental Fig. S4, C and D). There-fore, Pol �, Pol �, and Pol � may cooperate with each other tocarry out TLS across S-cdA and S-cdG in human cells.
DISCUSSION
It has been previously shown that S-cdA is a strong blockage tomammalian DNA polymerase � and T7DNA polymerase in vitro(23), and the bypass efficiencies of S-cdA and S-cdG during repli-cation are very low (less than �5%) in wild-type E. coli cells (25,29). In agreement with these findings, our results demonstratedthat both S-cdA and S-cdG constituted strong blockage to DNAreplicationmachinery in human cells.We also found that replica-tive bypass of S-cdA and S-cdG generated mutations in humancells. Similar to the observation in E. coli cells (25, 29), S-cdA was
weaklymutagenic and inducedA3T transversion at a frequencyof�10% in 293Tcells.On the other hand, themajor type ofmuta-tion induced by S-cdG is the G 3 A transition occurring at afrequency of �20%, although a low frequency (less than �5%) ofG3Ttransversionwasalsoobserved inE. colicells (25, 29).Com-paredwithE. colicells, replicativebypassofS-cdGwasmoremuta-genic in human 293T cells (�50%), where we observed a higherfrequency of G3T than the G3Amutation. The strong block-ing andmutagenic effects of the cPu lesions onDNAreplication inhuman cells, together with the abundant presence of these lesionsinmammalian tissues (13, 16, 18, 20, 21), suggest that these lesionsmay constitute significant endogenousDNA lesions thatmay playimportant roles in the development of human diseases.We further demonstrated a role for Pol � in promoting rep-
lication through the cPu lesions in vivo, by using human cellsthat are completely or partially deficient in Pol �. Consistentwith our results, depletion of its ortholog, i.e. Pol V, also causeddecreased bypass efficiencies of S-cdA and S-cdG in E. coli cells(25, 29). In addition, depletion of Pol� in human cells conferreda considerable reduction in mutation frequency of S-cdA, sug-gesting that Pol � may be involved in error-prone replicativebypass of this lesion inmammalian system. This result is some-what surprising, as our previous biochemical studies have sug-
FIGURE 7. Effects of siRNA knockdowns of TLS Pols on the replicative bypass of S-cdA and S-cdG in human cells. Shown in A and C are the RBE values ofS-cdA and S-cdG, respectively. Shown in B, D, and E are the RMF values of A3 T induced by S-cdA, G3 T and G3A mutations induced by S-cdG, respectively.The examined TLS polymerases include Pol � (POLH), Pol � (POLK), Pol � (POLI), and Pol � (REV3L). The data represent the mean � S.E. of results from threeindependent experiments. Significantly different RBE or RMF values from that of the corresponding control siRNA (siControl) are indicated with an asterisk (p �0.05). The p values were calculated by using unpaired, two-tailed t test.
Translesion Synthesis of 8,5�-Cyclopurine-2�-deoxynucleoside
28554 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 40 • OCTOBER 4, 2013

gested a role for Pol � in the error-free nucleotide insertionopposite cPu lesions (31). The difference in the fidelity of lesionbypass by Pol � between in vitro and in vivo studies may beattributed to two factors. First, it has been proposed that theTLS machinery requires the functional interaction betweenTLS Pols and other accessory proteins, which may modulatethe fidelity and/or efficiency of TLS Pols in lesion bypass in cells(6, 43, 51, 52). Second, the in vitro steady-state kinetic experi-ments were conducted in the presence of one nucleotide at atime, whereas the replication in cells occurs in themutual pres-ence of all four canonical nucleotides. Thus, the absence orpresence of competition in nucleotide incorporation under thetwo replication conditions may also result in different efficien-cies and fidelities of TLS (53).We also demonstrated, for the first time, that Pol � and Pol �
have important roles in translesion synthesis of cPu lesions.Our biochemical results demonstrated that human Pol � andyeast Pol � (REV3/REV7) could function efficiently in the inser-tion and extension steps, respectively, of TLS across S-cdA andS-cdG in vitro. Along this line, siRNA knockdown of Pol � andREV3L led to a substantial decrease in RBE values for S-cdAand S-cdG in human cells. Like in the case of Pol �, although invitro biochemical results showed that Pol � predominantlyincorporated the correct nucleotides opposite the cPu lesions,our in vivo replication studies showed that, siRNA knockdownof Pol � could cause a reduction in S-cdG-inducedG3Amuta-tion. This result suggested that Pol � may play a role in error-prone insertion opposite this lesion during replication inhuman cells.Moreover, we found that the simultaneous depletion of any
two of Pol �, Pol �, and Pol � had no additive effects on thereplicative bypass of cPu lesions, indicating that these threepolymerases might function in the same or partly overlappinggenetic pathways for bypassing cPu lesions in human cells. Onthe other hand, although human Pol � exhibited its ability toextend past these cPu lesions in vitro, knocking down theexpression of Pol � had no effect on TLS across S-cdA andS-cdG in human cells. This is in keeping with previous dataindicating that Pol IV, an ortholog of Pol �, may not be able tobypass these cPu lesions in E. coli cells (25, 29).It should be noted that we observed lower frequencies of the
G3 A mutation induced by S-cdG in Pol �-deficient humanfibroblast cells than the isogenic cells rescued with wild-typehuman Pol �; however, siRNA knockdown of Pol � had nosignificant effect on themutation frequencies of S-cdG in 293Tcells. This result may reflect a role for the residual Pol � in TLSacross S-cdG in cells. In this vein, it was suggested that residualTLS Pols might be sufficient for keeping themiscoding proper-ties of some DNA lesions unchanged, even though sometimeslesion bypass efficiencies could be reduced by the siRNAknockdown of some specific TLS Pols (37, 42, 43).Based on our findings, we proposed a model for the TLS
across cPu lesions in human cells. In this model, a replicativepolymerase stalled by cPu lesions on the template DNA is dis-placed by Pol� and/or Pol �, which inserts a nucleotide oppositethe lesion. A second polymerase switch can then be made,where another TLS polymerase (i.e. Pol �) accurately extendsthe primer terminus past the damage site. In keeping with our
study, two-polymerase mechanisms, in which insertion by one(or more) specific TLS Pol(s) is followed by extension withanother polymerase, has been suggested to play important rolesin the replicative bypass of several other DNA lesions in mam-malian cells. These included thymidine glycol, cis,syn-cyclobu-tane pyrimidine dimer, and pyrimidine(6–4)pyrimidone pho-toproducts in mammalian cells (6, 42, 43, 51, 52). It would beinteresting to unveil how different inserter and extender Pols asproposed here and in other studies (6, 42, 43, 51, 52) coordinatetheir actions during TLS across DNA damage in human cells.
Acknowledgments—We thank Profs. James E. Cleaver, RogerWoodgate, and Timothy R. O’Connor for providing cells and reagents.
REFERENCES1. Lindahl, T. (1993) Instability and decay of the primary structure of DNA.
Nature 362, 709–7152. Maslov, A. Y., and Vijg, J. (2009) Genome instability, cancer and aging.
Biochim. Biophys. Acta 1790, 963–9693. Friedberg, E. C., Aguilera, A., Gellert, M., Hanawalt, P. C., Hays, J. B.,
Lehmann, A. R., Lindahl, T., Lowndes, N., Sarasin, A., and Wood, R. D.(2006) DNA repair. From molecular mechanism to human disease. DNARepair 5, 986–996
4. Lange, S. S., Takata, K., and Wood, R. D. (2011) DNA polymerases andcancer. Nat. Rev. Cancer 11, 96–110
5. Sale, J. E., Lehmann, A. R., and Woodgate, R. (2012) Y-family DNA poly-merases and their role in tolerance of cellular DNAdamage.Nat. Rev.Mol.Cell Biol. 13, 141–152
6. Livneh, Z., Ziv, O., and Shachar, S. (2010)Multiple two-polymerasemech-anisms in mammalian translesion DNA synthesis. Cell Cycle 9, 729–735
7. Guo, C., Fischhaber, P. L., Luk-Paszyc,M. J., Masuda, Y., Zhou, J., Kamiya,K., Kisker, C., and Friedberg, E. C. (2003) Mouse Rev1 protein interactswith multiple DNA polymerases involved in translesion DNA synthesis.EMBO J. 22, 6621–6630
8. Ohashi, E., Murakumo, Y., Kanjo, N., Akagi, J., Masutani, C., Hanaoka, F.,and Ohmori, H. (2004) Interaction of hREV1 with three human Y-familyDNA polymerases. Genes Cells 9, 523–531
9. Murakumo, Y., Ogura, Y., Ishii, H., Numata, S., Ichihara,M., Croce, C.M.,Fishel, R., and Takahashi, M. (2001) Interactions in the error-pronepostreplication repair proteins hREV1, hREV3, and hREV7. J. Biol. Chem.276, 35644–35651
10. Masuda, Y., Ohmae, M., Masuda, K., and Kamiya, K. (2003) Structure andenzymatic properties of a stable complex of the human REV1 and REV7proteins. J. Biol. Chem. 278, 12356–12360
11. Makarova, A. V., Stodola, J. L., and Burgers, P. M. (2012) A four-subunitDNA polymerase Pol � complex containing Pol � accessory subunits isessential for PCNA-mediated mutagenesis. Nucleic Acids Res. 40,11618–11626
12. Johnson, R. E., Prakash, L., and Prakash, S. (2012) Pol31 and Pol32 subunitsof yeast DNApolymerase � are also essential subunits of DNApolymerase�. Proc. Natl. Acad. Sci. U.S.A. 109, 12455–12460
13. Jaruga, P., and Dizdaroglu, M. (2008) 8,5�-Cyclopurine-2�-deoxynucleo-sides in DNA. Mechanisms of formation, measurement, repair and bio-logical effects. DNA Repair 7, 1413–1425
14. Wang, Y. (2008) Bulky DNA lesions induced by reactive oxygen species.Chem. Res. Toxicol. 21, 276–281
15. Shaked, H., Hofseth, L. J., Chumanevich, A., Chumanevich, A. A., Wang,J., Wang, Y., Taniguchi, K., Guma, M., Shenouda, S., Clevers, H., Harris,C. C., and Karin, M. (2012) Chronic epithelial NF-�B activation acceler-ates APC loss and intestinal tumor initiation through iNOS up-regulation.Proc. Natl. Acad. Sci. U.S.A. 109, 14007–14012
16. Mitra, D., Luo, X., Morgan, A., Wang, J., Hoang, M. P., Lo, J., Guerrero,C. R., Lennerz, J. K.,Mihm,M. C.,Wargo, J. A., Robinson, K. C., Devi, S. P.,Vanover, J. C., D’Orazio, J. A., McMahon, M., Bosenberg, M. W., Haigis,K. M., Haber, D. A., Wang, Y., and Fisher, D. E. (2012) An ultraviolet-
Translesion Synthesis of 8,5�-Cyclopurine-2�-deoxynucleoside
OCTOBER 4, 2013 • VOLUME 288 • NUMBER 40 JOURNAL OF BIOLOGICAL CHEMISTRY 28555

radiation-independent pathway to melanoma carcinogenesis in the redhair/fair skin background. Nature 491, 449–453
17. Cadet, J., Douki, T., Gasparutto, D., and Ravanat, J. L. (2003) Oxidativedamage toDNA. Formation,measurement and biochemical features.Mu-tat. Res. 531, 5–23
18. Wang, J., Yuan, B., Guerrero, C., Bahde, R., Gupta, S., andWang, Y. (2011)Quantification of oxidative DNA lesions in tissues of Long-Evans Cinna-mon rats by capillary high-performance liquid chromatography-tandemmass spectrometry coupled with stable isotope-dilution method. Anal.Chem. 83, 2201–2209
19. Chatgilialoglu, C., Ferreri, C., and Terzidis, M. A. (2011) Purine 5�,8-cyclonucleoside lesions: Chemistry and biology. Chem. Soc. Rev. 40,1368–1382
20. Wang, J., Clauson, C. L., Robbins, P. D., Niedernhofer, L. J., andWang, Y.(2012) The oxidative DNA lesions 8,5�-cyclopurines accumulate with ag-ing in a tissue-specific manner. Aging Cell 11, 714–716
21. Brooks, P. J. (2008) The 8,5�-cyclopurine-2�-deoxynucleosides. Candidateneurodegenerative DNA lesions in xeroderma pigmentosum, and uniqueprobes of transcription and nucleotide excision repair. DNA Repair 7,1168–1179
22. You, C., Dai, X., Yuan, B., Wang, J., Wang, J., Brooks, P. J., Niedernhofer,L. J., and Wang, Y. (2012) A quantitative assay for assessing the effects ofDNA lesions on transcription. Nat. Chem. Biol. 8, 817–822
23. Kuraoka, I., Bender, C., Romieu, A., Cadet, J., Wood, R. D., and Lindahl, T.(2000) Removal of oxygen free-radical-induced 5�,8-purine cyclodeoxy-nucleosides from DNA by the nucleotide excision-repair pathway in hu-man cells. Proc. Natl. Acad. Sci. U.S.A. 97, 3832–3837
24. Pande, P., Das, R. S., Sheppard, C., Kow, Y. W., and Basu, A. K. (2012)Repair efficiency of (5�S)-8,5�-cyclo-2�-deoxyguanosine and (5�S)-8,5�-cy-clo-2�-deoxyadenosine depends on the complementary base.DNARepair11, 926–931
25. Jasti, V. P., Das, R. S., Hilton, B. A., Weerasooriya, S., Zou, Y., and Basu,A. K. (2011) (5�S)-8,5�-Cyclo-2�-deoxyguanosine is a strong block to rep-lication, a potent pol V-dependent mutagenic lesion, and is inefficientlyrepaired in Escherichia coli. Biochemistry 50, 3862–3865
26. Brooks, P. J., Wise, D. S., Berry, D. A., Kosmoski, J. V., Smerdon, M. J.,Somers, R. L., Mackie, H., Spoonde, A. Y., Ackerman, E. J., Coleman, K.,Tarone, R. E., and Robbins, J. H. (2000) The oxidative DNA lesion 8,5�-(S)-cyclo-2�-deoxyadenosine is repaired by the nucleotide excision repairpathway and blocks gene expression in mammalian cells. J. Biol. Chem.275, 22355–22362
27. Marietta, C., Gulam, H., and Brooks, P. J. (2002) A single 8,5�-cyclo-2�-deoxyadenosine lesion in a TATA box prevents binding of the TATAbinding protein and strongly reduces transcription in vivo.DNA Repair 1,967–975
28. Abraham, J., and Brooks, P. J. (2011) Divergent effects of oxidatively inducedmodification to theC8 of 2�-deoxyadenosine on transcription factor binding.8,5�(S)-Cyclo-2�-deoxyadenosine inhibits the binding of multiple sequencespecific transcription factors,while 8-oxo-2�-deoxyadenosine increases bind-ing of CREB and NF-�B to DNA. Environ. Mol. Mutagen. 52, 287–295
29. Yuan, B.,Wang, J., Cao, H., Sun, R., andWang, Y. (2011) High-throughputanalysis of the mutagenic and cytotoxic properties of DNA lesions bynext-generation sequencing. Nucleic Acids Res. 39, 5945–5954
30. Kuraoka, I., Robins, P.,Masutani, C., Hanaoka, F., Gasparutto,D., Cadet, J.,Wood, R. D., and Lindahl, T. (2001) Oxygen free radical damage to DNA.Translesion synthesis by human DNA polymerase � and resistance toexonuclease action at cyclopurine deoxynucleoside residues. J. Biol. Chem.276, 49283–49288
31. Swanson, A. L., Wang, J., and Wang, Y. (2012) Accurate and efficientbypass of 8,5�-cyclopurine-2�-deoxynucleosides by human and yeastDNApolymerase �. Chem. Res. Toxicol. 25, 1682–1691
32. Romieu, A., Gasparutto, D., and Cadet, J. (1999) Synthesis and character-ization of oligonucleotides containing 5�,8-cyclopurine 2�-deoxyribo-nucleosides, (5�R)-5�,8-cyclo-2�-deoxyadenosine, (5�S)-5�,8-cyclo-2�-deoxyguanosine, and (5�R)-5�,8-cyclo-2�-deoxyguanosine. Chem. Res.Toxicol. 12, 412–421
33. Thakur, M., Wernick, M., Collins, C., Limoli, C. L., Crowley, E., andCleaver, J. E. (2001) DNA polymerase � undergoes alternative splicing,
protects against UV sensitivity and apoptosis, and suppresses Mre11-de-pendent recombination. Genes Chromosomes Cancer 32, 222–235
34. de Feraudy, S., Limoli, C. L., Giedzinski, E., Karentz, D., Marti, T. M.,Feeney, L., and Cleaver, J. E. (2007) Pol � is required for DNA replicationduring nucleotide deprivation by hydroxyurea. Oncogene 26, 5713–5721
35. Goodman, M. F., Creighton, S., Bloom, L. B., and Petruska, J. (1993) Bio-chemical basis of DNA replication fidelity. Crit. Rev. Biochem. Mol. Biol.28, 83–126
36. Baker, D. J., Wuenschell, G., Xia, L., Termini, J., Bates, S. E., Riggs, A. D.,and O’Connor, T. R. (2007) Nucleotide excision repair eliminates uniqueDNA-protein cross-links from mammalian cells. J. Biol. Chem. 282,22592–22604
37. Yuan, B., You, C., Andersen,N., Jiang, Y.,Moriya,M.,O’Connor, T. R., andWang, Y. (2011) The roles of DNA polymerases � and � in the error-freebypass ofN2-carboxyalkyl-2�-deoxyguanosine lesions inmammalian cells.J. Biol. Chem. 286, 17503–17511
38. Ziegler, K., Bui, T., Frisque, R. J., Grandinetti, A., and Nerurkar, V. R.(2004)A rapid in vitropolyomavirusDNAreplication assay. J. Virol.Meth-ods 122, 123–127
39. Burns, J. A., Dreij, K., Cartularo, L., and Scicchitano, D. A. (2010) O6-Methylguanine induces altered proteins at the level of transcription inhuman cells. Nucleic Acids Res. 38, 8178–8187
40. Sanchez, J. A., Marek, D., and Wangh, L. J. (1992) The efficiency andtiming of plasmid DNA replication in Xenopus eggs. Correlations to theextent of prior chromatin assembly. J. Cell Sci. 103, 907–918
41. Taylor, E. R., and Morgan, I. M. (2003) A novel technique with enhanceddetection and quantitation of HPV-16 E1- and E2-mediated DNA repli-cation. Virology 315, 103–109
42. Ziv, O., Geacintov, N., Nakajima, S., Yasui, A., and Livneh, Z. (2009) DNApolymerase � cooperates with polymerases � and � in translesion DNAsynthesis across pyrimidine photodimers in cells fromXPV patients. Proc.Natl. Acad. Sci. U.S.A. 106, 11552–11557
43. Shachar, S., Ziv, O., Avkin, S., Adar, S., Wittschieben, J., Reissner, T.,Chaney, S., Friedberg, E. C., Wang, Z., Carell, T., Geacintov, N., andLivneh, Z. (2009) Two-polymerase mechanisms dictate error-free and er-ror-prone translesion DNA synthesis in mammals. EMBO J. 28, 383–393
44. You, C., Dai, X., Yuan, B., and Wang, Y. (2012) Effects of 6-thioguanineand S6-methylthioguanine on transcription in vitro and in human cells.J. Biol. Chem. 287, 40915–40923
45. Delaney, J. C., and Essigmann, J. M. (2004) Mutagenesis, genotoxicity, andrepair of 1-methyladenine, 3-alkylcytosines, 1-methylguanine, and 3-methyl-thymine in alkB Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 101,14051–14056
46. Delaney, J. C., and Essigmann, J. M. (2006) Assays for determining lesionbypass efficiency and mutagenicity of site-specific DNA lesions in vivo.Methods Enzymol. 408, 1–15
47. Gan, G. N., Wittschieben, J. P., Wittschieben, B. Ø., and Wood, R. D.(2008) DNA polymerase � (Pol �) in higher eukaryotes. Cell Res. 18,174–183
48. Levine, R. L., Yang, I. Y., Hossain, M., Pandya, G. A., Grollman, A. P., andMoriya,M. (2000)Mutagenesis induced by a single 1,N6-ethenodeoxyade-nosine adduct in human cells. Cancer Res. 60, 4098–4104
49. Yang, I. Y., Johnson, F., Grollman, A. P., andMoriya, M. (2002) Genotoxicmechanism for the major acrolein-derived deoxyguanosine adduct in hu-man cells. Chem. Res. Toxicol. 15, 160–164
50. Newton, C. R., Graham, A., Heptinstall, L. E., Powell, S. J., Summers, C.,Kalsheker, N., Smith, J. C., and Markham, A. F. (1989) Analysis of anypoint mutation in DNA. The amplification refractory mutation system(ARMS). Nucleic Acids Res. 17, 2503–2516
51. Yoon, J. H., Prakash, L., and Prakash, S. (2010) Error-free replicative by-pass of (6-4)-photoproducts by DNA polymerase � in mouse and humancells. Genes Dev. 24, 123–128
52. Yoon, J. H., Bhatia, G., Prakash, S., and Prakash, L. (2010) Error-free replica-tive bypass of thymine glycol by the combined action of DNA polymerases �and � in human cells. Proc. Natl. Acad. Sci. U.S.A. 107, 14116–14121
53. Andersen, N., Wang, J., Wang, P., Jiang, Y., and Wang, Y. (2012) In vitroreplication studies on O2-methylthymidine and O4-methylthymidine.Chem. Res. Toxicol. 25, 2523–2531
Translesion Synthesis of 8,5�-Cyclopurine-2�-deoxynucleoside
28556 JOURNAL OF BIOLOGICAL CHEMISTRY VOLUME 288 • NUMBER 40 • OCTOBER 4, 2013
Copyright © 2022 FDOKUMEN




















